
Acuaporinas humanos
32
Acuaporinas Humanos : Los reguladores de flujo de agua transcelular abstracto Palabras clave: Regulación Aquaporin, Flujo de agua transcelular, Homeostasis, Regulación del volumen celular, Aumento de volumen de Regulación, Disminución del volumen de Regulación Antecedentes: evidencia emergente apoya la opinión de que (AQP) canales de agua de acuaporina son reguladores de transcelular flujo de agua. Consistente con su expresión en la mayoría de los tejidos, AQPs están asociados con diversa fisiológico y procesos fisiopatológicos. Ofreview Alcance: estudios knockout AQP sugieren que el papel regulador de AQPs, en lugar de su acción como pasiva canales, es su función crítica. Transporte a través de todos AQPs se produce por un mecanismo pasivo común, pero su regulación y distribución celular varía significativamente dependiendo de células y tejidos tipo; el papel de AQPs en la celda regulación del volumen (CVR) es particularmente notable. Esta revisión examina el papel regulador de AQPs en transcelular el flujo de agua, especialmente en CVR. Nos centramos en los sistemas principales del cuerpo humano, abarcando procesos tan diversos como concentración de la orina en el riñón con el aclaramiento del edema cerebral. Principales conclusiones: AQPs son cruciales para la regulación de la homeostasis del agua, proporcionando poros selectivos para el rápido movimiento del agua a través de las membranas celulares diversos y jugar un papel regulador en la CVR. Mecanismos de compuerta se han propuesto para AQPs humanos, pero sólo se han reportado para AQPs vegetales y microbianas. En consecuencia, Es probable que la distribución y abundancia de AQPs en una membrana particular es el factor determinante de la membrana permeabilidad al agua y un regulador de flujo de agua transcelular. Significado general: Dilucidar los mecanismos que regulan el flujo de agua transcelular mejorará nuestra comprensión del cuerpo humano en la salud y la enfermedad. El papel central de AQPs específicas en la regulación de la homeostasis del agua proporcionará rutas a una gama de nuevas terapias. Este artículo es parte de un número especial titulado acuaporinas. 1. Introducción: el flujo de agua transcelular: un papel regulador para el consumo humano 1.1. Las acuaporinas
-
Upload
alexa-luna -
Category
Health & Medicine
-
view
23 -
download
5
Transcript of Acuaporinas humanos

Acuaporinas Humanos : Los reguladores de flujo de agua transcelular
abstractoPalabras clave:Regulación Aquaporin, Flujo de agua transcelular, Homeostasis, Regulación del volumen celular, Aumento de volumen de Regulación, Disminución del volumen de RegulaciónAntecedentes: evidencia emergente apoya la opinión de que (AQP) canales de agua de acuaporina son reguladores de transcelular flujo de agua. Consistente con su expresión en la mayoría de los tejidos, AQPs están asociados con diversa fisiológico y procesos fisiopatológicos.Ofreview Alcance: estudios knockout AQP sugieren que el papel regulador de AQPs, en lugar de su acción como pasiva canales, es su función crítica. Transporte a través de todos AQPs se produce por un mecanismo pasivo común, pero su regulación y distribución celular varía significativamente dependiendo de células y tejidos tipo; el papel de AQPs en la celda regulación del volumen (CVR) es particularmente notable. Esta revisión examina el papel regulador de AQPs en transcelular el flujo de agua, especialmente en CVR. Nos centramos en los sistemas principales del cuerpo humano, abarcando procesos tan diversos como concentración de la orina en el riñón con el aclaramiento del edema cerebral. Principales conclusiones: AQPs son cruciales para la regulación de la homeostasis del agua, proporcionando poros selectivos para elrápido movimiento del agua a través de las membranas celulares diversos y jugar un papel regulador en la CVR. Mecanismos de compuerta se han propuesto para AQPs humanos, pero sólo se han reportado para AQPs vegetales y microbianas. En consecuencia, Es probable que la distribución y abundancia de AQPs en una membrana particular es el factor determinante de la membrana permeabilidad al agua y un regulador de flujo de agua transcelular. Significado general: Dilucidar los mecanismos que regulan el flujo de agua transcelular mejorará nuestra comprensión del cuerpo humano en la salud y la enfermedad. El papel central de AQPs específicas en la regulación de la homeostasis del agua proporcionará rutas a una gama de nuevas terapias. Este artículo es parte de un número especial titulado acuaporinas.
1. Introducción: el flujo de agua transcelular: un papel regulador para el consumo humano1.1. Las acuaporinasUna propiedad notable de la mayoría de los tejidos humanos es su capacidad para transporte de fluidos extremadamente rápida. Esto se ejemplifica por la concentración de orina en el riñón [1], la rápida formación de una cavidad llena de fluido adyacente para el ovocito durante la foliculogénesis de ovario [2] y la secreción de saliva de las glándulas salivales [3]. Estos procesos son esenciales para la salud humana y se basan en el transporte altamente regulado de agua a través de los tejidos [4,5]. Tal flujo de agua trans-tejido es posible por dos vías: transcelular el flujo de agua a través de ambos basal y las membranas apicales, que se produce enrespuesta a los estímulos osmóticos [4] creado por transporte de sal [6]; o flujo paracelular a través de las uniones célula-célula en los espacios intercelulares, impulsado por la sal o soluto gradientes [6]. Flujo de agua juega un paracelular papel importante en el epitelio fugas tal como el endotelio corneal [7]. En esta revisión, nos centramos en el papel regulador de acuaporina (AQP) canales de

agua en la mediación de flujo de agua a través de transcelular celular membranas en los principales sistemas del cuerpo humano (Fig. 1). Los estudio de los mecanismos de regulación AQP humano que median flujo de agua transcelular se encuentra todavía en su infancia y por lo tanto este revisión también discutirá AQPs mamíferos como posibles modelos para la regulación de AQPs humanos (Tabla 1). Flujo de agua transcelular depende de la permeabilidad de la membrana plasmática a las moléculas de agua. El movimiento del agua por ósmosis puede ser a través de la bicapa lipídica, por co-transporte pasivo con otro iones y solutos [8] o a través de canales de agua AQP [9]. Muchos AQP canales se cree que tienen una especificidad exquisita para el agua y son capaz de transportar rápidamente en respuesta a cambios en la tonicidad; la evidencia sugiere que hacen una contribución fundamental para la regulación del flujo de agua transcelular [10]. Desde la primera AQP fue identificado por Peter Agre en 1988 [11], trece AQPs humanos han sido descubiertos. Resultados estructurales para varios familiares miembros [12-14] han establecido que los canales AQP comparten una común arquitectura estructural . Junto con los estudios bioquímicos , estos datos estructurales [ 15 ] han puesto de manifiesto que la unidad funcional es un AQP homotetrámero [ 15 ] y que cada monómero AQP se compone de seis transmembrana (TM) α -hélices conectadas por intracelular alterna ( ICL ) y extracelular ( ECL ) bucles . Los dominios TM forman un diestro agrupar alrededor del poro central de cada monómero AQP través que se produce el transporte de agua [ 16 ] . La especificidad del poro para el agua es un resultado de la unión directa de hidrógeno , de una manera por pares , entre una solo archivo de moléculas de agua y la firma de la familia AQP Asn – ProAla adorno en la parte más estrecha del poro [ 16 ] . El agua es la selectividad aún más con la ayuda de las interacciones con la constricción aromático / arginina sitio , que restringe físicamente el poro [ 16 ] . Mientras que la biología estructural de la familia AQP está , por tanto, ampliamente aceptado , los mecanismos que regulan la función fisiológica de AQPs están menos bien establecido [ 17 ] .
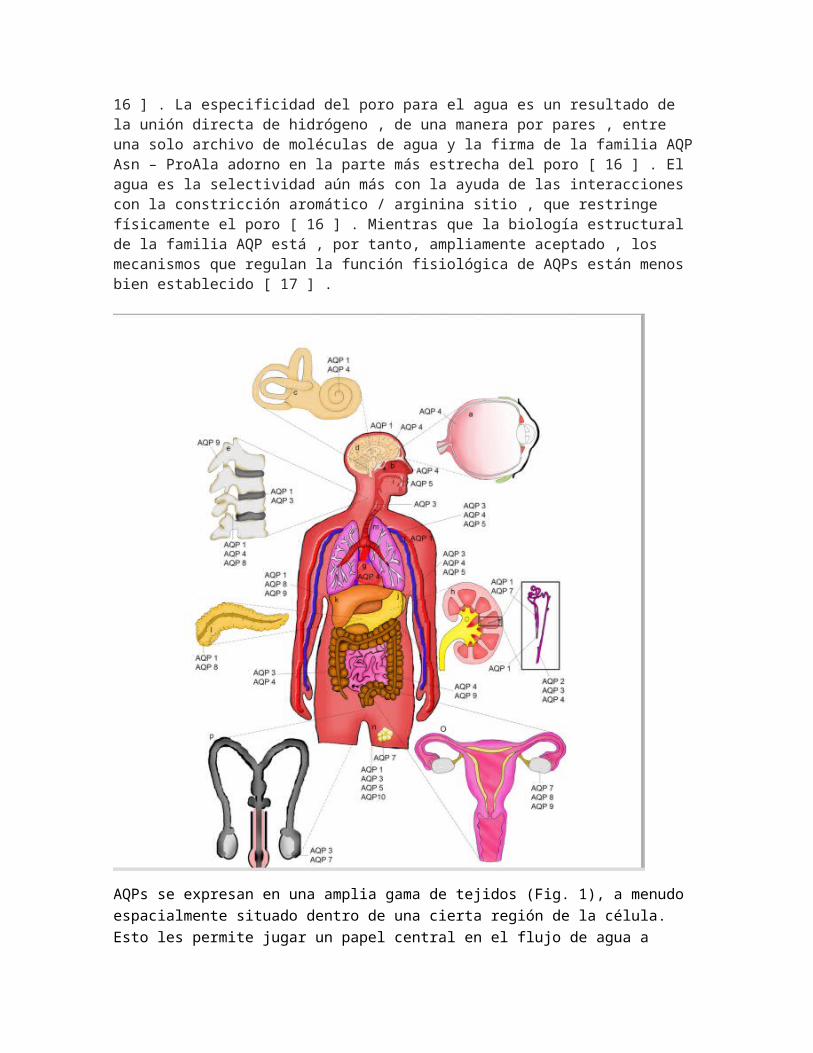
AQPs se expresan en una amplia gama de tejidos (Fig. 1), a menudo espacialmente situado dentro de una cierta región de la célula. Esto les permite jugar un papel central en el flujo de agua a través de los tejidos, que típicamente desencadena la regulación del volumen celular (CVR) mecanismos. Mientras que el transporte a través de todos AQPs utiliza un mecanismo pasivo común, su regulación y distribución celular varía significativamente entre sistemas, especificación camente en las células [17] y tejidos constituyentes. Esta revisión examina la regulación del flujo de agua transcelular en el cuerpo humano por AQP canales con un enfoque particular en la CVR. Flujo de agua
2. transcelular y la regulación del volumen celular
CVR es un componente necesario de la mecanicista transcelular mediada AQP el flujo de agua (Fig. 2). Se compone de disminución de volumen regulatorio (RVD), por lo general en respuesta a la inflamación celular inducida hipotonicidad-y aumento de volumen regulatorio (RVI), por lo general en respuesta a hypertonicityinduced encogimiento celular. Los mecanismos moleculares que subyacen a estos respuestas aún no se entienden completamente, pero es poco probable que

haya una único mecanismo común [18]. Las vías de señalización asociadas con CVR parecen ser dependientes del tipo de célula [18]. Sin embargo, el fin resultados de estas variadas vías son similares: RVD se basa en osmolito (Cloruro de potasio y taurina [19]) y flujo de salida de agua de la célula para reducir el volumen de la célula mientras que RVI se consigue osmolito y asociado afluencia de agua a través de la importación de sodio a la célula. Aunque la rápida RVI proceso siguiente encogimiento celular implica iones inorgánicos, después de horas de la exposición prolongada hipertónica, células animales a menudo reemplazan a los iones con no perturbar osmolitos orgánicos. Los mecanismos para esto incluyen transporte externo en la célula, la baja regulación de la degradación orgánica y sobre regulación de la síntesis orgánica [20]. Estos mecanismos facilitan la homeostasis de la osmolalidad dentro de la célula.
2.1. Disminución del volumen de Regulación
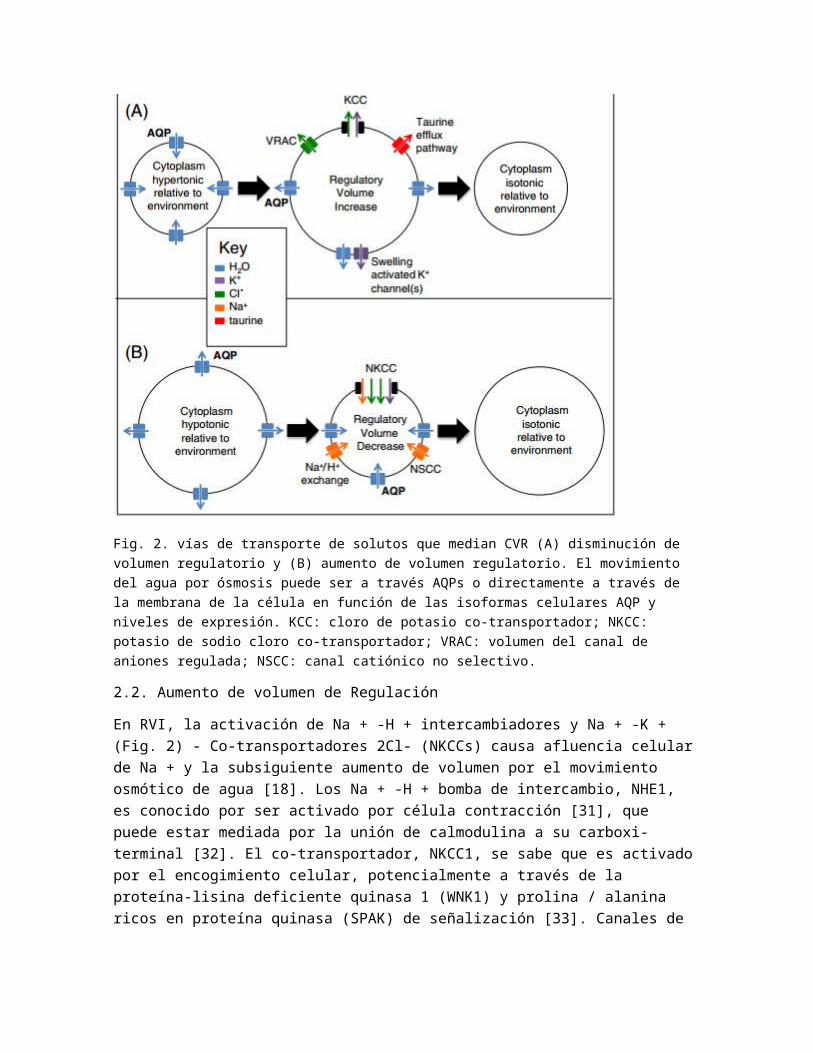
En RVD (Fig. 2), la activación de canales de K + permite el eflujo de K + de la célula y la posterior pérdida de agua por ósmosis ya sea a través AQPs o directamente a través de la bicapa lipídica; datos biofísicos muestran que Expresión AQP puede aumentar la permeabilidad al agua de la membrana hasta un ~ 50 veces [21,22]. Este flujo de salida de K + dependiente puede ser ya sea en intracelular la concentración de calcio [Ca2 +] (por ejemplo, en células de cáncer cervical humano [23]) o [Ca2 +] i independientes (por ejemplo, en células tumorales de ascitis de Ehrlich [24]). En la mayoría de los tipos celulares de un citoesqueleto de actina intacta es necesaria para hypotonicity inducida por K + eflujo. Sin embargo, en ganglio trigémino neuronas, citocalasina D (un inhibidor de la polimerización de actina) de tratamiento estimulado hinchazón activación de una corriente de K + [25] lo que demuestra que en estas células de una red de actina intacta no sólo es innecesario para RVD, sino que también parece tener efectos inhibidores. En algunas células, la proteína quinasa C (PKC) activación también se ha demostrado que induce una corriente hacia el exterior a través de K + los mismos canales que se activan en RVD [26]. El movimiento de K + fuera de la célula se ve favorecida por el gradiente de concentración, pero para mantener el potencial de membrana electrostática, canal de aniones volumen regulado (s) (VRAC) se mueven simultáneamente aniones Cl- (principalmente durante RVD) fuera de la célula, probablemente activado por tirosina quinasas [27,28]. También es pensaron que cuatro K + -CL- compañeros de transportistas (KCCs), conocidos por ser activados por la inflamación celular, pueden estar involucrados [29,30].

Fig. 2. vías de transporte de solutos que median CVR (A) disminución de volumen regulatorio y (B) aumento de volumen regulatorio. El movimiento del agua por ósmosis puede ser a través AQPs o directamente a través de la membrana de la célula en función de las isoformas celulares AQP y niveles de expresión. KCC: cloro de potasio co-transportador; NKCC: potasio de sodio cloro co-transportador; VRAC: volumen del canal de aniones regulada; NSCC: canal catiónico no selectivo.
2.2. Aumento de volumen de Regulación
En RVI, la activación de Na + -H + intercambiadores y Na + -K + (Fig. 2) - Co-transportadores 2Cl- (NKCCs) causa afluencia celular de Na + y la subsiguiente aumento de volumen por el movimiento osmótico de agua [18]. Los Na + -H + bomba de intercambio, NHE1, es conocido por ser activado por célula contracción [31], que puede estar mediada por la unión de calmodulina a su carboxi-terminal [32]. El co-transportador, NKCC1, se sabe que es activado por el encogimiento celular, potencialmente a través de la proteína-lisina deficiente quinasa 1 (WNK1) y prolina / alanina ricos en proteína quinasa (SPAK) de señalización [33]. Canales de cationes no selectivos sensible a amilorida (NSCCs) también pueden desempeñar un papel [34].
2.3. El papel regulador de las acuaporinas
La permeabilidad al agua de las membranas celulares puede no ser la limitante de la velocidad factor en la CVR [35], pero cualquier cambio rápido en el volumen celular es probable involucrar AQPs. Por ejemplo, el receptor transitorio activados por estiramiento potencial tipo vaniloide 4 (TRPV4) es un canal de Ca2 + -biased NSCC que es activada por la inflamación de células [36] y se ha implicado en osmosensing [37]. En algunos tipos de células TRPV4 se ha demostrado que
proporcionar una señal de Ca2 + que se correlaciona con la activación de la K + y Cl- canales responsable para la disminución de la osmolalidad celular asociada con RVD [38,39]. En células de la glándula salival humanos y murinos, TRPV4 tiene una interacción funcional con AQP5: en las células knockout AQP5, indujo hipotonicidad-la afluencia de calcio a través de TRPV4 fue atenuada y posterior RVD fue abolida. Hipotonicidad también aumentó la expresión de la superficie celular de tanto TRPV4 y AQP5 y aumentaron su co-localización [40]. En otro ejemplo, el esperma de AQP3 - / - ratones no sufrió su proceso de RVD normal y los ratones mostraron una reducción en la fertilidad [41,42]. A la entrada en el tracto reproductor femenino, espermatozoides normalmente encontrar una disminución en la osmolalidad extracelular, que se piensa que es la señal que activa la motilidad de esperma [41]. Sin embargo, este hipotónica el estrés también causa inflamación celular que, si no se corrige por RVD, cables a la fertilización con problemas causados por la flexión excesiva de la cola de los espermatozoides dentro del útero [42]. Si AQP3 simplemente estaba actuando de forma pasiva como el agua canal, RVD no sería abolida en AQP3 - / - ratones, sino más bien la escala de tiempo en el que la célula alcanza el equilibrio osmótico sería aumentado. Una explicación para la reducción de la fertilidad y RVD alterado en AQP3 - / - ratones, por tanto, es que AQP3, ya sea solo o como parte de una complejo macromolecular, está implicada en la vía de señalización que activa RVD en el esperma. Cuando se expone a una solución extracelular hipotónico, renal cultivaron células del conducto colector cortical renal (CCD), que no lo hacen de forma endógena expresar AQP2, hincharse en proporción al cambio en la osmolalidad extracelular pero no exhiben RVD. Sin embargo, cuando se transfectan con AQP2, estas células muestran un RVD de aproximadamente 40%. Contracción está mediada por Ca2 + afluencia través TRPV4, que activa Ca2 + - canales dependientes de K + y Cl- y Ca2 + dependiente de la liberación de Ca2 + intracelular de las tiendas. En las células renales CCD expresando AQP2, hipotónica estrés provoca la translocación de TRPV4 a la membrana plasmática, que es ausente en las células-AQP2 negativo. Cuando TRPV4 se transloca a la superficie de la célula antes de la exposición hipotónica usando 4α-forbol 12,13- didecanoato, RVD se recuperó en las células-AQP2 null, demostrando que es no sólo la alta permeabilidad al agua de AQP2 que permite RVD. Sin embargo, no parece haber ninguna co-localización entre endógeno TRPV4 y AQP2 sobreexpresada en este sistema, ya sea antes o después del choque hipotónico, lo que indica un funcional en lugar de física interacción [43]. Estas observaciones sugieren que AQP2 (y por lo tanto posiblemente, otros miembros de la familia AQP) forma parte de un sensorial y vía de señalización que resulta en la translocación TRPV4, posiblemente a través de la detección de la osmolalidad extracelular. Tomados en conjunto, estos ejemplos apoyan la idea de una señalización o función sensorial para AQPs en los mecanismos de RVD. Hemos sido incapaces de encontrar ninguna evidencia de AQPs juegan papeles similares en RVI. Sin embargo, dada que una variedad de AQPs podría estar implicado en el mecanismo de RVD y que RVI y RVD comparten un mecanismo básico común que implica la movimiento de osmolitos para provocar un cambio de volumen, no sería sorprendente descubrir un vínculo entre RVI y AQPs que va más allá de un mecanismo de conducción de agua pasivo.
3. Regulación de flujo de agua transcelular en el cuerpo humano
3.1. El cerebro y el sistema nervioso
3.1.1. Localización de AQPs

AQPs se expresan ampliamente en el sistema nervioso central (CNS); de hecho, más estudios se han realizado sobre AQPs en el SNC de en AQPs en los sistemas nerviosos periférico y entéricos (para una detallada crítica en AQPs en el sistema nervioso ver Papadopoulos et al. [44]). Los principales AQPs que se encuentran en el SNC son (i) AQP4 en la glía [45] y neuronas [46], (ii) AQP1 en el epitelio del plexo coroideo (que forma el líquido cefalorraquídeo (LCR) de barrera -Brain) [47], de la raíz dorsal ganglios [48] y las neuronas esofágicas [49] y (iii) en el AQP9 sustancia negra [50]. Las células gliales no son células nerviosas pero son esenciales para regulación y homeostasis del sistema nervioso central y comprenden aproximadamente 90% de las células en el cerebro [51]. Los astrocitos son los más abundantes glial células dentro del cerebro [51] y altos niveles de AQP4 se expresan en sus membranas plasmáticas. La distribución de AQP4 es altamente polarizada a los pies terminales astrocíticos, que entran en contacto con los vasos sanguíneos asociados con las interfaces de fluido cerebro-sangre-cerebro y [81]. El astrocítica Por lo tanto, los pies de gama funcionan como parte de la barrera hematoencefálica (BBB) mecanismo de intercambio de agua; esto ha sido establecido para ser AQP4 dependiente ya que la deleción de AQP4 en ratones resultó en una disminución de 31% en la captación de agua cerebral medido por el método de masa húmeda / seca determinar el contenido de agua del cerebro [52].
3.1.2. Papel regulador de AQPs
Como la principal AQP del CNS, se espera que AQP4 para jugar un papel importante en la regulación del flujo de agua en el cerebro, la médula espinal [53,54] e intersticial líquido que rodea las neuronas, manteniendo así las concentraciones de K + requerido para neuroexcitación [46,55]. En apoyo de esto, cuando AQP4 fue silenciada en astrocitos de interferencia por ARN, la aparente coeficiente de difusión (que es una medida del movimiento del agua dentro de tejidos, determinados por resonancia magnética) se redujo por 50% en cerebro de rata [56]. Es en este contexto que la baja expresión AQP4 niveles han sido asociados con ataques epilépticos [55]. Hay evidencia de que AQP translocación se produce en los astrocitos, que puede ser responsable de la regulación de AQP abundancia en el plasma membrana [57]. Aunque AQP4 se expresa principalmente en el plasma membrana, su abundancia superficie se incrementa por vesículas de transporte de AQP4 en respuesta a estímulos hipotónicas en astrocitos corticales de rata cultivadas: una correlación positiva entre la movilidad y la densidad de vesículas en el AQP4 Se observó membrana plasmática [58]. Así como AQP4, la stretchactivated Canal TRPV4 también se expresa con fuerza en extremo astrocítica pies [59]. Se ha sugerido que se coexpresarse AQP4 y TRPV4 y formar un complejo molecular, interactuando en la membrana plasmática para controlar CVR en astrocitos. Cuando los astrocitos primarios de ratón fueron poner bajo estrés hipotónico, se siguió un aumento de Ca2 + intracelular por una respuesta RVD; este mecanismo falló en células deficientes en AQP4 y en las células donde se utilizó la interferencia de ARN para silenciar TRPV4, incluso si se transfectaron con AQP1 [59,60]. Esto sugiere que en RVD astrocitos pueden ser AQP4 / TRPV4 específico, aunque la hinchazón de AQP1 células que expresan en este estudio no fue tan marcada como para AQP4- las células que expresan. Por lo tanto, si el volumen total de células es la cantidad biofísico ser detectado durante la respuesta RVD, es posible que la pérdida de RVD simplemente refleja el hecho de que un umbral de volumen no se ha cumplido. Si hay una interacción AQP4 / TRPV4, puede ser similar a la interacción entre AQP2 y TRPV4 en las células renales [43], discutidos anteriormente. AQP4 se ha asociado en gran medida con la fisiopatología de cerebro edema [61] de los cuales hay dos tipos principales: edema vasogénico es la

acumulación de agua en el espacio extracelular, generalmente debido a deterioro de la función de la acreditación; edema citotóxico es la hinchazón de astrocitos, mientras que el mantenimiento de su integridad celular [44]. Movimiento de agua de la sangre a través de los endotelios en astrocitos está mediada por canales AQP4; reducido edema citotóxico se observó en AQP4 - / - ratones [62]. AQP4 también se piensa para mediar en la reabsorción del exceso de líquido en el edema cerebral vasogénico por el movimiento transcelular de fluido desde el espacio extracelular a los pies terminales astrocíticos al vascular y regiones ventriculares del cerebro a través de la endotelial y epitelial barreras. En apoyo de esto, AQP4 ratones nulos muestran un aumento en el cerebro acumulación de agua debido a la reducción de la transferencia de agua transcelular a la sangre o LCR [61]. AQP1 expresión está restringida a la región del plexo coroideo de la cerebro en condiciones normales [47], pero se expresa en microvascular endoteliales y astrocitos reactivos de tumores cerebrales en los que se piensa para jugar un papel en el desarrollo de edema vasogénico [63]. Transporte de agua osmóticamente inducida se redujo 5 veces en AQP1 nula ratones junto con una disminución de la presión intracraneal y una disminución del 25% en la producción de CSF [47]. El mismo grupo también investigó la expresión y localización de AQPs en médula espinal de ratón, un área que no ha sido el foco de muchos estudios. Se detectó expresión de AQP4 en el astrocytic pies de gama de la materia gris en el contacto con los capilares y la procesos gliales fibrosos de la materia blanca, de nuevo en torno a la vasos sanguíneos. La distribución de varios otros AQPs todo el la médula espinal sugiere un papel en el mantenimiento del balance hídrico a través transcelular flujo de agua; fluido se mueve rápidamente de la subaracnoideo espinal espacio en el espacio perivascular y en todo el intersticio en el canal central. Este movimiento rápido de agua puede estar mediada por AQP4 y AQP9 en los limitans glia y AQP8 toda la capa ependimaria en el canal central [54]. Además de su localización en los astrocitos, AQP4 también se expresa en la retina [64], el epitelio olfativo [65] y dentro de Claudio y Células de Henson del oído interno [66]. K + grandes flujos desencadenan una AQP4- mediada por afluencia de agua transcelular que sale a través de canales localizados AQP4 en la membrana basal de las células de Claudio [66]; Participación AQP4 en CVR puede ser un importante componente mecanicista de la señal acústica transducción desde AQP4 ratones nulos son completamente sordos, pero no tienen alteraciones en la morfología del oído interno [66].
3.1.3. Conclusiones
Flujo de agua transcelular dentro del cerebro se asocia en gran medida con la formación y el aclaramiento del edema; la distribución altamente polarizada de AQP4 a los pies de extremo de los astrocitos es clave en la absorción de el exceso de líquido vasogénico para su aprobación por los capilares en el torrente sanguíneo. Sin embargo, AQP4 puede tener un efecto aditivo sobre la acumulación de agua en el edema citotóxico tal como se resume en la Fig. 3. El posicionamiento de AQP1 a lo largo de la barrera epitelial de la plexo coroideo que contiene CSF- indica un papel en el suministro de agua para la CSF a través de una ruta transcelular. Ello También es evidente que AQPs juegan un papel importante en la señalización a lo largo órganos periféricos y sensoriales del sistema nervioso por su contribución a los mecanismos de RCV y mantenimiento concentración de K +. Comprensión estos mecanismos serán esenciales en los tratamientos en desarrollo para el edema cerebral tras un accidente cerebrovascular, trauma y meningitis.
3.2. El sistema renal

3.2.1. Localización de AQPs
El papel de AQPs en el riñón es uno de los más ampliamente estudiado áreas en la investigación AQP [67]. El riñón es el sitio principal de la vasopresina arginina - (AVP; también llamada hormona antidiurética o ADH) mediada homeostasis agua corporal; en respuesta a estímulos osmóticos, agua reabsorción se produce a través de canales AQP que permiten la concentración de orina. AQPs en el riñón exhiben una localización altamente polarizado y específica patrón (Fig. 1): AQP1 se expresa en las células del túbulo proximal (PT); AQP2 es muy prominente en el conducto colector renal (CD renal) células principales y se encuentra en la membrana plasmática apical [68] y en el sub-apical vesículas [1]; AQP3 y AQP4 se expresan en células renales en el CD membrana basolateral [69,70]. AQP1 es responsable de la mayoría del agua reabsorción en el riñón [71] mientras AQP2 bien melodías reabsorción de agua; se transloca dentro de las vesículas subapicales al apical membrana después de la activación de AVP, el aumento de la permeabilidad al agua de la membrana [1]. Una vez que el agua ha entrado en una célula de CD renal, AQP3 y AQP4 proporcionan rutas de salida de la sangre [72].
3.2.2. Papel regulador de AQPs
Translocación activado por AVP AQP2 / es inducida por una proteína bien estudiada quinasa A (PKA) dependiente de la vía [73]. A partir del reconocimiento de un estímulo osmótico, AVP se libera y, posteriormente, se une el G receptor acoplado a proteína (GPCR), receptor V2 de vasopresina (V2R), en la membrana basolateral de las células principales de CD renal. Esto resulta en G activación de la proteína de la PKA y la posterior fosforilación de AQP2 en Ser266 dentro de vesículas intracelulares; esto facilita la translocación de la vesícula a lo largo de las redes de microtúbulos a la membrana apical [74] (Fig. 3A, B). El papel fundamental de este mecanismo de la multa control del agua reabsorción se ejemplifica por su disfunción en la diabetes nefrogénica insípida; en esta enfermedad, mutaciones en el receptor de vasopresina 2 o en Sí AQP2 conduce a una disminución de la capacidad para concentrar la orina [75]. Se necesita un Ca2 + intracelular afluencia para generar la necesaria RVD la respuesta después de la exposición hipotónica; en el riñón distal, esto sólo se produce en presencia de AQP2 [76]. Una interacción funcional entre el canal de Ca2 + iones TRPV4 -permeable y AQP2 se ha demostrado que se produzca en condiciones hipotónicas en células renales de CD y es involucrado en RVD. En particular, el canal TRPV4 sólo se activa en las células que AQP2 expresa. Cuando TRPV4 es bloqueado por el rojo de rutenio, Ca2 + la entrada es abolida junto con RVD. Esto confirma AQP2 y TRPV4 como esenciales componentes de la respuesta celular inducida por RVD hypotonicity- [43]. También se ha demostrado que AQP2 puede regular CVR mediante la interacción 5b con tropomiosina (TM5b) y la alteración de la dinámica del citoesqueleto. Ser256 fosforilación induce una interacción AQP2-TM5b líder a la despolimerización de filamentos de actina; este mecanismo es reversible [77]. AQP2 también está regulada independientemente de AVP en respuesta a hipertónica condiciones. Exposición hipertónica (600 mOsm / kg) se demostró que aumentar significativamente la actividad del promotor de AQP2, independiente de AVP, en células MDCK que expresan AQP2 murino. El elemento sensible se sugirió a residir entre -6,1 y -4,3 kb 5 'acompañamiento región del gen AQP2 [78]. Hypotonicity aguda también se ha demostrado que inducir la translocación de AQP2 hacia la membrana plasmática en ausencia de AVP: membrana plasmática rápida y la acumulación de la red trans-Golgi de AQP2 en las células CD renales de rata, se mostró a ser dependiente de MAPK, P38 y ERK1 / 2 la actividad [79]. Los primeros informes sugirieron que AQP1 se expresa

constitutivamente en Las membranas celulares del PT en el riñón [80] Sin embargo, estudios recientes han AQP1 demostrado que se expresa tanto en el citoplasma y en la membrana de las células cultivadas y puede ser inducida a someterse rápida y reversible translocación a la membrana plasmática tras la estimulación hipotónica mediado por los canales TRP, calcio, PKC y microtúbulos [57,81]. AQP1 También se ha demostrado que someterse a la translocación al plasma membrana de cholangiocytes en respuesta a la activación de la secretina específica Receptores acoplados a proteína G [82]. Otros estudios han demostrado que el efecto diurético de la acetazolamida implica AQP1 disparada translocación mediante la promoción de AQP1 y miosina interacciones de cadena pesada causando localización AQP1 a las membranas celulares túbulo proximal seguido por la degradación mediada por ubiquitina AQP1 a través de la proteasoma [83]. Ubiquitinación AQP2 También se ha demostrado estar implicado en AQP2 expresión y localización [84]. La fosforilación de AQP1 ha sido demostrado estar involucrados en la translocación de AQP1 a la membrana plasmática en ovocitos [85].
Fig. 3. AQPs Humanos son los reguladores de caudal de agua transcelular. (A) el flujo de agua paracelular y transcelular flujo de agua mediada AQP en el riñón conducto colector. (B) Mecanismos de AQP regulación en el riñón conducto colector a través de la translocación disparado. (C) La participación de AQPs en el flujo de agua transcelular en el edema cerebral. Flechas azules, movimiento H2O; flechas negras, AQP translocación; caja verde, unión estrecha; AVP / V2R, arginina vasopresina / receptor de vasopresina 2 estimulada AQP2 translocación; ?, Mecanismo de translocación Desconocido, y la flecha azul discontinua: papel regulador de AQPs en CVR (para más detalles ver Fig. 2).
3.2.3. Conclusiones

AQPs renales se forman una red altamente organizado que facilita la el mantenimiento de la homeostasis del agua. La localización específica de AQPs dentro de las células renales de CD proporciona una vía transcelular para que el agua sea reabsorbido de la orina a través de canales AQP2 en células renales de CD y de vuelta a la sangre a través de AQP3 y AQP4 en las membranas basolateral. La regulación de la expresión AQP1 y translocación media transcelular el flujo de agua en las células PT [83].
3.3. Tejidos secretores Especializados
3.3.1. Localización y funciones reguladoras de AQPs
Tejidos secretores especializados se basan en transcelular AQP-dependiente el flujo de agua para facilitar su homeostasis de fluidos. En la glándula salival, AQP5 facilita el flujo de agua transcelular tanto acinares y salival parótida células [86]; las células salivales aisladas de AQP5 - / - ratones tenían dramáticamente reducida permeabilidad al agua de la membrana tras la exposición a condiciones hipertónicas o hipotónicas [3]. La localización de AQP5 en el membrana luminal es consistente con alta permeabilidad al agua de la membrana y su papel en el transporte de agua osmótica de las células acinares al lumen de la glándula utilizado en la producción de saliva [87,88]. La inmunohistoquímica de las glándulas salivales humanas (HSG) ha demostrado AQP3 que está presente en las membranas basolateral de mucosa y células serosas acinares [89], lo que indica un posible papel de AQP3 así como AQP5 en el flujo de agua transcelular durante la formación de la saliva [89]. En HSG, AQP5 translocación se demostró que se produzca en un microtubuledependent manera; elevación de la [Ca2 +] i por estimulación con thapsigargin (A Ca2 + -ATPasa) y un ionóforo de Ca2 + resultaron en AQP5 localización en la membrana plasmática, que fue inhibida por el pretratamiento con los inhibidores de microtúbulos, la colchicina y vinblastina [90]. GFP-etiquetados AQP5 humano se ha demostrado que trasladar al membrana plasmática de las células HSG a la estimulación con carbacol (Un tipo muscarínico 3 receptores (M3R) agonista) [91], pero poco se sabe acerca de los mecanismos moleculares implicados. AQP5 translocación a la membrana apical de las células de la glándula parótida rata se ha demostrado que se producen después de la estimulación de la acetilcolina con M3R (ACh) y elevación posterior [Ca2 +] i. Además, después de la incubación con un ionóforo de Ca2 + translocación se activó sin ACh estimulación [92]. Aumento de los niveles de mRNA y proteína AQP5 se observaron cuando se expusieron líneas de células acinares de rata submandibular a las condiciones hipotónicas que causan inflamación celular rápida y más eficiente RVD. EGTA quelación de intracelular y extracelular de Ca2 + no lo hizo afectar CVR en una línea de células acinares de rata; de hecho CVR se encontró que era K + - y Cl - dependiente, con activada por mitógeno ERK-activación de quinasa (MEK) y el importante papel-β aminoácidos, taurina, jugando [93]. El aumento de la [Ca2 +] i se ha demostrado para desencadenar translocación AQP5 de los compartimientos citosólicos a la membrana plasmática, mientras que la eliminación de Ca2 + extracelular se ha demostrado que inhiben la translocación de AQP1 en células HEK293 [57]. Las células acinares de los ratones que carecen de la Canal TRPV4 o pantalla reducida Ca2 inducida hipotonicidad-AQP5 + afluencia y una respuesta RVD suprimido lo que sugiere un papel importante para TRPV4 y AQP5 interacciones en la generación de la respuesta de Ca2 + requerido para RVD efectiva después de estrés celular hipotónica [40]. Hay poco conocimientos sobre la regulación de AQP5 en las glándulas sudoríparas. Sudor ecrino glándulas tienen una alta permeabilidad al agua para apoyar su papel en el líquido secreción; sudor primaria se deposita en el lumen de la ecrino glándula antes de los

movimientos del agua permiten que el sudor para ser secretada a través la región ductal [94]. Recientemente, AQP5 se ha demostrado que se expresa en las membranas apicales, canalículos intracelulares de bobinas secretoras y en membrana basolateral de las glándulas sudoríparas ecrinas humanos. Apical Rápida translocación de AQP5 humana se produce en las células MDCK transfectadas de forma estable- después del tratamiento con el ionóforo de Ca2 +, A23187, lo que sugiere la participación de [Ca2 +] i en la translocación AQP5. Rápida translocación de AQP5 a la membrana plasmática apical de las células también se ha demostrado en ratón sudor glándulas durante la sudoración [95]. En los seres humanos, la activación ACh de los receptores muscarínicos aumentos [Ca2 +] i y induce sudar [94]. Es pensó que la regulación AQP5 por [Ca2 +] i contribuye a sudar liberación mediante el aumento de permeabilidad de la membrana apical, y que AQP5 puede colocalise con el canal de Ca2 +, ANO1, en la membrana plasmática [95]. Sin embargo, los mecanismos detrás de la translocación dentro de la AQP5 sudar glándula, incluyendo el papel de la fosforilación, todavía requieren más elucidación.
3.3.2. Conclusiones
AQP5 está implicado en la regulación de flujo de agua transcelular en el luminal regiones de glándulas. Esto facilita la formación de secreciones en el concentraciones y viscosidades necesarias requeridas para mantener el agua homeostasis.
3.4. El sistema tegumentario
3.4.1. Localización y funciones reguladoras de AQPs
El sistema tegumentario comprende la piel, el cabello y las uñas, pero nervios, ciertas glándulas y grasa a menudo también se clasifican como partes de la tegumento. La piel juega un papel fundamental en la homeostasis del agua y proporciona una función de barrera contra la pérdida excesiva de agua. Su agua y contenido de glicerol es esencial para la función normal; esto es en gran parte bajo el control de AQP3, que se expresa principalmente en el plasma membranas de la estrato basal de la epidermis, con la disminución expresión hacia el estrato granuloso y ninguno en el estrato corneum [96]. Sugiyama et al [97] la expresión primera reportado de AQPs 3 y 9 en queratinocitos cultivados humanos, aunque era AQP9 Sólo observado en la diferenciación de las células. Ratones de AQP3 nula muestran deterioro la función de barrera y reducida hidratación del estrato córneo, que era no corregido por la oclusión de la piel o la exposición a alta humedad. Sin embargo, la administración tópica u oral de glicerol ha demostrado que corregir muchos funciones de la piel defectuosas en-AQP3 nulo ratones [98]. AQP1 se ha detectado en biopsias de piel en un estudio del eccema atópico, aunque su expresión no era diferente en piel enferma o control, mientras que se encontró sobre regulación de AQP3 [99]. Enfermedades de la piel que expresión espectáculo reducido estrato córneo pantalla hidratación alterada niveles de AQP3, dependiendo de la enfermedad. En 2011, Voss y colegas mostraron que en la piel psoriásica, AQP3 fue preferencialmente expresa en el citoplasma en lugar de la membrana plasmática sugiere AQP3 que puede ser importante para el agua transcelular y glicerol transporte. AQP5 también se cree que está expresado en el plasma membrana en el estrato granuloso y puede desempeñar un papel en transcelular homeostasis del agua en la piel [100]. AQP7 es el transportador glicerol primaria en blanco (WAT) y marrón (MTD) tejidos adiposos. AQP7 se expresa abundantemente en el plasma membrana de los adipocitos y el ayuno se ha demostrado para arriba-regular ARNm AQP7 en los adipocitos de roedores [101]. En el modelo de ratón línea celular de los adipocitos, 3T3-L1,

aumenta en AQP7 mRNA expresión y liberación de glicerol se correlacionaron durante la diferenciación celular que indica que la permeabilidad de la membrana plasmática de glicerol puede mediar la acumulación de grasa en los adipocitos [102]. AQP3 y 9 también se expresan en tejido adiposo humano, con AQP3 localizada predominantemente en el citoplasma y AQP9 expresa constitutivamente en la membrana plasmática; una correlación positiva entre los niveles de transcripción de AQPs 3, 7 y 9 y el peso corporal (IMC) ha sugerido [103]. La expresión del gen AQP7 es el regulado en el WAT de los sujetos humanos obesos en comparación con los controles normales [104,105], pero sin cambios en la diabetes tipo 2 [104]. Sin embargo, el vínculo entre AQPs humanos y la obesidad es contradictoria; la revisión por Maeda et al. [106] describe algunos de estos estudios. Más AQP10 recientemente se ha propuesto ser una vía alternativa para eflujo de glicerol en los adipocitos humanos [107].
3.4.2. Conclusiones
AQP3, 7, 9 y 10 son aquaglyceroporins. Por tanto, es probable que la funciones de estos AQPs en el sistema tegumentario implican principalmente transporte de glicerol, aunque también pueden estar implicados en la regulación del flujo de agua transcelular; esto queda por determinar.
3.5. El sistema cardiovascular
3.5.1. Localización y funciones reguladoras de AQPs
AQPs cardíacos no han sido investigados en la misma medida como AQPs en el cerebro y el riñón. En el corazón, el agua se mueve desde el intersticialespacio, a través de los endotelios y en los vasos sanguíneos. Este proceso es típicamente atribuido al transporte de agua paracelular a través del endotelio de el corazón, ya que se considera que es "con fugas 'en comparación con el endotelio de otros órganos [108]. Aturdimiento miocárdico es la reducción de la producción de el corazón, a menudo visto después de la cirugía cardiaca como el bypass del corazón, y se ha asociado con la inflamación celular y edema. La expresión de miembros de la familia AQP en el miocardio ha sido mal caracterizado y su papel, en su caso, en la patología y la resolución de hinchazón de los cardiomiocitos, edema y función general dentro de la corazón no se ha investigado (para una revisión detallada y AQPs la gestión del agua infarto ver Egan [109]). AQP1 se ha detectado en el tejido humano, rata y ratón corazón [110], mientras que AQP4 sólo se ha detectado en el corazón de ratón [110]. La inmunofluorescencia imágenes revelaron que expresa abundantemente el- AQP1 se distribuye en la microvasculatura cardiaca humana y murina en altos niveles sólo a causa de la densa vascularización del músculo del corazón [110]. Inmunofluorescencia también mostró que AQP4 se localizó en las membranas plasmáticas de los cardiomiocitos del tejido del corazón de ratón. Los estudios funcionales en vesículas de membrana cardíacos desde AQP1 y AQP4 ratones knockout encontró que sólo AQP1 tuvo un papel en la permeabilidad al agua en el corazón; vesículas de ratones-AQP1 nula habían reducido la permeabilidad pero la supresión de AQP4 produjo ninguna reducción en la permeabilidad al agua [110]. Un estudio utilizando cardiomiocitos ventriculares de conejo sugirieron que el movimiento del agua en el corazón está mediada por el flujo de agua paracelular y no se produce a través de AQPs [111]. En las raras ocasiones que un osmótica gradiente está presente en el corazón (por ejemplo, durante celular hinchazón después cardiaca cirugía), AQP1 endotelial podría mediar en el flujo de agua desde la expandido espacio intersticial en los capilares [112]; de hecho es AQP1 la principal AQP expresado en células endoteliales vasculares [113]. Existe controversia sobre los

detalles de la expresión AQP4 en el corazón humano; algunos estudios han informado de la expresión de ARNm pero no o muy poca proteína [110]. Sin embargo, un estudio reciente [114] demostró la presencia de proteína AQP4 en el corazón humano utilizando Western secante; se localiza en la membrana de plasma con un muy débil señal en el citosol. Los análisis de inmunotransferencia de cardiomiocitos de ratón cultivadas también mostró que AQP4 estaba presente mientras inmuno-oro Imágenes de microscopía electrónica mostró proteína AQP4 en la membrana plasmática de los cardiomiocitos. Edema cardíaco surge cuando el tejido con una suministro de sangre reducido (isquémico) se convierte en hipertónica, haciendo que el agua fluir de los capilares (posiblemente a través de AQP1) en cardiomiocitos; esta provoca celular hinchazón y disminución del gasto cardíaco. Ratones isquémica tenía una disminuir en AQP4 ARNm, tal como se mide por RT-PCR. Ratones knockout AQP4 se presentó con lesión isquémica cardiaca menor, medida como el infarto, lo que sugiere que AQP4 puede ser una posible diana para el tratamiento de infarto miocardio.
3.5.2. Conclusiones
La principal AQP del sistema cardiovascular es AQP1 (Fig. 1), que probablemente regula la permeabilidad al agua de las redes de capilares del corazón mediando el flujo de agua a través de la capa endotelial en el sangre. AQP1 puede ayudar a la absorción del exceso de agua del intersticial espacio en los capilares, sin embargo, esto es controvertido. La investigación adicional por lo tanto, se requiere sobre el papel de AQPs en el corazón; esto podría proporcionar evidencia de AQP1 y / o AQP4 como dianas de medicamentos para el tratamiento de infarto de edema impresionante dentro del intersticio. AQP4 solamente tiene recientemente se ha detectado a nivel de proteínas dentro de los cardiomiocitos humanos, que podría conducir a la investigación en AQPs como transcelular transportadores de agua para el despacho y la formación de edema intersticial y cardiomiocitos hinchazón.
3.6. La vía aérea
3.6.1. Localización de AQPs
Hidratación de las vías respiratorias, secreciones sub-mucosas y transporte de fluidos alveolar todos requieren permeabilidad al agua de la epitelial y endotelial membranas de la vía aérea [115]. AQPs se expresan en consecuencia tejidos broncopulmonares (Fig. 1) y se regulan de una manera que facilita transporte de agua transcelular [116,117]. AQP1 se expresa predominantemente en los endotelios microvasculares en todo el pulmón y la parte superior de las vías respiratorias [117], mientras que AQP3 y AQP4 están presentes en el membranas basolateral de revestimiento epitelial de las vías respiratorias "[88]. AQP5 es expresado en la membrana apical de tipo I las células alveolares epiteliales de el pulmón distal y acinar células glandulares sub-epitelial, que proporciona La vía principal (junto con AQP1) para el flujo osmótico de agua dentro de todo el sistema de las vías respiratorias [118]. AQP5 se expresa también epitelio superficial de los bronquios y, como tal, es el lugar ideal para regular la hidratación de las superficies de las vías respiratorias [119]. De hecho, estudios con ratones knockout han mostrado que aumenta la secreción de fluido AQP5 [120]. Además, la expresión de AQP5 se reduce en las vías respiratorias inflamatoria condiciones, tales como la enfermedad pulmonar obstructiva crónica (EPOC), que se asocian con moco hipersecreción [119]. La modulación de AQP5 Por lo tanto, la expresión podría servir como terapia para las vías respiratorias inflamatoria condiciones,

aliviando los síntomas tales como el aumento de desecado moco que potencia las infecciones crónicas de las vías respiratorias típicas de quística fibrosis [121].
3.6.2. Papel regulador de AQPs
Cuando la expresión AQP dentro de la vía aérea se descubrió primero, varios Se publicaron los estudios murinos knockout AQP que investigó el hipótesis de que el agua puede ser transportada desde el espacio aéreo alveolar a través de los compartimentos intersticiales y capilares a través de osmoticallydriven Transporte AQP. Inicialmente, la permeabilidad al agua osmótica (Pf) de la pulmones se midió en AQP1 y AQP4 ratones knockout; Pf se redujo por 10 veces en los endotelios de AQP1 ratones nulos que sugiere un transcelular ruta. Sin embargo, se concluyó que AQP1 no proporciona la ruta para la absorción de fluido alveolar y poco efecto se observó en Pf en el Estudios knockout AQP4 [122]. Un año más tarde, el mismo grupo llevó a cabo un estudio muy similar, esta vez utilizando ratones knockout AQP5; el espacio aéreo-capilar Pf se redujo en 10 veces en el knockout AQP5 y otro 2-3 veces cuando un doble golpe de gracia de AQP1 / AQP5 era utilizado. Los autores concluyeron que AQP5 y AQP1 son las principales rutas para el flujo de agua transcelular en la vía aérea con la función primordial de AQP5 ser el transporte de agua a través de la membrana plasmática apical de tipo I células alveolares epiteliales [123]. TRPV4 se demostró posteriormente para regular la abundancia AQP5 bajo células epiteliales condiciones hipotónicas en pulmón de ratón (MLE) [124]. Después 2 h en medio hipotónico, una disminución en la abundancia AQP5 en las células MLE fue observado; este resultado fue observable después de 30 minutos, pero no antes de 10 min de exposición. Esta disminución en la AQP5 se bloqueó en presencia de rojo de rutenio, que es un inhibidor TRPV4, y también cuando las células se cultivaron en medio libre de Ca2 +, incluso cuando la osmolalidad era reducido a 127 mOsM (que es hipotónica); estos datos apoyan un papel de Ca2 + extracelular en la regulación de la abundancia AQP5. RT-PCR resultados mostraron que los niveles de ARNm no se vieron afectados cuando los niveles de proteína disminuido. Cuando se añadió un inhibidor de lisosomal, una reducción en AQP5 los niveles de ARNm no se observó lo que sugiere que la proteína AQP5 es probablemente la degradación que experimenta. Los mismos resultados se observaron en TRPV4- expresar y control de células HEK-293 transfectadas con AQP5; un AQP5 reducción en respuesta a hypotonicity sólo se observó en células que expresan TRPV4, que también fue bloqueado por el rojo de rutenio. La regulación de permeabilidad de la membrana por AQP5 abundancia se concluyó que ser fuertemente controlado por la osmolalidad y mediada por TRPV4 [124]. Otra prueba de que la osmolaridad regula la expresión AQP5 era proporcionado por Hoffert et al [125], que mostró que hipertónica estrés induce la expresión de AQP5 en las células cultivadas en MLE hipertónica medio (500 mOsM). Niveles de proteína AQP5 aumentaron después de las 8 hy alcanzó su punto máximo 24 horas después de la exposición, de regresar a la línea de base después de 6 h en isotónica medio. Solamente los solutos relativamente impermeables afectados expresión AQP5, lo que sugiere que un gradiente osmótico entre una célula y su entorno está implicado en la regulación de la expresión AQP5. Esta expresión mecanismo se demostró para requerir la activación de la extracelular señal-quinasa regulada (ERK) y la vía desde varios ERK inhibidores bloquearon la expresión AQP5; sin embargo, sin un hipertónica estímulo, la expresión de AQP5 se inhibió y activadores de ERK no pudo inducir la expresión. En el mismo estudio, las ratas que habían recibido diariamente inyecciones intraperitoneales de solución salina hipertónica tuvieron un aumento de 2 veces en AQP5 expresión en el pulmón en comparación con las ratas de control, lo

que sugiere la relevancia fisiológica de los mecanismos de regulación AQP5 en vivo [125]. AQP5 expresión también se ha demostrado ser regulada por una cíclico AMP / proteína quinasa A (cAMP / PKA) dependiente de la vía. En las células MLE, Además del análogo de cAMP, CPT-cAMP permeable a las células, causó un 4- fold aumento de AQP5 mRNA y los niveles de proteína en un dependiente de la dosis- manera; aumento de la síntesis de proteínas se realiza la ablación por la adición de el inhibidor de PKA, H89. Los estudios de inmunofluorescencia utilizando confocal microscopía en células MLE después de un tratamiento de 24 h CPT-cAMP revelado AQP5 que se transloca a la membrana plasmática apical. Creciente niveles de cAMP endógeno mediante tratamiento con forskolina y la β- agonista adrenérgico, isoproterenol, también indujo expresión de la proteína AQP5. El efecto forskolina también fue visto a trabajar ex vivo en el pulmón murino tejido lo que sugiere que este mecanismo molecular dependiente de AMPc puede ocurrir in vivo [126]. Sidhaye et al [127] estudiaron los efectos de cAMP en la regulación de la distribución y abundancia AQP5. Usando AQP5- expresando células epiteliales de pulmón de ratón, la distribución de AQP5 era observado por inmunofluorescencia y biotinilación superficie. Después tratamiento a corto plazo de las células con cAMP, AQP5 se internaliza y había una abundancia reducida de AQP5 en la membrana; a largo plazo exposición a cAMP (aproximadamente 8 h) dio como resultado de mayor thanbaseline Abundancia AQP5 en la membrana apical que indica una regulación al alza AQP5 de expresión en la membrana. Después de corto plazo la exposición, AQP5 abundancia fue también disminuyó temporalmente seguido por un marcado aumento después de una exposición 8 h. Cuando se trataron células con el agonista, tetrabutaline-β adrenérgico, que se sabe que aumenta los niveles intracelulares de cAMP, resultados idénticos a los observados después de cAMP Además se observaron en cuanto a la distribución y la abundancia de proteínas. Se observaron los mismos efectos in vivo utilizando ratones inyectados por vía subcutánea con el agonista. Cuando el inhibidor de PKA, H89, era agregado, todos los efectos de cAMP dependiente se inhibieron, adicionalmente purificada AQP5 se fosforiló por PKA pero no PKC o caseína, lo que indica que Se requiere la activación de PKA para la regulación AQP5 en las vías respiratorias. El contraste de los resultados se obtuvieron a partir de un estudio en el que humanos células epiteliales bronquiales fueron transfectadas establemente con el tipo salvaje (WT) AQP5 y dos mutantes AQP5 [128]. La primera mutante era una alanina sustitución de Ser156, el sitio PKA sustrato para la fosforilación de tanto AQP2 y AQP5, que está implicado en AQP2 membrana apical translocación. La segunda mutación estaba dentro de la segunda Asn-ProAla motivo, que forma el poro AQP. Estos experimentos fueron diseñados para examinar la importancia de la formación de poros y la fosforilación PKA en la expresión en la membrana de AQP5. WT-AQP5 ha demostrado ser expresado en la membrana plasmática apical y en vesículas de sub-apical, mientras que el mutante Ser156Ala también mostró expresión en la membrana. Esta indicó que el bloqueo del sitio de unión PKA no afectó translocación AQP5 como en el caso de AQP2. La expresión de un mutante era Asn185Asp localizada en todo el citoplasma lo que sugiere que al igual que AQP1, AQP5 requiere el motivo Asn-Pro-Ala para la formación de canal correcto y que que puede tener un papel en el plegamiento de proteínas, ya sea o oligomerización. Cuando el inhibidor de PKA, H89, se utilizó, no hay diferencia en la expresión membrana en comparación con el nivel basal fue visto después de 30 minutos, que también era el caso después de un tratamiento de 30 min cAMP en los tres tipos celulares estables. AQP2 translocación a la membrana apical fue inducida por la adición de cAMP y esto fue bloqueado por el tratamiento previo con H89. También cuando el AQP2 Ser156 se mutó, AQP2 se expresó en el citoplasma e incapaz de trasladar a la membrana. En

WT-AQP5 y Asn185Asp células mutantes, la fosforilación fue visto antes de la adición de cAMP. Sin embargo, en las células mutantes Ser156Ala, AQP5 nunca fue lo que indica que Ser156 fosforilada es el sustrato PKA en AQP5, que la localización de la membrana de la proteína no está regulado por PKA y que la estimulación de AMPc puede ser un evento separado en contraste con el Mecanismo AQP2 y los estudios previos realizados por otros grupos [128].
3.6.3. Conclusiones
La evidencia directa ha sido publicado por la participación de AQPs en flujo de agua transcelular a través del epitelio alveolar a través de un apical Ruta AQP5 y a través de AQP1 en el endotelio [118]. Este movimiento de agua entre el espacio aéreo y capilares alveolares es compartimentos esencial para la hidratación de la vía aérea, defensas eficaces vías respiratorias y reabsorción del exceso de líquido alveolar.
3.7. El sistema reproductivo
La permeabilidad de las membranas celulares a agua y hormonas en tanto los sistemas reproductores masculino y femenino es esencial para la foliculogénesis [2], la espermatogénesis [129] y el espermatozoide osmoadaptation [41]. Hay pocos estudios sobre la regulación de AQPs en el sistema reproductivo, pero perfiles de expresión se han esbozado y hasta a la fecha de revisión son disponible [41129130]. Nuevas evidencias sugieren funciones fisiológicas para AQPs dentro de los sistemas reproductivos humanos y que los cambios de agua son menudo transcelular y AQP mediada [2]; AQPs han sido recientemente asociado con la patogénesis de algunos trastornos reproductivos tales como el síndrome de ovario poliquístico (PCOS) [131] y el cáncer de ovario, donde AQP5 y AQP9 regulación se observaron en el tejido de tumores malignos de ovario [130].
3.7.1. El sistema reproductor femenino
El papel de AQPs en el ovario, específicamente el folículo ovárico, tiene ha estudiado bien. Durante la foliculogénesis, el antro es expandido por una gran afluencia rápida, de agua a través de la células de la granulosa (GC) de revestimiento; ello Se desconoce si este transporte de agua está mediada por paracelular mecanismos o por transcelular fluyen a través de canales de AQP. McConnell y colegas [2] demostraron que el movimiento del agua en el antro de folículos aislados de rata fue 3,5 veces mayor que la de C-inulina (Un azúcar complejo que se mueve a través de los tejidos a través de transferencia paracelular), lo que indica que la afluencia de agua en la cavidad antral tiene una transcelular componente. Cuando los folículos fueron pre-tratadas con HgCl2 (un AQP inhibidor), el movimiento del agua se redujo a la de inulina. Esta sugerido un papel para AQPs en la mediación de movimiento del agua durante foliculogénesis, especialmente a la luz de la detección de AQP7, AQP8 y AQP9 en GC por citometría de flujo [2]. AQP7 y AQP9 son aquaglyceroporins; su presencia dentro del folículo ovárico sugiere que la capacidad de solutos pequeños neutros para ser transportados rápidamente a través de la plasma la membrana puede ser un requisito de la foliculogénesis. En un estudio reciente en mujeres con síndrome de ovario poliquístico, inmunofluorescencia fue utilizado para confirmar la presencia de AQP9 en el núcleo, citoplasma y membrana plasmática de GCs humanos. En un estudio de 14 enfermos con SOP y 31 sujetos de control que eran infértiles de obstrucción de las trompas, GC y líquido folicular se obtuvieron de los participantes. Testosterona total (TT) y la hormona luteinizante niveles (LH)

fueron elevados en folicular fluido de PCOS en comparación con muestras de mujeres de control; sexo niveles de globulina transportadora de hormonas (SHBG) fueron menores en pacientes con SOP.RT-PCR resultados indicaron que los niveles de mRNA AQP9 disminuyeron en Enfermos de SOP y que no había una correlación significativa entre ARNm AQP9 y TT, los niveles de LH y SHBG en muestras SOP, pero sin correlaciones en muestras de control. Los estudios in vitro mostraron que la tratamiento de GCs con la dihidrotestosterona (DHT) tenía un inhibidor efecto sobre la expresión del ARNm AQP9 y que la adición de LY294002, un fosfatidilinositol 3-quinasa (PIK3) inhibidor, atenuado esta regulación a la baja de tal manera que los niveles de ARNm AQP9 se plantearon en comparación con los tratados con DHT solo. La adición de H89 y forskolina no rectificar la DHT iniciada AQP9 mRNA disminución sugiriendo que PKA y cAMP vías no están involucrados en este mecanismo. Esto sugiere que hiperandrogenismo (exceso de hormonas androgénicas) del fluido folicular se produce en pacientes con SOP y esto suprime Expresión AQP9 en GC a través de una vía folicular PIK3 afectando el desarrollo [131]. Seguir trabajando en los mecanismos detrás Regulación AQP9 en los ovarios sanos y patógenos pueden dar una idea sobre los posibles tratamientos para enfermedades en las que el hiperandrogenismo es un problema. Los datos mencionados anteriormente sugieren que el flujo de agua en transcelular el antro del folículo ovárico es un aspecto clave de la foliculogénesis y que el movimiento de agua para la expansión del antro es probable que se AQP mediada.
3.7.2. El sistema reproductor masculino
Cada vez hay más evidencia de que AQPs juegan un papel importante en RVD espermatozoide; esto asegura el mantenimiento de la estructura y función de los espermatozoides y la fertilidad tanto masculina. ADNc AQP7 fue aislado de rata testículo y fue demostrado que se expresa abundantemente en todo el testículo y en la membrana plasmática de spermatoids etapa tardía [132]. Aquaglyceroporins, AQP3 y AQP7, se han identificado dentro de los espermatozoides humanos [40,41,42,131,132,133] y sus roles investigados. AQP3 ha sido descrita como regulador de osmoadaptation espermatozoides durante hombre a mujer de transición, durante el cual los espermatozoides están expuestos a un ambiente hipo-osmótica con el potencial de dañar el esperma por el exceso de inflamación y menor la motilidad [41]. AQP3 se encuentra en la membrana plasmática de la flagelo de espermatozoides; Células mutantes muestran una disminución de la motilidad AQP3, aumentaron hinchazón y la cola flexión después de entrar en el medio ambiente hipotónica de Por lo tanto, el útero dificultando las posibilidades del esperma de alcanzar el oviducto y la mediación de un evento de la fertilización. Estos defectos son probablemente debido a los mecanismos de RVD ineficaces y la consiguiente inflamación después hipotónica el estrés [42]. Un estudio más reciente ha descrito una relación entre Localización AQP7 y características espermáticas; microscopía electrónica de transmisión imágenes mostraron expresión de AQP7 dentro del pericentriolar región del cuello, la región ecuatorial del acrosoma y una tinción difusa a lo largo de la cola. Muestras de esperma anormal, caracterizado por malformaciones de la cabeza, pieza intermedia, o la cola, que se muestra la intensidad más baja y tinción difusa en el citoplasma de cuerpos residuales, la cabeza y la cola. Especifico correlación entre el etiquetado AQP7 espermatozoides normales y el espermatozoide motilidad y morfología sugerido que AQP7 también tiene un papel en la regulación de los espermatozoides y la fertilidad masculina [134]. Varios miembros de la familia AQP se expresan dentro del epidídimo del tracto reproductor masculino [129]. Ellos están localizados en el capa epitelial y se cree que juegan un

papel importante en transepitelial transporte de agua y la concentración de espermatozoides [129]. AQP9 era la primera AQP identificó en el epidídimo [135] y ha sido etiquetado la principal AQP apical de las células principales del epitelio; Permite flujo transepitelial de solutos tales como glicerol, urea, manitol y sorbitol y es modulada por los andrógenos en ratas adultas de sexo masculino [136]. AQP3 se localiza exclusivamente a las membranas de las células basales del epidídimo y aunque AQP1 está ausente de las células epiteliales del epidídimo es expresado en el músculo liso y el endotelio vascular de la canales a través de los epidídimo [137], junto con AQP10 [138]. AQPs son importantes para facilitar una respuesta RVD en células de esperma especialmente después de la introducción con el medio ambiente hipotónico de la aparato reproductor femenino, que en el esperma-AQP3 deficiente puede causar hinchazón perjudicial y movilidad reducida. Si la regulación de los espermatozoides las células depende de mecanismos RVD, el trabajo futuro en este campo debe concentrarse en dilucidar estos mecanismos. Mecanismos RVD en otros tejidos a menudo implican complejos con AQPs, tales como la AQP4 compleja / TRPV4 en astrocitos [60] y el AQP2 / interacción TM5b en el riñón [77]. Por tanto, es factible que AQP3 y / o AQP7 en el esperma humano podría formar complejos moleculares con los canales iónicos tales como el volumen del canal de cloruro sensible CLC-3, que ha sido identificado en el esperma de mamífero y implicado en RVD [139140].
3.8. El sistema digestivo
3.8.1. Localización y funciones reguladoras de AQPs
Secreción y absorción, dos de las principales funciones del aparato digestivo sistema, tanto requiere la transferencia de fluido a través de las membranas celulares. Secreciones diarias en forma de saliva, jugos gástricos, mucosa intestinal, la bilis y el jugo pancreático comprenden un volumen total de aproximadamente 7,5 L de líquido en el sistema digestivo humano; aproximadamente 9 L de líquido se absorbe diario [141]. Varios miembros de la familia son AQP expresado en todo el sistema digestivo incluyendo AQP1 en el apical y las membranas basolateral y el citoplasma de cholangiocytes, la páncreas y a lo largo de las células endoteliales de los capilares responsables para la transferencia de agua transendotelial [142]; el revestimiento epitelial de la tracto gastrointestinal contiene AQP3 [143] y AQP4 [144,145]. Pancreático células de los conductos expresan AQP8 en la membrana plasmática apical [146] y AQP8 [147] y AQP9 [148] se encuentran en los hepatocitos del hígado; exámenes a fondo en la expresión y localización de AQPs en el sistema digestivo están disponibles en otros lugares [141,149]. En el tracto digestivo superior, AQP3 se expresa abundantemente en estratigráfica epitelios cado de la cavidad oral a la delantera del estómago. AQP3 se localiza en la membrana de células basales y el intermedio cada vez menos abundantes hacia la superficie epitelial y se cree que proporcionar un suministro de agua desde el lado sub-epitelial de estas células que se enfrentan a condiciones duras, tales como el bajo pH del estómago, para evitar que la deshidratación [143]. La inmunocitoquímica ha mostrado una fuerte expresión en AQP4 la membrana basolateral de las células parietales gástricas en ratones y Se formuló la hipótesis de jugar un papel en la secreción de ácido gástrico. Wang y colegas [144] utilizan varios agonistas de secreción gástrica para aumentar la producción de ácido en AQP4 ratones nulos y control; sin diferencias significativas fueron vistos en los niveles de secreción que sugieren que la supresión de AQP4 hizo no afecta la capacidad del estómago para secretar ácido gástrico [144]. En el colon distal y el recto, AQP3 se localiza en la basolateral membrana de las células epiteliales que recubren el lumen [143]. La inhibición de la AQP3 por

HgCl2 en ratas inducida por diarrea severa, lo que sugiere un papel para AQP3 en la regulación del contenido de agua fecal [150], a pesar de que debería ser observó que el mercurio es un tóxico inhibidor, no específica de AQP3 [151]. AQP3 puede mediar la reabsorción de agua de las heces por transportarlo desde el lumen, a través de la capa endotelial en el vasos sanguíneos a través de AQP1 [113]. El mecanismo por el cual esto es controlado se desconoce, pero una mayor comprensión podría llevar a tratamientos para problemas intestinales sobre- o bajo activo. Por ejemplo, la inhibición temporal de AQP3 podría tener un efecto laxante. AQP8 puede tener un papel en la secreción de bilis en los hepatocitos, que son responsable de la formación de la bilis antes de que sea secretado en la bilis conducto y modificado por cholangiocytes. AQP8 se detectó en el citoplasma y vesículas intracelulares de hepatocitos de rata por confocal microscopía. El tratamiento a corto plazo con cAMP inducida redistribución de AQP8 a la membrana plasmática y un aumento en la permeabilidad al agua dentro de 10 minutos. El inhibidor de microtúbulos, colchicina, bloqueó los efectos el tratamiento de cAMP que indica que la translocación AQP8 es estimulada por dependiente de cAMP y es microtúbulos [152].
3.8.2. Conclusiones
El sistema digestivo es un sitio importante de movimiento de fluido y tiene un perfil amplia expresión AQP dentro de su red de órganos. Un polarizada Patrón de expresión AQP sugiere que una ruta transcelular organizado para el agua es una función esencial de AQPs para facilitar la secreción de alto y las tasas de absorción.
3.9. El sistema musculoesquelético
3.9.1. Localización y funciones reguladoras de AQPs
El cartílago articular y el disco intervertebral (DIV) de tejidos especializados estructuras biomecánicas que están bajo cargas de compresión constantes [153,154]. Las células dentro de estos tejidos avasculares están expuestos a constantemente condiciones duras como el IVD es ~ 80% de agua [155] y articular tejido cartilaginoso es de alrededor de ~ 70% de agua [156]. El IVD se compone de tres regiones distintas: el núcleo pulposo gelatinoso (NP), que es encapsulado por el anillo fibroso y el extremo cartilaginoso placas de [153]. Las células nativas de la NP y el tejido de cartílago tanto secrete proteoglicanos y colágeno de tipo II; las trampas de la malla de colágeno proteoglicanos cargados negativamente (tales como aggrecan) que atraen cationes (principalmente K +, Na + y Ca +) resultantes en la afluencia de agua; esta proceso es responsable de la alta potencial osmótico de estos tejidos [157158] lo que les permite resistir biomecánico estático y dinámico cargas [154]. Tanto las células NP y los condrocitos deben regular su contenido y volumen de agua en estos osmótica que cambia rápidamente ambientes; Sin embargo poco se sabe acerca de los mecanismos que que emplean para hacer esto. Recientemente, los estudios se han realizado para identificar AQPs que se expresan en estos tejidos; AQP1 y bajos niveles de AQP3 también han sido identificados dentro de las células NP de la IVD humana [159] mientras AQP1 y AQP3 han demostrado que se expresa y colocalizaban en la membrana de cartílago articular equino condrocitos células [154] y los condrocitos de la rodilla humana [156]. AQP1 y AQP4 se expresan en el músculo esquelético y un estudio ha demostrado que los cambios de volumen de células que se producen durante la contracción muscular confiar en la rápida afluencia de agua [160]. AQP1 se encontró en el endotelial células de los capilares en el tejido muscular y en el plasma AQP4 membrana de las células de fibra muscular [160]. La localización de AQP1 y AQP4

dentro del tejido muscular sugiere una vía para transcelular el flujo de agua a través de la membrana de la célula endotelial y el sarcolema; estos AQPs pueden funcionar juntos como transportadores para el agua entre la sangre y miofibrillas durante la actividad muscular mecánica.
3.9.2. Conclusiones
No es de extrañar que las células nativas de la PN y articular cartílago AQPs expresas y es muy probable que AQPs son responsables para la CVR en estos ambientes altamente osmóticos. Se requiere más trabajo para dilucidar el papel funcional de las acuaporinas en estos tejidos; la número de estudios publicados han sugerido papeles para AQPs como componentes del aparato celular vital para el mantenimiento de la fisiológico homeostasis del sistema músculo-esquelético.
4. Conclusiones
La distribución constitutiva de AQPs se logra por el gen AQP expresión y / o la degradación de proteínas AQP en una escala de tiempo de horas al día [161]. Además rápida distribución espacial y temporal de AQPs es regulado por translocación estimulada o provocada de AQP-que contiene vesículas hacia y desde una membrana particular. Esto es particularmente así estudiado para V2R mediada AQP2 translocación en el conducto renal recogiendo células [74] (Fig. 3B). Mecanismos similares de translocación disparada tienen ha demostrado para otros AQPs [57]. Proponemos que la resultante distribución espacial y temporal de AQPs es crucial para la regulación de flujo de agua transcelular en los principales sistemas del cuerpo humano. Por ejemplo, la reabsorción de agua desde el lumen del riñón conducto colector implica el flujo de agua transcelular mediada por AQP2 en la membrana apical y AQP3 y AQP4 en la membrana basolateral. Flujo de agua transcelular a través de las células endoteliales de los capilares en la sangre es entonces mediada por AQP1 (Fig. 3A). En otro ejemplo, Ratones knockout AQP4 muestran cerca de los niveles normales de la presión intracranealy el contenido de agua, pero la acumulación reducida de agua en el cerebro siguiente edema citotóxico causado por el accidente cerebrovascular isquémico, lesión cerebral y meningitis [162163]. En los modelos vasogénicos edema ratón, AQP4 null ratones muestran una mayor acumulación de agua en el cerebro [61]. Estas Los estudios sugieren que la homeostasis del agua en un estado no patológico puede ser independiente del flujo de agua transcelular AQP4 mediada pero AQP4 que regula el flujo de agua transcelular en el edema cerebral (Fig. 3C). En el edema vasogénico, el agua se elimina de la extracelular espacio a través de AQP4 en los astrocitos intactos que componen la BBB. El agua sale entonces a través de la astrocito AQP4 en la membrana de astrocytic procesos de pie que rodean el capilar. Flujo de agua transcelular a través de células endoteliales capilares en la sangre está mediada por AQP1. Sin embargo, en el edema citotóxico, el flujo de agua desde el transcelular sangre en astrocitos está mediada por AQP4. En conclusión, parece que AQPs son cruciales para la regulación de homeostasis del agua, proporcionando poros selectivos para el movimiento rápido de agua, y otros solutos no cargados, a través de las membranas celulares diversas y jugar un papel regulador en la CVR. Mecanismos de compuerta, que permitan conformacionalmente estados abierto y cerrado distintas, se han propuesto para AQPs humanos a través de simulaciones moleculares dinámicas [164], pero Solamente se ha observado específicamente para AQPs vegetales y microbianas [165]. En consecuencia, es probable que la distribución y la abundancia de AQPs en una membrana particular, son los determinantes de la membrana permeabilidad al agua y un regulador de flujo de agua transcelular


















