
). UN ANÁLISIS ECOFISIOLÓGICO.132.248.9.195/ptd2014/enero/0707898/0707898.pdf · 2014. 1. 28. ·...
225
UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO POSGRADO EN CIENCIAS BIOLÓGICAS FACULTAD DE CIENCIAS ECOLOGÍA ASPECTOS SOBRE EL CRECIMIENTO Y REPRODUCCIÓN DEL ACOCIL Cambarellus (Cambarellus) montezumae (SAUSSURE, 1857). UN ANÁLISIS ECOFISIOLÓGICO. TESIS QUE PARA OPTAR POR EL GRADO DE: MAESTRA EN CIENCIAS BIOLÓGICAS PRESENTA: GARCÍA PADILLA GISELA MARINA TUTOR: Dr. José Román Latournerié Cervera ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM COMITÉ TUTOR: Dr. René de Jesús Cárdenas Vázquez ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM Dr. Ignacio Méndez Ramírez ENTIDAD DE ADSCRIPCIÓN: IIMAS, UNAM MÉXICO, D. F., ENERO DE 2014
Transcript of ). UN ANÁLISIS ECOFISIOLÓGICO.132.248.9.195/ptd2014/enero/0707898/0707898.pdf · 2014. 1. 28. ·...

UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
POSGRADO EN CIENCIAS BIOLÓGICAS
FACULTAD DE CIENCIAS
ECOLOGÍA
ASPECTOS SOBRE EL CRECIMIENTO Y REPRODUCCIÓN DEL ACOCIL Cambarellus
(Cambarellus) montezumae (SAUSSURE, 1857).
UN ANÁLISIS ECOFISIOLÓGICO.
TESIS
QUE PARA OPTAR POR EL GRADO DE:
MAESTRA EN CIENCIAS BIOLÓGICAS
PRESENTA:
GARCÍA PADILLA GISELA MARINA
TUTOR:
Dr. José Román Latournerié Cervera ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM
COMITÉ TUTOR:
Dr. René de Jesús Cárdenas Vázquez ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM
Dr. Ignacio Méndez Ramírez
ENTIDAD DE ADSCRIPCIÓN: IIMAS, UNAM
MÉXICO, D. F., ENERO DE 2014

UNAM – Dirección General de Bibliotecas
Tesis Digitales
Restricciones de uso
DERECHOS RESERVADOS ©
PROHIBIDA SU REPRODUCCIÓN TOTAL O PARCIAL
Todo el material contenido en esta tesis esta protegido por la Ley Federal del Derecho de Autor (LFDA) de los Estados Unidos Mexicanos (México).
El uso de imágenes, fragmentos de videos, y demás material que sea objeto de protección de los derechos de autor, será exclusivamente para fines educativos e informativos y deberá citar la fuente donde la obtuvo mencionando el autor o autores. Cualquier uso distinto como el lucro, reproducción, edición o modificación, será perseguido y sancionado por el respectivo titular de los Derechos de Autor.


UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
POSGRADO EN CIENCIAS BIOLÓGICAS
FACULTAD DE CIENCIAS
ECOLOGÍA
ASPECTOS SOBRE EL CRECIMIENTO Y REPRODUCCIÓN DEL ACOCIL Cambarellus
(Cambarellus) montezumae (SAUSSURE, 1857).
UN ANÁLISIS ECOFISIOLÓGICO.
TESIS
QUE PARA OPTAR POR EL GRADO DE:
MAESTRA EN CIENCIAS BIOLÓGICAS
PRESENTA:
GARCÍA PADILLA GISELA MARINA
TUTOR:
Dr. José Román Latournerié Cervera ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM
COMITÉ TUTOR:
Dr. René de Jesús Cárdenas Vázquez ENTIDAD DE ADSCRIPCIÓN: FACULTAD DE CIENCIAS, UNAM
Dr. Ignacio Méndez Ramírez
ENTIDAD DE ADSCRIPCIÓN: IIMAS, UNAM
MÉXICO, D. F., ENERO DE 2014

VNIVLI\.'.DAD NAqOJ.lAL AVFftI'MA D[
Mnm:,o
Dr. Isidro Ávila Martínez
POSGRADO EN CIENCIAS BIOLÓGICAS FACULTAD DE CIENCIAS
DIVISiÓN DE ESTUDIOS DE POSGRADO
Director General de Administración Escolar, UNAM Pr ese nt e
OFICIO FCIElDEPI030114
ASUNTO: Oficio de Jurado
Me permito informar a usted que en la reunión ordinaria del Comité Académico del Posgrado en Ciencias Biológicas, celebrada el día 28 de octubre de 2013 se aprobó el siguiente jurado para el examen de grado de MAESTRA EN CIENCIAS BIOLÓGICAS del (la) alumno (a) GARciA PADILLA GISELA MARINA con numero de cuenta 303177878 con la tesis titulada "Aspectos sobre el crec imiento y reproducci ón del acocil Cambarellus (Cambarelfus) montezumae (Saussure, 1857). Un análisis ecofisiológico", realizada bajo la dirección del (la ) DR. JOSÉ ROMÁN LATOURNERIÉ CERVERA:
Presidente:
Vocal:
Secretario:
Suplente:
Suplente:
DRA. MARIA LUISA FANJUL PEÑA
DR. RAMIRO ROMÁN CONTRERAS
DR. REN~ DE JESÚS CÁRDENAS VAzaUEZ
DR. LUIS HECTOR HERNANDEZ HERNANDEZ
DR. IGNACIO M~NDEZ RAMiREZ
Sin otro particu lar, me es grato enviarle un cordial saludo.
MCAAlMJFM/ASRlmnm
Atentamente "POR MI RAZA HABLARA EL EspiRITU"
Cd. Universitaria, D.F ., a 13 de enero de 2014
Dra. María del Coro Arizmendi Arriaga Coordinadora del Programa
~"..o DE O/,
.::Jv U ~1< O C¿ Il " • ....,. . ti)
OIVIS10"'t rtl:: ~~IJDfOS DE l· ..... "J

AGRADECIMIENTOS.
En primer lugar quisiera agradecer al Posgrado en Ciencias Biológicas de
la Universidad Nacional Autónoma de México, por haberme dado la
oportunidad de formarme tanto en el ámbito profesional como personal, por
haberme provisto de excelentes profesores, quienes me brindaron su
conocimiento y confianza en el transcurso de mis estudios.
Al Programa de Becas de Posgrado del Consejo Nacional de Ciencia y
Tecnología (CONACYT), por apoyarme económicamente durante mis estudios
de maestría y permitir hacer de este sueño una realidad.
Quisiera expresar mi más sincero agradecimiento a mi tutor el Dr. José
Román Latournerié Cervera por su ayuda, apoyo, paciencia y enseñanzas,
quien además de ser el director de esta tesis, es sin duda, una de las personas
que más ha contribuido en mi formación profesional. Por todos los años y lo
compartido, GRACIAS.
A los miembros de mi comité tutoral, mi reconocimiento y gratitud por todas las
enseñanzas que tan generosamente me han brindado, por llevarme de la mano
en la realización de esta investigación, al Dr. René de Jesús Cárdenas y al
Dr. Ignacio Méndez Ramírez.

Un agradecimiento muy especial a los miembros del jurado que revisaron esta
tesis: Dra. María Luisa Fanjul Peña, Dr. Ramiro Román Contreras y Dr.
Luis Héctor Hernández Hernández, quienes aportaron importantes
sugerencias para la culminación de este trabajo y sobre todo me brindaron su
apoyo y compresión en el proceso de titulación.
Al Dr. Fernando Arana Magallón y al Centro de Investigaciones
Biológicas y Acuícolas de Cuemanco (CIBAC), por el apoyo para la
realización del monitoreo del hábitat del acocil Cambarellus (C.) montezumae
en el lago de Xochimilco 2009 - 2012.
A la M en C. Yamel Nacif Osorio que con el paso del tiempo no solo se
convirtió en un ejemplo a seguir, sino en una GRAN AMIGA. Gracias por el
apoyo en esos días eternos de respirometría y de salidas al campo, por los
tantos consejos académicos y personales que me regalaste, pero sobre todo por
confiar en mi y ser parte ESENCIAL en vida.
A todos los miembros del Laboratorio de Acuacultura y Producción
Acuática, que de alguna manera han dejado huella en mi corazón, pero sobre
todo a Felipe Rojas Parra, a Silvia Hansen Bernal y a Erika Escalante
Garnelo por todo el apoyo que me han brindado, tanto en el trabajo de campo
como en el trabajo de laboratorio, sin ustedes este trabajo no sería una
realidad, gracias por los buenos momentos que hemos pasado trabajando,

viviendo y siendo cómplices en nuestras travesuras, POR SU AMISTAD Y
CARIÑO, GRACIAS.
A el Arq. Andrés Celis García por los días de pocilga que me hicieron
TANTO BIEN y por todos los momentos e historias compartidas, GRACIAS.
A la M. en C. Nubia Caballero Mendieta por ser mi primera Jefa y la mejor,
gracias por el apoyo en los momentos difíciles y durante el proceso de
titulación.
A la Universidad Nacional Autónoma de México, que siempre la llevare en
mi corazón a donde quiera que vaya, gracias por la oportunidad de formarme
como persona y como profesionista, gracias por permitirme hacer de mi hobby
mi trabajo.
Como no te voy a querer. . .

DEDICATORIA.
A mis padres:
A Zac = Piscis en maya.
Un hijo es un ser prestado para hacer un curso intensivo de cómo amar a alguien
más que a nosotros mismos y de cómo cambiar nuestros peores defectos para darle
los mejores ejemplos. Sí. ¡Eso es! Ser padre es el mayor acto de coraje que alguien
pueda tener, porque es exponerse a todo tipo de dolor, principalmente de la
incertidumbre de estar actuando correctamente y del miedo a perder algo tan
amado. Quiero que sepan que ambos lo hicieron maravillosamente, porque con
sus enseñanzas, amor y confianza, fortalecieron mi vida. Porque siempre
estuvieron para mí con palabras de apoyo, que me ayudaron a seguir adelante
(principalmente en estos últimos meses). Por todo esto, quiero que sientan que el
objetivo logrado, también es suyo, porque con su esfuerzo y sacrificio, logré el
triunfo que hoy les brindo. Con cariño y admiración. . .
Sólo quisiera ser uno de los motivos de tu sonrisa, quizá un
pequeño pensamiento de tu mente durante la mañana, o quizá
un lindo recuerdo antes de dormir. Sólo quisiera ser una fugaz
imagen frente a tus ojos, quizá una voz susurrante en tu oído, o
quizá un leve roce en tus labios. Sólo quisiera ser alguien que
quisieras tener a tu lado, quizá no durante todo el día, pero de
una u otra forma, vivir en ti.
GM

ÍNDICE
Agradecimientos
i
Dedicatoria
ii
Resumen
iii
Abstract
iv
1. Marco de referencia
1
2. Aspectos biológicos de la especie
5
2.1 Clasificación taxonómica
5
2.2 Morfología externa
6
2.3 Morfología interna
8
2.3.1 Sistema nervioso
8
2.3.2 Sistema circulatorio
8
2.3.3 Sistema respiratorio
9
2.3.4 Sistema excretor
9
2.3.5 Aparato digestivo
10
2.3.6 Aparato reproductivo
11
2.4 Ecdisis o proceso de muda
14
2.5 Reproducción
14
2.6 Hábitat y distribución natural
16
2.7 Alimentación
17
2.8 Requerimientos ambientales
18
2.9 Interacciones ecológicas
18
2.10 Valor nutrimental
19
3. Antecedentes
20
4. Justificación general
38

5. Objetivo general
38
6. CAPÍTULO 1. Estrategia reproductiva del acocil Cambarellus (Cambarellus)
montezumae (Saussure, 1857), en el Lago de Xochimilco, México D.F.
6.1 Introducción
39
6.2 Justificación
45
6.3 Objetivos
45
6.3.1 Objetivo general
45
6.3.2 Objetivos particulares
46
6.4 Material y método
47
6.4.1 Área de estudio
47
6.4.2 Trabajo de campo
53
6.4.2.1 Estacionalidad
54
6.4.2.2 Caracterización del hábitat 55
6.4.2.3 Colecta de los organismos 55
6.4.3 Trabajo de laboratorio
56
6.4.3.1 Caracterización de las colectas 56
6.4.3.1.1 Estructura poblacional 57
6.4.3.1.2 Proporción sexual 58
6.4.3.1.3 Relación peso-longitud 59
6.4.3.2 Indicadores del desempeño reproductivo 60
6.4.3.2.1 Época reproductiva 60
6.4.3.2.2 Índice de desove 60
6.4.3.2.3 Fecundidad 60
6.4.3.2.4 Esfuerzo reproductivo 61
6.4.3.3 Indicadores de la calidad del huevo 62
6.4.3.3.1 Volumen
62
6.4.3.3.2 Composición químico proximal 63
6.4.3.4 Desarrollo embrionario 63
6.4.3.4.1 Tiempo de incubación 63
6.4.2.4.2 Tamaño de las crías 64

6.4.3.5 Respuesta fisiológica de las hembras ovígeras 65
6.5 Resultados
67
6.5.1 Caracterización del hábitat
67
6.5.2 Caracterización de las colectas 70
6.5.3 Estructura poblacional
73
6.5.4 Proporción sexual
75
6.5.5 Relación Peso-Longitud
76
6.5.6 Indicadores de desempeño reproductivo 79
6.5.6.1 Época reproductiva
79
6.5.6.2 Índice de desove
81
6.5.6.3 Fecundidad
82
6.5.6.4 Esfuerzo reproductivo 86
6.5.7 Indicadores de la calidad del huevo 88
6.5.7.1 Volumen del huevo
89
6.5.7.2 Composición químico proximal del huevo 90
6.5.8 Desarrollo embrionario
93
6.5.8.1 Tiempo de incubación 94
6.5.8.2 Tamaño de las crías
95
6.5.9 Respuesta fisiológica de las hembras ovígeras 96
6.6 Discusión
97
6.7 Conclusiones
111
7. CAPÍTULO 2. Efecto de una dieta formulada sobre el crecimiento,
sobrevivencia y calidad de la puesta en crías de acocil Cambarellus
(Cambarellus) montezumae (Saussure, 1857), en condiciones controladas
7.1 Introducción
113
7.2 Justificación
118
7.3 Objetivos
118
7.3.1 Objetivo general
118
7.3.2 Objetivos particulares
119

7.4 Material y método
120
7.4.1 Trabajo de campo
120
7.4.1.1 Caracterización del hábitat y colecta de los organismos 120
7.4.2 Trabajo de laboratorio
121
7.4.2.1 Elaboración de las dietas 121
7.4.2.2 Análisis químico proximal de las dietas 122
7.4.2.3 FASE I. Crecimiento y sobrevivencia 123
7.4.2.3.1 Diseño experimental 123
7.4.2.3.2 Crecimiento y sobrevivencia 124
7.4.2.3.3 Tasa Instantánea de Crecimiento y Producción 126
7.4.2.4 FASE II. Indicadores de desempeño reproductivo 127
7.4.2.4.1 Diseño experimental 127
7.4.2.4.2 Talla mínima de madurez 128
7.4.2.4.3 Índice de desove 128
7.4.2.4.4 Fecundidad 129
7.4.2.4.5 Calidad de los reproductores 129
7.4.2.5 FASE III. Indicadores de la calidad de la puesta 132
7.4.2.5.1 Volumen y peso del huevo 133
7.4.2.5.2 Composición químico proximal de la puesta 133
7.4.2.5.1 Desarrollo embrionario 134
7.5 Resultados
136
7.5.1 Análisis de la calidad del agua 136
7.5.2 FASE I. Crecimiento y sobrevivencia 137
7.5.2.1 Crecimiento y sobrevivencia 138
7.5.2.2 Tasa Instantánea de Crecimiento y Producción 139
7.5.3 FASE II. Indicadores de desempeño reproductivo 142
7.5.3.1 Talla mínima de madurez 142
7.5.3.2 Índice de desove
144
7.5.3.3 Fecundidad
145
7.5.3.4 Calidad de los reproductores 147
7.5.4 FASE III. Indicadores de la calidad de la puesta 156
7.5.4.1 Volumen y peso del huevo 157

7.5.4.2 Composición químico proximal de la puesta 159
7.5.4.3 Desarrollo embrionario 160
7.6 Discusión
164
7.7 Conclusiones
176
8. Literatura citada
178

Cuadro
Página
1
Valor nutrimental del acocil Cambarellus (C.) montezumae de la
presa de Atlangatepec, Tlaxcala.
19
2
Distribución media mensual de la temperatura ambiente en la zona
lacustre de Xochimilco.
54
3
Características de cada edad ecológica del acocil Cambarellus (C.)
montezumae.
58
4
Características de los estadios de desarrollo embrionario del acocil
Cambarellus (C.) montezumae.
64
5
Parámetros físico-químicos del hábitat (Canal de Cuemanco) del
acocil Cambarellus (C.) montezumae en época cálida y fría del año.
68
6
Número de organismos del acocil Cambarellus (C.) montezumae
colectados mensualmente en el Canal de Cuemanco.
72
7
Abundancia de organismos del acocil Cambarellus (C.) montezumae
por edad ecológica en época cálida y fría del año.
74
8
Modelo potencial de crecimiento del acocil Cambarellus (C.)
montezumae por sexo y época del año.
76
9
Abundancia de hembras adultas del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
81
10
Características morfométricas y número de huevos de hembras
ovígeras del acocil Cambarellus (C.) montezumae colectas en época
cálida y fría del año.
83
11
Volumen y peso húmedo del huevo del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
89
12
Contenido químico proximal de los huevos del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
91

13
Contenido químico proximal de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.
92
15 Análisis químico proximal de las dietas.
122
16
Parámetros físico-químicos promedio registrados durante la fase
experimental para cada tratamiento.
136
17
Crecimiento (LT, LCT y Ph) y sobrevivencia (%) de crías de acocil
Cambarellus (C.) montezumae durante un periodo de 60 días.
139
18
Valores de TIC durante 60 días para crías de acocil Cambarellus (C.)
montezumae.
140
19
Producción (formación de tejido corporal en mg) de las crías del acocil
Cambarellus (C.) montezumae alimentados con la dieta formulada y
la dieta control durante 60 días.
141
20
Número de desoves de las hembras del acocil Cambarellus (C.)
montezumae alimentadas con la dieta formulada y la dieta control.
143
21
Índice de Desove las hembras del acocil Cambarellus (C.)
montezumae alimentadas con la dieta formulada y la dieta control.
144
22
Características morfométricas y número de huevos de hembras
ovígeras de Cambarellus (C.) montezumae para ambas dietas.
146
23
Índices morfométricos por tratamiento y sexo del acocil Cambarellus
(C.) montezumae al termino de los 150 días de experimentación.
148
24
Análisis químico proximal de las reproductores del acocil
Cambarellus (C.) montezumae al final del periodo experimental (150
días).
150
25
Gasto de energía promedio por tasa metabólica para ambos sexos y
tratamientos del acocil Cambarellus (C.) montezumae.
153

26
Gasto de energía promedio por excreción nitrogenada para ambos
sexos y tratamientos (Dieta formulada y control) del acocil
Cambarellus (C.) montezumae.
155
27
Volumen y peso húmedo del huevo de hembras alimentadas con la
dieta formulada y la dieta control del acocil Cambarellus (C.)
montezumae.
157
28
Composición químico proximal de los huevos de hembras alimentadas
con la dieta formulada y la dieta control del acocil Cambarellus (C.)
montezumae.
159
29
Sobrevivencia a los 15 días de crías de hembras alimentadas con la
dieta formulada y la dieta control del acocil Cambarellus (C.)
montezumae.
163
Figura
Página
1 Características biométricas del acocil Cambarellus (C.) montezumae.
56
2 Temperatura media mensual en el Canal de Cuemanco.
69
3 Oxígeno disuelto medio mensual en el Canal de Cuemanco.
69
4
Potencial de hidrogeno (pH) medio mensual en el Canal de
Cuemanco.
70
5
Abundancia del acocil Cambarellus (C.) montezumae en época cálida
y fría del año en el Canal de Cuemanco.
71
6
Número de organismos del acocil Cambarellus (C.) montezumae
colectados mensualmente en el Canal de Cuemanco.
72
7
Regresión lineal entre la temperatura hábitat y la abundancia del
acocil Cambarellus (C.) montezumae.
73
8 Abundancia de organismos por sexo del acocil Cambarellus (C.)
montezumae en época cálida y fría. 75

9
Modelo potencial de crecimiento del acocil Cambarellus (C.)
montezumae en época cálida (a=hembras y b=machos).
77
10
Modelo potencial de crecimiento del acocil Cambarellus (C.)
montezumae en época fría (a=hembras y b=machos).
78
11
Número de hembras ovígeras del acocil Cambarellus (C.) montezumae
colectadas mensualmente en el Canal de Cuemanco.
80
12
Relación entre temperatura del hábitat y la abundancia mensual de
hembras ovígeras del acocil Cambarellus (C.) montezumae.
80
13
Número total de huevos de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.
84
14
Relación del número de huevos respecto al tamaño de la hembra
ovígera del acocil Cambarellus (C.) montezumae.
85
15
Relación del número de huevos respecto a la amplitud del abdomen
de una hembra ovígera del acocil Cambarellus (C.) montezumae.
86
16
Relación entre el peso de la masa ovígera respecto al peso de la
hembra del acocil Cambarellus (C.) montezumae.
87
17
Peso de masa ovígera del acocil Cambarellus (C.) montezumae en
época cálida y fría del año.
88
18
Volumen del huevo del acocil Cambarellus (C.) montezumae en época
cálida y fría del año.
90
19
Contenido químico proximal de los huevos del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
91
20
Contenido químico proximal de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.
92
21 Desarrollo embrionario del acocil Cambarellus (C.) montezumae. 93

22
Tiempo de incubación del acocil Cambarellus (C.) montezumae en
época cálida y fría del año.
94
23
Peso de crías de época cálida y fría del año del acocil Cambarellus (C.)
montezumae.
95
24
Consumo de oxígeno de hembras ovígeras y no ovígeras del acocil
Cambarellus (C.) montezumae.
96
25
Diseño experimental para evaluar el crecimiento y sobrevivencia de
crías del acocil Cambarellus (C.) montezumae.
124
26 Características biométricas del acocil Cambarellus (C.) montezumae.
125
27
Diseño experimental para evaluar los indicadores del desempeño
reproductivo y la calidad de la puesta del acocil Cambarellus (C.)
montezumae.
126
28
Índices morfométricos registrados: LT, LCT y Ph de crías del acocil
Cambarellus (C.) montezumae en un periodo de 60 días.
138
29
TIC calculada para crías del acocil Cambarellus (C.) montezumae
alimentados con la dieta formulada y la dieta control.
141
30
Evolución del número de desoves de las hembras del acocil
Cambarellus (C.) montezumae alimentadas con la dieta formulada y
la dieta control.
143
31
Número de huevos portados por hembra del acocil Cambarellus (C.)
montezumae alimentadas con la dieta formulada y la dieta control.
147
32
Análisis discriminante (Modelo completo: LT, LCT, Ph, MO, MT,
Lípidos, Proteínas y Contenido calórico).
151
33
Análisis discriminante (Modelo reducido: Lípidos y contenido
calórico).
151

34
Gasto de energía promedio por tasa metabólica para ambos sexos y
tratamientos (Dieta formulada y control) del acocil Cambarellus (C.)
montezumae.
153
35
Gasto de energía promedio por excreción nitrogenada para ambos
sexos y tratamientos del acocil Cambarellus (C.) montezumae.
155
36
Volumen del huevo de hembras alimentadas con la dieta formulada y
la dieta control del acocil Cambarellus (C.) montezumae.
157
37
Peso húmedo del huevo de hembras alimentadas con dieta formulada
y dieta control del acocil Cambarellus (C.) montezumae.
158
38
Composición químico proximal de los huevos de hembras alimentadas
con la dieta formulada y la dieta control del acocil Cambarellus (C.)
montezumae.
160
39
Tiempo de incubación de hembras alimentadas con dieta formulada y
la dieta control del acocil Cambarellus (C.) montezumae.
161
40 Peso de crías de hembras alimentadas con la dieta formulada y la
dieta control del acocil Cambarellus (C.) montezumae. 162

iii
RESUMEN.
La presente investigación consta de dos capítulos, el primero de ellos trata
sobre la estrategia reproductiva del acocil Cambarellus (Cambarellus)
montezumae en el Lago de Xochimilco y el segundo, versa sobre el efecto de una
dieta formulada sobre el crecimiento, sobrevivencia y calidad de la puesta en
crías de este acocil bajo condiciones controladas.
En el primer capítulo, para determinar la estrategia reproductiva de la especie,
se llevó a cabo un muestreo anual en el Lago de Xochimilco, esto con el fin de
determinar las variaciones estacionales de la poblacional en cuanto a su
estructura poblacional y a su ecología reproductiva. Se encontró que
Cambarellus (C.) montezumae es una especie que se adapta a diferentes
condiciones ambientales; en los canales de Xochimilco este acocil encuentra
condiciones óptimas para su desarrollo, ya que los parámetros fisicoquímicos
del agua (temperatura, oxígeno y pH), se encuentran entre los valores de
tolerancia para la especie. La población de este acocil en el Lago de Xochimilco
es de tipo joven, ya que presenta una elevada proporción de crías y juveniles, lo
que confiere a la población una capacidad latente para su rápido crecimiento,
siempre y cuando existan las condiciones medioambientales apropiadas para su
desarrollo. Su estrategia reproductiva muestra que existe una época
reproductiva bien definida, la cual se lleva a cabo en la época cálida del año,
entre los meses de abril, mayo y junio principalmente, cuando la idoneidad de
las hembras es mayor para la reproducción reflejándose en la fecundidad, el
índice de desove y calidad de los huevos y crías.
En el segundo capítulo el objetivo principal de la investigación fue cuantificar
el efecto de una dieta formulada sobre la producción de este acocil, con la que se
pretendía mejorar el crecimiento, la sobrevivencia, la reproducción, la calidad

iii
de los reproductores y la puesta, por lo cual se decidió aumentar los niveles de
proteínas y ácidos grasos en la dieta, mediante el uso de harina de calamar y
aceite de pescado. Encontramos que existe un efecto diferencial en cuento a la
calidad de los reproductores y las crías de este acocil alimentadas con la dieta
formula, ya que los organismos alimentados con dicha dieta tuvieron una
menor edad mínima de primera madurez, así como un mayor índice de desove,
fecundidad, calidad de los huevos, sobrevivencia y tamaño de las crías.
Estos resultados pueden servir de base para el desarrollo de un sistema de
cultivo a nivel intensivo, ya que conocer los mecanismos de reproducción son
básicos y previos al funcionamiento de cualquier explotación acuícola, cuyo
objetivo principal es maximizar la producción de los organismos útiles, a través
de la producción de una progenie numerosa, saludable y homogenea.

iii
ABSTRACT.
The current research consists of two chapters, the first of which deals with the
reproductive strategy of the crayfish Cambarellus (Cambarellus) montezumae in the
Xochimilco Lake, and the second is about the effect of a formulated diet on growth,
survival and quality of the reproduction of this crayfish under controlled conditions.
In the first chapter, in order to determine the reproductive strategy of the species, an
annual sampling was collected in Xochimilco Lake, with the purpose of determining
the seasonal variations of the population regarding their population structure and
reproductive ecology. We found that Cambarellus (C.) montezumae is a species
adapting to different environmental conditions. Moreover, we found that this crayfish
found optimal conditions for its development in the canals of Xochimilco, as the
physicochemical parameters of the water (temperature, oxygen, and pH) are within
the tolerance values for the species. The population of the crayfish in Xochimilco Lake
is young, since it presents a high proportion of breeds and young specimens, which
gives the population a latent capacity for rapid growth as long as appropriate
environmental conditions for its development exist. Its reproductive strategy shows
that there is a well-defined reproductive season, which takes place in the warm
season, mainly in the months of April, May and June, when the fitness of females is
higher for reproduction, which is reflected in the fertility, the spawning rate, and
quality of eggs and breeds.
In the second chapter, the main objective of this research was to quantify the effect of
a formulated diet on the production of this crayfish. This diet was intended to improve
the growth, survival, reproduction, and quality of breeders and increase the spawning
rate. Therefore, it was decided to increase the protein and fatty acid levels in the diet
through the use of squid flour and fish oil.

iii
We found that there is a differential effect with regard to the quality of the breeders
and the breeds of this crayfish when they are fed with the formulated diet, since the
organisms fed with this diet had a lower minimum age at first maturity and a higher
spawning rate, fecundity, egg quality, survival, and breed size.
These results can serve as a basis for the development of an aquaculture system at
intensive level, as to know the reproductive mechanisms is critical and necessary to
start the operation of any aquaculture exploitation, whose main objective is to
maximize the production of useful organisms through the production of large, healthy,
and uniform offspring.

1
1. MARCO DE REFERENCIA.
México es un país multifacético, plural y diverso en numerosos aspectos. El
rasgo más distintivo del país es su gran heterogeneidad ambiental, albergamos
en nuestro territorio infinidad de variados paisajes, además de muchas y
singulares culturas. De la misma manera su inmejorable ubicación geográfica,
hace que nuestro país sea poseedor de una gran riqueza de recursos naturales,
con gran diversidad de flora y fauna tanto terrestres como acuáticas, las cuales
muestran en muchos casos un marcado endemismo, cuya preservación y
estudio resulta ser una necesidad actual y primordial, así como un serio
compromiso con las generaciones venideras, esto en el marco de la
sustentabilidad ambiental (Capital Natural, 2009).
La sustentabilidad ambiental se refiere a la administración eficiente y racional
de los recursos naturales, de manera tal, que sea posible mejorar el bienestar
de la población actual sin comprometer la calidad de vida de las generaciones
futuras. Uno de los principales retos que enfrenta México es incluir al medio
ambiente como uno de los elementos de la competitividad y el desarrollo
económico y social. Desafortunadamente, los esfuerzos de conservación de los
recursos naturales y ecosistemas suelen verse obstaculizados por un círculo
vicioso que incluye pobreza, agotamiento de los recursos naturales, deterioro
ambiental y más pobreza (Plan Nacional de Desarrollo, 2007-2012).
Indudablemente México enfrenta grandes retos en todos los aspectos de la
agenda ambiental. Esta agenda comprende temas fundamentales como la
conciliación de la protección del medio ambiente (la mitigación del cambio
climático, la reforestación de bosques y selvas, la conservación, uso del agua y
del suelo, la preservación de la biodiversidad, el ordenamiento ecológico y la

2
gestión ambiental) con la competitividad de los sectores productivos y con el
desarrollo social. Estos temas pueden atenderse desde tres grandes líneas de
acción, que son:
1. Aprovechamiento sustentable de los recursos naturales.
2. Protección del medio ambiente.
3. Educación y conocimiento para la sustentabilidad ambiental.
Desde la línea de acción del aprovechamiento sustentable de los recursos
naturales, en los últimos años ha sido necesario buscar fuentes que impulsen el
desarrollo de actividades que contribuyan con el crecimiento económico y social
del país, lo cual va de acuerdo con el Plan Nacional de Desarrollo 2007 – 2012
(PND), en el cual se considera estratégico establecer condiciones para que
México se inserte en la vanguardia tecnológica. Asimismo, el PND señala que
hay que aprovechar las nuevas tecnologías (biotecnología) y contribuir al
desarrollo de las mismas, lo cual no sólo implicaría un avance estructural sino
que repercutiría en un incremento en la competitividad de la economía
mexicana.
En este sentido la acuicultura se ha convertido en la actividad de producción de
alimentos con mayor crecimiento a nivel mundial en los últimos años. La
importancia de cubrir las necesidades alimentarias de la población aumenta
rápidamente y con ello, la necesidad de hacerlo de una manera sustentable.
En México la alimentación es un tema de alta prioridad; por un lado, se tiene el
problema de la desnutrición que prevalece en el sector de la población en
extrema pobreza y por otro lado, está el problema de la obesidad que existe en
gran parte de la población y con la cual se ha alcanzado el primer lugar de
obesidad infantil a nivel mundial (UNICEF, 2012).

3
Estos problemas pueden ser parcialmente compensados a través de un
incremento sustancial en el consumo de productos acuáticos. En la actualidad
México cuenta con varias especies de crustáceos y moluscos que están siendo
utilizadas en prácticas acuícolas como es el caso del “camarón” y el
“langostino”, las “ostras” y los “ostiones”, entre otras. Sin embargo, existen
especies que por sus características podrían emplearse en estas prácticas y a
las cuales se les ha puesto poca atención, tal es el caso del “acocil” o “cangrejo
de rio” (Huner, 1981; Huner & Avault, 1985; Holdich & Lowery, 1988; Villareal,
1991; Rodríguez-Serna, 1991).
De acuerdo con la Carta Nacional Acuícola (2012) elaborada por el INAPESCA,
el acocil Cambarellus (Cambarellus) montezumae (Saussure, 1857), es un
crustáceo decápodo de agua dulce susceptible a ser cultivado en el campo de la
acuicultura, por lo cual se ha considerado como una especie endémica con
potencial acuícola de alto valor nutricional.
Su valor nutricional se relaciona principalmente con la cantidad y calidad de
proteínas y lípidos que presenta, que tienen una influencia positiva en el
desarrollo del ser humano por su participación en el desarrollo muscular, la
regeneración de tejidos y enzimas, debido a su contenido de aminoácidos
esenciales, además de ser una fuente rica de ácidos grasos, principalmente del
tipo de los poliinsaturados (dentro de este grupo se encuentra el ácido
linolénico [omega 3] y el linoleico [omega 6]), los cuales son beneficiosos para el
ser humano (Monroy, 2011).
Asimismo, esta especie es de rápido crecimiento y se reproduce durante todo el
año con elevadas tasas de supervivencia, lo que hace posible generar suficientes
volúmenes para atender la demanda del mercado. El consumo de un producto

4
como este podría contribuir a disminuir problemas asociados a la desnutrición,
obesidad y a una alimentación desbalanceada.
Por todo lo anterior, surge el interés y la necesidad de estudiar aspectos sobre
el crecimiento y la reproducción de esta especie, lo cual nos puede ayudar a
sentar las bases para el desarrollo de un paquete tecnológico (protocolo de
producción), con el fin de facilitar la transferencia del conocimiento científico a
la sociedad y de esta manera, introducir esta especie en el mercado y ofrecer
una alternativa proteica de alta calidad y bajo costo.

5
2. ASPECTOS BIOLÓGICOS DE LA ESPECIE.
2.1 Clasificación taxonómica.
La especie Cambarellus (Cambarellus) montezumae pertenece a la familia
Cambaridae caracterizada por un dimorfismo cíclico en los machos, esta familia
colonizó las agua dulces de Norteamérica con dos subfamilias: Cambarellinae y
Cambarinae (Hobbs, 1989). En los cuerpos de agua continentales de nuestro
país la familia Cambaridae se encuentra representada hasta el momento por 57
especies, entre las que se encuentra la especie Cambarellus (C.) montezumae,
que fue recolectada en el lago de Chapultepec y descrita por primera vez para
la ciencia por Henry de Saussure en 1857 (Lám. 1).
Lámina 1. Acocil Cambarellus (Cambarellus) montezumae (Saussure, 1857).

6
Según DeGrave, et al., (2009) su clasificación taxonómica es:
PHYLUM Arthropoda
CLASE Crustacea
SUBCLASE Malacostraca
SUPERORDEN Eucarida
ORDEN Decápoda (Latreille, 1802)
SUBORDEN Pleocyemata (Burkenroad, 1953)
INFRAORDEN Astacidea (Latreille, 1802)
SUPERFAMILIA Astacoidea (De Haan, 1841)
FAMILIA Cambaridae (Hobbs, 1942)
GENERO Cambarellus (Ortmann, 1905)
ESPECIE Cambarellus (C.) montezumae (Saussure, 1857)
2.2 Morfología externa.
Cambarellus (C.) montezumae es un pequeño crustáceo decápodo de 30 a 50
mm de longitud total, con dimorfismo sexual claro ya que las hembras son de
mayor tamaño que los machos, aunque estos son de aspecto más robusto.
Su cuerpo se encuentra segmentado en tres tagmas: cabeza y tórax (cefalotórax)
y abdomen, el primero incluye los ojos pedunculados, el aparato bucal, la
cámaras branquiales y los sistemas: nervioso, circulatorio, digestivo y
reproductivo; en el abdomen se localizan los pleópodos, el ano y el telson.
Presentan cinco pares de apéndices torácicos, tres de ellos quelados, los cuales
se encuentran modificados para la defensa y el ataque, atrapar el alimento,
denominados quelípedos; y los otros dos pares de apéndices son más delgados y
cortos, sirven de locomoción, manipulación del alimento y limpieza; esto diez
apéndices se conocen como pereiópodos. Los apéndices abdominales se conocen

7
como pleópodos y sirven a la hembra para el transporte e incubación de los
huevos (Pennak, 1978), (Lám. 2).
Lámina 2. Morfología externa del acocil Cambarellus (C.) montezumae.
Cambarellus (C.) montezumae presenta dimorfismo sexual, en donde las
hembras se distinguen de los machos por ser de mayor tamaño, presentan un
abdomen más ancho y corto, sus quelas son cortas y anchas en contraparte con
las quelas de los machos que son largas y delgadas. Al ser más ancho el
abdomen de las hembras les permite una mayor superficie para portar los
huevecillos (Villalobos, 1955; Civera y Cruz, 1982).
En el macho el primer par de pleópodos esta transformado en gonópodos
copuladores, también se presentan pequeños ganchos llamados isquiopoditos
del segundo y tercer par de pereiópodos. Los gonópodos se encuentran en el
quinto par de pereiópodos y sus dos primeros pleópodos están modificados para
efectuar la cópula; el primer par puede tener dos formas: la forma I o madurez
sexual, la cual se caracteriza por presentar la punta córnea y más quitinizada,

8
y la forma II o de inmadurez sexual. Los machos pueden alternar las formas (I
y II) por medio de la muda. El segundo par de pleópodos es más largo y
contribuye para que los espermatóforos sean depositados alrededor del annulus
ventralis. Las hembras presentan los gonópodos en los coxopoditos de los
terceros pereiópodos y una estructura en forma de herradura denominada
annulus ventralis, ubicada entre el cuarto y quinto par de apéndices torácicos
(Villalobos, 1955; Civera y Cruz, 1982; Morones, 1991).
2.3 Morfología interna (Morones, 1991).
2.3.1 Sistema nervioso.
El sistema nervioso tiene una semejanza general a la lombriz de tierra, pero es
relativamente mayor; está formado por un par de ganglios cereboides o masa
cereboide, un collar esofágico, un ganglio infraesofágico y una cadena ventral
de ganglios; los ganglios supraesofágicos (cerebro) ubicados en la cabeza envían
nervios a los ojos, anténulas y antenas, y un par de conectivos las unen con los
ganglios subesofágicos situados detrás de la boca, en el extremo anterior del
cordón nervioso ventral.
A lo largo del cordón nervioso cada somita posee un par de ganglios fusionados
que envían nervios a los apéndices, músculos y otros ganglios. De la cadena
ventral parten nervios que van a inervar a los diferentes órganos del cuerpo.
Las fibras nerviosas gigantes surgen de los cuerpos celulares del cerebro y
contribuyen al reflejo de la huida rápida de este decápodo.

9
2.3.2 Sistema circulatorio.
Consta de un corto corazón muscular de forma irregular y de contorno
poligonal, y se halla suspendido y colocado en la región dorsal del tórax
mediante seis ligamentos que se insertan en las paredes del seno pericárdico
dorsal medio. La hemolinfa, oxigenada, incolora o ligeramente coloreada, es de
un tono verdoso que le comunica la hemocianina. En ella existen corpúsculos
celulares amibiodeos o amebocitos.
Al contraerse el corazón la hemolinfa es expulsada y corre por varias (seis)
arterias que se ramifican y llegan a diversos espacios o senos venosos que se
encuentran entre los órganos del cuerpo. Cuando la hemolinfa ha cedido su
oxígeno y recogido dióxido de carbono de las células, es conducida por varios
senos desde el seno esternal a las branquias en donde se oxigena y de allí
vuelve por el seno braquiocárdico al pericárdico y al corazón.
2.3.3 Sistema respiratorio.
La respiración se efectúa por varios pares de branquias, son delicadas
expansiones plumiformes de la pared del cuerpo, colocadas dentro de las
cámaras branquiales situadas a uno y otro lado de la región toráxica, contienen
conductos sanguíneos; la cámara branquial se encuentra cubierta por una
expansión del caparazón llamada branquiostegito, pero se abre ventralmente
en los dos extremos. Las regiones branquiales del caparazón que se extiende
sobre las branquias están separadas de la región cardiaca o areola, por los
surcos branquicardícos.

10
2.3.4 Sistema excretor.
Consta de un par de órganos bastante complicado llamado glándula verde,
situados en la región ventral de la cabeza y delante del esófago, sirven para
eliminar las substancias de desecho de la sangre y de los líquidos del cuerpo.
Cada glándula verde consta de una porción glandular de color verde que se
encarga de recoger los productos de excreción que lleva la hemolinfa que son
conducidos a una vejiga que las almacena y de un corto tubo que los expulsa al
exterior y que se abre por un orificio que está situado en la base del primer
artejo de la entena externa o antena del segundo par.
2.3.5 Aparato digestivo.
El aparato digestivo comprende la parte que se abre por encima de las
mandíbulas en la parte ventral de la cabeza y que continua con un esófago
corto tubular que termina en el estómago situado en el tórax, y que ocupa gran
parte de la porción cefálica. El estómago se divide en una porción ancha de
gran tamaño o cámara cardíaca, y otra más pequeña posterior o cámara
pilórica. En la porción anterior del estómago se encuentra un aparato
masticador, el molino gástrico, formado de piezas quitinosas calcificadas
denominadas oscículos. En la entrada de la cámara pilórica se localizan
numerosas cerdas parecidas a pelos, que solo permiten el paso de las partículas
finas y actúan como filtro.
El estómago no funciona como órgano de la digestión, sino como triturador
(masticador y filtrador) del alimento. Después se encuentra el intestino, que
es un tubo que se extiende en toda la longitud del resto del cuerpo y termina en
el ano (casi en la base del telson), en el que actúan los jugos digestivos. En el

11
tórax y a los lados del estómago se encuentran dos glándulas digestivas o
hepatopáncreas, con tres lóbulos a cada lado conteniendo muchos tubos que
vierten una secreción amarillenta en la parte pilórica del estómago
contribuyendo a la digestión de las sustancias nutritivas.
El alimento pasa a la boca mediante las quelas del segundo y tercer par de
patas en donde las mandíbulas lo parten en pequeños fragmentos, el alimento
es triturado y después de pasar por el esófago van a la cámara cardiaca, la cual
presenta salientes en las que se ubican los dientes fuertemente calcificados,
uno mediano y dos laterales movidos por músculos insertos fuera del estómago
que forman el molinillo gástrico que tritura más el alimento.
El hepatopáncreas secreta enzimas digestivas que almacenan glucógeno, grasa,
calcio y conjuntamente con el intestino medio, absorbe el alimento digerido.
Las partículas finas son empujadas por la boca o el molinillo gástrico al
intestino medio; los materiales no digeridos se transforman en heces en el
intestino para ser expulsadas por el ano. El tubo digestivo, excepto el intestino
medio esta tapizado por una delicada capa de quitina que en la boca y en el
ano, se continúan con la cutícula externa, en cada muda se cambia todo este
revestimiento.
2.3.6 Aparato reproductivo.
Aparato reproductor masculino. El aparato reproductor del macho, está
formado por tres pequeños testículos esferoides, los que miden
aproximadamente 180 micras de diámetro, situados debajo del corazón y atrás
del hepatopáncreas, en los tres lóbulos que forman los anteriores son cortos y
laterales en tanto que el posterior es largo y situado en la línea media. De uno

12
a otro lado del punto de unión de estos tres lóbulos parten los conductos
deferentes muy largos y estrechos en los existen numerosas vueltas antes de
llegar a los orificios que están situados en las coxas del último par de
periópodos; estos tres testículos están formados por un gran número de
pequeñas vesículas esferoidales originadas por dilataciones de los tubos
seminíferos, las células de los bordes de estas dilataciones de las células que
forman la pared de cada vesícula, al llegar la época de reproducción, se
multiplican y experimentan los cambios relativos a la espermatogénesis; de los
testículos laterales sale un conducto ancho de 2 mm de largo, que se une al
conducto del testículo anterior, al unirse estos dos sale un conducto largo y
ancho de donde salen dos conductos deferentes de aproximadamente 12 mm de
largo y de color lechoso que cuando tiene el contenido seminal en su interior
éste se escapa por ruptura. Los espermatozoides pasan después a los
conductos deferentes donde forma un material viscoso que se expulsa al
exterior en forma de espermatóforo filamentoso (Rosas, 1976).
Aparato reproductor femenino. El aparato reproductor de la hembra está
constituido por el ovario, órgano trilobulado situado inmediatamente debajo y
delante del corazón, entre el seno pericárdico y el tubo digestivo (volumen que
ocupa en el cefalotórax) de los tres lóbulos que lo forman, dos son anteriores,
laterales y uno medio impar; del tercio anterior de los lóbulos laterales y
ventralmente salen dos conductos cortos y gruesos: los oviductos, los cuales se
abren mediante orificios separados en la base del segundo par de patas
ambulacrales, por debajo del opérculo genital.
Las paredes del ovario están formadas por un epitelio formado por capas de
células cúbicas, de núcleo pequeño y cromatina compacta; entre ellas se
destacan las oogonias por su forma redondeada, su citoplasma claro y su núcleo

13
grande con la cromatina dispuesta en forma de red laxa. Estos elementos
sexuales al crecer, arrastran consigo a las células epiteliales de las partes del
ovario que acaban por formar en torno a cada oogonia una capa de células
aplanadas que se ponen en contacto directo con la membrana vitelina del ovulo,
dicha capa epitelial es semejante a la que presentan los folículos de primer
orden en los ovarios de los vertebrados.
El ovario tiene de 90 a 120 óvulos en promedio, cuando ya están maduros y
listos para la fecundación tienen un tamaño de 500 a 700 micras y son de color
verde olivo, la forma de los óvulos es casi esférica por la presión que ejercen
unos sobre otros. De la cara externa de los lóbulos salen los oviductos con un
diámetro de 600 micras y longitud de 12 mm, estos oviductos que atraviesan la
capa muscular son transparentes y elásticos y desembocan en las caras
internas de las coxas.
Cuando los óvulos se encuentran en el interior del ovario tienen diferente
tamaño y coloración que cuando ya se ha realizado la fertilización y han bajado
por los oviductos, pegándose en los pleópodos abdominales de la hembra, la
abertura de salida de los oviductos tienen forma de ojiva y se encuentra
escondida en las coxas (Rosas, 1976).
Por la cantidad y distribución de las reservas nutritivas, el huevo de los
acociles corresponde al tipo denominado centrolécito, en éste el citoplasma
formativo aparece como una capa muy fina por debajo de la membrana vitelina
pero la mayor parte se acumula en la porción central del vitelo y del citoplasma
formativo, lo que va a determinar el mecanismo de segmentación (Cantú,
1959), (Lám. 3).

14
Lámina 3. Morfología interna del acocil Cambarellus (C.) montezumae.
2.4 Ecdisis o proceso de muda.
El acocil debe pasar por la ecdisis o muda (que consiste en renovar su
caparazón o exoesqueleto, el cual está compuesto de quitina impregnada de
calcio) para crecer (crecimiento en escala, es decir que el acocil incrementa su
tamaño al mudar y lo detiene mientras que su exoesqueleto se endurece) o
regenerar alguna parte de su cuerpo. El acocil se desprende de su exoesqueleto
el cual sale con las antenas, ojos, mandíbulas, branquias y patas,
posteriormente se cubre de un nuevo caparazón (Santiago, 2001).
Durante la etapa de muda el acocil no se alimenta ya que en ese momento
también está regenerando sus órganos internos, especialmente el estómago
triturador. El endurecimiento del caparazón se realiza por reservas de calcio
en órganos especializados llamados gastrolitos (Santiago, 2001).
Antes de la muda se forman dos gastrolitos, que son piedras de carbonato
cálcico que se localizan a los lados del estómago. Cuando se aproxima la
muda, parte del calcio extraído del viejo exoesqueleto se almacena en los

15
gastrolitos durante la muda, el forro articulado del esófago y el estómago,
incluida la pared gástrica, se desplazan hacia adelante a través de la pared de
la boca. Los gastrolitos se depositan en la cavidad del estómago en donde se
disuelven; y el cuerpo absorbe ese carbonato de calcio para el endurecimiento
inicial del exoesqueleto y la boca (Coll, 1987).
2.5 Reproducción.
Los acociles se ubican entre los crustáceos decápodos de agua dulce que
presentan un marcado dimorfismo sexual y un ciclo de vida directo sin fases
larvales. La reproducción se lleva a cabo durante todo el año y depende de las
condiciones ambientales. En los acociles el acoplamiento es lento y difícil; el
macho busca a la hembra e intenta voltearla con la ayuda de sus quelas.
Después de varios intentos, cuando el macho domina a la hembra deposita el
líquido espermático, al principio sobre las placas externas de la aleta caudal y
después sobre el tórax y alrededor de los orificios externos de los oviductos en
donde la materia seminal se gelifica y los filamentos resultantes son como
espermatóforos tubulares resistentes que contienen el líquido seminal
(Espinoza, 1992).
La fertilización es externa y no es siempre inmediata, ya que la hembra puede
almacenar el saco espermático por más de seis meses, hasta que las condiciones
ambientales sean favorables. La hembra después de la fecundación se aísla en
una guarida donde proseguira la gestación.
El tiempo que transcurre desde la cópula hasta que los huevos son depositados
en el abdomen puede ser incluso de 3 ó 4 días (Morones, 1991). Cuando los
huevecillos descienden al abdomen son adheridos a los pleópodos en forma de

16
racimo por una sustancia mucilaginosa de color blanco que produce la hembra
días antes del desove. Cada huevo queda unido a los pleópodos por una
delgada membrana (Civera y Cruz, 1982). El número de huevos varía con la
calidad del medio, disponibilidad del alimento y el tamaño del abdomen de la
hembra (Santos, 1998).
La hembra provoca una corriente de agua que irriga los huevos fecundados
mediante contracciones repetidas de su cola. Los huevos no fecundados se
descomponen o son retirados por los apéndices abdominales los cuales limpian,
peinan y raspan constantemente al racimo. La duración de la incubación
depende de los parámetros del agua, llegando a ser de 35 a 40 días (Rosas,
1976).
La liberación no es sincrónica, sino se da de forma gradual en un lapso de 2-5
días. Los acociles tiene un desarrollo directo (no tienen estadios larvales) y
eclosionan como un pequeño acocil, el cual es muy parecido al adulto. La cría
tiene el caparazón blando y maleable, se alimenta de las reservas contenidas en
su cefalotórax, permanecen adheridas a la madre y regresan a refugiarse bajo
el abdomen durante aproximadamen diez días, hasta que concluyen dos mudas
(Espinoza, 1992; Avault & Huner, 1985).
2.6 Hábitat y distribución natural.
En México, Villalobos (1955) menciona la ubicación de esta especie en cuencas
cerradas del Eje Neovolcánico, que abarca desde Puebla hasta Jalisco y
Nayarit, siendo probablemente Cambarellus (C.) montezumae la especie de
distribución más occidental entre los Cambáridos mexicanos. Es evidente que
su área de distribución ha experimentado una merma considerable en los

17
últimos cuarenta años, debido principalmente a la desecación de los lagos que
ocuparon los Valles de México y Toluca y secundariamente a la contaminación
del Alto Lerma.
2.7 Alimentación.
Descripciones generales para Cambáridos indican que estos organismos en el
medio natural son omnívoros y depredadores ocasionales, donde el canibalismo
es común. Como en muchos decápodos, estos organismos se alimentan
principalmente de detritus enriquecido con microorganismos degradadores,
donde los fragmentos de hojas son predominantes así como de raíces y
microalgas, por lo que sus hábitos alimenticios se definen como politróficos
(Pennak, 1978; Huner, 1981; McHarney, 1984).
Los acociles juveniles son primordialmente cazadores y preferentemente
consumen proteína animal. En la edad adulta no depredan, pero consumen
pequeñas cantidades de animales aunque gran parte de su alimento consiste de
detritus vegetal (Huner, 1981; Rodríguez-Serna, 1991, 1999; Rodríguez-Serna,
et al., 1998).
De acuerdo a observaciones de Jones (1990) y Gherardi (2002), se sabe que
existen dos picos de actividad durante el día, los cuales están condicionados por
la intensidad luminosa; el más intenso se encuentra entre las 18 horas y la
media noche, y el segundo es justo antes de amanecer.

18
2.8 Requerimientos ambientales.
Los acociles requieren agua con un pH comprendido entre 7 y 9; en estas aguas
es imprescindible la presencia de calcio en altas concentraciones, necesario
para una adecuada calcificación en el proceso de muda, y un nivel mínimo de 4
ppm de oxígeno disuelto. El intervalo de temperatura es importante para el
desarrollo de esta especie y oscila entre los 12 y 26 °C (Moctezuma, 1996).
2.9 Interacciones ecológicas.
Cambarellus (C.) montezumae es presa de anfibios como ajolotes (Ambystoma
sp.); de ranas del género Rana; de peces como la carpa Cyprinus carpio, especie
introducida en México (Hinojosa & Zambrano, 2004), de los charales
Chirostoma jordani y de reptiles como la culebra de agua Tamnophis sp. Los
acociles juveniles son filtradores de materia orgánica finamente fragmentada y
consumidores de algas. Cuando crecen se adaptan para procesar el detritus,
fraccionan las raíces de la vegetación riparia y las hojas que caen de los árboles
favoreciendo su degradación por bacterias (Holdich, 2002). Cambarellus (C.)
montezumae es elemento fundamental como consumidor de especies macrófitas
como Elodea sp., planta acuática que se encuentra disponible en su hábitat
(Rodríguez-Serna y Carmona, 2002).
En su medio natural Cambarellus (C.) montezumae se alimenta de larvas de
invertebrados como los insectos de las clases Hemiptera (chinche y escorpión
acuático), Coleoptera (escarabajo acuático), Odonata (libélula) y Phasmida
(insecto palo); en el caso de los crustáceos, su alimentación incluye larvas de
anostracos, cladóceros (pulga de agua) y anfípodos (Hyalella sp.), e incluso

19
ingiere rotíferos (Moctezuma, 1996). Por lo tanto, se puede afirmar que
Cambarellus (C.) montezumae tiene una influencia significativa en varios
niveles tróficos, tanto como depredador y como presa (Rangel, 2009).
2.10 Valor nutrimental de la especie.
Fuente: Laboratorio de Bromatología del Centro de investigaciones en
Ciencias Biológicas, Universidad Autónoma de Tlaxcala, 1991.
Cuadro 1. Valor nutrimental del acocil Cambarellus (C.) montezumae de la
presa de Atlangatepec, Tlaxcala.
Composición nutrimental %
Materia humeda 73
Materia seca 27
Cenizas 16.8
Proteína 10.7
Lípidos 15.2
Fibra Cruda 26.6
Materia libre de N. 30.7

20
3. ANTECEDENTES.
El acocil Cambarellus (C.) montezumae es una especie endémica de nuestro
país, es un componente crítico en el ecosistema lacustre con un potencial
comercial importante, pese a esto, se conoce muy poco acerca de aspectos como:
su crecimiento, requerimientos nutricionales, procesos fisiológicos,
sobrevivencia, reproducción, producción y aspectos ecológicos tanto en
condiciones naturales como de laboratorio. Hasta ahora son escasos los
estudios que se han realizado; sin embargo, algunas investigaciones han
aportado aspectos significativos sobre la biología de este organismo; a
continuación se describen algunas de las investigaciones más importantes
sobre esta especie.
Uno de los primeros estudios sobre la especie fue realizado por Villalobos en
1955, en donde describe aspectos taxonómicos, morfológicos y de distribución de
los Cambarinos de la Fauna Mexicana (Crustacea: Decapoda); en este estudio
se menciona que dicha especie se distribuye en cuencas cerradas del Eje
Neovolcánico, que abarca desde Puebla hasta Jalisco, siendo probablemente
Cambarellus (C.) montezumae la especie de distribución más occidental entre
los Cambáridos mexicanos.
En relación a su biología, Cantú (1959) llevó a cabo un estudio sobre la
embriología de esta especie y realizó la colecta de los organismos en el Lago de
Chapultepec, de donde obtuvo hembras ovígeras de las cuales extrajo los
huevos para realizar cortes histológicos y observar los cambios superficiales del
blastodermo y su desarrollo. Cantú reporta que los huevos de Cambarellus
(C.) montezumae son de tipo centrolecito, con tres membranas (la membrana de

21
adhesión a las patas, el corion y la membrana vitelina). Su tipo de
segmentación es superficial y la gastrulación se produce por movimientos de
involución y convergencia.
En 1982, Civera y Cruz realizaron una investigación acerca de la posible
incubación artificial del acocil Cambarellus (C.) montezumae, en esta
investigación consideraron aspectos sobre el hábitat de este organismo, su
reproducción, desarrollo embrionario y fecundidad. Los autores concluyeron
que existe un modelo logarítmico de predicción de la fecundidad, basándose en
la relación entre la talla y/o el peso de la hembra progenitora y la fecundidad.
Asimismo, describen un mecanismo experimental de incubación, que
demuestra que es posible el desarrollo de los hueveos en un sistema de
incubación artificial.
Maldonado (1990) estudió la respuesta al estrés térmico del acocil Cambarellus
(C.) montezumae, comparando los métodos de temperatura crítica máxima
(TCM) y temperatura letal 50 (TL50). Encontró que la TCM para verano es de
35.1 °C y para invierno de 30.7 °C, la TL50 para organismos de verano fue de
41.8 °C y en invierno es de 41.5 °C, concluyendo que las altas temperaturas
modifican el medio interno de los organismos como consecuencia de la
búsqueda del equilibrio de temperaturas con el medio externo, afectando la
presión osmótica total.
Cornejo, en 1991, realizó estudios sobre la selección térmica de Cambarellus
(C.) montezumae y su correlación con algunos índices fisiológicos; encontrando
que existen diferencias en la selección térmica influenciadas por el sexo, talla y
época del año. Además encontró diferencias significativas en el desempeño de

22
los organismos debido al efecto que ejerce la temperatura como factor maestro
en el crecimiento, sobrevivencia y metabolismo.
En 1991 Rodríguez-Serna, determinó la influencia que tiene la dieta en la
eficiencia de asimilación, perdidas de energía por respiración y productos
nitrogenados, en el balance energético del acocil Cambarellus (C.) montezumae,
contrastando las temperaturas de 17 y 23 °C. En ese estudio se encontró que
los organismos mantenidos a 23 °C presentaron una mayor tasa metabólica, así
con un mayor consumo de alimento y una mayor excreción nitrogenada; a
diferencia de los organismos mantenidos a 17 °C, los cuales presentaron la
mayor eficiencia de consumo y crecimiento.
García en 1991, llevó a cabo un estudio sobre el efecto de diversas dietas sobre
la sobrevivencia y crecimiento de crías del acocil Cambarellus (C.) montezumae,
estas dietas consistieron en alimento balanceado, spirulina, alimento
balanceado-spirulina, Elodea densa, E. densa fresca y alimento balanceado-E.
densa fresca, el periodo de alimentación fue de 40 días. García concluye que el
mayor crecimiento se dio en las crías alimentadas con alimento balanceado - E.
densa fresca, seguido de las crías alimentadas con alimento balanceado –
spirulina, y finalmente de las crías alimentadas con alimento balanceado.
Por otro lado, Aguilar (1991) en su investigación, evaluó el crecimiento y
producción de juveniles de acocil Cambarellus (C.) montezumae,
alimentándolos con la macrófita Egeria densa usando cinco niveles de ración y
dos temperaturas 17 y 23 °C, con el fin de determinar el efecto de estas dos
variables sobre el crecimiento. Concluyendo que el crecimiento tiene una
relación significativa con respecto a la temperatura y los niveles de ración.

23
Morones, realizó un estudio en 1991 sobre aspectos reproductivos bajo
condiciones de laboratorio de Cambarellus (C.) montezumae, en el cual delimitó
las tallas a las cuales este organismo es capaz de reproducirse en condiciones
de cautiverio, demás efectuó observaciones sobre el comportamiento de las
hembras ovígeras. Los organismos utilizados en este experimento fueron
alimentados con un 4% de su peso promedio corporal de un macerado de
Poecillia reticulata y zanahoria cocida. La longitud total registrada para
hembras reproductoras fue de 3.0 cm en promedio en comparación con los
machos reproductores de una longitud total de 2.8 cm. La actividad de las
hembras ovígeras se vio reducida a permanecer ocultas dentro de sus refugios
hasta el periodo de eclosión de las crías.
En 1995 se llevó a cabo un proyecto llamado “Plan Lago de Xochimilco”, en el
cual Aragones realizó un estudio para conocer y aplicar biotecnología para
producir en cautiverio especies nativas del Lago de Xochimilco, entre estas
especie se consideró al acocil Cambarellus (C.) montezumae¸ ya que resultaba
relevante su fomento y rescate para las comunidades ribereñas debido a su
importancia económica y alimentaria. En este estudio se concluye que esta
especie es de fácil reproducción pero que hace falta mayor investigación en
torno a las condiciones físico-químicas del sedimento y la calidad del agua en el
Lago de Xochimilco, ya que este organismo en su hábitat natural presenta
elevadas tasas de mortalidad.
Moctezuma en 1996 estudió las bases biológicas y técnicas del cultivo de
Cambarellus (C.) montezumae bajo condiciones de laboratorio y en pequeños
estanques, realizó observaciones sobre su distribución natural, sus necesidades
de substrato y alimento, profundidad del agua, cobertura vegetal, oxigenación y
densidad de cultivo que promovieran su óptimo crecimiento.

24
En 1997, González comparó las concentraciones de los metales: Hierro (Fe),
Cobre (Cu), Manganeso (Mn), Zinc (Zn), Plomo (Pb), Cromo (Cr), Cobalto (Co),
Níquel (Ni) y Cadmio (Cd) en el acocil (Cambarellus (C.) montezumae), en el
ajolote (Ambystoma mexicanum) y en el charal (Chirostoma jordani), estas
determinaciones las realizó en diferentes sitios de la zona lacustre de
Xochimilco, encontrando que los organismos no son aptos para consumo
humano según la Organización Mundial de la Salud.
Arana y colaboradores en 1998, efectuaron una investigación sobre
Cambarellus (C.) montezumae en tres embalses del estado de Tlaxcala, con el
objetivo de delimitar la importancia que tienen estos organismos en la cadena
trófica, puesto que son depredadores activos que transforman la materia
orgánica en los cuerpos de agua dulce, además de ser aprovechado como
alimento autóctono por habitantes de las zonas rurales.
Barbosa (1998) realizó una revisión de estudios previos empleando varios
modelos estadísticos con la finalidad de observar el efecto que tienen la
temperatura y los distintos niveles de nutrientes (proteínas, lípidos y
carbohidratos), sobre el crecimiento de Cambarellus (C.) montezumae.
Concluyó que no hubo una relación significativa entre la preferencia de
temperatura y la talla de este organismo; asimismo, indica que los individuos
de menor talla exhiben una tasa de crecimiento superior que los de talla mayor;
y que el porcentaje óptimo de nutrientes es el siguiente: proteínas de 30% a
40%, lípidos no deben ser mayor al 10% y los carbohidratos entre 5 y 20%, con
base al peso corporal de un organismo.
En 1999 Salazar efectuó un experimento en el que uso organismos de la especie
Cambarellus (C.) montezumae para observar el daño tóxico y genotóxico del

25
pesticida dieldrín (hidrocarburo clorado) utilizando una técnica de medición
para los niveles de la lipoperoxidación, así como un bioensayo de electroforesis
unicelular en gel. En el cual se determinó que el pesticida dieldrín es
genotóxico y probablemente tóxico para este organismo, y se deduce que puede
tener un efecto potencial para sus depredadores.
Bárcena en el 2000, realizó un estudio sobre crecimiento del acocil Cambarellus
(C.) montezumae¸ utilizando dos dietas: la dieta A conformada por 40% de
semilla de amaranto, 30% de levadura de cerveza y 30% de pluma de pollo; y la
dieta B, constituida por 50% de alimento de engorda para conejo y 50% de
harina de carne. Los organismos se alimentaron al 4% de su peso húmedo
corporal promedio por día, se realizaron dos bloques: hembras y machos. Para
ambas dietas se observó un mayor crecimiento de las hembras en comparación
con los machos de su tratamiento.
En el año 2001 Santiago llevó a cabo una investigación sobre la tasa de
crecimiento de Cambarellus (C.) montezumae, usando diferentes temperaturas
en condiciones de laboratorio por un periodo de ochos meses. Las temperaturas
experimentales fueron 22 y 24 °C, no encontró diferencias significativas entre
éstas. Santiago reporta algunas observaciones sobre el ciclo de muda de estos
organismos, afirmando que durante los 2 primeros meses de vida estos
organismos tienen una mayor frecuencia de muda, observó que la madurez
sexual se presenta a una talla de 2.6 cm para hembras y de 2.06 cm para
machos, los apareamientos se llevaron a cabo durante todo el año y observó una
relación directa entre el peso de la hembra y el número de huevos que esta
porta.

26
Rodríguez-Serna & Carmona en el año 2002, estudiaron el balance energético
del acocil Cambarellus (C.) montezumae y las pérdidas de energía por
metabolismo. Se evaluó su efecto en la nutrición de los acociles, observando
que el metabolismo es modificado por la clase talla y el nivel de ración del
alimento, el factor limitante para el crecimiento de la especie es la
temperatura, encontrando los mejores resultados a una temperatura de 17 °C
en donde se presentó el mayor consumo de alimento, así como la mejor
eficiencia de asimilación de este.
En el 2003 Rojas realizó una revisión taxonómica de las especies de
Cambarellus (Cambarellus) (Cruatcea: Decapoda: Astacidae: Cambaridae), en
donde se encuentra ubicada taxonómicamente la especie de estudio. Esta
especie es aparentemente la especie con mayor distribución de las especies
pertenecientes al género Cambarellus. Muestra un alto grado de variación
morfológica, la cual puede atribuirse a cambios en las condiciones del agua, la
contaminación, el aislamiento en ciertas áreas o debido a la manipulación del
hombre en las áreas donde habita.
En el año 2003 Palacios realizó una investigación cuyo objetivo era optimizar la
tasa de reproducción en cautiverio del acocil Cambarellus (C.) montezumae, con
el fin de obtener un mayor número de hembras ovígeras por unidad de espacio
y tiempo. Realizó dos experimentos, uno en invierno (Noviembre a Febrero;
18.5 °C) y otro en primavera (Marzo a Mayo; 22 °C), en ambos experimentos se
utilizó para la alimentación de los organismos alimento balanceado con la
siguiente formulación: proteína 35%, grasa 5%, fibra cruda 25% y humedad
10%, además de alimento vivo como Tubifex, Artemia y Daphnia. El mayor
porcentaje de hembras ovígeras se obtuvo en el experimento de primavera.

27
Fragoso en el 2003, llevo a cabo un estudio sobre el aprovechamiento como
alimento de desechos orgánicos procedentes de las explotaciones pecuarias.
Durante este estudio se alimentó a acociles de la especie Cambarellus (C.)
montezumae con sólidos ensilados de cerdaza, por un periodo de cuatro meses.
Evaluando su crecimiento (ganancia de peso), mortalidad y número de mudas,
así como su reproducción. Fragoso concluye que el ensilado de cerdaza no
cubre las necesidades nutricionales de los acociles en cultivo.
Hinojosa & Zambrano (2004) realizaron una investigación sobre los efectos que
tiene la carpa Cyprinus carpio (especie exótica) sobre Cambarellus (C.)
montezumae lermensis (especie endémica) en embalses de bordos del municipio
de Acambay, Estado de México. Este estudio es trascendental para modificar
la percepción que se tiene sobre la carpa como depredador esencial del acocil,
ya que la conclusión que obtienen los investigadores, indica que el acocil no es
un alimento significativo para este animal. Sin embargo, se mantiene un
efecto negativo indirecto de la carpa hacia el acocil, porque ésta modifica el
entorno a través del incremento de nutrientes sobre el medio, generando
turbidez en el agua, destruyendo las plantas enraizadas que son el hábitat
apropiado de este organismo, causando consecuencias sobre su distribución y
abundancia.
Nacif en el 2004, llevó a cabo un estudio sobre la descomposición aeróbica de la
macrófita Egeria densa presente en el hábitat natural de Cambarellus (C.)
montezumae y su uso potencial como alimento para esta especie. Obtuvo
sustratos alimenticios, bajo los cuales se evaluó la respuesta de crías de este
acocil bajo un enfoque bioenergético. Concluyendo que el ensilado vegetal
promovió el crecimiento de las crías alcanzándose en un lapso de cinco semanas
tallas cercanas a la de un estadio de sub-adulto.

28
Félix y Gutiérrez (2005) realizaron un estudio poblacional del acocil
Cambarellus (C.) montezumae en dos sitios (El Boyecito y Las Rosas) del río
San Juan, Querétaro; El Boyecito, lugar con alta calidad del agua y sedimentos
y Las Rosas sitio altamente degradado en calidad debido a la contaminación
química y orgánica. El momento de máxima reproducción se localizó en
noviembre, ya que fue cuando se encontraron el mayor número de hembras
cargadas en ambas poblaciones; y durante los meses de diciembre y enero se
localizaron las tallas más chicas. Sólo se encontraron diferencias significativas
en las tallas mínimas de reclutamiento y máximas de crecimiento entre ambas
poblaciones, la población de Las Rosas tuvo menor talla de reclutamiento (0.4
cm) y mayor talla máxima (4.0 cm) indicando que los individuos presentes en el
lugar contaminado crecen más, debido posiblemente, a la mayor cantidad de
materia orgánica en el sustrato.
En el año 2006 Colín realizó un estudio sobre el crecimiento y ciclo de vida del
acocil Cambarellus (C.) montezumae de los canales de Xochimilco, empleando
tres diferentes densidades de población: 50, 100 y 150 organismos por metro
cuadrado. Los resultados indicaron un crecimiento acelerado entre la cuarta y
la sexta quincena respecto al peso y un crecimiento gradual respecto a la talla.
Entre la quinta y sexta quincena los machos presentaron cambios de forma II o
de inmadurez a forma I o de madurez sexual, durante esta etapa se pudo
observar un mayor aumento de talla en las hembras con respecto los machos y
un engrosamiento del abdomen. Además, se observó en esta investigación que
bajo condiciones apropiadas estos organismos pueden tener un ciclo continuo de
reproducción.
Álvarez y Rangel en el 2007, realizaron un estudio poblacional del acocil
Cambarellus (C.) montezumae en Xochimilco, basado en el análisis de 12

29
muestreos, en los que se colectaron 668 organismos. El intervalo de tallas
registrado fue de 21 a 41.5 mm de longitud total. La presencia constante de
machos forma I y de hembras ovígeras, así como la presencia de organismos de
tallas pequeñas en la mayoría de los muestreos, sugieren que existe una
reproducción continua a lo largo del año. Las tallas de la primera
reproducción fueron 24 mm de longitud total para hembras y 21 mm para
machos. No se encontró ninguna relación entre la abundancia de Cambarellus
(C.) montezumae con respecto a la temperatura, la concentración de oxígeno
disuelto y el pH.
Sánchez (2007) llevó a cabo un estudio sobre el aprovechamiento de los
ambientes reducidos en los canales de Xochimilco para desarrollar el cultivo del
acocil Cambarellus (C.) montezumae, para consumo humano. En este estudio
se evaluaron los parámetros fisicoquímicos del agua y sedimentos de los
canales de Xochimilco, además se determinó la presencia de metales pesados
tanto en el sedimento como en los organismos. Concluyendo que los canales de
Xochimilco cumplen con las características de un ambiente reducido, se
demostró que los niveles de metales pasados y carga bacteriana registrados, no
superan los límites máximos permisibles para crustáceos comestibles.
En el 2009 Vásquez realizó un estudio sobre la reproducción y el crecimiento
del acocil Cambarellus (C.) montezumae en condiciones de laboratorio, planteó
tres diseños experimentales; el primero consistió en cuatro tanques exteriores
de 800 L con 219 reproductores por 335 días, el segundo en condiciones de
laboratorio manejó dos machos y una hembra con diez réplicas, y en el tercero
se manejaron tres densidades, 77, 154 y 231 org/m2 con dos réplicas. En cada
caso, se controló la alimentación y se realizó monitoreo de la calidad del agua.
Para el primer diseño se obtuvieron 6 174 huevos y 3 162 larvas, con una

30
mortalidad del 51.2%. En el segundo diseño se obtuvieron 14 huevos en
promedio por hembra y en el tercer diseño se encontró que la mejor densidad es
con 77 org/m2. De acuerdo con los resultados, el acocil Cambarellus (C.)
montezumae es una especie que se adapta perfectamente a las condiciones de
cultivo y es posible mantener una reproducción constante a lo largo del año y
generar una buena cantidad de juveniles que pueden ser utilizados para
programas de recuperación de la especie, repoblación o bien cultivos
comerciales.
Rangel en el 2009, llevó a cabo un estudio sobre la dinámica poblacional de
Cambarellus (C.) montezumae, en la pista de Remo y Canotaje “Virgilio Uribe”
ubicada en Cuemanco; Xochimilco. Efectuó 24 muestreos quincenales donde
capturó 1,476 organismos; de los cuales 717 fueron hembras, 705 machos y 54
individuos indeterminados. Las principales características poblacionales las
determinó a través de los siguientes parámetros: abundancia, composición de
tallas por sexo y a lo largo del muestreo, tasa de crecimiento individual directa
y estimada, tallas de reproducción, fecundidad e intensidad de reclutamiento.
Calculó la talla máxima a la que puede llegar Cambarellus (C.) montezumae,
dentro del sitio de estudio, mediante el modelo de crecimiento de von
Bertalanffy teniendo como resultado 48.7 mm LT. Midió los principales
parámetros fisicoquímicos (temperatura, pH, concentración de oxígeno disuelto
y porcentaje de saturación de oxígeno disuelto) a fin de conocer si tienen una
relación con las principales variables demográficas. Los resultados indican que
la temperatura tiene un efecto significativo sobre la abundancia de
Cambarellus (C.) montezumae, por lo que el crecimiento poblacional aumenta
en los meses cálidos. Sin embargo, la reproducción y el reclutamiento se
presentaron en todo el año de colecta. La mortalidad es alta en individuos
jóvenes por lo que muy pocos llegan a edad reproductiva. Por lo tanto,

31
Cambarellus (C.) montezumae es una especie que presenta, principalmente,
características de la selección “r” para tener éxito en su hábitat.
Gayosso en 2009, determinó la cantidad de metales pesados (Cd, Cr, Fe, Ni y
Pb) en carpas (Cyprinus carpio) y acociles (Cambarellus (C.) montezumae) del
Lago de Xochimilco, encontrado que los acociles presentan una acumulación de
Cr, Cd y Pb en la talla grande, Fe en la talla mediana y Ni en la talla chica. Se
rebasó la concentración permisible para cada uno de los metales pesados
determinados, por lo que no se recomienda para consumo humano.
En el 2010, García-Padilla llevo a cabo la estimación del balance energético del
acocil Cambarellus (C.) montezumae en el lago de Xochimilco, realizó muestreos
en época cálida y fría del año, con el fin de determinar la influencia de la
estacionalidad sobre los elementos de la ecuación general del balance de
energía (C = P + R + F + U). El análisis de abundancia de los organismos,
mostró que en época cálida existe un mayor número de individuos con respecto
a los meses que son considerados de época fría, así como un mayor número de
individuos de tallas pequeñas, lo que nos indica que la reproducción se acentúa
en los meses cálidos del año. La estacionalidad no tuvo influencia en la
composición del tejido de los organismos. En cuanto a los parámetros de la
ecuación general del balance de energía, se obtuvo que Cambarellus (C.)
montezumae consume alrededor de 1,440 Calorías/ejemplar*día de alimento en
época cálida y 876 Calorías/ejemplar*día en época fría y pierde por evacuación
de heces 2 % de la energía total extraída del alimento, por respiración entre el
46 y 51 % y por excreción nitrogenada el 7%, por lo que el campo de crecimiento
o producción para la época cálida es del 45% de la energía obtenida del
alimento consumido y en época fría es del 40%, la importancia de conocer los

32
valores de la ecuación general del balance de energía, radica en la aplicación de
biotécnicas para acelerar la tasa de crecimiento y reproducción de esta especie.
En 2010 Cerón-Ortiz, et al., realizaron un estudio sobre la influencia del
alimento y la calidad del agua en el crecimiento de post-larvas del acocil
Cambarellus (C.) montezumae, diseñaron cuatro tratamientos: T1 (calidad del
agua alta y alimento inerte [Camaronina 35]), T2(calidad del agua baja y
alimento inerte[Camaronina 35]), T3 (calidad del agua alta y alimento vivo
[Daphnia pulex]) y T4 (calidad del agua baja y alimento vivo [Daphnia pulex]),
los organismos se alimentaron al 15% de su peso húmedo promedio. Los
resultaron mostraron los mejores crecimientos en largo y peso para
Camaronina 35 independientemente a la calidad del agua. Con relación al
ancho, se encuentran diferencias significativas tanto por la calidad del agua
como por el tipo de alimento. Los resultados expresaron la posibilidad que en
relación al crecimiento (peso y longitud) el efecto de la calidad del agua no
afecta directamente el desarrollo de este organismo, pero es necesario conocer
los impactos antropogénicos sobre aspectos básicos del hábitat como el alimento
y el refugio para hacer más efectiva la elaboración de una técnica de cultivo.
Ángeles-Monrroy, et al., en 2010, llevó a cabo un estudio sobre los parámetros
físico-químicos que afectan la población en medios naturales del acocil
Cambarellus (C.) montezumae en Tezontepec de Aldama, Hidalgo; para lo cual
se establecieron zonas de muestreo con el fin de obtener la talla (longitud y
ancho), peso y densidad de acociles. Se identificaron las posibles fuentes de
contaminación, los factores físico-químicos y se determinó el contenido y tipo de
microorganismos presentes en el agua. Los pesos y las tallas de los
organismos oscilaron en tres rangos: pesos menores a 0.09 gr, de 0.01 a 1.5 gr y
mayores a 1.6 gr. En talla los valores fueron menores a 9 mm, de 10 a 25 mm

33
y el tercero mayores a 26 mm. Los resultados descritos sugieren que el efecto
de la actividad antropogénica no es la afección directa al crecimiento del acocil,
sino más bien el efecto que tiene la interacción de los factores bióticos y
abióticos del hábitat, como el tipo de vegetación, el cual sirve de refugio y
alimento para este organismo.
En el 2010, Cortes realizó una investigación acerca de la relación entre los
organismos más representativitos del sistema lacustre de Xochimilco, el ajolote
(Ambystoma mexicanum) y el acocil (Cambarellus (C.) montezumae). Estos
organismos comparten el ambiente de forma muy estrecha, así como sus
respectivos papeles en la red de alimentos del ecosistema. El objetivo de este
trabajo fue determinar las posibles interacciones depredador-presa entre el
ajolote y el acocil en cuanto a la dirección de la interacción y a los tamaños
involucrados en ella. Encontrando que los ajolotes adultos depredan a los
acociles de diferentes tamaños, desde que éstos se separan del abdomen de la
hembra hasta que alcanzan la madurez sexual; por otro lado, los acociles
adultos son capaces de alimentarse de huevos y crías de ajolote.
Villa en 2010, lleva a cabo un estudio preliminar acerca de aspectos de la
dinámica poblacional del acocil Cambarellus (C.) montezumae en el Lago de
Xochimilco, específicamente en el canal del Bordo, los aspectos evaluados
fueron la abundancia de organismos, época reproductiva, picos de
reclutamiento y variaciones de talla dentro de la población. Los resultados
indican que la temperatura tiene un efecto significativo sobre la abundancia de
Cambarellus (C.) montezumae, los picos de reclutamiento de la población se
dieron en los meses de abril y junio, la época reproductiva comprendió los
meses de marzo y mayo.

34
Arredondo-Figueroa, et al., (2011), llevan a cabo un estudio aspectos
reproductivos del acocil Cambarellus (C.) montezumae en condiciones
controladas, con el objetivo de determinar la factibilidad de la producción de
huevos y juveniles y su potencial de aplicación a programas de rescate,
repoblación y cultivo. Se realizaron 2 ensayos experimentales, el primero en 4
estanques exteriores de 800 litros conectados a sistemas cerrados de
recirculación y el segundo en recipientes de plástico en laboratorio de 2 litros.
Para el primer ensayo se manejaron 219 reproductores durante 335 días
obteniendo 136 hembras ovígeras. Cada hembra produjo en promedio 45 ± 23
huevos por puesta. El 83.7% de los huevos eclosionaron obteniéndose un total
de 3 162 juveniles. Los resultados indican la factibilidad para su cultivo y
producción de huevos y juveniles.
En el 2011, Ávila determinó la calidad microbiológica del acocil Cambarellus
(C.) montezumae en las localidades: Presa Atlangatepec, Lagunas de Zempoala
y los acociles vendidos en el mercado de San Juan, México D.F., en los cuales
se determinaron coliformes totales, mesófitas aéreas y Salmonella sp., así como
bacterias asociadas. Encontrando que dichas muestras no cumplen con la
norma sanitaria de la NOM-242-SSA1-2009 para coliformes. El total de las
muestras cumplen con el límite que establece la NOM-242-SSA1-2009 para
bacterias mesófilas aerobias. Estos organismos no presentan un riesgo para el
consumidor debido a que los acociles se consumen cocidos; sin embargo, la
presencia de estas bacterias en el agua representa un indicador de la calidad
del agua de su hábitat y un riesgo para el pescador o comercializador de estos
organismos.
Cerón, et al., en 2011 obtuvieron resultados preliminares acerca de la
reproducción como un efecto secundario en cultivos controlados del acocil de rio

35
Cambarellus (C.) montezumae. En los bioensayos utilizados no se consideró la
separación se sexos, ya que no existen estudios que establezcan el efecto de esta
variable sobre los cultivos. Se utilizaron dos bloques, el primer bloque se
alimentó con Camaronina 35 y el segundo con Daphia sp. Se identificó a la
primera hembra ovígera a la semana 7 del cultivo (con 2.7 cm de longitud y un
peso de 1.2 g), dentro del bloque alimentado con Camaronina 35. Se
encontraron diferencias significativas en el número de hembras ovígeras en
ambos bloques, obteniendo el mayor número de hembras en el bloque
alimentado con Camaronina 35.
Santos, et al., (2011) evaluaron el aporte nutricional del acocil Cambarellus (C.)
montezumae que se encuentra en la presa Madin del Municipio de Atizapan de
Zaragoza encontrando valores del 77.57 % de humedad, proteína del 44.04 %,
20.17 % de carbohidratos, 3.56 % de lípidos, 8.93 % de fibra y 23.30 % de
minerales de los cuales solamente se cuantificaron: Na 0.608 g/100 g; K
0.608%; Fe 0.023%; Ca 11.85 g/100 g.
En el 2011 Aguilar-Román evaluó la influencia de la temperatura (20, 25 y 30
°C) en diferentes índices fisiológicos y crecimiento de crías de acocil
Cambarellus (C.) montezumae. Consideró los indicadores de supervivencia,
crecimiento total, tasa de crecimiento intrínseco (TIC) y producción (P). Los
organismos se alimentaron con una dieta comercial 40% de proteína, durante
un periodo de 60 días. Se obtuvo como resultado que el efecto de la
temperatura se refleja en la supervivencia, en la TIC y en la producción,
obteniendo los mejores resultados para los organismos mantenidos a 25 °C.
Considerando los resultados obtenidos en esta investigación se considera que el
rango de temperatura recomendado para obtener un rápido crecimiento y una
mayor producción de este organismo es de 20 a 25 °C.

36
Pacheco, et al., 2011 realizaron un experimento con acociles (Cambarellus (C.)
montezumae) obtenidos en los canales de Xochimilco (CIBAC), utilizando una
dieta artificial enriquecida con un probiótico (lactobacilo) y como control la
misma dieta pero sin probiótico. El experimento se realizó durante 24
semanas, la temperatura se mantuvo en 25±2ºC. Cada semana los organismos
fueron pesados y medidos para que al final del experimento se determinara la
Tasa de crecimiento relativo (TCR) y el Factor de condición (KM). Los valores
de TCR con la dieta con el probiótico indican que el mayor crecimiento se da en
las primeras 8 semanas, mientras que para la dieta control, se observa al final
del experimento. La dieta enriquecida permitió a los organismos tener un
grado de bienestar mejor a lo largo de todo el experimento, en cambio con la
dieta control, el beneficio apenas se observa en la semana 24.
En 2011 Gallardo llevó a cabo un estudio sobre el efecto de la temperatura en el
crecimiento, supervivencia, desove y viabilidad del acocil Cambarellus (C.)
montezumae, bajo condiciones de laboratorio. Se probaron tres temperaturas
18 ± 1 °C, 22 ± 1 °C y 26 ± 1 °C. Los resultados no mostraron diferencias
significativas entre tratamientos con respecto al crecimiento y supervivencia,
se encontraron diferencias significativas entre tratamientos respecto a la
supervivencia y no así en el porcentaje de desoves, a 26 °C se reportó la mayor
viabilidad, lo que demuestra que la temperatura tiene una marcada influencia
sobre los parámetros evaluados.
En el 2013, Escalante llevó a cabo una evaluación sobre el crecimiento de crías
de acocil Cambarellus (C.) montezumae alimentadas con detritus de las
macrófitas Eichhornia crassipes y Typha latifolia sometidas a un proceso de
descomposición aeróbico, dichas macrófitas se encuentran presentes en el

37
hábitat natural de esta especie. Para la evaluación del detritus de las
macrófitas como alimento en crías de acocil, se alimentaron a saciedad con el
detritus proveniente de tres tiempos de degradación de las macrófitas: T2, T3 y
T4, correspondientes a15, 21, 30 días respectivamente, y como alimento Control
se empleó Camaronina, la fase de crecimiento duró 90 días. Los resultados de
este trabajo muestran que entre tiempos de degradación para ambas
macrófitas no se encontraron diferencias significativas en el crecimiento y en el
análisis químico proximal de los acociles; pero se encontró un mejor desempeño
con el ensilado de E. crassipes tanto en el crecimiento como en la
supervivencia comparado con el de T. latifolia.

38
4. JUSTIFICACIÓN GENERAL.
El desarrollo de paquetes tecnológicos de especies endémicas como es el caso
del acocil Cambarellus (C.) montezumae, presenta un retraso en comparación
con los paquetes tecnológicos de especies comerciales como es el caso del
camarón, esto debido al poco o nulo desarrollo productivo a nivel piloto. Esto
establece la necesidad de que los estudios técnico-científicos que comprenden el
conocimiento de la biología (crecimiento, reproducción, nutrición y
alimentación) de esta especie, permitan optimizar y disminuir los costos
operativos de su cultivo, y de este modo generar un paquete tecnológico que
garantice un adecuado manejo del recurso y su aprovechamiento sostenible.
5. OBJETIVO GENERAL.
Contribuir al conocimiento sobre el crecimiento y la biología
reproductiva del acocil Cambarellus (Cambarellus) montezumae (de
Saussure, 1857), tanto en su hábitat natural como en condiciones
controladas.

CAPÍTULO 1.
ESTRATEGIA REPRODUCTIVA DEL ACOCIL Cambarellus
(Cambarellus) montezumae (SAUSSURE, 1857), EN EL LAGO
DE XOCHIMILCO. MÉXICO, DISTRITO FEDERAL.

39
6.1 INTRODUCCIÓN.
El valle de México se ubica en una gran cuenca cerrada (actualmente drenada
en forma artificial), que en la época prehispánica se acumulaba en el centro del
valle formando diversos lagos: al norte, Zumpango y Xaltocan; al centro, el gran
Lago de Texcoco; los tres contuvieron agua salobre. Al sur, el Lago de Chalco y
Xochimilco, de agua dulce y abastecidos por numerosos ríos y manantiales
(Ezcurra, 1990).
Una gran cantidad de productos tuvieron su origen en esta región
(principalmente en el Lago de Xochimilco), gracias a los beneficios de un medio
lacustre rico en cantidad y variedad de especies, a las técnicas de cultivo
intensivo por medio de chinampas, a la agricultura de temporal y a las piezas
de caza conseguidas en el pie de monte.
Aunque es difícil precisar la importancia absoluta que tuvo el aprovechamiento
de recursos alimenticios en cada etapa del desarrollo del México prehispánico,
es posible afirmar que en las más antiguas, la pesca, junto con la caza, la
recolección de animales y vegetales acuáticos, así como la caza de mamíferos y
recolección de plantas y frutos terrestres, fueron las actividades fundamentales
para garantizar la supervivencia de los asentamientos humanos localizados
alrededor del Lago de Xochimilco (Ezcurra, 1990).
Hasta la fecha, se han acumulado numerosos testimonios en códices y crónicas,
del consumo de una gran diversidad de productos. En el medio lacustre de
Xochimilco se consumían las tortugas de agua dulce, peces, las cuitlatl (ranas),
atepocatl (renacuajos), ahuauhtli (huevecillos del insecto acuático llamado

40
axayacatl), axolotl (ajolote) y acuitzil (acocil), entre otros productos (Ambrosio-
Arzate, et al., 2005).
Entre este grupo de productos el consumo de acocil (Cambarellus (C.)
montezumae) ha perdurado hasta nuestros días, aunque su consumo ya no es
tan frecuente, principalmente por su poca disponibilidad y su alto costo.
Su poca disponibilidad se debe a la merma que han sufrido sus poblaciones, ya
que se han visto disminuidas debido a la degradación, fragmentación y
contaminación de su hábitat, su sobreexplotación y la introducción de especies
exóticas. Su consumo actualmente es solo local en los estados del centro del
país como Tlaxcala, Hidalgo, Distrito Federal y Puebla (Rodríguez y Mendoza,
1999).
Este acocil representa un importante recurso biótico que posee un potencial
acuicultural importante en México, esto debido a que es un organismo tolerante
a los cambios de humedad y temperatura, así como a variaciones climáticas
extremas (Bückle, et al., 1994). Además, crece en un intervalo amplio de
temperatura, modula su tasa metabólica, ajusta su reproducción a las
condiciones imperantes en el ambiente y sus requerimientos nutricionales son
de bajo costo (Aguilar, 1991; Colín, 2006; Gutiérrez-Yurrita, 2008, Escalante,
2013). Sin embargo, existen escasos estudios destinados a comprender los
mecanismos de su biología, ecofisiología y cultivo.
En este sentido, es importante mencionar que los estudios encaminados al
conocimiento de su ciclo de vida y a las estrategias reproductivas de esta
especie en su hábitat natural, permiten señalar cómo este organismo ajusta sus
respuestas bioquímicas, fisiológicas y de comportamiento a las condiciones
imperantes en su hábitat, esta información nos puede ayudar a diseñar

41
programas de manejo con miras a su explotación sustentable y/o a la
conservación de la especie en su hábitat natural.
Una manera de estudiar estas respuestas es a través del cómo, cuándo y dónde
se reproducen estos organismos, con relación a los efectos de los factores
medioambientales, por lo que el objetivo básico de este capítulo será
comprender la ecología de la reproducción de esta especie en el Lago de
Xochimilco durante un ciclo anual.
Mientras los acociles son inmaduros toda la energía adquirida a través de la
ingesta de alimento se reparte entre el crecimiento y la sobrevivencia. Al
madurar gran parte de esta energía debe ser dedicada a los diversos procesos
reproductivos. El éxito reproductivo de la población dependerá enormemente
de dónde y cuándo se reproduzcan los acociles y de cómo los recursos
energéticos se administren para la reproducción (Saborido-Rey, 2005).
Es aquí donde surge una pregunta crucial: ¿Por qué es tan importante la
reproducción? La reproducción, es un proceso indispensable en la vida de los
organismos, ya que debido a ella perduran las especies a lo largo del tiempo,
por lo que todas las especies, incluido el acocil (Cambarellus (C.) montezumae),
buscaran maximizar su éxito reproductivo (Begon, et al., 2006).
Para maximizar el éxito reproductivo, un individuo debe repartir sus recursos
energéticos adecuadamente y además, procurar que su descendencia nazca en
las condiciones medioambientales adecuadas. Por tanto, la estacionalidad con
que se produce el desove es de vital importancia para el futuro desarrollo y
supervivencia de la descendencia.
Si los acociles buscan optimizar la producción de descendencia viable tienen
que procurar que ésta se desarrolle en el medio adecuado para favorecer su

42
sobrevivencia. Las crías deben eclosionar en un entorno que proporcione
alimento apropiado, protección de los depredadores y condiciones abióticas
favorables. La selección de la época de desove en cualquier especie se basa
también en la idoneidad de la condición de las hembras que deben haber
acumulado las suficientes reservas energéticas para este proceso, ya que como
en el caso de Cambarellus (C.) montezumae, las hembras ovadas permanecen
refugiadas durante todo el proceso de reproducción, desde el descenso de los
huevecillos hasta la liberación de las crías (Civera y Cruz, 1982; Begon, et al.,
2006).
En el Lago de Xochimilco las principales variaciones estacionales son la
temperatura y el fotoperiodo. Correlacionado con los cambios en estos factores
abióticos, hay normalmente variaciones en la abundancia y calidad del
alimento, así como en la densidad de la vegetación riparia que sirve de refugio
para el acocil Cambarellus (C.) montezumae (García-Padilla, 2010).
Las crías, debido a su tamaño y limitada capacidad locomotora, deben
alimentarse de alimento altamente nutritivo y de tamaño apropiado. Cuanto
más rápido se desarrollen los huevos y las crías, más rápido dejarán de ser
vulnerables a la depredación. Así pues, una primera estrategia es producir
huevos con gran contenido de vitelo y en las condiciones abióticas que permitan
un rápido desarrollo del huevo y la cría; y en un medio en el que exista la
suficiente cantidad de alimento para las crías (Saborido-Rey, 2005).
Otro aspecto muy importante es la compartamentalización de la energía
obtenida por el alimento que realizan los reproductores del acocil Cambarellus
(C.) montezumae, aproximadamente el 95% de la energía ingerida es digerida.
La energía digerida se subdivide en excreción nitrogenada y heces (10%) y
energía metabolizable (85%). A su vez, ésta tiene los siguientes destinos:

43
metabolismo basal y metabolismo activo (45%), crecimiento (0-40%) y
reproducción (0-40%), indicando entre paréntesis los porcentajes con respecto a
la energía ingerida (García-Padilla, 2010).
Con el comienzo de la actividad reproductora el individuo entra en una fase en
la que una parte importante de los recursos disponibles para crecimiento hasta
entonces, deben ser asignados a las tareas de la reproducción. Estas tareas
comprenden: formación de gónadas, búsqueda de pareja, acoplamiento,
búsqueda de refugio, cuidado maternal, etc. La energía destinada para la
formación de la gónada va a determinar la fecundidad y la calidad de las cría
(Saborido-Rey, 2005).
La fecundidad es el eslabón principal entre las estimaciones cuantitativas de
huevos y crías y la estimación del tamaño del stock o reclutamiento de la
población. Para calcular la biomasa reproductora el primer paso radica en
conocer la fecundidad específica, determinando el número de huevos puestos
por hembra. La segunda fase es determinar por mes o estación la probabilidad
de que una hembra adulta logre las condiciones necesarias para un desove
efectivo, es que se puede expresar por medio del índice de desove (Saborido-
Rey, 2005). La fecundidad también se utiliza para estimar la sobrevivencia,
para determinar el número de individuos necesario para mantener un
reclutamiento a nivel sostenible. Los datos de fecundidad generalmente se
encuentran relacionados con la talla, el peso o la edad de la hembra progenitora
(Begon, et al., 2006).
La fecundidad de una especie es el resultado de un proceso evolutivo, en el que
se optimiza el reparto de energía obtenida por la alimentación entre el consumo
metabólico, el crecimiento y la reproducción y tiene un carácter adaptativo en
relación a las estrategias vitales de r y k.

44
Las especies con estrategia “r” están adaptadas a ambientes muy variables e
impredecibles, ponen muchos huevos de pequeño tamaño, tienen una puesta
prolongada y una mortalidad larvaria muy elevada. Esta estrategia les
permite sobrevivir en cuanto se dan condiciones ambientales favorables. Las
especies estrategas “k” ocupan ambientes más estables o predecibles, ponen
menos huevos, de mayor tamaño y en puestas cortas. De alguna forma están
más protegidos y su mortalidad larvaria previsiblemente es menor (Begon, et
al., 2006).
Por tanto, las opciones posibles para asegurar la descendencia se reducen a dos:
aumentar el número de huevos o aumentar su tamaño. En este sentido el
acocil Cambarellus (C.) montezumae se ha considerado estratega “r” (Rangel,
2004 y 2009).

45
6.2 JUSTIFICACIÓN.
Es de amplio reconocimiento que los primeros estadios vitales (huevos, crías y
etapas tempranas de juveniles) son críticos para las fluctuaciones en la
abundancia de las poblaciones naturales. En los últimos años se ha extendido
una nueva visión acerca de la importancia maternal (fecundidad, rendimiento
reproductivo, calidad de los huevos y larvas, cuidado maternal y costos
bioenergéticos) en los procesos de reclutamiento, comprobándose que tienen
una importante influencia en las fluctuaciones de las poblaciones. El estudio
de la ecología de la reproducción del acocil Cambarellus (C.) montezumae es de
vital importancia para entender la dinámica de la población natural de este
acocil en el Lago de Xochimilco.
6.3 OBJETIVOS.
6.3.1 OBJETIVO GENERAL.
Conocer la estrategia reproductiva de la población de acocil Cambarellus
(C.) montezumae en el Lago de Xochimilco, desde el punto de vista de la
ecología reproductiva.

46
6.3.2 OBJETIVOS PARTICULARES.
Determinar la variación del número de organismos y su proporción
sexual en época cálida y fría del año.
Caracterizar la estructura poblacional de Cambarellus (C.) montezumae
en el lago de Xochimilco.
Establecer la evolución mensual del número de desoves de hembras
ovígeras de Cambarellus (C.) montezumae en época cálida y fría del año.
Evaluar la fecundidad de Cambarellus (C.) montezumae en función de su
talla, peso corporal y amplitud del abdomen.
Determinar la calidad del huevo y de las hembras ovígeras de
Cambarellus (C.) montezumae en época cálida y fría del año.
Evaluar la repuesta fisiológica de hembras ovígeras de Cambarellus (C.)
montezumae.

47
6.4 MATERIAL Y MÉTODO.
6.4.1 ÁREA DE ESTUDIO.
Antecedentes. El presente estudio se realizó en la zona lacustre de la
delegación Xochimilco, específicamente en el Canal de Cuemanco. La zona
lacustre de Xochimilco ha sido tradicionalmente reconocida como atractivo
turístico nacional e internacional, ya que representa un enorme patrimonio
cultural como jardín prehispánico y como vestigio histórico del sistema de
chinampas (forma de cultivo único en el mundo por su alto grado de integración
ambiental al antiguo funcionamiento hidrológico de esta cuenca), además de
albergar tradiciones y creencias religiosas únicas, representa también el último
testimonio precuauhtémico de lo que fueron las grandes culturas Xochimilca y
Mexica, así como uno de los últimos reductos del antiguo Lago del Valle de
México (Ezcurra, 1990).
Hoy sus remanentes más significativos están representados por las zonas
agrícolas y de chinampas de los ejidos de Xochimilco y San Gregorio Atlapulco.
A pesar del impacto ambiental al que ha sido sometida esta zona debido a la
presión demográfica y urbana, aún conserva características bióticas, abióticas y
culturales que la identifican como una de las reservas ecológicas más
importantes del Distrito Federal, ya que ofrece un gran número de servicios
ambientales a los habitantes del Valle de México, como son:
Dilución de contaminantes como el C02.
Un control de inundaciones.
Regula la temperatura del Valle de México y
Conserva la flora y fauna endémica.

48
Ubicación Geográfica. La zona lacustre de la delegación Xochimilco (Canal
de Cuemanco), se encuentra ubicada al sur de la Ciudad de México, entre las
coordenadas 19° 09’ de latitud norte y 98° 58’ y 99° 10’ de longitud oeste. La
delegación Xochimilco tiene una extensión territorial de 117.70 Km2, lo cual
representa el 7.9 % del territorio de la Ciudad de México. Se encuentra
delimitada al norte por las delegaciones de Coyoacán e Iztapalapa, al poniente
por Tlalpan, al sur por Milpa Alta y al oriente por Tláhuac (INEGI, 2008).
Clima. El clima presente en esta región se ha clasificado como C(w2)(w)b(i’),
templado semi-seco y sin estación invernal bien definida, este tipo de clima es
la variante menos húmeda de los templados (según la clasificación Köppen
modificada por García en 1988).
La precipitación pluvial en la región de estudio presenta niveles menores de
700 mm/año, la temporada de lluvias se ubica entre los meses de mayo y
octubre; en los demás meses sólo se registran lluvias esporádicas y aisladas.
La mayor precipitación pluvial se registra en junio con un valor que oscila
entre 100 y 110 mm y la mínima en febrero y diciembre, con valores menores a
5 mm. Las temperaturas mínimas se presentan en los meses de diciembre y
enero (7.5 °C), mientras que las más altas en los meses de abril, mayo y junio
(23.2 °C), la temperatura promedio en la zona es de 15.8 °C (INEGI, Inédito).
Hidrología. El lago de Xochimilco pertenece a la región del Pánuco, en la
cuenca hidrológica del Río Moctezuma y la subcuenca Lago Texcoco-Zumpango.
Se estima una longitud aproximada de 203 km de canales conectados entre sí;
entre los más importantes se encuentran: Cuemanco, Nacional, Chalco, Bordo,
Apatlaco, San Sebastián, Ampampilco, Texhuilo, Zacapa, Caltongo, Santa Cruz
y Japón. Las lagunas principales son Caltongo, Del Toro y el Lago de

49
Conservación de Flora, Fauna y Acuacultura de San Gregorio Atlapulco
(INEGI, 1998).
Actualmente, los canales y lagunas son alimentados artificialmente con agua
tratada de las plantas del Cerro de la Estrella, cuyo aporte es de 1 m³/s; San
Luis Tlaxialtemalco, que aporta 0.225 m³/s; además de la planta de San
Lorenzo Tezonco. Otra fuente de abastecimiento son los escurrimientos
superficiales originados por los ríos Santiago, San Lucas y San Gregorio que se
forman en las estribaciones de la Sierra Ajusco-Cuautzin (INEGI, 2008).
Vegetación. Vegetación Acuática y Subacuática. Está integrada por capas de
las especies Lemna minúscula (Lentejilla), Wolffia columbiana (chilacastle) y
Eichhornia crassipes (lirio acuático); el lirio es la especie más prolífica y su
abundancia depende de la época del año. En canales y lagunas son
característicos los “tulares” (Typha latifolia; Scirpus americanus y en menor
proporción Cyperus sp., Juncus sp., entre otras), constituyendo un elemento de
gran relevancia en la dinámica del Sistema (Ficha Ramsar, 2004).
Vegetación Terrestre. En esta zona existe vegetación asociada a las corrientes
de agua y común en las chinampas compuesta por especies leñosas como
Salixbon plandiana (ahuejote); asímismo, se registran especies introducidas
que se desarrollan en sitios propiamente secos, entre ellas Taxodium
mucronatum (ahuehuete), Salix babylonica (sauce llorón), Cassuarina
equisetifolia (casuarina), Eucaliptus sp. (eucalipto) y Fraxinu sudhei (fresno),
(Ficha Ramsar, 2004).
El cultivo sobre las chinampas es característico de esta región como la
floricultura (margarita, pensamiento, crisantemo, estate, rosa, alcatraz,

50
zempoalzúchitl y otros), también se cultivan verduras y legumbres (calabaza,
lechuga, espinaca, haba, frijol, rábano y acelga), así como cereales (Luna, 1988).
Fauna. En la zona lacustre de Xochimilco, la fauna terrestre, acuática y aérea
fue muy abundante; había gran cantidad de aves (Podilimbus podiceps, Anas
acuta, Anas platyrhynchos diazi, Accipiter striatus, Buteo jamaisensis,
Casmerodius albus, Egretta caerulea, Gallinula chroropus, Jacana spinosa,
Tyto alba, Calothorax pulcher, entre muchas otras especies), tlacuaches
(Didelphis virginiana), liebres, roedores, tigrillos, venados, tepexcuintles,
carpas de gran tamaño, ajolotes (Ambystoma mexicanum y Ambystoma
tigrinum), truchas, acociles (Cambarellus (C.) montezumae), ranas (Rana
montezumae, Rana tlaloci), culebras (Thamnophis eques, Thamnophis scaliger,
Toluca lineata), víboras (Crotalus polistictus y Sistrurus ravus) y tortugas
(Kinosternon hirtipes).
Desafortunadamente en la zona existen poblaciones animales que se han visto
reducidas como son las dos especies de ajolote (A. mexicanum y A. tigrinum; la
rana de Moctezuma y la rana de Xochimilco, así como el acocil (Cambarellus
(C.) montezumae). En la actualidad es rara su presencia a causa del deterioro
del medio ambiente y la excesiva pesca a que estuvieron sometidos durante
décadas (Ficha Ramsar, 2004).
Situación ecológica del Lado de Xochimilco. En 1936 y 1942 se emitieron
decretos que identificaban a Xochimilco “Zona Típica, Pintoresca de Belleza
Natural” cuya conservación es de interés público. El Departamento del
Distrito Federal, en sus Programas Parciales de Desarrollo Urbano señaló a la
región chinampera dentro de los límites de conservación ecológica. Por otra

51
parte, la UNESCO declaró la región chinampera de Xochimilco como
Patrimonio Mundial de la Humanidad el 11 de diciembre de 1987, esto tuvo
como propósito proteger, conservar y rehabilitar esta zona con la cooperación de
la comunidad internacional. Otro aspecto importante de la zona lacustre de
Xochimilco, es su incorporación a la clasificación de Humedales de Importancia
Internacional; el 2 de febrero de 2004 esta zona fue incorporada a la Lista de
Sitios Ramsar (Convenio Ramsar para la conservación y protección de
Humedales a nivel Mundial).
A pesar de los esfuerzos por conservar la zona lacustre de Xochimilco, se ha
perdido el control hidrológico debido al manejo inadecuado del acuífero que ha
generado hundimientos diferenciales en la zona, provocando el desnivel de los
canales y zanjas y la inundación de los terrenos. Asimismo, el agotamiento de
los manantiales que alimentaban el sistema y el aporte actual de aguas
residuales tratadas, ocasionaron la pérdida de la calidad del agua y la
degradación física y química del suelo (Ficha Ramsar, 2004).
El problema se ha acentuado debido al cierre de canales y zanjas. La
circulación del agua también está siendo afectada por el crecimiento excesivo
de vegetación acuática y por la acumulación de azolves. En general, la zona
lacustre de Xochimilco presenta eutroficación con alto contenido de sales y baja
proporción de sodio (Jiménez, 1989). En cuanto a la contaminación biológica se
han obtenido registros altos de coliformes fecales que rebasan los niveles
permisibles (Coutiño, 1981). Respecto a los metales pesados, en la zona
lacustre se han reportado elementos en concentraciones mayores a las
permisibles, tal es el caso de registros de cadmio y zinc (Rangel, 1981; Flores,
1982); asimismo se han registrado altos contenidos de bicarbonato, níquel y
cobre y de arsénico (Palacios et al. 1995).

52
La flora y la fauna nativas han sido afectadas por el deterioro del hábitat (agua
y suelo) y por la introducción de especies exóticas. Según Novelo y Gallegos
(1988) 26 especies de plantas acuáticas, de un total de 67 pertenecientes a 20
familias, están extintas de la región chinampera del sureste de la Cuenca de
México, y 8 especies tienen altas posibilidades de extinguirse en pocos años, si
continúan las condiciones de degradación existentes. La introducción del “lirio
acuático” a la zona lacustre de Xochimilco a fines del Siglo XIX, ha ido
sustituyendo a varias especies de plantas acuáticas nativas convirtiéndose en
una plaga por su alto grado de adaptabilidad a las condiciones físico-químicas
de los canales (Ficha Ramsar, 2004).
La vegetación terrestre también ha sido disminuida por la presencia de plagas
que dañan a los ahuejotes (árbol típico del paisaje chinampero), principalmente
el muérdago (Cladocolea loniceroide) y el malacosoma (Malacosoma incurvum
var. azteca). En cuanto a la introducción de especies exóticas se registran
varias especies de palmas (Phoenix canariensis y Washingtonia robusta),
casuarinas (Casuarina equisetifolia), pirul (Schinus molle), ficus (Ficus
benjaminia), higo (Ficus carica), hule (Ficus elastica), jacarandá (Jacaranda
mimosaefolia), ricino (Riccinus communis), fitolaca (Phytolaccai cosandra),
entre las más frecuentes. La pérdida del arbolado de la zona chinampera, por
las causas antes mencionadas, afecta el equilibrio ecológico y el paisaje típico
de la localidad dado que en el sistema agrícola de chinampas, las raíces del
ahuejote son las que retienen el sustrato de la misma (Ficha Ramsar, 2004).
Desde la época prehispánica la zona se distinguía por su gran productividad
agrícola que abastecía de alimentos a México-Tenochtitlán; sin embargo, por los
problemas citados, actualmente se utiliza aproximadamente solo un 15% de la
superficie de uso agrícola. Cerca de 350 hectáreas están afectadas por
inundación, una superficie aproximada de 500 hectáreas presenta degradación

53
por salinidad y sodicidad; cerca del 4% está ocupada por asentamientos
humanos irregulares y el resto ha sido abandonado por problemas de tenencia
de la tierra o por falta de interés. Por lo que respecta a la superficie en
producción, la problemática que se registra es debido al cambio en el tipo de
producción que se ha dado por la búsqueda de cultivos de mayor rentabilidad
(Ficha Ramsar, 2004).
El empleo de invernaderos ha conducido a un uso intensivo de agroquímicos, al
cierre de canales y zanjas, y en general al depósito de grandes cantidades de
residuos tóxicos. Por otro lado, existe la tendencia a la desaparición de la
agricultura en chinampas, que es una técnica prehispánica de bajo impacto
ambiental que da identidad a México.
6.4.2 TRABAJO DE CAMPO.
Se realizaron visitas mensuales al canal de Cuemanco en la zona lacustre de
Xochimilco, en los meses de marzo, abril, mayo, junio, julio, agosto, septiembre,
octubre, noviembre y diciembre de 2011 y enero y febrero de 2012; con el fin de
colectar organismos de la especie Cambarellus (C.) montezumae, la colecta de
estos organismos nos permitió establecer una comparación de los diferentes
parámetros evaluados tanto en época cálida como en época fría del año, lo cual
nos ayudó a disponer de información relevante para una adecuada comprensión
e interpretación de los factores que afectan el ciclo reproductivo de estos
organismos en su hábitat natural.

54
6.4.2.1 Estacionalidad.
Para establecer la estacionalidad en el área de estudio (época cálida y época
fría), se tomaron como referencia las temperaturas medias ambientales para
cada uno de los meses en la zona lacustre de Xochimilco, reportadas en el plan
Delegacional de Desarrollo Sustentable: Delegación Xochimilco (2010), las
temperaturas medias mensuales son las siguientes:
Cuadro 2. Distribución media mensual de la temperatura ambiente (°C) en la
zona lacustre de Xochimilco.
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
11.8 12.7 15.8 17.4 18.3 18.2 17.7 17.8 17.6 15.8 13.4 13.4
La época cálida está representada por los meses de abril, mayo, junio, julio,
agosto y septiembre, su temperatura media es 17.8 ± 0.35 °C y la época fría por
los meses de octubre, noviembre, diciembre, enero y febrero, con una
temperatura media de 13.8 ± 1.6 °C; la agrupación de los meses en cada una de
las épocas se llevó a cabo en base a datos reportados por García-Padilla en
2010, posteriormente se realizó una prueba de ANOVA para determinar si
existían diferencias significativas entre ambas época del año, la hipótesis nula
se analizó a un nivel de significancia de 0.05.

55
6.4.2.2 Caracterización del hábitat.
En virtud de que las condiciones del ambiente inciden en la distribución de los
recursos y usos de la energía contenida en el alimento ingerido de las especies
que habitan en los cuerpos de agua, y esto a su vez influye en parámetros como
la distribución de tallas y ciclo reproductivo, es básico entonces, efectuar un
monitoreo de las principales variables que pudieran afectarles. Por tal motivo
se midió en el sitio de colecta: la temperatura (± 0.05 °C) y el oxígeno disuelto (±
0.05 mg O2/L) con un oxímetro (YSI modelo 51B) y el pH con un potenciómetro
(HI 98129), tanto en el estrato fondo como en la superficie.
6.4.2.3 Colecta de los organismos.
Se colectaron los organismos que se obtuvieron tras 90 minutos de captura en
promedio, por medio de una red de cuchara de 1 mm de luz de malla de forma
triangular. La captura de los acociles se llevó a cabo entre las 10:00 y las
12:00 horas. Posteriormente fueron colocados en botes con agua del medio
para su traslado al área de acuarios del Laboratorio de Acuacultura y
Producción Acuática de la Facultad de Ciencias, UNAM.
Una vez en el área de acuarios se separaron las hembras ovígeras encontradas
en campo, se colocaron individualmente en botes de plástico de 2 L de
capacidad con refugios (para evitar la pérdida de huevos y apéndices) y
aireación constante a temperatura ambiente, se realizó un recambio de agua
del 25% cada tercer día, el resto de los organismos se colocaron en un reservorio
con capacidad de 75 L para promover el apareamiento. Este reservorio contó
con aireación constante y refugios, se realizó un recambio de agua una vez a la

56
semana del 20%. Tanto las hembras ovígeras como los organismos colocados
en el reservorio fueron alimentados ad libitum con alimento comercial
Camaronina (Purina, con un 40% de proteína).
6.4.3 TRABAJO DE LABORATORIO.
6.4.3.1 Caracterización de las colectas.
De los organismos colectados se obtuvieron las siguientes características
morfométricas: Longitud total (LT: desde la punta del rostro hasta la parte
final del telson), Longitud del cefalotórax (LCT: desde la punta del rostro, hasta
la parte final del cefalotórax en su región dorsal media) con un vernier (± 0.01
mm) Caliper y el peso húmedo (Ph) con una abalanza (± 0.01 gr) Ohaus, Scout
Pro (Fig. 1).
La determinación del sexo se llevó a cabo de manera visual, los datos obtenidos
se utilizaron para determinar la estructura poblacional de Cambarellus (C.)
montezumae en el Canal de Cuemanco.
Figura 1. Características biométricas del acocil Cambarellus (C.) montezumae.

57
De las hembras ovígeras obtenidas en el campo además de las medidas
mencionadas, se llevó a cabo el conteo aproximado del número de huevos, el
tiempo de incubación, el número de crías vivas eclosionadas, así como el peso
de las crías. Se tomó una hembra ovígera por cada mes de muestro para ser
congelada y posteriormente llevar a cabo análisis morfológicos y químicos
proximales de los huevecillos.
6.4.3.1.1 Estructura poblacional.
La determinación de la estructura de una población requiere uniformar
criterios para la clasificación de los organismos en clases o edades, en el caso
del desarrollo de Cambarellus (C.) montezumae aún no se cuenta con un
criterio para determinar en qué etapa de desarrollo se encuentra un
determinado individuo. Es un hecho que esta clasificación debería estar
vinculada con el concepto de edades ecológicas que, típicamente, considera la
existencia de una edad prereproductiva, reproductiva y posreproductiva
(Odum, 1987). El concepto es útil cuando es difícil establecer correlaciones
fehacientes con el tamaño del animal y la edad como es el caso de Cambarellus
(C.) montezumae.
Para determinar la estructura de la población de Cambarellus (C.) montezumae
en la zona lacustre de Xochimilco, específicamente en el Canal de Cuemanco, se
determinaron cuatro clases de tamaño que se muestran en el Cuadro 3.
Dentro de la edad prereproductiva encontramos las clases: cría, juvenil y
subadulto, en la edad reproductiva encontramos a la clase adulto en donde ya
se presentan encuentros reproductivos exitosos. Estas clases de tamaño
fueron determinadas en base a la experiencia en el manejo de la especie y en

58
observaciones realizadas en campo, así como en datos reportados en
investigaciones previas realizadas por otros autores (Rangel, 2004 y 2009;
García-Padilla, 2010; Villa, 2010).
6.4.3.1.2 Proporción sexual.
Para determinar la desviación de la proporción sexual esperada 1:1 se utilizó la
prueba Chi2. Tanto la estructura poblacional como la proporción sexual de los
organismos se analizaron por separado para cada época del año (época cálida y
época fría).
Cuadro 3. Características de cada edad ecológica del acocil Cambarellus (C.)
montezumae.
Etapa Edad Características
CríaOrganismos recién eclosionados de 2 mm hasta 10
mm de longitud total.
Prereproductiva JuvenilOrganismos de 11 mm a 18 mm de longitud total, no
presentan dimorfismo sexual aparente.
Subadulto
Organismos de 19 mm a 27 mm de longitud total, con
evidente dimorfismo sexual, pero que aun no
presentan maduración gonadal evidente.
Reproductiva Adulto
Organismos a partir de 28 mm de longitud total que
presentan fenómenos de maduración sexual (gónadas
evacuadas).

59
6.4.3.1.3 Relación peso-longitud.
Se estableció la relación peso-longitud de los organismos colectados para cada
clase talla y época de año con el fin de determinar las distintas fases de
crecimiento en peso y longitud de los acociles de acuerdo a la siguiente
ecuación:
Pt= a Lsb
En donde:
Pt= peso total (g) a una longitud estándar determinada.
Ls= longitud estándar.
a y b= valores constantes en la regresión.
El valor de la contante b permite definir el tipo de crecimiento que presentan
estos organismos, considerándose como isométrico un valor de b=3 y como
alométrico cuando presenta un valor diferente (Ricker, 1975), esta evaluación
se llevó a cabo mediante una prueba de t de Student.

60
6.4.3.2 Indicadores del desempeño reproductivo.
6.4.3.2.1 Época reproductiva.
La época reproductiva se determinó según la evolución mensual del número de
desoves, se reconoce actividad reproductiva cuando el número de desoves
alcanzan valores máximos en un ciclo anual.
6.4.3.2.2 Índice de desove.
Así mismo se calculó el índice de desove (ID) mediante la variación en el
porcentaje de hembras ovígeras en relación con el total de hembras adultas en
cada muestreo.
6.4.3.2.3 Fecundidad.
Entre los indicadores de desempeño reproductivo más importantes
encontramos a la fecundidad, que en términos biológicos se refiere a la
capacidad reproductora de un ser vivo. En este estudió la fecundidad (F) de
Cambarellus (C.) montezumae se determinó contando el número aproximado de
huevos que una hembra incuba en un momento dado durante su época
reproductiva.

61
Para representar la fecundidad se seleccionó el modelo potencial que ha sido
utilizado para numerosos crustáceos (Hines, 1988, 1991; Reid & Corey, 1991;
Palma & Arana, 1997):
F = a * Lsb
En donde:
F= fecundidad.
Ls= longitud estándar (Longitud Total).
a y b= valores constantes en la regresión.
Los datos fueron ajustados a un modelo de regresión lineal mediante la
transformación, logF =loga + blogLs, y los parámetros a y b fueron calculados a
través del método de mínimos cuadrados (Zar, 1996). Se utilizó análisis de
covarianza para comparar las pendientes entre las ecuaciones de las
regresiones lineales correspondientes a ambas épocas del año (Hamazaki, et al.,
2006).
6.4.3.2.4 Esfuerzo Reproductivo.
Otro indicador importante en el desempeño reproductivo de un organismo es el
esfuerzo reproductivo (ER), que es utilizado para determinar la relación entre
la inversión de energía en la reproducción y el tamaño de las hembras.
Este índice se estimó aplicando los índices propuestos por Parada, et al., (1987),
para invertebrados, quienes estiman el ER en base a la relación entre el peso
de la masa ovígera y el peso de la hembra. El esfuerzo reproductivo (ER) se
correlacionó linealmente con la talla corporal de las hembras tomando como

62
variable dependiente el esfuerzo reproductivo. Este índice se comparó entre
ambas épocas del año mediante una ANOVA unifactorial, la hipótesis nula se
examinó a un nivel de significancia de 0.05.
6.4.3.3 Indicadores de la calidad del huevo.
Para determinar la calidad de las masas ovígeras en ambas épocas del año se
llevó a cabo un análisis morfológico de los huevecillos (volumen del huevo y
peso húmedo) y el análisis químico proximal de su composición (composición de
lípidos y proteínas).
6.4.3.3.1 Volumen del huevo (Vh).
El volumen del huevo se calculó separando a una hembra ovígera proveniente
de cada mes de muestro, de cada uno de los huevecillos se midió el diámetro
mayor (a) y menor (b) con una rejilla milimétrica y una lupa, se descartaron los
huevos que presentaron algún grado de deterioro debido al periodo de
congelación. Los datos obtenidos se incorporaron en la fórmula Vh = 1/6 (a * b2
* π) propuesta por Reid & Corey (1991). De estos mismos huevecillos se
tomaron submuestras para llevar a cabo el análisis químico proximal.
6.4.3.3.2 Composición químico proximal del huevo.
El peso húmedo se determinó pesando cada huevecillo en una balanza digital
(± 0.001 gr) Ohaus, Scout Pro, de cada una de las hembras congeladas que se

63
utilizarían en el análisis químico proximal. Así mismo se llevó a cabo la
determinación del porcentaje de lípidos y proteínas de tres submuestras por
época del año.
El contenido de lípidos se evaluó mediante el método de Fofovainillina (2006);
para determinar la concentración de lípidos en las submuestras se realizó una
curva patrón con aceite de maíz, las muestras fueron leídas en un
espectrofotómetro Beckman DU 640 a 490 nm.
Para la determinación de proteína se utilizó el método de Nitrógeno Total de
Silicato 10023, para ello se utilizó un equipo HACH DR/780 y un reactor DRB
200, los valores obtenidos se multiplicaron por un factor de 6.25 para
determinar la cantidad de proteína total en los huevecillos, de acuerdo a lo
referido en el método Número 976.06 de la AOAC Analysis Methods (1995).
La comparación de los diversos índices en ambas épocas del año se realizó
mediante una ANOVA unifactorial, la hipótesis nula se examinó a un nivel de
significancia de 0.05.
6.4.3.4 Desarrollo embrionario.
6.4.3.4.1 Tiempo de incubación.
Durante el periodo de incubación-eclosión de la hembras ovígeras encontradas
en campo tanto en época cálida como en época fría, se realizó un monitoreo
diario de las características morfológicas de los huevecillos, se registró el
número de días en el que permanecían los huevecillos en cada uno de los

64
estadios de desarrollo embrionario. Para determinar en qué estadio del
desarrollo embrionario se encontraban, se desarrolló una clasificación basada
en la caracterización morfológica de las diferentes etapas de desarrollo del
embrión, durante el proceso de incubación-eclosión se describen 4 estadíos de
desarrollo en función de los cambios morfológicos ocurridos durante la
embriogénesis (Cuadro 4).
Cuadro 4. Características de los estadios de desarrollo embrionario del acocil
Cambarellus (C.) montezumae.
Estado de desarrollo Características
I
Huevo redondeado, vitelo uniforme y sin mancha ocular
visible.
II
Huevo ovoide, vitelo ocupa más de la mitad del huevo, mancha
ocular alargada y escasamente visible.
III
Huevo ovoide, poca cantidad de vitelo, ojos visibles bien
desarrollados e intensamente pigmentados, abdomen libre.
IV
Crías eclosionadas, completamente desarrolladas y con
movilidad.
6.4.3.4.2 Tamaño de las crías.
Una vez que las crías eran liberadas del abdomen de la madre y tenían
completa autonomía, se procedió a retirar a la hembra del reservorio y a tomar
sus características morfométricas. Aproximadamente a los 5 días posteriores a

65
la eclosión de las crías se llevó a cabo el registro de su peso húmedo, mediante
una balanza digital (± 0.001 gr) Ohaus, Scout Pro.
6.4.3.5 Respuesta fisiológica de las hembras ovígeras.
El cuidado que realizan las hembras de Cambarellus (C.) montezumae de la
masa ovígera que portan durante el periodo de incubación-eclosión es conocido
como cuidado parental, el cual se define como toda aquella conducta realizada
por uno o ambos progenitores que incrementa la supervivencia de la progenie
(Clutton-Brock, 1991). Si este incremento en la adecuación genera una
disminución en el éxito reproductivo residual de las hembras ovígeras, se le
conoce como inversión parental (Trivers, 1972). Esta disminución en el éxito
reproductivo residual puede ser debida a factores tales como: pérdidas de
oportunidad de apareamiento o alimentación, un aumento en el riesgo de
depredación y/o aumento en el gasto energético (Clutton-Brock, 1991; Tallamy
& Schaeffer, 1997; Gross, 2005).
En este sentido el propósito de este experimento fue determinar si existía un
incremento en el gasto energético de las hembras ovígeras de Cambarellus (C.)
montezumae en comparación con hembras que no se encontraban grávidas o
cargadas. Esto se determinó por medio de la cuantificación de la Tasa
Metabólica (R). Este índice fisiológico se evaluó a través del consumo de
oxígeno (QO2) de rutina expresado como una tasa, mg O2 g Ps-1 día-1 en
hembras ovígeras y hembras no grávidas bajo condición de ayuno,
pertenecientes al mes de abril en donde se presentó una mayor proporción de
hembras ovígeras en el hábitat natural.

66
Se llevó a cabo un monitoreo de 3 horas de medición del consumo de oxígeno,
debido a que no se deseaba someter a altos niveles de estrés a las hembras
ovígeras y evitar la pérdida de los huevecillos. El consumo de oxígeno se midió
en cámaras respirométricas de 120 mL de capacidad con una hembra por
cámara. Se emplearon 14 cámaras, 6 para hembras ovígeras, 6 para hembras
no grávidas y 2 cámaras control, todas sumergidas en una caja de plástico de
50 L, a manera de baño maría, para evitar los cambios bruscos de temperatura.
La medición de oxígeno se efectuó con un oxímetro YSI 51B (± 0.05 mg O2/L).
Se tomó una lectura inicial después de la cual se cerraron las cámaras, luego de
tres horas se tomó la muestra final. Se probó estadísticamente el efecto del
gasto energético por cuidado parental de las hembras ovígeras mediante una
prueba de ANOVA unifactorial.

67
6.5 RESULTADOS.
6.5.1 Caracterización del hábitat.
Se llevó a acabo un análisis de varianza de dos factores, para determinar si
existe estacionalidad (época cálida y época fría) y estratificación (estrato
superficie y estrato fondo) en el canal de Cuemanco de los parámetros físico-
químicos monitoreados: temperatura (T °C), oxígeno disuelto (O2) y potencial
de hidrogeno (pH), (Cuadro 5). Se encontraron diferencias significativas
(p<0.05) para la temperatura y el oxígeno disuelto tanto en la estacionalidad
como en la estratificación.
La época cálida presentó una temperatura media en el estrato superficie de
23.3 ± 1.3 °C y en el estrato fondo de 21.3 ± 1.2 °C, en contraste con la época
fría que presentó una temperatura media en el estrato superficie de 17.7 ± 2.4
°C y en el estrato fondo de 16.5 ± 2.1 °C. El mes más caluroso fue el mes de
junio con una temperatura media de 24.2 ± 1.6 °C y el mes más frío fue el mes
de enero con una temperatura media de 15.3 ± 0.2 °C (Fig. 2).
El oxígeno disuelto en época cálida presento valores promedio en el estrato
superficie de 9.5 ± 1.1 mg/L y en el estrato fondo de 4.1 ± 0.6 mg/L, en época
fría encontramos valores promedio para el estrato superficie de 10.5 ± 0.5 mg/L
y para el estrato fondo de 4.7 ± 0.6 mg/L (Fig. 3).
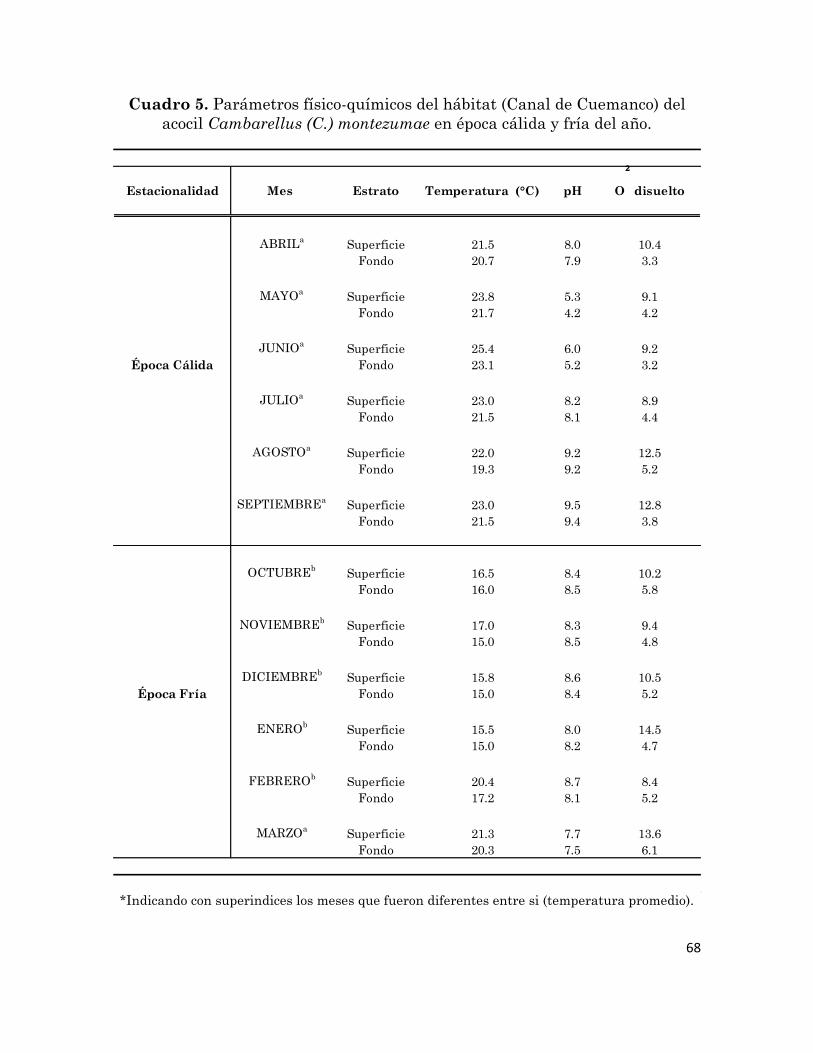
68
Cuadro 5. Parámetros físico-químicos del hábitat (Canal de Cuemanco) del
acocil Cambarellus (C.) montezumae en época cálida y fría del año.
Estacionalidad Mes Estrato Temperatura (°C) pH O
₂
disuelto
ABRILa
Superficie 21.5 8.0 10.4
Fondo 20.7 7.9 3.3
MAYOa
Superficie 23.8 5.3 9.1
Fondo 21.7 4.2 4.2
JUNIOaSuperficie 25.4 6.0 9.2
Época Cálida Fondo 23.1 5.2 3.2
JULIOaSuperficie 23.0 8.2 8.9
Fondo 21.5 8.1 4.4
AGOSTOaSuperficie 22.0 9.2 12.5
Fondo 19.3 9.2 5.2
SEPTIEMBREaSuperficie 23.0 9.5 12.8
Fondo 21.5 9.4 3.8
OCTUBREb
Superficie 16.5 8.4 10.2
Fondo 16.0 8.5 5.8
NOVIEMBREbSuperficie 17.0 8.3 9.4
Fondo 15.0 8.5 4.8
DICIEMBREbSuperficie 15.8 8.6 10.5
Época Fría Fondo 15.0 8.4 5.2
ENERObSuperficie 15.5 8.0 14.5
Fondo 15.0 8.2 4.7
FEBRERObSuperficie 20.4 8.7 8.4
Fondo 17.2 8.1 5.2
MARZOaSuperficie 21.3 7.7 13.6
Fondo 20.3 7.5 6.1
*Indicando con superindices los meses que fueron diferentes entre si (temperatura promedio).
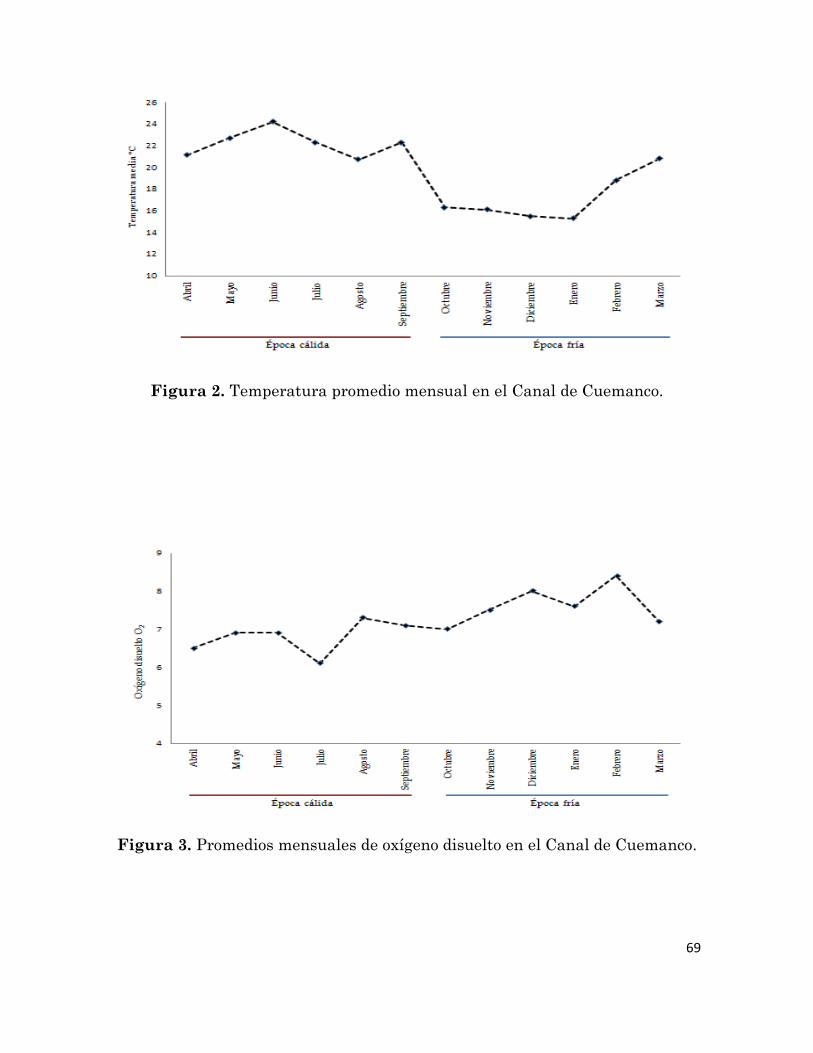
69
Figura 2. Temperatura promedio mensual en el Canal de Cuemanco.
Figura 3. Promedios mensuales de oxígeno disuelto en el Canal de Cuemanco.
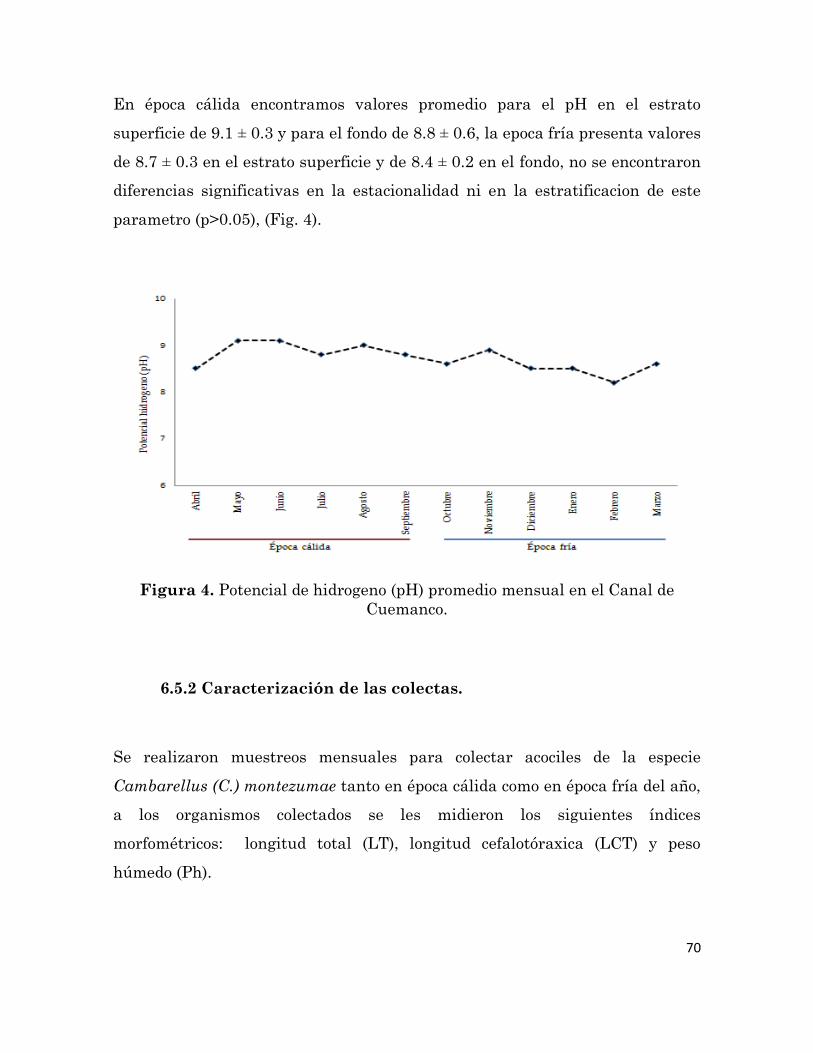
70
En época cálida encontramos valores promedio para el pH en el estrato
superficie de 9.1 ± 0.3 y para el fondo de 8.8 ± 0.6, la epoca fría presenta valores
de 8.7 ± 0.3 en el estrato superficie y de 8.4 ± 0.2 en el fondo, no se encontraron
diferencias significativas en la estacionalidad ni en la estratificacion de este
parametro (p>0.05), (Fig. 4).
Figura 4. Potencial de hidrogeno (pH) promedio mensual en el Canal de
Cuemanco.
6.5.2 Caracterización de las colectas.
Se realizaron muestreos mensuales para colectar acociles de la especie
Cambarellus (C.) montezumae tanto en época cálida como en época fría del año,
a los organismos colectados se les midieron los siguientes índices
morfométricos: longitud total (LT), longitud cefalotóraxica (LCT) y peso
húmedo (Ph).

71
Se colectaron 1,345 organismos a lo largo de los 12 muestreos; de los cuales 862
fueron organismos colectados en época cálida del año y 483 organismos
colectados en época fría del año, se encontraron diferencias significativas en el
número de organismos colectados, es decir en la abundancia, en ambas épocas
del año (p<0.05), (Fig. 5).
Figura 5. Abundancia del acocil Cambarellus (C.) montezumae en época cálida
y fría del año en el Canal de Cuemanco.
El número de organismos colectados varió en todas las colectas, sin embargo, se
puede obervar que en época cálida se colectaron 1.8 veces más organismos que
en época fría. La mayor captura se realizó en el mes de abril con 194
organismos, seguido por el mes de mayo en donde se capturaron 185
organismos; en el mes de enero se obtuvo la menor captura con 65 organismos
(Cuadro 6 y Fig. 6).
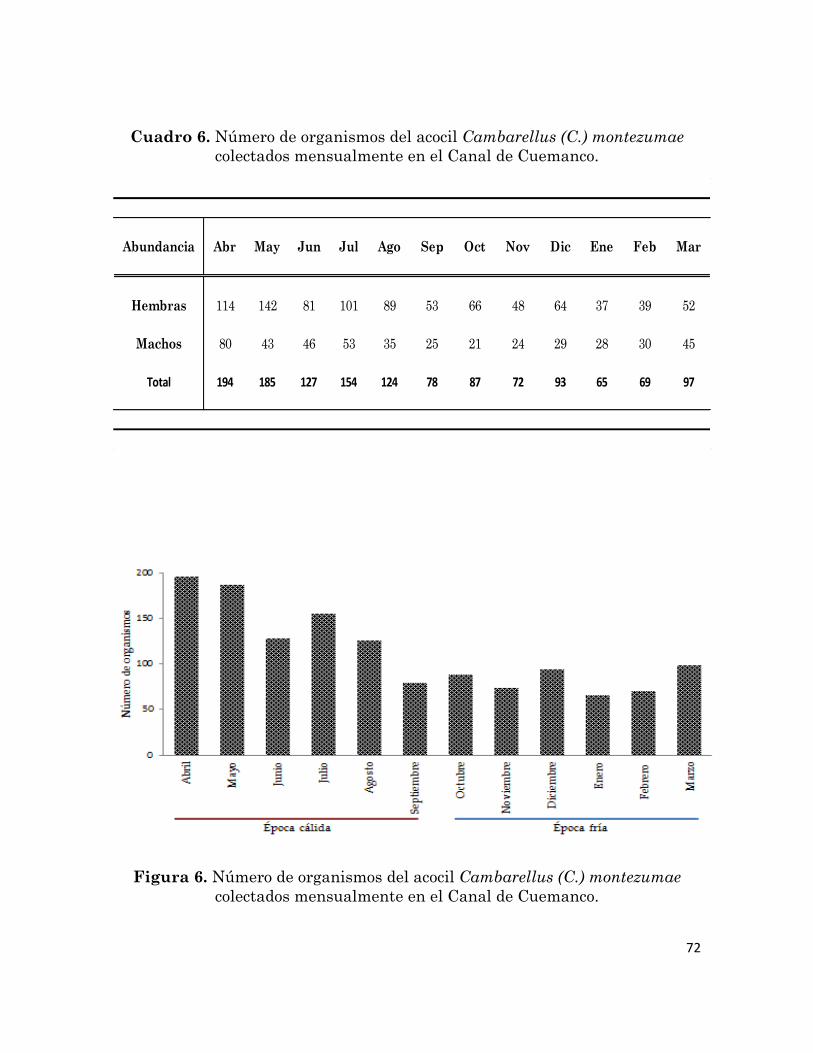
72
Cuadro 6. Número de organismos del acocil Cambarellus (C.) montezumae
colectados mensualmente en el Canal de Cuemanco.
Abundancia Abr May Jun Jul Ago Sep Oct Nov Dic Ene Feb Mar
Hembras 114 142 81 101 89 53 66 48 64 37 39 52
Machos 80 43 46 53 35 25 21 24 29 28 30 45
Total 194 185 127 154 124 78 87 72 93 65 69 97
Figura 6. Número de organismos del acocil Cambarellus (C.) montezumae
colectados mensualmente en el Canal de Cuemanco.
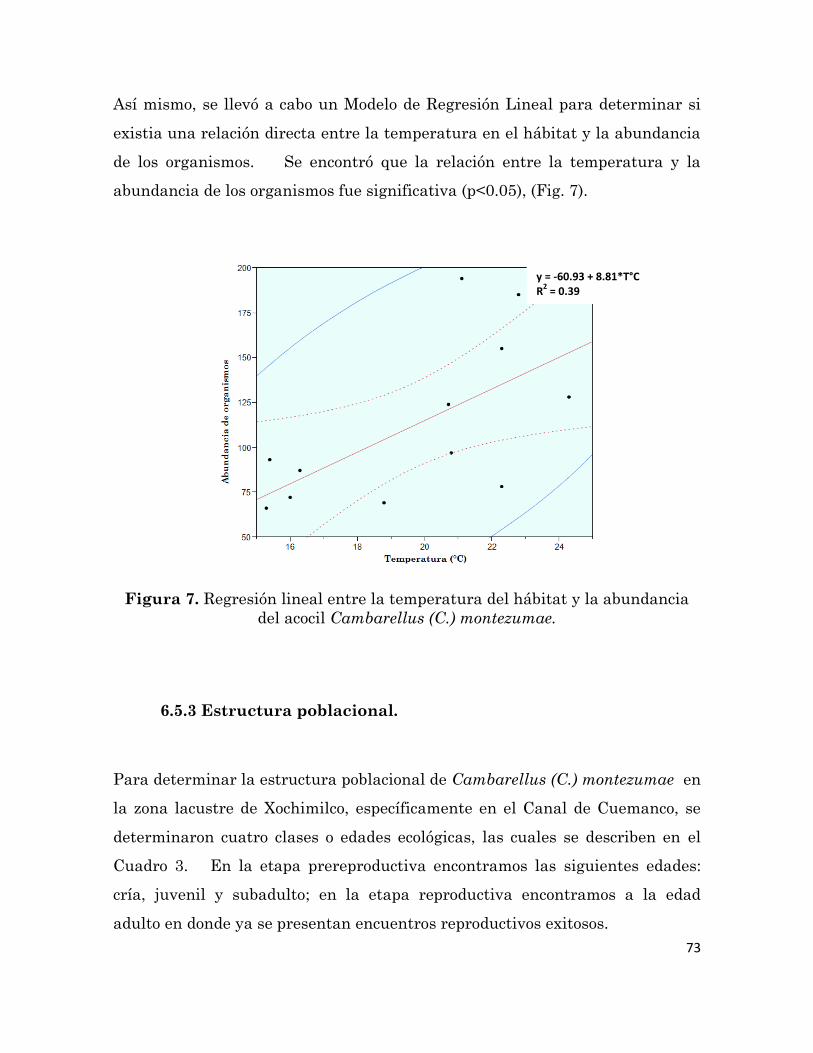
73
Así mismo, se llevó a cabo un Modelo de Regresión Lineal para determinar si
existia una relación directa entre la temperatura en el hábitat y la abundancia
de los organismos. Se encontró que la relación entre la temperatura y la
abundancia de los organismos fue significativa (p<0.05), (Fig. 7).
Figura 7. Regresión lineal entre la temperatura del hábitat y la abundancia
del acocil Cambarellus (C.) montezumae.
6.5.3 Estructura poblacional.
Para determinar la estructura poblacional de Cambarellus (C.) montezumae en
la zona lacustre de Xochimilco, específicamente en el Canal de Cuemanco, se
determinaron cuatro clases o edades ecológicas, las cuales se describen en el
Cuadro 3. En la etapa prereproductiva encontramos las siguientes edades:
cría, juvenil y subadulto; en la etapa reproductiva encontramos a la edad
adulto en donde ya se presentan encuentros reproductivos exitosos.
y = -60.93 + 8.81*T°C R
2 = 0.39
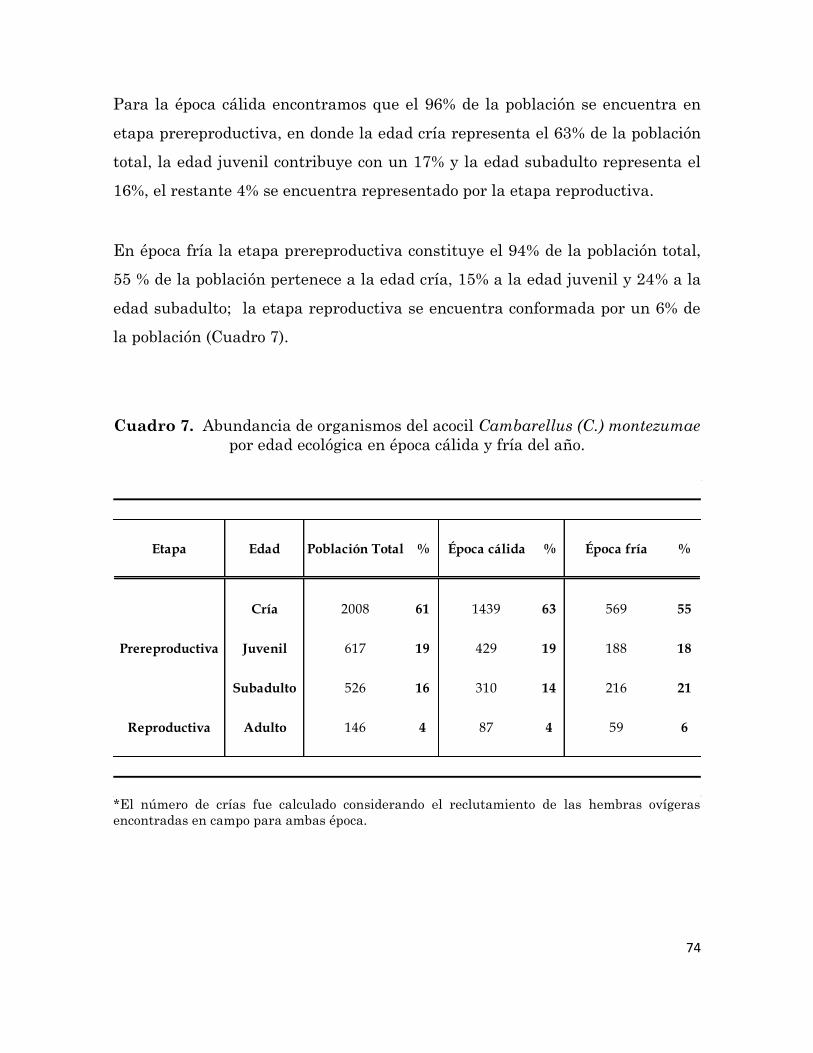
74
Para la época cálida encontramos que el 96% de la población se encuentra en
etapa prereproductiva, en donde la edad cría representa el 63% de la población
total, la edad juvenil contribuye con un 17% y la edad subadulto representa el
16%, el restante 4% se encuentra representado por la etapa reproductiva.
En época fría la etapa prereproductiva constituye el 94% de la población total,
55 % de la población pertenece a la edad cría, 15% a la edad juvenil y 24% a la
edad subadulto; la etapa reproductiva se encuentra conformada por un 6% de
la población (Cuadro 7).
Cuadro 7. Abundancia de organismos del acocil Cambarellus (C.) montezumae
por edad ecológica en época cálida y fría del año.
Etapa Edad Población Total % Época cálida % Época fría %
Cría 2008 61 1439 63 569 55
Prereproductiva Juvenil 617 19 429 19 188 18
Subadulto 526 16 310 14 216 21
Reproductiva Adulto 146 4 87 4 59 6
*El número de crías fue calculado considerando el reclutamiento de las hembras ovígeras
encontradas en campo para ambas época.

75
6.5.4 Proporción sexual.
La determinación del sexo se llevó a cabo de manera visual en cada uno de los
organismos, en la época cálida se encontramos 580 hembras y 282 machos, la
proporción sexual fue de 2:1 (2 hembras por cada macho) aproximandamente.
Para la época fría encontramos 306 hembras y 177 machos, la proporción
sexual fue la misma que para la época cálida de 2:1. Se llevó a cabo una
prueba de Chi2 para determinar si existía una desviación de la proporción
sexual esperada de 1:1 (1 hembra por cada macho), se encontraron diferencias
significativas (p<0.05), (Fig. 8).
Figura 8. Abundancia de organismos por sexo del acocil Cambarellus (C.)
montezumae en época cálida y fría.
En la figura anterior se puede observar que en todos los meses la abundancia
de hembras siempre fue superior que la de machos, en una proporción de 2:1, la
estacionalidad no tuvo influencia sobre esta proporción.

76
6.5.5 Relación Peso-Longitud.
Las relaciones talla (longitud total [mm]) y peso corporal (g) mostraron una
relación de tipo isometrico para hembras y machos en ambas épocas del año,
teniendo un mejor R2 para el modelo potencial (Pt= a Lsb), los organismos
colectados en epoca fría del año (Cuadro 8; Fig. 9 y 10).
Cuadro 8. Modelo potencial de crecimiento del acocil Cambarellus (C.)
montezumae por sexo y época del año.
Estacionalidad Sexo n LT (mm)* Peso (g)* Modelo R²
Hembras 580 23.87 ± 0.96 0.81 ± 0.04 P = 0.00002 Lt 3.02 ± 0.02
0.79
Época cálida
Machos 282 25.21 ± 1.39 0.77 ± 0.06 P = 0.00005 Lt 2.82 ± 0.08 0.81
Hembras 306 23.66 ± 1.38 0.71 ± 0.05 P = 0.00002 Lt 3.11 ± 0.01
0.95
Época fría
Machos 177 22.03 ± 1.38 0.51 ± 0.05 P = 0.00001 Lt 3.17 ±0.01 0.95
* media ± e.e.
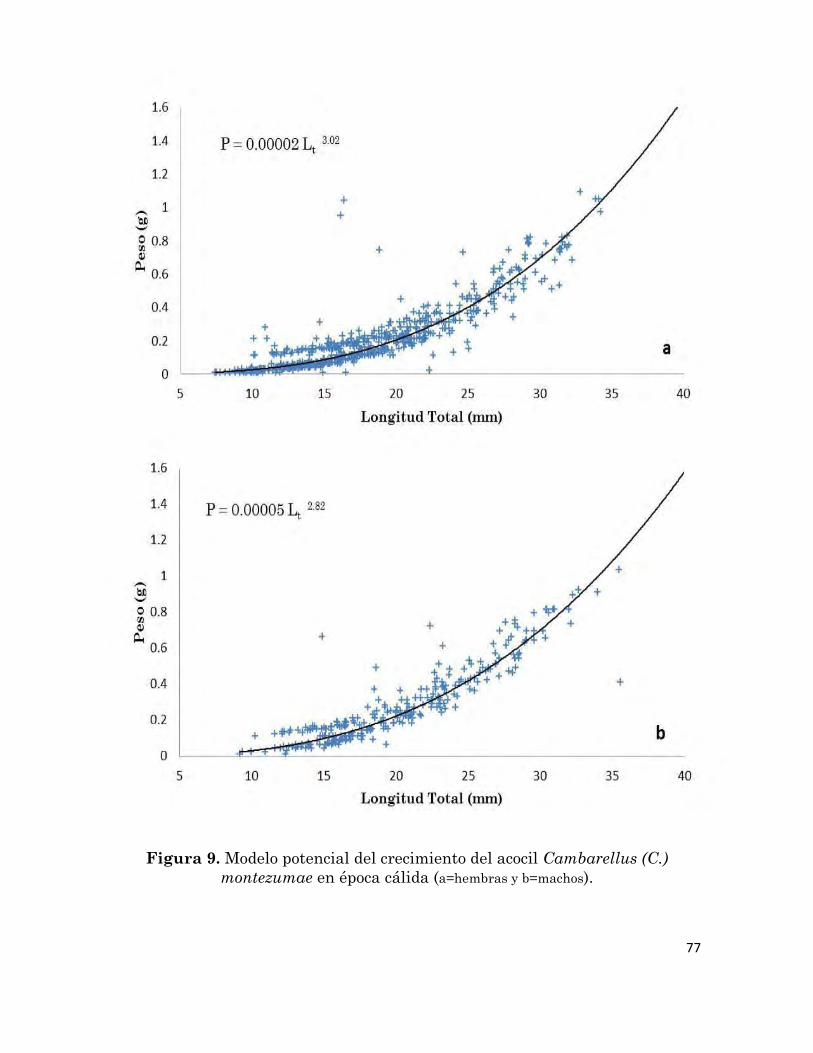
77
Figura 9. Modelo potencial del crecimiento del acocil Cambarellus (C.)
montezumae en época cálida (a=hembras y b=machos).
1.6
1.4 P = 0.00002 L, 302
1.2 +
1 + -" + ~
~ 0.8 + 1 + • +
Il. 0.6 +
OA +
+ 0.2 + +
+ a o
5 10 15 20 25 30 35 40
Longitud Total (mm)
1.6
1.4 P = 0.00005 L, 2.82
1.2
1 + - ++ " + ~
00.8 • + • Il. 0.6
0.4 +
0.2 + + .. b
O
5 10 15 20 25 30 35 40
Longitud Total <nun)
1.6
1.4 P = 0.00002 L, 302
1.2 +
1 + ~
" +
~
~ 0.8 + 1 + • +
1>. 0.6 +
OA +
+ 0.2 + +
+ a o
5 10 15 20 25 30 35 40
Longitud Total (mm)
1.6
lA P = 0.00005 L, 2.82
1.2
1 + ~
++ " + ~
0 0,8 +~ • + /'-+ + • 1>. 0.6
+ +1 +
+ + + .... OA + ., 0.2
+ + .. b O
5 10 15 20 25 30 35 40
Longitud Total <nun)
1.6
1.4 P = 0.00002 L, 302
1.2 +
1 + ~
" +
~
~ 0.8 + 1 + • +
Il. 0.6 +
OA +
+ 0.2 + +
+ a o
5 10 15 20 25 30 35 40
Longitud Total <nun)
1.6
lA P = 0.00005 L, 2.82
1.2
1 + ~
++ '" + ~
0 0,8 +~ • + /'-+ + • Il. 0.6
+ +1 + + + + t +" ....
OA + + + + ... 0.2
+ +". b O
5 10 15 20 25 30 35 40
Longitud Total <nun)
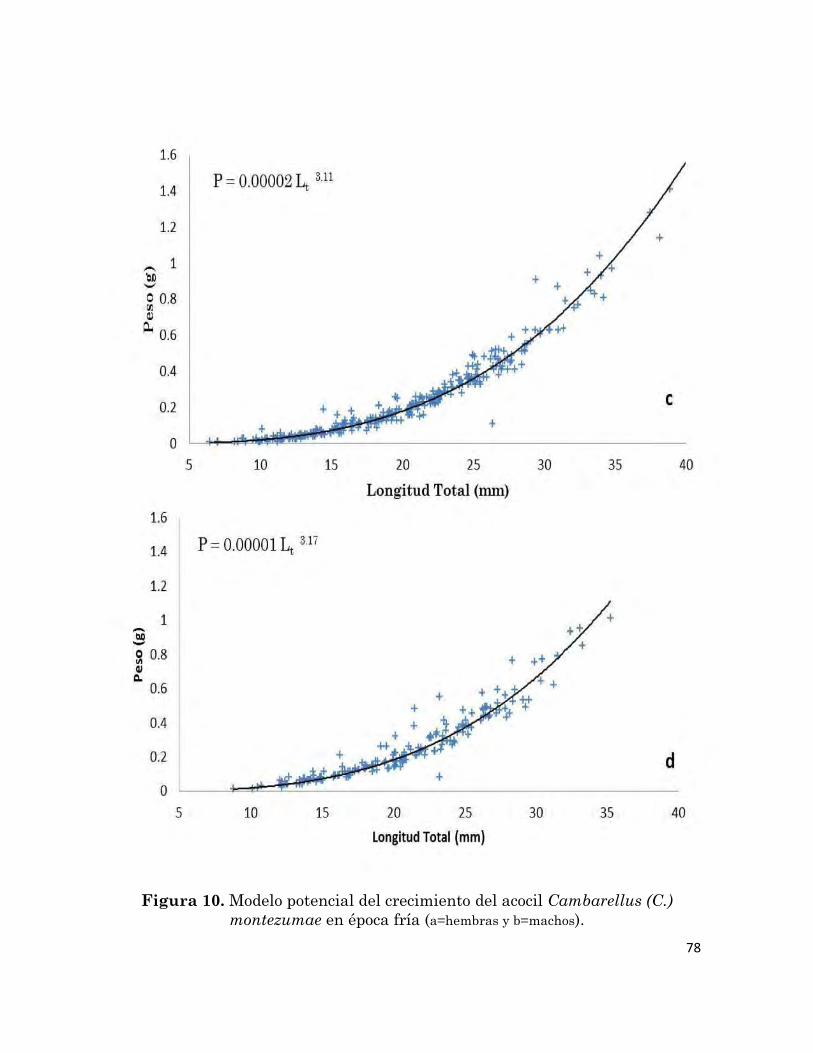
78
Figura 10. Modelo potencial del crecimiento del acocil Cambarellus (C.)
montezumae en época fría (a=hembras y b=machos).
1.6
1.4 P = O.OOOO2L, 3.11
1.2 +
1 + ~
" + + ~ + O 0.8 + ~ + • • Il. 0.6 ++
0.4 .... + e 0.2 + + +
+ + o
5 10 15 20 25 30 35 40
Longitud Total (mm)
1.6
1.4 P = 0.00001 L, 3.17
1.2
1 + ~ ++ " - + ~ 0.8 + ++ • ..
0.6
0.4
0.2 + d + + O
5 10 15 20 25 30 35 40
Longitud Total (mm)
1.6
1.4 P = 0.00002 L, 3.11
1.2 +
~ 1
" ~ + 0 0.8 • • Il. 0.6 ++
0.4 .... + e 0.2 + + +
+ + o
5 10 15 20 25 30 35 40
Longitud Total (uuu)
1.6
1.4 P = 0.00001 L, 3.17
1.2
1 + ~ ++ " - + ~ 0.8 + ++ • ..
0.6
0.4
0.2 + d +
O
5 10 15 20 25 30 35 40
Longitud Total (mm)
1.6
1.4 P = 0.00002 L, 3.11
1.2 +
~ 1
" ~ + 0 0.8 • • Il. 0.6 ++
0.4 .... + e 0.2 + + +
+ + o
5 10 15 20 25 30 35 40
Longitud Total (nun)
1.6
1.4 P = 0.00001 L, 3.17
1.2
1 + ~ ++ " - + ~ 0.8 + ++ • ..
0.6
0.4
0.2 + d +
O
5 10 15 20 25 30 35 40
Longitud Total (mm)

79
6.5.6 Indicadores de desempeño reproductivo.
6.5.6.1 Época reproductiva.
La época reproductiva se determinó según la evolución mensual del número de
desoves, se reconoce actividad reproductiva cuando el número de desoves
alcanza valores máximos en un ciclo anual.
En el ciclo anual encontramos un total de 42 hembras ovígeras en el hábitat
natural, de las cuales en época cálida se encontraron 35 hembras ovígeras que
representan el 83% de las hembras encontradas en todo el ciclo anual, el
restante 17% se encontró en época fría, con un total de 7 hembras ovígeras. El
mes que mayor actividad reproductiva fue el mes de mayo con un total de 10
hembras ovígeras, seguido del mes junio en donde se encontron 7 hembras
grávidas, tan sólo estos dos meses en su conjunto, representan el 41% de la
actividad reproductiva en el ciclo anual; en los meses de noviembre y diciembre
no se encontraron hembras ovígeras en el hábitat natural (Fig. 11).
Adicionalmente se llevo a cabo un Modelo de Regresión Lineal para determinar
si existía una relación directa entre la temperatura en el hábitat y la
abundancia de hembras ovígeras. Se encontró que la relación entre la
temperatura y la abundancia de las hembras ovadas fue significativa (p<0.05),
(Fig. 12).
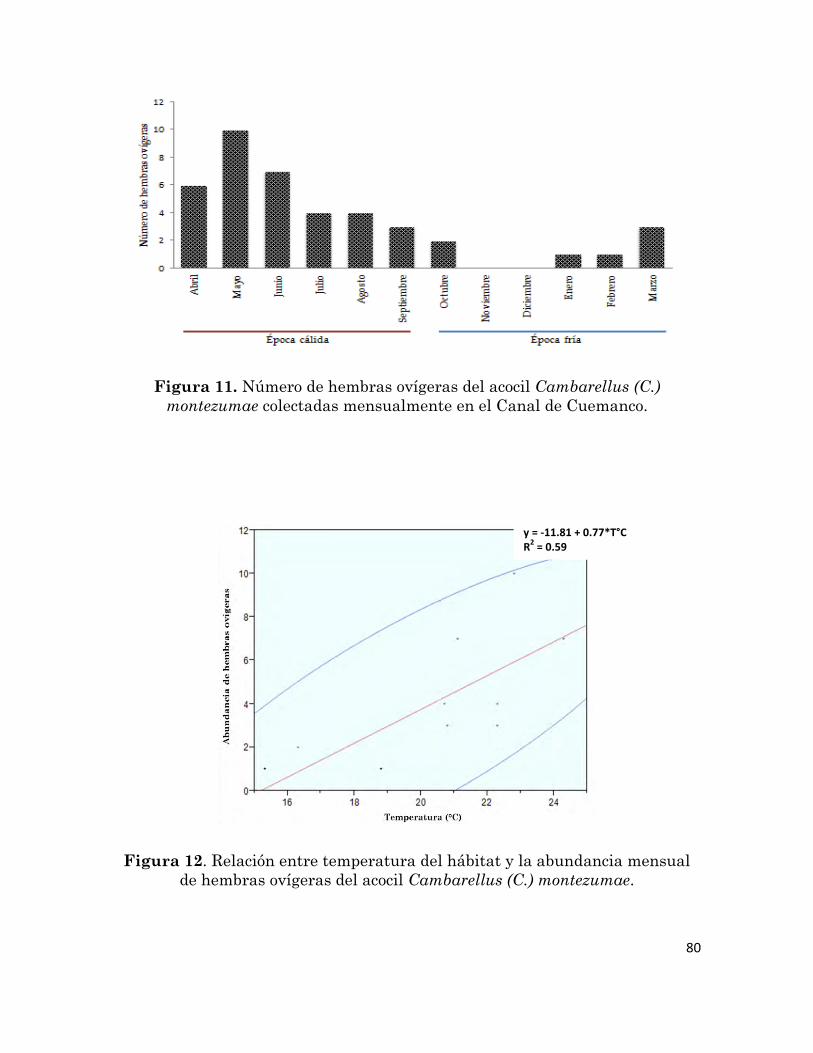
80
Figura 11. Número de hembras ovígeras del acocil Cambarellus (C.)
montezumae colectadas mensualmente en el Canal de Cuemanco.
Figura 12. Relación entre temperatura del hábitat y la abundancia mensual
de hembras ovígeras del acocil Cambarellus (C.) montezumae.
y = -11.81 + 0.77*T°C R
2 = 0.59

81
6.5.6.2 Índice de desove (ID).
Se calculó el índice de desove (ID) mediante la variación en el porcentaje de
hembras ovígeras en relación con el total de hembras en edad adulta en cada
una de la épocas del año. En época cálida encontramos un total de 49 hembras
en edad adulta, que pertenecen a la etapa reproductiva y que pueden tener
encuentros reproductivos exitosos y en época fría encontramos un total de 37
hembras; en el Cuadro 9 se muestra el número de individuos por sexo y edad
reproductiva para cada época cálida y fría del año.
Cuadro 9. Abundancia de hembras adultas del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
Estacionalidad Sexo Cría Juvenil Subadulto Adulto
Hembras 959 265 231 49
Época cálida
Machos 480 111 132 38
Hembras 379 102 147 37
Época fría
Machos 190 52 104 20
El ID para época cálida es de 0.41 y para época fría de 0.16, lo cual nos estaría
refiriendo que para época cálida el 41% de las hembras con potencial
reproductivo se están reproduciendo en comparación con el 16 % que lo hace en
época fría.

82
6.5.6.3 Fecundidad.
Debido a la importancia de la fecundidad como indicador de desempeño
reproductivo para la especie Cambarellus (C.) montezumae, se llevó a cabo una
caracterización morfométrica de las hembras ovígeras colectadas tanto en
época cálida como en época fría del año (Cuadro 10).
Las hembras ovígeras colectadas en época cálida presentaron una longitud
total promedio menor (34.31 ± 3.99 mm) que las hembras colectadas en época
fría (35.35 ± 3.65 mm), sin que esta diferencia en cuanto al tamaño (longitud
total) resultara significativa (p>0.05). El peso promedio de las hembras
ovígeras para época cálida fue de 0.97 ± 0.35 g y para época fría de 1.07 ±0.34 g,
no se encontraron diferencias significativas (p>0.05). La hembra ovígera más
grande se colecto en época cálida con una longitud total de 43.21 mm y un peso
de 1.91 g, así mismo la hembra ovígera más pequeña también se colecto en
época cálida con una longitud total de 28.31 mm y un peso corporal de 0.58 g.
En cuanto a la fecundidad (número de huevos por hembra), para época cálida
encontramos que en promedio una hembra porta 46 ± 25 huevos, en
comparación a una hembra de época fría que portaría en promedio de 49 ± 11
huevos, esta diferencia tampoco resulta ser estadísticamente significativa
(p>0.05), (Fig. 13).
En época cálida el mayor número de huevos portados por una hembra fueron
150, en comparación con una hembra de época fría donde el mayor número de
huevos fueron 65. El número total de huevos para época cálida fue de 1628 y
para época fría de 341, esto está estrechamente relacionado con el número de
hembras ovígeras encontradas en cada una de las épocas del año.
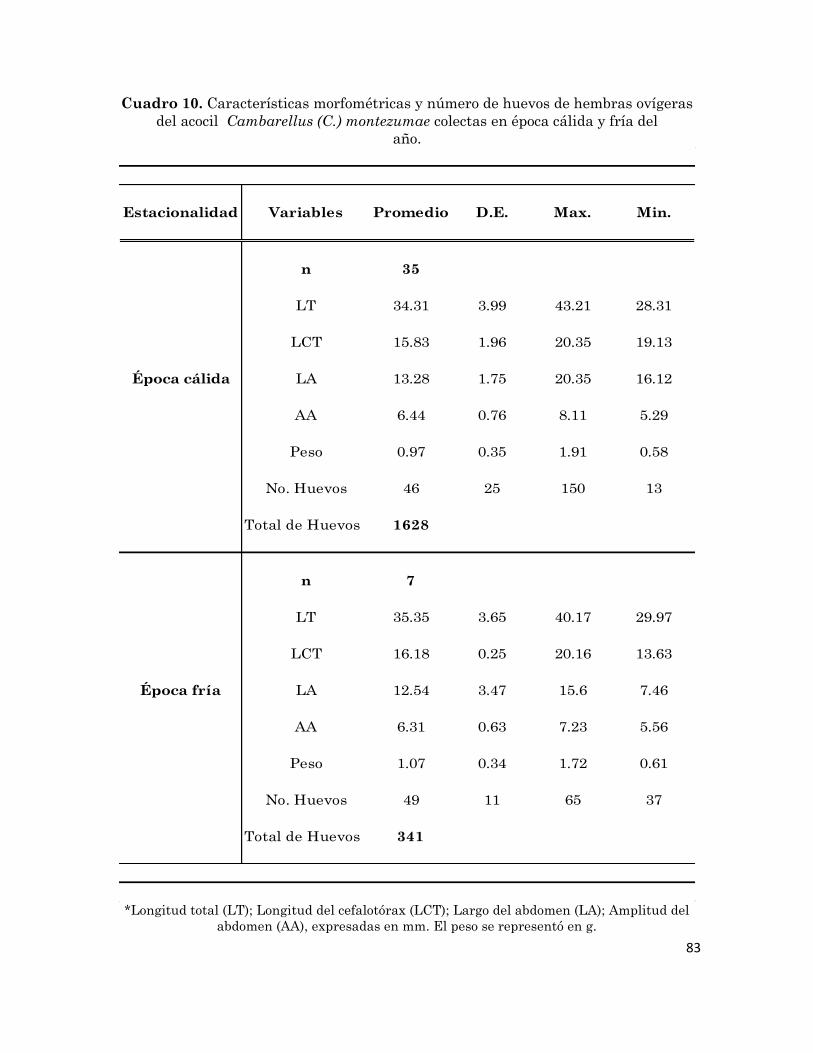
83
Cuadro 10. Características morfométricas y número de huevos de hembras ovígeras
del acocil Cambarellus (C.) montezumae colectas en época cálida y fría del
año.
Estacionalidad Variables Promedio D.E. Max. Min.
n 35
LT 34.31 3.99 43.21 28.31
LCT 15.83 1.96 20.35 19.13
Época cálida LA 13.28 1.75 20.35 16.12
AA 6.44 0.76 8.11 5.29
Peso 0.97 0.35 1.91 0.58
No. Huevos 46 25 150 13
Total de Huevos 1628
n 7
LT 35.35 3.65 40.17 29.97
LCT 16.18 0.25 20.16 13.63
Época fría LA 12.54 3.47 15.6 7.46
AA 6.31 0.63 7.23 5.56
Peso 1.07 0.34 1.72 0.61
No. Huevos 49 11 65 37
Total de Huevos 341
*Longitud total (LT); Longitud del cefalotórax (LCT); Largo del abdomen (LA); Amplitud del
abdomen (AA), expresadas en mm. El peso se representó en g.
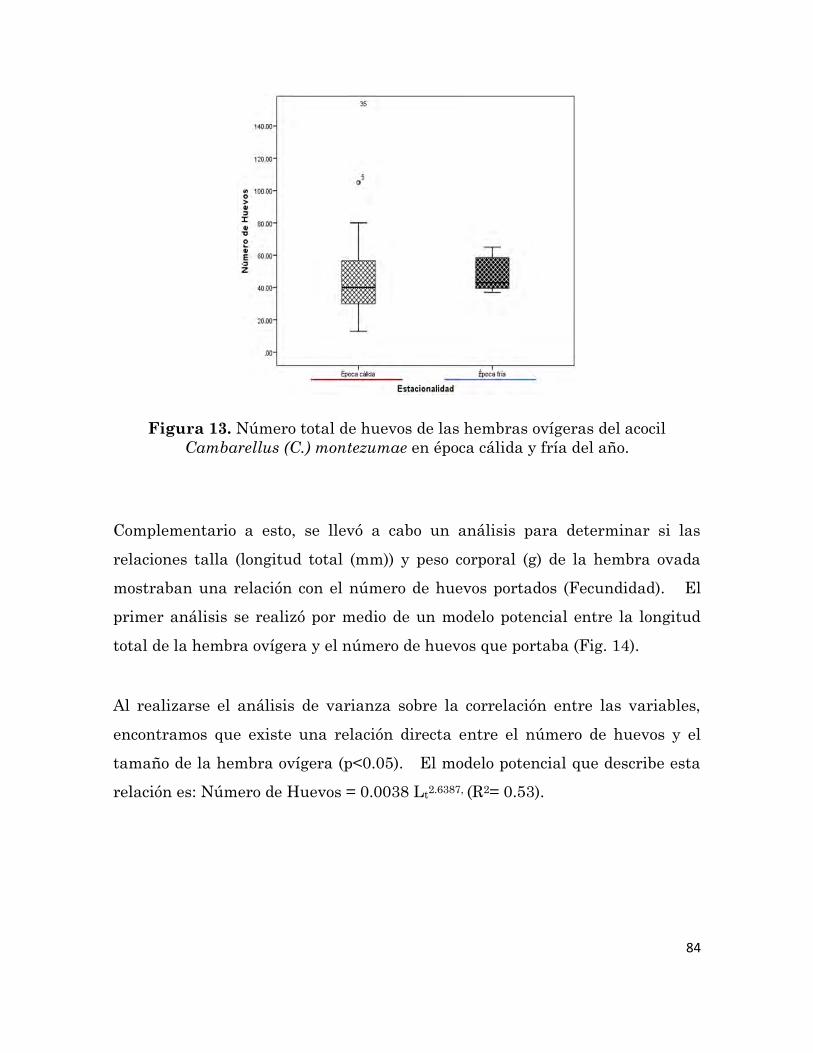
84
Figura 13. Número total de huevos de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.
Complementario a esto, se llevó a cabo un análisis para determinar si las
relaciones talla (longitud total (mm)) y peso corporal (g) de la hembra ovada
mostraban una relación con el número de huevos portados (Fecundidad). El
primer análisis se realizó por medio de un modelo potencial entre la longitud
total de la hembra ovígera y el número de huevos que portaba (Fig. 14).
Al realizarse el análisis de varianza sobre la correlación entre las variables,
encontramos que existe una relación directa entre el número de huevos y el
tamaño de la hembra ovígera (p<0.05). El modelo potencial que describe esta
relación es: Número de Huevos = 0.0038 Lt2.6387, (R2= 0.53).
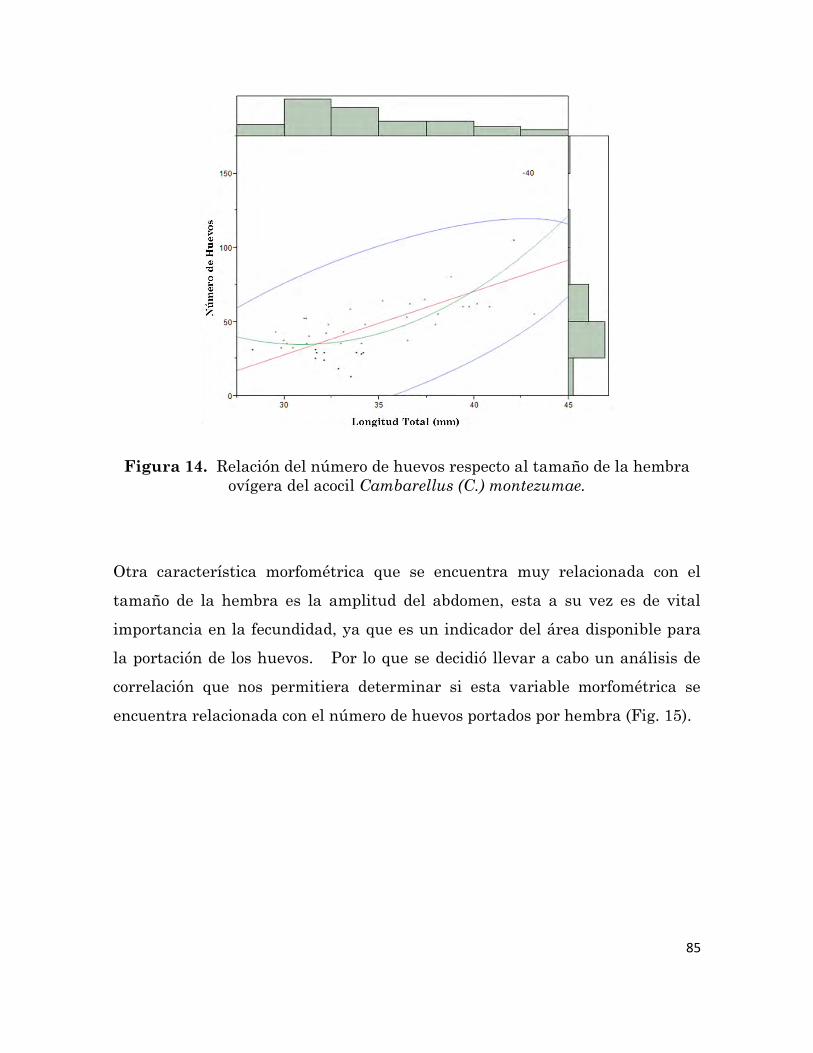
85
Figura 14. Relación del número de huevos respecto al tamaño de la hembra
ovígera del acocil Cambarellus (C.) montezumae.
Otra característica morfométrica que se encuentra muy relacionada con el
tamaño de la hembra es la amplitud del abdomen, esta a su vez es de vital
importancia en la fecundidad, ya que es un indicador del área disponible para
la portación de los huevos. Por lo que se decidió llevar a cabo un análisis de
correlación que nos permitiera determinar si esta variable morfométrica se
encuentra relacionada con el número de huevos portados por hembra (Fig. 15).
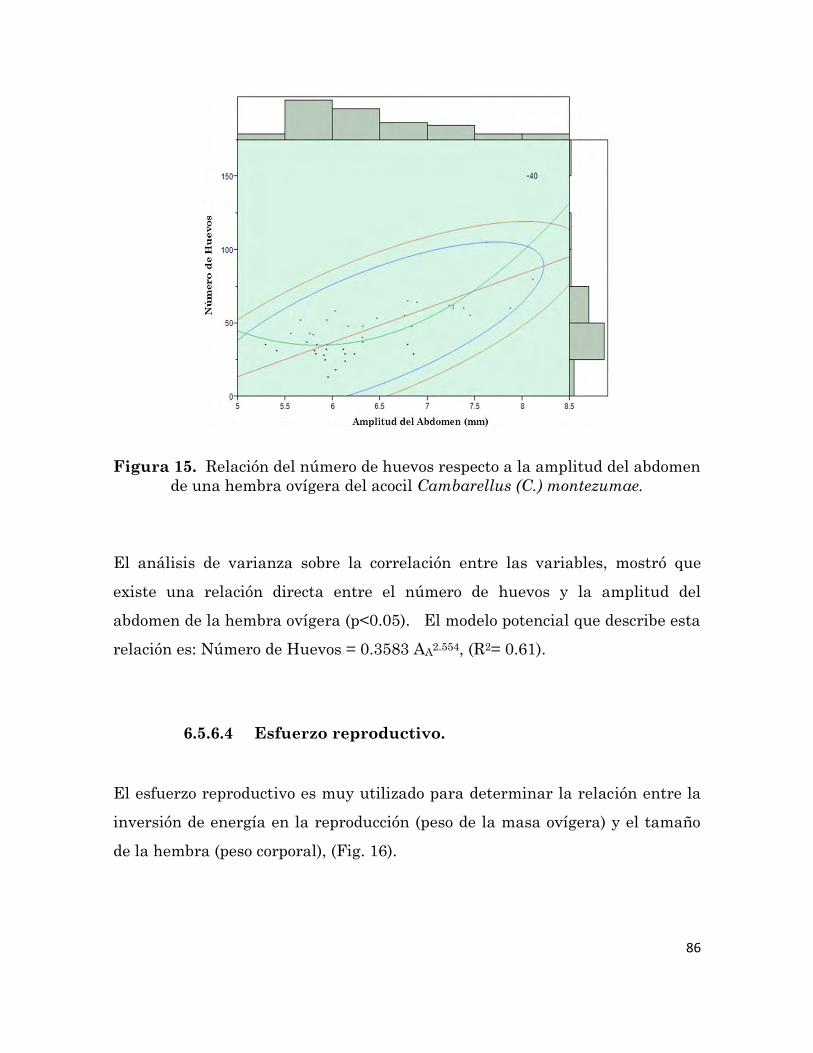
86
Figura 15. Relación del número de huevos respecto a la amplitud del abdomen
de una hembra ovígera del acocil Cambarellus (C.) montezumae.
El análisis de varianza sobre la correlación entre las variables, mostró que
existe una relación directa entre el número de huevos y la amplitud del
abdomen de la hembra ovígera (p<0.05). El modelo potencial que describe esta
relación es: Número de Huevos = 0.3583 AA2.554, (R2= 0.61).
6.5.6.4 Esfuerzo reproductivo.
El esfuerzo reproductivo es muy utilizado para determinar la relación entre la
inversión de energía en la reproducción (peso de la masa ovígera) y el tamaño
de la hembra (peso corporal), (Fig. 16).
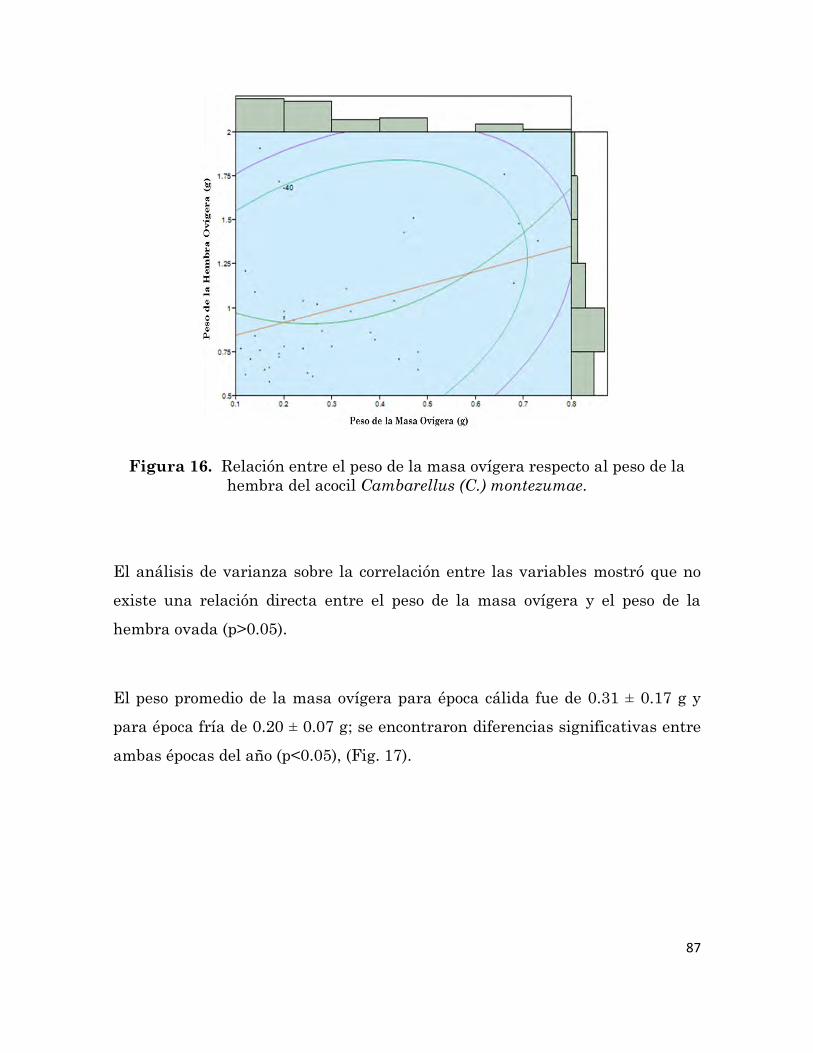
87
Figura 16. Relación entre el peso de la masa ovígera respecto al peso de la
hembra del acocil Cambarellus (C.) montezumae.
El análisis de varianza sobre la correlación entre las variables mostró que no
existe una relación directa entre el peso de la masa ovígera y el peso de la
hembra ovada (p>0.05).
El peso promedio de la masa ovígera para época cálida fue de 0.31 ± 0.17 g y
para época fría de 0.20 ± 0.07 g; se encontraron diferencias significativas entre
ambas épocas del año (p<0.05), (Fig. 17).

88
Figura 17. Peso de masa ovígera del acocil Cambarellus (C.) montezumae en
época cálida y fría del año.
6.5.7 Indicadores de la calidad del huevo.
Las características de la calidad del huevo están estrechamente relacionadas
entre sí, dado que a nivel ecológico están determinadas por el volumen, el peso,
la forma y el color, así como los parámetros internos directamente relacionados
con su composición química proximal. Para determinar esta calidad se
recurrió a la inspección de ciertos elementos del huevo tanto en su exterior
(volumen y peso) como en su interior (lípidos y proteínas).

89
6.5.7.1 Volumen del huevo.
El volumen del huevo mostró diferencias notorias entre ambas época del año,
encontrando volúmenes mayores para los huevos de hembras ovígeras
colectadas en época cálida. Al llevar a cabo la comparación estadística
encontramos diferencias significativas entre ambas épocas del año (p<0.05),
(Fig. 18). El peso húmedo de cada huevo presentó una alta relación con el
volumen del huevo, encontrando diferencias significativas entre ambas épocas
del año (Cuadro 11).
Cuadro 11. Volumen y peso húmedo del huevo del acocil Cambarellus (C.)
montezumae en época cálida y fría del año.
Estacionalidad Volumen del huevo mm³ Peso húmedo (mg)
Época cálida 0.45 ± 0.24 8.17 ± 0.79
Época fría 0.16 ± 0.09 4.35 ± 1.97
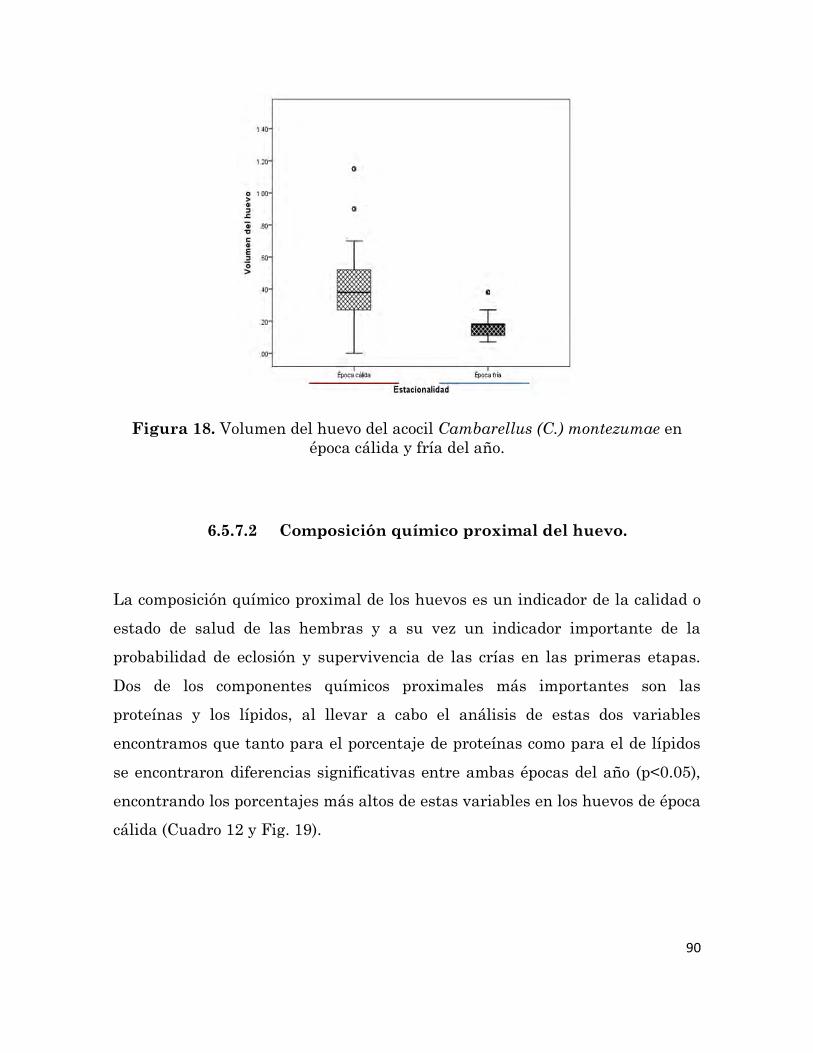
90
Figura 18. Volumen del huevo del acocil Cambarellus (C.) montezumae en
época cálida y fría del año.
6.5.7.2 Composición químico proximal del huevo.
La composición químico proximal de los huevos es un indicador de la calidad o
estado de salud de las hembras y a su vez un indicador importante de la
probabilidad de eclosión y supervivencia de las crías en las primeras etapas.
Dos de los componentes químicos proximales más importantes son las
proteínas y los lípidos, al llevar a cabo el análisis de estas dos variables
encontramos que tanto para el porcentaje de proteínas como para el de lípidos
se encontraron diferencias significativas entre ambas épocas del año (p<0.05),
encontrando los porcentajes más altos de estas variables en los huevos de época
cálida (Cuadro 12 y Fig. 19).
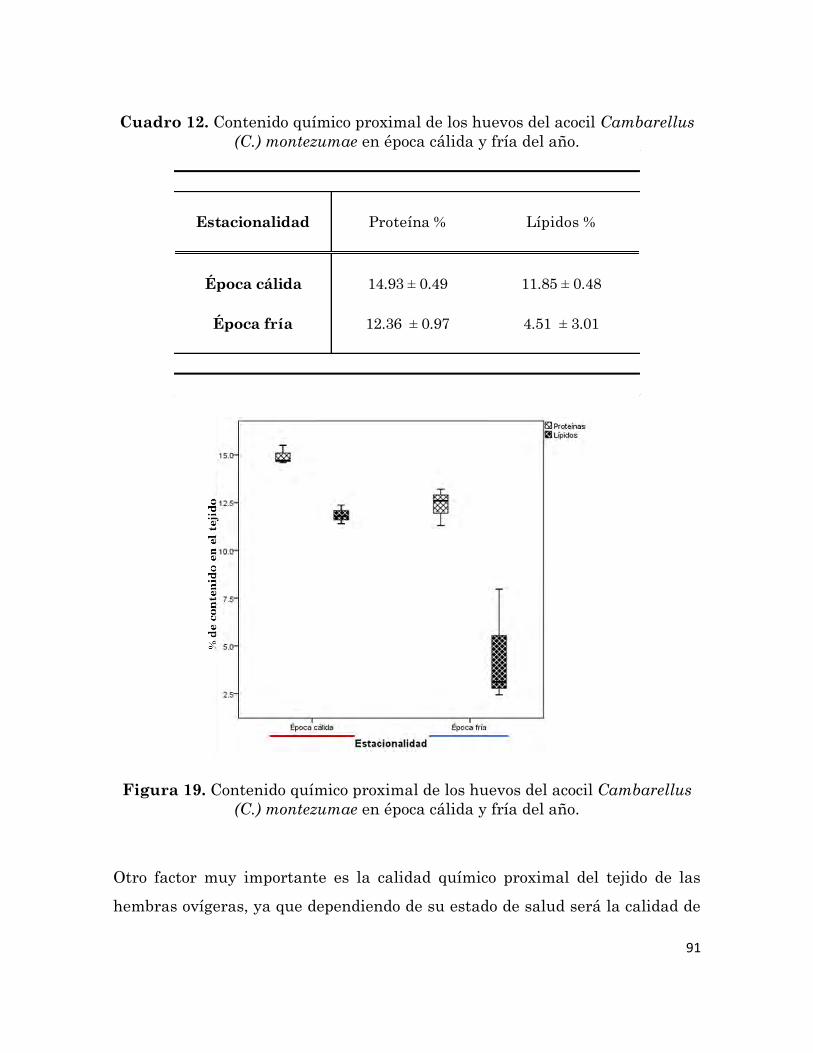
91
Cuadro 12. Contenido químico proximal de los huevos del acocil Cambarellus
(C.) montezumae en época cálida y fría del año.
Estacionalidad Proteína % Lípidos %
Época cálida 14.93 ± 0.49 11.85 ± 0.48
Época fría 12.36 ± 0.97 4.51 ± 3.01
Figura 19. Contenido químico proximal de los huevos del acocil Cambarellus
(C.) montezumae en época cálida y fría del año.
Otro factor muy importante es la calidad químico proximal del tejido de las
hembras ovígeras, ya que dependiendo de su estado de salud será la calidad de
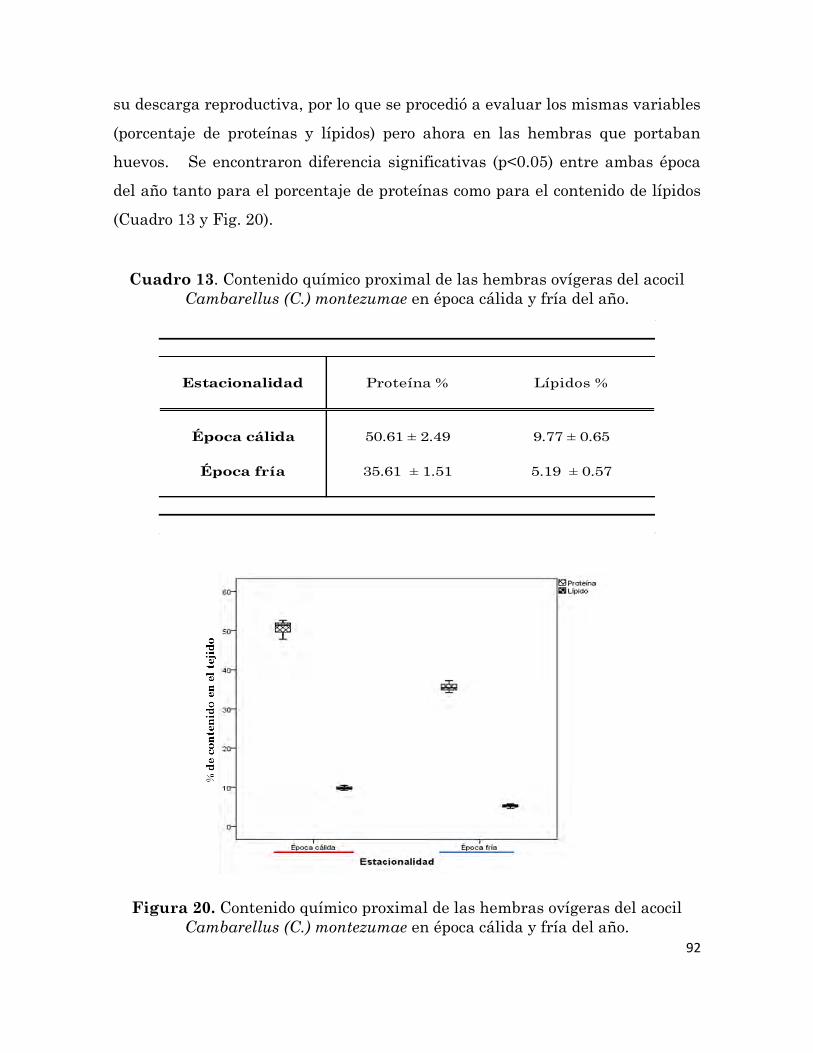
92
su descarga reproductiva, por lo que se procedió a evaluar los mismas variables
(porcentaje de proteínas y lípidos) pero ahora en las hembras que portaban
huevos. Se encontraron diferencia significativas (p<0.05) entre ambas época
del año tanto para el porcentaje de proteínas como para el contenido de lípidos
(Cuadro 13 y Fig. 20).
Cuadro 13. Contenido químico proximal de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.
Estacionalidad Proteína % Lípidos %
Época cálida 50.61 ± 2.49 9.77 ± 0.65
Época fría 35.61 ± 1.51 5.19 ± 0.57
Figura 20. Contenido químico proximal de las hembras ovígeras del acocil
Cambarellus (C.) montezumae en época cálida y fría del año.

93
6.5.8 Desarrollo embrionario.
El desarrollo embrionario del acocil Cambarellus (C.) montezumae para este
estudio se dividió en cuatro etapas que culminan con la eclosión (Fig. 21).
Estas etapas se describen en el Cuadro 4, este proceso va a estar determinado
por factores tantos bióticos como abióticos. Entre los factores bióticos que
influyen en este proceso se encuentran: la composición químico proximal de los
huevos, así como su volumen y tamaño, el cuidado que las hembras dan a la
masa ovígera y la interacción de éstas con los depredadores. Entre los factores
abióticos encontramos como factor principal a la temperatura, ya esta puede
influir de manera directa en el tiempo de incubación de los huevos, para este
estudio se monitoreo el desarrollo embrionario de Cambarellus (C.)
montezumae durante época cálida y fría del año, con temperaturas promedio de
23.3 y 17.7 °C respectivamente.
Figura 21. Desarrollo embrionario del acocil Cambarellus (C.) montezumae.

94
6.5.8.1 Tiempo de incubación.
Se monitoreo el desarrollo embrionario de 32 hembras ovígeras para época
cálida y de 4 hembras ovígeras para época fría, desde la etapa I en donde el
huevo es completamente redondo, presenta vitelo uniforme y aún no presenta
mancha ocular evidente, hasta la etapa IV, en donde el desarrollo embrionario
ha finalizado y las crías se encuentran completamente desarrolladas y con
movilidad.
Para la época cálida encontramos que el desarrollo embrionario se realiza en
un periodo promedio de 39 ± 2 días; para la época fría el período promedio de
incubación es de 46 ± 3 días; se encontraron diferencias significativas para
ambas época del año (p<0.05), (Fig. 22).
Figura 22. Tiempo de incubación del acocil Cambarellus (C.) montezumae en
época cálida y fría del año.
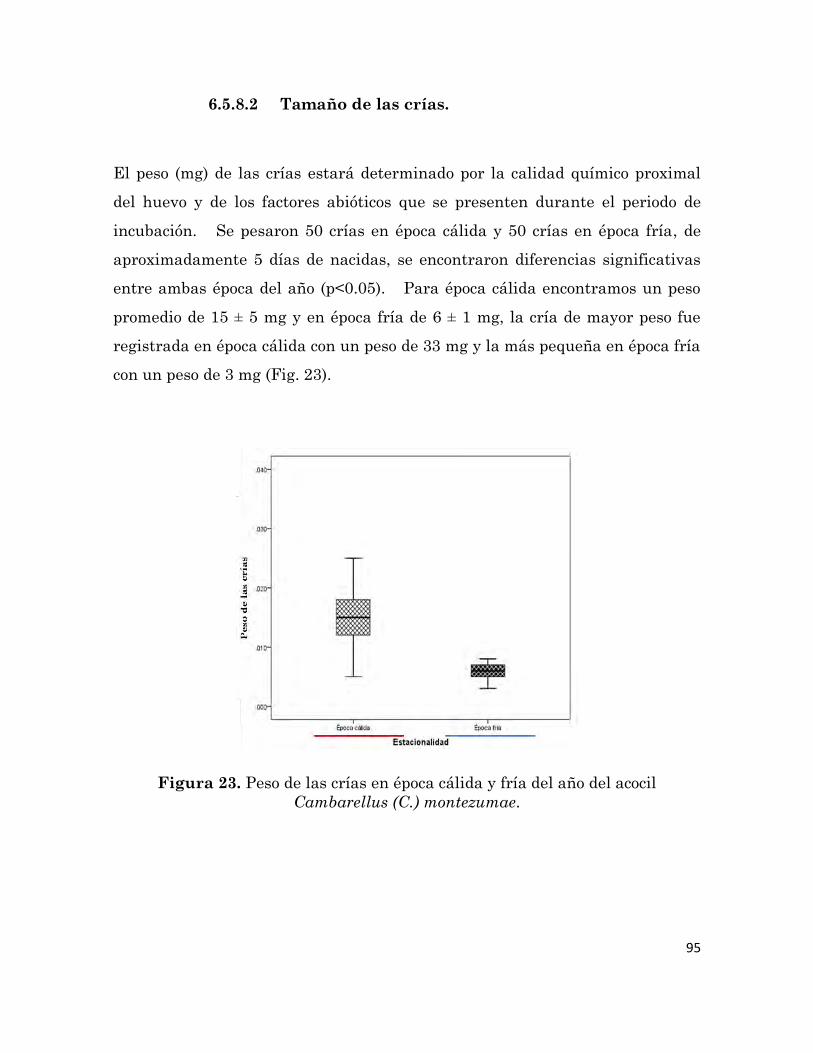
95
6.5.8.2 Tamaño de las crías.
El peso (mg) de las crías estará determinado por la calidad químico proximal
del huevo y de los factores abióticos que se presenten durante el periodo de
incubación. Se pesaron 50 crías en época cálida y 50 crías en época fría, de
aproximadamente 5 días de nacidas, se encontraron diferencias significativas
entre ambas época del año (p<0.05). Para época cálida encontramos un peso
promedio de 15 ± 5 mg y en época fría de 6 ± 1 mg, la cría de mayor peso fue
registrada en época cálida con un peso de 33 mg y la más pequeña en época fría
con un peso de 3 mg (Fig. 23).
Figura 23. Peso de las crías en época cálida y fría del año del acocil
Cambarellus (C.) montezumae.

96
6.5.9 Respuesta fisiológica de las hembras ovígeras.
El cuidado que realizan las hembras del acocil Cambarellus (C.) montezumae
de la masa ovígera que portan durante el periodo de incubación-eclosión, es
conocido como cuidado maternal, el cual se define como toda aquella conducta
realizada por las hembras que incrementa la supervivencia de la progenie
(Fernández, et al., 2003).
Se encontró que las hembras ovígeras consumen significativamente más
oxígeno que las hembras no ovígeras (p<0.05), las cuales consumen en
promedio 388.1 ± 36.9 cal/g Ps-1 día-1 y las hembras no ovígeras 233.4 ± 19.8
cal/g Ps-1 día-1 (Fig. 24).
Figura 24. Consumo de oxígeno de hembras ovígeras y no ovígeras del acocil
Cambarellus (C.) montezumae.

97
6.6 DISCUSIÓN.
La caracterización de hábitats acuáticos por medio del monitoreo de
parámetros físico-químicos como temperatura, pH y oxígeno disuelto, es tal vez
la forma más sencilla de identificar sus variaciones tanto espaciales como
temporales, resultantes de cambios en factores naturales como el clima de la
región. En general, es aceptado que dichos parámetros ejercen una influencia
notable sobre los procesos químicos y biológicos que ocurren dentro del
ecosistema (Hem, 1985; Manahan, 1972).
Las variaciones en los parametros físico-químicos en el Lago de Xochimilco
establecen una estacionalidad bien definida, de abril a septiembre son meses de
época cálida y los meses de octubre a marzo los correspondientes a época fría.
Dicha estacionalidad está determinada principalmente por las variaciones en la
temperatura (García-Padilla, 2010).
La temperatura desempeña un papel fundamental en el funcionamiento del
ecosistema al regular o afectar otros factores abióticos del sistema como son la
solubilidad de nutrientes, solubilidad de gases, el grado de toxicidad de ciertos
compuestos, el pH, el potencial redox, la densidad del agua, el estado físico y la
viscosidad del sustrato. Todas estas interacciones afectan a su vez la
distribución, composición y el grado de actividad metabólica de los organismos
que integran el ecosistema, por lo que la temperatura es un factor regulador de
procesos vitales para los organismos (factor controlador), así mismo afecta su
distribución espacial y temporal (factor directriz), (Johnson, 1987).
Estos cambios en la distribucón espacial y temporal de los individuos de una
población, estarán intimamente ligados a la estacionalidad, ya que en época

98
fría, las poblaciones decrecen, generalmente a causa de las bajas temperaturas,
de la escasez de alimento y refugio, generando cambios en los principales
parámetros demográficos (Chapman & Reiss, 1992).
En los ecosistemas lénticos el nivel de oxígeno puede variar tanto estacional
como diariamente, desde un estado sobresaturado hasta cerca de un estado
anóxico. Los acociles que habitan aguas cálidas o tropicales generalmente no
requieren altos niveles de oxígeno disuelto para sobrevivir. Sin embargo, su
crecimiento es lento en comparación con los que viven en aguas frías (Holdich,
2002). Este autor, indica que los acociles pueden soportar bajas
concentraciones de oxígeno (menos de 4.0 mg O2/L) ya que tienen una molécula
compensatoria que incrementa la eficiencia de transporte de oxígeno en la
hemolinfa, denominada hemocianina.
En este estudio, en la mayor parte de los muestreos la concentración promedio
de oxígeno en el estrato fondo y superficie se mantuvo por arriba de 6 mg O2/L
e incluso, en los meses de agosto, septiembre, enero y marzo se presentó una
sobresaturación de oxígeno disuelto con valores de 12 mg O2/L en el estrato
superficie. Las causas principales de sobresaturación de oxígeno pueden ser
debido al efecto acelerador de la temperatura sobre las altas concentraciones de
algas fotosintéticas que se encuentran en el cuerpo lacustre. Por lo anterior, se
deduce que Cambarellus (C.) montezumae puede soportar variaciones en los
niveles de oxígeno por lo que este parametro, no tiene un efecto significativo
sobre la abundancia de los organismos.
Uno de los factores importantes para el crecimiento de los acociles es el calcio,
(carbonato de calcio y bicarbonato de calcio), ya que necesitan una alta
concentración de estas sales durante la postmuda para reemplazar el calcio
cuticular que perdieron durante la fase de muda, por lo cual la mayoría de las

99
especies de acociles se ven afectados negativamente por aguas ácidas a un pH
de 5.5 (Appelberg, 1989).
En el Lago de Xochimilco encontramos valores de pH de entre 8.6 y 10, lo que
indica que el agua de los canales es muy alcalina, esto debido a que el lago
recibe agua de la planta tratadora de aguas negras del Cerro de la Estrella; en
consecuencia, ingresan nutrientes inorgánicos como sales, fósforo, nitratos y
nitritos, así como también metales pesados; dichos valores no inhiben el
crecimiento y abundancia de Cambarellus (C.) montezumae (Alva, 1996; Flores,
2009). En conclusión, a partir de los resultados obtenidos de los parámetros
físico-químicos, se demuestra que la temperatura es el único factor de los
considerados en este estudio que afecta la abundancia y crecimiento de
Cambarellus (C) montezumae en el Lago de Xochimilco.
Una de las principales preguntas en ecología es ¿qué determina la distribución
y abundancia de los organismos? La abundancia de individuos de una
población es producto de factores físicos del ambiente, de factores históricos y
de la relación entre sus individuos y con otras especies.
En este sentido se han realizado pocos estudios destinados a conocer la
interacción entre los factores limitantes del habitat (que son aquellos que
regulan el crecimiento y la expansión de las poblaciones), como la temperatura
(analizada a traves de la estacionalidad) y los principales parametros
demográficos del acocil Cambarellus (C.) montezumae en condiciones naturales.
Entre estos estudios se encuentran los elaborados por Rangel (2007 y 2009),
que se llevaron acabo en la pista de Remo y Canotaje “Virgilio Uribe”, la cual
presenta una mejor calidad del agua en comparación a los canales del Bordo y
Cuemanco en donde se han realizado estudios similares (Villa y García-Padilla,

100
2010), el común denominador en estos trabajos ha sido la relación que existe
entre la estacionalidad del habitat y la dinamica poblacional del acocil
Cambarellus (C.) montezumae.
En dichos trabajos se ha encontrado una relación significativa entre las
caracteristicas intrínsecas del medio (principalmente la temperatura) y la
abundancia de los individuos en la población, encontrando que en los meses
más calidos del año se encuentra una mayor abundancia del acocil Cambarellus
(C.) montezumae, lo cual concuerda con lo encontrado en este trabajo, en donde
los meses con mayor abundancia de individuos fueron abril, mayo y junio. En
los meses más fríos que fueron enero y febrero encontramos una reducción
considerable de la abundancia de individuos en la población.
Aunque la temperaura fue el único parámetro físico-químico que presentó una
relación directa con la abundancia del acocil Cambarellus (C.) montezumae, no
podemos afirmar que sea el único factor responsable de la abundancia de este
organismo, ya que existen otros factores que pueden estar incidiendo sobre el
tamaño de la población, estos factores pueden ser tanto abióticos como bióticos.
Los factores abióticos son todos aquellos relacionados con el ambiente físico del
Lago de Xochimilco y los factores bióticos (que de igual manera puede afectar la
distribución y abundancia de la población), son los relacionados con otros
organismos, es decir, las relaciones que puede presentar el acocil Cambarellus
(C.) montezumae con los integrantes de la misma población (ej. canibalismo) y
con las especies presentes en el habitat (ej. depredación).
Otro aspecto significativo dentro de la dinámica de una población es la
composición por edad y sexo de los individuos que la conforman, como
generalmente se denomina, la estructura poblacional. Conocer la estructura
poblacional es esencial en la investigación ecológica para la formulación de

101
propuestas de conservación y/o manejo sustentable de una población. El
principal motivo para analizar la distribución por edad y sexo de la población
del acocil Cambarellus (C.) montezumae en el Lago de Xochimilco, fue poder
realizar una inferencia de cuál era el estado actual del recurso.
La población de Cambarellus (C.) montezumae en el Lago de Xochimilco se
encuentra conformada por cuatro edades ecológicas: crías, juveniles, subadultos
y adultos, la proporción de individuos en cada una de estas edades no tuvo
variaciones significativas en relación a la época del año, observando que el 61%
de la población se encuentra en la edad de cría, el 19% en edad juvenil y el 16%
en edad de subadulto, estas edades corresponden a una etapa prereproductiva
en donde aún no existen encuentros sexuales exitosos. La etapa reproductiva
se encuentra conformada por la edad de adulto en donde sólo se encuentra el
4% de la población.
El examen de la estructura poblacional de este acocil, revela bastante
información acerca de la historia de la población y de sus perspectivas futuras.
La composición de la población va a depender de tres componentes
principalmente: mortalidad, fecundidad y migraciones, aunque el tercer
componente no va a tener influencia sobre esta población, ya que se encuentra
aislada, no existe un flujo de acociles entre una población y otra. Una población
con una elevada proporción de crías y juveniles como es este caso, se denomina
población joven, que generalmente es el resultado de una elevada fecundidad y
mortalidad alta.
Algunas poblaciones, con el paso del tiempo presentan cambios en la
fecundidad, es decir, que la fecundidad se reduce, con lo cual la composición de
la población inicia un proceso de maduración, esto es, hay un aumento en la
proporción de subadultos y adultos y una disminución de las proporciones de

102
crías y juveniles, este proceso continúa con el envejecimiento de la población
(Begon, et al., 2006).
La composición de una población aporta información valiosa sobre
características demográficas futuras, en el caso de la población de Cambarellus
(C.) montezumae en el Lago de Xochimilco, considerada una población joven
(por su elevada proporción de crías y juveniles), implica una capacidad latente
para un crecimiento rápido, siempre y cuando existan las condiciones
adecuadas para que esto suceda.
Aunque la población de Cambarellus (C.) montezumae presente características
favorables para presentar un rápido crecimiento de su población, esto no está
ocurriendo, debido muy probablemente a la influencia de la calidad del agua
del Lago de Xochimilco, ya que este lago era alimentado por 4 manantiales de
gran importancia y su calidad del agua era excelente. La sobreexplotación del
agua agotó el caudal de los manantiales, provocó hundimientos en los terrenos
y con ello la contaminación en las aguas del lago (INEGI, 2000); el nivel de
agua de los diversos canales ha bajado en forma alarmante desde noviembre de
1975 hasta la fecha (esta disminución del volumen del agua ha provocado que
en el Canal del Bordo donde en el 2011 aún se podían encontrar acociles hoy en
día ya no se encuentren); por ello, al final de la década de los ochenta el
gobierno del Distrito Federal puso en marcha el “Plan de Rescate Ecológico de
Xochimilco”, en el cual tenía como fin la recuperación en todos sentidos (flora,
fauna, etc.) de la zona lacustre.
Por otro lado, en la zona chinampera se han detectado 19 asentamientos
irregulares, que representan aproximadamente a 2,600 familias; esto implica
que todos los desechos que generan son vertidos al lago. Paralelamente en el
área turística se reciben a casi 30,000 visitantes cada fin de semana, en dicha

103
área se carece de una infraestructura sanitaria adecuada, por lo cual todo esto
provoca una grave contaminación por materia orgánica, dando como resultado
la eutrofización y contaminación del lago y con esto la disminución o
desaparición de fauna lacustre como es el caso de este acocil (Balanzario, 1982;
Arcos, et al., 1995).
Estas alteraciones en el hábitat han provocado una desviación de la proporción
sexual de este organismo, ya que en condiciones idóneas la proporción entre
hembras y machos en poblaciones naturales, debería ser cercano a 1:1 con igual
número de descendientes de uno y otro sexo (aunque las diferencias en la
mortalidad entre machos y hembras modifica esta proporción con el paso del
tiempo (Dajoz, 2002)). Sin embargo, en el caso del acocil Cambarellus (C.)
montezumae encontramos una proporción sexual de 2:1 (dos hembras por cada
macho), esta deviación en el valor esperado, puede deberse a algún tipo de
disruptor endocrino o alterador hormonal (Sustancias químicas que suplantan
a las hormonas naturales, trastornando los procesos normales de reproducción
y desarrollo), que puede estar presente en el hábitat de estos organismos,
provocando diferentes efectos adversos sobre la población, entre estos efectos
podemos considerar la desviación de la proporción sexual en la población y/o
cambios en su comportamiento, estos disruptores pueden ser plaguicidas
organoclorados, policlorobifenilos, agentes tensoactivos, ésteres de ácido ftálico
y/o monómeros plásticos, todos ellos provenientes de la actividad antropogénica
dentro del Lago de Xochimilco.
La proporción sexual influye decisivamente en la presión de la selección sexual
en el sentido de que establece la intensidad de la competencia por la cópula
dependiendo de la cantidad de parejas sexuales disponibles (Andersson, 1994).

104
En este trabajo se llevó a cabo el estudio de la ecologia reproductiva del acocil
Cambarellus (C.) montezumae, entendida desde el punto de vista de la
importancia maternal (epoca reproductiva, fecundidad, éxito reproductivo,
calidad de los huevos y juveniles) en los procesos de reclutamiento de la
población en el Lago de Xochimilco.
Como ya hemos hecho notar, las poblaciones naturales en general y en
particular la del acocil Cambarellus (C.) montezumae, viven inmersas en un
medio que experimenta cambios de luz, temperatura, pH, oxígeno disuelto,
calidad y cantidad de alimento disponible, entre otros factores, en donde la
supervivencia, reproducción y crecimiento van a depender en gran medida, de
la percepción de estos estímulos o factores ambientales por los individuos de la
población.
Uno de los principales factores ambientales que influyen en dichos procesos es
la temperatura, ya que no solo ejerce un efecto directo en la abundancia de
individuos en la población, sino que tambien ejerce un efecto significativo sobre
los patrones reproductivos. Para que la reproducción tenga éxito es preciso
que se produzca una sincronización de los reproductores entre sí y de éstos con
las variaciones de los factores ambientales. Esta sincronización permitirá que
existan individuos maduros en el momento más idóneo para garantizar una
mayor supervivencia de la progenie (Begon, et al., 2006).
Esta sincronización de los individuos con los factores ambientales resulta de
gran importancia en el ciclo reproductivo de Cambarellus (C.) montezumae, ya
que habita en un medio que experimenta variaciones estacionales y con estas
se presentan cambios en la cantidad y calidad de alimento disponible y en la
cobertura vegetal (la cual sirve de refugio para esta especie), dos de los

105
elementos más importantes para la subrevivencia de este organismo (Garcia-
Padilla, 2010).
Así, cada acocil debe disponer de un sistema que reciba la información
procedente tanto del exterior como del interior del organismo, que las integre y
determine el establecimiento de un estado endocrino idóneo, que regule a su
vez, todos los eventos fisiológicos que conducirán a lareproducción. Este
sistema (conformado por una serie de hormonas, el órgano X-Glándula del seno
y el órgano Y), va a encargarse de la traducción de los estimulos ambientales a
impulsos bioelectricos y hormonales y finalmente de la adaptación fisiológica de
la especie, que suele ser cíclica y en sincronía con el medio externo (Castelló,
1993).
Aunque se ha considerado que el acocil Cambarellus (C.) montezumae tiene una
reproducción contínua durante todo el año, en este trabajo se ha observado que
aun cuando encontramos desoves consecutivos (ecepto en los meses de
noviembre y diciembre), existe una estación bien definida para la reproducción,
y ésta comprende los meses correspondientes a la época cálida del año
(principalmente abril, mayo y junio), ya que en esta época se obtuvo el 83 % del
total de los desoves, con lo cual se concluye que esta especie en el Lago de
Xochimilco, presenta una reproducción cíclica y en sincronía con la epoca cálida
del año (donde las condiciones medio ambientales son las más favorables), lo
cual nos podría indicar que el acocil Cambarellus (C.) montezumae presenta
una estrategia reproductiva bien definida.
Una estrategia reproductiva es el conjunto de características que una especie
ha desarrollado para tener éxito en la reproducción (Vazzoler, 1996). Las
combinaciones con el hábitat, la fisiología y el comportamiento reproductivo
que lleva a cabo el acocil Cambarellus (C.) montezumae, va a determinar
directamente el éxito reproductivo de la especie en el Lago de Xochimilco.

106
Asi mismo, el éxito reproductivo va a depender de que las hembras puedan
asegurar la sobrevivencia de una porción considerable de huevos y/o crías, esto
por medio de una elección adecuada del lugar y tiempo del desove, ya que con
adecuadas condiciones ambientales el desarrollo del los huevos y crías se
llevará a cabo en un menor tiempo; y de esta manera, disminuirá el tiempo en
que son más vulnerables a los cambios ambientales y/o a la depredación
(Tresierra, 2002).
Los primeros estadios vitales son extremadamente vulnerables tanto a las
condiciones abióticas desfavorables como al ataque de los predadores. La
carencia de movilidad de estos estados tempranos significa que ellos puedan
aportar poco o nada ante esos peligros. Al igual que el momento de nacer es
importarte, el lugar elegido por la hembra ovígera para liberar a las crías
también lo será. De acuerdo a su comportamiento reproductivo el acocil
Cambarellus (C.) montezumae pertenece a la sección etológica de crustáceos
ovíparos con cuidado maternal, lo cual le da, en comparación a otras especies
de crustáceos que no presentan este comportamiento, una menor fecundidad y
una mayor probabilidad de supervivencia de la descendencia.
La fecundidad de Cambarellus (C.) montezumae se encuentra entre un rango
de 45 a 50 huevos, dicha fecundidad no se vio influenciada por la época del año
donde fue encontrada la hembra ovígera. La fecundidad más bien se relacionó
a otros factores como el tamaño y/o peso de la hembra (Civera y Cruz, 1982).
Hembras de menor tamaño portaron un número menor de huevos que las
hembras de mayor tamaño, lo cual puede estar relacionado directamente con el
área abdominal disponible de la hembra para portar los huevecillos, y su
capacidad para llevar a cabo la aireación de los mismos durante el periodo de
incubación-eclosión (Steven, et al., 1995).

107
Para este estudio encontramos a la hembra ovada más pequeña en época cálida
del año con una longitud total de 28.31 mm. Morones (1991), reporta una
longitud mínima de reproducción para las hembras de 27.01 mm y una
fecundidad de 47 huevos por hembra, lo cual es muy cercano a lo encontrado en
este trabajo, ambos estudios realizados en el lago de Xochimilco.
Civera y Cruz (1982), reportan fecundidades promedio de 110 huevos para
hembras ovigeras de 30.01 a 40.01 mm de longitud total y para hembras de
41.01 a 49.01mm de longitud total una fecundidad promedio de 161 huevos;
estos autores realizaron su estudio en la Presa “Barranca del Muerto”, en el
estado de Tlaxcala, donde la calidad del agua es mejor que en el Lago de
Xochimilco, y los parámetros físico-químicos (temperatura, pH, oxígeno
disuelto) son muy similares.
Álvarez y Rangel (2007), realizaron un estudio en la pista de Remo y Canotaje
“Virgilio Uribe” durante el año 2001, donde reportan actividad reproductiva en
los meses de enero, febrero, marzo, octubre y noviembre, estos meses se ubican
en la época fría del año, lo que contrasta con lo encontrado en este estudio
donde la mayor actividad reproductiva se dio durante los meses
correspondientes a época cálida. Encontraron tallas mínimas de reproducción
a los 24.01 mm de longitud total y una fecundidad promedio de 50 huevos,
tanto la talla mínima de reproducción como la fecundidad promedio son muy
parecidas a lo encontrado en este estudio.
Existe una homogeneidad en la talla de la primera reproducción en todos los
estudios [24.01 a 30.01 mm de longitud total para hembras (Holdich, 2002)].
Esta característica se encuentra relacionada con otros parámetros
poblacionales como la tasa reproductiva neta (Ro) y la tasa instantánea de

108
crecimiento (r) que explican el crecimiento poblacional (Begon, et al., 1996). La
talla de primera reproducción depende, además, de la condición medio
ambiental prevaleciente en ese momento de la historia de vida del organismo y
determina los procesos fisiológicos a seguir y, en última instancia, le permite
responder adaptativamente, a los cambios ambientales mediante cambios
fisiológicos y de comportamiento como repuesta a esa condición medio
ambiental.
Entre los parámetros ambientales de mayor influencia sobre la talla de
primera reproducción y la tasa de crecimiento en crustáceos se encuentra la
temperatura, junto con todos los cambios que conlleva en la dinámica del
ecosistema (Feist, et al., 2004), lo cual además afecta la temporada en que se
lleva a cabo la gametogénesis y establece la estacionalidad del ciclo reproductor
(Pankhurst, 1998).
Durante la gametogénesis se llevará a cabo la formación y de desarrollo de las
células generativas especializadas o gametos (células sexuales: ovocitos y
espermatozoides), permitiendo la preparación de las células sexuales para la
fecundación. Los ovocitos del acocil Cambarellus (C.) montezumae son ricos
en reservas nutritivas, las cuales aparecen acumuladas de muy diferentes
maneras y es la mayor o menor abundancia de vitelo y su particular
distribución en los óvulos lo que va a determinar en primer término, la
orientación de los planos de segmentación y las características de los
blastodermos, y posteriormente, el mecanismo de la gastrulación y de una
manera precisa el grado de desarrollo a que pueden llagar los nuevos
individuos antes de eclosionar (Cantú, 1959).
Las hembras de Cambarellus (C.) montezumae son ovíparas; los huevos por la
cantidad y distribución del vitelo pertenecen al tipo denominado centrolecito,

109
cada huevo presenta tres envolturas: la membrana de adhesión a las patas, el
corion y la membrana vitelina. El vitelo aparece formado por dos componentes
fundamentales: una sustancia homogénea y las gotas de materiales lipídicos.
Su segmentación es superficial y su gastrulación se produce por movimientos
de involución y convergencia (Cantú, 1959).
La cantidad y calidad de las reservas nutritivas de los huevos de este acocil van
a estar determinadas por varios parámetros que frecuentemente cambian a lo
largo de la estación reproductiva. Entre estos factores podemos mencionar el
estado endócrino de la hembra durante la ovogénesis, la cantidad y calidad del
alimento ingerido, parámetros físico-químicos del hábitat, estrés de los
reproductores, etc., (Campbell, et al., 1992; Bromage, et al., 1994; Brooks, et al.,
1997).
Debido a que se han encontrado evidencias de que la composición del huevo
cambia con respecto a la estación reproductiva de algunas especies, se decidió
analizar las características intrínsecas de los huevos, encontrando que el
tamaño de los huevos de época cálida fue mayor que el de los huevos de época
fría. Huevos de mayor tamaño dan lugar a larvas más grandes, con una mayor
eficiencia (tanto para desarrollarse como para alimentarse) y en consecuencia,
presentan una mayor sobrevivencia durante las fases tempranas de desarrollo
(Mylonas, et al., 2004).
Es de esperarse que el aumento en el volumen del huevo esté determinado por
una presencia mayor de sustancias de reserva, para este estudio se encontró un
mayor porcentaje de proteína y lípidos en los huevos de época cálida
reafirmando con esto, la importancia de la estacionalidad en el ciclo
reproductivo de Cambarellus (C.) montezumae.

110
Este acocil presenta un periodo de desarrollo embrionario corto en comparación
a otras especies de crustáceos y depende de la temperatura; este factor afecta
las tasas metabólicas y en consecuencia la velocidad de desarrollo por lo cual el
periodo de desarrollo fue menor en época cálida que en época fría. Kamler, et
al., (1998) mencionan que la temperatura modifica la eficiencia con la cual el
vitelo es utilizado por el embrión y que existe un patrón en el uso de la energía
relacionado con el incremento térmico presente en la época reproductiva.
Este efecto térmico también ha sido correlacionado con la eficiencia de
obtención de energía derivada del alimento en crías y juveniles (Hoffman y
Fisher, 2003) y que de manera general y en cualquier etapa de la vida de los
organismos, afecta la velocidad a la que ocurren sus reacciones bioquímicas,
por lo que la temperatura modifica el metabolismo de rutina y en consecuencia
del activo (Diana, 1995; Kamler, 1992). Se ha demostrado que entre los
factores ambientales la temperatura ejerce efectos sobre el crecimiento y el
desarrollo de la ontogenia de las especies (Mommsen, 1998; Ojanguren y
Braña, 2003; Robinson & Wardrop, 2002).
En síntesis, el efecto de la temperatura en la población del acocil Cambarellus
(C.) montezumae en el Lago de Xochimilco, denota no solo efectos sobre la
abundancia de organismos, sino también en la época de reproducción, en la
calidad del huevo y en el tiempo de desarrollo embrionario. Lo que pone de
manifiesto que este acocil posee una estrategia reproductiva bien definida.
Entender la estrategia reproductiva de una especie nos permitirá la
formulación de propuestas de conservación y/o de manejo sustentable.

111
6.7 CONCLUSIONES.
Las conclusiones que se obtuvieron en este capítulo sobre la ecología
reproductiva del acocil Cambarellus (C.) montezumae en el lago de Xochimilco
son:
El acocil Cambarellus (C.) montezumae es una especie que se adapta a
diferentes condiciones ambientales, en los canales de Xochimilco los
parámetros fisicoquímicos (temperatura, oxígeno y pH), se encuentran
entre los valores de tolerancia para la especie, sin embargo, esta especie
se encuentra sometida a un estrés constante debido a múltiples factores
como la contaminación crónica del cuerpo de agua, la depredación y la
actividad humana.
La composición de la población del acocil Cambarellus (C.) montezumae
en el Lago de Xochimilco es de tipo joven, ya que presenta una elevada
proporción de crías y juveniles que confiere a la población una capacidad
latente para su rápido crecimiento, siempre y cuando existan las
condiciones ambientales adecuadas.
La ecología reproductiva de este acocil nos muestra una marcada
estrategia donde existe una época reproductiva bien definida, la cual se
lleva a cabo en la época cálida del año (abril, mayo y junio) donde la
idoneidad de las hembras es mayor para la reproducción, reflejándose en
el índice de desove de esta especie y en el número de huevos producidos
por época.

112
La fecundidad a través de la época cálida y fría del año se mantiene
constante, viéndose afectada sólo por características intrínsecas de las
hembras, como el tamaño corporal y la amplitud del abdomen y no por
las condiciones ambientales.
Los huevos encontrados en época cálida del año presentan características
más favorables para su rápido desarrollo y sobrevivencia de las crías, ya
que presentan mayor tamaño, mayor volumen, mayor contenido de
lípidos y proteínas.
Las condiciones ambientales y la calidad del huevo se ven reflejadas en
el tiempo de incubación y en el tamaño de la cría al eclosionar, ya que en
época cálida el tiempo de incubación es menor y las crías de mayor
tamaño en comparación a época fría, en donde existe un retardo en el
periodo de incubación y las crías eclosionan de menor tamaño.
El incremento en la tasa metabólica de las hembras ovígeras pone de
manifiesto el cuidado maternal que ejercen sobre la masa ovígera, ya que
este comportamiento (movimiento de los pleópodos) aumenta la
adecuación biológica de la progenie, lo que implica un beneficio para las
crías pero tiene un costo para las hembras reflejado en el aumento del
consumo de oxígeno.

CAPÍTULO 2.
EFECTO DE UNA DIETA FORMULADA SOBRE EL
CRECIMIENTO, SOBREVIVENCIA Y CALIDAD DE LA PUESTA
EN CRÍAS DE ACOCIL Cambarellus (Cambarellus) montezumae
(SAUSSURE, 1857), EN CONDICIONES CONTROLADAS.

113
7.1 INTRODUCCIÓN.
La acuicultura en México se constituye como una alternativa económica muy
atractiva a mediano y largo plazo; sin embargo, en estos momentos el país
requiere de fuentes de trabajo productivas para su población, además de
alimentos de buena calidad, los cuales puedan ser alcanzados por la mayoría de
la sociedad sin que esto ocasione alteraciones en su economía familiar, por lo
que los cultivos acuícolas se deben considerar como una fuente complementaria
a la agricultura en la producción de alimentos.
En México la búsqueda y el desarrollo de tecnologías acuícolas útiles para
diversas especies tanto nativas como introducidas han sido poco estudiadas.
Para llevar a cabo una acuicultura productiva exitosa es necesario conocer las
especies nativas del país, y a partir de ellas, tratar de incorporar tecnologías
útiles para su explotación sustentable.
Uno de los aspectos más importantes que se debe de considerar, es el
conocimiento de la biología de la especie que se pretende explotar, con el fin, de
obtener perspectivas reales de producción; además, conociendo sus
requerimientos y potencial estas especies se pueden llevar a otras zonas
ambientales similares y a partir de organismos domesticados se pueden crear
nuevas zonas de cultivo económicamente activas. Por estos motivos se
considera pertinente realizar estudios a nivel experimental y de planta piloto,
para posteriormente desarrollar los proyectos más convenientes, para cada una
de las especies deseadas (Moss, et al., 1979).
Existe gran variedad de especies en las que se ha comprobado su productividad
a nivel experimental y que podrían tener buenos resultados en su producción,

114
este es el caso del acocil Cambarellus (C.) montezumae, el cual es una especie
endémica de la meseta central de nuestro país. Los cangrejos de río o acociles,
son especies que pueden considerarse como candidatas idóneas para
producciones en masa a bajo costo, además de que son conocidas por una parte
de la población mexicana (desde la época prehispánica) y, si se considera que
precisamente parte de esta población requiere de mayor cantidad de alimentos,
la producción acuícola de este acocil se vuelve una buena alternativa con un
mercado asegurado.
Debido a lo anterior, se vuelve de vital importancia la creación de un paquete
tecnológico que garantice el éxito del cultivo de esta especie. Uno de los
factores críticos en la explotación comercial de esta especie y de muchas otras
es el abastecimiento de crías para crecimiento, ya que las poblaciones de
acociles tanto naturales como cultivadas dependen de la producción de huevos
de buena calidad; pues las puestas de baja calidad constituyen uno de los
mayores impedimentos para la expansión de la acuicultura de cualquier
especie. En la industria de la acuicultura las puestas de buena calidad han
sido definidas como aquellas en las que los huevos tienen baja mortalidad en la
fecundación, formación del embrión y la eclosión (Bromage, et al., 1992).
Otros indicadores de la calidad de la puesta que se han utilizado son el tamaño
y la composición bioquímica del huevo. Aunque es conocido que huevos
grandes producen también crías grandes, no existen evidencias de que el
diámetro del huevo sea un aspecto determinante de su calidad (Kjorsvik, et al.,
1990). En cuanto a la composición bioquímica del huevo, en principio podría
esperarse que ese fuera el componente más determinante de la calidad. Sin
embargo, los resultados obtenidos en varias especies tantos marinas como
dulceacuícolas no han mostrado una clara relación entre la composición

115
bioquímica del huevo y la posterior supervivencia de huevos y larvas (Craik &
Harvey, 1984; Kjorsvik, et al., 1990).
En la mayoría de los trabajos de investigación sobre la calidad de la puesta, las
tasas de fecundación y de eclosión han sido utilizadas como criterios
importantes (Kjorsvik, et al., 2003). La supervivencia, hasta un determinado
estadio de desarrollo, y la producción final de larvas también se han utilizado
como medidas de la calidad (Fernández-Palacios, et al., 1995).
Existen numerosos factores que pueden afectar la calidad de la puesta, al igual
que el crecimiento gonadal y la fecundidad son muy susceptibles a las
influencias medioambientales, es claro que la calidad de los huevos producidos
podría también verse afectada limitando la producción de crías y su
sobrevivencia (Kjorsvik, et al., 1990). Factores tanto bióticos como abióticos
han sido señalados como posibles determinantes de la calidad del huevo o de su
supervivencia tanto en el medio natural como en cautiverio.
La calidad del huevo estaría determinada por varios parámetros físicos,
genéticos y químicos, así como por los procesos fisiológicos que suceden durante
el desarrollo inicial del huevo. Los factores que afectan la calidad del huevo
están determinados por sus propiedades intrínsecas y el entorno en el que son
fecundados y posteriormente incubados. Algunos de los factores que afectan a
la calidad del huevo son conocidos, pero muchos otros (probablemente más) son
desconocidos.
Factores que afectan la calidad del huevo incluyen el estado endocrino de la
hembra durante el crecimiento del oocito en el ovario. Existe la creencia
extendida de que los nutrientes aislados por el oocito y su procesamiento
durante el crecimiento y maduración, son los factores más importantes que

116
afectan la calidad del huevo (Craik & Harvey, 1984; Kjorsvik, et al., 1990;
Bromage, et al., 1992).
La fisiología del reproductor y su estado hormonal, que a su vez afecta la
incorporación de compuestos en los huevos, probablemente influyen en su
calidad, por ejemplo, el estrés en los reproductores tiene efectos perjudiciales
en la calidad de las puestas (Campbell, et al., 1994).
La dieta de los reproductores ha recibido gran atención con respecto a su efecto
en la calidad del huevo, y existen estudios que indican que las influencias
mayores sobre la calidad la ejercen sólo unos pocos de los componentes
dietéticos (Washburn, et al., 1990; Watanabe, et al., 1991a,b; Harel, et al.,
1994).
En acuicultura el manejo de los reproductores es probablemente el factor que
más influye en la calidad de la puesta; entre los numerosos factores que han
sido sugeridos como posibles determinantes de la calidad de la puesta, sólo
unos pocos han mostrado una clara evidencia en la calidad de la progenie
(Carrillo, et al., 2000) y pueden resumirse en los siguientes:
Factores que afectan a los reproductores
Genotipo
Alimentación
Inducción a la puesta
Estrés
Edad
Sobremaduración

117
Factores que afectan al huevo
Propiedades físicas y fisiológicas
Aberraciones cromosómicas
Colonización bacteriana
Propiedades físico-químicas del agua de la puesta e incubación
El efecto de los componentes de la dieta para reproductores ha mostrado tener
una relación directa con la calidad de la puesta, durante el periodo de
maduración o prepuesta es el factor que más influye en la calidad de los huevos
y la viabilidad de las crías, por lo cual en este capítulo, se evaluará el efecto de
una dieta formula sobre indicadores de la calidad de la puesta en crías del
acocil Cambarellus (C.) montezumae bajo condiciones controladas.
Existen componentes dietéticos como lípidos polares y no polares (Watnabe, et
al., 1991a, b; ácidos grasos (Harel, et al., 1994; Carrillo, et al., 1995; Bruce et
al., 1999; Marrozzaet al., 2003), ácido ascórbico (Mangor-Jensen, et al., 1991;
Dabroswki & Blom, 1994, 1995), vitamina A (Furuita, et al., 2003a), vitamina E
(Emata, et al., 2000), carotenos (Agius, et al., 2001), carbohidratos (Washburn,
et al., 1990; Mangor-Jensen & Brikeland, 1993), proteínas (Harel, et al., 2005;
Gunasekera, et al., 1996 a, b) y nucleótidos (González-Vecino, et al., 2004), que
han demostrado su efecto sobre la calidad de la puesta en reproductores de
peces, en el caso de los crustáceos aun no existen estudios concluyentes.
Lípidos y ácidos grasos son los componentes dietéticos que más influyen en la
calidad de las puestas, especialmente en aquellas especies de puesta continúa
como es el caso del acocil Cambarellus (C.) montezumae, que presentan cortos
periodos de vitelogenesis y que son capaces de incorporar estos componentes
dietéticos en los huevos incluso durante el periodo de puesta.

118
7.2 JUSTIFICACIÓN.
Buscando fuentes alternativas de producción de alimentos se ha encontrado en
el estudio del acocil Cambarellus (C.) montezumae una posibilidad de
producción acuícola, por lo que el estudio de los mecanismos de reproducción
son básicos y previos al funcionamiento de cualquier explotación acuícola, cuyo
objetivo principal es maximizar la producción de los organismos útiles a través
de la producción de una progenie numerosa, saludable y uniforme, mediante la
manipulación de la alimentación de los reproductores, del control de los
factores ambientales que los afectan y de la eliminación de los agentes
causantes de mortalidad.
7.3 OBJETIVOS.
7.3.1 OBJETIVO GENERAL.
Evaluar el efecto de una dieta formulada sobre el crecimiento,
sobrevivencia y calidad de la puesta de crías de acocil Cambarellus (C.)
montezumae en condiciones controladas.

119
7.3.2 OBJETIVOS PARTICULARES.
Determinar la tasa de crecimiento y sobrevivencia de crías de acocil
Cambarellus (C.) montezumae alimentadas bajo dos dietas (formulada y
control).
Establecer la talla mínima de madurez, el índice de desove y la
fecundidad de juveniles del acocil Cambarellus (C.) montezumae
alimentados bajo dos dietas (formulada y control).
Evaluar la calidad de los reproductores del acocil Cambarellus (C.)
montezumae alimentados bajo dos dietas (formulada y control) por un
periodo de 150 días.
Determinar la calidad del huevo de hembras ovígeras del acocil
Cambarellus (C.) montezumae alimentadas bajo dos dietas (formulada y
control).
Evaluar el tiempo de incubación, tamaño de las crías y su supervivencia,
de las hembras ovígeras del acocil Cambarellus (C.) montezumae
alimentadas bajo dos dietas (formulada y control).

120
7.4 MATERIAL Y MÉTODO.
7.4.1 TRABAJO DE CAMPO
7.4.1.1 Caracterización del hábitat y colecta de los organismos.
Se realizaron visitas mensuales al canal de Cuemanco en la zona lacustre de
Xochimilco en los meses de abril y mayo de 2011, con el fin colectar hembras
ovígeras de la especie Cambarellus (C.) montezumae. El canal de Cuemanco se
encuentra localizado dentro de la Área Natural Protegida (ANP) de la
delegación Xochimilco, al sur de la ciudad de México (Ver Cap. 1).
Los meses de colecta de las hembras ovígeras son considerados meses de época
cálida en donde la temperatura media es de 23.3 ± 1.3 °C, el pH se encuentra
en promedio alrededor de 8.5 ± 0.5, y la concentración de oxígeno promedio
varía según el estrato (superficie o fondo) de 9.5 a 4.1 mg/L.
La colecta de las hembras ovigeras se llevó a cabo por medio de una red de
cuchara de 1 mm de luz de malla de forma triangular. La captura de dichas
hembras se llevó a cabo entre las 10:00 y las 12:00 horas. Posteriormente
fueron colocadas en botes con agua del medio de una capacidad 2 L para su
traslado al área de acuarios del Laboratorio de Acuacultura y Producción
Acuática de la Facultad de Ciencias, UNAM.
Una vez en el área de acuario se llevó a cabo el monitoreo de las hembras
ovígeras colectadas en campo hasta la eclosión de las crías. Después de 10 días
de eclosionadas las crías fueron retiradas de la hembra y colocadas en un

121
reservorio de 120 L a temperatura ambiente, con aireación constante, tubos de
PVC de 5 cm de largo (que actuaron como refugio) y alimentación ad libitum
(Camaronina, Purina, 40 % de proteína) hasta alcanzar los 30 días de edad.
7.4.2 TRABAJO DE LABORATORIO.
7.4.2.1 Elaboración de las dietas.
Los ingredientes usados para la formulación de la dieta experimental como el
calamar, el aceite de hígado de pescado y la Camaronina comercial fueron
productos de uso comercial en la Ciudad de México. Los ingredientes
utilizados para cada una de las dietas se trituraron en un molino de martillo,
con el fin de que los ingredientes tuvieran un tamaño de partícula adecuado
para la alimentación de la especie en sus primeros estadios de vida.
El calamar se secó en una estufa a 60 °C por un periodo 48 horas con el fin de
aumentar su digestibilidad; una vez seco y convertido en harina se procedió a
agregar el aceite de pescado y la Camaronina triturada previamente, una vez
que se formó una pasta homogénea se procedió a secar la pasta en una estufa a
60 °C por un lapso de 24 horas. Esta mezcla de 47% harina de calamar, 47%
Camaronina y 6% de aceite de hígado de pescado fue lo que conformó la Dieta
Formulada = Acocilina. La Dieta Control = Camaronina, consistió en 100 % de
Camaronina (Purina, 40% de proteína), triturada a un tamaño de partícula
similar a la dieta formulada.

122
7.4.2.2 Análisis químico proximal de las dietas.
El análisis químico proximal de las dietas (formulada y control) se llevó a cabo
en el Laboratorio de Análisis Químicos para Alimentos, del Departamento de
Nutrición Animal y Bioquímica de la Facultad de Medicina Veterinaria y
Zootecnia, de la Universidad Nacional Autónoma de México, de acuerdo a los
métodos Oficiales de Análisis de la Asociación de Química Analítica (AOAC,
1990). Los resultados se muestran en el Cuatro 15 y se expresan en porcentaje
con respecto a la materia seca.
Cuadro 15. Análisis químico proximal de las dietas.
Dieta Formulada Dieta Control
Materia seca 98.1 94.7
Humedad 1.8 5.2
Proteína cruda 46.4 26.2
Extracto etéreo 31.5 12.2
Minerales totales 6.9 10.4
Fibra cruda 2.0 1.7
Extracto libre de nitrógeno 11.2 43.6
Contenido de energía (cal/g PS) 5591.0 4,265
*Los resultados se expresan en base húmeda
La evaluación de la dieta formulada (Acocilina) se llevó a cabo en tres fases: La
primera fase consistió en la evaluación del crecimiento y la supervivencia de
crías de acocil Cambarellus (C.) montezumae; la segunda consistió en la
evaluación de los indicadores de desempeño reproductivo; y la tercera en la

123
evaluación de la calidad de la puesta. Estas evaluaciones se llevaron a cabo en
las instalaciones del acuario del Laboratorio de Acuacultura y Producción
Acuática de la Facultad de Ciencias, UNAM.
7.4.2.3 FASE I. Evaluación del crecimiento y la sobrevivencia.
7.4.2.3.1 Diseño experimental.
Esta fase fue evaluada por un periodo de 60 días, el bioensayo se realizó en
peceras de 10 L de capacidad en las que se colocaron 15 peceras por
tratamiento, cada una tuvo una densidad de 10 organismos asignados
completamente al azar. Las crías tuvieron un peso promedio inicial de 25 ± 5
mg, no se encontraron diferencias significativas entre el peso inicial de las crías
para cada tratamiento (p>0.05). Los organismos fueron alimentados ad
libitum con las dietas de Acocilina y Camaronina, las peceras se mantuvieron a
una temperatura promedio de 23 ± 1 °C (Fig. 25).
Se colocó tubería de PVC de 5 cm de longitud y 3 cm de diámetro para que
actuara como refugio, las peceras tuvieron aireación contante y un calentador
de 5W para mantener la temperatura en un rango de 23 ± 1 °C, esta
temperatura fue seleccionada en base a los estudios previos de Santiago (2001)
y Aguilar (2011), los cuales señalan una temperatura óptima para el
crecimiento y la sobrevivencia de estos organismos entre 20 y 25 °C.

124
Figura 25. Diseño experimental para evaluar el crecimiento y sobrevivencia de
crías del acocil Cambarellus (C.) montezumae.
Se llevó a cabo el monitoreo de la calidad del agua de los acuarios utilizados en
cada fase experimental. Los parámetros físico-químicos monitoreados fueron:
la temperatura (± 0.05 °C) y el oxígeno disuelto (± 0.05 mg O2/L) con un
oxímetro (YSI modelo 51B) y el pH con un potenciómetro (HI 98129), con una
frecuencia de dos veces por semana en tres acuarios tomados al azar por cada
tratamiento. Se realizaron recambios de agua del 30% semanalmente.
7.4.2.3.2 Crecimiento y Sobrevivencia.
Para evaluar el crecimiento de un organismo se pueden realizar mediciones
para determinar los cambios en términos de longitud o peso. Dado que el
crecimiento ha sido registrado usualmente en términos del incremento en peso
(Jobling, 1994; Goddard, 1996), con el fin de evaluarlo se realizaron biometrías
cada 15 días de los organismos de cada tratamiento, registrando los índices

125
morfométricos de longitud total (LT: desde la punta del rostro hasta la parte
final del telson), longitud del cefalotórax (LCT: desde la punta del rostro hasta
la parte final del cefalotórax en su región dorsal media) con un vernier (± 0.01
mm) Caliper, y el peso húmedo (Ph) con una balanza (± 0.01 gr) Ohaus, Scout
Pro; la determinación del sexo se llevó a cabo de manera visual (Fig. 26).
Figura 26. Características biométricas del acocil Cambarellus (C.)
montezumae.
El análisis del crecimiento se realizó por medio de un ANOVA, para un diseño
factorial competo: Tratamiento (Dieta formula y control) X Tiempo (2 X 4), con
el paquete estadístico SPSS ver. 21 con el fin de determinar la influencia del
tipo de alimentación en los índices morfométricas considerados.
La sobrevivencia final fue calculada como el porcentaje (%) del número de
organismos vivos al término de los 60 días del experimento.

126
7.4.2.3.3 Tasa Instantánea de Crecimiento y Producción.
La Tasa Instantánea de Crecimiento (TIC) se calculó cada 15 días con los datos
obtenidos de peso húmedo (Ph) de cada una de las biometrías. De acuerdo con
Chapman (1971) que menciona que la Tasa Instantánea de Crecimiento (G) es
una medida de crecimiento exponencial; esta describe el crecimiento en un
instante particular del tiempo, y se estima con la siguiente ecuación:
G = [Ln (W2) – Ln (W1)] / t *100
Donde:
Ln (W2) = Logaritmo natural del peso en el momento t.
Ln (W1) = Logaritmo natural del peso inicial.
Es común que los datos de G se multipliquen por 100, y el resultado se exprese
como la tasa de crecimiento especifica (TEC) en % / día.
A su vez, se estimó el valor de producción usando los datos obtenidos de cada
una de las biometrías realizadas a los organismos mantenidos bajo la dieta
experimental y la dieta control. De acuerdo a la ecuación modificada por
Chapman (1971):
P = G * B
Donde:
P = Producción (g)

127
G = Tasa Instantánea de Crecimiento (g/día/%)
B = Biomasa promedio (g)
La producción debe entenderse como la formación de tejido corporal total en un
intervalo de tiempo, incluyendo el de los organismos que no sobreviven al final
del intervalo considerado. La producción puede ser medida en términos de
peso húmedo, peso seco, contenido de nitrógeno o contenido de energía. De
estos, la energía es el índice más flexible, realista y universal sobre todo cuando
la dinámica de un ecosistema entero está bajo estudio. Sin embargo, todas las
investigaciones de producción están basadas en el peso (usualmente peso
húmedo en g) en primera instancia, y el contenido de energía en segunda
instancia (Chapman, 1971).
Se calculó además la producción para los 60 días de fase experimental en
relación a la densidad de organismos, la cual para ambas dietas fue de 154
crías/m2.
7.4.2.4 FASE II. Indicadores del desempeño reproductivo.
7.4.2.4.1 Diseño Experimental.
La fase 2 fue evaluada por un periodo de 90 días adicionales a la fase
experimental 1, dicho bioensayo consistió en seguir alimentando a los
organismos de la primera fase hasta encontrar hembras ovígeras, en un diseño
por bloques al azar con quince repeticiones: los factores de contraste fueron:
tipo de alimento (Dieta Formulada = Acocilina VS Dieta Control =
Camaronina) y el sexo (Hembras vs Machos). En esta fase experimental se

128
evaluaron indicadores del desempeño reproductivo e indicadores de la calidad
del huevo. Los organismos continuaron siendo alimentados ad libitum con las
dietas de Acocilina y Camaronina, las peceras se mantuvieron a temperatura
promedio de 23 ± 1 °C.
7.4.2.4.2 Talla mínima de madurez.
La talla mínima de madurez de las hembras del acocil Cambarellus (C.)
montezumae se determinó según la evolución quincenal del número de desoves,
se reconoce la talla mínima de madurez en la cual el número de desoves
alcanza valores máximos dentro de un intervalo de tiempo, en este caso
durante los 150 días que duró la fase experimental.
7.4.2.4.3 Índice de Desove.
Así mismo, se calculó el índice de desove (ID) mediante la variación en el
porcentaje de hembras ovígeras en relación con el total de hembras adultas al
inicio del experimento.
Para determinar se existía una diferencia significativa en el número de desoves
entre ambas dieta, se llevó a cabo una chi2 de contingencia.

129
7.4.2.4.4 Fecundidad.
La fecundidad (F) de Cambarellus (C.) montezumae en este estudio se
determinó contando el número aproximado de huevos que una hembra incuba
en el momento dado de la fase experimental, durante el periodo de 60 a los 150
días.
7.4.2.4.5 Calidad de los reproductores.
Índices morfométricos.
Al concluir la fase experimental (150 días) se llevó a cabo una biometría final
de 10 organismos por cada sexo de ambas dietas experimentales, de los que se
tomaron los siguientes índices morfométricos: longitud total (LT) y longitud del
cefalotórax (LCT), con un vernier (± 0.01 mm) Caliper; y el peso húmedo (Ph)
con una balanza (± 0.01 gr) Ohaus, Scout Pro; la determinación del sexo se llevó
a cabo de manera visual.
Se sacrificó a los 40 organismos con shock hipotérmico a -20ºC por 3 horas, para
su posterior análisis químico proximal.
Composición químico proximal del tejido.
- Materia orgánica y Minerales totales.
Se determinó el contenido de materia orgánica (MO) y minerales totales (MT)
de una muestra de 3 individuos por sexo de cada una de las dietas, empleando

130
la técnica de incineración en un horno de alta temperatura (Thermolyne) a
550°C, durante un periodo de tres horas.
- Lípidos y Proteínas.
Para las determinaciones de lípidos y proteínas en los acociles se realizó un
homogenizado de una muestra de 3 individuos por sexo de cada una de las
dietas (Anexos - Procedimiento 1). Se llevó a cabo la determinación del
porcentaje de lípidos mediante el análisis de submuestras por época del año.
El contenido de lípidos se evaluó mediante el método de Fosfovainillina (2006);
para determinar la concentración de lípidos en las submuestras se realizó una
curva patrón con aceite de maíz, las muestras fueron leídas en un
espectrofotómetro Beckman DU 640 a 490 nm.
Para la determinación de proteína se utilizó el método de Nitrógeno Total de
Silicato 10023, para ello se utilizó un equipo HACH DR/780 y un reactor DRB
200, los valores obtenidos se multiplicaron por un factor de 6.25 para
determinar la cantidad de proteína total en los huevecillos, de acuerdo a lo
referido en el método Número 976.06 de la AOAC Analysis Methods (1995).
La comparación de los diversos índices en ambas épocas del año se realizó
mediante una ANOVA unifactorial, la hipótesis nula se probó a un nivel de
significancia de 0.05.

131
Índices fisiológicos.
- Tasa metabólica.
Este índice fisiológico se evaluó a través del consumo de oxígeno (QO2) de
rutina expresado como una tasa, mg O2g-1 PS día-1 en los organismos bajo
condiciones de ayuno en periodo de intermuda.
El consumo de oxígeno se midió en cámaras respirométricas de 120 ml de
capacidad con un individuo por cámara. Se emplearon 18 cámaras para estas
mediciones: 8 para la dieta formulada, 8 para la dieta comercial, y sus
respectivas cámaras control. Todas fueron sumergidas en un contenedor de 50
litros con un termostato marca Sunny de 100 W, el cual actuó como baño maría
y permitió conservar la temperatura a 23 ± 1 º C.
La medición del oxígeno se efectuó con un oxímetro YSI 51B (± 0.05 mg O2/L).
Se tomaron las lecturas iniciales de todas las cámaras previo a su cierre
hermético, y después de un periodo de 3 horas se abrieron secuencialmente
para registrar su concentración de oxígeno final.
- Excreción nitrogenada.
Las pérdidas por excreción de nitrógeno fueron evaluadas en las mismas
muestras en las que se midió el oxígeno consumido empleando alícuotas de 25
ml de c/u de ellas, la determinación de nitrógeno amoniacal se realizó mediante
el método de Azul de Indofenol (Rodier, 1981), donde la intensidad de este
compuesto leído a 640 nm con un espectrofotómetro es directamente

132
proporcional a la concentración de amonio, la cual fue expresada como mg N-
NH4g-1 PS h-1. Estos datos de absorbancia fueron transformados a valores de
concentración por medio de una curva patrón con estándares de concentración
conocido de NH4Cl, calculando la ecuación de regresión entre absorbancia (Abs)
vs concentración del estándar, con un modelo lineal (Abs = B0 + B1 [estándar]).
Tanto la tasa metabólica como la excreción de nitrógeno fueron estandarizadas
por medio de calorimetría indirecta (R y U), utilizando un coeficiente
oxicalórico (QOX) de 3.31 cal/mg O2 y un coeficiente de excreción (QEX) de 4.05
cal/mg N-NH4+ (Brafield & Solomon, 1972; Kay & Brafield, 1973). Con estos
valores se estimó la fracción de energía corporal destinada a respiración y a
excreción.
- Relación atómica O:N
Se calculó la relación atómica (O: N) para estimar las cantidades relativas de
proteínas, lípidos y carbohidratos oxidados por los organismos (Mayzaud &
Conover, 1988).
7.4.2.5 FASE 3. Indicadores de la calidad del huevo.
Para determinar la calidad de las masas ovigeras para ambas dietas se llevó a
cabo un análisis morfológico de los huevecillos (volumen del huevo y peso
húmedo), un análisis químico proximal de su composición (composición de
lípidos y proteínas) y un análisis de desarrollo embrionario (tiempo de

133
incubación, tamaño de las crías al eclosionar y % de sobrevivencia a los 15
días).
7.4.2.5.1 Volumen y peso húmedo del huevo.
El volumen del huevo se calculó separando a 5 hembras ovígeras por dieta, de
cada uno de los huevecillos se midió el diámetro mayor (a) y menor (b) con una
rejilla milimétrica y una lupa, se descartaron los huevos que presentaron algún
grado de deterioro debido al periodo de congelación. Los datos obtenidos se
incorporaron en la fórmula Vh = 1/6 (a * b2 * π) propuesta por Corey & Reid
(1991). De estos mismos huevecillos se tomaron submuestras para llevar a
cabo el análisis químico proximal.
El peso húmedo se determinó pesando cada huevecillo en una balanza digital
(± 0.001 gr) Ohaus, Scout Pro, de cada una de las hembras congeladas que se
utilizarían en el análisis químico proximal.
7.4.2.5.2 Composición químico proximal de la puesta.
Lípidos y Proteínas.
Así mismo se llevó a cabo la determinación del porcentaje de lípidos y proteínas
de tres submuestras para cada una de las dietas. El contenido de lípidos se
evaluó mediante el método de Folch (1959); para determinar la concentración
de lípidos en las submuestras se realizó una curva patrón con aceite de maíz,
las muestras fueron leídas en un espectrofotómetro Beckman DU 640 a 490
nm.

134
Para la determinación de proteína se utilizó el método de Nitrógeno Total de
Silicato 10023, para ello se utilizó un equipo HACH DR/780 y un reactor DRB
200, los valores obtenidos se multiplicaron por un factor de 6.25 para
determinar la cantidad de proteína total en los huevecillos, de acuerdo a lo
referido en el método Número 976.06 de la AOAC Analysis Methods (1995).
7.4.2.5.3 Desarrollo embrionario.
Tiempo de incubación.
Se realizó un monitoreo diario de las características morfológicas de los
huevecillos con el fin de determinar en qué estadio del desarrollo embrionario
se encontraban, durante el proceso de incubación-eclosión se describen 4
estadíos de desarrollo en función de los cambios morfológicos ocurridos durante
la embriogénesis (Cuadro 4 – Capitulo 1).
Tamaño de las crías.
Una vez que las crías eran liberadas del abdomen de la madre y tenían
completa autonomía, se procedió a retirar a la hembra del reservorio y a tomar
sus características morfométricas. Aproximadamente a los 5 días posteriores a
la eclosión de las crías se llevó a cabo el registro de su peso húmedo, mediante
una balanza digital (± 0.001 gr) Ohaus, Scout Pro.

135
Sobrevivencia de las crías a los 15 días.
La sobrevivencia final fue calculada como el porcentaje (%) del número de
organismos vivos al término de los 15 días de eclosionados.
Se analizaron estadísticamente los resultados entre la dieta formulada y la
dieta control, al igual que por sexo sobre las variables estimadas, mediante el
uso de ANOVAS utilizando el paquete estadístico SPSS ver. 21. A su vez se
realizó un análisis discriminante múltiple con las variables del análisis químico
proximal de los reproductores, y posteriormente uno de modelo reducido con las
variables de mayor importancia (lípidos y contenido de energía), con el paquete
estadístico JMP ver. 8.

136
7.5 RESULTADOS.
7.5.1 Análisis de la calidad del agua.
Durante los 150 días que comprendieron las tres fases experimentales se
registraron los parámetros físico-químicos: temperatura (± 0.05 °C), potencial
de hidrógeno (pH) con un potenciómetro HI 98129; y la concentración de
oxígeno disuelto (± 0.05 O2 mg/L) con un oxímetro YSI modelo 51B, cada tercer
día en tres acuarios tomados al azar para cada tratamiento.
Se llevó a cabo un análisis de varianza para determinar si existían variaciones
significativas de estos parámetros entre los tratamientos y a lo largo del
tiempo. No se encontraron diferencias significativas (p>0.05) para ninguno de
estos parámetros (Cuadro 16).
Cuadro 16. Parámetros físico-químicos promedio registrados durante la fase
experimental para cada tratamiento.
Tratamiento Temperatura (°C) O2 disuelto (mg/L) pH
Dieta Formulada 23.5 ± 0.1 5.9 ± 0.1 8.4 ± 0.1
Dieta Control 23.4 ± 0.2 6.0 ± 0.1 8.4 ± 0.1
*Media ± error estándar

137
7.5.2 FASE I. Crecimiento y Sobrevivencia.
7.5.2.1 Crecimeinto y Sobrevivencia.
Durante la primera fase experimental se midieron los indices morfometricos:
longitud total (LT), longitud cefalotoráxica (LCT) y peso húmedo (Ph) al inicio
del experimento y cada 15 días durante un período de 60 días. Se llevó a cabo
un análisis factorial completo para determinar si existían diferencias
significativas entre tratamientos y durante el periodo experimental.
Se encontraron diferencias significativas (p<0.05) tanto para el tratamiento
como para la biometría (tiempo) y su interacción,de cada uno de los índices
morfométricos medidos. El peso húmedo promedio (310 ± 22 mg), así como la
longitud total (22.71 ± 5.98 mm) y la longitud cefalotorácica (10.17 ± 2.82 mm)
de los organismos alimentados con la dieta control (Camaronina) después de los
60 días de experimentación fueron, mayores con respecto a los valores
obtenidos con la dieta formulada (Acocilina).
En cuanto a la sobrevivencia, se observó que el mayor poncentaje se obtuvo con
la dieta formulada (Acocilina), con un 69 % al cabo de los 60 días, en
comparación con la dieta control (Camaronina) donde se obtuvo una
sobrevivencia al final del experimento del 55%.
Se puede observar que la mayor mortalidad en la dieta formulada se dio
durante los 45 y 60 días en comparación a la dieta control donde existen dos
picos de alta mortalidad, el primero en los primeros 15 días del experimento y
el segundo al igual que para la dieta formulada en el periodo de los 45 a 60 días
(Cuadro 17 y Fig. 28).
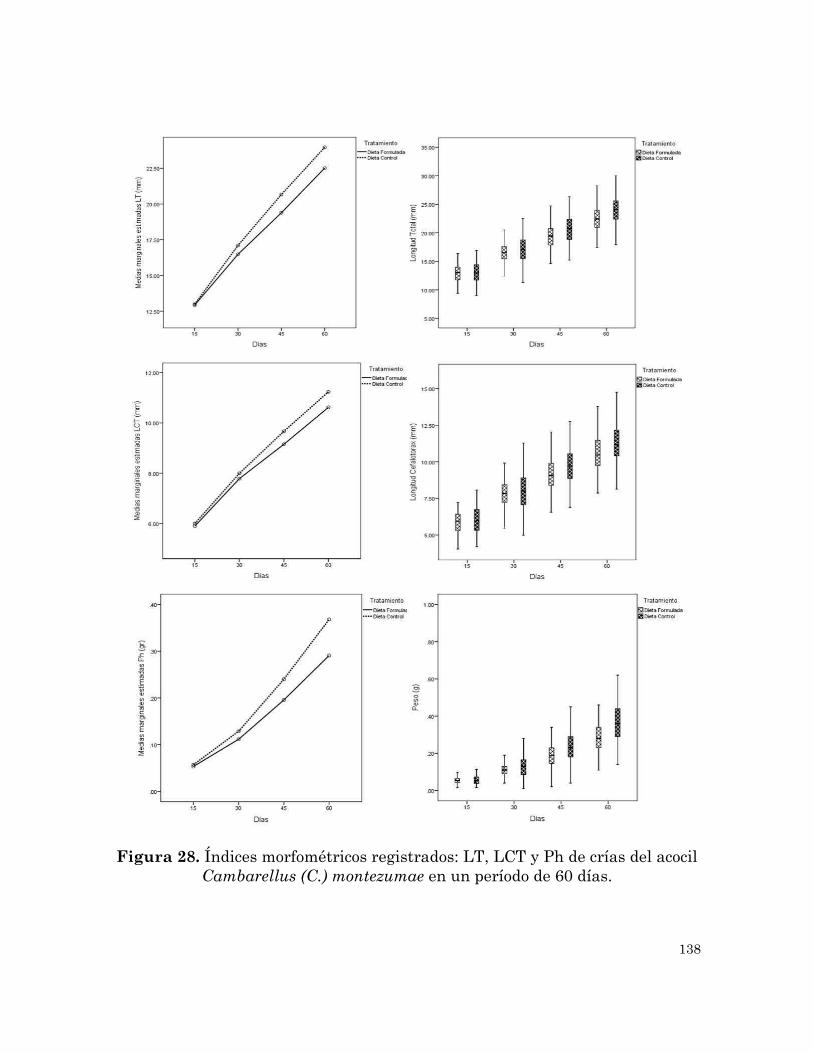
138
Figura 28. Índices morfométricos registrados: LT, LCT y Ph de crías del acocil
Cambarellus (C.) montezumae en un período de 60 días.
! ;:;
r .!! 11
t • . " ~
.,/
"
"
._ff
..,/
-' .... / ....
•
•
0"/
Días
./
-' ./
--.. ,/
••••••• S>
----_.-------_.-/ ....... .
.,/
..........
., ""
.,
1,"'a ..... mo r------------ ::::~"!"" --
¡¡¡¡¡jl l Oias
1,"'am,.mo T, •• ,ml.nto ... =~""!" ",-,, ~ .. ..---I'2 , j I {¡ '0 u ¡ 1 § , , t
" • ., Oías
1'310_0 y,,,,.móemo
-::-:=:=:' DicI. f.....-Diot.c-oI
" • ~
+ ~ , ~ I ¡ !
" • ., • Días
... ,i
./
.. /
" ~
........................
" ~
...-.... /
....
Días
• p
.... / ....
••••••• 1>
..........
•... -",.-...
. -./
.,/
"
"
T'3\. _o ... =~"'!""" '"
T'alarm.mo
... =:=
I'2 , .. 9 i 'o u
i ~ ,
Tmamienlo
=:==:-
§
• .r
" Días
Traraml9MO
~F .. ..---
t , ¡ I " ~ " Días
T'3\."""mo DioI. f ........ DIoI.eor.oI
+ ~ , ~ I ¡ !
" ~ " • Dias
./
....
0"/ ...
./
." ........ /
"'''
.. ' p .......... ....
" ~
"
.... /
.......................
~
....... /
• ....
....
. ...-",..
"
"
••••••• 1'
...... /
Y'3\a_o _ .. =~"'!""" ""
T'ala ...... mo
... =:=
I'2 , .. E -S .0 u
i ~ ,
Tmamitnto
=:=~=
§
• .r
" Oías
Tr:lramoento ~F .. ..---
t , ¡ I " ~ " Dias
y,,,u,,,,,,mo DioI. f ......-llIoI.c-oI
+ ~ , ~ I ¡ !
" ~ " • Olas

139
Cuadro 17. Crecimiento (LT, LCT y Ph) y sobrevivencia (%) de crías de acocil
Cambarellus (C.) montezumae durante un periodo de 60 días.
Tratamiento Días N LT (mm) LCT (mm) Ph (mg) Sobrevivencia (%)
0 150 - - 23 ± 5 100
15 144 12.92 ± 1.51 5.89 ± 0.18 53 ± 7 96
Dieta Formulada
30 137 16.47 ± 0.61 7.78 ± 0.71 111 ± 4 91
45 131 19.38 ± 7.26 9.15 ± 3.11 195 ± 36 87
60 104 21.95 ± 0.96 10.47 ± 0.51 275 ± 49 69
0 150 - - 21 ± 2 100
15 126 12.99 ± 0.64 6.01 ± 0.08 56 ± 11 84
Dieta Control 30 112 17.01 ± 3.41 7.99 ± 1.61 128 ± 5 75
45 108 19.16 ± 1.06 8.91 ± 1.13 180 ± 40 72
60 82 22.71 ± 5.98 10.17 ± 2.82 310 ± 22 55
*Media ± error estándar
7.5.2.1.2 Tasa Instantánea de Crecimiento (TIC) y Producción.
Los resultados de la Tasa Instantánea de Crecimiento (TIC) se presentan en el
Cuadro 19, donde se puede observar la estimación de la TIC utilizando el peso
húmedo de cada una de las biometrías, la TIC va siendo más baja con cada
biometría debido a que la velocidad de crecimiento de los organismos disminuye

140
con el aumento de talla. La TIC promedio fue mayor para los organismos
alimentados con la dieta control (Camaronina) con valor de 4.49 %/día, respecto
a la obtenida por los organismos alimentados con la dieta formulada (Acocilina)
de 4.14 %/día (Cuadro 18 y Fig. 29).
Cuadro 18. Valores de TIC durante 60 días para crías de acocil Cambarellus
(C.) montezumae.
Tratamiento N Tiempo (días) TIC %/días TIC promedio
144 T0- T15 5.57
137 T15 - T30 4.93
Dieta Formulada 4.14
131 T30 - T45 3.76
104 T45 - T60 2.29
126 T0- T15 6.54
112 T15 - T30 5.51
Dieta Control 4.49
108 T30 - T45 2.27
82 T45 - T60 3.62

141
Figura 29. TIC calculada para crías del acocil Cambarellus (C.) montezumae
alimentados con la dieta formulada y la dieta control.
La producción total después de los 60 días de fase experimental fue menor para
la dieta formulada (Acocilina), se tomó como base la dieta control (Camaronina)
para llevar a cabo la comparación; la estimación por metro cuadrado se hizo a
una densidad de 154 organismos (Cuadro 19).
Cuadro 19. Producción (formacion de tejido corporal en mg) de las crías del
acocil Camabarellus (C.) montezumae alimentados con la dieta formulada y la
dieta control durante 60 días.
Tratamiento mg/60 días/0.13m2 mg/m2 Relación prod./dieta
Dieta Formulada 171.42 1318.63 0.83
Dieta Control 206.57 1589.07 1

142
7.5.3 FASE 2. Indicadores de desempeño reproductivo.
7.5.3.1 Talla mínima de madurez.
La talla mínima de madurez de las hembras del acocil Cambarellus (C.)
montezumae se determinó según la evolución quincenal del número de desoves,
se reconoce la talla mínima de madurez en la cual el número de desoves
alcanza valores máximos dentro de un intervalo de tiempo, en este caso dentro
de los 150 días que duró la fase experimental.
Durante el periodo experimental encontramos un total de 21 hembras ovígeras
que fueron alimentadas con la dieta formulada (Acocilina) y 18 hembras
ovígeras que fueron alimentadas con la dieta control (Camaronina). El período
donde se encontró mayor actividad reproductiva para la dieta formulada fueron
entre los 75 – 90 días con un total de 8 hembras ovígeras en comparación con la
dieta control donde el período con mayor actividad reproductiva fue entre los 90
y 105 días, con un total de 9 hembras ovígeras. Para el período de los 120 a
135 días se habían dado ya el 100 % de los desoves para ambos tratamientos
(Cuadro 20 y Fig. 30).
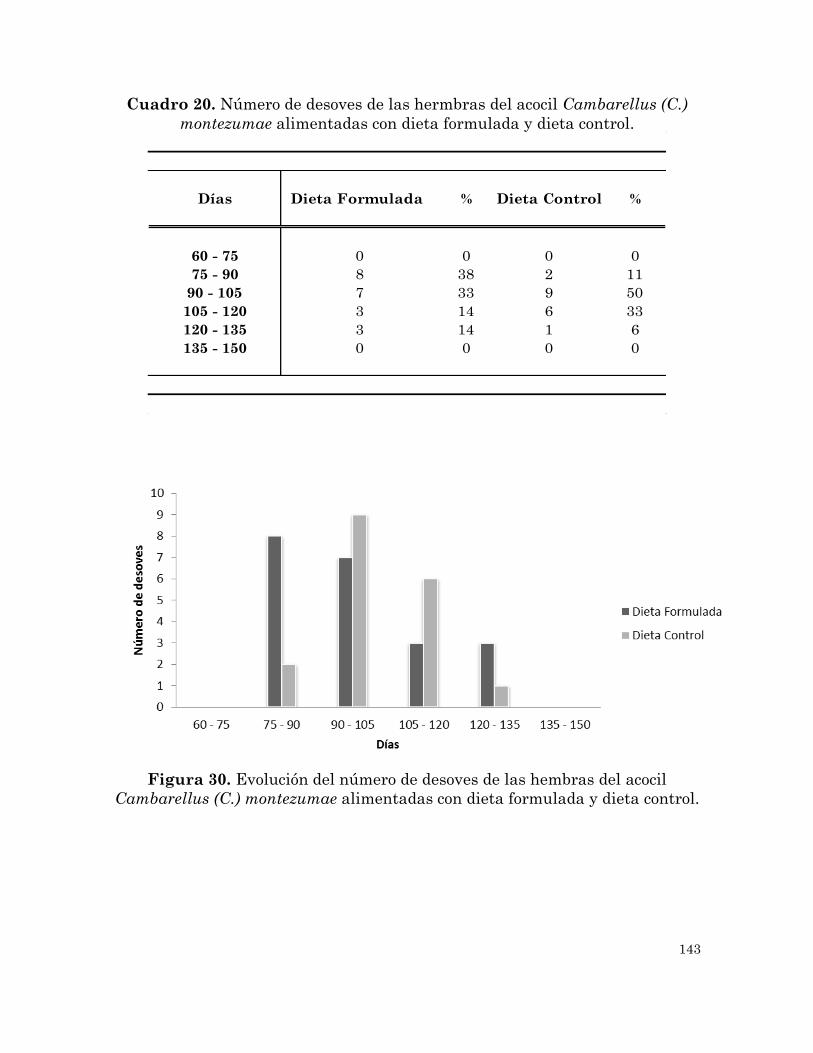
143
Cuadro 20. Número de desoves de las hermbras del acocil Cambarellus (C.)
montezumae alimentadas con dieta formulada y dieta control.
Días Dieta Formulada % Dieta Control %
60 - 75 0 0 0 0
75 - 90 8 38 2 11
90 - 105 7 33 9 50
105 - 120 3 14 6 33
120 - 135 3 14 1 6
135 - 150 0 0 0 0
Figura 30. Evolución del número de desoves de las hembras del acocil
Cambarellus (C.) montezumae alimentadas con dieta formulada y dieta control.

144
7.5.3.2 Índice de desove.
Se calculó el índice de desove (ID) mediante la variación en el porcentaje de
hembras ovígeras en relación con el total de hembras que iniciaron el
experimento para cada una de las dietas.
Los resultados muestran que el índice de desove (ID) presenta un porcentaje
mayor para la dieta formulada que para la dieta control, con 20% y 15 %
respectivamente; se encontraron diferencias significativas (p>0.05) al realizar
el Chi2 de contingencia. De igual manera, se encontró mayor número de
huevos (640) para la dieta formulada (Acocilina) que para la dieta control
(Cuadro 21).
Cuadro 21. Índice de Desove hembras del acocil Cambarellus (C.) montezumae
alimentadas con dieta formulada y dieta control.
Tratamiento Hembras No. de Desoves No. de Huevos Índice de Desove
Totales Totales (%)
Dieta Formulada 106 21 640 20
Dieta Control 118 18 563 15

145
7.5.3.3 Fecundidad.
Debido a la importancia de la fecundidad como indicador de desempeño
reproductivo para el acocil Cambarellus (C.) montezumae, se llevó a cabo una
caracterización morfométrica de las hembras ovígeras encontradas durante
toda la fase experimental para ambas dietas (Cuadro 22).
Las hembras ovígeras alimentadas con la dieta formula presentaron una
longitud total de mínima madurez o primera reproducción de 27.43 ± 2.05 mm,
en comparación con las hembras alimentadas con la dieta control que
alcanzaron una longitud total de 28.11 ± 1.61 mm, sin que esta diferencia en
cuanto al tamaño (longitud total) resultara significativa (p>0.05). El peso
promedio de las hembras ovígeras alimentadas con la dieta formulada fue de
0.54 ± 0.13 g y para la dieta control de 0.65 ±0.11 g, se encontraron diferencias
significativas (p<0.05) observándose mayor peso húmedo en las hembras
alimentadas con la dieta control.
En cuanto a la fecundidad (número de huevos por hembra), para la dieta
formulada encontramos que en promedio una hembra porta 30 ± 11 huevos, en
comparación a una hembra alimentada con la dieta control que porta en
promedio de 31 ± 9 huevos, esta diferencia no resultó ser estadísticamente
significativa (p<0.05), (Fig. 31).
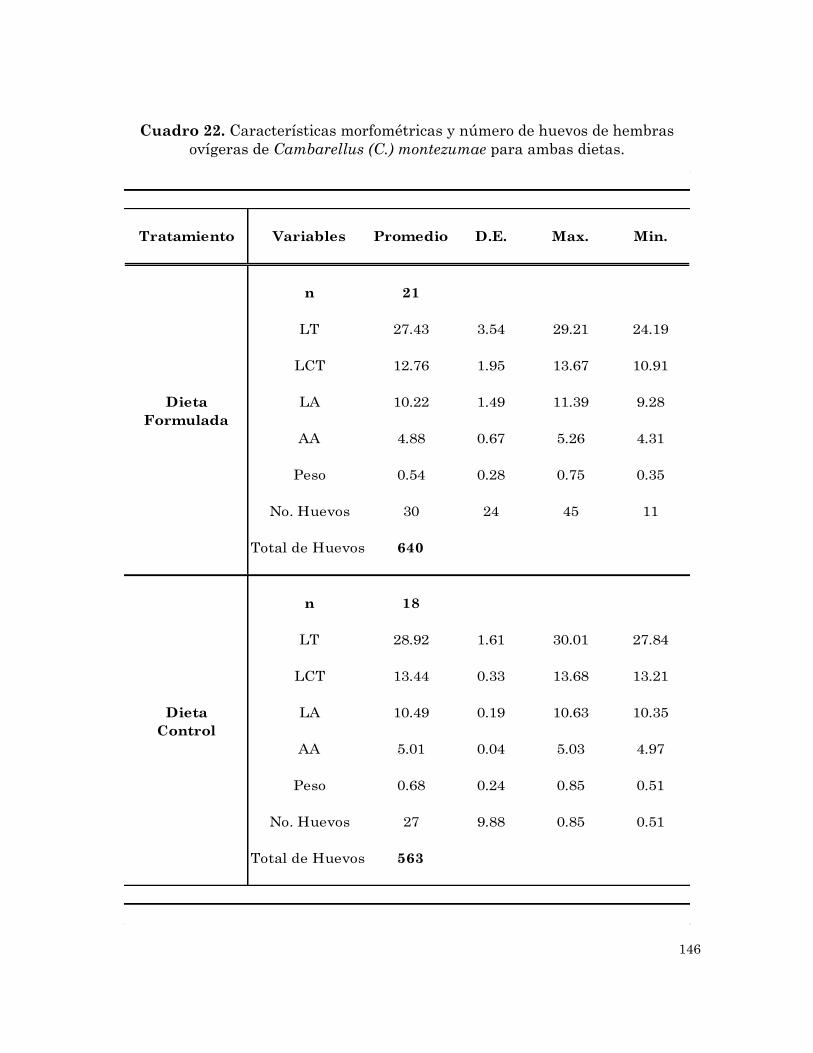
146
Cuadro 22. Características morfométricas y número de huevos de hembras
ovígeras de Cambarellus (C.) montezumae para ambas dietas.
Tratamiento Variables Promedio D.E. Max. Min.
n 21
LT 27.43 3.54 29.21 24.19
LCT 12.76 1.95 13.67 10.91
Dieta LA 10.22 1.49 11.39 9.28
Formulada
AA 4.88 0.67 5.26 4.31
Peso 0.54 0.28 0.75 0.35
No. Huevos 30 24 45 11
Total de Huevos 640
n 18
LT 28.92 1.61 30.01 27.84
LCT 13.44 0.33 13.68 13.21
Dieta LA 10.49 0.19 10.63 10.35
Control
AA 5.01 0.04 5.03 4.97
Peso 0.68 0.24 0.85 0.51
No. Huevos 27 9.88 0.85 0.51
Total de Huevos 563

147
Figura 31. Número de huevos portados por hembra del acocil Cambarellus (C.)
montezumae alimentadas con la dieta formulada y la dieta control.
7.5.3.4 Calidad de los reproductores.
Índices morfométricos.
Al final del tiempo experimental (150 días) se tomó una muestra de 10
organismos por sexo de cada uno de los tratamientos, y a los que se les
midieron los índices morfométricos: longitud total (LT), longitud cefalotoráxica
(LCT) y peso húmedo (Ph). Para cada tratamiento y sexo se probó
estadisticamente si existian diferencias significativas entre dichos índices. No
se encontraron diferencias significativas (p>0.05), para ninguno de los índices
morfométricos evaluados ni entre tratamientos ni entre los sexos (Cuadro 23).
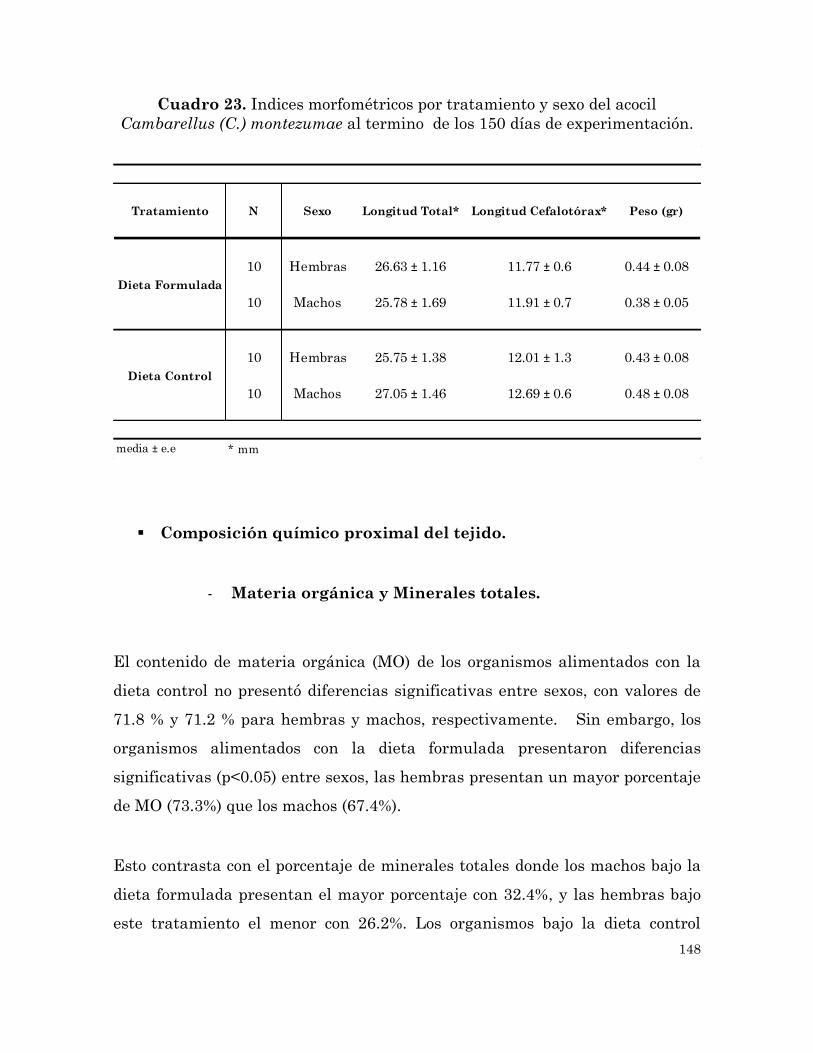
148
Cuadro 23. Indices morfométricos por tratamiento y sexo del acocil
Cambarellus (C.) montezumae al termino de los 150 días de experimentación.
Tratamiento N Sexo Longitud Total* Longitud Cefalotórax* Peso (gr)
10 Hembras 26.63 ± 1.16 11.77 ± 0.6 0.44 ± 0.08
Dieta Formulada
10 Machos 25.78 ± 1.69 11.91 ± 0.7 0.38 ± 0.05
10 Hembras 25.75 ± 1.38 12.01 ± 1.3 0.43 ± 0.08
Dieta Control
10 Machos 27.05 ± 1.46 12.69 ± 0.6 0.48 ± 0.08
media ± e.e * mm
Composición químico proximal del tejido.
- Materia orgánica y Minerales totales.
El contenido de materia orgánica (MO) de los organismos alimentados con la
dieta control no presentó diferencias significativas entre sexos, con valores de
71.8 % y 71.2 % para hembras y machos, respectivamente. Sin embargo, los
organismos alimentados con la dieta formulada presentaron diferencias
significativas (p<0.05) entre sexos, las hembras presentan un mayor porcentaje
de MO (73.3%) que los machos (67.4%).
Esto contrasta con el porcentaje de minerales totales donde los machos bajo la
dieta formulada presentan el mayor porcentaje con 32.4%, y las hembras bajo
este tratamiento el menor con 26.2%. Los organismos bajo la dieta control

149
obtuvieron registros de 28% y 31.7% para hembras y machos respectivamente
(Cuadro24).
Lípidos, Proteínas y Contenido calórico.
Los resultados para el contido de lípidos muestran que los organismos
alimentados con la dieta control tienen porcentajes de 10.2% y 19.7% para
hembras y machos, respectivamente, siendo este último el mayor porcentaje del
análisis. En cuanto a los organismos bajo la dieta formulada las hembras
presentaron 18.6% de contenido lipídico, mientras que los machos solo 15%. Al
realizar el analisis estadistico (ANOVA) se encontraron diferencias
significativas para este factor en la interacción de sexo y tratamiento (p<0.05),
(Cuadro 24).
En cuanto al contenido de proteína los organismos bajo la dieta formulada
presentan un mayor porcentaje con 43.2% y 48.1% para machos y hembras,
respectivamente, siendo las hembras de este tratamiento las que tienen el
mayor porcentaje en esta investigación. Los organismos bajo la dieta control
presentan porcentajes de 44.5% y 44.6% para hembras y machos,
respectivamente. Al realizar el analisis estadístico (ANOVA) no se
encontraron diferencias significativas (p>0.05) para el contenido de proteínas
en el tejido de los organismos (Cuadro 24).
Los resultados del análisis del contenido calorico del tejido muestran que los
organismos alimentados con la dieta formulada presentan un contenido calórico
mayor en comparación con los organismos alimentados con la dieta control, las
hembras presentan un contenido calórico de 6,294 cal-1 g PS, mientras que los
machos un contenido calórico de 5,784 cal g-1 PS. Sin embargo, los machos
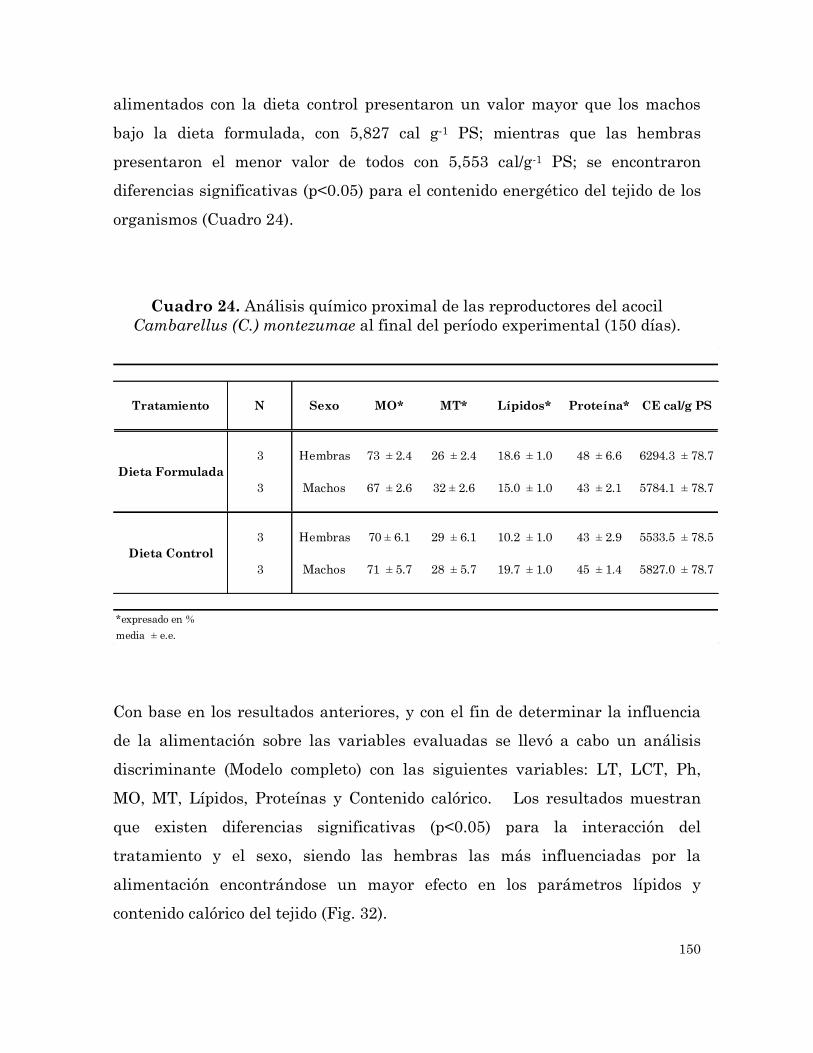
150
alimentados con la dieta control presentaron un valor mayor que los machos
bajo la dieta formulada, con 5,827 cal g-1 PS; mientras que las hembras
presentaron el menor valor de todos con 5,553 cal/g-1 PS; se encontraron
diferencias significativas (p<0.05) para el contenido energético del tejido de los
organismos (Cuadro 24).
Cuadro 24. Análisis químico proximal de las reproductores del acocil
Cambarellus (C.) montezumae al final del período experimental (150 días).
Tratamiento N Sexo MO* MT* Lípidos* Proteína* CE cal/g PS
3 Hembras 73 ± 2.4 26 ± 2.4 18.6 ± 1.0 48 ± 6.6 6294.3 ± 78.7
Dieta Formulada
3 Machos 67 ± 2.6 32 ± 2.6 15.0 ± 1.0 43 ± 2.1 5784.1 ± 78.7
3 Hembras 70 ± 6.1 29 ± 6.1 10.2 ± 1.0 43 ± 2.9 5533.5 ± 78.5
Dieta Control
3 Machos 71 ± 5.7 28 ± 5.7 19.7 ± 1.0 45 ± 1.4 5827.0 ± 78.7
*expresado en %
media ± e.e.
Con base en los resultados anteriores, y con el fin de determinar la influencia
de la alimentación sobre las variables evaluadas se llevó a cabo un análisis
discriminante (Modelo completo) con las siguientes variables: LT, LCT, Ph,
MO, MT, Lípidos, Proteínas y Contenido calórico. Los resultados muestran
que existen diferencias significativas (p<0.05) para la interacción del
tratamiento y el sexo, siendo las hembras las más influenciadas por la
alimentación encontrándose un mayor efecto en los parámetros lípidos y
contenido calórico del tejido (Fig. 32).
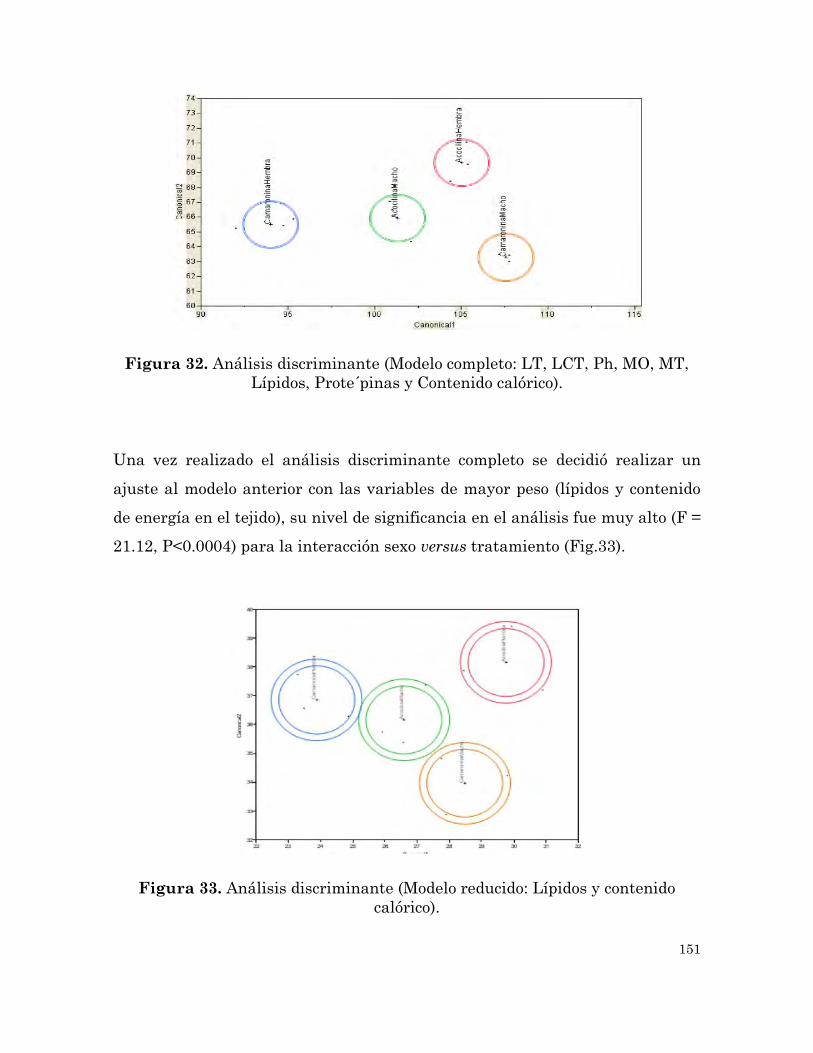
151
Figura 32. Análisis discriminante (Modelo completo: LT, LCT, Ph, MO, MT,
Lípidos, Prote´pinas y Contenido calórico).
Una vez realizado el análisis discriminante completo se decidió realizar un
ajuste al modelo anterior con las variables de mayor peso (lípidos y contenido
de energía en el tejido), su nivel de significancia en el análisis fue muy alto (F =
21.12, P<0.0004) para la interacción sexo versus tratamiento (Fig.33).
Figura 33. Análisis discriminante (Modelo reducido: Lípidos y contenido
calórico).

152
Índices Fisiológicos.
- Tasa Metabólica.
La tasa metabólica (QO2 mg O2 h-1 g-1 PS) fue determinada en un lapso de 3
horas en laboratorio al término de la fase experimental (150 días). Los valores
promedio de esta tasa fueron convertidos a cal día-1 PS utilizando un Qox =3.2
cal/mg O2 (Bradfield & Solomon, 1972).
No se encontraron diferencias significativas en el gasto de energía por
metabolismo respiratorio (p>0.05) entre la dieta formulada y la dieta control ni
entre sexos. El valor promedio para la dieta formulada fue de 630.8 cal/g
PS/día en conparación con el valor promedio de la dietacontrol que fue de 730.5
cal/g PS/día, se obtuvieron los valores del gasto de energía por metabolismo
respiratorio para cada sexo.
El porcentaje de energía destinado a respiración se calculó en base al contenido
calórico de un gramo de peso seco de tejido de un organismo por tratamiento y
sexo, encontrando valores para la dieta formulada de 10.4%y 10.5 % para
hembras y machos, respectivamente; para los organismos alimentados con la
dieta control encontramos valores de 11.6 % y 15.0 % para hembras y machos,
respectivamente (Cuadro 25 y Fig. 34).
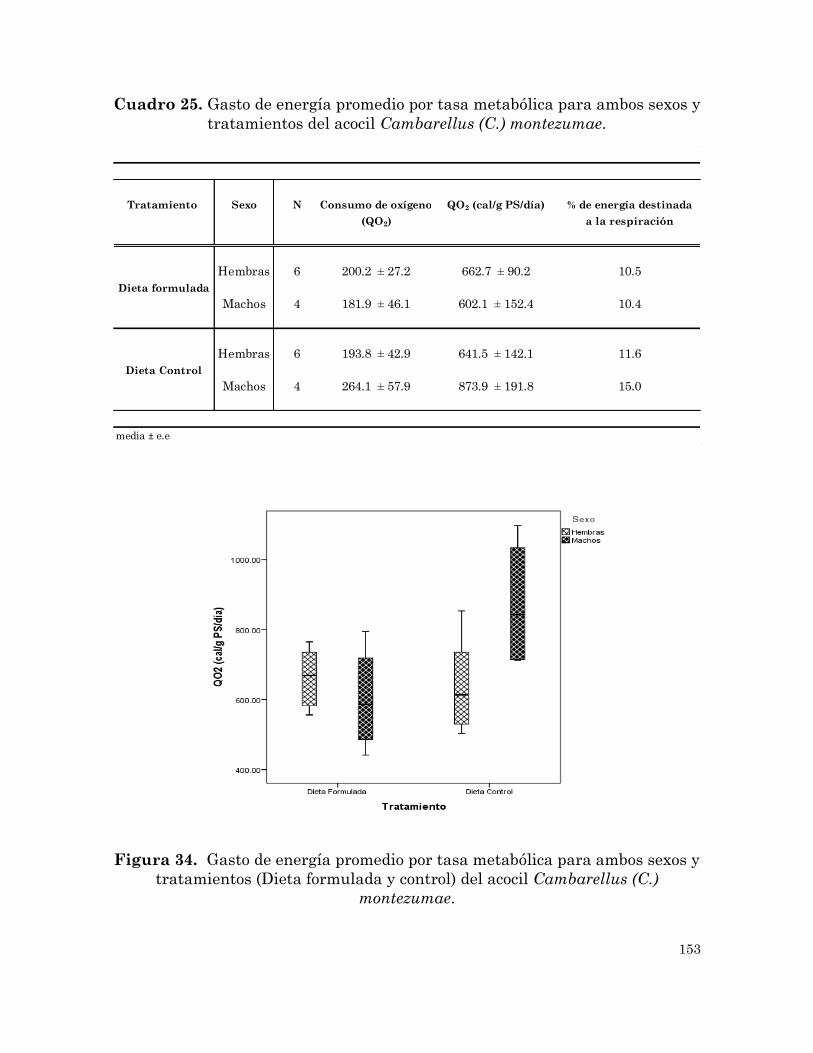
153
Cuadro 25. Gasto de energía promedio por tasa metabólica para ambos sexos y
tratamientos del acocil Cambarellus (C.) montezumae.
Tratamiento Sexo N Consumo de oxígeno QO2 (cal/g PS/día) % de energía destinada
(QO2) a la respiración
Hembras 6 200.2 ± 27.2 662.7 ± 90.2 10.5
Dieta formulada
Machos 4 181.9 ± 46.1 602.1 ± 152.4 10.4
Hembras 6 193.8 ± 42.9 641.5 ± 142.1 11.6
Dieta Control
Machos 4 264.1 ± 57.9 873.9 ± 191.8 15.0
media ± e.e
Figura 34. Gasto de energía promedio por tasa metabólica para ambos sexos y
tratamientos (Dieta formulada y control) del acocil Cambarellus (C.)
montezumae.

154
Excreción nitrogenada.
la tasa de excreción nitrogenada promedio se calculó para los organismos
sometidos a la dieta formulada y control en un lapso de 3 horas en laboratorio
al término de la fase experimental (150 días). Posteriormente se realizaron los
cálculos para obtener datos correspondientes a un ciclo de 24 horas. Los
valores promedio de la excreción nitrogenada fueron convertidos a cal día-1 PS
utilizando un QEX = 4-05 cal/mg N-NH4+(Bradfield & Solomon, 1972).
Los organismos que fueron sometidos a la dieta formulada presentaron una
menor tasa de excreción nitrogenada, 5.18 ± 0.15 mg N-NH4+/gPS/día, con
repecto a los organismos alimentados con la dieta control, lo cuales tuvieron
una mayor excreción nitrogenadas con valor promedio de 7.00 ± 0.21 mg N-
NH4+/gPS/día. Además, se calculo la pérdida de energía por excreción
nitrogenada para cada sexo por tratamiento, con el fin de determinar si existia
una influencia de la alimentación sobre el desempeño fisiológico para cada
sexo, al realizar la prueba de análisis de varianza no se encontraron diferencias
significativas(p>0.05), (Cuadro 26 y Fig. 35).
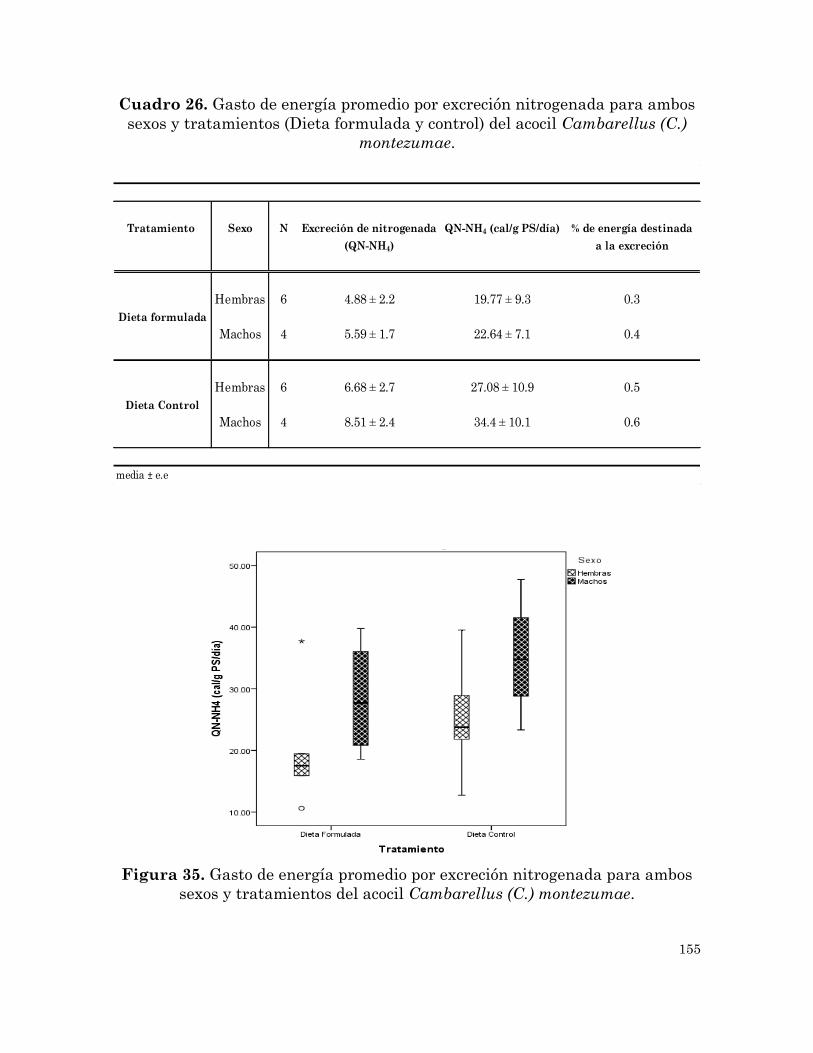
155
Cuadro 26. Gasto de energía promedio por excreción nitrogenada para ambos
sexos y tratamientos (Dieta formulada y control) del acocil Cambarellus (C.)
montezumae.
Tratamiento Sexo N Excreción de nitrogenada QN-NH4 (cal/g PS/día) % de energía destinada
(QN-NH4) a la excreción
Hembras 6 4.88 ± 2.2 19.77 ± 9.3 0.3
Dieta formulada
Machos 4 5.59 ± 1.7 22.64 ± 7.1 0.4
Hembras 6 6.68 ± 2.7 27.08 ± 10.9 0.5
Dieta Control
Machos 4 8.51 ± 2.4 34.4 ± 10.1 0.6
media ± e.e
Figura 35. Gasto de energía promedio por excreción nitrogenada para ambos
sexos y tratamientos del acocil Cambarellus (C.) montezumae.

156
Relación atómica O:N
La relación atómica O:N es un índice que permite determinar el tipo de
sustrato metabólico que es utilizado por los organismos acuáticos como fuente
de energía, ante variaciones de algunos parámetros ambientales como la
temperatura y la salinidad (Mayzaud & Conover, 1988, Díaz, et al, 1999;
Medina-Romo, 2005).
Para esta investigación se tomaron en cuenta que los valores de la relación O:N
entre 3 – 50 indican oxidación protéica, entre 50 – 60 una proporción igual de
proteínas y lípidos mientras que valores mayores a 60 un catabolismo de
carbohidratos, según lo reportado por Mayzaud & Conover (1988).
Los resultados para los organismos alimentados con la dieta formulada
presentan un índice de relación O:N de 29.19, mientras que los sometidos a la
dieta control de 25.02, en ambos casos denotan una oxidación de proteína, pero
en la dieta formulada el valor es mayor.
7.5.4 FASE 3. Indicadores de la calidad de la puesta.
Para determinar esta calidad se recurrió a la inspección de ciertos elementos
del huevo tanto en su exterior (volumen y peso) como en su interior (lípidos y
proteínas).

157
7.5.4.1 Volumen y peso del huevo.
El volumen del huevo mostró diferencias notorias entre ambas dietas,
encontrando volúmenes mayores en los huevos de hembras ovígeras
alimentadas con la dieta formulada (Acocilina). Al llevar a cabo la
comparación estadística encontramos diferencias significativas entre ambas
dietas (p<0.05), (Fig.36). El peso húmedo de cada huevo presentó una alta
relación con respecto al volumen del huevo encontrando diferencias
significativas entre ambas dietas (p<0.05), (Cuadro 27, Fig. 37).
Cuadro 27. Volumen y peso húmedo del huevo de hembras alimentadas con la
dieta formulada y la dieta control del acocil Cambarellus (C.) montezumae.
Tratamiento Volumen del huevo en (mm3) Peso húmedo en (mg)
Dieta Formulada 0.48 ± 0.24 0.010 ± 0.002
Dieta Control 0.32 ± 0.13 0.008 ± 0.001
media ± e.e
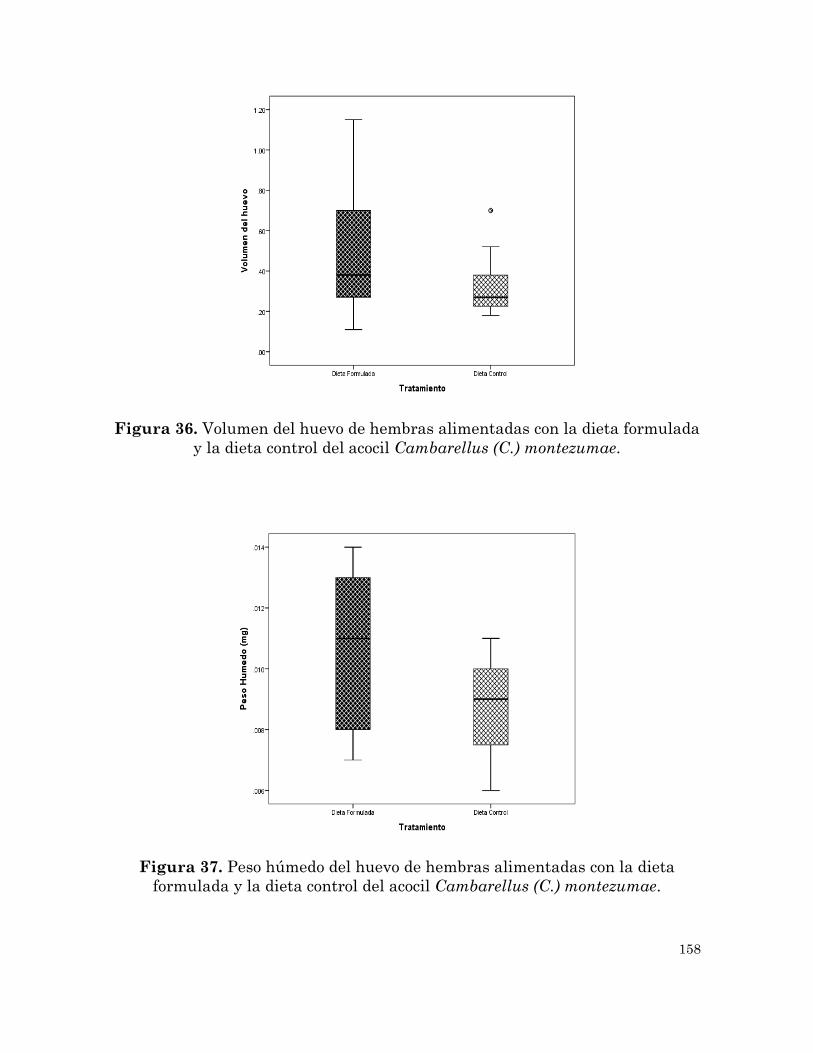
158
Figura 36. Volumen del huevo de hembras alimentadas con la dieta formulada
y la dieta control del acocil Cambarellus (C.) montezumae.
Figura 37. Peso húmedo del huevo de hembras alimentadas con la dieta
formulada y la dieta control del acocil Cambarellus (C.) montezumae.

159
7.5.4.2 Composición químico proximal de la puesta.
Lípidos y Proteínas.
La composición químico proximal de los huevos es un indicador de la calidad o
estado de salud de las hembras, y a su vez un indicador importante de la
probabilidad de eclosión y supervivencia de las crías en las primeras etapas.
Dos de los componentes químicos proximales más importantes son las
proteínas y los lípidos, al llevar a cabo el análisis de estas dos variables
encontramos que tanto para el porcentaje de proteínas como para el de lípidos
se encontraron diferencias significativas entre ambas dietas (formulada y
control), (p<0.05), encontrando los porcentajes más altos de estas variables en
los huevos de hembras ovígeras alimentadas con la dieta formulada (Acocilina),
(Cuadro 28 y Fig. 38).
Cuadro 28. Composición químico proximal de los huevos de hembras
alimentadas con la dieta formulada y la dieta control del acocil Cambarellus
(C.) montezumae.
Tratamiento Proteína % Lípidos %
Dieta Formulada 17.0 ± 1.4 27.9 ± 3.6
Dieta Control 11.3 ± 1.8 15.3 ± 3.0
media ± e.e
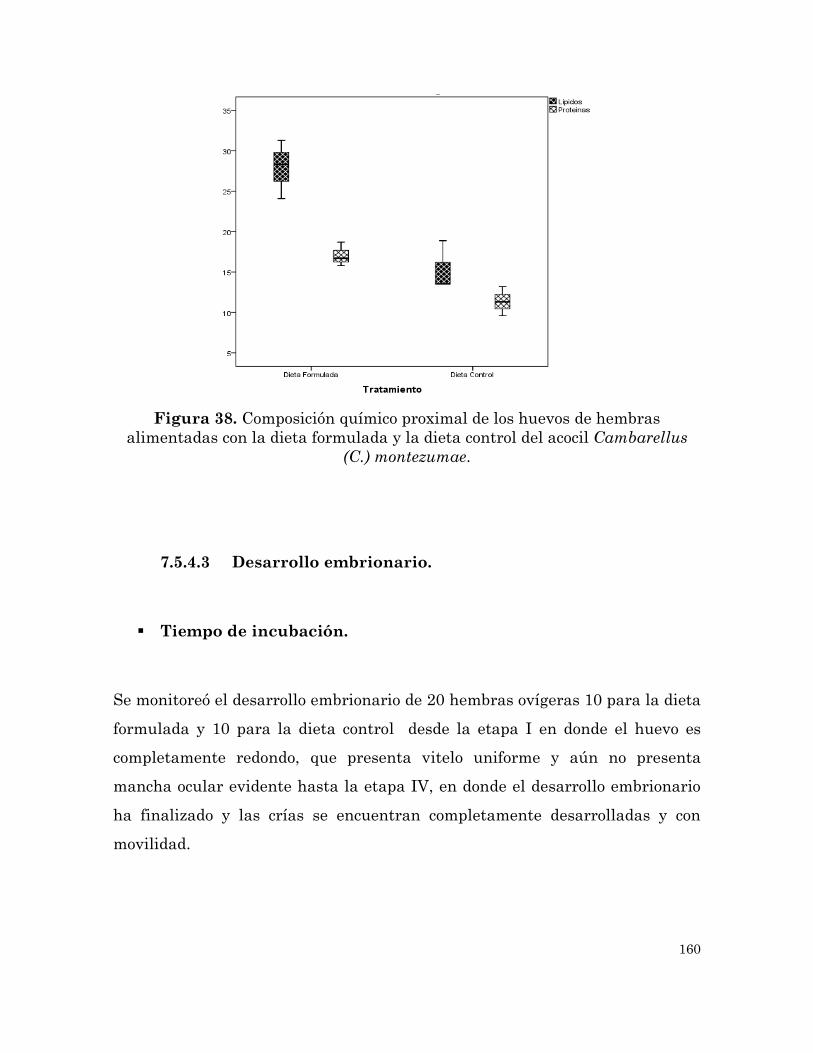
160
Figura 38. Composición químico proximal de los huevos de hembras
alimentadas con la dieta formulada y la dieta control del acocil Cambarellus
(C.) montezumae.
7.5.4.3 Desarrollo embrionario.
Tiempo de incubación.
Se monitoreó el desarrollo embrionario de 20 hembras ovígeras 10 para la dieta
formulada y 10 para la dieta control desde la etapa I en donde el huevo es
completamente redondo, que presenta vitelo uniforme y aún no presenta
mancha ocular evidente hasta la etapa IV, en donde el desarrollo embrionario
ha finalizado y las crías se encuentran completamente desarrolladas y con
movilidad.
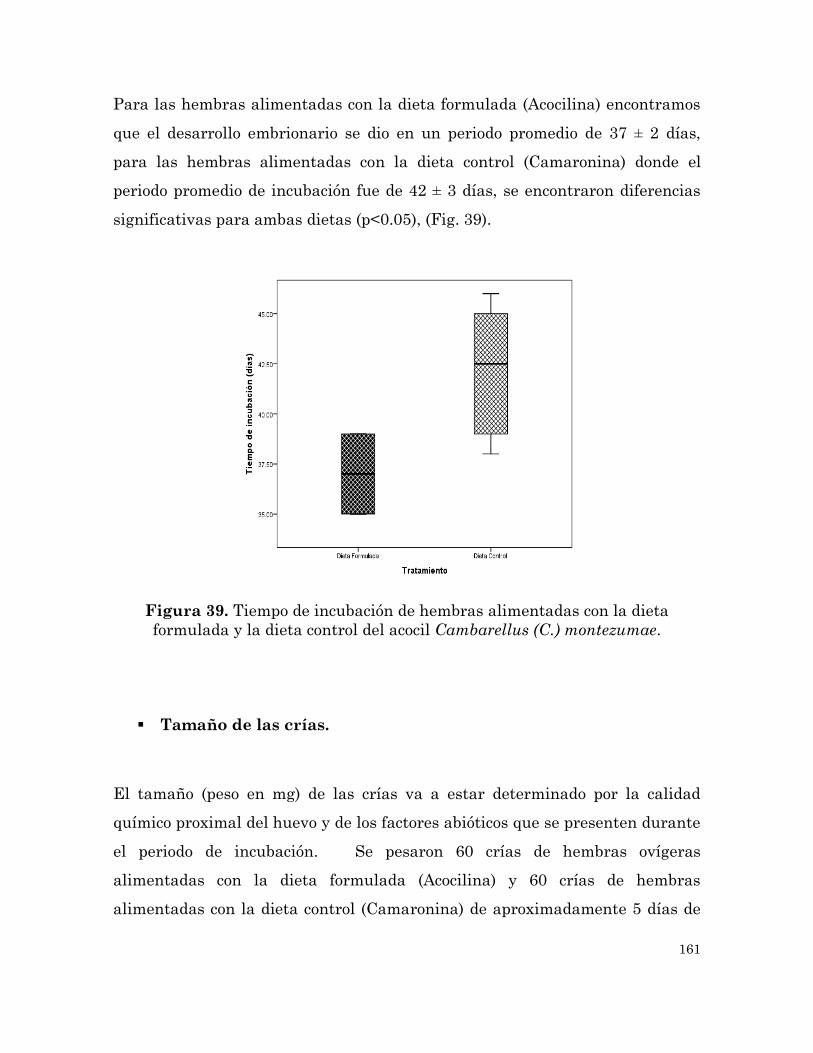
161
Para las hembras alimentadas con la dieta formulada (Acocilina) encontramos
que el desarrollo embrionario se dio en un periodo promedio de 37 ± 2 días,
para las hembras alimentadas con la dieta control (Camaronina) donde el
periodo promedio de incubación fue de 42 ± 3 días, se encontraron diferencias
significativas para ambas dietas (p<0.05), (Fig. 39).
Figura 39. Tiempo de incubación de hembras alimentadas con la dieta
formulada y la dieta control del acocil Cambarellus (C.) montezumae.
Tamaño de las crías.
El tamaño (peso en mg) de las crías va a estar determinado por la calidad
químico proximal del huevo y de los factores abióticos que se presenten durante
el periodo de incubación. Se pesaron 60 crías de hembras ovígeras
alimentadas con la dieta formulada (Acocilina) y 60 crías de hembras
alimentadas con la dieta control (Camaronina) de aproximadamente 5 días de
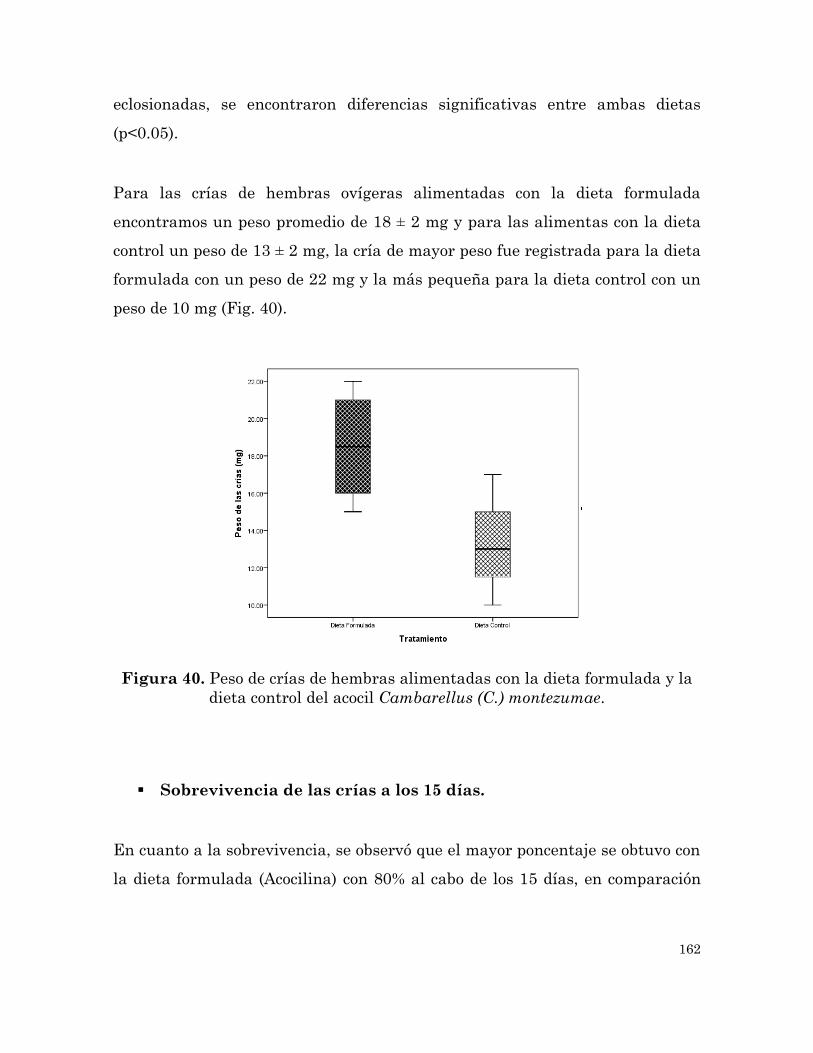
162
eclosionadas, se encontraron diferencias significativas entre ambas dietas
(p<0.05).
Para las crías de hembras ovígeras alimentadas con la dieta formulada
encontramos un peso promedio de 18 ± 2 mg y para las alimentas con la dieta
control un peso de 13 ± 2 mg, la cría de mayor peso fue registrada para la dieta
formulada con un peso de 22 mg y la más pequeña para la dieta control con un
peso de 10 mg (Fig. 40).
Figura 40. Peso de crías de hembras alimentadas con la dieta formulada y la
dieta control del acocil Cambarellus (C.) montezumae.
Sobrevivencia de las crías a los 15 días.
En cuanto a la sobrevivencia, se observó que el mayor poncentaje se obtuvo con
la dieta formulada (Acocilina) con 80% al cabo de los 15 días, en comparación

163
con la dieta control (Camaronina) donde se obtuvo una sobrevivencia del 58%
(Cuadro 29).
Cuadro 29. Sobrevivencia a los 15 días de crías de hembras alimentadas con la
dieta formulada y la dieta control del acocil Cambarellus (C.) montezumae.
Tratamiento 0 días 15 días % de sobrevivencia
Dieta Formulada 60 48 80
Dieta Control 60 35 58

164
7.6 DISCUSIÓN.
La calidad del agua tiene influencia directa en el desarrollo de los organismos,
y si estos parámetros (temperatura, oxígeno disueltoy pH) se mantienen a
niveles óptimos, el crecimiento de los organismos se lleva a cabo de mejor
manera. Los parámetros de la calidad del agua que fueron medidos durante la
fase experimental fueron: la temperatura (°C), el potencial de hidrogeno (pH) y
el oxígeno disuelto (O2), para ninguno de estos parametros se presentaron
diferencias significativas ni entre los tratamientos ni a lo largo del
experimento, lo que indicanría que el crecimiento del acocil Cambarellus (C.)
montezumae no se vió afectado por la variación de estos parametros.
La temperatura y el oxígeno disuelto tienen una relación inversa ya que al
aumentar la temperatura del agua disminuye la capacidad de está para captar
oxígeno. En la presente investigación esta relación no se vió afectada ya que
encontramos un promedio en la temperatura de 23.5 °C y una concentración de
oxígeno de 6.0 mg/L.
La temperatura es uno de los principales parámetros que desempeña un papel
primordial en el comportamiento, tolerancia y resistencia de las especies, ya
que puede llegar a ser un factor determinante en la respuesta fisiológica como
factor directriz (espacial y temporal) y controlador (funciones del organismo),
modificando las funciones nerviosas, el metabolismo respiratorio, la captación
de nutrientes e incidiendo en el desarrollo y la sobrevivencia. La capacidad del
acocil Cambarellus (C.) montezumae de tolerar amplios intervalos de
temperatura y evitar aquellos que pueden ser letales, le ha permitido una alta
plasticidad ecológica que se refleja en su gran capacidad de adaptación a
ambientes muy diversos.

165
En su habitát natural Cambarellus (C.) montezumae vive en temperaturas que
flúctuan a corto (24 horas) y largo plazo (estacional), (Cornejo, 1991), en el
intervalo de 10 a 30 °C y en condiciones extremas de -2 a 35 °C (Flores, 2009;
García-Padilla, 2010; Latournerié, com. pers.). La zona de preferencia
térmica para las crías, juveniles, subadultos y adultos oscila entre 19 y 25 °C,
siendo este rango térmico donde se optimizan los costos del metabolismo de
mantenimiento de la especie (Cornejo, 1991), de tal manera que una
temperatura de 23 ± 1 °C coloca a Cambarellus (C.) montezumae en un rango
óptimo para llevar acabo sus funciones vitales de sobrevivencia, crecimiento y
reproducción.
La concentración mínima de oxígeno requerida por una especie varía pero el
agua no debe presentar concentraciones menores a 70% de saturación de
oxígeno para organismos acuáticos (Gilbert, 1991). Con respecto a los niveles
de oxígeno disuelto disponible se observaron valores de 6.0 ± 0.1 mg/L.
Auvergne (1982) menciona que los valores de este factor para los acociles, se
deben ubicarse en el rango de 3 a 12 mg/L; ya que bajas concentraciones de
oxígeno pueden causar la muerte o suceptibilidad a patógenos, aun así, estos
organismos tienen gran tolerancia a niveles de 1 a 2 mg/L. Holdich (2002)
refiere que estos organismos pueden tolerar bajas concentraciones de oxígeno
gracias a que poseen un pigmento respiratorio (hemocianina) que incrementa la
eficiencia de transporte de oxígeno en su hemolinfa.
Los acociles requieren agua ligeramente alcalina con pH entre 6 y 8.5, donde es
imprescindible la presencia de calcio. Palacios (2003) señala que este elemento
es necesario para la calcificación del exoesqueleto de estos organismos durante
su crecimiento. A su vez, el pH también funciona como regulador en los
procesos respiratorios y enzimáticos, un valor por debajo de 5 o por arriba de 9

166
es peligroso o letal (Rosas, 1976). En este estudio los valores de pH se
mentuvieron en un intervalo de 8.4 ± 0.1.
La Acuicultura tiene como objetivos básicos la sobrevivencia, el crecimiento, y
la reproducción de las especies de mayor interés socio-económico. En la
práctica, esta actividad supone el mantenimiento de los organismos en
condiciones de cautividad con objeto de comercializarlos para su consumo, la
reintroducción de la especie a su habitat natural o con la intención de
incrementar su producción por encima de los niveles obtenidos en el medio
natural.
El cultivo intensivo de especies acuáticas y en particular de acociles introduce,
en la mayoría de los casos, variaciones respecto a las condiciones en las que se
encuentran en las poblaciones naturales, provocando alteraciones en su ciclo
reproductivo (Matsuyama, et al., 1991). Por tanto, el conocimiento de los
mecanismos que regulan la función reproductora de especies de interés es un
requisito indispensable para el desarrollo de la Acuicultura. Una vez
adquiridos los conocimientos necesarios sobre el proceso reproductor y su
regulación en una especie determinada (Cambarellus (C.) montezumae) es
posible controlar en cierta medida las distintas etapas de la reproducción.
La reproducción de los crustáceos es un proceso muy complejo por la diversidad
de elementos que determinan que un organismo se reproduzca. Los eventos
reproductivos están sujetos a múltiples estímulos del ambiente y de los propios
organismos. En esta investigación se evaluó el efecto que tiene una dieta
formulada (Acocilina) enriquecida en proteína y ácidos grasos sobre la
sobrevivencia, el crecimiento y la reproducción del acocil Cambarellus (C.)
montezumae bajo condiciones controladas.

167
A pesar de ser un grupo con alto potencial acuicultural y comercial, en la
actualidad es poco lo que se conoce en relación a los requerimientos
nutricionales de los acociles, y en particular del acocil Cambarellus (C.)
montezumae, por lo que la discusión en relación a sus requerimientos
nutricionales y sus efectos se presume pueden ser similares a las de otros
organismos acuáticos como camarones y peces (Bardach, et al., 1990).
Se puede señalar que los acociles en su hábitat son omnívoros, es decir,
ingieren materia vegetal o animal viva o muerta. Debido a que los acociles no
son depredadores activos no pueden capturar peces o insectos que nadan
rápidamente. Generalmente, cerca del 20% de la dieta del acocil consiste en
gusanos, larvas de insectos y otros tipos de materia relativamente inactiva, el
resto consiste en vegetación. El material vegetal consumido contiene grandes
cantidades de animales microscópicos particularmente cuando está en estado
de descomposición y durante el proceso el contenido de proteína es
considerablemente mayor debido al material de origen animal (Davis, 1987a;
García-Padilla, 2010, Escalante, 2013).
No pueden seguir desconociéndose las necesidades nutricionales indispensables
del acocil Cambarellus (C.) montezumae, si se pretende realizar producciones
comerciales con esta especie. Problema que poco a poco ha ido subsanándose,
ya que actualmente se inician estudios de especies nativas, como son los
acociles mexicanos del género Cambarellus, probándose dietas tanto naturales
como artificiales y estableciéndose las bases para el desarrollo de una
tecnología con alto potencial acuícola (García, 1991; Moctezuma, 1996;
Vásquez, 2009; Escalante, 2013).
Para desarrollar un cultivo a nivel intensivo es necesario contar con un
alimento de buena calidad que cubra los requerimientos de la especie y además

168
que su costo no sea elevado. En la elaboración de dietas para acociles, además
de utilizar subproductos agrícolas, también se pueden usar desperdicios de la
pesca (Cordero, 1988), tales como pescado de desecho, harina de sardina,
harina de calamar, algunos tipos de aceites (animales y vegetales) y
subproductos agrícolas.
De aquí la importancia de elaborar dietas que realmente cubran los
requerimientos de la especie, sin olvidar la parte económica de la misma; al
respecto para este trabajo se utilizó una dieta comercial Camaronina (Purina,
40% de proteína), la cual es la línea de alimentos completos pelletizados para
camarón de cultivo formulados y utilizados en sistemas de producción semi-
intensivo e intensivos. En la línea Camaronina se ofrecen alimentos con 35 y
40% de proteína, mismos que han sido formulados utilizando los últimos
avances tecnológicos y un balance adecuado de nutrientes, por lo cual se eligió
como dieta control. La dieta experimental denominada Acocilina, estuvo
conformada con un 47% de Camaronina (Purina, 40% de proteína), 47% de
harina de calamar y 6% de aceite de hígado de pescado.
Con la dieta experimental se pretendía mejorar el crecimiento, la
sobrevivencia, la reproducción, la calidad de los reproductores y la puesta, por
lo cual se decidió aumentar los niveles de proteínas y ácidos grasos en la dieta
mediante el uso de harina de calamar y el aceite de pescado.
La harina de calamar y el aceite de pescado son productos químicos de origen
natural obtenidos de la reducción de pesca pelágica con la consiguiente
producción de una línea de alimentos protéicos (la harina de calamar) y de una
línea de productos grasos (el aceite de pescado). Ambos productos son
utilizados en la formación de alimentos balanceados para la nutrición animal,

169
ya sea de productos acuícolas, de aves, de rumiantes, de cerdos y de estos
animales domésticos.
El calamar posee un alto nivel de proteínas, carbohidratos y grasas
(monoinsaturadas y poliinsaturadas), vitamina A, B3, B12, C, D y K, minerales
(Calcio, Hierro, Zinc, Yodo, Magnesio y Fosforo), aminoácidos (Ácido aspártico,
Ácido glutámico, Alanina, Arginina, Cisteína, Leucina, Metionina, Tirosina,
Triptófano y Valina) así como carotenos y retinol. El aceite de pescado aportó
a la dieta ácidos grasos omega-3 de cadena larga; eicosapentaenoico (C20:5,
EPA), docosapentaenoico (C22:5, DPA) y docosahexaenoico (C22:6, DHA),
(Valenzuela, 2012).
Otro aspecto importante fue la estabilidad de las dietas en el agua, ya que
contribuye a la conservación de su calidad; los crustáceos requieren de un
alimento de buena consistencia y alta estabilidad en el agua, ya que no lo
consumen rápidamente en el transcurso del día; por lo que los productores de
alimento para crustáceos recomiendan que su estabilidad en el agua sea de por
lo menos 24 horas (Tacos, 1987). Sin embargo, Zendejas (1991) indicó que no es
necesario que la dieta dure más de 6 horas en el agua ya que si ésta cuenta con
los atrayentes y palatabilidad adecuados, el organismo lo consumirá en un
lapso no mayor de 2 a 3 horas, que fue el lapso en el que los acociles consumían
la dieta formulada (Acocilina), en el caso de la dieta control (Camaronina), su
estabilidad en el agua y su palatabilidad da buenos resultados.
La nutrición de los reproductores sigue siendo una de las áreas más
pobremente entendidas e investigadas en el campo de la nutrición de
crustáceos, y los estudios realizados están limitados a unas pocas especies como
es el caso del camarón y el langostino (Brooks, et al., 1997; Izquierdo, et al,
2001). A grandes rasgos esto es debido a la necesidad de grandes

170
instalaciones, interiores o exteriores, para mantener grandes grupos de
crustáceos adultos y, consecuentemente, al alto costo necesario para poder
desarrollar largos experimentos de alimentación de reproductores.
Sin embargo, como ocurre en la nutrición humana y en ganadería
(Leboulanger, 1977), es obvio que los requerimientos nutricionales de los
reproductores se diferencian claramente de los que requieren los animales
juveniles en rápido crecimiento. Es más, como en otros animales, está claro
que muchos de los problemas y deficiencias que aparecen en las etapas
tempranas del desarrollo de las crías recién eclosionadas están directamente
relacionadas con el régimen alimenticio (tamaño de la ración, nivel de
nutrientes y periodo de alimentación) de los reproductores.
Varios estudios realizados sobre las principales especies cultivadas
(principalmente de camarón y peces), han demostrado que la reproducción y la
calidad del huevo son influenciadas por nutrientes como proteínas, lípidos,
minerales, vitaminas y carbohidratos, así como por el tamaño de la ración y por
el periodo de alimentación de los reproductores. Esto ha sido descrito en
especies como Procambarus clarkii (Hernández, 1996), Cherax tenuimanus
(Morrissy, 1989; Villareal, 1995), Cherax destructor (Mills & McCloud, 1983), y
Cherax quadricarinatus (Jones, 1995; Hutchings & Villarreal, 1996).
Una restricción en el nivel de ingesta afecta seriamente a la puesta. Así, la
reducción en la tasa de alimentación ha sido descrita como la causa de
inhibición de la maduración de las gónadas, la disminución de las tasas de
crecimiento, el retraso del período de puesta, los huevos y crías recién nacidas
son más pequeñas que las obtenidas de organismos que ingieren la ración
completa (Sasayama & Takahashi, 1972; Berglund, 1995).

171
Algunos estudios realizados en el medio natural han mostrado que la
disponibilidad de alimento es el principal factor ambiental determinante de la
fecundidad, que se puede definir como el número de huevos maduros que son
producidos por una hembra durante un ciclo reproductivo (Wotton, 1979;
Bromageeía, 1992). Algunos de estos estudios se han llevado a cabo sobre
especies dulceacuícolas que habitan en ríos con diferente disponibilidad de
alimento, mostrando que los ejemplares que viven en los ríos con más baja
productividad muestran una disminución en su fecundidad (McKay & Mann,
1969; Constanz, 1975; Ali & Wooton, 2000). Los estudios realizados en el
medio natural muestran ligeras evidencias de una relación directa entre el
tamaño de la ración y la reproducción, mostrando la necesidad de realizar
experimentos en cautividad que estudien los efectos del tamaño de la ración
sobre la fecundidad, los cuales aún no se han realizado en el acocil Cambarellus
(C.) montezumae, ya que para este experimento los organismos fueron
alimentados ad libitum.
Experimentos realizados en condiciones de cautividad, han mostrado que
restricciones en la ración tienen como consecuencia una reducción de la
fecundidad, y en algunos casos, de la proporción de ejemplares que llegan a la
maduración sexual; para el acocil Cambarellus (C.) montezumae alimentado ad
libitum con ambas dietas (formulada y control) encontramos una talla de
primera madurez entre los 75 y 90 días de experimentación para la dieta
formulada, y de 90 a 105 días para la dieta control.
Uno de los parámetros utilizados para evaluar la calidad de la puesta es el de
la fecundidad, la cual es considerada como el número medio de huevos
producidos por hembra (Wotton, 1979). La fecundidad se ve afectada por
deficiencias nutricionales en las dietas de los reproductores, fecundidades
reducidas han sido citadas en varias especies causadas por la influencia de un

172
desequilibrio de nutrientes que afecta el sistema endócrino, o por restricciones
en la disponibilidad de componentes bioquímicos para la formación de los
huevos.
El aumento en los niveles de lípidos dietéticos produce un aumento en la
fecundidad (Duray, et al., 1994), aunque este efecto también pudiera
relacionarse con el aumento gradual en los ácidos grasos esenciales de la dieta.
De hecho, uno de los factores nutritivos que más pueden afectar la calidad de la
puesta es el contenido en ácidos grasos esenciales (AGEs) de la dieta
(Watanabe, et al., 1984 a, b). La dieta formulada (Acocilina), al estar
adicionada con aceite de hígado de pescado, posee un alto contenido de ácidos
grasos omega-3 de cadena larga; eicosapentaenoico (EPA), docosapentaenoico
(DPA) y docosahexaenoico (DHA), lo cual se vió reflejado en la calidad
bioquímica de los reproductores, en el número de huevos totales y en la calidad
de la puesta, ya que todas estas variables tuvieron una mejor respuesta en
comparación a los organismos alimentados con la dieta control (Camaronina).
Asimismo, la composición protéica de la dieta de los reproductores influye en la
calidad de la puesta regulando la síntesis y selección de los componentes del
saco vitelino (Tandler, et al., 1995). Las proteínas actúan como fuente de
aminoácidos y como material de reserva utilizado durante muchas actividades
biosintéticas que son esenciales para las etapas tempranas de la embriogénesis
(Metcoff, 1986). El desarrollo embrionario depende del balance de aminoácidos
presentes en el huevo (Fynh & Serigstad, 1987; Fynh, 1989). El contenido de
proteínas para la dieta control fue del 46 % en comparación con la dieta control
con un 26 % de proteína cruda, el origen de la proteína de la dieta control
(Camaronina) se basa en el desperdicio de cabeza de camarón entre otros
subproductos; la proteína de la dieta formulada (Acocilina) fue de primera
calidad, la cual fue aportada principalmente por el harina de calamar que

173
además posee una fuente importante de aminoácidos como ácido aspártico,
ácido glutámico, alanina, arginina, cisteína, leucina, metionina, tirosina,
triptófano y valina (http://alimentos.org.es/calamar/).
En estudios llevados a cabo en especies acuáticas se ha estimado que el nivel
óptimo de proteínas en dietas, conteniendo harina de pescado como principal
fuente de energía, está alrededor del 45%; los reproductores alimentados por
debajo de ese nivel producen aproximadamente 30% menos de huevos
(Watanabe, et al., 1984). De la misma manera, se ha probado que
reproductores alimentados con dietas conteniendo diferentes porcentajes de
proteína, la fecundidad está positivamente correlacionada con el nivel de
proteína contenida en la dieta (El-Sayed, et al, 2003).
Otros nutrientes que pueden afectar la fecundidad son la vitamina E (Izquierdo
y Fernández-Palacios, 1997; Fernández-Palacios, et al., 1998), la vitamina C
(Blom & Dabrowski, 1995), la combinación de ambas vitaminas (Emata, et al.,
2000) y los carbohidratos. Los ácidos grasos poliinsaturados también influyen
de manera determinante en los procesos reproductivos (Moore, 1985),
incluyendo la producción de hormonas, el desarrollo gonadal y la ovulación.
Varios nutrientes son esenciales para el normal desarrollo del embrión y su
nivel óptimo en las dietas de los reproductores mejora la morfología del huevo y
el porcentaje de eclosión. Se ha encontrado que el porcentaje de huevos
morfológicamente normales se incrementa con un aumento de los niveles de
ácidos grasos esenciales (DHA y EPA), en las dietas de los reproductores y la
incorporación de estos ácidos grasos en los huevos (Fernández-Palacios, et al,
1995). Estos ácidos grasos desempeñan un papel estructural importante como
componentes de fosfolípidos en las biomembranas del organismo y están

174
asociados con su fluidez y el correcto funcionamiento de las funciones celulares
(Bell, et al., 1986).
En algunas especies los ácidos grasos esenciales se consideran la mayor fuente
de energía durante el desarrollo embrionario temprano (Falk-Peterseneí, et al.,
1989). No obstante, la composición en ácidos grasos de los lípidos de los
huevos no sólo es determinada por la dieta de los reproductores, sino que
también está relacionada con la calidad de los reproductores mismos (Pickova,
et al., 1997).
Los radicales libres pueden deteriorar las membranas del huevo y la integridad
de las mismas. La vitaminas E y C y los carotenos (por ejemplo la
astaxantina) se han mostrado como importantes agentes contra la acción de
radicales libres. Un aumento de los niveles dietéticos de vitamina E produce un
aumento en los porcentajes de huevos fecundados, de eclosión y de larvas
normales (Watanabe, et al, 1991 a). La función de la vitamina E como
antioxidante ínter e intracelular para mantener la homeostasis de metabolitos
lábiles en la célula y en el plasma tisular es bien conocida.
De nuevo, estos resultados sugieren que la composición bioquímica de los
huevos no debe ser usada como el único criterio para determinar su calidad, a
pesar del hecho de que varios autores (Sandnes, et al, 1984; Craik, 1985; Harel,
et al., 1994) han sugerido que la composición química de los huevos está
relacionada con el éxito de la puesta, desde el momento en que los nutrientes
almacenados en el huevo deben satisfacer las demandas nutricionales para el
desarrollo del embrión y su crecimiento.
Pocos estudios han podido demostrar la mejora de calidad de la puesta a través
de la dieta de los reproductores, en el caso de este estudio es factible señalar

175
que se puede demostrar que la alimentación de los reproductores juega un
papel esencial en la sobrevivencia, crecimiento, calidad de los reproductores y
de la puesta de crías de acocil Cambarellus (C.) montezumae, abriendo la
puerta para realizar investigaciones más detalladas acerca de los
requerimientos nutricionales de esta especie.

176
7.7 CONCLUSIONES.
Las conclusiones que se obtuvieron en este capítulo sobre el efecto de una dieta
formulada sobre el crecimiento, sobrevivencia y calidad de la puesta del acocil
Cambarellus (C.) montezumae bajo condiciones controladas son:
El efecto del aumento en los niveles de proteínas y lípidos (ácidos grasos)
en la dieta del acocil Cambarellus (C.) montezumae tuvo un efecto
significativo sobre la tasa de crecimiento y sobrevivencia de los
organismos, encontrando una menor tasa de crecimiento en los
organismos alimentados con la dieta formulada, lo cual se vio
compensado con una mayor sobrevivencia y menor talla de primera
madurez. Lo que pone de manifiesto que estos organismos realizan una
compartamentalización de la energía destinada al crecimiento y la
reproducción.
El efecto de los altos niveles de proteínas y ácidos grasos en la dieta se
vieron también reflejados en el índice de desove y la fecundidad de
hembras del acocil Cambarellus (C.) montezumae, obteniendo los mejores
resultados con la dieta la formulada. Lo cual estaría mostrando la
idoneidad de la dieta formulada (niveles de proteínas y lípidos) para
obtener mejores resultados sobre la producción de crías.
Se obtuvo una producción de progenie numerosa, saludable y uniforme
de los organismos alimentados con la dieta formulada, esto se observó en
la calidad del huevo, que presentaron mayor contenido de proteínas y

177
lípidos en comparación con los huevos producidos por los organismos
alimentados con la dieta control. Esto a su vez, se vió reflejado en menor
tiempo de incubación y mayor sobrevivencia de las crías de las hembras
ovígeras alimentadas bajo la dieta formulada.

178
8. LITERATURA CITADA.
Aguilar, E. R. 1991. Producción y crecimiento en Cambarellus montezumae
(Saussure), (Crustacea: Astacidae), empleando alimento vegetal. Tesis de
Licenciatura. Facultad de Ciencias. Universidad Nacional Autónoma de
México. 76 p.
Aguilar-Román, E. 2011. Efecto de la temperatura en el metabolismo
energético y crecimiento de crías de acocil Cambarellus montezumae
(Saussure). Tesis de licenciatura. Facultad de ciencias. Universidad
Nacional Autónoma de México. 89 p.
Agius, R.V., T. Watanabe, S. Satoh & V Kiron. 2001. Supplementation of
paprika as a carotenoid source in soft-drypellets for broodstock yellowtail
Serióla quinqueradiata (Temminck & Schlegel). Aquaculture Research.
32:1, 263-272 pp.
Alva, M. 1996. La calidad del agua en la Pista de Remo y Canotaje “Virgilio
Uribe”, México, D. F. con base en el contenido de nutrientes y factores
físicos. Tesis de Licenciatura. Universidad Autónoma Metropolitana –
Xochimilco, 35 p.
Álvarez, F. y R. Rangel. 2007. Estudio poblacional del acocil Cambarellus
montezumae (Crustacea: Decapoda: Cambaridae) en Xochimilco, México.
Revista Mexicana de Biodiversidad 78: 431-437 pp.
Andersson, M. 1999. Sexual selection. Princeton University Press. 218 p.

179
Appelberg, M. 1989. Evaluating water quality criteria for freshwater:
exemplified by the impact of acid – stress. In: J. Skurdal, K. Westman y
P. I. Bergan (eds.) Crayfish Culture in Europe. 476 p.
Aragones, R. G. 1995. Realizar investigación que permita conocer y aplicar la
biotecnología para producir en cautiverio acocil, rana, ajolote, desarrollo
de lotes. Informe servicio social. Universidad Autónoma Metropolitana -
Iztapalapa. 101 p.
Arana, M. F., R. Pérez-Rodríguez y A. Malpica-Sánchez. 1998. Cambaridos de
tres embalses del Estado de Tlaxcala, México (Crustacea: Decapoda).
Revista de la Sociedad Mexicana de la Historia Natural 48: 23 – 35 pp.
Arcos, R. E. y B. Barragán. 1995. Evaluación de la Calidad del Agua de los
Canales Chinamperos de Xochimilco, Posibles Alternativas de Usos y
Tratamiento. Memorias, II Seminario Internacional de Investigación de
Investigadores de Xochimilco. 76-82 pp.
Arredondo-Figueroa, J. L., A. Vásquez-González, L. Núñez-García, I. Barriga-
Sosa y J. Ponce-Palafox. 2011. Aspectos reproductivos del acocil
Cambarellus (Cambarellus) montezumae (Crustacea: Decapoda:
Cambaridae) en condiciones controladas. Revista Mexicana de
Biodiversidad 82: 168-178 pp.
Auvergne, M. 1982. El cangrejo de río. Mundi-Prensa. Madrid España. 119 p.

180
Avault, J. W. & J. V. Huner. 1985. Crawfish culture in the United States. In:
Huner J. V. & E. E. Brows (Eds.), Crustacean and mollusk aquaculture
in the United States. AVI. Publ. Co. Westpent, Connecticut, 1-61 pp.
Ávila, S. F. 2011. Determinación de la calidad microbiológica del acocil
Cambarellus montezumae. Tesis de Licenciatura. Facultad de Medicina
Veterinaria Zootecnista. Universidad Nacional Autónoma de México. 81
p.
Balanzario, Z. 1982. Contaminación de los Canales de Xochimilco y su
Repercusión en las Actividades Económicas. Boletín Mexicano de
Geografía y Estadística (Archivo Histórico de la Delegación Xochimilco).
247-284 pp.
Barbosa, M. 1998. Aplicación de modelos estadísticos al estudio de crecimiento
del acocil Cambarellus montezumae (Saussure) en condiciones de
laboratorio. Tesis de especialidad (Estadística Aplicada). Universidad
Nacional Autónoma de México. 32 p.
Barcena, B. L. 2000. Evaluación de dietas regionales en el crecimiento
(longitud y peso) del acocil Cambarellus montezumae, bajo condiciones de
cultivo. Tesis de licenciatura. Biología agropecuaria. Universidad
Autónoma de Tlaxcala. 96 p.
Begon, M., C. Townsend & J. Harper. 2006. Ecology, from Individuals to
Ecosystems. Blackwell Publishing Ltd., U.K. 698 p.

181
Brafield, A. E. & D. J. Salomon. 1972. Oxicalorific coefficients for animals
respiring nitrogenous substrates. Comparative Biochemistry Physiology
43:837-841 pp.
Bromage, J., J. Jones, C. Randall, M. Thrush & B. Davies. 1992. Broodstock
management, fecundity, egg quality and timing of egg production in the
rain brown trout (Onchorhynchus mykiss). Aquaculture.100: 141-166 pp.
Bromage, N. R., M. Bruce & N. Basavaraja. 1994. Egg quality determinants in
finfish: the role of overripening with special reference to the timing of
stripping in the Atlantic halihut Hippoglossus hippoglossus. Journal of
the World Aquaculture Society. 25: 13 -21 pp.
Brooks, S., C. Schreck, W. Contreras-Sanchez & M. S. Fitzpatrick. 1997. Egg
quality in fish: What makes a good egg? Reviews in Fish Biology and
Fisheries. 7: 387-416 pp.
Bruce, M. F., F. Oyen, G. Bell, F. Asturiano & B. Farndale. 1999. Development
of brood stock diets for the European sea bass (Dicentrarchus labrax)
with special emphasis on the importance of n-3 and n-6 HUFA to
reproductive performance. 117(1-4): 85-97 pp.
Bückle-Ramírez, L., F. Díaz, F. Correa, B. Barón & M. Hernández. 1994. Diel
thermoregulation of the crawfish Procambarus clarkia (Crustacea,
Cambaridae). Journal of Thermal Biology, 19(6):419-422 pp.
Campbell, P. M., M. Morgan, C. Wilson & L. Crim. 1994. Preliminary evidence
that chronic confínement stress reduces the quality of gametes produced
by brown and rainbow trout. Aquaculture.120: 151-169 pp.

182
Cantú, L. L. 1959. Contribución al conocimiento de la embriología de una
especie de acocil: Cambarellus montezumae montezumae (Saussure).
Crustacea: Decápoda. Tesis de licenciatura. Facultad de Ciencias.
Universidad Nacional Autónoma de México. 37 p.
Capital Natural de México. 2009. Acciones Estratégicas para su Valoración,
Preservación y Recuperación. Comisión Nacional para el Conocimiento y
Uso de la Biodiversidad. México. 95 p.
Carta Nacional Acuícola. 2012. Diario Oficial. Estados unidos Mexicanos.
Secretaría de Agricultura, Ganadería, Desarrollo Rural Pesca y
Alimentación. Segunda Sección. 80 p.
Carrillo, M., J. Cerdá, J. Navas, F. Oyen, J. Ramos & S. Zanuy. 2000. Some
criteria of the quality of the progeny as indicators of physiological brood
stock fitness. Cah. Options. Mediterr., 41: 61-13 pp.
Castelló, O. 1993. Acuicultura marina: fundamentos biológicos y tecnología de
la producción, Barcelona, España. 310 p.
Cerón, O. A., M. M. Ángeles y E. J. León. 2010. Evaluación de la influencia del
alimento y calidad del agua en el crecimiento de post-larvas del acocil
(Cambarellus montezumae). I Reunión Nacional de Innovación Acuícola y
Pesquera de Campeche. Nutrición Acuícola. 41 p.
Cerón, O. A., M. M. Ángeles y E. J. León. 2011. Resultados preliminares sobre
la reproducción como un efecto secundario en cultivos controlados del

183
acocil de río (Cambarellus montezumae). Memoria II Reunión Nacional
de Innovación Acuícola y Pesquera. León, Guanajuato. 59 p.
Chapman, W. D. 1971. Production. In: Bagenal, T. (Ed.) Methods for
assessment of fish production in fresh waters. 3er. Ed. IBP. No 3.
Blackwell Sci. Publ. Oxford. 365 p.
Civera, C. R. y S. L. Cruz. 1982. Bioensayo de incubación artificial del acocil
Cambarellus montezumae, (Saussure, 1858), (Decapoda: Astacidae).
Informe servicio social. Universidad Autónoma Metropolitana -
Iztapalapa. 75 p.
Colín, H. S. 2006. Estudio sobre el crecimiento y ciclo de vida del Cambarellus
montezumae montezumae (Saussure, 1857) de los canales de Xochimilco;
México, DF. Informe Servicio Social. Universidad Autónoma
Metropolitana - Iztapalapa. 57 p.
Coll, M. J. 1987. Cría del cangrejo de río. Editores Hispano Europea.
Barcelona-España. 158 p.
Cornejo, R. A. 1991. Selección térmica del acocil Cambarellus montezumae
(Saussure) (Crustacea: Astacidae) y su correlación con algunos índices
fisiológicos: diferencias estacionales. Tesis de licenciatura. Facultad de
ciencias. Universidad Nacional Autónoma de México. 71 p.
Cortés, J. E., H. Villareal-Colmenares y M. Rendón-Rumualdo. 2002. Efecto de
la frecuencia alimenticia en el crecimiento y sobrevivencia de juveniles
de langosta de agua dulce Cherax quadricarinatus (Decapoda:

184
Parastacidae). I Congreso Iberoamericano Virtual de Acuacultura. 882-
890 pp.
Cortés, O. A. 2010. Interacción de depredación entre el ajolote (Ambystoma
mexicanum) y el acocil (Cambarellus montezumae) especies nativas de
Xochimilco. Tesis Maestría en Ciencias Biológicas. Universidad Nacional
Autónoma de México. 64 p.
Coutiño, M. 1981. Evaluación Bacteriana en Vegetales Irrigados con Aguas
Negras en la Zona de San Gregorio, Xochimilco. Tesis de Licenciatura.
Facultad de Ciencias. Universidad Nacional Autónoma de México. 84 p.
Clutton-Brock, T. H. 1991. The Evolution of Parental Care. Princeton (NJ):
Princeton University Press.
Craik, J. C. & S. M. Harvey. 1984. Egg quality in rainbow trout. The relation
between egg viability, selected aspects of egg composition, and time of
stripping. Aquaculture, 40: 115-134 pp.
Dabrowski, K. & J. H. Blom. 1994. Ascorbic acid deposition in rainbow trout
(Oncorhynchus mykiss) eggs and survival of embryos. Comparative
Biochemistry and Physiology. 129-135 pp.
Dajoz, R. 2002. Tratado de ecología. Mundi-Prensa Libros. 600 pp.
De Gaves, S., N. Sammy & D. Pentcheff. 2009. A classification of living and
fossil genera of decapods crustaceans. National University Singapore.
Raffles bulletin of Zoology. Supplement No. 21: 1-109 pp.

185
Díaz, F., G. Escalante y E. Sierra. 2006. Fisiología energética de Cherax
quadricarinatus (Von Martens) alimentado con dos dietas, expuesto a un
régimen constante y fluctuante de temperatura. Hidrobiológica.
Universidad Autónoma Metropolitana - Iztapalapa.16: 1, 35-44 pp.
Emata, A., I. Borlongan & J. Damaso. 2000. Dietary vitamin C and E
supplementation and reproduction of milkfish Chanos chanos Forsskal.
Aquaculture Research, 31:7, 557-564 pp.
Escalante, G. E. 2013. Crecimiento de crías de acocil Cambarellus montezumae
(Saussure) con diferentes dietas vegetales. Tesis de Licenciatura.
Facultad de Ciencias. Universidad Nacional Autónoma de México. 99 p.
Espinoza, E. G. 1992. Evaluación de una calentador solar rustico con
funcionamiento nocturno para el cultivo del acocil mexicano
(Cambarellus montezumae) en clima templado. Colegio Nacional de
Educación Profesional Técnica el Zarco. 142 pp.
Ezcurra, E. 1990. De las Chinampas a la Megalopólis. México. Fondo de
Cultura Económica. (La Ciencia desde México). 65 p.
Feist, G., F. Grant F., Eenennaam J., Doroshov S., Schneider R. &
Fitzpatricke M. 2004. Early identification of sex in cultured White
sturgeon, Acipenser transmontanus, using plasma steroid levels.
Aquaculture, 232: 581-590 pp.
Félix, C. L. y Gutiérrez, Y. P. 2005. Influencia de los factores ambientales en la
ecología poblacional de Cambarellus montezumae (Crustacea, Decápoda).

186
Resumen. Memorias XVII Congreso Nacional de Zoología, Monterrey,
Nuevo León. 80-84 pp.
Fernández-Palacios, H., N. Sammy & D. Pentcheff. 1995. Effect of n-3 HUFA
level in broodstock diets on egg quality of gilthead sea bream (Sparus
aurata). Aquaculture.132 (3-4): 325-337 pp.
Flores, M. G. 1982. Determinación de algunos metales pesados en el sedimento
de los Canales del Lago de Xochimilco. Tesis Profesional. Escuela
Nacional de Ciencias Biológicas. Instituto Politécnico Nacional. México.
100 p.
Flores, R. A. 2009. Análisis preliminar de algunos factores físico-químicos
indicadores de la calidad del agua en Xochimilco. Tesis de Licenciatura.
Universidad Nacional Autónoma de México. 92 p.
Folch, J., M. Lees & G. H. Stanley. 1959. A simple method for the isolation and
purification of total lipids from animal tissues. The Journal of Biological
Chemistry. 226: 497-509 pp.
Fondo Internacional de Emergencia de las Naciones Unidas para la Infancia.
UNICEF. 2012. El doble reto de la malnutrición y la obesidad. ONU.
www.unicef.org/mexico/spanish/17047.htm
Fragoso, C. M., H. Martínez y M. Rodríguez. 2003. Estudio comparativo del
comportamiento productivo en acuario, del acocil (Cambarellus
montezumae) alimentado con excretas de cerdo ensiladas y empastillados

187
contra un alimento comercial para tilapia. REDVET - Revista
Electrónica de Veterinaria. Universidad Nacional Autónoma de México.
Furuita. H., H. Tanaka, T. Yamamoto, M. Shiraishi & T. Takeuchi. 2003a.
Supplemental effect of vitamin A in diet on the reproductive performance
and egg quality of the Japanese flounder Paralichthys olivaceus (T&S).
Aquaculture Research, 34(6): 461-468 pp.
García, O. A. 1991. Efecto de diversas dietas sobre la sobrevivencia y
crecimiento de crías del acocil Cambarellus montezumae (Saussure),
(Crustacea: Astacidae). Tesis de Licenciatura. Facultad de Ciencias.
Universidad Nacional Autónoma de México. 73 p.
García-Padilla, G. M. 2010. Estimación del balance energético del acocil
Cambarellus montezumae en el lago de Xochimilco. Tesis de
Licenciatura. Facultad de Ciencias. Universidad Nacional Autónoma de
México. 92 p.
Gilbert, B. 1991. Acuacultura. Eds. Omega, S.A. Barcelona. Vol. 1: 478 p.
Gherardi, F. 2002. Behaviour. In: D. M. Holdich (ed.), Biology of freshwater
crayfish. Blackwell Science. UK. 258 – 290 pp.
González, V. E. 1997. Presencia de metales pesados en Cambarellus
montezumae, Ambistoma mexicanum y Chirosthoma jordani, especies
endémicas comestibles del área lacustre de Xochimilco. Informe servicio
social. UAM Xochimilco. 63 p.

188
González-Vecino, J. L., T. Lyons & K. Jacques. 2004. Short & long term effects
of a nucleotide enriched broodstock diet on the reproductive performance
of haddock (Melanogrammusa eglefinus). 11 International Symposium
on nutrition and Feeding in Fish. Phuket. Thailand. 99 pp.
Guillaume, J., Bourne & N., Hodgson. 2004. Nutrición y alimentación de peces
y crustáceos. Editorial Mundi-Prensa, España. 475p.
Gutiérrez-Yurrita, P. J. 2008. Nuevos paradigmas y retos de la ecología. En:
Crustáceos de México estado actual de su conocimiento. Álvarez, F. N. y
R. A. Gabino. Universidad Autónoma de Nuevo León.301-329 pp.
Hamasaki, K. & F. Chu. 2006. Batch fecundity of the swimming crab Portunus
trituberculatus (Brachyura: Protunidae). Aquaculture. 253: 359-365 pp.
Harel, M., J. Bell, I. Uriarte, R. Henderson & J. Sargent, J. 1994. The kinetics
of nutrient, incorporation into body tissues of gilthead sea bream
(Sparus aurata) females and the subsequent effects on egg composition
and egg quality. British Journal of Nutrition, 72: 45-58 pp.
Hines, A. 1988. Fecundity and reproductive output in two species of deep-sea
crabs, Geryon fenneri and G. quinquedens (Decapoda: Brachyura).
Journal Crustacean Biology. 8 (4): 557-562 pp.
Hines, A. 1991. Fecundity and reproductive output in nine species of Cancer
crabs (Crustacea, Brachyura, Cangridae). Canadian Journal of Fisheries
and Aquatic Sciences. 48: 267-275 pp.

189
Hinojosa, D. & L. Zambrano. 2004. Interaction of common carp (Cyprinus
carpio) with bentic crayfish decapods in shallow ponds.
Hydrobiology.515: 115-122 pp.
Hem, J. D. 1985. Study and interpretation of the chemical characteristics of
natural water. 3rd Ed. U.S.Survey Water Supply Paper 2254.
Washington (USA). 1-263 pp.
Holdich, D. M. & R. S. Lowery (Eds.). 1988. Freshwater Crayfish, biology,
management and exploitation. Cromm Helm Press. London. 198 p.
Hobbs, H. H., Jr., 1989. An illustrated checklist of the American Crayfish
(Decapoda: Astacoidae: Cambaridae: Parastacidae) Smithsonian
Contributions to Zoology, 480: 1 – 236 pp.
Holdich, D. M. 2002. Biology of freshwater crayfish. Blackwell Science. Oxford
England. 702 p.
Huner, J. V. & J. E. Barr. 1984. Red swamp crawfish. Biology and exploitation.
The Louisiana Sea Grant College Program, Louisiana State University.
135 p.
Huner, J. V. 1981. Information about the biology and culture of red crawfish
Procambarus clarkii (Girad, 1852) (Decapoda: Cambaridae) for fisheries
managers in Latin America. Anales del Instituto de Ciencias del mar y
Limnología. Universidad Nacional Autónoma de México. 8(1):43-50 pp.

190
Hutchings, R. W. & H. Villarreal. 1996. Biología y Cultivo de la langosta de
agua dulce Cherax quadricarinatus. Manual de Producción. Navimar, S.
A. Guayaquil, Ecuador. 500 pp.
Inouye, L. S. & G. R. Lotufo. 2006. Comparison of macro-gravimetric and
micro-colorimetric lipid determination methods. Talanta 70: 584-587.
Instituto Nacional de Estadística y Geografía (INEGI). 2008. Cuaderno
estadístico delegacional de Xochimilco, Distrito Federal. Sistema
Nacional de Información Estadística y Geográfica.
http://www.inegi.org.mx Consultado en marzo 2013.
Instituto Nacional de Estadística y Geografía (INEGI). 2000. Cuaderno
Estadístico Delegacional, Xochimilco, Distrito Federal. 163 p.
Jiménez, O. E. 1989. Meso microclima de la Ciudad de México. Universidad
Nacional Autónoma de México. 95 pp.
Johnson, S. K. 1987. Calidad del agua en el Cultivo del Cangrejo de Rio
(Acocil). FONDEPESCA. Extensionismo. FOEXT/A3.
Jones, C. M. 1990b. General biology of Cherax quadricarinatus. In: C.C. Shelly
and M. Pearce, (Eds.) Farming the red-claw freshwater crayfish. NT
Department of Primary Industry and Fisheries Report No.21. Northern
Territory, Australia. 1 – 6 pp.
Jones, C. M. 1995. Evaluation of six diets for red claw, Cherax quadricarinatus,
von Martens, held in pond enclosures, Tenth International Symposium of

191
Astacology. Geddes, M. C., Fielde, D. R. and Richardson, A. M. M. eds.
Louisiana State University, U. S. A. 399-409 pp.
Kiewek, M. M. 2002. Efecto del nivel de lípidos en el desarrollo de juveniles de
langosta de agua dulce Cherax quadricarinatus (Von Martens 1868).
Tesis de Licenciatura. Universidad Nacional Autónoma de México.78 p.
Kjorsvik, E., Y. Marty, J. Moal & F. Samain. 1990. Egg quality in fishes.
Advances in Marine Biology, 26: 71-113 pp.
Kjorsvik, E., Y. Marty, J. Moal & F. Samain. 2003. Egg and larval quality
criteria as predictive measures for juvenile production in turbot
(Scophthalmus maximus). Aquaculture, 227 (4): 9-20 pp.
Klekowsky, R. Z. & A. Duncan. 1975. Phisiological approach to ecological
energetic. In Methods for ecological Bioenergetics. I.B.P. Blackwell
Science Oxford. 100 p.
Latournerié-Cervera, J. R., Y. Nacif y R. J. Cárdenas. 2006. Crecimiento,
producción y eficiencia de energía de crías de acocil Cambarellus
montezumae (Saussure) alimentadas con detritus de Egeria densa.
Revista electrónica de veterinaria REDVET. 7(12). 1-11 pp.
Lot, A., A. Novelo, M. Olvera y P. Ramírez. 1999. Catálogo de angiospermas
acuáticas de México. Instituto de Biología. Universidad Nacional
Autónoma de México. 161 p.

192
Manahan, S. E. 1972. Environmental Chemistry. Willard Grant Press Mass,
U.S.A., 393 pp.
McHarney, W. 1984. The freshwater aquaculture book. Hartley & Marks Press,
N.Y., 583 p.
Maldonado, R. G. 1990. Respuesta al stress térmico del acocil Cambarellus
montezumae (Saussure), (Crustacea: Astacidae). Comparación de
métodos. Tesis de Licenciatura. Facultad de Ciencias. Universidad
Nacional Autónoma de México. 75 p.
Mangor-Jensen, A., H. Fernández-Palacios & A. G. J Tacon. 1993. Effects of cod
brood stock dietary vitamin C on embryonic growth and survival.
Milestone. Rapp. Sent. Havbruk. Inst. Mar. Res. 18 – 8 pp.
Mazorra, C., K. Miller & G. Newkirk. 2003. Dietary lipid enhancement of brood
stock reproductive performance and egg and larval quality in Atlantic
halibut (Hippoglossus hippoglossus). Aquaculture, 227 (1-4): 21-33 pp.
Mills, B. J., J. Philip & H. McCould. 1983. Effects of stocking and feeding rates
on experimental pond production of the crayfish Cherax destructor, Clark
(Decapoda: Parastacidae). Aquaculture, 34: 51-72 pp.
Moctezuma, M. A. 1996. Bases biológicas y técnicas para el cultivo del acocil
Cambarellus montezumae. Tesis de Maestría en Acuacultura.
Universidad de Colima. 85 p.

193
Monroy, A. M., J. Carabias, C. Mapes y C. Toledo. 2010. Análisis de parámetros
físico- químicos que afectan la población en medio natural del acocil
(Cambarellus montezumae) en Tezontepec de Aldama, Hidalgo. I
Reunión de Innovación acuícola y Pesquera Campeche. Nutrición
acuícola. 43 pp.
Montemayor, L. J., R. Maldonado y E. Guarneros. 2010. Influencia de la
alimentación sobre la reproducción y crecimiento del acocil regio
(Procambarus regiomontanus) especie en peligro de extinción. Ciencias
Universidad Autónoma de Nuevo León, Vol. XVII. No. 3. 276-286 pp.
Morones, B. F. 1991. Aspectos reproductivos bajo condiciones de laboratorio de
Cambarellus montezumae (Saussure) proveniente de la zona lacustre de
Xochimilco, D.F. Informe servicio social. Universidad Autónoma
Metropolitana -Xochimilco. 94 p.
Morrissy, N. M., 1989. A standard reference diet for crustacean nutrition
research. IV. Growth of freshwater crayfish Cherax tenuimanus. Journal
of the World Aquaculture Society, 20: 114-117 pp.
Nacif, O. Y. 2004. Evaluación del proceso de descomposición aeróbica de la
macrófita Egeria densa presente el hábitat de Cambarellus montezumae
y su potencial como alimento para esta especie. Tesis de Licenciatura.
Facultad de Ciencias. Universidad Nacional Autónoma de México. 63 p.
Novelo, R. A. y M. M. Gallegos. 1988. Estudio de Flora y vegetación acuática
relacionada con el sistema de chinampas en el sureste del Valle de
México. Biota 13: 121-139 pp.

194
Odum, H. T. 1987. Energy systems in ecology. In Systems and Control
Encyclopedia - Theory, Technology, Applications, M. G. Singh, ed.
Pergamon Press, New York. 1458-1462 pp.
Palacios, S., E. Espinoza, G. García y C. Pérez. 1995. Absorción de Metales
Pesados por la Asociación de Azolla lemna (Chilacastle-Chilacastle) en el
Parque Ecológico Xochimilco, D. F. Resúmenes del Primer Seminario
Internacional de Investigadores de Xochimilco. México. 82 p.
Palacios, G. A. 2003. Aspectos de reproducción, alimentación y crecimiento en
cautiverio de Cambarellus montezumae, Saussure 1857, (Crustacea:
Decapoda) de Xochimilco. Informe Servicio Social. Universidad
Autónoma Metropolitana - Iztapalapa. 44 p.
Palma, S. y P. Arana. 1997. Aspectos reproductivos del langostino colorado
(Pleuroncodes monodon H. Milne Edwards, 1837), frente a la costa de
Concepción, Chile. Invest. Mar., Valparaíso. 25: 203-221 pp.
Parada, C. 1987. Influencia de factores físicos en la variabilidad inter-diaria de
la distribución de estados tempranos de Engraulis ringens en el Golfo de
Arauco. Tesis de Magíster en Ciencias, Mención Oceanografía. Facultad
de Oceanografía. Universidad de Concepción. 98 p.
Pennak, R. W. 1978. Freshwater invertebrates of the United States. Ed. Jhon
Wiley and Sons. Nueva York. EUA. 803 p.

195
Plan Delegacional de Desarrollo Sustentable: DelegaciónXochimilco.1991.
Rescate de Xochimilco. Universidad Autónoma Metropolitana -
Xochimilco. 310 pp.
Plan Nacional de Desarrollo. 2007-2012. Se presenta, en cumplimiento al
Artículo 26 de la Constitución Política de los Estados Unidos Mexicanos
y se ha elaborado de acuerdo a lo establecido en la Ley de Planeación.
319 p.
Rangel, M. R. 1981. Determinación de Algunos Metales Tóxicos, Boro, Cadmio,
Cobre, Plomo y Zinc en Aguas del Lago de Xochimilco Para Uso agrícola.
Tesis Profesional. Escuela Nacional de Ciencias Biológicas. Instituto
Politécnico Nacional. México. 89 p.
Rangel, R. 2004. Estudio poblacional de Cambarellus montezumae en la Pista
de Remo y Canotaje “Virgilio Uribe”, Xochimilco; México, D.F. Tesis de
Licenciatura. Facultad de Ciencias. Universidad Nacional Autónoma de
México. 50 p.
Rangel, R. 2009. Ecología poblacional de Cambarellus montezumae en
Xochimilco, México, D. F. Tesis de Maestría en Ciencias Biológicas.
Facultad de Ciencias. Universidad Nacional Autónoma de México. 72 p.
Reid, D. & S. Corey. 1991. Comparative fecundity of decapod crustaceans II.
The fecundity of fifteen species of Anomuran and Brachyuran crabs.
Crustaceana. 61: 175-189 pp.
Ricker, W. E. 1975. Computation and interpretation of biological statistics of
fish population. Bull. Fish. Res. Bd. Can., 191: 382 pp.

196
Rojas, P. Y. R. 2003. Revisión taxonómica de las especies de Cambarellus
(Cambarellus) (Crustacea: Decapoda: Astacidae: Cambaridae): análisis
morfológico. Tesis de Maestría. Instituto de Biología. Universidad
Nacional Autónoma de México. 102 p.
Rosas, M. M. 1976. Datos biológicos sobre el acocil del lago de Pátzcuaro (C.
montezumae patzcuarensis). Memorias del Simposio sobre pesca en
aguas continentales. Tuxtla Gutiérrez, Chiapas. 40-63 pp.
Rodríguez-Serna, M., 1991. Influencia de la dieta en la eficiencia de
asimilación: Pérdidas de energía por respiración y productos
nitrogenados en el balance energético del acocil Cambarellus
montezumae (Saussure) (Crustacea: Cambaridae). Tesina de
Licenciatura.Universidad Autónoma Metropolitana - Xochimilco. 43 p.
Rodríguez, S. M. 1999. Biología y sistemática de los Cambaridos del sudeste de
México y su potencial aprovechamiento en la acuacultura. Tesis de
Doctorado en Ciencias Biológicas. Universidad Nacional Autónoma de
México. 118 p.
Rodríguez, S. M. y Carmona, O. C. 2002. Balance energético del acocil
Cambarellus montezumae (Saussure) (Crustacea: Astacidae: Cambaride)
pérdida de energía en la tasa metabólica. Universidad Juárez Autónoma
de Tabasco, Villahermosa, México. Universidad y Ciencia 18:36. 182-134
pp.

197
Roy, E. 1985. El cangrejo de río, biología y nutrición. Acribia, S. A. Zaragoza
España. 185 p.
Saborido-Rey F. 2005. Review of population structure and ecology of S.
mentella in the Irminger Sea and adjacent waters. Reviews in Fish
Biology and Fisheries. 04-61 pp.
Salazar, I. 1999. Estudio sobre el efecto toxico y genotóxico sobre el acocil
Cambarellus montezumae. Tesis de Licenciatura. Facultad de Estudios
Superiores Cuautitlan. Universidad Nacional Autónoma de México. 50
p.
Sánchez, M. P. 2007. Aprovechamiento de los ambientes reducidos en los
canales de Xochimilco para el cultivo del acocil Cambarellus
montezumae, para consumo humano. Tesis de Maestría en Ciencias
Agropecuarias. Universidad Autónoma Metropolitana - Xochimilco. 44 p.
Santiago, M. C. 2001. Evaluación de la tasa de crecimiento de Cambarellus
montezumae (Saussure 1858) (Crustacea: Decapoda) a diferentes
temperaturas bajo condiciones de laboratorio y algunos aspectos de su
ciclo biológico. Informe Servicio Social. Universidad Autónoma
Metropolitana - Xochimilco. 50 p.
Santos, C. E. 1998. Importancia ecológica y socioeconómica del “acocil”
(Cambarellus montezumae Custacea: Decapoda), de dos embalses de
Tlaxcala. Informe Servicio Social. Universidad Autónoma Metropolitana
- Xochimilco. 45 p.

198
Santos, M. S., A. Celis y M. García-Rodríguez. 2011. Consumo de acocil
Cambarellus montezumae como fuente importante de macronutrientes.
Revista Latinoamericana de Química. 75 p.
Secretaría de Medio Ambiente y Recursos Naturales. Norma Oficial Mexicana
NOM – 059 – ECOL - 2001. Protección ambiental especies nativas de
México de flora y fauna silvestres-categorías de riesgo y especificaciones
para su inclusión, exclusión o cambio - lista de especies en riesgo.
Publicada en el Diario Oficial de la Federación el 6 de marzo de 2002.
153 p.
Tallamy, D. W. & C. W. Schaefer. 1997. Maternal care in the Hemiptera:
ancestry, alternatives, and current adaptive value, In: J.C. CHOE & B.J.
CRESPI (Eds). The evolution of social behavior in insects and arachnids.
Cambridge, Cambridge University Press. 94-115 pp.
Trivers, R. L. 1975. Natural selection of parental ability to vary the sex ratio of
offspring. Science, 179(68), 90-2 pp.
Valenzuela, B. A., G. Cifuentes y M. MAldonado. 2012. El aceite de pescado:
ayer un desecho industrial, hoy un producto de alto valor nutricional.
Revista Chilena de Nutrición 39:2. 201-209 pp.
Vazzoler, A. E. A. 1996. Biologia da reproduçao de peixes Teleósteos: Teoría e
práctica. EDUEM, Maringa, PR. 88 pp.
Villa, N. A. 2010. Aspectos preliminares de la dinámica poblacional del acocil
Cambarellus montezumae (Saussure) de Xochimilco. Tesis de

199
Licenciatura. Facultad de Ciencias. Universidad Nacional Autónoma de
México. 50 p.
Villarreal-Colmenares, H. 1991. A partial energy budget for the Australian
Crayfish Cherax tenuimanus. Journal of the World Aquaculture Society,
22:252-259 pp.
Villarreal, H. M., 1995. Evaluación del potencial de cultivo de la langosta de
agua dulce Australiana Cherax tenuimanusen función de su eficiencia
bioenergética. Memorias del 3er Simposium Internacional de Nutrición
Acuícola. Universidad Autónoma de Nuevo León, México. En prensa.
Vásquez, G. A. 2009. Aspectos sobre la reproducción y el crecimiento del acocil
Cambarellus (C.) montezumae (Saussure, 1857), en condiciones de
laboratorio. Tesis de Maestría en ciencias agropecuarias Universidad
Autónoma Metropolitana - Xochimilco. 101 pp.
Villalobos, F. A. 1955. Cambarinos de la Fauna Mexicana (Crustacea:
Decapoda). Tesis Doctorado. Facultad de Ciencias. Universidad Nacional
Autónoma de México. 290 p.
Washburn, B., B. Washburn, D. Frye & S. Hung. 1990. Dietary effects on tissue
composition, oogénesis and the reproductive performance of female
rainbow trout (Oncorhynchus mykiss). Aquaculture.90: 179-195 pp.
Zar, J. H. 1984. Biostatistical Analysis. Prentice-Hall. USA. 620 pp.


















