
Biología reproductiva mariposa - biblio.uabcs.mxbiblio.uabcs.mx/tesis/TE1297.pdf · especialmente...
45
11 1 I UNIVERSIDAD AUTNOMA DE BAJA CALIFORNIA SUR `REA INTERDISCIPLINARIA DE CIENCIAS DEL MAR DEPARTAMENTO DE BIOLOG˝A MARINA Biologa reproductiva de la raya mariposa Gymnura marmorata Cooper 1863 en Baha Almejas BC S MØxico TESIS DE LICENCIATURA Que para obtener el ttulo de Bilogo Marino PRESENTA JORGE DAVlLA ORTIZ La Paz Baja California Sur Abril de 2002
Transcript of Biología reproductiva mariposa - biblio.uabcs.mxbiblio.uabcs.mx/tesis/TE1297.pdf · especialmente...

111
I
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
`REA INTERDISCIPLINARIA DE CIENCIAS DEL MAR
DEPARTAMENTO DE BIOLOG˝A MARINA
Biología reproductiva de la raya mariposaGymnura marmorata Cooper 1863
en Bahía Almejas B C S MØxico
TESIS DE LICENCIATURA
Que para obtener el título de
Biólogo Marino
PRESENTA
JORGEDAVlLA ORTIZ
La Paz Baja California Sur Abril de 2002
051 7251
tJtV T
1
L 7J
çr
r zo
kt
I0 fi J
`A
tiijl li i
J A P1If
ti
Gymnura marmorataCooper 1863

Este trabajo esta dedicado
especialmente a YHWH
a mis padres Jorge y Ma del Carmen
a mis hennanas Diana y Brenda
Gracias por su paciencia y amor
Los amo

AGRADECIMIENTOS
Agradezco especialmenteatoøo mi comitØ revisor detesis al Dr Carlos Villavicencio al Dr Eduardo
Balart al B M Emelio Barjau al B M Everardo Mariano y al Dr Sergio Flores gracias por su apoyo
moral económico e intelectual
De igual manera agradezco infmitamente las atenciones recibidas en el campo pesquero Puerto
Viejo porparte delas familias SÆnchez y Escobar por las cuales esta tesis pudo llevarseacabo
Mis mas sinceras gracias al Laboratorio de Elasmobranquios y a todos sus integrantes Carlos
Everardo CarolinaJeanette AIfredo Ana LÆzaro Norma Arturo Lenin Jonathan Jorge Guadalupe
Sergio Luz Alberto EDT gracias por compartir su espacio sentimientos conocimientos opiniones y
cariæo Ya las siguientes generaciones de incautos les digo que es el mejor lab de la UA ß C
SMisreconocimientosa todos losmaestros que tuve enlacarrera gracias por todo amis
compa fierosde generación ami familia que aunque dedico este trabajo a ellos no tengo palabras de
cómo agradecerles todo su amory paciencia los amo mucho alprofe Carlos asu esposa Lety quesiempre fueron comomi familia en ellab gracias por todo su apoyo entodos aspectos ami familia aquí en la
Paz Paty Edgar AIine Priscilla Patsy que merecibieron cuando comenzó la aventura yatodos mis
hemla nos del C F COthoniel Mara Juvenal I1Øctor Silvia Nena Alicia Lucero Juan Gabriel
Alan Emmanue Luis Esteban Marisela Iris Israel IvÆn Elvira Etc Etc Etca todos mucllas
gracias Gracias al Doc Everporenseæarme la verdad ylo bueno que hay en todas las cosas gracias
portu paciencia cariæo amistad crØeme que no importa donde estemos siempre estarØ contigoy
recuerda quenosveremos en Sión Ami compadre al pelosaNonna yal sensei Lazarin gracias por todo chavo
s yamis anœgos Juan Eduardo Roberto Vidal Javiery Lula Øchenle galleta a todo loque
haganMi reconocimiento amory gratitudauna personita que estuvo ami lado durante todami
carrera que fuemi brazo derechoen muchas cosasy que espero vermi nombre enlos agradecimientos de su
tesis algœn día Gracias por todo Zuleyno importan nuestros destinos siempre ocuparÆs un lugar muyespecial enmi
corazón yalprimer taxónomo sobre la tierra Formó pues Dios dela tierra toda bestia del campo y
toda avede los cielosy tnijolasaAdÆm para queviese como les había de llamar y todo lo que AdÆm
llamó a los animales vivientes ese essu nombre Gen
219

íNDICE GENERAL PAG
RESUMEN 1
INTRODUCCIÓN 3
Antecedentes 3
Objetivos 4
`REA DE ESTUDIO 5
METODOLOG˝A 6
A Distribución temporal 6
B Biología Reproductiva 6
C DimorfIsmo sexual 8
RESULTADOS 9A Distribución temporal 9
Proporción de sexos 10
Composición por tallas 10
B Biología Reproductiva 11
Tallas de primera madurez 11
OvogØnesis 11
Fecundidad 11
Crecimiento embrionario 12
C Dimorfismo sexual 12
AnÆlisis morfomØtrico 12
`ngulos de apertura del disco 12
Pliegues caudales 12
Coloración 12
DISCUSIONES 14
A Distribución temporal 14
Proporción de sexos 15
Composición por tallas 15
B Biología Reproductiva 16
Tallas de primera madurez 16
Ciclo reproductivo 16
Fecundidad 18
Talla de nacimiento 18
C Dimorfismo sexual 19
CONCLUSIONES 22
LITERA TURA CITADA 23

˝NDICE GENERAL DE TABLAS PAG
Tabla 1 Aæos y meses de la captura comercial de G marmora a en Puerto Viejo 6
B C S
Tabla 2 Frecuencia de machos y hembras densidad relativa No Orgmes 100 9
por mes y total del muestreo de G marmora a
Tabla 3 Proporción sexual hembras por macho H M de G marmora a por mes y 10
total del muestreo
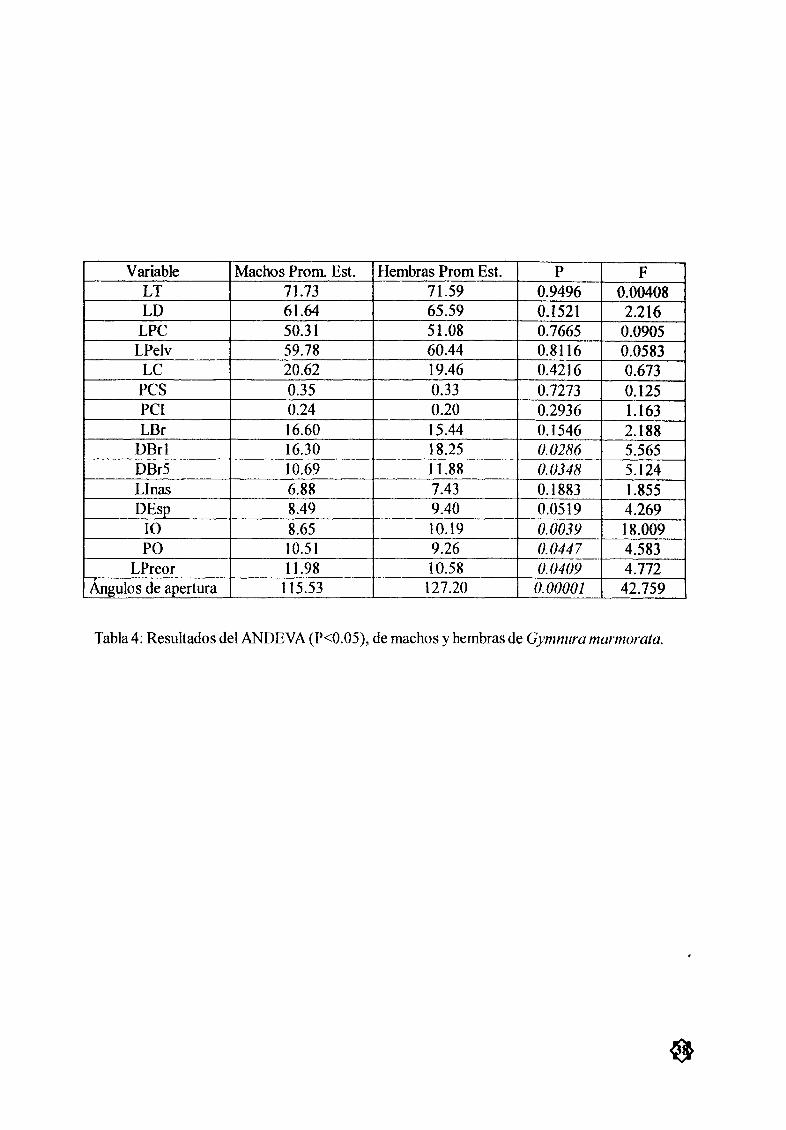
Tabla 4 Resultados del ANDEVA P 0 05 de machos y hembras de 38
G marmora a
˝NDICE GENERAL DE FIGURAS PAG
Figura 1 Localización del Ærea de estudio Puerto pesquero Puerto Viejo en el 26
complejo lagunar Bahía Magdalena Almejas
Figura 2 Medidas tomadas para la comparación morfomØtrica de Gymnura 27
marmora a
Figura 3a Tallas promedio mÆximas y mínimas por mes con referencia a la talla 28
de primera madurez para los machos
Figura 3b Tallas promedio mÆximas y mÚlimas por mes con referencia a la talla 28
de primera madurez para las hembras
Figura 4 a y b Distribución modal de cada frecuencia de tallas para hembras a y 29
machos b con respecto a la talla de primera madurez
Figura 5 Frecuencia relativa por intervalo de talla para machos y hembras 30
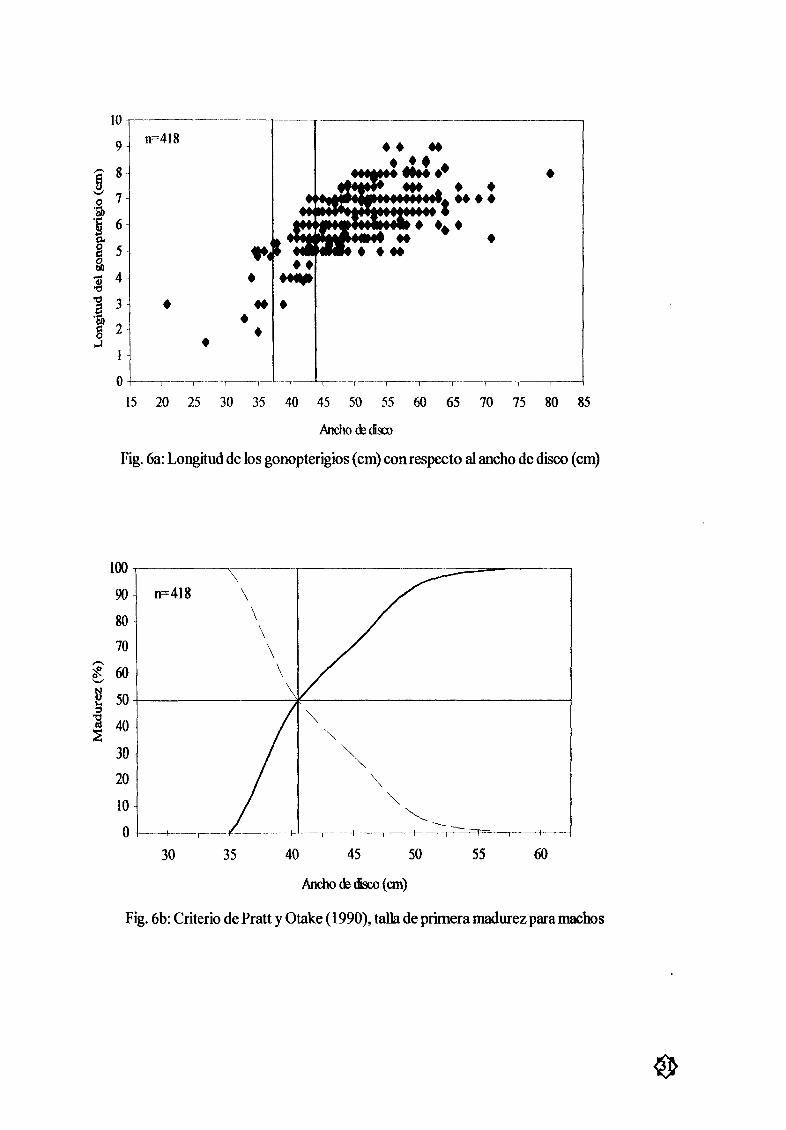
Figura 6a Longitud del gonopterigio cm con respecto al ancho de disco cm 31
Figura 6b Criterio de Pratt y Otake 1990 talla de primera madurez para machos 31
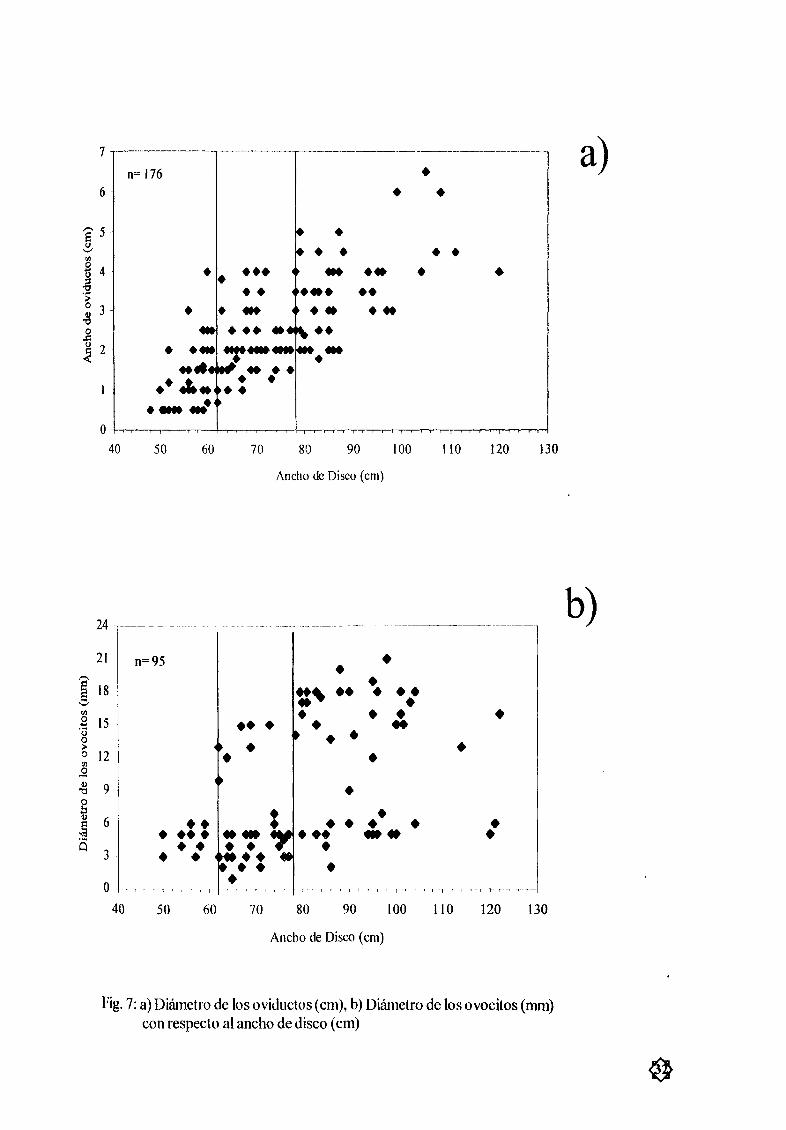
Figura 7 a DiÆmetro de los oviductos cm b DiÆn1etro de los ovocitos mm con 32
respecto al ancho de disco cm
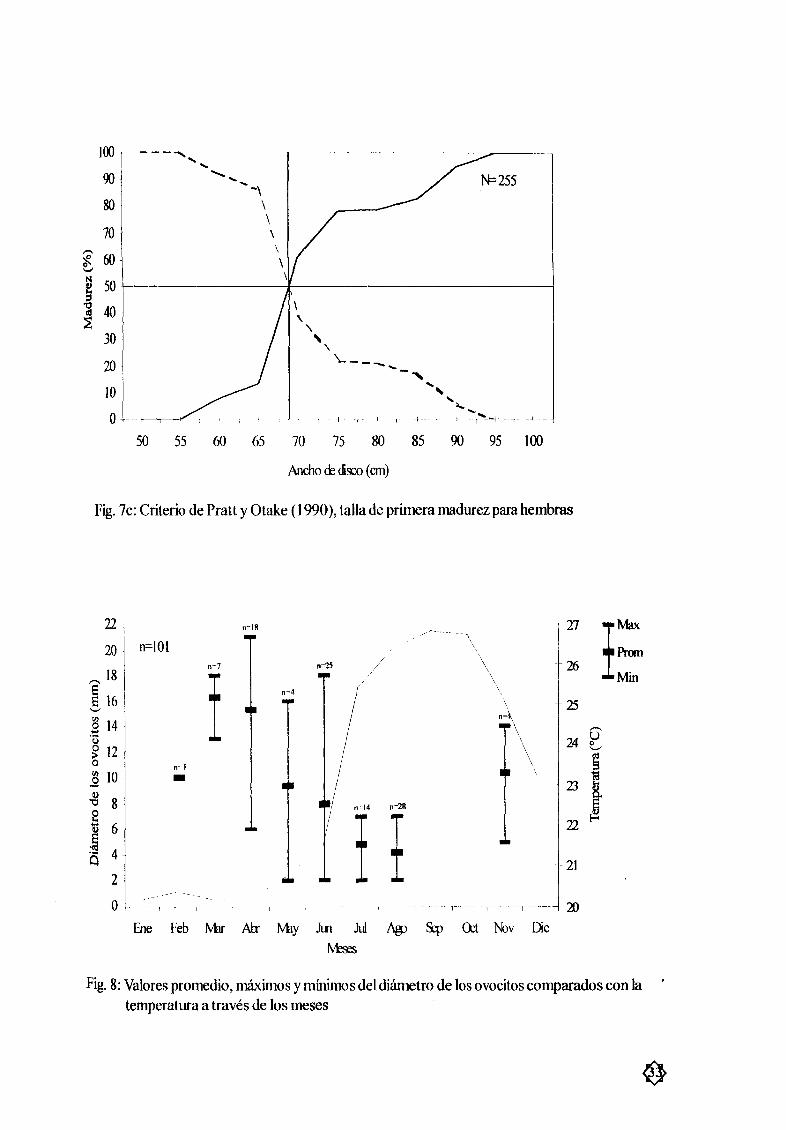
Figura 7c Criterio de PraU y Otake 1990 talla de primera madurez para hembras 33
Figura 8 Valores promedio mÆximos y mínimos del diÆmetro de los ovocitos 33
comparados con la temperatura a travØs de los meses
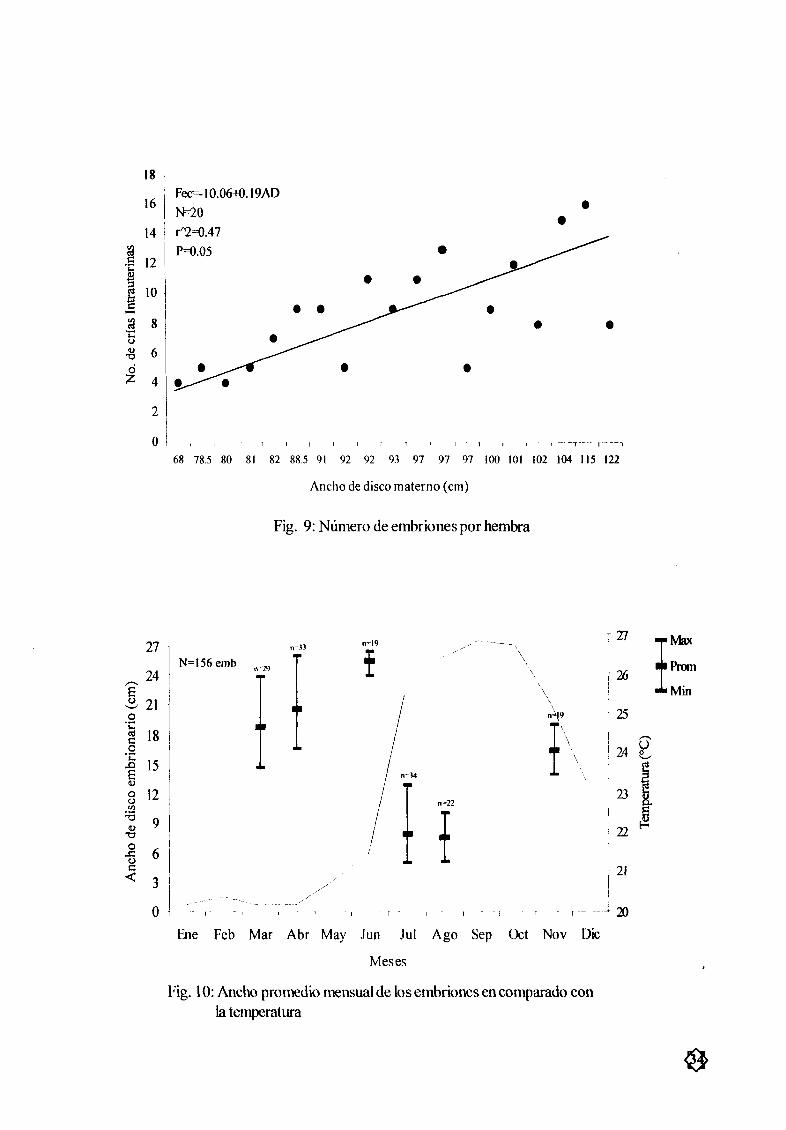
Figura 9 Nœmero de embriones por hembra 34
Figura 10 Ancho promedio mensual de los embriones comparado con la 34
temperatura
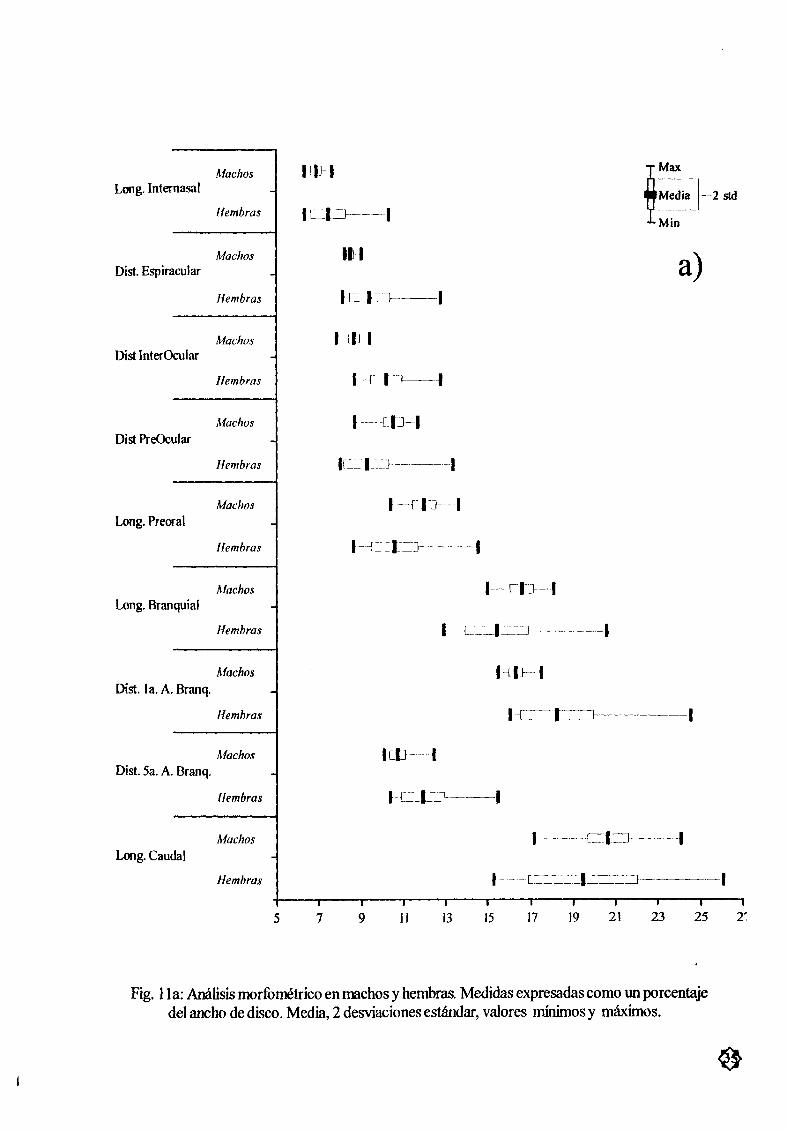
Figura 11 a AnÆlisis morfomØtrico en machos y hembras Medidas expresadas 35
como un porcentaje del ancho de disco Media 2 desviación estÆndar
valores mínimos y mÆximos
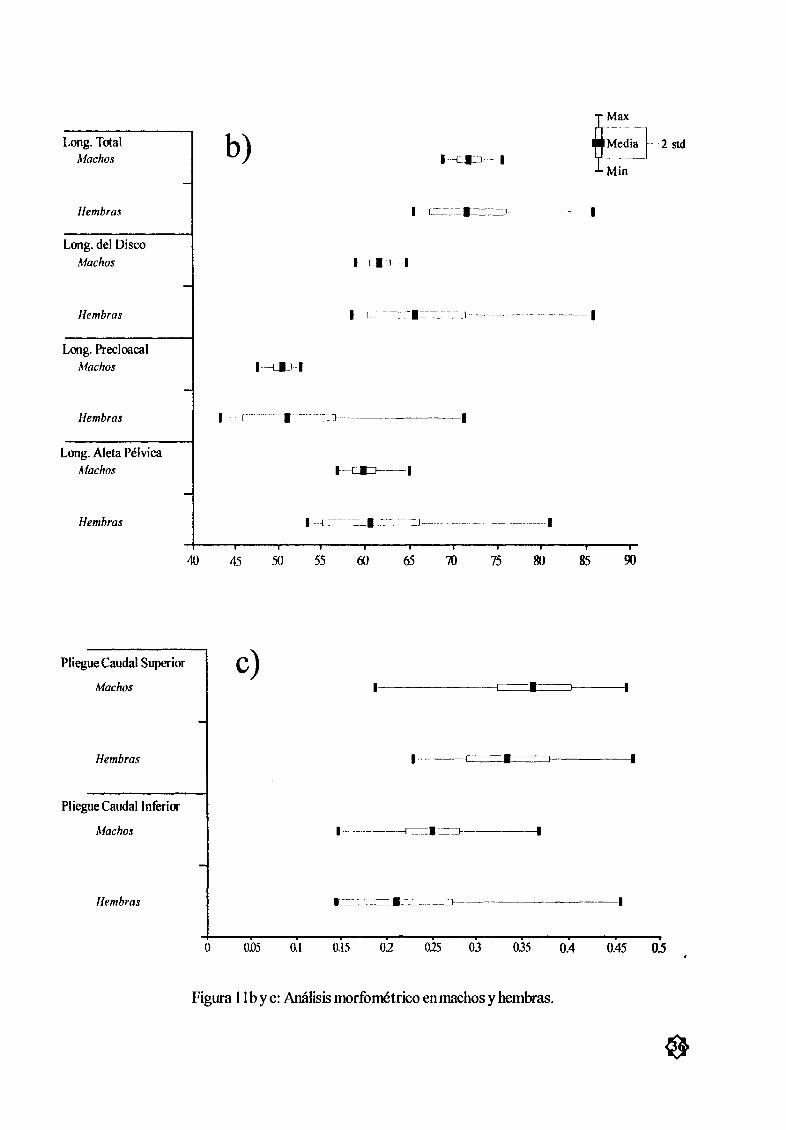
Figura I lb Y c AnÆlisis morfomØtrico en machos y hembras 36
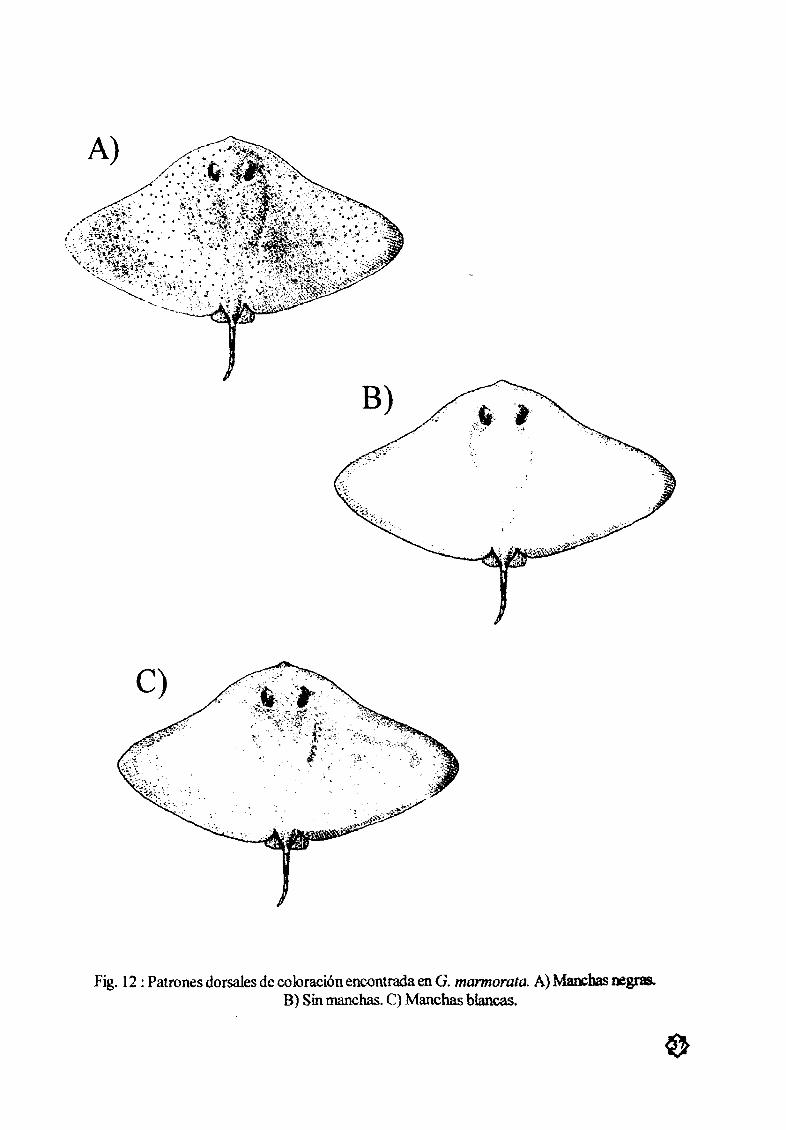
Figura 12 Patrones dorsales de coloración A Manchas negras B Sin manchas 37
C Manchas blancas

RESUMEN
La raya mariposa Gymnura marmorata fue descrita en la localidad de San Diego California
con una distribución que va desde Punta Concepción California hasta el norte de Perœ enel Pacífico
Occidental Pertenece a la familia Gymnuridae y en particular se hadestacado por presentar problemas de
sinonœnia con G crebripunctata sin embargo la información acerca de su biología y las características
utilizadas para su identificación no son claras Desde los aæos de 1992 hasta 1999 se observó la presenciade G marmorata enel complejo lagunar Bahía Almejas localizado en las costas del Pacífico Occidental
deBaja California Sur MØxico Este lugar ha sido reconocido por la presencia de otras 13 especies de
tiburones y 12 de rayas que penetran ala bahía con fines de reproducción alimentación y o protección El
objetivo general es Describir la biología reproductiva de la raya mariposa G marmorata en Bahía Alme
jas B C S MØxico así como establecer su estatus taxonómico Por medio de la captura comercial
realizada en un campo pesquero llamado Puerto Viejo en esta zona se obtuvieron datos acerca de la
composición de tallasy sexos estado de madurez sexual ovogØnesis fecundidad crecimiento embriona
rio morfometría del cuerpo y coloración analizando untotal de 1048 organismos de los cuales 610 fueron
hembras y 438 fueron machos La frecuencia en la captura presentó un aumento durante los meses de
marzo aagosto con un mÆximo en el mes de junio mientras que en los meses de septiembre a febrero su
captura fue casi nula Durante los mesesde marzo rrnyo y julio la proporción sexual demachos fue mayor
que la de hembras con unaproporció nsexual embrionaria de I 15 machos por hembra Las tallasmínimas
y mÆximas para los machos fueron de 21 y 91 5 cmde AD respectivamente con un promedio de 5 I 03 cm
AD mientras que para las hembras la menor fue de 25 5 cm de AD y la mayor de 122 cmAD con una
media de 6842 cm AD La talla de primera madurez para los machos fue de 4 I cm AD y para las
hembras de 62 cmAD El desarrollo de los ovocitos mostró los mayores promedios durante marzo abril y
los menores durantejulio agosto con unaprobable ovulación durante los mesesde abril ajunio ElnÚl1lero
de embriones In u ero por hembra varió de 4 a 16 siendo mas frecuentes entre 5 y 9 Los mayores
tamaæos embrionario sse observaron durante los meses demarzo abril y junio y los mas pequeæos enjulio
agosto
El anÆlisis morfomØtrico entre machos y hembras mostró por medio de una prueba ANDEVA
P 0 05 que tansolo 6 de las 18 características tomadas presentaron diferencias significativas incluyendopliegues caudales y Ængulo de apertura del disco En cuanto a la coloración de los organismos esta no
presentó diferencias por sexo talla o Øpoca del aæo con patrones de color similares entre embriones
juveniles y adultos Se puede decir que la presencia de machos y hembras de G marmora aen Bahía
Almejas se encuentra aparentemente regulada por la temperatura del agua con ciclos de visita an les
Las hembras fueron mas abundantes que los machos siendo estos œltimos mas pequeæos en talla La
mayoría de losmachos y hembras encontrados en la zonaeran maduros sexualmente con un estado de
ł

embarazo y desarrollo de ovocitos avanzado en las hembras y abundante semen en los machos Su
periodo de gestación es anual con una duración de 9 a12 meses y una expulsión embrionaria durante losmeses primavera verano Su fecundidad es de 2 a 9 embriones por hembra y la talla de nacimiento se
calculó entre los 21 y 265 cmAD El dimorfismo sexual secundario encontrado es comœn en otras
especies del gØnero por lo que debe considerarse aG crebripunctata como taxón invÆlido
ł

INTRODUCCIÓN
El grupo de los Elasmobranquios o peces cartilaginosos incluye WJa serie de organismos deno
minados comœnmente como tiburones y rayas Los peces batoideo s o rayas son derivados del grupo
NeoseIachii de tiburones que incluye de 494 a572 especies comprendidas en 5 órdenes 18 familias y 59
gØneros Compagno 1990 Dentro de las rayas Rajifonnes Myliobatoidea NeIson 1994 se encuentra
la fàmilia Gynmuridae conocida comœnmente con el nombre de rayas mariposa o tortillas McEachran
y Notarbartolo 1995 Esta familia se distribuye solo en la franja tropical y templado cÆlida del OcØano
AtlÆntico MarNegro India y OcØano PacífICO prefiriendo ambientes estuarinos de lagunas costeras y
bocas de dos BigeIow y Schroeder 1953 NeIson 1994 McEachran y Notarbartolo 1995 Esta familia
comprende dos gØneros Aetoplatea Müller y Henle 1841 con una pequeæa aleta dorsal en la mitad de
la cola y Gymnura van Hasselt 1823 sin dicha aleta dorsal El nœmero total de especies varía de lOa 12
dependiendo del autor Bígelow y Schroeder 1953 Nelson 1994 Hamlett 1999
En lo que respecta a las costas de MØxico el gØnero Gymnura es el œnico presente con tres
especies En el Pacífico se encuentran G marmorata Cooper 1863 Y G crebripunctata Peters 1869
Y en el AtlÆntico G micrura Bloch y Schneider 1801 aunque no se descarta la presencia de G altavela
Linnaeus 1758 en esta zona Castro Aguirre y Espinosa PØrez 1996
Inicialmente G marmorata fue descrita en la localidad de San Diego California con el nombre
específico de Pteroplatea marmorata Presenta una distribución que va desde Punta ConcepciónCalifornia hasta el norte de Perœ y se caracteriza por presentar un disco mÆs ancho que largo 1 5 2 0
veces ojos pequeæos ode igual tamaftoque los espirÆculos poros mucosos sobre la superficie del disco
y región branquial narinas separadas por un puente dØmøco piso de la boca sin papilas y aberturas
branquiales pequefias de las cuales la primera es aproximadamente igual al diÆmetro horizontal del ojoBeebe y Tee Van 1941 Castro Aguírre 1965 Castro Aguirre y Espinosa PØrez 1996 Esta especie
se ha destacado por presentar problemas de sinonímia con G crebripunctata Peters 1869 especiedescrita en MazatlÆn MØxico inicialmente conel nombre de Pteroplatea crebripunctata Esta œltima
presenta unadistribución que comprende desde la costa sur occidental de la Peninsula de Baja California
incluyendo el Golfo de California hastaEcuador Las características utilizadas para su identificación son
la presencia de un pliegue dØmøco en la parte superior Castro Aguirre yEspmosa PØrez 1996 o infe
rior McEachrany Notarbartolo 1995 de la cola nunca ambos una longitud preocular mayor que la
interocuIar y una forma delcuerpo mÆs triangular que G marmorata sin embargo la información acerca
de su biología es nula
ł

El primer autor enproponer esta sinonímia fue Breder 1928 basÆndose en los pliegues caudales
coloración y tamaæo de los organismos llegó a la conclusión de que eran similares en las dos especies A
partir de entonces autores como Beebe y Tee Van 1941 Bigelow y Schroeder 1953 Castro Aguirre
y colaboradores 1965 Y 1970 BeltrÆn FØlix el al 1986 Villavicencio Garayzar 1993 Y 1995b Y
Villavicencio Garayzar y Abitia CÆrdenas 1994 consideran aG marmorata como la œnica especie del
gØnero enel Pacífico Mexicano
En aæos mÆs recientes McEachran y Notarbartolo 1995 Castro Aguirre y Espinosa PØrez
1996 y De la Cruz Agilero 1997 redefmen aG crebripunctata como taxón vÆlido aportando claves
taxonómicas para su identificación fundamentadas en características de la longitud rostra pliegues cauda
les y su dístribución espacio temporal
La falta de datos biológicos como reproducción alimentación comportamiento y la coincidencia
de estas supuestas especies en un mismo lugar y tiempo ponen en duda su validez
Conbase enestos antecedentes se plantea el siguiente objetivo general
Describir la biología reproductiva de la raya mariposa Gymnura marmorata en Balœa Almejas B C
S MØxico y establecer su estatus taollÓmico
Objetivosparticulareso metas
Determinar la composición de tallas mensual y proporción de sexos
Determinar la talla de primera madurez para machos y hembras
Describir el periodo reproductivoDeterminar el periodo de gestación y talla de nacimiento
Defmir las diferencias morfológicas entre machos y hembras
Establecer el estatus taxonómico de G marmorata
ł

`REA DE ESTUDIO
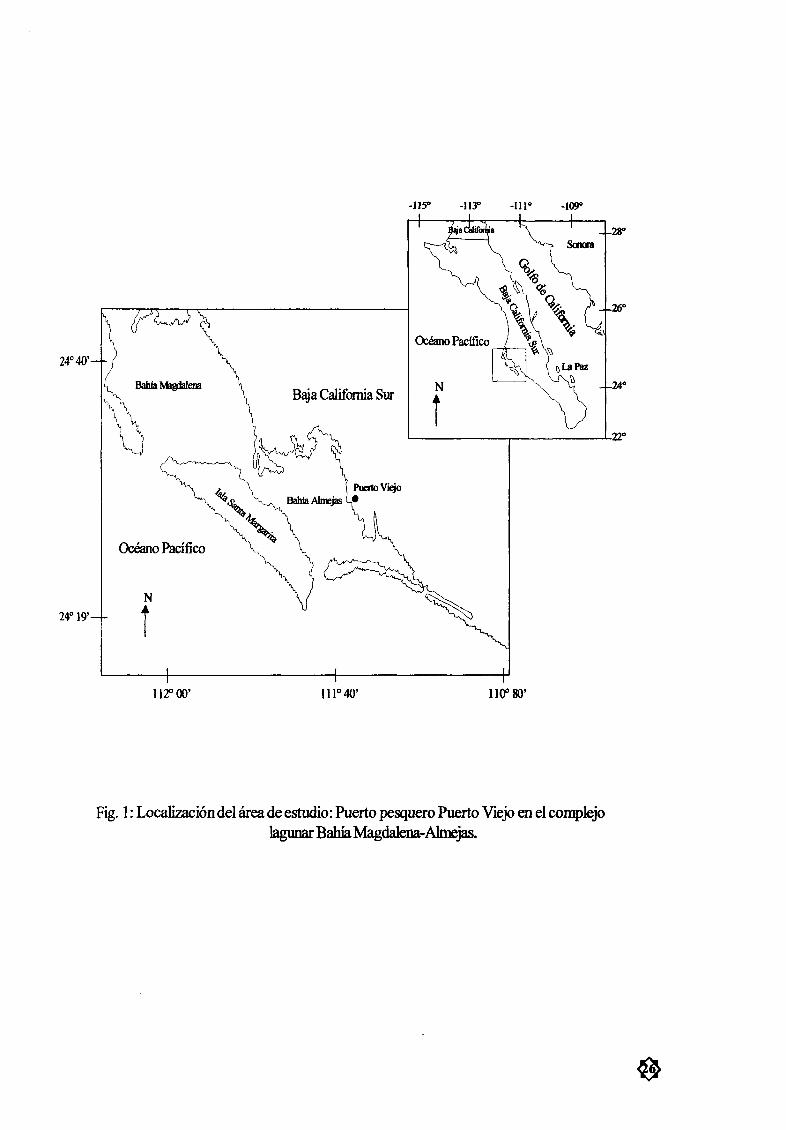
Los muestreos biológicos se realizaron en el campo pesquero de Puerto Viejo Fig 1 localizado
entre los 240 17 Y 240 37N Y 1 I 1 o 22 y 1110 57 W Mariano 1997 en el complejo lagunar Bahía
Magdalena Ahnejas de la costa occidental de la Península de Baja California ViIlavicencio Garayzar2000 Este complejo se encucntra fonnado y delimitado por la presencia de dos islas Isla Magdalena al
norte e Isla Margarita al sur Estas islas formanasu vez tres 7vnas principales1 Zona norte confonnada por esleros lagwlaS y canales con wllllÆximo de profwldidad de 3 5m
2 Zona central con Ula extensión de 883 Km2 conocida con el nombre deBahía Magdalena y
que se conecta con el ocØano Pacífico por medio de un canal denominado La Gaviota de 2 5 km de
longitud y una boca profunda entre las dos islas
3 Zona sureste COl W aextensión aproximada de 370 km2 denominada Ballía A1mcjas Esta Ærea
se encuentra limitada al sur por una zona de manglares llamada Isla Creciente y un canal de mareas
conocido como Canal de Rehœsa `lvarez el al 1975 ViJlaviccncio Garayzar 2000
Bahía Magdalena estÆ influenciada por doscorrientes principales la corriente de California en
primavera verano y la contracorriente costerade California enotoæo inviemo Lynn y Simpson 1987 La
productividad de la 7vnaes propiciada por las surgenciai que durante todo el aæo se presentan Bakun
1973 La temperatura del agua va de los 14 C durante mayo y junio hasta los 290C en los meses de
septiembre y octubre mientras que la temperatura anlbiental varía desde los 120C en diciembrey enero y
hasta los 300C enjulio y agosto COADS 1994
La bahía engeneral presenta poca incidencia de ciclones o tornlCntas tropicales y poca precipitación pluvial con dos etapade lIuviauna dejulio a octubre y otra de noviembre a febrero con un tiempode sequía entremarw y junio Salinas Zavala et al 1990 La mayor salinidad se registra en el noroeste
mientras que los niveles de oxígeno no varían significativamente durante el aæo yelpH se intensifica hacia
el interior de la Bahía `lvarez et al 1975
ZoogeogrÆficamente Ball˝a Magdalena representa el limite entre las provincias de San Diego y
Mexicana lo que ha favorecido al establecimiento de una gran riqueza y abwldancia de flora y fauna en el
Ærea Briggs 1974 Entre las especies de elasmobranquios encontradas 13 son de tiburones y 12 dc
rayas Villavicencio Garayzar y Abitia CÆrdenai 1994 de las cuales NOItine entemedor Rhinohatos
produclus Zapterix exasperata Carcharhinus ohscllrllsy Dasyatis longlls usan la bahía para alimen
tarse reproducirse y protegerse mientras que Dasyatis hrevis y Ileterodol1tlls mexical111s son espec ies
residentes del lugar Villavicencio Garayzar 2000
ł

METODOLOG˝A
A DISTRIBUCIÓN TEMPORAL
Los organismos analizados se obtuvieron de la pesca ribereæa realizada con redes agalleras de
monoftlarnento y multiftlamento luz de malla de entre 10 Y 30 cm tendidas por la mar1ana y revisadas al
día siguiente Tanto los meses como los aæos de muestreo se muestran en la tabla l
AIÌo Meses
1992 Enero a diciembre
1993 Marzo a agosto octubre y noviembre
1994 Marzo a junio agosto y octubre
1998 Mayo aagosto
1999 Junio a agosto
Tabla l Aæos y meses de la captura comercial de Puerto Viejo B C S
Una vezen el campo pesquero se registró el nœmero de organismos obtenidos como indicador de
abundancia ademÆs de su peso medidas y sexo lo que permitió realizar las observaciones de forma
coloración y reproducciÓn
B BIOLOG˝A REPRODUCTIVA
En las hembras se observó la flUlcionalidad de los ovariosy oviductos tomando el diÆmetro de los
ovocitos y ancho de los oviductos en su parte media ademÆs de observaciones sobre la presencia de
embrioneshuevos feClmdadoso semen en los œteros En losmachos se midió la longitud de los gonopterigiosdesde la parte anterior de la cloaca hasta la parte distaAdemÆs se realizaron observaciones del grado de
calcificación y la presencia de semen oprimiendo la parte anterior de las aletas pØlvicas para comprimir la
vesícula seminal
Con base en estas observaciones se clasificó a los organismos en categorías de madurez de acuer
do con Smithy Merriner 1986 Y Maríano 1997 considerando a los organismos como
ł

1 INMADURO Cuando las hembras presentan ovarios flÆcidos poco desarrollados y los oviductos
delgados yelongados En los machos los gonopterigios se encuentran pequeæos y flÆcidos sin la presen
cia de semen
2 MADURANDO En las hembras el ovario esta semi desarrollado con ovocitos pequeæos pero visibles
de color amarillo œteros mÆs desarrollados y con poco trofonemata vellos uterinos En machos los
gonopterigios no estÆn completamente calcificados pero muestran un alto grado de desarrollo sin la
presencia aœn de semen por lo que aÚn no se reproduccn
3 MADUROS Enlos machos los gonopterigios ya estÆn completamente calcificados y existe la presen
cia de semen en los conductos deferentes excepto despuØs de la cópula En las hembras los ovocitos
son de color amarillo producto de la vitelogØnesis con los œteros bien desarrollados y presencia de
tro fonemata
La talla de primera madure se estimÓ cuando el 50 de los organismos son sexualmentc maduros
en la frecuencia de clase Pratt y Otake 1990
La actividad ovÆrica se estahleciÓ al graficar la variaciÓn deldiÆnletro de los ovocitos por mes lo
que permitió estimar el periodo de ovulaciÓn de la especie
La fecundidad uterina se detelll1inó por medio dcl conteo de embrioncs in ulero por hembra Para
probar la relaciÓn entre la talla maternay el nœmero de embriones se realizó una regresiÓn lineal Y a bx
Simpfedorfer 1992 Downton 1996 donde
y Fecundidad
x Ancho de disco cm
a y b Constantes
En las ocasiones en que se presentaron crías Østas se midieron ancho de disco y longitud total y
se les determinó el sexo AdemÆs se calculó el promedio de la talla embrionaria mensual para obtener
información del periodo de gestación temporada de desarrollo de crías y estimar la talla de expulsión
ł

C DIMORFISMO SEXUAL
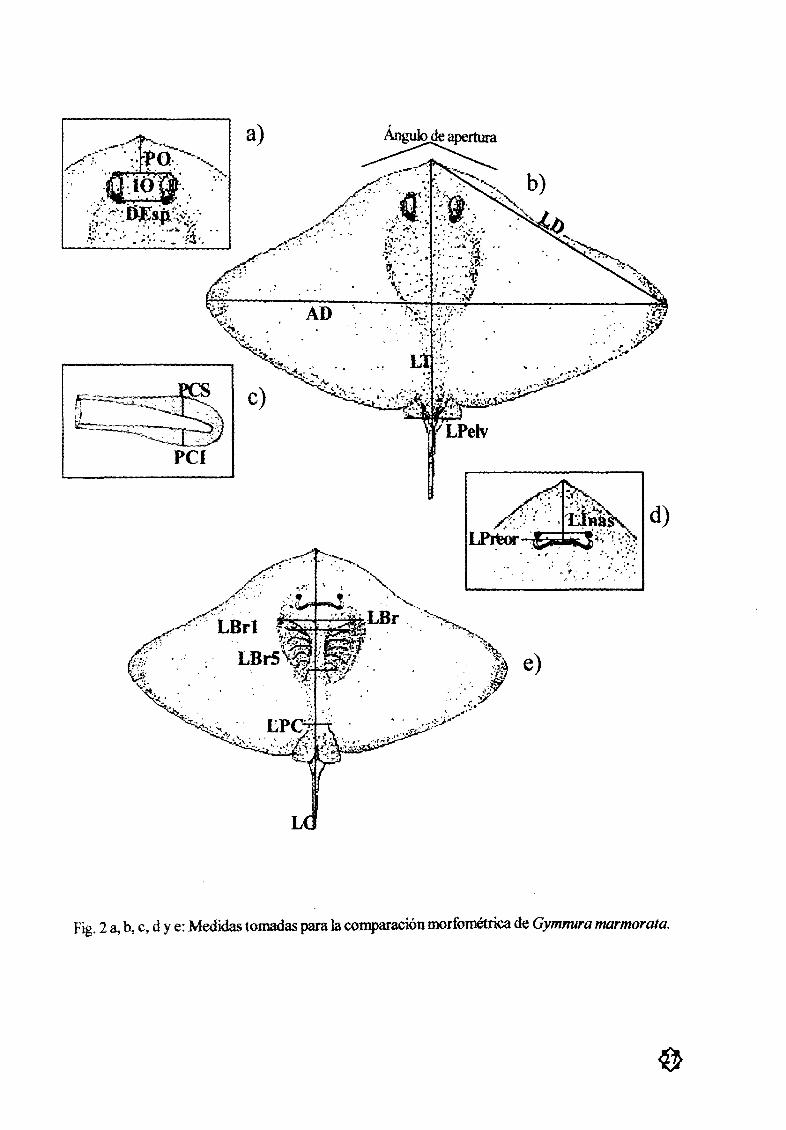
Para conocer las diferencias morfológicas entre machos y hembras se tomaron un total de 16
medidas Fig 2 de acuerdo con Bigelow y Schroeder 1953 WaUace 1967 Y Hubbs e lshiyama1968 las cuales son
Longitud Total LT cumprendida desde la punta del hocico hasta la punta de la cola
Ancho de Disco AD la medida mas grande entre los extremos de las aletas pectoralesLongitud delDisco LD distancia comprendida entre la punta del hocico y el margen posterior de
las aletas pectoralesLongitud Precloacal LPC desde la punta del hocico hasta el iniciu de la cloaca
Longitud de la Aleta PØlvica LPelv desde la parte anterior del hocico hasta la parte posterior de
las aletas pØlvicas
Longitud Caudal LC desde la punta de los pliegues caudales hasta el inicio de la cloaca
Pliegue Caudal Superior PCS la mayor anchura del pliegue caudal superiorPliegue Caudal 1nfcrior P I la mayor anchura del pliegue caudal inferior
Longitud Branquial LBr desde el extremu anterior delhocico hasta una línea imaginariaque une
las primeras abert uras branquialesDistancia entre las Primeras Aberturas Branquiales DBr 1 la separación entre los extremos inter
nos de las primeras aberturas branquialesDistancia entre las Quintas Abcrt mas Branquiales Dßr5 la separación entre los extremos internos
delas quintas aberturas branquiales Longitud
Internasal Unas la menor distancia entre los nostrilos Distancia
cntre EspirÆculos1 1 sr la menor distanciaentre los espirÆculos Longitud
Intcrocular10el menor espacio entre las órbitas oculares Longitud
Preocular PO longitud del hocico desde su punta hasta elmargen frontaldelojo Longitud
Preoral LPreor desde la punta del hocico hasta la hendidura bucal ensu parte media tomada
conlaposición cerrada Una
vez tomada las medidas cada una fuØ dividida entre su ancho dediscoy multiplicada por100
para la comparación entre ellos Wallace 1967 Las observacionesdelos plicgues dØrnœcos caudales
se realizaron deacuerdoa Breder 1928 McEachran y Notarbartolo 1995 y Castro Aguirre
yEspinosa PØrez 1996 para determinarsu posición ventra dorsal oambasy su tamaæo La
observaciónde la coloracióndelos organismos tanto para los adultos COTTK para los embriones fue con
baseen loreportado por ßreder 1928 Y McEachranyNotarbartolo 1995 Otro aspectoa considerar
fue el Ængulo de apertura del disco comprendido desdela punta del hocico hasta el inicio dela
región ocular segœn BigelowySchroeder 1953Fig 2 ł
RESULTADOS
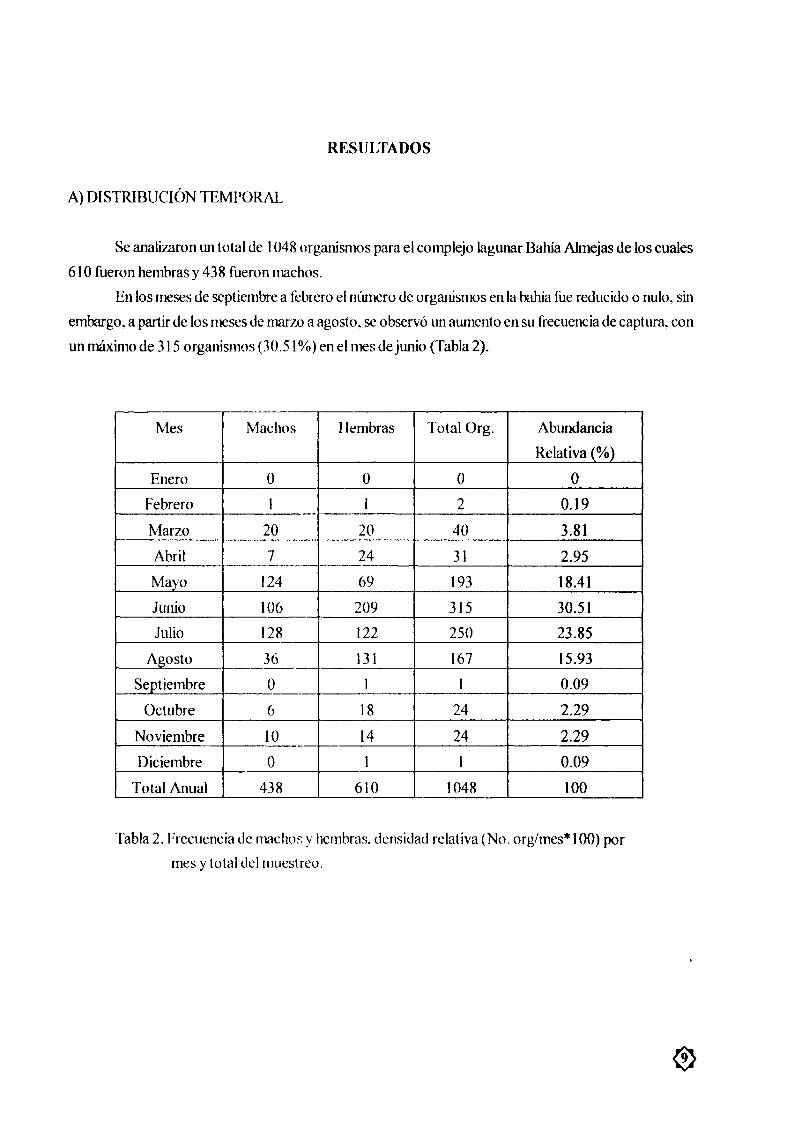
A DISTRIBUCIÓN TEMPORAL
Se analizaron tul total øe 1048 urgmœsmos para el complejo lagunar Barna Almejas de los cuales
610 fueron hembras y 438 fueron machos
En los meses de septiembre a febrero el nœmero de orgmœsmos en la balœa fue reducido o nulo sin
embargo a partir de los meses de marzoaagosto se observó un aumento ensu frecucncia de captura con
unmÆXinlO de 315 organismos 30 51 cn el mes de junio Tabla 2
Mes Machos J lembras Total Org Abundancia
Relativa
Enero O O O O
Febrero 1 1 2 0 19
Marzo 20 20 40 3 81f
Abril 7 24 31 2 95
Mayo 124 69 193 1841
Junio 106 209 315 30 51
Julio 128 122 250 23 85
Agosto 36 131 167 15 93
Septiembre O 1 1 0 09
Octubre 6 18 24 2 29
Noviembre 10 14 24 2 29
Diciembre O 1 1 0 09
Total Anual 438 610 1048 100
Tabla 2 Frecuencia de machos y hembras densidad relativa No orgmes 1 00 por
mes y total del muestreo
o
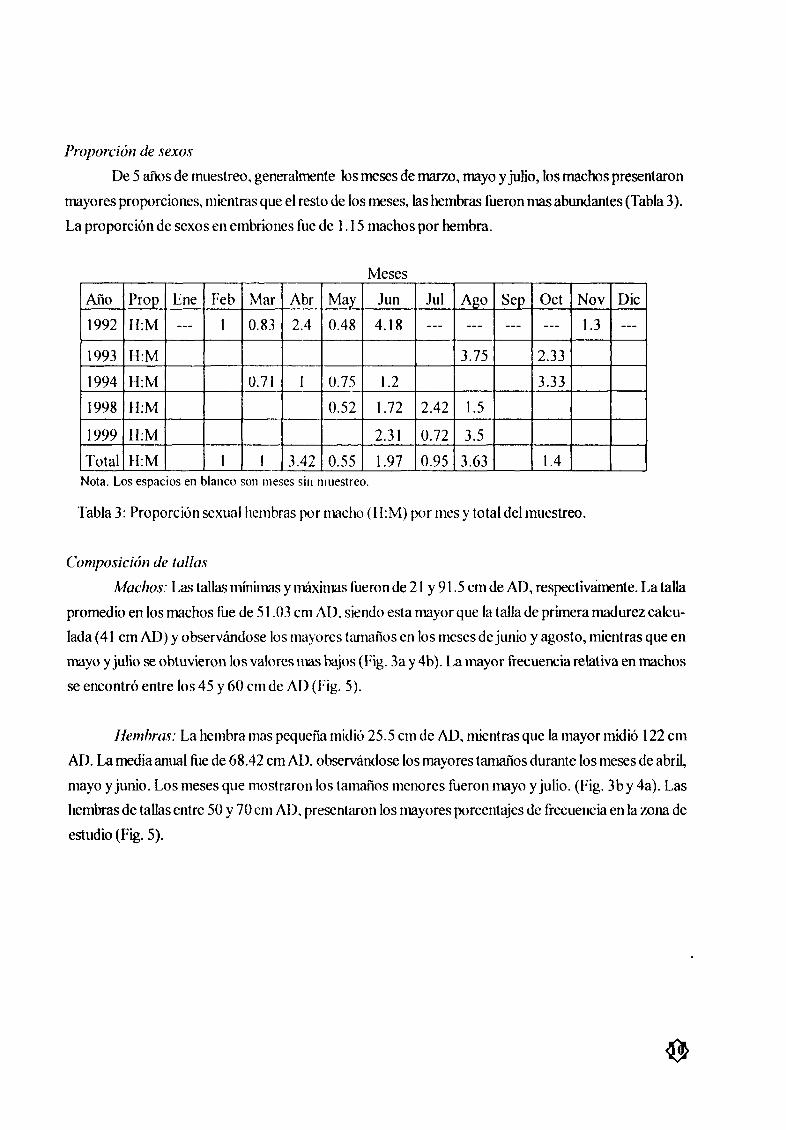
Proporción de sexos
De 5 aæos de muestreo generalmente los meses de marzo mayo y julio los machos presentaron
mayores proporciones mientras que el resto de los meses las hembras fueronmas abundantes Tabla 3
La proporción de sexos en embriones fue de 1 15 machos por hembra
Meses
Arlo Prop Ene feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
1992 I1 M 1 0 83 24 048 4 18 13
1993 H M 3 75 2 33
1994 H M 0 71 1 0 75 12 3 33
1998 H M 0 52 1 72 242 15
1999 H M 231 0 72 3 5
Total H M 1 1 342 0 55 197 0 95 3 63 14
Nota Los espacios en blanco son meses sin lIIuestreo
Tabla 3 Proporción sexual hembras por macho 11 M por mes y total del muestreo
ComposiciÓn de tallas
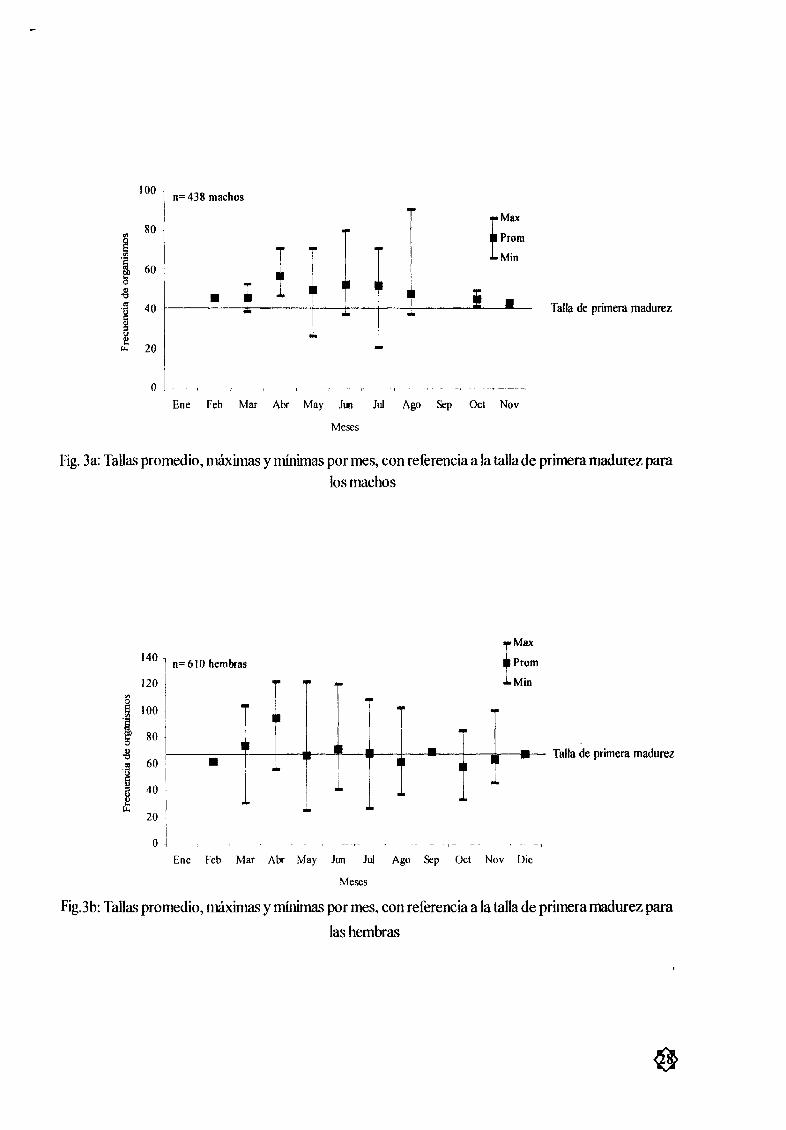
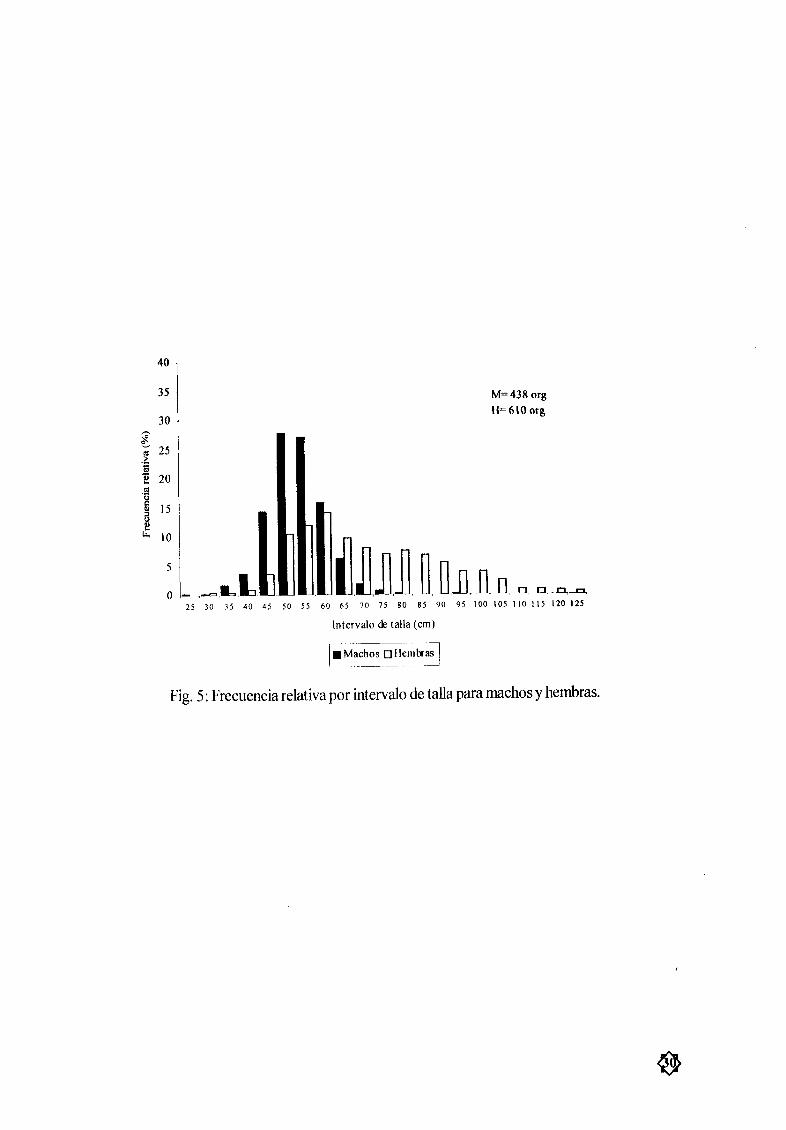
MacJlOs Las tallas mínimas y mÆximas fueron de 21 y 91 5 cm de AD respectivamente Latalla
promedio en los machos fue de 51 03 cm AD siendo esta mayor que la talla de primera madurez calcu
lada 41 cm AD y observÆndose los mayores tamaæos en los mesesde junio y agosto mientras que en
mayo y julio se obtuvieron los valores mas bajos Fig 3a y 4b La mayor frecuencia relativa en machos
se encontró entre los 45 y 60 cm de AD Fig 5
Hemhras La hembra mas pequeæa midió 25 5 cm de AD mientras que la mayor midió 122 cm
AD La mediaanual fue de 6842 cm AD observÆndose los mayores tamaæos durante los mesesde abril
mayo y junio Los meses que mostrarolllos tamarlos menores fueron mayo y julio Fig 3by 4a Las
hembras de tallasentre 50 y 70cm AD presentaron los mayores porcentajes de frecuencia en la zona de
estudio Fig 5

B BIOLOG˝A REPRODUCTIVA
Tallas deprimera madurez
Machos A partir de los 30cm de AD existió un marcado crecimiento de los gonopterigiosFig 6a encontrando poca calcificación hasta los 35 cm de AD DespuØs de los 37 cm AD se
observó la presencia de semen en las vesículas seminales con los gonopterígios semicalcificados Se
gœn el criterio de PraU y Otake 1990 la talla de primera madurez se estimó en 41 cm AD Fig 6b
cuando la presencia de semen en las vesículas seminales y el grado avanzado de calcificación de los
órganos copuladores fueron evidentes
Hembras Solo poseen el ovario izquierdo y los dosoviductos funcionales La relación entre el
ancho de disco con el diÆmetro de los oviductos Fig 7a y el diÆmetro de los ovocitos Fig 7b
reveló que antes de los 62 cm AD los oviductosy ovocitos se presentan pequeæos o indiferenciados
hembras completamente inmaduras DespuØs de esta talla se observa el desarrollo de ovocitos
vitelogØnesis trofonematay diÆmetros uterinos por lo que la talla deprimera madurez se calculÓ a los
68 cmAD Fig 7c
OvogØnesisEl desarrollo de los ovocitos Fig 8 roostró variaciones importantes durante el aæo con los
mayorespromedios durante marzo abril y los menores durante julio agosto mientrasque el resto del
aæo presentó un crecimiento constante Este desarrollo puede dividirse en tres partes principales La
primer parte se da durante los meses de mayo a agosto y esta definida por la presencia de ovocitos
muy pequeæos en los ovarios aprox 2 mmde diametro la segunda parte corresponde al crecimiento
de los ovocitos a partir del mes de agosto y hasta abril alcanzÆndose en este mes los mayores diÆmetros anuales La tercerparte es una combinación entreovocitos con diÆmetros grandes como pequeæoscomenzando desde el mes de abril hastajunio Esta combinación detamaæos sugiere que la ovulación
se lleva acabo durante estos meses
Fecundidad
El nœmero de embriones In utero por hembra varió de 4 a 16 siendo mas frecuentes entre 5 y
9 embriones Fig 9 El menor nœmero embrionario fue de 73 cmAD 4 embriones mientras que el
mayor se encontró en una hembras de 115 cmAD 16 embriones La comparación entre la longitudmaterna yel nœmero de embriones mostró una baja relación r2 047 sin embargo en los datos se
observa una ligera tendencia al aumento del nœmero de embriones con el awnento del ancho de disco
materno
e

Crecimiento embrionario
El tamaæo promedio de los embriones a travØs de los mesesen todos los aæos de muestreo Fig10 mostró los mayores valores durante marw 184 cmAD abril 20 62 cmAD y junio 25 55 cm de
AD Posterionnente el promedio disminuyó drÆsticamente durante julio y agosto 7 8 y 7 6 cmAD
encontrÆndose los tamaæos embrionarios mas pequeæos 5 Y 5 1 cm AD en dichos meses En noviembre
se observó un aumento en el tan1aæo de los embriones 1631 cmAD
C DIMORFISMO SEXUAL
AnÆlisis morfomØtricoEl anÆlisis estadístico entre machos y hembras con una prueba ANDE VA mostró que tansolo 5
de las 16 medidas tomadas presentaron diferencias significativas P 0 05 Estas medidas fueron distan
cia entre las primerasaberturas branquiales DBr1 distancia entre las quintas aærturasbranquiales DBr5
longitud interocular 10 longitud preocular PO y longitud preoral LPreor El resto de las medidas no
mostrarondiferencias estadísticas significativas sin embargo la variación en las hembras fue mayor que la
encontrada para los machos Tabla 4 y Fig 1 la by c
`ngulos de apertura del disco
El Ængulo de apertura del disco en los machos presentó valores desde los 1080a los 1220 con una
media de 115 530 en las hembras fue de 1260 a 1290 con una media de 127 20 de apertura La pruebaANDEVA expresó diferencias significativas P 0 05 entre machos y hembras Tabla 4
Pliegues caudales
La observación y anÆlisis de los pliegues caudales en 6 hembras 16 machos y 52 embriones 29
machos y 22 hembras embriones mostró que 70 organismos presentaron dos pliegues caudales en la
posición dorsal y ventral de la cola Solo 4 organismos no presentaron pliegues en ninguna de las posiciones cola desnuda 3 machos 46 a 50 cmAD y 1 embrión hembra de 6 7 cm de AD La pruebaANDEVA P 0 05 entre el PCS y el PCI no mostró diferencias significativas entre los orgæøsmosTabla 4 y Fig lIc
ColoraciÓn
En cuanto a la coloración de los org3Jøsmos esta nopresentó diferencias por sexo talla o Øpocadel aæo De esta forma los patronesde color embrionario s fueron sirrølares a los dejuveniles y adultos
Seobservaron tres patrones de coloración bien deftnidos en la parte dorsal del cuerpo 1 dorsQ
generalmente negro ode verde a cafØ obscuro con puntos de color claro desde puntos blancos hasta
verde claro o amarillo tenue 2 dorso cafØ claro o verde de claro a fuerte con pWltos de color obscuro

generalmente negro 3 dorso sin la presencia de puntos omanchas con varias tonalidades de cafØ y verde
desde claros aobscuros grisÆceo s e inclusive negro Los puntos tanto claros como obscuros no pre
sentaron unpatrón defuødo esto es pudieron verse en cualquier parte del dorso La cola tambiØn mostró
puntos de color claro plUltos de color obscuro y la ausencia de puntos independientemente de los patro
nes que presentaba el dorso El color de la cola fuØ el mismo que el dorsal El color ventral fue generalmente de color blanco a crema con coloraciones rojizas o parduscas en los bordes tanto de las aletas
pectorales como de las aletas pØlvicas y gonopterigios Fig 12

DISCUSIÓN
A ABUNDANCIA TEMPORAL
Las especies de la familia Gymnuridae no tienen una importancia económica relevanteyaque su
pesquería es incidental Bigelow y Schroeder 1953 con un uso marginal para consumo humano De la
Cruz Agüero el al 1997 No obstante esto en Balœa Almejas es capturada durante todo el aæo y
comercializada principalmente en forma seca salada junto con otras especies de rayas como Narcine
entemedor Rhinobatos productus Zapterix exasperata Dasyatis longus D brevis Myliobatis
caliJórnica M longirostris Rhinoptera steindaclmeri y tiburones como Carcharinus obscurus A
partir de estas capturas se pudo observar la variación mensual de su abundancia
En general la abundancia de la raya mariposa G marmorata tabla 2 mostró una variación anual
cíclica con la mayor abundancia de mayo aagosto primavera verano con una disminución de septiembre a febrero otoæo e invierno Con base en su experiencia los pescadoresde Puerto Viejo mencionan
que este fenómeno se repite cada aæo y que probablemente este ligado asu ciclo reproductivo A este
respecto Villavicencio Garayzar 1993 Y 1995b menciona que a partir de abril hasta agosto se dan las
mayoresabundancias de G marmorata enBalœa Abnejas reportando un porcentaje de captura comer
cialde 12 9 en segundo lugar despuØs de R productus 55 1 y antes de D brevis 11 3
Conbase en estos antecedentes se puede suponerque existe una inmigración de organismos en la
Balœa igual que el reportado por Mariano 1997 y Villavicencio Garayzar 2000 para D brevis y N
entemedor respectivamente Ellos mencionan una inmigración y emigración de los organismos reguladapor la temperatura del agua encontrando una entrada despuØs de marzo abril cuando las temperaturasson I1lÚÚmas hasta alcanzar los valores mÆximos en julio agosto cuando se presentan las temperaturas
mÆximas Posteriormente los organismos salen al descender la temperatura y regresan hasta el aæo si
guiente PrÆcticamente G marmorata se comporta de manerasimilar con una entrada de marzoa agosto
y una salida afmales de agosto y septiembre enel resto del aæo su abundancia es casi nula Este fenómeno
tambiØn es reportado por Villavicencio Garayzar J 993 que menciona que las hembras de G marmorata
entranaBahía Almejas a fmales de abril y principios de mayo cargadas y que disminuyen su abundancia
haciajunio permaneciendo solo algunas hembras grandes hasta agosto Por su parte BeltrÆn FØlix et al
1986 reporta que las mayoresabundancias de G marmorala para el estero de Punta Banda Ensenada
B C se presentan durante el mes de mayo y agrega que esta especie es considerada dentro delas cuatro
mas importantes tanto en abundancia como en biornasa AdemÆs tanto BeltrÆn FØlix elal 1986 como
Vtllavicencio Garayzar 2000 mencionan que la entrada y salida de estasespecies de elasmobranquios se

encuentran relacionadacon sus ciclos reproductivos sobre todo porque las hembrapresentaron estadíos
avanzados de embarazo y los machos fueron sexualmente maduros
Proporción de sexos
En la proporción de sexos el dominio de las hembras tambiØn es reportada por Villavicencio
Garayzar 1993 Y 1995b mencionando una proporción de 3 67 1 H M para esta zona sobre todo en los
meses de junio y julio agosto con untotal de 77 hembras por 21 machos Los cambios en las proporciones sexuales son explicadas por Wounns 1977 Pratt 1979 Y Smith y Merriner 1986 arglUI1entando
que las poblaciones de elasmobranquios presentan una segregación tanto por tallas como por sexos sobre
todo despuØs de alcanzar la madurez sexual
Los resultados ademÆs muestran una proporción sexual con embriones in u era no significativamentediferente P 0 05 de 1 15 machos porcada hembra en un total de 21 hembras analizadas Villavicencio
Garayzar 1993 tambiØn encontró unamayor proporción de machos 1 9 1 M H en 4 hembras grandes
Composición de tallas
La composición de tallas enel total de organismos capturados machos y hembras mostró un
mínimo de 21 cmy unmÆximo de 122 cm de AD Al respecto Breder 1928 reporta tallas de 24 3 cmy
39cm de LT mientras que Beebe y Tee Van 1941 menciona tallas mÆximas de 121 y 151 cmde AD al
igual que Castro Aguirre 1965 con 150 cm de AD Por otro lado BeltrÆn FØlix elal 986 menciona
que los organismos de 26 cma I 16 cm deAD fueron comunes en el estero de Punta Banda y Villavicencio
Garayzar 1993 Y 1995b entre los 40 5 cmy 109 cm AD para Bahía Almejas
El rango de tallas encontrado para los machos de 21 a91 5 cmAD fuØ mayor que el reportado
por ViIlavicencio Garayzar 1993 Y l995b de 40 5 a 60 cm de AD yel mostrado por McEachran y
Notarbartolo 1995 de 31 cm de LT y 39 cmde AD como mÆximo para machos maduros
Para el caso de las hembras el rango de tallasencontrado 25 5 y 122 cm AD tambiØn fue mayor
que el seæalado por ViIlavicencio Garayzar 1993 y 1995b de 42 a 109 cm AD Este autor tambiØn
reporta la captura de los ejemplares mayores entre marzo yagosto mientras que los resultados muestran
las mayores tallas en abril y julio McEachran y Notarbartolo 995 menciona que las tallas mÆximas son
de 100 cm de LT y ISO cm de AD para las hembras sin mencionar las tallas mínimas

B BIOLOG˝A REPRODUCTIVA
Tallas de primera madurez
Losresultados muestran una ta1Ia de primera madurez de 41 cmAD Villavicencio Garayzar 1993
Y 1995b reportauna talla de primera madurez de 45 cmADcon características similares para un total de
21 machos El reportar machos inmaduros entre los 40 5 a44 cmAD gonopterigios no calcificados sin
semen y maduros despuØs de los 45 cm AD Porotro lado McEachran y Notarbartolo 1995 reportan
machos sexualmente maduros a los 39cm de AD sin embargo no mencionan ninguna de las característi
cas de determinación Bigelow y Schroeder 1953 seæalan que los machos de G micrura maduran
despuØs de los 42 cm AV siendo esta muy similar a la propuesta en los resultados y la calculada por
Villavicencio Garayzar 1993
Encuanto al aparato reproductor de las hembras se observó que solo el ovario izquierdo y los dos
oviductos son funcionales condición general en la fumilia Gymnuridae Bigelow y Schroeder 1953 Daiber
y Booth 1960 Villavicencio Garayzar 1993 Conbase en la relación del diÆmetro del oviducto diÆmetro
de los ovocitos con el ancho de disco y la presencia de embriones o trofonemata en œteros la talla de
primera madurez se estimó en 68 cmde AD Al respecto VilIavicencio Garayzar 1993 menciona una
talla de primera madurez aproximada alos 74 cm AD con untotal de 77 hembras analizadas Ladiferen
cia en tallade este œltimo con respecto a los resultados parece estar bazado œnicamente en tamaæo de
muestra Por otro lado BeltrÆn FØlix el al 1986 reporta tallas grandes y maduras entre los 86 y 116 cm
AD para el estero de Punta Banda al observar la expulsión de embriones en el momento de la captura
Ciclo reproductivoComo se mencionó anteriormente los machos maduros ingresan a la bahía durante el mes de
mayo permaneciendo hasta el mesde julio agosto mientras que las hembras inician su llegada durante los
meses de mayo julÚo para salir en agosto Este comportamiento tambiØn fue observado tanto por
VilIavicencio Garayzar 1993 en Bahía Almejas como por BeltrÆn FØlix el al 1986 en el estero de
Punta Banda quienesjunto con los pescadores mencionan que la entrada de los organismos puede sercon
fmes reproductivos Se observó ademÆs deque las hembras adultas que entraban a la bahía presentabanun estado de embarazo y desarrollo de ovocitos coincidente con lo reportado para G marmorala en el
estero de Punta Banda BeltrÆn FØlix el al 1986
Los valores mÆximos en el diÆmetro de los ovocitos se alcanzaron desde el mes de abril hastael
mes de julio indicativo de que la ovulación ocurre enestosmeses DespuØs de este tiempo solo se obser
varon ovocitos pequeæos en los ovarios rangos de2 a7 mm de diÆmetro que posiblemente sean fecun

dados al aæo siguiente Bigelow y Schroeder 1953 hacen mención de la presencia de ovocitos pequeæos
rango de 5 a 10mm que inician su crecimiento antes del nacimiento de los embriones en desarrollo en
varias especies de raya mariposa
TambiØn durante los meses de abril ajulio se registraron los tamaæos mÆximos enembriones Fig10 infiriØndose que la expulsión de esta generación se lleva acabo en este tiempo la cual se observó por
la presencia de neonatosen las capturas Fig 3 y 4 ab Al mismo tiempo que las hembras eran portadoras de tallas embrionarias grandes otras presentaban seæales de cópula reciente como la presencia de
semen o embriones poco desarrolJados en los oviductos Este hecho fue evidente a finales de julio y
principios de agosto antes que las hembras abandonaran el lugar Posterior al mes de julio se observó un
crecimiento continuo de los embriones
Desde la entrada de los machos enmayo hasta su salida en agosto se observó un mayor nœmero
de machos maduros y con semen ademÆs de que el ancho dedisco encontrado permaneció por arriba de
la talla de primera madurez estimada Fig 5 y 3a esto es machos completamente maduros
Por su parte Villavicencio Garayzar 1993 reporta que la expulsión deembriones se realizaenel
mes de mayo y observa hembras con huevos fecundados enjunio y tallas embrionarias pequeæas durante
julio agosto con machos maduros presentes durante estosmeses BeltrÆn FØlix elal 1986 describe la
presencia de hembras de G marmorata con embarazo avanzado durante mayo
Bigelow y Schroeder 1953 mencionan que la duración delperiodo de gestación no es conocido
en la mayoría de las especies del gØnero Gymnura Al respecto Daiber y Booth 1960 proponen un
periodo de gestación de 5 a 6 meses para G micrura con base en la expulsión de los embriones en
verano y dos periodos para G altavela uno al fmalizar el invierno y otro despuØs del verano observan
do huevos fecundados y embriones poco desarrollados en los œteros durante los meses de febrero y julioagosto Villavicencio Garayzar 1993 menciona que solo serÆ posible determinar el o los periodos
reproductivos de G marmorata cuando se estudien otras Æreas sin embargo todo indica una posibleanualidad en el ciclo reproductivo con un periodo de gestación de julio amarzo de 9 a 12 meses Un
periodo de gestación anual ha sido propuesto por Joseph y Yerger In Daiber y Booth 1960 para G
micrura mientras que Wallace 1967 lo sugiere para G natalensis
Al respecto Martin y Cailliet 1988 proponen para Myliobatis californica unciclo reproductivoanual con base en tres evidencias que tambiØn se manifiestanen G marmora a 1 el œnico conocimiento
del periodo de ovulación se da en la mitad del verano 2 la finalización del crecimiento embrionario y l
captura de los neonatos se da afinales del verano 3 la confluencia de machos y hembras maduros en la
nœsma zona estuarina tambiØn se da durante los meses de verano
e

Villavicencio Garayzar 2000 mencionaparaN entemedor que la anualidaddel ciclo reproductivopuede ser probada por el desarrollo simultÆneo tanto de los ovocitos como de los embriones poniendo de
manifiesto que las hembras se reproducen todos los aæos durante su vida reproductiva
Cabe destacar que uno de los principales problemas en el estudio de los elasmobranquios es el
seguimiento de sus ciclos reproductivos ya que en ciertos estadíos de gravidez los organismos son inac
cesibles como resultado de su segregación y osus hÆbitos migrntorios Smith y Merriner 1986 Villavicencio
Garayzar 1993 siendo factible su estudio solo con el anÆlisis de organismos preservados en colecciones
ó por medio de las capturas de fines comerciales Luer y Gilbert 1991
Fecundidad
La fecundidad fue de 4 embriones y de 16 como mÆximo Al respecto Daiber y Booth 1960
sólo encontraron una fecundidad de 5 embriones por hembra en una muestra de cuatro hembras de G
micrura mientras que Wallace 1967 reporta de 5 a 9 embriones por hembra en G nata ensis TambiØn
se observó una ligera tendencia a incrementarse el nœmero de embriones con el ancho de disco materno
hecho que fue descrito tambiØn por Villavicencio Garayzar 1993 con unanÆlisis de solo cuatro hembras
con 82 cmAD a106 cmAD mientras que G allavela tambiØn muestra esta tendencia Un problema en
el estudio de la fecundidad lo manifiesta BeltrÆn FØlix elal 1986 al mencionar que las hembras expulsaban a los embriones cuando estas eran capturadas e izadas a bordo sin embargo se puede obtener una
aproximación del nœmero de embriones por hembra con un incierto margen de error
Talla de nacimiento
En organismos de vida libre el ancho de disco mas pequeæo perteneció aun macho y una hembra
de 21 y 25 5 cm AD respectivamente En embriones la talla mas grande In u era fue de 26 5 cmAD en
dos hembras de 101 cmy 122 cmAD Una talla similar reporta BellrÆn FØlix et al 1986 para el estero
de Punta Banda con organismos de 26 cm AD mientras que Castro Aguirre et al 1970 menciona una
talla de vida libre de 14 3 cmAD en el Golfo de California Bigelow y Schroeder 1953 reporta para G
m crura tallas entre los 15 a 23 cm AD mientras que Daiber y Boolh 1960 obtuvieron tallas de 24 2 cm
AD para la misma especie

C DIRMOFISMO SEXUAL
La presencia de dos especies en la zona ha sido mencionada por tres autores McEachran y
Notarbartolo 1995 Castro Aguirre y Espinosa PØrez 1996 y De la Cruz Agüero et al 1997 Ellos
utilizan la forma del cuerpo en espedallas longitudes pre e interoculares y la presencia y posición de los
plieguescaudales para discriminar entre ambas
En cuanto alos plieguescaudales McEachrany NotarOOrtolo 1995 mencionan que G marmorata
presenta dos pliegues bien desarrollados en la parte dorsal y ventral de la cola mientras que G
crebripunctata puede ono presentar un solo pliegue dØrmico enposición ventral Castro Aguirre y Espinosa PØrez 1996 mencionan que G crebripunctata solo presentaun pliegue dØrmico en el borde dorsal
de la cola mientras que G marmorata presenta ambos pliegues bien desarrollados en forma de una
pequeæa aleta caudal Apesar deque ambos autores contradicen susclaves coinciden con doscosas que
G crebripunctata presentaun solo pliegue caudal en unasola posición de la cola nunca en ambas y que
G marmorata cuenta con ambos pliegues caudales
McEachran y Notarbartolo 1995 utilizaron la forma del disco como característica discriminatoria
mencionando que G marmorata presentauna longitud preorbital menor que la distancia interorbital con
una forma del cuerpo mas redonda que G crehripunctata en la cual la longitud preorbital es mayor que
la distancia interorbital y por tanto su forma corporal es mas triangular Cabe seæalar que estos autores
tambiØn relacionan la formadel cuerpo con la presencia y posición de los pliegues caudales
El anÆlisis corporal de la raya mariposa del Pacífico ha sido realizado por autores corno Breder
1928 El menciona que los machos presentan ambos pliegues tanto en posición dorsal como ventral
menos desarrollados que en hembras y embriones Este autor tambiØn analiza otras características como
son los patrones de coloración y la forma general del cuerpo proponiendo ademÆs una sinonímia entre
ambas Con base en esta idea Beehe y Tee Van 1941 Bigelow y Schroeder 1953 Castro Aguirre y
colaboradores 1965 y 1970 BeltrÆn FØlix et al 1986 Villavicencio Garayzar 1993 y 1995b Y
Villavicencio Garayzar y Abitia CÆrdenas 1994 consideraron a G marmorata corno la œnica especieenel Pacífico Mexicano
Bigelow y Schroeder 1953 realizaron unestudio morfomØtrico muy similar para la especie G
micruraen el AtlÆntico Americano encontrando que la parte preocular prebucal y precloacal ademÆs del
ancho y forma del disco presentaron diferencias marcadas para los machos y las hembras denotando P9r
lo tanto un dimorfismo sexual Ellos tambiØn mencionan que la especie mÆs parecida tanto biológica como
morfológicamente aG micrura es G marmorata del Pacífico

Tal es así que si se analizan las ilustraciones presentadas porBigelow y Schroeder 1953 para G micrura
de Breder 1928 para G marmvrata y McEachran y Notarbartolo 1995 de G marmora a y G
crebripunctata estas no difieren en forma y sexo
La variación en la forma del disco ha sido reportada por otros autores para otras especies de
Gymnura como es el caso de Wallace 1967 para G natalensis El reporta que los juveniles no
reproductivos presentan una fonna deldisco mas triangular que la encontrada en los adultosreproductivos
y que este cambio estÆ relacionado con la madurez sexual El hecho de tener diferentes morfotipos antes y
despuØs de la madurez sexual tampoco es nuevo dentro delgrupo de los elasmobranquios ya que espe
cies como Gymnura micrura llalaelurus bivius y Myliobatus aquila lo presentan De aquí que
Gosztonyi 1973 Y WaUace 1967 se refieran aeste cambio como dimorfismo sexual secundario
Losresultados obtenidos del anÆlisis morfomØtrico muestran por un lado que la presencia y posición de los plieguescaudales no guarda ninguna relación con otras partes del cuerpo como son las longitudes preorbitales indicadas por McEachran y Notarbartolo 995 o con el sexo ancho de disco colora
ción formadel disco talla de nacimiento y madurez sexual mientras que las partes corporales que mos
traron diferencias significativas 5 de las 16 medidas ANDEVA P 0 05 porción preocular interocular
prebucal y la distancia nœnima entre la primera y quintaaberturas branquiales entre las dos especies no
difieren en mucho alas reportadas por Bigelow y Schroeder 1953 para la especie del AtlÆntico Tanto
machos como hembras en fase embrionaria presentaron la misma forma redondeada de la madre hecho
similar a las ilustraciones de Breder 1928 DespuØs delnacimiento y antes de la talla de primera madurez
la fOnlla de los machos varió en poco al de las hembras sin embargo cuando los machos alcanzaron la talla
de primera madurez se presentó un crecimiento mÆs acelerado de la parte preocular del cuerpo conclu
yendo con la forma triangularde los adultos observÆndose un dimorfismo sexual secundario como el
reportado en G micrura y G natalensis En las hembras el crecimiento preocular permaneció constan
te provocando que su forma redondeada no varíe desde la fase embrionaria hasta la adulta Estas diferen
cias en la forma del cuerpo se relacionan ademÆs con los Ængulos de apertura del disco medidas utilizadas
por Bigelow y Schroeder 1953 para G micrura Si comparamos los Ængulos obtenidosporestos auto
res de 1200 a 1300 para los machos y de 1300 a 1400 en las hembras de G micrura con los resultados
1080 a 1220 para machos y 1260 a1290parahembras de G marmura a se observa que los valores son
muy semejantes sobre todo porque los machos presentan Ængulos mas cerrados que las hembras en
ambas especies relacionÆndose esto directamente con su forma triangular
El anÆlisis dela coloración muestra patronesque coinciden con los reportes hechos Breder 928
Beebe y Tee Van 1941 Y Castro Aguirre 1965 no existiendo diferencias por sexos opor tallas pu
diØndose observar desde fases embrionarias hasta adultos reproductivos

Sumado aesto la identificación de los organismos obtenidos en Bahía Almejas condujo a que
todas las hembras fueran identificadas como G marmorata mientras que todos los machos fueran G
crebripunctata afirmando la idea de Breder 1928 Y Castro Aguirre 1970 de un dimorfismo sexual
Estas diferencias en la formadel cuerpo y pliegues caudales por lo tanto no presentan una confiabitidad
suficiente para la discriminación entre especies sobre todo antes de la fase reproductiva no obstante si
pueden ser ocupadas para la discriminación entre familias como lo presentan Bigelow y Schroeder 1953
Castro Aguirre 1965 Nelson 1994 Y McEachran y Notarbartolo 1995
051725

CONCLUSIONES
A Abundancia temporalLa presencia de machos y hembras de G marmorata enBahía Ahnejas es aparentemente regu
lada por la temperatura del agua entrando despuØs de marw abril cuando las temperaturas son mínimas
y alcanzando sus valores mÆximos en julio agosto cuando tambiØn se alcanzan las temperaturas mÆximas
Posterioffirente los organismos salen del lugar cuando la temperatura comienza de nuevacuenta su ascen
so y regresan hasta el próximo aæo Las hembras fueron mas ablUldantes que los machos casi durante
todo el aæo no así en fases embrionarias donde existió una mayor proporción de machos con respecto a
las hembras Los machos fueron mas pequeæos en talla que las hembras
B Biología reproductivaLa talla de primera madurez de los machos se estimó en 41 cmAD mientras que en las hembras
en 68 cm fue de AD con el ovario izquierdo y los dos oviductos funcionales Esta talla de madurez indica
que la mayorja de los machos y hembras encontrados en Bahía Almejas eran maduros sexualrnente Las
hembras presentaron un estado de embarazo y desarrollo de ovocitos avanzado y los machos presenta
ron abundante semen concluyendose que la presencia de esta especie en Bahía Almejas es con fmes
reproductivos Su periodo de gestación es anual con una duración de 9 a 12 meses y una expulsiónembrionaria durante los meses primavera verano Su fecundidad es de 2a9 embriones porhembra y la
talla de nacimiento se calcula entre los 2 I y 26 5 cm AD considerando los organismos de vida libre y la
tallaembrionaria mÆxima
C Dimorfismo sexual
Solo la porción preocular interocular prebucal la IllÚI˛ma distancia entre la primera y quinta aber
turas branquiales el ancho del disco y la forma de este difirió entre sexos despuØs de la talla de primeramadurez Sin embargo este dimorfismo sexual secundario es comœn en otrasespecies del gØnero por lo
que debeconsiderarse aG creblipunctata corno taxón invÆlido ademÆs de que las diferencias morfológicasencontradas no tienenuna confiabilidad suficiente en la discriminación entre especies sobre todo antes de
la fase reproductiva aunque si pueden serocupadas para la discriminación entre familias

LITERATURA cnADA
`LVAREZ B A GALINDO y A B CHEE 1975 Características hidroquímicas de Balœa MagdalenaB C S Ciencias Marinas 2 2 94 110
BAKUN A 1973 Costal upwelling indices west coast ofNorth America 1946 71 NOAA Tech
Rep NMFS SSRF 671 103p
BEEBE W y 1 TEE VAN 1941 Eastem Pacific expeditions ofthe New York loological Society XXVIll
Fishes from the tropical Eastem Pacific From Cedros Island lower California South to the
Galapagos island and Northern Peru Part 3 Rays mantas and chimeras Zoological 26 245
280p
BELTRAN FELIX J L M GREGORY A CHAGOYA y S `LVAREZ 1986 Ictiofauna del estero
de Punta Banda Ensenada BCÛa California MØxico antes de una operación de dragado Ciencias
Marinas 121 79 92
BIGELOW H B Y W C SCHROEDER 1953 Fishes ollhe weslern Norlh Allanlic Part 2 Saw
flShes guitarflShes skates and rays Sears Foundation for Marine Research New Haven 588 p
BREDER C M 1928 Elasmobranchii from Panama to lower California 171e Bingham OceanographicCollection 21 I 13 p
BRIGGS C 1 1974 Marine zoogeography McGraw Hill U S A 475 p
CASTRO AGUIRRE 1 L 1965 Peces Sierra Rayas Mantas y especies afmes de MØxico Anales delInstituto Nacional de Investigación Biológico Pesqueras MØxico Vol 1 259 p
CASTRO AGUIRRE 1 L ARVIZU M 1 y 1 P`EZ 1970 Contribución al conocimiento de los
peces del Golfo de California Revista de la sociedad Mexicana de Historia Natural 31 1 07
181p
CASTRO AGUIRRE 1 L y H ESPINOSA PÉREZ 1996 Listado Faunístico deMØxico VllCatÆ
logoSistemÆtico de las rayasy especies afines de MØxico Chondrichlhyes Elasmobranchii
RaRfòrmes Batoideim017Jha Instituto de Biología UN AM MØxico 75 p
COMPAGNO L 1 1990 Alternative life lœstory styles ofcartilaginous fishes in time and space Env
BioFish 28 33 75
COADS 1994 Data on CD ROOM ofthe Eastem Pacific ocean NOAAINMFS and ORSTOM
Monterey California Vol 5

DAIBER F c y BOOTH R A 1960 Notes on the biology ofthe butterfly rays Gymnura altavela andG micrura Copeia 2 137 139 p
DE LACRUZ A 1 1997 CatÆlogo de los peces marinos de Baja California Sur C ICIMAR I P
N MØxico 346 p
DOWNTON H C 1996 Estrategia reproductiva de la guitarra Rhinobatos productus Ayres 1856 en
la costa occidental de Baja California Sur MØxico Tesis de licenciatura U ABC S MØxico
32p
GOSlTONYI A E 1973 Sobre el dimorfismo sexual secundario en llalaelurus bivius Müller y Henle
1841 Garman 1913 Elasmobranchii Scyliorhinidae en aguas patagónico fueguinas Physis Secc
A Buenos Arres 32 85 317 323
HAMLETT W C 1999 Sharks Skates and Rays The biology of elasmobranch fishes The John
HopkinsUniversity press London 515 p
HUBBS Cy R ISHIYAMA 1968 Methods for the studyand description ofskates Rajidae CopeiaJ483 491
LUER C A y P W GILBERT 1991 Elasmobranch fish Oviparous Viviparous and OvoviviparousOceanus magazine 34 3 47 52
LYNN R1 Y 1 1 SIMPSON 1987 The California current system the seasonal variability ofits physicalcharacteristics Geophysic Research 9212 12947 12966
MARIANO M E 1997 Biología reproductiva de la raya lodera Dasyatis brevis Garman 1880 en
Babia Almejas B C S MØxico Tesis de licenciatura U ABC S MØxico 46 p
MARTIN L K Y G M CAILLlET 1988 Aspects ofthe Reproduction ofthe Bat Ray Myliobatiscalijòrnica in Central California Copeia 3 754 762
McEACHRAN 1 D Y G NOTARBARTOLO DI SCIARA 1995 Peces Batoideos Pertenece a
Guía FAO para la identificación de especies para los fines de la pesca Pacífico Centro
Oriental Volumen 11 Vertebrados parte 1 Organización de las Naciones Unidaspara la agricultura y la alimentación Roma 1200 p
NELSON 1 S 1994 Fishes orthe World John Wiley and Sons lnc U S A 600 p

PRAIT H L JR Y T K OTAKE 1990 Recomrnendations forwork needed to increase out knowledgeofreproductioll relative to fishery management p 509 510 In Pratt H L S H Gruber y T
Taniuchi eds Elasmobranchs as living resources Advances in the biology ecology systematicsand the status ofthe fIsheries Natiollal Oceanic and Atmospherics Administration National Ma
r˛ne Fisheries Service Tech Rep 90
PRAIT JR HL 1979 Reproduction in the blue shark Prionace glauca Fish Bull U S 77 2 445
470
SALINAS Z C A LEYVA CD LLUCH B y DIAZ RE 1990 Distribución geogrÆfica y variabilidadclimÆtica de los regímenes pluviomØtricos en Baja California Sur MØxico Atmó rera 3 217 237
SIMPFENDORFER C A 1992 Reprodm tive strategy ofthe Australian sharpnose shark Rhizoprionodontaylori Elasmobranchii Carcharhinidae from Cleveland Bay Northern Queensland A ust J
Mar Freshwater Res 43 67 75
SMITH 1 W y MERRlNER 1 V 1986 Observations on the reproductive biologyofthe Cownose ray
Rhinoptera bonasus in Chesapeake bayo Fishery bulletin 84 4 871 877 p
VILLAVICENCIO G C 1993 Notas sobre Gymnura marmorata Cooper Pises Dasyatidae en
BahiaAImejas B C S MØxico Rev Inv Cient U AB C S 41 91 94
VILLAVICENCIO G C y ABITIA C 1994 Elasmobranquios de Bahía Magdalena y Laguna San
Ignacio Baja California Sur MØxico Rev Inv Ciento U AB C S 5 2 63 67
VILLAVICENCIO G C 1995a Biología reproductiva de la guitarra pinta Zapterix exasperataPisces Rhinobatidae en Bahía Almejas Baja California Sur MØxico Ciencias Marinas U A
B C 21 2 141 153
VILLAVICENCIO G C 1995b Distribución temporal y condición reproductiva de las rayas PiscesBatoidei capturadas comercialmente en Bahía Almejas B C S MØxico Rev Inv Ciento
U ABC S 61 2 1 12
VILLAVICENCIO G C 2000 Taxonomía abundancia estacional edad y crecimiento y biologíareproductiva de Narcine entemedor Jordany Starks Chondrichtyes Narcinidae en Bahía Al
mejas B C S MØxico Tesis de Doctorado U AN L MØxico 138 p
WALLACE J H 1967 The Batoid fishes ofthe East Coast ofSouthern Africa Part 11 Manta EagleDuckbill Cownose Butterfly and Sting Rays Investigational report No 16 SouthAfrican As
sociatioll for marine biological research SudÆfrica 356 p
WüURMS 1 P 977 Reproduction and Development in Chondrichtyan fishes Amer Zool 17 379
410
24040
BahIa MaPJena
j
OcØano Pacífico
240 19
N
i
liZO 00
Baja California Sur
1Il0 40
J5 1130 1110 109
OcØano Pacifico
N
i
1Hf 80
Fig 1 Localización del Ærea de estudio Puerto pesquero Puerto Viejo enel complejolagunar Bahía Magdalena Almejas
2SO
UJO
240
220
nE I11 iJÞ i tk
f l k if
4
a
t1ìt
PCI
dJ
1tt
LlCIQJIt
y
o
d
f
1 e
jjflJ
Fig 2 ab e d y e Medidas tomadas para la comparación morfomØtrica de Gymnura marmorala
o
100n 438 machos
f80Prom
T Mini 60o
g Jd
40 Talla de primera madurez
120rr
O
Ene Feb Mar Abr May Jun JuI Ago p Ocl Nov
Meses
Fig 3a Tallas promedio mÆximas y mínimas pormes con referencia ala talla de primera madurez para
los machos
1401 n 610 hembras
120
Π100
tlI 80o
altl
60
gal
40 J8err
fMaxProm
Min
Talla de primera madurez
J
20
o jn
Ene Feb Mar Abr May lun luI Ago Sep Ocl Nov Die
Meses
Fig3b Tallas promedio mÆximas y mœømas por mes con referencia a la talla de primeramadurez para
las hembras

Hembras
alM zo
a70n21
eoM 9
50
40
30
20
10
o
2S 35 45 55 65 75 85 9S 105 lIS 125
10 A rd
70 11 24
60 M 95
50
40
30
20
10
O
25 35 45 55
1068
70MiO
60
50
40
30
20
10
O
25 35 85 95 105 115 125
1011 209
70M 60
60
50
t 40
30
1 2010
i O
25 35 45 55 65 75 85 95 lOS liS 125
Sl10
70 1F122
60M 58
50
40
30
20
10 Jo
25 35 45 55 65 75 85 95 105 115 125
10
70Il128
60M 54
50
40
30
20lI25 35 45
10Il18
70M 50
60
50
40
30
20
10
O
10
70
60
50
40
30
20
10
o
Ag SIO
75 85 95 105 115 125
re
Machos
lO Marzo b701120
60 M 50
50
40
30
20
10
O
25 35 45 55 65 75 85 95 lOS 115 125
lO Abra
70 07
60 M 51
SO
40
30
20
10
o
25 35 45 SS 65 75 85 95 lOS 115 125
lOMayo
70 068
60 M 60
SO
40
30
20
10
O
25 35 45 55 65 75 85 95 105 115 125
JwiolO
70 0106
60M 52
SO
40
30
20
J
O
25 35 45 55 65 75 85 95 lOS liS 125
lOJulio
7011 128
60M 48
SO
40
30
20
10
O85 95 lOS 115 125
lO
70
60
50
40
30
20
10
O
8S 95 105 115 125
lO
70
60
50
40
30
20
10
O
85 95 105 115 125
10
70
60
50
40
30
20
10
O
2S 35 45 55 65 75 85 95 lOS 115 125
Ì2 4Ancho de disco cm
a by DIstrjbución modal de cada frecuencia detallas para hembras a ymachos b con respecto ala talladeprimera madurez
25 35 95 105 115 125
n14
M 49
25 35 45 55 65 75 85 95 105 115 125
40 1
35
30
25
1 20
158e
10
5
O 1 LL25 30 35 40 45 50 55
M 438 orgH 610 org
l DJJFlü DO
60 65 70 75 80 85 90 95 100 105 110 115 120 125
Intervalo de talla cm
Ch S el I I
Fig 5 Frecuencia relativa por intervalo de talla para machos y hembras
10
90418
i 8v
7o
6
gs 5
48Ó 3
2l
O r 1 r
15 20 25 30 35 40 45 50 55 60 65 70 75 80 85
Ancho re dioco
Fig 6a Longitud de los gonopterigios cm conrespecto al ancho de disco cm
100
90
80
70
60
º 50gcd 40
30
20
10
0418
o r l I T
30 35 45 50 S5 6040
Ancho re ko cm
Fig 6b Criterio de Pratt y Otake 1990 talla de primera madurez para machos
55
j 4
3goc
2
7
n 176
6
1o
40 90 110 12010050 60 70 80
Ancho de Disco cm
24 n
21 0 95
Π18
o15
ßo
o 12ti
9
9tl
o
tlQ
6
Õ3
O TI 1 r r 1 rI
1 r Tr r 11 1 1 íl 1 r r T
40 50 60 70 80 90 100 110 120 130
Ancho de Disco cm
Fig 7 a DiÆmetro de los oviductos cm b DiÆmetro de los ovocitos mm
con respecto al ancho de øisco cm
a
130
b
100
IX
80
70i
60e
º 50
40
30
20
10
O l f 1 1
50 55 80 85 90 95 10060 65 70 75
Ancho ædiæo cm
Fig 7c Criterio de PraH y Otake 1990 talla de primera madurez para hembras
22 n 18 lJ
E20 nIOIn 7 n lb
18
Ì16n4
25
es 14uü 24 o
o 12n ifII 10Q 23
G8
81l
n 14 n48
e
í í22V 6
4es 212
o n 1 1 1 f r 21
Ene Feb Mlr Alr tvby Jm Jul Aw p Qt1fov Die
tvtæi
Fig 8 Valores promedio mÆximos y ll1Ú1Ì1noS del diÆmetro de los ovocitos comparadoscon la
temperatura a travØs de los meses
18
Fec10 06t19AD16
N20
14 r2047
g PO 05S 1210
l 10gg 8iull 6O
ÒZ 4
2
01 r T r l
68 785 80 8 82 885 91 92 92 93 97 97 97 100 101 102 104 115 122
Ancho de disco materno cm
Fig 9 Nœmero de embriones porhembra
n 19Z7
E27
I N 156 emb
n 33
fn 2924 2fj
EI
212So nà 9
i
rl 18e Uo 24i
o
o 15
1E n 34
o 12 23u n 22en
e9
f 12Ö
o6c
u
213
O I 1 20
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Die
Meses
Fig 10 Ancho promedio mensual delos embriones en comparado con
la temperatura
Machos 111H EMaxr J2 Id
Mín
Long Internasal
Hembras 1 L P
Machos 1IIIaDist Espiracular
Hembras IclJ I
Machos 1 JIIOíst InterOcular
Ilembras 1 L L I
Machos I IJIOíst PreOcular
Ilembras I I J I
Machos 1 clJ 1Long Preoral
Hembras I ct I
Machos 1 el1Long Branquial
Hembras 1 L t 1
Machos
Dist la A Branql U 1
Hembras I r tL
Machos
Dist 5a A BranqIcll I
Hembras l clJ 1
Machos I clJ 1Long Caudal
Hembras I r I T I
5
I
7
I
9
I
JI
I
13
I
15
I
17
I
19
I
21
I
23 25 2
Fig 1 la AnÆlisis morfomØtrico enmachos y hembras Medidas expresadas como un porcentajedel ancho de disco Media 2 desviaciones estÆndar valores mínimos y mÆximos
b1i 1J 1
EMaxr
2 md
Mio
Long Total
Machos
Hembras I c I
Long del Disco
Machos I LIJ I
Ilembras 1 e I J 1
Long Precloacal
Machos LIH
Hembras u c I J I
Long Aleta PØlvica
Machos 1 cIJ 1
Ilembras I L I J u 1
xl40 45 50 55 ro 65 70 75 00 85
Pliegue Caudal Superior eMachos I ctrI
Hembras 1 r I I
Pliegue Caudal Inferior
Machos J cIJ I
Hembras Ic Iu
I
o 0 05 01 015 02 025 03 035 04 045 05
Figura IIb Y e AnÆlisis morfomØtrico enmachos y hembras
At
i ff lir c
1 Úi
l i
t
B
C
Fig 12 Patrones dorsales de coloración encontrada en G marmorata A Manchas negrasB Sin manchas C Manchas blancas
ł
Variable Machos Prom Est Hembras Prom Est P F
LT 7173 7159 0 9496 0 00408
LD 6164 65 59 0 1521 2 216LPC 50 31 5108 0 7665 0 0905
LPelv 59 78 6044 0 8 116 0 0583
LC 20 62 1946 04216 0 673PCS 035 0 33 0 7273 0 125PCI 0 24 0 20 0 2936 1 163LBr 16 60 I 544 0 1546 2 188
DBrI 16 30 18 25 0 0286 5 565
DBr5 10 69 1188 0 0348 5 124
Unas 6 88 743 0 1883 1 855
DEsp 849 940 0 0519 4 26910 8 65 10 19 0 0039 18 009PO 10 51 9 26 0 0447 4 583
LPreor 11 98 10 58 0 0409 4 772
Angulos de apertura 115 53 127 20 0 00001 42 759
Tabla 4 Resultados del ANDEVA P 0 05 de machos y hembras de Gymnllfamarmorata


















