Anatomía y morfología comparada del desarrollo del … · A la Universidad Nacional de Colombia,...
70
Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu Cyphostyleae (Melastomataceae), y su interpretación en un contexto filogenético Támara Isabel Dobaiba Sandino Ramírez Universidad Nacional de Colombia Facultad de Ciencias, Departamento de Biología Bogotá, Colombia 2011
Transcript of Anatomía y morfología comparada del desarrollo del … · A la Universidad Nacional de Colombia,...

Anatomía y morfología comparada del desarrollo
del ovario y fruto en la tribu Cyphostyleae
(Melastomataceae), y su interpretación en un
contexto filogenético
Támara Isabel Dobaiba Sandino Ramírez
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento de Biología
Bogotá, Colombia
2011


Anatomía y morfología comparada del desarrollo
del ovario y fruto en la tribu Cyphostyleae
(Melastomataceae), y su interpretación en un
contexto filogenético
Támara Isabel Dobaiba Sandino Ramírez
Tesis presentada como requisito parcial para optar al título de:
Magister en Ciencias - Biología
Director:
Doctor Xavier Marquínez Casas
Línea de Investigación:
Sistemática
Grupo de Investigación:
Sistemática Biológica
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento de Bilogía
Bogotá, Colombia
2011


A mi familia, maestros y maestras


Agradecimientos
Agradezco a mis profesores Xavier Marquínez, Fabián Michelangeli y María Eugenia
Morales por su invaluable ayuda, paciencia y consejos en cada momento del desarrollo
teórico y experimental de este trabajo.
A mi familia y a Carlos por todo su apoyo y compañía de cada momento de este capítulo
de mi carrera y vida.
A Heriberto David y a Saúl Hoyos Gómez, por su colaboración en la colección del
material.
A la Universidad Nacional de Colombia, al departamento de Biología y al programa de
postgrado de Biología por haber aportado tanto en mi desarrollo profesional y personal.
A los encargados de los laboratorios biología de plantas y morfología de de plantas, del
departamento de biología, Mónica Cuervo y José Choachi por su apoyo durante el
desarrollo experimental del proyecto.
Al Jardín Botánico José Celestino Mutis por permitirme acceder a la colección de
Melastomataceae.
Al proyecto PBI Miconieae “NSF (DEB-0818399)”, del Jardín Botánico The New York
Botanical Garden y al Grupo Sistemática Biológica, UPTC-UNC por su apoyo económico
a este trabajo.


Resumen y Abstract IX
Resumen
En la familia Melastomataceae encontramos una correlación entre los caracteres
posición del ovario y el tipo de fruto así: tribus con ovario libre presentan fruto capsular y
tribus con ovario totalmente fusionado al hipanto, presentan fruto en baya. En la tribu
Cyphostyleae se presenta una condición atípica en donde las flores son epíginas y el
fruto es capsular. Mediante un estudio comparativo de la anatomía y morfología del
ovario y del fruto a lo largo del desarrollo, se evaluaron las homologías primarias
observadas en el proceso de desarrollo de botones, flores y frutos en dos especies de la
tribu Cyphostyleae, comparándolas con seis especies de las tribus Blakeeae,
Merianieae, Miconieae y Melastomeae. Se encontraron diferencias en el grado de fusión
entre ovario e hipanto; en las características y distribución de tejidos. Se realizó un
modelo de vascularización floral para cada especie. Los caracteres observados se
interpretaron en un contexto filogenético.
Palabras clave: Melastomataceae, Anatomía, Desarrollo, Flor, Fruto.
Abstract
In Melastomataceae family there is a correlation between the characters ovary position
and type fruit, then: tribes with ovary free presenting capsular fruit and tribes with ovary
completely fused to the hypanthium, presenting fruit berry. In the tribe Cyphostyleae
shows up an atypical condition where the flowers are epíginas and the fruit is capsular.
Primary homology observed in the process of development of bellboys, flowers and fruits
were evaluated by means of a comparative study of the anatomy and morphology of the
ovary and of the fruit throughout the development in two species of the tribe
Cyphostyleae, comparing them with six species of the tribes Blakeeae, Merianieae,
Miconieae and Melastomeae. There were differences in the degree of fusion of ovary and

X Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
hypanthium; the characteristics and distribution of tissues. We performed a floral
vascularization model for each species. The observed characteristics were interpreted in
a phylogenetic context.
Keywords: Melastomataceae, Anatomy, Development, Flower, Fruit.

XI
Contenido
Pág.
Resumen ......................................................................................................................... IX
Lista de figuras ............................................................................................................. XV
Lista de tablas ............................................................................................................ XVII
Lista de Símbolos y abreviaturas ............................................................................... XIX
Introducción .................................................................................................................... 1
1. Marco teórico ........................................................................................................... 3
1.1 Estructura de la flor y posición del ovario ............................................................... 5
1.2 Tipo de fruto ............................................................................................................ 8
2. Planteamiento del problema .................................................................................... 9
3. Objetivos ................................................................................................................. 12
3.1 Objetivo general............................................................................................. 12
3.2 Objetivos específicos ..................................................................................... 12
4. Metodología ............................................................................................................ 13
4.1 Fase de campo .............................................................................................. 13
4.2 Fase de laboratorio ........................................................................................ 15
6. Resultados .............................................................................................................. 18
6.1. Allomaieta hirsuta -------------------------------------------------------------------------------- 18
6.1.1. Descripción morfológica -------------------------------------------------------------- 18
6.1.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 18
Vascularización del pedicelo y el receptáculo ------------------------------- 19
Vacularización del ovario+hipanto --------------------------------------------- 20

XII Anatomía y morfología comparadas del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Vascularización del perianto----------------------------------------------------- 20
Vascularización del estilo y el estigma --------------------------------------- 20
6.1.3. Dehiscencia ------------------------------------------------------------------------------21
6.2. Allomaieta zenufanasana ----------------------------------------------------------------------- 24
6.2.1. Descripción morfológica -------------------------------------------------------------- 24
6.2.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 25
Vascularización del pedicelo y el receptáculo ------------------------------- 25
Vascularización del ovario+hipanto -------------------------------------------- 25
Vascularizacion del perianto ----------------------------------------------------- 27
Vascularizaciín del estilo y el estigma ----------------------------------------- 28
6.2.3. Desarrollo---------------------------------------------------------------------------------- 28
Cambios tisulares en el pedicelo y el receptáculo-------------------------- 29
Cambios tisulares en el ovario+hipanto -------------------------------------- 32
6.2.4. Dehiscencia ----------------------------------------------------------------------------- 35
6.3. Blakea granatensis ------------------------------------------------------------------------------ 36
6.3.1. Descripción morfológica -------------------------------------------------------------- 36
6.3.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 37
Vascularización del pedicelo, receptáculo y eje central del ovario ---- 37
Vascularización del ovario+hipanto y el perianto -------------------------- 38
Vascularizaciín del estilo y el estigma ---------------------------------------- 41
6.3.3. Desarrollo -------------------------------------------------------------------------------- 41
Cambios tisulares en el pedicelo y el receptáculo-------------------------- 43
Cambios tisulares en el ovario+hipanto--------------------------------------- 48
6.3.4. Dehiscencia ------------------------------------------------------------------------------ 49
6.4. Meriania nobilis ----------------------------------------------------------------------------------- 50
6.4.1. Descripción morfológica -------------------------------------------------------------- 50
6.4.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 50
Vascularización del pedicelo, el receptáculo y el hipanto ---------------- 52
Vascularización del ovario, el estilo y el estigma --------------------------- 52
Vascularización del perianto ---------------------------------------------------- 55
6.4.3. Desarrollo --------------------------------------------------------------------------------- 55
Cambios tisulares en el pedicelo y el receptáculo ------------------------- 56
Cambios tisulares en el ovario -------------------------------------------------- 60

Contenido XIII
Cambios tisulares en el hipanto ------------------------------------------------- 62
Dehiscencia -------------------------------------------------------------------------- 63
6.5. Miconia notabilis ---------------------------------------------------------------------------------- 65
6.5.1. Descripción morfológica -------------------------------------------------------------- 65
6.5.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 66
Vascularización del pedicelo, el receptáculo y el hipanto ---------------------- 66
Vascularización del ovario el estilo y el estigma----------------------------- 66
Vascularizaciín del perianto ------------------------------------------------------ 68
6.5.3. Desarrollo --------------------------------------------------------------------------------- 69
Cambios tisulares en el pedicelo y el receptáculo-------------------------- 71
Cambios tisulares en el ovario--------------------------------------------------- 74
Cambios tisulares en el hipanto ------------------------------------------------- 76
6.5.4. Dehiscencia ------------------------------------------------------------------------------ 77
6.6. Miconia squamulosa ----------------------------------------------------------------------------- 78
6.6.1. Descripción morfológica -------------------------------------------------------------- 78
6.6.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 78
Vascularización del receptáculo, el ovario y el hipanto ------------------- 80
Vascularización del estilo y el estigma ---------------------------------------- 80
6.6.3. Desarrollo --------------------------------------------------------------------------------- 83
Cambios tisulares en el pedicelo y el receptáculo-------------------------- 85
Cambios tisulares en el ovario--------------------------------------------------- 87
Cambios tisulares en el hipanto ------------------------------------------------- 89
6.6.4. Dehiscencia ------------------------------------------------------------------------------ 90
6.7. Tibouchina mollis --------------------------------------------------------------------------------- 91
6.7.1. Descripción morfológica -------------------------------------------------------------- 91
6.7.2. Estructura del ovario y el hipanto antes de antesis ---------------------------- 91
Vascularización del pedicelo, el receptáculo y el hipanto ---------------- 92
Vascularización del ovario -------------------------------------------------------- 93
Vascularizaciín del estilo y el estigma ----------------------------------------- 93
6.7.3. Desarrollo --------------------------------------------------------------------------------- 96
Cambios tisulares en el pedicelo y el receptáculo ------------------------- 98
Cambios tisulares en el ovario ------------------------------------------------- 100
Cambios tisulares en el hipanto y en los tabiques anterales ----------- 102
6.7.4. Dehiscencia -----------------------------------------------------------------------------103

XIV Anatomía y morfología comparadas del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
6.8. Tibouchina urvilleana ---------------------------------------------------------------------------105
6.8.1. Descripción morfológica -------------------------------------------------------------105
6.8.2. Estructura del ovario y el hipanto antes de antesis ---------------------------106
Vascularización del pedicelo, el receptáculo y el hipanto ---------------107
Vascularización del ovario ------------------------------------------------------108
Vascularizaciín del estilo y el estigma -------------------------------------- 110
6.8.3. Desarrollo ------------------------------------------------------------------------------ 111
Cambios tisulares en el pedicelo y el receptáculo ----------------------- 113
Cambios tisulares en el ovario ------------------------------------------------ 117
Cambios tisulares en el hipanto y en los tabiques anterales ----------- 117
6.8.4. Dehiscencia.----------------------------------------------------------------------------- 121
7. Discusión ............................................................................................................... 125
7.1 Posición del ovario y plan de desarrollo o bauplan ....................................... 125
7.2 Vascularización ............................................................................................ 133
7.3 Desarrollo del fruto ....................................................................................... 138
Divisiones celulares ...................................................................................... 140
Incremento del área celular .......................................................................... 143
Consistencia del fruto, disolución de tejidos y presecencia de esclereidas ... 144
8. Conclusiones ........................................................................................................ 147
Bibliografía ................................................................................................................... 149

XV
Lista de figuras
Pág.
Figura 1: Patrones de desarrollo floral temprano ................................................... 7
Figura 2: Allomaieta hirsuta, detalles de la morfología externa de la flor .............. 19
Figura 3: Allomaieta hirsuta, estructura y anatomia floral .................................... 22
Figura 4: Allomaieta zenufanasana, detalles de la morfología externa de la flor ... 24
Figura 5: Allomaieta zenufanasana, estructura y anatomia floral ......................... 26
Figura 6: Allomaieta zenufanasana, estadios del desarrollo ................................. 30
Figura 7: Allomaieta zenufanasana, desarrollo de tejidos en pedicelo y receptáculo.
. Cortes transversales ............................................................................ 31
Figura 8: Allomaieta zenufanasana, cortes transversales a nivel del ovario y el
. hipanto .................................................................................................. 33
Figura 9: Allomaieta zenufanasana, fruto dehiscente ........................................... 35
Figura 10: Blakea granatensis, detalles de la morfología externa de la flor............. 37
Figura 11: Blakea granatensis, estructura y anatomia floral .................................. 39
Figura 12: Blakea granatensis, estadios del desarrollo ........................................... 42
Figura 13: Blakea granatensis, desarrollo de tejidos en pedicelo y receptáculo.
Cortes transversales ............................................................................. 45
Figura 14: Blakea granatensis, cortes transversales a nivel del ovario y el hipanto..47
Figura 15: Meriania nobilis, detalles de la morfología externa de la flor .................. 51
Figura 16: Meriania nobilis, estructura y anatomia floral. ........................................ 53
Figura 17: Meriania nobilis, estadios del desarrollo ................................................ 57
Figura 18: Meriania nobilis, desarrollo de tejidos en pedicelo y receptáculo. Cortes
transversales ......................................................................................... 60
Figura 19: Meriania nobilis, cortes transversales a nivel del ovario y el hipanto ..... 62
Figura 20: Meriania notabilis, detalles de la morfología externa de la flor…………..65
Figura 21: Miconia notabilis, estructura y anatomia floral ...................................... 67
Figura 22: Miconia notabilis, estadios del desarrollo............................................... 70
Figura 23: Miconia notabilis, desarrollo de tejidos en pedicelo y receptáculo ......... 73

XVI Anatomía y morfología comparadas del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Figura 24: Miconia notabilis, cortes transversales a nivel del ovario y el hipanto .... 75
Figura 25: Miconia squamulosa, detalles de la morfología externa de la flor ........... 79
Figura 26: Miconia squamulosa, estructura y anatomia floral ................................. 81
Figura 27: Miconia squamulosa, estadios del desarrollo ......................................... 85
Figura 28: Miconia squamulosa, cortes transversales a nivel del pedicelo y el
receptáculo ............................................................................................ 87
Figura 29: Miconia squamulosa, cortes transversales a nivel del ovario y el hipanto ..89
Figura 30: Tibouchina mollis, detalles de la morfología externa de la flor ................ 92
Figura 31: Tibouchina mollis, estructura y anatomia floral ...................................... 94
Figura 32: Tibouchina mollis, estadios del desarrollo .............................................. 97
Figura 33: Tibouquina mollis, cortes transversales anivel del pedicelo.................... 98
Figura 34: Tbouchina mollis, desarrollo de tejidos en el ovario y el hipanto .......... 102
Figura 35: Tibouchina mollis, fruto en dehiscencia ................................................ 104
Figura 36: Tibouchina urvilleana, detalles de la morfología externa de la flor ........ 106
Figura 37: Tibouchina urvilleana, estructura y anatomia floral .............................. 109
Figura 38: Tibouchina urvilleana, estadios del desarrollo ...................................... 112
Figura 39: Tibouchina urvilleana, desarrollo de tejidos en pedicelo y receptáculo . 116
Figura 40: Tibouchina urvilleana, desarrollo de tejidos en el ovario y el hipanto ... 119
Figura 41: Tibouchina urvilleana, cortes transversales de tabuiques anterales ..... 122
Figura 42: Tibouchina urvilleana, fruto dehiscente ................................................ 123
Figura 43: Hipanto durante la dehiscencia ............................................................ 124
Figura 44. Optimización del carácter “posición del ovario”……………………………….131
Figura 45. Optimización del carácter “presencia y tamaño de bolsillos anterales”…..…133
Figura 46. Optimización de los caracteres relativos a la estructura y vascularización de la
flor. ………………………………………………………………………………………………136
Figura 47. Optimización del carácter “tipo de fruto”. ………………………………………140
Figura 48. Optimización de los caracteres relativos al desarrollo de la flor y el fruto…..142

XVII
Lista de tablas
Pág.
Tablas 1-3. Material Examinado…………………………………………………………….. 13
Tabla 4. Promedio de capas celulares y área promedio de las células en el pedicelo y
el receptáculo para los diferentes estadios de desarrollo en Allomaieta
zenufanasana…………………..………………………………………………...32
Tabla 5. Número promedio de capas celulares y área promedio celular en el ovario
para los diferentes estadios de desarrollo en Allomaieta
zenufanasana………………………..…………………………….……………..34
Tabla 6. Número promedio de capas celulares y área promedio de las células en el
pedicelo y el receptáculo para los diferentes estadios de desarrollo de
Blakea granatensis..…………………………………………………………….44
Tabla 7. Número promedio de capas celulares y área promedio de las células en los
tejidos del ovario+hipanto y en los tabiques anterales en Blakea
granatensis……………………….……….……………………………………..46
Tabla 8. Número promedio de capas celulares y área promedio de las células en el
pedicelo y el receptáculo para los diferentes estadios de desarrollo en
Meriania nobilis…………………………………………………………………..59
Tabla 9. Número promedio capas celulares y área promedio celular, en la pared
externa, los septos y el eje central del ovario a lo largo del desarrollo en
Meriania nobilis…………………………………………………………………...61
Tabla 10. Número promedio de capas celulares y área promedio de las células en el
hipanto en Meriania nobilis……..……………………………………………….63
Tabla 11. Promedio de capas celulares y área promedio de las células en el pedicelo y
el receptáculo para los diferentes estadios de desarrollo en Miconia
notabilis……………………………………………………………………………72

XVIII Anatomía y morfología comparadas del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Tabla 12. Promedio de células y área promedio celular, en la pared externa, los
septos y el eje central del ovario a lo largo del desarrollo de Miconia
notabilis…………………………………………………………………………...74
Tabla 13. Promedio de capas celulares y área promedio de las células en el hipanto
de Miconia notabilis………………………………………………………………76
Tabla 14. Promedio de capas celulares y área promedio de las células en el pedicelo y
el receptáculo para los diferentes estadios de desarrollo de Miconia
squamulosa………………………………………………………………………86
Tabla 15. Promedio de células y área promedio celular, en la pared externa, los septos y
el eje central del ovario a lo largo del desarrollo de Miconia squamulosa....88
Tabla 16. Promedio de capas celulares y área promedio de las células en el hipanto de
Miconia squamulosa......................................................................................90
Tabla 17. Promedio de células y área promedio celular, en la corteza y la médula del
pedicelo y el receptáculo a lo largo del desarrollo de Tibouchina mollis…..99
Tabla 18. Promedio de células y área promedio celular, en la pared externa, los septos y
el eje central del ovario a lo largo del desarrollo de Tibouchina mollis.......101
Tabla 19. Promedio de capas celulares y área celular promedio en los tabiques anterales
y el hipanto de Tibouchina mollis……………………………………………103
Tabla 20. Promedio de capas celulares y área promedio de las células en la médula y la
corteza del pedicelo y el receptáculo para los diferentes estadios de
desarrollo de Tibouchina urvilleana……………………………………….….115
Tabla 21. Promedio de células y área promedio celular, en la pared externa, los septos y
el eje central del ovario a lo largo del desarrollo de Tibouchina urvilleana...118
Tabla 22. Promedio de capas celulares y área celular promedio en los tabiques anterales
y el hipanto Tibouchina urvilleana……………………………………………120
Tabla 23. Caracteres variables referentes a la morfología y estructura floral…………..126
Tabla 24. Proporción de los lóculos del ovario que se encuentran por debajo del punto de
fusión entre el ovario y el hipanto, durante los nueve estadios del desarrollo,
para las especies en estudio……………………………………………….…130
Tabla 25. Caracteres anatómicos variables en la flor……………………………………..134

XIX
Lista de Símbolos y abreviaturas
Símbolo Término
at Antera
ba Bolsillos en donde se encuentran las enteras en el botón (bolsillos anterales).
br Brácteas y bractéolas.
c Carpelos.
cc Cilindro vascular central.
co Conectivo.
est Estambres.
fl Filamento.
hc Haces corticales.
hd Haces dorsales del ovario.
He Haces vasculares del estilo.
hi Hipanto.
hh Haces mayores del hipanto.
hh’ Haces secundarios del hipanto.
hl Haces laterales del os carpelos.
hm Haces mediales de los carpelos.
hv Haces ventrales del ovario.
hvp Haces vasculares de las placentas.
lc Lóbulos del cáliz.
lo Lóculos del ovario.

XX Anatomía y morfología comparadas del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Símbolo Término
med Haces medulares.
o Óvulos.
ov Ovario.
p Placentas.
pc Trazas centrales de los pétalos.
pe Pedicelo.
pl Trazas laterales de los pétalos.
pt Pétalos.
se Semilla.
st Estilo.
sc Trazas vasculares centrales de los sépalos.
sl Trazas vasculares laterales de los sépalos.
tb Trazas vasculares de las brácteas.
te Trazas vasculares de los estambres.
tp Trazas de los pétalos.
tq Tabiques que separan las anteras en el botón (Rabiques anterales).
ts Trazas vasculares de los sépalos.
tt Tejido de transmisión.

1
Introducción
Aunque la circunscripción y monofília de Melastomataceae es clara, encontrándose
discrepancias únicamente acerca de si se considera a Memecyloideae como una
subfamilia de Melastomataceae o como la familia hermana (Renner 1993; Clausing &
Renner 2001); la determinación de las relaciones de parentesco al interior de
Melastomataceae han resultado mucho más difíciles de establecer.
Entre los primeros esfuerzos por entender la diversidad en Melastomataceae, se
definieron entre 9-14 tribus (Triana 1871; Cogniaux 1891; Vliet et al. 1981; Renner 1993),
utilizándose en su taxonomía, como caracteres diagnósticos, el tipo de fruto (cápsula y
baya) y la posición del ovario (ínfero, semiínfero, súpero).
Sin embargo, estos dos caracteres están muy relacionados con la protección y dispersión
de las semillas, por lo que en su evolución se pueden ver afectados por selección
direccional (Clausing et al. 2000), con lo cual se espera el surgimiento independiente, en
forma de paralelismos y convergencias, de estas características que suponen una
ventaja adaptativa (Larson & Losos 1996). En diversas filogenias de angiospermas se
han detectado homoplasias para este tipo de caracteres así: en Myrtaceae (Johnson &
Briggs 1984); en Saxifragaceae (Soltis et al. 1990); en Rosaceae (Morgan et al. 1994); en
Ranunculaceae (Hoot 1991); en Lamiales s. l., (Wagstaff & Olmstead 1997); en

2 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae).
Asteraceae (Gustafsson & Albert 1999) y en Melastomataceae (Clausing et al. 2000;
Clausing & Renner 2001; Michelangeli 2000 y Fritsch et al. 2004).
Siendo los caracteres “tipo de fruto” y “posición del ovario”, frecuentemente
homoplásicos, pueden existir problemas en la identificación de homologías primarias, en
especial en un grupo tan diverso como Melastomataceae; adicionalmente, en esta familia
se presenta una fuerte correlación de presencia de ovario ínfero con fruto en baya, y
ovario súpero con fruto en cápsula. En el neotrópico se presenta excepción a esta norma
en cuatro géneros, tres de estos eventualmente ubicados o asociados con la tribu
Cyphostyleae, en los cuales se presenta ovario ínfero con fruto en cápsula. El presente
trabajo busca explorar esta correspondencia, en el desarrollo flor-fruto de distintos grupos
de Melastomataceae con el fin de revisar las homologías utilizadas en la taxonomía y en
las reconstrucciones filogenéticas de la familia.

1. Marco teórico
Melastomataceae es una familia muy diversa en el bosque tropical húmedo, presente
también en sabanas y bosque de alta montaña (Michelangeli et al. 2004), que cuenta con
cerca de 4500 especies, presenta su mayor diversidad en el neotrópico (~3000
especies), seguida por el sureste asiático (~1000), África (240) y Madagascar (225).
Aunque la circunscripción y monofília de la familia Melastomataceae han sido claramente
establecidas (Clausing & Renner 2001), la definición de taxones en su interior, así como
las relaciones taxonómicas y/o filogenéticas, entre estos han dado lugar a varias
propuestas de clasificación en los últimos 130 años (Triana 1871, Cogniaux 1891, Vliet et
al.1981, Renner 1993).
La familia Melastomataceae presenta dos subfamilias: Melastomatoideae, con diez tribus,
siete de las cuales tienen representación en Colombia; y Memecyloideae, conformada
por una sola tribu pantropical y seis géneros, de los cuales dos son neotropicales. Las
tribus están definidas principalmente por caracteres de la antera, forma de la semilla, tipo
de fruto y posición del ovario (Mendoza & Ramírez 2006). La familia presenta flores
epíginas o períginas en las que varía el grado de fusión entre el ovario y el hipanto
(Renner 1993), desde libre hasta totalmente fusionado. Las tribus Bertolonieae,
Melastomeae, Merianieae, Microlicieae y Sonereliae presentan ovario libre que da lugar a
un fruto en cápsula, mientras que en las tribus Blakeeae, Miconieae (Renner 1993) y
Henrietteae (Penneys et al. 2010), el ovario está fusionado al hipanto y da lugar a un
fruto en baya. Además de los frutos en baya y en cápsula la familia Melastomataceae
presenta numerosos tipos de fruto intermedios, como bayas secas, cápsulas carnosas y
bayas leñosas (Clausing et al. 2000).

4 Anatomía y Morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Como excepción a la aparente correspondencia, entre ovario libre con frutos capsulares y
ovario ínfero con frutos tipo baya; en el nuevo mundo se encuentran cinco géneros
(Alloneuron Pilg., Allomaieta Gleason, Physeterostemon R. Goldenberg & Amorim,
Tateanthus Gleason y Wurdastom B. Walln.), los cuales presentan frutos capsulares
derivados de flores con ovarios ínferos. Recientemente Michelangeli et al. (en
preparación) mediante un análisis filogenético con secuencias de ndhF y rbcL, han
sustentado la monofília de la tribu Cyphostyleae, ubicándola como grupo hermano de
Sonerileae+Dissochaetae, de fruto capsular y ovario libre; y a su vez este taxón queda
como hermano de un taxón más grande que incluye especies de Melastomeae y de
Bertolonieae, también de fruto capsular y ovario libre.
La tribu Cyphostyleae Gleason (Gleason 1929) originalmente estuvo conformada por los
géneros Allomaieta y Cyphostyla, Renner (1993) la incluyó en la tribu Miconieae por
presentar ovario ínfero y la asoció con Alloneuron; posteriormente el género Wurdastom
es segregado de Alloneuron por Wallnöfer (1996), en tanto que Lozano & Becerra (1999)
definen de nuevo a Cyphostyleae como una tribu independiente de Miconieae e incluyen
al género Cyphostyla dentro de la sinonimia de Allomaieta. En la filogenia de
Michelangeli et al. (en preparación) la especie muestreada de Alloneuron Pilg. queda
como grupo hermano del clado formado por las especies de Allomaieta.
El género Allomaieta está restringido a los Andes de Colombia (Lozano & Becerra 1999),
en tanto que Alloneuron está presente en Colombia (Wallnöfer 1996), Ecuador

Marco teórico 5
(www.trópicos.org 2008) y Perú (Wallnöfer 1996), al igual que Wurdastom
(www.trópicos.org 2008, León 2006). El otro género de Melastomataceae que presenta la
condición de ovario ínfero y fruto capsular es Physeterostemon, endémico de Brasil, cuyo
grupo hermano es Eriocnema (también de Brasil y con frutos capsulares, pero derivados
de ovarios libres); éstos dos géneros, a su vez son grupo hermano de Miconieae
(Goldenberg & Amorim 2006).
1.1. Estructura de la flor y posición del ovario
Los caracteres de la estructura de la flor y del fruto han sido ampliamente utilizados en la
taxonomía y sistemática de angiospermas, encontrándose desde el inicio de los sistemas
modernos de clasificación (cf. Linnaeus 1753). Jean-Baptiste Payer (1857, citado por
Leins & Erbar 1997) generó una vasta información acerca del desarrollo floral en
numerosas familias de angiospermas, impulsando el uso de la ontogenia en la
interpretación de las estructuras florales. Estos estudios del desarrollo contribuyeron
posteriormente en la reevaluación de homologías primarias al considerar las diferencias
en los patrones de desarrollo de estructuras aparentemente similares como evidencia de
homoplasia.
Varios autores (Boke 1964, 1968; Kaplan 1967; Simpson 1998; Kuzoff et al. 2001; Soltis
& Hufford 2002; Litt & Stevenson 2003; Soltis el al. 2003) han utilizado los estados
tempranos del desarrollo del meristema floral para la distinción entre flores hipóginas y
epíginas, así como entre diferentes formas de epigínia. Kuzoff et al. (2001) hicieron un

6 Anatomía y Morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
importante aporte en la evaluación de homologías primarias al reinterpretación el carácter
“posición del ovario”, definiéndolo con base en los patrones básicos de organogénesis en
el inicio del desarrollo floral, describiendo dos patrones de desarrollo (“bauplan”):
hipógino y epígino. La mayor parte de las flores que presentan ovario súpero en la flor
madura son el resultado de un patrón de desarrollo hipógino, en el cual, el ápice floral es
convexo durante toda la organogénesis. Por otra parte, la mayor parte de las flores que
presentan ovario ínfero en la flor madura son el resultado de un patrón de desarrollo floral
epígino-apendicular, en donde el ápice floral es cóncavo durante toda la organogénesis
(Fig. 1).
Una modificación en el estado final del desarrollo de un patrón de desarrollo hipógino es
el denominado epígino-receptacular, el cual surge cuando un ápice floral es inicialmente
convexo, pero justo después de que el primordio del gineceo emerge, la periferia del
ápice floral se expande y se levanta, creando una concavidad en el centro (Boke 1964,
1968; Fig. 1). El desarrollo floral epígino-receptacular es muy poco común,
encontrándose en Cactaceae y Aizoaceae (Kaplan 1967, Weberling 1992).
Las flores tradicionalmente descritas como períginas o de ovario medio pueden tener
patrones de desarrollo epíginos ó hipóginos, por lo que el uso del estado de carácter
perígino es errado en términos de homología primaria (Soltis et al. 2003). En
Melastomataceae sin embargo, es común la designación de estados como ½ o ¼ ínfero
(c.f. Penneys & Judd. 2005 para Charyanthus en Melastomataceae) para codificar el
grado de fusión entre el hipanto y el ovario.

Marco teórico 7
Figura 1. Patrones de desarrollo floral temprano, modificado de Kuzoff et al. 2001.
No todos los ovarios que aparentemente son súperos en la flor desarrollada son
realmente homólogos, porque el plan de desarrollo puede ser diferente aunque el
resultado final se parezca (Kuzoff et al. 2001, Soltis & Hufford 2002, Litt & Stevenson
2003, Soltis et al. 2003); lo anterior, sugiere la necesidad de estudiar en más detalle la
ontogenia en aquellos grupos donde haya indicios de ausencia de homología. El plan de
desarrollo ha sido estudiado en detalle en algunas Myrtales como Lythrum salicaria y
Fuchsia hybrida (Onagraceae, Sattler 1973 citado en Soltis et al. 2003) y en
Vochysiaceae (Litt & Stevenson 2003). En Haemodoraceae (Simpson 1998),
Saxifragaceae (Mort & Soltis 1999, Kuzoff et al. 2001, Soltis & Hufford 2002, Soltis et al.
2003) además de un estudio detallado de los planes de desarrollo de los ápices florales,
se han interpretado los resultados ontogenéticos en contextos filogenéticos. En
Melastomatáceas no se encuentran interpretaciones ni descripciones de los planes de
desarrollo.

8 Anatomía y Morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
1.2. Tipo de fruto
La definición del carácter “tipo de fruto” es compleja, lo cual se evidencia en los variados
esfuerzos que se han hecho con el fin de estabilizar su nomenclatura; en la filosofía
botánica de Linnaeus (1751, citado por Spjut 1994) se describen ocho clases de fruto:
cápsulas, silicuas, legumbres, folículos, drupas, pomos, bayas y estróbilos; en trabajos
posteriores esta clasificación se ha ampliado llegándose a utilizar hasta 150 nombres
para tipos de frutos (Spjut 1994). Schleiden (1849) expresa en forma muy crítica las
dificultades en cuanto a un acuerdo en la nomenclatura de frutos.
La asignación errónea de homologías para los estados del carácter tipo de fruto ha
generado numerosos grupos artificiales al interior de taxones como Apiaceae (Plunkett et
al. 1996), Lamiales s.l., (Wagstaff & Olmstead 1997), Myrtaceae (Johnson & Briggs
1984), Ranunculaceae (Hoot 1991), Rhamnaceae (Richardson et al. 2000), Rosaceae
(Morgan et al. 1994) y Saxifragaceae (Soltis et al. 1990).
Para frutos derivados de un ovario ínfero se ha encontrado que la epigínia puede
constreñir la dehiscencia del fruto (Gustaffson & Albert 1999). Clausing et al. (2000)
mediante el estudio de 52 caracteres morfológicos encontraron que la indehiscencia del
fruto en Melastomataceae está asociada con la fusión del ovario y los tejidos del hipanto,
la persistencia de la placenta, la carencia de un endocarpo persistente; en tanto que la
dehiscencia se correlaciona con los caracteres opuestos y con la persistencia del
exocarpo.

1. 2. PLANTEAMIENTO DEL PROBLEMA
En Melastomataceae, tanto la posición del ovario como el tipo de fruto han sido utilizados
en la definición de tribus; así: Miconieae, Henrietteae y Blakeeae presentan flores de
ovario ínfero, donde hay fusión entre el ovario y el hipanto, que se desarrollan en frutos
carnosos indehiscentes; Bertolonieae, Melastomeae, Merianieae y Microlicieae presentan
flores con ovario total o casi totalmente súpero, sin fusión entre ovario e hipanto, que dan
lugar a frutos secos dehiscentes (Cogniaux 1891, Renner 1993, Michelangeli et al. 2004;
Penneys et al. 2010).
La distribución de los caracteres posición del ovario y tipo de fruto muestra una
correlación entre ovario súpero (libre) y fruto dehiscente, y entre ovario ínfero
(completamente fusionado al hipanto) y fruto indehiscente; esta correlación podría
explicarse porque la condición ínfera limitaría la dehiscencia del fruto; sin embargo, se
encuentran excepciones en los géneros neotropicales Allomaieta, Alloneuron,
Wurdastom (Michelangeli & Morales P., com. per. 2008) y Tateanthus (Maguire 1970) y
el género brasilero Physeterostemon (Goldenberg & Amorim 2006), los cuales presentan
flores con ovario ínfero fusionado al hipanto que originan frutos capsulares.
Los caracteres tipo de fruto y posición del ovario frecuentemente se recuperan en las
filogenias como caracteres homoplásicos, con múltiples surgimientos independientes y/o

10 Anatomía y morfología comparada del ovario y fruto a lo largo del desarrollo en
la tribu Cyphostyleae
reversiones (Johnson & Briggs 1984; Soltis et al. 1990; Morgan et al. 1994; Hoot 1991;
Wagstaff & Olmstead 1997; Gustafsson & Albert 1999; Richardson et al. 2000). En
Melastomataceae al optimizar estos dos caracteres sobre filogenias construidas a partir
de caracteres morfológicos y/o moleculares, generalmente aparecen como homoplásicos
(Michelangeli 2000; Clausing et al. 2000; Clausing & Renner 2001; Fritsch et al. 2004);
esto podría estar mostrando que los estados de al menos uno de estos dos caracteres
podrían estar mal definidos o que las semejanzas en las estructuras del ovario y/o del
fruto en algunos taxones de Melastomataceae no serian resultado de ancestría en
común.
Las similitudes morfológicas pueden originarse por ancestros en común, pero también
por convergencias, paralelismos o reversiones (Wake 1991); en estos tres últimos casos
la asignación de estados de carácter basada en hipótesis de homología primaria
equivocadas puede disminuir la resolución y las medidas de soporte de los clados y/o
generar topologías incorrectas. En el caso de similitudes morfológicas aparentes, es
posible, a través de estudios más detallados, redefinir los caracteres y sus estados; sin
embargo, en ocasiones además de las similitudes aparentes, usualmente generadas por
similitudes funcionales, se pueden presentar formas completamente idénticas cuando
fenómenos particulares del desarrollo son activados por diferentes estímulos o cuando
existen constricciones que generan unas pocas opciones de expresión morfológica
(Wake 1991).

11
En esta investigación se estudió el desarrollo del ovario y del fruto, desde los primeros
estadios de desarrollo hasta el fruto maduro, comparando algunas especies de
Cyphostyleae con especies de Blakea, Miconia, Meriania y Tibouchina. Estudiando estos
caracteres no solamente como tipo de fruto y posición del ovario en la flor madura, sino
disgregándolos en caracteres anatómicos y morfológicos individuales para poder
compararlos con más precisión con los demás grupos de Melastomataceae. El marco
filogenético para esta investigación corresponde a las filogenias obtenidas por
Michelangeli et al. (en preparación).


2. 3. OBJETIVOS
2.1 Objetivo General
Interpretar el desarrollo ontogenético del ovario desde sus primeros estadios
hasta la dehiscencia del fruto, en un contexto filogenético en especies de la tribu
Cyphostyleae y géneros afines.
2.2 Objetivos específicos
Estudiar el desarrollo del ovario, desde los primeros estadios del ápice floral hasta
la dehiscencia del fruto en la tribu Cyphostyleae y géneros afines.
Evaluar en Cyphostyleae y géneros afines la homología primaria de las estructuras
morfológicas del ovario y del hipanto a lo largo del proceso ontogenético.

4. METODOLOGÍA
7.1. Fase de campo
De acuerdo con la disponibilidad, se seleccionaron dos especies de la tribu Cyphostyleae
(Allomaieta hirsuta y A. zenufanasana), una especie de la tribu Blakeeae (Blakea
granatensis), una de Merianieae (Meriania nobilis), dos especies de Miconieae (Miconia
notabilis y M. squamulosa) y dos especies de la tribu Melastomeae (Tibouchina mollis y
T. urvilleana).
A lo largo de 18 semanas se hizo un seguimiento de los ápices florales de los árboles
seleccionados, con el fin de establecer las etapas en el desarrollo de las muestras. De
cada ejemplar se colectaron especímenes para depositar en herbario y órganos
reproductivos (botones florales de diferentes edades, flores, frutos inmaduros en
diferentes fases del desarrollo y frutos en dehiscencia) en alcohol al 70%. El material
examinado se enumera en las tablas 1-3.
Tabla 1. Material Examinado.
No. Ejemplares de Herbario
1 Allomaieta zenufanasana Lozano. Colombia. Antioquia. Anorí, vereda los Sauses,
sector cerca a la Reserva Proaves, 7.0039N - 75.0937W. 1800 m, 24- Ene-2009
(fl). H. David et al., 2830 (COL, MEDEL).

14 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Tabla 2. Material Examinado (Continuación).
No. Ejemplares de Herbario
2 Allomaieta hirsuta (Gleason) Lozano. Colombia. Antioquia. Amalfi, vereda
Guayabito, parte alta de la quebrada Caracolí, 6.5121 N - 75.0621 W, 1850 m, 25-
Ene-2009. H. David et al. 2833 (MEDEL).
3 Allomaieta hirsuta (Gleason) Lozano. Colombia. Antioquia. Amalfi, vereda
Guayabito, parte alta de la quebrada Caracoli, 6.5121 N - 75.0621 W, 1850 m, 25-
ene-2009. H. David et al. 2834 (MEDEL).
4 Allomaieta hirsuta (Gleason) Lozano. Colombia. Antioquia Amalfi, vereda
Guayabito, parte alta de la quebrada Caracoli, 6.5121 N - 75.0621 W, 1850, m. 25-
ene-2009. H. David et al. 2835 (MEDEL).
5 Allomaieta hirsuta (Gleason) Lozano. Colombia. Antioquia. Amalfi, vereda
Guayabito, parte alta de la quebrada Caracoli, 6.5121 N - 75.0621 W, 1850 m., 25-
ene-2009. H. David et al. 2836 (MEDEL).
6 Blakea granatensis Naudin. Colombia. Cundinamarca. Bogotá, Jardín Botánico
José Celestino Mutis, Calle 63 No. 68-95, jardín de melastomatáceas hacia la zona
media de la pérgola, 1-jun-2010, T. Sandino 02 (COL).
7 Meriania nobilis Triana. Colombia. Cundinamarca. Bogotá, Campus de la
Universidad Nacional de Colombia, cerca a la entrada de la 26, al frente del
departamento de Filosofía, 28-jul-2010, T. Sandino 04 (COL).

Metodología 15
Tabla 3. Material Examinado (Continuación).
No. Ejemplares de Herbario
8 Miconia notabilis Triana. Colombia. Cundinamarca. Bogotá, Jardín Botánico José
Celestino Mutis, Calle 63 No. 68-95, Jardín de melastomatáceas, cerca a la
entrada de la pérgola, 1-Jun-2010, T. Sandino 01 (COL).
9 Miconia squamulosa Triana. Colombia. Cundinamarca. Bogotá, cerros de Suba,
avenida calatrava, por la entrada oriental del bosquecito que queda en medio de
los conjuntos San Jorge etapa I y San Jorge etapa II, 27-jul-2010, T. Sandino 03
(COL).
10 Tibouchina mollis (Bonpl.) Cogn. Colombia. Cundinamarca. Bogotá. Universidad
Nacional de Colombia. Entre la plaza Ché, la cafetería León de Greiff y el
departamento de artes plásticas, 28-jul-2010, T. Sandino 05 (COL).
11 Tibouchina urvilleana (DC.) Cogn. Colombia. Cundinamarca. Bogotá. Universidad
Nacional de Colombia. En los jardines del lado norte del departamento de
Arquitectura, 28-jul-2010, T. Sandino 06 (COL).
7.3. Fase de laboratorio
El material colectado en alcohol al 70%, correspondiente a botones florales, flores y
frutos se estudió bajo estereoscopio y microscopio óptico. En cada caso, se realizaron
observaciones, ilustraciones y fotografías.

16 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
El material seleccionado para estudio histológico bajo microscopio óptico fue tratado con
el protocolo de Johansen (1940) con modificaciones así: Deshidratación en series de
etanol (70 al 100%), de etanol absoluto-xilol 100% (90:10, 70:30, 50:50, 30:70, 10:90)
cada una por cuatro horas y de xilol 100% por 24 horas; imbibición en parafina (60°C),
tres cambios por 24 horas cada uno; secciones en micrótomo de rotación y fijación en
láminas con ayuda de adhesivo de Haupt; desparafinado en dos series de xilol 100%,
tres minutos cada uno, series de xilol 100%-etanol absoluto (50:50) tres minutos, etanol
(absoluto, 95%, 70%, 50%) dos minutos cada uno; tinción con safranina por 15-18 horas,
lavado en agua destilada, deshidratación en series de etanol (50%, 70%, 95%) por un
minuto cada una, contracoloración con fast green por 12 minutos, deshidratación en dos
series de etanol absoluto por un minuto, etanol-xilol (70:30 y 50:50), dos minutos cada
una y dos de xilol 100% por tres minutos cada uno, adición de una gota de citoresina, de
cubreobjetos y secado a temperatura ambiente.
Con base en características morfológicas externas observadas a lo largo del seguimiento
de árboles en campo, así como en las observaciones generadas en la disección y
análisis bajo estereoscopio y microscopio de inflorescencias, botones, flores y frutos. Se
establecieron entre ocho y nueve estadios de acuerdo al grado de transformación del
fruto al madurar, así: (1) botón durante la organogénesis floral; (2-4) botón desde el final
de la organogénesis cuando se presentan los primordios de los lóculos y placentas en el
gineceo hasta el inicio de la antesis; (5) botón durante el proceso de antesis; (6) flor
abierta; (7) fruto recién fecundado o gineceo no fecundado, después de la caída del

Metodología 17
perianto; (8) fruto en proceso de maduración, se presentan cambios en el volumen,
consistencia y/o color; (9) fruto maduro.
Mediante disecciones longitudinales y transversales, observadas bajo el estereoscopio,
se identificó el punto de unión entre el hipanto y el ovario de botones florales, justo antes
de antesis. Se realizaron cortes seriales que fueron observados bajo el microscopio
óptico en donde se observó la vascularización y la disposición de los diferentes tejidos en
el receptáculo, el hipanto y el ovario. Posteriormente se hizo un seguimiento de estas
observaciones en botones y frutos, colectados en los diferentes estados del desarrollo.
Finalmente se realizaron interpretaciones, para cada especie, de la secuencia de
observaciones en el desarrollo de la flor, un mapeo de caracteres sobre la filogenia de
Penneys et al (2010) y una interpretación de la historia evolutiva del fruto en el grupo de
estudio teniendo en cuenta el “patrón de desarrollo” de las flores y los procesos de
desarrollo de los frutos.

18 Anatomía y morfología de desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
5. Resultados
5.1. Allomaieta hirsuta (Gleason) Lozano
5.1.1. Descripción morfológica
Cimas tirsoides, 3-5 cm de longitud (Fig. 2 A); flores pentámeras, pedicelos 1-20 mm de
longitud, teretes, color verde, densamente cubiertos por tricomas simples blanquecinos;
dos brácteas lineado-lanceoladas 4-5 mm de longitud y 1-2 mm de ancho, caedizas entre
el estadio cuatro y el siete, color verde, de ápice agudo, margen entero, cubiertas por
tricomas blancos simples; hipanto campanulado de 4 mm de longitud y 2 mm de ancho,
verde, densamente cubierto por tricomas blancos (Fig. 2 B); cáliz caliptrado con cinco
lóbulos verdes, de ápice agudo, caducos al momento de la antesis, 6-9 mm de longitud,
2-3 mm de ancho, densamente cubiertos por largos tricomas simples; pétalos cinco, 23-
34 mm de longitud, 18-25 mm de ancho, espatulados, ápice obcordado, color magenta
con una pequeña zona amarilla en la base; estambres cinco, anteras oblongas de 2-3
mm de longitud, profundamente caniculadas ventralmente con dehiscencia transversal,
conectivo no prolongado, filamentos acintados de 7-8 mm de longitud; estilo de 6-7 mm
de longitud, curvo con estigma ensanchado (Fig. 2 C).
5.1.2. Estructura del ovario y el hipanto antes de antesis
Ovario pentalocular, se encuentra totalmente fusionado al hipanto (1.5-2 mm; Figs. 2 B, 3
A), en la parte superior del ovario se presentan cinco espacios anterales separados entre
sí por cinco tabiques anterales (menores a 0.3 mm de longitud; Fig. 3 O, R). Antes de la
antesis las anteras se encuentran encurvadas y los ápices de éstas se localizan al

Resultados 19
interior de los espacios entre los tabiques que unen al hipanto con el ovario (= “bolsillos
anterales”; traducción del término usado por Renner 1993, 2001 –pockets-). A partir de la
antesis, el ángulo entre el conectivo y el filamento de los estambres cambia y las anteras
pasan a tener una posición erecta y extrorsa (Fig. 1 C). El ovario es externamente glabro
y presenta cinco lóculos de aproximadamente 2.7 mm, de placentación axilar, con
placentas intrusivas que soportan numerosos óvulos anátropos (Figs. 3 N-O, Q-R).
Figura 2. Allomaieta hirsuta, detalles de la morfología externa de la flor. A.
Inflorescencia, vista lateral; B. Corte longitudinal botón estadio 5; C. Vista superior de la
flor; at: antera, fl: filamento, hi: hipanto, lc: lóbulos del cáliz, ov: ovario, pe: pedicelo, pt:
pétalos, st: estilo. Escala A = 20 mm; B-C = 2 mm.
Vascularización del pedicelo y el receptáculo: El pedicelo presenta un cilindro
vascular bicolateral (cc; Figs. 3 B-C, E-F) y 2-3 haces perifloemáticos medulares (med;
Figs. 3 D, F). Al nivel del receptáculo salen los 10 haces mayores del hipanto (hh) hacia
la periferia y 10 haces hacia el centro, que se unen en pares y conforman los cinco haces
ventrales del ovario (hv), los cuales quedan enfrentados a los septos (Figs. 3 D, G-M).

20 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Vascularización del ovario+hipanto: La vascularización del ovario+hipanto está
conformada externamente, en su parte inferior por los 10 haces mayores del hipanto y en
su parte superior por los 10 mayores y por 15-20 haces más pequeños (“= haces
menores del hipanto”; hh’) que se forman a partir de los haces mayores del hipanto.
Estos haces más pequeños se dirigen a través de los septos de los lóculos y de los
tabiques anterales hacia el centro y conforman, junto con los haces ventrales del ovario
la vascularización del estilo y el estigma (Figs. 3 J, M-L, Q-R). Hacia el centro se
presentan los cinco haces ventrales del ovario que circula por todo el eje central del
ovario y una serie de haces más pequeños (“=haces vasculares de las placentas”; hvp)
dispuestos longitudinalmente, que se forman a partir de los haces ventrales del ovario
(Figs. 3 N, Q).
Vascularización del perianto: Por encima del nivel del ovario a partir de los 5 haces
mayores del hipanto, opuestos a los lóculos, se forman un haz vascular por cada
estambre (te) ventralmente y desde cada uno de los 10 haces mayores del hipanto se
forman dos laterales que conforman 10 haces intermedios a los mayores del hipanto.
Dese cada uno de los haces mayores del hipanto que quedan enfrentados a los septos y
de los dos laterales a estos se forman tres haces vasculares que conforman la
vascularización de cada pétalo (pc y 2 pl). Los sépalos presentan también una traza
central (sc) proveniente de los haces mayores del hipanto enfrentados a los lóculos y dos
laterales (sl) provenientes de los laterales de los haces mayores del hipanto enfrentados
a los lóculos (Figs. 3 P, S).
Vascularización del estilo y el estigma: Está conformada por 10 haces vasculares
pequeños que rodean al tejido de transmisión el cual presenta forma de estrella; cinco de
estos haces provienen de los haces ventrales del ovario y cinco provienen de los haces
dorsales (Figs. 3 N-S).

Resultados 21
6.1.3. Dehiscencia
No se contó con material para realizar las observaciones acerca del desarrollo de esta
especie. Sólo se observaron frutos dehiscentes, en donde se presenta pérdida de los
tejidos no vasculares del hipanto, de forma que el fruto dehiscente presenta una
apariencia similar a una canastilla.

22 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)

Resultados 23
Figura 3 (Viene de la página anterior). Allomaieta hirsuta, estructura y anatomía floral.
A. Diagrama de sección longitudinal del botón antes de antesis (Estadio 5), B-S.
Diagramas y fotografías de cortes transversales a diferentes niveles; las líneas
trasversales en A indican el nivel del diagram correspondiente, mientras que los niveles
de corte de las fotografías corresponden a los del diagrama ubicado arriba de cada
fotografía; ba: bolsillos en donde se encuentran las anteras en el botón, cc: cilindro
vascular central, med: haces medulares, hh: haces mayores del hipanto, he: haces
vasculares del estilo, hh’: haces secundarios del hipanto, hv: haces ventrales del ovario,
hvp: haces vasculares de las placentas, lo: lóculos del ovario, p: placentas, pc: trazas
centrales de los pétalos, pl: trazas laterales de los pétalos, sc: trazas vasculares
centrales de los sépalos, sl: trazas vasculares laterales de los sépalos, te: trazas
vasculares de los estambres, tq: tabiques anterales. Escala A = 1 mm; B-S = 500 µm.
Cortes en micrótomo, coloración con safranina fast-green.

24 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
6.2. Allomaieta zenufanasana Lozano
6.2.1. Descripción morfológica
Cimas tirsoides, 3-4.5 cm de longitud (Fig. 4 A); flores pentámeras; pedicelos 1-18 mm de
longitud, teretes, color verde, densamente cubiertos por tricomas simples blanquecinos;
dos brácteas lineado-lanceoladas 5-9 mm de longitud y 1-2 mm de ancho, sésiles,
caedizas en el momento del antesis, verdes, ápice agudo, margen entero, cubiertas por
tricomas blancos simples; hipanto campanulado de 2 mm de longitud y 2 mm de ancho,
verde, densamente cubierto por tricomas blancos (Fig. 4 B); cáliz caliptrado con cinco
lóbulos verdes, de ápice agudo, caducos al momento de la antesis, 4-7 mm de longitud, 1
mm de ancho, densamente cubiertos por tricomas simples; pétalos cinco, 10-13 mm de
longitud, 6-8 mm de ancho, blancos; estambres cinco, anteras oblongas de 2 mm de
longitud con dehiscencia por poro en posición ventral apical, conectivo no prolongado,
filamentos acintados de 4 mm de longitud; estilo de 3 mm de longitud, curvo con estigma
ensanchado.
Figura 4. Allomaieta zenufanasana, detalles de la morfología externa de la flor. A.
Inflorescencia, vista lateral frutos inmaduros; B. Corte longitudinal botón estadio 5; C.
vista superior del ovario; at: antera, br: brácteas, c: cáliz, hi: hipanto, ov: ovario, pe:
pedicelo, pt: pétalos, st: estilo. Escala A = 10 mm; B-C = 2 mm.

Resultados 25
6.2.2. Estructura del ovario y el hipanto antes de antesis
Ovario pentalocular, de placentación axial, fusionado al hipanto en 5/6 de su longitud
(1.5-2 mm), en la parte superior del ovario (1 mm) la fusión es parcial, presentándose
cinco bolsillos anterales separados entre sí por cinco tabiques anterales (menores a 0.5
mm), la zona apical del ovario (0.5-1 mm) es libre (Figs. 4 C, 5 J, M). Antes de la antesis
las anteras se encuentran encurvadas en los bolsillos anterales. A partir de la antesis, el
ángulo entre el conectivo y el filamento de los estambres cambia y las anteras pasan a
tener una posición erecta. Los lóculos del ovario tienen aproximadamente 1.5 mm y
placentas intrusivas que soportan numerosos óvulos anátropos (Figs. 5 L).
Vascularización del pedicelo, el receptáculo y el eje central del ovario: El pedicelo
presenta un cilindro vascular bicolateral (cc) y 2-7 haces perifloemáticos medulares (Figs.
5 B-C, E). Al nivel del receptáculo desde el cilindro central salen hacia la periferia los 10
haces mayores del hipanto (hh) y hacia el centro 10 haces, que se unen en pares y
conforman los cinco haces ventrales del ovario (hv), los cuales quedan enfrentados a los
septos (Figs. 5 C-D, F-G).
Vascularización del ovario+hipanto: La vascularización del eje del ovario está
conformada por los cinco haces ventrales del ovario y una serie de haces más pequeños
(“=haces vasculares de las placentas”; hvp) dispuestos longitudinalmente, que se forman
a partir de los haces ventrales del ovario y se dirigen hacia las placentas donde se
ramifican e inervan los óvulos presentes en cada uno de los lóculos carpelares (Figs. 5
H-I, K-L).
La vascularización de pared del ovario+hipanto está conformada externamente por los 10
haces mayores del hipanto e internamente por los cinco haces mediales y los 10 laterales
de los carpelos, los cuales se originan a partir de los haces mayores del hipanto (Figs. 5
H-I, K-L). Hacia la parte superior del ovario los haces dorsales pasan a través de los
tabiques anterales y del techo del ovario hacia el estilo (Figs. 5 J, M).

26 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)

Resultados 27
Figura 5 (Viene de la página anterior). Allomaieta zenufanasana, estructura y anatomía
floral. A. Diagrama de sección longitudinal del botón antes de antesis (Estadio 5), B-S.
Diagramas y fotografías de cortes transversales a diferentes niveles, mientras que los
niveles de corte de las fotografías corresponden a los del dibujo ubicado arriba de cada
fotografía; las líneas trasversales en A indican el nivel del diagrama correspondiente; cc:
cilindro vascular central, hc: haces corticales, hh: haces mayores del hipanto, hm: haces
carpelares mediales, ba: bolsillos anterales, lo: lóculos del ovario, o: óvulos, p:
placentas, pc: trazas centrales de los pétalos, pl: trazas laterales de los pétalos, sc:
trazas vasculares centrales de los sépalos, sl: trazas vasculares laterales de los sépalos,
te: trazas vasculares de los estambres, tq: tabiques anterales. Escala A = 2 mm; B-S =
500 µm. Cortes en micrótomo, coloración con safranina – fast green.
Vascularización del perianto: Por encima del nivel del ovario e iniciando el estilo, en el
hipanto se encuentran los 10 haces mayores del hipanto y los 20 haces menores del
hipanto, que surgen a partir de los mayores quedando intercalados con estos (Figs. 5 N,
Q). Posteriormente los haces menores del hipanto se unen en pares de tal manera que
finalmente, al nivel de la base de los pétalos, se presentan 20 haces vasculares en el
hipanto (Figs. 5 O, R). Los cinco pétalos presentan tres trazas foliares cada uno, de estas
trazas las centrales corresponden a los cinco haces mayores del hipanto que a nivel de

28 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
los lóculos del ovario quedan opuestos a los septos y las trazas laterales provienen la
ramificación de los haces menores del hipanto (Figs. 5 O, R). Por encima del nivel de los
pétalos se encuentran los cinco haces mayores del ovario que quedan opuesto a los
lóculos y los 10 haces menores del hipanto, estos 15 haces conforman las tres trazas
foliares de cada uno de los cinco sépalos (Figs. 5 S, P).
Vascularización del estilo y el estigma: Está conformada por 10 haces vasculares
pequeños que rodean al tejido de transmisión el cual presenta forma de estrella, cinco de
estos haces provienen de los ventrales del ovario y cinco provienen de los mediales y
laterales de los carpelos (Figs. 5 N, Q).
7.2.3. Desarrollo
Se establecieron ocho estadios del desarrollo: (1) Desde el inicio del ápice meristemático
hasta la conformación de botones redondeados de 0.5 mm de longitud, en los que se da
la organogénesis floral, presenta bauplan de tipo epigineo apendicular, en donde el ápice
floral inicialmente cóncavo, en el momento de la emergencia de los primordios de los
sépalos se hace convexo y permanece así durante todo el desarrollo; así mismo el
gineceo es totalmente ínfero desde su inicio, quedando los lóculos del ovario inmersos en
los tejidos del hipanto durante todo el desarrollo (Figs. 6 A-B); (2) Botones de 0.5 a 2 mm,
más alargados longitudinalmente y con crecimiento principalmente del ovario (Figs. 6 C-
G); (3) Botones de 2 a 6 mm de longitud, externamente se observa una diferenciación
entre la zona del ovario y la del perianto en el botón, hay un incremento en el tamaño
relativo de las anteras y el estilo (Fig. 6 H); (4) Botones de 6 a 9 mm de longitud, a
diferencia del estadio anterior la zona del perianto es de mayor tamaño que la del ovario
(Fig. 6 I); (5) Antesis de la flor, incluye la apertura y caída del cáliz y la exposición de los
pétalos y las antera (Figs. 6 J-K); (6) Flor abierta hasta la caída del perianto (con o sin
fecundación); (7) Fruto (u ovario no fecundado, en este momento no se observa la
diferencia directamente) de color verde (Figs. 6 L-M); (8) Fruto seco, hacia el final de este

Resultados 29
estadio ocurre la dehiscencia, se rompen los tejidos no lignificados del hipanto y el fruto
adquiere una apariencia de canastilla (Fig. 6 N).
Cambios tisulares en el pedicelo y el receptáculo: Durante el desarrollo de la flor y el
fruto se observan cambios histológicos en los distintos tejidos de la flor-fruto. En el
pedicelo se presenta un incremento en el número promedio de capas celulares hacia el
inicio del desarrollo, desde 4.3 ± 0,71 hasta 8.3 ± 1,17, luego del estadio tres no hay
crecimiento significativo (Tabla 4). En la médula el incremento es desde 4.9 ± 0.64 capas
celulares en el estadio dos hasta 8.3 ± 1.17 capas de células en el estadio tres y 9.3 ±
0.64 en el estadio cinco. Mientras que en la corteza el número de capas celulares
aumenta desde 9.38 ± 1.3 hasta 12.88 ± 1.23 del estadio dos al tres; y luego se mantiene
más o menos constante hasta el estadio cinco (Tabla 4). En el receptáculo el aumento en
el número de capas es menos pronunciado que en el pedicelo. En la médula del
receptáculo el número promedio de capas celulares se incrementa desde 5.9 ± 1.4 en el
estadio uno hasta 10.3 ± 0.89 en el estadio cinco, y permanece más o menos constante
hasta fruto. En la corteza desde 7.3 ± 1.49 hasta 10.4 ± 1.51 en el estadio dos (Tabla 4).
En los tejidos parenquimáticos del pedicelo y el receptáculo se observaron células en
pares, en los tres primeros estadios, lo que constituye evidencia de división celular
reciente. El área celular promedio se incrementa a lo largo de todo el desarrollo tanto en
las células del pedicelo como en las del receptáculo (Tabla 4). Se presentan células no
lignificadas que sobretiñen de safranina a partir del estadio dos y cristales tipo drusa (Fig.
7 A-D). S presenta formación de esclereidas en la médula y la corteza, tanto en el
pedicelo como en el receptáculo; y formación de tejidos vasculares secundarios en el
pedicelo a partir del estadio cinco (Fig. 7 E-G).

30 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)

Resultados 31
Figura 6 (Página anterior) Allomaieta zenufanasana, estadios del desarrollo. A. Vista
externa estadio 1; B-C. Corte longitudinal, estadio 1 (corte a mano alzada, tinción con
tionina); D-F. Vista externa, estadio 2; G. Corte longitudinal, estadio 2; H. Vista externa,
estadio 3; I. Vista externa, estadio 4; J. Vista externa, estadio 5; K. Corte longitudinal,
estadio 5; L. Vista externa, estadio 7; M. Corte longitudinal, estadio 7; N. Vista externa,
estadio 8; at: anteras, br: brácteas, hi: hipanto, lc: lóbulos del cáliz, o: óvulos, ov:
ovario, pt: pétalos, st: estilo. Escala: A = 0.5 mm; B-N = 1 mm.
Figura 7. Allomaieta zenufanasana, desarrollo de tejidos en pedicelo y receptáculo.
Cortes transversales. A. De pedicelo de botón en estadio 2; B. Receptáculo en estadio 2;
C. Pedicelo de botón en estadio 3; D. Receptáculo en estadio 3; E. Receptáculo de
botón en estadio 5; F. Pedicelo en fruto en estadio 7; G. Receptáculo en fruto en estadio
7; cc: cilindro vascular central, med: haces medulares, hh: haces mayores del hipanto.
Escala: A-B = 100 µm; C-F = 500 µm. A-F. Coloración con safranina - fast green, cortes
en micrótomo.

32 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Tabla 4. Promedio de capas celulares y área promedio de las células en el pedicelo y el
receptáculo para los diferentes estadios de desarrollo en Allomaieta zenufanasana.
Estadio 1 2 3 5 7
Longitud (mm) 0-0.5 0.5-2 2-6 9-11
Fruto
Inmaduro
Número de capas celulares en
la médula del pedicelo.
4.3 ±
0,71 4.9 ± 0.64
8.3 ±
1.17
9.3 ±
0.64 -
Área promedio células médula,
pedicelo (µm²). -
289.6 ±
140.32
270.0 ±
147.22
454.1 ±
277.97 -
Número de capas celulares en
la corteza del pedicelo.
9.1 ±
1.36 9.4 ± 1.38
12.9 ±
1.23
12.8 ±
1.28 -
Área promedio células corteza,
pedicelo (µm²). -
164.4 ±
133.15
235.6 ±
93.87
412.3 ±
200.20 -
Número de capas celulares en
la médula del receptáculo.
5.9 ±
1.36
8.6 ±
0.92
8.1 ±
1.33
10.3 ±
0.89 9.6 ± 1.51
Área promedio células médula,
receptáculo (µm²). -
52.3 ±
26.15
287.2 ±
115.42
414.8 ±
196.75
490.9 ±
254.50
Número de capas de células en
la corteza del receptáculo.
7.3 ±
1.49
10.4 ±
1.51
11.8 ±
1.49
11.3 ±
1.58 11.5 ± 1.85
Área promedio células corteza,
receptáculo (µm²). -
58.8 ±
49.61
260.2 ±
120.01
365.7 ±
158.08
598.9 ±
346.03
Cambios tisulares en el ovario+hipanto: En la pared externa y en los septos del
ovario, el número promedio de capas celulares se mantiene constante a lo largo de todo
el desarrollo (Tabla 5); mientras que en el eje central del ovario se presenta un
incremento en el número promedio de capas celulares entre los estadios uno y cinco
desde 10.3 ± 0.89 hasta 15.8 ± 1.75 (Tabla 5). El área promedio celular se incrementa a

Resultados 33
lo largo de todo el desarrollo, principalmente entre los estadios uno a tres y cinco a siete
(Tabla 5). Al igual que en el pedicelo y el receptáculo, se observan células en pares,
células que sobretiñen con safranina entre los estadios dos y cuatro (Fig. 8 A-F);
formación de esclereidas dispersas en el mesocarpo y aumento en el grado de
lignificación de los haces mayores del hipanto y los ventrales del ovario a partir del
estadio siete (Fig. 8 G-H). La epidermis interna de los lóculos (endocarpo) se lignifica a
partir del estadio siete (Fig. 8 G-H). En el hipanto el número promedio de capas celulares
se mantiene más o menos constante a lo largo de todo el desarrollo (Tabla 5).
Finalmente se tiene que, el incremento en volumen que se observa entre la flor en
antesis (estadio 6) y el fruto maduro (estadio 8) se da como consecuencia de un aumento
en el área promedio de las células de la pared del fruto, entre un 100-200%; y de los
septos del ovario de alrededor de un 210% (Tabla 5).
Figura 8. Allomaieta zenufanasana, cortes transversales a nivel del ovario y el hipanto;
A, B. De botón en estadio 2; C, D. De botón en estadio 3; E, F. De botón en estadio 4; G,
H. En fruto inmaduro estadio 7, zona media y distal del ovario respectivamente; at:
anteras, ba: bolsillos anterales, c: carpelos, hd: haces dorsales del ovario, hh: haces
vasculares mayores del hipanto, hi: hipanto, hv: haces ventrales del ovario, lo: lóculos
del ovario, o: óvulos, p: placentas. Escala A-B = 100 µm; C-H = 500 µm. Coloración con
safranina fast-green, cortes en micrótomo.
34 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
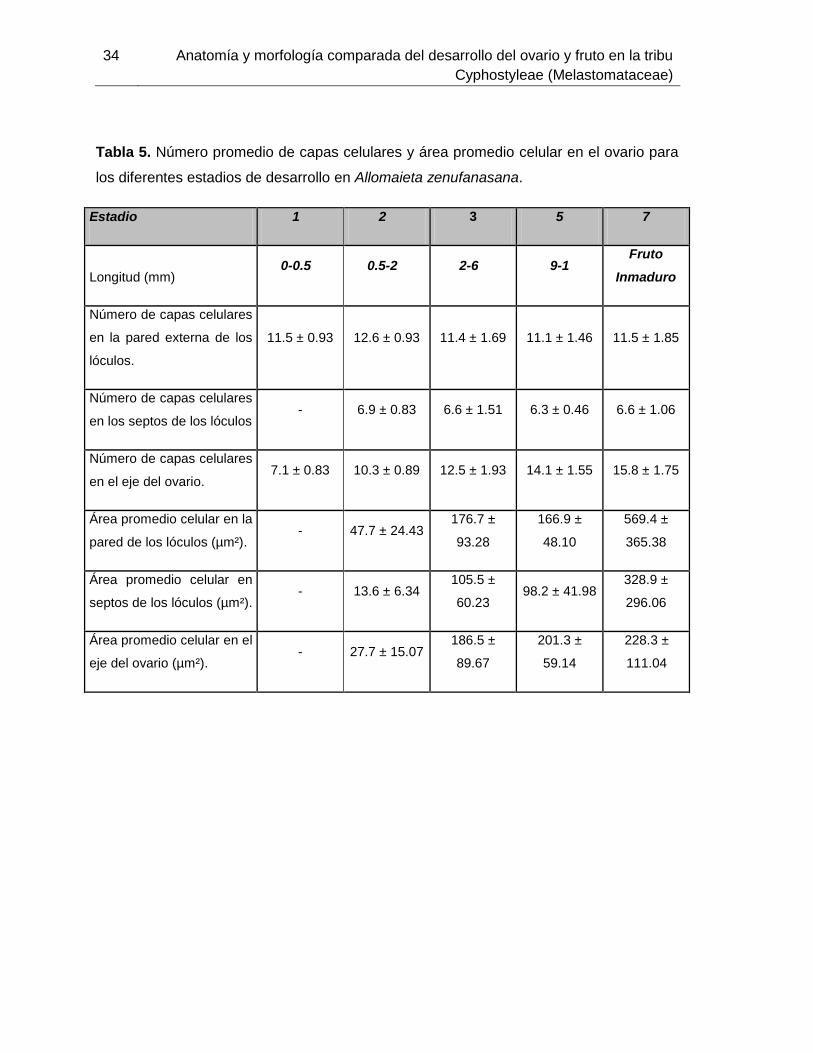
Tabla 5. Número promedio de capas celulares y área promedio celular en el ovario para
los diferentes estadios de desarrollo en Allomaieta zenufanasana.
Estadio 1 2 3 5 7
Longitud (mm) 0-0.5 0.5-2 2-6 9-1
Fruto
Inmaduro
Número de capas celulares
en la pared externa de los
lóculos.
11.5 ± 0.93 12.6 ± 0.93 11.4 ± 1.69 11.1 ± 1.46 11.5 ± 1.85
Número de capas celulares
en los septos de los lóculos - 6.9 ± 0.83 6.6 ± 1.51 6.3 ± 0.46 6.6 ± 1.06
Número de capas celulares
en el eje del ovario. 7.1 ± 0.83 10.3 ± 0.89 12.5 ± 1.93 14.1 ± 1.55 15.8 ± 1.75
Área promedio celular en la
pared de los lóculos (µm²). - 47.7 ± 24.43
176.7 ±
93.28
166.9 ±
48.10
569.4 ±
365.38
Área promedio celular en
septos de los lóculos (µm²). - 13.6 ± 6.34
105.5 ±
60.23 98.2 ± 41.98
328.9 ±
296.06
Área promedio celular en el
eje del ovario (µm²). - 27.7 ± 15.07
186.5 ±
89.67
201.3 ±
59.14
228.3 ±
111.04

Resultados 35
Figura 9. Allomaieta zenufanasana, fruto dehiscente. A. Vista lateral del fruto; B. Vista
superior de los carpelos; C. Vista lateral del fruto sin los carpelos; c: carpelos, hh: haces
mayores del hipanto, hv: haces ventrales del ovario; Escala = 1 mm.
5.2.4. Dehiscencia
La dehiscencia es apical loculicida, las paredes del ovario se rompen por la sutura medial
de cada lóculo, inicialmente por el ápice y luego a lo largo de toda su longitud; los septos
se separan del eje central del ovario. Por otra parte, los tejidos no lignificados de la pared
del fruto, que corresponde a la fusión de los tejidos del ovario y el hipanto, se rompen a
medida que se seca el fruto, quedando sólo los haces mayores del hipanto, los haces
ventrales del ovario, que conforman una columna en el centro del fruto; y las capas
internas del fruto, que incluyen a la epidermis interna lignificada (Fig. 6 N, 8 G-H, 9).

Resultados 36
5.3. Blakea granatensis Naud.
5.3.1. Descripción morfológica
Inflorescencias axilares unifloras, 1-(2) yemas seriales por axila; flores bisexuales,
actinomorfas, hexámeras; pedicelos de 4-6.5 mm de longitud, teretes, angostos en la
base, erectos, largos, verde-rojizos con tricomas dispersos, largos y simples; dos
brácteas de 24-26 mm de longitud, 16-22 mm de ancho; dos bractéolas de 22-25 mm de
longitud, 13-19 mm de ancho, ampliamente ovadas, caedizas en fruto inmaduro,
cóncavas, libres, rojizas a verde-rojizas, ápice largo-acuminado, con un nervio medio,
margen entera, con tricomas largos y simples, dispersos; hipanto de 3-4 mm de longitud,
6-7 mm de ancho, ampliamente campanulado, color blanco-verdoso, densamente
cubierto por tricomas simples pardo-rojizos; cáliz con 6 lóbulos de 4-3 mm de longitud y
7-8 mm de ancho, ampliamente ovados, rojizos, con base blanco-verdosa y un mucrón
en el centro del ápice, cubiertos por tricomas simples, perennes hasta fruto; seis pétalos
libres, dexocontortos en botón, 31-34 mm de de longitud, 26-30 mm de ancho,
espatulados a obovados, ápice cuspidado, blancos con tintes rosados en el ápice;
estambres 12, formando un semicircunferencia de más de 180º, filamentos de 10-13 mm
de longitud, libres, teretes, ligeramente arqueados, blancos y glabros, anteras basifijas,
con dos tecas, 7-8 mm de longitud, connadas lateralmente, blancas, con un conectivo de
9-10 mm de longitud, basal-dorsal, carnoso, triangular, elevado, termina en una
proyección aguda, amarillenta, la liberación del polen se da por dos pequeños poros
redondos en el ápice de la antera; estilo de 20-22 mm de longitud, curvo, blanquecino,
con tricomas simples; estigma angostamente cuspidado (Fig. 10 A).

Resultados 37
Figura 10. Blakea granatensis, detalles de la morfología externa de la flor. A. Vista
superior; B. Corte longitudinal de botón; at: antera, lc: lóbulos del cáliz, o: óvulos, ov:
ovario, pt: pétalos, st: estilo. Escala A = 20 mm; B = 5 mm.
5.3.2. Estructura del ovario y el hipanto antes de antesis
El ovario presenta seis lóculos y se encuentra totalmente unido al hipanto, los lóculos
tienen una longitud aproximada de 2 mm (Figs. 10 B, 11 A). Por encima del nivel del
ovario se presentan 12 tabiques anterales de 0.1-0.3 mm de longitud (Figs. 11 J, N). En
el botón, las anteras se encuentran arqueadas sobre el ovario, y los ápices de éstas se
encuentran en los espacios que quedan entre los tabiques anterales. A partir de la
antesis, el ángulo entre el conectivo y el filamento de los de los estambres cambia y las
anteras pasan a tener una posición erecta y extrorsa (Figs. 10 A-B). El ovario es
externamente glabro, de placentación axial, con placentas intrusivas que soportan
numerosos óvulos anátropos (Figs. 10 B, 11A).
Vascularización del pedicelo, receptáculo y del eje central del ovario: El pedicelo
presenta un cilindro vascular bicolateral continuo (cc) y numerosos haces perifloemáticos
medulares (med; Figs. 11 B, F). A diferencia de las otras especies observadas, en
Blakea encontramos dos brácteas y dos bractéolas justo por debajo del receptáculo

38 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
floral, lo que conlleva a que la vascularización en el pedicelo sea diferente,
presentándose tres trazas foliares (tb) que se dirigen a cada una de las brácteas y a
cada una de las bractéolas (Figs. 11 C-E, G-H). Inmediatamente por encima de la
separación de las trazas foliares de las bractéolas el tejido vascular se organiza de nuevo
en un cilindro vascular continuo del cual salen los 12 haces mayores del hipanto (hh)
hacia la periferia del receptáculo (Figs. 11 E, I-J, M ), los cuales se separan del cilindro
central en dos grupos intercalados de seis haces vasculares cada uno; primero se
separan los seis haces que a lo largo del botón floral quedan opuestos a los lóculos y en
seguida el otro grupo de seis haces que quedan opuestos a los septos del ovario. El
tejido restante del cilindro central, junto con los haces medulares, se anastomosa (Figs.
11 K, N) y rápidamente se divide en 12 haces (=“haces menores del hipanto”; hh’), que
quedan intercalados con los 12 haces mayores del hipanto, seis haces mediales de los
carpelos (hm), 12 haces laterales de los carpelos (hl), haces dorsales del ovario y un
gran número de haces ventrales (aproximadamente 84; hv) que recorren el eje central
del ovario y conforman la vascularización de las placentas y los óvulos (Figs. 11 L, O).
Vascularización del hipanto+ovario y el perianto: La vascularización del
hipanto+ovario está conformada por los 12 haces mayores intercalados con los 12
menores del hipanto, ubicados hacia la zona externa del hipanto; y los seis haces
mediales intercalados con los seis laterales de los carpelos, ubicados en la zona cercana
a los lóculos del ovario (Figs. 11 P, S). Por encima del nivel del ovario, desde el lado
adaxial de los haces mayores del hipanto surge una traza foliar por cada uno de los 12
estambres (te) y desde los lados, dos haces que se anastomosan formando los haces
menores del hipanto (Figs. 11 W-X, Za-Zb). Desde los seis haces mayores del hipanto
que a nivel del ovario quedan enfrentados a los septos, y desde 12 haces que provienen
de los 12 haces menores del hipanto, se forman tres trazas por cada uno de los seis
pétalos (tp; Figs. 11 Y, Zc). El tejido excedente (los seis haces mayores del hipanto que a
nivel del ovario quedan enfrentados a los lóculos y los 12 haces menores del hipanto) se
fracciona, se anastomosa y finalmente conforma tres trazas por cada uno de los seis
sépalos.

Resultados 39

40 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Figura 11 (Viene de la página anterior). Blakea granatensis, estructura y anatomía
floral. A. Diagrama de sección longitudinal del botón antes de antesis (Estadio 5), B-Zc.
Diagramas y fotografías de cortes transversales a diferentes niveles; las líneas
trasversales en A indican el nivel del diagram correspondiente, mientras que los niveles
de corte de las fotografías corresponden a los del diagrama ubicado arriba de cada
fotografía; at: anteras, ba: bolsillos anterales, br: bractéolas, cc: cilindro vascular central,
he: haces vasculares del estilo, hh: haces mayores del hipanto; hh’: haces menores del
hipanto, hl: haces laterales de los carpelos, hm: haces mediales de los carpelos, hv:
haces ventrales del ovario, lo: lóculos del ovario, o: óvulos, p: placentas, tb: trazas
foliares de las brácteas, te: trazas de los estambres, tp: trazas de los pétalos tq:
tabiques anterales, tt: tejido de transmisión. Escala A = 2 mm; B-Zc = 500 µm.
Coloración con safranina – fast green, cortes en micrótomo.

Resultados 41
Vascularización del estilo y el estigma: En el ápice del ovario, los haces mediales y
laterales de los carpelos se dirigen por el techo del ovario hacia el centro de éste
organizándose en 12 haces vasculares que rodean al tejido de transmisión en la zona
inferior del estilo. Hacia la parte superior del estilo los 12 haces vasculares se fraccionan
en 18 haces y forman un hexágono alrededor del tejido de transmisión (he; Figs. 11 Q-R,
T-Zc).
6.3.3. Desarrollo
Se establecieron ocho estadios del desarrollo: (1) menos de 7 mm de longitud y 4 mm de
ancho, las cuatro brácteas cubren totalmente al botón y se encuentran conniventes a
este, por lo que las dimensiones descritas en realidad se refieren al tamaño de las
brácteas, el botón propiamente dicho mide hasta 1 mm de longitud y 1.5 mm de ancho,
en este estadio ocurre la organogénesis floral (Figs. 12 A-C); el bauplan es de tipo
epigineo apendicular, de forma que el ápice floral inicialmente cóncavo, en el momento
de la emergencia de los primordios de los sépalos, se hace convexo permaneciendo así
durante todo el desarrollo; el gineceo es totalmente ínfero desde su inicio, encontrándose
los lóculos del ovario inmersos en los tejidos del hipanto durante todo el desarrollo floral;
(2) 7-13 mm de longitud y 5-6 mm de ancho, incluye las brácteas; el botón propiamente
dicho mide de 1 a 4 mm de longitud y 1.5-4 mm de ancho, se encuentra totalmente
cubiertos por las brácteas, aunque ya no tan cercanamente unidas a estas (Figs. 12 D-F);
(3) botones de 13-20 mm de longitud y 7-15 mm de ancho; el botón propiamente dicho
mide 4-7 mm de longitud y 4-8 mm de ancho, hacia la base su forma es más redondeada
que en los estadios anteriores; (4) botones casi esféricos de 20-25 mm de longitud y 16-
20 mm de ancho; sin las brácteas el botón mide 7-20 mm de longitud y 8-20 mm de
ancho (Figs. 11 G-J); (5) flor en antesis, se presenta la apertura de los pétalos y la
exposición de las anteras (Fig. 11 K); (6) flor abierta hasta la caída del perianto; (7) flor
sin perianto (con o sin fecundación; Fig. 12 L); (8) fruto inmaduro (verde; Fig. 12 M); (9)
fruto maduro (rojo; Fig.11 N).

42 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)

Resultados 43
Figura 12 (Página anterior). Blakea granatensis, estadios del desarrollo. A. Vista
externa, estadio 1; B-C. Corte longitudinal, estadio 1; D. Vista externa, estadio 2; E-F.
Corte longitudinal, estadio 2; G-I. Vista externa, estadio 4, con brácteas y bractéolas, sólo
con bractéolas; y sin brácteas ni bractéolas, respectivamente; J. Corte longitudinal,
estadio 4; K. Vista externa, estadio 5; L. Vista externa, estadio 7; M. Vista externa,
estadio 8; N. Vista externa, estadio 9; at: antera, br: brácteas y bractéolas, fl: filamento,
hi: hipanto, pe: pedicelo, lc: lóbulos del cáliz, ov: ovario, p: placentas, pt: pétalos, st:
estilo. Escala A-C = 0.5 mm; D-F = 1 mm; G-N = 5 mm.
Cambios tisulares en el pedicelo y el receptáculo: En la médula del pedicelo el
número promedio de capas de células se incrementa a lo largo del desarrollo,
principalmente entre los estadios dos y tres (desde 27,6 ± 1.6 hasta 36.2 ± 1.6; Tabla 6).
En la corteza el aumento en el número promedio de capas celulares se da principalmente
entre botones de los estadios uno y dos (desde 12.8 ± 2.4 hasta 24.8 ± 3.6) y luego se
mantiene casi continúo hasta el estadio ocho (28.0 ± 5.5). En la médula del pedicelo y el
receptáculo se observan células en pares entre los estadios uno y siete, mientras que en
las cortezas sólo entre los estadios uno y tres.
En el receptáculo el aumento en el número promedio de capas celulares es gradual a lo
largo de todo el desarrollo. En la médula desde 23.1 ± 2.1 (estadio 1) hasta 67.3 ± 4.4
(estadio 7) y en la corteza 17.0 ± 1.8 (estadio 1) hasta 43.8 ± 2.5 (estadio 8). En cuanto al
área celular promedio se presenta un incremento gradual tanto en el pedicelo como en el
receptáculo, entre los estadios uno y ocho (Tabla 6). Formación de braquioesclereidas,
tanto en el pedicelo como en el receptáculo a partir del estadio cinco y en la médula a
partir del estadio siete, inicialmente presentan mayor tamaño que las células que las
circundan; para el estadio nueve las células parenquimáticas pueden ser de mayor
tamaño que las esclereidas. Tanto en el receptáculo como en el pedicelo se observan
cristales tipo drusa desde el estadio uno siendo estos más abundantes a partir del
estadio seis, no hay cambios en el tamaño de espacios intercelulares ni tejidos
sobreteñidos en ningún estadio del desarrollo (Fig. 13).

44 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Tabla 6. Número promedio de capas celulares y área promedio de las células en el
pedicelo y el receptáculo para los diferentes estadios de desarrollo de Blakea
granatensis.
Estadio
1 2 3 5 7 8
Longitud 1-7 mm 7-13 mm 13-20 mm
Botón en
antesis
fruto
inmaduro
fruto
maduro
Número de capas celulares en
la médula del pedicelo.
25.7 ±
4.7
27.6 ±
1.6
36.2 ±
1.6
33.8 ±
4.71
58.3 ±
4.73
55.9 ±
6.96
Número de capas celulares en
la corteza del pedicelo.
12.8 ±
2.4
24.8 ±
3.6
22.3 ±
2.2
25.3 ±
3.40
21.9 ±
2.8 28.0 ± 5.5
Número de capas celulares en
la médula receptáculo
23.1 ±
2.1
30.6 ±
2.2
36.0 ±
2.9
41.7 ±
3.0
61.0 ±
7.2 67.3 ± 4.4
Número de capas celulares en
la corteza del receptáculo
17.0 ±
1.8
21.3 ±
2.4
36.8 ±
6.1
37.6 ±
3.46
39.3 ±
14.9 43.8 ± 2.5
Área promedio celular en
médula del pedicelo.
76.7 ±
70.3
92.5 ±
77.1
121.6 ±
115.4
323.6 ±
221.38
959.7 ±
558.4
915.5 ±
513.4
Área promedio celular en
corteza del pedicelo.
121.6 ±
111.7
128.7 ±
101.3
131.7 ±
112.8
186.4 ±
135.58
513.0 ±
440.1
1001.4 ±
906.0
Área promedio celular en
médula del receptáculo
23.1 ±
10.02
44.7 ±
28.9
64.9 ±
74.15
132.4 ±
93.01
287.2 ±
190.35
319.1 ±
265.67
Área promedio celular en
corteza del receptáculo
56.6 ±
47.24
76.4 ±
70.51
151.8 ±
144.82
102.1 ±
105.42
432.0 ±
424.51
412.4 ±
337.81
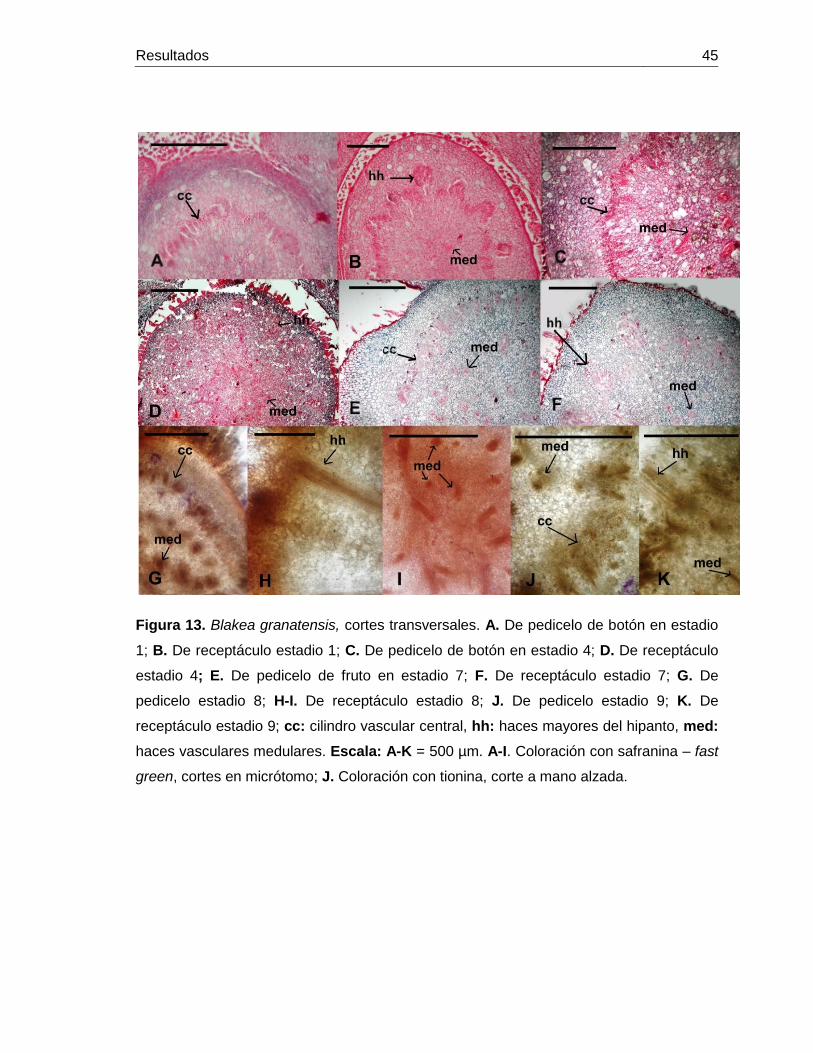
Resultados 45
Figura 13. Blakea granatensis, cortes transversales. A. De pedicelo de botón en estadio
1; B. De receptáculo estadio 1; C. De pedicelo de botón en estadio 4; D. De receptáculo
estadio 4; E. De pedicelo de fruto en estadio 7; F. De receptáculo estadio 7; G. De
pedicelo estadio 8; H-I. De receptáculo estadio 8; J. De pedicelo estadio 9; K. De
receptáculo estadio 9; cc: cilindro vascular central, hh: haces mayores del hipanto, med:
haces vasculares medulares. Escala: A-K = 500 µm. A-I. Coloración con safranina – fast
green, cortes en micrótomo; J. Coloración con tionina, corte a mano alzada.

46 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Tabla 7. Número promedio de capas celulares y área promedio de las células en los
tejidos del ovario+hipanto y en los tabiques anterales en Blakea granatensis.
Estadio 1 2 3 5 7 8
Longitud 1-7 mm 7-13 mm 13-20 mm
Botón en
antesis
fruto
inmaduro
fruto
maduro
Número de capas celulares en
los septos del ovario. 9.3 ± 1.5 8.6 ± 1.0 8.8 ± 1.3 9.0 ± 1.41 12.0 ± 3.2 -
Número de capas celulares en
la pared del ovario+hipanto. 27.8 ± 4.3 30.8 ± 1.5 33.3 ± 5.0
30.8 ±
5.85 29.1 ± 3.7 33.0 ± 3.8
Número de capas celulares en
el eje del ovario.
13.8 ±
2.49
13.4 ±
1.30
21.1 ±
3.83
44.5 ±
4.14
71.6 ±
4.41 -
Número de capas celulares en
los tabiques anterales. 6.0 ± 2.07 7.1 ± 1.64 8.1 ± 1.90 - - -
Área promedio células pared
lóculos (µm²)
71.4 ±
43.51
120.6 ±
114.22
267.4 ±
249.14
404.1 ±
341.56
346.1 ±
343.78
1364.6 ±
1044.27
Área promedio células septos
lóculos (µm²)
17.3 ±
10.02
38.5 ±
26.44
127.6 ±
83.30
120.0 ±
71.12
117.8 ±
87.18
314.2 ±
72.71
Área promedio células en el
eje (µm²)
33.2 ±
20.81
92.5 ±
27.86
142.8 ±
127.44
142.7 ±
98.32
134.7 ±
132.27
12410.4 ±
7414.53
Área promedio células en
tabiques anterales (µm²)
13.18 ±
6.71
13.95 ±
9.37
66.27 ±
72.67 - - -
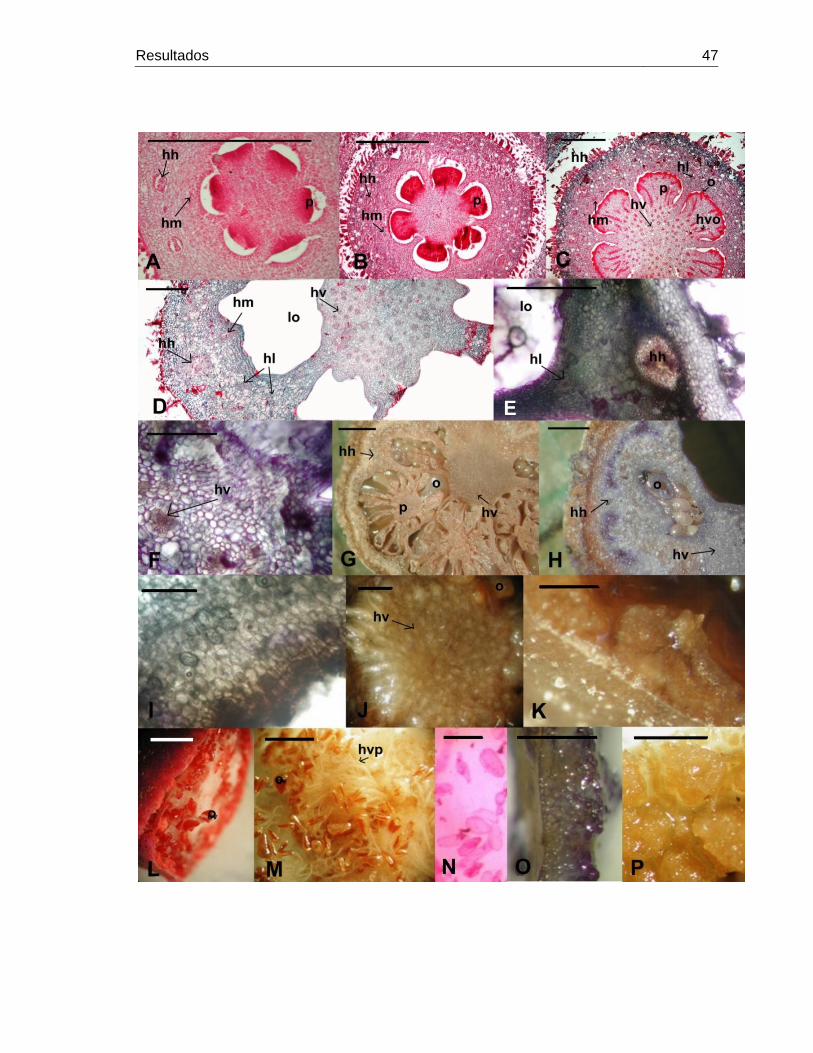
Resultados 47

48 Anatomía y morfología comparada del desarrollo del ovario y fruto en la tribu
Cyphostyleae (Melastomataceae)
Figura 14 (Página anterior) Blakea granatensis. Cortes transversales a nivel del
ovario+hipanto. A. En botón, estadio 1; B. En botón, estadio 2; C. En botón, estadio 3; D.
En flor, estadio 7; E-F. En fruto, estadio 8; detalles de la pared y del eje del fruto,
respectivamente; G-H. En fruto, estadio 8; I. En fruto, estadio 9, a nivel inferior de la
pared del fruto; J-K. En fruto, estadio 9 a nivel superior, detalles del eje y la pared del
fruto, respectivamente; del fruto respectivamente; L. Vista tangencial, estadio 9; M-N.
Tejido del eje central del ovario en fruto, estadio 9; O. Corte transversal de la pared de
fruto, estadio 9; P. Vista de la cara interna del ovario+hipanto en fruto, estadio 9; hh:
haces vasculares mayores del hipanto, hm: haces vasculares mediales de los carpelos,
p: placentas, hl: haces vasculares laterales de los carpelos, o: óvulos, hvp: haces
vasculares de las placentas, hv: haces ventrales del ovario, lo: lóculos del ovario. Escala
A-F, I, K = 500 µm; G-H, J, O, P = 1 mm; L-M = 3 mm; N = 200 µm. A-D. Coloración con
safranina-fast green, cortes en micrótomo; E-F, H-I, N-O. Coloración con tionina, cortes a
mano alzada. G, J-M, P. Cortes a mano alzada, sin tinción.
5.3.3.4. Cambios tisulares en el ovario+hipanto: Se presenta un aumento en el
número promedio de capas de células en el eje del ovario, desde 13.75 ± 2.49 (botones
de 7 mm) hasta 71.63 ± 4.41 (estadio 7) y en los septos de los lóculos desde 9.3 ± 1.5
(estadio 2) hasta 12.0 ± 3.2 (Tabla 7). En el hipanto, que incluye la pared del ovario, el
número promedio de capas celulares se mantiene constante a lo largo de todo el
desarrollo (Tabla 7). En estadio uno, el eje del ovario presenta tejido meristemático y a
partir del estadio 2 se observan células en pares hasta el estadio siete. En la pared del
ovario+hipanto se observan células en pares desde el estadio uno hasta el siete.
En cuanto al área promedio celular, hay un incremento en todas las regiones del ovario
observadas que se hace cada vez más acentuado a medida que madura el ovario (Tabla
7). La pared del ovario+hipanto presenta esclereidas dispersas a partir del estadio cinco,
que hacia el estadio siete conforman, en el mesocarpo, una zona más o menos continua
de entre dos y diez capas de células que le da rigidez al la región externa del mesocarpo;
en el eje del ovario se presentan esclereidas dispersas a partir del estadio siete. En la

Resultados 49
pared del hipanto+ovario se observan cristales tipo drusa a partir del estadio uno,
mientras que en el eje del ovario a partir del estadio tres (Fig. 14); células del eje del
ovario, las placentas, el endocarpo y las 5-7 primeras capas del mesocarpo, entre los
estadios siete y nueve pierden coherencia por disolución de la lámela media (Fig. 14 J, L-
N).
Finalmente se tiene que el incremento en el volumen del fruto se da por un aumento en
área promedio celular, en la pared del fruto y en los septos del ovario de entre 150 y
250%, entre los estadios siete a ocho; y en el eje central del ovario donde el área
promedio celular se incrementa alrededor de 100 veces entre los estadio siete y ocho
(Tabla 7).
5.3.4. Dehiscencia
Luego de la caída del perianto, el fruto permanece en estadio siete por entre seis y ocho
semanas, en estadio ocho por tres semanas y en estadio nueve por aproximadamente
una semana hasta que es consumido o se desprende de la planta.
En estadio 9, el tejido parenquimático de las placentas, el eje central del ovario y la parte
interna de la pared del ovario+hipanto se disgrega, probablemente por disolución de las
lamelas medias, en tanto que sus células aumentan de tamaño y se llenan de líquidos,
de tal forma que el interior del fruto en su madurez adquiere una consistencia de fluido
espeso, mientras que la zona más externa de la pared del ovario+hipanto presenta una
consistencia coriácea, dada por la presencia de abundantes esclereidas en la pared del
fruto (Fig. 14 E-P). En el Jardín Botánico José Celestino Mutis se observaron individuos
de Turdus fuscater alimentándose del fruto, lo perforaban y se comían la pulpa dejando la
mayor parte de la pared del fruto intacta en la planta.