
ANÁLISIS ESTACIONAL DE LAS CARACTERÍSTICAS …biblio.uabcs.mx/tesis/te3153.pdf · especial a M....
95
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA ANÁLISIS ESTACIONAL DE LAS CARACTERÍSTICAS QUÍMICO PROXIMALES DEL CALAMAR GIGANTE Dosidicus gigas, RECOLECTADO EN SANTA ROSALÍA, BAJA CALIFORNIA SUR, MÉXICO. TESIS DE LICENCIATURA PARA OBTENER EL TÍTULO DE: BIÓLOGA MARINA PRESENTA: DONAJI OCHOA TEPETLA DIRECTOR: DR. CÉSAR AUGUSTO SALINAS ZAVALA LA PAZ, BAJA CALIFORNIA SUR, 06 DE JUNIO DE 2014
Transcript of ANÁLISIS ESTACIONAL DE LAS CARACTERÍSTICAS …biblio.uabcs.mx/tesis/te3153.pdf · especial a M....

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
ANÁLISIS ESTACIONAL DE LAS CARACTERÍSTICAS QUÍMICO PROXIMALES DEL CALAMAR GIGANTE
Dosidicus gigas, RECOLECTADO EN SANTA ROSALÍA, BAJA CALIFORNIA SUR, MÉXICO.
TESIS DE LICENCIATURA PARA OBTENER EL TÍTULO DE:
BIÓLOGA MARINA
PRESENTA:
DONAJI OCHOA TEPETLA
DIRECTOR: DR. CÉSAR AUGUSTO SALINAS ZAVALA
LA PAZ, BAJA CALIFORNIA SUR, 06 DE JUNIO DE 2014

II

III
(HOJA DE DICTAMEN DE APROBACIÓN)

IV
A mi hijo Ulises Kiyari (Panky)
mi familia y todo el clan

V
AGRADECIMIENTOS
Del CIBNOR…
Esta Tesis es el reflejo de un trabajo de muchos años, obvio no todos seguidos, pero
los calamares siempre han estado presentes, desde aquella primera estancia de
investigación, donde empecé a conocer al equipo de calamareros, a la batuta de Dr.
César Salinas que confió en mí para la realización de la investigación y director de mi
tesis, muchas gracias a su paciencia, porque si me tardé un poco…
A quienes les tocó realizar la toma de muestras sin saber a quién iba a llegar, a todo
el personal del Laboratorio de Cefalópodos y a todos aquellos que pertenecieron y
aún pertenecen al grupo, con quienes he vivido tantas experiencias, recibido
consejos, ayuda y su amistad: Minda, Susy, Dana, Gastón, Rigo, Jorge… En
especial a M. en C. Arminda Mejía Rebollo que ha sido mi sinodal para esta tesis y
ha estado al pendiente en cada momento.
A MC. Sonia Gpe. Rocha y IBQ. Ma. Dolores Rondero del Laboratorio de Análisis
Químico Proximal, quienes me instruyeron y ayudaron para utilizar y comprender los
equipos durante el estudio de gabinete.
De la UABCS…
A quien me ha ayudado a formalizar mi escrito, quien me dio consejos de la tesis y
quien me hizo la pregunta más difícil: ¿Después qué? Dr. Carlos Sánchez Ortiz,
gracias por su tiempo. A mi sinodal experto porque la tesis se basa en análisis
bioquímicos, M. en C. Erika Torres Ochoa, gracias por todo el apoyo y consejos
sobre todo en esos momentos en que ya no sabía cómo plasmar mis ideas en el
escrito. A Dra. Liliana Hernández Olalde, que aceptó ser sinodal a pesar de que ya
estaba en la última fase de su embarazo.
En un momento existencialista cuando las gráficas eran kilométricas, M. en C.
Georgina Brabata Domínguez me ayudó en la parte estadística a darle formato y a
mejorar los resultados para que fueran mejor comprendidos.

VI
De la mejor carrera Biología Marina a mis compañeros y amigos: Charly, Mariposa,
Roy, Polo, Bruce, Arenita, Anet, Lizz, Rocho, Marce, Kass, Vaca, Natalia, Lina,
Armando, Carmina, Merit, Mel, Hanny, Fabián, Edgardo, Villalón, Isa, Hongos y los
que me faltaron… Por la compañía, aventuras, diversión, estudio, mar y escapadas.
Familia y Amigos…
Hubo un momento en que di una pausa a mi carrera de BM (tesis), me dediqué mi
segunda carrera: atención prehospitalaria y gracias a ello le pude dar a la tesis un
énfasis más humano y dirigido a la salud. También conocí a mi operador favorito de
ambulancias, que con una mirada sabemos qué ocurre y que necesitamos, un
excelente compañero en las guardias y gran amigo en la vida diaria, que ha estado
apoyando en los buenos y malos momentos, sin importar qué tan lejos hay que
llegar, muchas gracias Iván Campos.
A los amigos que se convirtieron en mi familia, a mi amá Rocio Reyes y sus angelitos
que en los últimos momentos que fueron difíciles de mi vida ha estado para
abrazarme, regañarme, enseñarme y siempre con un consejo.
A mi familia de sangre que me esperó, algo impaciente al final, pero siempre ha
estado ahí para apoyarme, a mi mamá Andrea, tíos: Julia, Valente, Gaspar, Fidel,
Pepe, Tina y Edna, primos Ale, Dany, Emy, Tania, Catty, Alan, Diego y Emiliano.
Gracias por todo este tiempo, los quiero.
A todos mis abuelos que están ya en otro plano, al igual que mi papá que estuvo la
primera mitad de mi vida y que me dejo a más familia, a mis hermanos Silvia y
Andrés con sus respectivos hijos y hasta ahora nuevos sobrinos.
Agradezco a Dios por mi personita ideal y príncipe azul, quien detuvo mi vida para
avanzar más lejos y desde que era un blastocisto inquieto (que rebotaba en mi útero)
supe que su energía me llenaría y espero me acompañe a más aventuras…
José Martí dijo: Hay tres cosas que cada persona debería hacer durante su vida:
plantar un árbol (), tener un hijo ( Kiyari) y escribir un libro ( tesis).
Estos objetivos los he cumplido. Y éste, es mi comienzo…

VII
RESUMEN
El calamar gigante Dosidicus gigas (D’Orbigny, 1835) se distribuye en el Pacífico
Oriental desde Alaska hasta Chile. Es un recurso pesquero de gran importancia en
México, Perú y Chile, con un consumo que gradualmente se ha incrementado en
nuestro país desde 1994 a la fecha (Luna-Raya et al. 2006). Debido al alto valor
nutricional que D. gigas presenta, este estudio se enfoca en conocer los porcentajes
de humedad, proteínas, lípidos y cenizas del músculo de esta especie en un ciclo
anual. Los especímenes fueron recolectados en Santa Rosalía, BCS y a partir del
Análisis Químico Proximal (AQP) se evaluó la calidad del músculo, ya que son los
fundamentos que ayudan a difundir y promover sus cualidades nutricionales e
incrementar la ingesta de los alimentos en general, ya sea para consumo humano o
para la elaboración de programas de nutrición. En el AQP se obtuvo de manera
general: humedad 78.9 %, proteínas 78.5 %, lípidos 1.8 % y cenizas 5.0 %. En los
machos se encontraron los valores mayores que en las hembras, pero únicamente
los porcentajes de humedad presentaron diferencia significativa (p<0.05); los
organismos maduros contienen mayor concentración de humedad (p<0.05) y en los
organismos en proceso de maduración se estimaron los porcentajes menores de
percentiles de proteínas y lípidos. Respecto a los valores mensuales de proteínas,
lípidos y cenizas, los meses de julio y agosto fueron elevados, mientras que en los
meses de septiembre y octubre estos disminuyen. Se realizó una comparación de los
valores nutricionales del calamar gigante con otros tipos de carnes (moluscos,
pescados y res) evidenciando que éste presenta una alta concentración de proteínas
y una baja concentración de lípidos. De acuerdo a los resultados obtenidos, el
calamar gigante es una excelente alternativa de alimento balanceado, el cual puede
ser consumido de manera directa o procesado, con una elevada calidad nutricional
para el consumo humano.

VIII
ÍNDICE
RESUMEN ................................................................................................................ VII
LISTA DE FIGURAS ................................................................................................... X
LISTA DE TABLAS .................................................................................................. XII
1. INTRODUCCIÓN ............................................................................................... 1
1.1. Antecedentes ........................................................................................................................... 3
1.2. Identificación y delimitación del problema .............................................................................. 7
1.3. Justificación .............................................................................................................................. 8
1.4. Objetivos .................................................................................................................................. 8
1.4.1. Objetivo general ..................................................................................................................... 8
1.4.2. Objetivos particulares ............................................................................................................ 9
1.5. Área de estudio ........................................................................................................................ 9
2. MÉTODOS ...................................................................................................... 12
2.1. Captura de organismos .......................................................................................................... 12
2.2. Preparación de la muestra ..................................................................................................... 13
2.3. Análisis Químico Proximal ...................................................................................................... 13
2.4. Análisis estadísticos ................................................................................................................ 14
3. RESULTADOS ................................................................................................ 15
3.1. Estructura de tallas ................................................................................................................. 15
3.2. Análisis Químico Proximal ...................................................................................................... 15
3.2.1. %Humedad ........................................................................................................................... 16
3.2.2. %Proteínas totales (Pt) ....................................................................................................... 19
3.2.3. %Lípidos totales ................................................................................................................... 21
3.2.4. %Cenizas .............................................................................................................................. 23
4. DISCUSIÓN..................................................................................................... 25
Estructura de tallas ................................................................................................................................ 25
Análisis Químico Proximal ...................................................................................................................... 25
%Humedad ............................................................................................................................................. 26
%Proteínas totales.................................................................................................................................. 27
%Lípidos totales ...................................................................................................................................... 28
%Cenizas ................................................................................................................................................ 29

IX
Variación del AQP por sección corporal ................................................................................................. 30
Variación del AQP por sexo .................................................................................................................... 31
Variación del AQP por estado de madurez ............................................................................................. 32
Variación del AQP entre meses .............................................................................................................. 32
Consumo del calamar gigante................................................................................................................ 33
5. CONCLUSIONES ............................................................................................ 37
6. RECOMENDACIONES ................................................................................... 38
7. LITERATURA CONSULTADA ........................................................................ 39
Hemerografía ......................................................................................................................................... 47
Recursos electrónicos............................................................................................................................. 48
ANEXO A. Marco Conceptual ................................................................................... 50
ANEXO B. Análisis Químico Proximal, ...................................................................... 58
ANEXO C. Tablas ANOVA y a posteriori DVS Tukey ............................................... 66
ANEXO D. Recetario ................................................................................................. 76

X
LISTA DE FIGURAS
FIGURA 1. Área de afloramiento en el Golfo de California: (A) vientos del noroeste,
(B) vientos del sureste, (C) mezcla de mareas (Tomado de Markaida, 2001).
FIGURA 2. Zona geográfica de muestreo de Dosidicus gigas en Santa Rosalía,
B. C. S, Golfo de California.
FIGURA 3. Estructura de las tallas de calamar gigante Dosidicus gigas, capturados
en Santa Rosalía B. C. S. durante 2004.
FIGURA 4. Parámetros nutricionales generales (Proteínas, Lípidos y Cenizas) del
músculo de calamar gigante Dosidicus gigas, mediante AQP.
FIGURA 5. Porcentajes de humedad del calamar gigante Dosidicus gigas capturado
en Sta. Rosalía B.C.S durante 2004. A: sección corporal, B: sexos, C: estadio de
madurez en machos, D: estadio de madurez en hembras y E: mes de colecta.
FIGURA 6. Porcentajes de proteínas del calamar gigante Dosidicus gigas capturado
en Sta. Rosalía B. C. S durante 2004. A: sección corporal, B: sexo, C: estadio de
madurez y D: mes de colecta.
FIGURA 7. Porcentajes de lípidos totales del calamar gigante Dosidicus gigas
capturado en Sta. Rosalía B. C. S durante 2004. A: sección corporal B: sexo,
C: estadio de madurez y D: mes de colecta.
FIGURA 8. Porcentajes de cenizas del calamar gigante Dosidicus gigas capturado en
Sta. Rosalía B. C. S durante 2004. A: sección corporal B: sexo, C: estadio de
madurez y D: mes de colecta.
FIGURA 9. AQP del calamar gigante Dosidicus gigas capturado en Sta. Rosalía,
B. C. S durante 2004. A: sección corporal, B: sexo, C: estadio de madurez y
D: mes de colecta.

XI
FIGURA 10. Calamar gigante Dosidicus gigas. A: vista dorsal de un ejemplar grande
(50-80 cm LM) B: vista ventral de un ejemplar de 25.4 cm LM; C: IV brazo
izquierdo hectocotilizado y pedúnculo tentacular (Tomado de Markaida, 2001).
FIGURA 11. Distribución geográfica del calamar gigante Dosidicus gigas en el
Pacífico Oriental. Se observa la expansión poblacional identificada en las últimas
décadas (Roper y Young, 1975 y Barnes, 1996; Nigmatullin et al., 2001 y Gilly et
al., 2005). Tomada de Bazzino, 2008.

XII
LISTA DE TABLAS
TABLA I. Composición químico proximal (expresado en porcentaje) de diferentes
organismos y alimentos, incluye conversión de materia húmeda a materia seca
de acuerdo a Martínez-Montes (2009), excepto para humedad.
TABLA II. Parámetros nutricionales de músculo de calamar gigante Dosidicus gigas,
de acuerdo a sus principales estructuras corporales. ANOVA de una vía y a
posteriori DVS de Tukey. * significativo p ≤ 0.05, ** significativo p ≤ 0.01, NS no
significativo p > 0.05.
TABLA III. Comparaciones estadísticas de la concentración de proteínas en músculo
de calamar gigante D. gigas de acuerdo a los meses de muestreo, con α = 0.05
y p < 0.05.
TABLA IV. Comparaciones estadísticas de los porcentajes de lípidos totales en los
meses de muestreo del calamar gigante D. gigas, con α = 0.05 y p < 0.05.
TABLA V. ANOVA de una vía en la prueba de %humedad según la sección corporal
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 3.1.
TABLA VI. Análisis a posteriori DVS de Tukey en la prueba de %humedad según la
sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA VII. ANOVA de una vía en la prueba de %humedad según el sexo de calamar
gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa
p > 0.05, α= 0.05, F= 10.6
TABLA VIII. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el
sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no
significativa p > 0.05, α= 0.05.

XIII
TABLA IX. ANOVA de una vía en la prueba de %humedad según el estado de
madurez de calamar gigante D. gigas macho. Diferencia significativa p < 0.05,
diferencia no significativa p > 0.05, α= 0.05, F= 6.33.
TABLA X. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el
estado de madurez de calamar gigante D. gigas macho. Diferencia significativa
p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa
p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
TABLA XI. ANOVA de una vía en la prueba de %humedad según el estado de
madurez de calamar gigante D. gigas hembra. Diferencia significativa p < 0.05,
diferencia no significativa p > 0.05, α= 0.05, F= 0.64.
TABLA XII. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el
estado de madurez de calamar gigante D. gigas hembra. Diferencia significativa
p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa
p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
TABLA XIII. ANOVA de una vía en la prueba de %humedad según el mes de colecta
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 45.9.
TABLA XIV. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el
mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XV. ANOVA de una vía en la prueba de proteínas según la sección corporal
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 1.74.

XIV
TABLA XVI. Análisis a posteriori DVS de Tukey en la prueba de proteínas según la
sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XVII. ANOVA de una vía en la prueba de proteínas según el sexo de calamar
gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa
p > 0.05, α= 0.05, F= 0.22.
TABLA XVIII. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el
sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no
significativa p > 0.05, α= 0.05.
TABLA XIX. ANOVA de una vía en la prueba de proteínas según el estado de
madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
no significativa p > 0.05, α= 0.05, F= 3.61.
TABLA XX. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el
estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XXI. ANOVA de una vía en la prueba de proteínas según el mes de colecta
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 29.6.
TABLA XXII. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el
estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.

XV
TABLA XXIII. ANOVA de una vía en la prueba de lípidos según el tipo de sección
corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
no significativa p > 0.05, α= 0.05, F= 6.5017
TABLA XXIV. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el
tipo de sección corporal de calamar gigante D. gigas. Diferencia significativa
p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa
p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
TABLA XXV. ANOVA de una vía en la prueba de lípidos según el sexo de calamar
gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa
p > 0.05, α= 0.05, F= 0.9414
TABLA XXVI. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el
sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no
significativa p > 0.05, α= 0.05.
TABLA XXVII. ANOVA de una vía en la prueba de lípidos según el estado de
madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
no significativa p > 0.05, α= 0.05, F= 3.0173
TABLA XXVIII. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el
estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XXIX. ANOVA de una vía en la prueba de lípidos según el mes de colecta de
calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 25.3489

XVI
TABLA XXX. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el
mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XXXI. ANOVA de una vía en la prueba de cenizas según la sección corporal
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 6.220
TABLA XXXII. Análisis a posteriori DVS de Tukey en la prueba de cenizas según la
sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.
TABLA XXXIII. ANOVA de una vía en la prueba de cenizas según el sexo de calamar
gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa
p > 0.05, α= 0.05, F= 0.8819
TABLA XXXIV. Análisis a posteriori DVS de Tukey en la prueba de cenizas según el
sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no
significativa p > 0.05, α= 0.05.
TABLA XXXV. ANOVA de una vía en la prueba de cenizas según el estado de
madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
no significativa p > 0.05, α= 0.05, F= 28.1792
TABLA XXXVI. Análisis a posteriori DVS de Tukey en la prueba de ceniza según el
estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.

XVII
TABLA XXXVII. ANOVA de una vía en la prueba de cenizas según el mes de colecta
de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no
significativa p > 0.05, α= 0.05, F= 86.231
TABLA XXXVIII. Análisis a posteriori DVS de Tukey en la prueba de cenizas según el
mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05,
diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001,
diferencia no significativa p > 0.05, α= 0.05.

1
1. INTRODUCCIÓN
El calamar gigante está clasificado dentro del grupo de los moluscos, el cual cuenta
con una diversidad morfológica bastante amplia, ya que está conformado por
animales que viven en el bentos (sustrato) como los caracoles y las almejas y
también los organismos que viven en el necton (columna de agua) como los
calamares (Ruppert y Barnes, 1996; Markaida, 2001; Boyle y Rodhouse, 2005).
La especie de calamar gigante Dosidicus gigas (D. gigas) se distribuye en el Pacífico
Oriental, desde Monterey, California hasta el sur de Chile (Nesis, 1983; Markaida,
2001; Nigmatullin et al, 2001), sin embargo desde 2004 se registró una expansión
hasta el Golfo de Alaska. Su distribución es semi-oceánica meridional tropical y
subtropical que coincide con zonas de alta productividad (Salinas-Zavala et al., 2005;
Wing, 2006; Zeidberg y Robison, 2007).
Se localizan en diferentes ambientes, desde fuentes hidrotermales hasta zonas
someras y desde latitudes polares hasta el Ecuador. También son depredadores
carnívoros, cazan en la columna de agua y en el sustrato; en particular los calamares
los han denominado como las criaturas más voraces del océano (Brusca y Brusca,
2003; Arancibia y Neira, 2006; Guerra, 2006). En cuanto a la evolución, los
calamares han modificado su concha reduciéndola a una estructura denominada
pluma o gladio, que de manera interna le sirve como un soporte corporal (Barnes et
al., 2001; Boyle y Rodhouse, 2005).
D. gigas es un organismo que realiza migraciones verticales diarias desde
profundidades entre 200 y 350 m y por la noche a aguas someras y cercanas a la
línea de costa. Mientras que las migraciones horizontales dentro del Golfo de
California tienen un desplazamiento en noviembre de Santa Rosalía a Guaymas y en
mayo de manera inversa (Markaida et al., 2005; Gilly et al., 2006; Bazzino, 2008).
En la actualidad existe un auge de la pesquería de calamar gigante, sobretodo en
países como México, Perú y Chile, que en su mayoría exportan el producto hacia
países asiáticos y una mínima parte se queda en sus respectivos territorios (Salinas-
Zavala et al, 2004; Blaskovic et al., 2008; CONAPESCA, 2011).

2
La pesca del calamar gigante es la más grande de cefalópodos del mundo y sus
principales productores son México y Perú. En nuestro país se pesca principalmente
en el Golfo de California, teniendo la disponibilidad del producto durante todo el año y
Baja California Sur es el estado principal como productor que aportó el 56.57 % del
total nacional. La mayoría del recurso capturado es exportado a países asiáticos,
europeos y norteamericanos (Markaida, 2001; Salinas-Zavala et al., 2004; INEGI,
2005; Luna-Raya et al., 2006; Blaskovic et al., 2008; CONAPESCA, 2013).
El calamar gigante como alimento ha sido catalogado como carne de buena calidad
nutrimental ya sea para el consumo humano o incluso en la elaboración de dietas en
la alimentación de organismos de cultivo (Abugoch et al., 1999; Martínez-Vega et al.,
2000b; Luna-Raya et al., 2006). Sin embargo en el calamar gigante, las
características bromatológicas reportadas anteriormente no distinguen entre las
diferentes secciones corporales, sexo, estado de madurez y mes de colecta. Los
cuales se cree que pueden ser variables por los factores endógenos y exógenos,
como la dieta, ontogenia, estado fisiológico y el ambiente (Bjerkeng et al, 1997). Tal
es el caso del salmón, el cual es un organismo con una dieta alta en grasa y como
consecuencia se encuentran grandes cantidades de lípidos corporales (Bell et al,
2001).
Por otro lado, el calamar al tener un valor nutricional alto, por sus grandes aportes de
proteínas y bajo en grasas (Luna-Raya et al., 2006), es un alimento ideal que puede
ayudar con el problema de salud más impactante en el mundo, la obesidad, ya que el
calamar es un producto balanceado de origen animal.
Al comparar el filete de calamar con carnes rojas, su diferencia inicia en el ambiente
de donde provienen (medio natural y ganadería, respectivamente). Doyle (2006) y
Lozano et al. (2008) mencionan que en la ganadería existen sustancias que pueden
provocar un riesgo de salud al consumidor y que, pueden ocurrir efectos tóxicos,
mutagénicos, carcinogénicos, desórdenes corporales, reacciones alérgicas y
resistencia bacteriana. Por ejemplo, el uso de anabólicos esteroides en el ganado y
antibióticos (Bavera et al., 2002).

3
Una forma de medir el valor nutricional de los seres vivos es mediante el Análisis
Químico Proximal (AQP), el cual es una serie de exámenes para evaluar y medir la
estructura de cualquier alimento, determinado a partir de los porcentajes de
humedad, proteínas, lípidos y cenizas (Badui-Dergal, 1993; Fox y Cameron, 2000).
1.1. Antecedentes
Los primeros autores que describen la distribución de D. gigas son Sato (1976) y
Nesis (1983), los cuales se basaron en organismos de la Costa Occidental de la
península de Baja California, posteriormente su estudio se ha incrementado (Klett,
1996 y Nigmatullin et al., 2001). Todos estos autores han hecho énfasis en el
calamar gigante como recurso pesquero de gran relevancia.
Existe una amplia gama de estudios acerca de biología, ecología y pesca del
calamar gigante. Entre los autores se destacan: Klett (1981, 1996), Ehrhardt et al.
(1986), Nevárez-Martínez et al. (2000, 2002), Morales-Bojórquez et al (2002),
Markaida (2001), Markaida et al. (2001, 2005, 2008), Bazzino et al. (2007). Estos
autores han generado tal información que amplía el conocimiento y comprensión del
mismo organismo y su interacción con el ambiente.
D. gigas es un recurso importante no solo para el ambiente, sino también para la
pesca, aunado a que es una fuente alimenticia importante, los estudios referidos a
este tema se encuentran sintetizados en la tabla I, donde se realiza un comparativo
nutricional dirigido al consumo humano. Posteriormente explicados en cada
apartado.

4
TABLA I. Composición químico proximal (expresado en porcentaje) de diferentes organismos y alimentos, incluye conversión de materia húmeda a materia seca de acuerdo a Martínez-Montes (2009), excepto para humedad.
Producto Especie %Humedad %Proteínas* %Lípidos* %Cenizas* Referencia
Calamar Dosidicus
gigas 82.23 86.12 4.90 7.37
Abugoch et al.,
1999
D. gigas 79.90 94.33 7.11 6.77 Gómez-Guillén et
al., 1997
D. gigas 2.09* 83.61 2.31 5.9 Martinez-Vega et
al., 2000b
Loligo plei 74.01 63.95 7.62 6.12 Cabello et. al.,
2004
Illex
illecebrosus 81.12 85.01 7.94 7.04
Krzynowek et al.,
1989
Pulpo Octopus
vulgaris 80.25 75.49 5.27 8.10
Schmidt-Hebbel
et al., 1990
O. vulgaris 79.65 76.86 5.31 8.80 Cabello et. al.,
2004
Abulón Haliotis spp 74.56 67.22 2.99 6.17 Agriculture
Handbook, 1987
Mejillón Perna
viridis 80.29 61.59 10.71 13.70
Cabello et. al.,
2004
Ostra Pinctada
imbricada 81.41 68.85 9.52 14.95
Cabello et. al.,
2004
Lenguado Syacium
gunteri 77.20 73.86 3.95 0.79
Castro- González
et al.¸1998
Filete
vacuno 72.70 77.66 14.29 4.03
Schmidt-Hebbel
et al., 1990
Huevo 73.70 49.05 43.73 --- Desrosier, 1999
Arroz 12.00 7.61 0.45 0.57 Desrosier, 1999
Uvas 17.00 3.34 0.60 2.53 Desrosier, 1999
* Determinación en ms
Se han realizado algunos estudios sobre el análisis de las características
nutricionales del calamar gigante. Algunos autores como Gómez-Guillén et al.
(1996a, 1996b, 1997), Gómez-Guillén y Montero (1997); realizaron análisis
proximales de acuerdo a los métodos oficiales de la Association of Official
Agricultural Chemists International 1975, (AOAC, por sus siglas en inglés),
reportando resultados similares en todos los casos en los porcentajes en humedad
(79.9 % + 0.16), lípidos (1.43 % + 0.12), proteínas (18.96 % + 0.15), cenizas (1.36 +
0.05). Realizaron una homogenización del músculo de calamar con hidrocoloides

5
(i-carragenina y fécula) y huevo blanco; y observaron la capacidad de gelación de la
carne de calamar gigante para realizar productos derivados como surimi.
En otros estudios sobre la carne de D. gigas, Abugoch et al. (1999) analizaron el
manto, mediante una caracterización químico proximal: humedad, proteínas (método
Kjeldhal), cenizas, lípidos y calorías (método Bligh y Dyer), obteniendo que el
calamar tiene buenas características nutricionales por el alto contenido de proteínas
(15.32%) y bajo en grasas (0.87 %) con 70 cal/100 g de carne que es más bajo en
grasas que la carne vacuna (proteínas 21.2 %, grasa 3.9 % y 129 cal/100). Además
elaboraron un gel, que puede ser útil en la producción de surimi comercial al
adicionar hidrocoloides como carragenina, alginato de sodio o albúmina de huevo.
Abugoch et al. (2000) analizaron las capacidades emulsionantes de la carne del
calamar D. gigas, almacenada durante 6 meses a de -25º C con la finalidad de
determinar las variaciones del AQP, obteniendo 2.82 g lípidos/g proteína, una
retención de agua de 3.64 g agua/g proteína, con una gelificación nula, lo cual fue
atribuido al tiempo de almacenado. También buscaron condiciones óptimas para la
separación y diferenciación de las proteínas miofibrilares sarcoplasmáticas con un
amortiguador fosfato pH=7.5 y fuerza iónica I= 0.05, de lo cual deducen que las
proteínas de sección corporal de calamar poseen una buena capacidad de
emulsionar y retener agua, por lo que la consideran como buena materia prima para
la elaboración de alimentos después de un periodo de congelación prolongado (seis
meses) excepto para la elaboración de geles.
En el cultivo de camarón (Litopenaeus vannamei) han utilizado al calamar como
fuente alimenticia, sin embargo se dieron a la tarea de determinar variables que
fueran más efectivas para la alimentación del mismo, con lo cual realizaron dos
bioensayos nutricionales de harina a base de calamar para alimentar al camarón, la
primera fue hecha con cada parte del calamar y la otra con cabeza y tentáculo, con
un contenido de proteínas rigurosamente uniforme en ambas dietas. A dichas
mezclas les hicieron el AQP de acuerdo a los métodos descritos por la AOAC, (1990)
donde encontraron proteína cruda de 71.86 % en vísceras y 86.55 % en aletas y en
cuanto a los lípidos, las vísceras presentaron la mayor concentración con un 9.04 %,

6
mientras que el manto tuvo un 2.38 %, la concentración de humedad de las harinas
fue de 0.02 % a 6.9 % (Martínez-Vega et al. 2000a).
Así mismo Martínez-Vega et al. (2000b) realizaron el AQP del calamar gigante
Dosidicus gigas después de un proceso de secado a temperatura ambiente, con el
sol y posteriormente en un cuarto especial, encontrando en el análisis bromatológico
general 85.35 % de proteínas, concluyen que los niveles proteicos se mantienen
elevados después de diferentes procesos de secado.
Es importante saber las características proximales de los alimentos, como por
ejemplo de los moluscos comerciales, entre los cuales destacan pulpos y calamares
(Loligo plei) ya que presentaron el mayor contenido de proteínas y el menor en
grasa, lo que los hace importantes para contribuir en el requerimiento proteico y de
calorías en el hombre; por ello los recomiendan como una alternativa alimenticia con
altos valores nutricionales. Sus resultados demuestran que los valores proteicos de
los moluscos se mantuvieron en un rango de 10.29 % (guacuco, bivalvo de la familia
Veneridae) a 16.62 % (Loligo plei) (Cabello et al., 2004).
Rosa et al. (2005) realizaron una comparación bioquímica (proteínas, lípidos,
glicógeno, colesterol, aminoácidos y ácidos grasos) del calamar Architeuthis sp
respecto a 14 especies de cefalópodos con diferentes estrategias de vida
(bentónicas, noectobentónicas, bentopelágicas y pelágicas) y distintos hábitat
(nerítico, demersal, oceánico y mar profundo). En donde la concentración de
proteínas (>60 %), lípidos (<10 %), aminoácidos (30-40 %) permanece de manera
similar entre las especies.
Finalmente Careche et al. (2006) realizaron un estudio que demuestra que el calamar
D. gigas es un organismo con el cual se pueden elaborar diferentes productos con
valor agregado, como la producción de surimi (extracto de proteínas miofibrilares)
que es de alto nivel nutricional y libre de grasas, tal información es publicada a
manera de nota periodística en el periódico Heraldo de Aragon III Milenio con fecha
16 de mayo de 2006.
Para determinar el consumo de la carne de calamar gigante en México, Luna-Raya et
al. (2006) efectuaron un diagnóstico económico, donde encontraron que si bien hay

7
un aumento de la oferta de este producto en el mercado, los niveles de consumo
siguen siendo muy bajos en el mercado nacional, a pesar de sus características
nutricionales y con un precio inferior a otros productos de fuente de proteínas de
origen animal.
1.2. Identificación y delimitación del problema
La toma de decisiones del ser humano, muchas veces se ve influenciada por la
mercadotecnia, en donde dicen que lo mejor es consumir carne (i.e. res, puerco), sin
embargo ésta tiene algunos puntos en contra que pueden provocar un riesgo para la
salud por el uso de anabólicos esteroides (Bavera et al., 2002).
El consumir en exceso la carne de res, aunado a la mala alimentación dan como
resultado el problema de obesidad y desnutrición. Por lo tanto es importante
determinar un cárnico ideal para consumirlo y promover nuevas alternativas para el
consumo de productos con los suficientes nutrientes para la población en general,
principalmente en el desarrollo de los infantes y para que continúen con una vida
plena durante la etapa adulta.
Los principales requerimientos alimenticios en la dieta humana son: altos niveles de
proteínas y su vez bajos niveles en grasa, sin embargo para personas que ganan un
salario mínimo diario, $67.29 M.N (área geográfica “A”) o $63.77 M.N. (área
geográfica “B”), estimado por la Comisión Nacional de los Salarios Mínimos
(CONASAMI, 2014), sería más factible la adquisición de un alimento rico en
proteínas y de bajo costo, en lugar de productos que cuentan con las características
nutricionales recomendadas, como la milanesa de res ($89.90/kg). Sin embargo, su
costo es elevado en el mercado, fue evaluado según lo estimado mediante
supermercado Soriana (2014) y por lo tanto al no consumir de manera regular una
fuente de proteínas ocasiona que existan deficiencias nutricionales.
La carne de calamar gigante es un producto ideal, ya que además de sus
propiedades nutricionales tiene un bajo costo en el mercado, en cualquiera de sus
presentaciones, filete de manto a $39.90/kg, cabeza a $48.90/kg y brazos cocidos
como imitación de pulpo $82.90/kg (supermercado Soriana, 2014).

8
Es importante estimar los porcentajes nutrimentales que contiene cada sección
corporal (aleta, brazo y manto), porque como en otros organismos como el salmón
que es un pescado con altos niveles de lípidos corporales porque consume en
mayores proporciones ya que es la manera en que se comercializa de manera
general y con ello verificar si existen diferencias bromatológicas entre las distintas
secciones.
Las características nutrimentales de algunos organismos pueden variar ya sea por la
misma ontogenia del animal o por la interacción con el medio ambiente, tal situación
no ha sido evaluada para el calamar gigante por lo que se busca determinar si existe
una variación estacional (mensual) o bien una diferencia nutricional entre machos y
hembras y su respectivo grado de madurez
1.3. Justificación
El reciente auge de la pesquería de calamar gigante y con el aprovechamiento del
75 % de su cuerpo (manto y brazos), así como el conocimiento previo del AQP que
consideran al calamar gigante D. gigas como un alimento de alto valor nutricional,
capaz de satisfacer las necesidades alimenticias, dan la importancia de este recurso
en México. Sin embargo aún no se han evaluado las diferencias estacionales entre
machos y hembras, así como entre su estadio de madurez. Por lo que la presente
tesis permitió conocer la composición químico proximal (%humedad, %proteínas,
%lípidos y %cenizas) de la aleta, brazo y manto en machos y hembras de los
distintos grados de madurez de calamar gigante capturados en 2004 en Santa
Rosalía, BCS.
1.4. Objetivos
1.4.1. Objetivo general
Estimar las variaciones estacionales de la composición químico proximal del
calamar gigante D. gigas en Santa Rosalía, Golfo de California.

9
1.4.2. Objetivos particulares
Comparar la composición químico proximal de cada sección corporal: aleta, brazo
y manto de calamar gigante.
Comparar la composición químico proximal entre machos y hembras de calamar
gigante.
Comparar la composición químico proximal entre los estados de inmadurez,
madurando y maduro de calamar gigante.
Comparar la composición químico proximal entre el mes de recolecta de calamar
gigante.
1.5. Área de estudio
El Golfo de California está ubicado al noroeste de México, con una longitud de 1000
km de largo y ancho promedio de 150 km. Se limita por la Península de Baja
California al oeste y los estados de Sonora, Sinaloa y Nayarit al este. Su batimetría
es tan compleja que incluye una gran cuenca de evaporación abierta al Océano
Pacífico al sur (Álvarez-Borrego, 1983; Álvarez-Borrego y Lara-Lara, 1991; Markaida,
2001).
Se formó por la separación de la falla de San Andrés y se divide en tres secciones, la
más al norte es somero y proveniente de la plataforma continental, mientras que el
resto forma cuencas que llegan hasta 3000 m de profundidad y separadas por
umbrales transversos hasta 1500 m. La costa occidental es rocosa y estrecha,
mientras que la oriental cuenta con lagunas costeras al sur; la más extensa de las
cuencas es la de Guaymas (240 x 60 km, profundidad 2100 m y relieve bajo el
umbral de 480 m) (Álvarez-Borrego, 2008; Markaida, 2001).
El Golfo de California es la única cuenca evaporítica del Pacífico, ya que está
rodeado por el desierto de Sonora, donde gana calor atmosférico y pierde humedad,

10
con lo cual existe una exportación de agua superficial caliente y salina (S 34.90 ‰)
hacia el Pacífico que a su vez se mezcla con agua profunda fría de baja salinidad
(S 34.60 ‰) que va ingresando. Al haber este flujo de los nutrientes y alcanzar la
zona eufótica por mareas y surgencias, se ve reflejado en una productividad alta
(Álvarez-Borrego y Lara-Lara, 1991; Markaida, 2001).
FIGURA 1. Área de afloramiento en el Golfo de California: (A) vientos del noroeste, (B) vientos del sureste, (C) mezcla de mareas (Tomado de Markaida, 2001).
Se ha propuesto un modelo de circulación de tres capas de agua en el Golfo, una
superficial que varía según los vientos de 0 a 50 m, otra hacia fuera de 50 a 250 m y
una hacia adentro de 250 a 500 m. Durante el invierno predominan los vientos del
noroeste y en el verano los del sur o sureste; con lo cual hay un patrón de surgencias
costeras de invierno en la costa continental y en verano en la península, se puede
observar en la Figura 1 (Álvarez-Borrego, 1983; Álvarez-Borrego y Lara-Lara, 1991).
En la cuenca de Guaymas la temperatura superficial es menor durante el invierno
frente a Guaymas, mientras que en el verano es menor en Santa Rosalía. En febrero
las isotermas, isohalinas e isobatas de oxígeno de las capas superficiales surgen

11
hacia el oriente. Se encuentra una termoclina entre abril y octubre, que más fuerte en
agosto (Markaida, 2001). El Golfo de California presenta alta productividad primaria
(0.382 g C m-2 día-1), fosfatos (>0.4 M) que aumenta de sur a norte y biológicamente
tiene una riqueza importante (Álvarez-Borrego y Lara-Lara, 1991, Markaida, 2001;
Álvarez-Borrego, 2008).
Tal es su riqueza, que en el ámbito pesquero que en México los estados de Baja
California, Baja California Sur, Sonora y Sinaloa son los de mayor producción, con el
99 % del total de la pesca de calamar (CONAPESCA, 2011). El calamar gigante se
localiza en dos zonas de pesca principales: Santa Rosalía, Baja California Sur y
Guaymas, Sonora, principalmente sobre el talud continental (200 a 2000 m), sobre
todo en zonas de convergencias (Sato, 1976).

12
2. MÉTODOS
2.1. Captura de organismos
Las muestras recolectadas del calamar gigante D. gigas fueron obtenidas de
muestreos mensuales (abril a octubre excepto junio) de 2004 en la principal zona
pesquera del país: Santa Rosalía, Baja California Sur (Fig. 3). Los organismos se
obtuvieron mediante la pesca artesanal, la cual se realiza en una embarcación menor
que generalmente lleva de dos a tres pescadores, generalmente en el periodo de
penumbra y oscuridad, ésta se encuentra detenida y presenta una fuente de luz que
atrae a los calamares, debido al fototropismo positivo característico de la especie
(Salinas-Zavala et al., 2005).
En el arte de pesca utilizan la potera, que es un anzuelo en forma de huso con
hileras de ganchos que al descender en el agua se mueven y con la luz forma una
bioluminiscencia que atrae al calamar, posteriormente jalan la potera con el calamar
capturado (Salinas-Zavala et al., 2005).
FIGURA 2. Zona geográfica de muestreo de Dosidicus gigas en Santa Rosalía, B. C. S, Golfo de California.

13
2.2. Preparación de la muestra
A cada calamar pescado se le obtuvieron los siguientes datos: Longitud Dorsal del
Manto (LDM) ±0.5 cm, sexo y catalogados de acuerdo a la escala de madurez
propuesta por Lipinski y Underhill (1995) donde los estadios 1-2 corresponden a
organismos inmaduros, 3 organismos en proceso de maduración y 4-5 a organismos
maduros. De cada individuo se separó una muestra de sección corporal (aleta, manto
y brazo) de 100 g, los cuales se mantuvieron congelados a -20º C hasta su
procesamiento en el Laboratorio de Cefalópodos del Centro de Investigaciones
Biológicas del Noroeste (CIBNOR). Posteriormente, las muestras fueron descongelas
por lote mensual, se procediendo a la obtención de la materia seca (ms), conforme a
la metodología del análisis %Humedad, que se describirá en el siguiente apartado.
Obtenidas dichas muestras secas se molieron y se almacenaron en frascos estériles
hasta el momento de sus respectivos análisis.
2.3. Análisis Químico Proximal
La serie de análisis se realizaron en el Laboratorio de Análisis Químico Proximal del
CIBNOR, siguiendo la metodología establecida por la AOAC (1995). A continuación
se resume la determinación de cada análisis y en el Anexo B se explica
específicamente los métodos empleados.
%Humedad: Se realizó por diferencia de peso seco, la muestra se colocó en un crisol
de porcelana y se dejó 24 horas a 70º C en la estufa.
%Proteínas totales: Se realizó mediante el Método Microkjeldahl-Tecator, donde se
obtuvo el producto del porcentaje de nitrógeno y para realizar la conversión se utilizó
el factor 6.25, el cual es el recomendado para este tipo de alimento.

14
Del cual:
%Lípidos totales: Se realizó la extracción de las muestras con 250 mL de éter de
petróleo y se dejaron 6 horas en el extractor.
%Cenizas: Se realizó por diferencia de peso carbonizado, la muestra se colocó en un
crisol de porcelana y se dejó 24 horas a 500º C en la mufla.
El análisis de humedad fue el único realizado con materia húmeda, el resto de los
análisis (%proteínas, %lípidos y %cenizas) fueron con ms.
2.4. Análisis estadísticos
Se utilizó el programa Statistic 10 donde se realizaron los Análisis de Variancia
(ANOVA) de una vía para encontrar la magnitud de cada fuente de variación (tipo de
sección corporal, sexo, madurez y mes de colecta).
Posteriormente se realizó una prueba DVS (Diferencia Verdaderamente Significativa)
de Tukey para identificar su fuente de variación, en donde el nivel de significancia
está en función de los valores de p: significativo p< 0.05, significativo p< 0.01,
altamente significativo p< 0.001 y no significativo p> 0.05.
V1 = ml de HCl gastados en la titulación muestra
V0 = ml de HCl gastados en la titulación blanco
N = normalidad del HCl
g = peso de la muestra

15
3. RESULTADOS
3.1. Estructura de tallas
Se analizó la sección corporal de 157 calamares gigantes (D. gigas), de los cuales 39
machos y 118 hembras. El rango de tallas fue de 15.0 a 80.0 LDM cm, así mismo se
identificaron dos modas en hembras (46 y 68 LDM cm) y dos en machos (46 y 56
LDM cm) (Fig. 3).
FIGURA 3. Estructura de las tallas de calamar gigante Dosidicus gigas, capturados en Santa Rosalía B. C. S. durante 2004.
3.2. Análisis Químico Proximal
Los resultados obtenidos del AQP fueron: %humedad, %proteínas, %lípidos y
%cenizas. A continuación se describe cada uno de los resultados encontrados.
El primer resultado que se obtiene en el AQP es el %humedad, en donde de manera
general el calamar gigante (D. gigas) tuvo una estimación de 78.34 % de humedad y
el resto fue considerado como ms (21.66 %).
En el presente trabajo se determinó que la carne de el calamar gigante es un
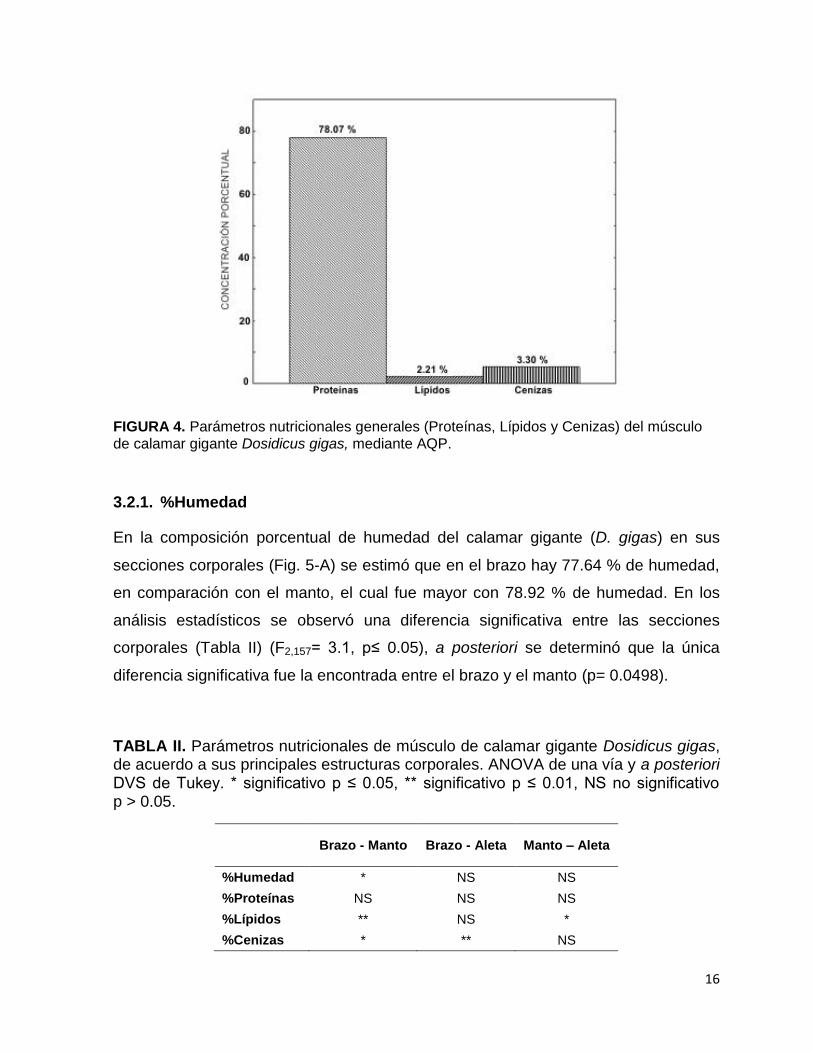
producto con alto nivel proteico (78.07 %) y bajo en grasa (2.21%) (Fig. 4), índices
que lo pueden clasificar como alimento altamente nutritivo, sin embargo también se
toman en cuenta los minerales (cenizas: 5.30 %).
16
FIGURA 4. Parámetros nutricionales generales (Proteínas, Lípidos y Cenizas) del músculo de calamar gigante Dosidicus gigas, mediante AQP.
3.2.1. %Humedad
En la composición porcentual de humedad del calamar gigante (D. gigas) en sus
secciones corporales (Fig. 5-A) se estimó que en el brazo hay 77.64 % de humedad,
en comparación con el manto, el cual fue mayor con 78.92 % de humedad. En los
análisis estadísticos se observó una diferencia significativa entre las secciones
corporales (Tabla II) (F2,157= 3.1, p≤ 0.05), a posteriori se determinó que la única
diferencia significativa fue la encontrada entre el brazo y el manto (p= 0.0498).
TABLA II. Parámetros nutricionales de músculo de calamar gigante Dosidicus gigas, de acuerdo a sus principales estructuras corporales. ANOVA de una vía y a posteriori DVS de Tukey. * significativo p ≤ 0.05, ** significativo p ≤ 0.01, NS no significativo p > 0.05.
Brazo - Manto Brazo - Aleta Manto – Aleta
%Humedad * NS NS
%Proteínas NS NS NS
%Lípidos ** NS *
%Cenizas * ** NS

17
El contenido de agua medida en ambos sexos de D. gigas difiere entre sí (Fig. 5-B).
En hembras se estimó con 77.95 % de humedad, mientras que los machos con 79.63
% de humedad. Esta tendencia tiene diferencias significativas entre ambos sexos
(F1,154= 10.59, p< 0.05). Al ser analizados, se comparó estadísticamente su
discrepancia dando una diferencia altamente significativa (p= 0.0011).
Debido a que se encontró la diferencia significativa de humedad entre los sexos de
calamar gigante (D. gigas), el análisis de estado de madurez se dividió en machos y
hembras. En machos (Fig.5-C), el porcentaje de humedad menor se registró en
individuos que estaban en proceso de maduración (77.35 %), mientras que el valor
máximo correspondió a los individuos maduros (80.91 %), lo cual mostró una
diferencia significativa (F2,36= 6.33, p< 0.01). Entre las comparaciones de los tres
estados de madurez se determinó que la diferencia fue significativa entre los
organismos en proceso de maduración y los maduros (p= 0.0047).
Por otro lado, en hembras D. gigas (Fig.5-D), el porcentaje de humedad menor se
encontró en hembras maduras (77.09 %) y los organismos que obtuvieron la
concentración máxima en hembras con proceso de maduración (78.24 %). Sin
embargo, no se identificó una diferencia significativa (F2,109= 0.64, p> 0.05).
En el análisis de %humedad correspondiente al mes de colecta de D. gigas (Fig. 5-E)
se encontró que mayo obtuvo el mínimo con 75.58 % y septiembre el máximo con
82.40 %. Se determinó que existieron diferencias significativas entre los seis meses
comparados (F5,154= 45.9, p< 0.001), las comparaciones que permanecieron
similares fueron entre los meses: abril y agosto, abril y septiembre, julio y agosto con
p< 0.05.

18
FIGURA 5. Porcentajes de humedad del calamar gigante Dosidicus gigas capturado en Sta. Rosalía B.C.S durante 2004. A: sección corporal, B: sexos, C: estadio de madurez en machos, D: estadio de madurez en hembras y E: mes de colecta.

19
3.2.2. %Proteínas totales (Pt)
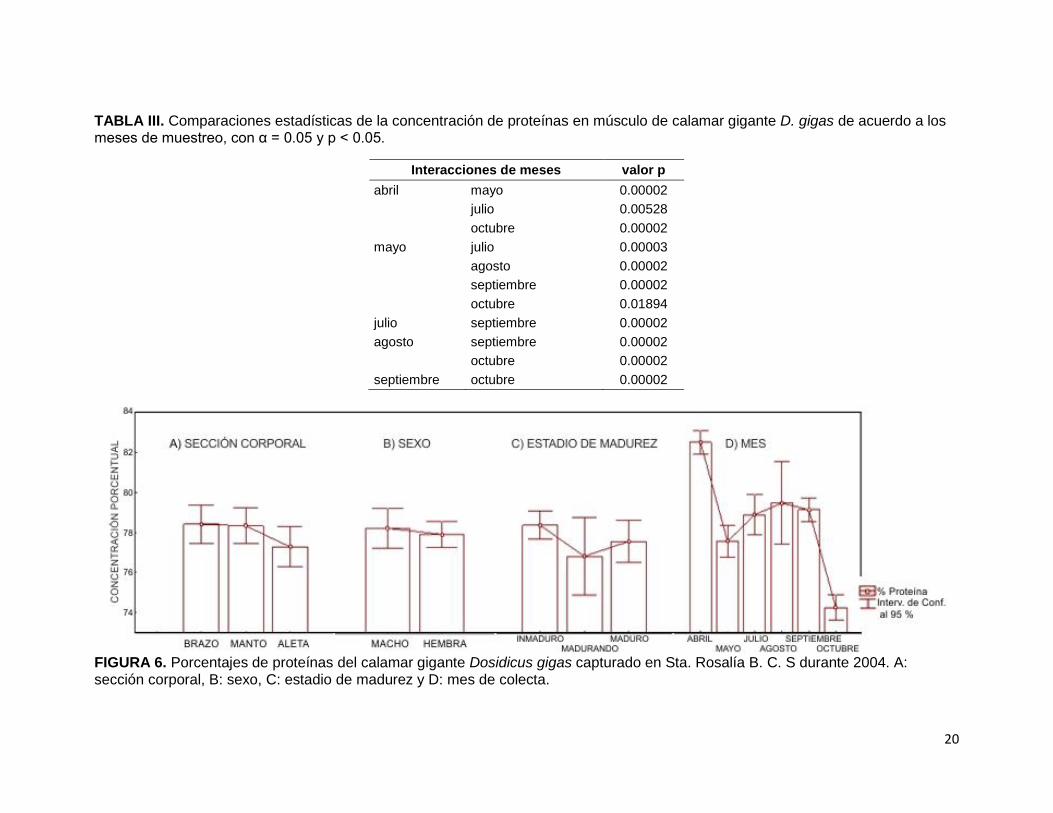
En la figura 6-A se muestran los porcentajes de proteínas en las diferentes secciones
corporales del calamar gigante. Se estimó que los valores mínimos fueron en aleta
(77.28 % de Pt), mientras que los máximos fueron en brazo (78.48 % Pt). Sin
embargo, al realizar las comparaciones estadísticas (Tabla II) se encontraron
similitudes en la concentración de proteínas entre la aleta, el brazo y el manto
(F2,136= 1.74, p> 0.05).
El percentil proteico determinado entre ambos sexos (Fig. 6-B) demostró que las
hembras presentan la concentración mínima (77.94 % Pt), mientras que los machos
presentan la concentración máxima (78.24 % Pt). En la comparación estadística
entre ambos sexos no existió diferencias significativas (F1,133= 0.22, p> 0.05) en la
concentración de proteínas.
Los porcentajes de Pt encontrada en D. gigas según su grado de madurez (Fig. 6-C)
indicaron que los organismos en proceso de maduración tienen el valor mínimo
(76.81 % Pt) y los inmaduros tienen el máximo (78.37 % Pt). De esta tendencia se
encontró diferencia significativa entre los grados de madurez (F2,130= 3.61, p< 0.05),
sin embargo la diferencia verdaderamente significativa se obtuvo entre los
organismos inmaduros y en proceso de maduración (p= 0.0418).
En la figura 6-E se observan los porcentajes de Pt del calamar gigante en los meses
de recolecta. El porcentaje mínimo fue en octubre con 74.70 %, mientras que el
máximo fue abril con 82.52 % y con diferencias significativas entre los meses
(F5, 133= 29.6, p< 0.001). Las relaciones entre los meses de mayo, julio, agosto y
septiembre permanecieron similares (p> 0.05) y el resto de las interacciones tuvieron
una diferencia significativa (p< 0.01) en el percentil proteico de calamar gigante D.
gigas, como se muestra en la tabla III.
20
TABLA III. Comparaciones estadísticas de la concentración de proteínas en músculo de calamar gigante D. gigas de acuerdo a los meses de muestreo, con α = 0.05 y p < 0.05.
Interacciones de meses valor p
abril mayo 0.00002
julio 0.00528
octubre 0.00002
mayo julio 0.00003
agosto 0.00002
septiembre 0.00002
octubre 0.01894
julio septiembre 0.00002
agosto septiembre 0.00002
octubre 0.00002
septiembre octubre 0.00002
FIGURA 6. Porcentajes de proteínas del calamar gigante Dosidicus gigas capturado en Sta. Rosalía B. C. S durante 2004. A: sección corporal, B: sexo, C: estadio de madurez y D: mes de colecta.

21
3.2.3. %Lípidos totales
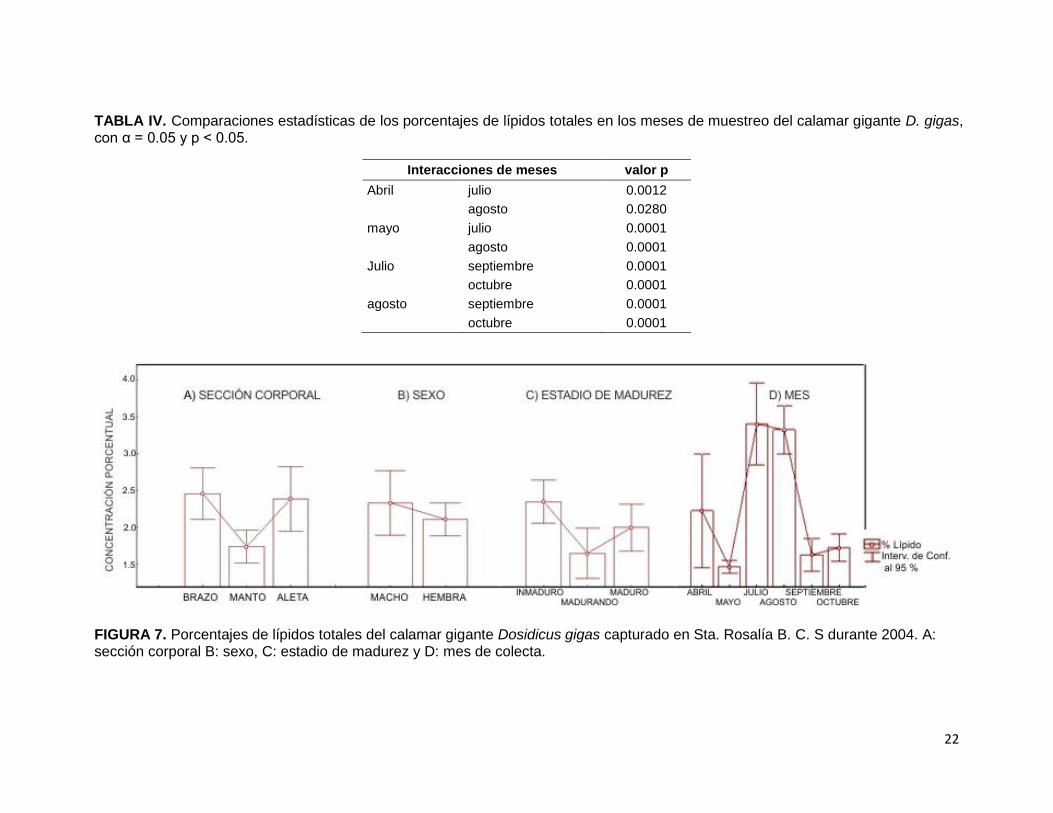
En el AQP, generalmente las grasas se encuentran en menores cantidades al resto
de los nutrientes. En el momento de realizar el estudio en los diferentes tipos de
sección corporal del calamar gigante D. gigas (Fig. 7-A) se encontró que el manto fue
la zona con el menor valor porcentual de lípidos totales (1.74 %), mientras que en el
brazo el mayor valor de lípidos (2.46 %). El análisis estadístico resultó que dichos
valores marcaron una diferencia significativa (Tabla II; F2,120= 6.50, p< 0.01). Esta
tendencia entre las secciones corporales de calamar gigante manifestó
estadísticamente diferencias significativas. Los resultados indicaron que los lípidos
entre el brazo y el manto son diferentes (p= 0.0043) y de igual forma el manto y la
aleta (p= 0.0165) difieren en la concentración de lípidos.
En el %lípidos totales se estimó que las hembras (Fig. 7-B) tienen la proporción
menor de concentración de lípidos (2.11 %), mientras que los machos cuentan con
una proporción mayor en la concentración de lípidos (2.34 %). Esta distinción,
prácticamente nula, estadísticamente hablando, no marcó algún tipo de diferencia
significativa (F1,117= 0.9414, p> 0.05) entre la determinación de lípidos por sexo.
De manera similar ocurrió con el análisis de grasas totales de los estadios de
madurez del calamar gigante (Fig. 7-C) se determinó estadísticamente que sus
diferencias son prácticamente nulas y no significativas (F2,115= 3.0173, p> 0.05). A
pesar de las distinciones, el valor mínimo corresponde a los organismos en estado
de maduración (1.65 %) y el valor máximo a los organismos maduros (2.55 %), es
decir la concentración de lípidos es independiente al estado de madurez de los
organismos.
En la figura 7-D se observan los porcentajes de los lípidos totales de D. gigas de los
mes de colecta, mayo presentó el valor mínimo (1.57 %), mientras que julio obtuvo el
valor máximo (3.39 %), se encontraron diferencias significativas (F5,117= 25.3489,
p< 0.001). Las comparaciones que hicieron tales diferencias se muestran en la tabla
IV, donde se evidencia que hay diferencia significativa en ocho de las 15
permutaciones para los diferentes meses.
22
TABLA IV. Comparaciones estadísticas de los porcentajes de lípidos totales en los meses de muestreo del calamar gigante D. gigas, con α = 0.05 y p < 0.05.
Interacciones de meses valor p
Abril julio 0.0012
agosto 0.0280
mayo julio 0.0001
agosto 0.0001
Julio septiembre 0.0001
octubre 0.0001
agosto septiembre 0.0001
octubre 0.0001
FIGURA 7. Porcentajes de lípidos totales del calamar gigante Dosidicus gigas capturado en Sta. Rosalía B. C. S durante 2004. A: sección corporal B: sexo, C: estadio de madurez y D: mes de colecta.

23
3.2.4. %Cenizas
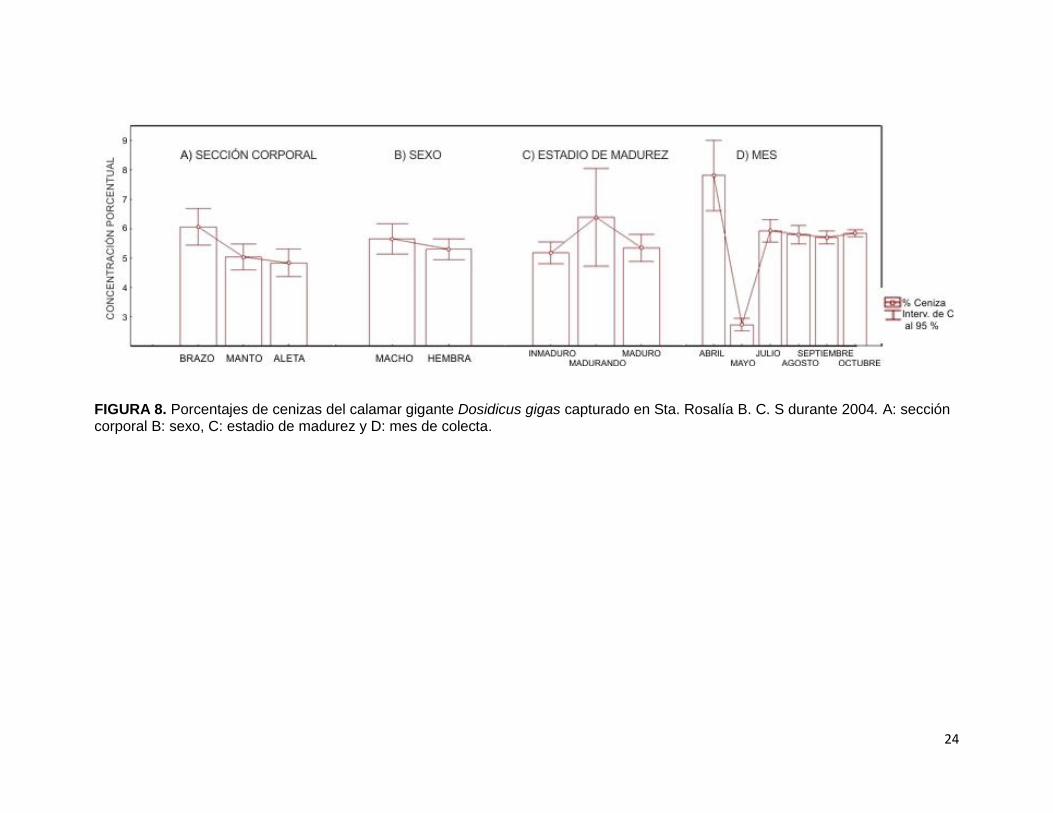
En la figura 8-A se observan los niveles del porcentaje de cenizas en las secciones
corporales de calamar gigante D. gigas, donde se puede observar que la aleta
presentó el nivel mínimo (4.82 %), mientras que el manto el nivel máximo (6.05 %).
Estas diferencias resultaron significativas (Tabla IV; F2,109= 6.2200, p< 0.01) y
específicamente entre brazo y manto (p= 0.0104) y entre brazo y aleta (p= 0.0066).
El percentil de cenizas entre ambos sexos de calamar gigante D. gigas (Fig. 8-B)
demostró que las hembras obtuvieron el porcentaje mínimo (5.29 %), mientras que
los machos el porcentaje máximo (5.65 %). Dichos valores dieron como resultado
una diferencia no significativa (F1,106= 0.8819, p> 0.05), por lo tanto hembras como
machos tienen proporciones similares de cenizas.
Los porcentajes de cenizas en los estadios de madurez de calamar gigante D. gigas
(Fig. 8-C) demuestran que los organismos inmaduros presentaron el valor mínimo
(5.17 %), mientras que los que están madurando presentaron el valor máximo
(6.38 %). Se encontraron diferencias significativas (F2,106= 28.1792, p< 0.001).
En las comparaciones de los grados de madurez marcaron dos tipos de diferencias
estadísticas, la primera diferencia significativa fue entre los calamares gigantes
inmaduros y los madurando (p= 0.0206) y los organismos que están madurando y los
maduros (p= 0.0447). La segunda diferencia altamente significativa (p= 0.0001) fue
entre los organismos inmaduros y maduros, por lo tanto se puede decir que la
concentración de cenizas difiere entre los tres estadios de madurez.
Al realizar el análisis de cenizas de calamar gigante D. gigas en los meses de colecta
(Fig. 8-D) se observó que el porcentaje menor fue mayo (2.72 %), mientras que el
porcentaje mayor fue abril (7. 78 %), esta tendencia provocó que existieran
diferencias significativas entre sí (F5, 106= 86.231, p< 0.001). Dichas diferencias
altamente significativas (p= 0.0001) presentaron que los porcentajes de cenizas de
abril y mayo fueron diferentes al resto de los meses.
24
FIGURA 8. Porcentajes de cenizas del calamar gigante Dosidicus gigas capturado en Sta. Rosalía B. C. S durante 2004. A: sección corporal B: sexo, C: estadio de madurez y D: mes de colecta.

25
4. DISCUSIÓN
Estructura de tallas
Las tallas de los organismos capturados para la realización de los AQP mostraron
dos modas en machos (46 y 56 LDM cm) y dos modas en hembras (48 y 68 LDM
cm), ambas pertenecientes al grupo de talla mediano de acuerdo a la propuesta de
Nigmatullin et al., 2001. A su vez mantienen la característica de la dinámica
poblacional en donde los machos son más pequeños que las hembras tal como ha
sido reportado por Nigmatullin et al., 1991; Nevárez-Martínez, et al. 2000; Bazzino, et
al., 2007; Mejía-Rebollo et al., 2008.
Cabe destacar, de acuerdo a los datos de CONAPESCA (2013), el calamar es una
especie de exportación y en México la pesquería de este recurso se encuentra en el
noveno lugar de extracción de 52 especies marinas, posicionado a Baja California
Sur como el principal productor de calamar (56.57 % del total de la pesca). Sin
embargo una gran parte de la pesca la dirigen hacia países asiáticos, europeos y de
norteamérica, donde posteriormente le dan un valor agregado transformándolo en
distintos alimentos tales como surimi, aritos empanizados o fritos, carne seca de
calamar, hamburguesa, botanas secas, entre otros.
Principalmente, la carne del calamar gigante se congela para su transporte antes de
ser procesado, Ramírez et al. (2004) y Dublán-García (2006) han corroborado que
los valores nutricionales se mantuvieron a pesar de ser congelado, por lo tanto en el
presente trabajo se procedió a congelar las muestras de calamar gigante D. gigas.
Análisis Químico Proximal
El porcentaje de humedad general del calamar gigante Dosidicus gigas fue 78.34 %,
el cual es un valor similar a los reportados con anterioridad para la especie, los
cuales van del 79 % al 82 % tal como se muestra en la tabla I. Por otro lado al
compararlo con otros alimentos considerados de alta calidad nutricional, el filete de
lenguado presenta un porcentaje de humedad de 77.2% (Castro-González et al.,
1998), concentración similar al calamar gigante. De igual forma el filete vacuno tiene

26
un porcentaje elevado de humedad con un 72.7 % (Schmidt-Hebbel et al., 1990) y
finalmente al comparar el porcentaje de humedad con un producto de origen vegetal
como el grano de arroz se pudo observar que el nivel de humedad de este último es
considerablemente menor ya que presenta un percentil de 12 % (Desrosier, 1999), lo
cual es de esperarse al tratarse de un grano que se comercializa seco. De acuerdo a
lo anterior, la carne de calamar gigante es considerada adecuada para el consumo
humano debido al porcentaje de humedad que contiene.
Tras analizar las macromoléculas utilizadas para evaluar la propiedad nutricional de
los alimentos destaca la interacción proteínas-lípidos ya que son de los componentes
más importantes para la nutrición de las personas, porque en un alimento alto en
proteínas y bajo en grasas es ideal para el sano desarrollo humano, según la OMS
(Food and Nutrition Board/Institute of Medicine, 2002).
El calamar gigante D. gigas cumple con los requisitos para ser una carne nutritiva por
el alto nivel proteíco (78.02 %) y bajo en grasas (2.32 %), tal como la carne de res
con 77.66 % de proteínas y 14.29 % de grasas (Schmidt-Hebbel et al., 1990) y el
pulpo con 76.86 % de proteína y 5.31 % de grasas (Cabello et al., 2004). Sin
embargo al comparar la carne de calamar con un alimento de la canasta básica
como el huevo, se puede corroborar que es más nutritivo ya que el huevo tiene 49.05
% de proteínas y 43.73 % de grasas (Desrosier, 1999). Estas referencias encuentran
sintetizadas en la tabla I.
%Humedad
El AQP inicia con la determinación del porcentaje de humedad, cabe mencionar que
a pesar de que el agua por sí sola no se cuenta como un agente nutritivo, la
humedad es una característica importante para la conservación del producto y su
procesamiento, razón por la cual se ha dedicado un apartado a este parámetro.
Relacionado con ello se sabe que un alimento de calidad debe contener por lo
menos de 18 a 22 % de humedad (Desrosier, 1999).

27
Los microorganismos necesitan del agua para desarrollarse, teniendo en cuenta ello,
entre mayor sea la concentración de agua, tiende a aumentar el crecimiento de
microorganismos (actividad de agua), Aunque existen otros factores que influyen
como el pH y temperatura (Frazier, 1978, Christian, 2000). En el caso del calamar
gigante, tan altos son los valores de %humedad que se ven reflejados en que
estando en refrigeración a 4º C solo se mantiene en buen estado para ser consumido
de 1 a 2 días (Dublán-García 2006; FDA, 2014).
En el presente trabajo se estimó el %humedad, en el manto de calamar fue 78.92 %,
resultado similar a lo reportado por Gómez-Guillén et al. (1997) con 79.9 % y
Abugoch et al. (1999) con 82.23 % de humedad. De igual manera en otros tipos de
moluscos la proporción de humedad se encuentran entre un rango de 70 % a 80 %
como es el caso del abulón Haliotis spp que tiene 74.56 % de humedad (Cabello et
al., 2004) y el pulpo Octopus vulgaris con 79.65 % de humedad (Agriculture
Handbook, 1987).
Por otro lado, al comparar cárnicos provenientes del mar con provenientes de
ganadería, se observó que el filete tiene 72.70 % de humedad (Schmidt-Hebbel et
al., 1990), se encuentra con valores similares al calamar gigante, al igual que el
huevo que contiene 73.70 % de humedad (Desrosier, 1999). Lo cual reafirma que
calamar gigante respecto a su cantidad de humedad es un producto de excelente
calidad para su consumo directo o bien para la elaboración de alimentos procesados
como los que antes se habían indicado.
%Proteínas totales
La proteína es un elemento sumamente importante dentro de los alimentos porque
son las que dan paso a diferentes actividades internas en el cuerpo como el
movimiento de los músculos, la tensión de la piel y la formación de anticuerpos. Al
determinarse el percentil proteico en el manto (78.48 %) de calamar gigante D. gigas,
se llega a la conclusión que es un cárnico con altos niveles de proteínas.

28
Al comparar el calamar gigante D. gigas con otros cárnicos se observa que Martínez-
Vega et al., (2000b) que obtuvieron 83.61 % de proteínas para calamar gigante, el
porcentaje en el pulpo Octopus vulgaris se mantuvo con 76.86 % de proteínas, el
lenguado Syacium gunteri con 73.86 % de proteínas (Castro-González et al., 1998),
a su vez también se conservó en un mismo parámetro la carne de res con 77.66 %
de proteínas (Schmidt-Hebbel et al., 1990). Se puede decir que estos tipos de carnes
se encuentran con percentiles similares, que quiere decir que tienen un nivel de
proteínas alto. Determinando que el filete de calamar gigante es un alimento con un
valor nutricional suficientemente alto para ser considerado como platillo principal en
una comida.
El calamar gigante al ser comparado con otros organismos marinos costeros
adheridos a sustratos duros como el abulón Haliotis sp que presentó 67.22 % de
proteínas (Agriculture Handbook, 1987) y el mejillón Perna viridis con 61.50 % de
proteínas (Cabello et al., 2004), se puede observar la diferencia proteíca entre un
organismo bentónico y un organismo del necton, ya que el calamar gigante al ser un
organismo nadador tiene más desarrollada la musculatura y por ende requiere de
mayor cantidad de proteínas para que dichos músculos operen correctamente.
%Lípidos totales
El porcentaje de lípidos se encuentra dentro del AQP; en el presente trabajo, el
%lípidos en el manto de calamar gigante D. gigas fue de 1.75 %, lo cual es similar a
lo reportado por Martínez-Vega et al. (2000b) con 2.31 %, mientras que Abugoch et
al. (1999) obtuvieron 4.90 % de lípidos. Todos estos porcentajes se encuentran en
menor proporción que el filete vacuno que contiene 14.29 % de lípidos (Schmidt-
Hebbel et al., 1990) y el huevo que contiene casi la mitad de su contenido nutricional
es de lípidos (43.73 %, Desrosier, 1999).
El filete del calamar gigante es comparable con otras carnes con similitudes
nutricionales, como es el filete de res, sin embargo éste último contiene una
proporción de grasa hasta 7 veces más que el calamar, por ello no es recomendable

29
un consumo excesivo. Se confirma que la carne del calamar gigante es un producto
saludable para consumo humano por contener mínimas proporciones de grasa.
%Cenizas
Por otro lado los minerales son bioelementos necesarios para la nutrición y de
manera general se considera que todos los seres vivos tenemos aproximadamente el
4.5 % de bioelementos (Badui-Dergal, 1993; Desrosier, 1999), los cuales se
determinan en conjunto dentro del análisis de cenizas representado como el
porcentaje de contenido inorgánico dentro de un alimento (Kirk, et al., 2000;
Hernández-Triana, 2004).
En el presente trabajo se estimó 5.01 % de cenizas en el manto de calamar gigante
D. gigas, dato que concuerda con los resultados obtenidos por Martínez-Vega et al.
(2000b) con 5.9 % de cenizas, Gómez-Guillén et al. (1997) con 6.77 % de cenizas y
Abugoch et al. (1999) con 7.37 % de cenizas.
Al realizar la comparación de cenizas de calamar con otros tipos de moluscos, se
observó que se encuentran en un rango similar de 5 % a 8 % (Tabla I), como por
ejemplo el pulpo Octopus vulgaris con 8.10 % de cenizas (Schmidt-Hebbel et al.,
1990) y el abulón Haliotis sp con 6.17 % (Agriculture Handbook, 1987). Al compararlo
con otro tipo de carne de origen diferente al marino como el filete vacuno se puede
observar que la concentración de micronutrientes en el calamar es superior al del
filete de res que contiene 4.03 % de cenizas.
El calamar gigante contiene un porcentaje de micronutrientes (cenizas) superior a la
media reportada para los organismos vivos en general. Además de ello el calamar
gigante puede cubrir prácticamente la totalidad de micronutrientes sugeridos (7 % a
9 %) para una dieta saludable por la FDA.

30
Variación del AQP por sección corporal
Las secciones corporales de calamar gigante D. gigas que se emplearon para el AQP son aleta, brazo y manto, puesto
que son las partes comerciadas para el consumo humano. Al analizar las cualidades nutricionales del calamar gigante
según el tipo de sección corporal (Fig. 9-A), se detectaron diferencias significativas entre el brazo con las demás
secciones analizadas (manto y aleta), así como incrementos en los percentiles de cenizas y lípidos para brazo (Tabla II).
FIGURA 9. AQP del calamar gigante Dosidicus gigas capturado en Sta. Rosalía B. C. S durante 2004. A: sección corporal, B: sexo, C: estadio de madurez y D: mes de colecta.

31
Esto se explica en términos fisiológicos, ya que el calamar gigante D. gigas utiliza las
principales secciones corporales de diferente manera. Al respecto se tiene que el
movimiento de los apéndices está relacionado con altas concentraciones de
minerales, debido a que el Calcio como micronutriente es el transmisor del impulso
nervioso que da paso a la contracción muscular (Hernández-Triana, 2004; Fernández
et al., 2011), por lo cual requieren una mayor concentración de nutrientes,
expresadas en %cenizas. Fisiológica y anatómicamente los brazos del calamar
gigante tienen una mayor actividad a diferencia del resto del cuerpo, puesto que los
utilizan en la alimentación, con los brazos atrapan a la presa y en la reproducción
(machos) con su hectocotilo (brazo especializado) transmiten el espermatóforo
(Barnes, et al., 2001). Por lo tanto en el brazo requieren una mayor concentración de
energía a diferencia del manto y ésta es obtenida de los lípidos (Layzer, 1991;
Barnes et al., 2001; Pérez-Ruiz y Lucía-Mulas, 2007).
Variación del AQP por sexo
En estudios previos donde han realizado AQP en el músculo de calamar gigante no
han diferenciado los organismos machos de las hembras, por lo tanto no se cuenta
con información comparable.
En el AQP (Fig. 9-B) se observó una ligera elevación porcentual de los machos,
donde los valores de proteínas, lípidos y cenizas se encontraron con una diferencia
de 0.30 % arriba de las hembras, sin embargo no se detectó diferencia significativa
(p> 0.05). En el caso de la humedad el incremento resultó mayor (1.66 %), el cual se
corroboró estadísticamente con una diferencia altamente significativa (p<0.001).
Los resultados obtenidos en el presente trabajo se pueden relacionar con sus
estrategias reproductivas y no sincronizadas, es decir que los machos maduran a
edades y tallas menores que las hembras (Paredi,1996; Markaida y Sosa-Nishizaki,
2001; Mejía-Rebollo, 2006), debido a ello hembras y machos redirigen la energía
necesaria para el proceso de la madurez hacia su respectivo aparato reproductor en
momentos diferentes, encontrando variaciones químico proximales en su organismo
(Paredi, 1996), lo cual concuerda con lo encontrado en el presente estudio.

32
Variación del AQP por estado de madurez
No existe algún estudio que permita relacionar el AQP del calamar gigante D. gigas
según su estado de madurez. De acuerdo a los resultados estimados (Fig. 9-C) en el
presente trabajo, el calamar gigante los organismos maduros (hembras o machos)
contienen una mayor concentración de humedad en un 1.86 % (p< 0.01), proteínas
en un 2.52 % (p> 0.05) y lípidos en un 0.90 % (p> 0.05) respecto a los que están en
proceso de maduración.
Sin embargo Paredi (1996) realizó un estudio al respecto para el calamar Illex
argentinus, en donde trató de identificar las fases de desnaturalización de la
proteínas del músculo de acuerdo al sexo y estado de madurez. Realizó una serie de
experimentos que se centran en elevar la temperatura en intervalos de 10º C por
minuto en un rango de 10º C a 100º C; sin embargo no encontraron diferencias
mayores, su aportación principal se centra en identificar distintas transiciones de
desnaturalización por tipo de proteínas (sarcoplasmática, actina, miosina y
paramiosina), más que por sexo o madurez.
Los machos contienen mayor concentración de agua, es que la formación de los
espermatóforos en machos maduros es el proceso más especializado en la biología
de los cefalópodos, lo cual le ha permitido producir una gran cantidad de esperma y
empaquetarla en los espermatóforos, transferirlos con el hectocotilo (brazo
especializado) hacia la hembra y mantenerlos viables hasta la fecundación (Morton,
1979; Mille-Pagaza, 2008). Ya que la parte interna de los espermatóforos está
constituida por una masa viscosa en su mayoría compuesta de agua, explicando con
ello los altos requerimientos de humedad en machos maduros.
Variación del AQP entre meses
Es importante llevar a cabo un análisis químico proximal comparativo de calamar
gigante de manera anual en Santa Rosalía B.C.S., puesto que nos encontramos en
un medio cambiante, en donde influyen varios factores como los climáticos, sin
embargo no existen trabajos de referencia con los que se comparen directamente.

33
Se detectaron variaciones en la concentración nutrimental a lo largo del tiempo (Fig.
9-D), específicamente en los análisis de proteínas, lípidos y cenizas, lo cuales
tuvieron valores elevados durante julio y agosto y disminuyeron en los meses de
septiembre y octubre, con lo cual se podría deducir que en Santa Rosalía se
encuentran las condiciones ambientales óptimas durante los meses cálidos (julio y
agosto), que permiten que el calamar tenga mayores concentraciones en los
parámetros nutricionales mencionados.
Relacionado con lo anterior Markaida et al. (2005) y López et al. (2006) mencionan
que existe una mayor concentración de calamares en la zona de Santa Rosalía en
los meses cálidos, ya que existe una migración horizontal dentro del Golfo de
California, en donde los calamares se mueven de Guaymas hacia Santa Rosalía,
debido a las surgencias presentes en la zona, que a su vez favorecen la alta
disponibilidad de presas para su alimentación. Esto implicaría que al tener una fuente
de alimento abundante el calamar estaría en mejores condiciones nutricionales, lo
podría ser detectado en el AQP.
Esto concuerda con los índices nutricionales obtenidos, en donde los calamares
capturados en julio y agosto tuvieron mayores porcentajes que los organismos de
septiembre y octubre.
Consumo del calamar gigante
Finalmente, cuando se realiza una comparación del análisis químico proximal se
puede determinar que el calamar gigante es un producto altamente nutritivo, por sus
niveles proteico alto y graso bajo, al igual que funge como una alternativa ideal como
ingrediente principal en una comida, aunado a que es alimento de costo comercial
bajo comparándolo con otros alimentos de origen marino como el pulpo ($84.90/kg) y
camarón chico ($175.90/kg), filete de pescado bagre ($56.90/kg) u otro más
comunes entre la población como la carne de res ($89.90/kg) costos provenientes del
supermercado Soriana (2014), además de ser utilizado en la fabricación de otros
productos procesados como el surimi.

34
El filete de res es un cárnico que se considera con un buen valor nutricional, sin
embargo también presenta un alto costo comercial ($89.90 kg), a comparación del
filete de calamar que es una carne con excelentes y mejores niveles nutricionales ya
que al mismo tiempo que presenta alta concentración de proteínas, su grasa es 7
veces menor que la de res siendo el filete de calamar un alimento nutritivo y de un
costo menor, de prácticamente la mitad ($39.90 kg).
Los costos de una clase de producto son variables, ya que ante la demanda existen
varios factores que influyen, Samuelson et al. (2004) mencionan que estos factores
son el ingreso medio (salario), la población (lugar donde viven), los precios de los
bienes relacionados (en este caso, costos de diferentes cárnicos), los gustos (como
algo subjetivo) y los elementos especiales (alergias, comida rápida, carnes
precocidas). En el caso específico del calamar el costo está afectado directamente
por la estructura de mercado de intermediarios, la cual está constituida por la
cooperativa pesquera, permisionario, venta directa a la planta y venta a otro
intermediario (De la Cruz-González, 2007).
Una estrategia para incrementar el consumo del calamar gigante en la región de ha
realizado anualmente desde 2004 a la fecha en la ciudad de Guaymas, Sonora;
dicha estrategia consiste en un evento de difusión denominado “Feria del Calamar
Gigante”. Inicialmente la feria de calamar empezó con la ayuda de los mismos
pescadores y sus familias, quienes hacían la invitación al consumo del calamar por
medio de una degustación de diferentes platillos en donde el ingrediente principal era
el calamar gigante en distintas modalidades como ceviche de calamar, escabeche de
calamar, machaca de calamar, barbacoa de calamar, calamar ranchero, calamar
ahumado, calamar al chipotle, calamar al tocino, calamar al horno, tamales de
calamar, salpicón de calamar, chorizo de calamar, entre otros (Luna-Raya, 2008).
Posterior a la degustación de los platillos se realizaron encuestas a los asistentes,
que indicaron que la frecuencia de consumo del calamar gigante fue baja, debido a
que el 30 % de los encuestados consumen calamar una vez al mes y se incrementó
hasta el 90 % de los asistentes (Luna-Raya, 2008).

35
En las encuestas derivadas de la Feria de Calamar, Luna-Raya (2008) observó la
relación de los costos comerciales del calamar con la disponibilidad de pagar por la
carne, después de haber asistido a la feria del calamar, en donde el precio promedio
que estaban dispuestos a pagar fue $18.00/kg de calamar gigante fresco y $26.00/kg
de calamar gigante cocido, cabe destacar que la disponibilidad de pago estuvo por
encima del precio real, ya que en marzo de 2004 el precio fue de $8.00/kg, en marzo
de 2005 de $11.00/kg y en marzo de 2006 de $12.00/kg. Lo cual fue el reflejo de que
los asistentes consideraran que el calamar gigante es un alimento de buena calidad
por el cual estarían dispuestos a pagar una cantidad mayor del precio de mercado
local.
Iniciativas, como la Feria del Calamar Gigante en Guaymas, Son. son las que
permiten la promoción de éste como alimento altamente nutritivo y en donde, hasta el
momento se ha venido celebrado dicho evento ininterrumpidamente desde 2004
hasta 2013, cuando se llevó a cabo la décima Feria de Calamar Gigante en el mes
de octubre, participando autoridades de los tres órdenes de gobierno. Destacando el
éxito del evento a nivel regional, que ha cumplido con el objetivo de difusión y
promoción del consumo de calamar gracias a las degustaciones y venta de platillos,
haciendo que una carne tan nutritiva como el calamar gigante pueda aparecer en la
lista de compras al momento de realizar la despensa por familia y tengan la certeza
que al consumirlo se puedan alimentar mejor.
Para la ciudad de La Paz se podría realizar un evento similar para promover el
consumo de calamar, sin embargo para fortalecer la cultura de consumo de calamar
a largo tiempo, la población objetivo podrían ser niños de nivel primaria y los
restauranteros a través del apoyo de la Cámara Nacional de la Industria
Restaurantera y Alimentos Condimentados (CANIRAC). Una primera fase incluiría la
impartición de talleres en las escuelas o comercios para que conocieran al calamar
gigante, su biología, hábitat, destacando principalmente sus características
nutricionales.

36
Como segunda fase y con ayuda de sus profesores (en el caso de las escuelas) y la
CANIRAC se podría realizar un concurso-degustación de platillos a base de calamar
gigante. Así abarcando dos principales sectores, las familias paceñas y los turistas,
en donde en miras posteriores ir incrementando esta celebración ante el consumo de
calamar gigante e ir involucrando a más y más población.
El calamar gigante presenta otros elementos de relevancia nutrimental como la
taurina, Klett-Traulsen (1996) lo menciona como una alternativa ante el
aprovechamiento del calamar; ya que la taurina es un aminoácido compuesto que
mejora el rendimiento mental (actúa como neurotransmisor), suprime la acumulación
de colesterol LDLR en el cuerpo, reduce la grasa neutra mejorando la absorción de
lípidos, mantiene controlada la presión arterial y previene de la diabetes (Kishida et
al., 2003; Sáez y Gómez, 2013); sin embargo dicha característica es poco conocida y
hasta la fecha no se ha tomado en cuenta en el aprovechamiento del calamar
gigante. Si bien la concentración de taurina no fue abordada en el presente estudio
es importante resaltar su existencia para considerarlo en estudios futuros.

37
5. CONCLUSIONES
Al conjugar los cuatro elementos del AQP (humedad, cenizas, proteínas y lípidos) se
obtiene la información nutricional principal de un alimento, en el caso del calamar
gigante es un producto el cual puede ser consumido de manera directa o procesado.
El análisis de humedad estimó en 78.92 % la carne de calamar gigante, además de
manera nutricional cuenta con concentraciones altas de minerales (5.01 %), y
proteínas (78.48 %), así como muy bajas de lípidos (1.75 %) siendo un producto
balanceado e ideal para el consumo humano.
Los calamares gigantes D. gigas machos presentaron valores más altos en el AQP
que las hembras, pero solo el análisis de humedad presentó diferencias estadísticas
(altamente significativa p<0.001). Porque los machos maduran a edades y tallas
menores que las hembras y redirigen su energía.
Los organismos maduros contienen mayor concentración de humedad (único con
diferencia significativa), mientras que en los organismos en proceso de maduración
presentaron los porcentajes menores de percentiles de proteínas y lípidos. Porque
los machos maduros requieren tales cantidades para la producción de
espermatóforos.
Los valores del AQP (proteínas, lípidos y cenizas) en el calamar gigante D. gigas
fueron elevados en los meses de julio y agosto y disminuyeron en los meses de
septiembre y octubre. En los meses cálidos existe una migración horizontal hacia
Santa Rosalía por las surgencias, con lo cual existe un aporte mayor de alimento.

38
6. RECOMENDACIONES
Para comprender con totalidad de la estacionalidad del calamar gigante, la
recolección de muestras mensuales debería ser la correspondiente a los
12 meses.
Se le podría dar continuidad a este trabajo con los análisis de otros elementos
como fibra y medición de calorimetría.
En estudios posteriores y para conocer aun más de los nutrientes del calamar
gigante Dosidicus gigas sería importante analizar la concentración y calidad
taurina que este contiene. Porque la taurina suprime la acumulación del colesterol
LDLR.
Después de un trabajo de investigación es importante darle continuidad a la
divulgación por lo que es importante como proyecto futuro retomar el tema de
talleres y concurso-degustación de platillos, con miras a promover de manera
activa el consumo de calamar gigante.

39
7. LITERATURA CONSULTADA
Abugoch, L.; A. Guarda; L. Pérez y M. Paredes. 1999 Determinación de la
composición químico-proximal y la formulación de un producto tipo gel jibia
(Dosidicus gigas). Arch. latin. nutr. 45(2): 156-161
Abugoch, L.; A. Guarda; L. Pérez y M. Donghi. 2000. Caracterización funcional y
bioquímica de la carne del manto de jibia (Dosidicus gigas). Arch. latin. nutr.
50 (4): 380-386.
Álvarez-Borrego, S. 1983. Gulf of California. En: B. H. Ketchum. Ed. Estuaries and
enclosed seas. Elsevier Amsterdam. 427-449
Álvarez-Borrego, S. 2008. Oceanografía de la región de las grandes islas. 2: 45-65.
En: G. Danemann y E. Ezcurra (Eds). Bahía de los Ángeles: Recursos
Naturales y Comunidad. Línea base 2007. SEMARNAT, Instituto Nacional de
Ecología, Pronatura noroeste, A.C. y San Diego Natural History Museum,
México, D.F.
Álvarez-Borrego, S. y J. R. Lara-Lara. 1991. The physical environment and
productivity of the Gulf of California. 47: 555-567 En: Dauphin and Simoneit
(Eds). The Gulf and Peninsula, Province of the Californias. America Asoc. of
Petroleum Geologist.
Agriculture Handbook No 8-15. 1987. Composition of Foods Finfish and Shellfish
Products. USDA. EUA. 192 pp.
Arancibia, H. y S. Neira. 2006. Assessing the potential role of predation by jumbo
squid (Dosidicus gigas) and fishing on small pelagics (common sardine
Strangomera bentincki and anchovy Engraulis ringens) and common hake
(Merluccius gayi) in central Chile, 33-39°S. En Olson RJ & JW Young (Eds).
The role of squid in open ocean ecosystems: 68-70. Report of a GLOBEC-
CLIOTOP/PFRP workshop, 16 - 17 November 2006, Honolulu, Hawaii, EUA.
GLOBEC Report 24.
Badui-Dergal, S. 1993. Química de los alimentos. 3ra. ed. Pearson Educación.
México. 648 pp.

40
Barnes, R. S.; P. Calow; P. J. Olive; D. W. Golding y J. I. Spicer. 2001. The
Invertebrates a Synthesis. Blackwell Science. 3ra. ed. EUA. 497 pp.
Bazzino, G. A. 2008. Uso, manejo y preservación de los recursos naturales
(Orientación Biología Marina). Tesis Doctoral en Ciencias. Centro de
Investigaciones Biológicas del Noroeste, S. C. México. 127 pp.
Bazzino, G. A.; C. Salinas-Zavala y U. Markaida. 2007. Variabilidad en la estructura
poblacional del calamar gigante (Dosidicus gigas) en Santa Rosalía, región
central del Golfo de California. Cienc. Mar. 33(2): 173-186.
Bell, J. G; J. McEvoy; D. R. Tocher; F. McGhee; P. J. Campbell y J. R. Sargent. 2001.
Replacement of Fish Oil with Rapeseed Oil in Diets of Atlantic Salmon (Salmo
salar) Affects Tissue Lipid Compositions and Hepatocyte Fatty Acid
Metabolism. J. Nutr., 131: 1535-1543.
Berk, Z. 1980. Introducción a la Química de los Alimentos de J. B. S. Braverman.
2da. ed. Editorial El Manual Moderno, S. A. de C. V. México. 358 pp.
Bjerkeng, B.; S. Refstie; K. T. Fjalestad; T. Storebakken; M. Rødbotten y. J. A. Roem.
1997. Quality parameters of the flesh of Atlantic salmon (Salmo salar) as
affected by dietary fat content and full-fat soybean meal as a partial substitute
for fish meal in the diet. Aquaculture. 157: 297-30.
Blaskovic, V.; A. Alegre y R. Tafur. 2008. Depredación del calamar gigante Dosidicus
gigas sobre los principales recursos pesqueros en el litoral peruano durante el
2007. Informe Técnico. IMARPE. 32 pp.
Boyle, P. R. 1983. Cephalopod Life Cycles. Vol I. Academic Press. EUA. 475 pp.
Boyle, P. R. y Rodhouse, 2005. Cephalopods: Ecology and fisheries. Blackwell
Publishing. EUA. 458 pp.
Brito-Castillo, L.; E. Alcántara-Razo; R. Morales-Azpeitia y C. A. Salinas-Zavala.
2000. Temperaturas del Golfo de California durante mayo y junio de 1996 y su
relación con las capturas de calamar gigante (Dosidicus gigas D’Orbigny,
1835). Cienc. Mar. 26(3): 413-440.

41
Brusca R. C. y G. J. Brusca. 2003. Invertebrates. Sinauer Associates. 2da. ed. EUA.
936 pp.
Cabello, A. M.; R. Villarroel-Lezama; B. E. Figueroa-García; M. C. Ramos-Marcano;
Y. Márquez-Figueroa y O. M. Vallenilla González. 2004. Parámetros de
Frescura de Moluscos. Rev. Cient. FCV-LUZ. 14(5): 457-466.
Careche, M.; A. J. Borderías e I. Sánchez-Alonso. 2006. Mejora del procedimiento de
elaboración de un concentrado proteico funcional a partir de músculo de
cefalópodos y producto así obtenido para ser utilizado en la elaboración de
productos análogos y otros nuevos productos. Consejo Superior de
Investigaciones Científicas (CSIC). España. Patente ES 2 231 023.
Castro-González, M. I.; J. L. Silencio-Barrita; M. E. Juárez-Silva; S. Montaño-
Benavides y F. Pérez-Gil. 1998. Composición química de la fauna de
acompañamiento del camarón de Veracruz (Golfo de México). Rev. Biol. Trop.
46(2): 249-256.
Christian, J. H. B. 2000. Drying and reduction of water activity. En: The
microbiological safety and quality of food. Vol. II Lund B. M. Ed. Aspen
Publishers. EUA. 146-156.
CONAPESCA 2011. Anuario Estadístico de Acuacultura y Pesca 2010. Comisión
Nacional de Acuacultura y Pesca. México. 310 pp.
De la Cruz-González, F. J. 2007. Evaluación económica de la captura de calamar
gigante (Dosidicus gigas D’Orbigny 1835) en Guaymas, Sonora. Tesis
Doctoral en Ciencias. Centro de Investigaciones Biológicas del Noroeste, S. C.
78 pp.
Desrosier, N. W. 1999. Elementos de tecnología de alimentos.14va. ed. Compañía
editorial continental, S. A. de C. V. México. 783 pp.
Dublán-García, O. 2006. Evaluación de los cambios estructurales y fisicoquímicos del
manto de calamar gigante (Dosidicus gigas) durante el almacenamiento en
refrigeración o congelación. Tesis Doctoral en Biotecnología. Universidad
Autónoma Metropolitana. México. 169 pp.

42
Ehrhardt, N; A. Solís; P. Jacquemin; j. Ortiz; P. Ulloa; G. González y F. García. 1986.
Análisis de la biología y condiciones del stock del calamar gigante Dosidicus
gigas en el golfo de California, México, durante 1980. Cienc. Pes. 5: 63-76.
Fernández, A; P. Sosa, D. Setton, V. Desantadina, M. Fabeiro, M. I. Martínez, N.
Piazza, P. Casavalle, M. Tonielli, V. Vacarezza, S. de Grandis, N. Granados y
J. Hernández. 2011. Calcio y nutrición. Sociedad Argentina de Pediatría.
Argentina. 19 pp.
Fisher, P. y A. E. Bender. 1976. Valor nutritivo de los alimentos. Editorial Limusa,
S.A. de C.V. Grupo Noriega (Eds). México. 205 pp.
Food and Nutrition Board/Institute of Medicine. 2002. Dietary Reference Intakes (DRI)
and Recommended Dietary Allowances (RDA) for energy, carbohydrate, fiber,
fats, fatty acids, cholesterol, proteins and amino acids. Institute of Medicine of
the National Academies. Washington DC. The National Academy Press En:
Hernández-Triana, M. 2004. Recomendaciones nutricionales para el ser
humano: actualización. Invest. Biomed. 23(4): 266-292.
Fox, B. A. y A. G. Cameron. 2000. Ciencia de los alimentos, nutrición y salud.
Editorial Limusa, S.A. de C.V. Grupo Noriega (Eds). México. 457 pp.
Frazier, W.C. y D. C. Westhoff. 1978. Food and Microorganisms. En: Food
Microbiology. 3ra ed. Editorial McGraw-Hill Inc. EUA.
Gilly, W. F., U. Markaida, C. H. Baxter, B. A. Block, A. Boustan, L. Zeidberg, K.
Reisenbichler, B. Robison, G. Bazzino y C. Salinas. 2006. Vertical and
horizontal migrations by the jumbo squid Dosidicus gigas revealed by
electronic taggin. Mar. Ecol. Prog. Ser. 324: 1-17
Gómez-Guillén, M. C.; T. Solas; J. Borderías y P. Montero. 1996a. Ultrastructtural
and rheological changes during the gelation of gigant squid (Dosidicus gigas)
muscle. Eur. Food Res. Tech. 202 (3): 215-220.
Gómez-Guillén, M. C.; T. Solas; J. Borderías y P. Montero. 1996b. Effect heating
temperature and sodium chloride concentration on ultrastructure and texture of

43
gels made from gigant squid (Dosidicus gigas) with addition of starch,
I- carrageenan and egg white. Eur. Food Res. Tech. 202 (3): 221-227.
Gómez-Guillén, M. C.; A. J. Borderías y P. Montero. 1997. Salt, nonmuscle proteins,
and hydrocolloids affecting rigidity changes during gelation of gigant squid
(Dosidicus gigas). J. Agric. Food Chem. 45: 616-621.
Gómez-Guillén, M. C. y P. Montero. 1997. Improvement of gigant squid (Dosidicus
gigas) muscle gelation by using gelling ingredients. Z. Leb. Unt. Fors. A. 204:
379-384.
Guerra, A. 2006. Estrategias evolutivas de los cefalópodos asociadas a la reducción
o pérdida de la concha, los cefalópodos actuales muestran una combinación
de características que explican su éxito evolutivo. Investigación y ciencia. 10
pp.
Hernández-Triana, M. 2004. Recomendaciones nutricionales para el ser humano:
actualización. Invest. Biomed. 23(4): 266-292.
Kirk, R. S., R. Sawyer y H. Egan. 2000. Composición y Análisis de Alimentos de
Pearson. 3ra. reimpresión. Compañía Editorial Continental, S. A. de C. V.
México. 777 pp.
Kishida, T., S. Miyazato, H. Ogawa y K. Ebihara. 2003. Taurine prevents
hypercholesterolemia in ovariectomized rats fed corn oil but not those fed
coconut oil. J. of nutr. 133(8): 2616-2621.
Klett-Traulsen, A. 1981. Estado actual de la pesquería del calamar gigante en el
estado de Baja California Sur. Departamento de pesca, INP. México. 33 pp.
Klett-Traulsen, A. 1996. Pesqueria de calamar gigante Dosidicus gigas. Pp 127-149
En Casas-Valdez, M. y G. Ponce-Diaz (Eds). Estudio del potencial pesquero y
acuícola de Baja California Sur. Vol. I, 350 pp.
Layzer R. B. 1991. How muscle use fuel. N. Engl. J. Med. 324: 411-6.
Lenhinger, A. L.; D. L. Nelson y M. M. Cox. 2001. Principios de Bioquímica. Omega
3ra. Edición. México. 1117 pp.

44
Lluch-Cota, S.; E. Aragón-Noriega; F. Arreguín-Sánchez; D. Aurioles-Gamboa, J,
Bautista-Romero; R. Brusca, R. Cervantes-Duarte; R. Cortés-Altamirano, P.
Del-Monte-Luna, A. Esquivel-Herrera, G. Fernández, M. Hendrickx; S.
Hernández-Vázquez; H. Herrera-Cervantes; M. Kahru; M. Lavín; D. Lluch-
Belda; D. Lluch-Cota; J. López-Martínez; S. Marinone, M. Nevárez-Martínez;
S. Ortega-García, E. Palacios-Castro, A.Páres-Sierra; G. Ponce-Díaz; M.
Ramírez-Rodríguez; C. Salinas-Zavala; R. Schwartzlose, A. Sierra-Beltrán.
2007. The Gulf of California: Review of ecosystem stratus and sustainability
changelles. Prog in Ocean. 1-26.
López, M.; Candela, J. y M. L. Argote. 2006. Why does the Ballenas Channel have
the coldest SST in the Gulf of California?. Geophys. Res. Lett. 33.
Lozano, M.; D. Arias. 2008. Residuos de fármacos en alimentos de origen animal:
panorama actual en Colombia. Rev. Colomb. Cienc. Pec. 21: 121-135.
Luna-Raya, M. C. 2008. Análisis de mercado del calamar gigante (Dosidicus gigas
D’Orbigny 1835) en México. Tesis Doctoral en Ciencias. Centro de
Investigaciones Biológicas del Noroeste, S. C. 68 pp.
Luna-Raya, M. C.; J. I. Urciaga-García; C. A. Salinas-Zavala; M. A. Cisneros-Mata y
L. F. Beltrán-Morales. 2006. Diagnóstico del consumo de calamar gigante en
México y en Sonora. Informe Técnico. México. 30 pp.
Macarulla, J. M. y F. M. Goñi. 1993. Biomoléculas: Lecciones de bioquímica
estructural. Editorial Reverté. México. 191 pp.
Markaida U. 2001. Biología del calamar gigante Dosidicus gigas Orbigny, 1835
(CEPHALOPODA: OMMASTREPHIDAE) en el Golfo de California, México.
Tesis doctoral en Ciencias. Centro de Investigación Científica y de Educación
Superior de Ensenada. 426 pp.
Markaida, U. y O. Sosa-Nishizaki. 2001. Reproductive biology of jumbo squid
Dosidicus gigas in the Gulf of California, 1995-1997. Fish. Res. 54(1): 63-82.

45
Markaida U., J. J. C. Rosenthal y W. F. Gilly. 2005. Tagging studies on the jumbo
squid, Dosidicus gigas, in the Gulf of California, México. Fish. Bull. 103(1):
219-226.
Markaida U., W. F. Gilly, C. A. Salinas-Zavala, R. Rosas-Luis y J. A. T. Booth. 2008.
Food and feeding of jumbo squid Dosidicus gigas in the Central Gulf of
California during 2005-2007. CalCOFI Rep. 49: 90-103.
Martínez-Vega, J. A.; L. E. Cruz-Suárez y D. Ricque-Marie. 2000a. Evaluación de las
diferentes partes corporales del calamar gigante (Dosidicus gigas) en forma de
harina, en dietas balanceadas para camarón (Litopenaeus vannamei). Cienc. y
Mar. 11-18.
Martínez-Vega, J. A.; L. E. Cruz-Suárez y D. Ricque-Marie. 2000b. Composición
corporal y el proceso de secado del calamar gigante Dosidicus gigas. Cienc. y
Mar. 35-38.
Mejía-Rebollo, A.; C. A. Salinas-Zavala; C. Quiñónez-Velázquez y U. Markaida. 2008.
Age, growth and matury of jumbo squid (Dosidicus gigas D’Orbigny, 1835) off
the western coast of the Baja California Peninsula. CalCOFI Rep. 49: 256-262.
Mille-Pagaza, S. R. 2008. Invertebrados. Instituto Politécnico Nacional. 691 pp.
Morales-Bojórquez, E. y M. O. Nevárez-Martínez. 2002. Estimación estocástica de la
captura y el reclutamiento del calamar gigante Dosidicus gigas (D’Orbigny,
1835) del Golfo de California, México. Cienc. Mar. 28(2): 193-204.
Morton, J. E., 1979. Molluscs, 5ª ed. Inglaterra, Hutchinson of London, 264 pp.
Nesis, K. N. 1983. Dosidicus gigas. En: Boyle P. R. (Ed.). Cephalopod Life Cycles.
Vol. I, Species Accounts. Academic Press. Londres. 475 pp.
Nevárez-Martínez, M. O.; A. Hernández-Herrera; E. Morales-Bojórquez; A. Balmori-
Ramírez; M. A. Cisneros-Mata y R. Morales-Azpeitia. 2000. Biomass and
distribution of the jumbo squid (Dosidicus gigas) in the Gulf of California,
México. Fish. Res. 49: 129-140.

46
Nevárez-Martínez, M. O. y E. Morales-Bojórquez. 2002. Estimación estocástica de la
capturabilidad y el reclutamiento del calamar gigante Dosidicus gigas
(D’Orbigny, 1835) del Golfo de California, México. Cienc. Mar. 28(2): 193-204.
Nigmatullin, Ch. M., A. V. Parfenyuk y V. N. Nicolski. 1991. Ecology and nektonic
epipelagic squid resources of the Atlantic and the southeastern Pacific oceans.
En Markaida U. 2001. Biología del calamar gigante Dosidicus gigas Orbigny,
1835 (CEPHALOPODA: OMMASTREPHIDAE) en el Golfo de California,
México. Tesis doctoral en Ciencias. Centro de Investigación Científica y de
Educación Superior de Ensenada. 426 pp.
Nigmatullin C.; K. N. Nesis y A. I. Arkhipkin. 2001. A review of the biology o jumbo
squid Dosidicus gigas (Cephalopoda: Ommastre phidae). Fish Res. 54: 9-19.
Paredi, M.E.; Tomas, M. C.; Crupkin, M. y M. C. Añón. 1996. Thermal denaturation of
muscle proteins from male and female squid (Illex argentinus) at different
sexual maturation stages. A differential scanning calorimetric study. J. Agric.
Food Chem. 44: 3812-3816.
Pérez-Ruiz, M. y A. Lucía-Mulas. 2007. Trastornos del metabolismo energético del
músculo: bases genéticas y bioquímicas de las miopatías que cursan con
intolerancia al ejercicio físico. Arch. Med. Dep. Vol. XXIV (122): 451-463.
Ramírez-Olivas, R.; O. Rouzaud-Sández; N. F. Haard; R. Pacheco-Aguilar y J. M.
Ezquerra-Brauer. 2004. Changes in firmness and thermal behavior of ice-
stored muscle of jumbo squid (Dosidicus gigas). Eur. Food Res. Tech. 219(4):
312-315.
Roper, C. F. E. y R. E Young. 1975. Vertical distribution of pelagic cephalopods.
Smith. Contr. Zool. 209: 1-51.
Rosa, R.; J. Pereira y M. L. Nunes. 2005. Biochemical composition of cephalopods
with different life strategies, with special reference to a gigant squid,
Architeuthis sp. Mar. Biol. 146: 739-751.
Ruppert E. E. y R. D., Barnes 1996. Zoología De Los Invertebrados. McGraw-Hill
Interamericana. 6ta edición. México. 1114 pp.

47
Sáez B., M. J. y M. Á. Gómez N. 2013. La protección del consumidor europeo en el
ámbito de los alimentos funcionales. Tesis doctoral. Universidad de Valladolid.
España. 457 pp.
Salinas-Zavala, C. A.; S. Sánchez-Hernández; E. A. Aragón-Noriega; C. Sánchez-
Verdugo; G. Soria-Martínez; G. Escoto-González; T. Moctezuma-Cano; S.
Camarillo-Coop; A. Mejía-Rebollo y G. Bazzino. 2005. Programa maestro de la
pesquería de calamar gigante (Dosidicus gigas). México. 81 pp.
Samuelson, P. A.; W. D. Nordhaus; J. Salazar-Cantú; R. C. Rodríguez-Guajardo.
2004. Microeconomía con aplicaciones a Latinoamérica. 17 ed. Mc Graw Hill.
México. 478 pp.
Sato, T. 1976. Results of exploratory fishing for Dosidicus gigas (D’ Orbigny) of
California and Mexico. FAO Fish. Rep. 170(1): 61-67
Schmidt-Hebbel, H.; Pennacchiotti, I.; Masson, L. y M. A. Mella. 1990. Tabla de
Composición química de alimentos chilenos. Facultad de Ciencias Químicas y
Farmacéuticas. Departamento de Ciencia de los Alimentos y Tecnología
Química. Santiago de Chile.
Stryer, L. 2001. Bioquímica. Editorial Reverté. 2da. Reimpresión. España. 459 pp.
Ulloa, P., M. Fuente y V. Ruiz. 2006. Hábitos alimentarios de Dosidicus gigas
(D’ Orbigny, 1835) (Cephalopoda: Teuthoidea) frente a la costa centro-sur de
Chile. Rev. Chil. Hist. Nat. 79:475 – 479.
Wing, B. L. 2006. Unusual invertebrates and fish observed in the Gulf of Alaska,
2004-2005. PICES Press 14(2): 26-28.
Zeidberg, L. D. y B. H. Robison. 2007. Invasive range expansion by the Humboldt
squid Dosidicus gigas, in the eastern North Pacific. PNAS 104: 12948-12950.
Hemerografía
Heraldo de Aragon. 2006. Industria alimentaria > ¿Es un calamar, es un cangrejo, es
abadejo? No, es… ¡surimi! (16 de mayo).

48
Recursos electrónicos
Acevedo-Sánchez, A. M. y D. C. López Nieto. 2011. Verificación de los métodos para
el análisis proximal en leche entera en el laboratorio de análisis de agua y
alimentos de la Universidad Tecnológica de Pereira.
http://repositorio.utp.edu.co/dspace/bitstream/11059/2834/1/63714H565.pdf.
16 de mayo de 2014, 14:46 h.
Bavera, G.; O. Bocco, H. Beguet y A. Petryna. 2002. Promotores del crecimiento y
modificadores del metabolismo. Cursos de Producción Bovina de Carne,
F.A.V. UNRC. www.produccion-animal.com.ar. 16 de mayo de 2014, 12:12 h.
CONAPESCA, 2013. Base de datos de producción Anuario 2012.
http://www.conapesca.sagarpa.gob.mx/wb/cona/anuario_2012_zip.
Consultado: 11 de febrero de 2014, 14:36 h.
CONASAMI, 2014. Comisión Nacional para el Salario Mínimo. Nuevos salarios
mínimos 2014, por área geográfica generales y profesionales.
http://www.conasami.gob.mx/nvos_sal_2014.html. Consultado 24 de febrero
de 2014, 08:57 h.
Doyle, M. E. 2006. Veterinary Drug Residues in Processed Meats – Potential Health
Risk. Fod Research Institute (FRI Briefings.
http://fri.wisc.edu/docs/pdf/FRIBrief_VetDrgRes.pdf. Consultado 16 de mayo
de 2014, 11:57 h.
FAO. 1993. Food and Agriculture Organization of the United Nationes.
Manual de técnicas para laboratorio de nutrición de peces y crustáceos.
http://www.fao.org/docrep/field/003/AB489S/ AB489S00.htm. Consultado: 11
de febrero de 2014, 20:33 h.
FAO. 1984. FAO Species catalogue VOL. 3. Cephalopods of the world an annotated
and illustrated catalogue of species of interest to fisheries. Clyde F.E. Roper
Michael J. Sweeney Cornelia E. Nauen 1984. FAO Fisheries Synopsis 125(3)
http://www.fao.org/fishery/species/2721
http://www.fao.org/fi/eims_search/advanced_s_reult.asp?JOB_NO=ac479
22 de abril de 2009, 15:47 h.

49
FDA, 2014. Food and Drug Administration
Tabla de almacenamiento en refrigerador y congelador.
http://www.fda.gov/downloads/food/resourcesforyou/HealthEducators/UCM148
133.pdf. Consultado 12 de marzo de 2014, 08:17 h.
Gilly, W. F. 2005. Spreading and stranding of jumbo squid. Ecosystems Observations
for the Monterrey Bay National Marine Sanctuary 2005: 20-22
http://www.montereybay.noaa.gov/reports/2005/eco/harvestedsp.html.
INEGI 2005. Perspectiva Estadística. Serie por Entidad Federativa. México.
http://cuentame.inegi.org.mx/monografias/informacion/BCS/Economia/default.a
spx?tema=ME&e=03#sp. Consultado 2 de abril de 2009, 13:39 h.
ITIS 2014. ITIS Report. Dosidicus gigas (D’Orbigny, 1835 in 1834-1847).
http://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_val
ue=82538&print_version=PRT&source=to_print. Consultado el 10 de febrero
de 2014, 21:10 h.
Soriana 2014. Carnes, pescados y mariscos. Sección: pescados y mariscos
http://www1.soriana.com/site/default.aspx?p=11329&nuor=0. Consultado el 25
de febrero de 2014, 8:38 h.
Martínez-Montes, J. M. 2009. Nutrición Animal I. Integración del Análisis Proximal.
http://www.oocities.org/mvz_jmtz/anprox.html. Consultado el 03 de febrero de
2014, 10:24 h.
Salinas-Zavala, C. A.; S. Sánchez-Hernández y S. Camarillo-Coop. 2004. Cadena
productiva de la pesquería de calamar gigante. Segunda etapa:
Aseguramiento de la materia prima y determinación de preferencias de
consumo en México y en el Mundo. 176 pp.
http://www.sagarpa.gob.mx/conapesca/organizacion/cadenas%20productivas/I
nforme-Final-Segunda-Etapa-Completo.pdf. Consultado el 29 de septiembre
de 2011, 15:07 h.

50
ANEXO A. Marco Conceptual

51
CARACTERÍSTICAS GENERALES DEL CALAMAR GIGANTE
El calamar gigante Dosidicus gigas se encuentra clasificados dentro de la clase
Cephalopoda (ITIS, 2014), grupo caracterizado, como su nombre lo indica: “tienen los pies
en la cabeza”. Estos organismos presentan un pie muscularizado que rodea a la cavidad
oral, dividido en una serie apéndices (ocho en pulpos, diez en calamares y más de 10 en
nautiloideos), habitan en todos los océanos de la Tierra y presentan una variedad
morfológica bastante amplia. A los cefalópodos como los calamares, se les ha considerado
como los nadadores campeones ya que han abandonado la vida sedentaria y se han
convertido en los predadores efectivos por su rápido desplazamiento hasta zonas
epipelágicas (Ruppert y Barnes, 1996; Abugoch et al., 1999; Markaida, 2001).
Dada la diversidad de ambientes en las cuales se localizan, se pueden encontrar desde
fuentes hidrotermales profundas hasta zonas someras y desde latitudes polares hasta el
Ecuador. Algunos de estos seres vivos tienen comportamientos complejos y son más
semejantes a los peces que a los demás moluscos (Ruppert y Barnes, 1996; Brusca y
Brusca, 2003; Arancibia y Neira, 2006; Guerra, 2006). Alcanzan grandes velocidades,
superiores a las de cualquier invertebrado e incluso superar a varias especies de teleósteos,
esto se debe a que presentan un sifón tubular ventral, altamente movible y manipulado a
cualquier dirección, el cual bombea agua hacia el exterior de la cavidad paleal para aumentar
su fuerza en la natación (Ruppert y Barnes, 1996; Boyle y Rodhouse, 2005).
En general los cefalópodos son predadores carnívoros, cazan en la columna de agua y en el
sustrato, en particular los calamares se denominan como las criaturas más voraces del
océano. Se extienden rápidamente con sus dos tentáculos para iniciar el ataque contra la
presa. Ulloa et al. (2006) y Markaida et al. (2008), afirman que en su dieta principal
predominan los teleósteos, seguida de moluscos, para finalizar con crustáceos.
Los calamares se consideran moluscos sin concha o aparentemente reducida ya que se ha
modificado a una manera interna como gladio (pluma) hecha de quitina (Boyle, 1983; Barnes
et al., 2001; Boyle y Rodhouse, 2005). El calamar gigante (Dosidicus gigas, D’Orbigny 1835)
es el representante de mayor tamaño de la familia Ommastrephidae (Steenstrup, 1857). Se
caracteriza por un manto cilíndrico, muy grande y robusto de paredes gruesas, dos aletas
romboides terminales y en la cabeza ocho brazos y dos tentáculos, sus brazos presentan de
60 a 200 pares de ventosas, su talla aproximada es de 50 a 90 cm y un peso de 10 a 30 Kg
(FAO, 1995; Abugoch et al., 1999).
CARACTERÍSTICAS DIAGNÓSTICAS
El calamar gigante Dosidicus gigas presenta dos aletas romboidales, musculosas y anchas,
del 49 al 65 % de la longitud del manto (LM), y su longitud del 41 al 49 % de la LM; ángulo de
la aleta es entre 50 y 65º. La Foveola con tiene de 5 a 9 pliegues junto con 2 a 8 bolsas
laterales; dáctilo de la maza tentacular con 4 hileras de ventosas; entre 60 y más de 200
pares de ventosas en el brazo I; membrana protectiva del brazo III igual al ancho del brazo,
membranas de los otros brazos nunca más altas que las ventosas; brazo IV derecho o
izquierdo hectocotilizado, sin ventosas y pedúnculos en la punta y membranas protectoras

52
perforadas y engrosadas; pequeños fotóforos subcutáneos en la superficie ventral del manto,
cabeza y brazos III y IV. La característica más conspicua es tal vez la prolongación de los
extremos distales de los brazos en filamentos largos y delgados, conteniendo entre 100 y
200 pares de ventosas diminutas muy agrupadas, 3 veces más numerosas que en los demás
Ommastrephidae (Fig. 10). Investigadores añaden la presencia de dos fotóforos intestinales
en su diagnóstico, sin embargo los colaboradores rusos opinan que éste carácter es
transitorio y no perdura en los adultos (Voss, et al., 1998; Markaida, 2001).
FIGURA 10. Calamar gigante Dosidicus gigas. A: vista dorsal de un ejemplar grande (50-80 cm LM) B: vista ventral de un ejemplar de 25.4 cm LM; C: IV brazo izquierdo hectocotilizado y pedúnculo tentacular (Tomado de Markaida, 2001).
SINONIMIAS
Dosidicus eschrichtii Steenstrup, 1857
Dosidicus steenstrupii Pfeffer, 1884
Ommastrephes giganteus
D'Orbigny, 1839-1842 en Férussac y
D'Orbigny, 1834-1848
Ommastrephes gigas D'Orbigny, 1835 en
1834-1847
Sepia nigra Bosc, 1802
Sepia tunicata Molina, 1782
NOMBRES COMUNES
Español: Calamar gigante (México, Perú),
jibia (Chile, Perú), pota (Perú), calamar
rojo (Chile).
Inglés: Jumbo squid, jumbo flying squid,
Humboldt Current squid, Peru squid,
giant squid.
Ruso: Kal'mar-dosidicus, gigantskij
peruano-chilijskij kal'mar.
Japonés: Amerika-ooakaika, Amerika-
oosurume.
CLASIFICACIÓN TAXONÓMICA
Reino Animalia Phylum Mollusca Clase Cephalopoda Cuvier, 1797 Subclase Coleoidea Bather, 1888 Superorden Decabrachia Boettger, 1952 Orden Teuthida Naef, 1916 Suborden Oegopsina D'Orbigny, 1845 Familia Ommastrephidae Steenstrup, 1857 Subfamilia Ommastrephinae Posselt, 1891 Género Dosidicus Steenstrup, 1857 Especie Dosidicus gigas (D'Orbigny, 1835 en 1834-1847)
C

53
DISTRIBUCIÓN DEL CALAMAR GIGANTE
Dosidicus gigas se distribuye en el Pacífico Oriental (Fig. 11), se mantiene como una
especie subtropical, que visita aguas tropicales, se ubica desde Monterey, California (EUA)
hasta el sur de Chile (Sudamérica), incluyendo el Golfo de California y particularmente
abundante en aguas de California, así como en el sistema de corrientes de Perú y Costa
Rica (Nesis, 1983; Markaida, 2001; Nigmatullin et al, 2001). Sin embargo en 2004 el Alaska
Department of Fish and Game recibió
reportes de capturas de calamar gigante
en el Golfo de Alaska, confirmándose
nuevamente la noticia en agosto de 2005.
Aunado a varamientos masivos en los
límites de la distribución, ampliándose la
misma, dichas expansiones poblacionales
han ocurrido durante periodos cálidos y de
manera simultánea, se observó un decline
de las poblaciones de atún y marlin en el
Pacífico, donde los grandes peces han
sido reemplazados por individuos
pequeños, por lo cual se plantea la
hipótesis de que en consecuencia a ello el
calamar tuvo que buscar otros peces que
satisficieran sus demandas alimenticias
(Salinas-Zavala et al., 2005; Wing, 2006;
Zeidberg y Robison, 2007).
FIGURA 11. Distribución geográfica del calamar gigante Dosidicus gigas en el Pacífico Oriental. Se observa la expansión poblacional identificada en las últimas décadas (Roper y Young, 1975 y Barnes, 1996; Nigmatullin et al., 2001 y Gilly et al., 2005). Tomada de Bazzino, 2008.
HÁBITAT DEL CALAMAR GIGANTE
El calamar gigante presenta una distribución semi-oceánica meridional tropical y subtropical,
coincide con las zonas de alta productividad, cuyas concentraciones de fosfatos son 0.8 mg
PO4 3/m2 entre la capa de 0-100 m de largo, así mismo desde zonas oceánicas y neríticas
en la superficie hasta profundidades mayores a 1500 m y son más abundantes en
Sudamérica, los adultos se han capturado desde aguas superficiales hasta profundidades de
60 a 80 m con temperaturas máximas de 15º C a 28º C hasta mínimas de 12.8º C a 16.5º C
(FAO, 1984; Brito-Castillo et al. 2000; Markaida, 2001; Nevárez-Martínez et al., 2000;
Nigmatullin et al., 2001).
D. gigas realiza migraciones verticales diarias donde viaja en el día a grandes profundidades
entre 200 y 350 m y por la noche en aguas someras o cercanas a la superficie, entre 50 y

54
150 m, ello de acuerdo al comportamiento acorde de los calamares omastréfidos (Gilly et al.,
2006; Bazzino, 2008).
La profundidad a la que se desplazan durante el día se asocia con el límite superior de la
Zona de Mínimo Oxígeno (ZMO), que en el Golfo de California se extiende de los 250 a los
800 m, presentando una zona hipóxica estable. D. gigas es un organismo singular, ya que
representa un desafío fisiológico porque es un predador activo con una tasa metabólica alta,
que de manera diurna se encuentra a profundidades >200 m (ZMO) donde se presentan <20
µM ≈ 0.5 ml/l oxígeno disuelto (Álvarez-Borrego y Lara-Lara, 1991; Bazzino, 2008).
Recientemente se han identificado ciertos patrones de migración horizontal dentro del Golfo
de California, donde en el mes de noviembre se desplaza de la región de Santa Rosalía a
Guaymas y de manera inversa de Guaymas a Santa Rosalía durante mayo, con una
velocidad promedio de 8 km/día (Markaida et al., 2005; Bazzino, 2008).
IMPORTANCIA ECONÓMICA Y PESQUERA DEL CALAMAR GIGANTE
El calamar gigante es el objetivo de la pesca de cefalópodos más grande del mundo, el cual
ha tuvo en 2004 800,000 t en desembarques (Gilly et al., 2006), esta pesquería es de gran
importancia en el mundo, sobre todo en países como México, Perú y Chile debido a los altos
niveles de capturas. Méxio y Perú han sido los primeros productores a nivel mundial de esta
espcie (Markaida, 2001; CONAPESCA, 2011; Blaskovic et al., 2008).
En el Golfo de California se encuentra un stock compuesto por varias cohortes y las
variaciones de captura están relacionadas con fluctuaciones ambientales, migraciones y por
su interacción con la pesquería de camarón según Morales-Bojórquez y Nevárez-Martínez
(2002), por lo que para estimar el reclutamiento requeriría de nuevas consideraciones en los
modelos para cohortes múltiples.
En México se ve beneficiada la región noroeste con este sector primario, ya que existe una
disponibilidad del recurso durante todo el año, en el Golfo de California se pescaron 100,000
t anuales entre 1996 y 1997 y durante el 2012 Baja California Sur aportó el 56.57 % del total
nacional con 12,546.26 t anuales y así, obteniendo el primer lugar nacional, de los ocho
productores (Salinas-Zavala et al., 2004; INEGI, 2005; CONAPESCA, 2013).
El impulso pesquero se asocia fuertemente a la demanda externa, ya que la mayoría del
recurso capturado es exportado a otros países como los asiáticos, seguidos por los de la
Unión Europea y de Norteamérica, sin embargo no todo el calamar es exportado ya que una
mínima porción permanece en nuestro territorio pero es aún menor el consumo de calamar.
Desde 1994 hasta la fecha, se está promoviendo su consumo en la alimentación humana
(Luna-Raya et al. 2006; Luna-Raya et al. 2008). Por lo que conocer las características
nutricionales de su carne es de gran relevancia, ya que estos compuestos son los que
determinan la calidad de la materia prima como alimento.

55
LOS ALIMENTOS
Los alimentos son sustancias capaces de aportar nutrientes, compuestos que cubren
requerimientos específicos en los organismos para el metabolismo, como proveer de
energía, crecimiento y restauración de secciones corporales. La constitución de los alimentos
procesados se puede dividir en dos grandes grupos, según la cantidad en que se
encuentran: agua, macronutrientes (proteínas, carbohidratos, fibra y lípidos) y
micronutrientes (minerales: calcio y fósforo, vitaminas: A, B, C, D, E y K y oligoelementos:
hierro, zinc, manganeso y selenio) (Berk, 1980; Badui-Dergal, 1993; Kirk et al, 2000).
La principal función de los macronutrientes es el aporte de energía para realizar las
actividades de cada sistema, mientras que los micronutrientes son los colaboradores para
llevar a cabo el metabolismo (Fisher y Bender, 1976; Fox y Cameron, 2000).
Para cumplir con el abastecimiento de nutrientes esenciales para el organismo se han
desarrollado los Rangos Aceptables de distribución de Macronutrientes (RAM) para dietas
saludables, donde se sugiere que la ingestión de grasas sea del 20 al 35 %, carbohidratos
del 45 al 65 % y proteínas del 10 al 35 % del total del consumo diario para el ser humano.
El Consejo de Alimentación y Nutrición de EUA (2002) propone que el consumo de dietas
con esta composición debe mantener en un valor mínimo el riesgo de enfermedad
degenerativas tales como enfermedad coronaria, obesidad y diabetes mellitus.
ANÁLISIS QUÍMICO PROXIMAL
El Análisis Químico Proximal (AQP) es un esquema de exámenes que son necesarios para
conocer la estructura de un alimento agrupado en principales nutrientes. Es utilizado
principalmente para evaluar la calidad de un alimento en función a los grupos de compuestos
con características físico-químicas semejantes, pero con diferente valor nutritivo. A pesar de
las limitaciones que tiene el análisis proximal, ha sido por más de un siglo el punto de partida
en la evaluación de un alimento y aunque los métodos de análisis hayan cambiado, el
fundamento permanece intacto (Berk, 1980; Badui-Dergal, 1993; Fox y Cameron, 2000; Kirk
et al., 2000). Dicho análisis consta de las siguientes mediciones:
%Humedad
El agua es esencial para el crecimiento de todas las células vivas y la mayoría de los
alimentos contienen suficiente agua apara favorecer el crecimiento, sin embargo, debido a
que el agua no contiene un valor energético, no se considerada como un nutrimento pero es
indispensable para la fisiología general del organismo, ya que la composición de nuestro
cuerpo es de un 70% de agua aproximadamente, bien sea localizado de manera intracelular
como intersticial, y sucede de igual manera en organismos marinos como los moluscos
(Badui-Dergal, 1993; Desrosier, 1999). Gracias a la determinación del agua en un alimento
se puede describir la composición y estimar el valor nutritivo de los distintos alimentos
(Badui-Dergal, 1993; Desrosier, 1999).

56
%Proteínas
Presentan un papel importante debido a que se encuentran prácticamente en todos los
procesos biológicos como la catálisis enzimática, transporte y almacenamiento de iones,
movimiento coordinado dentro del músculo, soporte mecánico (fuerza de tensión de la piel),
protección con anticuerpos, generación y transmisión de impulsos nerviosos y control del
crecimiento, así como la diferenciación génica. Realizan funciones en el organismo como la
regeneración y formación de secciones corporales, anticuerpos, síntesis de enzimas,
mecanismo de coagulación y hormonales. Las proteínas son el resultado de la
polimerización, mediante enlaces peptídicos de los 20 aminoácidos, (Berk, 1980; Macarulla y
Goñi, 1993; Desrosier, 1999; Stryer, 2001).
Los alimentos ricos en proteínas (como la carne de res, leche y huevos) son escasos en la
mayoría de los países en vía de desarrollo, además de ser más costosos de producir y
adquirir; sin embargo no es necesario producir más alimentos sino cuidar, procesar y
almacenar con los que ya se cuenta (Badui-Dergal, 1993).
El método de Kjeldahl es el utilizado para la obtención de las proteínas, el cual se basa la
evaluación del contenido de nitrógeno total en la muestra, posterior a ser digerida con ácido
sulfúrico en presencia de un catalizador y se registra el amoniaco obtenido (FAO, 1993;
Acevedo y López, 2011)
%Lípidos
Constituyen un grupo heterogéneo de biomoléculas, pero con características comunes de
solubilidad (poco o nada solubles en agua y solubles en disolventes orgánicos), la mayor
parte son aceites y grasas. Estas biomoléculas orgánicas que se encuentran en las células,
realizan funciones biológicas importantes: sirven como moléculas combustibles,
componentes estructurales de las membranas, formas de transporte y almacenamiento del
combustible catabólico, cubierta protectora y componentes de la superficie celular; sin
embargo hay algunas con intensa actividad biológica como ciertas vitaminas y hormonas
dentro de las cuales se localizan la progesterona, testosterona y ecdisona (Macarulla y Goñi,
1993; Lenhinger et al., 2001; Stryer, 2001).
Para determinar la cantidad de lípidos es mediante el método Soxhlet con éter de petróleo y
dando un porcentaje del peso después de que el solvente se evapora (FAO, 1993)
%Cenizas
Las cenizas son los residuos de un alimento, son compuestos inorgánicos (óxidos,
carbonatos, fosfatos, sulfatos y minerales como el sílice) que quedan después de quemar la
materia orgánica, sin embargo no siempre es la misma, ya que debido a la volatilidad hay
una pérdida de sustancias, considerándose como una medida general de calidad y a veces
es útil para identificar la autenticidad de un alimento o bien cuando se presenta un valor alto
se sugiere la presencia de un adulterante inorgánico y no idóneo para el consumo (FAO,
1993; Kirk, et al., 2000).

57
Los minerales y sus sales realizan funciones plásticas, ya que componen esqueleto,
cartílago y huesos (calcio, fósforo y magnesio), hemoglobina (fierro) y se encuentran otros
minerales en pequeñas proporciones como cobre, manganeso y cobalto. Los minerales
actúan como reguladores, mantienen la presión osmótica a través de las membranas
celulares, la relación alcalina, neutra o ácida de los sección corporals, en el sistema nervioso
ordena la excitabilidad y contractividad muscular, así como activar los procesos enzimáticos
de absorción (FAO, 1993; Hernández, 2004).
El calcio tiene como primera función (iónica) interviene en el proceso de coagulación
sanguínea, contracción muscular, transmisión del impulso nervioso y posteriormente la
osificación de los huesos, ya que el 99 % son compuestos insolubles en forma cristales de
hidroxiapatita y el resto se encuentra en los sección corporal blandos y líquido extracelular
(calcemia); es importante el consumo ya que mejora la acumulación en el sección corporal
óseo, aumentando la densidad ósea y evitando fracturas (Hernández, 2004, Fernández et
al., 2011).
De el fósforo total del cuerpo, ¾ partes se localizan en el esqueleto y dientes, mientras que
la otra ¼ parte en nucleoproteínas, son sustancias fundamentales de las membranas
celulares, además de ser un componente de enlaces de fosfato, fosfolípidos, fosfoproteínas,
ácidos nucleícos, y actúan como segundos mensajeros hormonales (Hernández, 2004;
Fernández et al., 2011).
El magnesio se moviliza junto con las proteínas de la sangre, y actúa como cofactor
enzimático del metabolismo de aminoácidos, lípidos y carbohidratos, estimula el crecimiento,
tiene una acción descalcificante, así como mantener el metabolismo del sodio, potasio y
calcio (Hernández, 2004).

58
ANEXO B. Análisis Químico Proximal,
siguiendo la metodología establecida por la A. O. A. C. (1995)

59
Determinación de %Humedad
Materiales y reactivos:
Cápsula o crisol de porcelana
Balanza analítica
precisión ±0.001
Estufa Thermolyne Serie 9000
Desecador Nalgene
Campana de extracción de humos
Silca gel
Procedimiento:
1. Pesar con exactitud de 2 a 5 g de muestra en una cápsula o crisol de porcelana,
previamente puesta a peso constante, extendiendo la muestra en una capa lo más fina
posible sobre la base de la cápsula o crisol.
NOTA: La cantidad de muestra a pesar dependerá de su humedad inicial, ya que a mayor
peso final mayor será la precisión. También es importante contar con cantidad suficiente
de muestra (2.5 g en base seca) si se piensa determinar cenizas. Es necesario utilizar
una balanza analítica con buena precisión (+ 0.001).
2. Colocar la cápsula y su contenido en una estufa a 70° C durante 24 h.
3. Retirar la cápsula de la estufa y dejar enfriar en un desecador durante 30 minutos.
4. Pesar la muestra seca en la balanza analítica.
5. Colocar la cápsula en la estufa por 1 hora aproximadamente a 100° C y desecar por 30
minutos.
6. Pesar, repetir el paso anterior hasta obtener un peso constante.
Cálculos:

60
Determinación de %Cenizas
Materiales y reactivos:
Cápsula o crisol de porcelana
Balanza analítica
Mechero Bunsen o parrilla de
calentamiento
Mufla
Desecador
Baño María
Campana de extracción de humos
Silca gel
Etanol
Glicerol
Procedimiento:
1. Pesar 5 g de muestra sólida (puede utilizarse el material residual de la determinación de
humedad) y colocarla en una cápsula de porcelana previamente puesta a peso constante
a 500° C por 24 h en la mufla.
NOTA: Debido a que en este procedimiento se quema todo el material orgánico en la
muestra, es importante tomar la máxima cantidad de muestra posible.
2. Carbonizar gradualmente la muestra sobre la llama del mechero Bunsen o la parrilla del
calentamiento, hasta obtener una masa carbonizada.
3. Incinerar en la mufla a 550-570° C (para quemar completamente el carbón) durante 24
horas.
4. Retirar la cápsula, colocarla durante 30 minutos en la estufa a 100° C y luego enfriar en
un desecador durante 30 minutos.
5. Pesar.
Cálculos:

61
Determinación de %Proteínas Totales: Método Microkjeldahl-Tecator
El principio sobre el cual se basa esta determinación es en la consideración de que la
mayoría de las proteínas tienen una cantidad aproximada del 16% de nitrógeno, y a partir de
este porcentaje se puede obtener el factor 6.25 y al multiplicar dicho factor por la cantidad de
nitrógeno obtenido nos indica el contenido de proteínas de la muestra analizada.
Cálculos:
Materiales y reactivos:
Sistema de digestión Tecator® (Mod.
DS12, serie 959)
Destilador automático Tecator®
Baño de agua refrigerado
Papel encerado para pesar
Balanza analítica
Tubos de digestión Tecator
Punta de pipeta de 50 ml Eppendorf
Vaso de precipitado 100 ml
Bomba de vacío
Campana de extracción de humos
Algodón
Metanol
Sulfato ferroso amoniacal
hexahidratado
Catalizador (Kjeltabs; (Mod. 1030,
serie 13363) K2SO4 y CuSO4)
Ácido sulfúrico concentrado
Ácido bórico
Rojo de metilo
Verde de bromocresol
Agua destilada
Hidróxido de sodio
Ácido clorhídrico 0.05 N
Procedimiento:
Digestión
1. Prender el sistema de digestión Tecator® (Mod. DS12, serie 959), el destilador
automático Tecator® y el sistema de enfriamiento del destilador.
2. Pesar de 0.05 g a 0.1 g de muestra y colocarla en los tubos de digestión Kjeldahl
secos y limpios. Pesar 0.1 g de sulfato ferroso amoniacial, cuyo contenido de
nitrógeno es conocido, para usarlo como estándar y colocarlo en otro tubo de
digestión. Colocarlo en un tubo independiente.

62
NOTA: Los tubos a utilizar deberán estar previamente enjuagados con agua destilada
y puestos a secar en la estufa, de no haber sucedido así, entonces se recomienda
hacerlo.
3. Adicionar a cada uno de los tubos 1.6 g de catalizador (Special Kjeltabs
C 3-5).
4. Añadir 3 ml de ácido sulfúrico concentrado a cada uno de los tubos.
5. Una vez que el digestor alcanzó los 400° C aproximadamente, meter los 12 tubos con
muestra a digerir colocando la tapa de succión sobre la boca de los tubos y prender
la bomba de succión.
NOTA: Mantener el flujo máximo de succión por 5 minutos, una vez transcurrido este
tiempo ajustar la intensidad de succión de manera tal que la nube de condensación
del ácido permanezca o se mantenga aproximadamente a 3/4 de la altura del tubo.
6. Una vez que la muestra se torna verde cristalina indicará que la digestión ha
terminado.
NOTA: El tiempo en que esto suceda dependerá de la cantidad de proteínas que
contenga la muestra, por lo que se recomienda estar al pendiente, ya que un exceso
en el tiempo de digestión puede ocasionar pérdida de nitrógeno.
7. Retirar los tubos del digestor y dejar enfriar las muestras con la tapa de succión aún
puesta.
8. Una vez frías las muestras, agregarles 10 ml de agua destilada y colocar la tapa de
succión sobre el baño de agua tibia o caliente y una succión máxima, esto con el fin
de lavar el sistema de succión. Una vez hecho lo anterior se apaga la bomba de
succión.
Destilación
1. Verificar que todos los recipientes periféricos del destilador automático Tecator®
(Mod. 1030, serie 13363) contengan sus respectivas soluciones (agua destilada,
solución receptora, álcali y ácido para titulación) en cantidad suficiente. En caso de
detectar algún faltante, preparar el reactivo.
2. En la posición HELP, lavar el sistema por medio de la inyección de vapor siguiendo
los siguientes pasos:
a. Colocar un tubo de digestión con aproximadamente 10 ml de agua destilada en la
posición de destilación con la puerta de seguridad cerrada y con el aparato en la
posición HELP, poner al switch de STEAM en la posición de encendido STEAM
por 5 minutos aproximadamente una vez pasado este tiempo volver el switch a la

63
posición OFF y entonces retirar el tubo para cambiar el agua. Repetir esta
operación hasta que el agua del tubo de destilación sea clara.
b. Verificar que la bureta de titulación no contenga burbujas, en caso de si
contenerlas sacarlas operando la bureta con el switch TITRANT (lo cual se
logrará desalojando aproximadamente 1/8 del volumen de la bureta y volviéndola
a llenar).
NOTA: Cuando la puerta de seguridad se encuentra arriba, la dirección de
inyección o succión de la bureta está dirigida hacia el recipiente contenedor de
HCl; cuando la puerta está cerrada entonces se dirige hacia el vaso (cristal) de
recepción.
c. Siguiendo en la posición HELP inyectar solución receptora repetidas veces hasta
que ésta presente un color rojo, vino.
3. Una vez verificado todo lo anterior, colocar el aparato en la posición AUTO para que
de esta manera empezar con la destilación.
4. Destilar en primer lugar un blanco (10 ml de agua destilada), y en caso de que la
lectura sea muy alta (mayor a 0.05) correr otros blancos hasta que esta lectura
disminuya y se mantenga constante.
NOTA: Debido a que el contenido en nitrógeno en el agua destilada dependerá de su
origen, se recomienda obtener un valor medio del mismo para tomarlo como calor de
referencia del blanco.
5. Continuar con el estándar (sulfato ferroso amoniacal) y después con las muestras
problema.
6. Una vez que se han finalizado las determinaciones, apagar el aparato y lavarlo según
el manual del destilador.
NOTA: Una vez lavado el aparato, llenar con agua destilada el recipiente receptor de
titulación. Ante cualquier duda consulte al encargado del laboratorio.
Cálculos:
Donde:
V1 = mililitros de HCl gastados en la titulación de la muestra
V0 = mililitros de HCl gastados en la titulación del blanco
N = normalidad del HCl
g = peso de la muestra en gramos
0.014 = miliequivalentes del HCl

64
Determinación de %Lípidos Totales
Materiales y reactivos:
Balanza analítica
Pinzas
Papel filtro Whatman No.1
Desecador Nalgene
Estufa Thermolyne
Cartucho de celulosa Whatman
(Cat. 2800258)
Campana de extracción de humos
Placas de cristalización
Sistema de cristalización
Soxhlet Glas-col
Matraz balón de fondo plano 500 mL
24/40
Refrigerante 55/50
Sifón 55/50
Éter de petróleo
Cordel
Procedimiento:
1. Poner a peso constante el papel filtro Whatman No. 1.
2. Manejar el papel con pinzas, pesar hoja por hoja en la balanza analítica, anotar el peso
en hoja de reporte, así como marcar el papel con lápiz, identificándolo preferentemente
con un número.
3. A continuación se deja el papel a la intemperie del cuarto para que éste equilibre su
humedad con el ambiente.
4. Sobre el papel filtro pesar de 2.5 a 3 g de la muestra seca a analizar.
NOTA: Esto en caso de que esta misma muestra se utilice también para determinar fibra
cruda, en caso de no ser así, se puede trabajar hasta con 0.5 g de muestra.
5. Una vez pesada la muestra junto con el papel filtro, anotar el peso de la muestra, y hacer
pequeños paquetes amarrando el extremo con un cordel y acomodar cada paquete
dentro de los cartuchos de celulosa.
6. Agregar 250 ml de éter de petróleo a cada uno de los matraces.
7. El cartucho y su contenido se introduce al depósito de extracción.
8. Abrir la llave de agua para hacer funcionar el sistema de refrigeración del sistema de
extracción.
9. Prender el aparato colocando el regulador de temperatura en la posición No. 5 por unos
minutos hasta que el éter empiece a ebullir, una vez que esto suceda mover el
controlador a la posición No. 1 en donde se deja en extracción durante 6 horas.
NOTA: Verificar periódicamente que el sistema de enfriamiento este funcionando
correctamente.

65
10. Una vez transcurridas las 6 horas se apaga el aparato, dejando funcionar solamente el
sistema de enfriamiento, y al cabo de 30 minutos aproximadamente se saca el cartucho
con su contenido del aparato Sxhlet y se escurre el exceso de éter, para luego dejarlo
aireando en un lugar limpio y libre de polvo durante unas 2 horas hasta que ya no se
perciba el olor a éter (realizarlo bajo la campana de extracción), para luego meterlo a la
estufa a 100° C durante 24 horas.
11. Con mucho cuidado se saca cada uno de los paquetitos que hay en los cartuchos, para
meterlos al desecador por 2 horas más.
12. Una vez secos los paquetitos, se procede a pesar cada uno de ellos.
13. Por diferencia de peso encontramos el porcentaje de lípidos de la muestra.
Cálculos:

66
ANEXO C. Tablas ANOVA y a posteriori DVS Tukey
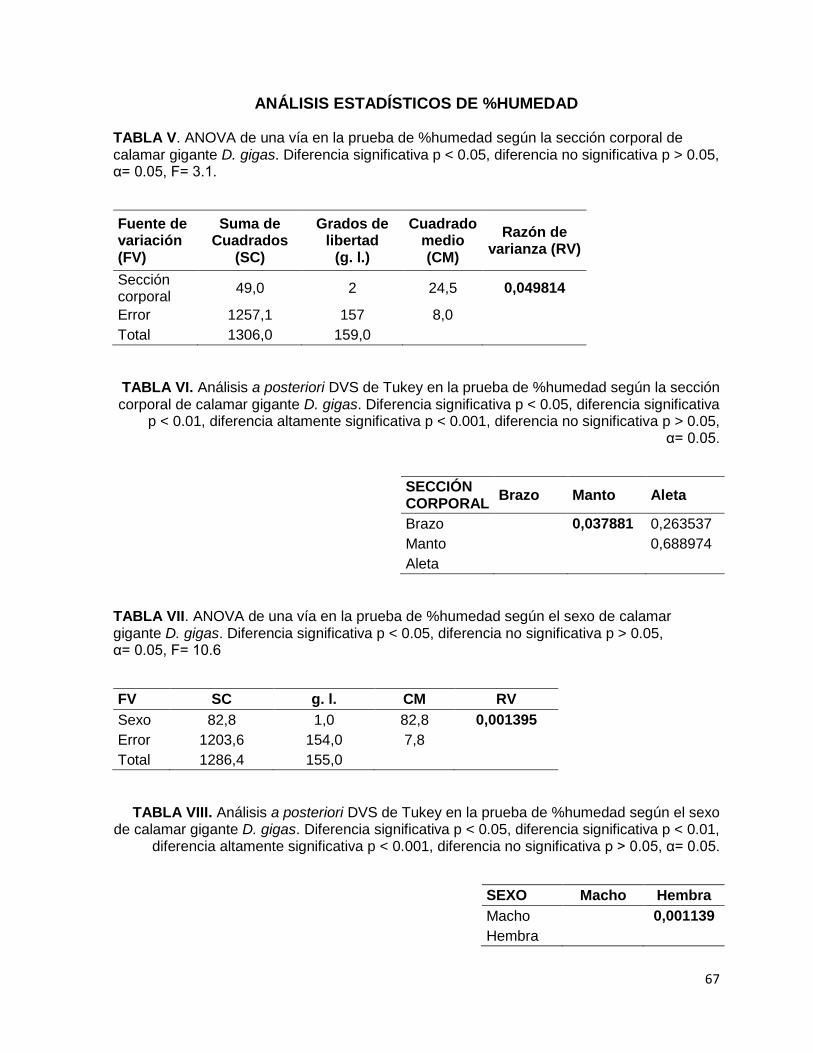
67
ANÁLISIS ESTADÍSTICOS DE %HUMEDAD
TABLA V. ANOVA de una vía en la prueba de %humedad según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 3.1.
Fuente de variación (FV)
Suma de Cuadrados
(SC)
Grados de libertad
(g. l.)
Cuadrado medio (CM)
Razón de varianza (RV)
Sección corporal
49,0 2 24,5 0,049814
Error 1257,1 157 8,0
Total 1306,0 159,0
TABLA VI. Análisis a posteriori DVS de Tukey en la prueba de %humedad según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa
p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SECCIÓN CORPORAL
Brazo Manto Aleta
Brazo 0,037881 0,263537
Manto 0,688974
Aleta
TABLA VII. ANOVA de una vía en la prueba de %humedad según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 10.6
FV SC g. l. CM RV
Sexo 82,8 1,0 82,8 0,001395
Error 1203,6 154,0 7,8
Total 1286,4 155,0
TABLA VIII. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa p < 0.01,
diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SEXO Macho Hembra
Macho 0,001139
Hembra

68
TABLA IX. ANOVA de una vía en la prueba de %humedad según el estado de madurez de calamar gigante D. gigas macho. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 6.33.
FV SC g. l. CM RV
Madurez 97,0 2,0 48,5 0.0044
Error 275,7 36,0 7,7
Total 372,7 38,0
TABLA X. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el estado de madurez de calamar gigante D. gigas macho. Diferencia significativa p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa
p > 0.05, α= 0.05.
MADUREZ Inmaduro Madurando Maduro
Inmaduro 0,682517 0,163019
Madurando 0,004696
Maduro
TABLA XI. ANOVA de una vía en la prueba de %humedad según el estado de madurez de calamar gigante D. gigas hembra. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 0.64.
FV SC g. l. CM RV
Madurez 9,3 2 4,6 0,529504
Error 789,3 109 7,2
Total 798,5155807 111
TABLA XII. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el estado de madurez de calamar gigante D. gigas hembra. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MADUREZ Inmaduro Madurando Maduro
Inmaduro 0,961019 0,536244
Madurando 0,619450
Maduro

69
TABLA XIII. ANOVA de una vía en la prueba de %humedad según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 45.9.
FV SC g. l. CM RV
Mes 781,9 5 156,4 0,00
Error 524,1 154 3,4
Total 1306,0 159,0
TABLA XIV. Análisis a posteriori DVS de Tukey en la prueba de %humedad según el mes de
colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05,
α= 0.05.
MESES Abril Mayo Julio Agosto Septiembre Octubre
Abril 0,000020 0,005284 0,708041 0,109466 0,000020
Mayo 0,000030 0,000020 0,000020 0,018942
Julio 0,117892 0,000020 0,076226
Agosto 0,000029 0,000021
Septiembre 0,000020
Octubre
ANÁLISIS ESTADÍSTICOS DE PROTEÍNAS
TABLA XV. ANOVA de una vía en la prueba de proteínas según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 1.74.
FV SC g. l. CM RV
Sección corporal
35,2 2 17,6 0,178902
Error 1372,1 136 10,1
Total 1407,3 138,0
TABLA XVI. Análisis a posteriori DVS de Tukey en la prueba de proteínas según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa
p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SECCIÓN CORPORAL
Brazo Manto Aleta
Brazo 0,992113 0,225479
Manto 0,240453
Aleta
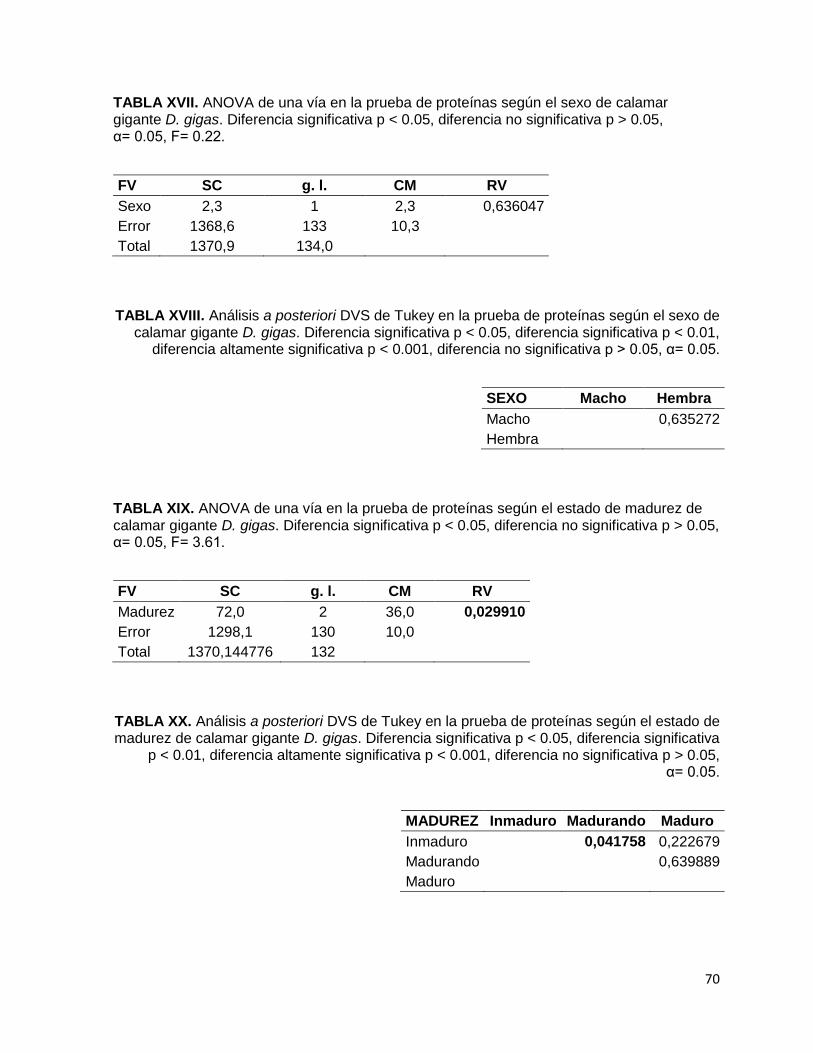
70
TABLA XVII. ANOVA de una vía en la prueba de proteínas según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 0.22.
FV SC g. l. CM RV
Sexo 2,3 1 2,3 0,636047
Error 1368,6 133 10,3
Total 1370,9 134,0
TABLA XVIII. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el sexo de
calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SEXO Macho Hembra
Macho 0,635272
Hembra
TABLA XIX. ANOVA de una vía en la prueba de proteínas según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 3.61.
FV SC g. l. CM RV
Madurez 72,0 2 36,0 0,029910
Error 1298,1 130 10,0
Total 1370,144776 132
TABLA XX. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa
p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MADUREZ Inmaduro Madurando Maduro
Inmaduro 0,041758 0,222679
Madurando 0,639889
Maduro

71
TABLA XXI. ANOVA de una vía en la prueba de proteínas según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 29.6.
FV SC g. l. CM RV
Mes 740,9 5 148,2 0,00
Error 666,4 133 5,0
Total 1407,3 138,0
TABLA XXII. Análisis a posteriori DVS de Tukey en la prueba de proteínas según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MESES Abril Mayo Julio Agosto Septiembre Octubre
Abril 0,000021 0,000594 0,012516 0,002229 0,000020
Mayo 0,360212 0,088866 0,197991 0,000021
Julio 0,965779 0,999156 0,000020
Agosto 0,997564 0,000020
Septiembre 0,000020
Octubre
ANÁLISIS ESTADÍSTICOS DE LÍPIDOS
TABLA XXIII. ANOVA de una vía en la prueba de lípidos según el tipo de sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 6.5017
FV SC g. l. CM RV
Sección corporal
13,9369 2 6,9685 0,002085
Error 128,6140 120 1,0718
Total 142,6 122,0

72
TABLA XXIV. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el tipo de sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SECCIÓN CORPORAL
Brazo Manto Aleta
Brazo 0,004287 0,953184
Manto 0,016480
Aleta
TABLA XXV. ANOVA de una vía en la prueba de lípidos según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 0.9414
FV SC g. l. CM RV
Sexo 1,1202 1 1,1202 0,333921
Error 139,2255 117 1,1900
Total 140,3 118,0
TABLA XXVI. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa p < 0.01,
diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SEXO Macho Hembra
Macho 0,334033
Hembra
TABLA XXVII. ANOVA de una vía en la prueba de lípidos según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 3.0173
FV SC g. l. CM RV
Madurez 7,0417 2 3,5208 0,052824
Error 134,1920 115 1,1669
Total 141,2336391 117

73
TABLA XXVIII. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MADUREZ Inmaduro Madurando Maduro
Inmaduro 0,475828 0,049812
Madurando 0,743390
Maduro
TABLA XXIX. ANOVA de una vía en la prueba de lípidos según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 25.3489
FV SC g. l. CM RV
Mes 74,1249 5 14,8250 0,00
Error 68,4260 117 0,5848
Total 142,6 122,0
TABLA XXX. Análisis a posteriori DVS de Tukey en la prueba de lípidos según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa
p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MESES Abril Mayo Julio Agosto Septiembre Octubre
Abril 0,098697 0,001178 0,027924 0,333862 0,462388
Mayo 0,000123 0,000123 0,981091 0,802862
Julio 0,999831 0,000123 0,000123
Agosto 0,000125 0,000125
Septiembre 0,997489
Octubre
ANÁLISIS ESTADÍSTICOS DE CENIZAS
TABLA XXXI. ANOVA de una vía en la prueba de cenizas según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 6.220
FV SC g. l. CM RV
Sección corporal
30,823 2 15,412 0,002768
Error 270,091 109 2,478
Total 300,9 111,0

74
TABLA XXXII. Análisis a posteriori DVS de Tukey en la prueba de cenizas según la sección corporal de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa
p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SECCIÓN CORPORAL
Brazo Manto Aleta
Brazo 0,010367 0,006644
Manto 0,851510
Aleta
TABLA XXXIII. ANOVA de una vía en la prueba de cenizas según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 0.8819
FV SC g. l. CM RV
Sexo 2,216 1 2,216 0,349822
Error 266,345 106 2,513
Total 268,6 107,0
TABLA XXXIV. Análisis a posteriori DVS de Tukey en la prueba de cenizas según el sexo de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia significativa p < 0.01,
diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
SEXO Macho Hembra
Macho 0,349947
Hembra
TABLA XXXV. ANOVA de una vía en la prueba de cenizas según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 28.1792
FV SC g. l. CM RV
Madurez 103,969 2 51,985 0,000000
Error 195,548 106 1,845
Total 299,5174157 108
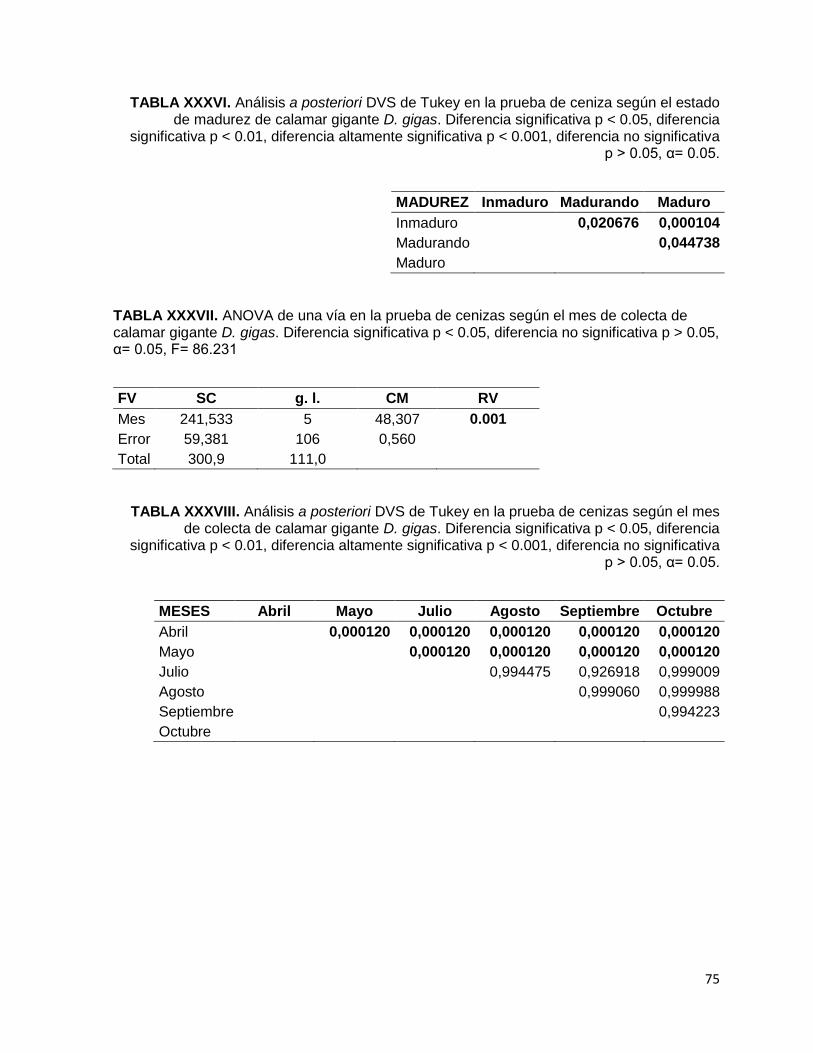
75
TABLA XXXVI. Análisis a posteriori DVS de Tukey en la prueba de ceniza según el estado de madurez de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MADUREZ Inmaduro Madurando Maduro
Inmaduro 0,020676 0,000104
Madurando 0,044738
Maduro
TABLA XXXVII. ANOVA de una vía en la prueba de cenizas según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia no significativa p > 0.05, α= 0.05, F= 86.231
FV SC g. l. CM RV
Mes 241,533 5 48,307 0.001
Error 59,381 106 0,560
Total 300,9 111,0
TABLA XXXVIII. Análisis a posteriori DVS de Tukey en la prueba de cenizas según el mes de colecta de calamar gigante D. gigas. Diferencia significativa p < 0.05, diferencia
significativa p < 0.01, diferencia altamente significativa p < 0.001, diferencia no significativa p > 0.05, α= 0.05.
MESES Abril Mayo Julio Agosto Septiembre Octubre
Abril 0,000120 0,000120 0,000120 0,000120 0,000120
Mayo 0,000120 0,000120 0,000120 0,000120
Julio 0,994475 0,926918 0,999009
Agosto 0,999060 0,999988
Septiembre 0,994223
Octubre

76
ANEXO D. Recetario

77
Spaghetti en salsa de calamar
por Donaji Ochoa Tepetla
Porciones: 6
Ingredientes
500 g de spaguetti cocido al dente en
agua con sal
½ taza de queso doble crema
chiapaneco
Ingredientes para el filete de calamar
1 kg de filete de calamar
2 cda. de vinagre de vino tinto
1 cda. de sal
1 cda. de azúcar
1 naranja (jugo)
3 hojas de laurel
Ingredientes para Salsa
1 cda. de mantequilla sin sal
Aceite de oliva cantidad necesaria (c/n)
200 g de cebolla
Ajo 2 dientes
500 g tomates
220 ml de puré de tomate
Sal y pimienta c/n
Hierbas de olor c/n: tomillo, orégano y
albahaca frescos
250 ml de vino tinto
3 gotas de tinta de calamar
30 g de filete de calamar previamente
hervido
Preparación
Hervir por 45 minutos el filete de calamar con abundante agua, añadir el vinagre, sal, azúcar,
jugo de naranja y las hojas de laurel. Sacar del agua el filete de calamar y retirarle una capa
delgada que lo recubre (porque eso es lo que amarga la preparación). Cortar en cubos
pequeños y reservar.
En una olla calentar la mantequilla y el aceite de oliva, sofreír la cebolla y el ajo. Se retirar
únicamente la cebolla y el ajo y mezclarlos en la licuadora junto con los tomates, el puré de
tomate y 30 g de filete de calamar (un par de cubos previamente cortados). Se reserva para
la salsa.
Con la mantequilla y el aceite de oliva restante se colocan los cubos de calamar hasta que se
sellen, salpimentar, añadir el tomillo y el orégano picados. Desglasar con vino tinto y dejar
evaporar el alcohol.
Incorporar la reserva de la salsa y cocinar por 30 minutos, añadir las gotas de tinta de
calamar y finalmente añadir la albahaca picada, revolver.
Servir la pasta al dente con la salsa y el queso doble crema por encima.
Acompañe el spaghetti en salsa de calamar con su bebida predilecta, invite a un comensal y
saboreen juntos cada uno de los aromas en conjunto y enamórense… Bon appetit.

78
El individuo ha luchado siempre para no ser absorbido por la tribu.
Si lo intentas, a menudo estarás solo, y a veces asustado.
Pero ningún precio es demasiado alto por el privilegio de ser uno mismo.
Friedrich Nietzsche





![ÁÁÁX µ Ç]v l] vX]v(} · Provisional Merit List of Candidates for admission in MBBS at KIMS against OPEN MERIT seats in session 2016-2017](https://static.fdocuments.es/doc/165x107/5f0a69327e708231d42b8033/x-v-l-vxv-provisional-merit-list-of-candidates-for-admission-in.jpg)












