
“CARACTERIZACIÓN DE LOS GENES AS1 Y GlsA … · CENTRO DE INVESTIGACION Y DE ESTUDIOS AVANZADOS...
80
CENTRO DE INVESTIGACION Y DE ESTUDIOS AVANZADOS DEL INSTITUTO POLITÉCNICO NACIONAL UNIDAD IRAPUATO “CARACTERIZACIÓN DE LOS GENES AS1 Y GlsA EN Agave tequilana” TESIS QUE PRESENTA Ing. José Alfredo Guzmán López PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS EN LA ESPECIALIDAD DE BIOTECNOLOGÍA DE PLANTAS DIRECCIÓN DE TESIS: Dra. June K. Simpson Williamson Irapuato, Gto., Julio del 2010
Transcript of “CARACTERIZACIÓN DE LOS GENES AS1 Y GlsA … · CENTRO DE INVESTIGACION Y DE ESTUDIOS AVANZADOS...

CENTRO DE INVESTIGACION Y DE ESTUDIOS AVANZADOS DEL
INSTITUTO POLITÉCNICO NACIONAL
UNIDAD IRAPUATO
“CARACTERIZACIÓN DE LOS GENES AS1 Y GlsA EN Agave tequilana”
TESIS QUE PRESENTA
Ing. José Alfredo Guzmán López
PARA OBTENER EL GRADO DE
MAESTRO EN CIENCIAS
EN LA ESPECIALIDAD DE
BIOTECNOLOGÍA DE PLANTAS
DIRECCIÓN DE TESIS:
Dra. June K. Simpson Williamson
Irapuato, Gto., Julio del 2010

ii
DEDICATORIA
A Dios por darme la vida y esta gran oportunidad.
A mi esposa:
Jazmín Abraham
Porque gracias a su cariño, su amor incondicional y su perseverancia he logrado
ser una mejor persona y avanzar en mi vida profesional.
A mis padres:
Alfredo Guzmán Hernández
María Guadalupe López Guerrero
Muchas Gracias por brindarme esta oportunidad mediante su sacrificio y esfuerzo,
este logro es suyo.
A mi hermana:
Elsa Guzmán López
Gracias por su apoyo y estar siempre conmigo.
A mi familia, porque siempre he recibido su apoyo y cariño en todo momento.

iii
AGRADECIMIENTOS
Agradezco al CONACYT por el apoyo financiero otorgado para la realización de
este trabajo de investigación, número de becario 219888.
Al CINVESTAV por la oportunidad de desarrollar este trabajo en la institución.
A la doctora June Simpson, por permitirme ser parte se de su grupo de trabajo, por
sus consejos, paciencia y enseñanzas.
A la doctora Alba Jofre y al doctor Stefan de Folter por su guía, sugerencias y
revisión de este trabajo.
A mi esposa: Jazmín Abraham, por todos sus consejos, su paciencia y tenacidad
para apoyarme en la realización de esta investigación.
Al doctor Octavio Martínez y a Fernando (El Güero), al doctor Edmundo Lozoya, a
la doctora Nayelli Marsch, a Marco Leyva, Cesar Aza, Hamlet Avilés, por su ayuda
prestada.
A Aurora Verver, por su asesoría técnica en los análisis microscópicos.
A mis compañeros y amigos de laboratorio: Katia, Emi, Miriam, Silvia, Celso,
Francisco, César, Uriel, Rocío, Juan Carlos, Paco, Hugo, Mario y Mariela por su
ayuda, compañía y apoyo.
A la familia Abraham Juárez, por toda su ayuda y apoyo.
A mis compañeros y amigos de generación, gracias por los buenos ratos.
A todo el personal administrativo del CINVESTAV unidad Irapuato por su apoyo.
A todas las personas que de alguna forma hicieron posible que se llevara a cabo
este trabajo.

iv
ÍNDICE GENERAL
DEDICATORIA ......................................................................................................... ii AGRADECIMIENTOS ............................................................................................. iii ÍNDICE GENERAL .................................................................................................. iv ÍNDICE DE TABLAS ............................................................................................... vi ÍNDICE DE FIGURAS ............................................................................................ vii RESUMEN. ........................................................................................................... viii ABSTRACT ............................................................................................................ iX I. INTRODUCCIÓN ................................................................................................. 1 II. ANTECEDENTES ............................................................................................... 2 II.1. Importancia de los Agaves ............................................................................ 2 II.2. Agave tequilana Weber var. Azul .................................................................. 4 II.2.1. Clasificación taxonómica ............................................................................ 4 II.2.2. Métodos de propagación ............................................................................ 5 II.3. Producción de bulbilos en Agave .................................................................. 6 II.4. Genes expresados durante el desarrollo de bulbilos .................................... 8 II.5. ASYMMETRIC LEAVES1 (AS1) ................................................................... 9 II.6. GlsA ............................................................................................................ 11 II.7. Amplificación rápida de extremos de DNA complementario (RACE) .......... 13 II.8. Tecnología Gateway® ................................................................................. 15 III. JUSTIFICACIÓN .............................................................................................. 16 IV. HIPÓTESIS ...................................................................................................... 18 V. OBJETIVO. ....................................................................................................... 18 V.1. Objetivo general. ......................................................................................... 18 V.2. Objetivos específicos .................................................................................. 18 VI. MATERIALES Y METODOS ............................................................................ 19 VI.1. Colecta de muestras .................................................................................. 19 VI.2. Selección de genes ................................................................................... 19 VI.3. Análisis de expresión por RT-PCR ............................................................ 20 VI.4. Hibridación in situ ...................................................................................... 21 VI.4.1. Preparación del templado de DNA ......................................................... 21 VI.4.2. Síntesis y marcaje de la ribosonda ......................................................... 22 VI.4.3. Hibridación .............................................................................................. 22 VI.4.4. Observación al microscopio .................................................................... 23 VI.5. Identificación de la clona con el gen AtqAS1 completo ............................ 23 VI.6. Expresión heterologa ................................................................................. 24 VI.6.1. Recombinación en vector de salida Gateway® pB7WG2D ..................... 26 VI.6.2. Transformación de E.coli ........................................................................ 26 VI.6.3. Transformación de Agrobacterium tumefaciens ..................................... 26 VI.6.4. Transformación de Arabidopsis .............................................................. 27 VI.6.5. Selección de líneas homocigotas con una sola inserción ....................... 27 VI.7. Obtención de la secuencia completa del cDNA de AtqGlsA ...................... 28 VI.7.1. RACE 5’ .................................................................................................. 28 VI.7.2. Clonación del gen AtqGlsA ..................................................................... 29 VI.8. Análisis de expresión por RT-PCR de los genes endógenos de Arabidopsis
AS1 y KNAT1 ..................................................................................................... 30

v
VI.9. Análisis de explantes in vitro con el gen AtqAS1 ....................................... 31 VI.9.1. Establecimiento de explantes in vitro ...................................................... 31 VI.9.2. Infección de los explantes ...................................................................... 32 VII. RESULTADOS ................................................................................................ 33 VII.1. Selección de genes .................................................................................. 33 VII.2. Análisis de expresión por RT-PCR del gen AtqAS1 ................................. 33 VII.3. Ensayo de Hibridación in situ de AtqAS1 ................................................. 34 VII.4. Identificación de la clona correspondiente al cDNA completo de AtqAS1 35 VII.5. Expresión heterologa ................................................................................ 38 VII.5.1. Transformación de Agrobacterium tumefaciens con el vector pB7WG2D
conteniendo el gen AtqAS1 ................................................................................ 38 VII.5.2. Expresión constitutiva del gen AtqAS1 en Arabidopsis thaliana ............ 39 VII.5.3. Complementación de la mutante as1 de A. thaliana .............................. 40 VII.5.3.1 Descripción de la mutante ................................................................... 40 VII.5.3.2 Complementación de la mutante as1 .................................................. 41 VII.6. Análisis de expresión por RT-PCR del gen AtqGlsA ................................ 42 VII.7. Obtención de la secuencia completa del cDNA de AtqGlsA ..................... 43 VII.7.1. RACE 5’ ................................................................................................. 44 VII.8. Expresión constitutiva de AtqGlsA en A. thaliana. .................................... 52 VII.9. Analisis de la mutante GlsA de A.thaliana ................................................ 52 VII.10. Análisis de expresión por RT-PCR de los genes endógenos de
Arabidopsis AS1 y KNAT1 ................................................................................. 53 VII.11. Establecimiento de explantes de Agave tequilana in vitro ...................... 54 VII.12. Infección de los explantes con Agrobacterium tumefaciens ................... 54 VIII. DISCUSIÓN ................................................................................................... 55 IX. CONCLUSIONES ............................................................................................ 64 X. PERSPECTIVAS .............................................................................................. 65 XI. REFERENCIA BIBLIOGRAFICA ..................................................................... 66

vi
ÍNDICE DE TABLAS
Tabla 1. Géneros y especies que forman la familia Agavaceae .............................. 2 Tabla 2. Clasificación taxonómica de Agave tequilana Weber var. Azul ................. 5 Tabla 3. Secuencias de genes relacionadas con la formación de bulbilos ............ 20 Tabla 4. Oligonucleótidos específicos usados para el análisis por RT-PCR en A.
tequilana ............................................................................................................ 21 Tabla 5. Reacción de digestión ............................................................................. 24 Tabla 6. Oligonucleótidos específicos usados para el análisis por RT-PCR en A.
thaliana .............................................................................................................. 31 Tabla 7. Porcentajes de identidad entre AtqAS1 y sus ortólogos de Arabidopsis y
de maíz .............................................................................................................. 37 Tabla 8. Condiciones de amplificación del RACE ................................................. 45

vii
ÍNDICE DE FIGURAS
Figura 1. Producción de tequila en México.............................................................. 3 Figura 2. Exportaciones de tequila en los principales países .................................. 4 Figura 3. Análisis de expresión por RT-PCR de genes KNOX en la mutante as1 y
silvestre .............................................................................................................. 11 Figura 4. Dominio J conservado en GlsA .............................................................. 12 Figura 5. Esquema de la reacción RACE 5’ .......................................................... 14 Figura 6. Sistema de recombinación Gateway® .................................................... 16 Figura 7. Tejido colectado durante la formación de bulbilos ................................. 19 Figura 8. Vectores Gateway® utilizados para clonación y expresión heteróloga ... 25 Figura 9. VectorPCR8®/GW/Topo® ....................................................................... 30 Figura 10. RT-PCR de AtqAs1 y AtqKNOX1 en A. tequilana ................................ 33 Figura 11. Detección del mRNA de AtqAS1 en la Hibridación in situ .................... 34 Figura 12. DNA plasmídico y Digestion de las clonas conteniendo el gen AtqAs1 35 Figura 13. Secuencia completa del cDNA de AtqAS1 con el ORF traducido ........ 37 Figura 14. Alineamiento de las secuencias de aminoácidos de AS1 .................... 37 Figura 15. DNA plasmídico y PCR de colonias de E.coli ...................................... 38 Figura 16. PCR de colonias de A. tumefaciens ..................................................... 39 Figura 17. Plantas de A. thaliana .......................................................................... 40 Figura 18. Comparación de fenotipos WT y as1 ................................................... 41 Figura 19. Fenotipo de la mutante as1 complementada con AtqAS1 .................... 41 Figura 20. Análisis de expresión por RT-PCR ....................................................... 42 Figura 21. Alineamiento en aminoácidos de las secuencias de AtqGlsA .............. 43 Figura 22. Prueba de integridad del cDNA ............................................................ 44 Figura 23. Prueba de integridad del cDNA por PCR ............................................. 45 Figura 24. PCR RACE5' anidado .......................................................................... 46 Figura 25. PCR RACE5' modificado ...................................................................... 47 Figura 26. Secuencia completa del ORF de AtqGlsA ............................................ 49 Figura 27. Alineamiento del ORF completo de AtqGlsA y sus ortólogos ............... 50 Figura 28. AtqGlsA completo ................................................................................. 51 Figura 29. RT-PCR de la interacción entre genes AS1 y KNOX .......................... 53 Figura 30. Modelo de la posible función de AtqAS1 y AtqGlsA en la formación de bulbilos en Agave tequilana…………………...………………………..………………63

viii
RESUMEN.
El Agave tequilana se puede reproducir de tres formas: semillas, hijuelos de
rizoma y bulbilos de la inflorescencia. Los bulbilos son hijuelos que se forman de
manera asexual en la inflorescencia cuando la producción de semillas es
interrumpida, asegurando la reproducción. Este fenómeno es raro en la naturaleza
y ha sido poco estudiado, las señales involucradas en iniciar el proceso de
inducción son desconocidas.
Con el propósito de identificar genes involucrados en este proceso,
en un trabajo previo fueron construidas tres bibliotecas de cDNA de tejido de
pedicelos florales en los que se indujo la formación de bulbilos y fue realizada una
búsqueda de genes que pudieran estar involucrados. La búsqueda reveló cDNAs
con identidad a GONIDIA FORMING PROTEIN (GlsA) de Lilium longiflorum y
ASYMMETRIC LEAVES1 (AS1) de Arabidopsis thaliana, se sabe que ambos
estan involucrados en la diferenciación celular. El objetivo de este trabajo fue
determinar el patrón de expresión de estos por RT-PCR e Hibridación in situ
durante la formación de bulbilos y realizar expresión ectópica en Arabidopsis
thaliana.
Los resultados de los RT-PCR mostraron que la expresión de AtqAS1 se
incrementa durante la formación de bulbilos en A. tequilana y también mostraron
una fuerte expresión en flores y meristemos laterales. Estos resultados están de
acuerdo con el patrón de expresión observado en A. thaliana, donde el nivel de
expresión más alto en órganos inmaduros sugiere un papel en la división y
diferenciación celular. La expresión de AtqGlsA en A. tequilana mostró altos
niveles en etapas iniciales de la formación de meristemos los cuales darán origen
a los bulbilos, disminuyendo en etapas en las cuáles los bulbilos ya están
formados. Esto sugiere una importante función de AtqGlsA durante la etapa de
inducción, quizá asociada con la diferenciación celular, como fue reportado en
Lilium longiflorum.

ix
El cDNA de longitud completa de AtqAS1 fue expresado ectópicamente y
usado para complementar líneas mutantes as1 de Arabidopsis thaliana para inferir
su función en Agave tequilana. Las plantas expresando ectópicamente AtqAS1
mostraron fenotipo similar a las plantas 35S::AS1 y la mutante as1 fue
exitosamente complementada, los resultados mostraron que AtqAS1 es
equivalente en función a AS1. El cDNA completo de AtqGlsA fue obtenido usando
RACE 5´, fue expresado ectópicamente en A. thaliana y está en proceso el
análisis de las transformantes. Por ultimo fue intentada la transformación y
sobreexpresión de AtqAS1 en A. tequilana.
ABSTRACT.
Agave tequilana can reproduce by three forms: seeds, offsets and bulbils.
Bulbils are asexual offsets on the inflorescences that are produced when seed
production is interrupted, thus assuring reproduction. This phenomenon is rare in
nature and little studied, the signals involved in initiating the process of induction
are unknown.
With the aim of identifying genes involved in this process, in previous work,
three cDNA libraries of floral stems induced to produce bulbils were constructed
and a search for genes that could be involved in this process was carried out. The
search revealed cDNA´s showing homology to GONIDIA FORMING PROTEIN
(GlsA) of Lilium longiflorum and ASYMMETRIC LEAVES1 (AS1) of Arabidopsis
thaliana, both of which are known to be involved in cellular differentiation. In this
work, the objective was to determine the expression pattern of these genes by RT-
PCR and in situ hybridization during bulbil formation and ectopic expression in
Arabidopsis thaliana.
RT-PCR data showed that expression of AtqAS1 increases during bulbil
formation in A. tequilana, and also showed strong expression in flowers and lateral
meristems. These results agree with the expression pattern observed in A.

x
thaliana, where a higher level of expression in immature organs suggests a role in
cellular division and differentiation. AtqGlsA expression in A. tequilana showed
high levels at the initial stage of meristem formation which gives rise to bulbils,
decreasing in stages in which bulbils already are formed. This suggests an
important function for AtqGlsA during the induction stage, perhaps associated with
cellular differentiation, as reported for Lilium longiflorum.
The AtqAS1 full length cDNA was ectopically expressed and used to
complement mutant as1 lines of Arabidopsis thaliana in order to infer their function
in Agave tequilana. Plants ectopically expressing AtqAS1 showed similar
phenotypes to 35S::AS1 and the as1 mutant was successfully complemented.
Results showed that AtqAS1 is equivalent in function to AS1. AtqGlsA full length
cDNA was obtained using RACE 5´ and ectopically expressed in A. thaliana.
Analysis of transformants is in process. Transformation and AtqAS1
overexpression in A. tequilana also was attempted.

1
I. INTRODUCCIÓN.
La familia Agavaceae cuenta con 9 géneros y 330 especies, es endémica
de América y su centro de origen y diversidad se encuentra en México donde se
localizan 76 % de las especies descritas en el mundo. Desde tiempos
prehistóricos el hombre ha aprovechado esta familia como alimento, bebidas,
herramientas, fibras, medicamentos y como material de construcción. Su
importancia económica actual deriva principalmente de la obtención de fibras y
bebidas alcohólicas como lo son el tequila y mezcal (García-Mendoza, 2007).
De la familia Agavaceae el Agave tequilana Weber var. Azul se encuentra
entre los de mayor importancia económica ya que se han cultivado cientos de
hectáreas en el oeste de México con la finalidad de producir tequila ya que La
Denominación de Origen del tequila, establece que sólo la variedad Azul de Agave
tequilana puede ser utilizada para la elaboración de tequila, debido a sus
cualidades deseables como lo son el alto contenido de azúcares reductores, y lo
prolífico de sus hijuelos (Valenzuela, 1997).
El Agave tequilana Weber var. Azul se puede reproducir de forma sexual
por medio de semillas y de forma asexual mediante hijuelos de rizoma y bulbilos.
Dado que en la práctica se suprime la floración del cultivo y a que las semillas
tienen un bajo porcentaje de germinación y las plantas son muy heterogéneas, se
suele usar los hijuelos de rizoma como método de propagación para plantaciones,
este método de propagación se ha usado por 200 años o más por la rapidez con
que se obtienen plántulas de buen tamaño y la cantidad que se producen. Los
bulbilos son pequeños hijuelos que se presentan en la inflorescencia al fallar la
reproducción sexual (Arizaga y Ezcurra, 1995). Sin embargo son un método de
reproducción muy exitoso, pues cada planta puede llegar a producir hasta 3000
bulbilos, además se establecen fácilmente en suelo y estudios preliminares
mostraron que el porcentaje de variabilidad genética entre ellos es bajo (Simpson,
datos no publicados). Hasta ahora no hay reportes acerca de los mecanismos
moleculares que se llevan a cabo para la formación de los bulbilos en Agave.

2
II. ANTECEDENTES. II.1. Importancia de los Agaves
En la actualidad se reconocen en la familia Agavaceae nueve géneros y
cerca de 300 especies (Tabla 1. Eguiarte et al., 2000; Rocha et al., 2006). El gran
número de especies es notable, si se toma en cuenta la gran similitud morfológica
y ecológica que existe entre la mayoría de las especies del género (Eguiarte et al.,
2000).
Tabla 1. Géneros y especies que forman la familia Agavaceae.
Géneros No. de especies No. en México
Yucca 49 29
Hesperoyucca 1 1
Hesperaloe 5 5
Beschorneria 7 7
Furcraea 25 11
Agave 166 125
Manfreda 28 27
Polianthes 13 13
Prochnyanthes 1 1
Total 293 217
Las plantas de este género son perennes, rizomatosas, de tallos acaules,
hojas grandes dispuestas en roseta y suculentas-fibrosas que terminan en una
espina; los márgenes de las hojas presentan pequeñas espinas ganchudas o
rectas; inflorescencia en espiga o panoja con escapo largo semileñoso; las flores
son de color amarillo verdoso, protándricas con perianto infundiliforme de tubo de
longitud variable y seis segmentos casi iguales; seis estambres filamentosos
filiformes, más largos que los segmentos del perigonio, con anteras amarillentas;
ovario ínfero trilocular, tricarpelar, con placentación axilar, multiovulada; fruto

3
capsular leñoso alargado, con tres alas con numerosas semillas aplanadas algo
triangulares de testa negra (Gentry, 1982).
Los agaves son monocárpicos, esto es, que sólo tienen una floración al
cabo de la cual la planta muere. Aún cuando exista alta producción de semillas en
la reproducción sexual, debido a su gran depredación y también a que las
condiciones de germinación no son siempre muy adecuadas, su reproducción es
principalmente en forma asexual, por hijuelos de rizoma (Granados, 1993).
México es el centro de origen y diversidad del género Agave y tiene una
clara importancia económica en el país, principalmente debido a la industria
tequilera.
En las siguientes gráficas se muestra la producción de tequila en México y
la tasa de exportación, hasta el año 2006.
Fig. 1. Producción de tequila en México.
4
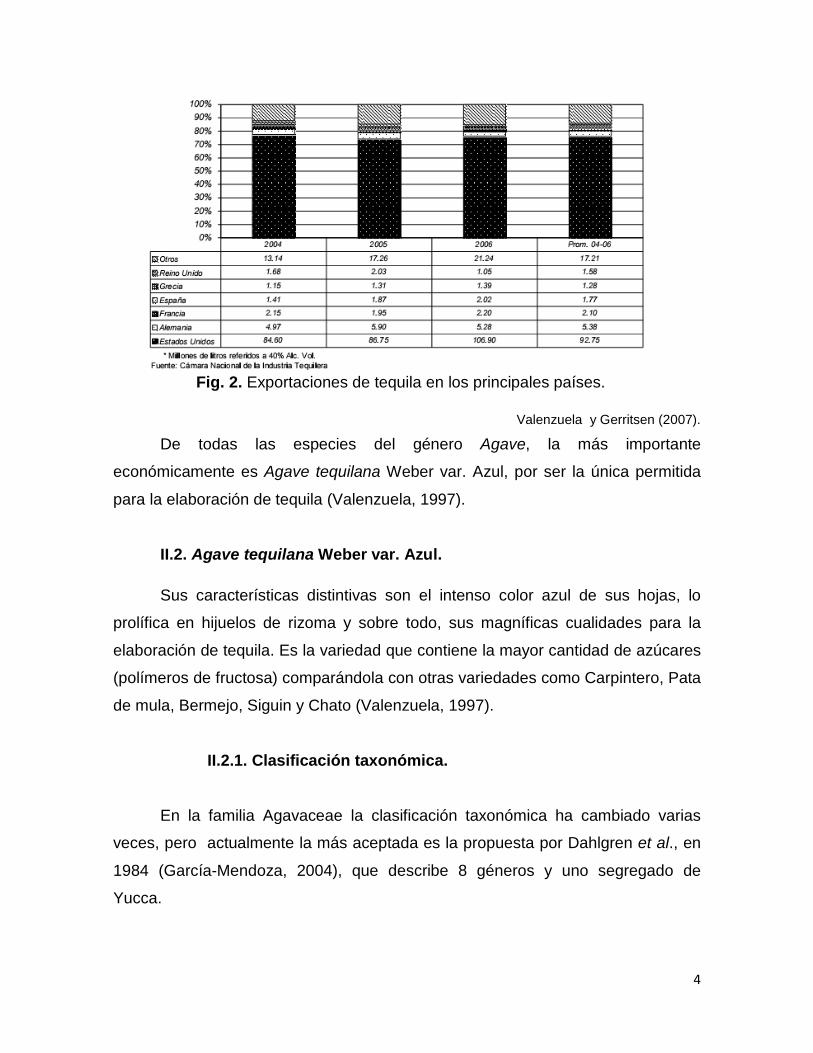
Valenzuela y Gerritsen (2007).
De todas las especies del género Agave, la más importante
económicamente es Agave tequilana Weber var. Azul, por ser la única permitida
para la elaboración de tequila (Valenzuela, 1997).
II.2. Agave tequilana Weber var. Azul.
Sus características distintivas son el intenso color azul de sus hojas, lo
prolífica en hijuelos de rizoma y sobre todo, sus magníficas cualidades para la
elaboración de tequila. Es la variedad que contiene la mayor cantidad de azúcares
(polímeros de fructosa) comparándola con otras variedades como Carpintero, Pata
de mula, Bermejo, Siguin y Chato (Valenzuela, 1997).
II.2.1. Clasificación taxonómica. En la familia Agavaceae la clasificación taxonómica ha cambiado varias
veces, pero actualmente la más aceptada es la propuesta por Dahlgren et al., en
1984 (García-Mendoza, 2004), que describe 8 géneros y uno segregado de
Yucca.
Fig. 2. Exportaciones de tequila en los principales países.

5
Tabla 2. Clasificación taxonómica de Agave tequilana Weber var. Azul.
División Angiospermae
Clase Monocotyledonae
Orden Asparagales
Familia Agavaceae
Subfamilia Agavoideae
Género Agave
Subgénero Agave
Especie Tequilana
Variedad Azul
García-Mendoza, 2004; Colunga-García et al., 2007.
II.2.2. Métodos de propagación.
El agave tequilero tiene tres formas de propagarse: por vía sexual en forma
de semillas, por hijuelos de rizomas y por bulbilos (pequeños hijuelos) de la
inflorescencia o quiote, las dos últimas son vías asexuales. La vía sexual, por
semillas, no es utilizada y pocas veces se ve, ya que se suprime la floración del
cultivo. Las semillas tienen bajo porcentaje de germinación, su crecimiento es
lento y las plántulas resultantes son muy heterogéneas para ser utilizadas en el
cultivo. Los hijuelos de rizoma son usados comúnmente para el establecimiento de
plantaciones. Ésta ha sido la forma de propagación más común que se ha
practicado durante mucho tiempo (doscientos años o más). La ventaja de esta
forma de propagación es la rapidez con que se obtienen plántulas de buen tamaño
y la cantidad de éstas que produce la planta. Los bulbilos son hijuelos pequeños
que emergen en el quiote, junto con las flores no fecundadas que caen
posteriormente sin formar frutos. Son poco frecuentes y casi desconocidos porque
la floración se reprime. Estos hijuelos no se utilizan en la propagación de material
vegetativo para plantación. También existe la micropropagación de agave por

6
cultivo de tejidos, como una propagación masiva, aunque los costos son mayores
por lo que sólo la utilizan las grandes empresas tequileras (Valenzuela, 1997).
La reproducción sexual se logra mediante la polinización que efectúan
algunos animales, principalmente murciélagos nectarívoros y en menor grado,
insectos diurnos y nocturnos (palomillas, abejas, abejorros) y aves (colibríes, aves
percheras). Pero la mayoría de los agaves se propaga de manera asexual,
produciendo hijuelos en diferentes partes de la roseta o la inflorescencia (García-
Mendoza, 2007).
En ambientes extremos como en los que se desarrollan las especies de
Agave y otras plantas semélparas, la propagación vegetativa funciona como una
estrategia segura para perpetuar el genotipo, ya que la reproducción sexual
parece involucrar grandes riesgos y la probabilidad de éxito en el establecimiento
de nuevas plántulas con frecuencia es extremadamente baja por estar sujeta a la
limitada oportunidad en espacio y tiempo. En contraste, los propágulos vegetativos
tienen la ventaja de estar unidos a la planta madre durante las etapas tempranas
de su ciclo de vida, un hecho que les da una probabilidad mucho más alta de
sobrevivir en condiciones ambientales desfavorables (Arizaga y Ezcurra 2002).
II.3. Producción de bulbilos en Agave.
Una de las adaptaciones del género Agave es la diversidad de mecanismos
de reproducción. Este género ha adquirido mecanismos eficientes que le permiten
un escape a la semelparidad, la propagación clonal es muy importante en la
mayoría de las especies del género. En la propagación clonal, al producir nuevas
rosetas de manera vegetativa, un genotipo no muere al reproducirse, ya que
quedan copias más jóvenes pero geneticamente identicas. Aunque existen
algunos géneros que han escapado a la semelparidad, produciendo
inflorescencias anuales, como Manfreda, Polianthes y Prochnyanthes, no siendo el
caso en Agave (Colunga-García et al., 2007).

7
La producción de bulbilos es prolífica en A. angustifolia Haw. var.
marginata, A. fourcroydes Lem., A. murpheyi, A. sisalana y A. wercklei. Cuatro de
estas especies no producen semillas germinables y pueden representar híbridos
estériles (Szarek et al., 1996). Las plantas que producen un gran número de
cápsulas con semillas, usualmente no forman bulbilos o muy pocos y las que
producen pocas cápsulas forman grandes cantidades de bulbilos, por lo que se ha
sugerido que la formación de bulbilos está inversamente relacionada al éxito de la
fructificación (Arizaga y Ezcurra, 1995).
Valenzuela (1997) indujo con el corte de botones florales la propagación de
bulbilos y encontró que no sólo los botones cortados emiten bulbilos, sino también
las flores que no reciben polen extraño. La propagación por bulbilos es una
estrategia utilizada en otras especies de agave como en A. angustifolia y A.
sisalana, de esta manera se obtienen tantos bulbilos como botones florales hayan
existido, de 2000 a 3000 plántulas por planta madre se han observado en A.
sisalana. Esta técnica es relativamente más barata para la propagación masiva,
sin embargo, habría que estudiar la homogeneidad genética que se produce y
tomar en cuenta el tiempo de maduración de cada especie (Valenzuela, 1997).
A pesar de que la formación de bulbilos es un proceso relativamente poco
frecuente, se sabe que su formación no es exclusiva de la familia Agavaceae,
también está presente en otras familias de monocotiledóneas y aún en
dicotiledóneas (Arizaga y Ezcurra, 1995).
Un ejemplo de formación de bulbilos similar a lo que se observa en el
género Agave, es Titanotrichum oldhamii que pertenece a la familia
Gesneriaceae. En esta planta un solo meristemo floral es reemplazado por un
grupo de 50 a 70 bulbilos, formados en las axilas de las brácteas. Al final de la
época de floración, la mayor parte de los meristemos florales se convierten en
meristemos vegetativos, dando lugar a los bulbilos, así decenas de miles de
bulbilos pueden producirse de una sola planta y el porcentaje de viabilidad de

8
éstos es muy alto (alrededor de 95 %) comparado con el de las semillas (75 %
aproximadamente) por lo que los bulbilos son el principal medio de propagación
de Titanotrichum (Wang y Kronk, 2003). En Agave tequilana el porcentaje de
germinación de las semillas en campo es apenas de alrededor de 12 % (Escobar-
Guzmán, et al. 2008), lo que sugiere que requiere vías alternativas de
propagación.
II.4. Genes expresados durante el desarrollo de bulbilos. Hasta ahora no existen reportes sobre genes directamente relacionados
con la formación de bulbilos. Entre los escasos reportes que describen la
existencia de bulbilos está el de Wang, et al. (2004) en Titanotrichum oldhamii,
donde se evaluó la expresión del gen GFLO (ortólogo de FLORICAULA de
Antirrhinum majus) un gen que induce la floración, se observó que la expresión
disminuye durante la formación de bulbilos hasta desaparecer. Los autores
propusieron que los genes responsables de la formación de bulbilos debían ser los
que regulan negativamente a los genes que inducen la floración como GFLO, sin
embargo no se ha identificado algún gen con esta función en plantas que formen
bulbilos.
El primer estudio realizado en Agave sobre la formación de bulbilos a nivel
histológico y molecular se llevó a cabo en Agave tequilana (Abraham-Juárez et al.,
2010), esta especie forma bulbilos de manera inducida, es decir sólo cuando se
propician las condiciones adecuadas, que en general incluyen métodos para evitar
la formación de semillas. Se observó que la forma en que ocurre el desarrollo de
bulbilos es a través de un programa con características de organogénesis, a partir
de formación de meristemos de novo y se propuso que probablemente existía un
reclutamiento de programas de desarrollo de tipo organogénico que confería a un
grupo de células determinadas la capacidad de adquirir una nueva identidad
meristemática. En este trabajo se llevó a cabo la elaboración de bibliotecas de
cDNA como una estrategia para la búsqueda de genes que pudieran estar

9
involucrados en este proceso de desarrollo. Entre los genes propuestos se
encontraron algunos reportados en otras plantas que tienen la función de inicio y
desarrollo de meristemos, un segundo grupo identificado está compuesto por
genes expresados específicamente durante el desarrollo de bulbilos en Agave
tequilana. Para analizar el efecto de algunos de estos genes en el desarrollo, se
llevó a cabo expresión heteróloga en Arabidopsis thaliana, con lo cuál se pudo
comprobar que los ortólogos de genes KNOXI de Agave tienen funciones
equivalentes a los de Arabidopsis. En base a los análisis de expresión por RT-
PCR en tiempo real e Hibridación in situ y la expresión heteróloga, se propuso un
modelo para formación de bulbilos en Agave tequilana que involucra señales de
desarrollo vegetativo y la posible participación de los genes AtqKNOX1,
AtqKNOX2, AtqUNK1 y AtqUNK2.
En el grupo de genes identificados en las bibliotecas de cDNA durante la
formación de bulbilos en Agave tequilana también se encuentran AtqGlsA y
AtqAS1. Estos genes no han sido caracterizados durante este proceso por lo que
su estudio es de gran importancia en la construcción del modelo propuesto.
AtqGlsA parece muy interesante por su expresión exclusiva en los pedicelos
durante la formación de bulbilos y AtqAS1 por la función de su ortólogo AS1 de
Arabidopsis thaliana en la formación de primordios de órganos laterales. Esta
información sugiere que estos dos genes participan en la formación de bulbilos.
II.5. ASYMMETRIC LEAVES1 (AS1).
El gen AS1 de Arabidopsis, ortólogo de RS2 de maíz, reprime los genes
KNOXI durante la formación de hojas, en mutantes rs2 se observa expresión
ectópica alrededor de la vasculatura de la hoja de proteínas KNOXI, los genes
KNOXI se expresan en regiones meristemáticas y tienen la función de iniciar y
mantener los meristemos. Así los genes AS1 y KNOXI se expresan en dominios
mutuamente excluyentes (Tsiantis et al., 1999). AS1 está involucrado en el

10
mantenimiento de las células que formarán las hojas. En dicotiledóneas la
sobreexpresión de genes KNOXI produce hojas lobuladas, estipulas distales y
meristemos ectópicos ocasionales en la parte adaxial de la hoja similar a la
mutante as1 (Chuck et al., 1996).
En la mutante as1 se interrumpe el desarrollo de cotiledones, hojas y
órganos florales. En plantas mutantes as1 adultas se ha observado callos en la
superficie de la hoja y brotes ectópicos en la parte adaxial del peciolo similar a las
sobreexpresantes KNAT1. En la mutante as1 cambia el patrón de crecimiento de
las células extendiéndose en múltiples direcciones del peciolo. En el meristemo
además se expresan otros genes que son necesarios para su funcionamiento
como CLV, WUS y CUC2. AS1, PHAN y RS2 (de Arabidopsis, Antirrhinum y maíz
respectivamente) regulan negativamente genes KNOXI en primordios de órganos,
lo que demuestra un grado de conservación funcional (Byrne et al 2000).
La actividad del meristemo apical es especificado en parte por los genes
KNOXI y por un complejo formado por AS1 y AS2 que crea un asa en el promotor
de KNOXI y crean un estado de la cromatina que bloquea la actividad del
enhancer durante la organogénesis (Guo et al 2008).
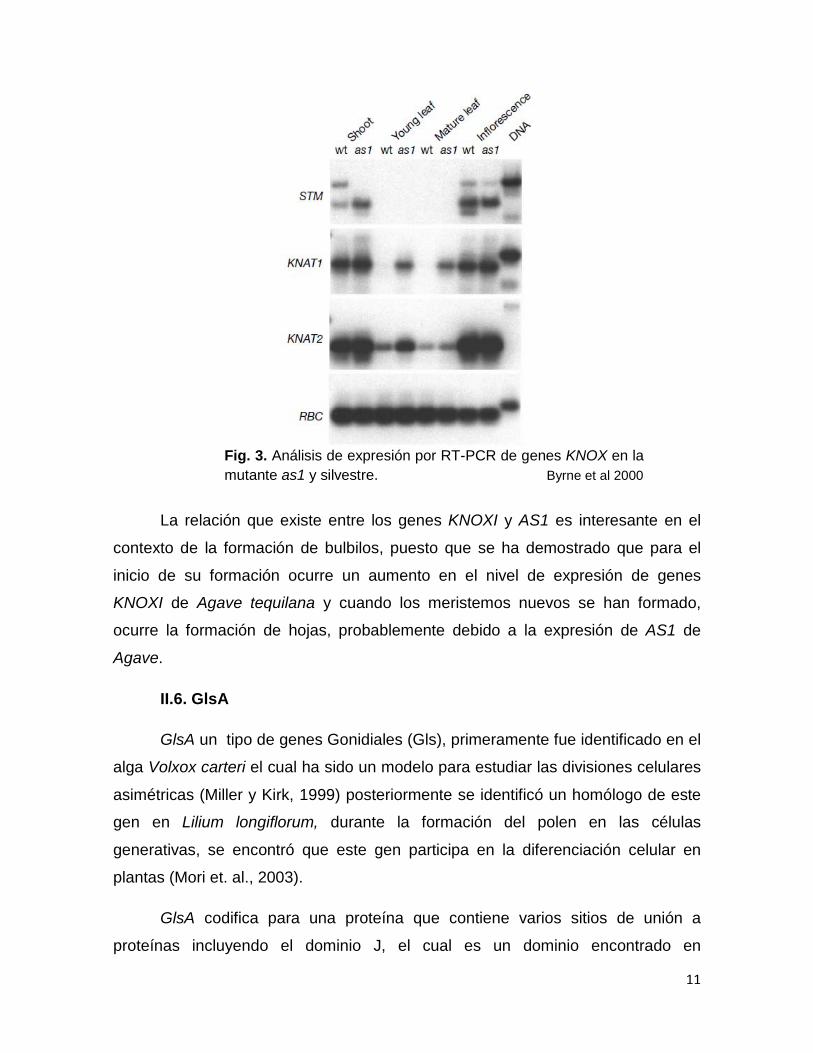
Análisis de expresión por RT-PCR en plantas mutantes as1 y WT mostraron
que en los genes de la clase KNOXI, STM, KNAT1 y KNAT2 aumentan su
expresión en la mutante as1, demostrando sus patrones antagonistas, además se
reportó que la mutante as1 puede rescatar el fenotipo de la mutante stm en
meristemos vegetativos y de embriones (Byrne et al 2000).
11
La relación que existe entre los genes KNOXI y AS1 es interesante en el
contexto de la formación de bulbilos, puesto que se ha demostrado que para el
inicio de su formación ocurre un aumento en el nivel de expresión de genes
KNOXI de Agave tequilana y cuando los meristemos nuevos se han formado,
ocurre la formación de hojas, probablemente debido a la expresión de AS1 de
Agave.
II.6. GlsA
GlsA un tipo de genes Gonidiales (Gls), primeramente fue identificado en el
alga Volxox carteri el cual ha sido un modelo para estudiar las divisiones celulares
asimétricas (Miller y Kirk, 1999) posteriormente se identificó un homólogo de este
gen en Lilium longiflorum, durante la formación del polen en las células
generativas, se encontró que este gen participa en la diferenciación celular en
plantas (Mori et. al., 2003).
GlsA codifica para una proteína que contiene varios sitios de unión a
proteínas incluyendo el dominio J, el cual es un dominio encontrado en
Fig. 3. Análisis de expresión por RT-PCR de genes KNOX en la mutante as1 y silvestre. Byrne et al 2000

12
chaperonas activadas por choque térmico y es esencial para que se puedan llevar
a cabo las divisiones asimétricas, pero este dominio no afecta las divisiones
simétricas (Miller y Kirk 1999).
Se han encontrado ortólogos de GlsA en diferentes especies desde
microorganismos, invertebrados hasta animales y plantas; en Arabidopsis thaliana
existen dos ortólogos del gen y existen dos mutantes insercionales, una para cada
gen, sin embargo hasta ahora no se ha reportado ningún fenotipo para estas
mutantes.
Lilium
Dominio J Dominio M
Volvox
Arabidopsis
Chaperona J
Arabidopsis
Fig.4. Dominio J conservado en GlsA. El dominio J seencuentra conservado en ortologos de GlsA, en L. longiflorum,A. thaliana, V carteri y en E. coli, se encuentra en la chaperonade choque térmico DNAJ.
Mori et al., 2003
A partir del análisis de las bibliotecas de cDNA de la formación de bulbilos
en Agave tequilana, se identificó un ortólogo del gen GlsA cuya expresión fue
exclusiva de este tejido y estado de desarrollo (Abraham-Juárez, 2010. Tesis de
doctorado); en base a los reportes en Lilium longiflofum, es probable que esté
involucrado en las divisiones celulares que ocurren en las primeras etapas de la
formación de bulbilos. El cDNA correspondiente a GlsA que se identificó no cuenta
con la región codificante completa, por lo que será necesario completarla usando
la tecnología de RACE PCR. Esto permitirá realizar análisis de expresión

13
heteróloga por expresión constitutiva y complementación de mutantes en
Arabidopsis thaliana para evaluar su función.
II.7. Amplificación rápida de extremos de DNA complementario (RACE).
El RACE es un método usado para obtener la parte desconocida de
secuencias, ya sea hacia el 5’ ó el 3’ del cDNA, usando una parte de la secuencia
conocida proveniente de algún método de escrutinio de bibliotecas, con este
método se asegura la amplificación de transcritos completos, mediante la
eliminación de RNA mensajero truncado y en general de todo RNA que no tenga
Cap y cola de poliA, mediante la acción de enzimas como la fosfatasa intestinal de
ternera (CIP), dejando la posibilidad de clonar el producto del RACE y secuenciar
por métodos convencionales (Zhang y Frohman, 2008).
El RACE comúnmente es usado para identificar las secuencias
desconocías de cDNA hacia los extremos 5’ ó 3’ incluyendo las secuencias no
traducidas (UTR), para estudiar los sitios de inicio de la transcripción, caracterizar
las regiones promotoras y obtener la secuencia de cDNA completa de un gen.
A continuación se presenta en forma esquemática la técnica de RACE 5´.

14
Primera cadena cDNA
Oligo de RNA
Transcriptasa reversa
3’ Cola de poli A
Oligo anidado Gene Racer 5’
Primera cadena cDNA
Oligos GSP anidado y GSP
Oligo Gene Racer 5’
Oligo de RNA 3’ Cola de poli A
RNA ligasa
RNA no mensajero
RNA truncado AAAAAAAA
mRNA m7G-p-p-PO4 AAAAAAAA 3’ cola de poli A 5’ CAP
TAP
RNA no mensajero
RNA truncado AAAAAAAA
mRNA m7G-p-p-PO4 AAAAAAAA 3’ cola de poli A 5’ CAP
CIP PO4
CIP PO4
Fig.5. Esquema de la reacción RACE 5’. A Defosforilación con la CIP para eliminar todo lo que no sea RNA mensajero completo. B Eliminación de la Cap. C Adición del sitio conocido gene RACER 5’. D Síntesis de cDNA por la transcriptasa reversa E Amplificación del fragmento desconocido por PCR con el oligo reverso específico.
A
B
C
D
E

15
La obtención de las secuencias completas de los cDNAs permite utilizarlos
para análisis de expresión y evaluar su función biológica. Para esto se dispone de
la alternativa de expresión heteróloga en Arabidopsis thaliana, puesto que el
método de transformación genética de Agave aún no está establecido.
Para llevar a cabo la expresión heteróloga un método eficiente es la
utilización de vectores de clonación con la tecnología Gateway® (Invitrogen).
II.8. Tecnología Gateway®.
La tecnología Gateway® es un método de clonación que aprovecha las
ventajas de recombinación sitio específico del bacteriófago λ para mover un gen
de interés a otros sistemas como vectores de expresión de una forma más sencilla
y rápida. Los métodos convencionales de clonación en vector tienen algunas
desventajas como el que algunas enzimas de restricción no se pueden utilizarse,
ya que podrían cortar dentro del gen de interés, los métodos de limpieza
necesarios reducen la eficiencia de recuperación de clonas conteniendo
fragmentos grandes, además se requiere de mayor tiempo para seleccionar las
colonias que tienen el inserto adecuado. La tecnología Gateway® ofrece un
método de clonación más rápido y hasta con una eficiencia del 99 %, se mantiene
la orientación y el marco de lectura, no se necesitan enzimas de restricción, y
ofrece la flexibilidad de pasar el gen de interés de un vector a otro de expresión de
forma reversible.
Para que se pueda llevar a cabo la recombinación sitio específico es
necesario que se cuente con la secuencia de recombinación att y la clonasa que
es la enzima que media la reacción de recombinación, los sitios de recombinación
se denominan attB, attP, attL y attR y estos sitios solamente se pueden
recombinar de forma especifica entre los sitios attB y attP y los sitios attL y attR y
a su vez son mediadas por las enzimas clonasa LR y clonasa BP respectivamente,
en la reacción LR se forma un vector de expresión transfiriendo fragmentos de un
vector de entrada con los sitios attL a un vector destino con los sitios attR,

16
produciéndose un vector de expresión con los sitios attB y un vector producto con
los sitios attP Fig.6.
Fig.6. Sistema de recombinación Gateway® . A, reacción LR para dar origen a un vector de expresión. B, reacción BP para dar origen a un vector de entrada.
A
B
Fig.6. Sistema de recombinación Gateway® . A, reacción LR para dar origen a un vector de expresión. B, reacción BP para dar origen a un vector de entrada.
A
B
III. JUSTIFICACIÓN.
Multiplicar plantas sin necesidad de que estas se reproduzcan sexualmente,
ha sido una de los principales objetivos de la biotecnología vegetal desde su
origen. Conocer y entender el mecanismo por el cual algunas especies logran
clonarse naturalmente traería grandes beneficios para la propagación de plantas
que presentan problemas para reproducirse de forma sexual.
Agave tequilana Weber var. Azul es un buen modelo para estudiar la
formación de bulbilos, por presentar este fenómeno de forma inducible y por la
gran cantidad de tejido disponible para el análisis debido al tamaño de las
inflorescencias.
El análisis de las bibliotecas de cDNA que se generaron a partir de tejido de
pedicelos florales durante la formación de bulbilos como una fuente de información
de expresión de genes es esencial para entender este fenómeno.

17
En base a estudios previos realizados en el grupo de trabajo, se
seleccionaron los genes AtqAS1 y AtqGlsA para ser caracterizados durante la
formación de bulbilos. Su análisis es de gran importancia para entender el proceso
y poder aportar información útil a los estudios de plantas que se propagan en
forma vegetativa.

18
IV. HIPÓTESIS.
Los genes AtqAS1 y AtqGlsA están involucrados en el proceso de desarrollo de
bulbilos en Agave tequilana.
V. OBJETIVO.
V.1. Objetivo general.
Caracterizar los genes AtqAS1 y AtqGlsA de Agave tequilana durante la formación de bulbilos en términos de su expresión y función en un sistema heterólogo.
V.2. Objetivos específicos.
• Analizar los patrones de expresión por RT- PCR e Hibridación in situ durante la formación de bulbilos, de los genes seleccionados.
• Completar la secuencia del gen AtqGlsA mediante RACE-PCR.
• Verificar en un sistema de expresión heterólogo (Arabidipsis thaliana) la función de los genes AtqAS1 y AtqGlsA, por sobreexpresión y complementación.
• Determinar la relación en patrones de expresión entre AtqAS1 y genes KNOX en líneas expresando AtqAS1.
• Infectar explantes de Agave tequilana in vitro mediante infiltración con Agrobacterium tumefaciens transformada con el gen AgAS1.
• Verificar la transformación genética de Agave tequilana mediante expresión del gen de GFP, resistencia a fosfinotricina y PCR.

19
VI. MATERIALES Y METODOS.
VI.1. Colecta de muestras.
Para los análisis de expresión por RT-PCR e Hibridación in situ y para el
RACE 5´, se colectó tejido de pedicelos florales durante la formación de bulbilos
en Agave tequilana Weber var. Azul, correspondientes a las mostradas en la Fig.7.
En T0-T1 no se observa aún algún cambio morfológico en los pedicelos, en T2 y
T3 inicia la expansión del tejido debajo del bractéolo, en T4 se puede observar
crecimiento de tejido indiferenciado y en T5 los meristemos formados de novo han
adquirido la forma de los bulbilos. También se colectó tejido de raíz, hoja, anteras,
ovarios, tépalos, meristemos de umbelas y meristemo apical reproductivo (tomado
de la punta del eje floral de 1.5 m de altura).
Fig.7. Tejido colectado durante la formación de bulbilos. A, T0/T1. B, T2, C. T3, D. T4, E. T5. El círculo en A indica la zona donde se formarán los bulbilos, el rectángulo en D indica el tejido meristemático indiferenciado. Las barras son de 0.5 cm. Imagen tomada de Abraham-Juárez, 2010.
VI.2. Selección de genes.
Los genes con los que se trabajó se seleccionaron a partir de un proyecto
de análisis del transcriptoma de Agave tequilana para el cual se realizaron 15
bibliotecas de cDNA a partir de diferentes tejidos como, hoja, piña, ovarios,
anteras, raíz, meristemo vegetativo, meristemo floral y bulbilos. Con respecto a la
formación de bulbilos se realizaron tres bibliotecas de diferentes etapas de su
desarrollo. Hasta el momento se cuenta con más de 35000 secuencias, en
promedio de 1200 pb y 300,000 reads de secuenciación 454 (Martínez-Hernández
et al., datos no publicados).
Los criterios que se usaron para su selección fueron:

20
Secuencias que se encontraran únicamente durante el desarrollo de bulbilos
Secuencias que se asociaran con el desarrollo de bulbilos
Con estos criterios se encontraron los siguientes genes.
Tabla 3. Secuencias de genes relacionadas con la formación de bulbilos.
Nombre Criterio de selección
Descripción del gi
UNK1
Secuencias encontradas solo en bibliotecas de bulbilos
gi|23197966|gb|AAN15510.1| expressed protein [Arabidopsis thaliana]
UNK2 gi|50919935|ref|XP_470328.1| expressed protein [Oryza sativa]
UNK3 gi|50920103|ref|XP_470412.1| unknown protein [Oryza sativa]
UNK4 gi|54287600|gb|AAV31344.1| unknown protein [Oryza sativa]
UNK5 gi|6714413|gb|AAF26101.1| unknown protein [Arabidopsis thaliana]
GlsA gi|31442292| gonidia forming protein GlsA [Lilium longiflorum]
WIP gi|34908460|ref|NP_915577.1| WIP zinc finger - like protein [Oryza sativa]
KNAT1
Secuencias asociadas con el desarrollo de meristemos
gi|1256575|gb|AAC49251.1| KNAT1 homeobox-like protein[Solanum lycopersicum]
KNAT2 gi|7581979|emb|CAB88029.1| knotted1-like homeobox protein [Dendrobium grex]
AS1 gi|145360723|ref|NM_129319.3| AS1 (Asymmetric leaves1) [Arabidopsis thaliana]
YABBY2 gi|4928751|gb|AAD33716.1| YABBY2 [Arabidopsis thaliana]
MADSbox gi|33309882|gb|AAQ03227.1| MADS box protein [Elaeis guineensis]
Abraham-Juárez, 2010.
VI.3. Análisis de expresión por RT-PCR.
Para realizar los análisis de expresión de los genes AtqAS1, AtqGlsA y
AtqKNOX1 se colectó tejido de 5 etapas de la formación de bulbilos que van
desde la etapa T0 donde no se ven cambios morfológicos obvios hasta la etapa T5
donde ya se pueden observar embriones somáticos, se extrajo RNA de tejidos de
estas etapas así como de tejido de raíz, ovario, antera, hoja, meristemo joven,
meristemo reproductivo y meristemo de umbela utilizando Trizol (Invitrogen) de
acuerdo a las recomendaciones del fabricante. Con el RNA total se sintetizó cDNA
con la enzima transcriptasa reversa (Fermentas) de acuerdo con las

21
recomendaciones del fabricante y se realizó amplificación por PCR utilizando
oligonucleótidos específicos para los genes seleccionados. Tabla 4.
Tabla 4. Oligonucleótidos específicos usados para el análisis por RT-PCR en A.
tequilana.
VI.4. Hibridación in situ.
El protocolo que se usó está basado en el reportado por Jackson et al.
(1994) y Ruiz-Medrano et al. (1999).
Antes de llevar a cabo el procedimiento de la hibridación, es necesario
sintetizar las ribosondas, estas se pueden almacenar a -80°C durante un mes
hasta su uso. A continuación se describe el procedimiento.
VI.4.1. Preparación del templado de DNA.
Se amplificó un fragmento de 355 pb, que comprende del nucleótido 169 al
523 de la secuencia codificante, incluyendo la mitad del domino MYB y una parte
no conservada entre la secuencia de Agave y Arabidopsis, utilizando los
Nombre ID Secuencia
AtqAS1-F 5’-ATCAAGAAGGGGTCTCTCACC-3’
AtqAS1-F 5’- CTTCCGTTGCTCTGACTCTGT-3’
AtqGlsA-F 5´-GGGTCCGGAATCTAGTTGACA-3´
AtqGlsA-R 5´-TTCTTCCTTCCCCTCCATCTC-3´
AtqKNOX1-F 5´-CCTCTGCAGGAGGCCATGAAT-3´
AtqKNOX1-R 5´-GAGTTCGTGCCTAGGCTGCTG-3

22
oligonucleótidos específicos para AtqAS1, tabla 4, a partir de DNA plasmídico de
la clona aamhmqmeq011_g04 que corresponde a este gen. El producto de PCR
se clonó en el vector pGEM-T Easy (Promega) que contiene los promotores T7 y
SP6. Para determinar el sentido en el que se clonó el producto de AtqAS1 se llevó
a cabo una digestión del DNA plasmídico con la enzima PstI. Esta enzima tiene un
sitio de corte único en un extremo del fragmento AtqAS1 y en el vector pGEM-T
Easy, lo cual permitió identificar clonas con el fragmento en sentido y en
antisentido.
Se eligió una clona en cada orientación para usar el DNA como templado
para las ribosondas. El DNA fue purificado con fenol:cloroformo:alcohol isoamílico
(25:24:1) y precipitado con etanol al 100 % y NaOAc, se midió su concentración en
nanodrop y se almacenó a -20°C hasta su uso.
VI.4.2. Síntesis y marcaje de la ribosonda.
Se utilizó el sistema de Transcripción in vitro Ribroprobe® (Promega) y se
siguió el protocolo de acuerdo a las recomendaciones del fabricante. Al final se
removió el templado de DNA y se purificaron las ribosondas, se midió su
concentración en nanodrop y se almacenaron a -80°C hasta su uso.
VI.4.3. Hibridación.
El procedimiento para la hibridación se describe a continuación en forma
general, está basado en el protocolo adaptado a tejidos de Agave tequilana
descrito por Abraham-Juárez, 2010 (Tesis de doctorado).
a) Pretratamiento del tejido: Remoción de la parafina e hidratación del tejido.
b) Prehibridación: En la solución de hibridación (sin ribosonda) durante una
hora.

23
c) Hibridación de ribosondas: Se usaron 200 ng de ribosonda para cada
laminilla disueltos en la solución de hibridación.
d) Lavados: Con una solución de SSC de alta astringencia.
e) Bloqueo y adición del anticuerpo: Con una solución de leche
descremada (Difco) en PBS 1X y anticuerpo Anti-Digoxigenin-AP Cat. No.
11093274910 (Roche) a 1:2000.
f) Lavados y adición de sustrato. Con PBS 1X, el sustrato fue NBT/BCIP
(Nitro blue tetrazolium chloride/ 5-bromo-4-chloro-3-indolyl phosphate Cat.
No. 11697471001 (Roche), se incubaron a temperatura ambiente cubiertas
de la luz por 2 días.
g) Lavados y deshidratación el tejido. Con PBS 1X y series de etanol hasta
el 100%.
h) Montaje de las laminillas. Con citoseal 60.
VI.4.4. Observación al microscopio.
Se utilizó un microscopio OLYMPUS BX60 y las imágenes fueron tomadas
con la cámara OLYMPUS DP71 acoplada al microscopio.
VI.5. Identificación de la clona con el gen AtqAS1 completo.
De los respaldos de los bancos en bacteria se pusieron a crecer en medio
LB con kanamicina dos clonas que contenían parte de la secuencia del gen
AtqAS1 incubándose a 37°C por 18h en agitación. Se extrajo DNA plasmídico, se
ajustó la concentración de DNA a una concentración de 150ng/µl y se realizó una
digestión usando la enzima de restricción BsrGI incubando a 37°C 3h según la
siguiente reacción:

24
Tabla 5. Reacción de digestión.
Reactivo µl DNA 2.0 BsrGI 0.5 Buffer 2 (1X) 1.0 BSA 10X 1.0 H2Odd 5.5 ∑ 10.0
Además para confirmar el resultado se mandó a secuenciar la clona
seleccionada por los dos lados utilizando los oligonucleótidos universales M3.
Cuando se obtuvo la secuencia de la clona seleccionada se realizó un
análisis in silico para tratar de inferir si la secuencia obtenida estaba completa,
para lo cual se realizaron alineamientos y determinación del porcentaje de
identidad con respecto a genes ortólogos, utilizando el programa Bio Edit. Una vez
teniendo secuencias en nucleótidos de tamaño y secuencia similar se procedió a
buscar el ORF y traducirlo a aminoácidos mediante el programa DNA Star para
realizar nuevamente un alineamiento y saber su porcentaje de identidad en
aminoácidos con respecto a sus genes ortólogos de Arabidopsis y de maíz.
VI.6. Expresión heteróloga.
Para poder analizar la función de los genes seleccionados fue necesario
realizar sobreexpresión y complementación heteróloga en Arabidopsis dado que
no se cuenta con un método estandarizado para la transformación de Agave, para
lo cual una vez teniendo el gen seleccionado completo en el vector de entrada se
procedió a pasarlo mediante recombinación al vector de salida Gateway®
pB7WG2D Fig.8. (Karimi et al., 2002) el cual tiene el promotor constitutivo 35S, el
gen de resistencia al antibiotico espectinomicina, el gen de resistencia al herbicida
fosfinotricina, el gen suicida ccdB y el gen de la proteína verde fluorescente GFP.

25
Fig. 8. Vectores Gateway® utilizados para clonación y expresión heteróloga. A, vector usado para la clonación de las bibliotecas, B, vector usado para la
expresión en A. thaliana.
A
B

26
VI.6.1. Recombinación en vector de salida Gateway® pB7WG2D.
Para llevar a cabo la recombinación se mezclaron en un tubo de PCR:
150 ng de vector pB7WG2D, 150 ng de vector de entrada pDONR222
conteniendo el gen AtqAs1 y en su caso 150 ng de vector PCR®8/GW/Topo®
conteniendo el gen AtqGlsA, 1.0 µL de mezcla de la enzima LR II Clonasa de
Invitrogen y agua desionizada estéril hasta completar 10 µL de volumen final, se
incubó a 25°C por 16 horas, se agregó 1.0 µL de Proteinasa K (20mg/mL) y se
incubó a 25°C por 10 min.
VI.6.2. Transformación de E.coli.
Se dializó la mezcla de recombinación usando una membrana millipore de
0.025 µm por 20 min. Sobre agua desionizada estéril, se mezcló todo el volumen
de la recombinación en un tubo de 1.5 ml con un vial de 50 µL de células
electrocompetentes de E.coli Top 10, se pasó la mezcla a una celda de
electroporación de 2 mm y se electroporó usando el programa EC2 en el
electroporador MicroPulser Electroporator de Biorad. Inmediatamente después se
agregaron 300 µL de medio soc, se regresó el contenido a un tubo de 1.5 ml y se
incubó a 37°C por 1h. En agitación a 200RPM. Se sembró en medio LB sólido
conteniendo 100 mg/L de espectinomicina, se incubó a 37°C por 16h. Se
escogieron 4 colonias, se inoculó medio LB líquido, se extrajo DNA plasmídico y
se verificó que tuvieran el inserto mediante PCR con oligonucleótidos específicos
para el gen de interés.
VI.6.3. Transformación de Agrobacterium tumefaciens.
Después de verificar que el DNA plasmídico seleccionado tuviera el gen de
interés se procedió a transformar la cepa GV2260 de A. tumefaciens por
electroporación bajo las mismas condiciones que para transformar E. coli. Se
sembró en medio YEB sólido con Espectinomicina, Carbenicilina y Rifampicina
100 mg/L de cada uno y se incubó a 28°C durante 48 horas, se seleccionaron tres

27
colonias, se inoculó medio liquido conteniendo los mismos tres antibióticos, se
extrajo nuevamente DNA plasmídico y se comprobó que tuvieran el gen de interés
mediante PCR con oligonucleótidos específicos y se seleccionó una clona, se
mezcló en glicerol a una concentración final de 50 % (v/v), se congeló en nitrógeno
líquido y se guardó a -80°C como respaldo, para ser usados en las
transformaciones de Arabidopsis siguientes.
VI.6.4. Transformación de Arabidopsis.
Se pidieron al stock center ABRC semillas de la mutantes as1 en fondo Col
0, línea CS3374 para ser complementadas con el gen AtqAS1 bajo el control del
promotor 35S del virus del mosaico de la coliflor.
Se inocularon 5 ml de medio YEB con espectinomicina, carbenicilina y
rifampicina 100 mg/L se centrifugaron y se lavó el paquete celular 3 veces con
medio MS al 0.5X, 5 % de sacarosa y al final se resuspendió en 300 µL de medio
de infiltración con 0.02 % de Silwet L-77 y se inocularon los botones florales de
plantas mutantes as1 y silvestres de Arabidopsis, Col 0, mediante la modificación
del método “Floral dip” reportada por Martínez-Trujillo et al., 2004. Los botones
florales se inocularon 3 veces a intervalos de aproximadamente 4 días, cuando se
incrementó el número de botones.
VI.6.5. Selección de líneas homocigotas con una sola inserción.
Una vez que las plantas infectadas llegaron al termino de su ciclo y que las
semillas están maduras se cosecharon, se esterilizaron y se sembraron en medio
MS al 0.1X conteniendo 20 mg/L de fosfinotricina para seleccionar las líneas
transformadas que contienen el gen de resistencia a fosfonitricina y se
mantuvieren hasta dos semanas después de su germinación en un cuarto de
crecimiento a 25°C y 16h de luz por 8h de obscuridad. Una vez seleccionadas las
plantas resistentes T0 se pasaron a tierra y se llevaron a invernadero, nuevamente
se cosecharon sus semillas y se seleccionaron las líneas que tuvieran
descendencia en donde hubiera 3 plantas vivas por cada muerta T1, para
seleccionar las que sólo tuvieran una sola copia del gen AtqAS1. Nuevamente se

28
volvieron a sembrar algunas descendientes de estas líneas T1 en medio de
selección (20 mg/l de fosfinotricina) para buscar líneas que todas las semillas
sobrevivan debido a que sean homocigotas para el gen AtqAS1 de esta manera
buscamos seleccionar plantas que únicamente tengan una sola inserción y que
sean homocigotas para el gen AtqAS1. Este método se llevó a cabo partiendo de
plantas silvestres para llevar a cabo expresión constitutiva del genAtqAS1 y
partiendo de la mutante as1 para realizar su complementación.
VI.7. Obtención de la secuencia completa del cDNA de AtqGlsA.
Por análisis realizados previamente (Abraham-Juárez, 2010. Tesis de
doctorado) se sabia que el gen GlsA se encontraba incompleto en todas las clonas
de los bancos que contenían parte de su secuencia, por lo que se escogió una
clona para poder diseñar oligonucleótidos específicos y poder amplificar la parte
de la secuencia faltante usando el kit “GeneRacer. For full-length, RNA ligase-
mediated rapid amplification of 5´ and 3´ cDNA ends (RLM-RACE)” de Invitrogen.
VI.7.1. RACE 5’.
La reacción de RACE 5’ consiste de varios pasos en los que se busca
asegurarse que se van a amplificar sólo los transcritos, específicos del gen
seleccionado, que se encuentren completos.
Para comenzar la reacción RACE se extrajo RNA total con Trizol® de tejido
de la etapa 3 de la formación de bulbilos, ya que es en esta etapa donde se
encuentra una mayor expresión del gen AtqGlsA. Después se seleccionó el RNA
mensajero mediante Dynabeads, siguiendo el protocolo del fabricante. Una vez
teniendo el mensajero se sometió a una defosforilación por acción de la enzima
fosfatasa intestinal de ternera (CIP), para seleccionar únicamente transcritos que
tengan 7-metil-guanosina trifosfatada (Cap) y cola de poli A, eliminando
mensajeros truncados y RNA que no sea mensajero asegurando de esta manera
que todos los mensajeros restantes estén completos. A continuación se elimina el
CAP mediante la pirofosfatasa ácida de tabaco (TAP) para poder ligarle al
mensajero una secuencia conocida de RNA (oligo RACE 5’), para poder usarla

29
como sitio de unión de un oligonucleótido especifico al amplificar por PCR,
después se realiza una transcripción reversa usando la enzima SuperScript™ III
RT y el oligonucleótido dT para obtener la primera cadena lista para RACE, que
contiene sitios conocidos en los extremos 5’ y 3’. Para obtener el extremo
desconocido 5’ se amplificó por PCR usando como oligonucleótido directo el oligo
RACE 5’ (homólogo al RACE 5’ de RNA) y como reverso un oligonucleótido
especifico para el gen de interés (oligo GSP), también se realizó una segunda
amplificación usando oligonucleótidos anidados para amplificar la secuencia
desconocida.
Teniendo una banda seleccionada en gel de agarosa al 1 %, se procedió a
cortarla y a purificar el DNA usando el kit “GFXTM PCR and gel band purification
kit” de General Electric y después de verificar su calidad, se clonó en el vector
pGEM T-Easy de Promega, se transformó E. coli por electroporación, se
seleccionaron colonias resistentes a ampicilina, se crecieron en medio liquido y se
extrajo DNA plasmídico, para mandar a secuenciar. Después los resultados de la
secuenciación se alinearon con sus ortólogos de A. thaliana y L. longiflorum y se
identificaron sus dominios para saber si ya se contaba con la secuencia completa.
VI.7.2. Clonación del gen AtqGlsA. Una vez conociendo la secuencia completa del gen se identificó el marco de
lectura abierto (ORF) del AtqGlsA y se diseñaron oligonucleótidos específicos para
amplificar el ORF completo, una vez teniendo una banda consistente con el
tamaño del ORF completo, en gel de agarosa al 1 %, se cortó la banda, se purificó
y se clonó en el vector PCR8®/GW/TOPO® de Invitrogen, el cual cuenta con la
tecnología Topo® y Gateway®, lo que permite clonar un fragmento de PCR y
después pasarlo a un vector de salida por recombinación LR, también tiene
resistencia a espectinomicina y los sitios de secuenciación universales M13 Fig.9.

30
Fig.9. Vector PCR8®/GW/TOPO®. Vector usado para clonar las bandas del RACE 5’.
VI.8. Análisis de expresión por RT-PCR de los genes endógenos de Arabidopsis AS1 y KNAT1.
Para realizar un análisis de la posible forma en que AtqAS1 está afectando
la expresión de los genes AS1 y KNAT1 se realizaron reacciones de RT-PCR con
oligonucleótidos específicos, para lo cual se tomó tejido de hoja de planta
silvestre, de las mutantes knat1, as1, la sobreexpresante KNAT1, la planta
expresando constitutivamente AtqAS1 en fondo silvestre y la planta expresando el
gen AtqAS1 en fondo mutante as1 (complementada). De estos tejidos se extrajo
RNA utilizando el método de Trizol (Invitrogen), se sintetizó cDNA y se realizó la
amplificación de los genes por PCR, usando la actina como control de carga.

31
Tabla.6. Oligonucleótidos específicos usados para el análisis por RT-PCR en A.
thaliana.
VI.9. Análisis de explantes in vitro con el gen AtqAS1.
VI.9.1. Establecimiento de explantes in vitro.
Para tratar de transformar A. tequilana se estableció in vitro tejido del
meristemo de la planta en medio de multiplicación Eriksson (Eriksson, 1965) con
compuestos orgánicos de Linsmaier y Skoog (Linsmaier y Skoog, 1965)
conteniendo auxinas y citocininas como reguladores de crecimiento estandarizado
para Agave por Nava (1988), para lo cual se seleccionaron hijuelos de rizoma de
aproximadamente 1 a 2 años de edad, se les cortó la raíz y las hojas dejando
solamente la piña de aproximadamente 8cm de diámetro, se enjuagó con agua
para eliminar la tierra, se fue cortando la parte externa de la piña hasta dejar un
cilindro del centro de aproximadamente de 3cm de diámetro y 4cm de largo, se
Nombre ID Secuencia
KNAT1-F 5´-CGATGTTGAAGCCATGAAGG-3´
KNAT1-R 5´-GCTGTTGTCGAGCCTCAAAG-3´
AS1-F 5´-CTGCGCCTCAACCGCCAATC-3´
AS1-R 5´-CCTTACATTACATTACAAGTTAC-3´
AtqAS1-F 5’-ATCAAGAAGGGGTCTCTCACC-3’
AtqAS1-F 5’- CTTCCGTTGCTCTGACTCTGT-3’
ACT2-F 5´-GTACAACCGGTATTGTGCTGGAT-3´
ACT2-R 5´-GCTTGGTGCAAGTGCTGTGATTTC-3´

32
dejaron 2 horas en solución fungicida (Benomilo 1.2 g/l, Benlate 1.2g/l y Ridomil
bravo 3g/l), se cambiaron a una solución fungicida nueva y se dejaron 5 horas con
agitación ligera, en campana en condiciones asépticas se dejaron en cloro al 40 %
(v/v) de blanqueador comercial 20 minutos, luego en cloro al 20 % (v/v) 10
minutos, al 10 % (v/v) 5 minutos hasta dejarlos solamente en agua, se enjuagaron
con agua estéril 3 veces, 5 minutos cada vez, luego se cortó el tejido hasta dejar
un cilindro de aproximadamente 1.5cm de diámetro y se cortaron rodajas de 3mm
de grosor las cuales se pusieron en el medio de multiplicación.
VI.9.2. Infección de los explantes.
Una vez teniendo establecidos los explantes, se infectaron dejando caer en
cada explante 1 a 2 gotas de la solución de infiltración, preparada de la misma
manera que para infectar A. thaliana pero con acetosiringona, conteniendo
Agrobacterium transformada con el gen AtqAS1 bajo el control del promotor 35S,
se incubaron en obscuridad a 28°C por 48 horas, en condiciones asépticas, se
lavaron los explantes con antibiótico cefotaxima a 500mg/L, para eliminar el
Agrobacterium, se enjuagaron con agua estéril y se pasaron a medio de
multiplicación con cefotaxima a 250mg/L, se dejaron crecer los brotes 3 semanas,
se separaron, se observaron en el estereoscopio de fluorescencia y se pasaron a
medio de selección con fosfinotricina.

33
VII. RESULTADOS.
VII.1. Selección de genes.
Para este estudio se seleccionaron dos genes, el AtqAs1 y el AtqGlsA
debido a que se ha reportado que sus ortólogos están relacionados con la
formación de meristemos y además el AtqGlsA se encontró únicamente en las
bibliotecas de cDNA que se realizaron a partir de tejido tomado durante la
formación de bulbilos, lo cual lo hace un excelente candidato para este estudio.
VII.2. Análisis de expresión por RT-PCR del gen AtqAS1.
Los análisis de expresión del gen AtqAS1 mostraron que en A. tequilana la
expresión del gen se incrementa conforme avanza el desarrollo de los bulbilos,
además de que en general su expresión se incrementa en tejidos en desarrollo
como lo son los meristemos, ovario y anteras, lo cual sugiere tener un papel en la
división y diferenciación celular, además, concuerda con lo observado en A.
thaliana. Además se realizó el análisis con el gen AtqKNOX1 ya que se sabe que
el AtqAS1 puede interactuar negativamente con algunos genes de esta familia
(Hay et al., 2006) y se pudo observar que para raíz, ovario, antera y hoja así es.
Sin embargo parece haber una expresión similar en los meristemos jóvenes,
reproductivos y de umbela Fig.10.
Fig.10. RT-PCR de AtqAS1 y AtqKNOK1 en A. tequilana. T0-T5 etapas en la formación de bulbilos. R, raíz. O, ovario. A, anteras. H, hoja. MJ, meristemo joven. MR, meristemo de la raíz. MU, meristemo de umbelas.

34
VII.3. Ensayo de Hibridación in situ de AtqAS1. El resultado obtenido en los análisis de expresión por RT-PCR mostró que
AtqAS1 aumenta su expresión en las últimas tres etapas colectadas de la
formación de bulbilos y en meristemo reproductivo y de umbelas. Se observó el
mismo patrón en la expresión de AtqKNOX1, sin embargo esta técnica no permite
determinar a nivel tisular el lugar de la expresión. Por lo que fue necesario realizar
un ensayo de Hibridación in situ, para identificar en qué parte de los meristemos
se expresa específicamente AtqAS1 y establecer si existe la relación de expresión
excluyente con respecto a los transcritos de AtqKNOX1.
Como un primer ensayo para el establecimiento de las condiciones para la
Hibridación in situ de AtqAS1, se tomaron muestras de bulbilos en la etapa T5,
meristemos de umbelas y meristemo apical reproductivo. La Fig.11 muestra que la
expresión con la sonda antisentido se localiza en primordios de órganos laterales,
con la sonda sentido no se observó expresión. Aunque hace falta realizar el
ensayo con muestras de tejidos de todas las etapas del desarrollo de bulbilos y
determinar el lugar de la expresión durante las etapas tempranas.
Fig.11. Detección del mRNA de AtqAS1 en la Hibridación in situ. La detección se hizo con
digoxigenina, produciendo coloración morada, las puntas de flecha indican los sitios de expresión.
La escala en todas las imágenes corresponde a 100 µm.

35
La expresión de AtqAS1 se observa en hojas jóvenes en la etapa T5 de
formación de bulbilos, en cambio AtqKNOX1 no se expresa en este tejido
únicamente en el domo meristemático, consistente con los reportes en Arabidopsis
y maíz (Hake et al., 2004). Así mismo en meristemos de umbelas se observó una
clara expresión de AtqAS1 en primordios de órganos laterales.
VII.4. Identificación de la clona correspondiente al cDNA completo de AtqAS1.
En la base de datos de los bancos realizados se encontraron dos
secuencias incompletas independientes del gen AtqAS1, las cuales están
contenidas en las clonas aamhflovo009_n08 y aamhmqmeq011_g04, que se
encontraron en tejido de flor y meristemo reproductivo respectivamente, se extrajo
DNA plasmídico se realizó la digestión para liberar el inserto y se separó en gel de
agarosa al 1 % Fig.12.
Fig.12. DNA plasmídico y Digestion de las clonas conteniendo el gen AtqAs1. A DNA plasmídico de las clonas de los bancos conteniendo el gen AtqAs1. B Digestión de las mismas clonas con la enzima BsrGI, M.P marcador de peso molecular 100pb (Invitrogen). Flovo, Clona encontrada en tejido de ovario. Meq, clona encontrada en tejido de meristemo reproductivo.
M.P. M.P. Flovo Meq Flovo Meq
900pb
1300pb
A B

36
Se eligió la clona aamhmqmeq011_g04 para mandarla a secuenciar ya que
contiene el fragmento más grande y comparando la secuencia con su ortólogo de
Arabidopsis se esperaría un fragmento de aproximadamente 1157 pb.
Una vez obtenida la secuencia se comparó mediante alineamientos, usando
el software Clustal W (Larkin et al., 2007), con las secuencias del gen AS1 de
Arabidopsis y de maíz, luego se identificó su ORF Fig.13 y se tradujo a secuencia
de aminoácidos para poder realizar un nuevo alineamiento de aminoácidos, se
identificó su domino MYB de unión a DNA Fig.14 y se obtuvo su porcentaje de
identidad.
>AtqAS1
TTGGTGTTCTGTGAGATGCCCCAGGTTTTCTCATCAATAAAAAAAAAAAAGAAATGTAGATTTATTCCTTCATTAATAACAATTTAGTGATTTTCCTGTTCAGATCCGTCGGCGATGGGCGGCGGCGGCAGCAG ATGAAGGAGAGACAGCGGTGGGGATCTGAGGAAGATGGGATCCTGCGGGCCTACGTGAAG M K E R Q R W G S E E D G I L R A Y V K CAGTACGGCCCCCGCGAGTGGCACCTCGTCTCGCAGCGCATGAACGTCGCCCTCAACCGT Q Y G P R E W H L V S Q R M N V A L N R GACGCCAAGTCCTGCCTCGAGCGCTGGAAGAACTACCTCAAGCCCGGCATCAAGAAGGGG D A K S C L E R W K N Y L K P G I K K G TCTCTCACCGAGGACGAGCAGAGGCTCGTCATCCGCCTGCAGGCCAAGCACGGCAACAAG S L T E D E Q R L V I R L Q A K H G N K TGGAAGAAGATCGCCGCCGAGGTCCCCGGCCGCACGGCCAAGAGGCTCGGCAAGTGGTGG W K K I A A E V P G R T A K R L G K W W GAGGTCTTCAAGGAGAAGCAACAGAGGGAGTTGAAAGAGAGCGGTGTCAGTCCTGCGATT E V F K E K Q Q R E L K E S G V S P A I GAGCCCAGCAAGTACGATCGCATTTTGGAGAATTTCGCCGAGAAATTGGTAAAAGAGAGG E P S K Y D R I L E N F A E K L V K E R CAAATTGTGATGCCGACGCCCATGCTGCCCCCTTGGCTTTCGACCAATTGCCCCAGTGGC Q I V M P T P M L P P W L S T N C P S G GGTGGCGGTGGCGGCAGCAGCCCTTCCGTTGCTCTGACTCTGTCGCCGTCCACTGGGCCG G G G G G S S P S V A L T L S P S T G P CCGCCCCCTGCGGCAGCTGGCCCTGGGCCTTGGGGGAACGGGGGCCCCAGCAGCAGCAGC P P P A A A G P G P W G N G G P S S S S AGCAGCAGCATTGTCCTGTCCGAGCTCGGGGAGTGCTGCAGGGAGGTGCAGGAGGGGCAC S S S I V L S E L G E C C R E V Q E G H CAGGCGTGGGCCGCGCACAGGAAGGAGGCGGCGTGGAGGCTGAACAGGGTGGAGCTGCAG Q A W A A H R K E A A W R L N R V E L Q CTGGAGTCAGAGAAGGCCTGCAAGAGGAGGGAGAAGATGGAGGAGGTGGAGGCGAAGATA L E S E K A C K R R E K M E E V E A K I CGGGCGCTCAAAGAGGAGGAGAAGGCGGTGCTGGAGAGGATCGAGGCCGAGTACAGGGAG R A L K E E E K A V L E R I E A E Y R E

37
CAGGTGTTGGCGTTGAGGAGGGACGCGGAGGCCAAAGAGCAGAAGCTGGCAGAGCAGTGG Q V L A L R R D A E A K E Q K L A E Q W GCGGCGAAGCACGCTAGGCTGGGCAAGTTGATGGAGCAGCTGGGCTGTCATCACCAGTGG A A K H A R L G K L M E Q L G C H H Q W ATGCCGGCCGCGGCCGAGGCCAACGGCCGGTGA M P A A A E A N G R * GGCCTTTTTTACGAGGATTTCTTGTTCAGTGTTTTAGTTTTTTACTTGGTGTATGATGATGGATTGCCTTGTTGGAAAAACTCTTTATGTTG
Fig.13. Secuencia completa del cDNA de AtqAS1 con el ORF traducido.
AS1arabidopsis 1 MKERQRWSGEEDALLRAYVRQFGPREWHLVSERMNKPLNRDAKSCLERWKNYLKPGIKKGSLTEEEQRLVIRLQEKHGNKWKKIAAEVPGRTAKRLGKWW 100AS1AGAVEcompleta 1 MKERQRWGSEEDGILRAYVKQYGPREWHLVSQRMNVALNRDAKSCLERWKNYLKPGIKKGSLTEDEQRLVIRLQAKHGNKWKKIAAEVPGRTAKRLGKWW 100RS2Zeamays 1 MKERQRWRPEEDAVLRAYVRQYGPREWHLVSQRMNVALDRDAKSCLERWKNYLRPGIKKGSLTEEEQRLVIRLQAKHGNKWKKIAAEVPGRTAKRLGKWW 100
AS1arabidopsis 101 EVFKEKQQREEKESNK---RVEPIDESKYDRILESFAEKLVKERSNVVPAAAAAATVVMANSNGGFLHSEQQVQPPNPVIPPWLATSN-NGNNVVA---- 192AS1AGAVEcompleta 101 EVFKEKQQRELKESG----VSPAIEPSKYDRILENFAEKLVKER---------------------------QIVMPTPMLPPWLSTNCPSGGGGGG---- 165RS2Zeamays 101 EVFKEKQQRELRDSRRPPPEPSPDERGRYEWLLENFAEKLVGERPQQAAAAPS------------------PLLMAAPVLPPWLSSNAGPAAAAAAAVAH 182
AS1arabidopsis 192 ---RPPSVTLTLSPSTVAAAAPQPPIPWLQQQQPERAENGPGGL----VLGSMMPSCSGSSESVFLSELVECCRELEEGHRAWADHKKEAAWRLRRLELQ 285AS1AGAVEcompleta 165 ---SSPSVALTLSPST----GPPPP-----------AAAGPG------PWGNGGPSSSSSS-SIVLSELGECCREVQEGHQAWAAHRKEAAWRLNRVELQ 240RS2Zeamays 183 PPPRPPSPSVTLSLAS-AAVAPGPPAPAPWMPDRAAADAAPYGFPSPSQHGGAAPPGMAVVDGQALAELAECCRELEEGRRAWAAHRREAAWRLKRVEQQ 281
AS1arabidopsis 286 LESEKTCRQREKMEEIEAKMKALREEQKNAMEKIEGEYREQLVGLRRDAEAKDQKLADQWTSRHIRLTKFLEQQMGCRLDRP--------- 367AS1AGAVEcompleta 241 LESEKACKRREKMEEVEAKIRALKEEEKAVLERIEAEYREQVLALRRDAEAKEQKLAEQWAAKHARLGKLMEQ-LGCHHQWMPAAAEANGR 330RS2Zeamays 282 LEMEREMRRREVWEEFEAKMRTMRLEQAAAAERVERDHREKVAELRRDAQVKEEKMAEQWAAKHARVAKFVEQMGGCSRSWSSATDMNC-- 370
Dominio MYB
Fig.14. Alineamiento de las secuencias de aminoácidos de AS1 de
Arabidopsis thaliana, AS1 de Agave tequilana y RS2 de Zea mays. Sombreado en
color anaranjado se indica la secuencia del dominio MYB.
Porcentajes de identidad en aminoácidos
Sequence 1: AS1 Arabidopsis Sequence 1: AtqAS1
Sequence 2: AtqAS1 Sequence 2: RS2 Zea mays
Identities: 61.0 % Identities: 56.4 %
Similarities: 61.0 % Similarities: 56.4 %
Tabla.7. Porcentaje de identidad entre AtqAs1 y sus ortólogos de Arabidopsis y de maíz.

38
De acuerdo a los resultados anteriores, se tiene la secuencia codificante
completa de AtqAS1, se aprecia que el dominio MYB está altamente conservado
en las secuencias de Arabidopsis, Agave y maíz y el porcentaje de identidad de
las secuencias en aminoácidos indica que AtqAS1 tiene mayor similitud a AS1 que
a RS2, a pesar de que maíz y Agave son monocotiledóneas.
VII.5. Expresión heteróloga.
Después de identificar una clona en las bibliotecas con el cDNA completo
de AtqAS1, se procedió a realizar una recombinación para transferir el cDNA de la
biblioteca con clonas Gateway® de entrada (pDONR222) a un vector Gateway® de
salida (pB7WG2D).
VII.5.1. Transformación de Agrobacterium tumefaciens con el vector pB7WG2D conteniendo el gen AtqAS1.
Después de realizar la recombinación se transformó E.coli mediante
electroporación se extrajo DNA plasmídico, se llevó a cabo la identificación del gen
de interés AtqAS1 PCR con oligonucleótidos específicos para comprobar que las
clonas fueran positivas y se separó en gel de agarosa al 1 % Fig.15.
Fig.15. DNA plasmídico y PCR de colonias de E.coli. A, DNA plasmídico de 4 colonias de E.coli B, PCR con oligonucleótidos específicos de AtqAS1 de las mismas clonas, M.P marcador de peso molecular 100pb (Invitrogen).
A B M.P. M.P. 1 2 3 4 1 2 3 4
355pb

39
Se seleccionó una clona positiva y con ese DNA plasmídico se transformó
la cepa GV2260 de A. tumefaciens por electroporación, se seleccionaron 3
colonias, se extrajo DNA plasmídico, se realizó PCR con oligonucleótidos
específicos para el gen AtqAS1 y el fragmento se separó en gel de agarosa al 1 %
Fig.16.
VII.5.2. Expresión constitutiva del gen AtqAS1 en Arabidopsis
thaliana.
Para analizar la función del gen de agave se realizó la expresión
constitutiva del gen AtqAS1 en A. thaliana. Se seleccionó la clona 3 para
transformar Arabidopsis para lo cual se infectaron 6 plantas silvestres con A.
tumefaciens conteniendo el gen AtqAS1 bajo el control del promotor constitutivo
35S del virus del mosaico de la coliflor, de las semillas recuperadas de las plantas
infectadas se sembraron aproximadamente 6,000 en medio de selección (20 mg/l
de fosfinotricina) y sólo se encontraron 40 líneas resistentes T0, esto corresponde
a 0.67 % de eficiencia de transformación. De las 40 líneas resistentes se
escogieron las 12 líneas que mostraron el fenotipo más visible, en base a los
reportes de la sobreexpresión de AS1 de Arabidopsis, cuyas características más
sobresalientes son hojas de la roseta alargadas y peciolos muy largos en
Fig.16. PCR de colonias de A. tumefaciens. PCR con oligonucleótidos específicos de AtqAS1 de 3 clonas, M.P marcador de peso molecular 100pb (Invitrogen). C+, control positivo.
M.P. 1 2 3 C+
355pb

40
comparación con plantas WT. Se sembraron aproximadamente 200 semillas de la
descendencia de cada planta T1, para buscar segregación 3:1, encontrándose 5
líneas independientes con una sola inserción de cada línea. Se sembraron 12
plantas y de la descendencia de estas líneas se sembraron nuevamente
aproximadamente 200 semillas, en medio de selección para identificar líneas
homocigotas T2. Estas líneas se usaron para los análisis de expresión por RT-
PCR.
Se pudo observar que algunas (8 de las 40 líneas independientes en la T0)
de las líneas descendientes de las plantas infectadas, mostraron fenotipo similar al
de las plantas sobreexpresando el gen AS1 reportada previamente (Chua et al.,
2005) Fig.17.
VII.5.3. Complementación de la mutante as1 de A. thaliana.
VII.5.3.1 Descripción de la mutante.
La mutante as1 se encuentra en fondo Col 0 y tiene una mutación de
carácter recesivo del ORF por la eliminación de una G en la posición 691 del locus
AT2G37630 por rayos X, mostrando hojas de la roseta asimétricas y lobuladas,
bordes de la hoja ligeramente enrollados hacia abajo y algo triangulares (Ori et al.,
2000), roseta pequeña, silicuas levemente afectadas con apariencia desigual,
A B C
Fig.17. Plantas de A. thaliana. A planta de A. thaliana silvestre, B Planta de A.thaliana sobreexpresando AS1 (Chua et al., 2005), C Planta expresando constitutivamente el gen de agave AqtAS1 mostrando fenotipo similar a las plantas sobreexpresantes AS1.

41
replum alargado por contener más células que en la silvestre, (Alonso-Cantabrana,
et al., 2007), Fig.18.
VII.5.3.2 Complementación de la mutante as1.
Para la complementación se infectaron los botones florales de la mutante
as1 con A. tumefaciens conteniendo el gen AtqAS1 bajo el control del promotor
35S del virus del mosaico de la coliflor, desde la descendencia T1 se logró
observar plantas (18 de 40 líneas independientes en la T0) que muestran un
fenotipo silvestre e incluso en algunos casos se pudo observar fenotipo similar al
de plantas sobreexpresando el gen AS1, Fig.19.
Fig.18. Comparación de fenotipos WT y as1. A planta de A. thaliana silvestre, B Planta de A.thaliana mutante as1.
A B
Fig.19. Fenotipo de la mutante as1 complementada con AtqAS1. A planta de A. thaliana silvestre, B Planta de A.thaliana mutante as1, C Planta mutante as1 complementada con el gen de Agave AtqAS1 mostrando fenotipo similar a las plantas sobreexpresantes AS1. Las plantas pequeñas en C son mutantes as1 que no fueron complementadas.
A B C

42
VII.6. Análisis de expresión por RT-PCR del gen AtqGlsA.
A partir de la investigación previa a este trabajo (Abraham-Juárez, 2010.
Tesis de doctorado) se determinó el perfil de expresión del gen AtqGlsA por RT-
PCR semicuantitativo y en tiempo real Fig.20.
Fig.20. Análisis de expresión por RT-PCR semicuantitativo, A y en tiempo real,
B de AtqGlsA, durante las seis etapas colectadas del desarrollo de bulbilos y otros
órganos de la planta.
Se puede observar que la expresión del gen AtqGlsA es exclusiva de la
formación de bulbilos y su expresión es mayor en la etapa 3, cuando comienza la
proliferación celular que dará origen a los bulbilos.
A
B

43
VII.7. Obtención de la secuencia completa del cDNA de AtqGlsA.
Se sabía que la secuencia del gen AtqGlsA estaba incompleta en las clonas
de las bibliotecas en las que se identificó la secuencia, las clonas fueron
encontradas en tejido de la formación de bulbilos correspondientes a las etapas 2
y 3 (ajswble02014_h03 y ajswble02013_f10). A continuación se muestra el
alineamiento de los fragmentos de estas secuencias proteinitas con sus ortólogos
de L. longiflorum y de A. thaliana (Abraham-Juárez, 2010).
Fig.21. Alineamiento en aminoácidos de las secuencias de AtqGlsA de A. tequilana y sus ortólogos de L. longiflorum y A.thaliana.

44
Se observa que las secuencias de Agave tequilana no están completas
comparándolas con la secuencia de L. longiflorum, a la secuencia más grande
correspondiente a la clona ajswble02013_f10 le faltan aproximadamente 234
aminoácidos hacia el extremo 5´.
VII.7.1. RACE 5’.
Para completar la secuencia se recurrió a una reacción RACE 5’ para lo
cual se extrajo RNA total con Trizol (Invitrogen) a partir de tejido de la etapa 3 de
la formación de bulbilos, se purificó el RNA mensajero utilizando el kit de
Dynabeads (Dynal) y se sintetizó el cDNA mediante el RACE5’.
Una vez teniendo el cDNA del RACE5’ se realizó una primera prueba de
amplificación para ver si el cDNA se encontraba íntegro, esta prueba consistió en
realizar PCR sin uno de los oligonucleótidos para comprobar que no se estuvieran
obteniendo artefactos que pudieran confundir los resultados, así como amplificar
una secuencia interna del gen con oligonucleótidos específicos para comprobar
que el mensajero del gen realmente se encuentre en el cDNA del RACE y usando
oligonucleótidos para el gen de actina como control positivo Fig.22 y Fig.23.
Fig.22. Prueba de integridad del CDNA.M.P marcador de 100pb, 1 amplif icaciónusando oligos gene RACER 5’ y GSP, 2Control negatico sin cDNA, 3 Sin el oligoGSP, 4 Sin el oligo Gen RACER 5’.
1 2 43M.P

45
Fig.23. Prueba de integridad del CDNA. M.P marcador de100pb, 1 amplif icación usando oligos gene RACER 5’ y GSP, 2gene RACER 5’ y GSP nested, 3 gene RACER y GlsA interno, 4GlsA directo y reverso internos 5’, 5 actina, 6 gene RACER concDNA diferente del RACE, 7 GlsA internos cDNA diferente delRACE, 8 actina cDNA diferente del RACE.
1 2 43M.P 5 6 87
cDNAcDNA RACE
Para realizar la amplificación de la parte de la secuencia desconocida se
realizó un primer PCR con las condiciones como siguen:
Tabla 8. Condiciones de amplificación del RACE.
Reactivos µl Proceso Temp. [°C]
Tiempo Ciclos
Buffer 10X 2.5 Desnaturalización 94 4 min. 1
MgCl2 (25mM) 1.0 Desnaturalización 94 30s 25
dNTP’s (10mM) 0.5 Alineamiento 60 30s
Oligo gene RACER 5’
(10mM)
1.5 Extensión 72 1min.
Oligo GSP (10mM) 1.0 Elongación final 72 10min. 1
cDNA RACE 0.5 Mantenimiento 4 ∞ 1
Taq DNA pol 0.4
H2Odd 17.6
∑ 25.0
395 pb GlsA 455 pb ACT2

46
Después se repitió un PCR usando los oligonucleótidos gene RACER5’ y
GSP anidados, se corrió en gel de agarosa al 1 % y se encontraron 3 bandas de
aproximadamente 450 pb, 800 pb y de 1100 pb, se intentó cortarlas, purificarlas y
clonarlas para mandarlas secuenciar, sin embargo por la poca cantidad de DNA
que se recuperó, en un primer intento sólo se logró clonar y secuenciar la banda
más abundante de 450 pb. Fig.24.
Fig.24. PCR RACE5’Anidado M.Pmarcador de 100pb, 1 oligos geneRACER5’ y GSP, 2 oligos geneRACER5’ y GSP anidados
M.P 1 2M.P
1100pb800pb450pb
Aunque con la secuenciación de la banda de 450 pb, la secuencia conocida
del gen AtqGlsA se incrementó en 346 pb no fue suficiente para completar toda la
secuencia del gen, ya que seguían faltando aproximadamente 500 pb, incluso, la
mayor parte del dominio J, el cual se sabe que es esencial para el funcionamiento
del gen GlsA, también faltaba.

47
Por lo anterior se procedió a modificar las condiciones del PCR para poder
cortar la banda de 1,100 pb, clonarla y mandarla secuenciar, las modificaciones
que se hicieron fueron: aumentar la cantidad de dNTP’s 10 mM, de 0.5 a 1.0 µl, el
tiempo de alineamiento de 1.0 a 2.0 minutos y el número de ciclos se aumentó a
40, con lo que se logró tener una banda de aproximadamente 1,100 pb más
concentrada y prácticamente única, por lo que fue más fácil de cortar, purificar,
clonar y secuenciar Fig.25.
Con este nuevo producto de PCR se logró conocer la secuencia completa
del gen AtqGlsA Fig. 26, comparándola con sus ortólogos de L. longiflorum, A.
thaliana y V. carteri, como se observa en la Fig.27.

48
>AtqGlsA GAGACCTCTTTCGCGCAGCCCCAGCCCTAGCTGACAGACTCCTCCTCTCTCCTCTCCTCTTTCCTTTCGCTGCCGCCGGCGCCCCTTCGCTGCTCAAACCCTAAATCAGCCACCCAGGTTGTTCATCTGAAGAAACTGGTTCACGGAGTGATATTTGCTACTTTGAACACCTTTCATTATCTGTAGTTAATACTGCTTCAGCGTT ATGGATGCCCAGACAAGTACCCTGCTAATCACATATTCGCCAGAGATTCTTGATGGTGAA M D A Q T S T L L I T Y S P E I L D G E CCTATCCTTGTCTCATCAAATTCCCTCCCTGTGAAAGCACCACATTTTGAACCAGCTGGA P I L V S S N S L P V K A P H F E P A G CATGCATTTCATTTAGCTGCACTTAAACTTTCGGGTTTTGATGAGGAAGAAGATGCAGAC H A F H L A A L K L S G F D E E E D A D ACAGATGGGCAAAGTGAAAAGTCAGATGACAAAGGACAAACTTCTAAGCCGTCTTTTGAT T D G Q S E K S D D K G Q T S K P S F D TCCTATAGCACTAAAGGCAAAAAGAAATCAGGCGATGGAAGCAAGCAACAAGATCATTAT S Y S T K G K K K S G D G S K Q Q D H Y GCACTATTAGGCTTGGGCCATTTACGGTTTTTGGCTACCGAGGAACAGATACGAAAGAGT A L L G L G H L R F L A T E E Q I R K S TACCGTGAGACTGCCTTGAGGCATCATCCTGACAAGCAGGCTGCTATTCTTCTCAATGAA Y R E T A L R H H P D K Q A A I L L N E GAAACAGAAGATGCAAAGCAGGCAAAGAAGGACGAGATAGAAAACCACTTCAAAGCCATT E T E D A K Q A K K D E I E N H F K A I CAAGAAGCATATGAGGTTCTAATTGATCCTGTGAAGAGAAGAATTTATGACTCTACAGAT Q E A Y E V L I D P V K R R I Y D S T D GAGTTTGATGATGAAATTCCAACTGACTGTGCCCCGCATGACTTTTTCAAGGTATTTGGT E F D D E I P T D C A P H D F F K V F G CCGGCATTTATGAGAAACGGTAGGTGGTCTGTTAATCTTCCAGCTCCCTCTCTAGGGGAT P A F M R N G R W S V N L P A P S L G D GAGAAAACTTCATTGGAAGAAGTGGATAGTTTCTATAATTTCTGGTATGCTTTTAAGAGC E K T S L E E V D S F Y N F W Y A F K S TGGAGGGAATTCCCGCACGCAGACGAGTTTGATCTCGAACAAGCTGAATCTCGGGACCAT W R E F P H A D E F D L E Q A E S R D H AAGAGATGGATGGAGAGGCAAAATGCAAAGCTACGAGAAAAGGCCAGGAAGGAAGAGCAT K R W M E R Q N A K L R E K A R K E E H GCACGGGTCCGGAATCTAGTTGACAATGCATACAAGAAAGATCCAAGAATACTTCGGAGG A R V R N L V D N A Y K K D P R I L R R AAAGAGGAAGAAAAGGCAGAGAAGCAAAGGAAAAAGGAGGCAAAATTTTTGGCTAAGAAG K E E E K A E K Q R K K E A K F L A K K TTGCAGGACGAAGAGGCAGCAAGAGCTGCGGAAGAGGAGAGACGCCGGAAAGAAGAGGAT L Q D E E A A R A A E E E R R R K E E D GATAAGAAAGCAGCTGAAGCGGCTTTGAACCAGAAGAAGCAGAAGGAGAAGGACAAGAAG D K K A A E A A L N Q K K Q K E K D K K CTCTTGCGCAAAGAAAGAACTCGCTTACGTACGCTCTCTTCATCTGTTGTGTCGAAAACC L L R K E R T R L R T L S S S V V S K T TCTAGTGATATTTCAGAAGATGCTGTGGAGAGTCTATGCATGTCATTTGACATGGATCTG S S D I S E D A V E S L C M S F D M D L CTTAGATGTCTGTGTGATGAGATGGAGGGGAAGGAAGAAAATGAGAGAGGTCAACTGCTA L R C L C D E M E G K E E N E R G Q L L AAAGATGCACTGAACAAAGGGAACTCAAATGGAACAAAACAGGAAGAGAAGAGTTCACAT

49
K D A L N K G N S N G T K Q E E K S S H TCAAACGGTTCTCTAGGTTCAGGTTCGAAAGTAAATGGAGTTGTGACTCAGGTTAAGCCA S N G S L G S G S K V N G V V T Q V K P GGAAGCGTGCTAAGCAAACACGAGAAGAAAGAGAAGCCTTGGGGAAAAGAAGAAATTGAA G S V L S K H E K K E K P W G K E E I E TTGCTGAGAAAGGGGATGCAGAAATATCCAAAGGGAACATCAAAGAGGTGGGAGGTTATC L L R K G M Q K Y P K G T S K R W E V I TCAGAGTACATCGGCACTGGGAGATCAGTTGAAGAGATCCTGAAGGCCACAAAGACTGTT S E Y I G T G R S V E E I L K A T K T V CTCCTCCAGAAACCAGATTCTGCAAAAGCTTTTGATTCTTTCCTTGAGAAAAGAAAGCCT L L Q K P D S A K A F D S F L E K R K P GCACCGTCCATTGCGTCACCTCTGACAACTAGAACTGAGTCAGACGGTGTACCTCTTACT A P S I A S P L T T R T E S D G V P L T GGAATTCAAGAGAGCTCTTCTGAAGTCTCTAAGCAGTCTTCAACCGGTGATGGCACTTCC G I Q E S S S E V S K Q S S T G D G T S CAGAATTCAGTAGGTGGACAGGTTCACGCACCTAATGGAGTTTCCTCAAGTGCCGAACAA Q N S V G G Q V H A P N G V S S S A E Q GACACATGGTCAGCTATTCAAGAAAGAGCGCTTATTCAGGCTCTAAAGACATTCCCAAAG D T W S A I Q E R A L I Q A L K T F P K GAAACCGCCCAACGGTGGGAGAGAGTTTTTGCGGCAGTTCCTGGAAAAACTGTTAACCAA E T A Q R W E R V F A A V P G K T V N Q TGCAAAAAGAAATTCGCATTGATGAAGGAGAGCTTCCGCAGCAAGAAAAATGCTGATTAA C K K K F A L M K E S F R S K K N A D *
TATGCATGTTTTACATTACATAGAGGGAGGGGTTTTGAGAATAGAATATATCGGGCTATAAGATTTAGTTTATTTTTGGCAGTTGAACTTTATTAACCATTTTATGGGAAAAAAAAAAAAAAAAAAAAAA
Fig.26. Secuencia completa del ORF de AtqGlsA en nucleótidos y en aminoácidos.
La secuencia codificante completa de AtqGlsA mide 1980 pb, produciendo una proteína de 660 aminoácidos, como se muestra en la figura 26.
Después se llevó a cabo el alineamiento de las secuencias de aminoácidos,
se lograron identificar todos los dominios completos incluyendo el dominio J el cual
se sabe es indispensable para las divisiones asimétricas y diferenciación celular,
además, estimula la actividad de proteínas, por medio de activación
postraduccional. Proteínas que contienen este dominio actúan en la
termoprotección de la célula doblando y previniendo la agregación de proteínas sin
plegamiento, está involucrado en la síntesis, dirección y translocación de proteínas
en variaos organelos, además el dominio J actúa junto con el dominio M, sin

50
GlsA Agave tequilana ORF 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - M D A QT S T L L I T Y S P E I L D G E P - - - I L V S S N S L 29GlsA Lilium longif lorum 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - M D A K S - C L L L T Y S S E V L N GV P - - - L L F A S N C L 28GlsA Volvox car ter i 1 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - M L L S R H L S L E F V E E L R P GT P D GR L V T C S V A G 31Arabidops is -thaliana60203 1 M L N H C D R N QGGY S L GV R C S S Y K L F I M P S R R S E S A I R L I T Y S E E L V D GK P - - - F F A F S N C L 57Arabidops is -thaliana67593 1 - - - - - - - - - - - - - - - - - - - - - - - - - M QS WG I N S A I K L L T Y S S E L K D GQA - - - L Y A S S N C H 32
GlsA Agave tequilana ORF 30 P V K A P H F E P A GH A F H L A A L K L S G F D E E E D A D T D GQS E K S - D D K GQT S K P S F D S Y S T - - - - 84GlsA Lilium longif lorum 29 P V K A V N Q E P A GH A F H D A A L K L C GL - F E E D V V A D D R S E S S - D D R GP A FM A S S D S Y R S - - - - 82GlsA Volvox car ter i 32 P V P Y GA K D P A GY Y F H L K A L QQA GL W E D P T P T E D A D A D A N GT A D A A A S A A A A A GA D S D K P P 91Arabidops is -thaliana60203 58 P V K A L N R E P A GH A F H S A A L K L H GC A E E P T D - D E GGD K K V GD D K E K E Y V P S F N S Y A N - - - - 112Arabidops is -thaliana67593 33 P V K A L N R E P A GH A F H S A A L K L R GC A K E A T S K N E D T D K K V P K E K D G E Y I P S Y D S H N I - - - - 88
GlsA Agave tequilana ORF 84 - - - - - K G - - - - K K K S GD GS K QQD H Y A L L GL GH L R F L A T E E Q I R K S Y R E T A L R H H P D K QA A 135GlsA Lilium longif lorum 82 - - - - - K S - - - - K K K S A GK S D QQD H Y A L L GL GH I R F L A T E E Q I R K S Y R E T A L K H H P D K QA A 133GlsA Volvox car ter i 92 P GV DWK GS K A R K R K A V K K K D GS D P Y S L L GL A N E RWT A S E A Q I R A A Y R K T C L E N H P D K A L I 151Arabidops is -thaliana60203 112 - - - - - K G - - - - K K K S G - - T QQQD H Y A L L GL S N L R Y L A T E D Q I R K S Y R E A A L K H H P D K L A - 160Arabidops is -thaliana67593 88 - - - - - K G - - - - K K K S G - - K L QH D H Y A L L GL GN L R Y L A T D D Q I R K S Y R D A A L K H H P D K L A T 137
GlsA Agave tequilana ORF 136 I L L N E E T E D A K QA K K D E I E N H F K A I Q E A Y E V L I D P V K R R I Y D S T D E F D D E I P T D C A P H D F 195GlsA Lilium longif lorum 134 L L L T E K T E A A K QA K K D E I E N H F K D I Q E A Y E V L I D P V R R R V Y D S T D E F D D E V P S D C A P QD F 193GlsA Volvox car ter i 152 N - - - - - - - V T D E A E R E R I V E H F K T I QD A Y D I L S D P A K R R E F D S T D E F D D T L P L QC D P K D F 204Arabidops is -thaliana60203 160 - - - - - - - - K A K E A K K D E I E S R F K A I Q E A Y E V L M D P T R R R I F D S T D E F D D E V P S D C L P QD F 212Arabidops is -thaliana67593 138 L L L L E E T E E A K QA K K D E I E S H F K L I Q E A Y E V L M D S T K R R I F D S T D E F D D K V P T D C A P QD F 197
GlsA Agave tequilana ORF 196 F K V F GP A FM R N GRWS V N L P A P S L GD E K T S L E E V D S F Y N FWY A F K S WR E F P H A D E F D L E QA 255GlsA Lilium longif lorum 194 F K V F GP A FM R N GK WS V V QP V P S L GD D K T S L E E V D N F Y D FWY A F K S WR E F P H A D E F E L E QS 253GlsA Volvox car ter i 205 L K V F GP A F R R N A RWS T V E P V P D V GD D S T P WGD V S K F Y D FWY T F K S WR E F P H P D E E D V E A A 264Arabidops is -thaliana60203 213 F K V F GP A F K R N A RWS V N QR I P D L GD E N T P L K D V D K F Y N FWY A F K S WR E F P D E E E H D L E QA 272Arabidops is -thaliana67593 198 F K V F GP A F K R N A RWS - N S P L P D L GD E N T P L K E V D R F Y S T WY T F K S WR E F P E E E E H D I E QA 256
GlsA Agave tequilana ORF 256 E S R D H K RWM E R QN A K L R E K A R K E E H A R V R N L V D N A Y K K D P R I L R R K E E E K A E K QR K K E A K 315GlsA Lilium longif lorum 254 E S R D H K RWM E R QN A K L R E K A R K E E Y A R V R S L I D N A Y K R D P R L L R R K E Q E K A E K QR R K E A K 313GlsA Volvox car ter i 265 E S R E H R RW I E R N N A K L R E K GK K E E GR R L R E F V D A A Y K H D P R V L R K K E E D R L E R E R K K A E K 324Arabidops is -thaliana60203 273 D S R E E R RWM E K E N A K K T V K A R K E E H A R I R T L V D N A Y R K D P R I V K R K E E E K A E K QQK K D A K 332Arabidops is -thaliana67593 257 E S R E E K RWM E R E N A R K T QK A R K E E Y A R I R T L V D N A Y K K D I R I QK R K D D E K A K K L QK K E A K 316
GlsA Agave tequilana ORF 316 F L A K K - - L QD E E A A R A A E E E R R R - K E E D D K K A A E A A L N QK K QK E K D K K L L R K E R T R L R T L 372GlsA Lilium longif lorum 314 Y M A K K - - L Q E E E A A R A V E E E R L R - K E E D E K K A A E A A V I N K K I K E K E K K L L R K E K T R L R T L 370GlsA Volvox car ter i 325 E E A R R R A Q E E E E A R K A A E E E A R R V A E E E A K R V A E E A R - - - K QR E A V K A QL K QL R K R L R T V 381Arabidops is -thaliana60203 333 I QA K K - - K Q E E D A A I A A E E E K R R - K E E E E K R A A E S A QQQK K T K E R E K K L L R K E R N R L R T L 389Arabidops is -thaliana67593 317 V M A K R - - QQ E E A A A A A I E E E K R R - K E E E A K R A A E A A QL H K R A K E R E K K L L R K E R S R L R V L 373
GlsA Agave tequilana ORF 373 S S S V V S K T S S D I S E D A V E S L CM S F DM D L L R C L C D EM E G - - - K E E N E R GQL L K D A L N K GN S 429GlsA Lilium longif lorum 371 S V S L V S D S L L D L T E D D V E K T C N S F GL E QL R N L C E GM E G - - - R E G I E R A QL L K A A V S GDM L 427GlsA Volvox car ter i 382 A E - - GS GGA R L V S E D D V E K L I T K L E P E A L A V L V E A L S S C D I S E G E T A A A A QK GL L T E A L R 439Arabidops is -thaliana60203 390 S A P L V A QR L L D I S E E D I E N L CM S L N T E QL QN L C D K M GN - - - K E GL E L A K V I K D GC N S S R N 446Arabidops is -thaliana67593 374 S A P V L S QR L L G I S D E H V E D L CM S L N T E QL R K L C D K M E N - - - K E GM A L A K V I K N GS N I D D D 430
GlsA Agave tequilana ORF 430 N GT K Q E E K S S H S N GS L GS GS K V N GV V T - - - QV K P GS V L S K H E K K E K P WGK E E I E L L R K GM 486GlsA Lilium longif lorum 428 E I S K K E P N D L K P N GS T N S GL K S N GS V T - - - S A K P V I MM S S Y E K K E K P WGK E E I E L L R K G I 484GlsA Volvox car ter i 440 G I D QR E E E A A R T R E A A K R E A E L A A K V A - - - - - - A R E E H R R K M A A M R EWS E E E L R L L D K A C 493Arabidops is -thaliana60203 446 - - D E A E S K - E K V S K K T N GGT E P T T R V S - - - - - - - QL D S S T QK K Q - - P WS K E E I DM L R K GM 494Arabidops is -thaliana67593 431 K I E I E E E E - V QV A V K QN GH I E A N GH V E A N GH V E A K V D T A T H E K K E K P WS K E E I DM L R K GT 489
GlsA Agave tequilana ORF 487 QK Y P K GT S K RW E V I S E Y I GT GR S V E E I L K A T K T V L L QK P D S A K A F D S F L E K R K P A P S I A S 546GlsA Lilium longif lorum 485 QK Y QK GT S R RW E V I S E Y I GT GR S V E E I L K A T K T V L L QK P D S S K A F D S F L E K R K P A K A I V S 544GlsA Volvox car ter i 494 N K F P M GT P K RW E A V A A F V R T - R T L D E V L L M V K D R QGA S A T RM K A Q E DWK GA QK K P A E V R A 552Arabidops is -thaliana60203 495 I K Y P K GT S R RW E V I S E Y I GT GR S V E E I L K A T K T V L L QK P D S A K A F D S F L E K R K P S A S I T S 554Arabidops is -thaliana67593 490 T K F P K GT S QRW E V I S E Y I GT GR S V E E I L K A T K T V L L QK P D S A K A F D S F L E N R K P A A S I N S 549
GlsA Agave tequilana ORF 547 P L T T R T E - - - - - - - - - - - - - - - - - - - - - - - - - S D GV P L T G I Q E S S S E V S K QS S T GD GT S Q 581GlsA Lilium longif lorum 545 P L T T R L E - - - - - - - - - - - - - - - - - - - - - - - - - S E GS T V E A GD A S S K S I P T P S L S - S S S P E 578GlsA Volvox car ter i 553 QA D T R A E A F T D V E V K L T G E A A I K L M QP A A K S A S GA T A A T A T A A A S A A P A A K A A A S GGS S T 612Arabidops is -thaliana60203 555 P L S T R E E L - - - - - - - - - - - - - - - - - - - - - - - - G E S - L P T M T T T T N A K P S K E T V V GK S S S S 589Arabidops is -thaliana67593 550 P L S T R E E L - - - - - - - - - - - - - - - - - - - - - - - - G E P I I P T K A H E D N N S T K T E T A E QN GK T K 585
GlsA Agave tequilana ORF 581 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - N S V GGQV H A P N GV S S S A E 599GlsA Lilium longif lorum 578 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - K P D GT P V S L P N GV P S V P E 596GlsA Volvox car ter i 613 GT P A A P A S P S S S S A R S K A A K T P A A T A T A A GA P S E GT T E A A A GA T T A S S A A A A K GV K K A A V 672Arabidops is -thaliana60203 589 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - QS S D N N G E - - - - V GGS S D 603Arabidops is -thaliana67593 585 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - E N N N S N GN S E P A A A S GS D 603
GlsA Agave tequilana ORF 600 QD T - - WS A I Q E R A L I QA L K T F P K E T - A QRW E R V F A A V P GK T V N QC K K K F A L M K E S F R S K K 656GlsA Lilium longif lorum 597 QD T - - WS A T Q E R A L I QA L K T F P K D V - N QRW E R V A A A I P GK T M N QC R K K F L S M K E D F R S K K 653GlsA Volvox car ter i 673 A D S GA WS E A Q E L A L V A A L K QC P K E L GA E RWD A V A V L V P GK T K A QC F K R F K E L R E A F R S K K 732Arabidops is -thaliana60203 604 A D S - - WS T V Q E R A L V QA L K T F P K E T - S QRW E R V A A A V P GK T M N QC K K K F A E L K E I I R N K K 660Arabidops is -thaliana67593 604 P D G - - WS A V Q E R A L V QA L K T F P K E T - N QRW E R V A T A V P GK T M N QC K K K F A D L K D V I R T K K 660
GlsA Agave tequilana ORF 657 N A D - - - - - - - - - - - - - 659GlsA Lilium longif lorum 654 S G - - - - - - - - - - - - - - 655GlsA Volvox car ter i 733 QA A G E GGGGGGD D GD D 748Arabidops is -thaliana60203 661 T GV - - - - - - - - - - - - - 663Arabidops is -thaliana67593 661 P T A - - - - - - - - - - - - - 663
Dominio J
***Residuos esenciales para la función del Dominio J
Dominio M
Dominios WR1 y WR2
***
embargo no se conoce la función de los dominios WR (repetidos de triptófano)
(Miller y Kirk, 1999) Fig.27.
Fig.27. Alineamiento del ORF completo de AtqGlsA y sus ortólogos de L. longiflorum, A. thaliana y V. carteri en aminoácidos. Se muestran los dominios funcionales.
La secuencia de AtqGlsA tiene mayor similitud a GlsA de L. longiflorum, con
un porcentaje de identidad en aminoácidos de 71.3 %.

51
Una vez conocida la secuencia completa del cDNA de AtqGlsA se procedió
a diseñar oligonucleótidos específicos para amplificar su ORF completo, clonarlo,
pasarlo a un vector de expresión y realizar su expresión constitutiva en A. thaliana.
Para amplificar el ORF completo se sintetizó cDNA a partir de la etapa 3 de
la formación de bulbilos y se realizó PCR usando el oligonucleótido directo ORF
GlsA dir: 5´AATACTGCTTCAGCGTTATGGATGC 3´ y el reverso ORF GlsA rev:
5´CAAAACCCCTCCCTCTATGTAATGT 3´, se corrió en gel de agarosa al 1 % y
sólo se observó una banda del tamaño esperado muy tenue por lo que se realizó
una reamplificación con los mismos oligonucleótidos, incrementando la cantidad
de cloruro de magnesio 25 mM de 1.0 µl a 1.5 µl, se cortó una banda consistente
con el tamaño esperado de 2,034 pb, se purificó y se clonó en el vector
PCR8®/GW/Topo® de Invitrogen y se digirió con EcoRI obteniéndose tres
fragmentos consistentes con los sitios de corte de EcoRI en la secuencia del gen
AtqGlsA y se mandó secuenciar para asegurarse de que contiene el cDNA
completo.
1kb 1 2 C+ 1kb 4 5
2kb955pb751pb
274pb
Fig.28. AtqGlsA completo. 1 Amplif icación delORF completo, 2 reamplif icación con los mismosoligos, C+ amplif icación con los oligos internos, 4 y5 digestión de DNA plasmídico con EcoRI enPCR8/GW/TOPO.

52
La secuencia completa en el vector PCR8®/GW/Topo® se transfirió al vector
de expresión pB7WG2D mediante recombinación LR, se transformó A.
tumefaciens por electroporación. Como tanto el vector de entrada
PCR8®/GW/TOPO® como el de salida pB7WG2D tienen resistencia a
espectinomicina, se extrajo DNA plasmídico para comparar el tamaño del vector
ya que el pB7WG2D mide 12,542 pb y el PCR8®/GW[/TOPO® mide 2,817 pb,
además se amplificó por PCR el gen de resistencia a fosfinotricina (Bar), el cual
sólo se encuentra en el vector de expresión y se comprobó por PCR y digestión
que el cDNA estuviera completo.
VII.8. Expresión constitutiva de AtqGlsA en A. thaliana.
Con A. tumefaciens transformada con el vector pB7WG2D llevando el cDNA
de interés, se infectaron plantas silvestres de A. thaliana, de la misma manera que
para el gen AtqAS1, sin embargo hasta el momento apenas se cuenta con la
generación T0 de las plantas transformadas y aun no se a podido observar algún
fenotipo sobresaliente distinto de la silvestre, por lo que se continuará examinando
su descendencia en busca de algún fenotipo consistente con la expresión
constitutiva del gen.
VII.9. Análisis de la mutante glsa de A.thaliana.
Se pretendía realizar complementación de la mutante glsa de A. thaliana con el
gen AtqGlsA de A. tequilana, sin embargo, no se observó ningún fenotipo diferente
al de las plantas silvestres, en las mutantes pedidas al stock center, las líneas
pedidas fueron las líneas que tienen una inserción independiente en los dos genes
ortólogos a GlsA en A. thaliana, líneas: CS805009 que contiene una inserción de
T-DNA en el locus AT5G06110, localizada en la región 300-UTR3’, coordenadas
C/1841009-1843000, la línea SALK_070956 con una inserción de T-DNA en el
locus AT3G11450, localizada en un exón, coordenadas, C/3605459-
3606969,3606997-3607477 y la línea SALK_070965 con una inserción en el locus
AT3G11450, localizada en un exón, coordenadas, C/3605459-3606969,3606997-

53
ACT2
KNAT1
AtqAS1
AS1
WT knat1 35SKNAT1 as1 35SAtqAS1 CompAS1
3607477. Como es probable que no veamos ningún fenotipo en las mutantes
debido a que en las mutantes hay un locus del gen funcional y puede ser
suficiente para ocultar el fenotipo mutante, por redundancia de genes, se
realizaron cruzas entre la línea CS805009 y las otras dos líneas, de forma
independiente, de forma que en una cruza la línea CS805009 fuera donadora de
polen y en otra fuera la receptora. En la descendencia de estas cruzas se
observaron algunos fenotipos distintos de la planta silvestre, que pueden deberse
a plantas que tengan en los dos loci la inserción del T-DNA, pero aun faltaría
genotipificarlas para comprobar que realmente se debe a esto. Estas plantas
podrían ser usadas para complementarse con el gen AtqGlsA, sin embargo por
cuestiones de tiempo se dejarán pendientes.
VII.10. Análisis de expresión por RT-PCR de los genes endógenos de Arabidopsis AS1 y KNAT1.
Dado los resultados anteriores se buscó hacer un análisis más exacto de la
forma en que se encuentran interactuando el gen AtqAS1 y los genes KNOX
endógenos de Arabidopsis, por lo que se realizó un análisis de expresión por RT-
PCR en hojas de A.thaliana de las plantas mutantes as1, knat1, expresando los
genes AtqAS1, AtqKNOX1 y de la mutante as1 complementada con el gen AtqAS1
Fig.29.
Fig.29. RT-PCR de la interacción entre genes AS1 y KNOX. WT, planta silvestre. knat1, mutante knat1. 35SKNAT1, sobreexpresante KNAT1 as1, mutante as1. 35SAtqAS1, planta expresando constitutivamente AtqAS1. CompAS1, planta mutante complementada con AtqAS1.
355 pb
293 pb
152 pb
524 pb

54
En la figura 29, el eje vertical representa los genes evaluados y el eje
horizontal las plantas utilizadas para el análisis.
Se puede observar que en el gen AtqAS1 sólo se expresa en las plantas
complementadas y la que lo expresa constitutivamente, como era de esperarse.
En las plantas mutantes knat1 se observa un ligero aumento de la expresión de
AS1 con respecto a la silvestre, sin embargo, en la sobrexpresante KNAT1 se
incrementó su expresión contrariamente a lo que se esperaría y se ve algo de
expresión en la mutante as1, que puede ser debido a que la mutante fue generada
por la eliminación de una base recorriendo el marco de lectura. Con respecto al
KNAT1 se encontró expresión en la sobreexpresante de este gen, por estar bajo el
control del promotor 35S y en la mutante as1 se observa un ligero incremento.
VII.11. Establecimiento de explantes de Agave tequilana in vitro.
El establecimiento de explantes in vitro resultó satisfactorio ya que de cada
planta se logró obtener aproximadamente 5 explantes y cada explante dio varios
brotes en cantidades muy variables, ya que algunos no dieron brotes, otros sólo
unos cuantos y otros explantes dieron aproximadamente 200 brotes, sin embargo
por la poca cantidad de hijuelos de rizoma disponibles se probó estableciendo
explantes a partir de meristemos de bulbilos que tuvieran piñas relativamente
grandes aproximadamente 3 a 4 cm de diámetro. Para establecer explantes a
partir de estas plantas fue necesario omitir el primer lavado con cloro al 40 %
dejándose el resto del procedimiento sin modificar, con este método en
aproximadamente 3 semanas se obtuvieron grandes cantidades de brotes.
VII.12. Infección de los explantes con Agrobacterium tumefaciens.
Para infectar los explantes se utilizaron varias cepas de A. tumefaciens,
probándose las cepas AGL0, GV2260 y LBA4404. La cepa AGL0 parece ser la
más agresiva y en la mayoría de los casos el tejido infectado se necrosó y tuvo

55
que ser descartado, con la cepa LBA4404 ocurrió algo similar, sin embargo en
menor cantidad y con la cepa GV2260 prácticamente el tejido no se dañó, también
se probó infectando una sola vez cuando se realizaron los cortes y cuando
comenzaban a formarse los brotes, infectando 2 y 3 veces a intervalos
aproximados de 15 días y se probó infiltración al vacio, para ayudar a la bacteria a
entrar al tejido. Para verificar la transformación se utilizó el estereomicroscopio de
fluorescencia para detectar expresión de GFP, sin embargo no se logró observar
fluorescencia que indicara claramente la transformación ya que los brotes en que
se observó fluorescencia el tejido estaba dañado y probablemente se debía al
efecto de la lignina. Otro método para verificar la transformación fue la resistencia
a fosfinotricina, se estableció la concentración mínima letal de fosfinotricina para
brotes silvestres en 20mg/l. Los brotes regenerados de los explantes infectados,
se pasaron a medio de multiplicación con esta concentración de fosfinotricina, sin
embargo todos los brotes que se probaron de esta manera murieron en
aproximadamente tres semanas. Hasta ahora no se han obtenido brotes
resistentes.
VIII. DISCUSIÓN.
La mayoría de las plantas se reproducen por medio de la producción de
semillas, evolutivamente esto tiene la ventaja de generar variabilidad genética
para la adaptación de las especies a los cambios que ocurren en el medio en el
que se desarrollan (Hamrick et al., 1992). Sin embargo, existen algunas especies
que se reproducen mayormente en forma vegetativa, este es el caso en Agave
tequilana. La estrategia de reproducción asexual también tiene algunas ventajas,
en Agave las reservas de energía que la planta ha acumulado a través de su largo
ciclo de vida (6 a 8 años en Agave tequilana) son utilizadas para la producción de
una espectacular inflorescencia que producirá miles de semillas. Pero si la
producción de semillas se ve interrumpida, esta especie recurre a un cambio en la
estrategia de reproducción. Cuando ocurre una falla en la polinización o es

56
interrumpida la formación de semillas, se forman los bulbilos de la inflorescencia
como una estrategia de rescate (Szarek et al., 1996). Cuando las señales de
formación de frutos son interrumpidas, hay un reclutamiento de genes y
probablemente un cambio en el balance de hormonas que favorecerán la
formación de meristemos de novo que darán origen a la formación de bulbilos
(Abraham-Juárez et al., 2010).
El estudio de las señales que favorecen la formación de bulbilos está en sus
inicios, ya que en general las Agavaceas han sido muy poco estudiadas a nivel
molecular y el fenómeno de la formación de bulbilos es relativamente poco
frecuente en la naturaleza, por lo que existen muy pocos reportes al respecto.
En el trabajo realizado por Abraham-Juárez et al. (2010), se propuso un
grupo de genes candidatos para estar involucrados en la formación de bulbilos en
Agave tequilana, dentro del cuál se encuentran algunos factores de transcripción y
genes relacionados con el desarrollo de meristemos; se evaluó la expresión de
dos genes de la familia KNOXI y dos genes que parecen ser marcadores de la
actividad meristemática que no han sido caracterizados en otras plantas,
situándolos en una red de interacción de señales que involucra desdiferenciación
celular para la formación de estructuras vegetativas que formarán las nuevas
plántulas. Dentro del grupo de los genes que se propusieron como candidatos, se
encuentran dos cDNA con identidad significativa a los genes AS1 de Arabidopsis
thaliana y a GlsA de Lilium longiflorum. AS1 es un factor de transcripción con un
dominio MYB que en Arabidopsis tiene la función de formar primordios de hojas,
reprimiendo a los genes KNOXI que se expresan en el meristemo apical (Hay et
al., 2006). GlsA en Lilium longiflorum se identificó durante la formación del polen
durante las divisiones celulares asimétricas y se propuso que tiene la función de
llevar a cabo diferenciación celular en plantas (Mori et al., 2003).
Con estos antecedentes se eligieron los genes AtqAS1 y AtqGlsA para
llevar a cabo su caracterización durante la formación de bulbilos y tratar de

57
situarlos en el modelo de interacción de los factores que están involucrados en
este proceso.
Los genes AtqAS1 y AtqGlsA presentan patrones de expresión que los relacionan con la formación de bulbilos.
De acuerdo a los resultados de los análisis de expresión por RT-PCR, se
determinó que AtqAS1 tiene un aumento de expresión en la etapa T3 de la
formación de bulbilos y aún más en T4 y T5. Estas etapas corresponden a estados
de desarrollo avanzados, es decir cuando ya están formados los domos
meristemáticos y ya existen primordios de hojas. Con respecto a los otros órganos
de la planta, no se detectó expresión en raíz, esto sugiere que AtqAS1 solamente
se expresa en la formación de órganos laterales aéreos y no en raíces laterales,
contrario a lo que ocurre en Arabidopsis, donde sí existe expresión en raíz (Sun et
al., 2002). La más alta expresión se detectó en anteras, meristemo de umbelas y
meristemo apical reproductivo, sugiriendo un papel en la diferenciación celular,
como fue propuesto en Arabidopsis, por Byrne et al., 2002. Para hacer una
comparación del patrón de expresión de AtqAS1 con los genes KNOXI, se realizó
el RT-PCR de AtqKNOXI; se observó que AtqKNOXI también presenta expresión
elevada en las etapas T3, T4 y T5. Estos resultados no permiten identificar el
dominio de expresión específico, ya que la muestra que se está evaluando
corresponde a todo el pedicelo floral con la región en la que inicia la formación de
bulbilos. Por lo que fue necesario llevar a cabo un ensayo de Hibridación in situ,
esta técnica permite identificar el grupo de células donde se está expresando el
gen de interés y aunque se propuso en los objetivos llevar a cabo la Hibridación in
situ a través de todas la etapas de formación de bulbilos de los dos genes AtqAS1
y AtqGlsA, el tiempo sólo permitió realizar un primer ensayo con AtqAS1 en tejido
de meristemos de umbelas, meristemo apical reproductivo y la ultima etapa de
formación de bulbilos T5. Sin embargo este ensayo proporcionó información sobre
el lugar de expresión en los meristemos evaluados y se pudo determinar que el
mRNA de AtqAS1 se localiza en los primordios de órganos laterales como hojas y
umbelas. En los bulbilos en T5 se observa expresión en las hojas, lo cuál no

58
ocurre con AtqKNOX1, este último sólo se expresa en el domo del meristemo
apical, lo cual sugiere que AtqKNOX1 está involucrado en el mantenimiento del
meristemo y AtqAS1 en la formación de las hojas. Aunque es necesario realizar la
Hibridación in situ en todas las etapas de la formación de bulbilos para determinar
el lugar de la expresión desde el inicio de su formación.
El análisis de las bibliotecas de cDNA junto con el RACE PCR proporcionó herramientas adecuadas para la identificación de cDNAs completos.
Para la obtención de la secuencia completa del cDNA de AtqAS1, fue
suficiente con el análisis del tamaño de los insertos de las clonas correspondientes
y la resecuenciación por los dos extremos de los cDNAs. En trabajos anteriores
del análisis de las bibliotecas de cDNA de Agave tequilana se ha observado que
secuencias de 1,200 pb (tamaño promedio de insertos de las bibliotecas) o menos
tienen una alta probabilidad de estar completas, en este caso la secuencia de
AtqAS1 que se buscaba mide 1,127 pb por lo que está dentro del rango del
tamaño promedio de los insertos.
No ocurrió así en la búsqueda del cDNA completo de AtqGlsA, el cual ahora
sabemos que mide 1,980 pb. El fragmento de la secuencia que se tenía era de
1,450 pb, es decir, faltaban 530 pb hacia el extremo 5´ para completarlo. La
técnica de RACE PCR 5´ resultó adecuada para la obtención de la secuencia
completa, aunque durante la metodología se llevaron a cabo varias modificaciones
para la amplificación específica de la banda de 1,100 pb obtenida por el PCR
RACE con el oligonucleótido Gene Racer 5´ y el oligonucleótido específico para la
secuencia conocida de AtqGlsA, hasta lograr amplificar una banda única de este
tamaño y la clonación para su secuenciación. Al final, cuando se amplificó la
secuencia completa del cDNA que medía 2,034 pb y se clonó en el vector
Gateway® de entrada TOPO® PCR8®, se tuvo que analizar una gran cantidad de
clonas para verificar la presencia del cDNA completo, pues en la mayoría se
habían clonado solamente fragmentos mas pequeños, pero se lograron identificar
5 clonas con el inserto de 2,034 pb, las cuales se mandaron a secuenciar para

59
verificar su identidad y poder utilizarlas para la expresión heteróloga en
Arabidopsis thaliana.
AtqAS1 y AtqGlsA tienen identidad significativa con sus ortólogos de A. thaliana y de L. longiflorum. Los cDNAs completos fueron traducidos a aminoácidos usando el programa
DNA star Edit seq y después fueron comparados por alineamiento con sus
ortólogos más cercanos. AtqAS1 tiene un porcentaje de identidad de 61 % con
AS1 de Arabidopsis y de 56 % con RS2 de maíz, estos niveles de similitud
permiten suponer que son ortólogos. La secuencia de Agave tequilana es más
similar a la de Arabidopsis que a la de maíz, al contrario de lo que se esperaría,
por la relación más cercana que existe taxonómicamente con maíz,
probablemente debido a la alta conservación que existe en este tipo de genes de
desarrollo en las plantas en general, pues el dominio MYB es casi idéntico para las
tres secuencias. Existen reportes de que el dominio MYB es esencial para la
función de estos genes, pues tanto AS1 como RS2 complementan totalmente la
mutante as1 cuando se usa la secuencia codificante completa, pero cuando se
usa la secuencia en la que el dominio MYB ha sido interrumpido ya no se logra la
complementación con ninguno de los dos genes (Theodoris et al., 2003).
El alineamiento de la secuencia codificante traducida a aminoácidos de
AtqGlsA con las de L. longiflorum, A. thaliana y V. carteri. Muestra que tiene mayor
similitud con la de Lilium, con un porcentaje de identidad de 71 % en aminoácidos,
por lo que se puede considerar que son ortólogos; el alineamiento también
muestra que todas las secuencias comparadas tienen los cuatro dominios
conservados (J, M, WR1 y WR2), de estos dominios el J es el más importante
para la función de la proteína, pues se ha demostrado que es indispensable para
las divisiones asimétricas y diferenciación celular (Miller y Kirk, 1999).

60
La expresión heteróloga en A. thaliana demostró que AtqAS1 tiene la misma función que AS1.
La expresión de AtqAS1 bajo el control del promotor constitutivo 35S en
plantas silvestres de Arabidopsis produjo un fenotipo similar al que se ha
reportado en plantas sobreexpresantes de AS1 (Chua et al., 2005). El cual
consiste en hojas de la roseta y peciolos alargados y floración tardía, con respecto
a plantas silvestres. Este fenotipo se observó desde la generación T0 en 8 de las
40 líneas transformadas, pero en la generación T1 todas las plantas, heterocigotas
y homocigotas presentaron el fenotipo, aunque en un gradiente de severidad,
siendo las homocigotas las que lo presentaron en mayor grado. Este resultado
sugirió que AtqAS1 podía tener la misma función que AS1.
La suposición de la función de AtqAS1 se confirmó con su expresión en la
mutante as1. De las 40 líneas transformadas obtenidas en la generación T0, 18
mostraron complementación total de la mutante as1, con respecto a la morfología
de las hojas y el tiempo de floración. as1 presenta hojas de la roseta relativamente
pequeñas y en forma de corazón, floración tardía, superficie rugosa en las silicuas
y tamaño pequeño de la planta en general (Ori et al., 2000). Las líneas
complementadas recuperaron el tamaño de la planta como las silvestres, la forma
de las hojas y en algunos casos con forma alargada como las sobreexpresantes
de AS1, así como el tamaño y forma de las silicuas. Estos resultados demostraron
que AtqAS1 está tomando la función de AS1 para restituir el fenotipo.
Los RT-PCR que se realizaron con tejido de hoja de plantas WT, knat1,
35S::KNAT1, 35S::AtqAS1, as1 y as1cAtqAS1 confirmaron que AS1 y KNAT1 se
reprimen mutuamente y que AtqAS1 tiene el mismo efecto sobre KNAT1 que AS1.

61
Los análisis de expresión y la expresión heteróloga sugieren que AtqAS1 está situado en la red de interacción de genes de desarrollo para la formación de bulbilos, después que los genes KNOXI.
Los resultados de los RT-PCR y el ensayo de la Hibridación in situ
mostraron que AtqAS1 aumenta su nivel de expresión el las etapas de formación
de bulbilos en las que se están formando los primordios de hojas y en los
meristemos en los que se están formando órganos laterales. Pero su expresión se
eleva después que la de los genes AtqKNOX1 y AtqKNOX2 y se observa en
primordios de hojas, a diferencia de estos genes. Además la expresión constitutiva
en Arabidopsis y la complementación total de la mutante as1 muestran que
AtqAS1 tiene la función de AS1 en A. thaliana. Por lo que se puede suponer que
AtqAS1 interviene en la formación de bulbilos en las etapas tardías, es decir,
cuando se forman las hojas que formarán la roseta de la nueva planta de Agave.
Se requiere profundizar en el análisis de las mutantes glsA de A. thaliana para establecer su función en esta planta y poder realizar la complementación con AtqGlsA.
Para poder utilizar las mutantes insercionales glsA de A. thaliana como
herramienta para determinar la función de AtqGlsA, es necesario caracterizar las
mutantes por genotipificación y establecer cual es la función de este gen en
Arabidopsis, por análisis del fenotipo de las cruzas que se realizaron. Por su
función en V. carteri y L. longiflorum (Miller y Kirk, 1999), (Mori et. al., 2003), así
como el patrón de expresión que presenta en Agave tequilana, parece un gen
muy interesante para determinar su función durante la formación de bulbilos. El
avance que se logró en este trabajo es significativo, pues se logró identificar la
secuencia completa del cDNA, lo cual permitió definir a AtqGlsA como un ortólogo
más de GlsA con el dominio funcional J y se puede considerar que también está
involucrado en la formación de bulbilos por su patrón de expresión.

62
Es necesario realizar más pruebas para la obtención de plantas de Agave
tequilana transformadas con Agrobacterium.
Aunque es posible caracterizar funcionalmente los genes de Agave en el
sistema heterólogo de A. thaliana, será muy útil tener la opción de analizar en
detalle la expresión de los genes de Agave en la misma especie. Por esta razón
iniciamos el desarrollo de un protocolo de regeneración y transformación. Los
intentos de transformación que se llevaron a cabo no resultaron en un método
eficiente para la obtención de transformantes a partir de explantes de meristemos.
Sin embargo proporcionaron alternativas a seguir, por ejemplo el uso de
meristemos de bulbilos como explantes para la regeneración de brotes in vitro.
Esta fuente de explantes es adecuada para el propósito de la transformación por
ser fácil de obtener y ser una fuente abundante por la cantidad de bulbilos que se
obtienen de una sola planta. También el uso de las diferentes cepas de
Agrobacterium tumefaciens proporcionó información sobre el grado de agresividad
en los explantes que se usaron, probablemente la siguiente modificación a seguir
sea el uso de diferentes vectores de expresión.
Posible función de los genes AtqGlsA y AtqAS1 en la formación de bulbilos en Agave tequilana.
Por su patrón de expresión es probable que AtqGlsA tenga actividad en
etapas tempranas de la formación de bulbilos, en base a la función de su ortólogo
de Lilium longiflorum (Mori et. al., 2003), probablemente esté llevando a cabo
divisiones celulares asimétricas esenciales para el inicio de los meristemos que
darán origen a los bulbilos. A nivel molecular en base a la presencia del dominio J
puede ser que esta proteína esté actuando en la activación de otras proteínas
involucradas en este proceso, a través de procesamiento postraduccional. Dado
que AtqAS1 completó la mutante as1 y su expresión constitutiva en Arabidopsis
thaliana provoca un fenotipo semejante a la sobreexpresión del gen AS1 (Chua et
al., 2005), se puede concluir que AtqAS1 es un ortólogo de AS1, su mayor
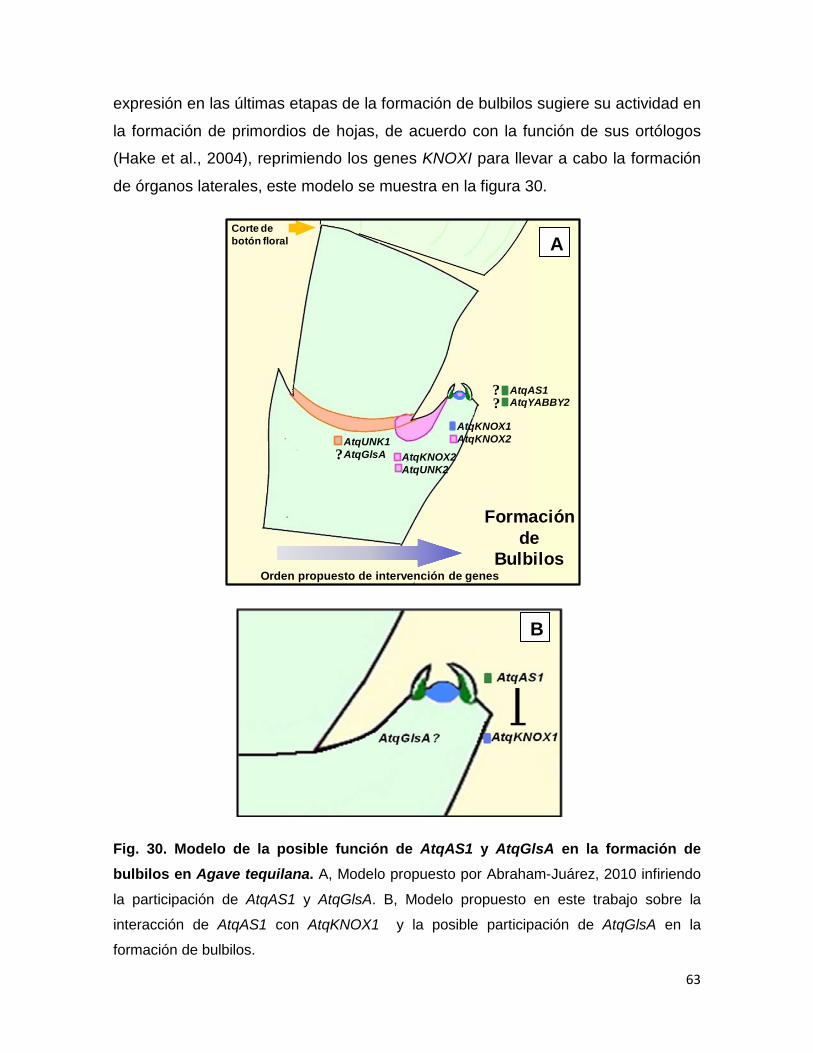
63
expresión en las últimas etapas de la formación de bulbilos sugiere su actividad en
la formación de primordios de hojas, de acuerdo con la función de sus ortólogos
(Hake et al., 2004), reprimiendo los genes KNOXI para llevar a cabo la formación
de órganos laterales, este modelo se muestra en la figura 30.
Orden propuesto de intervención de genes
AtqUNK1AtqGlsA AtqKNOX2
AtqUNK2
AtqKNOX1AtqKNOX2
?
? ?
Corte de botón floral
AtqAS1AtqYABBY2
Formación de
Bulbilos
Fig. 30. Modelo de la posible función de AtqAS1 y AtqGlsA en la formación de bulbilos en Agave tequilana. A, Modelo propuesto por Abraham-Juárez, 2010 infiriendo
la participación de AtqAS1 y AtqGlsA. B, Modelo propuesto en este trabajo sobre la
interacción de AtqAS1 con AtqKNOX1 y la posible participación de AtqGlsA en la
formación de bulbilos.
A
B

64
IX. CONCLUSIONES.
Se determinó el patrón de expresión por RT-PCR de AtqAS1.
Se identificó la secuencia completa del cDNA de AtqAS1 por secuenciación
de la clona por los dos extremos.
Se completó la secuencia del cDNA de AtqGlsA por RACE 5´.
Se llevó a cabo la expresión constitutiva de AtqAS1 en plantas silvestres de
Arabidopsis thailana, obteniendo un fenotipo similar al reportado para
plantas sobreexpresantes de AS1.
Se logró la complementación total de la mutante as1 con el cDNA de
AtqAS1, confirmando que el gen de Agave tiene la misma función que AS1.
Se comprobó la interacción negativa entre KNAT1 y AtqAS1 por RT-PCR en
Arabidopsis thaliana.
Se realizó la expresión constitutiva de AtqGlsA en plantas silvestres de
Arabiopsis thaliana.
Se probaron varias condiciones para la infección de explantes de Agave
tequilana con Agrobacterium tumefaciens con el propósito de sobreexpresar
AtqAS1, pero hasta ahora no se han obtenido transformantes.

65
X. PERSPECTIVAS.
• Completar el análisis de Hibridación in situ con AtqAS1 y llevar a cabo el
análisis con AtqGlsA.
• Realizar la genotipificación de las dos mutantes insersionales glsA de
Arabidopsis thaliana y caracterizar el fenotipo de las cruzas entre ellas.
• Caracterizar el fenotipo de las plantas de Arabidopsis expresando
constitutivamente AtqGlsA.
• Completar los análisis de expresión de los genes KNOXI en las plantas as1,
as1 complementada con AtqAS1 y 35S::AS1.
• Llevar a cabo análisis de interacción de proteínas correspondientes a los
genes identificados como participantes en la formación de bulbilos en
Agave tequilana, por ensayos de dos híbridos en levadura.
• Realizar más pruebas para la transformación de Agave tequilana por
infección con Agrobacterium tumefaciens, utilizando otros vectores de
expresión.

66
XI. REFERENCIA BIBLIOGRAFICA.
• Abraham-Juárez M J. 2010. Análisis de la formación de bulbilos en Agave
tequilana a nivel histológico y molecular. Tesis de Doctorado. Centro de
Investigación y de Estudios Avanzados del Instituto Politécnico Nacional.
Irapuato, México. 118p.
• Abraham-Juárez M J, Martínez-Hernández A, Leyva-González M A, Luis Herrera-Estrella L, Simpson J. 2010. Class I KNOX genes are associated
with organogenesis during bulbil formation in Agave tequilana. Journal of
Experimental Botany, doi:10.1093/jxb/erq215.
• Alonso-Cantabrana H, Ripoll J J, Ochando I, Vera A, Ferrándiz C, Martínez-Laborda A. 2007. Common regulatory networks in leaf and fruit
patterning revealed by mutations in the Arabidopsis ASYMMETRIC
LEAVES1 gene. Development, 134, 2663-2671.
• Arizaga S, Ezcurra E. 1995. Insurance against reproductive failure in a
semelparous plant: bulbil formation in Agave macroacantha flowering stalks.
Oecologia, 101, 329 – 334.
• Arizaga S, Ezcurra E. 2002. Propagation mechanisms in Agave
macroacantha (Agavaceae), a tropical arid-land succulent rosette. American
Journal of Botany, 89 (4), 632– 641.
• Byrne M E, Barley R, Curtis M, Arroyo J M, Dunham M, Hudson A, Martienssen R A. 2000. Asymmetric leaves1 mediates leaf patterning and
stem cell function in Arabidopsis. Nature, 408, 967-971.
• Byrne M E, Simorowski J, Martienssen R A. 2002. ASYMMETRIC
LEAVES1 reveals knox gene redundancy in Arabidopsis. Development,
129, 1957-1965.
• Colunga-García P, Larqué-Saavedra A, Eguiarte E, Zizumbo-Villarreal D (eds). 2007. En lo ancestral hay futuro: del tequila, los mezcales y otros
agaves. CICY, CONACYT, CONABIO, SEMARNAT, INE. México. pp. 3-18.
• Chua Y L, Channelière S, Mott E, Gray J C. 2005. The bromodomain
protein GTE6 controls leaf development in Arabidopsis by histone

67
acetylation at ASYMMETRIC LEAVES1. GENES & DEVELOPMENT, 19,
2245–2254.
• Chuck G, Lincoln C, Hake S. 1996. Knat1 induces lobed leaves with
ectopic meristems when overexpressed in Arabidopsis. Plant Cell, 8, 1277-
1289.
• Eguiarte L E, Souza V, Silva-Montellano A. 2000. Evolución de la familia
Agavaceae: Filogenia, biología reproductiva y genética de poblaciones.
Boletín de la Sociedad Botánica de México, 66, 131-150.
• Eriksson T. 1965. Studies on the growth requirements and growth
measurements of cell cultures of Happlopappus gracilis. Physiol Plant, 18,
976-993.
• Escobar-Guzmán RE, Zamudio-Herández F, Gil-Vega K, Simpson J. 2008. Seed production and gametophyte formation in Agave tequilana and
Agave americana. Botany. 86, 1343-1353.
• García-Mendoza A. 2004. Agaváceas. En: A. J. García –Mendoza, M. J.
Ordóñez y M. Briones (eds). Biodiversidad de Oaxaca. Universidad
Autónoma de Oaxaca. Instituto de Biología. UNAM-Fondo Oaxaqueño para
la Conservación de la Naturaleza-World Wildlife Fund. México. pp 159-169.
• García-Mendoza A. 2007. Los agaves de México. CIENCIAS, 87, 14-23.
• Gentry H S. 1982. Agaves of Continental North America, The University of
Arizona Press, Tucson. pp. 668.
• Granados S D. 1993. Los Agaves en México, Universidad Autónoma
Chapingo, México. pp. 31-35.
• Guo M, Thomas J, Collins G, Timmermansa M C P. 2008. Direct
Repression of KNOX Loci by the ASYMMETRIC LEAVES1 Complex of
Arabidopsis. The Plant Cell, 20, 48-58.
• Hake S, Smith H, Holtan H, Magnani E, Mele G, Ramirez J. 2004. The
role of KNOX genes in planta development. Annu. Rev. Cell Dev. Biol. 20,
125–51
• Hamrick J L, Godt M J, Sherman-Broyles S L. 1992. Factors influencing
levels of genetic diversity in woody plant species. New Forests, 6, 95-124.

68
• Hay A, Barkoulas M, Tsiantis M. 2006. ASYMMETRIC LEAVES1 and
auxin activities converge to repress BREVIPEDICELLUS expression and
promote leaf development in Arabidopsis. Development, 133, 3955-3961.
• Jackson D, Veit B, Hake S. 1994. Expression of maize KNOTTED1 related
homeobox genes in the shoot apical meristem predicts patterns of
morphogenesis in the vegetative shoot. Development, 120, 405-413.
• Karimi M, Inze D, Depicker A. 2002. Gateway® vectors for Agrobacterium-
mediated plant transformation. Trends in Plant Science 7, 193-195.
• Larkin MA, Blackshields G, Brown NP, et al. 2007. Clustal W and Clustal
X version 2. Bioinformatics 23, 2947–2948.
• Linsmaier E M, Skoog F. 1965. Organic Growth Factor Requirements of
Tobacco Tissue Cultures. Physiologia Plantarum, 18 (1), 100–127.
• Martínez-Trujillo M, Limones-Briones V, Cabrera-Ponce JL and Herrera-Estrella L. 2004. Improving Transformation Efficiency of Arabidopsis
thaliana by Modifying the Floral Dip Method. Plant Molecular Biology
Reporter 22, 63–70.
• Miller S M, Kirk D L. 1999. glsA, a Volvox gene required for asymmetric
division and germ cell specification, encodes a chaperone-like protein.
Development, 126, 649-658.
• Mori T, Kuroiwa H, Higashiyama T and Kuroiwa T. 2003. Identification of
higher plant GlsA, a putative morphogenesis factor of gametic cells.
Biochemical and Biophysical Research Communications 306, 564–569.
• Nava A. 1988. Agave tequilana Weber “Azul” in vitro: un modelo para
estudios en morfogénesis. Tesis de Maestría. Instituto de Enseñanza e
Investigación en Ciencias Agrícolas. Centro de Genética. Colegio de
Postgraduados. Montecillo, México. 233pp.
• Ori N, Eshed Y, Chuck G, Bowman J L, Hake S. 2000. Mechanisms that
control KNOX gene expression in the Arabidopsis shoot. Development, 127,
5523-5532.
• Rocha M, Good-Ávila S V, Molina-Freaner F, Arita H T, Castillo A, García-Mendoza A, Silva-Montellano A, Gaut B S, Souza V, Eguiarte l E.

69
2006. Pollination biology and adaptive radiation of Agavaceae, with special
emphasis on the genus Agave. Aliso, 22, 329-344.
• Ruiz-Medrano R, Xoconostle-Cazarez B and Lucas WJ. 1999. Phloem
long-distance transport of CmNACP mRNA: implications for supracelular
regulation in plants. Development, 126, 4405-4419.
• Sun Y, Zhou Q, Zhang W, Fu Y, Huang H. 2002. ASYMMETRIC
LEAVES1, an Arabidopsis gene that is involved in the control of cell
differentiation in leaves. Planta, 214, 694–702.
• Szarek S R, Driscoll B, Shohet C, Priebe S. 1996. Bulbil production in
Agave (Agavaceeae) and related genera. The Southwestern Naturalist, 41
(4), 465-469.
• Theodoris G, Inada N y Freeling M. 2003. Conservation and molecular
dissection of ROUGH SHEATH2 and ASYMMETRIC LEAVES1 function in
leaf development. PNAS, 100(11), 6837-6842.
• Tsiantis M, Schneeberger R, Golz J F, Freeling M, Langdale J A. 1999. The Maize rough sheath2 Gene and Leaf Development Programs in
Monocot and Dicot Plants. Science, 284 (5411), 154 – 156.
• Valenzuela Z A. 1997. El agave tequilero su cultivo e industria, Litteris
editores, México. pp. 31-41, 49-51, 123-134.
• Valenzuela Z A, Gerritsen P R W. 2007. Memorias del 1er Seminario
Internacional de Tequila: Ambiente, Cultura y Sociedad. Autlán: Universidad
de Guadalajara, Centro Universitario de la Costa Sur, Instituto Manatlán de
Ecología y Conservación de la Biodiversidad/Signo Tequila, A.C.
• Wang C N, Cronk Q C. 2003. Meristem fate and bulbil formation in
Titanotrichum (Gesneriaceae). American Journal of Botany, 90 (12), 1696-
1707.
• Wang C N, Moller M, Cronk Q C. 2004. Altered expression of GFLO, the
Gesneriaceae homologue of FLORICAULA/LEAFY, is associated with the
transition to bulbil formation in Titanotrichum oldhamii. Dev. Genes Evol
214, 122-127.

70
• Zhang Y, Frohman M A. 1997. Using rapid amplification of cDNA ends
(RACE) to obtain full-length cDNAs. Methods in Molecular Biology, 69, 61-
87.


















