CARACTERIZACIÓN DE LAS PROTEÍNAS DE LA MEMBRANA CELULAR ... · Esta membrana está formada por...
16
CARACTERIZACIÓN DE LAS PROTEÍNAS DE LA MEMBRANA CELULAR Resumen La membrana celular o membrana citoplasmática está formada por una bicapa lipídica que delimita a la célula. Esta membrana está formada por fosfolípidos, glucolípidos y proteínas. La función de la membrana citoplasmática es regular el intercambio de moléculas entre el interior y el exterior celular, servir de barrera, contener el citosol y los distintos organelos celulares. Las proteínas que integran la membrana plasmática de las células son diversas y se agrupan en tres clases: integrales, periféricas y las proteínas ancladas a los lípidos. Estas proteínas actúan como conductos o como anclas en la superficie extracelular de la membrana. En este proyecto se analizó la composición de las proteínas de membrana de eritrocitos de carnero por corrimiento electroforético. El análisis de las proteínas de membrana se realizó en células rojas sanguíneas de carnero, que obtuvieron por punción de la yugular. La sangre se recibió en solución de Alsevers que proporcionó inhibición de la coagulación sanguínea, amortiguación del pH y tonicidad fisiológica. Para obtener las membranas celulares o fantasmas de eritrocitos se retiró la solución de Alserves por centrifugación. La parte celular de la sangre se lavó por centrifugación con una solución isotónica de fosfatos para retirar el plasma sanguíneo (suero y factores de coagulación). Después, el paquete celular se resuspendió en una solución hipotónica de fosfatos de sodio para romper las células y liberar la hemoglobina. Los fantasmas de los eritrocitos se lavaron por tres veces más con la solución isotónica hasta retirar totalmente los restos de hemoglobina. Las membranas celulares obtenidas se analizaron SDS-PAGE. Las proteínas de membrana se desnaturalizaron con calor, bajo condiciones reductoras utilizando el reactivo buffer de Laemmli con β- mercaptoetanol. Debido a la presencia de detergente SDS, que carga a las moléculas negativamente, las proteínas migraron de manera proporcional inversa a su peso molécular, en el campo eléctrico de 100 mV a través del gel de matriz porosa de acrilamida. Para revelar las proteínas, los geles se tiñeron con azul de Coomassie y se destiñeron con una solución de ácido acético-etanol. Los geles al 10 % por su consistencia fueron más manejables, sin embargo, se observó una menor separación entre las proteínas en los geles al 6% por una mejor visualización por la mayor separación de las bandas de las proteínas espectrinas α y β de masas moleculares de 240 y 220 kDa, la anquirina de 116 KDa, banda 3 de 90 a 100 KDa y la actina de 44 KDa. La tinción de azul de Coomassie no revela las glicoforinas A y C que se ven con tinción de plata.
Transcript of CARACTERIZACIÓN DE LAS PROTEÍNAS DE LA MEMBRANA CELULAR ... · Esta membrana está formada por...

CARACTERIZACIÓN DE LAS PROTEÍNAS DE LA MEMBRANA CELULAR Resumen
La membrana celular o membrana citoplasmática está formada por una bicapa lipídica que delimita a
la célula. Esta membrana está formada por fosfolípidos, glucolípidos y proteínas. La función de la
membrana citoplasmática es regular el intercambio de moléculas entre el interior y el exterior celular,
servir de barrera, contener el citosol y los distintos organelos celulares. Las proteínas que integran la
membrana plasmática de las células son diversas y se agrupan en tres clases: integrales, periféricas
y las proteínas ancladas a los lípidos. Estas proteínas actúan como conductos o como anclas en la
superficie extracelular de la membrana. En este proyecto se analizó la composición de las proteínas
de membrana de eritrocitos de carnero por corrimiento electroforético. El análisis de las proteínas de
membrana se realizó en células rojas sanguíneas de carnero, que obtuvieron por punción de la
yugular. La sangre se recibió en solución de Alsevers que proporcionó inhibición de la coagulación
sanguínea, amortiguación del pH y tonicidad fisiológica. Para obtener las membranas celulares o
fantasmas de eritrocitos se retiró la solución de Alserves por centrifugación. La parte celular de la
sangre se lavó por centrifugación con una solución isotónica de fosfatos para retirar el plasma
sanguíneo (suero y factores de coagulación). Después, el paquete celular se resuspendió en una
solución hipotónica de fosfatos de sodio para romper las células y liberar la hemoglobina. Los
fantasmas de los eritrocitos se lavaron por tres veces más con la solución isotónica hasta retirar
totalmente los restos de hemoglobina. Las membranas celulares obtenidas se analizaron SDS-PAGE.
Las proteínas de membrana se desnaturalizaron con calor, bajo condiciones reductoras utilizando el
reactivo buffer de Laemmli con β- mercaptoetanol. Debido a la presencia de detergente SDS, que
carga a las moléculas negativamente, las proteínas migraron de manera proporcional inversa a su
peso molécular, en el campo eléctrico de 100 mV a través del gel de matriz porosa de acrilamida.
Para revelar las proteínas, los geles se tiñeron con azul de Coomassie y se destiñeron con una
solución de ácido acético-etanol. Los geles al 10 % por su consistencia fueron más manejables, sin
embargo, se observó una menor separación entre las proteínas en los geles al 6% por una mejor
visualización por la mayor separación de las bandas de las proteínas espectrinas α y β de masas
moleculares de 240 y 220 kDa, la anquirina de 116 KDa, banda 3 de 90 a 100 KDa y la actina de 44
KDa. La tinción de azul de Coomassie no revela las glicoforinas A y C que se ven con tinción de
plata.

CARACTERIZACIÓN DE LAS PROTEÍNAS DE LA MEMBRANA CELULAR
INTRODUCCIÓN
La estructura y función de las células dependen de las características de sus membranas
que separan el interior de las células del exterior y definen los compartimentos internos de
las células eucariotas, los organelos, el núcleo y el citoplasma. La formación de las
membranas biológicas se basa en las propiedades fisicoquímicas de los lípidos, y todas las
membranas celulares comparten una misma organización estructural: bicapas de fosfolípidos
con proteínas asociadas. Las proteínas de membrana son responsables de muchas
funciones especializadas; como receptores que permiten a la célula responder a señales
externas, el transporte selectivo de moléculas a través de la membrana, y otras participan en
el transporte de electrones y de la fosforilación oxidativa. Además, las proteínas de
membrana controlan las interacciones entre células de organismos pluricelulares. La
organización estructural común de las membranas se relaciona con una gran variedad de
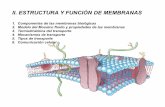
procesos biológicos y funciones especializadas, son superficies delgadas y flexibles. Fig. 1
Todas las membranas poseen una composición común, pero tienen propiedades diferentes.
Las membranas de las células tienen una estructura general común, contiene una bicapa de
fosfolípidos con proteínas distribuidas en la superficie o que la cruzan completamente y son
una estructura fluida (Avers, C., 1994). Fig. 1. Los fosfolípidos que forman a la membrana
tienen la propiedad fisicoquímica de ser ambifílica; una parte que es hidrofóbica (porción de
cola lipídica) y la otra parte es hidrofílica (porción de cabezas de fosfato). Estas moléculas en
solución acuosa concentran sus porciones hidrofóbicas para que estén en contacto estrecho
y puedan esconderse del agua, mientras las partes hidrofílicas están expuestas. Los
fosfolípidos en solución acuosa se agregan para formar una bicapa que forma pequeñas
vesículas cerradas. (Alberts, 2010). Según el tipo celular y el organelo particular de la célula,
una membrana puede contener cientos de proteínas diferentes. Cada proteína de membrana
tiene una orientación definida en relación con el citoplasma, por lo que las propiedades de
una y otra superficie difieren mucho. Las proteínas de membrana pueden agruparse de tres
clases distintas: Las proteínas integrales penetran en la capa lipídica, por lo que tienen
dominios que sobresalen por los lados extracelular y citoplásmico de la membrana. Por sus
dominios hidrófobos transmembrana son difíciles de aislar en una forma soluble. Su
extracción requiere el uso de un detergente como el detergente iónico (cargado) SDS que
desnaturaliza las proteínas o el detergente no iónico (sin carga) Tritón X-100. La mayoría de

proteínas celulares funcionan como receptores que se unen con sustancias específicas en la
superficie de la membrana; como conductos o transportadores participantes en el
desplazamiento de solutos a través de la membrana, o como agentes que transfieren
electrones durante los procesos de fotosíntesis y respiración. (Karp, 2011)
Figura 1. Estructura de la membrana plasmática, modelo del mosaico fluido.
Las proteínas periféricas-extrínsecas se sitúan completas fuera de la capa bicapa lipídica en
el lado citoplasmático o en el extracelular. Fig. 1. Casi siempre se pueden solubilizar con
soluciones salinas muy concentradas que debilitan los enlaces electrostáticos que mantienen
unidas a las proteínas periféricas de la membrana. Estas proteínas brindan soporte
estructural y actúan como anclas para las proteínas integrales de la membrana, funciona
fluidamente. Otras funcionan como enzimas, cubiertas especializadas, o factores que
transmiten señales a través de la membrana. (Karp, 2011)
Las proteínas ancladas a los lípidos se localizan fuera de la bicapa lipídica, en la superficie
extracelular o citoplasmática. Muchas proteínas presentes en la cara externa de la
membrana plasmática están unidas a la membrana mediante un oligosacárido vinculado con
la molécula de fosfatidilinositol, que está sepultada en la hoja externa de la bicapa lipídica.

Las proteínas periféricas de la membrana tienen el enlace glucosilfosfatidilinositol y se
conocen como Proteínas ancladas por GPI. Otro grupo de proteínas presentes en el lado
citoplasmático de la membrana plasmática se fija mediante una o más cadenas largas
hidrocarbonadas incrustadas en la hoja interna de la bicapa lipídica. (Karp,2011).
MARCO TEÓRICO
Los eritrocitos se derivan de las células madre denominadas hemocitoblastos, surgen de
progenitores nucleados o eritroblastos, que se diferencian y expulsan terminalmente su
núcleo para formar el pirenocito. Este último produce el reticulocito. Los reticulocitos
continúan sintetizando proteínas y contienen orgánulos intracelulares, que se eliminan
durante varios días por procesos complejos de remodelación y degradación de la membrana
para producir glóbulos rojos bicóncavos maduros sin organelos intracelulares o citoesqueleto
transcelular (Fig. 2). Los eritrocitos son células altamente especializadas; en humanos y en
carnero son células bicóncavas, conformadas de aproximadamente 8 µm de diámetro y 2 µm
de espesor en el borde, no contienen núcleos u organelos celulares, y están llenos con
aproximadamente 450 mg/ml de hemoglobina en su citoplasma para la entrega de O2 y la
remoción del CO2, son notablemente deformables sin sufrir fragmentación celular,
increíblemente estables, atraviesan repetidamente capilares sanguíneos, incluso más
pequeños que su diámetro, en todos los tejidos periféricos. La vida media es de
aproximadamente 120 días en humanos y 40 días en ratón. Para realizar sus funciones
circulatorias, los eritrocitos contienen en la membrana iones especializados como
transportadores de gases para regular el intercambio O2/CO2, un pH intracelular, y agua en
homeostasis, así como proteínas glicosiladas que forman las bases del sistema de los
grupos sanguíneos antigénicos. Las proteínas de membrana están ancladas a una capa
delgada del citoesqueleto de aproximadamente 100 nm de espesor. El citoesqueleto, que
cubre el lado citoplasmático de la membrana como una red poligonal, es el principal
determinante de la forma de la célula y de estas propiedades mecánicas. (Rodi, 2010).

Figura 2. Maduración del eritroblasto para la producción de eritrocitos.
Su composición lipídica es asimétrica: La hemicapa exterior contiene, moléculas lipídicas que
contienen azúcares, denominadas glicolípidos. Los glicolípidos más complejos, son los
gangliósidos, que contienen oligosacáridos con uno o más residuos de ácido siálico, lo que
les proporciona una carga neta negativa. Exponiendo las células a un medio hipoosmótico
(en el que la concentración de sal sea mucho menor que la del interior celular), resulta fácil
preparar membranas celulares de eritrocito vacías o “fantasmas”. (Rodi, 2010). Cuando la
hemoglobina se extrae de los eritrocitos por lisis osmótica y lavado para obtener fantasmas,
esa membrana plasmática tiene dominios disponibles para análisis bioquímico, biofísico,
estructural y funcional.
Las proteínas en el límite entre el citoesqueleto y la membrana plasmática controlan la forma
de la célula, delimitan los dominios de membrana especializados y estabilizan las uniones a
otras células y al sustrato. Estas proteínas también regulan la locomoción celular y las
respuestas citoplásmicas a factores de crecimiento y otros estímulos externos. Esta
diversidad de funciones celulares se corresponde con la gran cantidad de mecanismos
bioquímicos que median las conexiones entre las proteínas de membrana y el citoesqueleto
subyacente se llama esqueleto de membrana o citoesqueleto membranal.
Análisis bioquímico de las proteínas. Se conocen diferentes técnicas para el análisis
experimental de las proteínas, sin embargo, la más empleada y más eficiente es la
electroforesis, la cual es uno de los métodos bioquímicos para analizar complejos
moleculares como las proteínas, en función del tamaño, carga eléctrica y otras propiedades
físicas, ya que emplea geles de poliacrilamida. La electroforesis en una dimensión es el
método de elección, donde las proteínas se separan de acuerdo a su masa molecular y las

proteínas se solubilizan en duodecil sulfato de sodio (SDS). Es una técnica sencilla,
reproducible y permite la separación de proteínas de 10-300 KDa. Una de las aplicaciones
más comunes de la electroforesis en una dimensión es la caracterización de proteínas
después de realizar algún tipo de purificación previa.
La separación y purificación de proteínas son necesarias para llevar a cabo el estudio y
caracterización del proteoma de los sistemas biológicos. Para ello, éstas deben ser
separadas selectivamente a partir de muestras complejas mediante un procedimiento de
fraccionamiento adecuado. Los métodos de separación se basan en diferentes
características de las proteínas tales como solubilidad, tamaño o carga, entre otras. El
término electroforesis se introdujo por primera vez en 1907 por Michaelis para definir el
fenómeno por el cual una molécula que posee carga neta se desplaza en respuesta a la
aplicación de un campo eléctrico. La velocidad de migración o movilidad a través del campo
eléctrico dependerá de varios factores como son: la intensidad de dicho campo; la carga
neta, tamaño y forma de las moléculas; así como la fuerza iónica, viscosidad y temperatura
del medio en el cual las moléculas se están moviendo. La electroforesis es una herramienta
analítica simple, rápida y muy sensible, lo que la convierte en una técnica de gran utilidad
para la separación y el estudio de moléculas cargadas tales como proteínas y ácidos
nucleicos. La electroforesis es un sistema bioquímico desnaturalizante y discontinuo. Es
discontinuo porque está conformado por dos geles de poliacrilamida de resolución y
empacamiento que presentan diferente concentración, composición y pH. Ambos geles se
encuentran unidos pero limitados por una fase de separación visible al trasluz. Debido a que
en estado líquido ambos geles pueden mezclarse fácilmente, deben ser preparados por
separado esperando la total polimerización del primero para continuar con la polimerización
del otro. El gel de empacamiento generalmente se localiza en la parte superior del sistema
formando los pocillos donde se depositarán las muestras. El gel de resolución por su parte
forma el cuerpo del gel por donde migrarán y se separarán las proteínas.
La electroforesis bidimensional 2D-PAGE permite separar hasta miles de proteínas en un
solo ensayo y constituye actualmente el método más eficiente para la separación de mezclas
muy complejas de proteínas. Se basa en una separación de proteínas en función de la carga,
seguida de una separación de las proteínas en función de su masa molecular. Por otra parte,
al aplicar un campo eléctrico a un medio que contiene partículas cargadas, las partículas
cargadas negativamente migran hacia el ánodo o polo positivo mientras que, las cargadas
positivamente migran hacia el electrodo negativo o cátodo. Este principio se puede utilizar

para separar las fracciones de proteínas puesto que los aminoácidos constituyentes de las
proteínas, y por tanto las proteínas, son compuestos anfóteros que se comportan como
ácidos cediendo protones y quedan con carga negativa o bases que captan protones y
quedan con carga positiva dependiendo del medio. En este estudio se utilizó la electroforesis
en una dimensión para analizar las proteínas intrínsecas y extrínsecas de la membrana
plasmática de los eritrocitos.
OBJETIVO GENERAL
Analizar bioquímicamente las proteínas de la membrana citoplásmica expresadas en los
eritrocitos.
OBJETIVOS ESPECÍFICOS
1. Obtener membranas celulares de los eritrocitos
2. Extraer proteínas intrínsecas y extrínsecas de las membranas plasmáticas de
eritrocitos.
3. Examinar las proteínas presentes en las membranas citoplásmicas de eritrocitos
bioquímicamente.
PLANTEAMIENTO DEL PROBLEMA
La dificultad en separar las proteínas intrínsecas y extrínsecas de la membrana plasmática
de las células, es la presencia de otras proteínas semejantes pertenecientes a las
membranas de otros organelos de una o dos membranas. Por lo que se requiere de una
estirpe celular que permita obtener las proteínas específicas de la membrana plasmática.
Para este trabajo se consideró que el eritrocito es una célula que posee el sistema estructural
de las membranas más simple de todas las membranas celulares que permitirá la obtención
de las proteínas de interés donde se analizaran las proteínas que pertenecieran en forma
exclusiva de la membrana citoplásmica utilizando un análisis bioquímico con electroforesis en
una dimensión.
HIPÓTESIS
Si se libera el contenido citoplásmico del eritrocito entonces se obtendrán proteínas de
membrana plasmática.

DESARROLLO
Obtención de eritrocitos y las membranas plasmáticas “fantasmas”. La extracción de la
sangre conteniendo los eritrocitos se obtuvo por punción directa en la vena yugular de un
carnero adulto. La sangre obtenida se colocó en un matraz Erlenmeyer estéril conteniendo
una solución de Alsevers como anticoagulante y conservador para los eritrocitos. La solución
consistió en glucosa 2.05 %, cloruro sódico, 0.42 %, citrato sódico 0.8 % y citrato, 0.055 % en
agua destilada y se esterilizó previamente por calor en autoclave 15 min a 115 lb de presión.
Posteriormente, la solución conteniendo los eritrocitos se mantuvo a 4°C hasta su uso.
Se tomaron alícuotas de 30 ml en tubos cónicos de 50 ml (Falcon); se nivelaron en una
balanza granataria de dos platos con las camisas de la centrífuga para nivelarlos. Se
centrifugaron a 1500 rpm, a 4°, por 20 minutos.
El sobrenadante se retiró por decantación y los eritrocitos se obtuvieron en la parte de inferior
del tubo. Los eritrocitos se resuspendieron en una solución isotónica y se lavaron 3X para
retirar los restos de Alsevers en las mismas condiciones de centrifugación.
Figura 3. Obtención de las proteínas intrínsecas y extrínsecas de eritrocitos.
En seguida se lavaron 3X con una solución hipotónica para liberar la hemoglobina de las
membranas. Cuando el sobrenadante no mostró restos de hemoglobina, las muestras se
lavaron nuevamente 3X con solución isotónica, se eliminó el sobrenadante y el pellet se
resuspendió con 1 ml de solución de PBS con glicerol al 10% y se congelaron a -20°C hasta
su uso.

Análisis de las proteínas extrínsecas e intrínsecas de la membrana celular de los
eritrocitos.
Se empleó la técnica de electroforesis para la separación y visualización de las proteínas a
través de la migración de los polipéptidos como solutos iónicos bajo la influencia de un
campo eléctrico; estas partículas migran hacia el cátodo o ánodo (electrodos - y +), en
dependencia de una combinación de su carga, peso molecular y estructura tridimensional. Se
prepararon dos geles de poliacrilamida con SDS, Acrilamida-bis acrilamida-SDS (SDS-
PAGE) Fig. 4 y 5, considerando las proporciones del gel resolutor y del gel concentrador,
para concentraciones de 5% y 10 % Figs. 4 y 5. El gel concentrador funcionó como la matriz
de poliacrilamida que retendría a las proteínas manteniéndolas uniformes antes que ellas
migraran hacia el gel de resolución. Fig. 5.
Figura 4. Secuencia del ensamblaje del sistema electroforético de los geles. Limpieza de los vidrios, introducción en el soporte de vidrios, cierre hermético, montaje en el soporte del conjunto del marco de sujeción y electrodo
El gel resolutor fue la matriz de poliacrilamida donde las moléculas de proteínas migraron
para generar un perfil de bandas o patrón electroforético, el patrón varió de acuerdo al peso
molecular de cada una de las moléculas y a la concentración de los geles, se utilizó una guía
para ensamblar los vidrios para polimerizar los geles Figs. 4, 5 y 6.

Figura 5. Gel de acrilamida conteniendo el gel separador y el gel concentrados para corrimiento electroforético.
Después se ensamblaron en los marcos y se introdujeron en la placa de sujeción, la cual se
colocó en la cámara, se conectó a la fuente de poder y se aplicó una corriente de 100 mV,
Fig. 6.
Los geles se ensamblaron en el equipo conteniendo la placa de sujeción, el soporte, la
cámara con amortiguadores de corrimiento. El corrimiento se realizó hasta justo antes de que
el frente de corrimiento saliera de los geles dentro de las placas de sujeción.
Terminado el corrimiento los geles se tiñeron con el colorante de Azul de Coomassie en
solución con ácido acético- alcohol durante toda la noche. Al día siguiente, los geles se
extrajeron y se colocaron en solución desteñidora ácido acético-etanol hasta el revelado de
las bandas proteicas.

Figura 6. Equipo utilizado para el corrimiento electroforético. Gel que fue introducido en la cámara y conectado a la fuente de poder con un voltaje de 100 mV.
RESULTADOS
Con la metodología experimental empleada utilizando un choque osmótico se liberó el
contenido del citoplasma de los eritrocitos y se obtuvieron las membranas de las células
eritrocíticas libres de otras estructuras celulares. Así mismo, la técnica de electroforesis en
geles de poliacrilamida con SDS (SDS-PAGE) en una dimensión fue capaz de separar la
mezcla compleja de proteínas integrales y extrínsecas presentes en la membrana plasmática
de los eritrocitos de carnero. Este proceso permitió separar las proteínas en base a su peso
molecular, ya que el detergente SDS las carga negativamente y todas migran hacia el polo
positivo Figs. 7 A y B.
En la misma Fig. 7 se pueden observar dos perfiles electroforéticos, donde se muestran las
bandas teñidas con azul de Coomassie que marcan cada proteína presente en las
membranas de los eritrocitos. En A. se observa el corrimiento al 6% que permitió el mejor
corrimiento por la consistencia del gel. En B. se observaron 15 proteínas de membrana, pero
tomando como referencia la espectrina, que es la proteína más abundante de los eritrocitos,

sus isoformas corrieron más separadas en el gel al 6% respecto al 10 %, sin embargo, se
aprecia menor número de proteína que en el gel al 10 %.
A B
Figura 7. Electroforesis en gel de acrilamida. A. al 6 % B. Al 10 %. Teñidas con azul de Coomassie.
Las proteínas extraídas de las membranas de eritrocitos de carnero se analizaron por SDS-
PAGE al 6 y 10 % en presencia de β-mercaptoetanol y se tiñeron con azul de Coomassie y
se revelaron los componentes proteicos principales. Las espectrinas α y β localizadas al
inicio en ambos geles, al 6% se observó mayor separación entre ambos polipéptidos debido
a la porosidad de la matriz del gel que a dicha concentración permitió una mejor migración de
las proteínas respecto al corrimiento al 10%. Las espectrinas α y β, se encontraron muy
cercanas en el gel al 10%, y se observaron proteínas de menores masas moleculares como
las Anquirinas de masa molecular de 116 KDa, la proteína de intercambio aniónico Banda 3,
las Glucoforinas con pesos moleculares que oscilan entre 72 y 131 KDa, la Actina de 44
KDa, la Globina . Las Espectrinas α y β son las proteínas principales del citoesqueleto en los
eritrocitos, son un tetrámero constituido por dos cadenas polipeptídicas diferentes se localiza
en la superficie de la membrana interna en asociación con glicoproteínas que atraviesan la
membrana y con partículas intramembrana, y tienen masas moleculares de 220 y 240 KDa
respectivamente. La proteína de unión a la membrana para espectrina se denominó
anquirina, y se ha purificado y caracterizado. Se ha demostrado que la anquirina está
estrechamente asociada en extractos detergentes de vesículas con banda 3, un polipéptido
que abarca la membrana principal, y se une directamente a un fragmento proteolítico
derivado del dominio citoplasmático de la banda 3. Cuadro 1.

Al estudiar las proteínas de la membrana plasmática del eritrocito humano mediante
electroforesis en gel de poliacrilamida con SDS, se detectan aproximadamente 15 bandas
proteicas principales, cuyas masas moleculares van de 15 y 250 KDa. Tres de estas
proteínas (la espectrina, la glucoforina y la banda 3) constituyen más del 60% (en peso) de la
proteína total de la membrana. [Alberts, 2007].
Para mantener la forma característica del eritrocito, el citoesqueleto de espectrina-actina se
une firmemente a la membrana plasmática por dos proteínas periféricas, cada una de las
cuales se une a una proteína de membrana específica y a los fosfolípidos de membrana. La
anquirina conecta el centro de la espectrina a la proteína integral banda 3, una proteína de
transporte de aniones en la membrana.
Cuadro 1. Los resultados muestran las masas moleculares aparentes de las principales bandas de proteínas observadas en electroforesis SDS-PAGE en condiciones reductoras al 6 y 10 % en estos ensayos.
PROTEÍNA PM kDa
Espectrina α 240
Espectrina β 220
Anquirina y 3 Isoformas 116
Glucoforinas A,B,C,D 131-72
Banda 3 90-100
Actina 44
La proteína banda 4.1, un componente de los vértices de la red, se une a la proteína integral
glucoforina. Por otra parte, tanto la anquirina como la banda 4.1 contienen motivos de unión
a lípidos, que ayudan a unirlas a la membrana. Esta unión doble a través de la anquirina y la
banda 4.1 garantiza que la membrana está conectada tanto a los lados como a los vértices
de la red del citoesqueleto de espectrina y actina [Lodish, 2008].
El tipo celular con el que se realizó la técnica de electroforesis, los eritrocitos tienen ventajas
por ser los elementos cuantitativamente más numerosos de la sangre, son fáciles de obtener
libres de otros tipos celulares. Al carecer de núcleo y de otras organelos intracelulares
rodeados por membrana los convierte en un sistema ideal para el estudio de las membranas
biológicas, siendo la membrana plasmática la única membrana que poseen y no están

contaminados por otros tipos celulares. Se encuentran como células individuales y no es
necesario separarlas de un tejido complejo. Además de que se pueden obtener membranas
plasmáticas purificadas de manera intactas, tan solo con colocar las células en una solución
salina diluida (hipotónica). (Karp,2011)
La espectrina, junto con la actina, son los componentes principales del citoesqueleto cortical
en los eritrocitos. Las espectrinas eritrocitarias, denominadas α y β, tienen masa molecular
de 240 y 220 kDa, respectivamente. La cadena β presenta un único dominio de unión a la
actina en la región amino terminal. Las cadenas α y β se asocian lateralmente para formar
dímeros que se unen cabeza con cabeza para formar tetrámeros con dos dominios de unión
a la actina separados aproximadamente por 200 nm. También se observó el intercambiador
aniónico Banda 3. Los extremos de los tetrámeros de espectrina se asocian con filamentos
cortos de actina, dando como resultado una red de espectrina-actina que forma el
citoesqueleto cortical de los glóbulos rojos. (Rodi, 2010). El análisis bioquímico de proteínas
de membranas de eritrocitos de carnero en SDS-PAGE teñidas con plata, revelan otras
proteínas no se observan con tinción de Azul de Coomassie.
ANÁLISIS E INTERPRETACIÓN DE RESULTADOS
El componente de mayor masa molecular en la membrana del eritrocito fue la espectrina, los
tetrámeros de espectrinas están unidos con la membrana por una proteína llamada
anquirina, la cual está conectada a la banda 3. La banda 4.2 estabiliza la unión entre la
anquirina y el intercambiador aniónico banda 3. La espectrina se une también con la
glicoforina C mediante la banda 4.1; este entramado se ancla en múltiples sitios de la
membrana. La banda 4.1 y la aducina, estabilizan la asociación de la espectrina con la
actina. Las subunidades de la actina forman microfilamentos con la tropomiosina, a los que
se asocia la proteína tropomodulina. La banda 4.9 o dematina, produce el entrecruzamiento
de estos microfilamentos de actina. La estructura de la doble capa lipídica es fundamental en
la organización del citoesqueleto en la membrana del eritrocito.
Otros de los componentes de la membrana del eritrocito anclada a red de proteínas que
forman el citoesqueleto son lípidos el 95% de lípidos son fosfolípidos, colesterol no
esterificado. Los fosfolípidos son la fosfatidilcolina (lecitina), 13% de esfingomielina 26%
fosfatidiletanolamina (cefalina), 27% fosfatildilserina, 13% fosfatidilinositol 5% Colesterol.
Ácidos grasos. Glucolípidos.

Las proteínas de la membrana son proteínas integrales, total o parcialmente sumergidas en
la bicapa lipídica. Las proteínas periféricas ubicadas fuera de la bicapa. Las más importantes
que forman el esqueleto, tienen interés clínico y las enfermedades asociadas se llaman
membranopatías.
CONCLUSIONES
El análisis bioquímico de las proteínas de membrana del eritrocito con la técnica de
electroforesis desnaturalizante o SDS-PAGE permitió la identificación 15 proteínas en geles
al 10 % y menor cantidad en geles al 6 %. Las espectrinas α y β, banda 3, glicoforinas, actina
y tubulina, fraccionadas de acuerdo con su masa molecular en una matriz de poliacrilamida,
usando un campo eléctrico.
PERSPECTIVAS
Las proteínas de membrana plasmática en células eucariontes tienen importancia en los
mecanismos de invasión por microorganismos. Se sabe que toxina codificada en plásmido
(Pet) de Escherichia coli enteroagregativa (EAEC) produce daño intestinal. Pet pertenece a la
subfamilia de proteínas autotranspotadoras con actividad de serina proteasa de
Enterobacteriaceae (SPATEs). Esta toxina de induce efectos enterotóxicos (secreción de
líquidos) y citotóxicos (daño celular). El blanco celular de Pet son proteínas del citoesqueleto
membranal que funcionan de anclaje a los microfilamentos de actina. Ensayos con
membranas de eritrocitos o de células epiteliales HEp-2 purificadas y tratadas con toxina de
Pet, se ha observado proteólisis de las cadenas de α y β espectrina y fodrina (espectrina no
eritroide), generándose un producto de degradación de 120 kDa, que alterá la estructura de
los microfilamentos de actina. Este es un nuevo modelo de daño celular mediado por
SPATEs de EAEC, que incluye la internalización de la proteasa, la inducción de algunos
pasos de señalización intermedios desconocidos y, finalmente, la degradación de la
espectrina/fodrina para destruir a la célula (Villaseca, 2000).
BIBLIOGRAFÍA
1. Alberts, B. (2010). Biología Molecular de la Célula. Ed. Omega, Barcelona.

2. Avers. J CH. (1994). Biología Celular. 2da. Ed. Iberoamericana. Madrid.
3. Karp, G. (2011). Biología Celular y Molecular McGraw-Hill Interamericana, 746 pág.
4. Salazar M. AM, Sandoval R. AS, Armendariz B. JS. (2016). Fundamentos de Biología
molecular Ed. McGraw-Hill Interamericana. México.
5. Villaseca, Jorge M., Fernando Navarro-García, Guillermo Mendoza-Hernández, James P.
Nataro, Alejandro Cravioto y Carlos Eslava. Pet Toxin from Enteroaggregative Escherichia
coli Produces Cellular Damage Associated with Fodrin Disruption Infection and immunity,
2000, p. 5920–5927 Vol. 68, No. 10
CIBERGRAFÍA
http://cbs.izt.uam.mx/nacameh/v9n2/Nacameh_v9n2_077_PerezCh-etal.pdf http://www.ciens.ucv.ve:8080/generador/sites/lab-bioq-gen/archivos/Lomonte%20-%20Cap13%20PAGE.pdf http://www.fcn.unp.edu.ar/sitio/quimicabiologica1/wp-content/uploads/2011/04/SDS-PAGE.pdf https://ibcmunq.files.wordpress.com/2010/03/tp3.pdf https://www.uco.es/dptos/bioquimica-biol-mol/pdfs/16%20ELECTROFORESIS%20GELES%20PAA.pdf