
Articulo Fermentados
10
3 3 5 2 Research Article R ec ei ve d: 1 6 N ov em ber 20 12 R ev is ed: 9 Apr il 20 13 Ac cept ed a rt ic le p ub l is hed: 19 Apr il 20 13 P ub li sh ed online in Wi ley Onli n e L ib ra r y: 17 Jun e 2 01 3 (wileyonlinelibrary.com) DOI 10.1002/jsfa.6184 App lic abi lit y of Lact obaci llus plant arum IMDO 788 as a starter culture to control vegetable fermentations Dor rit Wou ter s, a Silvia Grosu -Tudo r, b Med ana Zamfir b and Luc De Vuyst a∗ Abstract BACKGROUND:Fermentat ion of vegetables and fruits is a tradit ional preser vat iontechni que, e.g. in Eastern Eur ope. Although usually spontaneous fermentation processes are applied, the addition of lactic acid bacteria (LAB) starter cultures could acc elerate proces sin g and improve the consis tency and qualit y of the end -pr oducts. RESULTS: The application of Lactobacillus plantarum IMDO 788 as a starter culture strain for cauliflower and mixed vegetable fermentations resulted in accelerated acidification as compared with the spontaneous fermentations. The strain dominated the background microbiota throughout the process, whereas the spontaneous fermentations were characterised by widely variable species diversity. During the spontaneous fermentations, almost all carbohydrates were converted into lactic acid, ethanol, mann itol and ace tic aci d, ind ica ting the par tic ipatio n of bot h het erofer mentat ive and homofer ment ati ve LAB spe cie s. During the starter cul ture-added fer mentations , residual carboh ydr ates were found and lac tic acid and eth anol were the mai n end-me tabolites.Vegetable- assoc iatedaromas, ethyl aceta te and isoamyl acetat e wereproduced durin g all fermen tation s. The high concentration of ethanol and the production of ethyl acetate and isoamyl acetate suggested the involvement of yeasts durin g all fermentation s. CONCLUSION: Lactobacillus plantarum IMDO 788was an ade quate starter cul tur e str ainfor vegeta blefermen tat ions, pre vai ling over endogenous LABcommuniti es. Fur the r opt imi sat ion of the starter cul ture formula tion is nec ess ary to avoid yeast growth. c 2013 Soc iet y of Chemic al Indust ry Keywords: vegetable fermentatio n; lactic acid bacteria; Lacto bacillus plantarum; starter cultures INTRODUCTION Ferment ed veg et abl es and fru its have been pro duc ed for tho usands of yea rs mainl y for pr ese rva tio n purpos es, as fre sh veg - etables and fruits are very perishable. 1,2 In addition, fermenta tion deli versend-pro duct s thatare appr eciat ed for thei r typic al senso ry properties. 3 Fermented vegetables and fruits of economic impor- tance include sauerkraut and its Asian variant kimchi, cucumbers and olives, although many fermentations performed on a small scale or at household level are attracting interest as well. 4 These small-scale fermentatio ns comprise a wide diversity of vegetables and fru its suc h as art ichoke s, 5 caper berr ies, 6 carrots, 7 – 11 cauliflowers, 11 – 13 cherries, 14 eggplants, 15 garlic, 16,17 ginger, 18,19 gre en bean s, 7 green tomatoes, 11,20 leeks, 21 mangoes, 19 marrows, 7 onions, 22 – 25 peppers, 26 pineapples 27 and turnips 28 and are often performe d in Eastern and Southern European countries. At present, many vegetable and fruit fermentation processes st i ll r el y on t he n at ur al ly oc cu r ri ng m ic r ob io ta of the fre sh raw ma ter ial s, among whi ch lac tic aci d bac ter ia (L AB) overgrow the undesirable communitie s of Enterobact eriaceae, Pseudomonad ace ae and yeasts, at least whe n appropria te fermentation conditions are set up. 3 This involves the addition of salt to the fresh vegetables and fruits (dry salting or brining), aft er which the mixture is pr ote cte d fro m light and oxygen and left to fer ment at tempera tur es bet ween 16 and 35 ◦ C. 2 However, spont aneous fermenta tion proc esses suff er from a lack of control, often resulting in end-products of inconsistent quality. 29 In thecase of chee se and fermente d sausa ge prod uctio n, these problems are avoided by the use of starter cultures. 29,30 Star ter culture s are micr obial prepar ation s of a larg e numb er of cells of one or more microbial strains, which are added to the raw material to acce lera te and cont rol the ferment ation process. 31 With regar d to fermented veg etables and fru its , however, several difficulties, e.g. the inability to inactivate the endo geno usmicrobiota by heattreatmentwithoutcausingdrastic texture changes, and the desired succession of different micro- organismsupon fermenta tion, somewhatimpede the introduction of starter cultures in these cases. 29,32 Nevertheless, an increasing number of studies concerning starter culture use for fermented ∗ Corre spond ingto:Luc De Vuyst , Resea rchGroup of Indus trialMicrobio logyand Food Biotechnology (IMDO), Department of Bioengineering Sciences, Faculty of Sciences and Bioengineering Sciences, Vrije Universiteit Brussel, Pleinlaan 2, B-105 0 Bruss els, Belgi um E-ma il: ldvuys [email protected] a Research Group of Industrial Microbiology and Food Biotechn ology (IMDO), DepartmentofBioengineeringSciences,FacultyofSciencesandBioengineering Scien ces, VrijeUniversi teitBrussel, Plein laan2, B-10 50 Brusse ls, Belg ium b Instit uteof Biolo gy Buch arest(IBB),Romania n Acade my,SplaiulIndepend ente i 296, 060031 Buch arest , Roman ia J Sci Food Agric 2013; 93: 3352 – 3361 www.soci.org c 2013 Society of Chemical Industry
-
Upload
gabriela-tellez-chavez -
Category
Documents
-
view
228 -
download
0
Transcript of Articulo Fermentados

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 1/10
Research Article
Received: 16 November 2012 Revised: 9 April 2013 Accepted article published: 19 April 2013 Published online in Wiley Online Library: 17 June 2013
(wileyonlinelibrary.com) DOI 10.1002/jsfa.6184
Applicability of Lactobacillus plantarum IMDO
788 as a starter culture to control vegetablefermentations
Dorrit Wouters,a Silvia Grosu-Tudor,b Medana Zamfirb and Luc De Vuysta∗
Abstract
BACKGROUND: Fermentation of vegetables and fruits is a traditional preservation technique, e.g. in Eastern Europe. Althoughusually spontaneous fermentation processes are applied, the addition of lactic acid bacteria (LAB) starter cultures couldaccelerate processing and improve the consistency and quality of the end-products.
RESULTS: The application of Lactobacillus plantarum IMDO 788 as a starter culture strain for cauliflower and mixed vegetable
fermentations resulted in accelerated acidification as compared with the spontaneous fermentations. The strain dominatedthe background microbiota throughout the process, whereas the spontaneous fermentations were characterised by widelyvariable species diversity. During the spontaneous fermentations, almost all carbohydrates were converted into lactic acid,ethanol, mannitol and acetic acid, indicating the participation of both heterofermentative and homofermentative LAB species.During the starter culture-added fermentations, residual carbohydrates were found and lactic acid and ethanol were the mainend-metabolites.Vegetable-associatedaromas, ethyl acetate and isoamyl acetate were produced during all fermentations. Thehigh concentration of ethanol and the production of ethyl acetate and isoamyl acetate suggested the involvement of yeastsduring all fermentations.
CONCLUSION: Lactobacillus plantarum IMDO 788was an adequate starter culture strainfor vegetablefermentations, prevailingover endogenous LAB communities. Further optimisation of the starter culture formulation is necessary to avoid yeast growth.c 2013 Society of Chemical Industry
Keywords: vegetable fermentation; lactic acid bacteria; Lactobacillus plantarum; starter cultures
INTRODUCTIONFermented vegetables and fruits have been produced for
thousands of years mainly for preservation purposes, as fresh veg-
etables and fruits are very perishable.1,2 In addition, fermentation
deliversend-products thatare appreciated for their typical sensory
properties.3 Fermented vegetables and fruits of economic impor-
tance include sauerkraut and its Asian variant kimchi, cucumbers
and olives, although many fermentations performed on a small
scale or at household level are attracting interest as well.4 These
small-scale fermentations comprise a wide diversity of vegetables
and fruits such as artichokes,5 caper berries,6 carrots,7– 11
cauliflowers,11–13 cherries,14 eggplants,15 garlic,16,17 ginger,18,19
green beans,7 green tomatoes,11,20 leeks,21 mangoes,19 marrows,7
onions,22–25 peppers,26 pineapples27 and turnips28 and are often
performed in Eastern and Southern European countries.
At present, many vegetable and fruit fermentation processes
still rely on the naturally occurring microbiota of the
fresh raw materials, among which lactic acid bacteria (LAB)
overgrow the undesirable communities of Enterobacteriaceae,
Pseudomonadaceae and yeasts, at least when appropriate
fermentation conditions are set up.3 This involves the addition
of salt to the fresh vegetables and fruits (dry salting or brining),
after which the mixture is protected from light and oxygen
and left to ferment at temperatures between 16 and 35 ◦C.2
However, spontaneous fermentation processes suffer from a
lack of control, often resulting in end-products of inconsistent
quality.29 In thecase of cheese and fermented sausage production,
these problems are avoided by the use of starter cultures.29,30
Starter cultures are microbial preparations of a large number
of cells of one or more microbial strains, which are added to
the raw material to accelerate and control the fermentation
process.31 With regard to fermented vegetables and fruits,
however, several difficulties, e.g. the inability to inactivate the
endogenous microbiota by heattreatmentwithoutcausingdrastic
texture changes, and the desired succession of different micro-
organismsupon fermentation, somewhatimpede the introduction
of starter cultures in these cases.29,32 Nevertheless, an increasing
number of studies concerning starter culture use for fermented
∗ Correspondingto:Luc De Vuyst, ResearchGroup of IndustrialMicrobiologyand
Food Biotechnology (IMDO), Department of Bioengineering Sciences, Faculty
of Sciences and Bioengineering Sciences, Vrije Universiteit Brussel, Pleinlaan 2,
B-1050 Brussels, Belgium E-mail: [email protected]
a Research Group of Industrial Microbiology and Food Biotechnology (IMDO),
DepartmentofBioengineeringSciences,FacultyofSciencesandBioengineering
Sciences, VrijeUniversiteitBrussel, Pleinlaan2, B-1050 Brussels, Belgium
b Instituteof Biology Bucharest(IBB),Romanian Academy,SplaiulIndependentei
296, 060031 Bucharest, Romania
J Sci Food Agric 2013; 93: 3352–3361 www.soci.org c 2013 Society of Chemical Industry

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 2/10
Lactobacillus plantarum IMDO 788 as a starter culture for vegetable fermentations www.soci.org
vegetable and fruit production are being published. Furthermore,
the search for appropriate starter cultures is directed towards
functional starter cultures, which are not only able to accelerate
and control the fermentation process but can also contribute to
food safety and/or offer sensory, technological, nutritional and/or
health-associated advantages.31 Goodcandidatefunctionalstarter
cultures are mainly wild-type strains that originate from the
naturalecosystem, as these usually exert more elaborate metabolic
activities in comparison with industrial bulk starters, often from
diversesources.31 This has indeedbeen shownforthefermentation
of carrots, green beans and marrows, forwhich an autochthonous
starter culture performed better with regard to acidification rate,
inhibition of undesired microbial communities, vitamin C content
andsensoryattributesin comparison withan allochthonousstarter
culture consisting of the same species.7 The use of starter cultures
also allows one to reduce the added salt concentration necessary
fore.g. cucumberand sauerkraut fermentations andto reduce the
biogenic amine content of sauerkraut and kimchi.33–39
The frequent appearance of Lactobacillus plantarum as the
prevailing LAB species during vegetable and fruit fermentations
makes this common species a promising candidate for starter
culture trials. In this direction, various Lb. plantarum strains havebeen successfully applied as starter cultures for olive, cucumber
and garlic fermentations.16,36,40–43 Also, Lb. plantarum IMDO 788
has been used as a starter culture for leek fermentations.44
Furthermore, several Lb. plantarum strains possess functional
properties such as thecapacity to degrade oleuropein, to produce
bacteriocins and/or certain flavouring compounds, to tolerate
salt and/or acid environments and to resist gastrointestinal
conditions.41,42,45 All these features make their application as
starter cultures even more interesting.
This study was aimed at the implementation and validation
of Lb. plantarum IMDO 788 as a starter culture for vegetable
fermentations in general. It is a competitive strain isolated from
spontaneously fermented leek that has been tested as a starterculture for fermented leek production.44 The work was performed
through an extensive study of the microbiology and metabolite
kinetics of fermentations with cauliflower and with a mixture of
carrot, cauliflower and green tomato carried out in Romania at
household level.
MATERIALS AND METHODSStarter culture preparation
Lactobacillus plantarum IMDO 788 was used as starter culture
throughout this study. The strain was originally isolated from
a spontaneous leek fermentation process and selected on the
basis of a comparison of 12 Lb. plantarum strains from fermentedleek owing to its fast growth and acidification as well as its
capability to produce diacetyl.21 The strain was stored at −80 ◦C
in de Man/Rogosa/Sharpe (MRS) medium (Oxoid, Basingstoke, UK)
supplemented with 250 g L−1 glycerol (Sigma-Aldrich, Steinheim,
Germany) as cryoprotectant.
For biomass production a monoculture fermentation with Lb.
plantarum IMDO 788 was carried out in MRS mediumat 30 ◦C o n a
200mL scale in glass bottles. After 12 h of fermentation, cells were
harvested through centrifugation (16 000× g, 20min, 4 ◦C) and
washed twice with saline (8.5g L−1 NaCl), each time followed by
centrifugation (16 000×g, 20 min, 4 ◦C). The fresh cell paste was
resuspended in 10 mL of saline, and 5 mL of this cell suspension
was subsequently added to the respective fermentation vessels.
Experimental design
The vegetable fermentations carried out in this study were
performed at household level in Romania. They involved
a spontaneous fermentation with cauliflower alone (further
referred to as the spontaneous cauliflower fermentation) and a
spontaneous fermentation with a mixture of carrot, cauliflower
and green tomato (further referred to as the spontaneous
mixed vegetable fermentation), which served as the control
fermentations. In addition, two fermentations with the same
ingredients as the spontaneous ones (further referred to as the
starter culture-added cauliflower fermentation and the starter
culture-added mixed vegetable fermentation respectively) were
inoculated with Lb. plantarum IMDO 788 as starter culture strain
to obtain a final cell density of ca 6.0 log(colony-forming units
(CFU) g−1 vegetables). The fermentations were performed in 15 L
glass bottles, which were filled with whole (green tomato) or
cut (other vegetables) pieces of freshly harvested vegetables
that were submerged in tap water at room temperature, to
which 35 g L−1 NaCl, 20 g L−1 sucrose and/or starter culture were
added. All fermentations were initiated in the kitchen (ambient
temperature 25 ◦C)and left therefor 3 days, afterwhichthe bottles
were placed in the cellar (ambient temperature 16◦
C) until theend of the fermentations (5 weeks). Brine and vegetable particle
samples were withdrawn at specific time points throughout the
fermentations, namely at days 0, 1, 2, 3, 5, 8, 11, 14, 21, 28 and
35. All samples were immediately analysed for acidification by pH
measurements with an InoLab 720 pH meter (Wissenschaftlich-
Technische Werkstatten GmbH, Weilheim, Germany) and used for
plating andLAB isolation, whilepartsof thesampleswerestoredat
−20 ◦C and transported to Belgium for culture-independent and
metabolite target analyses.
Microbiological analysis
Isolation
For the isolation of LAB, 0.1 mL aliquots of appropriate decimaldilutions of the brine samples prepared in saline were plated on
MRS agar medium (Oxoid). Colonies (10%) from MRS agar were
picked up randomly, purified through plating after growth in MRS
medium, and 1.5 mL of each overnight culture was transferred
to cryovials containing glycerol (250 g L−1) for storage at −80 ◦C.
The isolates were transported on dry ice to Belgium for further
identification.
Bacterial DNA extraction
After having checked their catalase activity, 628 catalase-
negative MRS agar isolates (101, 213, 115 and 199 isolates
from the spontaneous cauliflower, spontaneous mixed vegetable,
starter culture-added cauliflower and starter culture-added mixedvegetablefermentationsrespectively) were grown in MRSmedium
overnight. Then 2 mL of each culture was centrifuged (8000×g,
15min,4 ◦C) and thecell pellets were subjected to DNAextraction
with a Nucleospin 96 tissue kit (Macherey Nagel GmbH, Duren,
Germany) following the manufacturer’s instructions.
For culture-independent analysis, total DNA was extracted
directly from the brine and vegetable particle samples as
described previously.11 Briefly, the actual DNA extraction from
cell pellets was carried out by a combination of enzymatic,
chemical and mechanical lysis of the cells, followed by
phenol/chloroform/isoamyl alcohol (49:49:1 v/v/v) extraction and
purification of the aqueous phase with a Nucleospin food kit
(Macherey Nagel) as described previously.11
J Sci Food Agric 2013; 93: 3352–3361 c 2013 Society of Chemical Industry wileyonlinelibrary.com/jsfa

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 3/10
www.soci.org D Wouters et al.
Culture-dependent identification of bacteria
Classification andidentification of theLAB isolateswere performed
by (GTG)5-PCR fingerprinting of their genomic DNA, numerical
analysis of the fingerprints using BioNumerics 5.1 software
(Applied Maths, Sint-Martens-Latem, Belgium) and verification
of the identity of the (GTG)5-PCR clusters through 16S rRNA gene
sequencing of representative isolates as described previously.11
Culture-independent community dynamics analysis and identification
Culture-independent analysis of the fermentation samples (brine
as well as vegetable particle samples), including community
dynamics and identification, was carried out through denaturing
gradient gel electrophoresis (DGGE) of PCR amplicons targeting
the V3 region of the 16S rRNA gene, using the forward 357f and
reverse 518r universal primer pair.46,47 A GC clamp was attached
to the 5 end of the forward primer. The DGGE conditions (such as
a 35–60% denaturing gradient) and analysis methods, including
DGGE bandsequencing and BLAST analysis of the sequences, were
as described previously.11
Metabolite target analysis
Carbohydrate consumption and end-metabolite production
Metabolite target analysis was performed on the brine samples
of the fermentations. Therefore cells and vegetable particles were
removed from the brines by a 15 min centrifugation at 8000×g
(4 ◦C). The resulting cell-free brine supernatants were analysed by
various chromatography techniques.
High-performance liquid chromatography was applied for the
determination of the concentrations of lactic acid, acetic acid and
ethanol as described previously.11 Calibration was performed with
external standards. Samples were analysed in triplicate and the
results arerepresentedas theaverages of these three independent
measurements. The errors on the measurements are represented
as standard deviations.High-performance anion exchange chromatography with
pulsed amperometric detection was used for the determination of
glucose, fructose, sucrose, glycerol and mannitol concentrations
as described previously.11 Quantifications were performed with
standard addition; the original concentrations in the cell-free
brine supernatants with corresponding errors were calculated as
described previously.48
Volatile aroma production
Static headspace gas chromatography coupled with mass
spectrometry was used for the determination of the evolution of
different volatile aroma compounds throughout the time course
of thefermentations. Forthis, 5 g of cell-freebrine supernatant wastransferred into a headspace vial, to which 1 g of NaCl was added
to enhance volatility.49 As an internal standard, a known amount
of 2-butanol (1mL L−1, Sigma-Aldrich) was added to each sample.
Analyses were performed on an Agilent 6890 gas chromatograph
(Agilent Technologies, Santa Clara, CA, USA) coupled to an Agilent
5973 N mass spectrometer (Agilent Technologies) equipped with
an MPS2 Gerstel autosampler (Gerstel GmbH & Co. KG, Mulheim-
an-der-Ruhr, Germany). The capillary column was a DB-WAXetr
column (Agilent Technologies) with a length of 30 m, an internal
diameter of 0.25 mm and a film thickness of 0.5µm. Helium was
used as carrier gas at a flow rate of 1 mLmin−1. Samples were
equilibrated by agitation at 60 ◦C for 30 min prior to injection.
The injection port was in split mode (40:1 split ratio) and the
needle temperature was 150 ◦C. The injection volume was 1 mL
at a rate of 500 µL s−1. After optimisation, the oven temperature
programme consisted of an initial step at 40 ◦C for 5 min, followed
by a linear increase from 40 to 120 ◦C at 20 ◦Cmin−1 and a linear
increasefrom120to225 ◦Cat10 ◦Cmin−1.Finally,thetemperature
was kept constant at 225 ◦C for 5 min. The temperature of the
transfer tube was held at 280◦C. Detection was done with
an MS detector (ionisation energy 70 eV, 4.1 scans s−1, source
temperature 230 ◦C, scan range m / z 29–200). Identification of the
detectedcomponents was achieved by comparison with standard
compounds that were injected separately and with library data
(NIST 08 database, http://www.nist.gov).
Samples of each time point were analysed in triplicate and the
concentrations of volatile components of interestoriginating from
plant, bacterial or yeast metabolism are represented as arbitrary
units (AU) in relation to the internal standard (2-butanol). The
displayed values represent the means of the three independent
measurements and the respective standard deviations.
RESULTS
Acidification patterns The application of Lb. plantarum IMDO 788 as a starter
culture resulted in accelerated acidification of the vegetables
in comparison with the spontaneous fermentations (Table 1). The
pH values of the starter culture-added cauliflower and mixed
vegetable fermentations dropped within 1 day. Afterwards, only
slight decreases in the pH values occurred. In the spontaneous
fermentations, themajorpH drop occurred during thefirst 2 days,
after which the pH values decreased slightly.
LAB species diversity
The LAB species diversity was less elaborate during the starter
culture-added fermentations than during the spontaneous
fermentations. The starter culture-added fermentations weredominated by Lb. plantarum throughout the entire fermentation
processes. The (GTG)5-PCR fingerprints of the Lb. plantarum
isolates, except for those of some isolates appearing at the
end of the starter culture-added mixed vegetable fermentations,
concurred with the fingerprint of the applied starter culture,
Table 1. Acidification patterns in brine of vegetable fermentationsstudied
pH value
Time
(days)
Spontaneous
cauliflower
Starter
culture-added
cauliflower
Spontaneous
mixed
vegetable
Starter
culture-added
mixed vegetable
0 6.43 6.36 6.58 6.54
1 4.48 3.65 5.76 3.87
2 3.75 3.55 3.87 3.55
3 3.74 3.45 3.49 3.46
5 3.78 3.45 3.50 3.50
8 3.72 3.38 3.42 3.42
11 3.64 3.37 3.36 3.39
14 3.54 3.32 3.29 3.33
21 3.54 3.33 3.23 3.29
28 3.54 3.35 3.16 3.23
35 3.53 3.35 3.16 3.23
wileyonlinelibrary.com/jsfa c 2013 Society of Chemical Industry J Sci FoodAgric 2013; 93: 3352–3361

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 4/10
Lactobacillus plantarum IMDO 788 as a starter culture for vegetable fermentations www.soci.org
indicating that the starter culture strain Lb. plantarum IMDO 788
prevailed during these fermentations. In contrast, a succession of
micro-organisms occurredduring the spontaneousfermentations.
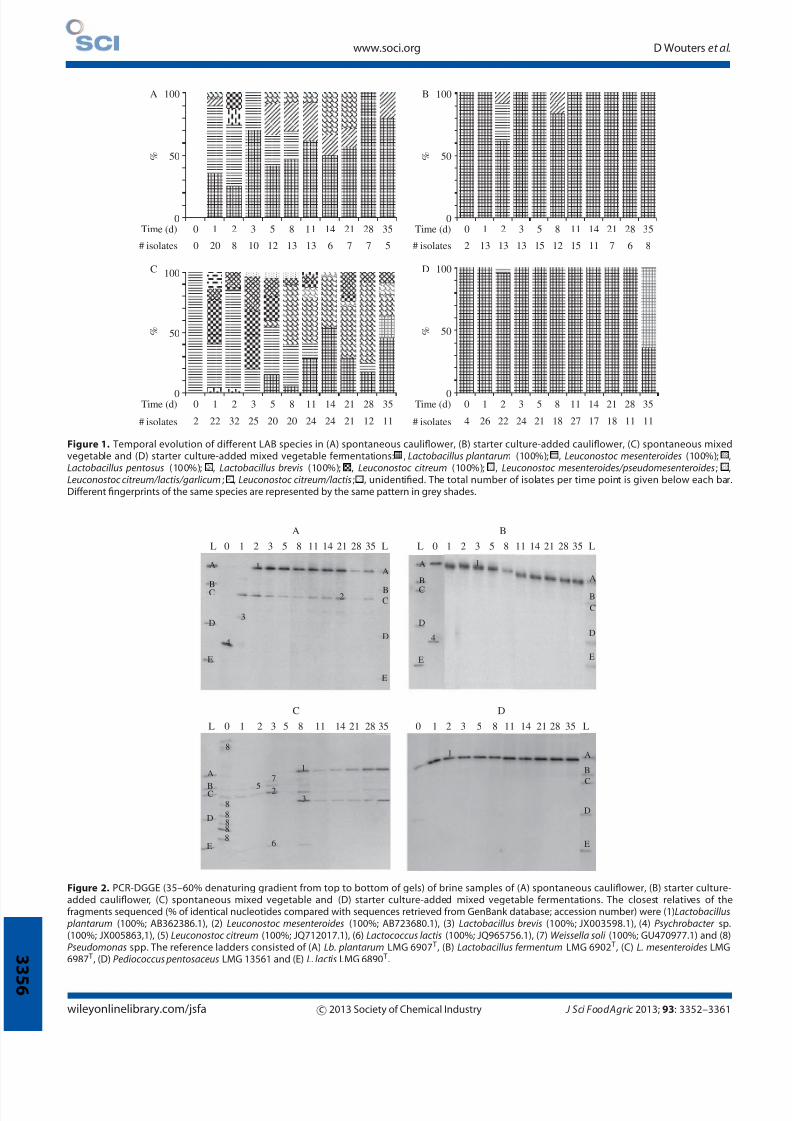
Lactobacillus plantarum was the main species found during the
spontaneous cauliflower fermentation (Table 2). Also, Leuconostoc
mesenteroides, Lactobacillus pentosus and, to a lesser extent,
Lactobacillus brevis were important members of the fermentation
microbiota of the spontaneous cauliflower fermentation. At the
start of this fermentation, Lb. plantarum and L. mesenteroides
prevailed (Fig. 1). After 5 days of fermentation, the share of
L. mesenteroides decreased and Lb. pentosus together with Lb.
plantarum took over. After 5 weeks of fermentation, Lb. plantarum
was the prevailing LAB species in the spontaneous cauliflower
fermentation. Except for Lb. pentosus, the same LAB species as
those found during the spontaneous cauliflower fermentation
were present in the spontaneous mixed vegetable fermentation
(Table 2). However, the presence of Lb. plantarum was not
as abundant. The most frequently isolated species during the
spontaneous mixed vegetable fermentation was L. mesenteroides,
followed by Lb. brevis. In addition, Leuconostoc citreum was often
isolated from this fermentation. Together with L. mesenteroides,
it prevailed at the onset of the spontaneous mixed vegetablefermentation (Fig. 1). After 5 days, a shift of the fermentation
microbiota took place, with Lb. brevis and L. mesenteroides as the
prevalentLAB species.Upon furtherfermentation, L.mesenteroides
disappeared and Lb. brevis together with Lb. plantarum took over
the fermentation process.
PCR-DGGE gels of brine and vegetable particle fermentation
samples were similar for all fermentations carried out; an overview
of the PCR-DGGE analyses of the brine samples of the different
fermentations is presented in Fig. 2. Culture-independent analysis
of thestarterculture-added fermentations exposed theubiquitous
presence of Lb. plantarum. Culture-independent analysis of
the spontaneous fermentations revealed the presence of L.
mesenteroides, Lb. brevis and Lb. plantarum, with L. mesenteroides
and Lb. plantarum as the prevailing LAB species throughout
the spontaneous cauliflower fermentation and Lb. plantarum
together with Lb. brevis as the prevailing LAB species from day 8
onwards during the spontaneous mixed vegetable fermentation.
The presence of L. mesenteroides during the spontaneous mixed
vegetable fermentation was restricted to the first 8 days of the
fermentation. During the initial phase of the spontaneous mixed
vegetable fermentation, L. citreum, Weissella soli and Lactococcus
lactis were present as well. The latter two species, however, were
not found during the culture-dependent diversity analysis. Besides
LAB, Pseudomonas species and a Psychrobacter species were
found at the start of the spontaneous mixed vegetable and the
spontaneous and starter culture-added cauliflower fermentations
respectively.
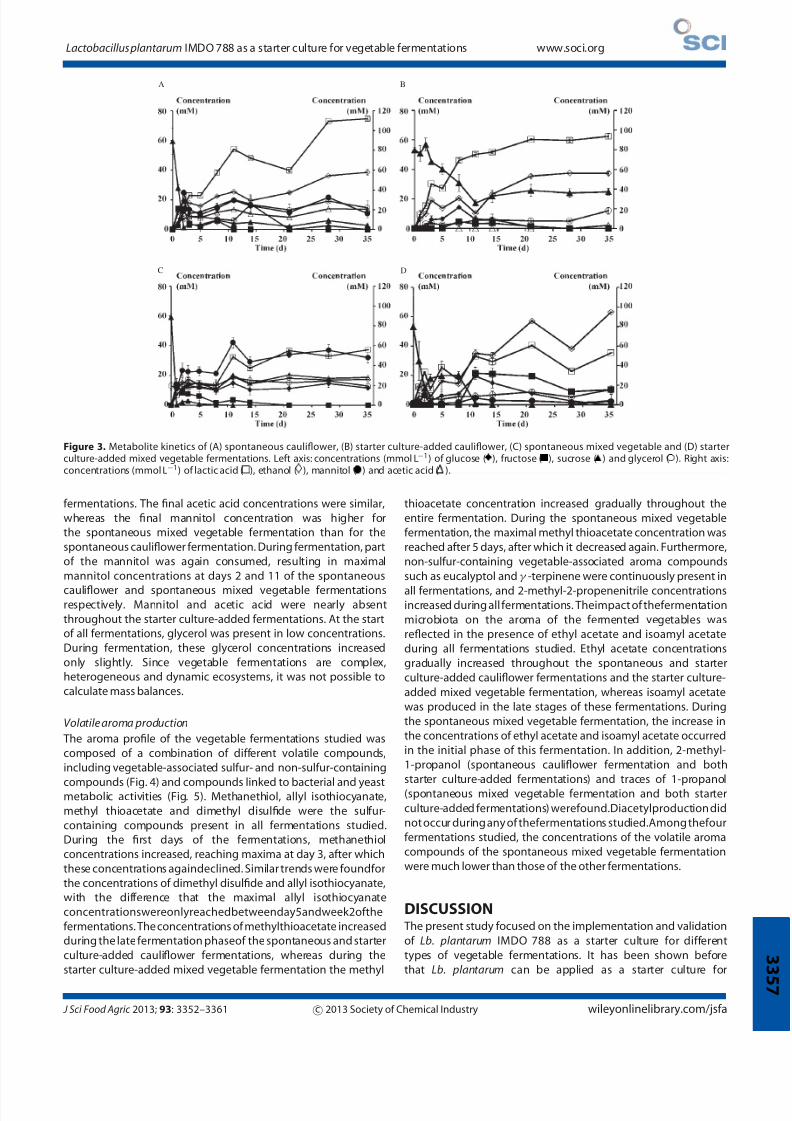
Metabolite kineticsCarbohydrate consumption and end-metabolite production
At thestart of allfermentations, sucrose was thesolecarbohydrate
in the brine samples (Fig. 3). Upon further fermentation, a
clear distinction could be made between the carbohydrate
consumption patterns for the spontaneous and starter culture-
added fermentations. Sucrose was depleted at days 1 and 2 of
the spontaneous mixed vegetable and spontaneous cauliflower
fermentations respectively, whileitwas onlydepletedafter2 weeks
during the starter culture-added mixed vegetable fermentation.
During the starter culture-added cauliflower fermentation, only
part of the sucrose was used. Glucose and fructose were present
in nearly equimolar concentrations (±13mmolL−1)atday1ofthe
spontaneous fermentations, after which fructose was consumed
gradually, and it was depleted after 11 days and 3 weeks of the spontaneous cauliflower and spontaneous mixed vegetable
fermentations respectively. During the spontaneous cauliflower
fermentation, glucose concentrations increased slightly until day
11 of the fermentation, after which it was further consumed to
become depletedin week 3. During the entire spontaneous mixed
vegetable fermentation, the glucose concentrations remained
nearly constant. Glucose and fructose concentrations of the
starter culture-added fermentations were initially low. During
fermentation, they first increased slightly, reaching their maximal
values at day 11, and decreased again upon further fermentation.
Glucose and fructose were depleted after 3 weeks of the starter
culture-added cauliflower fermentation, whereas only glucose
was depleted at the end of the starter culture-added mixedvegetable fermentation. Lactic acid was a major end-metabolite
of carbohydrate metabolism during all fermentations. However,
final lactic acid concentrations of the spontaneous and starter
culture-added cauliflower fermentations were approximately
twice those of the spontaneous and starter culture-added
mixed vegetable fermentations. Ethanol was produced during
all fermentations, reaching divergent final concentrations. Acetic
acid and mannitol were produced during both spontaneous
Table 2. LAB species diversity of vegetable fermentations studied as revealed through (GTG)5-PCR fingerprinting of genomic DNA of 628 isolates. The numbers represent the percentages of the to tal amount of isolates of the particular fermentations
Species
Spontaneous cauliflower
(101 isolates)
Starter culture-added
cauliflower (115 isolates)
Spontaneous mixed
vegetable (213 isolates)
Starter culture-added
mixed vegetable (199 isolates)
Lb. plantarum 52.5 93.9 18.3 96.0
Lb. plantarum fingerprint type (FT) 2 — — 0.9 3.5
L. mesenteroides 23.8 3.5 28.2 0.5
L. mesenteroides/pseudomesenteroides 1.0 — 0.5 —
Lb. pentosus 13.9 3.6 — —
Lb. brevis 7.9 — 23.0 —
Lb. brevis FT2 — — 2.3 —
L. citreum 1.0 — 12.7 —
L. citreum/lactis/garlicum — — 11.3 —
L. citreum/lactis — — 1.4 —
Unidentified — — 1.4 —
J Sci Food Agric 2013; 93: 3352–3361 c 2013 Society of Chemical Industry wileyonlinelibrary.com/jsfa
8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 5/10
www.soci.org D Wouters et al.
100
50
0
100
50
0
100
50
0
100
50
0
0 20 8 10 12 13 13 6 7 7 5# isolates
% %
% %
Time (d) 0 1 2 3 5 8 11 14 28 35
2 22 32 25 2020 24 24 21 12 11# isolates
Time (d)
# isolates
Time (d)
2 13 13 13 15 12 15 11 7 6 8
# isolates 4 26 22 24 21 18 27 17 18 11 11
A
C
B
D
0 1 2 3 5 8 11 14 21 28 3521
0 1 2 3 5 8 11 14 28 35 Time (d) 0 1 2 3 5 8 11 14 21 28 3521
Figure 1. Temporal evolution of different LAB species in (A) spontaneous cauliflower, (B) starter culture-added cauliflower, (C) spontaneous mixedvegetable and (D) starter culture-added mixed vegetable fermentations: , Lactobacillus plantarum (100%); , Leuconostoc mesenteroides (100%); ,Lactobacillus pentosus (100%); , Lactobacillus brevis (100%); , Leuconostoc citreum (100%); , Leuconostoc mesenteroides/pseudomesenteroides; ,Leuconostoc citreum/lactis/garlicum; , Leuconostoc citreum/lactis; , unidentified. The total number of isolates per time point is given below each bar.Different fingerprints of the same species are represented by the same pattern in grey shades.
A B
A
D
B
E
C
4 D
B
E
A
1
C
C
A
D
B
E
D
B
E
A
C 2
3
1
4
C
A
D
B
E
C
1
23
5
6
7
8
8
8
88
8
D
1
D
B
E
A
C
L 0 1 2 3 5 8 11 14 21 28 35 L L 0
0
1
1
2
2
3
3
5
5
8
8
11
11
14
14
21
21
28
28
35
35
L
L0 1 2 3 5 8 11 14 21 28 35L
Figure 2. PCR-DGGE (35–60% denaturing gradient from top to bottom of gels) of brine samples of (A) spontaneous cauliflower, (B) starter culture-added cauliflower, (C) spontaneous mixed vegetable and (D) starter culture-added mixed vegetable fermentations. The closest relatives of thefragments sequenced (% of identical nucleotides compared with sequences retrieved from GenBank database; accession number) were (1) Lactobacillus plantarum (100%; AB362386.1), (2) Leuconostoc mesenteroides (100%; AB723680.1), (3) Lactobacillus brevis (100%; JX003598.1), (4) Psychrobacter sp.(100%; JX005863,1), (5) Leuconostoc citreum (100%; JQ712017.1), (6) Lactococcus lactis (100%; JQ965756.1), (7) Weissella soli (100%; GU470977.1) and (8)Pseudomonas spp. The reference ladders consisted of (A) Lb. plantarum LMG 6907 T , (B) Lactobacillus fermentum LMG 6902 T , (C) L. mesenteroides LMG6987 T , (D) Pediococcus pentosaceus LMG 13561 and (E) L. lactis LMG 6890 T .
wileyonlinelibrary.com/jsfa c 2013 Society of Chemical Industry J Sci FoodAgric 2013; 93: 3352–3361
8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 6/10
Lactobacillus plantarum IMDO 788 as a starter culture for vegetable fermentations www.soci.org
A B
C D
Figure 3. Metabolite kinetics of (A) spontaneous cauliflower, (B) starter culture-added cauliflower, (C) spontaneous mixed vegetable and (D) starterculture-added mixed vegetable fermentations. Left axis: concentrations (mmol L−1) of glucose ( ), fructose ( ), sucrose ( ) and glycerol ( ). Right axis:concentrations (mmol L−1) of lactic acid ( ), ethanol ( ), mannitol ( ) and acetic acid ( ).
fermentations. The final acetic acid concentrations were similar,
whereas the final mannitol concentration was higher for
the spontaneous mixed vegetable fermentation than for the
spontaneous cauliflower fermentation. During fermentation, part
of the mannitol was again consumed, resulting in maximal
mannitol concentrations at days 2 and 11 of the spontaneous
cauliflower and spontaneous mixed vegetable fermentationsrespectively. Mannitol and acetic acid were nearly absent
throughout the starter culture-added fermentations. At the start
of all fermentations, glycerol was present in low concentrations.
During fermentation, these glycerol concentrations increased
only slightly. Since vegetable fermentations are complex,
heterogeneous and dynamic ecosystems, it was not possible to
calculate mass balances.
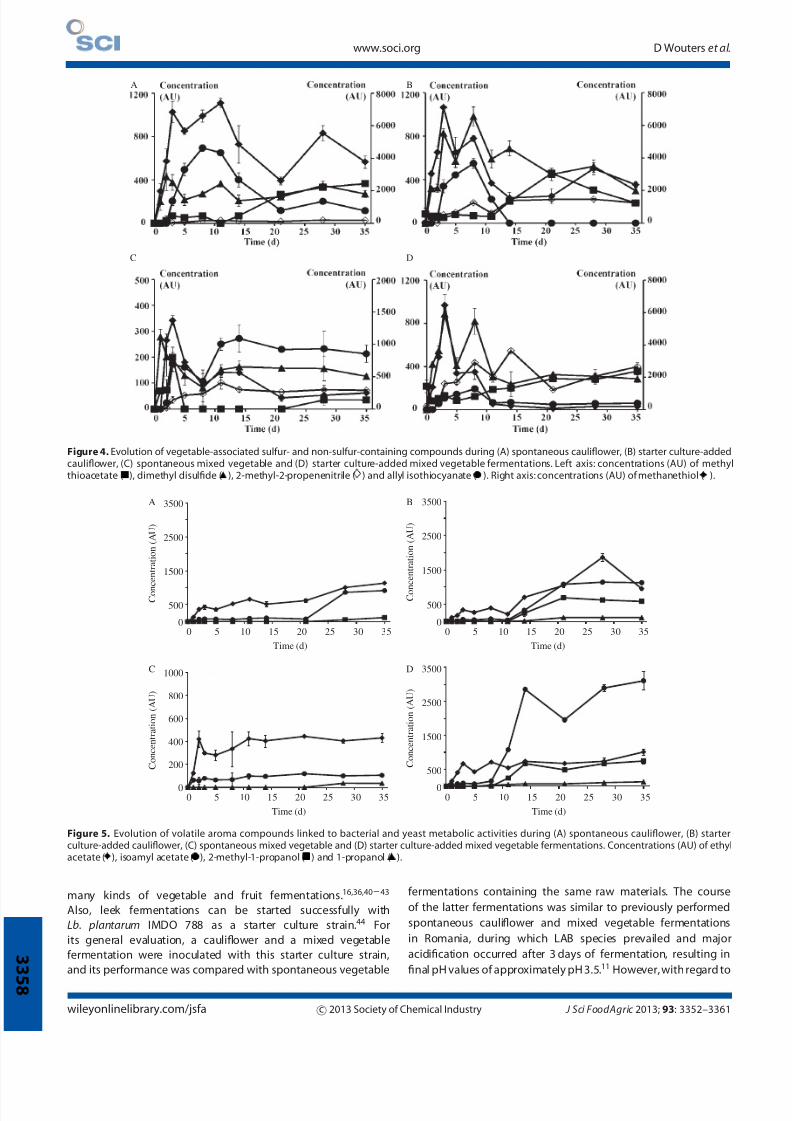
Volatile aroma production
The aroma profile of the vegetable fermentations studied was
composed of a combination of different volatile compounds,
including vegetable-associated sulfur- and non-sulfur-containing
compounds (Fig. 4) and compounds linked to bacterial and yeastmetabolic activities (Fig. 5). Methanethiol, allyl isothiocyanate,
methyl thioacetate and dimethyl disulfide were the sulfur-
containing compounds present in all fermentations studied.
During the first days of the fermentations, methanethiol
concentrations increased, reaching maxima at day 3, after which
these concentrations againdeclined. Similar trends were foundfor
the concentrations of dimethyl disulfide and allyl isothiocyanate,
with the difference that the maximal allyl isothiocyanate
concentrationswereonlyreachedbetweenday5andweek2ofthe
fermentations. The concentrations of methylthioacetate increased
during the late fermentation phaseof the spontaneous and starter
culture-added cauliflower fermentations, whereas during the
starter culture-added mixed vegetable fermentation the methyl
thioacetate concentration increased gradually throughout the
entire fermentation. During the spontaneous mixed vegetable
fermentation, the maximal methyl thioacetate concentration was
reached after 5 days, after which it decreased again. Furthermore,
non-sulfur-containing vegetable-associated aroma compounds
such as eucalyptol and γ -terpinene were continuously present in
all fermentations, and 2-methyl-2-propenenitrile concentrationsincreased during all fermentations. Theimpact of thefermentation
microbiota on the aroma of the fermented vegetables was
reflected in the presence of ethyl acetate and isoamyl acetate
during all fermentations studied. Ethyl acetate concentrations
gradually increased throughout the spontaneous and starter
culture-added cauliflower fermentations and the starter culture-
added mixed vegetable fermentation, whereas isoamyl acetate
was produced in the late stages of these fermentations. During
the spontaneous mixed vegetable fermentation, the increase in
the concentrations of ethyl acetate and isoamyl acetate occurred
in the initial phase of this fermentation. In addition, 2-methyl-
1-propanol (spontaneous cauliflower fermentation and both
starter culture-added fermentations) and traces of 1-propanol
(spontaneous mixed vegetable fermentation and both starter
culture-added fermentations) werefound.Diacetylproduction did
not occur during any of thefermentations studied.Among thefour
fermentations studied, the concentrations of the volatile aroma
compounds of the spontaneous mixed vegetable fermentation
were much lower than those of the other fermentations.
DISCUSSION The present study focused on the implementation and validation
of Lb. plantarum IMDO 788 as a starter culture for different
types of vegetable fermentations. It has been shown before
that Lb. plantarum can be applied as a starter culture for
J Sci Food Agric 2013; 93: 3352–3361 c 2013 Society of Chemical Industry wileyonlinelibrary.com/jsfa
8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 7/10
www.soci.org D Wouters et al.
A B
DC
Figure 4. Evolution of vegetable-associated sulfur- and non-sulfur-containing compounds during (A) spontaneous cauliflower, (B) starter culture-addedcauliflower, (C) spontaneous mixed vegetable and (D) starter culture-added mixed vegetable fermentations. Left axis: concentrations (AU) of methylthioacetate ( ), dimethyl disulfide ( ), 2-methyl-2-propenenitrile ( ) and allyl isothiocyanate ( ). Right axis: concentrations (AU) of methanethiol ( ).
Time (d)
A
C
B
D
C o n c e n t r a t i o n ( A U )
3500
2500
1500
500
00 10 15 20 25 30 35
C o n
c e n t r a t i o n ( A U )
3500
2500
1500
500
0
C o n c e n t r a t i o n ( A U )
1000
800
600
400
200
0
C o n
c e n t r a t i o n ( A U )
3500
2500
1500
500
0
5
Time (d)
0 10 15 20 25 30 355
Time (d)
0 10 15 20 25 30 355
Time (d)
0 10 15 20 25 30 355
Figure 5. Evolution of volatile aroma compounds linked to bacterial and yeast metabolic activities during (A) spontaneous cauliflower, (B) starterculture-added cauliflower, (C) spontaneous mixed vegetable and (D) starter culture-added mixed vegetable fermentations. Concentrations (AU) of ethylacetate ( ), isoamyl acetate ( ), 2-methyl-1-propanol ( ) and 1-propanol ( ).
many kinds of vegetable and fruit fermentations.16,36,40–43
Also, leek fermentations can be started successfully with
Lb. plantarum IMDO 788 as a starter culture strain.44 For
its general evaluation, a cauliflower and a mixed vegetable
fermentation were inoculated with this starter culture strain,
and its performance was compared with spontaneous vegetable
fermentations containing the same raw materials. The course
of the latter fermentations was similar to previously performed
spontaneous cauliflower and mixed vegetable fermentations
in Romania, during which LAB species prevailed and major
acidification occurred after 3 days of fermentation, resulting in
final pH values of approximately pH 3.5.11 However, with regard to
wileyonlinelibrary.com/jsfa c 2013 Society of Chemical Industry J Sci FoodAgric 2013; 93: 3352–3361

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 8/10
Lactobacillus plantarum IMDO 788 as a starter culture for vegetable fermentations www.soci.org
the composition of the fermentation microbiota, the spontaneous
cauliflower fermentation of the present study differed from the
previously performed cauliflower fermentation, as Lb. pentosus, L.
mesenteroides and, in particular, Lb. plantarum prevailed during
the different fermentation stages instead of Weissella kimchii and
Lb. brevis. Yet, such a sequential prevalence of L. mesenteroides
and Lb. plantarum has been shown for cauliflower fermentations
performed according to a traditional recipe in Northern Greece
before.13 The spontaneous mixed vegetable fermentation of thepresent study harboured fewer Weissella species compared with
the one performed before.11
The application of the Lb. plantarum IMDO 788 starter culture
strain affected the fermentation courses in different ways. For
example, its addition in high numbers ensured a nearly immediate
prevalence of this strain, which was maintained throughout
the entire starter culture-added fermentation processes. Such
a persistence of the added starter culture strain is one of the
main criteria for its applicability, which has been demonstrated
for the use of Lb. plantarum starter culture strains in the
production of, among others, kimchi and olives before.50,51 As
a result of the fast prevalence of Lb. plantarum IMDO 788 in
the fermentations studied, the pH dropped very rapidly duringday 1 of the fermentations, which is beneficial for the quality of
the end-products, as rapid acidification minimises the influence
of the background microbiota that may contain spoilage and
pathogenic bacteria.7,26,42 Regarding carbohydrate consumption
patterns, one of the main influences of the application of the
Lb. plantarum IMDO 788 starter culture strain was the low
presence of mannitol and acetic acid due to a low abundance
of heterofermentative LAB species. In contrast, residual sucrose
was retrieved for a long time during the starter culture-added
fermentations, even until the end of the starter culture-added
cauliflower fermentation. Residual glucose and fructose were
found in the case of the spontaneous and starter culture-added
mixed vegetable fermentations respectively. The presence of
residual carbohydrates may cause problems with regard to thestability of the end-products, since it may lead to secondary
fermentation by yeasts, as has been shown during carrot
fermentations. 8 Secondary fermentation by yeasts can result in
tissue softening due to pectinolytic activity and in the production
of off-flavours through lipolytic and saccharolytic activities.3,52
Nevertheless, yeasts are not always considered undesirable, as
they can contribute to the generation of a versatile aroma profile,
as, for instance, in the case of fermented olives, artichokes
and caper berries.5,6,53,54 Yet, it remains unclear if yeasts are
desirable in vegetable and fruit fermentations or whether they are
primarily a threat to the stability and quality of the end-products.
Indications of a strong involvement of yeasts in the fermentations
studied were reflected in the ethanol concentrations, which weregenerally much higher than those measured in the case of
Romanianvegetablefermentationsthatwerestudiedpreviously. 11
Aroma analysis revealed this large participation of yeasts in the
fermentation processes as well, as ethyl acetate and isoamyl
acetate were among the main aroma compounds found. Since
such esters may provide fruity notes, their presence may be
desirable, although excessive concentrations lead to off-flavours.
Also, glycerol formation during these fermentations pointed
towards the activity of yeasts. In many other cases, however,
the addition of a starter culture is sufficient for microbial control
of the fermentation process, as has been shown, for instance, for
kimchi fermentations.55 Presumably, the addition of sucrose at
the start of the fermentations is one of the main reasons for the
presence of yeasts, since sucrose is not preferred as an energy
sourceby the added Lb. plantarum starter culture strain. However,
in view of a fast prevalence of LAB, the addition of carbohydrates
occurs occasionally at the start of traditional vegetable and fruit
fermentations, such as in the case of olive, onion and Thai mixed
vegetable and fruit fermentations.19,25,56 For example, for the
fermentation of Bella di Cerignola table olives, the addition of a
starter culture consisting of multiple strains of Lb. plantarum and
of small amounts of glucose assures a correct fermentationcourse,
which includesa decreasein the pHto a safevalue and the control
of yeast growth, whereas the addition of glucose alone stimulates
yeast growth.56 Besides the contribution of yeasts to the aroma of
fermented vegetables, theraw ingredientsand metabolic activities
of LAB influence the final aroma. Sulfur-containing compounds
such as methanethiol and allyl isothiocyanate and the non-sulfur-
containing compound 2-methyl-2-propenenitrile belong to the
breakdown products of glucosinolates, present in all cabbage
types; they are partly responsible for the characteristic aroma of
cabbages.57 Besides through the formation of lactic acid, acetic
acid, mannitol and/or ethanol, the influence of the metabolic
activities of LAB on the aroma of fermented vegetables could be
expressed through the presence of volatile compounds such as1-propanol and methanethiol, of which the former is presumably
produced via lactic acid conversion and the latter via amino acid
conversions.58,59 Overall, no clear influence of the application of
the Lb. plantarum starter culture strain was noticed with regard
to the aroma of the end-products, as no diacetyl production,
characteristic for this strain, was noticed.
To conclude, the applied starter culture strain Lb. plantarum
IMDO 788 wasadequateto dominatethe background LABspecies
of vegetables, thereby ensuring rapid acidification and thus
microbiologically safe end-products.However, yeastgrowthcould
not be suppressed, perhaps because of sucrose addition, which
makes the additionof small amounts of glucose instead of sucrose,
if any, and the introduction of, preferably, a heterofermentative
LAB strain to the starter culture advisable for future experiments,
to limit residual carbohydrate concentrations and to obtain end-
products without yeasts.
ACKNOWLEDGEMENTS The authors acknowledge the financial support of the Research
Council fromthe VrijeUniversiteit Brussel, the Romanian Academy
and the Research Foundation Flanders (FWO). DW is the recipient
of a post-graduate grant from the Agency for Innovation by
Science and Technology (IWT) in Flanders. Part of this work was
supported by the post-doctoral research project PD33 of the
Romanian National Research Plan (PNII-RU) and by the project
RO1567-IBB05/2012 of the Institute of Biology Bucharest of theRomanian Academy.
REFERENCES1 Gorny J, Microbial contamination of fresh fruits and vegetables, in
Microbiology of Fruits and Vegetables, ed. by Sapers GM, Gorny JRand Yousef AEY. CRC Press, Boca Raton, FL, pp. 3–32 (2006).
2 Li KY, Fermentation: principles and microorganisms, in Handbook of Fermented Food and Beverage Technology , ed. by Hui YH, Meunier-GoddikL, HansenAS, Josephsen J, NipWK, Stanfield PS, etal. MarcelDekker, New York, NY, pp. 595–609 (2004).
3 Hutkins RW, Fermented vegetables, in Microbiology and Technologyof Fermented Foods, ed. by Hutkins RW. Blackwell Publishing, Oxford,pp. 223– 259 (2006).
J Sci Food Agric 2013; 93: 3352–3361 c 2013 Society of Chemical Industry wileyonlinelibrary.com/jsfa

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 9/10
www.soci.org D Wouters et al.
4 Breidt FJR, McFeeters RF and Dıaz-Muniz I, Fermented vegetables, inFoodMicrobiology:Fundamentalsand Frontiers,ed.byDoyleMPandBeuchat LR. ASM Press, Washington, DC, pp. 783–793 (2007).
5 YokoiK, Kawasaki K,NishitaniG, Taketo A andKodaira K,Fermentationof Jerusalem artichoke with or without lactic acid bacteria startercultures. FoodSci Technol Res 12:231– 234 (2006).
6 Pulido RP, Ben Omar N, Abriouel H, Lopez RL, Canamero MM andGalvez A, Microbiological study of lactic acid fermentation of caperberries by molecular and culture-dependent methods. Appl EnvironMicrobiol 71:7872–7879 (2005).
7 Di Cagno R, Surico RF, Siragusa S, De Angelis M, Paradiso A, MinerviniF, et al., Selection and use of autochthonous mixed starter for lacticacid fermentation of carrots, French beans or marrows. Int J Food Microbiol 127:220– 228 (2008).
8 Gardner NJ, Savard T, Obermeier P, Caldwell G and Champagne CP,Selection and characterization of mixed starter cultures for lacticacid fermentation of carrot, cabbage, beet and onion vegetablemixtures. Int J Food Microbiol 64:261– 275 (2001).
9 Joshi VK, Sharma S and Thakur NS, Effect of temperature, saltconcentration and fermentation type (inoculated vs. natural) onlacticacidfermentationbehaviourandqualityofcarrot. Acta Aliment 37:205–219 (2008).
10 Niketic-Aleksic GK, Bourne MC and Stamer JR, Preservation of carrotsby lactic acid fermentation. J FoodSci 38:84–86 (1973).
11 Wouters D, Grosu-Tudor S, Zamfir M and De Vuyst L, Bacterialcommunity dynamics, lactic acid bacteria species diversity,
and metabolite kinetics of traditional Romanian vegetablefermentations. J Sci FoodAgric 93:749– 760 (2013).
12 Desai P and Sheth T, Controlled fermentation of vegetables usingmixed inoculum of lactic cultures. J Food Sci Technol 34 :155–158(1997).
13 Paramithiotis S, Hondrodimou OL and Drosinos EH, Developmentof the microbial community during spontaneous cauliflowerfermentation. Food ResInt 43:1098–1103 (2010).
14 Di Cagno R, Surico RF, Minervini G, Rizzello CG, Lovino R, Servili M,et al., Exploitation of sweet cherry (Prunus avium L.) puree addedof stem infusion through fermentation by selected autochthonouslactic acid bacteria. Food Microbiol 28:900– 909 (2011).
15 Sanchez I, Palop L and Ballesteros C, Biochemical characterizationof lactic acid bacteria isolated from spontaneous fermentation of Almagro eggplants. Int J Food Microbiol 59:9–17 (2000).
16 De Castro A, Montano A, Sanchez AH and Rejano L, Lactic acidfermentation and storage of blanched garlic. Int J Food Microbiol
39:205–211 (1998).17 MontanoA, CasadoFJ, De CastroA, SanchezAH andRejano L, Vitamin
content and amino acid composition of pickled garlic processedwith and without fermentation. J Agric Food Chem 52:7324–7330(2004).
18 Chang CH, Chen YS and Yanagida F, Isolation and characterisation of lactic acid bacteria from yan-jiang (fermented ginger), a traditionalfermented food in Taiwan. J Sci FoodAgric 91:1746–1750 (2011).
19 Tanganurat W, Quinquis B, Leelawatcharamas V and BolotinA, Genotypic and phenotypic characterization of Lactobacillus plantarum strains isolated from Thai fermented fruits andvegetables. JBasic Microbiol 49:377– 385 (2009).
20 Laleye LC and Simard RE, Lactic acid fermentation of sliced greentomatoes. Dev IndMicrobiol 23:601– 609 (1982).
21 Wouters D, Bernaert N, Conjaerts W, Van Droogenbroeck B, DeLoose M and De Vuyst L, Species diversity, community dynamics,
and metabolite kinetics of spontaneous leek fermentations. Food Microbiol 33:185–196 (2013).22 Bisakowski B, Atwal AS, Gardner N and Champagne CP, Effect of lactic
acid fermentation of onions ( Allium cepa) on the composition of flavonol glucosides. IntJ FoodSci Technol 42:783– 789 (2007).
23 KarovicovaJ,Drdak M, Simko P andVollekV, Organic acids productionby lactic acid fermentation of vegetables (green pea and onion).Nahrung 36:500– 502 (1992).
24 Millet A, Lamy E, Jonas D, Stintzing F, Mersch-Sundermann V andMerfort I, Fermentation enhances the biological activity of Alliumcepa bulb extracts. J Agric Food Chem 60:2148–2156 (2012).
25 Roberts JS and Kidd DR, Lactic acid fermentation of onions. Food Sci Technol 38:185– 190 (2005).
26 Di Cagno R, Surico RF, Minervini G, De Angelis M, Rizzello CG andGobbettiM,Useofautochthonousstarterstofermentredandyellowpeppers (Capsicumannum L.)to be stored at room temperature. Int J Food Microbiol 130:108– 116 (2009).
27 Di Cagno R, Cardinali G, Minervini G, Antonielli L, Rizzello CG, RicciutiP, et al., Taxonomic structure of the yeasts and lactic acid bacteriamicrobiota of pineapple ( Ananas comosus L. Merr.) and use of autochthonous starters for minimally processing. Food Microbiol 27:381– 389 (2010).
28 Maifreni M, MarinoM andConte L, Lacticacid fermentation of Brassicarapa: chemical and microbial evaluation of a typical Italian product(Brovada). Eur Food ResTechnol 218:469– 473 (2004).
29 JosephsenJ andJespersenL, Starter cultures andfermented products,in Handbook of Fermented Foodand Beverage Technology , ed. byHuiYH, Meunier-Goddik L, Hansen AS, Josephsen J, Nip WK, StanfieldPS, et al. Marcel Dekker, New York, NY, pp. 23–49 (2004).
30 Leroy F,VerluytenJ andDe VuystL, Functional meatstarterculturesforimproved sausage fermentation. Int J Food Microbiol 106:270–285(2006).
31 Leroy F and De Vuyst L, Lactic acid bacteria as functional startercultures for the food fermentation industry. TrendsFoodSci Technol 15:67–78 (2004).
32 HarrisLJ, Themicrobiology of vegetable fermentation,in Microbiology of Fermented Foods, ed. by Wood BJB. Blackie Academic andProfessional, London, pp. 45–72 (1998).
33 Halasz A, Barath A and Holzapfel WH, The influence of starter cultureselection on sauerkraut fermentation. Z Lebensm Unters Forsch208:434–438 (1999).
34 Johanningsmeier S, McFeeters RF, Fleming HP and ThompsonRL, Effects of Leuconostoc mesenteroides starter culture on
fermentation of cabbage with reduced salt concentrations. J Food Sci 72:M166–M172 (2007).
35 Lee H, Yoon H, Ji Y, Kim H, Park H, Lee J, et al., Functional propertiesof Lactobacillus strains isolated from kimchi. Int J Food Microbiol 145:155–161 (2011).
36 McFeeters RF and Perez-Dıaz I , Fermentation o f cucumbers brinedwith calcium chloride instead of sodium chloride. J Food Sci 75:C291–C296 (2010).
37 Rabie MA, Siliha H, El-Saidy S, El-Badawy AA and Xavier Malcata F,Reduced biogenic amine contents in sauerkraut via addition of selected lactic acid bacteria. Food Chem 129:1778–1782 (2011).
38 Spicka J, Kalac P, Bover-Cid S and K rizek M, Application of lactic acidbacteria starter cultures for decreasing the biogenic amine levels insauerkraut. Eur Food ResTechnol 215:509– 514 (2002).
39 Wiander B and Ryhanen EL, Laboratory and large-scale fermentationof white cabbage into sauerkraut and sauerkraut juice by usingstarters in combination with mineral salt with a low NaCl content.
EurFood ResTechnol 220:191–195 (2005).40 Leal MV, Baras M, Ruiz-Barba JL, Floriano B and Jimenez-Dıaz
R, Bacteriocin production and competitiveness of Lactobacillus plantarum LPCO10 in olive juice broth, a culture medium obtainedfrom olives. Int J Food Microbiol 43:129– 134 (1998).
41 Leal-SanchezMV, Ruiz-Barba JL,SanchezAH, RejanoL, Jimenez-Dıaz RandGarrido A,Fermentationprofile andoptimizationof green olivefermentation using Lactobacillus plantarum LPCO10 as a starterculture. Food Microbiol 20:421– 430 (2003).
42 Romeo F and Poiana M, Abilityof commercially available Lactobacillusstrains as starter in brining and debittering of table olives. Acta Aliment 36:49– 60 (2007).
43 Sanchez AH, Rejano L, Montano A and de Castro A, Utilization at highpH of starter cultures of lactobacilli for Spanish-style green olivefermentation. Int J Food Microbiol 67:115– 122 (2001).
44 Wouters D, Bernaert N, Anno N, Van Droogenbroeck B, De Loose M,
Van Bockstaele E and De Vuyst L, Application and validation of autochthonous lactic acid bacteria starter cultures for controlledleekfermentations andtheir influence on theantioxidantpropertiesof leek. International Journal of Food Microbiology 165:121–133(2013).
45 Karasu N, Simsek O and Con AH, Technological and probioticcharacteristics of Lactobacillus plantarum strains isolated fromtraditionally produced fermented vegetables. Ann Microbiol 60:227– 234 (2010).
46 Ercolini D, Moschetti G, Blaiotta G and Coppola S, Behavior of variableV3 region from 16S rDNA of lactic acid bacteria in denaturinggradient gel electrophoresis. Curr Microbiol 42:199– 202 (2001).
47 Vasilopoulos C, Ravyts F, De Maere H, De Mey E, Paelinck H, DeVuyst L, et al., Evaluation of the spoilage lactic acid bacteria inmodified-atmosphere-packaged artisan-type cooked ham usingculture-dependent and culture-independent approaches. J Appl Microbiol 104:1341–1353 (2008).
wileyonlinelibrary.com/jsfa c 2013 Society of Chemical Industry J Sci FoodAgric 2013; 93: 3352–3361

8/12/2019 Articulo Fermentados
http://slidepdf.com/reader/full/articulo-fermentados 10/10
Lactobacillus plantarum IMDO 788 as a starter culture for vegetable fermentations www.soci.org
48 Vrancken G, Rimaux T, De Vuyst L and Leroy F, Kinetic analysis of growth and sugar consumption by Lactobacillus fermentum IMDO130101 revealsadaptation to theacidic sourdough ecosystem. IntJ Food Microbiol 128:58– 66 (2008).
49 Kolb B and Ettre LS, Static Headspace-GasChromatography: Theoryand Practice (2nd edn). Wiley, Hoboken, NJ (2006).
50 Lee K and Lee Y, Effect of Lactobacillus plantarum as a starter onthe food quality and microbiota of kimchi. Food Sci Biotechnol 19:641– 646 (2010).
51 Ruiz-Barba JL, Cathcart DP, Warner PJ and Jimenez-Dıaz R, Use of Lactobacillus plantarum LPC010, a bacteriocin producer, as a starterculture in Spanish-style green olive fermentations. Appl EnvironMicrobiol 60:2059–2064 (1994).
52 Bevilacqua A, Perricone M, Cannarsi M, Corbo MR and Sinigaglia M, Technological and spoiling characteristics of the yeast microfloraisolated from Bella Di Cerignola table olives. Int J Food Sci Technol 44:2198–2207 (2009).
53 Arroyo-LopezFN, Querol A, Bautista-Gallego J and Garrido-FernandezA, Role of yeasts in table olive production. Int J Food Microbiol 128:189–196 (2008).
54 Nisiotou AA, Chorianopoulos N, Nychas GJE and Panagou EZ,Yeast heterogeneity during spontaneous fermentation of black
Conservolea olives in different brine solutions. J Appl Microbiol 108:396– 405 (2010).
55 Chang JY and Chang HC, Improvements in the quality and shelf-lifeof kimchi by fermentation with the induced bacteriocin-producingstrain,Leuconostoccitreum GJ7asastarter. JFood Sci 75:M103–M110(2010).
56 Perricone M, Bevilacqua A, Corbo MR and Sinigaglia M, Use of Lactobacillus plantarum and glucose to control the fermentationof Bella di Cerignola table olives, a traditional variety of Apulian
region (southern Italy). J FoodSci 75:M430–M436 (2010).57 MithenRF,DekkerM, Verkerk R,RabotS andJohnsonIT,The nutritionalsignificance, biosynthesis and bioavailability of glucosinolates inhuman foods. J Sci FoodAgric 80:967–984 (2001).
58 Landaud S, Helinck S and Bonnarme P, Formation of volatilesulfur compounds and metabolism of methionine and othersulfur compounds in fermented food. Appl Microbiol Biotechnol 77:1191–1205 (2008).
59 Oude-Elferink SJWH, Krooneman J, Gottschal JC, Spoelstra SF, FaberF and Driehuis F, Anaerobic conversion of lactic acid to aceticacid and 1,2-propanediol by Lactobacillus buchneri . Appl EnvironMicrobiol 67:125–132 (2001).
J Sci Food Agric 2013; 93: 3352–3361 c 2013 Society of Chemical Industry wileyonlinelibrary.com/jsfa


















