3. Componente Neuromuscular
26
3. Componente Neuromuscular Editar 0 5 … Componente Neuromuscular: Músculos Masticatorios El tejido muscular es el órgano primario del cuerpo humano, constituyendo el 70-80% del peso corporal (exceptuando los casos de obesidad). Su función es contraerse para posibilitar la locomoción, constricción, bombeo y otros movimientos de propulsión en un organismo vivo. Características generales de los músculos: 1. Proporciona soporte y postura al esqueleto. 2. Es el mayor consumidor de energía en el cuerpo; sin el adecuado suplemento energético y la adecuada remoción de productos de desecho metabólico, sobreviene la fatiga y cese de función. 3. Los músculos no pueden empujar, expandirse o aumentar de tamaño activamente. Fijándose a un origen inmóvil y a una inserción móvil, solo pueden contraerse para producir un tirón (movimiento) o tensión (fuerza). 4. Son capaces de activarse por regiones y variar el grado de contracción según sea la acción a ejecutar. 5. Acompañan a otros músculos ya sea para estabilizar bajo tensión o servir de fijación durante el movimiento. 6. Su funcionamiento nunca es aislado o unilateral, sino como una estructura de vectores de fuerza que afectan a una estructura o sistema determinado, en nuestro caso, el SE. Según sus características, los músculos se clasifican en 3 tipos: 1. Músculo esquelético o estriado: es el músculo más abundante del cuerpo, está formado por fibras rodeadas de tejido conectivo, que se conectan a los huesos del esqueleto por medio de los tendones para producir movimiento. Reciben su nombre por su aspecto bajo el microscopio donde muestran estrías transversales. Su contracción es controlada voluntariamente por el sistema nervioso somático. Todos los músculos masticatorios son esqueléticos. 2. Músculo liso: reciben este nombre porque sus células no tienen estrías. Forma parte importante de las paredes de la mayoría de vasos sanguíneos y diversos órganos huecos o tubulares como los intestinos y vejiga. Su contracción es involuntaria y está regulada por el Sistema Nervioso Autónomo. 3. Músculo cardiaco: está constituido por células estriadas de control involuntario. Se encuentra en gran parte del corazón y mínimamente en las paredes de las venas pulmonares y cava superior. La musculatura esquelética se desarrolla a partir de los mioblastos, con una cantidad predeterminada de fibras musculares previamente establecidas
-
Upload
estrella-jc -
Category
Documents
-
view
423 -
download
3
Transcript of 3. Componente Neuromuscular

3. Componente Neuromuscular Editar 0 5…
Componente Neuromuscular: Músculos MasticatoriosEl tejido muscular es el órgano primario del cuerpo humano, constituyendo el 70-80% del peso corporal (exceptuando los casos de obesidad). Su función es contraerse para posibilitar la locomoción, constricción, bombeo y otros movimientos de propulsión en un organismo vivo. Características generales de los músculos:
1. Proporciona soporte y postura al esqueleto.2. Es el mayor consumidor de energía en el cuerpo; sin el adecuado suplemento energético y la adecuada
remoción de productos de desecho metabólico, sobreviene la fatiga y cese de función.3. Los músculos no pueden empujar, expandirse o aumentar de tamaño activamente. Fijándose a un origen
inmóvil y a una inserción móvil, solo pueden contraerse para producir un tirón (movimiento) o tensión (fuerza).
4. Son capaces de activarse por regiones y variar el grado de contracción según sea la acción a ejecutar.5. Acompañan a otros músculos ya sea para estabilizar bajo tensión o servir de fijación durante el movimiento.6. Su funcionamiento nunca es aislado o unilateral, sino como una estructura de vectores de fuerza que
afectan a una estructura o sistema determinado, en nuestro caso, el SE.
Según sus características, los músculos se clasifican en 3 tipos:
1. Músculo esquelético o estriado: es el músculo más abundante del cuerpo, está formado por fibras rodeadas de tejido conectivo, que se conectan a los huesos del esqueleto por medio de los tendones para producir movimiento. Reciben su nombre por su aspecto bajo el microscopio donde muestran estrías transversales. Su contracción es controlada voluntariamente por el sistema nervioso somático. Todos los músculos masticatorios son esqueléticos.
2. Músculo liso: reciben este nombre porque sus células no tienen estrías. Forma parte importante de las paredes de la mayoría de vasos sanguíneos y diversos órganos huecos o tubulares como los intestinos y vejiga. Su contracción es involuntaria y está regulada por el Sistema Nervioso Autónomo.
3. Músculo cardiaco: está constituido por células estriadas de control involuntario. Se encuentra en gran parte del corazón y mínimamente en las paredes de las venas pulmonares y cava superior.
La musculatura esquelética se desarrolla a partir de los mioblastos, con una cantidad predeterminada de fibras musculares previamente establecidas desde el nacimiento. A partir de ahí, cambios considerables toman lugar en cuanto a la cantidad y calidad de los músculos esqueléticos hasta la edad adulta: su diferenciación y crecimiento se produce por influencias neurales. A medida el músculo va creciendo en tamaño y fuerza, sus células aumentan en longitud y diámetro.Según su forma, los músculos esqueléticos pueden ser rectos (el origen y la inserción tienen el mismo ancho) o en abanico (el origen es más ancho que la inserción). Según sus características, los músculos esqueléticos pueden ser:
Simples: son músculos que se sobreponen a poca resistencia durante la contracción, por lo que no requieren producir altas tensiones durante función y sus líneas de acción no son complicadas porque tienen brazos de palanca favorables con respecto a la ATM. Sin embargo tienen que contraerse considerablemente para mover la mandíbula a distancias relativamente grandes. Se caracterizan por tener fibras que corren paralelas desde su origen a inserción, con un área transversal de volumen modesto excepto su zona media que puede ser un poco más gruesa. Actúan como correas que producen movimientos exclusivos de traslación. Ejemplos son el Digástrico y pterigoideo externo.

Complejos: el tamaño y localización de sus inserciones es muy variable para permitir la activación general o regional de las fibras musculares, lo cual es muy conveniente en la ejecución y combinación de movimientos (lateralidad y protrusión) durante la masticación. Por las características de estos patrones de movimiento, la necesidad de producir tensión considerable durante función a menudo bajo condiciones casi isométricas y en espacios restringidos, es importante una arquitectura en penachos que permite rotación de sus fibras alrededor de su inserción, aumentando su ángulo a medida se acortan y trasladan al tendón en la dirección deseada. Si todos los haces se adhieren a un lado del tendón, se describe como unipenacho; si se adhieren a ambos lados es bipenacho y multipenacho cuando existen múltiples tendones centrales. Ejemplos son el masetero y pterigoideo medial.
En el sistema estomatognático podemos encontrar funcionando 3 grupos de músculos:
Músculos masticatorios (ver Fig. No.2.8): Los músculos masticatorios están encargados de controlar la posición y desplazamiento mandibular durante la masticación, habla o deglución, y producir fuerzas en grados y velocidades variables para triturar alimentos. Por su anatomía funcional especial y todo lo mencionado anteriormente, los músculos masticatorios son los más complejos y poderosos del cuerpo. Todos los músculos masticatorios trabajan simultáneamente ya que cuando unos están actuando de forma primaria, otros lo hacen de manera antagonista. Las consecuencias de sus acciones son pertinentes para la prótesis dental, ortodoncia y cirugía maxilofacial. Pueden ser de 2 tipos:
Elevadores: son los encargados del cierre mandibular durante la masticación y otras funciones relacionadas. Poseen fibras musculares cortas y gruesas, dispuestas en abundantes penachos, que les permite generar grandes cantidades de fuerza sin cambiar mucho su longitud durante la contracción muscular. Se consideran músculos complejos que tienen aponeurosis que se desplazan y deforman durante la función. Entre ellos tenemos al masetero, temporal y pterigoideo Interno.
Depresores: están encargados de la apertura mandibular durante la masticación y otras funciones relacionadas. Son músculos simples con fibras paralelas y delgadas, que pueden contraerse más pero sin producir mucha fuerza. Entre ellos tenemos al digástrico, los suprahioideos, y pterigoideo externo.
Musculos cervicales: estabilizadores del cráneo
Esternocleidomastoideo (ECM): rotar el cráneo. Trapecio: extender el cuello y fijar la base del cráneo. Escalenos: fijar o inclinar la columna vertebral.
Músculos accesorios:
Cutáneo del cuello: levanta la piel superior del tórax y ayuda a la apertura mandibular.

Buccinador: sonrisa, ubicación de los alimentos en las tablas oclusales durante masticación. Orbicular de los labios.
Se conoce como línea de acción muscular (ver Fig. No.2.9) a la dirección del movimiento generado por la contracción y es determinada por la geometría del músculo dentro del sistema. Desde un plano sagital y frontal se puede observar que los músculos elevadores tienen una línea de acción dirigida hacia arriba.
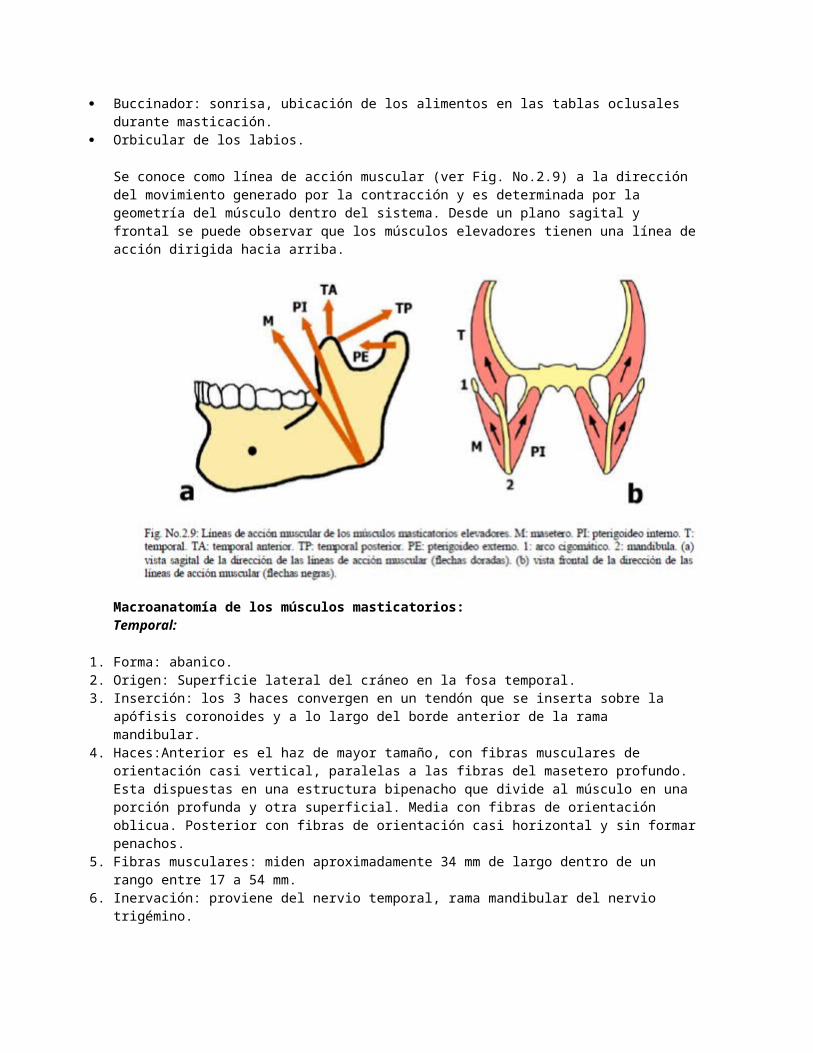
Macroanatomía de los músculos masticatorios:Temporal:
1. Forma: abanico.2. Origen: Superficie lateral del cráneo en la fosa temporal.3. Inserción: los 3 haces convergen en un tendón que se inserta sobre la apófisis coronoides y a lo largo del
borde anterior de la rama mandibular.4. Haces:Anterior es el haz de mayor tamaño, con fibras musculares de orientación casi vertical, paralelas a las
fibras del masetero profundo. Esta dispuestas en una estructura bipenacho que divide al músculo en una porción profunda y otra superficial. Media con fibras de orientación oblicua. Posterior con fibras de orientación casi horizontal y sin formar penachos.
5. Fibras musculares: miden aproximadamente 34 mm de largo dentro de un rango entre 17 a 54 mm.6. Inervación: proviene del nervio temporal, rama mandibular del nervio trigémino.7. Irrigación: proviene de las arterias temporal anterior, media y posterior.8. Función: es más posicionador y estabilizador que productor de fuerza, sirviendo de guía para los músculos
elevadores y ATM. Los 3 haces musculares son activos durante todos los movimientos mandibulares y vectorialmente, tienen la capacidad de funcionar en distintas líneas de acción: elevación (haz anterior) y retrusión (haz medio y posterior). El haz anterior es el área más prominente e importante del músculo porque desde un punto de vista mecánico, tiene una línea de acción vertical que le permite traccionar la mandíbula hacia arriba y limitadamente hacia medial, asentando el cóndilo firmemente contra el disco articular en la eminencia articular para mantener juntas las superficies articulares; por lo tanto, el músculo temporal es responsable de la posición más superior del ensamblaje cóndilo-disco dentro de la cavidad glenoidea. Los haces medio y posterior son activos en movimientos de lateralidad y durante el paso de protrusión a retrusión. En general, cuando participan en las lateralidades, son más activos en el lado de trabajo. Finalmente, tiene participación en los movimientos finos del habla.
Masetero:
1. Forma: rectangular.2. Origen: arco cigomático.3. Inserción: en la superficie lateral de la mandíbula, desde la región inferior a la 2 molar hasta el ángulo
mandibular.4. Haces: anatómicamente se reconocen un haz superficial, que es el más fuerte, con fibras delgadas, de
orientación oblicua en disposición multipenachos, y otro profundo. Funcionalmente se reconocen 4 sectores: superficial y profundo anterior, y superficial y profundo posterior.
5. Fibras: miden aproximadamente 26 mm de largo dentro de un rango de 14 a 38 mm, siendo las fibras más largas las que están en el sector superficial anterior (son 35% mas grandes con respecto a las posteriores). La diferencia de tamaño está en la capacidad del masetero de activarse diferencialmente dependiendo de la función. Cuando se contraen, las fibras se engrosan y aumentan sus diámetros transversales. El masetero no tiene fibras paralelas insertadas en ángulos fijos con respecto a sus aponeurosis como generalmente se asume, sino una estructura multipenacho que permiten activarse en grados diferentes, con vectores variables de fuerza que distribuyen la tensión irregularmente
6. Estructura: tiene 4 aponeurosis unidas individualmente al arco zigomático, cada uno de ellos con una orientación tridimensional particular y con haces multipenachos para un mayor poder de contracción.
7. Inervación: sensitivo por nervio auriculotemporal y plexo cervical superficial. Motor por nervio maseterino rama del nervio maxilar inferior.
8. Irrigación: ramas superficiales de la arteria transversal de la cara, arteria facial y arteria maseterina rama de la maxilar interna.
9. Función: está encargado de la elevación y cierre mandibular, proporcionando fuerza durante la masticación y posicionamiento oclusal al final del ciclo masticatorio; es normal que este musculo tenga una alta actividad EMG. Colabora con la protrusión simple y lateralidades extremas. El haz superficial tiene fibras orientadas oblicuamente que se activan durante elevación, elevación con protrusión y movimientos en lado de balance. El haz profundo tiene una acción estabilizadora del cóndilo dentro de la cavidad glenoidea y aunque contribuye con la elevación, es más vigoroso hacia la retrusión. Neuromuscularmente hablando, el contacto oclusal posterior estimula la contracción del masetero para que genere fuerza triturante. El masetero es el musculo que sufre más por la actividad parafuncional, estrés emocional y puntos gatillo.
Pterigoideo Interno o Medial:
1. Forma: rectángulo pequeño y abultado con orientación diagonal.2. Origen: principalmente en la fosa pterigoidea, tuberosidad maxilar y proceso piramidal del hueso palatino.3. Inserción: Parte posterior e inferior de la superficie interna de la rama y ángulo mandibular donde termina
frente a las inserciones del masetero.4. Fibras: cortas de 16 mm aproximadamente con orientación oblicua.5. Estructura: densos penachos septados que generan máxima tensión dentro de su región anatómica
confinada, lo cual es necesario para mantener un balance entre espacio y tensión necesaria para la función.6. Inervación: Pterigoideo interno, rama del nervio maxilar inferior.7. Irrigación: Arteria pterigoidea, rama de la arteria facial.8. Función: Participa en la elevación, formando una especie de cabestrillo mandibular con el masetero. Aunque
produzca fuerza, su funcionamiento principal es durante los movimientos laterales de la mandíbula. Muy activo en la protrusión, colabora con los movimientos de rotación mandibular. Cuando se combinan movimientos protrusivos y laterales, su actividad domina sobre el temporal.
Pterigoideo Externo o Lateral:
1. Forma: cono.2. Origen: El haz superior se origina en el ala mayor del esfenoides mientras que el inferior proviene de la placa
pterigoidea lateral.

3. Inserción: ambos haces se insertan en la fóvea pterigoidea del cóndilo a la altura de su cuello con excepción de una parte del haz superior, donde un aproximado del 20% de sus fibras se insertan sobre el disco y la cápsula articular.
4. Haces: hay un haz superior, que es pequeño y delgado. Aproximadamente el 22% de las fibras del pterigoideo externo se insertan en el disco, siendo la mayoría del haz superior. Cuanto menos sea la superficie de inserción del pterigoideo externo al cóndilo, más difícil será que se sobreponga a desplazamientos discales. Y hay un haz Inferior el cual es más grande y más grueso que el haz superior, cuyas fibras miden aproximadamente 22 mm. Las fibras están adecuadas mas para contracciones isotónicas que isométricas y tienen una línea de acción casi paralela con su masa muscular.
5. Inervación: nervio pterigoideo externo, rama del temporobucal, rama del maxilar inferior.6. Irrigación: arteria pterigoidea rama de la maxilar interna.7. Función: El pterigoideo externo no funciona solo y como esta envuelto en casi todos los movimientos
mandibulares, cuesta definir sus verdaderas funciones. Se ha sugerido que en general, el pterigoideo lateral es un musculo de soporte para la estabilización del ensamblaje cóndilo-disco. Se ha sugerido que ambos haces del pterigoideo externo son independientes y de funciones antagonistas. El haz superior es activo durante cierre, retrusión, máxima intercuspidación y movimientos en lado de trabajo. El haz inferior es activo durante la apertura, protrusión y movimientos de lado de balance; además asiste durante la apertura mandibular a los músculos suprahioideos para mover el cóndilo hacia abajo y adelante sobre la eminencia articular. Durante el cierre mandibular, el haz inferior se relaja mientras que el haz superior se contrae para posicionar y estabilizar el disco articular contra la eminencia articular. La contracción simultanea de ambos músculos lleva hacia adelante el disco articular en protrusión mandibular; la contracción aislada de un músculo produce los movimientos de lateralidad. Murray confirma las funciones del haz inferior pero no acredita las del haz superior o la acción reciproca entre haces. De hecho, pareciese que ambos haces desempeñan la misma función como una unidad con cantidades variables de actividad finamente graduadas y una distribución acorde a las demandas biomecánicas de la tarea.
8. Particularidades: el juego de los 2 haces del pterigoideo externo permiten una relación constante entre disco y cóndilo. Si este músculo sufre cambios patológicos, suceden respuestas anómalas en la relación disco-cóndilo como los ruidos articulares. El músculo no posee receptores de estiramiento lo cual explica porque se contractura asintomáticamente.
Suprahiodeos:El grupo muscular suprahioideos tienen en común su inserción en el hueso hioides el cual participa la apertura, balance craneal y de unión a la mandíbula.Genihiodeo: Músculo de forma alargada.
1. Origen: Apófisis geni inferiores2. Inserción: Superficie anterior del cuerpo del hioides.3. Inervación: nervio genihioideo rama del hipogloso mayor.4. Irrigación: arteria lingual y sublingual.5. Función: Elevador del hueso hioides y lengua. Si esta fijo al hioides, ayuda a deprimir y retruir la mandíbula.
Milohioideo: Músculo de forma aplanada. La unión del músculo derecho e izquierdo forma el piso anatómico de la boca.
1. Origen: Línea oblicua interna.2. Inserción: Parte anterior del hioides.3. Inervación: nervio milohioideo, rama del maxilar inferior.4. Irrigación: arteria submentoneana rama de la arteria facial.5. Función: Eleva el hioides, piso de la boca y la base de la lengua. Si esta fijo al hioides, en conjunto con el
genihioideo ayuda a deprimir y retruir la mandíbula.
Estilohioideo: Músculo delgado.
1. Origen: Borde posterior de la apófisis estiloides.

2. Inserción: Asta mayor del hueso hioides.3. Inervación: rama estilohioidea del nervio facial.4. Irrigación: ramas de la arteria auricular posterior.5. Función: Eleva el hioides y piso de la boca.
Digástrico: Músculo alargado en forma de arco que tiene 2 vientres: anterior y posterior, conectados por un tendón intermedio.
1. Origen: El vientre posterior se origina en la apófisis mastoides y el vientre anterior proviene de la fosita digástrica de la mandíbula.
2. Inserción: El vientre posterior se inserta en el hioides y el anterior en el borde inferior mandibular a nivel de la línea media. El tendón intermedio se inserta en el hueso hioides a través de las fibras de la fascia cervical externa.
3. Inervación: el vientre posterior esta inervado por una rama del nervio facial y el vientre anterior por una rama del nervio milohioideo de la división mandibular del trigémino.
4. Irrigación: vientre anterior por arteria submentoneana, rama de la arteria facial y posterior por ramas de la arteria occipital y articular posterior.
5. Función: es muy activo durante la apertura, estabilizando y regulando la posición del hioides. Ambos vientres pueden tener una acción conjunta o separada; por ejemplo, en conjunto elevan al hueso hioides y parte de la lengua. Muestra una actividad moderada o marcada durante protrusión, retrusión, lateralidades y al final de la depresión mandibular. Son activos durante la parafunción y deglución.
Infrahioideos:
Esternotirohioideo: Se origina en el manubrio esternal y primer cartílago costal y se inserta en el cartílago tiroides.
Tirohioideo: Músculo plano y corto. Parece continuar hacia el hueso hioides el trayecto del esternohioideo. Se origina el cartílago tiroides para insertarse en el asta mayor del hueso hioides.
Esternocleidohioideo: Se origina en la clavícula, manubrio esternal y en el primer cartílago costal, para insertarse en el cuerpo del hueso hioides.
Omohioideo: Tiene 2 vientres y un tendón intermedio. El vientre posterior se origina en el omoplato para terminar en el tendón intermedio. El vientre anterior que se desprende de este se dirige hacia el hueso hioides.
La inervación de los músculos infrahioideos proviene del asa descendente del hipogloso. La irrigación es provista por ramas de la arteria tiroidea superior.Función: descienden el hioides y la laringe. Si fijan el hioides, permiten la acción de los suprahioideos para deprimir la mandíbula. Tienen una relación con los suprahioideos a través del hioides.Los músculos supra e infrahioideos son antagónicos de los músculos masticatorios y de los posteriores del cuello, y sinérgicos de los músculos profundos anteriores del cuello. La estática de la cabeza depende del balance de los supra e infrahioideos.Músculos del cuelloSon muchos los músculos del cuello pero solo nos dedicaremos a los más importantes:
1. Trapecio: Musculo ancho, plano y triangular con una apariencia de “colgador de abrigos”, que cubre la superficie lateral y posterior del cuello. El origen de este musculo se encuentra en la superficie externa del hueso occipital (a nivel de la línea nucal superior) y en la línea media posterior de las regiones cervicales y torácicas. A partir de ahí, se forman 3 haces de fibra que se insertan en las siguientes localizaciones para realizar sus funciones: el haz superior se inserta en la cara anterior y borde posterior de la clavícula, actuando de forma unilateral para que sus fibras superiores elevan el hombro, inclinan el cuello y la cabeza lateralmente hacia el mismo lado que se contrae y ayudan en la rotación extrema de la cabeza hacia el lado opuesto. Es sinergista del esternocleidomastoideo y antagonista con el elevador de la escapula. Actuando de forma bilateral, las fibras superiores extienden la cabeza y el cuello pero solo contra resistencia. El haz medio se inserta en el acromion del omoplato para retraer la escapula (la mueven hacia la línea media),

asistiendo a la flexión y abducción del brazo en el hombro y ayudando a rotar la escapula. El haz inferior se inserta en la mitad final de la espina del omoplato para retraer la escapula y rotar la fosa glenoidea hacia arriba, deprimiendo el borde vertebral de la escapula y asistiendo a la flexiona y abducción del brazo. En general, el trapecio eleva, desciende y retrae la escapula y eleva la clavícula. La inervación está dada por la porción espinal del nervio accesorio espinal. La irrigación está asegurada por la arteria dorsal de la escápula. Rama de la arteria subclavia, penetra en el músculo a nivel de la rama Terminal del nervio accesorio. Presentan dolor referido en el área occipital y temporal detrás de los ojos, y detrás del oído.
2. Esternocleidomastoideo (ECM): Musculo robusto de la cara lateral del cuello que se origina en el manubrio esternal y 1/3 medio de la clavícula, atravesando en sentido diagonal la región anterolateral del cuello por debajo del musculo cutáneo para insertarse en la apófisis mastoides del hueso temporal y línea nucal superior. Se divide en 2 fascículos: el esternal, más próximo a la superficie, es cilíndrico y unipenacho que se inserta en el manubrio del esternón y clavicular, aplanado y multipenacho que toma su inserción en la clavícula. En la zona media del músculo, se encuentra una zona en la que convergen multitud de nervios, el punto de Erb, de gran importancia quirúrgica.Cuando la contracción es bilateral, flexionan el cuello y llevan la cabeza hacia delante, acercando el mentón al tórax y controlan la hiperextensión del cuello. Son sinergistas con el musculo trapecio para estabilizar y fijar la posición de la cabeza en el espacio durante los movimientos mandibulares de la masticación y habla. Elevan la caja torácica durante la inspiración en sinergismo con los músculos escalenos. En contracción unilateral sinérgicamente con el trapecio homolateral, inclina la cabeza y el cuello hacia el mismo lado. También son sinérgicos para limitar el movimiento de inclinación contralateral de la cabeza y del cuello. El ECM es el musculo que resiste el efecto de latigazo durante un accidente. La irrigación está dada por las arterias esternocleidomastoideas superiores, medias y algunas veces inferiores y la inervación por el nervio espinal y el segundo cervical. El ECM siempre está activo en los pacientes bruxistas. Su dolor es referido al oído y frente de la cabeza (propio del fascículo clavicular).
3. Escaleno: Tiene una estructura triangular que se extiende de los procesos transversos de las vertebras cervicales a las 2 primeras costillas, existiendo 3 haces unipenacho: anterior, medio y posterior. La acción de los músculos escalenos anterior y medio es la elevación de la primera costilla y la rotación del cuello hacia el mismo lado, la acción del escaleno posterior es elevar la segunda costilla e inclinar el cuello hacia el lado opuesto. También actúan como músculos respiratorios accesorios en la inspiración, junto con el ECM. Los escalenos son importantes estabilizadores del cuello. Esta irrigado por la arteria escalenica e inervado por las ramas anteriores del 3, 4 , 5 y 6 nervios cervicales.
Microestructura del Músculo esqueléticoLas fibras musculares constituyen la unidad estructural del músculo esquelético. Son células multinucleadas, cilíndricas o en forma de huso, de tamaño variable según su localización en el cuerpo, sexo y edad del sujeto. Las fibras musculares tienen la característica principal de contraer por estimulación nerviosa para desarrollar tensión y movimiento. El número de fibras para un músculo esta preestablecido durante el nacimiento. Desde la infancia pasando por la adolescencia hasta llegar a edades adultas, cambios considerables toman lugar en cuanto a la calidad muscular: diámetro, longitud y fuerza. La masa muscular total y la fuerza va disminuyendo progresivamente después de la quinta década de vida, volviéndose más marcada en mujeres después de la menopausia y en hombres posterior a la sexta década de vida.Características de las fibras musculares de los músculos masticatorios:
1. Son inervadas por una sola terminación nerviosa.2. Están dispuestas en dirección paralela u oblicua a su eje de acción.3. Vistas longitudinales en el microscopio óptico muestran dos estructuras: las miofibrillas de disposición
longitudinal y las estriaciones, de disposición perpendicular en la fibra.4. Poseen una irrigación abundante por parte de vasos sanguíneos del tejido conectivo que las envuelve.
Comúnmente se asume que todas las fibras musculares tienen el mismo tamaño, corriendo paralelas entre si y el eje mayor del musculo desde su origen hasta inserción; esto es cierto en músculos potentes de otras regiones del cuerpo pero en el SE, donde el espacio es pequeño, hay músculos cortos con fibras de longitud

de aproximadamente 15-40 mm y diámetro de 10-80 m que aumenta durante la etapa de crecimiento o por efecto del ejercicio, que corren paralelas a la línea de acción del músculo, en ángulo u oblicuamente desde una aponeurosis.Las fibras musculares poseen una capacidad de regeneración limitada; en casos de lesiones que superen esta capacidad, la reparación será realizada por el tejido conjuntivo, dando origen a una fibrosis que en algunos casos influir en la función.Las fibras musculares usualmente están adheridas al hueso por medio de tendones y tienen aponeurosis internas a las que se adhieren las fibras musculares (ver Fig. No.2.10).
Cada fibra muscular está recubierta por una matriz delgada de tejido conectivo llamada endomisio. Grupos de 10 a 100 fibras están unidos en fascículos rodeados por otra capa que es el perimisio, cuyos extremos están fijados a los tendones. Todo el musculo está rodeado por el epimisio. Las capas de tejido conectivo son necesarias por varias razones:
Proveen un punto fijo de unión para ejercer tracción y generar libre movimiento. Unir fibras entre sí, reduciendo la fricción entre ellas. Su red capilar es responsable del aporte de oxígeno y nutrientes.
Estructura de las fibras musculares:Las fibras musculares están rodeadas de una membrana celular o sarcolema que funciona como una barrera altamente selectiva, conveniente para mantener el potencial de reposo. Dentro del sarcolema, se encuentra el sarcoplasma donde están suspendidos organélos como las mitocondrias, el sistema tubular, miofibrillas, iones calcio, potasio y magnesio, todos ellos participes importantes de la contracción muscular (ver Fig. No.2.11).El sistema tubular es un grupo de túbulos longitudinales y transversos, interconectados entre si para formar una red alrededor de cada miofibrilla. Se conoce como triada cuando 2 túbulos longitudinales engloban a uno transverso.

La función primordial del sistema tubular es la de regular la concentración de iones de calcio en las miofibrillas para permitir la interacción de filamentos durante la contracción por acción de la enzima ATPasa (Adenosintrifosfatasa). El calcio es almacenado y bombeado según necesidad desde el retículo sarcoplásmico.En el sarcoplasma de las fibras musculares, se encuentran las miofibrillas que a su vez están compuestas por 2 tipos de filamentos proteicos (Ver Fig. No.2.12):
Actina: cada miofibrilla contiene 3000 filamentos finos de actina. Su unidad básica es una proteína globular llamada actina G, que en conjunto conforman cadenas de actina F, las cuales forman espirales dobles en donde se adhieren proteínas como la tropomiosina en las hendiduras entre las cadenas de actina F y la troponina en un extremo de la molécula de tropomiosina.
Miosina: cada miofibrilla contiene 1500 filamentos gruesos de miosina. Está formada por dos unidades proteicas de distinto peso molecular unidas flexiblemente entre si para permitir movimiento: una cabeza (llamada meromiosina pesada) y una cola (llamada meromiosina ligera), asemejando a un palo de golf. La miosina de los músculos masticatorios es única con respecto a otros músculos y se conoce como “súper rápida” por su alta actividad de ATPasa que permite una contracción bastante rápida y fuerte.
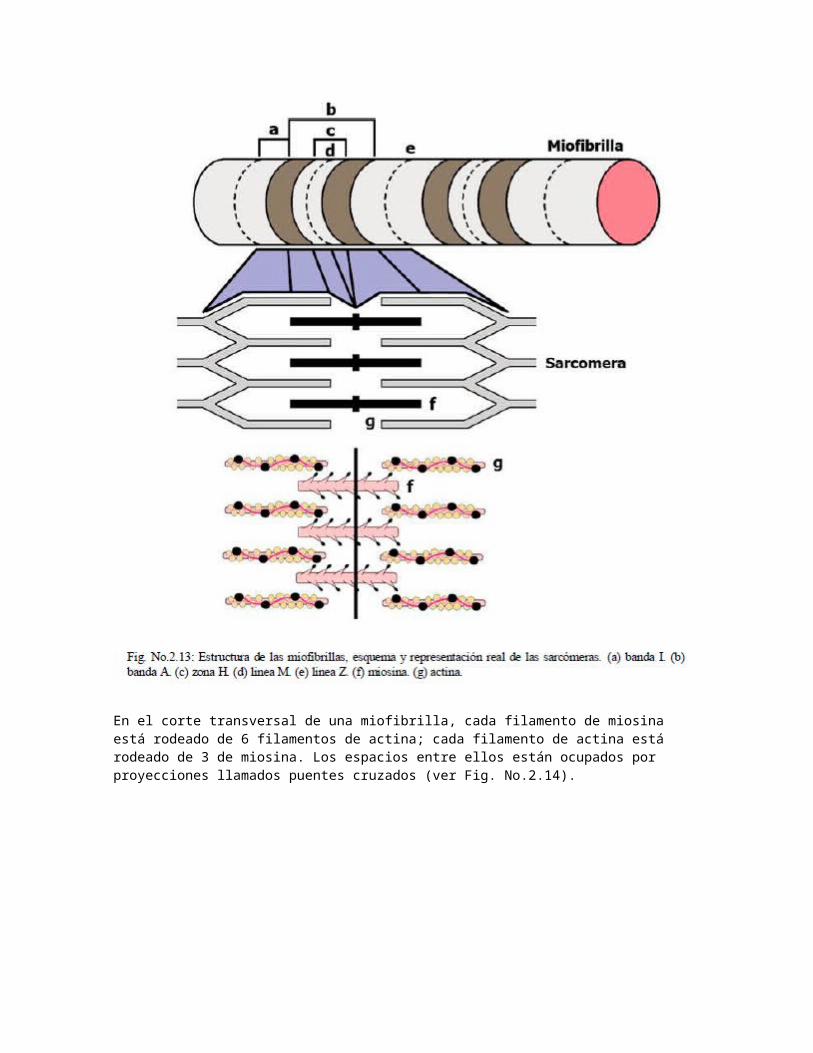
El 80% del volumen de las fibras musculares corresponde a las miofibrillas; la cantidad de miofibrillas dentro de una fibra pueden ser cientos o miles dependiendo de su grosor. La disposición de actina y miosina dentro de las miofibrillas conforman patrones repetitivos repartidos en segmento corto llamadas sarcómeras que constituyen la unidad funcional contráctil del musculo esquelético. Al observarse en un microscopio de luz polarizada, se distinguen las siguientes bandas:

Bandas I: de aspecto claro, contienen solo actina. Bandas A: de aspecto oscuro, contienen la misma superposición de actina y miosina. En la zona de
superposición, cada filamento de miosina está rodeado de 6 filamentos de actina. Dentro de la banda A, hay una porción de aspecto claro conocido como zona H dentro de la cual esta una línea al centro de la zona, la línea M que representa la unión entre filamentos.
Las sarcómeras miden de 2 a 3 µm, de los cuales la banda A representa 1.5 m y la banda I 1m en reposo. En máxima extensión mide 3m y en su mínima contracción 1.5 m. La longitud de la banda A es constante mientras que la banda I es variable: cortas durante la contracción, largas en relajación y muy largas en estiramiento. Las líneas Z son sitios de unión donde los filamentos de miosina se intercalan con otros filamentos de actina, dando el aspecto estriado característico del músculo esquelético. La porción de la fibra muscular contenida entre líneas Z conforman los límites de una sarcómera (ver Fig. No.2.13).
En el corte transversal de una miofibrilla, cada filamento de miosina está rodeado de 6 filamentos de actina;

cada filamento de actina está rodeado de 3 de miosina. Los espacios entre ellos están ocupados por proyecciones llamados puentes cruzados (ver Fig. No.2.14).
Tipos de fibras muscularesLos músculos están formados por fibras musculares que presentan distintas características. A grandes rasgos las podemos clasificar según:
1. Forma de acción: Tónica que son fibras de activación y relajación lenta, que inducen movimientos lentos y sostenidos o mantienen una posición por periodos largos de tiempo. Fásicas que son fibras de activación y relajación rápida. Se adaptan bien a movimientos rápidos de breve duración.
2. Color: Fibras rojas que se caracterizan por su diámetro reducido, abundancia de mioglobina, citocromos en sus numerosas mitocondrias, irrigación sanguínea más rica y líneas Z más gruesas. Están relacionadas con un comportamiento tónico. Fibras blancas que son más anchas pero contienen menos mioglobina y mitocondrias. Están relacionadas con un comportamiento fásico. Intermedias que presentan características intermedias respecto a las fibras rojas y blancas.Los músculos compuestos principalmente por fibras rojas pueden sostener una contracción durante periodos más largos que las blancas, debido a su disponibilidad continua de energía.
3. Estudios inmunohistoquimicos: I o posturales, caracterizadas por la presencia de una enzima llamada adenosintrifosfatasa. II o rápidas, caracterizadas por la abundancia de la enzima acido succínico deshidrogenasa. Las tipo II a su vez se pueden subdividir en A y B.
4. Fatiga: cuando un musculo se fatiga, pierde fuerza. Si la disminución es poca, se dice que la fibra es resistente a la fatiga y si la disminución es mucha, la fibra es fatigable. La resistencia a la fatiga tiene que ver con el sustrato energético que utilizan y la presencia de enzimas metabólicas aeróbicas/oxidativas o anaeróbicas/glicolíticas. Según lo anterior, las fibras se clasifican en O (oxidativa) porque utilizan oxigeno como sustrato energético procesado por vías aeróbicas/oxidativas. OG (oxidativa-glicolítica) porque tienen un comportamiento intermedio aeróbico/anaeróbico; G (glicolitica) porque utilizan glucógeno como sustrato energético procesado por vías anaeróbicas/glicolíticas.
5. Velocidad de contracción: definida por el tiempo que se tarda un musculo para producir una nueva fuerza nueva luego de haber producido una anterior. La velocidad de contracción depende de la cantidad de meromiosina pesada que poseen las fibras musculares ya que esta permite mayor actividad de la ATPasa para proveer energía para generar contracciones más rápidas y con mayor fuerza que fibras lentas. Sin embargo, aunque sean menos poderosas, las fibras lentas manejan más eficientemente la energía que las rápidas. De esta manera, las fibras pueden ser clasificadas en S (contracción lenta), FR (contracción rápida y resistente a la fatiga) y FF (contracción rápida y fatigable).
Combinando todas las clasificaciones, las fibras musculares pueden ser:
I-SO: contracción lenta, metabolismo oxidativo, resistente a la fatiga. Son de color rojo, contracción lenta y débil, abundantes mitocondrias y enzimas oxidativas que mantienen activo el metabolismo para reponer el

ATP de forma lenta pero continua, resistentes a la fatiga porque son capaces de mantener una cantidad constante de energía en la fibra muscular, pobres en glucógeno, en actividad de las fosforilasas y ATPasa y son comunes en músculos posturales como aquellos de la espalda y cuello.
IIB-FFG: contracción rapida, metabolismo glicolitico, fatigable. Son de color blanco, contracción extremadamente rápida debido a su miosina ATPasa rápida, fácilmente fatigables por su capacidad de ejercer fuerzas bastante grandes y son ricas en glucógeno y por lo tanto usan mecanismos anaeróbicos (o glicoliticos) para restaurar el almacenamiento de energía (ATP). La glicólisis es un proceso que genera energía rápidamente para producir grandes fuerzas pero por un período bastante corto de tiempo. Al no usar vías metabólicas aeróbicas, tienen pocas mitocondrias comparadas con las de tipos S. Consumen todas las reservas energéticas a corto plazo y su reemplazo es relativamente lento al igual que el retorno del poder contráctil del músculo. Alta de fosforilasa y de ATPasa pero pobre en actividad oxidativa.
IIA-FROG: contracción rapida, metabolismo oxidativo-glicolitico, resistente a la fatiga. Color rojo, características intermedias: contracción relativamente rápida y resistente a la fatiga, ejercen cantidades moderadas de fuerza aunque continuas por periodos prolongados, tienen una cantidad considerable de mitocondrias, metabolismo oxidativo-glucolítico y alta actividad oxidativa, glucolítica y de ATPasa.
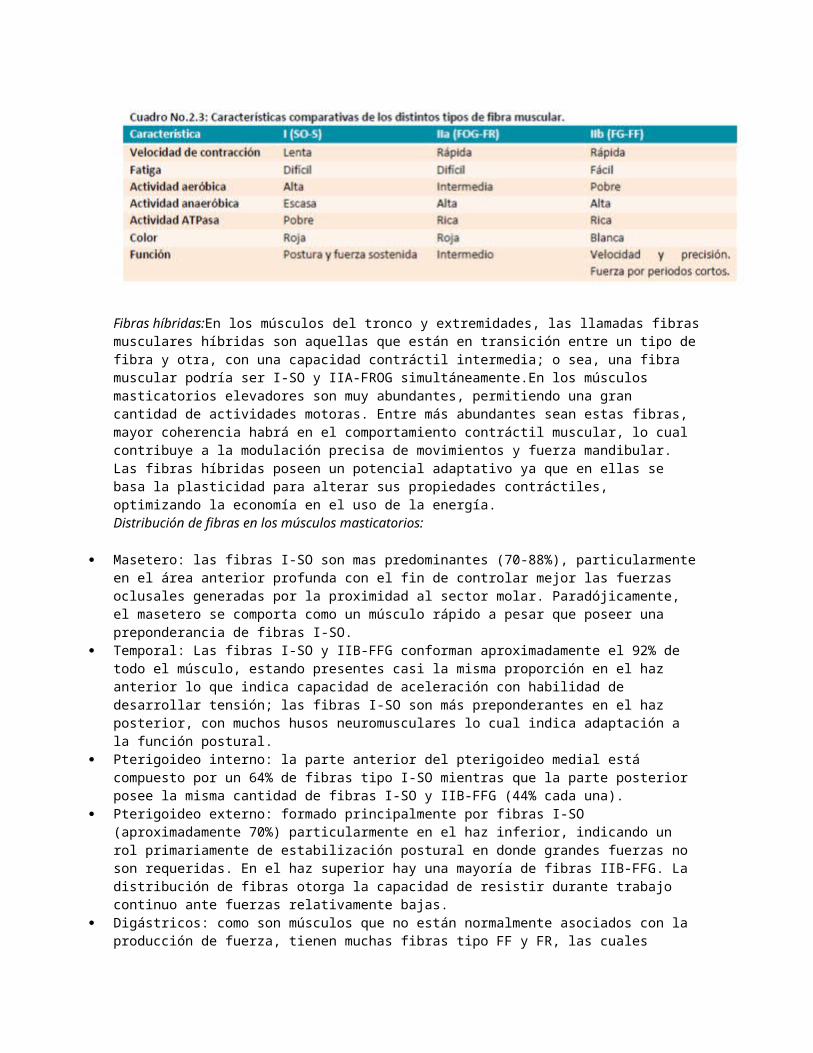
Una comparación entre los distintos tipos de fibra puede ser vista en el cuadro No.2.3.
Fibras híbridas:En los músculos del tronco y extremidades, las llamadas fibras musculares híbridas son aquellas que están en transición entre un tipo de fibra y otra, con una capacidad contráctil intermedia; o sea, una fibra muscular podría ser I-SO y IIA-FROG simultáneamente.En los músculos masticatorios elevadores son muy abundantes, permitiendo una gran cantidad de actividades motoras. Entre más abundantes sean estas fibras, mayor coherencia habrá en el comportamiento contráctil muscular, lo cual contribuye a la modulación precisa de movimientos y fuerza mandibular. Las fibras híbridas poseen un potencial adaptativo ya que en ellas se basa la plasticidad para alterar sus propiedades contráctiles, optimizando la economía en el uso de la energía.Distribución de fibras en los músculos masticatorios:
Masetero: las fibras I-SO son mas predominantes (70-88%), particularmente en el área anterior profunda con el fin de controlar mejor las fuerzas oclusales generadas por la proximidad al sector molar. Paradójicamente, el masetero se comporta como un músculo rápido a pesar que poseer una preponderancia de fibras I-SO.
Temporal: Las fibras I-SO y IIB-FFG conforman aproximadamente el 92% de todo el músculo, estando presentes casi la misma proporción en el haz anterior lo que indica capacidad de aceleración con habilidad de desarrollar tensión; las fibras I-SO son más preponderantes en el haz posterior, con muchos husos neuromusculares lo cual indica adaptación a la función postural.
Pterigoideo interno: la parte anterior del pterigoideo medial está compuesto por un 64% de fibras tipo I-SO mientras que la parte posterior posee la misma cantidad de fibras I-SO y IIB-FFG (44% cada una).

Pterigoideo externo: formado principalmente por fibras I-SO (aproximadamente 70%) particularmente en el haz inferior, indicando un rol primariamente de estabilización postural en donde grandes fuerzas no son requeridas. En el haz superior hay una mayoría de fibras IIB-FFG. La distribución de fibras otorga la capacidad de resistir durante trabajo continuo ante fuerzas relativamente bajas.
Digástricos: como son músculos que no están normalmente asociados con la producción de fuerza, tienen muchas fibras tipo FF y FR, las cuales confieren rapidez para los movimientos mandibulares durante el habla. Al ser embriológicamente diferentes a los músculos masticatorios, son iguales que los demás músculos del cuerpo y tienen una proporción similar de fibras I, IIA y IIB.
Cuando un músculo está compuesto por una mayoría de algún tipo de fibra, significa que esta optimizado para ejecutar una actividad en particular pero lo incapacita a ejecutar otro tipo de actividad. Pero en realidad, para poder realizar una gran variedad de movimientos a diferentes velocidades y duraciones, los músculos masticatorios tienen distintas proporciones de fibras con las que juegan para realizar la función: entre más variada sea la composición de fibras, más capacitado esta el músculo para realizar diferentes actividades motoras. En general, músculos poderosos poseen un alto contenido de fibras de contracción rápida mientras que los músculos posturales que deben de contraerse indefinidamente sin fatiga, contienen fibras mucho más lentas.La diferencia en la composición de fibras entre los músculos elevadores y depresores sugieren una diferencia en la cantidad de actividad diaria: la alta proporción de fibras lentas en los músculos elevadores indican que tienen una mayor función en el día al mantener la posición postural de reposo contra la gravedad.Reclutamiento de las fibras musculares:Para activar selectivamente los distintos tipos de fibra según sea la necesidad, solamente es necesario un sistema basado en el reclutamiento en orden jerarquico: al momento de la contracción muscular cuando se requiere de poca fuerza, son las fibras I-SO las que se activan primero ya que permiten ejercer fuerzas indefinidamente sin fatigarse. A medida que sea necesario aumentar la cantidad de fuerza, se van activando progresivamente más fibras musculares tipo I-SO. Si sucede un incremento de fuerza arriba del 20% del máximo posible, hace que la presión intramuscular sea mayor, disminuyendo la perfusión de sangre oxigenada e interfiriendo con el desempeño de las fibras I-SO que son netamente aeróbicas. Entonces se activan las fibras musculares IIA-FROG cuyas capacidades aeróbicas-anaeróbicas permiten al músculo ejercer fuerza aún cuando la perfusión sanguínea está limitada. Cuando la fuerza es realmente grande y la presión intramuscular impide el paso de sangre oxigenada casi en su totalidad, se termina reclutando fibras tipo IIB-FFG que se activan mecanismos alternos para producir energía como las vías anaeróbicas de producción de ATP; paralela a la activación de las vías anaeróbicas, sobreviene la acumulación de productos metabólicos secundarios a estas vías (acido láctico y pirúvico) que estimulan las terminaciones sensitivas. Entonces sobreviene el dolor y fatiga que obligan a la caída en la cantidad de fuerza y el cesamiento de la contracción. Un músculo completamente fatigado tiende a contracturarse en forma de espasmo, o sea que la tensión y el acortamiento del músculo se mantienen prolongadamente por la unión rígida de la actina y miosina dentro de la sarcómera. La transición entre la utilización de sustratos energéticos de naturaleza aeróbica hacia anaeróbica ocurre antes de los 120 segundos y es muy probable que desde los 60 segundos, los cambios ocurren de forma paulatina en la medida que las reservas de oxigeno van disminuyendo.Tono muscular:El tono es una actividad de contracción parcial y asincrónica de algunas unidades motoras para mantener activos los músculos posturales por largos periodos de tiempo con poca o ninguna fatiga, para soportar peso o posiciones contra la fuerza de gravedad y mantener una tensión muscular constante. El tono proporciona 2 cosas:
Firmeza y tensión constante. Resistencia a la gravedad, emociones y movimiento.
Es el SNC quien activa diferentes unidades motoras en distintos momentos dentro de un mismo grupo muscular para que la postura muscular sea transferida de una unidad motora a otra, en forma suave y continua, para mantener en alerta continua que mantiene al musculo listo para actuar y evitar la fatiga. Los músculos esqueléticos funcionan adecuadamente solo cuando la distancia normal se mantiene entre el
origen y la inserción de la fibra muscular. Los músculos pierden el tono y potencia cuando la longitud muscular se acorta y se incrementa si la longitud aumenta. No está claro si se debe a un reflejo monosináptico o a las propiedades viscoelásticas musculares.Contracción MuscularUnidad motora:El cerebro no controla las fibras musculares individualmente. En cambio, una sola motoneurona organiza y controla grupos de varias fibras por medio de las llamadas unidades motoras (ver Fig.No.2.15). Las unidades motoras varían de tamaño, manejando grupos desde 10 hasta más de 1000 fibras musculares. En sí, el número de fibras musculares por unidad motora gradúa la fuerza muscular y precisión de movimientos. Unidades motoras pequeñas con pocas fibras musculares bajo su comando realizan movimientos finos y de poca potencia como sucede en el globo ocular o el haz inferior del pterigoideo externo. Unidades motoras grandes con muchas fibras musculares realizan movimientos toscos pero de gran potencia como ocurre con los músculos masetero o temporal. Las grandes unidades motoras pueden generar hasta 200 veces más tensión que la que produce las unidades más pequeñas, siendo la tensión directamente proporcional a la cantidad de unidades motoras activadas.
Las unidades motoras de los músculos masticatorios son pequeñas y se caracterizan por la contracción muscular regional, con una acción muy focalizada, sin cruzar los límites de las aponeurosis internas musculares. Durante la función, las motoneuronas van reclutando unidades motoras secuencialmente según sea la tarea, comenzando por aquellas unidades motoras pequeñas con descargas lentas. Cerca de la mitad de las unidades son reclutadas al 10-20% de la contracción máxima voluntaria, siendo un proceso bastante rápido. Al igual que las fibras nerviosas, las fibras musculares son excitables por estímulos externos, permitiendo que el sarcolema se vuelva permeable a los iones sodio (proceso llamado despolarización), produciendo un potencial de acción, o sea un cambio en las cargas eléctricas propagada al interior de la fibra con una duración de unos cuantos milisegundos (ver Fig. No.2.16); las unidades motoras responden bajo el principio de “todo o nada”, es decir que ante un estimulo nervioso, todas se contraen sin existir la posibilidad de una contracción parcial.

Un potencial de acción transmitido por una motoneurona generará un potencial de acción simultáneamente en todas las fibras musculares conectadas a la unidad motora para contraer exactamente al mismo tiempo. Para ello, el axón de la motoneurona contacta con la superficie de la fibra muscular a través de un ramillete de terminaciones agrupadas que forman la placa neuromotora (ver. Fig. No.2.15).Proceso de contracción muscular:Contracción es la actividad ejercida por la fibra muscular para generar fuerza mecánica dentro del musculo. La fuerza ejercida sobre un objeto es denominada tensión y el objeto a su vez ejerce una fuerza conocida como carga. La tensión y la carga son fuerzas opuestas de manera tal que si se quiere mover el objeto, la tensión debe ser mayor a la carga.Básicamente, la contracción de las fibras musculares es un proceso complejo impulsado por ATP en el cual la actina y miosina dentro de las sarcómeras se entrecruzan, deslizándose entre si en respuesta a los "trenes" continuos de potenciales de acción provenientes del SNC. Cuando un estímulo es lo suficientemente fuerte para producir un potencial de acción, estimulara toda la fibra. No existe un impulso débil que produzca una respuesta débil: se produce el potencial o no.Una placa neuromotora comprende de una zona presináptica ubicada en la motoneurona la cual contiene acetilcolina y una zona postsináptica en la fibra muscular que posee receptores de membrana para fijar la acetilcolina. Un potencial de acción desencadena la liberación de acetilcolina la cual busca unirse con su receptor en la zona postsináptica para modificar la permeabilidad del sarcolema a los iones sodio y potasio. Cuando las cantidades de acetilcolina son elevadas, el sarcolema se despolariza con la subsecuente transmisión del potencial de acción para liberar iones calcio concentrados en los túbulos T del retículo sarcoplásmico (ver Fig. No.2.17). El aumento de la concentración de calcio desencadena la interacción de la actina y miosina de la sarcómera, con la consiguiente contracción muscular.

En la membrana postsináptica existe un sistema de degradación del neurotransmisor constituido por un grupo de enzimas llamadas acetilcolina-esterasa. La actuación de las enzimas determina la destrucción de la acetilcolina al cabo de unos pocos milisegundos, evitando la persistencia de su acción a nivel del receptor. Durante el estado de reposo, se cree que el complejo troponina-tropomiosina bloquea los sitios de inserción de las cabezas de miosina sobre el filamento de actina. La teoría más consistente al respecto sostiene que cuando se unen los iones calcio a la troponina, desaparece el bloqueo sobre la actina. Inmediatamente, las cabezas de miosina movidas por la energía provista por la ATPasa se mueven para liberarse de su unión con la actina, deslizándose a través de los puentes cruzados para unirse a un sitio más alejado de su inserción original en la actina; esto produce movimiento entre filamentos (ciclos repetidos de unión y desunión). La contracción continuará mientras existan iones calcio en concentraciones elevadas. En sí, la longitud de los filamentos no cambia durante la contracción sino que al deslizarse una sobre la otra, la sarcómera disminuye en longitud.Los cambios dimensionales que se producen durante la contracción muscular no son iguales en todos los músculos, inclusive no lo son en el interior de un mismo musculo. Por ejemplo, el musculo temporal puede sufrir un acortamiento de entre el 50-80% de su tamaño inicial, el masetero se acorta entre 3-54%, el pterigoideo externo entre 9.6-45% y el pterigoideo interno y digástrico al menos 10%.Una característica que puede afectar la contracción muscular es la frecuencia de aplicación de estímulos a la fibra. Una sucesión rápida de potenciales de acción a una fibra muscular es un fenómeno conocido como sumación, o sea, la adición de más ondas de contracción para logar un movimiento más fuerte, ya sea incrementando el número de unidades motoras que se contraen simultáneamente o incrementando la frecuencia de descargas de las unidades motoras individuales. La máxima respuesta que puede dar el musculo a una estimulación de alta frecuencia se llama tétano que ocurre cuando las contracciones sucesivas se fusionan y no puede distinguirse una de otra.Longitudes muscularesEn general, el musculo adopta 3 longitudes:
1. Reposo: corresponde a la longitud adoptada durante tono muscular.
2. Contracción: a partir de la longitud de reposo, cuando la fibra muscular es estimulada, esta responde de forma máxima porque la eficacia de la interacción entre actina y miosina está a su nivel máximo.
3. Estiramiento: cuando el músculo es estirado pasivamente más allá de su posición de reposo, genera un nivel de tensión proporcional a la cantidad estirada como consecuencia de su comportamiento elástico. Si la tensión es removida, el musculo regresara a su longitud de reposo gracias a sus propiedades viscoelásticas.
El comportamiento global del musculo depende de la integración de los comportamientos antes mencionados, o sea, cuando se contrae y a la vez es sometido a un cambio de longitud. Por encima o debajo de la longitud de reposo, la cantidad de tensión que genera una sarcómera no es la óptima, sobretodo si el cambio de longitud es muy grande, porque la capacidad de interacción entre la actina y miosina es menos efectiva. El máximo nivel de efectividad todavía es posible cuando el músculo es estirado hasta un 110-120 % respecto a su longitud de reposo, longitud en que la respuesta contráctil sigue siendo satisfactoria aunque se haya sobrepasado ligeramente la respuesta elástica.Si el musculo es estirado más allá de sus propiedades viscoelásticas, tendrá menor capacidad de retornar a su longitud de reposo y mayor tendencia al desgarro.Tipos de contracción muscular:
Isotónica: término que significan "misma fuerza", o sea, es una contracción o acortamiento muscular general bajo una tensión constante, estimulada por un gran número de unidades motoras contra una resistencia. Ocurre durante el habla o masticación, donde hay movimiento tratando de vencer la resistencia de la gravedad y bolo alimenticio respectivamente (ver Fig. No.2.18.b).
Isométrica: término que significa "mismo largo", o sea, desarrollo de tensión sin contracción o acortamiento muscular, que sucede por ejemplo cuando se apretan los dientes en máxima intercuspidación. Su función es el de sostener una posición o carga. La fuerza ejercida durante las contracciones isométricas puede ser bastante alta. Este tipo de contracción es evidente en pacientes bruxistas (ver fig. No.2.18.c). La tensión muscular que se genera en una contracción isométrica depende del recrutamiento y la sumación de fibras musculares.
Contracciones excéntricas o de alargamiento: los músculos pueden ejercer fuerza mientras su longitud aumenta controladamente.
A pesar que es académicamente conveniente el pensar sobre contracciones isométricas e isotónicas separadamente, la mayoría de los movimientos mandibulares es una mezcla de ambas. Con estos tipos de contracciones, los músculos de la cabeza y cuello mantienen constantemente una posición adecuada de la cabeza. Si se gira la cabeza a la derecha, algunos músculos deben ajustarse (contracción isotónica) y otros deben de estabilizar o mantener la posición (contracción isométrica).

Electromiografía:La electromiografía (EMG) es una técnica para grabar y exhibir los potenciales de acción de las fibras musculares, y que nos permite observar la actividad muscular en reposo o movimiento, condiciones de tonicidad muscular, etc. El electromiógrafo es un aparato eléctrico, muy parecido a un voltímetro, que recoge, amplifica y registra la actividad muscular. Su funcionamiento está basado en los potenciales de acción de cada fibra muscular, sumados espacial y temporalmente para formar lo que se conoce como unidad motora de potencial de acción (MUAP). La recolección de la actividad muscular es hecha a partir de censores o electrodos que graban poblaciones de MUAP cuya magnitud de energía es grabada en el rango de microvolts. Los electrodos recogen la actividad de las fibras musculares más superficiales; en la medida que las fibras estén localizadas a mayor profundidad, la señal captada será más pequeña por la cantidad de tejido corporal que deberá atravesar. Generalmente se colocan 3 electrodos: dos de grabación sobre el musculo a estudiar y uno de referencia en cualquier lugar del cuerpo. La señal a estudiar que es exhibida en un monitor, conocida como señal EMG, es el resultado de la comparación entre las señales de los electrodos de grabación y el de referencia para eliminar señales innecesarias de otros músculos, amplificación y filtración de ruidos ambientales que surgen durante la grabación. Finalmente, la señal EMG ampliada y filtrada, se muestra ya sea de forma no procesada (toda la señal) o procesada (convertida a una señal de fácil interpretación.Adaptación MuscularLos músculos son entidades versátiles capaces de cambiar, sus características anatómicas, fisiológicas, histoquímicas y bioquímicas para adaptarse a demandas funcionales alteradas y funcionar bajo condiciones ambientales cambiantes. La naturaleza dinámica del musculo le permite cambiar su fenotipo para optimizar su función de contracción y utilización de energía lo cual se manifiesta en la velocidad de contracción y fuerza máxima generada por el musculo. La respuesta adaptativa es individual e independiente para cada musculo y varía según la naturaleza del estímulo.Las adaptaciones incluyen:
1. Cambios en las características anatómicas de las fibras musculares en su tamaño y tipo, asociadas con cambios de intensidad, duración y frecuencia de la actividad muscular.

2. Modificaciones de sus propiedades contráctiles (velocidad de contracción y generación de fuerza) en respuesta a demandas funcionales variantes.
La sobrecarga y actividad muscular aumentada provoca transición hacia fibras lentas y resistentes a la fatiga, junto a un incremento del área transversal de la fibra muscular, tienen mayor contenido mitocondrial y mayor potencial oxidativo-aeróbico para producir energía, aumentando la cantidad de fuerza máxima que un músculo puede ejercer. La actividad muscular reducida provoca la transición hacia fibras rápidas y más fatigables. Este proceso que influye el comportamiento motor de la fibra se llama "plasticidad".