Análisis de estabilidad en un modelo de remodelación ósea...
80
Equation Chapter 1 Section 1 Trabajo Fin de Grado Grado en Ingeniería Química Análisis de estabilidad en un modelo de remodelación ósea basado en poblaciones celulares Autor: Clara Benito Gallego Tutor: Javier Martínez Reina Dep. Ingeniería Mecánica Escuela Técnica Superior de Ingeniería Universidad de Sevilla Sevilla, 2015
Transcript of Análisis de estabilidad en un modelo de remodelación ósea...

Equation Chapter 1 Section 1
Trabajo Fin de Grado
Grado en Ingeniería Química
Análisis de estabilidad en un modelo de
remodelación ósea basado en poblaciones
celulares
Autor: Clara Benito Gallego
Tutor: Javier Martínez Reina
Dep. Ingeniería Mecánica
Escuela Técnica Superior de Ingeniería
Universidad de Sevilla
Sevilla, 2015

Trabajo Fin de Grado
Grado en Ingeniería Química
Análisis de estabilidad en un modelo de
remodelación ósea basado en
poblaciones celulares
Autor:
Clara Benito Gallego
Tutor:
Javier Martínez Reina
Dep. Ingeniería Mecánica
Escuela Técnica Superior de Ingeniería
Universidad de Sevilla
Sevilla, 2015

Índice
Índice
1 Introducción/Objetivos 1
2 Biología ósea 2 2.1. Funciones del tejido óseo 2 2.2. Tipos de tejidos óseos 2
2.2.1 Tejido óseo compacto 2 2.2.2 Tejido óseo esponjoso o trabecular 3
2.3. Tipos de huesos 4 2.3.1 Huesos largos 4 2.3.2 Huesos cortos 5 2.3.3 Huesos planos 5 2.3.4 Huesos irregulares 5
2.4. Composición del hueso 5 2.4.1 Matriz ósea 5 2.4.2 Células óseas 6
3 Remodelación ósea 10 3.1 Etapas de la remodelación ósea 10
3.1.1 Activación 10 3.1.2 Reabsorción 10 3.1.3 Inversión 12 3.1.4 Formación 12 3.1.5 Mineralización 12 3.1.6 Final 12
3.2 Factores que influyen en la remodelación ósea 13 3.2.1 Factores bioquímicos 13 3.2.2 Factores biomecánicos 15 3.2.3 Factores geométricos 15
4 Modelo matemático 16 4.1 Mecanismos de regulación que tiene en cuenta el modelo 16
4.1.1 Mecanismos bioquímicos 16 4.1.2 Mecanismos biomecánicos 20 4.1.3 Mecanismos geométricos 22
4.2 Desarrollo del modelo matemático 23 4.2.1 Hipótesis consideradas en el modelo 23 4.2.2 Modelo matemático 24 4.2.3 Parámetros del modelo 24
5 Estabilidad del sistema 30 5.1 Fundamentos de la linealización 30 5.2 Particularización de la linealización 32

5.2.1 Parámetros del modelo en función de OBp, OBa, OCp y OCa 32 5.2.2 Ecuaciones del modelo en función de OBp, OBa, OCp y OCa 36 5.2.3 Cálculo del punto estacionario 40 5.2.4 Cálculo de la matriz jacobiana 41
6 Análisis de variables 48 6.1 Simplificaciones tenidas en cuenta para el análisis 48
6.1.1 Regulación geométrica 48 6.1.2 Regulación biomecánica 48 6.1.3 Función de activación de la diferenciación de OCu en OCp 48 6.1.4 Tasa de producción de RANKL por preosteoblastos 49
6.2 Análisis de constantes del modelo 49 6.2.1 Poblaciones de osteoblastos y osteoclastos precursores 49 6.2.2 Constantes de diferenciación 51 6.2.3 Constantes de apoptosis 54 6.2.4 Índice de proliferación de OBp, POBp 55 6.2.5 Constantes de disociación 56 6.2.6 Constantes de asociación 61 6.2.7 Función de activación de la diferenciación de OCu en OCp, πact, OCuMCSF 62 6.2.8 Índices de producción 63 6.2.9 Índices de degradación 65 6.2.10 Volumen diario de matriz ósea 67 6.2.11 Densidad de TGFβ almacenado en la matriz ósea, ηtgf βbone 68 6.2.12 Densidad OPG a la que para la producción endógena, βOPGmax 69 6.2.13 Máximo número de RANKL/OBp, NOBprankl 69 6.2.14 Receptores RANK/OCp, NOCprank 70
7 Aplicaciones a los casos inestables 71
8 Conclusiones 74
Bibliografía 76

1. INTRODUCCIÓN/OBJETIVOS
1
1 INTRODUCCIÓN/OBJETIVOS
El hueso está sometido a un proceso continuo de renovación de manera que se van reparando las zonas
dañadas. Este proceso se conoce con el nombre de remodelación ósea. Dicha remodelación se lleva a
cabo mediante la acción de diferentes poblaciones celulares: osteoclastos, que contribuyen a la
resorción del tejido dañado y los osteoblastos que contribuyen a la formación de un nuevo tejido óseo,
además de los osteocitos y las células de borde que también participan de este proceso.
En este trabajo se va a revisar el modelo matemático propuesto por Pivonka et al. que da forma a la
dinámica de las poblaciones celulares durante la remodelación ósea. El modelo en cuestión tiene en
cuenta distintos tipos de mecanismos de regulación: i) bioquímicos, mediante la segregación de
hormonas y citoquinas que permiten su comunicación, ii) biomecánicos, los osteocitos responden al
esfuerzo mecánico convirtiendo la señales mecánicas en señales bioquímicas, iii) geométricos, que
tienen en cuenta la influencia de la disponibilidad de superficie microscópica ósea en el desarrollo y
actividad de las poblaciones celulares.
El objetivo del trabajo es realizar un estudio de estabilidad del modelo analizando cada una de las
constantes que lo conforman y estudiar los casos en los que el sistema sea inestable.

2. BIOLOGÍA ÓSEA
2
2 BIOLOGÍA ÓSEA
El sistema óseo o sistema esquelético es uno de los principales sistemas del cuerpo humano. Los
huesos, unidos entre sí mediante articulaciones, forman el esqueleto.
2.1. Funciones del tejido óseo
El tejido óseo cumple varias funciones:
1. Entrega soporte y forma al cuerpo. Actúa de sostén contra la fuerza de la gravedad.
2. Sirve de protección al cuerpo, protege los órganos internos frente a impactos.
3. Por su interacción con el sistema muscular tiene un importante rol en la generación de
movimiento. Gracias a los músculos que se insertan en los huesos a través de los tendones y
su contracción sincronizada se produce el movimiento.
4. El tejido óseo almacena una serie de minerales que cuando son necesarios los libera a la
sangre y ésta los distribuye a otras partes del organismo. Actúa de reserva de calcio.
5. Aloja la médula ósea, fuente de células madre y células sanguíneas.
2.2. Tipos de tejidos óseos
Todos los tipos de huesos estarán compuestos por dos tipos de tejidos:
2.2.1 Tejido óseo compacto
El tejido óseo compacto es una masa compacta de tejido óseo donde las cavidades sólo son visibles al
microscopio. Es el tejido óseo que observaríamos en la diáfisis. El hueso compacto presenta una
estructura formada por láminas concéntricas alrededor de un conducto central que se denomina
conducto de Havers. Los osteocitos, las células maduras del hueso, se disponen normalmente entre
estas láminas concéntricas. El conducto de Havers es el conducto por donde van a pasar los vasos
sanguíneos que nutren al hueso y los nervios. La forma en que se orientan los sistemas de Havers en
cada zona del hueso compacto guarda estricta relación con los requerimientos mecánicos a que el
hueso está sometido. Al conjunto formado por el conducto de Havers y las laminillas óseas con los
osteocitos se denomina osteona. La osteona es la unidad estructural del hueso compacto. Además de
estos canales existen otros transversales, los canales de Volkman, que conectan los canales de Havers

2. BIOLOGÍA ÓSEA
3
entre sí y con el periosteo.
Figura 1
Tejido óseo compacto. Se observan las osteonas
2.2.2 Tejido óseo esponjoso o trabecular
El tejido óseo esponjoso o trabecular es el hueso que observaríamos en la epífisis. Está constituido por
una red tridimensional de espículas o trabéculas óseas ramificadas que limita un laberinto de espacios
intercomunicados ocupados por médula ósea con vasos sanguíneos. Dichas trabéculas se disponen
según la dirección de la carga para trabajar de un modo más eficaz. El tejido óseo esponjoso tiene otra
estructura histológica en la que las láminas óseas no están organizadas de forma concéntrica sino que
forman trabéculas dispuestas en red observándose como un mosaico de piezas angulares que dejan
espacios internos dónde se sitúa la médula ósea. Las laminillas no forman osteonas. En este caso los
vasos sanguíneos no penetran en las trabéculas así que los osteocitos se nutren de los propios vasos
que hay en la médula ósea.
Figura 2
Tejido óseo esponjoso. Se observan las trabéculas

2. BIOLOGÍA ÓSEA
4
2.3. Tipos de huesos
Los huesos pueden clasificarse según su forma en largos, cortos, planos e irregulares.
2.3.1 Huesos largos
Los huesos largos son brazos de palanca para ejecutar movimientos. En los huesos largos podemos
distinguir tres regiones anatómicas: a) los extremos o epífisis que están constituidos por hueso
esponjoso recubierto por una capa delgada de hueso compacto, la epífisis es la parte encargada de
interaccionar con otros huesos para formar las articulaciones mediante el cartílago articular que se
encuentra en su extremo, b) la diáfisis que es un cilindro hueco de paredes gruesas formadas por tejido
óseo compacto. En la cavidad central de la diáfisis se encuentra la médula ósea. La diáfisis se ensancha
en los extremos, a la vez que disminuye el espesor de la capa de cortical y comienza a aparecer hueso
trabecular. A esta zona del hueso se le denomina c) metáfisis.
Figura 3
Estructura de un hueso largo

2. BIOLOGÍA ÓSEA
5
2.3.2 Huesos cortos
Los huesos cortos dan sostén y soportan pesos. Están formados por una capa cortical de tejido óseo
compacto y una zona central de hueso esponjoso. Son aquellos que no tienen una dimensión
predominante. Se localizan agrupados y habitualmente permiten el movimiento de las
extremidades, como los huesos del carpo y del tarso.
2.3.3 Huesos planos
Los huesos planos se encargan de proteger órganos. Son aquellos que tienen una dimensión bastante
más pequeña que la de los huesos largos o cortos. Apenas soportan carga, como en el caso del
cráneo. Están compuestos de una capa de hueso esponjoso entre dos capas delgadas de hueso
compacto. Su forma es aplanada, no redondeada. Tienen médula, pero no una cavidad de
médula ósea.
2.3.4 Huesos irregulares
Representan todos aquellos huesos que por su forma no se pueden clasificar en otro tipo.
2.4. Composición del hueso
El tejido óseo es una variedad de tejido conjuntivo. Como todos los tejidos conjuntivos, el tejido óseo
está formado por células y por una matriz ósea.
2.4.1 Matriz ósea
La matriz ósea presenta una parte orgánica que forma el 20% de esta matriz, una parte inorgánica que
representa el 70 %y el 10 % restante es agua
La fase orgánica está formada por sustancia fundamental (proteoglicanos y glucoproteínas) y
fibras principalmente colágenas, siendo el colágeno tipo I la más abundante aunque también
encontramos colágeno tipo III y tipo VI. La estructura helicoidal del colágeno permite que se
soporten cargas atracción y cargas a compresión. También encontramos otras proteínas no
colágenas como la osteocalcina, la osteonectina o la osteopontina.
La fase inorgánica está formada por fosfato de calcio qué forma los cristales de hidroxiapatita
Ca10(PO4)6(OH)2 encargados de proporcionar resistencia y dureza al tejido óseo. El cristal de
hidroxiapatita puro tiene simetría hexagonal, pero se agrupa con otros cristales y forma una
superceldilla con simetría ortorrómbica. Tiene impurezas que hacen que el cristal sea amorfo.

2. BIOLOGÍA ÓSEA
6
Figura 4
Fibra de colágeno
En estos huecos ( ) se empieza a atraer el mineral. Éste empieza a crecer y a rodear el tropocolágeno.
El agua contenida en el hueso forma parte de la médula ósea que rellena los poros. Al
principio, cuando se forma el hueso, éste está formado por fase orgánica y por agua. El agua
es sustituida por mineral de forma progresiva durante la mineralización ósea.
2.4.2 Células óseas
El tejido óseo está formado por células óseas especializadas las cuales poseen diferentes funciones.
Podemos encontrar entre ellas: los osteocitos, células que se encuentran en el interior del tejido óseo,
los osteoblastos, células formadoras del tejido óseo, los osteoclastos, las cuales destruyen el tejido óseo
y las células de borde, encargadas de proteger la matriz ósea. Los osteoblastos y osteoclastos trabajan
conjuntamente para la regulación del crecimiento óseo.
Las células osteoprogenitoras o células madre ósea son células indiferenciadas con carácter de
fibroblastos. Durante la formación de los huesos está células sufren división y diferenciación a células
formadoras de hueso (osteoblastos) mientras que los preosteoclastos darán origen a los osteoclastos.
Estas células osteoprogenitoras se encuentran en la capa interna del periostio en el endostio y en los
canales del hueso que contienen los vasos sanguíneos.

2. BIOLOGÍA ÓSEA
7
Figura 5
2.4.2.1 Osteoclastos
Son células multinucleadas, tienen alrededor de 10 o 20 núcleos. Son grandes, miden alrededor de 100
μm. Son ricas en mitocondrias y vacuolas, así como en lisosomas. Están encargadas de la reabsorción
del tejido óseo, tanto del dañado como del inútil por su poca eficiencia. Son producidas por la médula
ósea, y la cara del osteoclasto pegada a la matriz ósea segrega ácidos que hacen disminuir el PH y
disuelven la fase mineral del tejido. A continuación reabsorben la fase orgánica con la acción de
enzimas, avanzando hacia el interior del tejido a una velocidad de decenas de micras al día. Al finalizar
la reabsorción reciben una señal de tipo química y sufren apoptosis (suicidio celular programado).
2.4.2.2 Osteoblastos
Son los responsables de la síntesis de la mayoría de los componentes de la matriz ósea y de la
activación de los osteoclastos. Los osteoblastos proceden de la diferenciación de células madre
mesenquimales. Éstos están sometidos a una diferenciación a través de diversos pasos teniendo como
objetivo final su diferenciación a osteocitos. Se diferencian (2 a 3 días) de células madre
mesenquimales, pero para ello necesitan un cierto nivel de estímulo mecánico, mientras no haya una
determinada fuerza no terminan de diferenciarse. Los osteoblastos son producidos tanto en la medula
ósea como en la capa interna del periostio (membrana dura que recubre la mayor parte de los huesos)
bajo ciertos niveles de estímulo mecánico. Los principales productos proteicos de un osteoblasto son:
colágeno tipo I y varias proteínas no colágenas entre las que se incluyen el RANKL, la osteocaicina,
necesaria para la mineralización ósea, la osteopontina, que interviene en la formación de la zona de
sellado y la sialoproteína ósea que interviene en la unión de los osteoblastos con la matriz extracelular.

2. BIOLOGÍA ÓSEA
8
Los osteoblastos siguen tras el hueco producido por los osteoclastos (probablemente siguiendo un
rastro químico) rellenándolo con osteoide a una velocidad de 1μm/día. Una vez ejercida su función la
mayoría de los osteoblastos desaparecen por un proceso de apoptosis o muerte celular programada, no
obstante, parte de los osteoblastos quedan rodeados y enclaustrados por la matriz que han sintetizado.
En este momento cesa su actividad anabólica y se convierten en osteocitos. Además, algunos
osteoblastos quedan retenidos en la superficie ósea y conforman las denominadas células de borde.
2.4.2.3 Osteocitos
Son osteoblastos que han quedado atrapados dentro de la matriz ósea. El osteoblasto va echando capas
y capas de osteoides de manera que estos se estrujan y se diferencian a osteocitos. Al diferenciarse
cambian su forma y aparecen unas ramificaciones que ocupan los túneles de la matriz y les permite
conectarse entre sí y con las células de borde. Más del 90% de las células óseas son osteocitos.
2.4.2.4 Células de borde
De los osteoblastos que han finalizado su misión de segregación de osteoide en la superficie de la
matriz ósea, unos sufren apoptosis y otros se diferencian y cambian su forma a una más plana y
alargada, cubriendo toda la matriz. Tienen dos funciones principales: protegen la matriz ósea de
diversos agentes químicos presentes en la sangre que pueden disolver el mineral y servir de control de
la remodelación iniciándola, mantienen la comunicación con los osteocitos y tienen receptores de
hormonas (estrógeno, PTH) y otros “mensajeros químicos”, para iniciar la remodelación ósea en caso
necesario. Cuando se inicia la remodelación deben retirarse de la superficie para que los osteoclastos
actúen disolviendo la matriz. Para ello forman un conducto con los vasos cercanos.

2. BIOLOGÍA ÓSEA
9
Figura 6
Células óseas

3. REMODELACIÓN ÓSEA
10
3 REMODELACIÓN ÓSEA
La remodelación ósea se basa principalmente en la acción de dos tipos de células: los osteoclastos son
los responsables de la resorción del hueso, mientras que los osteoblastos tienen el papel de la
formación mediante la secreción de osteoide, una sustancia rica en colágeno que al mineralizarse
forma una nueva matriz ósea.
Ambas poblaciones de células trabajan conjuntamente en las llamadas Unidades Multicelulares
Básicas (BMUs). Una BMU consiste en un conjunto de células en diferentes fases de maduración.
Ésta incluye osteoclastos, osteoblastos y osteocitos dentro de la matriz ósea y células de borde
cubriendo la superficie del hueso. Cada BMU pasa por las diferentes fases que conforman la
remodelación ósea.
3.1 Etapas de la remodelación ósea
El funcionamiento biológico del tejido óseo trata de una compleja sucesión de señales bioquímicas y
actividades celulares que conllevan la reabsorción de hueso (por ser innecesario o por estar dañado) y
la posterior formación de tejido nuevo a través de la activación de las BMU. A esta secuencia de
sucesos se le conoce como Secuencia ARF y puede ser dividida en 6 fases:
3.1.1 Activación
En el tejido óseo existe un entramado de osteocitos conectados entre sí y con las células de borde por
medio de ramificaciones que atraviesan los poros y canales del hueso junto con el fluido sanguíneo
que riega todo el tejido proveyéndolo de nutrientes. Estos filamentos son sensibles a las tensiones
tangenciales producidas por dicho flujo, el cual fluye a través de los canalículos gracias al estímulo
mecánico que los deforma. Si dicho flujo se interrumpe (bien por falta de estímulo mecánico, bien por
la presencia de microgrietas que actúan de “embalse” para el fluido), los osteocitos dejan de emitir la
señal inhibitoria a las células de borde.
3.1.2 Reabsorción
Las células de borde son las encargadas de comenzar la remodelación. Según la teoría inhibitoria, su
tendencia es la de promover la remodelación en todo momento, pero la señal inhibitoria que reciben
de los osteocitos se lo impide. Al dejar de sentirla segregan una sustancia química, el RANK-L, que
es detectada por la proteína RANK de los precursores de los osteoclastos presentes en la médula ósea
provocando su diferenciación y fusión en osteoclastos maduros. Conjuntamente, las células de borde
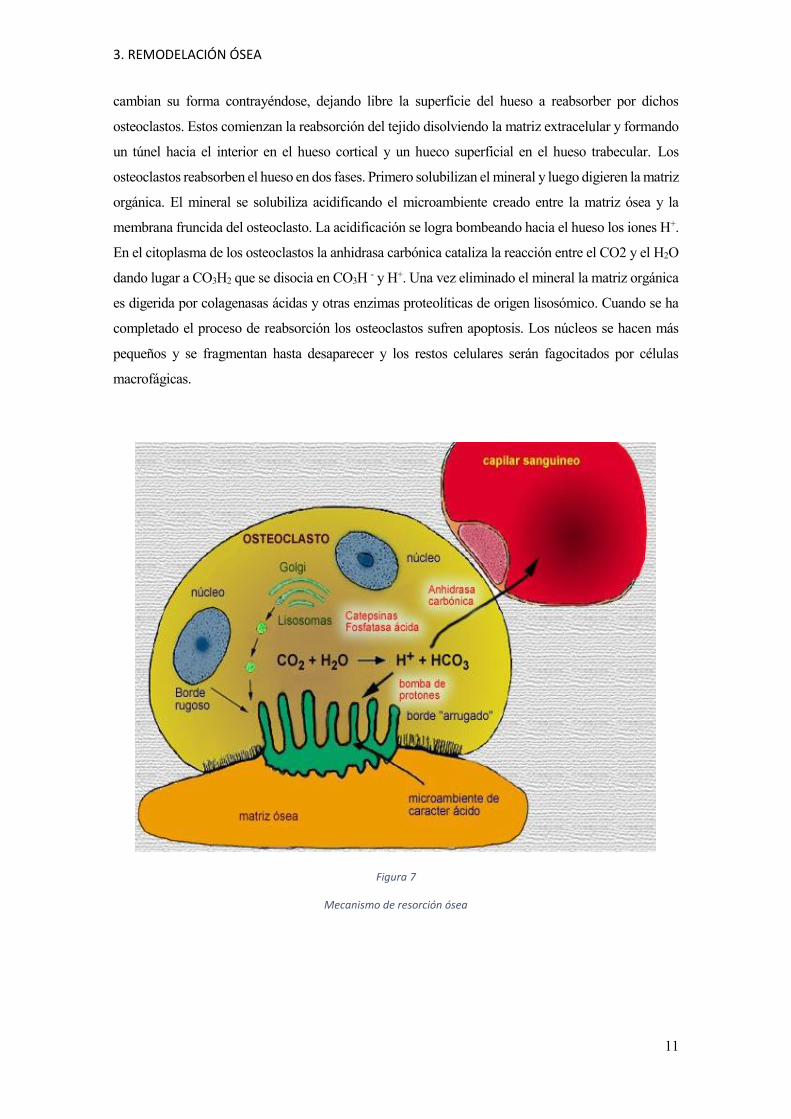
3. REMODELACIÓN ÓSEA
11
cambian su forma contrayéndose, dejando libre la superficie del hueso a reabsorber por dichos
osteoclastos. Estos comienzan la reabsorción del tejido disolviendo la matriz extracelular y formando
un túnel hacia el interior en el hueso cortical y un hueco superficial en el hueso trabecular. Los
osteoclastos reabsorben el hueso en dos fases. Primero solubilizan el mineral y luego digieren la matriz
orgánica. El mineral se solubiliza acidificando el microambiente creado entre la matriz ósea y la
membrana fruncida del osteoclasto. La acidificación se logra bombeando hacia el hueso los iones H+.
En el citoplasma de los osteoclastos la anhidrasa carbónica cataliza la reacción entre el CO2 y el H2O
dando lugar a CO3H2 que se disocia en CO3H - y H+. Una vez eliminado el mineral la matriz orgánica
es digerida por colagenasas ácidas y otras enzimas proteolíticas de origen lisosómico. Cuando se ha
completado el proceso de reabsorción los osteoclastos sufren apoptosis. Los núcleos se hacen más
pequeños y se fragmentan hasta desaparecer y los restos celulares serán fagocitados por células
macrofágicas.
Figura 7
Mecanismo de resorción ósea

3. REMODELACIÓN ÓSEA
12
3.1.3 Inversión
La formación del nuevo tejido corre a cargo de los osteoblastos, pero la activación de estos no es
inmediata, sino que en el ser humano ocurre unos 30 días después de la activación de los osteoclastos.
El período de inversión es un periodo de aparente inactividad.
3.1.4 Formación
Los preosteoblastos son formados por diferenciación de células madre mesenquimales y atraídos a las
paredes de la cavidad dejada por los osteoclastos por factores químicos. Allí maduran, dando lugar a
los osteoblastos que comienzan a depositar osteoide (colágeno y agua) rellenando la cavidad. La
formación de estos preosteoblastos está supeditada a la existencia de un cierto nivel de estímulo
mecánico. Los osteoblastos sintetizan el componente orgánico de la matriz ósea (colágeno tipo I,
proteoglicanos, proteínas implicadas en la adhesión celular, osteocalcina y factores de crecimiento) y
controlan el depósito de las sales minerales. Los osteoblastos pasan sucesivamente por tres estadios
funcionales: a) proliferación celular y síntesis de los componente orgánicos de la matriz ósea, b)
maduración de la matriz ósea (cambios en la composición y organización de la matriz que la hacen
competente para ser mineralizada) y c) depósito de mineral.
3.1.5 Mineralización
Los osteoblastos solo depositan osteoide, no mineral. Este comienza a aparecer unos 10 días después
de haber sido depositado el osteoide al desplazar al agua contenido en este.
3.1.6 Final
Los osteoclastos sufren apoptosis y la BMU se detiene. Los osteoblastos terminan de rellenar la
cavidad y se diferenciaran a osteocitos o células de borde en función de donde hayan quedado
atrapadas. Estas células se encargarán de iniciar futuras remodelaciones en esta zona si fuera necesario.
La actividad de la BMU termina mucho antes de que el tejido se mineralice por completo, puesto que
la vida media de una BMU es de unos 100 días (unos 3 meses).

3. REMODELACIÓN ÓSEA
13
Figura 8
Etapas de la remodelación ósea
3.2 Factores que influyen en la remodelación ósea
3.2.1 Factores bioquímicos
Entre los factores bioquímicos involucrados en la remodelación ósea los más importantes son los
factores locales, autocrinos y paracrinos. En este apartado se van a comentar los factores más
importantes para posteriormente, desarrollarlos de manera más detallada cuando se explique el modelo
matemático que gobierna el comportamiento de las poblaciones celulares en el hueso.
3.2.1.1 Estimulación a la diferenciación de los osteoclastos
Los osteoclastos son estimulados a diferenciarse desde sus progenitores hematopoyéticos por el factor
estimulante de colonias de macrófagos, M-CSF. Una vez que las células hematopoyéticas se
comprometen, el factor M-CSF y el factor nuclear kappa beta ligando, RANKL, actúan conjuntamente
para favorecer la diferenciación de los osteoclastos precursores en algo más parecido a los osteoclastos
activos. Finalmente, la diferenciación a osteoclastos activos está promovida por el factor RANKL.
En la estimulación a la diferenciación de los osteoclastos el sistema RANK/RANKL/OPG juega un
papel muy importante. Éste sistema es un mecanismo de señalización esencial involucrado en la

3. REMODELACIÓN ÓSEA
14
comunicación entre las distintas poblaciones celulares óseas. Los osteoclastos precursores segregan el
receptor RANK y en la superficie de los preosteoblastos se encuentra el ligando RANKL. La unión
de RANKL con su receptor, RANK, promueve la diferenciación de preosteoclastos a osteoclastos
activos. Sin embargo, puede intervenir una tercera proteína, la osteoprotegerina (OPG) emitida por los
osteoblastos activos, actuando como receptor señuelo, secuestrando el RANKL y evitando la
formación de complejos RANK-RANKL por lo que dejan de diferenciarse osteoclastos activos,
reduciéndose la resorción ósea.
Además de la llamada señalización RANK-RANKL-OPG existen otros factores influyentes en la
remodelación ósea como las hormonas PTH, la calcitonina, los estrógenos y la vitamina D3.
Figura 9
Esquema del proceso de diferenciación que se produce en los osteoclastos y los correspondientes factores que intervienen
3.2.1.2 Estimulación a la diferenciación de los osteoblastos
Existen factores de crecimiento, como el TGF-β, que se encuentran almacenados en la matriz ósea y
son liberados durante la resorción llevada a cabo por los osteoclastos, promoviendo la activación de
los pre-osteoblastos en osteoblastos activos.
Además del factor TGF-β, existen otros potenciadores de la población de pre-osteoblastos como son
las proteínas Wnts.

3. REMODELACIÓN ÓSEA
15
Figura 10
Esquema del proceso de diferenciación que se produce en los osteoblastos y los correspondientes factores que intervienen. Incluye miembros de la ruta de señalización Wnt.
3.2.2 Factores biomecánicos
La estimulación mecánica del hueso acelera la formación de periostio en las regiones sometidas a
esfuerzos extremos, es decir, exceso o defecto de carga.
Las vías de señalización Wnt juegan un papel fundamental en la regulación biomecánica de la
remodelación ósea, transducciendo señales mecánicas en señales bioquímicas.
Las vías de señalización Wnt son un grupo de vías de transducción de señales mecánicas en señales
bioquímicas. Están formadas por proteínas encargadas de transferir las señales que proceden del
exterior de una célula desde la superficie receptora de ésta hasta su interior. Este mecanismo por el
cual los osteocitos convierten los estímulos mecánicos en señales bioquímicas recibe el nombre de
secuencia de mecanotransducción.
3.2.3 Factores geométricos
La disponibilidad de superficie ósea influye en el proceso de remodelación ósea ya que dicho
factor mantiene una fuerte dependencia con el reclutamiento de preosteoblastos y osteoclastos.

4. MODELO MATEMÁTICO
16
4 MODELO MATEMÁTICO
Distintos autores han propuesto modelos matemáticos que dan forma a la dinámica celular en la
remodelación ósea. En este apartado se va a desarrollar el modelo que propone Pivonka et al. que tiene
en cuenta la regulación bioquímica, biomecánica y geométrica. La gran novedad que propone este
modelo es la consideración de la regulación geométrica en las diferentes etapas del desarrollo de las
poblaciones celulares.
4.1 Mecanismos de regulación que tiene en cuenta el modelo
4.1.1 Mecanismos bioquímicos
En el modelo se consideran dos líneas celulares, osteoblastos y osteoclastos.
Las células de linaje osteoblástico proceden de células madre mesenquimales. Se consideran tres
estados de osteoblastos Se consideran tres estados en el desarrollo de los osteoblastos. Las piscinas de
células madre mesenquimales que son capaces de dar lugar a células de carácter osteoblástico,
adipocitos u otros tipos de células la denominamos osteoblastos precursores (OBUS). Una vez que estas
células se comprometen a osteoblastos, sin ser aún un sólo tipo de células sino un conjunto de varios
fenotipos que comparten características similares, las denominamos preosteoblastos (OBPS) y en tercer
lugar, los osteoblastos activos (OBAS).
La segunda línea celular considerada en el modelo es la de los osteoclastos estos proceden de células
madre hematopoyéticas. Estos bancos de células madre hematopoyéticas que pueden dar lugar a
osteoclastos recibe el nombre de osteoclastos precursores (OCUS). Del mismo modo que ocurre con el
linaje osteoblástico, el modelo cuenta con preosteoclastos (OPPS) y osteoclastos activos (OCAS).

4. MODELO MATEMÁTICO
17
Figura 11
Ilustración esquemática del modelo de poblaciones celulares
Los factores bioquímicos involucrados en el proceso de remodelación ósea son:
Sistema RANK/RANKL/OPG
El sistema RANK/RANKL/OPG es un mecanismo de señalización esencial involucrado en la
comunicación entre las distintas poblaciones celulares óseas. Esta secuencia está formada por tres
componentes principales:
i. RANK
El receptor RANK es expresado en la superficie de los preosteoclastos (OPPS). Su activación
conlleva a la reorganización del citoesqueleto del osteoclasto y a cambios fundamentales para
su activación, movilidad y establecimiento en la superficie ósea a resorber. También
desencadena una señal de supervivencia en el osteoclasto maduro.
ii. RANKL
RANKL es una citoquina (proteína responsable de la comunicación celular) que se encuentre
en la superficie de las células de linaje osteoblástico que juega un papel muy importante en la
formación, actividad y supervivencia de los osteoblastos a través de la interacción con su
receptor RANK.
iii. OPG

4. MODELO MATEMÁTICO
18
La osteoprotegerina, OPG, otra proteína segregada por los osteoblastos activos, es un inhibidor
natural de los RANKL y juegan un papel muy importante en la regulación de la resorción del
hueso Ya que actúa como receptor señuelo, secuestrando el RANKL y evitando la activación
de RANK.
Figura 12
RANK/RANKL/OPG
Cuando se dan situaciones que incrementan la remodelación ósea como puedan ser fracturas o
estímulos mecánicos, el ligando RANKL se une a su receptor RANK de manera que crea un impacto
sobre la diferenciación de los osteoclastos precursores y la subsiguiente activación de los osteoclastos
maduros en osteoclastos activos dando comienzo a la reabsorción del hueso. Por otro lado, la proteína
OPG da lugar a la formación del complejo RANKL-OPG. La presencia de OPG implica una menor
formación de complejos RANKL-RANK y por lo tanto una menor estimulación de osteoclastos para
su diferenciación. Un alto ratio de RANKL/OPG favorece la reabsorción del hueso mientras que un
bajo ratio reduce la actividad de los osteoclastos y por tanto la reabsorción de hueso.
Existen otros factores que actúan como elementos reguladores de OPG y RANKL. Entre ellos se

4. MODELO MATEMÁTICO
19
encuentran la hormona paratiroidea (PTH), glucocorticoides, estrógeno y vitamina D3. Las células de
estroma son tomadas como transductores de las señales de citoquinas/ hormonas para los osteoclastos
donde la entrada es la señal citoquina/hormona/mecánica y la salida es el ratio RANKL/OPG. Este
mecanismo de control de los niveles RANKL/OPG por citoquinas y hormonas es de suma
importancia.
Las principales hormonas que regulan la formación de osteoclastos son la hormona paratiroidea
(PTH), la vitamina D3 y la calcitonina. Los receptores de la hormona paratiroidea (PTH) y de la
vitamina D sólo son expresados en las células de linaje osteoblástico de manera que estas hormonas
participan en la formación de osteoclastos activos de manera indirecta a través de los osteoblastos. La
tercera hormona involucrada, la calcitonina, actúa directamente sobre los osteoclastos inhibiendo la
acción de la hormona paratiroidea (PTH) y de la vitamina D, inhibiendo por tanto, la resorción ósea.
La PTH (hormona paratiroidea) es una hormona proteica secretada por las glándulas paratiroideas. La
PTH actúa estimulando los osteoclastos y favoreciendo la resorción ósea, una acción ligada a la
presencia de vitamina D. La PTH favorece la proliferación de RANKL y disminuye OPG de manera
que se forma un mayor número de complejos RANK-RANKL y se incrementa la población de
osteoclastos activos resorbiéndose mayor cantidad de hueso.
Figura 13
Estimulación de osteoclastos activos en presencia de la hormona paratiroidea, PTH
En el modelo se asume que la fracción de receptores ocupados en una célula determina la fuerza de la
señal recibida por la célula y por tanto, la fuerza de la respuesta por parte de la célula a dicha señal.
Por otra parte, en el modelo, el factor de crecimiento transformante TGF-β se une con sus receptores
en los osteoblastos precursores promoviendo su diferenciación a preosteoblastos mientras que si se
unen a sus receptores en los preosteoblastos inhiben su diferenciación a osteoblastos activos. La acción
del TGF-β en los osteoblastos depende del estado de maduración de las células. Las principales
funciones del TGF-β son:
Presencia
PTH
↑RANKL
↓OPG↑↑RANKL
↑↑complejos RANK-RANKL
↑OCa

4. MODELO MATEMÁTICO
20
- Provocar la diferenciación de los osteoblastos precursores a preosteoblastos
- Inhibir la diferenciación de los preosteoblastos a osteoblastos activos
- Provocar la apoptosis de los osteoclastos
Además del factor TGF-β, existen otros potenciadores de la población de pre-osteoblastos como son
las proteínas Wnts cuyas funciones principales son:
- Promover la diferenciación de osteoblastos desde células madre mesenquimales
- Estimular la proliferación de pre-osteoblastos
- Mejorar la supervivencia de osteoblastos y osteocitos.
4.1.2 Mecanismos biomecánicos
En la actualidad, la respuesta mecánica está aceptada como un importante mecanismo regulatorio para
mantener la masa ósea. El punto de vista más comúnmente extendido es el de que los osteocitos actúan
como mecanotransductores. Los osteocitos y las células de borde son las células más abundantes en
el hueso. Los osteocitos están distribuidos a través del hueso y tienen la habilidad de comunicarse con
otras células óseas. Las células de borde cubren más del 90% de la superficie del hueso de un adulto
y mantiene conexiones con osteoblastos y osteocitos. Los osteocitos realizan la transducción de
señales bioquímicas en respuesta a los estímulos mecánicos. Éstos no responden directamente al
esfuerzo mecánico sino que lo hace indirectamente por el flujo de fluido extracelular provocado por
la carga. Además de la red de osteocitos, los osteoblastos también responden al estímulo mecánico
incluso en ausencia de osteocitos secretando mensajeros y expresando numerosos factores de
crecimiento. Los osteoclastos también se ven afectados por el estímulo mecánico. Cuando se aplica
una cierta carga, el banco de osteoclastos precursores reduce la expresión de RANKL, reduciéndose
el número de osteoclastos activos.
El modelo en cuestión incluye un mecanismo regulatorio biomecánico que desencadena respuestas en
las células óseas. Este modelo utiliza un novedoso enfoque basado en la homogeneización
micromécanica multiescala de la rigidez ósea. Esto permite calcular las tensiones a nivel de la matriz
ósea extravascular que aloja osteocitos. Éstos convierten la señal micromecánica en señales
bioquímicas para las células óseas en el espacio vascular produciendo una respuesta biomecánica a la
remodelación ósea.
La respuesta bioquímica al estímulo mecánico (mecanotransducción) implica una serie de sucesos que
incluye señalización ATP y Ca2+, liberación de prostaglandina y óxido nítrico y señalización Wnt/Lrp5
los cuáles resultan en la formación de hueso.

4. MODELO MATEMÁTICO
21
Figura 14
Mecanotransducción
- Sobreuso mecánico
Los osteocitos responden de manera indirecta a los esfuerzos mecánicos (deformación) del
hueso. Una acción biomecánica sobre el hueso provoca la apoptosis de los osteocitos de
manera que no segregan esclerostina (una proteína que inhibe a Wnt, incrementándose los
niveles de RANKL/OPG). Al no segregar esclerostina se incrementan los niveles de Wnt y
se produce el inicio del ciclo remodelador por diferenciación de preosteoblastos a osteoblastos
activos.
- Desuso mecánico
De lo contrario, ante el desuso mecánico se incrementa el ratio RANKL/OPG elevándose la
resorción de hueso. Esto ocurre porque no hay estimulo mecánico que inhiba la producción
de esclerostina por parte de los osteocitos de manera que la producción de esclerostina se

4. MODELO MATEMÁTICO
22
traduce en una señal inhibitoria a las células de borde por la cual las BMUs no son activadas.
El estímulo mecánico puede cuantificarse mediante un valor escalar que representa la densidad de
energía de deformación.
4.1.3 Mecanismos geométricos
La idea de que la morfología del hueso, es decir, de que su estructura geométrica influye de alguna
forma en la regulación de las poblaciones celulares no adquiere gran importancia en las últimas
publicaciones sobre remodelación ósea. No obstante, dicho modelo tiene en cuenta que el factor
geométrico influye de manera considerable en el proceso de remodelación ósea y para ello incluye dos
factores geométricos en el modelo.
Para los procesos bioquímicos, la morfología de los poros juega un papel muy importante además de
determinar la superficie específica (uno de los principales factores geométricos implicados en la
remodelación ósea). Las células óseas requieren una superficie para realizar sus funciones así como
iniciar el proceso de remodelación o resorber y formar tejido óseo.
Los parámetros geométricos y morfológicos más importantes son la porosidad vascular (fvas) y la
superficie específica (Sv).
La porosidad vascular puede definirse como la fracción de volumen de poros vasculares por volumen
de tejido total.
𝑓𝑣𝑎𝑠 =𝑉𝑣𝑎𝑠
𝑉𝑇
(4. 1)
La fracción volumétrica de matriz ósea se define del mismo modo:
𝑓𝑏𝑚 =𝑉𝑏𝑚
𝑉𝑇
(4. 2)
La superficie específica de un material poroso, la cantidad de superficie ósea disponible en un volumen
representativo, se define como:
𝑆𝑉 =𝑆𝑝
𝑉𝑇
(4. 3)
Basándose en datos experimentales, Martin establece una importante relación entre la superficie
específica (Sv) del hueso y la porosidad vascular (fvas) representativa para los distintos tipos de hueso:
2 3 4 5( )V vas vas vas vas vas vasS f af bf cf df ef (4. 4)

4. MODELO MATEMÁTICO
23
Con esta ecuación, el modelo tiene en cuenta la regulación geométrica. La evolución de las
poblaciones celulares predice la evolución de la porosidad vascular debido a la resorción del hueso
por parte de los osteoclastos y a la formación de hueso por parte de los osteoblastos. La ecuación
también permite estimar cambios en la superficie específica asociados a cambios en la porosidad y
por último, se asume que este cambio en la disponibilidad de superficie ósea influye en la evolución
de las poblaciones celulares.
Figura 15 Posible
Posible efecto del factor geométrico en la evolución de la porosidad vascular en osteoporosis de acuerdo con la ecuación (4.4) de Martin. Las curvas punteadas indican el incremento lineal de la porosidad sin tener en cuenta la respuesta
geométrica mientras que la otra curva si tiene en cuenta dicha respuesta. En la parte superior se representa el hueso trabecular y en la inferior el hueso cortical.
4.2 Desarrollo del modelo matemático
4.2.1 Hipótesis consideradas en el modelo
A continuación se van a detallar las hipótesis que tiene en cuenta el modelo:
- Se consideran osteoblastos y osteoclastos en diferentes etapas de desarrollo.
- Las poblaciones de osteoblastos precursores (OBUs) y de osteoclastos precursores (OCUs) se
asumen constantes y no se consideran variables de estado. Se asume que las piscinas de
células precursoras, tanto de linaje osteoblástico como osteoclástico, se autorenuevan de
manera ilimitada.
- La tasa proliferación de preosteoblastos, POBp es asumida como un parámetro constante a
pesar de ser un factor que está regulado por múltiples factores como la administración de
hormonas tales como PTH, factores de crecimiento o señalización Wnt.

4. MODELO MATEMÁTICO
24
- El modelo asume constante la cantidad de factor estimulante de colonia de macrófagos, M-
CSF
4.2.2 Modelo matemático
Teniendo en cuenta los mecanismos de regulación bioquímicos, biomecánicos y geométricos, las
ecuaciones que gobiernan las densidades de las poblaciones celulares en el modelo pueden expresarse
como:
, , ,( ) ( )bm
u u u p p p pact act
TGF TGF
p OB V OB O OBuB u OB p OB V OB re OB pp
dOB g S D OB P OB g S D OB
dt
(4. 5)
,( )p p p are
TGF
a OB V OB OB p Op B a
dOB g S D OB A OB
dt
(4. 6)
, , ,( ) ( )u u u u p p P
MCSF RANKL RANKL
p OC V OC OC OC u OC V OCact ac OC Pt act
dOC g S D OC g S D OC
dt
(4. 7)
, ,( )p p P a a
RANKL TGF
a OC V OC OC P OC OC aact act
dOC g S D OC A OC
dt
(4. 8)
4.2.3 Parámetros del modelo
A continuación se van a describir uno a uno los parámetros que conforman este conjunto de
ecuaciones.
4.2.3.1 Poblaciones celulares
En el modelo, las poblaciones de preosteoblastos (OBP), osteoblastos activos (OBA), preosteoclastos
(OPP) y osteoclastos activos (OCA), medida en número de células por unidad de volumen son las
variables de estado del sistema.
4.2.3.2 Tasas de diferenciación, proliferación y apoptosis
- Los parámetros DOBu, DOBp, DOCu, DOCp representan la tasa de diferenciación de osteoblastos
precursores, preosteoblastos, osteoclastos precursores y preosteoclastos respectivamente.
- POBp es la tasa proliferación de preosteoblastos.
- AOBa y AOCa representan tasas de apoptosis de osteoblastos activos y osteocastos activos
respectivamente.

4. MODELO MATEMÁTICO
25
4.2.3.3 Osteoblastos y osteoclastos precursores
OBu y OCu son las poblaciones de osteoblastos y osteoclastos precursores que el modelo toma
como constantes.
4.2.3.4 Funciones de activación y represión
Las respuestas celulares como la diferenciación, proliferación y la apoptosis están relacionadas con
las interacciones de algunos receptores con sus ligandos. Alguna de estas interacciones son
estimulantes, mientras que otras son inhibidoras.
La manera de tener en cuenta las respuestas de las poblaciones celulares a la concentración de
determinadas moléculas es mediante la llamada ‘función de Hill’ que puede actuar de activador (si al
aumentar la concentración de la molécula considerada, también crece la población celular) o de
represor (a mayor concentración, decrece la población celular).
Activador:
𝛽𝜋𝑎𝑐𝑡,𝑐é𝑙𝑢𝑙𝑎𝑚𝑜𝑙é𝑐𝑢𝑙𝑎 = 𝛽
𝑚𝑜𝑙é𝑐𝑢𝑙𝑎
𝑘𝑎𝑐𝑡 + 𝑚𝑜𝑙é𝑐𝑢𝑙𝑎
(4. 9)
Represor:
𝛽𝜋𝑟𝑒𝑝,𝑐é𝑙𝑢𝑙𝑎𝑚𝑜𝑙é𝑐𝑢𝑙𝑎 = 𝛽
1
1 +𝑚𝑜𝑙é𝑐𝑢𝑙𝑎
𝑘𝑟𝑒𝑝
(4. 10)
Donde,
- 𝑘𝑎𝑐𝑡 es un coeficiente de activación (nº moléculas/V)
- 𝑘𝑟𝑒𝑝 es un coeficiente de represión (nº moléculas/V)
- 𝑚𝑜𝑙é𝑐𝑢𝑙𝑎 es la concentración de una determinada proteína u hormona (nº moléculas/V)
- β suele ser el nivel máximo de la tasa de diferenciación o apoptosis de un tipo de población
celular (unidad de tiempo)-1
RANKL-RANK
Las funciones activadoras relacionadas con el aumento de la población de preosteoclastos (por mayor
tasa de diferenciación de OCu) y de la población de osteoclastos activos (por mayor tasa de
diferenciación de OCp) debido a la unión RANKL a RANK ( , ,,u pact a
RANKL RANKL
OC OCct ) son:
𝐷𝑂𝐶𝑢𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑅𝐴𝑁𝐾𝐿 = 𝐷𝑂𝐶𝑝
𝑅𝐴𝑁𝐾𝐿
𝐾𝑂𝐶𝑅𝐴𝑁𝐾𝐿 + 𝑅𝐴𝑁𝐾𝐿
(4. 11)

4. MODELO MATEMÁTICO
26
𝐷𝑂𝐶𝑝𝜋𝑎𝑐𝑡,𝑂𝐶𝑝𝑅𝐴𝑁𝐾𝐿 = 𝐷𝑂𝐶𝑝
𝑅𝐴𝑁𝐾𝐿
𝐾𝑂𝐶𝑅𝐴𝑁𝐾𝐿 + 𝑅𝐴𝑁𝐾𝐿
(4. 12)
Siendo
𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑅𝐴𝑁𝐾𝐿 = 𝜋𝑎𝑐𝑡,𝑂𝐶𝑝
𝑅𝐴𝑁𝐾𝐿
Donde,
- 𝐾𝑂𝐶𝑅𝐴𝑁𝐾𝐿es la constante de disociación de la unión RANKL con su receptor RANK expresado en
OCuS Y OCpS.
- RANKL es la concentración libre de RANKL:
[ ] [ ]1
bm
RANKL RANKL
RANKLRANKL
RANKL OPG RANKL RANK
PRANKLeft
D RANK
K K
LeftRANKL
OPG RANK
(4. 13)
Donde,
- RANKLeft es la máxima concentración de RANKL que está regulada por la hormona
paratiroidea PTH,
, p
RANKL P
OBp p act
TH
OBRANKLeft N OB (4. 14)
- OPG es la concentración de moléculas OPG de señalización bioquímica
,
,
max
a
a
OBa rep
OBa re
OPG PTH
a OB
OPG PTH
a OBpOPG
OPG
DO
OB
PG
OB
(4. 15)
- RANK es la concentración de moléculas de receptor RANK
RANK
OCp pRANK N OC (4. 16)

4. MODELO MATEMÁTICO
27
TGFβ – OBu/ OBp/Oca
Las funciones de activación de la unión TGF-β con sus receptores ( , u
TGF
OBact
, , a
TGF
OCact
) son:
, uOBu act OBu t
TGF
OB gf
OBu
TGFD D
k TGF
(4. 17)
𝐴𝑂𝐶𝑎𝑂𝐶𝑎𝜋𝑎𝑐𝑡,𝑂𝐶𝑎𝑇𝐺𝐹𝛽
= 𝐴𝑂𝐶𝑎𝑂𝐶𝑎
𝑇𝐺𝐹 − 𝛽
𝑘𝑂𝐶𝑎𝑡𝑔𝑓𝛽
+ 𝑇𝐺𝐹 − 𝛽
(4. 18)
Como 𝑘𝑂𝐶𝑎𝑡𝑔𝑓𝛽
= 𝑘𝑂𝐵𝑢𝑡𝑔𝑓𝛽
, 𝜋𝑎𝑐𝑡,𝑂𝐶𝑎𝑇𝐺𝐹𝛽
= 𝜋𝑎𝑐𝑡,𝑂𝐵𝑢𝑇𝐺𝐹𝛽
La función de represión de la unión TGF-β con su receptor , p
TGF
OBrep
es:
,
1
1pOBp p rep OBp p
tgf
OBp
TGF
OBD OB D OBTGF
k
(4. 19)
Donde
res a TGF
TGF
k OC STGF
D
(4. 20)
4.2.3.5 Regulación geométrica
La regulación geométrica es incluida en el modelo mediante las funciones gcélula (SV), dependientes de
la superficie específica. Estas funciones incluyen la regulación geométrica en varios estados de
desarrollo de osteoblastos y osteoclastos. Se distinguen dos tipos de acción geométrica:
A. Inicio de la remodelación, creación de una nueva BMU
Cuando se inicia un nuevo evento de remodelación se crea una nueva BMU. Se piensa que los
primeros pasos para la creación de una nueva BMU es el reclutamiento de preosteoblastos y
preosteoclastos en la superficie ósea. Dicho reclutamiento está controlado por los osteocitos que
comienzan a actuar por estimulo mecánico. La disponibilidad de superficie ósea presenta una
compleja dependencia del reclutamiento de preosteoblastos y osteoclastos que es modelada por
las funciones gOBu (SV) y gOCu (SV).
B. Modelado de la resorción y formación en una BMU existente

4. MODELO MATEMÁTICO
28
La maduración, es decir, la diferenciación de preosteoblastos y preosteoclastos en osteoblastos y
osteoclastos activos respectivamente depende de la disponibilidad de superficie ósea ya que, tanto
los osteoblastos como los osteoclastos se activan sólo si disponen de una superficie sobre la cual
realizar las funciones para la que han sido activadas, resorber o formar tejido óseo. Esta
dependencia está modelada por las funciones gOpu (SV) y gOCp (SV). Además, estas funciones
determinan cuantos de los osteoblastos y osteoclastos de una BMU están remodelando el hueso y
la cantidad de hueso resorbido y formado en una BMU.
En la actualidad no existe una determinación experimental de la influencia de la superficie específica,
SV, en el desarrollo de las poblaciones celulares óseas por lo que la forma específica de dichas
funciones es desconocida. A pesar de desconocer la función exacta que gobierna la regulación
geométrica se puede afirmar que la diferenciación celular aumenta con SV y que hay que tener en
cuenta no linealidades debido a los complejos mecanismos microscópicos que intervienen en el
proceso. Teniendo en cuenta estas asunciones, las funciones se pueden modelar siguiendo una ley de
potencia:
𝑔𝑐é𝑙𝑢𝑙𝑎(𝑆𝑉) = (𝑆𝑣
𝑆𝑣(𝑡0))
𝐾𝑐é𝑙𝑢𝑙𝑎
(4. 21)
Dónde,
𝐾𝑐é𝑙𝑢𝑙𝑎 ≥ 0, célula = OBu, OBp, OCu, OCp
𝑆𝑣(𝑡0) es la superficie específica en un estado de remodelación ósea estable, sin ganancia ni pérdida
de volumen.
De esta manera, en un estado estacionario se cumple que gcélula=1. Si deseamos liberar al modelo de la
regulación geométrica se elegirá 𝐾𝑐é𝑙𝑢𝑙𝑎 = 0
4.2.3.6 Regulación biomecánica
La densidad de energía de deformación es un factor importante que determina la capacidad de
adaptación del hueso a distintas cargas mecánicas.
La densidad de energía de deformación microscópica 𝜓𝑏𝑚incrementa la proliferación de
preosteoblastos a consecuencia de la muerte de osteocitos y el correspondiente descenso de
esclerostina. El modelo incluye esta relación entre el estímulo mecánico y la auto-expansión de
preosteoblastos mediante la siguiente función de activación:

4. MODELO MATEMÁTICO
29
*
*
1( ),
2
11 , ( ) ,
2 2 ( )
,
0, ,
bm bm o
bmbm o bm bm
bm obm
p
bm bm
t
tt
act OB
(4. 22)
Donde,
λ es la constante que cuantifica la transducción biomecánica en el caso de sobreuso
𝜓𝑏𝑚(t0) es el valor de 𝜓𝑏𝑚en el estado estacionario
𝜓𝑏𝑚∗ es el umbral a partir del cual se cumple que П𝑎𝑐𝑡,𝑂𝐵𝑝
𝜓𝑏𝑚 = 1

5. ESTABILIDAD DEL SISTEMA
30
5 ESTABILIDAD DEL SISTEMA
En este apartado se va a analizar la estabilidad del sistema matemático que gobierna el
comportamiento de las poblaciones celulares en el hueso. Nos enfrentamos a un sistema de ecuaciones
no lineales por lo que la estabilidad del sistema deberá ser calculada mediante linealización.
Para analizar la estabilidad del modelo matemático que propone Pivonka se van a tomar las constantes
que conforman el modelo y se va a analizar cuán estable es el sistema si se toma una de las constantes,
sin modificar el resto de ellas, y se varía su valor.
En los siguientes subapartados se va a desarrollar paso a paso la linealización del sistema con sus
cálculos pertinentes para poder determinar su estabilidad.
5.1 Fundamentos de la linealización
El cálculo de la estabilidad del sistema mediante linealización se ha basado en el Teorema de
Linealización de Liapunov y Poincaré.
Se considera el sistema autónomo 5.1,
( , )
( , )
dxF x y
dtdy
G x ydt
(5. 1)
con un punto crítico en (x0, y0), tal que las funciones F(x, y) y G(x, y) sean de clase C1 (R2). Entonces,
aproximando z = F(x, y) y z = G(x, y) (cerca del punto (x0, y0)) por sus respectivos planos tangentes
en dicho punto,
0 0 0 0 0 0
0 0 0 0 0 0
( , ) ( , ).( ) ( , ).( )
( , ) ( , ).( ) ( , ).( )
F FF x y x y x x x y y y
x y
G GG x y x y x x x y y y
x y
(5. 2)

5. ESTABILIDAD DEL SISTEMA
31
Se puede escribir
0
0
( , )
( , )
x xF x yA
y yG x y
Donde A es la matriz jacobiana del campo (F(x, y), G(x, y))t en el punto (x0, y0), es decir,
0 0 0 0
0 0 0 0
( , ) ( , )
( , ) ( , )
F Fx y x y
x yA
G Gx y x y
x y
(5. 3)
De esta manera, el sistema 5.1 se encuentra próximo al sistema lineal.
Teorema 5.1.1 (Teorema de Linealización de Liapunov y Poincaré)
1. El punto crítico del sistema es asintóticamente estable si y sólo si todos los
autovalores de la matriz A poseen parte real negativa.
2. El punto crítico del sistema es inestable si y sólo si la matriz A del sistema posee
un autovalor con parte real positiva.
Más aún, si los autovalores de A son distintos entre sí y distintos de cero se puede
decir lo siguiente:
1. Si λ1 < λ2 < 0, entonces (x0, y0) es un nodo asintóticamente estable.
2. Si λ1 > λ2 > 0, entonces (x0, y0) es un nodo inestable.
3. Si λ1 < 0 < λ2, entonces (x0, y0) es un punto de silla.
4. Si λ1 no es real y Re (λ1) < 0, entonces (x0, y0) es un foco asintóticamente
estable.
5. Si λ1 no es real y Re (λ1) > 0, entonces (x0, y0) es un foco inestable.

5. ESTABILIDAD DEL SISTEMA
32
5.2 Particularización de la linealización
En este apartado se incluyen los diferentes pasos que se han dado hasta poder determinar si el sistema
es estable o inestable.
En primer lugar se presenta el sistema de ecuaciones con el que se va a trabajar en función de las
diferentes poblaciones celulares que intervienen en el modelo. En segundo lugar se calculan sus puntos
estacionarios. A continuación se obtiene la matriz jacobiana del sistema en dichos puntos
estacionarios. Finalmente se calculan los autovalores de dicha matriz y se evalúa su parte real, siendo
estable el sistema si la parte real de todos los autovalores es negativa.
Figura 16
Diagrama de flujo para determinar la estabilidad del sistema
Las ecuaciones del modelo matemático a linealizar son las ecuaciones (4.5), (4.6), (4.7) y (4.8).
5.2.1 Parámetros del modelo en función de OBp, Oba, OCp y OCa
Para facilitar los cálculos, acortando el nombre de las distintas variables, se ha asignado una letra a
cada variable. En la siguiente tabla se recoge la lista de las distintas variables con sus letras
correspondientes:
SISTEMA DE ECUACIONES
CÁLCULO PTOS ESTACIONARIOS
DEL SISTEMA
CÁCULO MATRIZ
JACOBIANA
CÁLCULO AUTOVALORES
PARTE REAL AUTOVALORES
•S. ESTABLE-
•S. INESTABLE+

5. ESTABILIDAD DEL SISTEMA
33
LETRA CTE LETRA CTE
a1 gOBu (Sv) n1 KD1,TGFβ
a2 gOBp (Sv) n2 𝑘𝑂𝐵𝑝𝑡𝑔𝑓𝛽
a3 gOcu (Sv) n3=n1 𝑘𝑂𝐶𝑎𝑡𝑔𝑓𝛽
a4 gOCp (Sv) p 𝑘𝑂𝐶𝑟𝑎𝑛𝑘𝑙
b1 DOBu q βrankl
b2 DOBp r1 �̃�RANKL
b3 DOCu r2 �̃�PTH
b4 DOCp r3 �̃�OPG
c1 OBU s1 K[RANKL-OPG]
c2 OCU s2 K[RANKL-RANKL]
f ,bm
pOa Bct
t1 𝑁𝑂𝐵𝑝
𝑟𝑎𝑛𝑘𝑙
g 𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑀𝐶𝑆𝐹 t2 𝑁𝑂𝐶𝑝
𝑟𝑎𝑛𝑘
h1 AOBa u 𝑃𝑅𝐴𝑁𝐾𝐿𝜑𝑏𝑚
h2 AOCa v1 βPTH
i POBp v2 β𝑂𝐵𝑎𝑜𝑝𝑔
j 𝜂𝑡𝑔𝑓 𝛽𝑏𝑜𝑛𝑒 w1 𝑘𝑂𝐵,𝑎𝑐𝑡
𝑝𝑡ℎ
k KRES w2 𝑘𝑂𝐵,𝑟𝑒𝑝𝑝𝑡ℎ
l STGF-β x PPTH
m1 �̃�TGF-β y βOPGmax
Tabla 1
Las ecuaciones del modelo están conformadas por distintos parámetros de los cuáles unos son
constantes y otros dependen de OBp, OBa, OCp y OCa. Los parámetros que no son constantes deben
ser desarrollados para poder integrarlos en las ecuaciones del modelo y realizar las correspondientes

5. ESTABILIDAD DEL SISTEMA
34
derivadas.
A continuación se van a desarrollar los parámetros del modelo que dependen de OBp, OBa, OCp y
OCa:
i. Funciones de activación/ represión de la unión TGF-β con sus receptores ( , u
TGF
OBact
, , a
TGF
OCact
,
, p
TGF
OBrep
):
Funciones de activación,
Tomando la ecuación (4.17) y (4.20) se obtiene:
, u
res a TGF
act tgfTGFO
TGF
OB
Bu res a TGF
k OC S
k D k OC S
(5. 4)
, ,a u
TGF TGF res a TGF
act OC act tgfTGFOBu r
O
es a TGF
B
k OC S
k D k OC S
(5. 5)
Poniendo las ecuaciones en función de cómo se han nombrado en la tabla,
, ,
1 1a u
TGF TGF
OBa
act OC act
a
jkOC l
n m jkOC l
(5. 6)
Funciones de represión,
Tomando la ecuación (4.19) y (4.20) se obtiene:
, p
tgfTGFOBp
rep tgfTGFO
TGF
OB
Bp res a TGF
k D
k D k OC S
(5. 7)
Poniendo la ecuación en función de cómo se ha nombrado en la tabla,
2 1,
2 1p
T
rep
a
GF
OB
n m
n m jkOC l
(5. 8)

5. ESTABILIDAD DEL SISTEMA
35
ii. Función de activación de la unión RANKL con su receptor ( , ,,u pact a
RANKL RANKL
OC OCct ):
Teniendo en cuenta las ecuaciones (4.11) y (4.12) donde RANKL se define por la ecuación
(4.13). Los términos de ésta ecuación,
- RANKLeft es la máxima concentración de RANKL que está regulada por la hormona
paratiroidea PTH y queda definida por la ecuación (4.14):
, p
RANKL P
OBp p act
TH
OBRANKLeft N OB
En esta ecuación la función de activación de la unión PTH con su receptor, , p
PTH
OBact es:
,
,p
PTH
OBact pth
OB act
PTH
k PTH
(5. 8)
siendo
PTH PTH
PTH
PPTH
D
(5. 9)
Y el parámetro RANKL
OBpN es el número máximo de moléculas RANKL que pueden ser
expresadas por un único preosteoblasto.
- OPG es la concentración de moléculas OPG de señalización bioquímica que queda
definida por la ecuación (4.15)
- RANK es la concentración de moléculas de receptor RANK que queda definida por la
ecuación (4.16)
Introduciendo las expresiones RANKLeft, OPG y RANK en la ecuación del ligando RANKL y
renombrando las distintas variables como indica la tabla 1 para facilitar el manejo de las expresiones,
11
2 1 1 11
2 1 1
2 2 21 2 2
2 2 2 3 1 2 2
1( )
p
p
ap
a
v x q ut OB
r w v x v xq r OB
r w v xRANKL
yv r w OBS S t OC
v r w OB yr v x r w
(5. 10)

5. ESTABILIDAD DEL SISTEMA
36
Finalmente, considerando las expresiones anteriores y nombrando las distintas constantes como indica
la tabla 1 para su simplificación, la función de activación de la unión RANKL con su receptor,
, ,u p
RANKL
act ac
RANKL
OC OCt , se puede expresar del siguiente modo:
11
2 1 1 11
2 1 1
2 2 211 1 2 2
2 1 1 2 2 2 3 1 2 211
2 1 1
,
1( )
u
p
p
RANKL
OC
a
ac
p
p
t
p
a
v x q ut OB
r w v x v xq r OB
r w v x
yv r w OBv x q ut OB p S S t OC
r w v x v r w OB yr v x r wv xq r OB
r w v x
(5. 11)
5.2.2 Ecuaciones del modelo en función de OBp, OBa, OCp y OCa
Las ecuaciones del sistema matemático que gobierna el comportamiento de las poblaciones celulares
en el hueso están formadas por varios parámetros. Unos son constantes, mientras que otros son
dependientes de las concentraciones de las distintas poblaciones celulares OBp, OBa, OCp y OCa.
Tras desarrollar en el apartado anterior las expresiones de dichos parámetros dependientes ( , u
TGF
OBact
,
, a
TGF
OCact
, , p
TGF
OBrep
, , ,,u pact a
RANKL RANKL
OC OCct ) podemos obtener las ecuaciones del modelo en función de OBp,
OBa, OCp y OCa. Como se ha hecho con el resto de ecuaciones, éstas se presentan renombrando las
distintas variables como indica la tabla 1.

5. ESTABILIDAD DEL SISTEMA
37
1 12
2 21
1 1 2 1
aup p p
a a
da b OB if a b
jk l n m
n m jk l n m jk l
OCOB OB OB
OC Cd Ot
(5. 12)
22 1
2
2
1
1a p a
a
n m
n m jkOB OB OB
da b h
dt C lO
(5. 13)
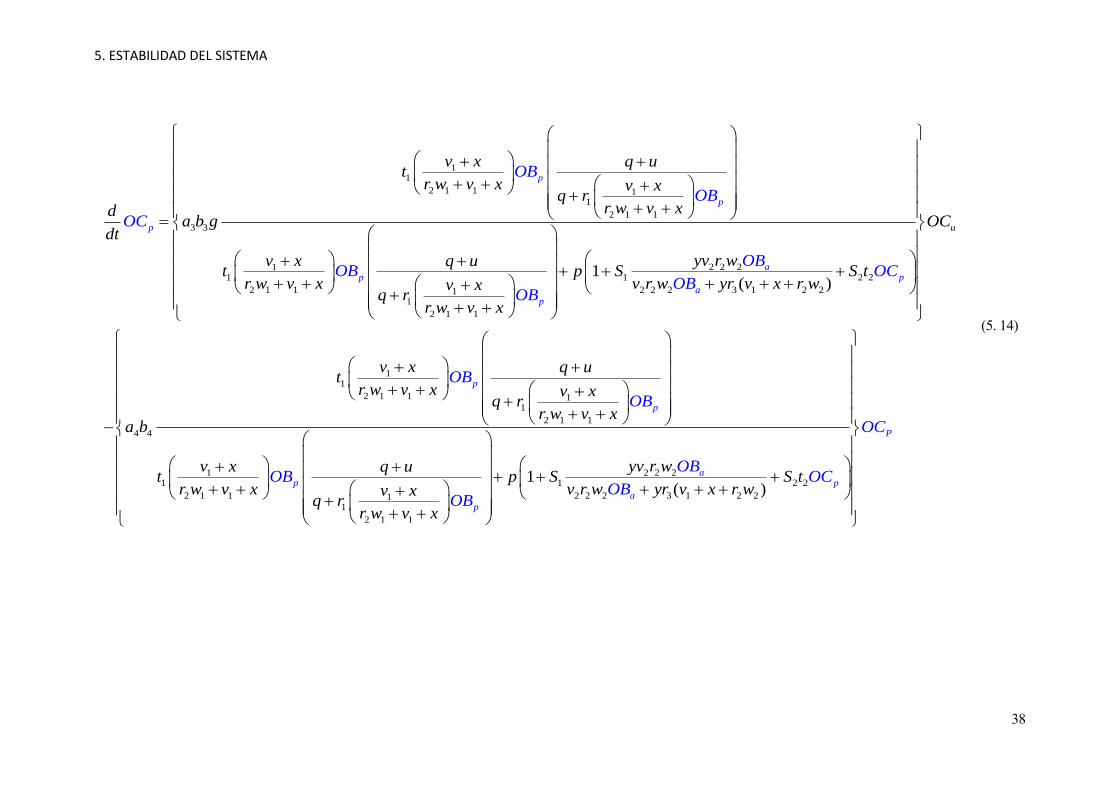
5. ESTABILIDAD DEL SISTEMA
38
11
2 1 1 11
2 1 1
3 3
2 2 211 1 2 2
2 1 1 2 2 2 3 1 2 211
2 1 1
1( )
p
p
p
ap p
a
p
OB
OB
OC
OBOB O
v x q ut
r w v x v xq r
r w v xda b g
dt
yv r wv x q ut p S S t
r w v x v r w yr v x r wv xq r
r w v
COB
OBx
11
2 1 1 11
2 1 1
4 4
2 2 211 1
2 1 1 2 2 2 3 111
2 1 1
1(
p
p
ap
p
u
a
OC
v x q ut
r w v x v xq r
r w v xa b
yv r wv x q ut p S
r w v x v r w yr vv xq r
OB
OB
OBOB
O
r v x
BOB
w
2 2
2 2 )
P
p
OC
S tx w
OCr
(5. 14)

5. ESTABILIDAD DEL SISTEMA
39
11
2 1 1 11
2 1 1
4 4
2 2 211 1 2 2
2 1 1 2 2 2 3 1 2 211
2 1 1
1( )
p
p
a
ap p
a
p
OB
OB
OC
OBOB OC
OBO
v x q ut
r w v x v xq r
r w v xda b
dt
yv r wv x q ut p S S t
r w v x v r w yr v x r wv xq r
r w v xB
1 1
2
P
aa
a
h
OC
OCOC
OC
jk l
n m jk l
(5. 15)

5. ESTABILIDAD DEL SISTEMA
40
5.2.3 Cálculo del punto estacionario
En este apartado se explica cómo se calculan los puntos estacionarios del sistema. Para su cálculo se
ha implementado una función con la herramienta de software matemático MATLAB. Los puntos
estacionarios del sistema van a cambiar en función de la constante que se esté analizando, es decir,
que se esté variando.
Para el cálculo del punto estacionario se anulan todas las derivadas parciales del sistema y se despejan
los valores de OBp, OBa, OCp y OCa.
El sistema a resolver por la herramienta MATLAB para cada una de las constantes que se va a analizar
es el siguiente:
1 1 2 22 1
1 1 2 1
0 ap
a
u p
a
a b OB ifOC
OBjk l n m
n m jk l n m jk la b OB
OC OC
(5. 16)
2 1
1
2
2
2 10 p a
a
OB OBOC
n m
n m k la b h
j
(5. 17)
11
2 1 1 11
2 1 1
3 3
2 2 211 1 2 2
2 1 1 2 2 2 3 1 2 211
2 1 1
0
1( )
p
p
ap p
a
p
v x q ut
r w v x v xq r
r w v xa b g
yv r wv x q ut p S S t
OB
OB
OBOB OC
OBOB
r w v x v r w yr v x r wv xq r
r w v x
11
2 1 1 11
2 1 1
4 4
2 2 211 1
2 1 1 2 2 2 3 1 211
2 1 1
1(
u
p
p
ap
a
p
OB
O
OC
v x q ut
r w v x v xq r
r w v xa b
yv r wv x q ut p S
r w v x v r w yr v x rv
B
OBOB
OBxq r
r w v xOB
2 2
2 )
P
p
OC
S Ctw
O
(5. 18)

5. ESTABILIDAD DEL SISTEMA
41
11
2 1 1 11
2 1 1
4 4
2 2 211 1 2 2
2 1 1 2 2 2 3 1 2 211
2 1 1
0
1( )
p
p
ap p
a
p
v x q ut
r w v x v xq r
r w v xa b
yv r wv x q ut p S S t
r w v x v r w yr v x r wv xq
O
rr w
B
OB
OBOB OC
v
OBB
xO
1 1
2
P
aa
a
jk l
n m j
OC
OCOC
C lh
kO
(5. 19)
5.2.4 Cálculo de la matriz jacobiana
En este apartado se va a calcular la matriz jacobiana del sistema de ecuaciones que gobierna el
comportamiento matemático de las poblaciones celulares. Éste cálculo se hace necesario puesto que
la estabilidad del sistema se evalúa en función de los valores de la parte real de los autovalores de
dicha matriz.
La matriz jacobiana está formada por las derivadas parciales de las distintas ecuaciones del modelo,
calculadas en su punto estacionario. Los puntos estacionarios del sistema de ecuaciones, como se ha
explicado en el apartado anterior, van variando en función de la constante que se esté analizando. La
matriz jacobiana varía del mismo modo. Para cada variación de la constante que está siendo analizada
se tendrá una matriz jacobiana.
El cálculo de las derivadas parciales que conforman la matriz jacobiana se ha realizado sin emplear
ninguna herramienta de cálculo matemático. Posteriormente, al igual que se ha procedido para calcular
los puntos estacionarios, los resultados de las derivadas parciales se han integrado en un programa de
cálculo utilizando MATLAB. Dichas derivadas parciales son función de los puntos estacionarios OBp,
OBa, OCp y OCa.
La matriz jacobiana a calcular es la siguiente:

5. ESTABILIDAD DEL SISTEMA
42
0 00 0
0 00 0
0 0
0
p p p p
p a p a
a a a a
p a p a
P P
p a
aa
ap
fOB fOB fOB fOB
OB OB OC OC
fOB fOB fOB fOBJ
OB OB OC OC
fOC fOC
OB OB
fOCfOC
OBOB
0 0
0 00
P P
p a
aa
ap
fOC fOC
OC OC
fOCfOC
OCOC
(5. 20)
A continuación se van a calcular las distintas derivadas parciales que conforman dicha matriz. Los
cálculos se van a realizar fila a fila.
I. Fila 1
2 1
2 1
2 2
0 a
p
p
fOBif a b
O
n m
n m kB Oj lC
(5. 21)
0
0p
a
fOB
OB
(5. 22)
0
0p
p
fOB
OC
1 1 2 1
1 1
1 1 1 2 22 2
0 2 1
( )( )a ap
a a
p
a
n m jk l jk jk l n m jk
n m
fOB jka b c a b
OC
jk l n m jk
OCOB
OC lC OCO
(5. 23)

5. ESTABILIDAD DEL SISTEMA
43
II. Fila 2
2 1
2 1
2 2
0
a
p a
fOBa b
O
n m
n m jk lCB O
(5. 24)
1
0
a
a
fOBh
OB
(5. 25)
0
0a
p
fOB
OC
(5. 26)
2 1
2 2 2
2 10
a
a
p
a
n m jk
n m jOB
O
fOB
k la
OC Cb
(5. 27)
III. Fila 3
( ) ( )
3 3 2 2
0
( ) ( )
4 4 2
AD AD PCBAD PCB AD
OB OB OBfOC p p pP a b c gOB AD PCBp
AD AD PCBAD PCB AD
OB OB OBp p p
a b OCp
AD PCB
(5. 28)

5. ESTABILIDAD DEL SISTEMA
44
0
( ) ( )
3 3 2 2
( ) ( )
4 4 2
P
a
AD AD PCBAD PCB AD
OB OB OBfOC a a aa b c gOB AD PCB
AD AD PCBAD PCB AD
OB OB OBa a aa b OC
pAD PCB
(5. 29)
0
( ) ( )
3 3 2 2
( ) ( )
4 4 2
P
p
p
AD AD PCBAD PCB AD
OC OC OCfOC p p p
a b c gOC AD PCB
PCBAD AD PCB AD OC
OCp
a bAD PCB
(5. 30)
0
0P
a
fOC
OC
(5. 31)
IV. Fila 4
0
( ) ( )
4 4 2a
p
AD AD PCBAD PCB AD
OB OB OBfOC p p p
a b OCpOB AD PCB
(5. 32)
0
( ) ( )
4 4 2a
a
AD AD PCBAD PCB AD
OB OB OBfOC a a aa b OCpOB AD PCB
(5. 33)

5. ESTABILIDAD DEL SISTEMA
45
0
( ) ( )
4 4 2
p
a
p
PCBAD AD PCB AD OC
OCfOC p
a bOC AD PCB
(5. 34)
2
1 1
2 2
0 1 1
2 a a a aa
a a
jkOC l n m jkOC l jkOC lOC jkfOCh
OC n m jkOC l
(5. 35)
Los cálculos necesarios para el desarrollo de las derivadas parciales de las filas 3 y 4 son bastante
largos y complejos. Para simplificar los cálculos y por cuestiones de espacio se han creado los términos
AD y PCB con sus correspondientes derivadas. A continuación se definen éstos términos:
1 13 1 1 2 2 3 1 1 2 2
2 1 1 2 1 1
1 12 2 2 1 2 2 2 1
2 1 1 2 1 1
p
p a
v x v xAD qyr t v x r w yr ut v x r w OB
r w v x r w v x
v x v xqv r w t uv r w t OB OB
r w v x r w v x
(5. 36)
11 3 1 1 2 2
2 1 1
2 2 2 1 2 2 2 2 2 3 1 2 2
1 11 2 2 2 1 1 1 2 2 2 1
2 1 1 2 1 1
2 2 2 2 2 1 3
p
a p
p a
v xPCB pyr r t v x r w OB
r w v x
pqv r w pqys v r w OB pqs t yr v x r w OC
v x v xprv r w t pr s yv r w t OB OB
r w v x r w v x
pqs t v r w pyr r s
12 2 1 1 2 2
2 1 1
11 2 2 2 2 2 1 3 1 2 2
2 1 1
p p
p a p
v xt t v x r w OB OC
r w v x
v xpr s t v r w t OB OB OC pqyr v x r w
r w v x
(5.37)
Derivadas de AD con respecto a OBp, OBa y OCp:

5. ESTABILIDAD DEL SISTEMA
46
1 12 2 2 1 2 2 2 1
2 1 1 2 1 1
1 13 1 1 2 2 3 1 1 2 2
2 1 1 2 1 1
a
p
v x v xADqv r w t uv r w t OB
OB r w v x r w v x
v x v xqyr t v x r w yr ut v x r w
r w v x r w v x
(5. 38)
1 12 2 2 1 2 2 2 1
2 1 1 2 1 1
p
a
v x v xADqv r w t uv r w t OB
OB r w v x r w v x
(5. 39)
0p
AD
OC
(5. 39)
Derivadas de PCB con respecto a OBp, OBa y OCp:
1 11 2 2 2 1 1 1 2 2 2 1
2 1 1 2 1 1
12 2 2 2 2 1 3 2 2 1 1 2 2
2 1 1
11 2 2 2 2 2 1
2 1 1
a
p
p
a p
v x v xPCBprv r w t pr s yv r w t OB
OB r w v x r w v x
v xpqs t v r w pyr r s t t v x r w OC
r w v x
v xpr s t v r w t OB OC
r w v x
11 3 1 1 2 2
2 1 1
v xpyr r t v x r w
r w v x
(5. 40)
1 11 2 2 2 1 1 1 2 2 2 1
2 1 1 2 1 1
11 2 2 2 2 2 1 2 2 2 1 2 2 2
2 1 1
p
a
p p
v x v xPCBprv r w t pr s yv r w t OB
OB r w v x r w v x
v xpr s t v r w t OB OC pqv r w pqys v r w
r w v x
(5. 41)

5. ESTABILIDAD DEL SISTEMA
47
12 2 2 2 2 1 3 2 2 1 1 2 2
2 1 1
11 2 2 2 2 2 1 2 2 3 1 2 2
2 1 1
p
p
p a
v xPCBpqs t v r w pyr r s t t v x r w OB
OC r w v x
v xpr s t v r w t OB OB pqs t yr v x r w
r w v x
(5. 42)

6. ANÁLISIS DE VARIABLES
48
6 ANÁLISIS DE VARIABLES
En este trabajo se ha realizado un análisis de las constantes del modelo de comportamiento de las
poblaciones celulares en el proceso de remodelación ósea que propone Pivonka. En cada análisis se
estudia la estabilidad del sistema. Se estudia de qué manera queda afectado el sistema al variar el valor
de las constantes.
6.1 Simplificaciones tenidas en cuenta para el análisis
Con objeto de simplificar el análisis de variables no se han tenido en cuenta todos los mecanismos de
regulación del proceso de remodelación ósea que tiene en cuenta Pivonka en su modelo.
6.1.1 Regulación geométrica
Se va a considerar que nos encontramos en un estado de remodelación ósea estable, es decir, sin
ganancia ni pérdida de volumen. De este modo se cumple que:
𝑆𝑣(𝑡0) = 𝑆𝑣
De manera que,
𝑔𝑐é𝑙𝑢𝑙𝑎(𝑆𝑉) = (𝑆𝑣
𝑆𝑣(𝑡0))
𝐾𝑐é𝑙𝑢𝑙𝑎
= 1 (6. 1)
6.1.2 Regulación biomecánica
Para realizar dicho análisis no se va a considerar el estímulo mecánico de manera que:
П𝑎𝑐𝑡,𝑂𝐵𝑝𝜓𝑏𝑚 = 1
(6. 2)
6.1.3 Función de activación de la diferenciación de OCu en OCp
El modelo asume constante la cantidad de factor estimulante de colonia de macrófagos, M-CSF, por
lo que la función que regula la diferenciación de osteoclastos precursores a preosteoclastos, 𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑀𝐶𝑆𝐹 ,
es constante.

6. ANÁLISIS DE VARIABLES
49
6.1.4 Tasa de producción de RANKL por preosteoblastos
La tasa de producción de RANKL por los preosteoblastos, 𝑃𝑅𝐴𝑁𝐾𝐿𝜓𝑏𝑚 , se ve favorecida durante el desuso
mecánico:
1 , ( )( )
0, ( )
bmbm bm o
bm obm
bm bm o
k tt
RANKL
t
P
(6. 3)
Para el análisis de las variables se asume que la densidad de energía de deformación microscópica es
mayor que el valor de la misma en estado estacionario, 𝜓𝑏𝑚 > 𝜓𝑏𝑚(𝑡0). Por tanto,
𝑃𝑅𝐴𝑁𝐾𝐿𝜓𝑏𝑚 = 1
6.2 Análisis de constantes del modelo
En este apartado se van a analizar las diferentes constantes que presenta el modelo. Para ello se va a
tomar una a una las constantes que conforman el modelo y se va a variar su valor entre [valor original
del parámetro]*10-p hasta [valor original del parámetro]*10p, siendo p=5 en intervalos de 0.5 (p=[-
5:0.5:5]). En las gráficas que se presentan se representa la parte real de los autovalores de la matriz
jacobiana explicada en el apartado de Linealización frente a la variación de p.
6.2.1 Poblaciones de osteoblastos y osteoclastos precursores
6.2.1.1 Población de osteoblastos precursores, OBu
El número de osteoblastos precursores se asume mucho mayor que el resto de las poblaciones
celulares. Al variar este parámetro el sistema permanece estable, ya que para todas las variaciones que
se han hecho, los valores de la parte real de los autovalores de la matriz jacobiana son negativos.

6. ANÁLISIS DE VARIABLES
50
GRÁFICA 1
6.2.1.2 Población de osteoclastos precursores, OCu
Para una variación entre [valor original del parámetro]*104 hasta [valor original del
parámetro]*105 el sistema se hace inestable. Los valores de las constantes para los que el sistema
se hace inestable están demasiados alejados del valor nominal. Esto hace pensar que el sistema se
hace inestable porque valores tan altos de la población de osteoclastos precursores no son
biológicamente factibles.
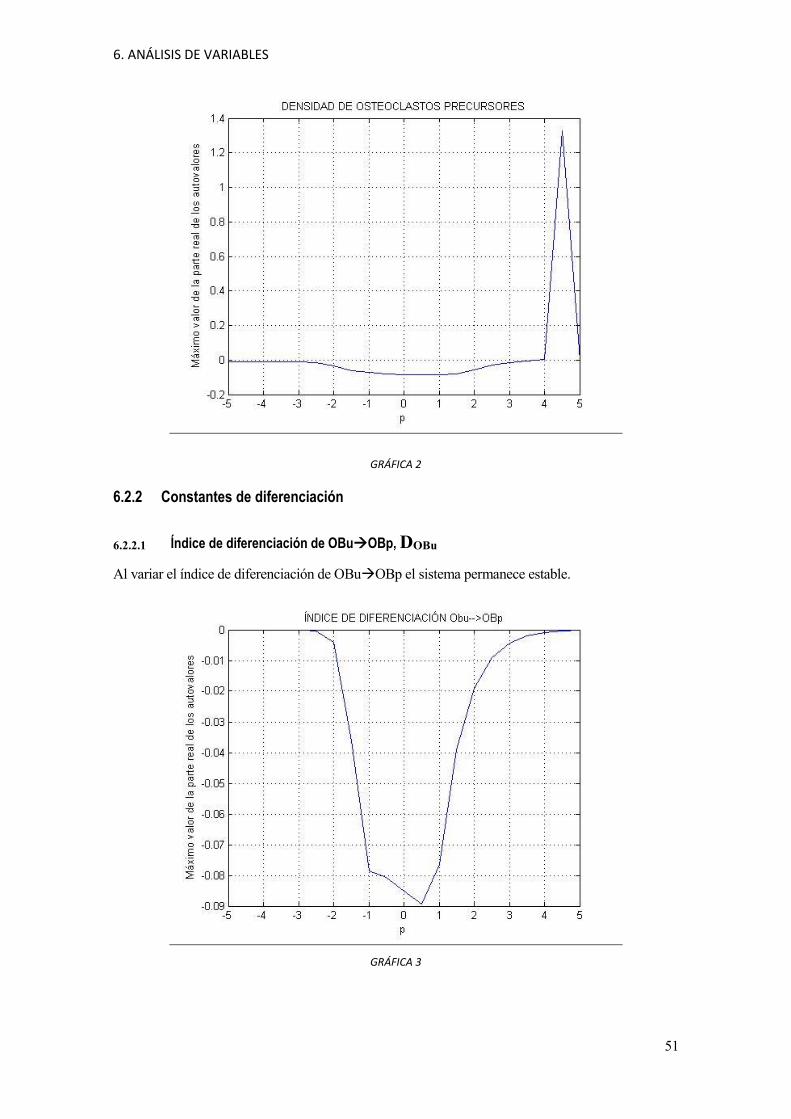
6. ANÁLISIS DE VARIABLES
51
GRÁFICA 2
6.2.2 Constantes de diferenciación
6.2.2.1 Índice de diferenciación de OBuOBp, DOBu
Al variar el índice de diferenciación de OBuOBp el sistema permanece estable.
GRÁFICA 3

6. ANÁLISIS DE VARIABLES
52
6.2.2.2 Índice de diferenciación de OBpOba, DOBp
Para una variación entre [valor original del parámetro]*10-5 hasta [valor original del
parámetro]*10-3 el sistema se hace inestable. Algunos valores de DOBp para los que el sistema
se hace inestable están muy próximos al valor nominal. Esto hace pensar que el sistema se hace
inestable porque el modelo falla o porque el valor de esta constante está muy ajustado y hay que
prestar mucha atención su valor exacto, ya que cualquier mínimo cambio en dicho valor hace
inestable el sistema. Para valores pequeños de DOBp la población de osteoclastos activos supera
a la de osteoblastos activos ya que para valores bajos de DOBp se inhibe la diferenciación de
preosteoblastos precursores a osteoblastos activos. Al haber mayor población de osteoclastos
activos, éstos producen más TGFβ de manera que se inhibe aún más la diferenciación OBpOBa.
Esta mayoría de osteoclastos y la fuerte inhibición de la diferenciación de OBa traen como
consecuencia una disminución de la masa ósea, de tal manera que ya no se recupera el estado normal
del proceso remodelatorio.
GRÁFICA 4
6.2.2.3 Índice de diferenciación de OCuOCp, DOCu
Para una variación entre [valor original del parámetro]*104 hasta [valor original del
parámetro]*105 el sistema se hace inestable. . Los valores del índice de diferenciación de
OCuOCp para los que el sistema se hace inestable están muy alejados del valor nominal. Esto
hace pensar que el sistema se hace inestable porque valores tan altos de esta constante solo se
darían en situaciones anormales de enfermedad, ya que en una persona sana serían biológicamente
inviables.

6. ANÁLISIS DE VARIABLES
53
GRÁFICA 5
6.2.2.4 Índice de diferenciación de OCpOca, DOCp
La variación de este parámetro no afecta a la estabilidad del sistema.
GRÁFICA 6
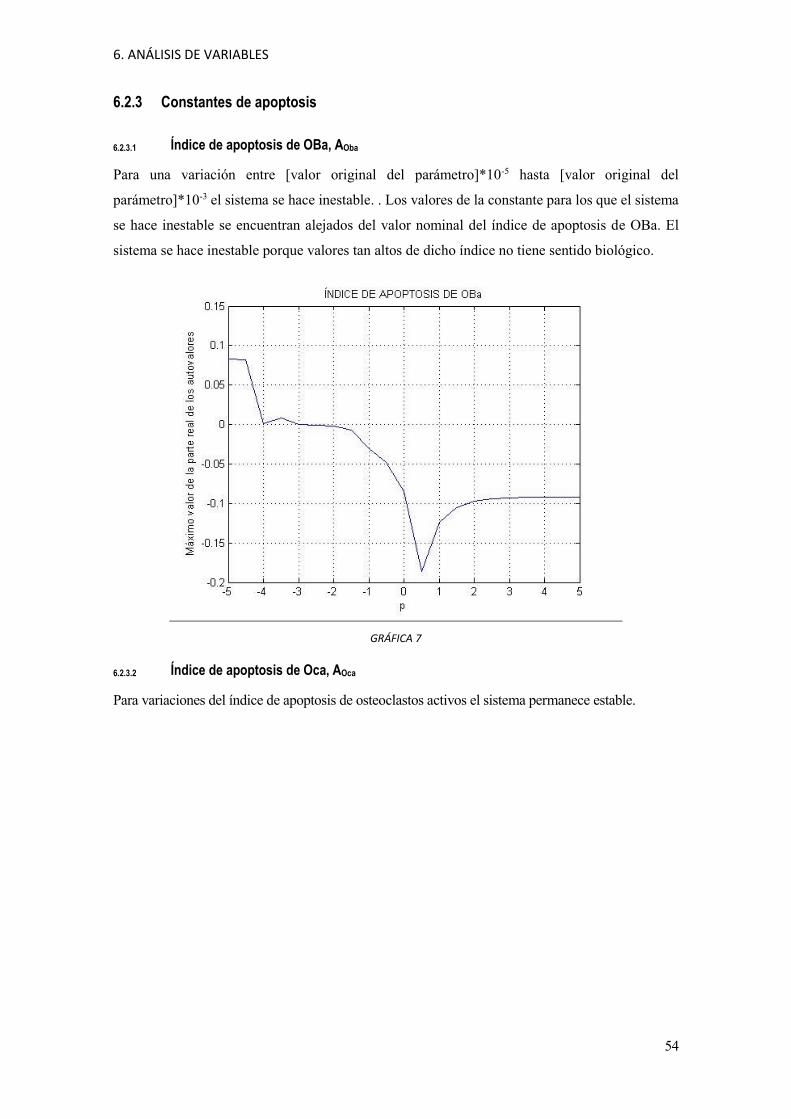
6. ANÁLISIS DE VARIABLES
54
6.2.3 Constantes de apoptosis
6.2.3.1 Índice de apoptosis de OBa, AOba
Para una variación entre [valor original del parámetro]*10-5 hasta [valor original del
parámetro]*10-3 el sistema se hace inestable. . Los valores de la constante para los que el sistema
se hace inestable se encuentran alejados del valor nominal del índice de apoptosis de OBa. El
sistema se hace inestable porque valores tan altos de dicho índice no tiene sentido biológico.
GRÁFICA 7
6.2.3.2 Índice de apoptosis de Oca, AOca
Para variaciones del índice de apoptosis de osteoclastos activos el sistema permanece estable.

6. ANÁLISIS DE VARIABLES
55
GRÁFICA 8
6.2.4 Índice de proliferación de OBp, POBp
Para todo el rango de variación (desde [valor original del parámetro]*10-5 hasta [valor original
del parámetro]*10-5) el sistema se hace inestable. Esto es así porque se dispara la proliferación de
osteoblastos precursores y con ello la población de osteoblastos activos. Este gran aumento en la
población de osteoblastos podría traer como consecuencia un aumento en la masa ósea, de tal manera
que ya no se recupera el estado anterior. Por tanto, el sistema es muy sensible a cambios en el índice
de proliferación de osteoblastos precursores.
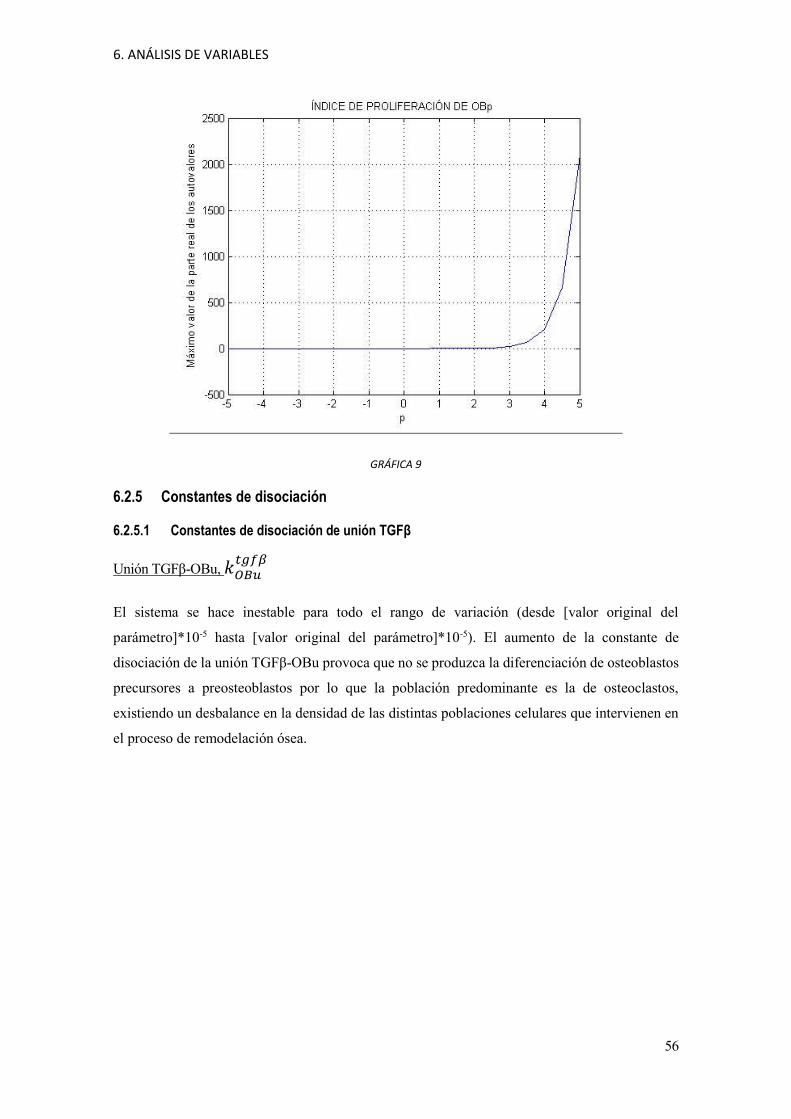
6. ANÁLISIS DE VARIABLES
56
GRÁFICA 9
6.2.5 Constantes de disociación
6.2.5.1 Constantes de disociación de unión TGFβ
Unión TGFβ-OBu, 𝑘𝑂𝐵𝑢𝑡𝑔𝑓𝛽
El sistema se hace inestable para todo el rango de variación (desde [valor original del
parámetro]*10-5 hasta [valor original del parámetro]*10-5). El aumento de la constante de
disociación de la unión TGFβ-OBu provoca que no se produzca la diferenciación de osteoblastos
precursores a preosteoblastos por lo que la población predominante es la de osteoclastos,
existiendo un desbalance en la densidad de las distintas poblaciones celulares que intervienen en
el proceso de remodelación ósea.

6. ANÁLISIS DE VARIABLES
57
GRÁFICA 10
Unión TGFβ-OBp, 𝑘𝑂𝐵𝑝𝑡𝑔𝑓𝛽
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*10-5 hasta [valor original del parámetro]*10-2.5. Los valores de la constante para los
que el sistema se hace inestable están alejados del valor nominal. El sistema se hace inestable ya
que valores tan bajos de dicha constante no tiene sentido biológico.

6. ANÁLISIS DE VARIABLES
58
GRÁFICA 11
Unión TGFβ-OCa, 𝑘𝑂𝐶𝑎𝑡𝑔𝑓𝛽
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*10-5 hasta [valor original del parámetro]*10-3. En este rango no se produce la
apoptosis de osteoclastos. Algunos valores de 𝑘𝑂𝐶𝑎𝑡𝑔𝑓𝛽
para los que el sistema se hace inestable
están muy próximos al valor nominal. Esto hace pensar que el sistema se hace inestable porque
el modelo falla en alguna de sus ecuaciones o parámetros o porque el valor de esta constante está
muy ajustado y hay que prestar mucha atención su valor exacto, ya que cualquier mínimo cambio
en dicho valor hace inestable el sistema.

6. ANÁLISIS DE VARIABLES
59
GRÁFICA 12
6.2.5.2 Constantes de disociación de unión PTH-OB
En 𝜋𝑎𝑐𝑡,𝑂𝐵𝑝𝑡ℎ
, 𝑘𝑂𝐵,𝑟𝑒𝑝𝑝𝑡ℎ
Para este parámetro el sistema se mantiene estable en todas sus variaciones.
GRÁFICA 13

6. ANÁLISIS DE VARIABLES
60
En 𝜋𝑟𝑒𝑝,𝑂𝐵𝑝𝑡ℎ
, 𝑘𝑂𝐵,𝑎𝑐𝑡𝑝𝑡ℎ
Para variaciones este parámetro el sistema permanece estable.
GRÁFICA 14
6.2.5.3 Constantes de disociación de unión RANKL, 𝒌𝑶𝑪𝒓𝒂𝒏𝒌𝒍
Para variaciones de la constante de disociación de unión RANKL el sistema permanece estable.

6. ANÁLISIS DE VARIABLES
61
GRÁFICA 15
6.2.6 Constantes de asociación
6.2.6.1 Constante de asociación de unión RANKL-OPG, K[RANKL-OPG]
El sistema permanece estable para todo el rango de variación de la constante de asociación de unión
RANKL-OPG
GRÁFICA 16

6. ANÁLISIS DE VARIABLES
62
6.2.6.2 Constante de asociación de unión RANKL-RANK, K[RANKL-RANK]
El sistema permanece estable para todo el rango de variación de la constante de asociación de unión
RANKL-RANK
GRÁFICA 17
6.2.7 Función de activación de la diferenciación de OCu en OCp, 𝝅𝒂𝒄𝒕,𝑶𝑪𝒖𝑴𝑪𝑺𝑭
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*103 hasta [valor original del parámetro]*105. Al aumentar este parámetro se dispara
la población de preosteoclastos. Los valores de 𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑀𝐶𝑆𝐹 para los que el sistema se hace inestable
están demasiados alejados del valor nominal. Esto hace pensar que el sistema se hace inestable
porque valores tan altos de dicha constante no tienen sentido biológico en el caso de una persona
sana.
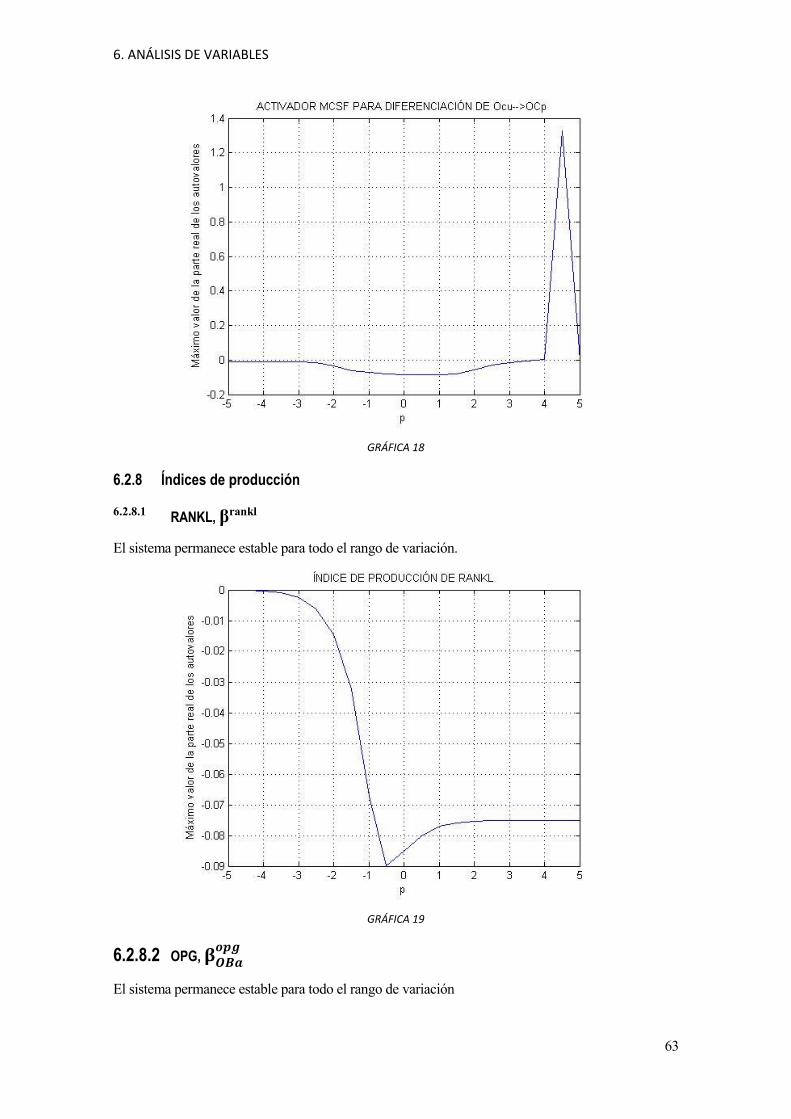
6. ANÁLISIS DE VARIABLES
63
GRÁFICA 18
6.2.8 Índices de producción
6.2.8.1 RANKL, βrankl
El sistema permanece estable para todo el rango de variación.
GRÁFICA 19
6.2.8.2 OPG, 𝛃𝑶𝑩𝒂𝒐𝒑𝒈
El sistema permanece estable para todo el rango de variación

6. ANÁLISIS DE VARIABLES
64
GRÁFICA 20
6.2.8.3 PTH, βPTH
El sistema permanece estable para todo el rango de variación
GRÁFICA 21

6. ANÁLISIS DE VARIABLES
65
6.2.9 Índices de degradación
6.2.9.1 RANKL, �̃�RANKL
El sistema permanece estable para todo el rango de variación
GRÁFICA 22
6.2.9.2 OPG, �̃�OPG
El sistema permanece estable para todo el rango de variación
GRÁFICA 23

6. ANÁLISIS DE VARIABLES
66
6.2.9.3 PTH, �̃�PTH
El sistema permanece estable para todo el rango de variación
GRÁFICA 24
6.2.9.4 TGFβ, �̃�TGF-β
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*10-5 hasta [valor original del parámetro]*10-2.5. Los valores de �̃�TGF-βpara los que el
sistema se hace inestable se encuentran alejados del valor nominal. Esto hace pensar que el
sistema se hace inestable porque tales valores no tienen sentido biológico en el caso de una
persona sana.
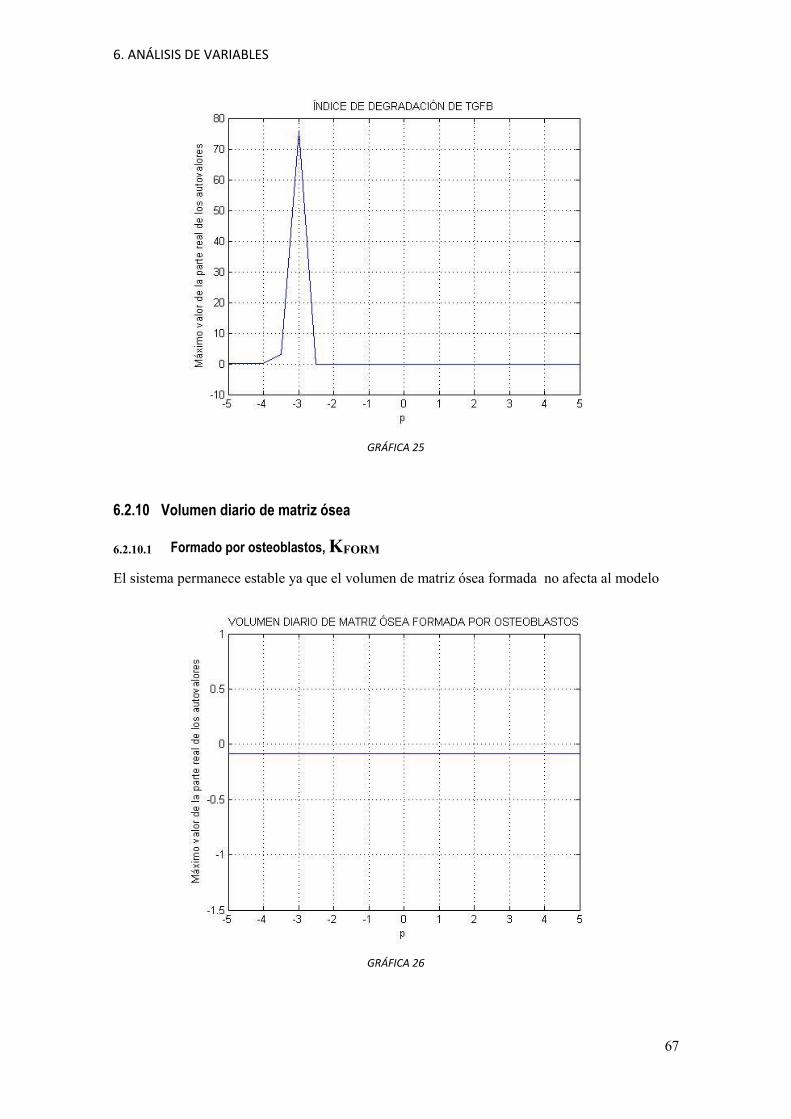
6. ANÁLISIS DE VARIABLES
67
GRÁFICA 25
6.2.10 Volumen diario de matriz ósea
6.2.10.1 Formado por osteoblastos, KFORM
El sistema permanece estable ya que el volumen de matriz ósea formada no afecta al modelo
GRÁFICA 26

6. ANÁLISIS DE VARIABLES
68
6.2.10.2 Resorbido por osteoclastos, KRES
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*102.5 hasta [valor original del parámetro]*105. Cuando aumenta este parámetro se
dispara la pérdida de masa ósea. Los valores de KRES para los que el sistema se hace inestable se
encuentran alejados del valor nominal. Esto hace pensar que el sistema se hace inestable porque
tales valores no tienen sentido biológico en el caso de una persona sana.
GRÁFICA 27
6.2.11 Densidad de TGFβ almacenado en la matriz ósea, 𝜼𝒕𝒈𝒇 𝜷𝒃𝒐𝒏𝒆
El sistema se hace inestable para el rango de variación que comprende desde [valor original del
parámetro]*103 hasta [valor original del parámetro]*105. Los valores de 𝜂𝑡𝑔𝑓 𝛽𝑏𝑜𝑛𝑒
para los que el
sistema se hace inestable se encuentran alejados del valor nominal. Esto hace pensar que el
sistema se hace inestable porque tales valores no tienen sentido biológico en el caso de una
persona sana.

6. ANÁLISIS DE VARIABLES
69
GRÁFICA 28
6.2.12 Densidad OPG a la que para la producción endógena, βOPGmax
El sistema permanece estable para todo el rango de variación
GRÁFICA 29
6.2.13 Máximo número de RANKL/OBp, 𝑵𝑶𝑩𝒑𝒓𝒂𝒏𝒌𝒍
El sistema permanece estable para todo el rango de variación

6. ANÁLISIS DE VARIABLES
70
GRÁFICA 30
6.2.14 Receptores RANK/OCp, 𝑵𝑶𝑪𝒑𝒓𝒂𝒏𝒌
El sistema permanece estable para todo el rango de variación
GRÁFICA 31

7. APLICACIONES A LOS CASOS INESTABLES
71
7 APLICACIÓN A UN CASO INESTABLE
En este apartado se van a analizar las densidades de las poblaciones celulares para una de las
constantes que en alguno de los rangos de variación hacen inestable el sistema, es decir, se va a
estudiar qué población se dispara al infinito provocando que el sistema no vuelva a su estado
anterior de estabilidad.
Para analizar este caso se parte de la siguiente población inicial:
OBp(1) = 1x10-3
OBa(1) = 1x10-3
OCp(1) = 1x10-3
OCa(1) = 1x10-3
Como ejemplo se ha tomado el índice de proliferación de OBp, POBp
GRÁFICA 9
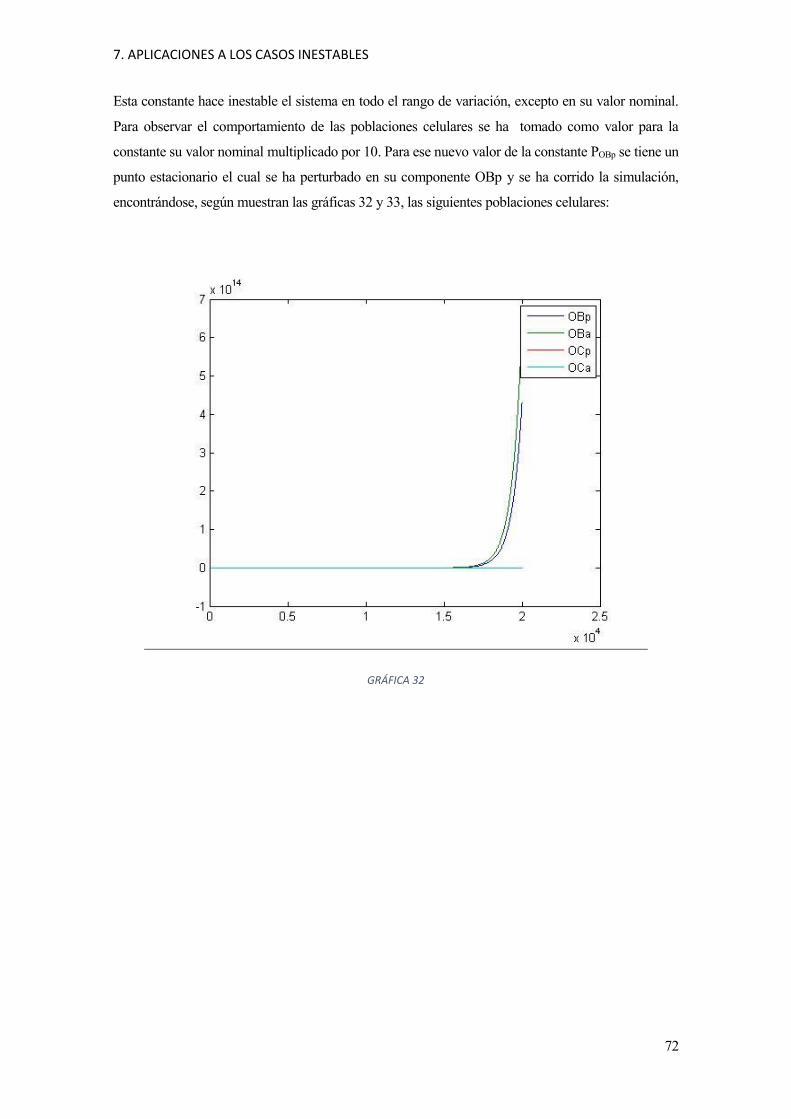
7. APLICACIONES A LOS CASOS INESTABLES
72
Esta constante hace inestable el sistema en todo el rango de variación, excepto en su valor nominal.
Para observar el comportamiento de las poblaciones celulares se ha tomado como valor para la
constante su valor nominal multiplicado por 10. Para ese nuevo valor de la constante POBp se tiene un
punto estacionario el cual se ha perturbado en su componente OBp y se ha corrido la simulación,
encontrándose, según muestran las gráficas 32 y 33, las siguientes poblaciones celulares:
GRÁFICA 32
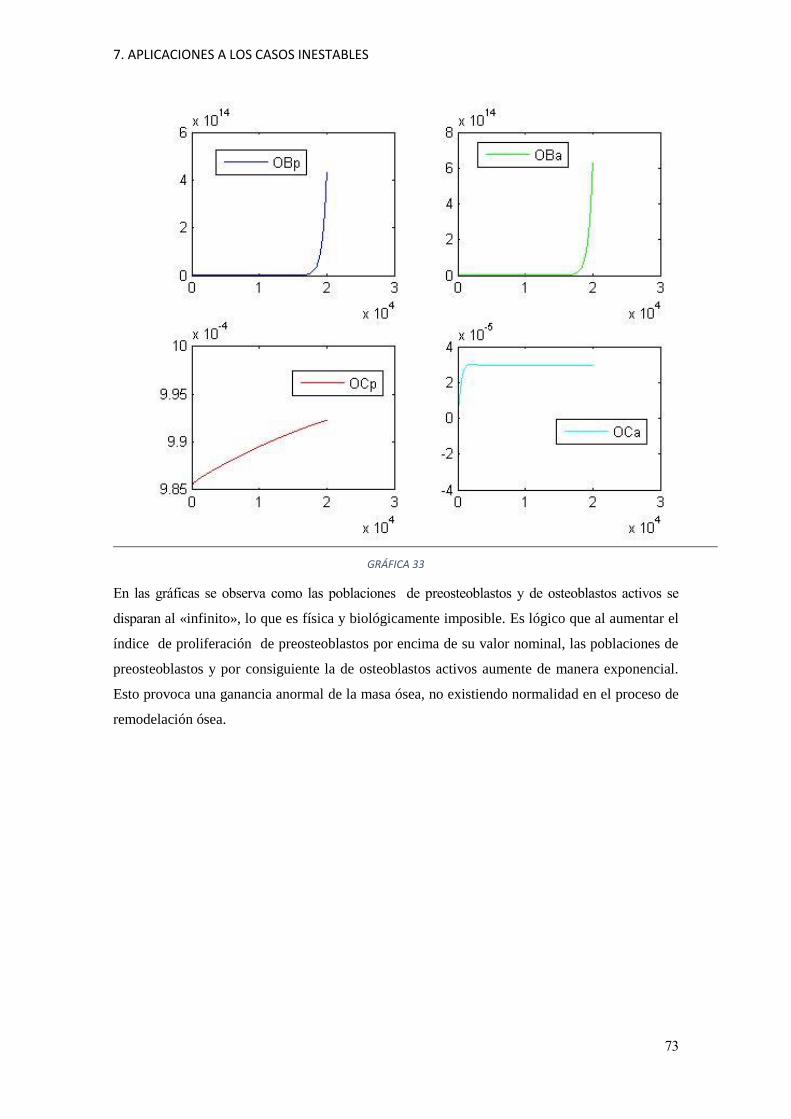
7. APLICACIONES A LOS CASOS INESTABLES
73
GRÁFICA 33
En las gráficas se observa como las poblaciones de preosteoblastos y de osteoblastos activos se
disparan al «infinito», lo que es física y biológicamente imposible. Es lógico que al aumentar el
índice de proliferación de preosteoblastos por encima de su valor nominal, las poblaciones de
preosteoblastos y por consiguiente la de osteoblastos activos aumente de manera exponencial.
Esto provoca una ganancia anormal de la masa ósea, no existiendo normalidad en el proceso de
remodelación ósea.

74
8 CONCLUSIONES
El modelo que se ha estudiado y analizado muestra una alta sensibilidad ante cambios en algunas
de las constantes que constituyen el modelo. Dichos cambios provocan importantes
modificaciones en la respuesta dinámica de la población de osteoclastos y osteoblastos. En
algunas ocasiones el sistema puede regresar a su estado normal en cuanto a los ciclos de
generación de osteoclastos y osteoblastos pero la masa ósea puede verse afectada en su volumen
total.
En este trabajo se estudian algunas variaciones de las constantes propias del proceso de
remodelación para el modelo de Pivonka, que llevan a posibles alteraciones de los procesos del
metabolismo óseo.
A continuación se muestra una tabla con las distintas constantes del modelo, que al ser variadas dentro
del rango especificado, hacen inestable el sistema. También se representa el rango de variación para
el que se produce la inestabilidad.
CONSTANTES RANGO DE VARIACIÓN
Densidad de osteoclastos precursores, [OCu] [(OCu)*104 – (OCu)*105]
Índice de diferenciación de OBpOBa, [DOBp] [(DOBp)*10-5 – (DOBp)*10-0.5]
Índice de diferenciación de OCuOCp, [DOCu] [(DOCu)*104 – (DOCu)*105]
Índice de apoptosis de OBa, [AOBa] [(AOBa)*10-5 – (AOBa)*10-2]
Índice de proliferación de OBp, [POBp] [(POBp)*10-5 – (POBp)*105]
Cte. de disociación de unión TGFβ-OBu, [𝑘𝑎𝑐𝑡,𝑂𝐵𝑢𝑇𝐺𝐹β
] [𝑘𝑎𝑐𝑡,𝑂𝐵𝑢𝑇𝐺𝐹β
*10-5 – 𝑘𝑎𝑐𝑡,𝑂𝐵𝑢𝑇𝐺𝐹β
*105]
Cte. de disociación de unión TGFβ-OBp, [𝑘𝑟𝑒𝑝,𝑂𝐵𝑝𝑇𝐺𝐹β
] [𝑘𝑟𝑒𝑝,𝑂𝐵𝑝𝑇𝐺𝐹β
*10-5 – 𝑘𝑟𝑒𝑝,𝑂𝐵𝑝𝑇𝐺𝐹β
*10-0.5]
Cte. de disociación de unión TGFβ-OBa, [𝑘𝑎𝑐𝑡,𝑂𝐶𝑎𝑇𝐺𝐹β
] [𝑘𝑎𝑐𝑡,𝑂𝐶𝑎𝑇𝐺𝐹β
*10-5 – 𝑘𝑎𝑐𝑡,𝑂𝐶𝑎𝑇𝐺𝐹β
*10-0.5]
Activador MCSF diferenciación de OCuOCp, [𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑀𝐶𝑆𝐹 ] [𝜋𝑎𝑐𝑡,𝑂𝐶𝑢
𝑀𝐶𝑆𝐹 *104 – 𝜋𝑎𝑐𝑡,𝑂𝐶𝑢𝑀𝐶𝑆𝐹 *105]

75
Índice de degradación de TGFβ, 𝐷𝑇𝐺𝐹𝛽 [(𝐷𝑇𝐺𝐹𝛽)*10-5 – (𝐷𝑇𝐺𝐹𝛽)*10-3.5]
Volumen diario de matriz ósea reabsorbida, [𝐾𝑅𝐸𝑆] [(𝐾𝑅𝐸𝑆)*102.5 – (𝐾𝑅𝐸𝑆)*105]
Densidad de TGFβ almacenado en la matriz ósea, [𝜂𝑏𝑜𝑛𝑒𝑇𝐺𝐹𝛽
] [(𝜂𝑏𝑜𝑛𝑒𝑇𝐺𝐹𝛽
)*102.5 – (𝜂𝑏𝑜𝑛𝑒𝑇𝐺𝐹𝛽
)*105]
Tabla 2
Con respecto a los rangos de variación de las constantes para los que el sistema se hace inestable
pueden ocurrir dos situaciones
1. El rango para el cual el sistema se hace inestable está muy próximo al valor nominal de la
constante. En este caso, el sistema es inestable por un posible fallo del modelo matemático en
sí.
2. El rango para el cual el sistema se hace inestable se encuentra bastante alejado del valor
nominal de la constante. En este caso, se considera que tales valores de las constantes son
física y biológicamente imposibles y que sólo se darían bajo situaciones de enfermedades
relacionadas con los huesos o algunas alteraciones metabólicas.

76
Bibliografía
[ 1]
Pivonka P, Zimak J, Smith DW, Gardiner BS, Dunstan CR, Sims NA, Martin TJ,
Mundy GR. Model structure and the control of bone remodeling: a theoretical study.
Bone 2008; 43:249.
[ 2]
Pivonka P, Zimak J, Smith DW, Gardiner BS, Dunstan CR, Sims NA, Martin TJ,
Mundy GR. Theoretical investigation of the role of the RANK–RANKL–OPG system
in bone remodeling. J Theor Biol 2010; 262: 306–16.
[ 3]
Pivonka P, Buenzli PR, Sheiner S, Hellmich C, Dunstan CR. The influence of bone
Surface availability in bone remodelling: A mathematical model including coupled
geometrical and biochemical regulations of bone cells. Engineering Structures 2013;
47:134–147.
[ 4] Robling AG, Castillo AB, Turner CH. Biomechanical and Molecular Regulation of Bone
Remodeling, Ann. Rev. Biomed. Engrg., 8 (2006), pp. 455-498
[ 5]
S. V. Komarova, Mathematical model of paracrine interactions between osteoclasts
and osteoblasts predicts anabolic action of parathyroid hormone on bone, J.
Endocrinol., 146 (2005), pp. 3589–3595.
[ 6]
S. V. Komarova, R. J. Smith, S. J. Dixon, S. M. Sims, and L.M.Wahl, Mathematical
model predicts a critical role for osteoclast autocrine regulation in the control of bone
remodeling, Bone, 33 (2003), pp. 206–215.
[ 7]
M. D. Ryser, N. Nigam, and S. V. Komarova, The celular dynamics of bone
remodelling: A Mathematical Model. Siam J. Appl. Math. Vol. 70, No. 6 (2010), pp.
1899–1921.
[ 8] Martin RB. Porosity and specific surface of bone. Crit Rev Biomed Eng 1984; 10:
179–222.
[ 9] Martin TJ. Paracrine regulation of osteoclast formation and activity: milestones in
discovery. J Musculoskel Neuron Interact 2004; 4: 243–53.