“CARACTERIZACIÓN AMBIENTAL Y ESTRUCTURA TRÓFICA … · A todos los compañeros que integraron e...
138
“CARACTERIZACIÓN AMBIENTAL Y ESTRUCTURA TRÓFICA DEL HÁBITAT DE LA VAQUITA MARINA, Phocoena sinus”. TESIS QUE PARA OBTENER EL GRADO DE DOCTORA EN CIENCIAS MARINAS PRESENTA MÓNICA YANIRA RODRÍGUEZ PÉREZ LA PAZ, B.C.S., DICIEMBRE DE 2013 INSTITUTO POLITÉCNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
Transcript of “CARACTERIZACIÓN AMBIENTAL Y ESTRUCTURA TRÓFICA … · A todos los compañeros que integraron e...

“CARACTERIZACIÓN AMBIENTAL Y ESTRUCTURA TRÓFICA DEL HÁBITAT DE LA
VAQUITA MARINA, Phocoena sinus”.
TESIS
QUE PARA OBTENER EL GRADO DE
DOCTORA EN CIENCIAS MARINAS
PRESENTA
MÓNICA YANIRA RODRÍGUEZ PÉREZ
LA PAZ, B.C.S., DICIEMBRE DE 2013
INSTITUTO POLITÉCNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS



“Courage isn’t about having the strength or perseverance to go on……..it’s
about going on when you don´t have the strength but only the sheer will to live”
Ken Allan Dronsfield
Gracias por darme la fuerza para no perder la
voluntad…….………….
Para Ofe y Sayu

AGRADECIMIENTOS PERSONALES Quiero gradecer muy especialmente a mis queridos amigos por haber enriquecido mi
vida y permitirme formar parte de la suya: Vilma, Cris, Dany, Paty, Dieguito, Fer, Mar,
Sand, Ramón, Lyg, Kary, Haniel, Susan. Espero que esta amistad perdure por
siempre…………
A mis más grandes amores por haber soportado mis ausencias y olvidos pero
siempre a mi lado dándome la fortaleza para seguir adelante: Mom, Kaly, Laiky, Ofe,
Sayu, a mis hermanas, a José………

AGRADECIMIENTOS
Agradezco al Centro Interdisciplinario de Ciencias Marinas (CICIMAR), del Instituto
Politécnico Nacional (IPN) por la oportunidad que me brindo durante estos cuatro
años para poder dar un paso más en mi formación académica.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el apoyo económico
otorgado durante mis cuatro años de doctorado, así como por la Beca Mixta para
movilidad en el extranjero.
Al Programa Institucional de Formación de Investigadores por su apoyo durante los
dos primeros años de este proceso.
A la Fundación Rositter: Cetacean Society International, por su apoyo económico
durante mi estancia en la Universidad de Laramie, Wyoming.
A la Fundación David y Lucile Packard (contrato 2010-36137) a traves de la
Universidad de Arizona por el proyecto Pesca Artesanal del Norte del Golfo de
California: Ambiente y Sociedad (PANGAS) (http://pangas.arizona.edu), porque sin
su apoyo incondicional en la logística de este proyecto nada hubiera sido posible.
Al proyecto SEP-CONACyT (contrato 2008-105922) y al Instituto Politécnico Nacional
(Proyecto multidisciplinario 2013-1581), por su apoyo económico para la realización
de este estudio.
Quiero expresar un profundo agradecimiento a mis directores de tesis: Al Dr. David
Aurioles Gamboa y a la Dra. Laura Sánchez Velasco por su guía y su confianza, por
compartir su experiencia enseñándome que la perseverancia y la disciplina son el
camino para llegar a las metas planteadas.

Al Dr. Miguel Lavín por su apoyo logístico y su confianza incondicional durante este
proceso, así como por sus valiosas aportaciones hechas a este trabajo.
Al Dr. Sergio Martínez Díaz, Dra. Claudia Janetl Hernández Camacho y Dr. Francisco
García Rodríguez por el seguimiento, revisión y valiosas observaciones hechas a
este trabajo.
Al equipo de trabajo de PANGAS: con especial énfasis al Dr. Víctor Godínez por las
facilidades otorgadas para la obtención de las muestras de variables ambientales, así
como por su ayuda en la toma de muestras durante los cruceros.
A toda la tripulación del Buque Oceanográfico Francisco de Ulloa por las facilidades
otorgadas durante los muestreos.
Al Laboratorio de Química Marina, especialmente a la M en C. Sonia Futema
Jiménez por su ayuda durante el procesado de las muestras.
Al personal del Laboratorio de Computo por todas las facilidades otorgadas durante
el doctorado.
Al Laboratorio de Ictiología por su valiosa ayuda durante la identificación de todas las
especies de peces colectadas, con especial énfasis al Dr. Victor Manuel Cota Gómez
por su paciencia, apoyo y guía.
De manera muy especial agradezco toda la ayuda otorgada, no sólo en el ámbito
académico, sino también en el personal, por todos sus consejos y guía durante este
periodo al C.P. Humberto Ceseña Amador, gracias por haberme apoyado
incondicionalmente aun a larga distancia y en tus días más saturados, siempre con
una sonrisa amable.

Al C. Cesar Servando Casas Nuñez, por toda la ayuda otorgada para hacer posible
que todos los tramites salieran a tiempo en cada paso del doctorado.
Al Dr. Armando Jaramillo-Legorreta y a su esposa Edwina por la aportación
bibliográfica otorgada.
Al Dr. Seth Newsome por sus valiosas enseñanzas y su paciencia infinita durante mi
estancia en Laramie, por enseñarme a ver la Ciencia de una manera diferente……….
Pero sobre todo por tu apoyo…….
A la Dra. Sora Kim, por su guía y confianza, por haberme hecho sentir en casa
durante mi estancia en Wyoming.
A la Universidad de Wyoming por abrirme sus puertas y darme todo el apoyo para
poder enriquecer mi vida profesional y personal al tener la oportunidad de compartir
con personas que nunca imagine como el Dr. Carlos Martínez del Río, Dr. Mark
Clementz, Dr. Paul Koch…….
A todos los compañeros que integraron e integran los dos equipos de trabajo en los
cuales participé y que me enseñaron tanto durante esta fase de mi formación
académica: Al Laboratorio de Ecología de Pinnípedos “Burney J. LeBoeuf” y al
Laboratorio de Plancton, pero sobre todo gracias por su amistad y compañía.

ÍNDICE
ÍNDICE DE FIGURAS i
ÍNDICE DE TABLAS v
GLOSARIO vii
RESUMEN xii
ABSTRACT xiii
1. INTRODUCCIÓN GENERAL 1
2. ANTECEDENTES
2.1 Phocoena sinus 5
2.1.1 Morfología 5
2.1.2 Biología 6
2.1.3 Ecología 7 2.1.4 Alimentación 7
2.1.5 Caracterización del hábitat 8
3. ÁREA DE ESTUDIO 9
3.1 Mareas 10
3.2 Hidrografía y Corrientes 11
3.3Sedimentos 11
4. JUSTIFICACIÓN 12
5. HIPÓTESIS 13
6. OBJETIVOS 14

7. MÉTODO GENERAL 15
7.1 Obtención y procesamiento de muestras 16
7.1.1 Muestras ambientales 16
7.1.2 Sedimento 16
7.1.3 Fitoplancton 17 7.1.4 Zooplancton 18
7.1.5 Procesamiento de macrofauna y tejido óseo de Phocoena sinus 19
8. CAPÍTULO 1
CARACTERIZACIÓN AMBIENTAL DEL ÁREA DE MAYORES AVISTAMIENTOS DE
Phocoena sinus
8.1 INTRODUCCIÓN 20
8.2 METODOLOGÍA 22
8.2.1 Tratamientos de datos ambientales 22
8.2.2 Análisis estadísticos entre variables ambientales 22
8.2.3 Caracterización del hábitat potencial de Phocoena sinus 23
8.3 RESULTADOS
8.3.1 Diferencias espaciales 24
8.3.2 Variaciones temporales en el Alto Golfo de California 24
8.3.3 Delimitación del hábitat potencial de Phocoena sinus 32
8.4 DISCUSIÓN
8.4.1 Caracterización espacial del Alto Golfo de California 42
8.4.2 Delimitación del hábitat potencial de Phocoena sinus 44
8.4.3 Variaciones temporales de las variables ambientales en el 46
Alto Golfo de California

9. CAPÍTULO 2
REGIONES ISOTÓPICAS DE CARBONO Y NITRÓGENO EN EL ALTO GOLFO DE
CALIFORNIA COMO TRAZADORES DE LA LINEA BASE DE Phocoena sinus
9.1 INTRODUCCIÓN 49
9.2 METODOLOGÍA
9.2.1 Diferencias temporales en δ13C y δ15N de la línea base del 52
Alto Golfo de California
9.2.2 Isomapas de δ13C y δ15N en sedimento y zooplancton del 52
Alto Golfo de California
9.2.3 Delimitación isotópica del área de alimentación de Phocoena sinus 53
9.3 RESULTADOS
9.3.1 Diferencias temporales de δ13C y δ15N en plancton del 54
Alto Golfo de California
9.3.2 Características isotópicas del sedimento 58
9.3.3 Diferencia espacial de δ13C y δ15N en plancton y sedimento 59
9.3.4 Área de alimentación de Phocoena sinus a partir de δ13C y δ15N 63
9.4 DISCUSIÓN
9.4.1 Diferencias isotópicas temporales en plancton del 66
Alto Golfo de California
9.4.2 Diferencias espaciales en las regiones isotópicas obtenidas del 67
sedimento y plancton en el Alto Golfo de California
9.4.3 Diferencias isotópicas temporales en la línea base del 70
Alto Golfo de California
9.4.4 Diferencias espaciales de δ13C and δ15N en la trama trófica 71

10. CAPÍTULO 3
ESTRUCTURA TRÓFICA Y AMPLITUD DE NICHO DEL HÁBITAT DE Phocoena
sinus
10.1 INTRODUCCIÓN 74
10.2 METODOLOGÍA
10.2.1 Determinación del nivel trófico 76
10.2.2 Contribución de presas potenciales a la dieta de Phocoena sinus 77
10.2.3 Análisis de amplitud de nicho 78
8.3 RESULTADOS
8.3.1 Isótopos estables de δ13C y δ15N en Phocoena sinus 79
8.3.2 Estructura de la trama trófica en el hábitat de Phocoena sinus 81
a partir de isótopos de δ13C y δ15N
8.3.3 Amplitud de nicho y determinación de dieta de la Phocoena sinus 83
8.4 DISCUSIÓN
8.4.1 Señales de δ13C y δ15N en Phocoena sinus 87
8.4.2 Estructura de la trama trófica de Phocoena sinus 88
8.4.3 Nicho trófico y proporción de dieta de Phocoena sinus 90
11. CONCLUSIONES GENERALES 92
12. BIBLIOGRAFÍA 94

i
INDICE DE FIGURAS
Figura 1: Reserva de la Biósfera del Alto Golfo de California y Delta del Río Colorado,
su límite sur lo representa la línea punteada roja. El polígono azul muestra el área de
refugio para la vaquita marina y las líneas negras representan la batimetría del área.
5
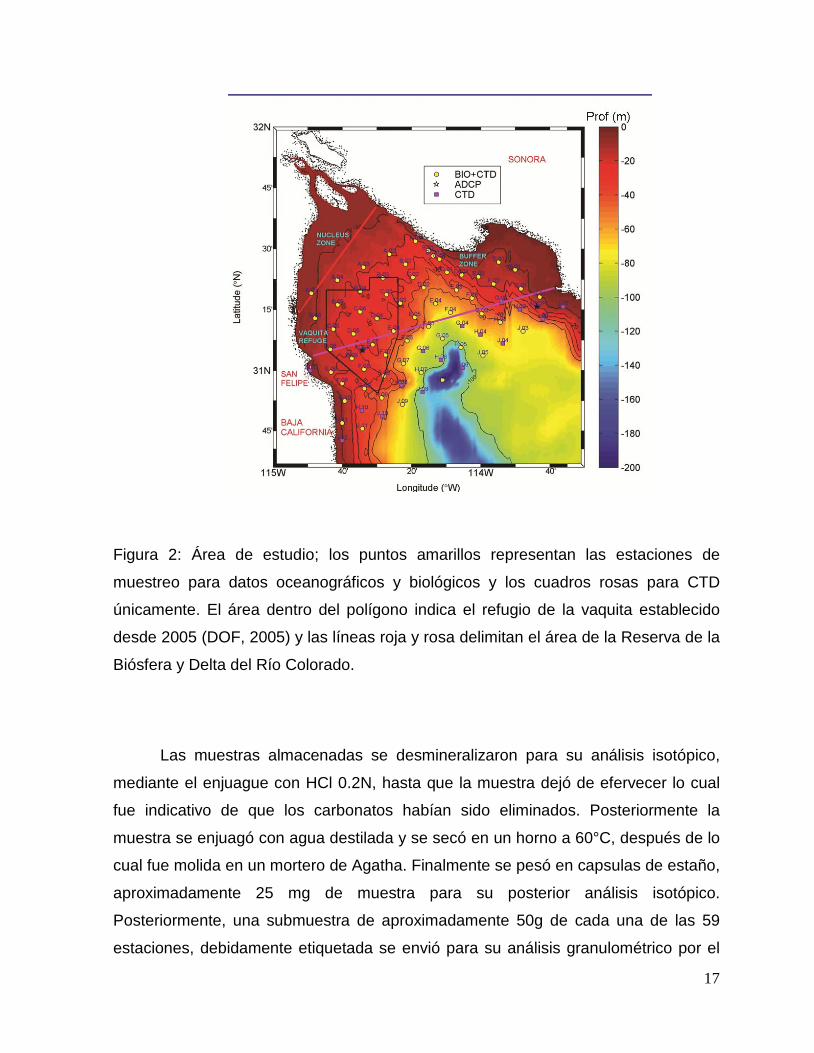
Figura 2: Área de estudio; los puntos amarillos representan las estaciones de
muestreo para datos oceanográficos y biológicos y los cuadros rosas para CTD
únicamente. El área dentro del polígono indica el refugio de la vaquita establecido
desde 2005 (DOF, 2005) y las líneas roja y rosa delimitan el área de la Reserva de la
Biósfera y Delta del Río Colorado. 17 Figura 3. Resultado del análisis de grupos mediante el programa SIMPROF (Similarity
profiles) con base en temperatura (°C), Salinidad (ups), Turbidez (mg/L), Granulometría (%),
O2 (Disuelto) (mL/L), Chl “a” (μg/L) y Profundidad (m) para el mes de junio-2010 (A) y marzo-
2011 (B). La definición de los ambientes resultantes se muestra en C y D. Los colores en el
mapa corresponden a las estaciones marcadas con el corchete del mismo color en los
clúster. Los círculos en el mapa corresponden a los avistamientos por intervalos de
frecuencia de aparición desde el 1992 hasta 2007 (Tomado de Villa-Ramírez et al., 1995;
Guerrodette et al., 1995; Jaramillo-Legorreta, 2008). Por fuera del refugio se enmarca en
negro el área propuesta en el presente trabajo, con una extensión de ~530km2 más que el
refugio actual. 26
Figura 4. Batimetría del Alto Golfo de California. Los puntos negros señalan las estaciones
de muestreo y el polígono corresponde al área de refugio de la vaquita marina. 30
Figura 5. Granulometría del Alto Golfo de California. A). Análisis TERNPLOT que define tres
grupos, los cuales se enmarcan dentro de los recuadros. El color de cada uno de los
recuadros corresponde a los observados al tamaño de partícula señalado en sus vértices. B).
Grupos granulométricos arrojados por el análisis TERNPLOT. 31
Figura 6. Concentración de oxígeno disuelto promedio en la columna de agua durante en
junio (A) y marzo (B) en el Alto Golfo de California. 35

ii
Figura 7. Gradiente promedio de turbidez en la columna de agua durante en junio (A) y
marzo (B) en el Alto Golfo de California. 36
Figura 8. Gradiente promedio de temperatura en la columna de agua durante junio (A) y
marzo (B) en el Alto Golfo de California. 38
Figura 9. Gradiente promedio de salinidad en la columna de agua durante en junio (A) y
marzo (B) en el Alto Golfo de California. 39
Figura 10. Concentración promedio de clorofila “a” en la columna de agua durante junio (A) y
marzo (B) en el Alto Golfo de California. 40
Figura 11. Biomasa zooplanctónica en el Alto Golfo de California durante junio (A) y marzo
(B). 42
Figura 12. Series de tiempo de A). Clorofila y B). Temperatura superficial del mar. Ambas
abarcan un periodo de 10 años y cada color representa las tres áreas arrojadas por el
análisis SIMPROF para el Alto Golfo de California. Verde (Sur), azul (Oeste) y rojo (Este).
Fuente: Aqua/MODIS; http://oceancolor.gsfc.nasa.gov. 43
Figura 13. Isomapas de δ13C en fitoplancton (A y B) y zooplancton (C y D) durante junio
(2010) y marzo (2011). Los puntos negros representan las estaciones de muestreo y el
polígono la zona de refugio de la vaquita marina. 60
Figura 14: Isomapas de δ15N en fitoplancton (A y B) y zooplancton (C y D) durante junio
(2010) y marzo (2011). 61
Figura 15: Gradiente isotópico de carbono (A) y nitrógeno (B) en sedimento muestreado
durante junio 2010. 63
Figura 16: A) Análisis clúster de afinidad por grupo de estaciones obtenido mediante el índice
de disimilitud de Bray Curtis y B) distribución de las regiones arrojadas por el análisis. 65

iii
Figura 17: ACC donde se relacionan las variables ambientales con las regiones isotópicas
66
Figura 18. Media y desviación estándar de los valores de δ13C y δ15N en sedimento y
muestras biológicas colectadas en el Alto Golfo de California. Los diamantes blancos,
representan las muestras colectadas en la región Este y los diamantes negros la región
Oeste. El círculo gris, representa la media y desviación estándar de la señal isotópica
encontrada en colágeno óseo del cráneo de vaquita marina. La línea punteada conecta el
valor real de la especie con el valor corregido (círculo blanco) por un factor de discriminación
de 3.1‰ y 2.04‰ para δ13C y δ15N respectivamente, de acuerdo a Borell et al. (2012).
Sedimentos (Sed), fitoplancton (Phy), zooplancton (Zoo), Chione (Chi), Atrina (Atr), Mugil
(Mgl), Anchoa (Ach), Oligoplictes (Olg), Cynoscion (Cyc). 70
Figura 19. Valores de δ13C y δ15N de materia orgánica en sedimentos, fitoplancton y varios
organismos marinos colectados en los alrededores de Rocas Consag. En el eje Y se indican
las ubicaciones de nivel trófico y en diamante azul la posición de la vaquita marina. 89
Figura 20. Amplitud de nicho obtenida mediante el área Convex-Hull de los valores
isotópicos de la vaquita marina. 90
Figura 21. Diagramas de cajas obtenidos mediante el modelo de mezcla SIAR, que
representan el rango de contribución de cada uno de los grupos de presas potenciales a la
dieta de la vaquita marina. 92

iv
ÍNDICE DE TABLAS
Tabla 1: Resultados arrojados por el análisis Kruskal-Wallis para las variables muestreadas
durante la campaña de junio del 2010 y marzo del 2011. 27
Tabla 2: Análisis U Mann-Whitney aplicado a las variables muestreadas en el Alto Golfo de
California durante junio del 2010. 28
Tabla 3: Análisis U Mann-Whitney aplicado a las variables muestreadas en el Alto Golfo de
California durante marzo del 2011. 29
Tabla 4. Diferencias espaciales en las regiones ambientales arrojadas por el análisis
SIMPROF para junio (2010). En negritas se señalan aquellas que presentan mayor
variabilidad entre áreas. 32
Tabla 5. Diferencias espaciales en las regiones ambientales arrojadas por el análisis
SIMPROF para marzo (2011). En negritas se señalan aquellas que presentan mayor
variabilidad entre áreas. 33
Tabla 6. Promedio y rango de valores de las variables ambientales muestreadas durante
junio (2010) y marzo (2011) en el Alto Golfo de California. 34
Tabla 7. Características ambientales del área de refugio de la vaquita marina encontradas en
ambas temporadas de muestreo (junio-2010 y marzo-2011). 44
Tabla 8. Coordenadas del hábitat potencial de la vaquita marina en el Alto Golfo de
California. 46
Tabla 9. Diferencias estacionales en los valores de δ13C y δ15N en fitoplancton y zooplancton
del Alto Golfo de California. Las primeras dos filas corresponden a los valores isotópicos
encontrados en el sedimento muestreado en junio 2010. 59
Tabla 10. Correlaciones entre las variables ambientales y las regiones isotópicas de las dos
estaciones muestreadas (junio 2010 y marzo 2011). 68

v
Tabla 11. Media y desviación estándar de los valores de δ13C y δ15N encontrados en
sedimentos y muestras biológicas colectadas en los alrededores de Rocas Consag (Oeste) y
encontrados en la bibliografía pertenecientes a Bahía Adair (Este; Spackeen, 2009) del Alto
Golfo de California. 71
Tabla 12. Media y desviación estándar de los diferentes grupos biológicos muestreados en
los alrededores de Rocas Consag. 86
Tabla 13. Contribución de las diferentes especies de presas potenciales a la dieta de
Phocoena sinus a partir del modelo de mezcla Bayesiano (SIAR). 92

vi
GLOSARIO Abundancia: Número de individuos por unidad de superficie o de volumen.
Amplitud trófica. Selección de presas que conforma la dieta de un consumidor.
Área marina protegida. Área de terreno intermareal o submareal que junto con sus aguas
de superficie, flora, fauna y rasgos históricos y culturales acompañantes, haya sido
reservada bajo la ley u otros medios efectivos para proteger la totalidad o parte del entorno
comprendido.
Bentónico: Organismo que vive en el fondo oceánico o de cuerpos de agua dulce (bentos).
El bentos está formado por los organismos vegetales y animales que viven sobre el fondo,
semienterrados, fijos o que pueden moverse sin alejarse demasiado de él.
Cadena trófica: La cadena trófica es el diagrama lineal que describe el proceso de
transferencia de sustancias nutritivas a través de las diferentes especies de una comunidad
biológica, en el que cada uno se alimenta del precedente y es alimento del siguiente.
Una cadena alimenticia es la ruta alimenticia desde un productor hasta un consumidor final.
Competencia: Proceso mediante el cual organismos de la misma especie o distinta utilizan
un mismo recurso. La competencia puede darse por alimento, agua, territorio, o cualquier
otro que se vea limitado al ser utilizado por más de una especie.
Cetáceo: Orden de mamíferos adaptados a la vida acuática con cuerpo fusiforme,
extremidades delanteras transformadas en aletas, con la aleta caudal dispuesta
horizontalmente, sus orificios nasales comunican al exterior a través de un respiradero o
espiráculo, situado en la parte superior de la cabeza. El orden de los cetáceos se divide en
dos subórdenes: Odontocetos y mistacocetos.
Clorofila-a. Es uno de los pigmentos más comunes presente en los organismos que realizan
fotosíntesis, cuyo resultado es la liberación de oxígeno. Se usa como un estimador indirecto
de la biomasa fitoplanctónica o algal que se presenta en un ecosistema.

vii
(δ) Delta. Notación adoptada para expresar diferencias relativas en las razones de isótopos
entre una muestra y un estándar. Se expresa en unidades de partes por mil (‰).
δ13C. Relación del isótopo pesado (13C) y el isótopo ligero (12C) de una muestra expresada
en partes por mil con respecto al del estándar Pee Dee Belemnite. Generalmente en
estudios de alimentación se usa para definir el tipo de ambiente que utilizan los
consumidores.
δ15N. Relación del isótopo pesado (15N) y el isótopo ligero (14N) de una muestra expresada
en partes por mil, con respecto al nitrógeno atmosférico.
Demersal: Organismo que vive muy próximo al fondo de cuerpos de agua marina o dulce.
Depredador especialista: Organismo que utiliza un número reducido de recursos a lo largo
del tiempo y presenta mayor preferencia por algunos componentes en particular.
Depredador generalista: Organismo que utiliza una gran variedad de recursos alimenticios
sin ninguna selección aparente.
Diversidad: Es la relación entre el número de especies y la abundancia relativa de las
mismas.
Ecosistema: Unidad fundamental de la biosfera que representa un nivel de organización que
funciona como un sistema, en el cual se manifiestan conjuntamente los factores abióticos
(físicos y químicos), así como los factores bióticos (los que provienen de los seres vivos).
Enriquecimiento isotópico. Término utilizado para describir la cantidad de un isótopo X con
respecto a una cantidad previa. Por ejemplo, una sustancia enriquecida en 15N o 13C estará
disminuida isotópicamente en 14N o en 12C respectivamente.
Especie: Grupo de poblaciones naturales real o potencialmente inter-reproductivas y
aislados reproductivamente de otros grupos análogos. Es un concepto que destaca la
capacidad de reproducción dentro de un sistema de poblaciones y el aislamiento
reproductivo frente a otras.

viii
Factor de discriminación isotópica. Aumento en la concentración de un isótopo estable
que se presenta en un consumidor respecto a su presa, resultado de una discriminación
metabólica hacia el isótopo pesado.
Flujo de materia y energía en el ecosistema: La combinación por donde fluye la materia y
circula la energía entre los componentes bióticos y abióticos en la naturaleza, es en el seno
mismo del ecosistema; la ruta de entrada de la energía se establece desde que los
autótrofos (generalmente fotosintéticos) producen materiales orgánicos que resultan la
fuente energética para los heterótrofos o consumidores, iniciando así el flujo de materia y de
energía en los sistemas vivos.
Hábitat: Espacio que reúne las características físicas y biológicas necesarias para la
supervivencia y reproducción de una o varias especies.
Isótopo. Átomo de un mismo elemento con igual número de protones (Z+) y
electrones (e-) pero diferente número de neutrones (N).
Isótopo estable. Isótopo que no cambia sus características químicas a lo largo del tiempo.
Nicho ecológico: El papel o función de una especie dentro del ecosistema. El nicho
ecológico se define en términos de las condiciones físicas y los recursos necesarios para la
supervivencia de la especie y su posicionamiento en la red trófica.
Nivel trófico. Posición o categoría en la que se clasifica un determinado organismo dentro
de la cadena trófica; dicha posición está dada de acuerdo a la manera en la cual obtiene su
materia y energía.
Protección: El conjunto de políticas y medidas para mejorar el ambiente y controlar su
deterioro (Ley GEEPA, 2000).
Superposición o traslape trófica: Uso compartido de recursos por dos especies, sexos o
poblaciones. Este traslape trófico no necesariamente se encuentra relacionado directamente
con la competencia.

ix
Tasa de recambio isotópico: Periodo en el que las razones isotópicas de un tejido reflejan
la señal o firma isotópica de una dieta en particular. Los tejidos con una tasa de recambio
isotópica rápida reflejarán una dieta reciente mientras que aquellos tejidos con una tasa de
recambio lenta reflejarán la dieta promedio representativa de un periodos mucho más
extenso.
Zooplancton: Organismos heterótrofos que viven en la columna de agua y que se alimentan
de otros organismos. Son los principales consumidores primarios

x
RESUMEN La vaquita marina (Phocoena sinus) es catalogada como uno de los cetáceos más
amenazados a nivel mundial, con una reducción en su población del 57% en los
últimos 11 años, registrando actualmente un numero poblacional de ~245 individuos
en el Alto Golfo de California. Su investigación se ha enfocado en estudios
demográficos dejando de lado aspectos ecológicos. En el presente estudio se
pretendió reforzar este aspecto con base en dos objetivos principales: 1) Caracterizar
y delimitar la extensión real de su hábitat y 2) Determinar su papel ecológico dentro
del ecosistema. Para esto se utilizará información de variables físicas, químicas y
biológicas, recolectadas en dos cruceros oceanográficos (julio-agosto 2010 y marzo-
abril 2011), en conjunto con la climatología de la región mediante imágenes
satelitales temperatura y clorofila; todo esto sumado a un análisis isotópico de la
especie y diversos componentes de su trama trófica. El análisis clúster basado en el
índice Bray-Curtis definió 3 regiones ambientales dentro del Alto Golfo de California
(Este, Oeste y Sur), además de 3 provincias granulométricas (Este, Centro y Oeste).
El análisis de perfiles de similitud arrojo 3 regiones isotópicas (Este, Oeste y Costa)
presentando diferencias significativas en sus proporciones isotópicas de δ 15N
(p>0.05), con los valores más enriquecidos en la región Oeste (10.7±2.0), con
relación a la región Este (7.2±2.3). Tanto las variables físicas, como las isotópicas
definieron las mismas dos regiones Oeste y Este. El mayor número de avistamientos
de la vaquita presenta una correlación positiva con sedimentos limos-arcillosos
(r2=0.4, p>0.05), con las proporciones isotópicas más enriquecidas (r2=0.6, p>0.05) y
con la mayor turbidez. Se obtuvo un fraccionamiento isotópico promedio del δ15N de
2.7‰ entre los 4 niveles tróficos que conforman la trama trófica de la vaquita, la cual
ocupó un nivel trófico de 4.0. Finalmente se puede concluir, que el área de mayores
avistamientos de la vaquita marina está bien delimitada en la región Oeste y presenta
características distintivas con respecto a la región Este y la Sur, las cuales se
conservan a lo largo del año.
Palabras clave: Alto Golfo de California, depredador tope, regiones ambientales,
isótopos estables.

xi
ABSTRACT
The vaquita (Phocoena sinus) is listed as one of the most endangered cetaceans
worldwide, with a reduction in its population of 57% in the last 11 years, currently
recording a population size of ~ 245 individuals in the Upper Gulf of California.
Research on it has focused on demographic studies, leaving aside important
ecological aspects. In this study part of the ecology of P. sinus is addressed focussing
on two main objectives: 1) To characterize and define the actual extent of the
vaquita’s habitat and 2) to determine its ecological role in the ecosystem. For this
study, we used physical, chemical and biological variables measured during two
cruises (July-August 2010 and March-April 2011). Additionally, we analized the
variability in the region through satellite images of salinity and temperature, all this
coupled with an isotopic analysis of the species and various food web components of
the vaquita´s habitat. A cluster analysis, based on the Bray-Curtis Index, defined
three environmental regions in the Upper Gulf of California (East, West and South),
plus 3 granulometric provinces (East, Central and West). The analysis of similarity
also produced three isotopic regions (East, West and Costa) showing significant
differences in δ15N isotope ratios (p > 0.05), with more enriched values in the West
region (10.7‰ ± 2.0‰) compared to the East region (7.2‰ ± 2.3‰). Both the
physical and the isotopic variables defined two large regions, the West and East. The
largest number of sightings of vaquita, positively correlated with silt-clay sediments (r2
= 0.4, p > 0.05), characterized with more enriched isotope ratios (r2 = 0.6, p > 0.05)
and higher turbidity. The estimated average of δ15N isotopic fractionation between 4
trophic levels was 2.7‰, with the vaquita occupying the trophic level 4.0. It can be
concluded that the largest area of vaquita sightings is well defined within the West
region and exhibits distinct characteristic with respect to the East and South regions,
which are persistent throughout the year.
Keywords: Palabras clave: Alto Golfo de California, depredador tope, regiones
ambientales, isótopos estables.

1
1. INTRODUCCIÒN GENERAL
México ocupa el tercer lugar entre los países con mayor biodiversidad en el
mundo; siendo a su vez el segundo país donde habita el mayor número de especies
de mamíferos (Mittermeier y Goettsch, 1992; Ceballos y Brown, 1995). En cuanto a
los mamíferos marinos, el Golfo de California alberga cerca del 40% de las especies
que habitan en el mundo (Enríquez-Andrade et al., 2005). Entre estas especies, una
de las que presenta mayor notoriedad a nivel mundial es la vaquita marina,
Phocoena sinus, debido a su carácter de especie rara y a que actualmente presenta
serios problemas de conservación encontrándose en critico peligro de extinción
(IUCN, 2008; NOM-059-ECOL-1994).
Debido a la vulnerabilidad de P. sinus y otras especies, principalmente por la
sobre-explotación pesquera, en 1993 se decretó oficialmente la Reserva de la
Biosfera del Alto Golfo de California y Delta del Río Colorado. Esta área se delimitó
con el fin de atender problemas como la disminución de la productividad pesquera y
la urgente protección de la vaquita y de la especie de sciaénido, Totoaba macdonaldi
(DOF, 1993). El área protegida está constituida por una zona núcleo denominada
Delta del Río Colorado dentro de una zona de amortiguamiento, cuyo límite al Sur
abarca desde San Felipe, Baja California hasta Puerto Peñasco, Sonora (DOF,
1993). Posteriormente, en el año 2005 se estableció el refugio para la protección de
la vaquita marina, el cual abarca una amplia porción del área reconocida como la
zona de mayor concentración de la especie, con una extensión de 1,263.85 km2
(DOF 2005; Figura 1). Pero desafortunadamente el establecimiento del refugio no ha
podido evitar que la población de la vaquita siga disminuyendo (Jaramillo-Legorreta,
2008).
A pesar de los amplios esfuerzos para su conservación, la vaquita marina es
la especie de cetáceo marino más amenazado a nivel mundial, con una población de
~245 individuos (CV=73%, 95%CI 68-884; Gerrodette et al. 2011). Los estudios
muestran que aunque en las últimas décadas se ha realizado un gran esfuerzo por

2
conocer el estatus y la ecología de la especie, los 25 años de desconocimiento
desde que se descubrió (1959) hasta que los estudios científicos iniciaron (1984),
fueron cruciales para que la población de P. sinus fuera diezmada y llevada casi a su
extinción (Silber, 1990). Dando como resultado que aspectos tan importantes como
su tamaño poblacional, la extensión real de su hábitat o su ecología trófica, hasta el
momento permanezcan desconocidos.
La vaquita marina, es una de las seis especies que actualmente conforman la
familia Phocoenidae; cuatro de las cuales pertenecen al género Phocoena (Gaskin et
al., 1984). En general, las marsopas presentan una distribución anti-tropical, teniendo
preferencia por aguas frías con menos de 17°C, costeras y altamente productivas
donde el alimento es muy abundante (Read, 1999; Ridgway y Harrison, 1999). A
diferencia del resto de las marsopas, la vaquita marina restringe su distribución a la
porción oeste del Alto Golfo de California (AGC), donde las temperaturas exceden los
30°C en verano (Silber, 1990; Gallo-Reynoso, 1998; Read, 1999). Debido a lo
anterior es considerada como el cetáceo marino con el área de distribución más
reducida a nivel mundial (Barlow, 1986; Brownell; 1986, Silber, 1990; Gerrodette et
al., 1995; Barlow et al., 1997; Gallo-Reynoso, 1998; Jaramillo et al., 1999; Vidal et al.,
1999; Jaramillo-Legorreta, 2008; Gerrodette et al., 2011). Esta restricción de hábitat
hace a la especie más vulnerable a cualquier impacto por la sobreexplotación
pesquera o por la modificación de su hábitat (Gallo-Reynoso, 1998).
Además de lo anterior; su estatus de protección, su pequeño tamaño
poblacional que origina la formación de pequeños grupos (2 vaquitas por cada 1000
km vs 47 individuos por cada 1000 km en Phocoena phocoena) (Silber, 1990; Silber
y Norris, 1991; Barlow et al., 1993), su pequeña talla corporal y su bajo perfil
superficial (visibles únicamente por 3 segundos, en intervalos superficiales que van
de 1 a 1.5 minutos) (Silber et al., 1988), dificultan estudios sobre su ecología.
Principalmente aquellos basados en técnicas convencionales como el análisis de
contenido estomacal u observaciones in situ. (Silber y Norris, 1991; Gerrodette et al.,
1995; Barlow et al., 1997; Jaramillo-Legorreta et al., 1999; D’ Agrosa et al., 2000).

3
1.1. Caracterización de hábitat y ecología trófica en depredadores tope
La caracterización del hábitat de los depredadores, con base en el análisis de
diversas variables físicas, químicas y biológicas son elementos clave para sustentar
medidas de conservación (López-Martín et al. 1998). Al respecto, estudios previos
han revelado que la abundancia de los depredadores marinos se asocia con factores
oceanográficos como: batimetría, distancia a la costa, velocidad de la corriente,
profundidad de la termoclina, temperatura, turbidez, sólidos disuelto y suspendidos,
concentración de clorofila y salinidad. Estos factores propician características físico-
químicas específicas que generan condiciones óptimas para la agregación de
especies que satisfacen la demanda alimenticia de los depredadores tope (Tershy,
1992; Jaquet y Whitehead, 1996; Croll et al. 1998; Davis et al., 1998; Springer et al.,
1999; Ortega-Ortiz, 2002; Yen et al., 2004; Tynan et al., 2005; Hastie et al., 2006).
En cuanto a los estudios tróficos convencionales, la dificultad principal se debe
a que la información obtenida se restringe al periodo cercano al consumo de presas,
donde la digestión del consumidor puede imposibilitar su identificación, sesgando los
resultados hacia una dieta poco representativa, como han mostrado diversos autores
(Ridoux, 1994; Lesage et al., 2001; García-Rodríguez y Aurioles, 2004).
Buscando complementar la información arrojada por técnicas convencionales,
en las últimas décadas se ha utilizado cada vez con mayor frecuencia el análisis de
isótopos estables de δ13C y δ15N, el cual se basa en la premisa de que existe un
enriquecimiento predecible en las proporciones isotópicas de δ 13C y δ15N (0.5-1‰ y
3-5‰ respectivamente) de presa a depredador (Schoeninger y DeNiro, 1984; Fry,
1988). El pequeño fraccionamiento observado en el δ13C permite determinar la fuente
de carbono del cual parte la producción primaria (planctónicos vs bentónicos y
pelágicos vs oceánicos), y que se refleja en las señales isotópicas de los tejidos de
los consumidores (Hobson y Welch, 1992; Rau et al. 1992; France, 1995; Achituv et
al., 1997; Walker et al., 1999; Walker y Macko, 1999).

4
Con respecto al δ15N el enriquecimiento refleja la posición a lo largo de la red
trófica desde el primer nivel hasta los depredadores tope (Minegawa y Wada, 1984,
Hobson y Welch, 1992; Abend y Smith, 1997; Griffin y Valiela, 2001). Esta
aproximación metodológica, ha arrojado información relevante sobre la estructura y
función de distintos ecosistemas sin recurrir a números grandes de muestras.
Además, para el caso de especies para las cuales se dificulta obtener muestras, ya
sea por su escasez o dificultad logística, es posible usar organismos existentes en
colecciones científicas y museos como hueso, plumas, dientes, etc (Hobson y Welch,
1992; Hobson et al., 1997; Walker y Macko, 1999; Kelly, 2000; Lesage et al., 2001).
Es importante considerar que al ser la composición isotópica de los tejidos el
resultado de la integración de un número n de dietas en un tiempo X en el pasado, al
usar tejido con diferente tasa metabólica, se puede obtener información de la dieta
promedio de un animal en diferentes intervalos de tiempo (Tieszen et al., 1983). De
lo anterior se espera, que a medida que la tasa metabólica de los tejidos es más
rápida, la ventana temporal que se obtiene de los mismos es más corta, lo cual
representa implicaciones en el análisis de dietas. Esto resulta en que tejidos con tasa
metabólica alta como hígado, páncreas, grasa reflejen periodos más cortos de la
dieta en comparación con tejidos metabólicamente menos activos como hueso o
dientes (Tieszen et al. 1983; Hobson et al. 1996; Koch, 2007; Newsome et al. 2007;
Wolf et al. 2009; Riofrío-Lazo y Aurioles 2013).
En lo que corresponde a P. sinus, la implementación de planes de manejo
exitosos requiere ampliar la información sobre su ecología, los aspectos fundamental
para su conservación son: a) la protección de su hábitat, lo que requiere definir las
dimensiones de este espacio vital y b) la integridad de las funciones ecológicas en
ese hábitat, lo que demanda un mejor conocimiento del papel de la vaquita y sus
relaciones tróficas. En función de lo anterior, en el presente estudio se plantearon las
siguientes metas: 1) La caracterización ambiental del AGC para delimitar el área que
conforma el hábitat real de la especie, más allá del área de protección vigente y 2).
La modelación de la estructura trófica mediante el análisis de isótopos estables de

5
δ13C y δ15N en los diferentes componentes del área oeste del AGC que corresponde
al mayor número de avistamientos en las últimas décadas. Este último punto con el
fin de obtener información sobre los hábitos alimentarios de la vaquita y su papel en
el ecosistema lo cual no ha sido posible con métodos tradicionales.
Figura 1: Reserva de la Biósfera del Alto Golfo de California y Delta del Río Colorado;
su límite sur lo representa la línea punteada roja. El polígono azul muestra el área de
refugio para la vaquita marina y las líneas negras representan la batimetría del área
en metros.

6
2. ANTECEDENTES 2.1. Phocoena sinus 2.1.1 Biología
A pesar del conocimiento tan limitado de esta especie, muchos aspectos de
su biología son claramente únicos (Villa-Ramírez et al., 1995). Una de las
características más sobresalientes de la vaquita marina es que a diferencia de otros
Phocoenidos, los cuales presentan una distribución confinada a aguas frías (Gaskin
et al., 1984), a esta especie la podemos encontrar en temperaturas promedio que
exceden los 30°C en Verano (Álvarez-Borrego, 1983). Si la temperatura corporal de
la vaquita marina es como la de otros mamíferos (cercana a 37°C), existiría muy
poca diferencia entre su temperatura interna y la de su ambiente durante el periodo
más caluroso, situación que podría generar problemas serios en la termorregulación
especialmente en episodios de gran actividad física (depredación, reproducción)
(Silber y Norris, 1991).
La vaquita presenta una longevidad de más de 20 años aproximadamente,
alcanzando la madurez sexual entre los 3 y 6 años de edad; aunque hasta la fecha
se desconoce la edad exacta ya que no se han podido obtener organismos de esta
edad (Hohn et al., 1996). El periodo reproductivo es estacional aunque bianual (la
mayoría de los nacimientos se presentan entre febrero y marzo), lo que podría
significar una tasa de crecimiento poblacional menor en relación a otros cetáceos
(Hohn et al., 1996).
2.1.3 Ecología
Se ha corroborado la presencia de P. sinus durante todo el año en el AGC; lo
cual confirma que es el cetáceo con el área de distribución más reducida a nivel
mundial (Rojas-Bracho et al., 2006). Generalmente, la vaquita marina se observa en

7
aguas entre 10-60 m y alrededor de 11 a 25 km de la costa (Silber y Norris, 1991).
Frecuentemente los individuos de esta especie se dispersan en solitario o en
pequeños grupos de no más de siete (Wells et al, 1981; Vidal et al. 1987; Silber et
al., 1988; Silber et al. 1994). Generalmente, P. sinus muestra preferencia por aguas
de alta turbidez que son producidas por la existencia de fondos lodosos y corrientes
de marea intensas. Estas mareas promueven la agregación de presas facilitando el
éxito de captura por parte de depredadores incluyendo a la vaquita marina, lo que
podría explicar su presencia en esta área particular del AGC durante todo el año
(Silber, 1990).
2.1.4 Alimentación
La escasa información que se tiene sobre la alimentación de la vaquita marina
proviene de estudios basados en análisis de contenido estomacal, los cuales han
sido muy limitados tanto por el escaso número de especímenes disponibles como por
el típico problema de encontrar estómagos cuyo contenido presenta avanzado grado
de digestión (Brownell, 1983; Fitch y Brownell, 1986; Vidal, 1995; Pérez-Cortes,
1996; Findley y Nava, 1996). La información generada, sugiere que es una especie
oportunista que consume aproximadamente 21 especies registradas; dentro de las
cuales los peces son las presas principales (87.5%), seguidos por calamares (37.5%)
y como componentes incidentales los crustáceos (12.5%) (Pérez-Cortés, 1996;
Findley y Nava, 1996).
2.2. Caracterización del Hábitat
Los cetáceos presentan una alta capacidad de desplazamiento, cuyos
patrones de distribución se relacionan con factores oceanográficos que propician
características específicas que satisfacen la demanda alimenticia de los
consumidores (Tershy, 1992; Jaquet y Whitehead, 1996; Doniol-Valcroze et al., 2007;
Panigada et al., 2008).

8
Por ejemplo autores como Davis et al. (1998), Ballance et al. (2006) y Stockin
et al. (2008) relacionan los patrones de distribución de varias especies de mamíferos
marinos con variables ambientales como batimetría, temperatura, salinidad y sus
gradientes horizontales. De manera más extensa, estudios como el de Springer et al.
(1999) y Tynan et al. (2005) han encontrado un vínculo directo con procesos
oceanográficos tales como surgencias, frentes oceánicos, giros, etc. Las variables
que influyen en la distribución de los mamíferos marinos difieren entre áreas, como
se demuestra en los trabajos de Jaquet y Whitehead (1996) quienes observaron que
las zonas de alimentación de los cachalotes en las Islas Galápagos está altamente
correlacionada con las regiones donde se presentan frentes oceánicos, una
batimetría prominente y una alta productividad secundaria, los cuales son esenciales
en la agregación de calamares y peces. Por otro lado, en el Golfo de California,
Jaquet y Gendron (2002) encontraron una relación entre la distribución y abundancia
relativa del cachalote con la profundidad y temperatura del mar, lo que a su vez se
relacionó cercanamente a la abundancia del calamar gigante, Dosidicus gigas. Los
autores de estos estudios señalan que la segregación de especies en las diferentes
temporadas parece responder en gran medida a las características ambientales que
rigen la distribución y disponibilidad de las presas preferenciales de cada especie de
cetáceo. También los cambios en las comunidades de mamíferos marinos
responden a variables ambientales como lo demostró Salvadeo (2008), quien asoció
10 especies de odontocetos observados dentro del Golfo de California con la
disponibilidad y abundancia de presas, como consecuencia de los patrones
migratorios de sardinas y calamares, asociados a su vez con la variación ambiental
del área.
En el caso particular de la vaquita, su presencia se ha corroborado a partir de
múltiples censos poblacionales, únicamente entre Puertecitos, Baja California y
Puerto Peñasco, Sonora (Figura 1) (Barlow, 1986; Brownell, 1986; Silber y Norris,
1991; Jaramillo-Legorreta 2008; Gerrodette et al., 2011), más específicamente en
las cercanías de las Rocas Consag (Villa-Ramírez et al., 1995; Jaramillo-Legorreta,

9
2008; Gerrodette et al., 2011). Aparentemente, los factores que definen su
distribución son muy poco conocidos, entre ellos se encuentra: una preferencia por
aguas relativamente someras (entre 20-100 m) y cercanas a la costa (entre 10-40
Km de distancia), donde predominan sustratos arcillosos-limosos o arcillosos-
limosos-arenosos (Silber, 1990; Villa-Ramírez et al., 1995; Gallo-Reynoso, 1998).
Pero nada de esto es concluyente ya que no existe hasta ahora ningún estudio que
incluya muestreos intensivos de múltiples variables ambientales en un periodo
especifico, ni el análisis de las mismas con el que se puedan determinar las
condiciones que prevalecen en la zona de mayor concentración de la especie.
3. Área de Estudio El AGC se ubica en el extremo norte del Golfo de California entre los
31°41'00'' N y 114°30'00'' W y los 31°40'42'' N y 114°47'00'' W, con una extensión
aproximada de 5000 km2, se localiza en los límites de una línea imaginaria entre San
Felipe, Baja California y Puerto Peñasco, Sonora (Álvarez-Borrego et al., 1975; Lavín
et al., 1997) (Figura 1). Esta región se encuentra rodeada por las tierras áridas de
Sonora y Baja California, debido a lo cual predomina un clima desértico continental,
con una precipitación media anual de 68 mm en el Golfo de Santa Clara y 100 mm
en San Felipe (Miranda et al., 1990), la cual es excedida por una evaporación media
de 1.1 m año-1 (Lavín et al., 1998). Esta área presenta dos estaciones climáticas bien
marcadas: a). Invierno, durante el cual se presentan escasas diferencias entre el
Golfo de California y la costa del Océano Pacífico y b). Verano, cuando la
temperatura máxima del aire puede exceder los 40oC (Roden, 1964). De igual
manera los vientos presentan una marcada estacionalidad, con magnitudes de entre
8-12 ms-1 provenientes del noroeste en invierno y de 2-5 ms-1 del sudeste en verano
(Lavín et al., 1997).
El AGC es un sistema relativamente somero, con profundidades menores a 30
m y un relieve del fondo marino irregular, que forma depresiones que cruzan la
plataforma somera hasta el borde de Cuenca Wagner (Thompson 1968, Álvarez,

10
2003, Álvarez et al. 2009). La mayor parte del AGC se caracteriza por una planicie de
sedimentos donde la profundidad incrementa gradualmente hacia la cuenca Wagner.
Por el contrario, muy cerca de la costa de Sonora los cambios en profundidad se
hacen abruptos evidenciando la presencia de un canal (Thompson, 1969).
3.1. Mareas
Las mareas vivas tienen un rango máximo de 7 a 8 m y generan corrientes
superficiales máximas de 0.7 a 0.8 m s–1 (Lavín et al. 1998, Carriquiry y Sánchez
1999, Álvarez y Jones 2002). La intensa mezcla por marea crea un ambiente
verticalmente mezclado, excepto durante eventos cortos de estratificación cerca del
fondo y en la superficie debido a corrientes de gravedad y calentamiento solar,
respectivamente (Lavín et al. 1998). La re-suspensión crea un ambiente de alta
turbidez con concentraciones de sedimento de hasta 60 a 80 mg/L–1 a 1 m sobre el
fondo durante las corrientes de marea más intensas. Las concentraciones mayores
se encuentran cerca de la cabeza del AGC y decrecen hacia el sureste, en aguas
más profundas (Alvarez y Jones 2002, 2004).
3.2. Hidrografía y Corrientes
Después del represamiento y desvío del Río Colorado para uso agrícola y
urbano en los años 20`s (Thompson, 1968), el AGC llegó a ser un estuario inverso
(Lavín y Organista, 1988; Miranda-Reyes et al., 1990). Las características que
predominan son; el incremento hacia el limite noroeste de la salinidad y la densidad,
alcanzando hasta 38.8 ups y 24.4 kg/m3 en verano y 36.7 ups y 27.2 kg/m3 en
invierno, respectivamente. Aunque se ha comprobado que en años de alta
precipitación el AGC desarrolla clásicas condiciones estuarinas (Lavín y Sánchez,
1999). En el caso de la temperatura, ésta incrementa en verano hasta un máximo de
32°C y decrece hasta los 15°C en invierno (Álvarez-Borrego et al., 1975; Hernández-
Ayon et al., 1993; Lavín et al., 1998).

11
Debido a las altas temperaturas alcanzadas en el AGC, Lavín y Organista
(1988) sugieren que para evitar una acumulación de calor en el AGC, este debe
presentar una circulación termohalina Norte-Sur a partir de la cual se exporte calor y
sal hacia el sur a partir de una circulación anticiclónica. Esto tiene importantes
implicaciones ecológicas, ya que estas corrientes de gravedad se convertirían en un
mecanismo natural de fertilización para el AGC debido al alto contenido de nutrientes
inorgánicos que fluyen hacia la superficie (Hernández-Ayón et al., 1993).
3.3. Sedimentos
Antes de su represamiento, el Río Colorado suministraba al AGC 20X109
m3/año de agua dulce y 180x106 tons/año de sedimentos; pero al desviar su cauce,
la fuente de sedimentos terrígenos se redujo drásticamente (Álvarez y Jones, 2002).
Los sedimentos del fondo son mezclas de arena, limo y arcilla, encontrando el
tamaño de partícula más gruesa, en las áreas someras frente a la antigua
desembocadura del Río Colorado y en la costa este. Los limos y arcillas, predominan
en el resto del AGC, especialmente en el lado oeste donde los depósitos lodosos
forman extensas planicies inter y submareales (Thompson, 1968, Carriquiry y
Sánchez, 1999).

12
4. JUSTIFICACIÓN La vaquita marina enfrenta graves problemas de conservación ya que el
refugio implementado para su protección no ha sido una medida suficiente para
recuperar a su población (Rojas-Bracho y Jaramillo-Legorreta, 2002). En conjunto
con lo anterior, su pequeño número poblacional, distribución geográfica restringida y
su alto grado de endemismo, dan como resultado que su distribución no sea
conocida con exactitud. De acuerdo a Jaramillo-Legorreta (2008) la implementación
de un plan viable de recuperación de la especie, requiere un conocimiento amplio
de identificación de áreas y temporadas críticas para la vaquita. Por lo anterior, se
hace indispensable el estudio de las condiciones ambientales y relaciones
ecológicas presentes en el AGC, que determinan la presencia de la especie en un
área tan reducida. Esto es un aspecto relevante, si se pretende el desarrollo de una
estrategia eficiente para el manejo y la conservación, no sólo de la única población
existente de vaquita marina, sino de un área como el AGC que alberga a varias
especies económica y ecológicamente importantes.

13
5. HIPÓTESIS De acuerdo a lo reportado para el resto de las marsopas en el presente
estudio se espera que el hábitat de distribución potencial de la vaquita marina, que
reúna las características necesarias para su supervivencia, este delimitado por
características ambientales como tipo de fondo, temperatura, batimetría y turbidez y
que estas características influyan en el tipo de alimento e interacciones tróficas de la
vaquita, reflejándose en su firma isotópica.

14
6. OBJETIVO GENERAL
Definir el hábitat de distribución potencial y área de alimentación de la vaquita marina
(Phocoena sinus), así como describir las interacciones tróficas de la especie a partir
de variables ambientales e isotópicas.
6.1 Objetivos Específicos
CAPITULO 1
1. Caracterizar ambientalmente el AGC con base en las variables
muestreadas durante cruceros realizados en junio de 2010 y marzo de 2011.
2. Determinar que parámetros ambientales se relacionan con la
distribución del mayor número de avistamientos de la vaquita marina y en
función de esto delimitar la extensión de su hábitat potencial.
CAPITULO 2
3. Determinar la existencia de áreas isotópicas a partir de δ13C y
δ15N en el AGC y evaluar las posibles diferencias entre junio de 2010 y marzo
de 2011.
4. Delimitar la extensión isotópica del área de alimentación de la
vaquita marina a partir de los valores de δ13C y δ15N.
CAPITULO 3
5. Establecer el número de niveles en la trama trófica del área de
mayores avistamientos de la vaquita marina a partir de δ13C y δ15N.

15
6. Definir la amplitud del nicho trófico de la vaquita marina con
base en los valores isotópicos de carbono y nitrógeno en colágeno óseo de la
especie.
7. Determinar la contribución proporcional de diferentes presas
potenciales a la dieta de la vaquita marina muestreadas alrededor de Rocas
Consag, mediante un modelo de mezcla Bayesiano.

16
7. METODOLOGIA GENERAL
7.1. Obtención y procesamiento de muestras
La obtención de datos hidrográficos y muestras biológicas se llevó a cabo en
dos fases dentro del proyecto Pesca Artesanal del Norte del Golfo de California:
Ambiente y Sociedad (PANGAS):
1). Para el muestreo de datos ambientales, sedimentos, fitoplancton,
zooplancton, se realizaron dos cruceros oceanográficos a bordo del Buque Francisco
de Ulloa del CICESE en el AGC. Uno durante la temporada cálida (1 al 9 de junio del
2010) y uno en la estación fría (25 de marzo al 02 de abril), los cuales cubrieron la
Reserva de la Biosfera del Alto Golfo de California incluyendo el área Refugio de la
vaquita marina (Figura 2).
7.1.1 Datos ambientales: Para la obtención de las variables oceanográficas se
muestrearon 75 estaciones durante la temporada cálida y 77 durante la temporada
fría. En cada una de las cuales obtuvieron perfiles de temperatura, conductividad,
fluorescencia, turbidez y oxígeno disuelto utilizando un CTD SeaBrid 911plus con
sensores acoplados de los parámetros mencionados, los cuales fueron procesados
por el equipo CIRCUS del CICESE- Campus Ensenada.
7.1.2 Sedimento: Se obtuvieron 59 muestras de sedimento por duplicado, del
primer centímetro superficial durante la campaña de julio, mediante una draga
VanVeen, manteniéndose a -20ºC perfectamente bien etiquetados para su posterior
análisis isotópico, elemental y granulométrico.
17
Figura 2: Área de estudio; los puntos amarillos representan las estaciones de
muestreo para datos oceanográficos y biológicos y los cuadros rosas para CTD
únicamente. El área dentro del polígono indica el refugio de la vaquita establecido
desde 2005 (DOF, 2005) y las líneas roja y rosa delimitan el área de la Reserva de la
Biósfera y Delta del Río Colorado.
Las muestras almacenadas se desmineralizaron para su análisis isotópico,
mediante el enjuague con HCl 0.2N, hasta que la muestra dejó de efervecer lo cual
fue indicativo de que los carbonatos habían sido eliminados. Posteriormente la
muestra se enjuagó con agua destilada y se secó en un horno a 60°C, después de lo
cual fue molida en un mortero de Agatha. Finalmente se pesó en capsulas de estaño,
aproximadamente 25 mg de muestra para su posterior análisis isotópico.
Posteriormente, una submuestra de aproximadamente 50g de cada una de las 59
estaciones, debidamente etiquetada se envió para su análisis granulométrico por el

18
método de Taylor (1948), en el Laboratorio de Edafología en el Centro de
Investigaciones Biológicas del Noroeste (CIBNOR).
7.1.3 Fitoplancton: Las muestras se obtuvieron de una red de 75 estaciones
en verano y 77 en invierno. Los muestreos se realizaron durante el día y la noche
utilizando una red cónica con 30 cm de diámetro de boca y luz de malla de 64 μm.
Los arrastres fueron horizontales en los primeros siete metros de la capa superficial
del mar y de forma circular por un periodo de 10 minutos aproximadamente, a una
velocidad de 1.5 nudos. La muestra fue pasada por un tamiz de 505 μm para
desechar el material zooplanctónico que pudiera haber quedado atrapado y
posteriormente por un tamiz de 64 μm para mantener la muestra con la menor
cantidad de agua posible como lo recomiendan Chanton y Lewis (1999) y
Santamaría del Ángel, com. pers. Finalmente se congeló a -20ºC hasta su procesado
en el laboratorio de Ecología de Pinnípedos en el CICIMAR-IPN
7.1.4 Zooplancton: Las muestras se obtuvieron de la misma red de estaciones
para ambas temporadas que en el caso del fitoplancton. El muestreo se llevó a cabo
mediante una red superficial con un diámetro de boca de 60 cm y un tamaño de
malla de 505μm, a partir de arrastres circulares a una velocidad de 2.5 nudos,
durante el día y la noche.
Para la obtención de la biomasa zooplanctónica se utilizó el método de
desplazamiento de volumen, estandarizando la muestra a mL/1000m3 (Kramer et al.,
1972). Este procedimiento lo realizó personal del Laboratorio de Plancton y Biología
Marina del CICIMAR-IPN, y los resultados obtenidos fueron ingresados a la base de
datos del presente estudio. La muestra se almacenó a -20ºC hasta su procesado y
análisis isotópico de δ13C y δ15N en el Laboratorio de Ecología de Pinnípedos del
CICIMAR.

19
Para el análisis isotópico, tanto las muestras de fitoplancton como de
zooplancton fueron desmineralizadas con ácido clorhídrico 0.2 N en repetidas
ocasiones para eliminar los carbonatos inorgánicos presentes en la misma. La
muestra se consideró completamente desmineralizada cuando el material dejó de
efervecer. Posteriormente, las muestras se enjuagaron con agua destilada para
retirar el exceso de ácido y se secaron a 60ºC, moliéndose finalmente en un mortero
de Ágata para homogenizarlas (Bouillon et al. 2002).
2). Un total de 40 muestras de peces y macro-invertebrados (Caracoles,
bivalvos, esponjas, crustáceos, moluscos) se obtuvieron a partir de: a)
pescadores locales del área de San Felipe, BCS; b) adquisición directa en
mercados de la localidad y c) la colaboración en la colecta de peces por el
personal de investigaciones ictiológicas del CEDO. Todas las muestras obtenidas
se almacenaron a -20ºC hasta su posterior identificación taxonómica en el
laboratorio de ictiología y procesado en el laboratorio de Ecología de Pinnípedos
del CICIMAR-IPN.
3). Colágeno óseo de P. sinus: Se muestreó un trozo de cráneo de 34
vaquitas marinas, los cuales fueron proporcionadas por la Colección Osteológica
de Mamíferos Marinos del CIAD-ITESM Campus Guaymas. Adicionalmente se
usaron 4 datos isotópicos de colágeno óseo de vértebras de vaquitas obtenidos
de la tesis de Rodríguez-Pérez (2007). Cabe mencionar, que debido a que no fue
posible extraer los lípidos por cuestiones logísticas, únicamente se utilizaron 15
datos en el presente estudio, las cuales presentaron una proporción (C/N) menor
a 3.5, la cual está dentro del rango de proteína pura (Ambrose, 1990).

20
7.1.5. Procesamiento para análisis isotópico de macrofauna y tejido óseo de P.
sinus
Las muestras de musculo y hueso de los diferentes grupos se lavaron con
agua destilada con el fin de retirar cualquier impureza que pudiera estar presente en
el tejido. Cuando la muestra provenía de estructuras que presentaban carbonatos, se
enjuagó por triplicado con HCl al 10%, durante un periodo de 24 a 48 h, después de
las cuales la muestra se enjuagó con agua destilada y se secó con el fin de evaporar
el ácido.
Posteriormente la muestra se trituró en un mortero de Ágata y una fracción del
polvo obtenido (entre 0.8 y 1.2 mg) se colocó en cápsulas de estaño para su
posterior análisis isotópico de δ13C y δ15N. El análisis isotópico se llevó a cabo en un
espectrómetro de masas Carlo-Erba Finnigan Delta Plus XL en interfase con un
analizador elemental NC 2500 el cual tiene una precisión de 0.2‰, en el Laboratorio
de Isótopos Estables en la Universidad de Davis California.
Las lecturas de las concentraciones isotópicas, para cada una de las
especies se reportaron con relación al estándar que para carbono es
PeeDeeBelemnita y para nitrógeno se utilizará el N2 (Aire atmosférico). La
composición de isótopos se describió con la definición δ delta convencional como
partes por mil (‰) como sigue (DeNiro y Epstein, 1978):
δ= (13C/12C ó 15N/14N)= [(Rm/ Rs) - 1] X 1000 (ec. 1)
Donde:
Rm= Proporción del número de átomos del isótopo pesado sobre el ligero de
la muestra (13C/12C ó 15N/14N).
Rs = Proporción del número de átomos del isótopo pesado sobre el ligero del
estándar (13C/12C ó 15N/14N).

21
8. CAPÍTULO 1
CARACTERIZACIÓN AMBIENTAL DEL ÁREA DE MAYORES AVISTAMIENTOS DE LA VAQUITA MARINA
8.1. INTRODUCCIÓN
Los mamíferos marinos se desplazan en áreas con características
oceanográficas que influyen en la agregación de presas, lo que les permite ser
exitosos en su ambiente (Gaskin, 1983; Luseau et al., 2004). Algunas de estas
características son la temperatura, concentración de clorofila, batimetría, la presencia
de frentes, etc, (Baumgartner et al., 2001; Etnoyer, 2004; Luseau et al., 2004; Jaquet
y Gendron, 2002).
La vaquita marina, como consumidor de alto nivel trófico no es la excepción y
aunque hasta el momento son pocos los estudios que se han realizado sobre el uso
y extensión real de su hábitat, se ha observado que la especie presenta dos
características que la distinguen del resto de las marsopas: a) la preferencia por
aguas que pueden exceder los 30°C de temperatura en verano y b) un área de
distribución más reducida. En años recientes se ha determinado que la especie se
limita a la región Oeste del AGC, más específicamente a los alrededores de Rocas
Consag (Silber, 1990; Gerrodette et al., 1995; Jaramillo-Legorreta et al., 1999;
Gerrodette et al., 2011).
Hasta el momento se conoce que el AGC presenta gradientes ambientales en
factores como tamaño de grano, temperatura, salinidad y profundidad (Thompson,
1969; Lavín et al. 1998; Carriquiry y Sánchez, 1999). Además, esta variabilidad en
las características físicas, se ve reflejada en la parte biológica como lo demuestran
Millan-Nuñez et al. (1999) quienes reportan que el área adyacente a la península de
Baja California presenta una mayor productividad que la registrada para el área este,
cerca de la costa de Sonora. Valdez-Muñoz et al. (2010), encontraron que la

22
distribución espacial de juveniles de totoaba presentó la mayor abundancia al
suroeste de la Isla Montague y parte oeste del AGC, infiriendo que la alta turbidez de
esa región provee protección y alimento a los individuos. Por el contrario, Sánchez-
Velasco et al. (2012) indican que de los tres hábitats de larvas de peces encontrados
en el alto Golfo, el menos diverso es el que corresponde a la zona oeste, sugiriendo
que los cambios antropogénicos sobre la región al cortar el flujo de agua dulce y
volver al AGC un sistema hipersalino pudo haber reducido el hábitat de algunas
especies.
Actualmente, uno de los puntos urgentes para el establecimiento de un plan
de manejo exitoso para la conservación de la vaquita marina es la identificación del
área en la cual habita alrededor del año (Jaramillo-Legorreta, 2008). Por lo anterior,
uno de los resultados más importantes de este estudio será caracterizar el hábitat
de la vaquita y determinar: a) si dicho hábitat está restringido al área donde
actualmente se observan la mayoría de los registros de animales o b) si este es
más amplio y explicar porque hay varios registros a mayor distancia de este núcleo
actual.

23
8.2. METODOLOGÍA
8.2.1Tratamientos de datos ambientales
Para realizar una exploración preliminar de los datos, se hicieron mapas con el
programa Surfer v9.0 a partir de las variables ambientales de biomasa
zooplanctonica, concentración de clorofila, oxígeno, turbidez, salinidad, temperatura,
profundidad y granulometría. Para delimitar las regiones sedimentológicas, se utilizó
el programa TERNPLOT, que permitió agrupar los resultados granulométricos de
acuerdo con la similitud en el tamaño de grano basado en el diagrama ternario de
Marshall (1996; www.geologynet.com/programs/ternplot.xls ).
8.2.2 Análisis estadísticos entre variables ambientales:
Para evaluar si existía una regionalización en función de las variables
ambientales de temperatura (°C), salinidad (ups), clorofila (μg/L), oxígeno disuelto
(mL/L), granulometría (%) y turbidez (mg/L), se realizó un análisis de perfiles de
similitud (SIMPROF) con el programa PRIMER v6.0 (Clarke et al. 2008). Esta prueba
se basa en un análisis de grupos (cluster) y asocia las diferentes variables de cada
estación de acuerdo al grado de similitud de los datos entre las mismas.
Posteriormente para determinar si entre los grupos definidos por el análisis existían
diferencias significativas, se aplicó un análisis de similitud de una vía (ANOSIM), que
no requiere suposición de normalidad de los datos usando el software PRIMER v6.0.
Esta prueba estadística describe la separación entre grupos, asignando un valor
numérico R, donde el valor más positivo indica mayor separación (Clarke y Gorley,
2006).
Como el valor R obtenido del ANOSIM nos señala si existen diferencias
significativas entre los grupos (regiones), pero no cuales variables en específico son
las que nos marcan esas diferencias, se realizó un análisis multivariado Kruskall-

24
Wallis, con el fin de definir que variables presentaban diferencias significativas entre
las regiones .
Para definir diferencias significativas entre regiones se procedió a realizar una
prueba U Mann-Withney. Para determinar si existían diferencias significativas entre
junio (2010) y marzo (2011) de las variables ambientales se utilizó una prueba U
Mann-Whitney mediante el programa STATISTICA v8.0.
A partir de las regiones ambientales obtenidas del análisis SIMPROF, se
realizaron series de tiempo de promedios mensuales de datos satelitales de
temperatura superficial del mar (SST) y clorofila (SSChl “a”), como un aproximado de
la productividad primaria (http://oceancolor.sfc.nasa.gov). Estas imágenes tuvieron
una resolución de 4X4 km. Lo anterior con el fin de respaldar la permanencia de la
vaquita en su hábitat potencial como consecuencia de la estabilidad del sistema a
través del tiempo.
8.2.3 Caracterización del hábitat potencial de P. sinus
Con base en los censos realizados durante los últimos 15 años (1992-2007)
recopilados de la bibliografía (Villa-Ramírez et al., 1995; Gerrodette et al., 1995;
Jaramillo-Legorreta, 2008), que incluyen registros tanto visuales como acústicos, se
caracterizó ambientalmente la región que en la última década ha concentrado la
mayor cantidad de avistamientos de la especie. Posteriormente, se determinó la
extensión de las variables ambientales de la región arriba mencionada y sus
diferencias con respecto a otras regiones ambientales definidas por el análisis
SIMPROF. Estas diferencias, se evaluaron mediante la comparación de los valores
de las variables ambientales encontrados entre las regiones encontradas.

25
8.3. RESULTADOS 8.3.1 Diferencias espaciales:
En función de las variables de profundidad, temperatura, salinidad, oxígeno,
concentración de clorofila “a”, granulometría y turbidez, se realizó un análisis cluster
mediante el cual se definieron 3 regiones (Figura 3 A y B):
1). Este: La cual corresponde a la región costera continental, aledaña al
estado de Sonora.
2). Oeste: Región costera peninsular, aledaña a la Península de Baja
California.
3). Sur: Región oceánica, que corresponde a la Cuenca Wagner.
Con respecto a esta última sólo se pudo obtener información limitada en
cuanto a las características granulométricas debido a que únicamente se pudieron
muestrear 4 estaciones, a diferencia de la región Este con 19 estaciones
muestreadas y la Oeste con 30. La presencia de estas tres regiones se mantuvo en
las dos temporadas de muestreo (junio 2010 y marzo 2011), aunque se observa una
mayor extensión de la región Oeste durante marzo.
El análisis ANOSIM mostró diferencias significativas (0.436 y 0.471, para junio
y marzo, respectivamente) entre las tres regiones encontradas. Para definir cuáles
eran las variables que determinaban las diferencias obtenidas del análisis, se realizó
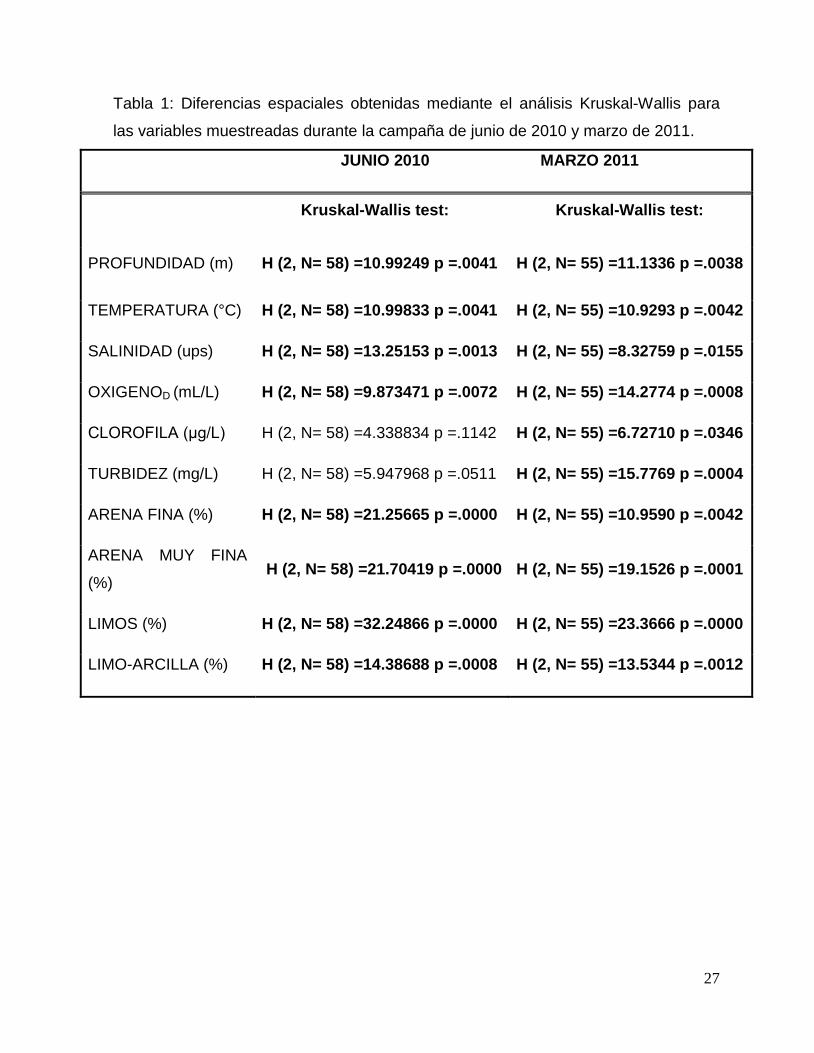
un análisis Kruskal-Wallis que mostró diferencias significativas entre todas las
variables analizadas excepto para la clorofila “a” y la turbidez durante marzo (Tabla
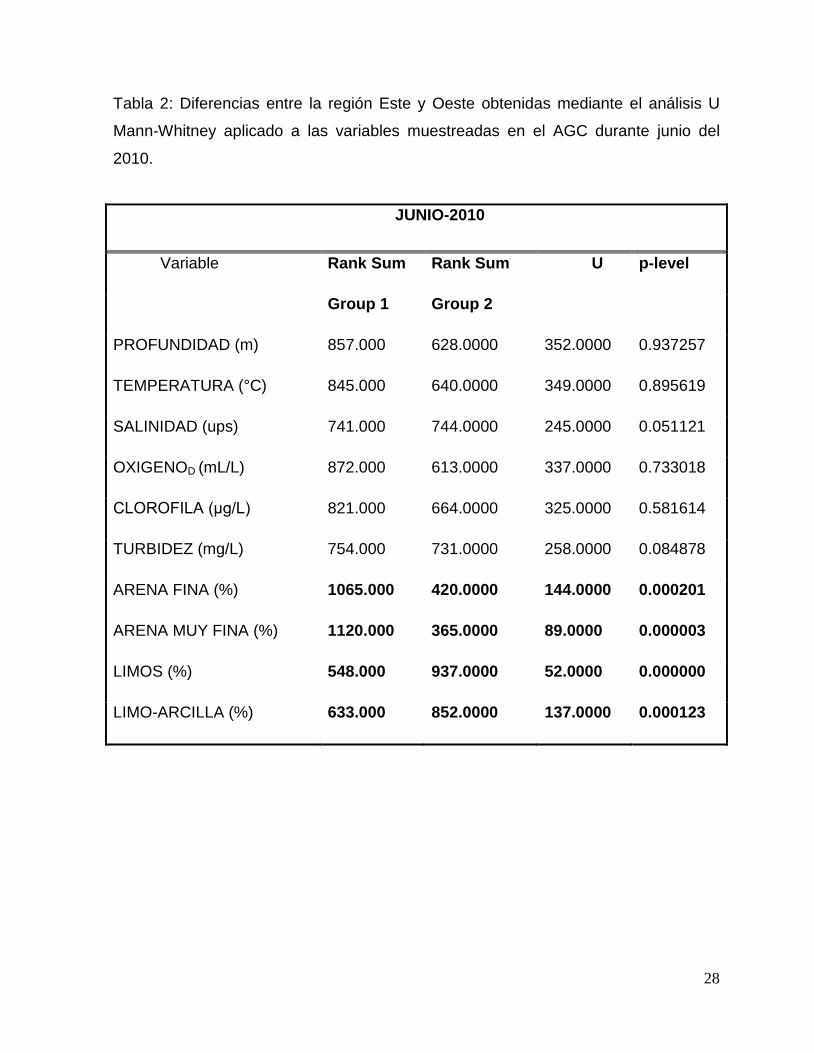
1). Para confirmar lo anterior y determinar si entre la región Este y Oeste existían
diferencias significativas se realizó una prueba pareada U Mann-Whitney,
encontrando que las variables significativas fueron las características del sedimento
26
(granulometría y turbidez; Tabla 2). Adicionalmente durante marzo también el
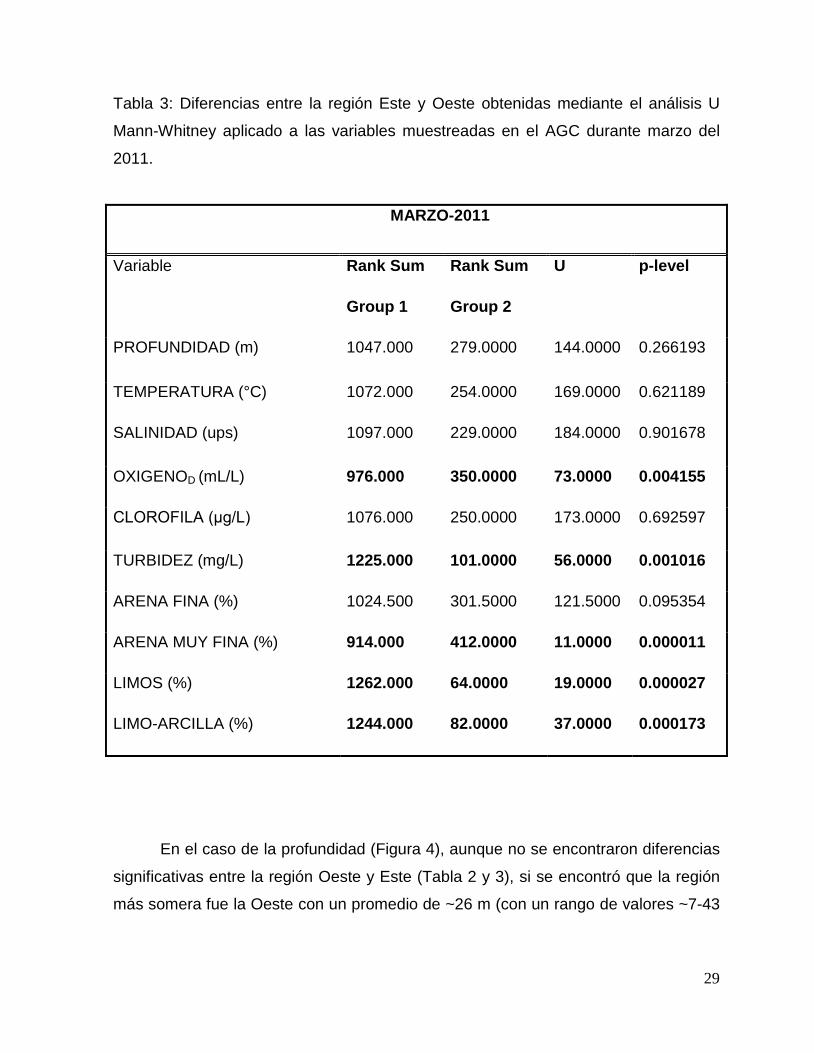
oxígeno presentó una diferencia espacial significativa (Tabla 3).
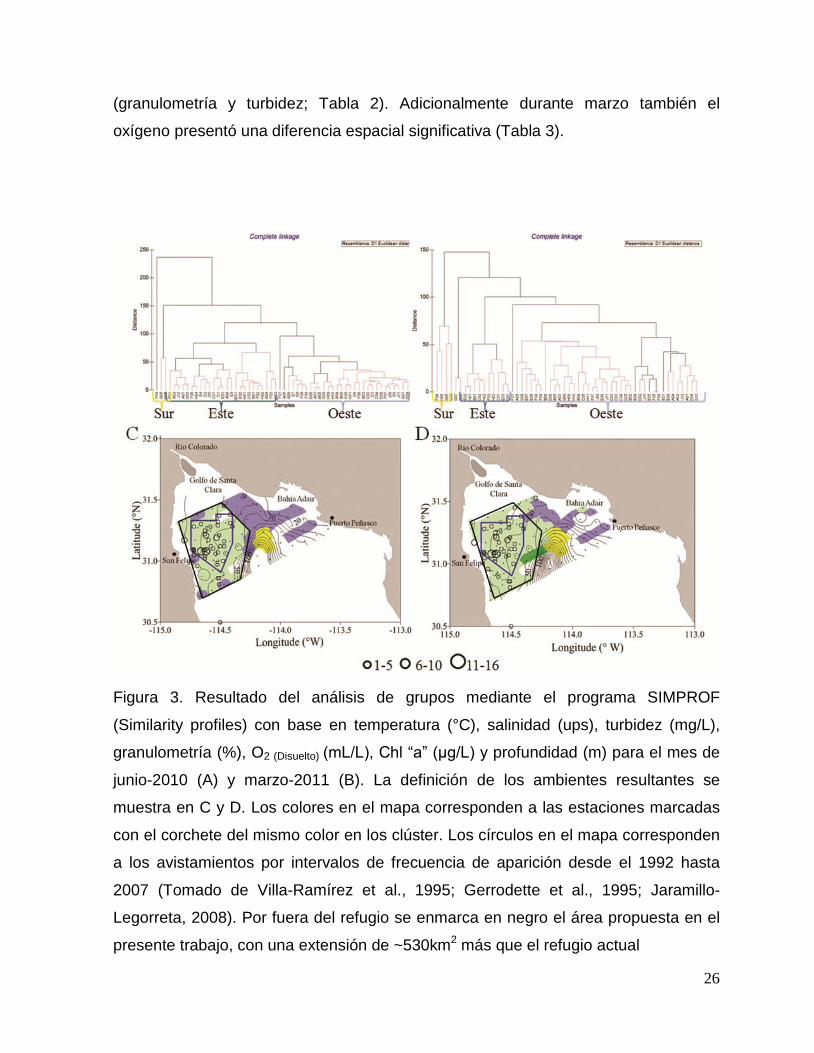
Figura 3. Resultado del análisis de grupos mediante el programa SIMPROF
(Similarity profiles) con base en temperatura (°C), salinidad (ups), turbidez (mg/L),
granulometría (%), O2 (Disuelto) (mL/L), Chl “a” (μg/L) y profundidad (m) para el mes de
junio-2010 (A) y marzo-2011 (B). La definición de los ambientes resultantes se
muestra en C y D. Los colores en el mapa corresponden a las estaciones marcadas
con el corchete del mismo color en los clúster. Los círculos en el mapa corresponden
a los avistamientos por intervalos de frecuencia de aparición desde el 1992 hasta
2007 (Tomado de Villa-Ramírez et al., 1995; Gerrodette et al., 1995; Jaramillo-
Legorreta, 2008). Por fuera del refugio se enmarca en negro el área propuesta en el
presente trabajo, con una extensión de ~530km2 más que el refugio actual
27
Tabla 1: Diferencias espaciales obtenidas mediante el análisis Kruskal-Wallis para
las variables muestreadas durante la campaña de junio de 2010 y marzo de 2011.
JUNIO 2010 MARZO 2011
Kruskal-Wallis test: Kruskal-Wallis test:
PROFUNDIDAD (m) H (2, N= 58) =10.99249 p =.0041 H (2, N= 55) =11.1336 p =.0038
TEMPERATURA (°C) H (2, N= 58) =10.99833 p =.0041 H (2, N= 55) =10.9293 p =.0042
SALINIDAD (ups) H (2, N= 58) =13.25153 p =.0013 H (2, N= 55) =8.32759 p =.0155
OXIGENOD (mL/L) H (2, N= 58) =9.873471 p =.0072 H (2, N= 55) =14.2774 p =.0008
CLOROFILA (μg/L) H (2, N= 58) =4.338834 p =.1142 H (2, N= 55) =6.72710 p =.0346
TURBIDEZ (mg/L) H (2, N= 58) =5.947968 p =.0511 H (2, N= 55) =15.7769 p =.0004
ARENA FINA (%) H (2, N= 58) =21.25665 p =.0000 H (2, N= 55) =10.9590 p =.0042
ARENA MUY FINA
(%) H (2, N= 58) =21.70419 p =.0000 H (2, N= 55) =19.1526 p =.0001
LIMOS (%) H (2, N= 58) =32.24866 p =.0000 H (2, N= 55) =23.3666 p =.0000
LIMO-ARCILLA (%) H (2, N= 58) =14.38688 p =.0008 H (2, N= 55) =13.5344 p =.0012
28
Tabla 2: Diferencias entre la región Este y Oeste obtenidas mediante el análisis U
Mann-Whitney aplicado a las variables muestreadas en el AGC durante junio del
2010.
JUNIO-2010
Variable Rank Sum Rank Sum U p-level
Group 1 Group 2
PROFUNDIDAD (m) 857.000 628.0000 352.0000 0.937257
TEMPERATURA (°C) 845.000 640.0000 349.0000 0.895619
SALINIDAD (ups) 741.000 744.0000 245.0000 0.051121
OXIGENOD (mL/L) 872.000 613.0000 337.0000 0.733018
CLOROFILA (μg/L) 821.000 664.0000 325.0000 0.581614
TURBIDEZ (mg/L) 754.000 731.0000 258.0000 0.084878
ARENA FINA (%) 1065.000 420.0000 144.0000 0.000201
ARENA MUY FINA (%) 1120.000 365.0000 89.0000 0.000003
LIMOS (%) 548.000 937.0000 52.0000 0.000000
LIMO-ARCILLA (%) 633.000 852.0000 137.0000 0.000123
29
Tabla 3: Diferencias entre la región Este y Oeste obtenidas mediante el análisis U
Mann-Whitney aplicado a las variables muestreadas en el AGC durante marzo del
2011.
MARZO-2011
Variable Rank Sum Rank Sum U p-level
Group 1 Group 2
PROFUNDIDAD (m) 1047.000 279.0000 144.0000 0.266193
TEMPERATURA (°C) 1072.000 254.0000 169.0000 0.621189
SALINIDAD (ups) 1097.000 229.0000 184.0000 0.901678
OXIGENOD (mL/L) 976.000 350.0000 73.0000 0.004155
CLOROFILA (μg/L) 1076.000 250.0000 173.0000 0.692597
TURBIDEZ (mg/L) 1225.000 101.0000 56.0000 0.001016
ARENA FINA (%) 1024.500 301.5000 121.5000 0.095354
ARENA MUY FINA (%) 914.000 412.0000 11.0000 0.000011
LIMOS (%) 1262.000 64.0000 19.0000 0.000027
LIMO-ARCILLA (%) 1244.000 82.0000 37.0000 0.000173
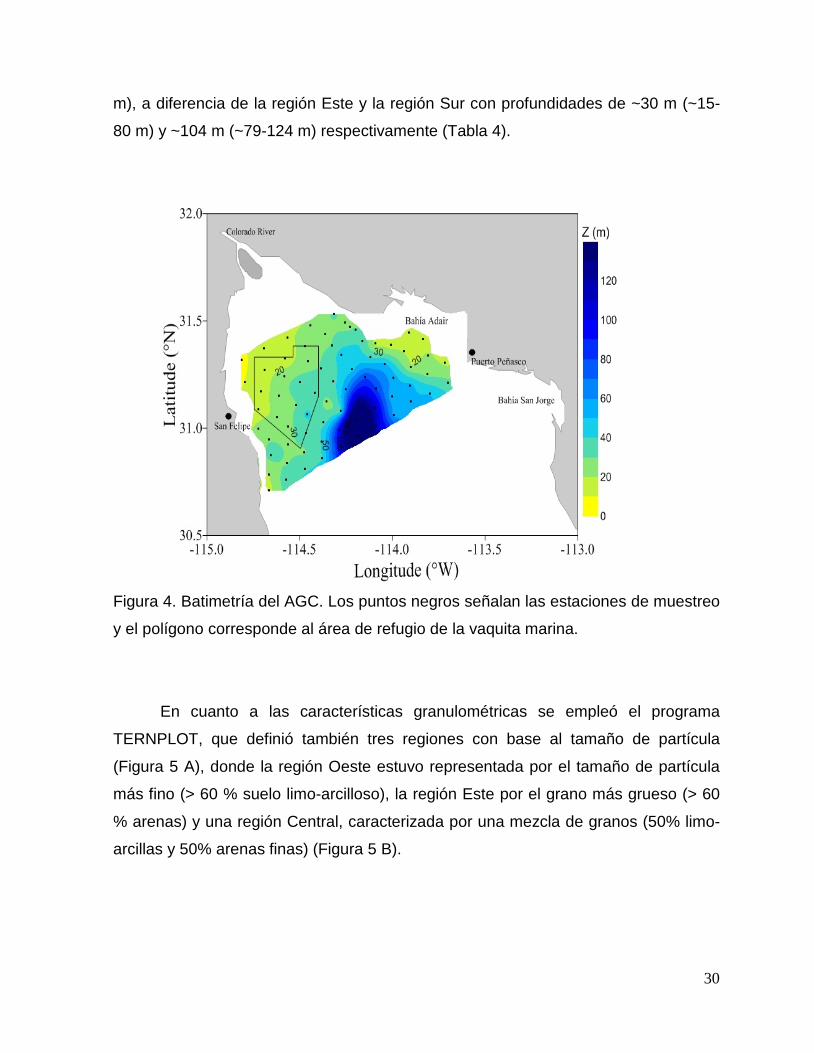
En el caso de la profundidad (Figura 4), aunque no se encontraron diferencias
significativas entre la región Oeste y Este (Tabla 2 y 3), si se encontró que la región
más somera fue la Oeste con un promedio de ~26 m (con un rango de valores ~7-43
30
m), a diferencia de la región Este y la región Sur con profundidades de ~30 m (~15-
80 m) y ~104 m (~79-124 m) respectivamente (Tabla 4).
Figura 4. Batimetría del AGC. Los puntos negros señalan las estaciones de muestreo
y el polígono corresponde al área de refugio de la vaquita marina.
En cuanto a las características granulométricas se empleó el programa
TERNPLOT, que definió también tres regiones con base al tamaño de partícula
(Figura 5 A), donde la región Oeste estuvo representada por el tamaño de partícula
más fino (> 60 % suelo limo-arcilloso), la región Este por el grano más grueso (> 60
% arenas) y una región Central, caracterizada por una mezcla de granos (50% limo-
arcillas y 50% arenas finas) (Figura 5 B).
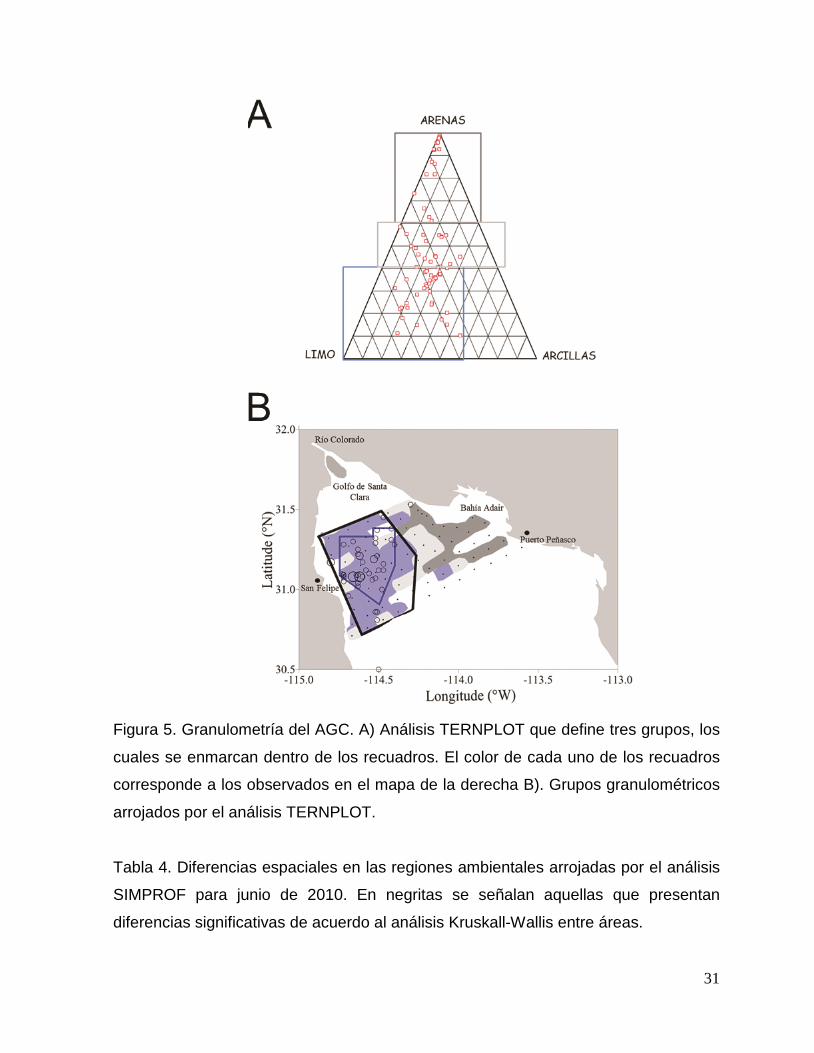
31
Figura 5. Granulometría del AGC. A) Análisis TERNPLOT que define tres grupos, los
cuales se enmarcan dentro de los recuadros. El color de cada uno de los recuadros
corresponde a los observados en el mapa de la derecha B). Grupos granulométricos
arrojados por el análisis TERNPLOT.
Tabla 4. Diferencias espaciales en las regiones ambientales arrojadas por el análisis
SIMPROF para junio de 2010. En negritas se señalan aquellas que presentan
diferencias significativas de acuerdo al análisis Kruskall-Wallis entre áreas.

32
ESTE OESTE SUR
VARIABLES PROMEDIO (MIN-MAX)
PROMEDIO (MIN-MAX)
PROMEDIO (MIN-MAX)
PROFUNDIDAD (m) 33.9 (15-79.7) 26.4 (7.3-43.1) 105.3 (78.9-124.4)
TEMPERATURA (°C) 23.2 (21-25.6) 23.7 (22-25.2) 20.6 (20.4-21.01)
SALINIDAD (ups) 36.0 (35.6-36.8) 36.4 (35.8-37.5) 35.6 (35.6-35.7)
OXIGENOD (mL/L) 4.4 (3.5-4.9) 4.5 (4.2-4.8) 3.8 (3.7-3.9)
CLOROFILA (μg/L) 0.18 (0.03-0.65) 0.28 (0.03-0.71) 0.07 (0.04-0.10)
TURBIDEZ (mg/L) 12.4 (9.9-24.6) 15.2 (10.1-39.4) 10.4 (10.4-10.4) ARENA FINA (%) 16.2 (3.8-39.3) 9.6 (0.6-61.9) 31.9 (21.3-42.8) ARENA MUY FINA (%) 53.7 (24.8-83.8) 22.8 (8.3-39.03) 32.0 (20.6-52.8) LIMOS (%) 19.1 (0.6-41.4) 40.2 (0.9-65.9) 15.8 (1.8-23.6) LIMO-ARCILLA (%) 8.3 (0-25.6) 26.3 (0.3-55.3) 19.3 (0.4-34.0)
Para el resto de las variables (temperatura, salinidad, oxígeno disuelto,
clorofila y turbidez) se encontraron los menores valores en la región Sur para ambas
temporadas de muestreo (Tabla 4 y 5). En el caso de la región Oeste y Este los
valores de las diferentes variables fueron muy similares para ambas temporadas de
muestreo, excepto para la turbidez, la cual presenta diferencias significativas
encontrando la mayor concentración en la región Oeste (Tabla 4 y 5)
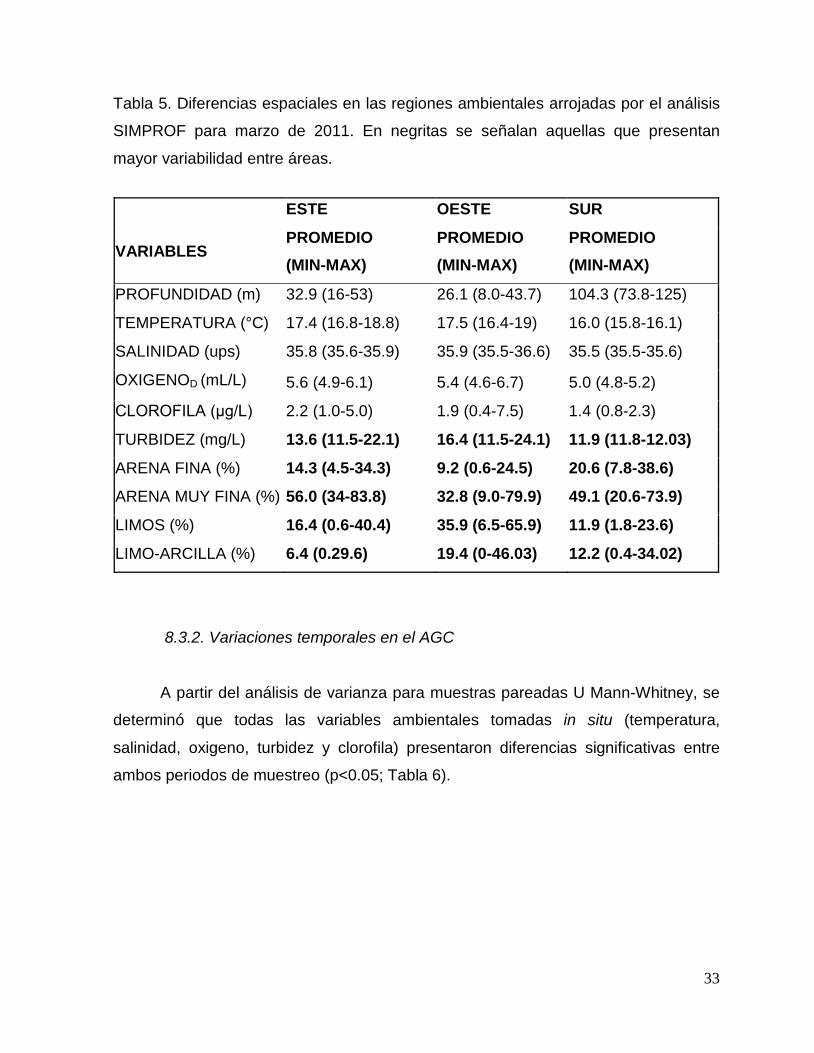
33
Tabla 5. Diferencias espaciales en las regiones ambientales arrojadas por el análisis
SIMPROF para marzo de 2011. En negritas se señalan aquellas que presentan
mayor variabilidad entre áreas.
8.3.2. Variaciones temporales en el AGC
A partir del análisis de varianza para muestras pareadas U Mann-Whitney, se
determinó que todas las variables ambientales tomadas in situ (temperatura,
salinidad, oxigeno, turbidez y clorofila) presentaron diferencias significativas entre
ambos periodos de muestreo (p<0.05; Tabla 6).
ESTE OESTE SUR
VARIABLES PROMEDIO (MIN-MAX)
PROMEDIO (MIN-MAX)
PROMEDIO (MIN-MAX)
PROFUNDIDAD (m) 32.9 (16-53) 26.1 (8.0-43.7) 104.3 (73.8-125)
TEMPERATURA (°C) 17.4 (16.8-18.8) 17.5 (16.4-19) 16.0 (15.8-16.1)
SALINIDAD (ups) 35.8 (35.6-35.9) 35.9 (35.5-36.6) 35.5 (35.5-35.6)
OXIGENOD (mL/L) 5.6 (4.9-6.1) 5.4 (4.6-6.7) 5.0 (4.8-5.2)
CLOROFILA (μg/L) 2.2 (1.0-5.0) 1.9 (0.4-7.5) 1.4 (0.8-2.3)
TURBIDEZ (mg/L) 13.6 (11.5-22.1) 16.4 (11.5-24.1) 11.9 (11.8-12.03)
ARENA FINA (%) 14.3 (4.5-34.3) 9.2 (0.6-24.5) 20.6 (7.8-38.6)
ARENA MUY FINA (%) 56.0 (34-83.8) 32.8 (9.0-79.9) 49.1 (20.6-73.9)
LIMOS (%) 16.4 (0.6-40.4) 35.9 (6.5-65.9) 11.9 (1.8-23.6)
LIMO-ARCILLA (%) 6.4 (0.29.6) 19.4 (0-46.03) 12.2 (0.4-34.02)
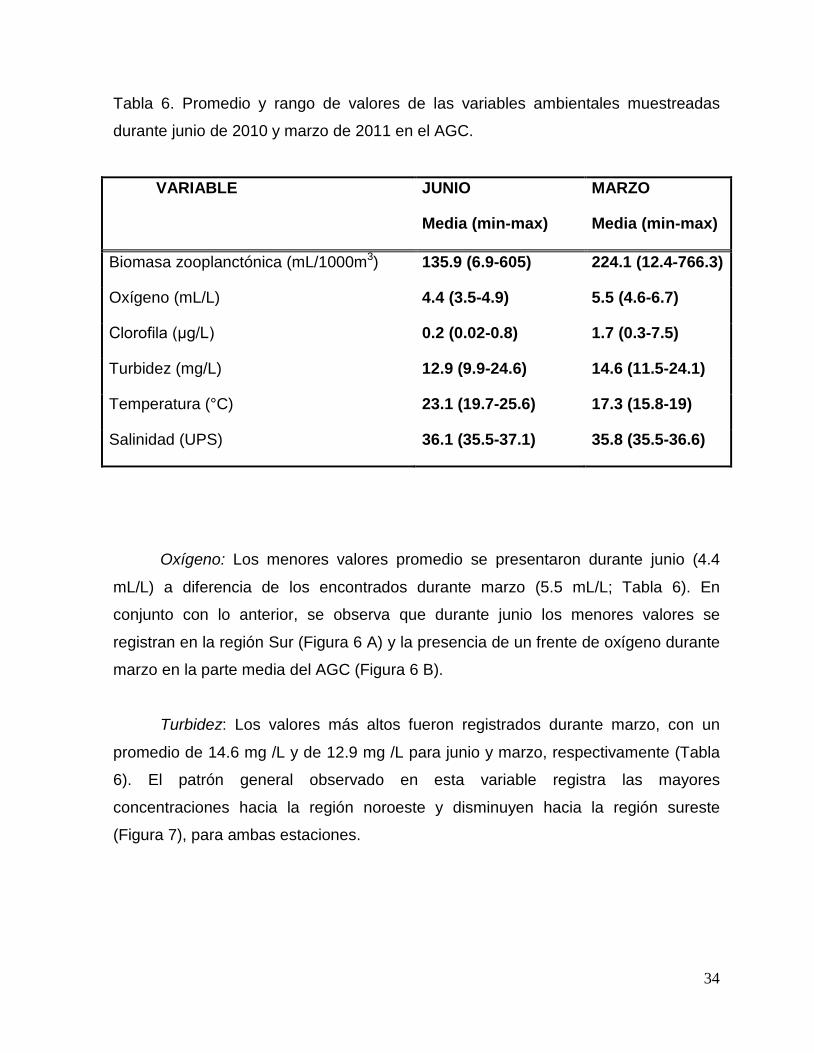
34
Tabla 6. Promedio y rango de valores de las variables ambientales muestreadas
durante junio de 2010 y marzo de 2011 en el AGC.
VARIABLE JUNIO
Media (min-max)
MARZO
Media (min-max)
Biomasa zooplanctónica (mL/1000m3) 135.9 (6.9-605) 224.1 (12.4-766.3)
Oxígeno (mL/L) 4.4 (3.5-4.9) 5.5 (4.6-6.7)
Clorofila (μg/L) 0.2 (0.02-0.8) 1.7 (0.3-7.5)
Turbidez (mg/L) 12.9 (9.9-24.6) 14.6 (11.5-24.1)
Temperatura (°C) 23.1 (19.7-25.6) 17.3 (15.8-19)
Salinidad (UPS) 36.1 (35.5-37.1) 35.8 (35.5-36.6)
Oxígeno: Los menores valores promedio se presentaron durante junio (4.4
mL/L) a diferencia de los encontrados durante marzo (5.5 mL/L; Tabla 6). En
conjunto con lo anterior, se observa que durante junio los menores valores se
registran en la región Sur (Figura 6 A) y la presencia de un frente de oxígeno durante
marzo en la parte media del AGC (Figura 6 B).
Turbidez: Los valores más altos fueron registrados durante marzo, con un
promedio de 14.6 mg /L y de 12.9 mg /L para junio y marzo, respectivamente (Tabla
6). El patrón general observado en esta variable registra las mayores
concentraciones hacia la región noroeste y disminuyen hacia la región sureste
(Figura 7), para ambas estaciones.
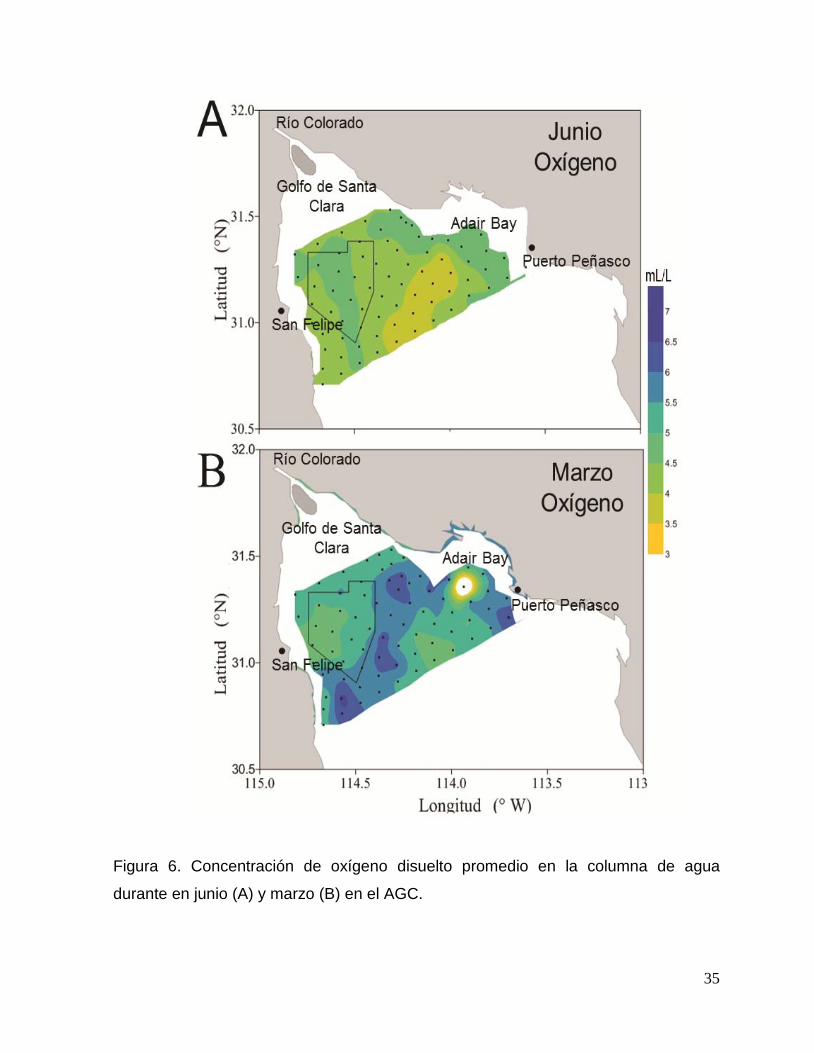
35
Figura 6. Concentración de oxígeno disuelto promedio en la columna de agua
durante en junio (A) y marzo (B) en el AGC.
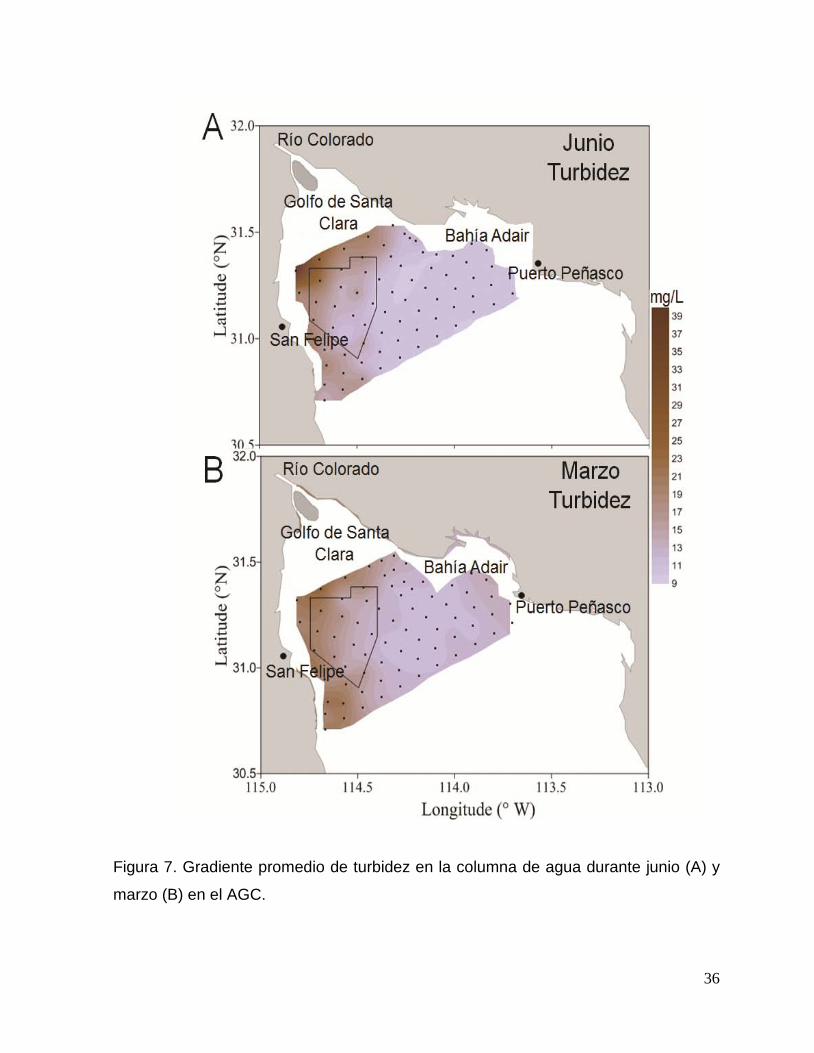
36
Figura 7. Gradiente promedio de turbidez en la columna de agua durante junio (A) y
marzo (B) en el AGC.

37
Temperatura y salinidad: De manera contrastante al resto de las variables, la
temperatura y salinidad presentaron diferencias significativas con los mayores
valores en el mes de junio (23.1°C y 36.1 ups, respectivamente), a diferencia de
marzo con valores promedio de 17.3°C y 35.8 ups (Tabla 6). Ambas variables
presentaron un gradiente SE-NO con los mayores valores hacia el oeste (Figura 8 y
9).
Clorofila “a”: Los mayores valores se registraron durante el mes de marzo, con
valores promedio de 1.7 μg/L; a diferencia de junio donde los valores promedio
fueron de 0.20 μg/L respectivamente (Tabla 6). Durante junio los mayores valores se
encontraron al norte (Figura 10 A), a diferencia de marzo donde se observan valores
más altos y homogéneos a lo largo del AGC a excepción de la región Sur, que
presenta las menores concentraciones (Figura 10 B).

38
Figura 8. Gradiente promedio de temperatura en la columna de agua durante junio
(A) y marzo (B) en el AGC.

39
Figura 9. Gradiente promedio de salinidad en la columna de agua durante junio (A) y
marzo (B) en el AGC.

40
Figura 10. Concentración promedio de clorofila “a” en la columna de agua durante
junio (A) y marzo (B) en el AGC.

41
Biomasa zooplanctónica: De igual manera que el resto de las variables, las
mayores concentraciones se encontraron en marzo, con valores promedio de 224
mL/1000m3, a diferencia de junio donde los valores promedio estuvieron alrededor de
los 136 mL/1000m3 (Tabla 6). En el caso de la biomasa zooplanctónica, en junio no
se encuentra ningún patrón evidente (Figura 11 A), aunque durante marzo las
mayores concentraciones se observan hacia la región Sur (Figura 11 B).
De acuerdo a las series de tiempo de SST (Figura 12 A) y SSChl “a” (Figura
12 B) las tres áreas mencionadas muestran una fuerte señal anual, pero permanecen
fuera de fase por alrededor de seis meses. El máximo de SST se presenta en
agosto-septiembre (~32.2°C) y el mínimo en enero-febrero (~14.7°C), mientras la
SSChl “a” presenta su máximo en marzo-abril (9.2μg/L)) y mínimo en julio-septiembre
(0.3μg/L). El máximo de SST durante los meses cálidos, lo presenta la región Oeste
(32.2°C), seguida de la Este (31.8°C) y Sur (31.6°C), con valores similares. El
mínimo durante los meses fríos, lo presenta la región Sur (14.7°C), seguidos de la
región Oeste (14.9°C) y Este (15.4°C).
En cuanto a los máximos de SSChl “a” durante los meses cálidos los presenta
la región Oeste (0.9μg/L), seguido de la Este (0.5μg/L) y la Sur (0.3μg/L), pero para
los meses fríos la Sur (6.3μg/L), Oeste (4.7μg/L) y Este (3.9μg/L). A pesar de las
diferencias en la concentración de SSChl “a” en las series de tiempo (2002-2012), se
observa una relativa estabilidad en el sistema (Figura 12 A), presentando los picos
de productividad, siempre durante la misma temporada. De igual manera la
temperatura muestra estabilidad a través de los años (Figura 12 B).
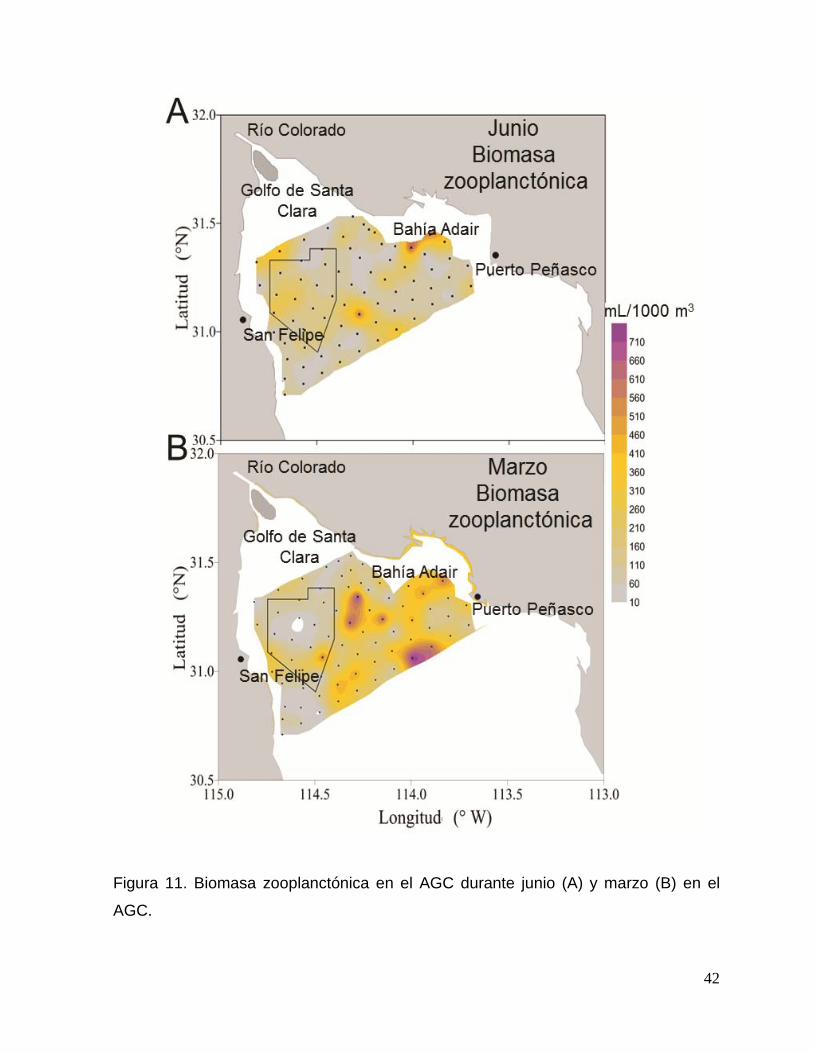
42
Figura 11. Biomasa zooplanctónica en el AGC durante junio (A) y marzo (B) en el
AGC.

43
Figura 12. Series de tiempo de A). Clorofila y B). Temperatura superficial del mar.
Ambas abarcan un periodo de 10 años y cada color representa las tres áreas
arrojadas por el análisis SIMPROF para el AGC. Verde (Sur), azul (Oeste) y rojo
(Este). Fuente: Aqua/MODIS; http://oceancolor.gsfc.nasa.gov.
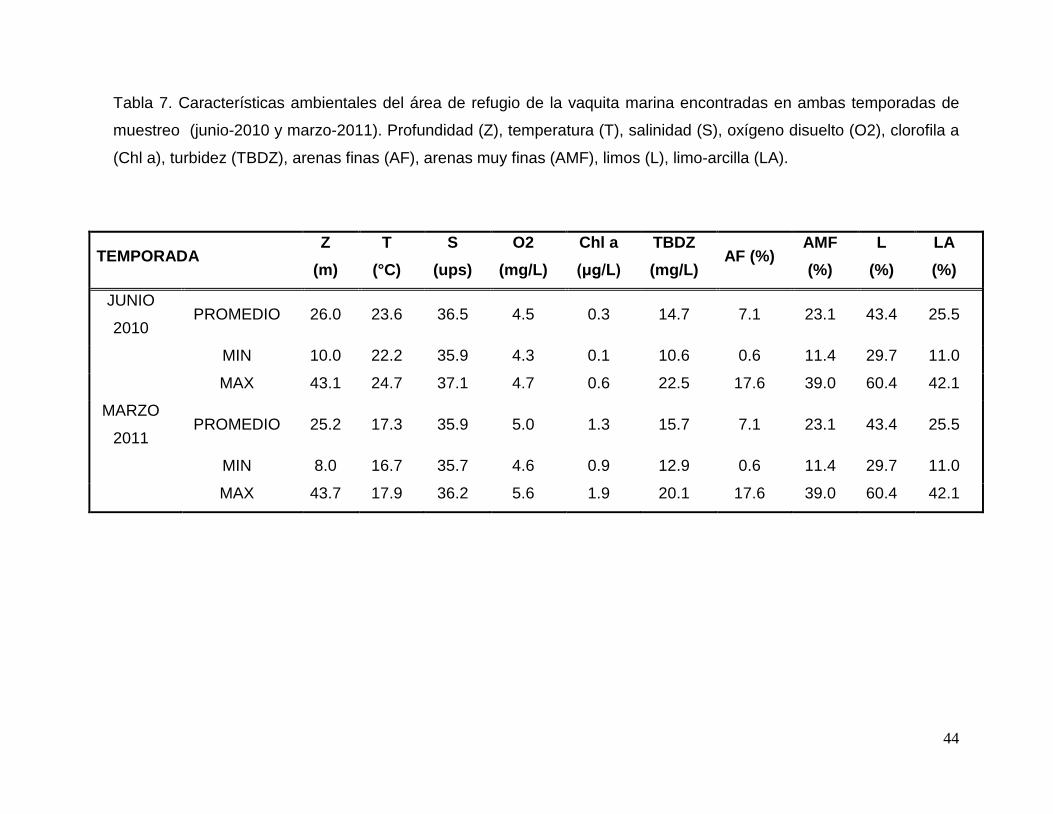
44
Tabla 7. Características ambientales del área de refugio de la vaquita marina encontradas en ambas temporadas de
muestreo (junio-2010 y marzo-2011). Profundidad (Z), temperatura (T), salinidad (S), oxígeno disuelto (O2), clorofila a
(Chl a), turbidez (TBDZ), arenas finas (AF), arenas muy finas (AMF), limos (L), limo-arcilla (LA).
TEMPORADA Z
(m) T
(°C) S
(ups) O2
(mg/L) Chl a (μg/L)
TBDZ (mg/L)
AF (%) AMF (%)
L (%)
LA (%)
JUNIO
2010 PROMEDIO 26.0 23.6 36.5 4.5 0.3 14.7 7.1 23.1 43.4 25.5
MIN 10.0 22.2 35.9 4.3 0.1 10.6 0.6 11.4 29.7 11.0
MAX 43.1 24.7 37.1 4.7 0.6 22.5 17.6 39.0 60.4 42.1
MARZO
2011 PROMEDIO 25.2 17.3 35.9 5.0 1.3 15.7 7.1 23.1 43.4 25.5
MIN 8.0 16.7 35.7 4.6 0.9 12.9 0.6 11.4 29.7 11.0
MAX 43.7 17.9 36.2 5.6 1.9 20.1 17.6 39.0 60.4 42.1

45
8.3.3. Delimitación del hábitat potencial de P. sinus
La figura 3 es el resultado de la recopilación bibliográfica de 154
avistamientos de vaquita principalmente durante primavera y verano en los últimos
15 años (mayo-1992 a abril-2007).
El área de mayores avistamientos de la vaquita presenta como características
conservativas, porque no presentan diferencias significativas entre las dos
temporadas de muestreo (Tabla 7):
1). Una profundidad promedio ~26 m.
2). Un tipo de fondo constituido en más del 68% de sedimento limo-arcilloso.
3). Una turbidez >14.7 mg/L (con un rango de valores entre 10.6-22.5 mg/L).
Por otro lado las características del área con diferencias significativas entre los
dos periodos de muestreo fueron: la temperatura con un promedio de 23.6°C y
17.3°C para junio y marzo respectivamente, la salinidad con un promedio de 36.5 ups
y 35.9 ups; el oxígeno disuelto ~4.5 mL/L y 5.0 mL/L y la concentración de clorofila
0.3 y 1.3 μg/L, respectivamente (Tabla 7).
A partir de las estaciones que limitaban la región Oeste (Figura 3), por ser la
que presentó el mayor número de avistamientos en las última década, se definió un
polígono cuyas coordenadas fueron introducidas a la Calculadora de Distancias
Latitud/Longitud de la NOAA (http://www.nhc.noaa.gov/gccalc.shtml). La extensión
del hábitat potencial de la vaquita marina obtenida fue de 1794 Km2, cuyas
coordenadas se observan en la tabla 8 (Figura 3). Para describir el hábitat potencial
de la vaquita marina, se utilizaron únicamente las variables conservativas (Ver la
última sección de resultados).
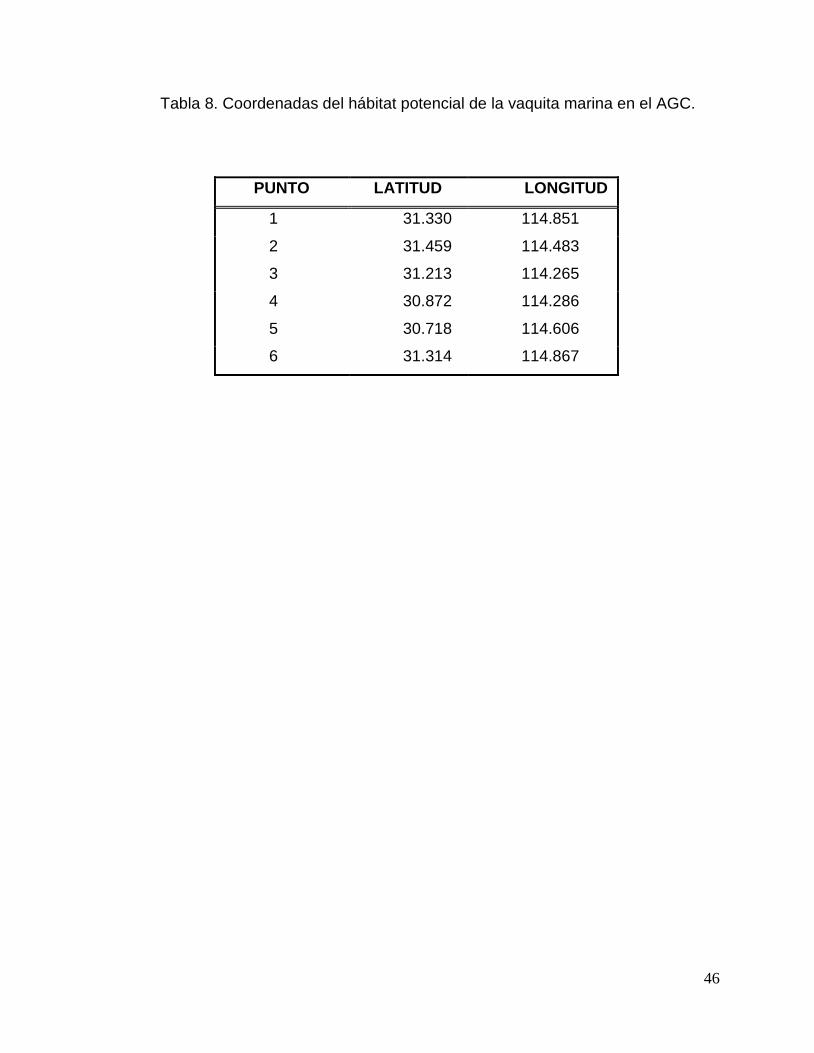
46
Tabla 8. Coordenadas del hábitat potencial de la vaquita marina en el AGC.
PUNTO LATITUD LONGITUD
1 31.330 114.851
2 31.459 114.483
3 31.213 114.265
4 30.872 114.286
5 30.718 114.606
6 31.314 114.867

47
8.4. DISCUSIÓN
8.4.1. Caracterización espacial del AGC:
Basados en el análisis SIMPROF (Figura 3 A y B) se regionalizó el AGC en
tres regiones: Este, Oeste y Sur. Al compararse las tres regiones, se encontraron
diferencias significativas para cada una de las variables (profundidad, temperatura,
salinidad, clorofila a, oxígeno disuelto, granulometría y turbidez) entre regiones, a
excepción de la turbidez y la clorofila “a” durante la campaña de junio 2010 (Tabla 1).
Estas diferencias fueron marcadas para la región Sur con respecto a las otras dos
regiones, la cual muestra características distintivas con respecto a las regiones
Oeste y Este (Tabla 4 y 5). Lo anterior confirma estudios previos (Sánchez-Velasco
et al. 2012) donde se encontraron características tanto hidrográficas como biológicas
(clorofila “a”, biomasa zooplanctónica y larvas de especies de peces dominantes) que
distinguen a la región de Cuenca Wagner del resto del AGC.
Sin embargo, al comparar las regiones Este y Oeste se encontraron
diferencias menos evidentes, las cuales se basaron principalmente en características
granulométricas y de turbidez que se repitieron en las dos temporadas de muestreo,
a excepción de las arenas finas durante marzo (Tabla 2 y 3). La diferencia nula en la
composición de arenas finas durante marzo, probablemente se debe a la ampliación
de la región Oeste (Figura 3 D), por la homogenización de las diferentes variables en
la columna de agua producida por la entrada de vientos del noroeste. Estos vientos
alcanzan velocidades de hasta 12 ms-1 provocando la mezcla intensa de la columna
de agua durante los meses fríos, a diferencia de la época cálida con vientos del
sureste cuyas velocidades no son mayores a 5 ms-1 (Lavín et al. 1997).
La granulometría (Figura 5) y la turbidez (Figura 7), mantienen el patrón
sugerido en estudios previos donde existe una mayor turbidez y predominancia de
grano fino en la región Oeste (Thompson, 1969; Carriquiry y Sánchez, 1999). Estas
características parecen ser consecuencia del aporte de más de 180 X 106 tons/año

48
de sedimentos arrojados por el Rio Colorado antes de su represamiento (Thompson,
1968; Álvarez y Jones, 2002), conservándolo hoy en día como un ambiente de alta
turbidez. Lo anterior, debido a que su régimen macromareal continúa reciclando las
planicies de marea y los depósitos poco profundos de la plataforma (Durazo-Arvizu,
1989; Argote et al. 1995; Alvarez y Jones, 2002).
8.4.2. Delimitación del hábitat potencial de P. sinus
Las características definidas para la zona de mayores avistamientos de la
vaquita marina en el presente estudio se dividieron en dos tipos:
1) Conservativas, es decir, aquellas que cambian entre las dos estaciones
de muestreo como la profundidad, un tipo de fondo de tipo limo-arcilloso y la
turbidez (Tabla 7).
2) No conservativas, aquellas que presentaron variación entre las dos
temporadas de muestreo. Dentro de estas se consideran la temperatura, la
salinidad; el oxígeno disuelto y la concentración de clorofila “a” (Tabla 7).
A partir de la relación entre la extensión de las regiones obtenidas mediante el
análisis SIMPROF basado en las variables ambientales del AGC, así como en la
recopilación de información sobre los avistamientos de la vaquita marina en la última
década (Figura 3 C y D), se definió un hábitat potencial para la vaquita. El presente
estudio, se refiere como hábitat potencial, a la región en la cual la especie pudiera
moverse por fuera del refugio para su protección o bien en caso de que los esfuerzos
para su recuperación fueran exitosos y su población aumentara.
Dentro de las variables descritas, una de las más importantes en la definición
de regiones ambientales fue la profundidad (Tabla 4 y 5). Esta variable, aunque no
tuvo diferencias significativas entre la región Oeste y Este, si parece tener influencia
sobre la permanencia de la especie en la región más somera, con no más de 60 m

49
de profundidad (Tabla 7; Figura 4). Esta preferencia fue mencionada anteriormente
tanto para la vaquita (Silber, 1990), como para la mayoría de las especies de
marsopas, las cuales presentan preferencia por áreas costeras poco profundas y
generalmente menores a 100 m (Ridgway y Harrison, 1999).
De estas características conservativas que definen el hábitat actual de la
vaquita, la mayor turbidez se corresponde con las preferencias generales de las
marsopas que tienden a concentrarse en áreas con influencia de descarga de ríos y
gran aporte de nutrientes (Watts y Gaskin, 1985; Palka, 1995; Ridgway y Harrison,
1999; Read, 1999). Estas zonas permiten la agregación de una abundante
concentración de presas (Watts y Gaskin, 1985; Palka, 1995; Read, 1999), lo cual
explicaría porque la especie se mantiene en este ecosistema capaz de sostener
grandes y complejas tramas tróficas (Millán- Núñez et al. 1999).
Por otra parte Silber (1988), sugiere que la preferencia de la especie en un
área con gran turbidez, le proporciona protección a la detección visual de los
depredadores. Esto debido a que la longitud de onda de sus vocalizaciones se
encuentra por debajo del rango auditivo de la orca y los tiburones, considerados
como sus depredadores. Adicionalmente, este ambiente de alta turbidez facilitaría a
la vaquita la captura de presas al no ser detectada de manera visual por estas,
mientras ella utiliza el sistema de ecolocación para ubicarlas (Silber, 1988).
Con respecto a la granulometría (Figura 2B), el predominio de sustratos
suaves (limo-arcillas) en el área, puede estar relacionado con la explotación de la
vaquita hacia recursos bentónicos de manera importante, que habitan este tipo de
ambientes (Findley y Nava, 1994; Pérez-Cortes et al. 1996).
Tomando en cuenta las características ambientales que predominan en la
región con el mayor número de avistamientos de la vaquita en la última década, se
determinó hasta donde se replicaban las variables conservativas. Estas últimas, se
consideran las de mayor influencia para la permanencia de la vaquita en esta región,

50
tanto por ser características reportadas para definir el hábitat de la mayoría de las
especies de marsopas (Watts y Gaskin, 1985; Ridgway y Harrison, 1999; Read,
1999), como por permanecer constantes a lo largo del tiempo.
De lo anterior se obtuvo una región con una extensión de 1794 km2,
considerada como el hábitat potencial para la especie y cuyas dimensiones se
extienden por 530.15 km2 más allá del área definida actualmente como refugio para
la vaquita (Figura 3 C y D).
La definición de este hábitat potencial para la vaquita marina presenta gran
relevancia, ya que al ser más amplio que la zona donde habita actualmente,
permitiría considerar con mayor precisión el área para proteger a esta especie y
mejorar sus posibilidades de conservación. Es importante considerar que la
delimitación de áreas naturales protegidas demasiado pequeñas, pueden resultar
poco efectivas como se ha mostrado también en el caso del delfín de Nueva Zelanda
(Cephalorhynchus hectori; Slotten, 2012). Este factor alcanza más relevancia, al
reconocerse que uno de los puntos señalados como críticos en la última década para
un plan de manejo exitoso, es la identificación del área que ocupa la especie durante
todo el año (Jaramillo-Legorreta, 2008).
8.4.3. Variaciones temporales de las variables ambientales en el AGC
El hábitat potencial para la vaquita marina, encontrado en el presente estudio,
se extiende hacia el Este y Sur por fuera del polígono establecido como zona de
refugio para la especie. Esta área guarda las características que al parecer están
influyendo en la permanencia de la especie en la región. Lo anterior podría otorgarle
a la vaquita una mejor protección, en caso de que su población se recupere y su
distribución espacial se amplíe. Al mismo tiempo, puede servir como protección de
los individuos de esta especie, que eventualmente se mueven fuera del polígono y se
encuentran vulnerables a las actividades pesqueras que se llevan a cabo fuera de

51
este.
Contrario a lo reportado por otros estudios sobre la poca importancia que
representa para la vaquita el nulo aporte del Rio Colorado al área (Farfán y Álvarez-
Borrego, 1992; Cupul-Magaña, 1994; Santamaría-del-Ángel et al., 1994; Álvarez-
Borrego, 2003), en el presente estudio se recomienda no minimizar la influencia de
sus aportes en la selección de hábitat de la vaquita. Esto puede ser respaldado por
registros hechos durante el año 1993, cuando se observó un aumento en la descarga
del flujo hacia el AGC asociadas a un mayor deshielo y precipitación, lo cual originó
que el área exhibiera condiciones clásicas estuarinas (Lavín y Sánchez, 1999), en
este año también se registró el avistamiento de la vaquita al sur de Puertecitos, BC
(Barlow et al., 1997)
Aunque no es posible relacionarlo de manera directa por falta de datos, es
importante mencionar que otros avistamientos reportados en la parte suroeste del
AGC (1986-1988 y 1992) por Barlow et al, (1997) y Silber (1990), coinciden con
eventos ENSO (El Niño Southern Oscillation;
http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.sht
m). Estos eventos se caracterizan por periodos de alta precipitación, causando que
las presas en Estados Unidos abran sus compuertas para la liberación de flujo. Lo
anterior aumenta la llegada de grandes cantidades de sedimento y un aumento en la
turbidez del área, ampliando el hábitat de la especie. El aumento en el flujo del Rio
Colorado hacia el AGC y su relación con eventos ENSO fue reportado y
correlacionado previamente con la mayor captura de camarón en el área, sugiriendo
que el decrecimiento en la descarga del Rio Colorado hacia el delta y estuario puede
afectar negativamente la captura de estos organismos, debido entre otros factores al
aumento en la salinidad del agua reduciendo así su hábitat de crianza (Galindo-Bect
et al. 2000). Lo anterior es importante porque a pesar de que en ninguno de los
estudios sobre los hábitos alimentarios de la vaquita marina, basados en análisis de
contenido estomacal se han reportado al camarón como presa de la especie, si se
han registrado muertes incidentales en redes camaroneras (D’Agrosa et al. 1995;

52
Vidal, 1995). Esto podría sugerir que la vaquita complementa su dieta con este tipo
de organismos, haciéndola vulnerable tanto por la modificación de su hábitat como
por la disminución de algunos tipos de las presas que consume.
Basados en análisis in situ de las diferentes variables ambientales, como con
series de tiempo de clorofila y SST (Figura 12 A y B), además de reportes previos
sobre la granulometría que muestran que la constitución del tipo de suelo ha
permanecido estable durante al menos la última década, (Thompson, 1979;
Carriquiry y Sánchez, 1999) se puede inferir, la permanencia de la vaquita en el área
noroeste durante todo el año. Esto respalda lo reportado por Silber (1988 y 1991) y
Gerrodette et al (2011).
Finalmente es importante mencionar, que aunque el AGC presenta
variabilidad importante entre ambas temporadas de muestreo en cuanto a la
temperatura, salinidad, oxígeno y clorofila “a” (Tabla 7), las características más
relevantes en el hábitat de la vaquita parecen ser permanentes a lo largo del año.

53
9. CAPÍTULO 2 REGIONES ISOTÓPICAS DE C Y N EN EL AGC COMO TRAZADORES DE
LA LINEA BASE DE Phocoena sinus 9.1. INTRODUCCIÓN
Uno de los principales problemas que actualmente presentan los planes de
manejo de la vaquita marina, es la escasa información sobre sus movimientos,
distribución, uso de hábitat y conducta de alimentación que permitan delimitar las
áreas críticas para la recuperación exitosa de la especie (Jaramillo-Legorreta, 2008).
Actualmente, una de las técnicas más utilizadas en ecología que puede aportar más
información necesaria de este tipo, es el análisis de isótopos estables, sobre todo de
carbono y nitrógeno. Actualmente, la construcción de mapas de distribución
geográfica de valores isotópicos (ISOMAPAS) en la línea base (es decir de la fuente
de nutrientes, sedimentos superficiales, materia orgánica, fitoplancton y zooplancton
que constituye los primeros niveles de una trama trófica), nos permiten obtener
información sobre los movimientos de las especies de interés (Graham et al. 2010).
Lo anterior origina, que los consumidores que hacen uso de los recursos de esa
región, presenten una señal isotópica similar en sus tejidos, que nos permita trazar la
ruta trófica de la cual provienen (Takai et al. 2000; Aurioles et al. 2009; Graham et al.
2010; Paez-Rosas et al. 2012).
La construcción ISOMAPAS y su uso para trazar los movimientos de los
consumidores marinos, requiere que se tomen en cuenta algunos parámetros para
entender la variación en el valor δ13C y δ15N de la línea base. En el caso del carbono
isotópico esta variación es asociada a múltiples factores tales como la concentración
de CO2 atmosférico disuelto en la columna de agua, el cual presenta valores
empobrecidos en 13C (Rau et al., 1992; Goericke y Fry, 1994). En este caso cuando
la concentración de CO2(aq) aumenta en el medio, la alta disponibilidad para los
productores primarios permite que estos discriminen el isótopo pesado (13C) sobre el

54
ligero (12C), enriqueciendo sus tejidos con este último (Rau et al., 1992; Goericke y
Fry, 1994). Como consecuencia, cuando los productores encuentran las condiciones
ideales para aumentar su tasa de crecimiento, el CO2(aq) será asimilado en mayor
proporción, originando un enriquecimiento en el CO2 residual enriquecido en el
isótopo pesado (Goericke y Fry, 1994; Popp et al. 1998). Otros factores que influyen
en el fraccionamiento isotópico por el fitoplancton son el tamaño de las células, la
composición taxonómica (Fry y Wainright, 1991; Popp et al. 1998) y la influencia de
carbono bentónico proveniente de macrofitas costeras, las cuales son enriquecidas
en comparación con el fitoplancton pelágico (Michener y Schell, 1994; France, 1995).
En el caso de la composición de δ15N de los productores, esta es dependiente
del valor isotópico de la fuente de nutrientes, siendo los aportes de nitrógeno
“regenerado” (NO3, NH4) más enriquecidos en 15N que nitrógeno “nuevo” (N2)
(Montoya, 2007). Lo anterior como consecuencia del grado en el cual estos fueron
reciclados por procesos microbianos (Montoya, 2007; Graham et al. 2010). Además
de lo anterior, la proporción isotópica del nitrógeno pesado (15N) en los tejidos del
consumidor, también se ve influida por la posición trófica de las presas que consume
(Minagawa y Wada, 1984). Se ha observado que en las tramas tróficas, cuando los
organismos se alimentan y asimilan el nitrógeno de su dieta, se excreta
preferencialmente el 14N y los consumidores empiezan a enriquecerse en 15N
respecto a su alimento (Steele y Daniel, 1978). Debido a lo anterior, la principal
utilidad de los isótopos de nitrógeno en estudios ecológicos, es como trazador de los
niveles tróficos que ocupan los organismos en su ambiente (Minagawa y Wada,
1984).
Aunque en el AGC, no se conocen estudios sobre regiones isotópicas que
puedan caracterizar el área, si se han observado valores enriquecidos desde la base
(sedimentos; Aguiñiga, 1999) hasta consumidores de niveles tróficos altos del área,
como el lobo marino de California, Zalophus californianus (Porras-Peters et al. 2008;
Aurioles et al. 2013), sobre todo en cuanto al nitrógeno. En el caso particular de la
vaquita marina, se han observado valores enriquecidos para ambos isótopos,

55
ubicándola como un depredador costero-estuarino de alto nivel trófico (Rodríguez-
Pérez et al. 2010). Teniendo en cuenta la utilidad de este tipo de información para
delimitar las áreas tróficas que son explotadas por especies depredadoras, en el
presente capitulo se propone la construcción de isomapas de las proporciones de
carbono (δ13C) y nitrógeno (δ15N) en la línea base a partir de un extenso muestreo de
sedimento, fitoplancton y zooplancton a lo largo del AGC; con el fin de utilizar esta
señal isotópica como una etiqueta natural para trazar la línea base del área de
forrajeo de la cual parte la vaquita marina.

56
9.2. MÉTODO
9.2.1 Diferencias temporales en las proporciones de δ13C y δ15N en la línea
base del AGC
Para evaluar las diferencias significativas en las proporciones isotópicas de
carbono y nitrógeno en la línea base del área de estudio entre las temporada
muestreadas (junio-2010 y marzo-2011), se realizaron mapas de gradiente isotópicos
de las muestras de sedimento, fitoplancton y zooplancton, usando el programa Surfer
v9.0. Las diferencias temporales (entre la estación cálida y fría) en el valor promedio
de δ13C y δ15N de sedimento, fitoplancton y zooplancton fueron evaluadas usando
una prueba estadística T-student. Los análisis estadísticos fueron realizados con el
programa STATISTICA 8.0.
9.2.2 Isomapas de δ13C y δ15N en sedimento y zooplancton del AGC
Para obtener las regiones isotópicas formadas a partir de los grupos que
conforman la línea base, se utilizó la matriz sin estandarizar correspondiente a los
valores de δ13C y δ15N del sedimento y zooplancton muestreados durante las dos
estaciones de muestreo. En este sentido el fitoplancton no fue utilizado para el
análisis por presentar un patrón menos claro de distribución de los datos. A los
resultados obtenidos, se les aplicó el Índice de disimilitud de Bray Curtis (1957) que
se basa en el grado en que dos muestras se diferencian una de la otra. Esta
disimilitud entre las muestras se convierte en distancias entre los resultados de la
muestra en un “mapa”. A partir de los datos obtenidos del Índice de disimilitud de
Bray Curtis se obtuvo un dendrograma mediante el método de unión flexible (β -.25),
con ayuda del programa PCORD v 6.0.
Para evaluar si existían diferencias significativas entre regiones, se utilizó la
prueba no-paramétrica de permutación multi-respuesta (ANOSIM) usando el

57
programa PRIMER v6.1.4, el cual no requiere supuesto de normalidad de varianza.
Esta prueba estadística describe la separación entre grupos, arrojando un valor
numérico R, donde el valor más positivo indica mayor separación (Clarke y Gorley
2006).
Con respecto a la relación de las variables ambientales y las regiones
isotópicas obtenidas a partir del análisis cluster, se elaboraron matrices
considerando: el valor promedio de cada una de las estaciones, para la columna de
agua de temperatura (°C), salinidad (UPS), Clorofila ”a” (mg/L), turbidez (Kg/m),
oxígeno disuelto (mL/L) y la granulometría. Dichos parámetros fueron tomados in situ
y representan los dos periodos de muestreo, a partir de las cuales se hizo el Análisis
Canónico de Correspondencias (ACC). La solución del ACC se muestra en un
diagrama de ordenación con las estaciones muestreadas y regiones obtenidas
representadas por puntos y las variables ambientales representadas por flechas (Ter
Braak, 1986). Finalmente las diferencias espaciales de δ13C y δ15N específicas en los
valores de sedimento fueron estimadas a partir de un análisis multivariado Kruskal-
Wallis.
9.2.3 Delimitación isotópica del área de alimentación de P. sinus
A partir de datos isotópicos de los diferentes grupos (sedimento, fitoplancton,
zooplancton, macrofauna, vaquita marina) muestreados en los alrededores de rocas
Consag (considerada como la región Oeste), se realizó una comparación con grupos
similares correspondientes a Bahía Adair (Spackeen, 2009). Los grupos
pertenecientes a la base de la trama trófica se obtuvieron mediante muestreo directo
y los datos de macrofauna se obtuvieron de la bibliografía. Lo anterior con el fin de
determinar el área que constituye el nicho trófico preferencial para la vaquita marina.

58
9.3. RESULTADOS
9.3.1 Diferencias temporales de δ13C y δ15N en plancton del AGC
Tanto en fitoplancton como en zooplancton se encontraron diferencias
significativas en los valores de δ13C y δ15N entre periodos de muestreo (T-test,
P=<0.05). Como se observa en las Figuras 13 A y C, las muestras de fitoplancton
colectadas durante marzo (-19.5‰ ±1.2‰) tienen valores más enriquecidos de δ13C
que las colectadas en junio (-21.4‰±1.1‰). En cuanto a los valores de nitrógeno
(Figura 14 A y C), se encontró un patrón similar con valores promedio de
13.3‰±1.2‰ en marzo y de 12.0‰±1.6‰ para las muestras colectadas durante el
crucero de junio (Tabla 9).
En cuanto al zooplancton (Figura 13 B y D), los valores más enriquecidos en
δ13C se encontraron en marzo (-19.1‰±1.1‰) en comparación a los registrados para
las muestras colectadas en la estación cálida (-20.2‰±1.2‰). La excepción al patrón
descrito anteriormente fue el δ15N en zooplancton (Figura 14 B y D), ya que no
presentó diferencias estacionales significativas (T-test, P=0.4556), aunque durante
junio los valores fueron ligeramente enriquecidos (15.1‰±1.4‰) con respecto a los
encontrados en marzo (14.9‰±1.4‰; Tabla 9).

59
Tabla 4. Diferencias estacionales en los valores de δ13C y δ15N en fitoplancton y
zooplancton del AGC. Las primeras dos filas corresponden a los valores isotópicos
encontrados en el sedimento muestreado en junio 2010. Sedimento (SED),
Fitoplancton (fito), Zooplancton (zoo).
JUNIO MARZO
VARIABLE PROMEDIO (MIN-MAX)
(‰)
PROMEDIO (MIN-MAX)
(‰)
δ13CSED -21.7 (-27.9--12.2)
δ15NSED 9.1 (2.6- 13.1)
δ13Cfito -21.4 (-23.4--18.3) -19.5 (-17.6--22.8)
δ15Nfito 12.0 (7.8-15.3) 13.3 (8.2-15.7)
δ13Czoo -20.2 (-23.9--18.1) -19.1 (-23.7--17.3)
δ15Nzoo 15.0 (12.2-18.5) 14.9 (12.1-18.7)
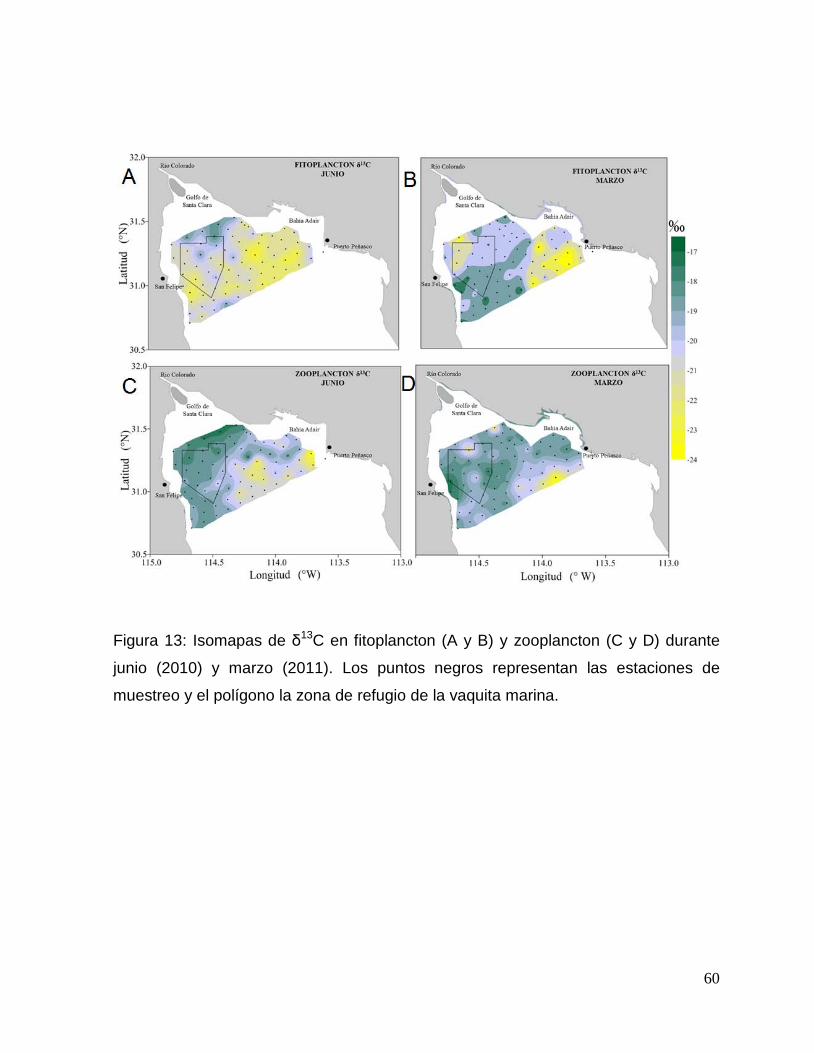
60
Figura 13: Isomapas de δ13C en fitoplancton (A y B) y zooplancton (C y D) durante
junio (2010) y marzo (2011). Los puntos negros representan las estaciones de
muestreo y el polígono la zona de refugio de la vaquita marina.
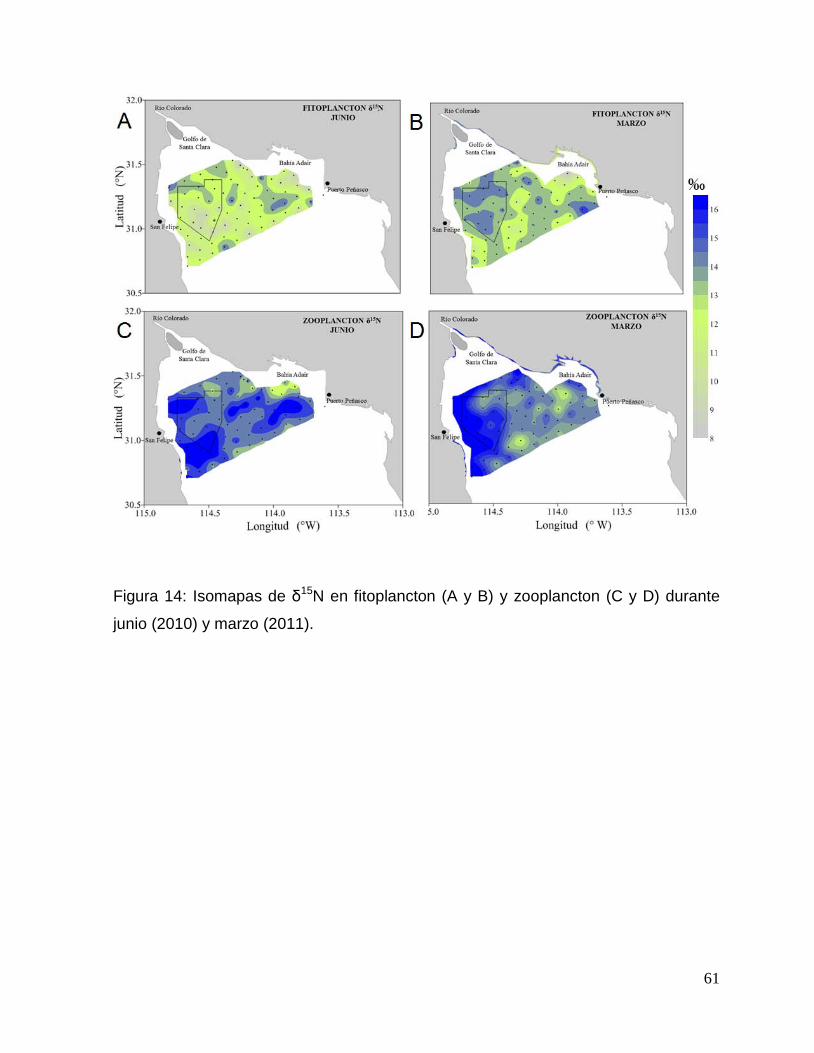
61
Figura 14: Isomapas de δ15N en fitoplancton (A y B) y zooplancton (C y D) durante
junio (2010) y marzo (2011).

62
9.3.2 Características isotópicas del sedimento
Con el propósito de definir si existían diferencias significativas en los valores
isotópicos de carbono y nitrógeno, se realizó un análisis multivariado a partir de las
regiones granulométricas determinadas por el programa TERNPLOT. Este análisis
determinó tres regiones definidas por su tamaño de grano: Oeste, Media y Este
(Figura 5).
De acuerdo a los valores obtenidos en el área de estudio para la razón
isotópica del nitrógeno se encontraron diferencias significativas entre las tres
provincias granulométricas mencionadas arriba (Kruskal-Wallis p<0.05). Además, los
resultados granulométricos están directamente correlacionados con los valores de
δ15N obtenidos del sedimento (r2> 0.05; p< 0.01), encontrando los valores más altos
en la región Oeste (10.7‰±1.6‰), con el tamaño de partícula más fino, dominada
por limos y arcillas. La parte media presenta valores intermedios 9.5‰±2.3‰, tanto
de isótopos estables como de tamaño de partícula, presentando una mezcla de
sedimento limoso con arenas finas (Figura 15B) y los más empobrecidos
(7.3‰±2.2‰) en la zona este donde predominan arenas finas.
Para definir si las diferencias en los valores de δ13C entre las tres regiones
granulométricas fueron significativas, se realizó una prueba multivariada, obteniendo
como resultado que no se presentaron diferencias significativas entre las regiones
(Kruskal-Wallis p= 0.8540). El valor promedio de δ13C obtenido para las tres regiones
granulométricas fue muy similar: oeste (-21.6‰±3.2‰), media (-21.9‰±2.9‰) y este
(-21.7‰±4.1‰). A pesar de lo anterior, se observó un enriquecimiento de δ13C en las
estaciones más costeras del AGC (-16.4‰±3.8‰; Figura 15A).

63
Figura 15: Gradiente isotópico de carbono (A) y nitrógeno (B) en sedimento
muestreado durante junio 2010.
9.3.3 Diferencias espaciales de δ13C y δ15N en plancton y sedimento del AGC

64
A la matriz transformada correspondiente a los datos de δ13C y δ15N en
sedimentos y zooplancton, por presentar la menor variabilidad espacial y entre
estaciones (junio-2010 y marzo-2011), se le aplicó el Índice de disimilitud de Bray
Curtis (1957). A partir de los resultados arrojados por el análisis se obtuvo el
dendograma que se muestra en la Figura 16A. Este dendograma es un diagrama en
forma de árbol que indica cómo se relacionan dichas variables y permite detectar las
tres regiones dentro del área de estudio que se ilustran en la Figura 16B.
Las regiones encontradas fueron denominadas de acuerdo a su distribución
en el área de estudio como sigue:
1). Oeste: Adyacente a la Península de Baja California
2). Este: Adyacente a las costas de Sonora
3). Costa: Son algunos puntos encontrados en la zona más costera
determinados sobre todo por los valores de δ13C
Las tres regiones mostraron diferencias significativas en sus valores isotópicos
(ANOSIM test, R=0.569), con el mayor enriquecimiento de δ13C en la región Costera
y para el δ15N en la región Oeste.
Las regiones isotópicas (ISOMAPAS) mencionadas, fueron también
detectadas por el análisis canónico de correspondencia (ACC), donde el 75% de la
varianza es explicada por los dos primeros ejes (Figura 17). Como se observa en la
figura 17 el ACC define claramente la región Este, principalmente correlacionada con
la mayor profundidad y el tipo de suelo arenoso (>60%); a diferencia de la región
Oeste la cual se correlaciona principalmente con la mayor temperatura, salinidad y
un predominio (>60%) de suelo de tipo limo-arcilloso. Con respecto a la región
Costera, que se define únicamente por 5 puntos marcados con rojo en la figura 16,
se caracteriza por presentar los mayores valores de δ13C en sedimento (-
14.8‰±3.3‰)
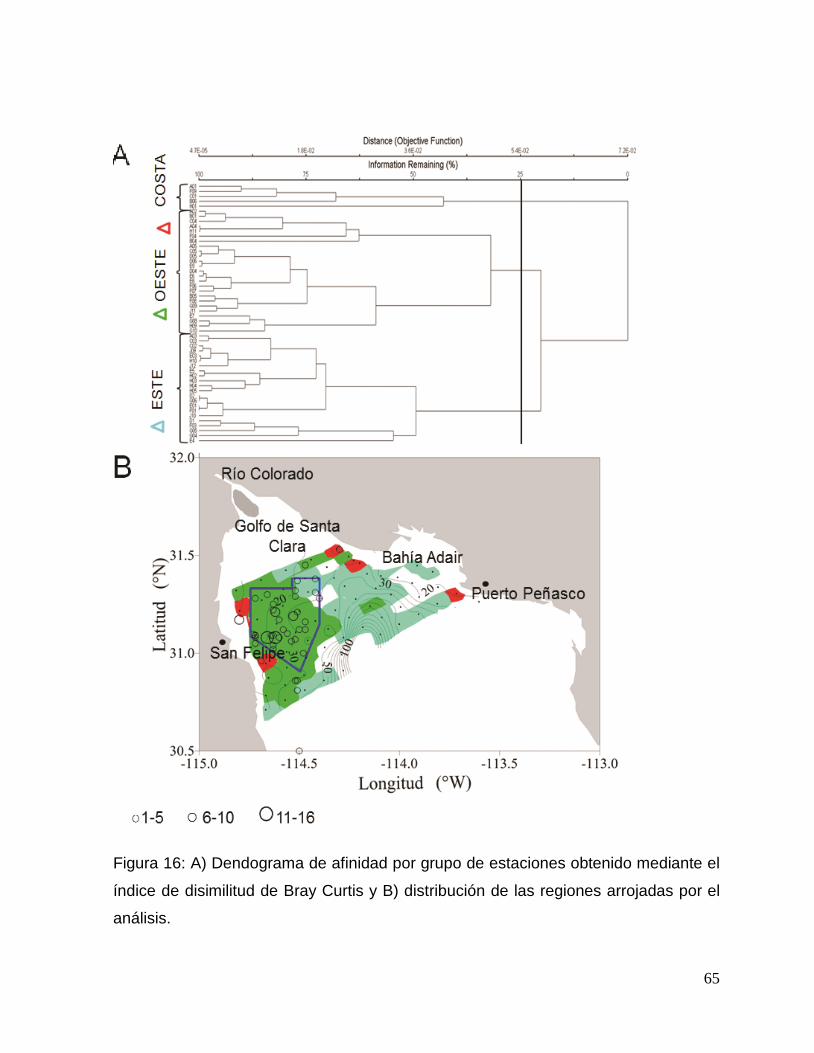
65
Figura 16: A) Dendograma de afinidad por grupo de estaciones obtenido mediante el
índice de disimilitud de Bray Curtis y B) distribución de las regiones arrojadas por el
análisis.

66
Figura 17: ACC donde se relacionan las variables ambientales con las regiones
isotópicas.
Finalmente, como se observa en la Tabla 10, la mayor correlación entre los
valores isotópicos y las variables ambientales se presentó entre la profundidad en
ambas temporadas (cálida, r=0.621 y fría, r=0.606) y el tamaño de grano (r=-0.5), así
como entre la temperatura (r=-0.649) y la salinidad (r=-0.744) durante la temporada
cálida.
Aunque para la regionalización mediante el análisis de Bray-Curtis no fue
considerado el fitoplancton, porque este presenta una variabilidad tan alta que
enmascara las variaciones menores de otros parámetros. Es importante resaltar dos

67
factores principales en los valores isotópicos de este: 1) El mayor enriquecimiento de
carbono fue encontrado en el norte (-19.0‰±0.6‰) y oeste (-18.4‰±0.7‰) para
verano e invierno, respectivamente. En contraste los valores isotópicos que sugieren
plancton oceánico se encontraron en el sureste (-21.9‰±0.7‰ y -21.2‰±1.0‰,
respectivamente. Figura 13 A y B) y 2).
No se encontró un claro patrón en los valores de δ15N durante el verano, solo
se observan parches de enriquecimiento a través del AGC (Figura 14 A y B). Aunque
en marzo el mayor enriquecimiento se encontró en el noroeste, como se reportó
anteriormente para el sedimento y el zooplancton. Aunque el enriquecimiento de δ13C
en el fitoplancton no se correlacionó con la concentración de clorofila (r2= 0.0028; P=
0.8783), ambos factores muestran un patrón similar con los mayores valores en el
noroeste (0.6±0.5 μg/l; Figura 10).
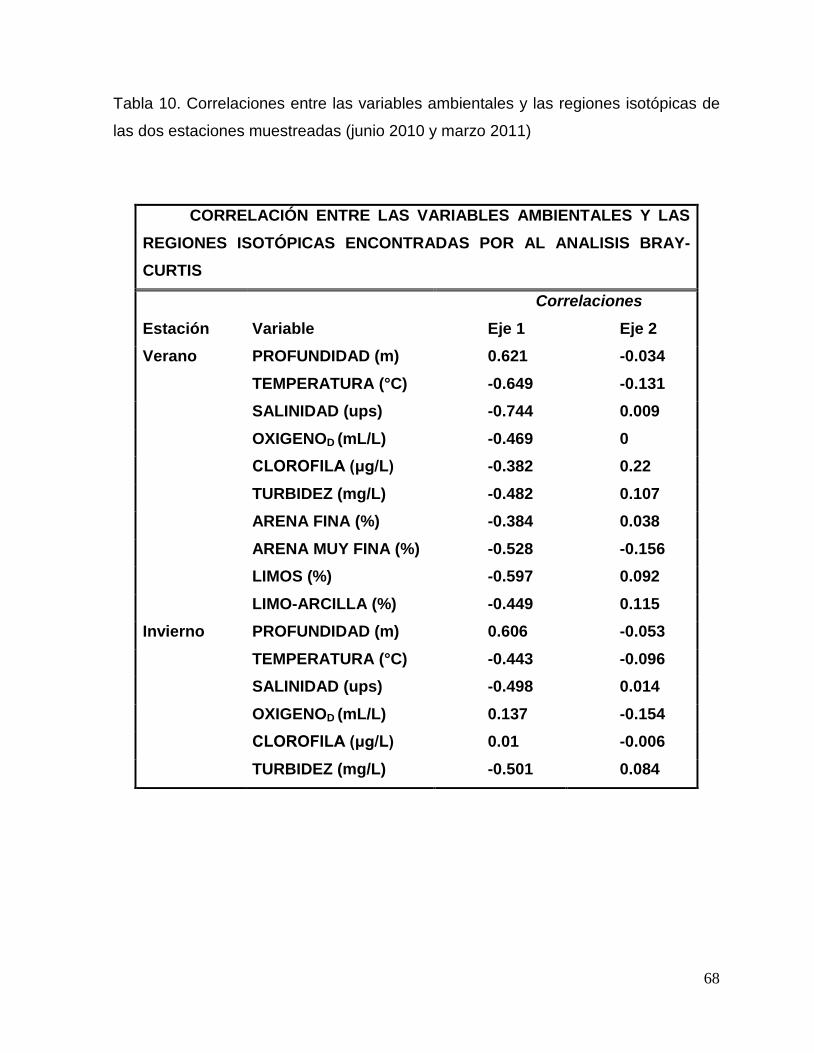
68
Tabla 10. Correlaciones entre las variables ambientales y las regiones isotópicas de
las dos estaciones muestreadas (junio 2010 y marzo 2011)
CORRELACIÓN ENTRE LAS VARIABLES AMBIENTALES Y LAS REGIONES ISOTÓPICAS ENCONTRADAS POR AL ANALISIS BRAY-CURTIS
Correlaciones Estación Variable Eje 1 Eje 2 Verano PROFUNDIDAD (m) 0.621 -0.034
TEMPERATURA (°C) -0.649 -0.131 SALINIDAD (ups) -0.744 0.009 OXIGENOD (mL/L) -0.469 0 CLOROFILA (μg/L) -0.382 0.22 TURBIDEZ (mg/L) -0.482 0.107 ARENA FINA (%) -0.384 0.038 ARENA MUY FINA (%) -0.528 -0.156 LIMOS (%) -0.597 0.092 LIMO-ARCILLA (%) -0.449 0.115
Invierno PROFUNDIDAD (m) 0.606 -0.053 TEMPERATURA (°C) -0.443 -0.096 SALINIDAD (ups) -0.498 0.014 OXIGENOD (mL/L) 0.137 -0.154 CLOROFILA (μg/L) 0.01 -0.006 TURBIDEZ (mg/L) -0.501 0.084

69
9.3.4 Determinación del área de alimentación de P. sinus a partir de δ13C y
δ15N
La característica tan particular del enriquecimiento en los valores de δ15N en la
región oeste del AGC se ve reflejada en los valores isotópicos de toda la trama
trófica. Como se observa en la figura 18 las diferencias más notables entre los
valores de la región Este (Spackeen, 2009) y Oeste (Tabla 11), se encuentran en: El
sedimento (3.5‰), fitoplancton (0.5‰), zooplancton (1.1‰), Chione (3.1‰), para los
gasterópodos bentónicos Atrina y Nassarius (6.4‰), Cynoscion (4.1‰), Mugil
(4.3‰), Oligoplictes (4.8‰) y Paralabrax (4.1‰). Una excepción fue la anchoa, la
cual presenta valores promedio de carbono y nitrógeno con una diferencia entre
áreas de sólo 0.6‰, con un ligero enriquecimiento en la región Este. En el caso del
δ13C, no se observó un patrón tan consistente, aunque en muchos casos si se
observó un ligero enriquecimiento en la región Este: sedimento (0.2‰), Atrina y
Nassarius (0.1‰), Anchoa (1.3‰) Cynoscion (0.8‰), Oligoplictes (2.0‰). Aunque el
resto de los grupos, presentan valores enriquecidos hacia la región Oeste:
fitoplancton (1.1‰), zooplancton (0.5‰) y Mugil (0.1‰). En el caso de los bivalvos,
no existió diferencia entre áreas.
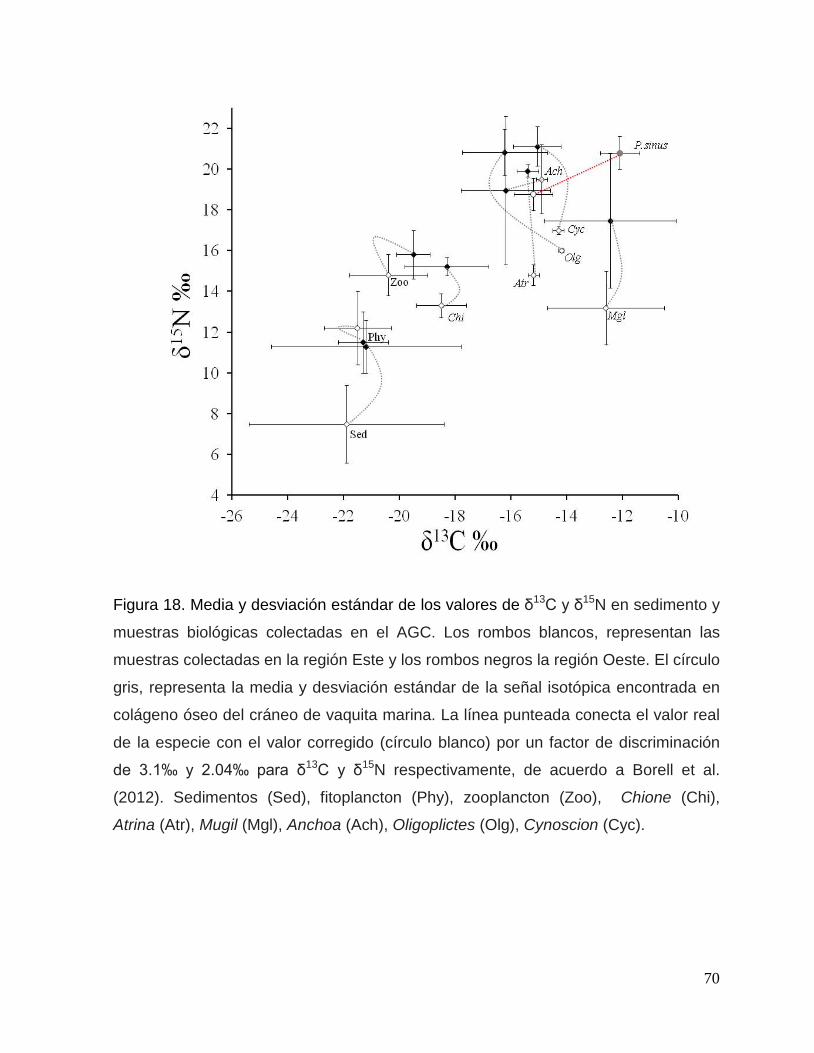
70
Figura 18. Media y desviación estándar de los valores de δ13C y δ15N en sedimento y
muestras biológicas colectadas en el AGC. Los rombos blancos, representan las
muestras colectadas en la región Este y los rombos negros la región Oeste. El círculo
gris, representa la media y desviación estándar de la señal isotópica encontrada en
colágeno óseo del cráneo de vaquita marina. La línea punteada conecta el valor real
de la especie con el valor corregido (círculo blanco) por un factor de discriminación
de 3.1‰ y 2.04‰ para δ13C y δ15N respectivamente, de acuerdo a Borell et al.
(2012). Sedimentos (Sed), fitoplancton (Phy), zooplancton (Zoo), Chione (Chi),
Atrina (Atr), Mugil (Mgl), Anchoa (Ach), Oligoplictes (Olg), Cynoscion (Cyc).

71
Tabla 11. Media y desviación estándar de los valores de δ13C y δ15N encontrados en sedimentos y muestras biológicas
colectadas en los alrededores de Rocas Consag (Oeste) y encontrados en la bibliografía pertenecientes a Bahía Adair
(Este; Spackeen, 2009) del AGC.
OESTE ESTE
GRUPO N δ13C±SD (‰) δ15N±SD (‰) C/N N δ13C±SD (‰) δ15N±SD (‰) C/N Sedimento 21 -21.4±3.4 10.7±2.0 9.1 10 -21.2±4.4 7.2±2.3 8.4
Fitoplancton 2 -20.1±1.5 12.7±1.6 6.0 20 -21.2±1.2 12.2±2.0 6.5
Zooplancton 42 -19.2±0.8 15.9±15.9 6.5 20 -19.7±1.4 14.8±1.0 6.5
Chione 4 -18.3±1.5 15.2±0.5 4.3 14 -18.3±0.6 12.1±0.9 ND
Atrina (Oeste), Nassarius (Este)
3 -14.9±0.7 19.3±0.6 3.7 8 -14.8±0.2 15.2±0.4 ND
Anchoa 2 -16.2±1.6 18.9±3.6 3.9 3 -14.9±0.2 19.5±1.7 ND
Cynoscion 3 -15.1±0.8 21.1±1.0 3.4 3 -14.3±0.2 17.0±0.2 ND
Mugil 2 -12.5±2.4 17.5±3.3 3.4 4 -12.6±2.1 13.2±1.8 ND
Oligoplites -16.2±1.5 20.8±1.1 3.5 3 -14.2±0.1 16.0±0.1 ND
P. sinus 15 -12.1±0.7 20.8±0.8 3.3

9.4. DISCUSIÓN
9.4.1 Diferencias temporales en los valores isotópicos de plancton en el AGC
La construcción de regiones isotópicas en el ambiente marino sobre escalas
temporales y espaciales ha constituido una valiosa herramienta para obtener
información sobre la relación de los consumidores tope son su ambiente. Los
resultados encontrados en el presente estudio muestran un mayor enriquecimiento
tanto de δ13C (Figura 13 A y B) y δ15N (Figura 14 A y B) en fitoplancton durante el
periodo frio probablemente sugiriendo una mayor mezcla por mareas y la entrada de
vientos del noroeste (Delgado-González et al., 1994). Estudios previos han mostrado
un marcado ciclo estacional en el patrón de vientos con la mayor intensidad en
invierno con vientos provenientes del norte (8 a 12ms-1) y con una menor intensidad
en verano cuando se presenta un cambio en la entrada de los vientos desde el
sureste (2 a 5ms-1) (Lavín et al., 1997). Esto puede causar constante mezcla en la
columna de agua, principalmente en los meses fríos (Lavín et al., 1998), originando
una mayor disponibilidad de los nutrientes del fondo para los productores primarios
resultando en mayores afloramientos durante la estación fría.
En conjunto con lo anterior los valores enriquecidos de δ13C pueden asociarse
a tasas rápidas de crecimiento en el fitoplancton, porque durante los procesos
fotosintéticos las células toman preferencialmente el 12C disuelto. Sin embargo, en
zonas de afloramiento, la demanda de CO2 acuoso por el fitoplancton supera su
disponibilidad, originando como consecuencia un menor fraccionamiento durante la
fotosíntesis (Goericke y Fry 1994; Graham et al., 2010). Esto enriquece los valores
de δ13C a lo largo de toda la trama trófica, como se observa en el zooplancton
(Figura 13 C y D).
En el AGC los valores de productividad primaria presentan una tendencia a
incrementarse durante las mareas vivas y los meses fríos, alcanzando valores de 76
mgC m–3 h–1, mostrando que la variación en la concentración de clorofila, la

73
abundancia de fitoplancton y la productividad primaria son fuertemente influenciados
por la mezcla producida por las corrientes de marea y por los efectos en la
estacionalidad (Millán-Núñez et al., 1999). Al respecto, en el muestreo realizado en
marzo, se observan las mayores concentraciones de clorofila (1.85 μg/L), en
contraste con la estación cálida donde el promedio fue de 0.40 μg/L (Figura 10 A y
B). Esto parece asociarse con la transición de mareas vivas durante el crucero de
marzo y la permanencia de mareas muertas durante el crucero de junio
(http://predmar.cicese.mx).
En cuanto a los valores de δ15N en fitoplancton, la mezcla por mareas puede
promover la entrada de nitrato a la zona eufótica reflejándose en el enriquecimiento
de los valores de 15NO3. Esto ocurre porque la mayor disponibilidad de nitrato
proveniente del fondo es enriquecido en δ15N. Además, estos afloramientos de
nutrientes, originan las condiciones idóneas para que la tasa de productividad
fitoplanctónica aumente, al ser asimilado por el fitoplancton la disponibilidad de 14NO3
disminuye y el valor de δ15N se incrementa acercándose al valor de δ15N de la fuente
de nitratos enriquecidos en el isótopo pesado (Montoya 2007). Un caso particular fue
el del δ15N en zooplancton que no presentó diferencia entre estaciones (Figura 14 C
y D), probablemente debido al consumo una fuente de proteína distinta proveniente
del fondo donde la actividad bacteriana es mayor.
9.4.2 Diferencias espaciales en las regiones isotópicas obtenidas del
sedimento y plancton en el AGC
Las tres regiones obtenidas del análisis Bray-Curtis (Figura 16 A), mostraron
diferencias significativas entre los valores de carbono y nitrógeno. El mayor
enriquecimiento de δ13C se presenta en la región costera, el cual es determinado
principalmente por los valores de carbono del sedimento, encontrando un valor
promedio de -15.4‰±2.5‰ y un rango de -11.9‰ a -17.9‰ (Figura 16 B), que está
dentro de valores encontrados para algas bénticas marinas (~-17.0‰±4.0‰), pero

74
más enriquecidos que los encontrados en especies fitoplanctónicas (~-21.0‰±3.0‰;
France, 1995). Este enriquecimiento en los sedimentos puede indicar la entrada de
materia orgánica de una fuente macrofitica, como macroalgas o pastos marinos más
que una fuente fitoplanctonica.
Como se observa en la Figura 15 A los valores de carbono en sedimento en la
zona media y suroeste presentan señales isotópicas promedio -21.8‰±1.1‰; con un
rango de valores entre -19.8‰ a -24.4‰. Finalmente, el δ13C en la zona norte
muestra un valor promedio de -26.6‰±0.9‰, con un rango de valores entre -25.4‰
a -27.9‰, similar al esperado para materia orgánica de origen terrestre (-26‰;
Cifuentes et al. 1988). La mayoría de los valores de δ13C en el AGC sin embargo son
similares (-21.7‰±3.3‰), a los reportados generalmente para el fitoplancton marino
(-22‰; France, 1995). La variación en los valores de δ13C en las distintas áreas del
AGC sugiere la influencia diferencial de al menos tres fuentes de MO (macrofitas,
fitoplancton y materia orgánica terrestre).
El δ15N en la región Oeste fue más enriquecido en el sedimento y el
zooplancton coincidiendo con el tamaño de grano más pequeño (suelo de tipo limo-
arcilloso); la región Este tuvo mayor proporción de arenas finas lo que se reflejó en el
ACC donde hubo correlación más alta entre los valores isotópicos con la
temperatura, la salinidad y el tamaño de grano (Figura 17; Tabla 10).
El presente estudio nos permite inferir sobre las diferentes fuentes de
nitrógeno en el AGC, usando como indicador el δ15N en los sedimentos, debido a que
los múltiples estados de oxidación del nitrógeno y las rutas metabólicas de bacterias
heterótrofas y quimio-autotróficas, permiten distinguir entre suplemento de nitrógeno
“nuevo” y “reciclado” a la zona eufótica (Dugdale and Goering 1967). Los resultados
encontrados muestran una diferencia de ~3.5‰ entre la región Oeste y Este, lo cual
podría ser explicado por dos factores principales entre otros; 1). El patrón de
circulación del AGC y 2). Las diferencias en el origen de los aportes de materia
orgánica hacia los sedimentos.

75
Considerando el primer factor, algunos estudios previos sugieren que la fuente
de sedimentos en el noroeste es derivado del reciclamiento por re-suspensión de la
estructura deltaica del Río Colorado (Carriquiry y Sánchez, 1999) cuyo transporte
posterior es consistente con una circulación ciclónica (Carriquiry et al. 2001). Esta
circulación podría generar la acumulación de la MO en la región Oeste y su
constante remineralización y reciclamiento por quimio y heterótrofos (Orozco-Borbón,
1995), lo que resultaría en un enriquecimiento del δ15N. En cuanto al segundo punto,
generalmente los valores elevados de δ15N en sedimentos coinciden con suelos de
tipo limo arcillosos (10.7‰ ± 2.0‰) que normalmente se asocian a estuarios con
influencia marina (10‰ a 33‰) y podrían reflejar la dominancia de material re-
procesado, donde el δ15N se enriquece durante la degradación (Fogel y Cifuentes,
1993).
En contraste los valores más empobrecidos en δ15N, se presentaron en la
región Este dominada por arenas finas (7.2‰ ± 2.3‰), más similares a MO de origen
planctónico (~8‰; Cifuentes et al., 1988). El rango de valores encontrado (2.6‰ a
13.1‰) muestra las diferencias en la fuente de MO que se deposita en los
sedimentos, encontrando los menores valores para la fuente terrestre de nitrógeno
orgánico, los cuales están en un promedio de ~3‰ para δ15N (Schoeninger and
DeNiro, 1984).
En el caso del fitoplancton, se puede observar que el mayor enriquecimiento
de δ13C, se presenta hacia la parte norte durante junio extendiéndose hacia el
suroeste en el mes de marzo. Se observa también la entrada de agua menos
enriquecida del sur con características oceánicas en ambas estaciones (Figura 13 A
y B). Sánchez-Velasco et al. (2012) reportaron un rango decreciente de clorofila
superficial de 0.45 mg/L hacia el norte a 0.1mg/L hacia el sur en el AGC, asumiendo
que esto se debe a la intrusión de agua baja en clorofila (<0.1 mg/L) con
características oceánicas proveniente del sur. También, Millan-Nuñez et al. (1999)
encuentran la mayor concentración de clorofila y abundancia fitoplanctónica hacia la

76
región peninsular de Baja California (2.6–18.2 mg m–3 y 274 cells mL–1) en
comparación con el lado continental de las costas de Sonora (1.8–12.7 mg m–3 y 166
cells mL–1).
9.4.3 Diferencias temporales en los valores isotópicos de la línea base del
AGC
Los resultados encontrados en el presente estudio muestran un mayor
enriquecimiento en el AGC, tanto de δ13C y δ15N en fitoplancton durante el periodo
frio probablemente sugiriendo una mayor mezcla por mareas y la entrada de vientos
del noroeste (Delgado-González et al., 1994). Estudios previos han mostrado un
marcado ciclo estacional en el patrón de vientos con la mayor intensidad en invierno
con vientos provenientes del norte (8 a 12ms-1) y con una menor intensidad en
verano cuando se presenta un cambio en la entrada de estos desde el sureste (2 a 5
ms-1) (Lavín et al., 1997). Esto puede causar constante mezcla en la columna de
agua, principalmente en los meses fríos (Lavin et al. 1998), originando una mayor
disponibilidad de los nutrientes del fondo para los productores primarios resultando
en mayores afloramientos durante la estación fría.
En conjunto con lo anterior los valores enriquecidos de δ13C pueden asociarse
a tasas rápidas de crecimiento en el fitoplancton, porque durante los procesos
fotosintéticos las células toman preferencialmente el 12C disuelto. Sin embargo en
zonas de afloramiento, la demanda de CO2 acuoso por el fitoplancton supera su
disponibilidad, originando como consecuencia un menor fraccionamiento durante la
fotosíntesis (Goericke and Fry 1994; Graham et al. 2010). Esto enriquece los valores
de δ13C a lo largo de toda la trama trófica.
En el AGC los valores de productividad primaria presentan una tendencia a
incrementarse durante las mareas vivas y los meses fríos, alcanzando valores de 76
mgC m–3 h–1, mostrando que la variación en la concentración de clorofila, la
abundancia de fitoplancton y la productividad primaria son fuertemente influenciados

77
por la mezcla producida por las corrientes de marea (Millán-Núñez et al. 1999). Al
respecto, en el muestreo realizado en marzo, se observan los mayores
concentraciones de clorofila (1.85 μg/L; Figura 10 B), en contraste con la estación
cálida donde el promedio fue de 0.40 μg/L (Figura 10 A). Esto parece asociarse con
la transición de mareas vivas durante el crucero de marzo y la permanencia de
mareas muertas durante el crucero de junio (http://predmar.cicese.mx).
En cuanto a los valores de δ15N en fitoplancton, la mezcla por mareas puede
promover la entrada de nitrato a la zona eufótica reflejándose en el enriquecimiento
de los valores de 15NO3. Esto ocurre porque la mayor disponibilidad de nitrato
enriquecido en δ15N proveniente del fondo y su completa asimilación por el
fitoplancton (Montoya 2007). Si la disponibilidad de nitrato disminuye, el valor de δ15N
puede incrementar acercándose al valor de δ15N de la fuente de nitratos (Graham et
al. 2010). Un caso particular fue el del δ15N en zooplancton que no presentó
diferencia entre estaciones, probablemente debido al consumo de una fuente de
proteína distinta proveniente del fondo donde la actividad bacteriana es mayor.
9.4.4 Diferencias espaciales de δ13C and δ15N en la trama trófica
La composición isotópica de los consumidores en sistemas marinos se
relaciona a la fuente de alimento y agua que el animal ingiere, de tal manera que
cuando estas muestran gradientes isotópicos, los datos pueden ser usados para
estudiar preferencia de hábitat, movimientos entre ecosistemas y los patrones
migratorios en el océano (Newsome et al. 2007).
El AGC presenta diferencias evidentes en la señal de δ13C y δ15N entre la
región Oeste y Este. Esto se observa en la Figura 18, la cual muestra estas
diferencias, por un lado la región Oeste representada por las Rocas Consag
(diamantes negros) y Bahía Adair, que representa la región Este (diamantes
blancos). Esta imagen muestra diferentes grupos partiendo de la base de la trama
trófica (sedimentos, fitoplancton y zooplancton), hasta los depredadores de alto nivel

78
trófico, en este caso representados por la vaquita marina o marsopa del Golfo de
California.
Las diferencias entre el δ13C y δ15N en la línea base (sedimento y fitoplancton;
δ13C: 0.2‰ y 1.1‰ and δ15N: 3.5‰ y 0.5‰, respectivamente); sugieren, en el caso
del sedimento, que la fuente de carbono es similar en la región Oeste y Este. En este
caso, si la circulación ciclónica predomina, entonces esperaríamos que el resto del
material biológico originado en el Este y Norte fuera transportado al oeste por efecto
de la misma, como se ha reportado en el caso del sedimento (Carriquiry et al. 2001).
En el caso del fitoplancton, la mayor diferencia del δ13C puede estar reflejando
la dinámica del sistema en constante movimiento que mantiene una resuspensión
constante de los nutrientes del fondo causando afloramientos en la región Oeste
(Millan-Nuñez et al. 1999). Si los valores de δ13C están relacionados con la tasa de
crecimiento del fitoplancton, entonces se esperaría que en la zona de mayor
productividad primaria, los organismos discriminen menos 13C enriqueciéndose en
comparación con la región Este, con una tasa de crecimiento fitoplanctónico menor.
En conjunto con lo anterior, la región Este refleja características menos costeras con
valores isotópicos empobrecidos. Estas características oceánicas en la masa de
agua han sido observadas en hábitats larvarios, previamente definidos (Sánchez-
Velasco et al. 2012). En el caso del δ15N las diferencias más pequeñas se
encontraron en el fitoplancton.
El zooplacton sigue la misma tendencia que el sedimento con valores más
enriquecidos en la región Oeste, con una diferencia con la región Este de 0.5‰ y
1.1‰ para δ13C y δ15N, respectivamente. El hecho de que el zooplancton fue similar
al patrón de enriquecimiento de 15N en los sedimentos, es más consistente con el
fraccionamiento isotópico de nitrógeno durante el crecimiento microbiano (Macko y
Estep 1984; Hoch et al. 1996), que con características fitoplanctónicas.

79
Los diferentes niveles tróficos (NT) presentan diferencias mínimas para
carbono en Chione and Mugil (0.2‰) y un mayor enriquecimiento en la región Este
para Atrina y Nassarius (-0.7‰), Oligoplictes (-2.0‰), Cynoscion (-0.8‰) y Anchoa (-
0.8‰). Esto coincide con la mayor abundancia de macrofitas para Bahía Adair, la
cual origina una mayor proporción de 13C en la trama trófica que se refleja en los
valores isotópicos de los organismos que hacen uso de esta.
En el caso del nitrógeno existe un patrón de enriquecimiento en la región Este
(Chione 1.9‰, Atrina 6.4‰, Oligoplictes 4.8‰, Cynoscion 4.1‰, Mugil 4.3‰), el cual
es soportado por los valores enriquecidos desde la línea base, indicando que todas
son especies que se alimentan en estas áreas el tiempo suficiente para que los
valores isotópicos de la fuente queden impresos en sus tejidos. En el caso de Atrina
y Nassarius aunque son bentónicos, los valores de nitrógeno pueden ser diferentes
en el lado oeste tanto por diferencias en la línea base, como porque su dieta es
constituida por diferentes fuentes de proteína. Un caso especial es la anchoa, la cual
muestra valores similares de δ15N en ambas zonas con una diferencia de 0.6‰ con
el mayor enriquecimiento en el Este indicando un posible movimiento de esta especie
entre ambas regiones.
En el caso particular de la vaquita marina como depredador tope y especie
indicadora de enriquecimiento del área, en la Figura 18 se muestran los valores
reales (círculo gris) y corregidos (δ13C: -15.2‰±0.5‰, δ15N: 18.6‰±0.8‰; círculos
blancos) mediante un valor de discriminación isotópica de 3.11 y 2.04 para δ13C y
δ15N respectivamente (Borell et al. 2012). El valor corregido permanece cercano a los
valores de las especies presa que corresponden a la región Oeste. Este
enriquecimiento respalda lo reportado en avistamientos históricos ubicando su
hábitat en el lado oeste del AGC (Barlow, 1986; Brownell; 1986, Silber, 1990;
Gerrodette et al., 1995; Barlow et al., 1997; Gallo-Reynoso, 1998; Jaramillo et al.,
1999; Vidal et al., 1999; Jaramillo-Legorreta. 2008; Gerrodette et al. 2011).

80
CAPÍTULO 3
ESTRUCTURA TRÓFICA Y AMPLITUD DE NICHO DEL HÁBITAT DE Phocoena sinus
10.1 INTRODUCCIÓN
La necesidad de conocer la estructura y función del ecosistema como
antecedente para logar la sustentabilidad de recursos acuáticos vivos ha sido
extensamente reconocida, particularmente en lo que se refiere a la estructura trófica
y el flujo de biomasa a través de la interacción de especies (Christensen y Pauly,
1993). En el caso de la vaquita marina, el estudio sobre su ecología presenta
dificultades importantes, debido a que técnicas convencionales como el análisis de
contenido estomacal u observaciones in situ, a pesar de proporcionar información
importante sobre su ecología, se han visto limitadas por el pequeño número de
muestra obtenido. Lo anterior, debido al grado de digestión de los organismos
recuperados y a factores como su pequeño tamaño poblacional y al poco tiempo que
pasa en superficie (Silber, 1988).
Por tales razones se propone como alternativa el análisis de isótopos
estables de δ13C y δ15N, ya que esta aproximación metodológica permite realizar
estudios sobre muestras existentes en colecciones científicas y de museos como
hueso, plumas, dientes, etc., además por el hecho de que no son necesarios
tamaños de muestra elevados (Hobson y Welch, 1992; Hobson et al., 1997; Walker
y Macko, 1999; Kelly, 2000; Lesage et al., 2001). Cabe mencionar que este es el
segundo estudio isotópico para conocer los hábitos alimentarios de la vaquita y
determinar su papel en la trama trófica que se presentan en su hábitat.
El análisis de isótopos estables en mamíferos marinos del AGC, ha probado
ser efectivo para el estudio de la diferenciación trófica entre colonias (Aurioles et al.
2009; Elorriaga-Verplancken y Aurioles-Gamboa, 2009), clases de edad (Niño-Torres

81
et al., 2006; Newsome et al. 2006; Porras-Peters et al. 2008; Elorriaga-Verplancken,
2009), así como para definir el grado de superposición trófica entre colonias de lobo
marino (Porras-Peters et al. 2008).
El área que comprende Rocas Consag se ha considerado como una anomalía
debida al característico enriquecimiento de δ15N encontrado en lobos marinos de los
alrededores de Rocas Consag (22.4‰ ± 0.5‰), que se presenta probablemente
desde la base de la trama trófica, en comparación con colonias de la misma especie
en el resto del Golfo de California (Porras-Peters et al. 2008).
En el caso específico de la vaquita, un habitante restringido al AGC, solo
existe un trabajo en el que se analizaron las proporciones isotópicas en vértebras y
dientes (Rodríguez-Pérez et al. 2010). En ese estudio, se observaron valores
promedio de δ13C de –11.99‰ ±1.59‰ (n= 12) y de δ15N de 21.98‰±2.04‰ (n= 12).
Estos valores muestran que P. sinus se ubica tróficamente en el nivel de carnívoro
terciario-cuaternario y con valores similares a otras especies de mamíferos marinos
de la misma región, sugiriendo que esta marsopa se puede alimentar de presas
comunes a otros depredadores de esta región. En el presente capitulo se utiliza esta
técnica con el objetivo de obtener un modelo estructural trófico del hábitat de la
vaquita.

82
10.2. MÉTODO
10.2.1 Determinación del nivel trófico:
Para determinar la estructura trófica y el papel que juega cada una de las
especies estudiadas en el ecosistema del AGC, la señal de δ 15N será convertida a
posición trófica (PT) usando la ecuación 2 (Pinnegar et al., 2002; Post, 2002).
PT= λ + (δ15Ndepredador - δ15Nbase)/∆n (ec. 2)
Dónde:
PT= Posición trófica de cada una de las especies a estudiar
λ= Posición trófica a la cual pertenecen los organismos utilizados como δ 15N
base, por ejemplo; en el caso del fitoplancton el valor de λ=1
δ15N= Valor delta del tejido tanto del consumidor como de la base de la trama
trófica. En el presente estudio el valor delta correspondería tanto al fitoplancton como
a los valores encontrados en sedimento, lo cual se asume como el inicio de la trama
trófica superficial y bentónica, respectivamente.
∆n= Valor asumido para el fraccionamiento que se presenta de un nivel a otro
en la trama trófica. Este valor se obtendrá a partir de las diferencias encontradas de
las observaciones hechas para los valores isotópicos entre los consumidores y sus
presas potenciales (Post, 2002).
10.2.2 Contribución porcentual de presas potenciales a la dieta de P. sinus
El papel trófico que desempeñan las especies muestreadas, así como si existe
o no impacto de la vaquita sobre las especies comerciales, se determinará utilizando

83
el modelo Bayesiano SIAR propuesto por Parnell et al. (2008) de acuerdo a la
ecuación 3:
(ec. 3)
Donde:
Xij= Valor observado del isótopo j del consumidor i. sjk= Valor del isótopo j en fuente k. Con la media (μjk) y la varianza (wjk2)
normalmente distribuidas.
cjk= factor de fraccionamiento trófico, con la media (λjk) y la varianza (τjk 2)
normalmente distribuidas.
pk= Proporción en la dieta de la fuente k, estimada por el modelo.
qjk= Concentración del isótopo j en la fuente k de acuerdo a Phillips y Koch
(2002)
εij= Error residual, describe la varianza adicional entre las observaciones
no consideradas y es estimado por el modelo.
Este modelo estima la probabilidad de distribución de la contribución de un
número n de fuentes a una mezcla, mientras evalúa la incertidumbre asociada a las
mismas, al fraccionamiento y a las señales isotópicas. Además, este modelo permite
la incorporación de información previa sobre la dieta del depredador al análisis. A
partir de los valores porcentuales obtenidos en el modelo se pretende inferir la
contribución de las diferentes especies a la dieta de la vaquita.

84
10.2.3 Análisis de amplitud de nicho
Se determinó la amplitud trófica de la vaquita, a partir del análisis de área
Convex-hull y la posterior comparación de la varianza encontrada entre los dos
grupos, de acuerdo a lo reportado por Jaeger et al. (2009). Finalmente, los resultados
fueron corroborados mediante la prueba estadística no paramétrica U- de Mann-
Whitney

85
10.3. RESULTADOS
10.3.1 Isótopos estables de δ13C y δ15N en P. sinus
La muestra de colágeno óseo de vaquita marina utilizada para el presente
estudio corresponde a una mezcla de juveniles (n=4), adultos (n=7) e individuos para
los cuales la edad no pudo ser determinada (n=4). Al aplicar la prueba estadística
multivariada Kruskal-Wallis no se encontraron diferencias significativas para δ13C o
δ15N (p= 0.1596 y p= 0.5552, respectivamente), o entre hembras (n=5), machos
(n=7) y organismos de sexo desconocido (n=3) (Kruskal-Wallis, p= 0.9663 y p=
0.9728, respectivamente), por lo cual todos los datos de los individuos muestreados
se integraron a un solo promedio. El valor promedio para los valores de δ13C y δ15N
en vaquita marina fue de -12.1‰±0.7‰ y 20.8‰±0.8‰ respectivamente. Los valores
encontrados para las diferentes especies y grupos muestreados en los alrededores
de Rocas Consag se muestran en la Tabla 12. Cabe mencionar que los valores de
vaquita utilizados pertenecen únicamente al número de muestra que no presenta
influencia de los lípidos sobre los isótopos de δ13C, a pesar de no haber sido
extraídos durante el procesado.
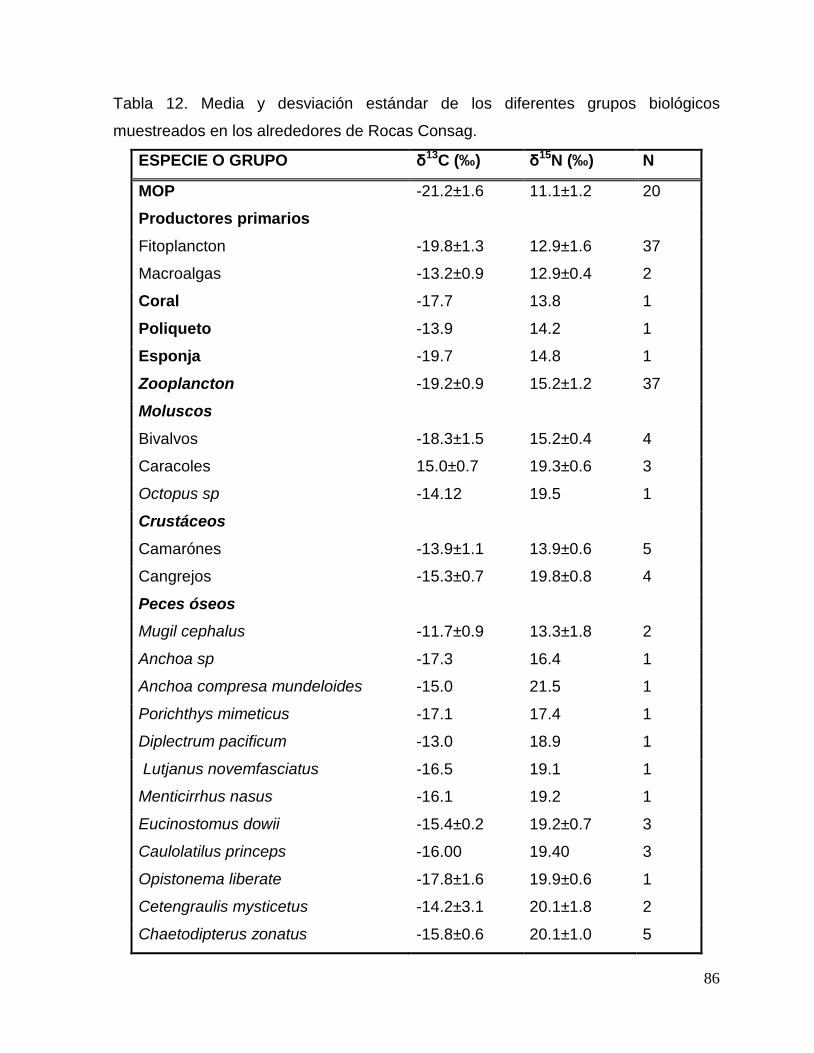
86
Tabla 12. Media y desviación estándar de los diferentes grupos biológicos
muestreados en los alrededores de Rocas Consag.
ESPECIE O GRUPO δ13C (‰) δ15N (‰) N
MOP -21.2±1.6 11.1±1.2 20
Productores primarios
Fitoplancton -19.8±1.3 12.9±1.6 37
Macroalgas -13.2±0.9 12.9±0.4 2
Coral -17.7 13.8 1
Poliqueto -13.9 14.2 1
Esponja -19.7 14.8 1
Zooplancton -19.2±0.9 15.2±1.2 37
Moluscos
Bivalvos -18.3±1.5 15.2±0.4 4
Caracoles 15.0±0.7 19.3±0.6 3
Octopus sp -14.12 19.5 1
Crustáceos
Camarónes -13.9±1.1 13.9±0.6 5
Cangrejos -15.3±0.7 19.8±0.8 4
Peces óseos
Mugil cephalus -11.7±0.9 13.3±1.8 2
Anchoa sp -17.3 16.4 1
Anchoa compresa mundeloides -15.0 21.5 1
Porichthys mimeticus -17.1 17.4 1
Diplectrum pacificum -13.0 18.9 1
Lutjanus novemfasciatus -16.5 19.1 1
Menticirrhus nasus -16.1 19.2 1
Eucinostomus dowii -15.4±0.2 19.2±0.7 3
Caulolatilus princeps -16.00 19.40 3
Opistonema liberate -17.8±1.6 19.9±0.6 1
Cetengraulis mysticetus -14.2±3.1 20.1±1.8 2
Chaetodipterus zonatus -15.8±0.6 20.1±1.0 5
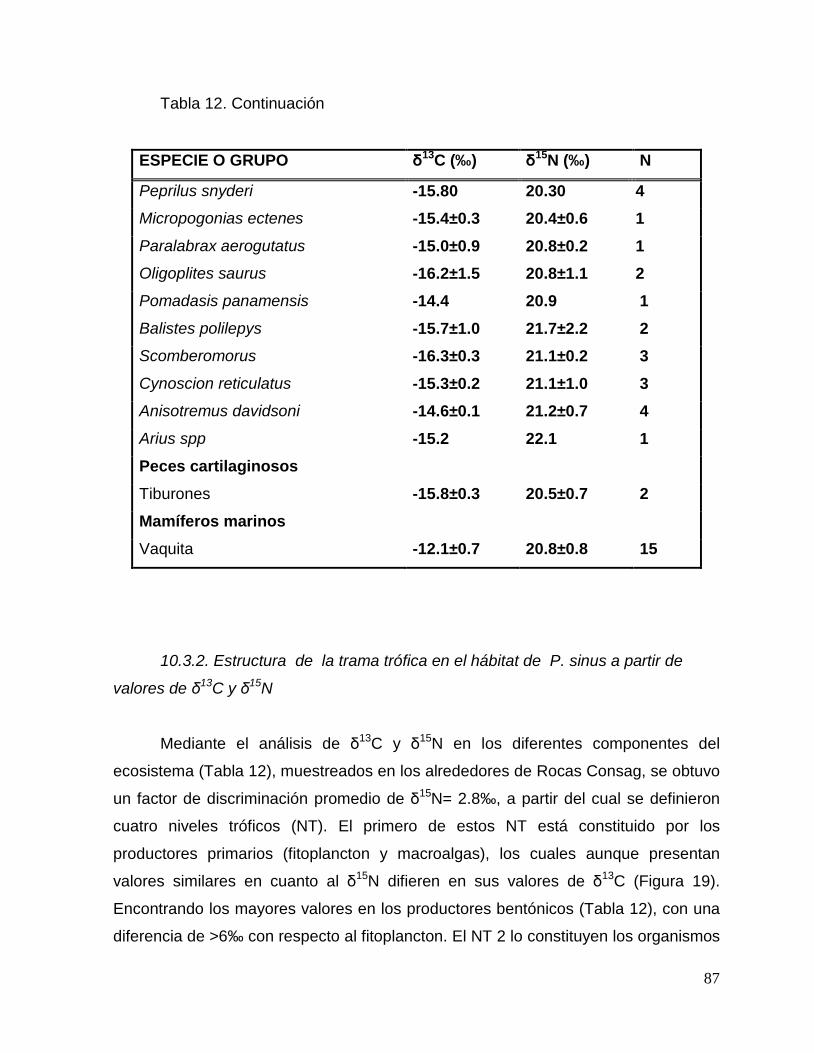
87
Tabla 12. Continuación
ESPECIE O GRUPO δ13C (‰) δ15N (‰) N
Peprilus snyderi -15.80 20.30 4 Micropogonias ectenes -15.4±0.3 20.4±0.6 1 Paralabrax aerogutatus -15.0±0.9 20.8±0.2 1 Oligoplites saurus -16.2±1.5 20.8±1.1 2 Pomadasis panamensis -14.4 20.9 1 Balistes polilepys -15.7±1.0 21.7±2.2 2 Scomberomorus -16.3±0.3 21.1±0.2 3 Cynoscion reticulatus -15.3±0.2 21.1±1.0 3 Anisotremus davidsoni -14.6±0.1 21.2±0.7 4 Arius spp -15.2 22.1 1 Peces cartilaginosos Tiburones -15.8±0.3 20.5±0.7 2 Mamíferos marinos Vaquita -12.1±0.7 20.8±0.8 15
10.3.2. Estructura de la trama trófica en el hábitat de P. sinus a partir de
valores de δ13C y δ15N
Mediante el análisis de δ13C y δ15N en los diferentes componentes del
ecosistema (Tabla 12), muestreados en los alrededores de Rocas Consag, se obtuvo
un factor de discriminación promedio de δ15N= 2.8‰, a partir del cual se definieron
cuatro niveles tróficos (NT). El primero de estos NT está constituido por los
productores primarios (fitoplancton y macroalgas), los cuales aunque presentan
valores similares en cuanto al δ15N difieren en sus valores de δ13C (Figura 19).
Encontrando los mayores valores en los productores bentónicos (Tabla 12), con una
diferencia de >6‰ con respecto al fitoplancton. El NT 2 lo constituyen los organismos

88
filtradores (esponjas y bivalvos), así como zooplancton y algunas especies de peces
como la Anchoa sp y Mugil cephalus (Figura 19). Estos últimos presentan valores de
δ15N menores de 17‰ (Tabla 12), aunque se encontraron representantes de las
mismas especies con valores de δ15N mayores a 20‰ que los ubica en un NT ≥4.0
(Figura 19). En el NT 3 se ubican organismos como el pulpo, los gasterópodos y
diferentes géneros de peces (Opistonema, Peprilus, Micropogonias, Diplectrum,
Cetengraulis, Menticirrhus, Caulolatilus, Lutjanus). Por último, el NT lo constituyen el
resto de los géneros de peces, tiburones y la vaquita marina (Figura 19), con valores
superiores a 20‰.
Debido a que estudios realizados sobre la alimentación de la especie indican
que la vaquita hace uso tanto de especies pelágicas como bentónicas, en el presente
estudio, siguiendo los criterios de Post (2002) se determinó el NT de la especie
tomando en cuenta por un lado la materia orgánica del sedimento (10.7‰±2.0‰) y
por el otro el bivalvo filtrador Chione (15.2‰±0.5‰). Este último se incluyó ya que de
acuerdo a Post (2002), este tipo de organismos presentan una menor variabilidad a
través del tiempo en su señal isotópica que el fitoplancton. Lo anterior debido
principalmente a que son organismos sésiles de larga vida que registran la señal del
ambiente pelágico de una manera más confiable. El factor de discriminación de δ 15N
utilizado para la obtención del NT de la vaquita marina fue de 2.04‰, de acuerdo a lo
reportado por Borrell et al. (2012) para colágeno óseo en cetáceos. El NT
determinado para la vaquita marina fue de 4.0 (Figura 19).
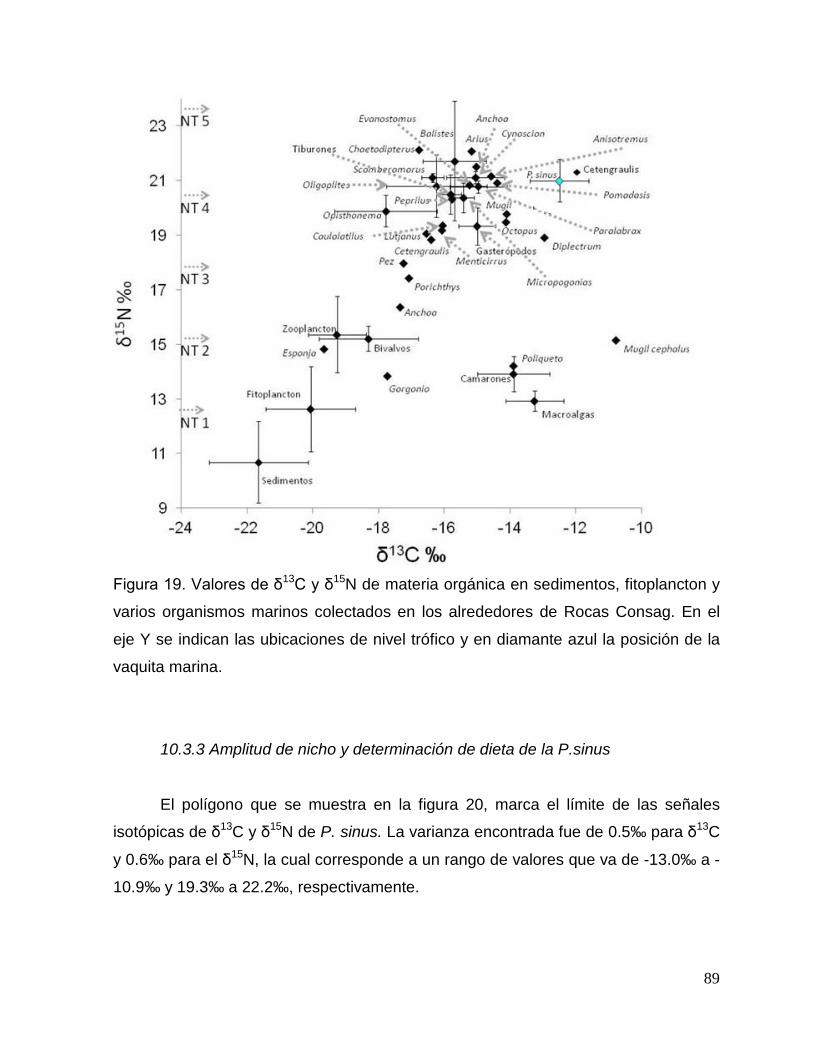
89
Figura 19. Valores de δ13C y δ15N de materia orgánica en sedimentos, fitoplancton y
varios organismos marinos colectados en los alrededores de Rocas Consag. En el
eje Y se indican las ubicaciones de nivel trófico y en diamante azul la posición de la
vaquita marina.
10.3.3 Amplitud de nicho y determinación de dieta de la P.sinus
El polígono que se muestra en la figura 20, marca el límite de las señales
isotópicas de δ13C y δ15N de P. sinus. La varianza encontrada fue de 0.5‰ para δ13C
y 0.6‰ para el δ15N, la cual corresponde a un rango de valores que va de -13.0‰ a -
10.9‰ y 19.3‰ a 22.2‰, respectivamente.

90
Figura 20. Amplitud de nicho isotópico obtenida mediante el área Convex-Hull de los
valores δ13C y δ15N de la vaquita marina.
Finalmente, para determinar la contribución de diferentes presas potenciales a
la dieta de la vaquita, se utilizó el modelo de mezcla Bayesiano SIAR (Parnell et al.,
2008), que indicó una baja probabilidad de contribución de cada una de las especies
que se introdujeron el modelo, con no más de un 8%. De acuerdo a los resultados
obtenidos, las especies M. cephalus y O. libertate (Tabla 13), contribuyeron con un
porcentaje mayor de 29.2% y 15.9%, respectivamente (Figura 21).
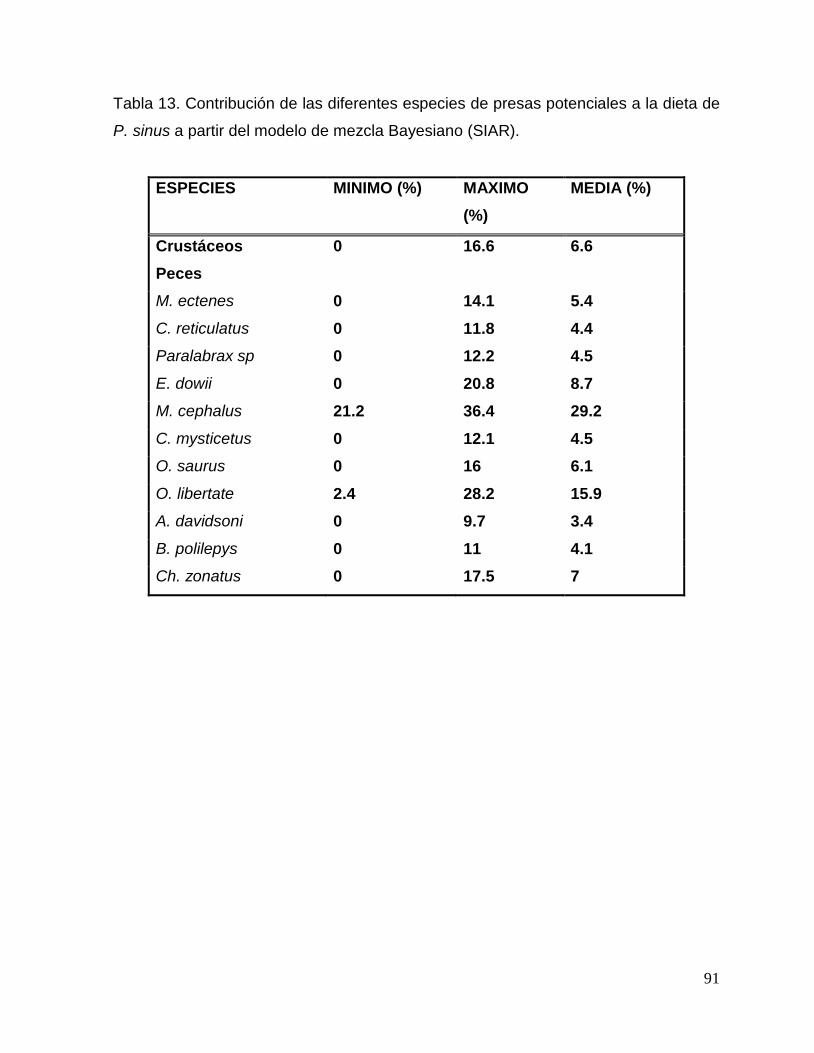
91
Tabla 13. Contribución de las diferentes especies de presas potenciales a la dieta de
P. sinus a partir del modelo de mezcla Bayesiano (SIAR).
ESPECIES MINIMO (%) MAXIMO (%)
MEDIA (%)
Crustáceos 0 16.6 6.6 Peces M. ectenes 0 14.1 5.4 C. reticulatus 0 11.8 4.4 Paralabrax sp 0 12.2 4.5 E. dowii 0 20.8 8.7 M. cephalus 21.2 36.4 29.2 C. mysticetus 0 12.1 4.5 O. saurus 0 16 6.1 O. libertate 2.4 28.2 15.9 A. davidsoni 0 9.7 3.4 B. polilepys 0 11 4.1 Ch. zonatus 0 17.5 7
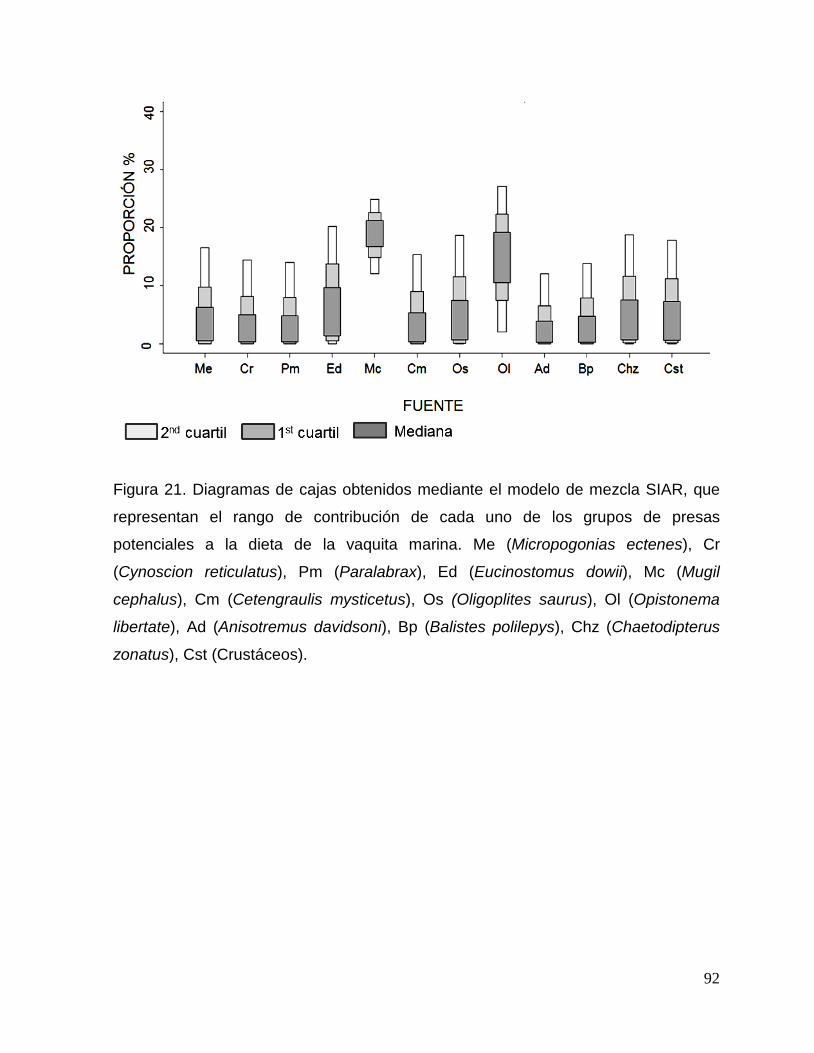
92
Figura 21. Diagramas de cajas obtenidos mediante el modelo de mezcla SIAR, que
representan el rango de contribución de cada uno de los grupos de presas
potenciales a la dieta de la vaquita marina. Me (Micropogonias ectenes), Cr
(Cynoscion reticulatus), Pm (Paralabrax), Ed (Eucinostomus dowii), Mc (Mugil
cephalus), Cm (Cetengraulis mysticetus), Os (Oligoplites saurus), Ol (Opistonema
libertate), Ad (Anisotremus davidsoni), Bp (Balistes polilepys), Chz (Chaetodipterus
zonatus), Cst (Crustáceos).

93
10.4. DISCUSIÓN
10.4.1 Señales de δ13C y δ15N en P. sinus
El análisis de isótopos estables en tejidos animales reflejan aquellos valores
de las tramas tróficas de las cuales parten, arrojando de esta manera, información
sobre su dieta y sobre su área de forrajeo (Rubinstein y Hobson, 2004). Los
resultados arrojados a partir de colágeno óseo en P. sinus muestran en el caso del
carbono (-12.1‰±0.7‰; Tabla 6) valores característicos de consumidores costeros,
como lo reportan Aurioles-Gamboa et al., (2013), quienes documentan valores en
Zalophus californianus pertenecientes a todas las colonias del Golfo de California de
-12.3‰±0.4‰, que también tiene hábitos costeros. Otro ejemplo de organismos
costeros lo muestra Díaz-Gamboa (2003), quien reporta dos ecotipos de Tursiops
truncatus, donde el ecotipo costero presenta valores de -12.9‰±0.4‰. Por otro lado,
los resultados de δ13C respaldan el uso de un hábitat similar a otras especies de
marsopas que se han reportado con hábitos costeros y cuyos valores son muy
similares por ejemplo; Toperoff (2002), reporta valores en Phocoena phocoena de -
13.4‰±0.5‰ y Schoeninger y DeNiro (1984), de -12.5‰±1.3‰, los cuales a pesar
de las diferencias latitudinales que presentan con respecto a la vaquita marina,
permanecen cercanos a los registrados para la especie.
.
En cuanto al nitrógeno, este muestra un enriquecimiento en P. sinus
(20.8‰±0.8‰), en comparación con P. phocoena (15.7‰±0.7‰) y Phocoenoides
dalli (16.2‰±0.6‰), las cuales no pueden ser comparables debido a las diferencias
latitudinales en sus ecosistemas respectivos. De acuerdo a Graham et al. (2010),
cuando los consumidores se alimentan en un ambiente específico por un tiempo
relativamente largo, adquieren la composición isotópica de sus presas locales. Por lo
anterior, la composición isotópica de un animal puede ser usada como un marcador
natural de su permanencia en un hábitat o de los movimientos a través de hábitats
diferentes. De tal manera que en el caso de la vaquita marina y los otros

94
Phocoenidos mencionados, los valores isotópicos nos indican un enriquecimiento
mayor en el área que ocupa la primera, debido al origen de nutrientes del cual parten
sus ambientes respectivos. A diferencia de lo anterior, la vaquita marina presenta
valores de δ15N similares a Z. californianus (21.0‰±0.9‰), el cual presenta hábitos
también una dieta predominantemente piscívora. Estos resultados, muestran la
importancia de conocer los valores de δ15N en la base de la trama trófica de la cual
parten los individuos al momento de inferir sobre hábitos tróficos, con el fin de no
subestimar o sobreestimar su papel dentro del ecosistema. Como se muestra en el
caso de la vaquita marina, los altos valores de nitrógeno, se deben más al origen de
nutrientes enriquecidos del cual parte la trama trófica que al consumo de presas de
niveles tróficos superiores a los que consumen otras especies de marsopas.
10.4.2 Estructura de la Trama Trófica de P. sinus
Actualmente es imperativo entender la extensión y estructura de las tramas
tróficas, para poder comprender como se relacionan las comunidades en los
ecosistemas. El análisis de isotopos estables, en particular de carbono y nitrógeno,
ha probado ser una valiosa herramienta para definir este tipo de cuestiones (Post et
al. 2000). Muchas investigaciones estudian los niveles tróficos porque estos son
sencillos de definir, caracterizan el papel funcional de los organismos y facilitan la
estimación de flujos de masa o energía a través de las comunidades ecológicas
(Post, 2002). A partir de las señales isotópicas de P. sinus y del resto de las especies
muestreadas (peces, macroinvertebrados, macroalgas, zooplancton, fitoplancton)
para el área oeste del AGC (Tabla 6), se determinó un fraccionamiento de δ 15N de
2.6‰±0.9‰ , respectivamente, mediante el cual se obtuvieron 4 niveles tróficos para
el área muestreada (Figura 14).
Como se observa en la figura 19, los valores isotópicos de carbono nos
permiten observar que la mayoría de los organismos presentan una tendencia hacia
un origen fitoplanctónico. A diferencia de lo anterior, especies como los camarones y
la vaquita marina presentan cierta influencia hacia un origen enriquecido en δ13C,

95
que parece reflejar el hábitat bentónico, cuyos valores parecen tener un aporte
importante de productores primarios bentónicos como las macroalgas. Lo anterior se
ve reflejado en la diferencia de 6.6‰ encontrada en los valores de δ13C (Figura 19)
entre productores pelágicos (fitoplancton) y bentónicos (macroalgas), aunque con
valores de δ15N similares (Tabla 12).
Debido a que el número de muestra de especies conocidas como presas de la
vaquita fue reducido, en el presente estudio se utilizó para la obtención de su NT un
factor de discriminación de 2.04, obtenido de colágeno óseo en cetáceos (Borell et al.
2012). Los resultados obtenidos definieron un NT para la vaquita de 4.0, muy
cercano al 4.2 obtenido por Pauly et al. (1998) para la especie, cuyos resultados se
basaron en el análisis de contenido estomacal.
Como se observa en la figura 19, la vaquita marina presenta valores de δ15N
similares a varias especies de peces consideradas como presas potenciales, así
como a las muestras obtenidas de especies reportadas como presas en estudios
tróficos de la especie (Anchoa sp y Cynoscion reticulatus). A pesar de estas
similitudes en cuanto al nitrógeno, en el caso del δ 13C difiere por más de 3‰ con
respecto a los organismos mencionados anteriormente. Lo anterior puede ser debido
a las diferencias en el tipo de tejido muestreado. Varios autores han documentado
que el factor de discriminación dieta/tejido varía de acuerdo a las características
como tipo de dieta (calidad nutricional) metabolismo (enrutamiento isotópico, tasa de
crecimiento, síntesis de aminoácidos), así como a la variación de la composición de
aminoácidos o lípidos en los tejidos muestreados (Hobson et al. 1996; Koch, 2007;
Newsome et al. 2010). El colágeno óseo en particular presenta gran diferencia en el
factor de discriminación de la proteína comparado con otros tejidos (Hobson et al.
1996; Koch, 2007; Borrell et al. 2012), que debe ser tomada en cuenta cuando éste
tipo de tejido es usado para determinar nivel trófico o para evaluar la dieta de los
organismos en reconstrucciones isotópicas a largo plazo (Borrell et al. 2012).

96
La diferencia encontrada en los valores isotópicos de vaquita en cuanto al
δ13C, con respecto a otros organismos del ecosistema como los peces, respalda la
idea de que el colágeno óseo presenta un factor de discriminación mayor que el
encontrado en otros tipo de tejidos (Koch, 2007; Newsome et al. 2010; Borell et al.
2012; Riofrío-Lazo et al. 2012). De tal manera que mientras tejidos suaves como
musculo, hígado y piel se enriquecen únicamente por 1-2‰ con relación a la dieta, el
colágeno óseo típicamente tiene valores entre 4-5‰ mayores que su alimento. Esto
se debe principalmente a su composición particular de aminoácidos donde la glicina
constituye en un 33% de la proteína total y este aminoácido está enriquecido en 13C
(+8‰) (Koch, 2007).
En cuanto al δ15N el factor de discriminación reportado se encuentra entre
2.0‰ y 3.0‰ para colágeno óseo de mamíferos (Koch, 2007; Newsome et al. 2010;
Borell et al. 2012). Asumiendo un factor de discriminación similar para la vaquita
marina, así como el hecho de que el hueso es una integración del alimento
consumido por un extenso periodo de tiempo (Tieszen et al. 1983; Hobson et al.
1996; Kurle 2002; Kurle and Worthy, 2002), se esperan valores similares de δ15N
entre la vaquita y algunas especies de peces (Figura 19), ya que aunque algunos
probablemente constituyan parte de su dieta, el colágeno óseo estaría resumiendo la
integración de esta información más la de organismos con valores isotópicos
menores (probablemente de NT inferiores), que pudieron no haberse muestreado
durante los cruceros.
10.4.3 Nicho trófico y proporción de dieta de P.sinus
Las aproximaciones a nivel poblacional usan frecuentemente tanto la posición
trófica como la amplitud de nicho, para entender su interacción en la trama trófica así
como su papel en el ecosistema (Bearhop et al., 2004). En el presente estudio se
utilizó el área Convex-hull evaluada por el análisis de varianza, una herramienta que
ha probado ser útil en la obtención de este tipo de información. A partir del análisis
de amplitud de nicho basado en la obtención de la varianza en la vaquita marina, se

97
encontraron varianzas de δ13C= 0.5 y δ15N= 0.6 (Figura 20). Si se considera, de
acuerdo a Bearhop et al. (2004) y Jaeger et al. (2009), que una varianza de más de
1‰ sugiere a un consumidor generalista y una varianza menor a menor a 1‰ a un
especialista, los valores en la vaquita indican un comportamiento con tendencia hacia
el especialismo. Lo anterior contradice lo reportado por Findley y Nava (1994) y
Pérez-Cortés et al. (1996) que la catalogaron como especie generalista.
Sin embargo es necesario tomar con reserva estos resultados ya que si bien la
vaquita podría presentar una alimentación generalista basados en el número de
especies de peces de las cuales se alimenta, estas especies podrían presentar
valores isotópicos similares probablemente por sus hábitos tróficos parecidos (por
ejemplo pelágicos, bentónicos, etc.,), otorgándole a la vaquita valores isotópicos con
baja variabilidad.
Este punto es respaldado por los resultados del modelo de mezcla (SIAR;
Figura 21), que sugiere a Mugil cephalus (~29.2%; Tabla 13) como presa principal.
Es importante mencionar, que aunque esta especie no ha sido reportada como presa
importante de la vaquita, los resultados arrojados por el modelo y el hecho de que M.
cephalus se alimente de detritus y algas en el fondo, esto parece indicar la
preferencia de la vaquita sobre recursos bentónicos. Los resultados del modelo de
mezcla indican además, que este tipo de especies predominan en fondos suaves
(www.fishbase.org), reflejando de esta manera la influencia que tienen sus hábitos
alimentarios en la selección de hábitat por la especie. El resto de las especies que
alimentaron el modelo de mezcla contribuyeron con menos del 8%.
Los resultados encontrados en el presente estudio sobre los hábitos
alimentarios de la vaquita son relevantes ya que si bien estudios previos basados en
el análisis de estómagos han arrojado datos importantes, también se han visto
limitados por el número de muestra y por el grado de digestión de los especímenes,
el cual ha sido muy avanzado en varias ocasiones impidiendo la obtención de
resultados confiables (Findley y Nava, 1994; Pérez-Cortés, 1996).

98
11. CONCLUSIONES GENERALES
A partir de diferentes variables ambientales se definieron tres regiones
bien delimitadas dentro del AGC: Oeste, Este y Sur, de las cuales el área donde se
han registrado la mayoría de los avistamientos de P. sinus en la última década es la
región Oeste. Esta región se caracteriza por presentar un tipo de suelo con más del
60% de limo-arcillas, la mayor turbidez de las tres regiones encontradas (15.2 y 16.4
mg/L, para junio y marzo, respectivamente), así como la menor profundidad
promedio (~26 m).
A partir de las características que predominan en la región de mayores
avistamientos de la especie se pudo definir un hábitat potencial para la misma con
una extensión de 1794 km2, que es al menos 530 km2 mayor al área de refugio de la
vaquita (1264 km2).
De acuerdo a los valores isotópicos de carbono y nitrógeno en plancton
se encontró un gradiente, con valores enriquecidos en la parte noroeste (donde se
presentan los mayores avistamientos de vaquita), así como la entrada de valores
empobrecidos de la región sur, probablemente influenciados por las aguas
provenientes de la región norte del Golfo de California.
La región Oeste presenta un claro enriquecimiento de δ15N con respecto
a la región Este en sedimento y plancton, el cual se refleja en toda la trama trófica,
permitiendo delimitar el área de alimentación de la vaquita y los organismos
analizados en una zona bien delimitada en la región Oeste.
El valor de δ13C (-12.1‰±0.7‰) encontrado en la vaquita marina la ubica
como una especie costera y con un importante aporte bentónico a su dieta.

99
Considerando los valores de δ15N en distintos organismos del hábitat de
la vaquita, se encontraron 4 niveles tróficos y un fraccionamiento isotópico de 2.8‰
entre esos niveles, dentro de los cuales, la vaquita ocupo un NT de 4.0.
A partir de los resultados obtenidos del análisis Convex-hull, la vaquita
marina presentó una varianza de 0.5‰ y 0.6‰ para δ13C y δ15N, respectivamente,
considerados para poblaciones especialistas.
De acuerdo al modelo de mezcla SIAR y basados en un factor de
discriminación isotópica de 2.04 (Borell et al. 2012), la especie detritívora Mugil
cephalus presentó la mayor contribución a la dieta de la vaquita marina.

100
12. BIBLIOGRAFÌA
http://oceancolor.sfc.nasa.gov
http://predmar.cicese.mx
http://www.fishbase.org
http://www.geologynet.com/programs/ternplot.xls
http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears
http://www.nhc.noaa.gov/gccalc.shtml
Abend, A. y Smith, T. 1997. Differences in stable isotope ratios of δ13C and δ15N
between long-finned pilot whales (Globicephala melas) and their primary prey
in the western north Atlantic. Journal of Marine Science, 54: 500-503.
Achituv, Y., Brickner, I y J. Erez. 1997. Stable carbon isotope ratios in Red sea
barnacles (Cirripedia) as an indicator of their food source. Marine Biology. 130:
243-247.
Aguíñiga, S. 1999. Geoquímica de la Cuenca Estuarina del Río Colorado: δ13C, δ15N
y biomarcadores lipídicos en sedimentos superficiales. Tesis de Doctorado.
Instituto de Investigaciones Oceanológicas. Universidad Autónoma de Baja
California. Ensenada, B. C.
Alvarez-Borrego, S. 1983. Gulf of California. En: Estuaries and enclosed seas,
Ketchurn, H (Ed). Eelsevier, New York. 427-449 pp.
Álvarez-Borrego, S., Flores-Báez, B. y Galindo Bect. 1975. Hidrología del Alto Golfo
de California-II. Condiciones durante Invierno, Primavera y Verano. Ciencias
Marinas. 2: 21-36.
Alvarez-Borrego, S. 2003. Physical and biological linkages between the Upper and
Lower Colorado Delta. In: Rapport DJ, Lasley WL, Rolston DE, Nielsen NO,
Qualset CO, Damania AB, editors. Managing for healthy ecosystems. Boca
Raton (FL): Lewis, A CRC Press Company; 1077–1089 p.
Álvarez, L. y Jones, S. 2002. Factors influencing suspended sediment flux in the
upper Gulf of California, Estuarine, Coastal and Shelf Science (2002), 54, 747-
759.

101
Álvarez Sánchez, L G. y S. E. Jones. 2004. Short-term observations of suspended
particulate matter in a macro-tidal inverse estuary: the Upper Gulf of California.
Journal of Coastal Research 20(3): 645-654
Alvarez LG, Suárez-Vidal F, Mendoza-Borunda R, Escobar- González M. 2009.
Bathymetry and active geological structures in the Upper Gulf of California.
Boletin de la Sociedad Geológica Mexicana. 61(1): 129–141.
Ambrose, SH. 1990. Preparation and characterízation of bone and tooth collagen for
stable carbon and nitrogen isotope analysls. J Archaeol Sci. 17:430-451
Argote, M. L., A. Amador., M. F. Lavín y J. R. Hunter. 1995. Tidal dissipation and
stratification in the Gulf of California. Journal of Geophysical Research 100: 16103–16118.
Aurioles-Gamboa, D., Newsome, S., Salazar-Pico, S., y Koch, P.2009. Stable Isotope
Differences Between Sea Lions (Zalophus) from the Gulf of California and
Galapagos Island. Journal Mammalogy. 90 (6): 1410-1420.
Aurioles-Gamboa, D., Rodríguez-Pérez, M., Sánchez-Velásco, L., Lavín, M. 2013.
Habitat, trophic level, and residence of marine mammals in the Gulf of
California assessed by stable isotope analysis. Marine Ecology Progress
Series.
Ballance, L. 2006. Habitat use patterns and ranges of the bottlenose Dolphin in the
Gulf of California, Mexico. Marine Mammal Science. 8: 262–274
Barlow, J. 1986. Factors affecting the recovery of Phocoena sinus, the vaquita or Gulf
of California harbor porpoise. Administrative Report. E.U. National Marine
Fisheries Service. La Jolla, California. 19 p.
Barlow, J., Fleischer, K., Forney y Maravilla, O. 1993. An experimental aerial survey
for vaquita (Phocoena sinus) in the northern Gulf of California, Mexico. Marine
Mammal Science 9:89-94.
Barlow, J., Gerrodette, T y Silber, G. 1997. First estimates of vaquita abundance.
Marine Mammal Science 13:44-58.
Baumgartner MF, Mullin KD, Nelson LM y Leming TD. 2001. Cetacean habitats in
the northern Gulf of Mexico. Fish. Bull. 99:219–239.

102
Bearhop, S., Adams, C.E., Waldron, S., Fuller, R.A. y MacLeod, H. 2004 Determining
trophic niche width: a novel approach using stable isotope analysis. Journal of
Animal Ecology, 73, 1007–1012.
Borell, A., Abad-Oliva, N., Gómez-Campos, E., Giménez, J y Aguilar, A. 2012.
Discrimination of stable isotopes in fin whale tissues and application to diet
assessment in cetaceans. Rapid Communication in Mass Spectrometry. 26:
1596–1602.
Bouillon, S., Raman, A., Dauby, P y Dehairs, F. 2002. Carbon and Nitrogen Stable
Isotope Ratios of Subtidal Benthic Invertebrates in an Estuarine Mangrove
Ecosystem (Andhra Pradesh, India). Estuarine, Coastal and Shelf Science. 54:
901–913
Bray, J. R. and Curtis, J. T. (1957) An ordination of the upland forest communities of
Southern Wisconsin. Ecol. Monogr., 27, 325–349.
Brownell, R. 1983. Distribution of the vaquita Phocoena sinus in Mexican waters.
Marine Mammal Science, 2(4): 299-305.
Brownell, L., Findley, T., Vidal, O., Robles, A y Manzanilla S. 1987. External
morphology and pigmentation of the vaquita, Phocoena sinus (Cetacea:
Mammalia). Marine Mammal Science 3:22-30.
Brownell, R. 1986. Distribution of the vaquita Phocoena sinus in Mexican waters.
Marine Mammal Science. 2(4): 299-305.
Carriquiry, J. D. y A. Sánchez. 1999. Sedimentation in the Colorado River delta and
Upper Gulf of California after a century of discharge loss. Marine Geology 158: 125–145.
Carriquiry JD, Sanchez A, Camacho-Ibar V (2001) Sedimentation in the Northern Gulf
of California after the elimination of the Colorado River discharge. Sedim Geol
144: 37-62.
Ceballos, G. y J.H. Brown. 1995. Global patterns of mammalian diversity, endemicity,
and endangerment. Conservation Biology, 9:559-568.
Chanton, J Y Lewis, G. 1999. Plankton and Dissolved Inorganic Carbon Isotopic
Composition in a River-Dominated Estuary: Apalachicola Bay, Florida.
Estuarios. 22 (3): 575-583.

103
Christensen V y D. Pauly. 1993. Trophic models of aquatic ecosystems. ICLARM
Conf Proc. 26:390. Clarke, K.R., R.N. Gorley. 2006. PRIMER v6: User
Manual/Tutorial. PRIMER-E. Plymouth.
Cifuentes, L. A., Sharp, J. H., and Fogel. M. L. 1988, Stable carbon and nitrogen
isotope biogeochemistry in the Delaware estuary, Limnol. Oceanogr. 33:1102-
1115.
Clarke, K.R., P. J. Somerfield., R. N. Gorley. 2008. Testing of null hypotheses in
exploratory community analyses: similarity profiles and biota-environment
linkage. Journal of Experimental Marine Biology and Ecology. 366: 56–69.
Croll, D., Tershy, B., Hewitt, R., Demer, D., Fiedler, P., Smith, S., Armstrong, W.,
Popp, J., Kiekhefer, T., López, V., Urban, J y Gendron, D. 1998. An integrated
approach to the foraging ecology of marine birds and mammals. Deep-Sea
Reserch. 45: 1353-1371.
Cupul Magaña, A. L. 1994. Flujos de sedimentos en suspensión y de nutrientes en la
cuenca estuarina del Río Colorado. Tesis de Maestría. Facultad de Ciencias
Marinas UABC., Ensenada, B.C., México. 117 p.
Davis, R., Fargion, G., May, N., Leming, T., Baumgartner, M., Evans, W., Hansen, L.,
Mullin, K. 1998. Physical Habitat of Cetacean Along the Continental Slope in
the North-Central and Western Gulf of Mexico. Marine Mammal Science. 14(3):
490-507.
D'Agrosa, C., O. Vidal y W. C. Graham. 1995. Mortality of the vaquita (Phocoena
sinus) in gillnet fisheries during 1993-94. Reports of the International Whaling
Commission Special Issue 16:283-294.
D'Agrosa, C., C. E. Lennert-Cody y O. Vidal. 2000. Vaquita bycatch in Mexico's
artisanal gillnet fisheries: driving a small population to extinction. Conservation
Biology 14:1110-1119.
Delgado-González, O., Ocampo-Torres, F., Larios-Castillo, S. 1994. Las brisas
durante algunos meses de primavera y verano durante en el noroeste del
Golfo de California. Ciencias Marinas. 20 (3): 421-440.
DeNiro, M. y Epstein, S. 1978. Influence of diet on the distribution of carbon isotopes
in animals. Geochimica et Cosmochimica Acta, 42: 495- 506.

104
Díaz‐Gamboa, R. 2003. Diferenciación entre tursiones, Tursiops truncatus, costeros y
oceánicos en el Golfo de California por medio de isótopos estables de carbono
y nitrógeno. Tesis de Maestría CICIMAR‐IPN. 62 p.
DOF 1993. Decreto por el que se declara área natural protegida con el carácter de
Reserva de la Biosfera, la región conocida como Alto Golfo de California y
Delta del Río Colorado, ubicada en aguas del Golfo de California y los
municipios de Mexicali, Baja California. Diario Oficial de la Federación, jueves
10 de junio de 1993,México, D.F. 24-28 p.
DOF. 2005. Programa de protección de la vaquita dentro de área de Refugio ubicada
en la porción occidental del Alto Golfo de California. Diario Oficial de la
Federación, Septiembre del 2005.
Doniol-Valcroze, T., Berteaux, D., Larouche, P. y Sears, R. 2007. Influence of thermal
fronts on habitat selection by four rorqual whale species in the Gulf of St.
Lawrence. Marine Ecology Progress Series. 335: 207-216.
Dugdale R. C. y J. J. Goering (1967). Uptake of new and regenerated forms of
nitrogen in primary productivity. Limnology and Oceanography, 12, 196-206.
Durazo-Arvizu, R. 1989 Frentes térmicos de verano en el Alto Golfo de California.
Tesis de Maestría. CICESE. Ensenada, B.C. México.
Elorriaga-Verplancken, F., y Aurioles-Gamboa, D. 2009. Trace metal concentrations
in the hair of Zalophus californianus pups and their relation to feeding habits.
Biological Trace Element Research 126:148–164.
Enríquez-Andrade, R., G. Anaya-Reyna, J.C. Barrera-Guevara, M.A, Carvajal-
Moreno, M.E. Martínez-Delgado, J. Vaca-Rodríguez & C. Valdés-Casillas.
2005. An analysis of critical areas for biodiversity conservation in the Gulf of
California Region. Ocean & Coastal Management. 48: 31-50
Etnoyer, P., Canny, D., Mate, B., Morgan, L.E., 2004. Persistent Pelagic habitat in the
Baja California to Bering Sea (B2B) Ecoregion. Oceanography 17 (1), 90–101.
Farfán, C. y S. Alvarez-Borrego. 1992. Zooplankton Biomass of the Northemmost
Gulf of California. Ciencias Marinas. 18 (3):17-36

105
Findley, L., J. Nava., y J. Torre. 1994. Food habits of Phocoena sinus (Cetacea:
Phocoeanidae). Resúmenes XI Conferencia Bienal Sobre Biología de Los
Mamíferos Marinos. E.U. 37 p.
Fitch, J y Brownell, R. 1986. Fish otoliths in cetacean stomachs and their importance
in interpreting feeding habits. Journal of the Fisheries Research Board of
Canada 25:2561-2574.
Fogel, M. L. y Cifuentes, L. A. 1993. Isotope fractionation during primary production.
In: Organic Geochemistry. Engel, M. H. y Macko, S. A (eds). Plenum Press,
New York, 73–98.
France, R. L. 1995. Carbon-13 enrichment in benthic compared to planktonic algae :
foodweb implication. Marine Ecology Progress Series. 124: 307-312.
Fry, B. 1988. Food web structure on Georges Bank from stable C, N and S isotopic
composition. Limnology Oceangraphy, 33: 1182-1190.
Fry, B. y Wainright, SC. 1991. Diatom sources of %-rich carbon in marine food webs.
Marine Ecology Progress Series 76: 149-157.
Gallo-Reynoso, J. P. 1998. La vaquita y su hábitat crítico en el Alto Golfo de
California. Gaceta Ecológica 47-29-44. Instituto Nacional de Ecología.
SEMARNAP. 44 p.
García-Rodríguez, F. J. y Aurioles, G. D. 2004. Spatial and temporal variation in the
diet of the California sea lion (Zalophus californianus) in the Gulf of California,
Mexico. Fishery Bulletin. 102 (1): 47-62.
Gaskin D. 1983. The ecology of whales and dolphins. Heineman. Inglaterra. 459p.
Gaskin, E., Smith, G., Watson, A., Yasui, W y Yurick, D. 1984. Reproduction in
porpoises (Phocoenidae): implications for managment. Reports of the
International Whaling Commission Special Issue 6:135-148.
Gerrodette,T., Fleischer, L. A., Pérez-Cortés, H., y Villa-Ramírez, B. (1995).
Distribution of the Vaquita, Phocoena sinus, Based on Sightings from
Systematic Surveys. Rep. Int. Whal. Commn. Special Issue. 16: 273-281.
Gerrodette, T., B. Taylor., R. Swift., S. Rankin., A. Jaramillo-Legorreta y L. Rojas-
Bracho. 2011. A combined visual and acoustic estimate of 2008 abundance,

106
and change in abundance since 1997, for the vaquita, Phocoena sinus. Marine
Mammal Science
Goericke R, Fry B (1994) Variations of marine d13C with latitude, temperature, and
dissolved CO2 in the world ocean. Global Biogeochem Cycles 8:85‐90
Graham, B. S., P. L. Koch, S. D. Newsome, K.W. McMahon y D. Aurioles. 2010.
Using isoscapes to trace the movements and foraging behavior of top
predators in oceanic ecosystems. In J. B. West, G. J. Bowen, T. E. Dawson
and K. P. Tu, (eds). Isoscapes: Understanding movement, pattern, and
process on earth through isotope mapping. Springer, Berlin, Germany.
Griffin y Valiela. 2001. δ15N isótope studies of life history and trophic position of
Fundulus heterolictus and Menidia menidia. Marine Ecology Progress Series.
214: 299-305.
Hastie, G., Rosen, D. y Trites, A. 2006. The influence of depth on a breath-hold diver:
Predicting the diving metabolism of Steller sea lions (Eumetopias jubatus).
Journal of Experimental Marine Biology and Ecology. 336 (2): 163-170.
Hernández-Ayón, M., Galindo-Bect, S., Flores- Báez, B y Álvarez-Borrego, S. 1993.
Nutrient concentrations are high in the turbid waters of the Colorado River
Delta. Estuarine Coastal and Shelf Science 37: 593-602.
Hobson KA, Schell DM, Renouf D, Noseworthy E. 1996. Stable carbon and nitrogen
isotopic fractionation between diet and tissues of captive seals: Implications for
dietary reconstructions involving marine mammals. Canadian Journal of
Fisheries & Aquatic Sciences 53:528‐533
Hobson, K., Sease, J. y Piatt, J. 1997. Investigating trophic relationship of pinnipeds
in Alaska and Washington using stable isotope ratios of nitrogen and carbon.
Marine Mammal Science, 13: 114-132.
Hobson, K. y Welch, H. 1992. Determination of trophic relationship within a high Artic
marine food web using 13C and 15N analysis. Marine Ecology Progress Series,
84: 9-18.
Hohn, A., Read, A., Fernandez, S., Vidal, O. y Findley, L. 1996.Life history of the
vaquita, Phocoena sinus (Phocoenidae, Cetacea). Journal Zoological London,
239: 235-251.

107
IUCN. 2008. Red list of threatened species: A global species assessment. Cabridge.
1-185 p. Eds. Baillie, E., Hilton-Taylor, C y Stuart, S.
Jaeger, A., Blanchard, P., Richard, P y Cherel, Y. 2009. Using Carbon And Nitrogen
Isotopic Values Of Body Feathers To Infer Inter‐ And Intra‐Individual Variations
Of Seabird Feeding Ecology During Moult. Marine Biology. 156: 1233–1240
Jaquet N y Gendron D. 2002. Distribution and relative abundance of sperm whales in
relation to key environmental features, squid landings and the distribution of
other cetacean species in the Gulf of California, Mexico. Marine Biology. 141:
591–601.
Jaquet, N., y Whitehead, H. 1996. Scale-dependent correlation of sperm whale
distribution with environmental features and productivity in the South Pacific.
Marine Ecology Progress Series. 135: 1-9.
Jaramillo-Legorreta, A., Rojas-Bracho, L y Gerrodette, T. 1999. A new abundance
estimate for vaquitas: first step for recovery. Marine Mammal Science 15:957-
973.
Jaramillo-Legorreta, A.M. 2008. Estatus actual de una especie en peligro de
extinción, la vaquita (Phocoena sinus): Una aproximación poblacional con
métodos acústicos y bayesianos. Doctoral Dissertation. Facultad de Ciencias
Marinas. Universidad Autónoma de Baja California. Ensenada, Baja California,
México. 115 pp.
Kelly, J. 2000. Stable Isotopes of carbon and nitrogen in the study of avian and
mammalian trophic ecology. Canadian Journal of Zoology, 78: 1-27.
Kramer, D., M. J. Kalin, E. G. Stevens., J. R. Thrailkill, y J. R. Zweifel. 1972.
Collecting and processing data on fish and larvae in the California Current
region. NOAA Tech. Rep. NMFS CIRC-370:l-38.
Kurle CM (2002) Stable-isotope ratios of blood components from captive northern fur
seals (Callorhinus ursinus) and their diet: applications for studying the foraging
ecology of wild otariids. Can J Zool 80:902–909
Kurle, C y Worthy, G. 2001. Stable Isotope Assessment Of Temporal And Geographic
Differences In Feeding Ecology Of Northern Fur Seals (Callorhinus ursinus)
And Their Prey. Oecologia. 126: 254–265

108
Lavín, M., Alvarado, R., Alvarez, L., Godínez, V., Washburn, L. 2004. Mixing, Gravity
Currents And Circulation In The Upper Gulf Of California. PECS 2004 – Mérida
- México
Lavín, M. F., E. Beier y A. Badán. 1997. Estructura hidrográfica y circulación del
Golfo de California: Escalas estacional e interanual. En: M. F. Lavín Ed.
Contribuciones a la Oceanografía Física en México. Unión de Geofísica
Mexicana. Monografía 3, 41-171 p.
Lavín, M., Gaxiola-Castro, G., Robles, J. y Richter, K. 1995. Winter Water Masses
and Nutrients in the Northern Gulf of California. Jounal of Gophysical
Research. 100: 8587-8605.
Lavín, M., Godínez, V., y Alvarez, L. 1998. Inverse-estuarine Features of the Upper
Gulf of California. Estuarine, Coastal and Shelf Science. 47: 769–795.
Lavín, M. y Organista, S. 1988. Surface heat flux in the Northern Gulf of California.
Journal of Gophysical Research. 93: 14033-14038.
Lavín, M. F. y S. Sánchez. 1999. On how the Colorado river affected the hydrography
of the upper Gulf of California. Continental Shelf Research 19, 1545–1560.
Lessage, V; Hamill, M. O y K. Kovacs. 2001. Marine mammals and the community
structure of the estuary and Gulf of St. Lawrence, Canada: evidence from
stable isotope analysis. Marine Ecology Progress Series. 210: 203- 221.
López-Martín1, J., Jiménez, J y Ruiz-Olmo, J. 1998. Caracterización Y Uso Del
Hábitat De La Nutria Lutra Lutra (Linné, 1758) En Un Río De Carácter
Mediterráneo. Galemys. 10: 175-190.
Lusseau, D., Willians, R., Wilson, B., Grellier, K., Hammond, P.S. y P.M.Thompson.
2004. Parallel influence of climate on the behaviour of Pacific killer whales and
Atlantic bottlenose dolphins. Ecology Letters 7: 1068- 1076.
Macko, S. A. and M. L. F. Estep. 1984. Microbial alteration of stable nitrogen and
carbon isotopic compositions of organic matter. Organic Geochemisty. 6:
787‐790.
Marshall D (1996) TERNPLOT: An excel spreadsheet for ternary diagrams. Comp
Geo 22(6): 697-699.

109
Michener RH, Schell DM. 1996. Stable isotope ratios as tracers in marine aquatic
food webs, Vol. Blackwell, Boston
Millán Núñez, R., E. Santamaría del Angel, R. Cajal Medarano y O. A. Barocio León.
1999. El delta del Río Colorado: Un ecosistema con alta productividad
primaria. Ciencias Marinas 25(4):509-540 p.
Miranda, F., S. Reyes Coca y J. García López. 1990. Climatología de la región
noroeste de México. Parte I: Precipitación. Reporte Técnico. EBA No.3
CICESE, Ensenada, B.C., México, 160 p.
Mittermeier, R. A. y C. Goettsch de M. 1992. La importancia de la diversidad
biológica de México. México ante los retos de la biodiversidad. Sarukhán, J y
Dirzo, R (eds.). Comisión Nacional para el Conocimiento y Uso de la
Biodiversidad. México, D. F. 63-73 pp.
Minegawa, M y Wada, E. 1984. Stepwise enrichment of δ15N along food chains:
further evidence and the relation between δ15N and animal age. Geochimica et
Cosmochimica Acta, 48: 1135- 1140.
Montoya JP (2007) Natural abundance of 15N in marine planktonic ecosystems. En:
Michener R y K. Lajtha (eds) Stable isotopes in ecology and environmental
science, 2nd ed. Blackwell, Malden, MA, pp 176–201
Newsome S, Clementz M, Koch P (2010) Using stable isotope biogeochemistry to
study marine mammal ecology. Mar Mamm Sci 26(3): 509–572
Newsome, S., Koch, P., Etnier, M y Aurioles-Gamboa, D. 2006. Using carbon and
nitrogen isotope values to investigate maternal strategies in northeast Pacific
Otariids. Marine Mammal Science, 22(3): 556-572.
Newsome, S., Martínez del Rio, C., Phillips, D.L. y Bearhop, S. 2007. A Niche for
Isotopic Ecology. Frontiers in Ecology and the Environment. 5: 429– 436.
Niño-Torres C., Gallo-Reynoso, J. P., Galván-Magaña, F., Escobar-Briones, E. y
Macko, S. 2006. Isotopic analysis of δ13C, δ15N y δ34S “a feeding tale” in teeth
of the longbeaked common dolphin, Delphinus capensis. Marine Mammal
Science.

110
Páez-Rosas, D., Aurioles-Gamboa, D., Alava, J., Palacios, D. 2012. Stable isotopes
indicate differing foraging strategies in two sympatric otariids of the Galapagos
Islands.
Palka, D. 1995. Abundance estimate of Gulf of Maine harbor porpoise. The Biology of
Phocoenidae. Report of International Whaling Commission Special Issue 16.
28-50.
Panigada, S., Zanardelli, M., MacKenzie, M., Donovan, C., Mélin, F., Hammond, P.
2008. Modelling habitat preferences for fin whales and striped dolphins in the
Pelagos Sanctuary (Western Mediterranean Sea) with physiographic and
remote sensing variables. Remote Sensing of Environment. 112: 3400–3412.
Parnell A, Inger R, Bearhop S, Jackson AL. 2008. SIAR: Stable isotope analysis in R.
http://cran.rproject. org/web/packages/siar/index.html
Pérez-Cortés, H. 1996. Contribución al conocimiento de la biología de la vaquita
Phocoena sinus. Tesis de Maestría. Instituto de Ciencias del Mar y Limnología.
UNAM. 69 p.
Phillips, D.L. y Koch, P.L. 2002. Incorporating Concentration Dependence In Stable
Isotope Mixing Models. Oecologia. 130: 114–125.
Pinnegar, J., Jennings, S., O’brien, C y Polunin, V. 2002. Long-term changes in the
trophic level of the Celtic Sea fish community and fish market price distribution.
Journal of Applied Ecology 39:377–390.
Popp BN, Laws EA, Bidigare RR, Dore JE, Hanson KL, Wakeham SG (1998) Effect
of phytoplankton cell geometry on carbon isotopic fractionation. Geochim
Cosmochim Acta 62:69–77
Porras-Peters, H., Aurioles-Gamboa D., Cruz-Escalona V., Koch P. 2008. Trophic
level and overlap of sea lions (Zalophus californianus) in the Gulf of California,
Mexico. Marine Mammal Science. 24(3): 554-576.
Post, D. 2002. Using stable isotopes to estimate trophic position: models, methods,
and assumptions. Ecology. 83 (3): 703-718.
Post, D. M., Pace, M. y Hairston, N. 2000. Ecosystem Size Determines Food‐Chain
length In Lakes. Nature. 405: 1047–1049.
Rau, G., Ainley, D., Bengtson, J., Torres, J. y Hopkins, T. 1992. 15N/14N and 13C/12C

111
in Weddell seabirds, seals and fish: implications for diet and trophic structure.
Marine Ecology Progress Series, 84: 1-8.
Read, A. 1999. Porpoises World Life Library. Voyageur Press. Vancouver, Canada.
Ridgway S. H. y R. Harrison. 1999. Handbook of marine mammals. The second book
of marine mammals. Vol. 6. Academic Press. 473p.
Ridoux, V. 1994. The diets and dietary segregation of seabirds at the subantartic
Crozet Island. Mar. Ornithol. 22: 1-192
Ríofrío,- LazoM., Aurioles-Gamboa, D y Le Boeuf, BJ. 2012. Ontogenetic changes in
feeding habits of the northern elephant seals revealed by δ15N and δ13C
analysis of growth layers in teeth. Marine Ecology Progress Series. 450: 229–
241
Roden, G.I., Oceanographic aspects of the Gulf of California, Marine Geology of the
Gulf of California: A Symposium, Mem. Am. Assoc. Pet. Geol., 3, 30-58, 1964.
Rodríguez -Pérez MY, Escobar-Briones EG, Gallo-Reynoso JP, Morales-Puente P,
Cienfuegos-Alvarado, E (2010). Composición elemental e isotópica estable de
δ13C y δ15N en Phocoena sinu. In: 60 años de la colección nacional de
mamíferos del instituto de biología, UNAM: Aportaciones al conocimiento y
conservación de los mamíferos mexicanos. Instituto de Biología, UNAM. 147-
154p.
Rojas-Bracho, L. y A.M. Jaramillo-Legorreta. 2002. Vaquita Phocoena sinus. pp.
1277-1280 In: Perrin, W.F., B. Würsig y J.G.M.Thewissen (Eds.) Encyclopedia
of Marine Mammals. Academic Press, San Diego, California, USA. 1414 pp.
Rojas-Bracho, L.,. Reeves, R y Jaramillo-Legorreta, A. 2006. Conservation of the
vaquita, Phocoena sinus. Mammal Rev. 36 ( 3): 179–216. Printed in
Singapore.
Rojas-Bracho, L y Taylor, B. 1999. Risk factors affecting the vaquita (Phocoena
sinus). Marine Mammal Science 15:974-989.
Rubenstein, D. R. y Hobson K. 2004.From birds to buterflies: Animal movement
paterns and stable isotopes. Trends in Ecology and Evolution 19:256–263.

112
Salvadeo, Ch. 2008. Análisis de la Comunidad de Odontocetos y la Relación con su
Ambiente, en el Extremo Suroccidental del Golfo de California, México (2003-
2006). Tesis de Maestria. La Paz, BCS. Mèxico.
Sánchez Velasco, L., M. F. Lavin Peregrina, S. P. A. Jiménez Rosenberg, J. M.
Montes Aréchiga y P. Turk Boyer. 2012. Larval fish habitats and hydrography
in the biosphere reserve of the Upper Gulf of California (june 2008).
Continental Shelf Research 33(2012): 89-99 p.
Santamaría del Ángel, E., S. Álvarez-Borrego y E. Müller-Krager. 1994. Gulf of
California biogeographic regions based on coastal zone color scanner imagery.
Journal of Geophysical Research 99: 7411-7421.
Schoeninger, M. y DeNiro, M. 1984. Nitrogen and carbon isotopic composition of
bone collagen from marine and terrestrial animals. Geochimica et
Cosmochimica Acta, 48: 625- 639.
Shanks, A. L. 1983. Surface slicks associated with tidally forced internal waves may
transport pelagic larvae of benthic invertebrates and fishes shoreward. Marine
Ecology Progress Ser. 13: 311-315.
Silber, G.K.1988. Recent Sightings of the Gulf of California Harbor Porpoise,
Phocoena sinus. J. Mamm. 69 (2): 430-433.
Silber, G. K 1988. Observations on the behavior and ventilation cycles of the vaquita,
Phocoena sinus. Marine Mammal Science. 4 (1): 62-67.
Silber, G. 1990. Occurrence and distribution of the vaquita, Phocoena sinus, in the
northern Gulf of California. Fishery Bulletin, 88: 339-346.
Silber, G y Norris, K. 1991. Geographic and seasonal distribution of vaquita
Phocoena sinus. Anales del Instituto de Biología. UNAM. Serie Zoología,
62(2): 263-268.
Silber, G. K., Newcomer, M., Silber, P., Pèrez-Cortès, H y Ellis. G. 1994. Cetaceans
of the northern Gulf of California: distribution, occurrence, and relative
abundance. Marine Mammal Science 10:283-298.
Slotten, E. 2012. Size matters – marine protected areas can solve bycatch problems
for the New Zealand dolphin and Vaquita, if they are large enough. Report of
International Whaling

113
Springer, A. M; Piatt, J. F; Shuntov, V. P; Van Vliet, G. B; Vladimirov, V. L; Kuzin, A.
E; Perlov, A. S. 1999. Marine Birds and Mammals of the Pacific Subarctic
Gyres. Progress in Oceanography ISSN 0079-6611. 43 (2): 443-487.
Steele, K y Daniel, R. 1978. Stable isotope analysis of human remains: a further
complication to the use of variation in the natural abundance of δ15N for tracer
studies. Journal of Agricultural Sciences, 90: 7‐9.
Stockin, K.A., G.J. Pierce, V. Binedell, N. Wiseman & M.B. Orams. 2008. Factors
Affecting the Occurrence and Demographics of Common Dolphins (Delphinus
sp.) In the Hauraki Gulf, New Zealand. Aquatic Mammals.34(2): 200-211.
Takai N, Onaka S., Ikeda, Y., Yatsu, A., Kidokoro, H y Sakamoto, W. 2000.
Geographical Variations in Carbon and Nitrogen Stable Isotope Ratios in
Squid. Journal Marine Biology. 80: 675–684.
Taylor, D.W. 1948. Fundamentals of soil mechanics. New York, J. Wiley. 700p.
Ter Braak, C.J.F., 1986. Canonical correspondence analysis: a new eigenvector
technique for multivariate direct gradient analysis. Ecology 67 (5), 60–71.
Tershy, B.R. 1992. Body Size, Diet, Habitat Use, and Social Behavior of
Balaenoptera Whales in the Gulf of California. Journal of Mammalogy. 73 (3):
477-486.
Thompson R.W. 1968. Tidal flat sedimentation on the Colorado River delta,
Northwestern Gulf of California. Bulletin of the Geological Society American
Memoirs. 107: 133
Thompson, R. W. 1979. Tidal-flat sediments of the Colorado River delta,
northwestern Gulf of California. In: Ginsburg, R.N. (Ed.). Tidal Deposits.
Springer, New York, pp. 57–65.
Tieszen, L., Boutton, W., Tesdahl, G y Slade, N. 1983. Fractionation and Turnover of
Stable Carbon Isotopes in Animal Tissues: Implications for δ13C Analysis of
Diet. Ocologia. 57: 32–37.
Toperoff, A. 2002. Examination of diet of harbor porpoise (Phocoena phocoena) from
central California using stomach content and stable isotope analysis from
multiple tissues. San Jose State University. Tesis de Maestria. 1-120p.

114
Tynan, C., Ainley, D., Barth, J., Cowles, T., Pierce, S y Spear, L. 2005. Cetacean
distributions relative to ocean processes in the northern California Current
System. Deep-Sea Research II 52:145–167.
Valdez-Muñoz, C., Aragón-Noriega, EA., Ortega-Rubio, A., Salinas-Zavala, CA.,
Arreola-Lizárraga, JA., Hernández-Vázquez, SY., Beltrán-Morales, LF. 2010.
Distribución y abundancia de juveniles de totoaba, Totoaba macdonaldi, y la
salinidad del hábitat de crianza. Interciencia. vol. 35.
Vidal O, Aguayo A, Findley L, Robles A, Bourillón L, Vomend I, Turk P, Gárate K,
Maroñas I, Rosas J. 1987. Avistamientos de mamíferos marinos durante el
crucero “Guaymas I” en la región superior del Golfo de California, primavera
de 1984. X Reunión Internacional para el Estudio de los Mamíferos Marinos.
Marzo de 1987. México, DF.
Vidal, O., Brownell, R y Findlay, L. 1999. Vaquita, Phocoena sinus (Norris and
mcfarland, 1958). Handbook of Marine Mammals The second book of dolphins
and the porpoises. S. H. Ridgway y R. Harrison (eds). Academic Press. Vol
(6):Pp. 357-378.
Villa-Ramírez, B., Delgado-Estrella, A. Y Peralta-Pérez, M. 1995. Aportaciones
recientes al conocimiento de la biología y distribución de Phocoena sinus
(Cetácea: Phocoenidae). Revista de la Sociedad Mexicana de Historia Natural,
46: 163-172.
Walker, J. Y Macko, S. 1999. Dietary studies of marine mammals using stable C y N
isotopic ratios of teeth. Marine Mammal Science, 15(2):314-334.
Walker, J., Potter CH. Y Macko, S. 1999. The diets of modern and historic bottlenose
dolphin populations reflected through stable isotopes. Marine Mammal
Science, 15(2): 314-334.
Watts, P. y D.E. Gaskin. 1985. Habitat index analysis of the harbour porpoise
(Phocoena phocoena) in the southern coastal Bay of Fundy, Canada. Journal
of. Mammalogy 66(4):733-744.
Wells S., Würsig G., Norris S. 1981. Un reconocimiento de los mamíferos marinos en
el alto Golfo de California, México. VI Reunión Internacional para el Estudio de

115
los Mamíferos Marinos de la Península de Baja California. Febrero de 1981.
La Paz, BCS.
Wolf, B., Carleton, S y Martínez del Río, C. 2009. Ten Years of Experimental Isotopic
Ecology. Functional Ecology. 23: 17–26.
Worthy, G y Edwards, E. 1990. Morphometric and Biochemical Factors Affecting Heat
Loss in a Small Temperate Cetacean (Phocoena phocoena) and a small
tropical cetacean (Stenella attenuata). PhysiologicaI Zoology. 63(2):432-442.
Yen, P., Sydeman, W., Hyrenbach, K. 2004. Marine bird and cetacean associations
with bathymetric habitats and shallow-water topographies: implications for
trophic transfer and conservation. Journal of Marine Systems 50 (2004) 79–
99.