CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS … · por esos momentos de alegría que pasamos. A todos...
149
CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS DEL INSTITUTO POLITÉCNICO NACIONAL UNIDAD DE BIOTECNOLOGÍA E INGENIERÍA GENÉTICA DE PLANTAS DEPARTAMENTO DE BIOTECNOLOGÍA Y BIOQUÍMICA Identificación y cuantificación de metabolitos secundarios en plantas de tabaco (Nicotiana tabacum) transformadas con sistemina y prosistemina de jitomate en respuesta a daño mecánico, herbivoría con Manduca sexta e infestación con mosquita blanca (Bemisia tabaci). T E S I S QUE PARA OBTENER EL GRADO DE: MAESTRÍA EN CIENCIAS CON ESPECIALIDAD EN: BIOTECNOLOGÍA DE PLANTAS PRESENTA: I.B. Flor Citlally Alcántar Aguirre DIRECTOR DE TESIS: DR. JOHN PAUL DÉLANO FRIER IRAPUATO, GUANAJUATO DICIEMBRE DE 2005
Transcript of CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS … · por esos momentos de alegría que pasamos. A todos...

CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS
DEL INSTITUTO POLITÉCNICO NACIONAL
UNIDAD DE BIOTECNOLOGÍA E INGENIERÍA GENÉTICA DE PL ANTAS
DEPARTAMENTO DE BIOTECNOLOGÍA Y BIOQUÍMICA
Identificación y cuantificación de metabolitos secundarios
en plantas de tabaco (Nicotiana tabacum) transformadas con sistemina
y prosistemina de jitomate en respuesta a daño mecánico, herbivoría con
Manduca sexta e infestación con mosquita blanca (Bemisia tabaci).
T E S I S QUE PARA OBTENER EL GRADO DE:
MAESTRÍA EN CIENCIAS
CON ESPECIALIDAD EN:
BIOTECNOLOGÍA DE PLANTAS
PRESENTA:
I.B. Flor Citlally Alcántar Aguirre
DIRECTOR DE TESIS:
DR. JOHN PAUL DÉLANO FRIER
IRAPUATO, GUANAJUATO DICIEMBRE DE 2005

DEDICATORIAS
A mi nana Lolita: por recordarme su gran fortaleza de lucha y amor a la vida, por tus
cuidados, atención, amor tierno y sonrisa que me regalabas en cada regreso.
A ti madre: por darme lo mejor de ti, por otorgarme tu apoyo y confianza, por estar
siempre conmigo.
A mis hermanos, David, América Anahí y Daryl Ivan: por sus grandes cualidades y
enseñanzas.
A mis pequeños sobrinos, Sebastián y Carlos: por mantener con gran alegría la
casa de la abuela.
A Humberto Valenzuela Soto, por su apoyo incondicional en todo momento.
A mis queridas tías y primas, que me han alentado en este camino.
A mis amigas de la infancia: por encontrarnos siempre con gusto y cariño a pesar
de la distancia.

AGRADECIMIENTOS
A Dios: por permitirme alcanzar este anhelo, por ser el único y verdadero camino de
vida.
A mi asesor, Dr. John Paúl Délano Frier: por su aceptación en su laboratorio,
paciencia, constante dedicación y consejos otorgados durante mi estancia; gracias.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT): por el apoyo económico
otorgado a través de la beca con número de registro 174677.
A los doctores Jorge Molina Torres y Neftalí Ochoa Alejo: por formar parte del
comité evaluador, como por su crítica constructiva y sugerencias que aportaron al
presente trabajo de tesis.
A la Q.F.B. Norma Martínez Gallardo: por su experiencia e invaluable apoyo para
concluir este trabajo; mil gracias.
Al Q.F.B. Enrique Ramírez Chávez: por sus consejos y aporte metodológico en la
identificación de metabolitos secundarios.
A la Dra. Ma. Del Carmen Rocha Granados: por su buena disponibilidad en el
trayecto de este trabajo.
A la Q. Yolanda Rodríguez Aza y María Isabel Cristina Elizarraraz Anaya: por su
atención y tiempo recibido.
Al I.A. Javier Luevano Borroel: por su buena disponibilidad y consejos en el manejo
de M. sexta.
A mis amigos y compañeros de laboratorio: Carla, Humberto, Gloria, Hamlet y
Miriam, por su gran solidaridad, apoyo y palabras alentadoras en tiempos difíciles;
por esos momentos de alegría que pasamos.
A todos mis compañeros de generación: Victor, Pedro, Frank, Marco, Alfredo,
Javier, Gloria, Amparo y Cecilia.
A Nancy Barojas Pérez: por la amistad que se formó entre nosotras, por
escucharme y aconsejarme después de un mal momento.
A mi amigo Carlos Montoya: por compartirme su experiencia y apoyo en momentos
difíciles.

El presente trabajo de tesis fue realizado en el
Laboratorio de Fisiología de la Defensa, del
Departamento de Biotecnología y Bioquímica
de Plantas del CINVESTAV- IPN, Unidad de
Biotecnología e Ingeniería Genética de Plantas,
bajo la asesoría del Doctor John Paúl Délano Frier.

ÍNDICE GENERAL
Pág.
ABREVIATURAS i
ÍNDICE DE FIGURAS iii
RESUMEN X
1. INTRODUCCIÓN 1
2. ANTECEDENTES 4
2.1. Mecanismos de defensa 4
2.1.1. Tipos de respuestas de defensa en planta 5
2.1.1.1. Respuesta innata y respuesta específica, gen por gen. 6
2.1.1.2. Respuesta local 8
2.1.1.3. Respuesta sistémica 9
2.2. Mecanismos de defensa en tabaco 10
2.3. Respuestas de resistencia en plantas contra el ataque de plagas 13
2.3.1. Inhibidores de proteasas 13
2.3.2. Polifenol oxidasas 14
2.3.3. Oligosacáridos 15
2.4. Sistemina 16
2.4.1. Actividad y translocación de sistemina 17
2.4.2. Modelo de la ruta de señalización activada por sistemina 20
2.4.3.Glicopéptidos ricos en hidroxiprolina (“sisteminas”) de tabaco
(TobHypSys)
22
2.5. Prosistemina 25
2.5.1. Expresión y supresión de la actividad de la prosistemina 26
2.5.2. Homología de la prosistemina de jitomate en plantas solanáceas 28
2.6. Metabolitos secundarios en tabaco 30
2.6.1. Alcaloides (nicotina, nornicotina) 31
2.6.2. Compuestos fenólicos 33
2.6.3. Terpenos 34

2.7. Modelo de estudio 37
2.7.1. Plantas de tabaco transformadas con prosistemina y sistemina
de jitomate Nt -LePS 08, Nt -LeSYS 03 y la cruza WT X Nt -LeSYS 03
37
2.7.2. Alteraciones morfológicas y fisiológicas en la línea Nt -LeSYS
03
39
2.7.3. Resistencia a herbivoría por larvas de Manduca sexta en plantas
Nt-LePS y Nt -LeSYS
43
3. HIPÓTESIS 46
4. OBJETIVOS 47
4.1. Objetivo general 47
4.2. Objetivos específicos 47
5. MATERIALES Y MÉTODOS 48
5.1. Cultivo de plantas 48
5.2. Cultivo in vitro de la línea Nt -LeSYS 03 48
5.3. Preparación de X-Gluc 49
5.4. Ensayos de daño mecánico 50
5.5. Bioensayos 51
5.5.1. Condiciones del experimento de herbivoría con Manduca sexta 51
5.5.2. Infestación con mosquita blanca (Bemisia tabaci) 52
5.6. Preparación de extractos 52
5.7. Análisis de fenoles totales 53
5.8. Condiciones para cuantificar e identificar metabolitos secundarios por
GC/MS
53
5.8.1. Derivatización 53
5.8.2 Cromatografía de gases acoplada a espectrometría de masas
(GC/MS)
54
5.9. Curvas de calibración 54

5.10. Análisis estadístico 55
6. RESULTADOS 56
6.1. Identificación de metabolitos secundarios de interés por GC/MS, en
plantas jóvenes y adultas.
56
6.2. Cuantificación de nicotina en plantas jóvenes y/o adultas 67
6.2.1. Tratamiento: daño mecánico 67
6.2.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas) 68
6.2.3. Tratamiento: infestación por mosquita blanca (plantas adultas) 70
6.3. Cuantificación de ácido clorogénico en plantas jóvenes y/o adultas 72
6.3.1. Tratamiento: daño mecánico 72
6.3.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas) 78
6.3.3. Tratamiento: infestación por mosquita blanca (plantas adultas) 78
6.4. Cuantificación de dihidrocapsenona en plantas jóvenes y/o adultas 80
6.4.1. Tratamiento: daño mecánico 80
6.4.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas) 83
6.4.3. Tratamiento: infestación por mosquita blanca (plantas adultas) 87
6.5. Cuantificación de ácido nornicotina en plantas jóvenes y/o adultas 88
6.5.1. Tratamiento: daño mecánico 88
6.6. Cuantificación de fenoles totales en plantas jóvenes y/o adultas 92
6.6.1. Tratamiento: daño mecánico 92
6.6.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas) 93
6.6.3. Tratamiento: infestación por mosquita blanca (plantas adultas) 96
7. DISCUSIÓN DE RESULTADOS 98
8. CONCLUSIONES 113
9. PERSPECTIVAS 114
10. BIBLIOGRAFÍA 116
11. APÉNDICE 132

ABREVIATURAS
aa Aminoácidos
ABA Ácido abscísico
ACG Ácido clorogénico
AJ Ácido jasmónico
AMPC Adenosina Monofosfato Cíclica
AOC Ciclasa de óxido de aleno
AS Ácido salicílico
Avr Gen de Avirulencia
C-F Consumidor - Fuente
cm Centímetro
FAL Fenilalanina amonio liasa
EAS 5-epi-aristoloqueno sintasa
Et Etileno
g Gramo
GC/MS Cromatografía de gases acoplada a espectrometría de
masas
GRHP Glicoproteínas ricas en hidroxiprolina
GTP Guanidina trifosfato
FPP Farnesil bifosfato
HDJ Hoja distal joven (apical)
HDV Hoja distal vieja (basal)
HL Hoja local (dañada)
h Hora
HS Hoja sistémica
IP Inhibidor de proteasa
JA Jasmonato
L Litro
LOX Lipooxigenasa i

MB Mosquita blanca
MeJA Éster metílico del ácido jasmónico
mg Miligramo
ml Mililitro
ng Nanogramo
NN Nornicotina
OGA Oligosacáridos
PF Peso fresco
PFO Polifenol oxidasa
PG Poligalacturonasa
Pin II Inhibidor de proteasa II
R Gen de Resistencia
RH Respuesta hipersensible
RP Relacionadas a patogénesis
rpm Revoluciones por minuto
RSA Resistencia sistémica adquirida
RSI Resistencia sistémica inducida
µg Microgramo
µl Microlitro
ii

LISTA DE FIGURAS
Pág. Figura 1. Reconocimiento del ataque de M. sexta por N. attenuata;
incremento de los niveles de AJ y etileno (Et), e inducción de defensas
directas e indirectas (modificado de Baldwin et al., 2003).
12
Figura 2. Estructura primaria de la sistemina de jitomate.
17
Figura 3. Efecto de la sustitución de cada uno de los 18 aminoácidos de
la sistemina de jitomate, por alanina, sobre la inducción de la síntesis de
inhibidores de proteasa tipo I en hojas de jitomate.
18
Figura 4. Modelo hipotético de la ruta de transducción de señales en
células de jitomate activada por sistemina.
21
Figura 5 . Estructura primaria de la sistemina de jitomate y de los
glicopéptidos ricos en hidroxiprolina presentes en jitomate (TomSys y
TomHypSys I, II y III) y tabaco (TobHypSys I y II), respectivamente.
23
Figura 6 . Actividad de TobHypSys I y TobHypSys II sintéticas, carentes
de modificaciones con cadenas de oligosacáridos y con sustituciones de
alanina en cada uno de sus aminoácidos.
25
Figura 7. Niveles de inhibidores de proteasas I y II en plantas de jitomate
sin transformar (1) y transformadas (2) con el transgen de la prosistemina
en orientación “sentido”.
27
Figura 8. A) Acumulación de inhibidores de proteasas en hojas de
jitomate tratadas con distintas concentraciones de prosisteminas
recombinantes. B) supresión de inhibidores de proteasas en hojas de
29
iii

jitomate tratadas con distintas concentraciones de prosisteminas
recombinantes (Dombrowski et al., 1999).
Figura 9. Estructura de la nicotina y nornicotina.
32
Figura 10. Estructuras de compuestos fenólicos involucrados en defensa.
34
Figura 11. Estuctura del capsidiol y su producto de degradación,
dihidrocapsenona, y otros terpenos involucrados en defensa.
36
Figura 12. Niveles de inhibidores de tripsina (IT), PFO y
poligalacturonasas (PG), inducibles por daño en hoja local (HL) y hoja
sistemica (HS) en hojas de tabaco silvestres (Wt), y en plantas
transformadas con LePS y LeSYS.
38
Figura 13 . I. Fenotipo mostrado por plantas de tabaco transformadas con
sistemina de jitomate (Nt-LeSYS).
40
Figura 14 . II. Fenotipo mostrado por plantas de tabaco transformadas con
sistemina de jitomate (Nt-LeSYS).
41
Figura 15. III. I. Fenotipo mostrado por plantas de tabaco transformadas
con sistemina de jitomate (Nt-LeSYS).
42
Figura 16. Larvas de Manduca sexta antes y 6 días después de haber
sido alimentadas con plantas de tabaco sin transformar y con plantas Nt-
LePS y Nt-LeSYS.
44
Figura 17 . Análisis del crecimiento de larvas de M. sexta alimentadas con
plantas sin transformar y plantas Nt-LePS y Nt-LeSYS.
44
iv

Figura 18. Vista general de la magnitud del daño causado por larvas de
M. sexta en plantas de tabaco sin transformar (control) y transformadas
(Nt-LePS y Nt-LeSYS).
45
Figura 19. Cromatograma típico obtenido al separar extractos
metanólicos derivatizados de hoja de Nicotiana tabacum por GC/MS. La
nicotina emergió después de 15.83 min (*); timol después de 15.71 min
(*); nornicotina después de 20.05 min (*); el posible producto de
degradación del capsidiol (capsenona) después de 32.78 min (*), y el
ácido clorogénico después de 48.83 min (*).
57
Figura 20. a) Cromatograma de GC/MS del estándar puro de nicotina,
con un tiempo de retención de 15.80 min (*), y b) su correspondiente
espectro de masas.
58
Figura 21. a) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando a nicotina con un tiempo de retención de
15.83 min (*), y b) su correspondiente espectro de masas.
59
Figura 22. a) Cromatograma de GC/MS del estándar puro del ácido
clorogénico, con un tiempo de retención de 48.90 min (*), y b) su
espectro de masas.
60
Figura 23. a) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando al ácido clorogénico con un tiempo de
retención de 48.83 min (*), y b) su correspondiente espectro de masas.
61
Figura 24. a) Cromatograma de GC/MS del estándar puro de capsidiol
[32.35 min (*)] derivatizado y un producto de degradación estable
posiblemente dihidrocapsenona, con un tiempo de retención de 32.18
min (*), y b) el espectro de masas correspondiente al pico surgido a los
62
v

32.18 min.
Figura 25. a) Cromatograma de GC/MS del estándar puro de capsidiol
[32.35 min (*)] derivatizado y un producto de degradación estable
posiblemente dihidrocapsenona, con un tiempo de retención de de 32.18
min (*), y b) el espectro de masas correspondiente al pico surgido a los
32.35 min.
63
Figura 26. a) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando a un posible producto de degradación
del capsidiol con un tiempo de retención de 32.78 (*), y b) su
correspondiente espectro de masas.
64
Figura 27. a) Cromatograma de GC/MS del estándar puro de nornicotina
derivatizado, con un tiempo de retención de 20.54 min (*), y b) su
correspondiente espectro de masas.
65
Figura 28. a) Cromatograma típico de GC/MS de un extracto metanólico
derivatizado de hojas de Nicotiana tabacum, identificando a la nornicotina
con un tiempo de retención de 20.57 min (*), y b) su correspondiente
espectro de masas.
66
Figura 29. Niveles de nicotina en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas
a daño mecánico (5 día). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
69
Figura 30. Comparación entre los niveles de nicotina en plantas WT y
transgénicas adultas, 5 días después de haber sido expuestas a
herbivoría por larvas de M. sexta durante 24 h. Los asteriscos indican la
diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
71
vi

Figura 31 . Comparación entre los niveles de nicotina en plantas WT y
transgénicas adultas medidos al cumplirse el ciclo de vida de Bemisia
tabaci (28 días después de la oviposición). Los asteriscos indican la
diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
73
Figura 32. Niveles de ácido clorogénico en plantas (jóvenes y adultas)
WT y transgénicas intactas (0 h) y cinco días después de haber sido
sometidas a daño mecánico (5 día). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
77
Figura 33. Comparación entre los niveles de ácido clorogénico en plantas
WT y transgénicas adultas, 5 días después de haber sido expuestas a
herbivoría por larvas de M. sexta durante 24 h. Los asteriscos indican la
diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
79
Figura 34. Comparación entre los niveles de ácido clorogénico en plantas
WT y transgénicas adultas medidos al cumplirse el ciclo de vida de
Bemisia tabaci (28 días después de la oviposición). Los asteriscos indican
la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
81
Figura 35. Niveles de capsidiol (cuantificado indirectamente en forma de
un posible producto de degradación) en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 día) y cinco días después de haber sido
sometidas a daño mecánico (5 día). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
84
Figura 36. Comparación entre los niveles de capsidiol (cuantificado
indirectamente en forma de un posible producto de degradación), en
plantas WT y transgénicas adultas, 5 días después de haber sido
expuestas a herbivoría por larvas de M. sexta durante 24 h. Los
86
vii

asteriscos indican la diferencia estadísticamente significativa (P ≤ 0.05*; P
≤ 0.01 **).
Figura 37. Comparación entre los niveles de capsidiol (cuantificado
indirectamente en forma de un posible producto de degradación) en
plantas WT y transgénicas adultas, medidos al cumplirse el ciclo de vida
de Bemisia tabaci (28 días después de la oviposición). Los asteriscos
indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
Figura 38 . Niveles de nornicotina en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas
a daño mecánico (5 día). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
89
91
Figura 39. Fenoles solubles totales en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas
a daño mecánico (5 día). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
94
Figura 40. Comparación entre los niveles de fenoles totales solubles en
plantas WT y transgénicas adultas, 5 días después de haber sido
expuestas a herbivoría por larvas de M. sexta durante 24 h. Los
asteriscos indican la diferencia estadísticamente significativa (P ≤ 0.05*; P
≤ 0.01 **).
95
Figura 41. Comparación entre los niveles de fenoles solubles totales en
plantas WT y transgénicas adultas, medidos al cumplirse el ciclo de vida
de Bemisia tabaci (28 días después de la oviposición). Los asteriscos
indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
97
Figura 42. Curva de calibración de nicotina. 132
viii

Figura 43. Curva de calibración de ác. clorogénico.
Figura 44. Curva de calibración de capsidiol.
Figura 45. Curva de calibración de nornicotina.
133
134
135
ix

RESUMEN
Las plantas de tabaco del género Nicotiana (Solanaceae) contienen diversos
metabolitos secundarios. Los grupos más importantes son los alcaloides, terpenos y
componentes fenólicos. Se sabe que la síntesis de novo de muchos de estos
compuestos se incrementa dramáticamente como respuesta al daño mecánico o
ataque por herbívoros y/o patógenos y que este cambio es inducido en muchos
casos por ácido jasmónico, el cual funciona como una señal mediadora de cambios
en el metabolismo secundario en respuesta a muchos tipos de agobio biótico y
abiótico (Gundlach et al., 1992; Hildman et al., 1992; Reinbothe et al., 1994).
Investigaciones previas demostraron que la transformación de plantas de tabaco (N.
tabacum) con prosistemina (LePS) y sistemina (LeSYS) de jitomate, resultaron ser
altamente resistentes a herbivoría por larvas de Manduca sexta. Se sugirió que la
sobreexpresión LePS y LeSYS induce cambios en las respuestas de defensa contra
estos insectos en tabaco, entre los cuales podría incluirse la acumulación de
metabolitos secundarios considerados como fitoalexinas o fitoanticipinas en esta
especie. Así mismo, una de las líneas analizadas, la Nt-LeSYS 03, presentó un
fenotipo anormal caracterizado por enanismo, floración temprana, presencia de
regiones necróticas similares a una reacción hipersensible en la superficie de la
hoja y una germinación muy lenta, lo cual también podría estar relacionado con
alteraciones en la síntesis de estos metabolitos, en particular compuestos
fenilpropanoides. Para explorar esta posibilidad, se analizaron los niveles
constitutivos e inducibles de metabolitos relacionados con defensa contra
herbívoros y/o patógenos (nicotina, nornicotina, ácido clorogénico, capsidiol y
fenoles solubles totales) en follaje y raíz. Al analizar y cuantificar los niveles de
estos metabolitos por distintos tratamientos (daño mecánico, herbivoría con
Manduca sexta e infestación con mosquita blanca), se detectaron variaciones de
acuerdo a la etapa de desarrollo de la planta, tipo de tratamiento y genotipo.
En el tratamiento con daño mecánico en plantas adultas, los niveles de nicotina y
capsidiol, medido como uno de sus productos de degradación más estables x

(posiblemente capsenona), fueron más altos en las plantas transformadas que las
plantas WT (con algunas excepciones en la línea Nt-LeSYS 03). También se
detectó un aumento significativo de ácido clorogénico (ACG) en todos los tejidos
analizados de las plantas WT. A su vez, la herbivoría por larvas de M. sexta redujo
significativamente, en ocasiones a niveles no detectables, los niveles de nicotina,
nornicotina y ACG. El efecto fue particularmente marcado en las plantas
transformadas con LePS y LeSYS. La nornicotina no se detectó en los diferentes
genotipos de plantas sometidos a herbivoría o infestación por B. tabaci, mientras
que el capsidiol aumentó considerablemente en plantas infestadas con mosquita
blanca principalmente en la línea transgénica Nt-LePS.
Estos resultados sugieren que, de algún modo no identificado aún, la expresión de
LePS o LeSYS modifica las rutas biosintéticas del metabolismo secundario en
plantas de N. tabacum.
xi

1
I. INTRODUCCIÓN
Las plantas están continuamente expuestas a diversos cambios
ambientales, incluyendo el ataque por numerosos insectos herbívoros. El daño
mecánico o por herbívoros y la invasión por patógenos activan sistemas de
defensa de la planta. Para que éstos se activen, las señales provocadas por
tales ataques son transmitidas desde el tejido dañado hacia el resto de la planta
(respuesta sistémica) o son diseminadas sólo localmente, induciendo cambios
en la vecindad del sitio dañado (respuesta local). Estas señales pueden
provocar reacciones de defensa y cambios específicos en la planta que
conduzcan hacia una mayor resistencia contra organismos invasores (Baldwin et
al., 1994, 1997; Walling, 2000).
Las solanáceas constituyen una familia de plantas en las cuales los
diversos mecanismos de defensa empleados contra insectos dañinos y
patógenos han sido estudiados con más detalle. Un ejemplo de ello son los
diversos metabolitos secundarios que se acumulan en diferentes especies de
esta familia, ya sea constitutivamente o en respuesta a un ataque. Entre los tipos
de metabolitos más importantes presentes se encuentran los alcaloides,
terpenos y compuestos fenólicos. Al parecer, la acumulación de estos
metabolitos se encuentra bajo control genético y es dependiente de las
condiciones ambientales (Tso, 1972). Los metabolitos secundarios con función
defensiva actúan de modos tan diversos como sus estructuras. Por ejemplo, la
nicotina y los alcaloides relacionados a esta pueden funcionar como toxinas
defensivas ya que interfieren con el funcionamiento de receptores de acetil colina
en el sistema nervioso de los herbívoros (Shoji et al., 2000; Liu et al., 2005),
mientras que el ácido clorogénico tiene un impacto sobre la calidad nutricia de
las proteínas vegetales ingeridas por insectos herbivoros debido a su capacidad
de reaccionar con aminoácidos alquilatables, como la cisteína, metionina,
histidina y lisina, una vez que ha sido oxidado a su forma quinoide por polifenol
oxidasas (PFO) (Felton et al., 1992; Duffey y Scout, 1996).
Se conoce que diferentes genotipos de tabaco (Nicotiana tabacum) y
especies silvestres de Nicotiana varían en su habilidad para acumular y

2
almacenar nicotina (Saunders y Bush, 1979; Sinclair et al., 2004). Además, se ha
demostrado que el ataque por insectos herbívoros en N. sylvestris y N. attenuata
incrementa dramáticamente la síntesis y acumulación de nicotina (Baldwin et al.,
1998,1999). Ahora se sabe que la acumulación de nicotina inducida por herida
es regulada por el ácido jasmónico (AJ), este último se produce inicialmente en
la hoja y posteriormente transportado hacia la raíz, que es el órgano dónde se
sintetiza la nicotina y de donde se distribuye al resto de la planta (Baldwin et al.,
1997; Zhang y Baldwin, 1997). En N. attenuata, el tratamiento con jasmonato
indujo la acumulación de nicotina en la planta, la cual se relacionó positivamente
con una resistencia durable hacia el herbívoro (Baldwin et al., 1998). Estos
resultados están de acuerdo con la multiplicidad de funciones que se le han
asignado al AJ, considerado como una hormona vegetal capaz de modular, entre
otras respuestas, el metabolismo secundario en respuesta a estrés biótico y
abiótico (Creelman et al., 1997). Esta capacidad fue ampliamente confirmada en
estudios en los cuales se reportó que la aplicación exógena de AJ indujo la
acumulación de una gran variedad de metabolitos secundarios en cultivos
celulares y en plantas de diversas especies, incluyendo alcaloides (Gundlanch
et al., 1992; Aerts et al., 1994) y compuestos fenólicos. (Lee et al., 1997).
Además de la nicotina y ACG, otros reportes han mostrado una estrecha
relación entre la síntesis y acumulación de los diterpenos glicosilados en tabaco,
como los 16-hidroxigeranil linalool A y B, y la resistencia contra Heliothis
virescens. En concordancia con su función defensiva, este metabolito solo ha
sido detectado en genotipos resistentes a este herbívoro (Snook et al., 1997).
En jitomate (Lycopersicon esculentum) se descubrió que la sistemina,
liberada a partir de la prosistemina, su proteína precursora, es un polipéptido
involucrado en la inducción sistémica de proteínas en respuesta al daño, las
cuales se sugiere que confirieren resistencia contra agresores bióticos. Al igual
que la respuesta al daño mecánico y a otros evocadores activos como
oligogalacturónidos derivados de la fragmentación de la pared celular, la actividad
biológica de la sistemina es dependiente de AJ. En un trabajo previo, del cual se
derivó la presente investigación, se generaron plantas transgénicas de tabaco
capaces de sobreexpresar constitutivamente tanto la prosistemina (LePS) como la
sistemina (LeSYS) de jitomate. Estas plantas mostraron diferentes grados de
resistencia a herbivoría por larvas de Manduca sexta, la cual curiosamente no se

3
correlacionó muy claramente con la acumulación de altos niveles de proteínas
defensivas, como polifenoloxidasas (PFO), inhibidores de tripsina (IT) y
quimotripsina (IQT), solo se detectaron niveles altos en una de las líneas
sobreexpresantes de LeSYS. Éstas últimas presentaron, además, un fenotipo
severamente anómalo, caracterizado por enanismo, morfología alterada de las
hojas y flores, floración precoz y esterilidad (Rocha-Granados, 2004). De manera
que en este trabajo se analizó la posibilidad, previamente demostrada en otras
especies de plantas, de que la resistencia diferencial contra larvas de insectos
herbívoros así como las alteraciones morfológicas y fisiológicas observadas en
una de las líneas de N. tabacum sobreexpresantes de LeSYS, pudieran haberse
debido a una modificación en la síntesis y acumulación de metabolitos
secundarios causada por la expresión de LePS y LeSYS. Para ello, se
compararon los patrones de acumulación de nicotina, ACG, nornicotina (NN),
dihidrocapsenona y compuestos fenólicos totales entre plantas no modificadas
(WT) y plantas transformadas en diferentes estadios de desarrollo y en respuesta
a daño mecánico o infestación por insectos. Los resultados obtenidos indican que
la expresión de LePS y LeSYS en N. tabacum modifica el patrón de acumulación
de metabolitos secundarios, lo que podría estar relacionado con la resistencia
diferencial observada contra larvas de insectos herbívoros, e inclusive, con el
fenotipo severo que caracteriza a una de las líneas sobreexpresantes de
sistemina (Nt-LeSYS 03).

4
2. ANTECEDENTES
2.1. Mecanismos de defensa
Las plantas, en su medio natural han desarrollado un gran número de
estrategias defensivas contra una gran variedad de organismos invasores y de
agentes abióticos causantes de una diversidad de agobios ambientales. Las
estrategias defensivas están basadas tanto en la erección de barreras físicas y
químicas preformadas, así como en la síntesis de metabolitos defensivos en
respuesta a estímulos asociados con el organismo invasor (Ebel y Cosio, 1994).
El primer grupo lo constituyen las defensas constitutivas, las cuales se
encuentran presentes durante todo el desarrollo de la planta. Éstas comprenden
barreras tanto de tipo físico como químico incluyendo espinas, tricomas, la
cutícula superficial de la hoja y ceras protectoras de diversos órganos, así como
metabolitos secundarios existentes en plantas sanas, conocidos como
fitoanticipinas. Entre éstos están las saponinas, los glicósidos cianogénicos,
cumarinas y los glucosinolatos, los cuales son tóxicos a atacantes no
especializados (Osbourn, 1996).
Se ha sugerido que los metabolitos secundarios, más que subproductos
metabólicos, tienen una función en la interacción de las plantas con su ambiente
y otros organismos para proveer una defensa contra la infección, depredación o
agobio ambiental (Rhodes, 1994). Cabe mencionar que las defensas químicas
se dividen en dos categorías: los metabolitos secundarios y proteínas (Roberts y
de Bruxelles, 2001).
Un segundo grupo lo comprenden las llamadas defensas inducibles; esto
significa que sólo se producirán como respuesta al reconocimiento del organismo
agresor. Este reconocimiento requiere la interacción entre evocadores o
inductores (compuestos liberados a partir de los organismos agresores o de las
propias plantas a causa del daño que éstos les provocan) y sus receptores
específicos en las plantas (Agrawal et al., 1999). Numerosas proteínas
relacionadas con defensa, cuya acumulación es regulada por AJ y que incluyen

5
enzimas involucradas en la síntesis de alcaloides, inhibidores de proteasas,
enzimas de carácter defensivo como la polifenol oxidasa (PFO) y péptidos
antimicrobianos como las defensinas y tioninas, pertenecen al grupo de defensas
inducibles.
Las respuestas defensivas inducibles pueden activarse de manera
localizada, en el área de daño, o bien tener efecto sistémico al activarse en la
planta completa. La eficiencia de los mecanismos defensivos depende
estrictamente de la velocidad de proliferación del agresor y del tiempo necesario
para el reconocimiento del mismo por el huésped y de la subsiguiente activación
de las defensas (Dong, 1998). Como ejemplo de las defensas inducibles se
encuentra la síntesis y acumulación de especies reactivas de oxígeno,
metabolitos secundarios como compuestos fenólicos, alcaloides, terpenos, etc.
(agrupados bajo el término fitoalexinas), enzimas hidrolíticas como quitinasas y
glucanasas, y la acumulación de proteínas relacionadas con patogénesis,
conocidas como proteínas RP (Osbourn., 1996; Agrawal et al., 1999; Richael y
Gilchrist., 1999).
Estos tipos de mecanismos de defensa se mencionarán más
ampliamente en los siguientes subíndices.
2.1.1. Tipos de respuestas de defensa en planta
Se han identificado diversas respuestas de defensa en varias especies de
plantas empleadas como modelo de estudio. Éstas pueden clasificarse en tres
clases: (1) innata, la cual está involucrada en el reconocimiento y/o generación
de señales internas de la planta, (2) local inducida, que tiene lugar en el sitio de
daño así como en la periferia de éste, teniendo efectos adversos directos sobre
el organismo agresor y (3) sistémica inducida, la cual abarca el tejido distante
y/o un órgano no afectado de la planta (Halbrock et al., 1995).

6
2.1.1.1. Respuesta innata y respuesta específica, gen por gen
La respuesta innata, también conocida como respuesta independiente del
huésped, confiere una protección robusta, principalmente contra patógenos. Esta
se puede expresar durante diferentes etapas del proceso invasivo del patógeno.
La primera línea de defensa la constituyen las barreras físicas preformadas (e.
g., tricomas y espinas) junto con características topograficas y químicas (e. g.,
ceras) de la superficie de los tejidos vegetales, particularmente de hojas. Si estas
barreras son superadas, las plantas activan una serie de cambios de índole
defensivo en respuesta a evocadores ampliamente reconocidos por ellas, los
cuales son liberados durante el proceso invasivo, como fragmentos de la pared
celular de plantas y hongos, y enzimas hidrolíticas (e. g., celulasas, proteasas,
pectinasas). Éstas también son capaces de identificar ciertos patrones
moleculares asociados exclusivamente con patógenos (PMAP), como
glicoproteínas y polipéptidos derivados de éstas (e. g., el péptido PEP-13 que se
genera a partir de una transglutaminasa de Phytophthora sojae), componentes
del flagelo y lipopolisacáridos de bacterias Gram negativas, peptidoglicanos y
ácido lipoteico de bacterias Gram positivas, fragmentos de ADN no metilados,
proteínas bacterianas (e. g., harpinas) y de oomicetos (e. g., elicitinas). Si todo
esto es superado por el invasor y como última opción, la planta activa un
mecanismo de resistencia mediado por la interacción entre los productos de los
genes de avirulencia (Avr) y de resistencia (R) de patógenos y plantas,
respectivamente. En insectos, plantas y vertebrados, la identificación de algunos
PMAP, como por ejemplo la flagelina, está mediada por cinasas similares a
receptores (“Toll-like”), caracterizadas por una estructura en la que abundan
secuencias repetidas ricas en leucina.
La resistencia innata involucra a todo un género o especie de plantas; es
compleja, ancestral, estable y heredable. Por el contrario, la resistencia
específica o del tipo gen por gen, está limitada a ciertos genotipos en una
especie normalmente susceptible, es parásito específica y conferida por un solo
gen R que reconoce a un único gen Avr y, por lo mismo, es menos duradera
(Gómez-Gómez y Boller, 2002; Thordal-Christensen, 2003; Parker, 2003).
Una vez que se establece la unión del evocador o PMAP al receptor, los
cambios más importantes a través de la membrana plasmática son la activación

7
del flujo de iones H+,
K+, Cl- y Ca2+ desde y hacia la célula y la formación de
H2O2 (estallido oxidativo) (Apostol et al., 1989; Nürberger et al., 1994; Roberts y
de Bruxelles, 2001). Después de 5 a 30 minutos, se inicia la fosforilación
dependiente de calcio de algunas proteínas, que inducen una síntesis rápida de
etileno y la activación transcripcional de muchos genes relacionados con la
defensa (Dietrich et al., 1990). De manera temporal, el estallido oxidativo y la
fosforilación/desfosforilación de proteínas, parecen estar controlados por la
activación de canales iónicos, pero su posible relación causal con otros
componentes de la cadena de transducción de señales que conducen a la
activación de respuestas, como por ejemplo proteínas que enlazan GTP, inositol,
AMPc, etc., aún se desconoce. Las células vegetales tienen la capacidad de
reconocer un patógeno potencial y de transmitir esta información dentro de la
célula y células vecinas, para mantener y optimizar las respuestas de defensa
(Basse et al., 1993; Nürberger et al., 1994). La interacción entre evocador-
receptor es el punto de partida para la traducción de señales que activa la
expresión de genes involucrados en la resistencia, ya sea del tipo “no especifico”
que como se indicó anteriormente se induce por diversos evocadores no
específicos reconocidos por numerosas especies, e inclusive géneros, de
plantas resistentes a toda una raza de patógenos, o los de tipo “especifico”,
donde el evocador es el producto de un gen de avirulencia (Avr) del patógeno, y
el receptor, el correspondiente producto del gen de resistencia R, en la planta
(Grant y Loake, 2000).
En la mayoría de los casos, la interacción entre los productos de los
genes R y Avr inicia cascadas de señalización que dan lugar a la reacción
hipersensible (RH) y resistencia sistémica adquirida (RSA). La RH consiste en
una necrosis rápida y localizada en el sitio de daño, actúa directamente contra el
patógeno, mediante la citotoxidad de las especies reactivas de oxígeno, como el
anión superóxido (O2-) y el peróxido de hidrógeno (H2O2), que participan en el
estallido oxidativo (Apostol et al., 1989). Además, se caracteriza por la
acumulación de compuestos fenólicos alrededor de las células atacadas,
fragmentación del ADN, condensación citoplasmática y muerte celular
programada, procesos similares a la apoptosis que se observa en células
animales (Birch et al., 1999).

8
Junto con la RH, en la planta se pueden presentar diferentes cambios
estructurales, moleculares, bioquímicos y/o fisiológicos que ocurren poco tiempo
después del ataque patogénico y tienen lugar tanto en el sitio de daño como en
la periferia (respuestas locales) o, más tardíamente, en zonas distantes a éste
(respuestas sistémicas).
2.1.1.2. Respuesta local
La respuesta local consiste en una rápida acumulación de enzimas,
proteínas estructurales y/o metabolitos, como consecuencia de una activación
transcripcional que es mediada por el reconocimiento del patógeno. Esta
activación local de genes es restringida a una pequeña área definida alrededor
del sitio de ingreso del patógeno (Bowles, 1990; Geoffroy et al., 1990; Ryan y
Jagendorf, 1995).
Una respuesta típica en este sitio localizado es la síntesis de novo de
enzimas biosintéticas de compuestos tóxicos para el patógeno, tales como las
enzimas del metabolismo de los fenilpropanoides, como la fenilalanina amonio
liasa (FAL), la 4-cumarato: CoA ligasa (4CL) y otras enzimas involucradas en la
síntesis de fitoalexinas y otros metabolitos secundarios (Nicholson y
Hammerschmidt, 1992; Freytag, 1994).
Tambien mediante este mecanismo son sintetizadas otras proteínas,
como las glicoproteínas ricas en hidroxiprolina (GPRH), proteínas ricas en glicina
(PRG), proteínas relacionadas con la patogénesis (RP), intra y extracelulares,
así como quitinasas, β-1,3-glucanasas, peroxidasas, lipoxigenasas, proteasas,
inhibidores de proteasas y amilasas, poligalacturonasas y otras proteínas
antimicrobianas y/o tóxicas a insectos (Geoffroy et al., 1990; Linthorst, 1991;
Stintzi et al., 1993).
Como ya se mencionó, el reconocimiento del evocador ocasiona la
activación de una o más rutas de transducción de señales, y por último la
síntesis de compuestos relacionados con defensa. De manera rápida y local
surgen eventos comunes a distintas rutas de transducción de señales, donde se
incluyen: apertura de canales iónicos y entrada de calcio, alteración del potencial
de membrana (despolarización o hiperpolarización). Estos eventos tempranos

9
son acompañados posteriormente por la síntesis de AS, AJ y/o otros compuestos
que son señales necesarias para la activación génica (Hammond-Kosack y
Jones, 1996).
Los diferentes cambios estructurales, moleculares, bioquímicos y/o
fisiológicos que ocurren poco tiempo después del ataque patogénico, tienen
lugar tanto en el sitio de daño como en su periferia (respuestas locales) o, más
tardíamente, en zonas distantes a éste (respuestas sistémicas).
2.1.1.3. Respuesta sistémica
Uno de los aspectos más elusivos y fascinantes en respuesta a daño es la
habilidad de generar señales en el sitio de herida y que se transmitan a partes
distantes de la planta. La respuesta a patógenos mejor caracterizada es la
resistencia sistémica adquirida (RSA), la cual le confiere resistencia a la planta
contra un amplio rango de patógenos e involucra la inducción de distintas
proteínas RP (Ryals et al., 1996).
El ácido salicílico (AS), un producto de la ruta de los fenilpropanoides, ha
mostrado ser esencial en la inducción de la RSA (Gaffney et al., 1993; Vernooij
et al., 1994; Pallas et al., 1996). Se ha demostrado que un incremento en los
niveles de AS precede a la RSA, y que dicho incremento es necesario y
suficiente para el establecimiento de la misma. Recientes evidencias han
sugerido que también existen rutas independientes de AS (Heo et al., 1999).
Tanto en tabaco como en Arabidopsis, el AS es suficiente y necesario
para la inducción de RSA. Esto ha sido aprovechado en la agricultura, en la cual
se emplean inductores sintéticos como el BTH y el INA, que presentan gran
similitud estructural con AS, para inducir un estado de resistencia contra
patógenos en numerosos cultivos de interés comercial (Lawton et al., 1996;
Friedrich et al., 1996; Gorlach et al., 1996). El papel del AS en la inducción del
RSA se comprobó en plantas de tabaco capaces de expresar el gen nahG, que
codifica una salicilato hidrolasa bacteriana; estas plantas presentaron bajos
niveles de AS, fueron incapaces de expresar RSA y presentaron más
susceptibilidad a una gran variedad de patógenos (Delaney et al., 1997; Gaffney
et al., 1993).

10
Además de la RSA, existe otro tipo de respuesta denominada Resistencia
Sistémica Inducida (RSI) que también provee protección contra una amplia gama
de patógenos. Esta respuesta, que es inducida por rizobacterias promotoras del
crecimiento, no depende de AS, no está asociada a la RH ni a la síntesis de
proteínas RP, está regulada por el ácido jasmónico, el etileno y la proteína
reguladora NPR-1 (Pieterse et al., 1996, 1998; Van Loon et al., 1998; Knoester et
al., 1998).
Se cree que el etileno funciona como inductor en la síntesis de
compuestos, genes o estructuras defensivas como lignina, proteínas RP y
fenilalanina amonio liasa (FAL). Por otra parte, existe una ruta defensiva
comúnmente conocida como la ruta octadecanoica que involucra AJ o su éster
metílico (MeJA) para inducir, principalmente, la síntesis de inhibidores de
proteasas (IP) en respuesta a herbivoría (Peña-Cortés et al., 1989; Farmer y
Ryan, 1992).
Existen varios ejemplos de la inhibición de una vía por la otra. Por
ejemplo, en plantas heridas tratadas simultáneamente con AS, se inhibe la
acumulación de IP y PFO, que son respuestas típicas producidas por herbivoría,
y reguladas por AJ (Stout et al., 1999). A su vez, también se ha observado la
interferencia del AJ en la resistencia mediada por AS contra ciertos patógenos,
manifestándose como la disminución de la síntesis de algunas proteínas RP
(Sano y Ohashi, 1995; Niki et al., 1998).
2.2. Mecanismos de defensa en tabaco
El tabaco es atacado en cualquier etapa de su desarrollo y en todas los
órganos de la planta por diversos agresores incluyendo hongos, bacterias, virus,
nemátodos, afidos (Myzus nicotianiae; M. persicae) y otros insectos chupadores
e insectos masticadores, entre los que se encuentra Manduca sexta, que es una
especie tolerante a la nicotina y Heliothis virescens (Johnson et al., 1992; Snook
et al., 1997).
Como ya se ha descrito, la resistencia o tolerancia de las plantas hacía los
insectos herbívoros y patógenos está mediada por mecanismos de defensa
inducidos o constitutivos (Mauricio et al., 1997; Buell, 1998). En tabaco, como en

11
otras plantas, las defensas inducibles pueden clasificarse como directas e
indirectas. Dentro del primer tipo se incluyen los inhibidores de proteasas, (van
Dam et al., 2001; Glawe et al., 2003) diterpenos glicosilados (Keinänen et al.,
2001) y metabolitos secundarios como la nicotina (Winz y Baldwin, 2001;
Steppuhn et al., 2004) capsidiol (Guedes et al., 1982; Bohlmann et al., 2002) y
ácido clorogénico (Tanguy y Martin, 1972; Lou y Baldwin, 2003).
A su vez, la defensa indirecta se considera como aquella que depende de
un tercer nivel trófico, representado por insectos depredadores o parásitos de los
insectos herbívoros, los cuales son atraídos hacia la planta atacada mediante la
emisión inducida de compuestos volátiles (Halitschke et al., 2000). En N.
attenuata, el daño mecánico y el ataque por M. sexta ocasiona un incremento de
los niveles de AJ, el cual está relacionado con la acumulación de metabolitos
secundarios, inhibidores de proteasas y la síntesis de novo de volátiles. En esta
especie, la inducción de defensas se presenta tanto a nivel local como sistémico,
particularmente en las hojas conectadas filotácticamente, mientras que los
volátiles se inducen en toda la planta. Sin embargo, el ataque a N. attenuata por
M. sexta no origina un aumento de nicotina ya que el incremento simultáneo de
etileno inhibe la expresión de putrescina N-metiltransferasa, que es una de las
enzimas clave en la biosíntesis de nicotina. Este patrón de expresión de
respuestas defensivas ha sido estudiado con particular detalle en especies
silvestres de Nicotiana (e.g., N. sylvestris y N. attenuata) (McCloud y Baldwin,
1997; Lou y Baldwin, 2003; Qu et al., 2004), (Ver Figura 1).

12
DEFENSAS EN TABACODEFENSAS EN TABACO
ATAQUE CON
Manduca sexta
Baldwin y col., 2003
nicotina
Defensas indirectas
Defensas directas
Metabolitos secundarios
Diterpenosglicosilados,etc.
VolatilesMeJA
AJ
Et
daño
regurgitante
Putrescina PMT
IPs
Figura 1. Reconocimiento del ataque de M. sexta por N. attenuata, incremento
de los niveles de AJ y etileno e inducción de defensas directas e indirectas
(modificado de Baldwin et al., 2003).

13
2.3. Respuestas de resistencia en plantas contra el ataque de plagas
El daño implicado por herbívoros requiere de múltiples estrategias de
defensa en la planta. El sitio de daño debe protegerse y la respuesta de defensa
aumentar contra el herbívoro y patógenos oportunistas (Roberts y de Bruxelles,
2001). En el caso específico de los insectos herbívoros, se ha observado que
éstos inducen una serie de defensas en las plantas tanto a nivel local como a
nivel sistémico (Gatehouse, 2000). Por ejemplo, se ha demostrado que el daño
causado por el ataque de insectos masticadores activa respuestas de defensa
típicas en ciertas especies de plantas. Entre estas respuestas, se encuentran la
activación de genes que codifican para inhibidores de proteasas y polifenol
oxidasas, así como de proteínas asociadas a proteólisis controlada, como son
las proteínas tipo ubiquitina (Bowles, 1990; Ryan, 1990).
2.3.1. Inhibidores de proteasas
Los IP son proteínas con gran afinidad por enzimas proteolíticas, capaces
de inhibir fuerte y específicamente su actividad enzimática (Richardson, 1991).
Los inhibidores de proteasas se acumulan de manera constitutiva, pueden ser
inducibles por daño causado mecánicamente, por insectos o por patógenos. Las
evidencias experimentales indican que su efecto protector radica en la inhibición
de enzimas que son esenciales para la digestión de proteínas (Ryan, 1981; de
Bruxelles y Roberts, 2001). Éstos fueron primeramente descritos con detalle en
plantas solanáceas. Sin embargo, hoy en día se ha encontrado que los IP son
expresados en semillas, tubérculos y tejidos vegetativos de más de 100 especies
diferentes de plantas, muchos de los cuales se acumulan después de sufrir algún
tipo de daño (Kessler et al., 2002).
La clasificación de los IP se basa en el tipo de proteasa (o proteasas) que
son capaces de inhibir: serina, cisteína, aspártico y las metalo proteasas. Los
inhibidores de estas enzimas producidas por plantas afectan el crecimiento y
desarrollo de plagas al interferir en la degradación de proteínas de origen foliar.
El argumento más aceptado es que el efecto defensivo se produce como
consecuencia de los efectos antinutricios que se presentaran en los insectos

14
alimentados con plantas que sintetizan altas concentraciones de IP, los cuales
están relacionados con la pérdida de la asimilación de algunos aminoácidos
esenciales, especialmente los sulfurados, debido a una secreción excesiva de
tripsina (Ryan, 1990).
La síntesis de estas enzimas se inicia con la liberación de evocadores,
inducidos por algún tipo de daño; la respuesta se lleva a cabo de manera local y
al mismo tiempo se inicia la activación de mecanismos de transducción de
señales para iniciar la respuesta sistémica (Schaller y Ryan, 1995).
Se considera que la amplia distribución de los IP en tejidos de
almacenamiento y en distintas familias de plantas, su inducibilidad por daño
mecánico y herbivoría, así como su efecto protector en plantas transgénicas, son
criterio suficiente para atribuirles una función como proteínas de defensa contra
herbívoros.
2.3.2. Polifenol oxidasas
Las polifenol oxidasas son un grupo de enzimas inducibles que oxidan un
amplio rango de compuestos fenólicos en plantas (Vaughn et al., 1988). La
enzima polifenol oxidasa (PFO) se clasifica como proteína antinutricional dentro
de las llamadas SWRPs (de sus siglas en inglés, “Systemic Wound Response
Proteins”) o proteínas en respuesta a daño que son proteínas que se expresan
como respuesta al ataque por herbívoros (Ryan, 2000).
La enzima PFO, al ser ingerida por los herbívoros, junto con compuestos
fenólicos y la combinación de inhibidores de proteasas, forman una barrera muy
efectiva para la digestión de proteínas por insectos (Graham et al., 1986; Felton
et al., 1992; Bergey et al., 1996). El efecto antinutritivo se basa en la capacidad
que poseen para destruir nutrientes esenciales en las dietas de los insectos. La
eficiencia de las enzimas oxidativas, fenol oxidasas, peroxidasas y
lipooxigenasas (LOX), depende del ambiente químico existente en el intestino
del insecto, del potencial Redox y de los niveles de antioxidantes. Además, la
calidad y cantidad de proteína presente en la dieta ejercen influencia sobre los
efectos oxidativos en las enzimas de los herbívoros (Stout y Duffey, 1996).

15
Durante el proceso de alimentación del insecto, las PFO tienen acceso a
sus sustratos y generan orto-quinonas a partir de compuestos fenólicos
presentes en la dieta. Estas tienen un efecto nocivo en el huésped ya que
pueden formar enlaces covalentes con grupos sulfhídrilo de aminoácidos
sulfurados y con aminoácidos básicos, como la lisina. La modificación química
de estos aminoácidos provoca eventualmente precipitación de las proteínas, lo
cual altera de manera significativa la digestión y la asimilación de nitrógeno en el
insecto (Felton et al., 1992).
2.3.3. Oligosacáridos
La mayor parte de los patógenos y herbívoros que atacan plantas,
secretan numerosas enzimas degradativas, cuya acción provoca la generación
de fragmentos de componentes celulares (como de la pared celular), que actúan
como evocadores bióticos (Stout y Bostock, 1999). Se ha observado que tanto
los oligogalacturónidos (OGA), provenientes de los fragmentos de la pared
celular, como los fragmentos de quitina y quitosanas, derivados de las paredes
celulares de patógenos, pueden activar a los genes IP. En algunos bioensayos
realizados con diferentes OGA, se observó que moléculas con un grado de
polimerización de 2 unidades de ácido galacturónico fueron capaces de inducir a
los genes IP. Se ha sugerido que los OGA actúan como inductores en la
expresión de los genes IP solamente a nivel local. Esto se atribuye a su poca
movilidad, relacionada con su alta afinidad por la pared celular, y al hecho de
que sólo pueden ser liberados en el sitio donde el daño ha sido inflingido en las
plantas por insectos o por patógenos y donde se generan las enzimas
necesarias para hidrolizar fragmentos activos de la pared celular (Bowles, 1998).
La aplicación de OGA da como resultado una rápida despolarización de la
membrana plasmática, flujo de iones de calcio dentro del citoplasma (Thain et
al., 1990; Mathieu et al., 1991; Messiaen y van Cutsem, 1994), la generación de
especies reactivas de oxigeno (Orozco-Cárdenas y Ryan, 1999) y la inducción
de AJ (Doares et al., 1995a) y etileno (O’Donnell et al., 1996).

16
2.4. Sistemina
La sistemina es un polipéptido de señalización que fue descubierto
durante investigaciones enfocadas a detectar la señal sistémica que regula la
expresión de genes de defensa en respuesta al ataque de insectos y daño
mecánico en hojas de jitomate (Lycopersicon esculentum). Este polipéptido
consta de 18 aminoácidos y no está modificado por cadenas laterales de
oligosacáridos. Se encontró que induce la actividad de inhibidores de proteasas
al ser aplicado en concentraciones muy bajas, del rango picomolar a fentomolar,
a plantas jóvenes de jitomate (Figura 2). El polipéptido fue purificado por medio
de una laboriosa secuencia de separación, la última de las cuales se hizo por
cromatografía líquida de alta resolución en fase reversa (RP-HPLC, por sus
siglas en inglés). La actividad del polipéptido fue ensayada aplicando fracciones
eluídas directamente de las columnas a plantas jóvenes a través de soluciones
acuosas absorbidas directamente por la corriente de transpiración de las
plántulas cuyos tallos habían sido cortados previamente por debajo del
hipócotilo. Una vez purificado, se detectó que el polipéptido sintético, de
secuencia idéntica al aislado y marcado radiactivamente con 14C, era
transportado de las hojas heridas a tejidos distales (Pearce et al., 1991).
Además, su movimiento a través del floema correlacionó con la velocidad de la
señal sistémica que se produce en respuesta a daño, que se estimó como de
aproximadamente de 3 cm/h (Nelson et al., 1983; Pearce et al., 1991; Narváez-
Vasquez et al., 1995). Debido precisamente a la movilidad sistémica de este
polipéptido a través del floema se le llamó “sistemina”. La sistemina no solo
induce la expresión de inhibidores de proteasa sino que también incrementa la
actividad de PFO, que también puede ser inducida por MeJA, además de un
amplio espectro de proteínas relacionadas a defensa, como lo son las SWRP.
Este polipéptido es muy polar y posee numerosos aminoácidos cargados
en su estructura primaria; posee, además, un enlace dibásico entre los
aminoácidos 9 y 10 que podría ser reconocido por proteasas tipo subtilisina para
su degradación y/o procesamiento (Schaller et al., 1999) y dos pares de prolinas
en las posiciónes 6 y 7, y 12 y 13. Por medio de estudios de dicroismo circular se
detectó una leve estructura secundaria en la región central del polipéptido
(Toumadje y Jonhson, 1995). Se estima que la conformación enrollada (“coil-

17
coiled”) es debido a estos dos pares de residuos de prolina presentes en su
secuencia. Se sabe, también, que las poliprolinas son motivos estructurales
empleados para la unión con otras proteínas (Ferris et al., 2001); se especula
que esta característica podría ser importante para el reconocimiento de
sistemina por su receptor.
+H3N-AVQSKPPSKRDPPKMQTD-COO-
1 5 10 15 18
Figura 2. Estructura primaria de la sistemina de jitomate.
2.4.1. Actividad y translocación de sistemina
Mediante la sustitución con alanina y deleciones en la secuencia de
sistemina, Pearce y colaboradores (1993), demostraron que se requería la
secuencia completa de sistemina para permitir la inducción máxima de los genes
de inhibidores de proteasas. Deleciones en el extremo carboxilo terminal de la
secuencia de sistemina causaron una disminución o pérdida total de la actividad
del polipéptido, principalmente al eliminar el ácido aspártico presente en la última
posición desde el extremo carboxilo terminal. Se demostró trambién que son
necesarios por lo menos los cuatro últimos aminoácidos (Met-Gln-Thr-Asp) para
que se manifieste una mínima actividad de sistemina. Al ser deletados estos
cuatro aminoácidos en orden progresivo se inactivó por completo la actividad
inductora de inhibidores de proteasas. Asimismo, la deleción de la alanina del
extremo amino-terminal redujo hasta 300 veces su actividad, y deleciones
progresivas de esta región causaron incrementos en la pérdida de actividad. Sin
embargo, se observó que podían ser eliminados hasta 14 aminoácidos de la
región NH2-terminal sin que se perdiera por completo la actividad del polipéptido.
Para determinar si las modificaciones en los aminoácidos de la sistemina

18
afectaban su actividad inductora, se utilizó una sistemina sintética análoga que
contenía una sustitución con Ala en cada aminoácido del polipéptido. Se observó
que las sustituciones por Ala en los residuos 2-6, 8, 9 ,10, 14 y 15 producían
pequeños cambios en la actividad, indicando que estos aa eran relativamente
poco importantes en la actividad de la sistemina, mientras que las sustituciones
en los residuos 12 y 13 correspondientes a uno de los dos pares de prolinas
causó una reducción drástica de la actividad biológica, mientras que la
sustitución en la posición 17 (Thr) abolió completamente la actividad inductora de
la sistemina (Figura 3). De este resultado surgió la hipótesis de que la treonina
es fosforilada como parte del proceso de señalización, la cual no ha podido hasta
el momento ser validada o desechada.
Figura 3. Efecto de la sustitución de cada uno de los 18 aminoácidos de la
sistemina de jitomate por alanina, sobre la inducción de la síntesis de inhibidores
de proteasa tipo I en hojas de jitomate.

19
En experimentos en los que se utilizaron sólo los 14 aa del extremo amino
terminal de una sistemina análoga, se detectó la unión a la membrana celular de
Lycopersicon peruvianum, la cual compitió con la sistemina natural por el sitio de
unión, mientras que la secuencia (Met-Gln-Thr-Asp) del extremo carboxilo no
presentó este comportamiento (Meindl et al., 1998). De este comportamiento
surgió un modelo para explicar el mecanismo de unión ligando-receptor, en el
cual se sugiere que el extremo amino terminal es el que se une inicialmente al
receptor para permitir la generación de los cambios conformacionales que
permitirán la posterior unión del extremo carboxilo terminal y el inicio de las
cascada de señalización (Meindl et al., 1998).
Análisis autorradiográficos y el uso del microscopio de luz, revelaron que
al aplicar sistemina marcada con 14C o 3H en hojas heridas de plantas de
jitomate, ésta se difundía a través de toda la hoja después de 30 min, mientras
que a los 90 min la radioactividad era detectada en las hojas apicales de las
plantas. A su vez, se observó que la aplicación de sistemina marcada con 3H se
concentró en el xilema y el floema de las nervaduras de la hoja herida 15
minutos después de su aplicación. Después de 30 min, la radioactividad se
encontró principalmente en los tejidos vasculares del peciolo, y a los 90 min, la
marca fue encontrada en el floema del tallo principal de las plantas. Los análisis
autorradiográficos, bioquímicos e histológicos, sugerían fuertemente que la
sistemina marcada radioactivamente y aplicada a hojas de jitomate heridas, era
translocada inicialmente al apoplasto o espacio intercelular, posteriormente al
tejido vascular de las nervaduras menores y al simplasto de las células
acompañantes del complejo de tubos cribosos, y finalmente a los tejidos distales
a través del floema. Con estos resultados se obtuvieron fuertes evidencias a
favor del transporte de la sistemina desde el sitio de la herida hacia los tejidos
distales de la planta (Narváez-Vázquez et al., 1995).

20
2.4.2. Modelo de la ruta de señalización activada p or sistemina
El sistema modelo para el estudio de la ruta de señalización activado por
sistemina es el jitomate. En esta especie, el modelo propone la percepción de la
sistemina por un receptor tipo cinasa de repetidos ricos en leucina, de 160-kDa,
localizado en la membrana plasmática de las células de jitomate (Scheer y
Ryan, 2002). La percepción rápida y reversible da como resultado un incremento
en la concentración del Ca2+ libre en el citosol debido a la apertura de canales de
la membrana plasmática y de almacenes internos. El Ca2+ estimula la actividad
de una proteín cinasa (PK) aun no identificada, resultando en la inhibición de la
H+-ATPasa de la membrana plasmática y, por lo tanto, en la despolarización del
potencial de la membrana plasmática y en la alcalinización del medio. El Ca2+
liberado también puede activar una fosfolipasa A2 (PLA2) y mediar la fosforilación
de una MAPK, resultando en la hidrólisis de lípidos y la liberación del ácido
linolénico de la membrana, con la consecuente producción de ácido jasmónico y
la activación de genes defensivos (Schaller, 1999) (Figura 4).

21
MP
Ca2++++
RS-160
Ca2+
Ca2+ H+
MAPK
PLA2
PK
K+, Cl
ActivaciActivaciActivaciActivacióóóónnnnggggéééénicanicanicanica
vacuola
H-ATP
P
Ác. jasmónico
ReducciReducciReducciReduccióóóónnnn
β----OxidaciOxidaciOxidaciOxidacióóóón (3)n (3)n (3)n (3)
COOHCOOHCOOHCOOH
oooo
Ca+
SISTEMINASISTEMINA
ÁÁcc. . linollinol ééniconico
Figura 4. Modelo hipotético de la ruta de transducción de señales en células de
jitomate activada por sistemina. MP: membrana plasmática; PLA2: Fosfolipasa
A2: MAPK: cinasas activadas por mitógenos; RS: receptor de la sistemina de 160
kDa (Schaller, 1999).

22
2.4.3. Glicopéptidos ricos en hidroxiprolina (“sist eminas”) de tabaco
(TobHypSys)
Mediante el empleo de cromatografía en columna se facilitó el aislamiento
de dos polipéptidos de 18 aa de las hojas de tabaco llamados TobHypSys I y
TobHypSys II. El análisis de su estructura reveló que ambos tienen residuos de
prolinas hidroxiladas, los cuales se encuentran glicosilados por cadenas laterales
de oligosacáridos, ricas en pentosas, con un grado de polimerización de 6 a 9
unidades (Pearce et al., 2001). Al igual que la sistemina en jitomate, ambos
mostraron la capacidad de inducir la síntesis de inhibidores de tripsina en plantas
de tabaco y de activar a MAP cinasas asociadas con el daño en cultivos
celulares en suspensión de tabaco (Pearce et al., 2001). Se detectó un tercer
polipéptido en tabaco, pero a muy bajas concentraciones. Este resultó casi
idéntico a TobHypSys II, excepto por la presencia adicional de leucina en la
región C-terminal. La aplicación de sistemina de jitomate a suspensiones
celulares de tabaco no causó la típica alcalinización en el medio del cultivo
celular. Asimismo, cuando se aplicó sistemina exógena a través del tallo de
plantas jóvenes de tabaco, tampoco se activó la síntesis de inhibidores de
tripsina en las hojas de tabaco, confirmándose que la sistemina de jitomate no es
percibida por las células de tabaco. Esto se debe, quizás, a sus marcadas
diferencias estructurales (Figura 5). Sin embargo, experimentos posteriores
indicaron que la sistemina de jitomate resultó capaz de inducir la alcalinización
del medio extracelular de suspensiones celulares de tabaco preparadas a partir
de plantas de tabaco transformadas con el ADNc del receptor de sistemina de
jitomate (SR160) bajo el promotor constitutivo 35S. De manera que posiblemente
existen uno o varios componentes implicados en la señalización intracelular
capaces de interactuar con el receptor SR160 para facilitar la señalización
mediada por sistemina en las células de tabaco (Scheer et al., 2003). Otra
posibilidad es que el motivo de poliprolina conservado en todas las sisteminas, el
cual produce una estructura conocida como PP II, podría ser importante para el
reconocimiento por el receptor SR 160, en forma similar a lo que se propone que
ocurre durante la interacción entre las glicoproteínas ricas en hidroxiprolina y
otras proteínas de la pared celular de plantas (Ferris et al., 2001). Sin embargo,
TobHypSys I y TobHypSys II inducen la alcalinización del medio en

23
suspensiones celulares de jitomate y la síntesis de inhibidores de proteasas en
plantas de jitomate, indicando que los receptores de jitomate puedan reconocer a
las llamadas sisteminas de tabaco (Glicopéptidos ricos en hidroxiprolina),
activando la misma señalización activada por la sistemina de jitomate (LeSYS).
Debido a que el precursor de las sisteminas de tabaco, proTobHypSys, se
encuentra hidroxilado y glicosilado, puede asociarse con la pared celular, como
se demostró para el precursor proTomHypSys en jitomate (Narváez-Vázquez et
al., 2005), y posiblemente puede ser procesado en el sitio de daño para inducir
señales de amplificación en la síntesis de oxilipinas (Ryan y Pearce, 2003).
Recientemente se aislaron de hojas de jitomate tres polipéptidos glicosilados e
hidroxilados de 15, 18 y 20 aa (TomHypSys), similares en la estructura básica a
TobHypSys I y II, los cuales inducen fuertemente la alcalinización cuando se
adicionan a cultivos celulares de jitomate y tabaco. En tabaco, la respuesta a
daño activa sistemicamente la síntesis de inhibidores de tripsina los cuales son
homólogos a los inhibidores II de jitomate (Pearce et al., 1993b).
Figura 5 . Estructura primaria de la sistemina de jitomate y de los glicopéptidos
ricos en hidroxiprolina presentes en jitomate (TomSys y TomHypSys I, II y III) y
tabaco (TobHypSys I y II), respectivamente.
TomSys +H2N- A V Q S K P P S K R D P P K M Q T D- COO-
TomHypSys I +H2N- R T O Y K T O O O O T S S S O T H H -COO-
TomHypSys II +H2N- G R H D Y V A S O O O O K P Q D E Q R Q-COO-
TomHypSys III +H2N- G R H D S V L P O O S O K T D -COO-
TobHypSys I +H2N- R G A N L P O O S O A S S P O O S K E - COO-
TobHypSys II +H2N- N R K P L S O O S O K P A D G O R P- COO-

24
La utilización de dos TobHypSys sintéticas, carentes de carbohidratos,
indujo una débil alcalinización en cultivos celulares de tabaco en suspensión y
una débil inducción de inhibidores de tripsina en hojas de tabaco, indicando que
la modificación con cadenas laterales de carbohidratos favorece la actividad
biológica de los glicopéptidos ricos en hidroxiprolina. Al sustituir con alanina la
arginina (R) en la región amino terminal, ácido glutamico (E) en el extremo C-
terminal y en la hidroxiprolina en posición 15 de TobHypSys I, se eliminó por
completo la inducción de actividad de este polipéptido. Además, sustituciones en
las hidroxiprolinas en posición 10 y 14 redujeron drásticamente la actividad del
glicopéptido. A su vez, las sustituciones con alanina en arginina (R) y asparagina
(N) en la región amino terminal redujeron severamente la actividad de
TobHypSys II sintéticas (medida como la capacidad de causar la alcalinización
del medio extracelular al ser aplicados a cultivos celulares de N. tabacum), así
como también la sustitución en el extremo carboxilo terminal de la arginina en la
posición 17, hidroxiprolina en la posición 10 y prolina posición 18,
respectivamente (Pearce et al., 2001) (Figura 6).

25
Figura 6. Actividad de TobHypSys I y TobHypSys II sintéticas, carentes de
modificaciones con cadena de oligosacáridos y con sustituciones de alanina en
cada uno de sus aminoácidos.
2.5. Prosistemina
La prosistemina, considerada el precursor de la sistemina, es una
proteína hidrofílica de aproximadamente 23 kDa que contiene un alto porcentaje
de aa cargados y pocos aa hidrofóbicos (McGurl et al., 1992). Este polipéptido
de 200 aa, no posee una secuencia señal típica de las proteínas que son
exportadas al exterior de la célula o a diferentes organelos a través del retículo
endoplásmico, ya que no hay evidencia de que esta proteína esté glicosilada
(Deláno-Frier et al., 1999). Se ha determinado que el sitio de síntesis de la
prosistemina se localiza en las células del parénquima del haz vascular de hojas,
pecíolos y tallos; se ha especulado que la localización tan estricta de la síntesis
de prosistemina en el tejido vascular puede representar una ventaja para la

26
planta, al facilitar el procesamiento de la prosistemina en respuesta a heridas y
promover una rápida carga de la sistemina en el floema, donde puede ser
rápidamente transportada a través de la planta (Jacinto et al, 1997; Li et al.,
2002a; Ryan y Moura, 2002; Narváez-Vázquez y Ryan, 2004; Scilmiller y Howe,
2005).
2.5.1. Expresión y supresión de la actividad de la prosistemina
La función de la prosistemina fue demostrada al transformar plantas de
jitomate con el ADNc de la prosistemina en orientación antisentido, la cual causó
una reducción severa de la inducción de inhibidores de proteasas y, por
consiguiente, una mayor susceptibilidad a herbivoría por larvas de Manduca
sexta (McGurl et al., 1992).
A su vez, la expresión del gen de la prosistemina en orientación sentido,
condujo a una sobre-expresión constitutiva de los genes de inhibidores de
proteasas, ya que aun sin ser heridas, las plantas transgénicas presentaron
niveles de alrededor de 87 µg/ml del inhibidor I y 75 µg/ml del inhibidor II,
mientras que las plantas control no mostraron acumulación alguna de los
inhibidores. Además, los niveles de los inhibidores I y II se incrementaron hasta
2.5 veces más en las plantas transgénicas en respuesta a daño mecánico
(Figura 7). La sobre-expresión de la prosistemina en plantas de jitomate no sólo
dió como resultado la acumulación constitutiva de inhibidores de proteasas, sino
también la acumulación de otras proteínas involucradas en mecanismos
defensivos, entre las cuales se identificaron una polifenol oxidasa, inhibidores de
cistein proteasas y de catepsina D, exopeptidasas (serin-carboxipeptidasa y
leucina aminopeptidasa), una aspártico proteasa, e isoformas específicas de
lipoxigenasa, calmodulina y cistatina (Bergey et al., 1996). Como consecuencia
de esta sobre-expresión múltiple de genes relacionados con defensa, estas
plantas resultaron altamente resistentes al daño causado por Manduca sexta y
otros insectos (Li et al., 2002b).

27
Figura 7. Niveles de inhibidores de proteasas I y II en plantas de jitomate sin
transformar (1) y transformadas (2) con el transgen de la prosistemina en
orientación “sentido” (según McGurl et al., 1994).
Dombrowski et al., (1999), estudiaron la inducción de inhibidores de
proteasas en plantas de jitomate tratadas con varias prosisteminas
recombinantes truncadas a lo largo de su secuencia aminoácidica. Éstas tenían
deleciones en los primeros 15 (PS 185) y 87 (PS 113) aa y otra sin la región
codificante para sistemina l78 (PS ∆SYS). Utilizaron como controles a la
sistemina (LeSYS) y prosistemina (LePS) con su secuencia aminoácidica
completa, y a la sistemina (A17) y prosistemina (PS A195) mutadas en la
treonina 17 y prolina 195, respectivamente, que como se ha mencionado es
esencial para su actividad biológica (ver Figuras 8 A y B). Al medir los niveles de
actividad del inhibidor I en hojas de plantas de jitomate tratadas con las
diferentes sisteminas y prosisteminas, observaron que la inducción de la
actividad del Inh I fue similar en las plantas tratadas con sistemina y
prosistemina. Esta actividad se mantuvo cuando se usó la PS 185 y disminuyó
ligeramente en las plantas que fueron tratadas con PS 113. Sin embargo, la
inducción de actividad inhibitoria se abatió por completo al usar PS ∆SYS, SYS
A17 y PS A195. Estos resultados coincidieron con los reportados previamente
por Pearce et al., (1993) y sugerían fuertemente que la actividad biológica de la
020406080
100120140160180200
1 2 1 2
Con
c. d
el in
hibi
dor (
ug/m
l)
Inh I Inh II

28
prosistemina radica básicamente en la región carboxilo terminal, donde está
ubicada la sistemina. Sin embargo, no descartaban la posibilidad de que la
región amino terminal podría estar involucrada en otros procesos de señalización
o bioquímicos, así como en el reconocimiento y anclaje de la prosistemina al
receptor SR 160, localizado en la membrana plasmática de las células de
jitomate.
2.5.2. Homología de la prosistemina de jitomate en plantas
solanáceas
La acumulación de inhibidores de proteasas en respuesta a daño se ha
observado en muchas especies de plantas incluyendo la papa, tabaco, alfalfa,
maíz, álamo y sauce, pero en ninguna de éstas se conocía la naturaleza de la
señal sistémica. Usando la técnica de reacción en cadena de la polimerasa
en combinación con transcripción reversa (RT-PCR) se aislaron y caracterizaron
secuencias de ADNc que codifican a homólogos de prosistemina en papa
(Solanum tuberosum), papa silvestre (S. nigrum) y chile (Capsicum annuum). El
análisis de la secuencia deducida de las prosisteminas de estas plantas mostró
un alto grado de similitud (de 85 a 94%) e identidad (de 73 a 88%) entre estas
especies, siendo la prosistemina de chile la que presentó la más baja identidad y
similitud en comparación con las otras especies. Las secuencias predichas de la
prosistemina de papa, papa silvestre y chile no mostraron diferencias
significativas con la secuencia de la prosistemina de jitomate. Por tal motivo, se
investigó si las prosisteminas sintéticas homólogas poseían actividad inductora
de inhibidores de proteasas en plantas de jitomate. Los resultados obtenidos
indicaron que la prosistemina de papa silvestre fue 10 veces menos activa que la
de jitomate, mientras que las prosisteminas de papa y chile fueron similares a la
de jitomate. Al utilizar estas prosisteminas en su respectiva especie fue posible
detectar la producción de ARNm de inhibidores I y II en papa y papa silvestre,
mientras que en chile sólo fue detectado el ARNm del inhibidor II (Constabel et
al., 1998).
El tabaco carece de un gen homólogo a la prosistemina de jitomate
(Pearce et al., 2001). Sin embargo, produce una vigorosa respuesta localizada al

29
daño y tiene, además, un sistema bien definido de señalización entre las hojas y
la raíz mediado por ácido jasmónico, el cual es responsable de la acumulación
de nicotina en raíz (Zhang y Baldwin, 1997).
Figura 8. A y B) Acumulación de inhibidores de proteasas en hojas de jitomate
tratadas con distintas concentraciones de prosisteminas recombinantes.
(Dombrowski et al., 1999).
0
20
40
60
80
100
120
140
160
180
200
2.5
pM
250
fM25
fM2.
5 pM
250
fM25
fM25
pM
2.5
pM25
pM
2.5
pM
250
fM H2O
PS 185 PS 113 PS∆SYS PS A195
Inhi
bido
r I
(ug/
ml e
xtra
cto)
A
0
20
40
60
80
100
120
140
160
180
2.5
0.25
0.025 2.
50.
250.0
25 2.5
0.25
0.025 2.
50.
250.0
25 250
Buffer
PS SYS PSA17 SYS A17 A17
Inhi
bido
r I
(ug/
ml e
xtra
cto)
B

30
Como se indicó anteriormente, el análisis de varias genotecas de ADNc
de hojas de tabaco reveló la presencia de dos prosisteminas, denominadas pro-
TobSys-A (165 aa) y pro-TobSys-B (164 aa), a partir de las cuales son liberadas
dos sisteminas mayoritarias (TobSys I y TobSys II) y una minoritaria (TobSys Ia)
(Pearce et al., 2001). En tabaco, el precursor ppTobHS-A se indujo en respuesta
a múltiples estímulos, incluyendo daño mecánico, AJ, ácido abscísico, herbivoría
por M. sexta e infestación con B. tabaci. Su expresión generalmente precedió a
la acumulación de transcritos de inhibidores de proteasa del tipo I y II y otros
genes de indole defensiva en tabaco, como una dioxigenasa inducida por
patógenos, por lo que se sugiere que al igual que LeSYS, TobSys juega un papel
importante en la regulación de respuestas defensivas en N. tabacum, tanto a
nivel local como sistémico (Rocha-Granados et al., 2005).
2.6 Metabolitos secundarios
Los metabolitos secundarios proveen defensas contra la infección,
depredación o agobio ambiental. La producción de metabolitos secundarios
ocurre frecuentemente a concentraciones relativamente bajas en la mayoría de
las familias de plantas. Por eso, parece razonable asumir que éstos pudieran
estar implicados en relaciones de evolución en aquellas especies que los
acumulan. Una característica fundamental del metabolismo secundario, es que
los productos no están uniformemente distribuidos a través de la planta, sino que
frecuentemente están limitados a órganos particulares, células y tejidos
particulares dentro del órgano (Rhodes, 1994).
El órgano en el cual los productos secundarios se acumulan es con
frecuencia diferente de aquel en donde ocurre la biosíntesis. Por ejemplo, la
capacidad de biosíntesis de nicotina está limitada a las raíces de la planta de
tabaco, aunque la nicotina se acumula tanto en hojas como en raíces (Baldwin et
al., 1997).
Las solanáceas contienen diversos metabolitos secundarios, siéndo los
grupos más importantes los alcaloides, terpenos y componentes fenólicos. La
síntesis de estos metabolitos está bajo control genético y depende críticamente

31
de las condiciones ambientales (Saitoh et al., 1985; Snook et al., 1986; Sisson y
Severson, 1990). Como se mencionó líneas arriba, el AJ juega un papel
fundamental en la regulación de la síntesis de novo de numerosos metabolitos
secundarios en cultivos celulares y en plantas de varias especies incluyendo los
alcaloides, terpenos, y compuestos fenólicos (Rickauer et al., 1997; Memelink et
al., 2001).
En N. tabacum y otras especies de Nicotiana, se ha comprobado el papel
fundamental de ciertos metabolitos secundarios en la defensa contra patógenos
e insectos plaga. Un ejemplo muy evidente de esta función se obtuvo de plantas
transgénicas de tabaco capaces de expresar constitutivamente la fitoalexina
resveratol, las cuales exhibieron una marcada resistencia contra el hongo
Botrytis cinerea (Hain et al., 1993).
2.6.1. Alcaloides (nicotina, nornicotina)
Más de 12,000 alcaloides han sido aislados desde el descubrimiento de la
morfina. Alrededor del 20% de las especies de las plantas que florecen producen
alcaloides. Varios alcaloides son tóxicos para insectos o parásitos, aunque
algunas especies de insectos se han adaptado a los alcaloides pirrolizidinos que
se acumulan en las plantas. Para ello han desarrollado mecanismos para
utilizarlos para su propio beneficio, modificandolos como monoesteres por medio
de procesos enzimaticos o acumulándolos en tejidos especializados, llegando
inclusive a utilizarlos como precursores de feromonas para atraer a la hembra,
como es el caso del lepidóptero Creatonotos transiens, o como sustancias de
defensa contra sus depredadores, tal como se ha observado en las larvas de
Tyria jacobaea (Buchanan et al., 2000; Wittstock y Gershenzon, 2002). El ataque
por herbívoros en tabaco silvestre estimula la biosíntesis de nicotina. Se sabe
que diferentes genotipos de tabaco (N. tabacum) varían en su habilidad de
acumular y almacenar nicotina. Además, en las especies nativas N. sylvestris, N.
attenuata, N. quadrivalvis y N. clevelandii se ha encontrado que el ataque por
insectos herbívoros incrementa dramáticamente la síntesis y acumulación de
nicotina. (Baldwin et al., 1998, 1999; Lou y Baldwin, 2003; Qu et al., 2004).
Cuando N. attenuata es atacada por diferentes herbívoros en su habitat natural,
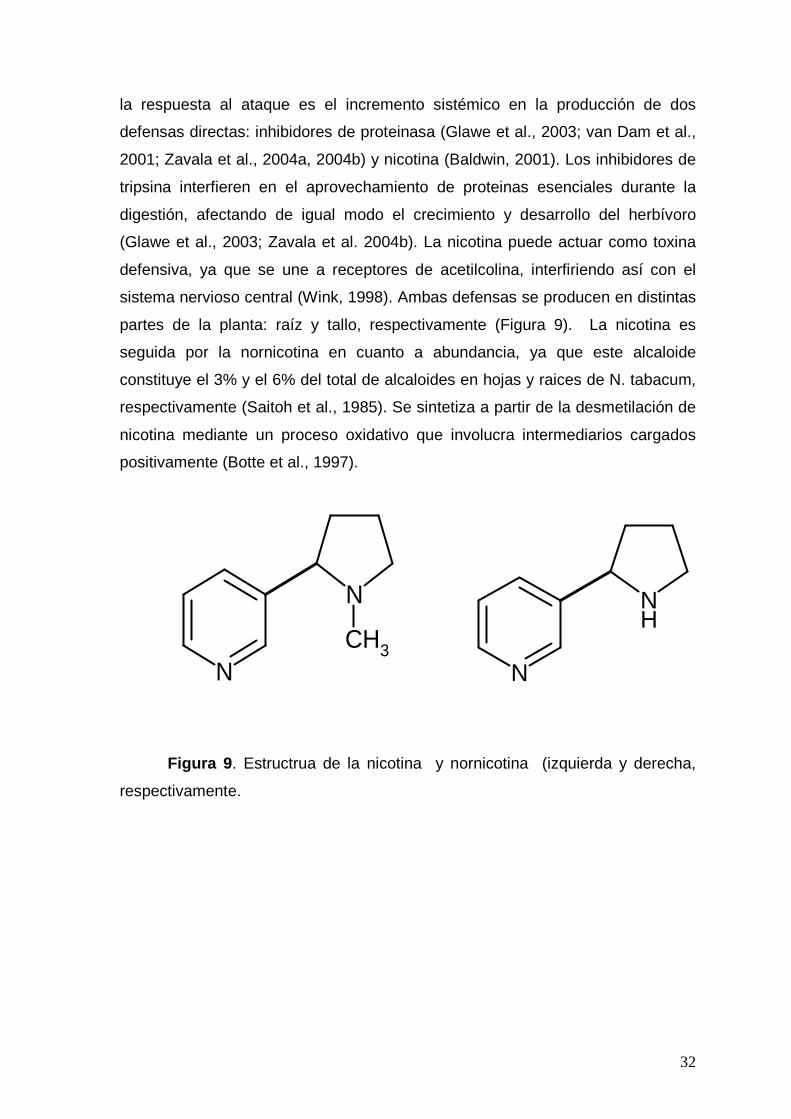
32
la respuesta al ataque es el incremento sistémico en la producción de dos
defensas directas: inhibidores de proteinasa (Glawe et al., 2003; van Dam et al.,
2001; Zavala et al., 2004a, 2004b) y nicotina (Baldwin, 2001). Los inhibidores de
tripsina interfieren en el aprovechamiento de proteinas esenciales durante la
digestión, afectando de igual modo el crecimiento y desarrollo del herbívoro
(Glawe et al., 2003; Zavala et al. 2004b). La nicotina puede actuar como toxina
defensiva, ya que se une a receptores de acetilcolina, interfiriendo así con el
sistema nervioso central (Wink, 1998). Ambas defensas se producen en distintas
partes de la planta: raíz y tallo, respectivamente (Figura 9). La nicotina es
seguida por la nornicotina en cuanto a abundancia, ya que este alcaloide
constituye el 3% y el 6% del total de alcaloides en hojas y raices de N. tabacum,
respectivamente (Saitoh et al., 1985). Se sintetiza a partir de la desmetilación de
nicotina mediante un proceso oxidativo que involucra intermediarios cargados
positivamente (Botte et al., 1997).
N
NCH3
NH
N
Figura 9 . Estructrua de la nicotina y nornicotina (izquierda y derecha,
respectivamente.

33
2.6.2. Compuestos fenólicos
Los compuestos fenólicos derivados de la ruta fenilpropanoide,
sintetizados a partir de fenilalanina y tirosina, que a su vez se derivan de la ruta
del ácido shiquímico, son metabolitos áromaticos que tienen uno o más grupos
hidroxilo unidos a un anillo áromatico. El aumento en el contenido de fenoles
inhibe el crecimiento y regeneración de plántulas; además, éstos actuan como
antioxidantes y como sustancias estructurales en los tejidos suberosos,
protegiendo de la desecación y ataque de patógenos.
La formación de suberina en tubérculos de papa dañada, está dada por
un aumento de componentes fenólicos como ACG (Buchanan et al., 2000). Este
compuesto fenólico se encuentra parcialmente en los tricomas glandulares, los
cuales presentan actividad de PFO. Asimismo, el ACG aumenta después de un
tratamiento con MeJA en N. attenuata y N. sylvestris (Keinänen et al., 2001).
La cafeoilputrescina, que es otro metabolito fenólico que se acumula en
hojas apicales, ovarios y flores de plantas de tabaco. Está implicado en
respuestas de resistencia a diversos tipos de estrés. Por ejemplo, los niveles de
este metabolito aumentan después de exponer hojas de tabaco a ozono
(Langebartels et al., 1991); también se ha observado su aumento en cultivos
celulares de papa tratados con evocadores e infecciones fungosas (Keller et al.,
1996).
Sin embargo, otros fenoles, como el AS y su éster acetílico, inhiben la
expresión de los genes IP en L. esculentum debido al antagonismo con la ruta de
transducción de señales dependiente de ácido jasmónico y sistemina (ƠDonnell
et al., 1996).

34
H
OH
OH
NNH2
OO
CO2H
OH
OH
OO
OH OHOH
COOH
HO
Ácid o clorogénico
Figura 10. Estructuras de compuestos fenólicos involucrados en defensa.
2.6.3 Terpenos
Los isoprenoides o terpenos es el grupo de metabolitos secundarios en
plantas con mayor número de estructruras reportadas hasta la fecha, que es de
aproximadamente 24,000 (Gershenzon y Kreis, 1999). Los isoprenoides
clasificados como metabolitos secundarios incluyen monoterpenos,
sesquiterpenos y diterpenos. Son considerados como secundarios porque
aparentemente no son necesarios para las funciones vitales de la célula; sin
embargo, participan en importantes interacciones entre plantas y su medio
ambiente. Ejemplos de esto son la interacción planta-planta (Rocha-Granados et
al., 2005), planta-insecto (Dicke, 1994; Paré y Tumlinson, 1997) y planta-
patógeno (Croft et al., 1993).
Cafeoilputrescina Ácido salicílico

35
Los terpenos poseen un número importante de funciones en las plantas
(Kleining, 1989; Rodríguez-Concepción y Gruissem, 1999). Con respecto a
funciones defensivas en tabaco, varios reportes indican que los diterpenos
glicosilados 16-hidroxigeranilinalool A y B en tabaco implican antibiosis contra
Heliothis virescens F. ya que sólo se encuentran presentes en variedades
resistentes de N. tabacum (Snook et al., 1986).
Un sesquiterpeno de gran importancia en chile (Capsicum annuum),
tabaco y otras plantas solanáceas es el capsidiol. Su acumulación en estas
plantas, la cual es inducida por factores bióticos y abióticos, limita la dispersión
de patógenos en los tejidos infectados. Sin embargo, es un metabolito inestable
ya que sus niveles en plantas decaen rápidamente después de su síntesis
(Lozoya-Gloria, 2004). Numerosos reportes indican que la acumulación de
capsidiol en las hojas de plantas de N. tabacum se debe casi exclusivamente en
respuesta a patógenos (sin embargo consultar Bohlmann et al., 2002).
A su vez, algunos diterpenos (duvanos y labdanos) son componentes de
la capa de cutícula en hojas de tabaco cuya presencia se ha asociado con
resistencia a insectos masticadores (H. virescens) y chupadores (el áfido M.
persicae) (Johnson y Severson, 1982). Recientemente, se descubrió también
que un diterpeno tipo labdano llamado WAF-1, actúa como una señal endógena
durante la activación de respuestas defensivas contra daño mecánico y la
infección por el virus del mosaico del tabaco (Ohashi et al., 2003).
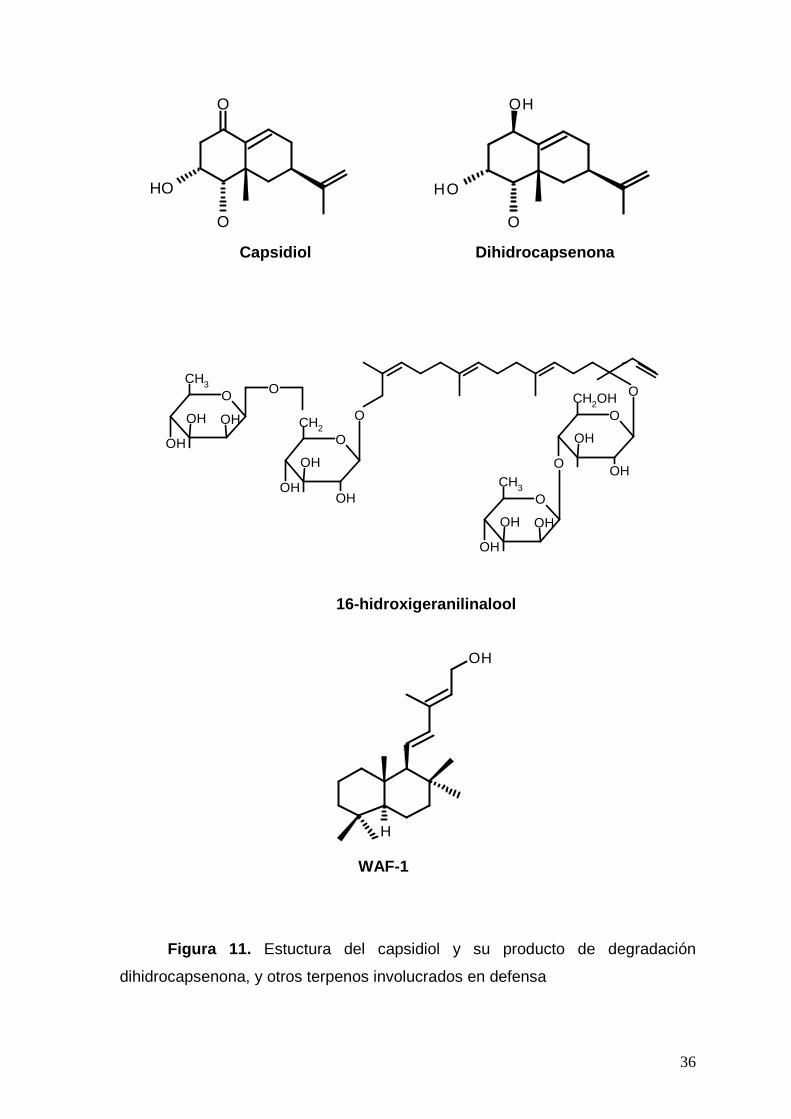
36
O
O
OH
O
OH
OH
Capsidiol Dihidroca psenona
OCH3
OH
OH OH
O
OCH2
OH
OH
O
OH
OCH2OH
O
OH
O
OH
OCH3
OH
OH OH
16-hidroxiger anilinalool
OH
H
WAF-1
Figura 11. Estuctura del capsidiol y su producto de degradación
dihidrocapsenona, y otros terpenos involucrados en defensa

37
2.7. Modelo de estudio
2.7.1. Plantas de tabaco transformadas con prosiste mina y sistemina
de jitomate ( Nt-LePS 08, Nt-LeSYS 03 y WT ×××× Nt-LeSYS 03)
El genero Nicotiana es dividido en tres subgéneros, 14 secciones y 66
especies; pertenece a la sub-tribu Nicotianae de la familia de las solanáceas
(Goodspeed, 1954). Se sabe que a diferencia de varias especies de la sub-tribu
Solaneae, el tabaco no posee homólogos de la prosistemina de L. esculentum
(LePS) (Pearce et al., 2001).
En un trabajo previo (Rocha-Granados, 2004) se generaron varias líneas
de plantas de tabaco sobre-expresantes de prosistemina (LePS) y sistemina
(LeSYS) de jitomate. De éstas, se selecciono una línea sobreexpresantes de
LeSYS: Nt-LeSYS 03 pues el análisis bioquímico mostro los mayores niveles de
actividad de inhibidores de proteasas, PFO y poligalacturonasas basales e
inducidos por daño (Fig. 12). Ademas se selecciono una sola sobreexpresante
de LePS: Nt-LePS, 08, pues presento una acumulación mayor de IT y PG
basales. A su vez, la línea Nt-LeSYS 03 mostró un fenotipo alterado,
caracterizado por anomalías tanto morfológicas como fisiológicas (ver más
adelante), las cuales desaparecieron en la cruza WT × Nt-LeSYS 03. Esta última
cruza también se incluyó en este estudio para ser comparada con la línea Nt-
LeSYS 03.

38
Figura 12. Niveles de inhibidores de tripsina (IT), PFO y
poligalacturonasas (PG) inducibles por daño en hoja local (HL) y hoja sistemica
(HS) en hojas de tabaco silvestres (Wt), y en plantas transformadas con LePS y
LeSYS (Rocha-Granados, resultados no publicados).
IT
0
300
600
900
1200
1500
1800
2100
0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24
Wt S1 S3 CS3 Ps1 Ps8 Ps10
AC
TIV
IDA
D E
SP
EC
IFIC
A.
HL
HS
PFO
0
2
4
6
8
10
12
14
16
0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24
Wt S1 S3 CS3 Ps1 Ps8 Ps10
D.O
./min
/mg
pro
teín
a
HL
HS
PG
0
20
40
60
80
100
120
140
160
180
200
0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24 0 16 24
Wt S1 S3 CS3 Ps1 Ps8 Ps10
nm
ol/m
in/m
g p
rote
ína
HL
HS

39
2.7.2. Alteraciones morfológicas y fisiológicas en la línea Nt -LeSYS
03.
La línea Nt-LeSYS 03 mostró una acumulación altamente significativa de
actividad inhibitoria contra tripsina (IT), quimotripsina (IQT) y PFO. Además, esta
línea se caracterizó por desarrollar un fenotipo con numerosas alteraciones
morfológicas y fisiológicas, las cuales de describen a continuación: hojas más
pequeñas y lanceoladas (Figura 13, B y D), con numerosas regiones necróticas
sobre la superficie de sus hojas, semejantes a las generadas durante la reacción
hipersensible en una interacción planta-patógeno incompatible; senescencia más
acelerada de sus hojas; desarrollo y crecimiento más rápido de los brotes
laterales, aunque de mucho menor tamaño que la planta sin transformar (Figura
13 A y 14 B); ausencia de un pecíolo bien definido, el cual más bien semeja una
prolongación de la hoja (Figura 15 B) y alteraciones morfológicas en flores, que
fueron más pequeñas y con estambres de talla menor al pistilo, lo cual contrastó
con las flores de plantas de tabaco WT, que normalmente desarrollan estambres
largos que sobresalen del pistilo para permitir la caída del polen maduro sobre
éste y así asegurar la autofecundación (Figura 14 A, Figura 15 D y G a la
derecha, a la izquierda en 15 F y 15 G). Además, los sacos polínicos de las
plantas Nt-LeSYS 03 produjeron muy poco polen. Es muy probable que estas
dos anomalías se combinaron para impedir su autofecundación natural. La
fecundación fue altamente ineficiente, ya que la mayoría de las cápsulas
producidas estaban vacías y solamente en algunas se encontró semilla.
Además, las pocas semillas producidas por las plantas Nt-LeSYS 03 germinaron
mucho más lentamente que las semillas de las plantas no transformadas. De
modo que no se observó emergencia después de un mes de haber sembrado las
semillas de las plantas transformadas, mientras que la germinación de las
semillas de plantas control no tardó más de 3 semanas (Figura 15 A) (Rocha-
Granados, 2004). Por estas características se optó por cultivar esta línea in vitro
para agilizar la generación de las plantas necesarias para los experimentos.

40
CRECIMIENTO
Figura 13 . I. Fenotipo mostrado por plantas de tabaco transformadas con
sistemina de jitomate (Nt-LeSYS). En los paneles A, B, C se pueden observar las
diferencias entre plantas Nt-LeSYS 03 y WT. En el panel (A), se muestran
plantas de la misma estatura pero de diferente edad. Mientras que en los
siguientes páneles se observa clararmente la diferencia en tamaño entre plantas
de la misma edad (B) y en la morfología de las hojas (C y D).
B
C
D
WT Nt-LeSYS 03
WT Nt-LeSYS 03
Nt-LeSYS 03
WT Nt-LeSYS 03
A

41
Figura 14 . II. Fenotipo mostrado por plantas de tabaco transformadas con
sistemina de jitomate (Nt-LeSYS). Observese en el panel (A) hacia la izquierda
que el pístilo sobresale de los estambres. En el panel B se muestra la
generación espontánea en la superficie de las hojas de regiones necróticas
localizadas semejantes a las que se forman durante la reacción hipersensible.

42
GERMINACIÓN FLORACIÓN ANORMAL
DESARROLLO DEL PECIOLO
A
B
D
E
F
G
B C
Figura 15. III. Fenotipo mostrado por plantas de tabaco transformadas con
sistemina de jitomate (Nt-LeSYS). En el panel (A), la semilla de las plantas Nt-
LeSYS 03 presentó un claro retardo en la tasa de germinación, y el desarrollo
subsiguiente de las plántulas fue muy lento con respecto al control, mientras que
en los paneles (B) y (C) las flechas señalan las diferencias en el desarrollo de los
pecíolos de las hojas entre plantas control (B) y las plantas Nt-LeSYS 03 (C). En
los paneles D, E y F, se muestran diferencias morfológicas observadas en las
flores de plantas Nt-LeSYS 03 con respecto al testigo. La fotografía del panel (G)
muestra que las flores de las plantas Nt-LeSYS 03 (derecha) son más pequeñas
que las flores de la planta WT (izquierda). Además, los estambres de las plantas
Nt-LeSYS 03 fueron mas cortos (E, a la Izquierda, F, a la derecha), que los
desarrollados en las plantas control (D, a la derecha, F, a la izquierda) y no
produjeron suficiente polen (tomado de Rocha-Granados, 2004).

43
2.7.3. Resistencia a herbivoría por larvas de Manduca sexta en
plantas Nt-LePS y Nt-LeSYS
Como se mencionó líneas arriba, estudios previos demostraron que las
plantas de jitomate capaces de expresar el gen de la prosistemina en orientación
antisentido fueron más susceptibles al daño causado por larvas de Manduca
sexta que las plantas control (Orozco-Cárdenas et al., 1993), mientras que
aquellas capaces de expresarlo en orientación sentido presentaron una mayor
resistencia a insectos que se alimentan del contenido celular (Li et al., 2002b).
Tomando en consideración estos antecedentes, era posible esperar que las
plantas Nt-LePS y Nt-LeSYS presentaran igualmente una mayor resistencia a la
depredación por larvas de Manduca sexta, debido a los altos niveles
constitutivos y/o inducibles de IT, IQT y PFO, que son proteínas involucradas en
las defensas de las plantas contra el ataque de insectos, detectados en algunas
de las líneas (Figura 12). De modo que las plantas Nt-LePS 01, 08, 10 y 32 y Nt-
LeSYS 01, fueron expuestas al ataque continuo de larvas de M. sexta durante 6
días. Los resultados mostrados en las Figuras 16 (A y B) y 17 indican que las
larvas que se alimentaron de las plantas Nt-LePS y Nt-LeSYS presentaron una
reducción significativa en peso y tamaño en relación a las larvas que se
alimentaron de las plantas control. En la Figura 18 se aprecia también que la
magnitud del daño causado por las larvas en las plantas Nt-LePS y Nt-LeSYS
fue claramente menor al que se produjo en las plantas control, las cuales fueron
prácticamente defoliadas. Sin embargo, la clara resistencia a la herbivoría por M.
sexta mostrada por todas las líneas de plantas Nt-LePS y Nt-LeSYS 01 no
coincidió con los resultados obtenidos con la línea LeSYS 03, la cual resultó
altamente susceptible a la herbivoría por larvas de M. sexta (Rocha-Granados,
resultados no publicados) a pesar de que los ensayos bioquímicos mostraron
una acumulación robusta de proteínas relacionadas con defensa en respuesta a
daño mecánico (Figura 12). Esto sugiere que otros factores pudieran estar
involucrados en la resistencia a la herbivoría observada en las plantas
transgénicas; una posibilidad podría ser la acumulación de nicotina y/u otros
metabolitos secundarios asociados con resistencia a insectos, como el ACG y
nornicotina.

44
Figura 16. Larvas de Manduca sexta antes (A) y 6 días después (B) de haber
sido alimentadas con plantas de tabaco sin transformar (B, a la izquierda) o con
plantas Nt-LePS y Nt-LeSYS (B, a la derecha) (Tomado de Rocha-Granados,
2004).
Figura 17. Análisis del crecimiento de larvas de M. sexta alimentadas con
plantas de tabaco sin transformar y plantas Nt-LePS y Nt-LeSYS. La gráfica
muestra el peso y longitud de las larvas alimentadas en plantas transgénicas y
control, en donde el promedio, error estándar, análisis de varianza y prueba de
significancia se determinaron con una confiabilidad del 95%, usando el paquete
estadístico SPSS 10.0 para Windows. Con un nivel de significancia P≤0.01
(Tomado de Rocha-Granados, 2004).
Control Transformadas
BA
Peso
Longitud
0
2
4
8
6
10
Control Transform adas
Pes
o (
g)
y lo
ng
itu
d (
cm)
a
b
c
d

45
Figura 18. Vista general de la magnitud del daño causado por larvas de M. sexta
en plantas de tabaco sin transformar (control) y transformadas (Nt-LePS y Nt-
LeSYS) (Tomado de Rocha-Granados, 2004).
Control
LePSLePS

46
3. HIPÓTESIS
La resistencia diferencial ante la folivoría por larvas de Manduca sexta y
las alteraciones fenotípicas observadas en plantas transgénicas de Nicotiana
tabacum capaces de sobreexpresar la sistemina o la prosistemina de
Lycopersicon esculentum puede ser el reflejo de modificaciones en la síntesis y
acumulación de metabolitos secundarios relacionados con defensa y/o
desarrollo.

47
4. OBJETIVOS
4.1. Objetivo general
Identificación y cuantificación de metabolitos secundarios en plantas de
tabaco (Nicotiana tabacum) transformadas con sistemina y prosistemina de L.
esculentum en respuesta a daño mecánico, herbivoría por Manduca sexta e
infestación con mosquita blanca (Bemisia tabaci).
4.2. Objetivos específicos
1. Identificación y cuantificación de los niveles basales de metabolitos
secundarios (nicotina, nornicotina, ácido clorogénico y capsidiol),
mediante GC/MS en hoja y raíz de plantas transformadas de tabaco (Nt-
LePS 08, Nt-SYS 03 y WT × Nt-LeSYS 03) en dos etapas distintas de
desarrollo.
2. Cuantificación de fenoles totales en hoja y raíz de plantas transformadas
de tabaco (Nt-LePS 08, Nt-SYS 03 y WT × Nt-LeSYS 03) en dos etapas
distintas de desarrollo.
3. Identificación y cuantificación de nicotina, nornicotina, ácido clorogénico y
capsidiol, en hoja y raíz de plantas transformadas de tabaco (Nt-LePS 08,
Nt-SYS 03 y WT x Nt-LeSYS 03) sometidas a daño mecánico, folivoría
por larvas de Manduca sexta e infestación por Bemisia tabaci.
4. Cuantificación de fenoles totales en hoja y raíz de plantas transformadas
de tabaco (Nt-LePS 08, Nt-LeSYS 03 y WT × Nt-LeSYS 03) sometidas a
daño mecánico, folivoría por larvas de Manduca sexta e infestación por
Bemisia tabaci.

48
5. MATERIALES Y MÉTODOS
5.1. Cultivo de plantas
Las semillas de plantas Nt-LePS 08 y WT × Nt-LeSYS 03, fueron
germinadas en charolas de germinación y después de 2 semanas se
transplantaron a macetas de 750 ml para plantas jóvenes y macetas de 2.35 l
para plantas adultas con una mezcla general de tierra formada por 3 partes de
Sunshine Mix 3 (SunGro Horticulture, Canada), 1 parte de vermiculita (SunGro
Horticulture, USA) y 1 parte de perlita (Termolita S.A., México). Previamente a la
realización de los experimentos, se realizó la prueba histoquímica de GUS para
la detección de las transformantes (homo y heterocigotas; ver más adelante); las
plántulas que resultaron positivas fueron seleccionadas. Una vez seleccionadas,
se creció una planta por maceta, fertilizandose cada semana con una solución
nutritiva 20-10-20 (N-P-K) (Peters Professional; Scotts-Sierra Horticultural
Products Co, Marysville, OH, EUA).
5.2. Cultivo in vitro de la línea Nt-LeSYS 03
Para el desarrollo de este trabajo se partió de las hojas más jóvenes de
tabaco de la línea Nt-LeSYS 03 las cuales fueron lavadas con jabón comercial y
desinfectadas, por espacio de 10 min en agua destilada estéril + Tween 20 al
0.1% e hipoclorito de sodio (cloro comercial) al 10% (v/v). Posteriormente, fueron
enjuagadas varias veces con agua destilada estéril. Las hojas se fragmentaron
en trozos de 0.5 cm × 0.5 cm; posteriormente se cultivaron in vitro en frascos de
vidrio de 300 ml, con tapa de rosca, que contenían 40 ml de medio MS sólido
(Murashige y Skoog, 1962), el cual contiene lo siguiente:

49
Composición del medio de cultivo Sacarosa o azúcar comercial 30 g Sales de Murashige y Skoog 4.32 g Mioinositol 1 g Vitaminas de Murashige y Skoog 1 ml Bencil amino purina 0.5 mg/l Agar 8 g Nota: ajustar el pH=5.7 ± 1, antes de agregar el agar, cristalizar el medio por 10 minutos en el horno de microondas y esterilizar a 121 ºC por 20 min.
La regeneración a partir de hoja tardó aproximadamente 2 meses y
medio, hasta la obtención de plántulas a una temperatura de 25 ºC, con un
fotoperiodo de 16 h luz y 8 h oscuridad, cambiando una sola vez el tejido a
cultivo nuevo. A partir de tallos de plántulas la regeneración tarda de 15 a 20
días. Después de este lapso de tiempo, las plántulas de esta línea se utilizaron
para posteriores experimentos.
5.3. Preparación de X-Gluc
Para realizar la prueba histoquímica de GUS, se utilizó el método descrito
por Jefferson et al., (1987). El ensayo se realizó cuando las plántulas
presentaban un par de hojas bien desarrolladas. Para ello se tomaron pequeños
segmentos de hoja de plantas transformadas de tabaco Nt-LePS 08, Nt-SYS 03,
así como de la cruza de WT × Nt-LeSYS 03 los cuales fueron sumergidos en la
solución X-Gluc y puestos en la oscuridad a 37 °C durante toda la noche. La
clorofila de los tejidos fue removida lavando varias veces con etanol al 70%.
Finalmente se lavaron con una mezcla de metanol: acetona, 3:1 (v/v), hasta que
pudieron verse claros. Los tejidos ya desteñidos se visualizaron en un
microscopio estereoscópico.
La solución “stock” para la preparación del X-Gluc consistió de los siguientes
reactivos:

50
Solución “stock” Concentración final Mezcla (ml)
NaH2PO4 al 1 M 100 Mm 10.0 EDTA 0.5 M pH 8 10 Mm 20.0 Ferrocianuro de potasio 0.005 M, pH 7
0.5 Mm 10.0
X-Gluc 0.02 M* 1 mM 1.0 Triton X-100 10% 0.1 % 0.1 Agua destilada _ 58.9 *en 50 mg/ml de dimetilformamida o DMSO
Una vez preparada la solución “stock”, se esterilizó por filtración (0.22 µm) y se
almacenó en la oscuridad a 4 °C.
5.4. Ensayos de daño mecánico
Para realizar los experimentos de daño mecánico se utilizaron tres plantas
de tabaco de cada una de las líneas en estudio, Nt-LePS 08, Nt-SYS 03 y WT ×
Nt-LeSYS 03. Como controles se usaron tres plantas WT por cada línea
transformada utilizada.
Plantas jovenes de cuatro semanas que presentaban 4 hojas bien
desarrolladas fueron heridas en su 3ª hoja (considerando como la primera hoja a
la más desarrollada, en la base de la planta) utilizando unas pinzas metálicas de
disección estériles y presionando cuatro veces a lo largo de la hoja y
perpendicularmente a la vena principal. Después del quinto día se colectaron las
hojas dañadas (respuesta local) y la hoja adyacente superior sin dañar
(respuesta sistémica). De igual forma, el ensayo se realizó en plantas adultas de
siete semanas, con 16-20 hojas bien desarrolladas. Para ello, la hoja media (hoja
8, 9 o 10), que representa la transición entre tejido fuente-consumidor en N.
tabacum (Masclaux et al., 2000) fue herida; esta hoja (respuesta local), las hojas
distales intactas, inferior (hacia la base) y superior (hacia el ápice), localizadas
filotácticamente en un ángulo de 0° con respecto a la hoja dañada y la raíz

51
fueron colectadas 5 días después de haber aplicado el daño (respuestas
sistémicas).
5.5. Bioensayos
5.5.1. Condiciones del experimento de herbivoría co n M. sexta
Para este experimento se utilizaron las mismas líneas de plantas
transgénicas de tabaco empleadas anteriormente para los ensayos de daño
mecánico. Las plantas se mantuvieron en una cámara de crecimiento (Conviron,
USA) a una temperatura constante de 28 °C y bajo un fotoperiodo de 16 h luz y
8 h oscuridad. Las larvas de Manduca sexta Linnaeus empleadas se
encontraban en el quinto instar de desarrollo. Estas larvas fueron crecidas en el
insectario del Cinvestav-Unidad Irapuato y alimentadas con una dieta artificial, a
base de germen de trigo, caseína, azúcar, levadura y una mezcla de sales,
suplementada con vitaminas (Yamamoto, 1964). Las larvas permanecieron sin
alimento durante un período de 12 h previo al inicio del experimento. El ensayo
de herbivoría se realizó cuando las plantas alcanzaron una longitud de 40 cm y
tenían entre 16 y 20 hojas bien desarrolladas; este bioensayo se inició colocando
una larva de M. sexta por planta, en la parte media de la planta (hoja de
transición entre tejido fuente-consumidor). Las larvas se alimentaron libremente
por un periodo de 24 horas. Transcurrido este tiempo, las larvas se retiraron de
las plantas; la colección del tejido (hoja dañada, hojas distales en la región apical
y basal y raíz) fue hasta cinco días después.

52
5.5.2. Infestación con mosquita blanca ( Bemisia tabaci)
Para realizar estos ensayos se utilizaron 70 mosquitas (Bemisia tabaci
Gennadius) libres de virus y recién emergidas para infestar 2 hojas por planta
(35 mosquitas/hoja). Las plantas que se utilizaron fueron plantas jóvenes de
cuatro hojas, en las cuales se colocaron 2 pequeñas jaulas (“clip cages”; de 4 cm
de diámetro y 3.5 cm de altura) fabricadas a partir de vasitos de unicel del
número cero, adheridas a las hojas 2 y 3, contando desde la base de la planta.
En éstas se introdujeron las mosquitas para obligarlas a ovipositar en un área
delimitada de la hoja por un período de 24 h. Una vez transcurrido este tiempo,
las mosquitas fueron retiradas y las plantas se mantuvieron por 28 días
adicionales en un cuarto de crecimiento (2.3 m [ancho] × 4.2 m [largo] × 3 m
[alto]), bajo condiciones controladas de luz, temperatura y fotoperíodo (16 h luz a
28°C/8 h oscuridad a 20° C), para permitir la terminación del ciclo de vida de la
mosquita blanca. Una vez cumplidos los 28 días se consideró a las dos hojas
infestadas originalmente como hojas locales, mientras que las hojas distales
localizadas filotácticamnte en un ángulo de 0° (considerado el tejido con la
máxima conección vascular con las hojas infestadas), en la parte basal y apical
de la planta, respectivamente, y la raíz, fueron colectadas para medir la
respuesta sistémica.
5.6. Preparación de extractos
La extracción de metabolitos secundarios, se llevó a cabo a partir de tejido
molido (con N2 líquido) y congelado (a − 80 °C) de hoja y raíz, de las líneas
seleccionadas Nt-LePS 08, Nt-SYS 03 y WT × Nt-LeSYS 03 sometidas a los
diferentes tratamientos anteriormente descritos. Para ello, se pesaron 200 mg de
tejido fresco en 1 ml de una solución de metanol acuoso (40% metanol v/v)
conteniendo 0.5% (v/v) de ácido acético y 0.1 mg/ml de timol, el cual se empleó
como estandar interno, para el análisis por GC/MS. Las mezclas se agitaron
vigorosamente por 2 horas a 4 °C y se centrifugaron por 15 minutos a 12,000

53
rpm y a la misma temperatura. Los sobrenadantes obtenidos se filtraron y fueron
utilizados inmediatamente para el análisis de los metabolitos de interés.
5.7. Análisis de fenoles totales
La cuantificación de fenoles totales presentes en los extractos
metanólicos se realizó de acuerdo al método de Folin (Swain y Hillis, 1959)
adaptado para su uso en microplacas de 96 pozos. Para ello, se tomaron 10 µl
de muestra y se aforaron a 50 µl con agua desionizada. Se agregarón 50 µl del
reactivo de Folin (diluído en una proporción 1:2) y 100 µl de carbonato de sodio
al 7.5% (w/v). La mezcla se incubó a 45 ºC por 15 minutos y se dejó enfriar a
temperatura ambiente por 10 a 15 min. Las lecturas se hicieron en un lector de
placas (Ultramark; Microplate Imaging System; BioRad Laboratories, Hercules,
CA, EUA) por triplicado a una longitud de onda de 750 nm y se compararon con
una curva patrón preparada con ácido caféico (en un rango de 15 a 500 µg/ml).
5.8. Condiciones para cuantificar e identificar met abolitos secundarios por
GC/MS
5.8.1 Derivatización
Para el análisis de metabolitos secundarios por GC/MS, fue necesario
producir derivados silanizados de menor polaridad con la finalidad de
incrementar su volatilidad y su estabilidad térmica. Para ello, se siguió la
metodología descrita por Fiehn et al., (2000). De modo que los extractos
metanólicos acidificados (250 µl) se evaporaron a sequedad empleando un
sistema de concentración al vacío (Heto Maxi Dri Lyo, Termo Electron
Corporation, Allerød, Dinamarca). Posteriormente, los extractos secos fueron
derivatizados inicialmente en una solución de 50 µl de piridina en 20 µl
metoxiamina, la segunda reacción de derivatización se inició inmediatemente
después de este tiempo mediante la adición directa de 80 µl de metil-trimetil-

54
sililtrifluoracetamina (MSTFA). La nueva mezcla reactiva se incubó a 37 °C por
30 minutos. Las mezclas derivatizadas se dejaron reposar por dos horas a 25 ºC
antes de inyectar la muestra al sistema GC/MS. La metoximación y la
trimetilsilanización están diseñadas para permitir la derivatización y análisis de
hidroxi y amino ácidos, alcoholes, azúcares, poliaminas y ácidos aromáticos,
entre otros (Fiehn et al., 2000).
5.8.2 Cromatografía de gases acoplada a espectrome tría de masas
(GC/MS).
Los compuestos derivatizados fueron analizados por medio de
cromatografía de gases usando un cromatógrafo de gases (Hewlett-Packard
5890 Series II), equipado con una columna capilar HP-1 de 30 m de largo, 0.25
mm de diámetro interior y 0.25 µm de película fina a base de dimetilpolisiloxano y
acoplado a un detector selectivo de masas (Hewlett-Packard/MSD 5972 series).
Se empleó helio como gas acarreador, a un flujo constante de 1 ml/min. La
temperatura del inyector fue mantenida a 230 °C y a una presión de 8.75 psi.
Para realizar las separaciones, se usó el siguiente perfil de tiempo/temperatura:
la temperatura inicial del horno fue de 70 °C, seguido por un incremento gradual
de la temperatura hasta 310 °C, a una velocidad 1.0 °C/min. El tiempo total de
corrida fue de 54 minutos; cada una se inició después de la inyección de 1 µl de
las muestras derivatizadas. Los picos fueron identificados mediante la
comparación con los espectros de masas y tiempos de retención de estándares
comerciales de referencia.
5.9. Curvas de calibración
Se realizaron curvas de calibración para nicotina, nornicotina, ácido
clorogénico y capsidiol. Las curvas se hicieron usando estandares comerciales
(Sigma-Aldrich Chemical Co., St. Louis, MO, EUA) con excepción del capsidiol,
el cual se purificó, por medio de cromatografía fina, a partir de extractos de fruto

55
de chile ancho (Capsicum annuum) tratado con hemicelulasa de Aspergillus
niger (Brooks et al., 1985). Los estándares se derivarizaron igual que las
muestras, a excepción de la nicotina, la cual se diluyó con metanol e inyectó
directamente. Para la derivatización, se tomaron 10 µl de cada una de las
muestras empleadas para generar las curvas tipo, las cuales se evaporaron a
sequedad, para ser posteriormente derivatizadas en las mismas condiciones que
se emplearon para las muestras problema. Cada punto de la curva de calibración
se preparó por duplicado y se evaluó por GC/MS.
Los resultados obtenidos fueron analizados para obtener las curvas de
calibración de cada uno de los metabolitos, incluyendo sus respectivas
pendientes y coeficientes de correlación r (que fueron superiores a 0.95 en todos
los casos). Con estas curvas se obtuvieron las ecuaciones para cuantificar cada
uno de los metabolitos de interés (ver apéndice).
5.10. Análisis estadístico
Se analizaron tres plantas por línea seleccionada para el experimento de
daño mecánico y mosquita blanca, y de tres a cinco plantas por línea para el
experimento de herbivoría. Las lecturas se hicieron por duplicado en todos los
análisis por GC-MS. Para el análisis de fenoles totales, cada extracto se midió
por triplicado. Los datos fueron sometidos a un análisis de varianza de una vía
(ANOVA), utilizando para ello el programa JMP, versión 3.1.6.2., para Windows.
Los datos se reportaron como medias. Éstas se compararon entre sí para
detectar diferencias significativas empleando el método de Tukey, que es un
estimador que permite diferenciar valores calculados con un valor tabulado que
se determina en base a los grados de libertad y a una probabilidad de error.

56
6. RESULTADOS
6.1. Identificación de metabolitos secundarios de i nterés por GC/MS, en
plantas jóvenes y adultas
El análisis de los metabolitos de interés se realizó por GC/MS, que resultó
ser una técnica analítica más versátil para el análisis de un elevado número de
muestras que la cromatografía líquida de alta resolución (HPLC), cuyo uso se
exploró también en este trabajo (resultados no mostrados). La identificación se
hizo por comparación con el tiempo de retención y espectro de masas de
compuestos de referencia disponibles comercialmente (ej., nicotina, nornicotina,
y ácido clorogénico) o bien purificados a partir de tejidos vegetales (e.g.,
capsidiol). Esta estrategia se empleó debido a la relativa complejidad de los
cromatogramas (se muestra un ejemplo típico en la Fig. 19), en los cuales se
podían apreciar numerosos picos cuyos espectros de masa resultaron de difícil
interpretación debido a que muchos de ellos se encontraban aparentemente
modificados, especialmente por oligosacáridos. La comparación entre el tiempo
de retención y espectro de masas de los compuestos de interés presentes en las
soluciones de referencia y en los extractos metanólicos crudos de tabaco
analizados, se muestran a continuación para nicotina (Figs. 20 y 21), ácido
clorogénico (Figs. 22 y 23), capsidiol (Figs. 24 a 26) y nornicotina (Figs. 27 y 28).
En este último caso, sólo se pudo detectar un producto de degradación del
capsidiol en los extractos, lo que coincidió con la conocida inestabilidad de este
compuesto in vivo (Ward et al., 1977; Guedes et al., 1982; Whitehead et al.,
1987; Threlfall y Whitehead, 1988).
C

57
Figura 19. Cromatograma típico obtenido al separar extractos metanólicos
derivatizados de hoja de Nicotiana tabacum por GC/MS. La nicotina emergió
después de 15.83 min (*); timol (T), empleado como estándar interno) después
de 15.71 min (*); nornicotina después de 20.05 min (*); el posible producto de
degradación del capsidiol (dihidrocapsenona) después de 32.78 min (*), y el
ácido clorogénico después de 48.83 min (*).
5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00 50.00
1000000
2000000
3000000
4000000
5000000
6000000
7000000
8000000
9000000
1e+07
1.1e+07
1.2e+07
Time-->
Abundance
TIC: NT193b.D
* T
*
32.78 48.83 20.05 *
* 15.83

58
Figura 20. A) Cromatograma de GC/MS del estándar puro de nicotina, con un
tiempo de retención de 15.80 min (*), y B) su correspondiente espectro de
masas.
5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00 50.00
200000
400000
600000
800000
1000000
1200000
1400000
1600000
1800000
2000000
2200000
2400000
Time-->
Abundance
TIC: nic102a.D 15.80
30 40 50 60 70 80 90 100 110 120 130 140 150 160 1700
50000
100000
150000
200000
250000
300000
350000
400000
450000
m/z-->
Abundance
Scan 2173 (15.811 min): nic102a.D84
133
161
42119926551 77 105 14530 58 99
A
B
*

59
Figura 21. A) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando a nicotina con un tiempo de retencion de 15.83
min (*) , y B) su correspondiente espectro de masas.
14.50 15.00 15.50 16.00 16.50 17.00 17.50 18.00 18.50 19.00
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
Time-->
Abundance
TIC: NT193b.D
14.20
14.84
15.35
15.39
15.70
15.83 16.74
16.82 18.57
19.02
A
40 60 80 100 120 140 160 180 200 220 240 260 280 3000
50000
100000
150000
200000
250000
300000
350000
400000
450000
m/z-->
Abundance
Scan 2360 (15.821 min): NT193b.D84
133
161
42119
65105 145 299207
٭
B
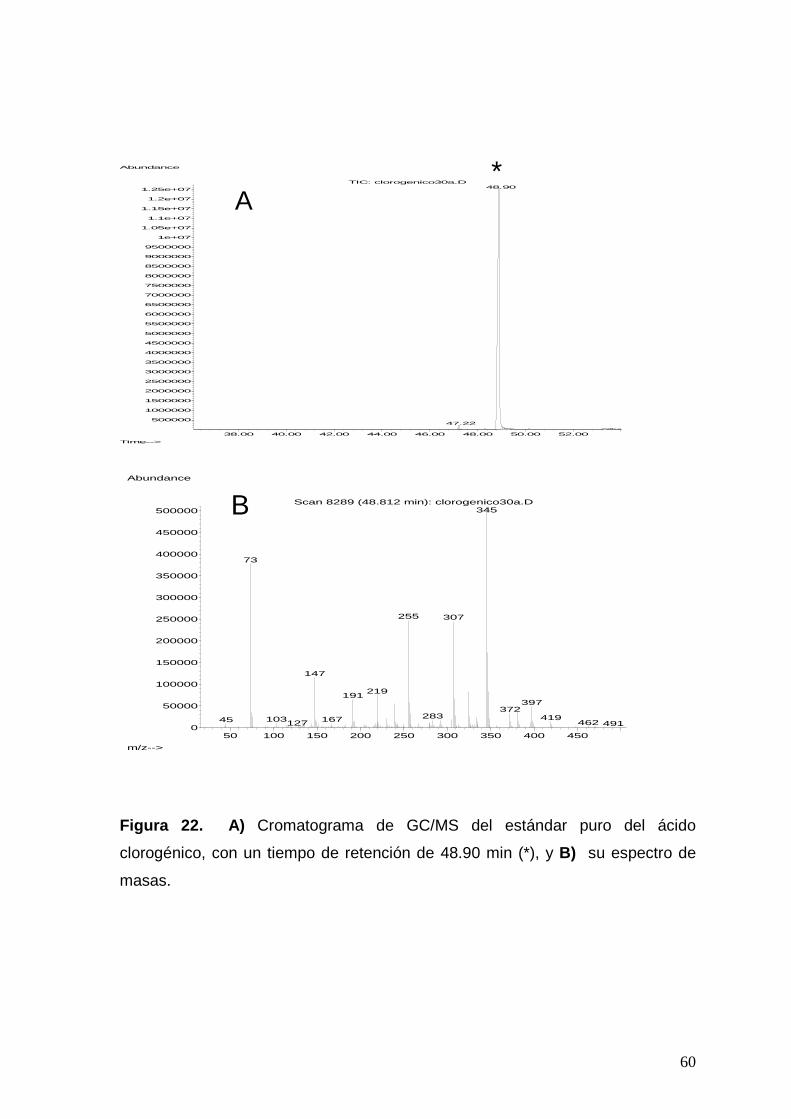
60
Figura 22. A) Cromatograma de GC/MS del estándar puro del ácido
clorogénico, con un tiempo de retención de 48.90 min (*), y B) su espectro de
masas.
38.00 40.00 42.00 44.00 46.00 48.00 50.00 52.00
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
4500000
5000000
5500000
6000000
6500000
7000000
7500000
8000000
8500000
9000000
9500000
1e+07
1.05e+07
1.1e+07
1.15e+07
1.2e+07
1.25e+07
Time-->
Abundance
TIC: clorogenico30a.D
47.22
48.90
A
50 100 150 200 250 300 350 400 4500
50000
100000
150000
200000
250000
300000
350000
400000
450000
500000
m/z-->
Abundance
Scan 8289 (48.812 min): clorogenico30a.D345
73
255 307
147
219191
397372
283 419103 16745 462127 491
*
B

61
Figura 23. A) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando al ácido clorogénico con un tiempo de retencion
de 48.83 min (*), y B) su correspondiente espectro de masas.
50 100 150 200 250 300 350 400 4500
5000
10000
15000
20000
25000
30000
35000
40000
45000
50000
55000
m/z-->
Abundance
Scan 8479 (48.829 min): NT193b.D345
73
255 307
147
219191397
372283 41916745 103 326 462123
46.50 47.00 47.50 48.00 48.50 49.00 49.50 50.00 50.50 51.00 51.50 52.00
50000
100000
150000
200000
250000
300000
350000
400000
450000
500000
550000
600000
650000
700000
750000
800000
Time-->
Abundance
TIC: NT193b.D
48.83*
A
B

62
Figura 24. A) Cromatograma de GC/MS del estándar puro de capsidiol [32.35
min (*)] derivatizado y un producto de degradación estable [posiblemente
capsenona, con un tiempo de retención de 32.18 min (*),, y B) el espectro de
masas correspondiente al pico surgido a los 32.18 min.
31.8031.9032.0032.1032.2032.3032.4032.5032.6032.7032.8032.9033.0033.100
100000
200000
300000
400000
500000
600000
700000
800000
900000
1000000
1100000
Time-->
Abundance
TIC: estCAP1.D
32.18
32.35
50 100 150 200 250 300 350 400 4500
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
22000
24000
26000
28000
30000
32000
m/z-->
Abundance
Scan 5393 (32.187 min): estCAP1.D73
319
147
220
189103 243
45 169 361271 451291
*
A
B
*
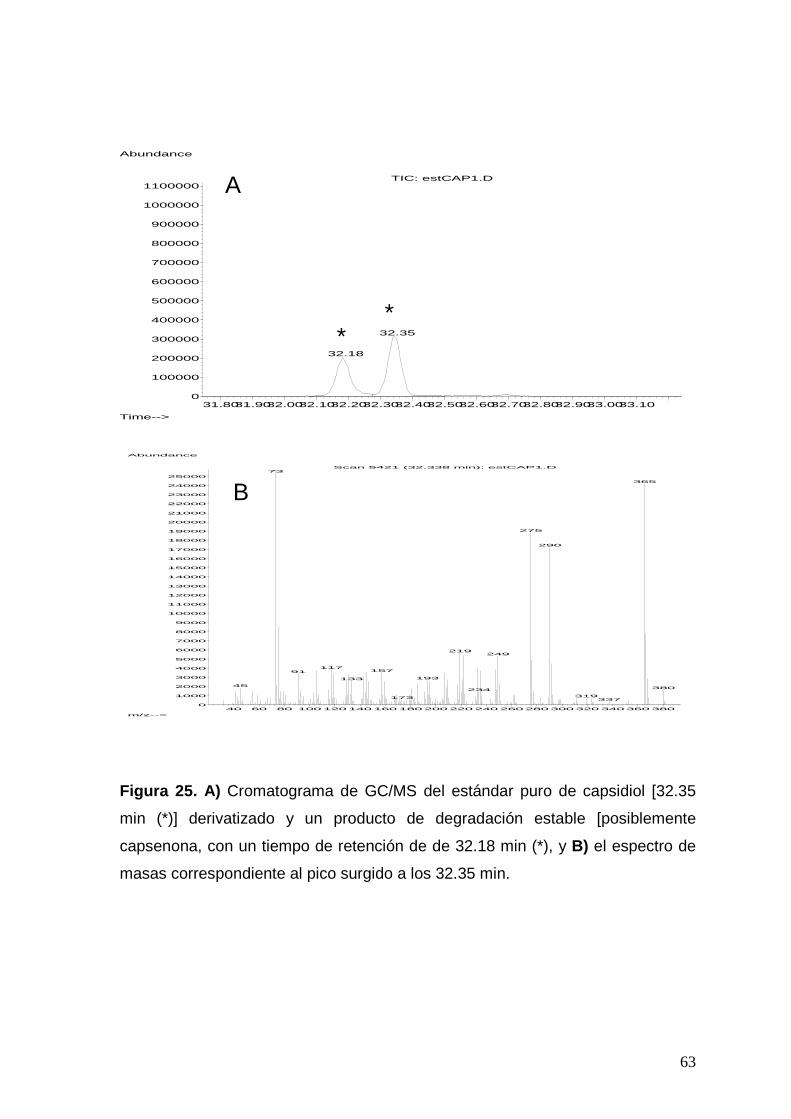
63
Figura 25. A) Cromatograma de GC/MS del estándar puro de capsidiol [32.35
min (*)] derivatizado y un producto de degradación estable [posiblemente
capsenona, con un tiempo de retención de de 32.18 min (*), y B) el espectro de
masas correspondiente al pico surgido a los 32.35 min.
31.8031.9032.0032.1032.2032.3032.4032.5032.6032.7032.8032.9033.0033.100
100000
200000
300000
400000
500000
600000
700000
800000
900000
1000000
1100000
Time-->
Abundance
TIC: estCAP1.D
32.18
32.35
A
* *
40 60 80 100 120 140 160 180 200 220 240 260 280 300 320 340 360 3800
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
m/z-->
Abundance
Scan 5421 (32.338 min): estCAP1.D73
365
275
290
219249
11715791
193133
45380234
319173 337
B
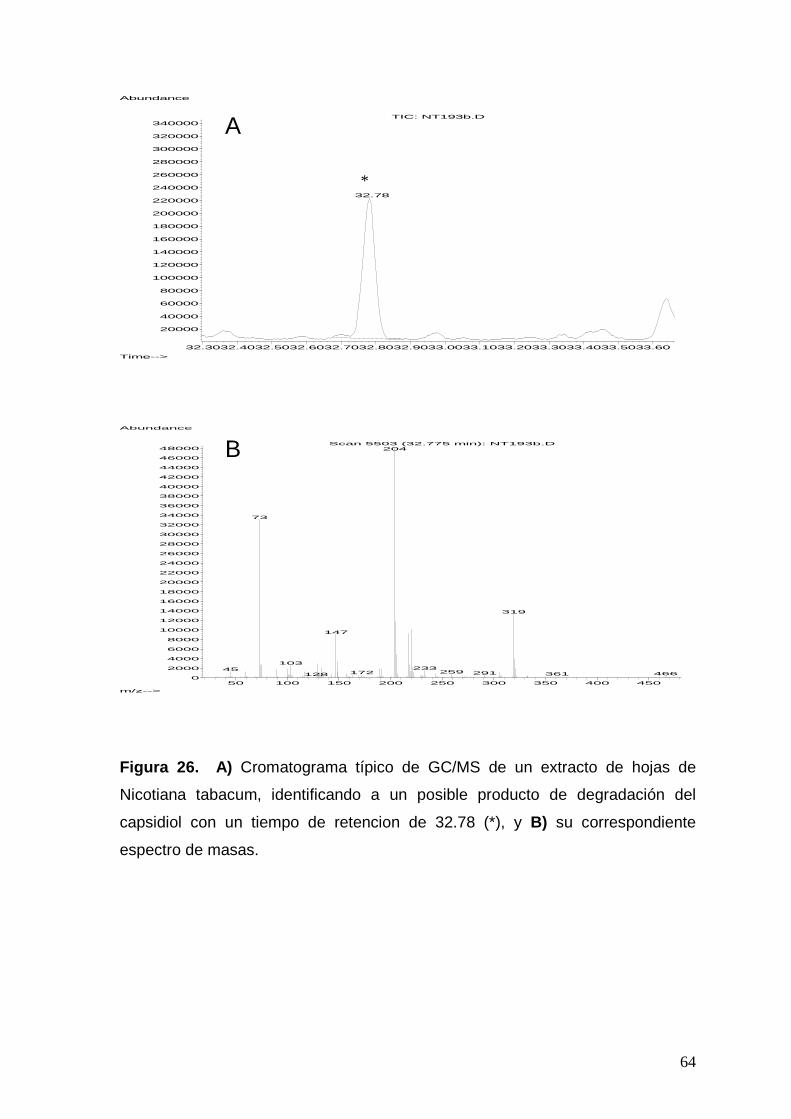
64
Figura 26. A) Cromatograma típico de GC/MS de un extracto de hojas de
Nicotiana tabacum, identificando a un posible producto de degradación del
capsidiol con un tiempo de retencion de 32.78 (*), y B) su correspondiente
espectro de masas.
32.3032.4032.5032.6032.7032.8032.9033.0033.1033.2033.3033.4033.5033.60
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
220000
240000
260000
280000
300000
320000
340000
Time-->
Abundance
TIC: NT193b.D
32.78
50 100 150 200 250 300 350 400 4500
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
22000
24000
26000
28000
30000
32000
34000
36000
38000
40000
42000
44000
46000
48000
m/z-->
Abundance
Scan 5503 (32.775 min): NT193b.D204
73
319
147
10323345 259172 291 466361128
*
A
B

65
Figura 27. A) Cromatograma de GC/MS del estándar puro de nornicotina
derivatizado, con un tiempo de retención de 20.57 min (*), y B) su
correspondiente espectro de masas.
30 40 50 60 70 80 90 1001101201301401501601701801902002102200
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
m/z-->
Abundance
Scan 3063 (20.618 min): nornicotinastd3a.D142
73
205220
191
45 5911832 17710688 132
16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00 32.00 34.00
50000
100000
150000
200000
250000
300000
350000
400000
450000
500000
550000
600000
Time-->
Abundance
TIC: nornicotinastd3a.D 20.54A
B
*
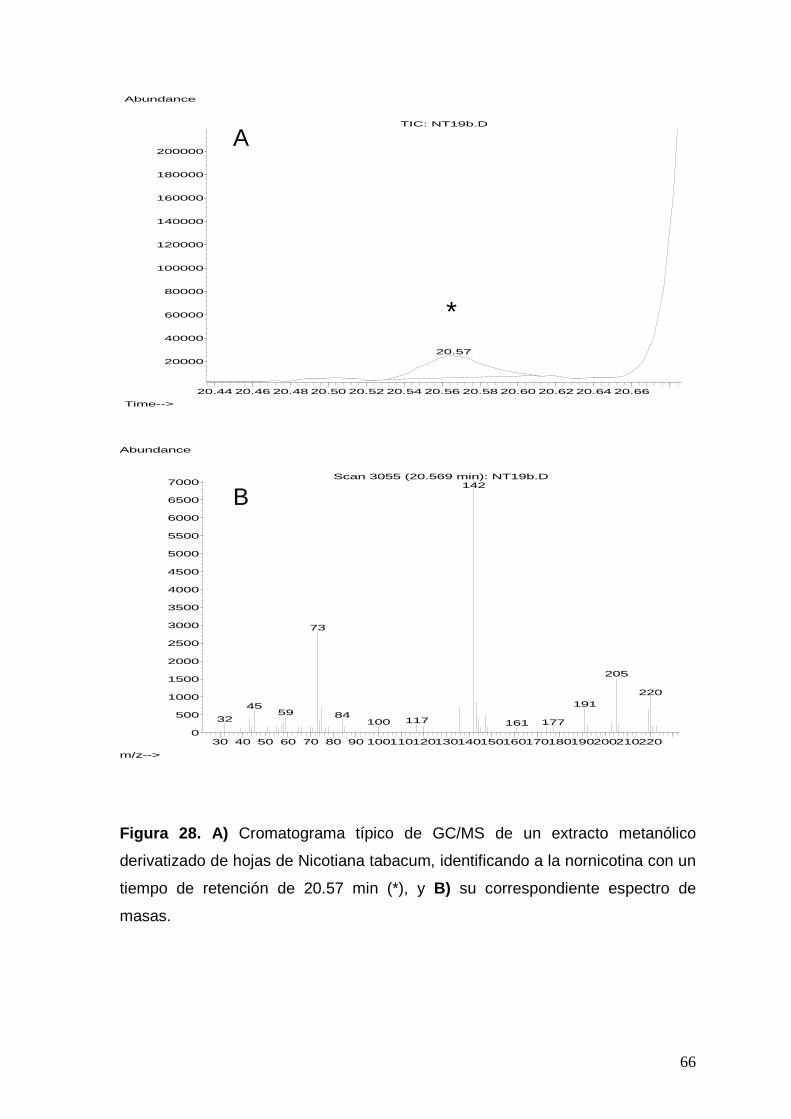
66
Figura 28. A) Cromatograma típico de GC/MS de un extracto metanólico
derivatizado de hojas de Nicotiana tabacum, identificando a la nornicotina con un
tiempo de retención de 20.57 min (*), y B) su correspondiente espectro de
masas.
30 40 50 60 70 80 90 1001101201301401501601701801902002102200
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
6000
6500
7000
m/z-->
Abundance
Scan 3055 (20.569 min): NT19b.D142
73
205
220
1914559 8432 117100 177161
20.44 20.46 20.48 20.50 20.52 20.54 20.56 20.58 20.60 20.62 20.64 20.66
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
Time-->
Abundance
TIC: NT19b.D
20.57
A
B
*

67
6.2. Cuantificación de nicotina en plantas jóvenes y/o adultas
6.2.1. Tratamiento: daño mecánico
Los resultados obtenidos se muestran en la Figura 29. En plantas WT
adultas (Fig. 29 B), se detectó un aumento altamente significativo en los niveles
de nicotina en la hoja dañada (de 2.2 a 5.9 µg/mg peso freso [PF]), hojas distales
intactas (superior: de 1.4 a 5.2 µg/mg peso freso [PF]; inferior: de 2.0 a 2.7
µg/mg peso freso [PF]) y raíz (de 0.96 a 1.22 µg/mg peso freso [PF]). Aunque
todas las diferencias se produjeron con un nivel de significancia p <0.01, éstas
fueron más marcadas en la hoja dañada y en la hoja distal joven. La inducción
en la hoja distal vieja está de acuerdo con la generación de señales
bidireccionales que podrían ser de diferente naturaleza química, en coincidencia
con los resultados reportados por Schittko y Baldwin (2003).
En plantas de la cruza transgénicas WT × Nt-LeSYS 03, se observó un
patrón de inducción casi idéntico al de plantas WT, excepto por la falta de una
inducción local significativa, quizás como consecuencia de los niveles basales
más altos que los observados en plantas WT. La acumulación de nicotina
inducida por daño en raíces de esta línea de plantas fue la más alta detectada
(de 0.88 a 2.12 µg/mg PF).
En plantas de las líneas Nt-LeSYS 03 y Nt-LePS 08, los niveles basales
también resultaron más altos que en plantas WT. Además, se observó una
acumulación de nicotina inducida por daño más intensa, en particular en la hoja
dañada (respuesta local: de 3.3 y 3.7 a 7.3 y 7.2 µg/mg PF, respectivamente) y
la hoja distal joven (respuesta sistémica en dirección hacia el ápice: de 4.62 y
4.69 a 9 y 8.23 µg/mg PF, respectivamente). No se observó inducción sistémica
de nicotina en respuesta al daño en la hoja distal inferior (en dirección hacia la
base) de plantas Nt-LeSYS 03. En estas plantas se observaron los niveles de
nicotina más bajos en raíz, tanto basales como inducidos por daño (0.6 y 0.85
µg/mg PF, respectivamente).
En resumen, los resultados obtenidos en plantas adultas dañadas indican
que: i) los niveles basales de nicotina en plantas adultas intactas fueron más
altos en plantas transformadas que en plantas WT, excepto en la raíz de plantas

68
Nt-SYS 03, y ii) la inducción local y sistémica (en hojas distales apicales) de
nicotina por daño mecánico fue más intensa en las líneas Nt-LePS 08 y Nt-
LeSYS 03.
Los niveles basales de nicotina en hoja y raíz de plantas WT jóvenes
resultaron similares a los detectados en plantas adultas (2.2 y 0.9 µg/mg PF,
respectivamente) (Fig. 29 A). Sin embargo, el daño mecánico en plantas jóvenes
no indujo una acumulación local de nicotina, mientras que en hojas distales se
detectó una reducción significativa en los niveles de nicotina (de 3 a 2.1 µg/mg
PF).
En general, el efecto causado por daño mecánico en plantas transgénicas
jóvenes fue similar (neutro o negativo) al observado en plantas WT. Sólo se
detectaron niveles superiores a los de las plantas intactas en la respuesta local
en la línea Nt-LeSYS 03 y en la respuesta sistémica en plantas WT × Nt-LeSYS
03. En contraste, los niveles de nicotina, locales y sistémicos (en hoja y raíz),
detectados en las plantas Nt-LePS 08 fueron significativamente inferiores a los
detectados en plantas intactas (1.12 vs 1.21, 1.0 vs 1.6 y 0.73 vs 0.54 µg/mg PF
en hoja dañada, hoja distal y raíz de plantas dañadas e intactas,
respectivamente).
6.2.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas)
En plantas WT, el aumento en los niveles de nicotina causado por
herbivoría en la hoja dañada fue similar al observado en plantas dañadas
mecánicamente, mientras que en la raíz, la acumulación de nicotina fue
aproximadamente 5 veces mayor a la inducida por daño mecánico (1.2 vs. 6.2
µg/mg PF). A su vez, no se detectaron diferencias en la acumulación sistémica
de nicotina entre ambos tratamientos (Fig. 30).
En todas las plantas transgénicas dañadas por larvas de M. sexta se
observó una marcada reducción en la acumulación de nicotina en todos los
tejidos analizados, con respecto a niveles basales detectados en plantas
transformadas no dañadas de la misma edad, particularmente en raíz. Este
efecto fue mucho más evidente en las líneas Nt-LePS 08 (Fig. 30 B), y en WT ×

69
PLANTA JÓVEN PLANTA ADULTA
Figura 29. Niveles de nicotina en plantas (jóvenes y adultas) WT y transgénicas
intactas (0 h) y cinco días después de haber sido sometidas a daño mecánico (5
día). En hoja local (HL), hoja sistémica (HS), hoja distal vieja (HDV), hoja distal
jóven (HDJ) y raíz. Los asteriscos indican la diferencia estadísticamente
significativa (P ≤ 0.05*; P ≤ 0.01 **).
WT
0
1
2
3
4
HL HS RAÍZug N
icot
ina/
mg
PF
0 h
5 día
*
A
WT X Nt-LeSYS 03
0
1
2
HL HS RAÍZug N
icot
ina/
mg
PF
0 h
5 día **
C
Nt-LePS 08
0
1
2
HL HS RAÍZug N
icot
ina/
mg
PF
0 h
5 día
**
*****
*
E
Nt-LeSYS 03
0
1
2
3
4
HL HS RAÍZug N
icot
ina/
mg
PF
0 h
5 día *
G
WT
0
2
4
6
8
HL HDV HDJ RAÍZug N
icot
ina/
mg
PF
0 h
5 día
**
**
*
**
B
WT X Nt-LeSYS 03
0
2
4
6
8
HL HDV HDJ RAÍZug N
icot
ina/
mg
PF
0 h
5 día
****
**
D
Nt-LePS 08
02468
10
HL HDV HDJ RAÍZug N
icot
ina/
mg
PF
0 h
5 día
*
****
**
F
Nt-LeSYS 03
02468
10
HL HDV HDJ RAÍZug N
icot
ina/
mg
PF
0 h
5 día
**
****
H

70
Nt-LeSYS 03 (Fig. 30 A) que en la línea Nt-LeSYS 03 (Fig. 30 C). Esto se reflejó
en la detección de contenidos de nicotina en las plantas transgénicas dañadas
por M. sexta que fueron marginal o significativamente inferiores a los observados
en plantas WT igualmente tratadas.
Los resultados indicaron que la sobreexpresión de LePS y LeSYS en N.
tabacum, causó una represión generalizada de la acumulación de nicotina en
respuesta a herbivoría por M. sexta. Este efecto se observó en todos los tejidos
analizados, particularmente en raíz, y en menor proporción en las hojas dañada
y distal superior. Mientras tanto, la acumulación de nicotina inducida por
herbivoría en plantas WT se detectó únicamente en las hojas dañadas y la raíz.
En raíz, la inducción fue sumamente fuerte: 6.5 × los niveles detectados en raíz
de plantas intactas, y 5.1 × los niveles detectados en raíz de plantas dañadas
mecánicamente.
6.2.3. Tratamiento: infestación por mosquita blanca (plantas
adultas)
Los resultados obtenidos al analizar las plantas en estudio al término del
ciclo de vida de B. tabaci (28 días), en los cuales se compararon los niveles de
nicotina acumulados en plantas WT con cada una de las líneas transgénicas, se
muestran en la Figura 31 (A-C). Los niveles de nicotina detectados en la hoja
infestada y en la raíz de las plantas WT resultaron menores a los niveles basales
de nicotina en plantas intactas de edad similar, especialmente en raíz, donde se
observó una reducción a niveles no detectables. A su vez, se observó un ligero
incremento de nicotina (×1.5) en la hoja distal superior.
En forma similar a lo observado en los ensayos con M. sexta, la
infestación con B. tabaci en las tres líneas de plantas transgénicas redujo
marcadamente los niveles de nicotina en relación a plantas transgénicas intactas
de la misma edad. Este efecto se reflejó más claramente en la hoja infestada y la
raíz. En dos líneas (WT × Nt-LeSYS 03 y Nt-LePS 08), ver Figs. 31 A y B, la
nicotina en raíz se redujo a niveles no detectables, mientras que ésta se
mantuvo sin cambio en la línea Nt-LeSYS 03 (Fig. 31 C).
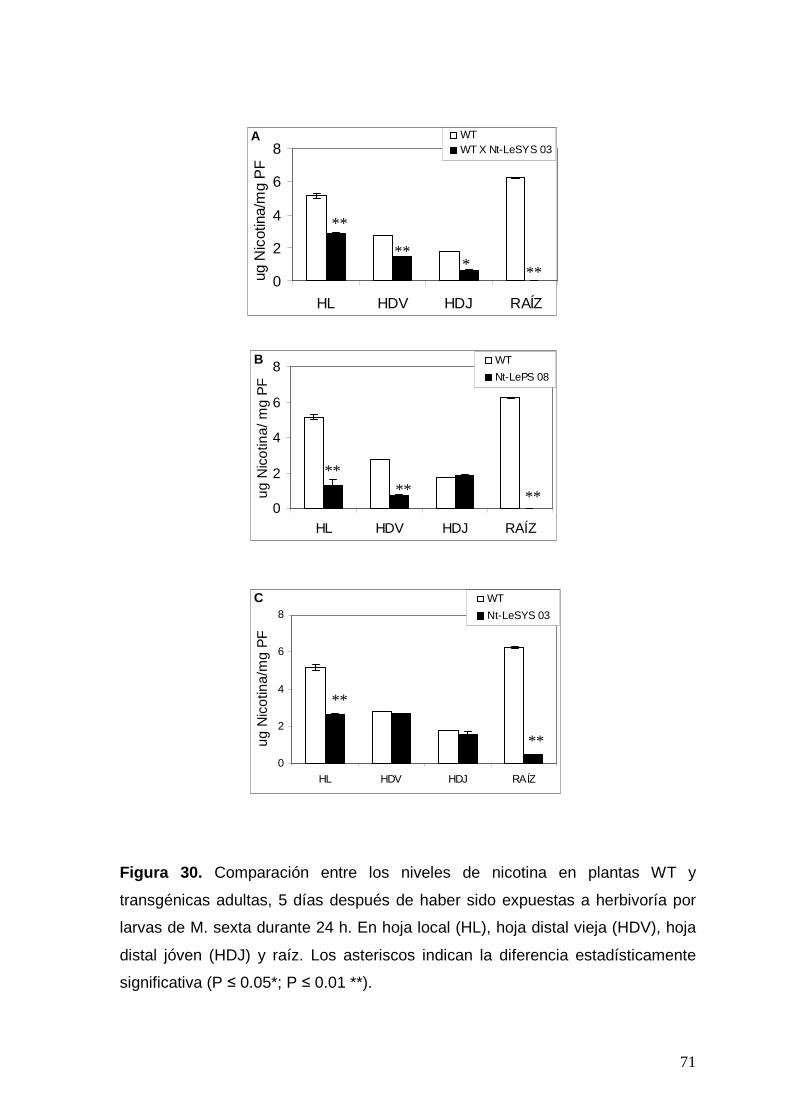
71
Figura 30. Comparación entre los niveles de nicotina en plantas WT y
transgénicas adultas, 5 días después de haber sido expuestas a herbivoría por
larvas de M. sexta durante 24 h. En hoja local (HL), hoja distal vieja (HDV), hoja
distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia estadísticamente
significativa (P ≤ 0.05*; P ≤ 0.01 **).
0
2
4
6
8
HL HDV HDJ RAÍZ
ug N
icot
ina/
mg
PF
WT
Nt-LePS 08
**** **
B
0
2
4
6
8
HL HDV HDJ RAÍZ
ug N
icot
ina/
mg
PF
WT
Nt-LeSYS 03
**
**
C
0
2
4
6
8
HL HDV HDJ RAÍZ
ug N
icot
ina/
mg
PF
WTWT X Nt-LeSYS 03
**
*** **
A

72
Al comparar los niveles de nicotina presentes en plantas infestadas WT y
transgénicas, se observó un incremento significativo en la respuesta local y
sistémica en follaje de plantas Nt-LePS 08 (Fig. 31 B). En las plantas Nt-LeSYS
03 (Fig. 31 C), la acumulación sistémica de nicotina fue significativamente
superior a la observada en plantas WT, particularmente en raíz, mientras que el
efecto represor de la infestación por MB fue muy fuerte en plantas WT × Nt-
LeSYS 03 (Fig. 31 A), cuyos niveles de nicotina en hoja infestada y hoja distal
superior fueron significativamente inferiores a los detectados en plantas WT
infestadas.
Los resultados obtenidos indica que la infestación por B. tabaci no tuvo un
efecto evidente sobre la inducción de la acumulación de nicotina en hojas de
plantas WT (±1.5× los niveles basales detectados en plantas intactas de la
misma edad), pero redujo drásticamente la acumulación de nicotina en raíz (a
niveles no detectables). Curiosamente, la ausencia de nicotina en la raíz de
plantas infestadas no se observó en la línea Nt-LeSYS 03 (Fig. 31 A), mientras
que los niveles de nicotina en las hojas de las plantas transgénicas infestadas
Nt-LePS 08 (respuesta local y sistémica; Fig. 30 B) y Nt-LeSYS 03 (sólo
respuesta sistémica; Fig. 31 C), fueron significativamente superiores a los
detectados en plantas WT, pero menores a los niveles basales detectados en las
plantas transgénicas intactas de la misma edad. Finalmente, en las plantas WT ×
Nt-LeSYS 03 infestadas se observó una represión de la acumulación de nicotina
en hojas, la cual fue significativamente menor a la observada en plantas WT.
6.3. Cuantificación de ácido clorogénico en plantas jóvenes y/o adultas
6.3.1. Tratamiento: daño mecánico
Los resultados obtenidos se muestran en las Figuras 32 A (plantas
jóvenes) y 32 B (plantas adultas). En general, el contenido de ácido clorogénico
(ACG) detectado tanto en hoja como en raíz fue más alto que el reportado en la
literatura, inclusive al ser comparado con los niveles detectados en plantas de N.
tabacum capaces de altas tasas de síntesis de fenilpropanoides causada po

73
Figura 31. Comparación entre los niveles de nicotina en plantas WT y
transgénicas adultas medidos al cumplirse el ciclo de vida de Bemisia tabaci (28
días después de la oviposición). En hoja local (HL), hoja sistémica (HS) y raíz.
Los asteriscos indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤
0.01 **).
0
2
4
6
HL HS RAÍZ
ug N
icot
ina/
mg
PF
WT
Nt-LePS 08
**
**
B
0
1
2
3
HL HS RAÍZ
ug N
icot
ina/
mg
PF
WT
Nt-LeSYS 03**
**
**
C
0
1
2
3
HL HS RAÍZ
ug N
icot
ina/
mg
PF
WT
WT X Nt-LeSYS 03
****
A

74
redireccionamiento de la ruta biosintética de los fenilpropanoides debido a la
sobreexpresión de FAL (Howles et al., 1996), de una enoil-CoA hidratasa
bacteriana (Mayer et al., 2001) o del gen C58-6b de Agrobacterium tumefaciens
(Gális et al., 2002). En estos casos, los niveles máximos reportados fueron de
0.3 a 5 µg/mg PF. Esta discrepancia posiblemente refleje diferencias entre
variedades de N. tabacum, en forma similar a lo que se ha reportado entre
especies de Nicotiana. Por ejemplo, en la especie silvestre N. attenuata los
niveles de ACG son extremadamente bajos, de aproximadamente 27 µg/g PF
(Keinänen et al., 2001). Las diferencias observadas también podrían haberse
debido al empleo de GC/MS, en lugar de otras técnicas como HPLC/MS, para su
aislamiento y cuantificación.
En plantas WT adultas (Fig. 32 B), los niveles basales promedio de ácido
clorogénico (ACG) detectados fueron los siguientes: 9.01 µg/mg PF (hoja de
transición entre tejido consumidor-tejido fuente [C-F], u hoja local), y 4.3 y 4.1
µg/mg PF en las hojas distales vieja y joven, respectivamente. En raíz, los
niveles de ACG fueron más bajos (1.7 µg/mg PF). El daño mecánico indujo un
incremento significativo en los niveles de ACG tanto local como sistémico y en
todos los tejidos analizados: es decir, se detectó una acumulación de ACG que
fue 1.5, 1.2, 1.3 y 1.18 veces más alta que en plantas intactas en la hoja dañada,
en hojas distales inferior y superior y en raíz, respectivamente.
En la línea WT × Nt-LeSYS 03 los niveles basales de ACG en todos los
tejidos analizados, excepto la hoja de transición tejido consumidor-tejido fuente
(C-F), fueron superiores a los detectados en las plantas WT (entre 1.95 [raíz] y
3.8 [hoja distal inferior] veces más altos). Sin embargo, el daño mecánico causó
una disminución significativa en los niveles de ACG en todos los tejidos
analizados de estas plantas (entre 2.4 [respuesta sistémica en raíz] y 5.3
[respuesta sistémica en hoja distal superior] veces más bajos).
Al comparar los niveles basales de ACG en plantas WT y Nt-LePS 8, se
observó una drástica reducción (27× menores), a valores casi indetectables, en
la hoja de transición C-F (hoja local) de ésta última, mientras que en las hojas
distales inferior y superior, y en raíz, los niveles basales de ACG fueron
superiores a los de las plantas WT (1.5× [hoja distal inferior], 4.1× [hoja distal
superior] y 1.3× [raíz] veces más altos). Curiosamente el daño mecánico causó

75
un incremento significativo en ACG solamente en los tejidos aéreos más viejos
de plantas de esta línea (hoja C-F o local [25×] y hoja distal inferior [1.6×]); en
raíz y en la hoja distal superior se observó una disminución significativa, la cual
fue mucho más marcada en éste último tejido (13×). En plantas WT × Nt-LeSYS
03 se detectó una disminución altamente significativa de ACG en todos los
tejidos de las plantas dañadas.
En la línea Nt-LeSYS 03 se observó una disminución generalizada en los
niveles basales de ACG, los cuales fueron entre 2.5 (raíz) y 6.2 (hoja C-F) más
bajos que los detectados en plantas WT intactas. Al igual que la línea WT × Nt-
LeSYS 03, el daño mecánico provocó una disminución altamente significativa en
los niveles de ACG en tejido aéreo, particularmente la hoja distal inferior
(respuesta sistémica basal) donde el ACG disminuyó a valores no detectables.
En cambio, esta línea fue la única en la cual se detectó una acumulación
significativa de ACG en raíces de plantas dañadas.
Los niveles basales de ACG en plantas WT jóvenes fueron menores a los
detectados en las plantas adultas, particularmente en la hoja inferior, u hoja
“local” (0.56 µg/mg PF) (Fig. 32 A). Los niveles de este metabolito fueron 16
(hoja inferior), 3.3 (hoja superior) y 4.5 veces más bajos que los detectados en la
hojas C-F y distal inferior, y raíces de plantas adultas, respectivamente. Un
comportamiento similar se observó en las tres líneas de plantas transgénicas. A
su vez, el daño mecánico en plantas WT sólo produjo una acumulación sistémica
significativa de ACG en las hojas inferiores adyacentes intactas y en la raíz.
Los niveles basales de ACG en hojas de las tres líneas de plantas
transgénicas analizadas fueron menores a los detectados en las plantas WT,
particularmente en las hojas de la línea Nt-LeSYS 03, donde no se detectó este
metabolito en las hojas inferiores. En hojas jóvenes adyacentes superiores los
niveles se redujeron, entre 3.0 y 3.6 veces, con respecto a hojas similares en
plantas WT. La respuesta sistémica al daño mecánico en la línea WT × Nt-
LeSYS 03 fue similar a la observada en plantas WT, mientras que en las raíces
de plantas dañadas se observó una desaparición de ACG. La ausencia de ACG
en raíces de plantas dañadas también se observó en la línea Nt-LePS 08.
Asimismo, el daño mecánico causó una reducción significativa de ACG a nivel
local (Nt-LePS 08) y sistémico (Nt-LeSYS 03). Resultó interesante observar la

76
completa falta de respuesta local al daño mecánico en esta última línea, lo cual
contrastó con la acumulación de ACG en raíz, cuyos niveles fueron los más
elevados de todas las plantas en estudio.
En síntesis, los niveles de ACG en tabaco están al parecer regulados por
el estado de desarrollo de la planta, ya que se incrementaron notablemente en
plantas WT adultas. El estado de desarrollo de la planta también ejerció una
influencia sobre la acumulación de ACG en follaje en respuesta al daño
mecánico en plantas WT, ya que ésta fue detectada tanto en forma local como
sistémica en plantas adultas, mientras que en plantas jóvenes solo se observó
en forma sistémica, incluyendo la raíz. Excepto en casos aislados, se observó
una disminución generalizada en los niveles de ACG basales e inducidos por
daño en las plantas transformadas, la cual también se vio fuertemente influida
por el estado de desarrollo de la planta. El ejemplo más claro de esta tendencia
se observó en la línea WT × Nt-LeSYS 03, que en hojas de plantas jóvenes se
comportó en forma similar a las plantas WT, detectándose un incremento
significativo de ACG en respuesta al daño en tejido sistémico, mientras que en
plantas adultas el daño causó una disminución significativa en los niveles de
ACG en todos los tejidos analizados. Asimismo, los niveles de ACG, tanto
basales como inducidos por daño, se redujeron drásticamente tanto en plantas
jóvenes (líneas Nt-LePS 08 y Nt-LeSYS 03) como adultas (línea Nt-LeSYS 03).
Esta última línea se distinguió por la alta acumulación constitutiva de ACG en
raíz, cuyos niveles se incrementaron aún más en plantas adultas dañadas
mecánicamente. Esta característica contrastó con lo observado en las raíces de
plantas jóvenes y adultas de las dos líneas restantes, en las cuales el daño
mecánico redujo el contenido de ACG en forma altamente significativa,
especialmente en plantas jóvenes, donde no pudo ser detectado.

77
PLANTA JÓVEN PLANTA ADULTA
Figura 32. Niveles de ácido clorogénico en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas a daño
mecánico (5 día). En hoja local (HL), hoja sistémica (HS), hoja distal vieja (HDV),
hoja distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
WT
0
1
2
3
HL HS RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
*
A
WT X Nt-LeSYS 03
0.0
0.2
0.4
0.6
HL HS RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
C
Nt-LePS 08
0.0
0.2
0.4
0.6
HL HS RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
*
E
Nt-LeSYS 03
0.0
0.2
0.4
0.6
HL HS RAÍZ
ug C
loro
géni
co/m
g P
F
0 h
5 día
G
WT
0
5
10
15
20
HL HDV HDJ RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
*
*
*
***
B
WT X Nt-LeSYS 03
0
5
10
15
20
HL HDV HDJ RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
*
**
*
**** **
D
Nt-LePS 08
0
5
10
15
20
HL HDV HDJ RAÍZug C
loro
géni
co/m
g P
F0 h
5 día
*
**
***
**
F
Nt-LeSYS 03
0
1
2
3
HL HDV HDJ RAÍZug C
loro
géni
co/m
g P
F
0 h
5 día
****
**
H

78
6.3.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas)
Los resultados obtenidos se muestran en la Figura 33. En plantas WT
sometidas a herbivoría con larvas de M. sexta solo se detectó ACG en las hojas
dañadas. Además, los niveles detectados en éstas disminuyeron drásticamente
con respecto a lo observado en hojas similares en plantas intactas (21×
menores) y dañadas mecánicamente (31× menores).
El efecto negativo sobre los niveles de ACG causado por la folivoría de
larvas de M. sexta en plantas WT se agudizó en las tres líneas de plantas
transgénicas analizadas, ya que en todos casos, el ACG disminuyó a niveles no
detectables en todos los tejidos analizados.
En resumen, la folivoría por M. sexta provocó una disminución muy
marcada en los niveles de ACG en plantas WT y una supresión de su síntesis y
acumulación en las plantas transformadas, donde este metabolito se redujo
hasta niveles no detectables.
6.3.3. Tratamiento: infestación por mosquita blanca (plantas adultas)
Los resultados obtenidos se muestran en la Figura 34. En plantas WT se
observó una disminución en los niveles de ACG, con respecto a los niveles
basales en plantas no infestadas de la misma edad, en las hojas infestada con B.
tabaci (1.8× menores) y en raíces (2.1× menores); mientras que en la hoja distal
superior (respuesta sistémica, dirección acropetal), los niveles de ACG fueron
ligeramente superiores (1.9× más altos).
Los niveles de ACG detectados en tejidos de plantas infestadas de la
línea Nt-LePS 08 no resultaron estadísticamente diferentes a los detectados en
plantas WT, mientras que en la línea Nt-LeSYS 03, el contenido de ACG en
follaje se redujo hasta niveles no detectables, mientras que en raíz la cantidad
detectada fue significativamente menor a la detectada en raíces de plantas WT
infestadas. A su vez, el nivel de ACG en raíces de plantas Nt-LeSYS 03
infestadas fue 1.7× menor a la detectada en plantas transgénicas de la misma
edad no infestadas.

79
Figura 33. Comparación entre los niveles de ácido clorogénico en plantas WT y
transgénicas adultas, 5 días después de haber sido expuestas a herbivoría por
larvas de M. sexta durante 24 h. En hoja local (HL), hoja distal vieja (HDV), hoja
distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia estadísticamente
significativa (P ≤ 0.05*; P ≤ 0.01 **).
0.0
0.5
1.0
HL HDV HDJ RAÍZug c
loro
géni
co/m
g P
F
WT
Nt-LePS 08
**
B
0.0
0.5
1.0
HL HDV HDJ RAÍZug c
loro
géni
co/m
g P
F
WT
Nt-LeSYS 03
**
C
0.0
0.5
1.0
HL HDV HDJ RAÍZug c
loro
géni
co/m
g P
F
WT
WT X Nt-LeSYS 03
**
A

80
La misma tendencia se observó en la línea WT × Nt-LeSYS 03. Sin embargo, la
disminución con respecto a plantas WT infestadas, aunque significativa, no fue
tan drástica, al menos en follaje. La infestación con B. tabaci causó también una
disminución considerable en ACG en todos los tejidos analizados de plantas WT
× Nt-LeSYS 03 con respecto a los niveles basales de ACG en tejidos
equivalentes de plantas de esta línea no infestadas de la misma edad.
Los resultados obtenidos indican que la infestación con B. tabaci en
plantas WT indujo un incremento sistémico (en hoja) y una disminución local y
sistémica (en raíz) en los niveles de ACG en relación a los niveles detectados en
tejidos de plantas WT no infestadas. En forma similar a los resultados obtenidos
de plantas dañadas por M. sexta (Fig. 33), la infestación con B. tabaci causó una
disminución generalizada en los niveles de ACG en plantas WT y transgénicas.
Sin embargo, el efecto fue mucho más marcado en plantas transformadas con
LeSYS (líneas Nt-LeSYS 03 y WT × Nt-LeSYS 03).
6.4. Cuantificación de dihidrocapsenona en plantas jóvenes y/o adultas
6.4.1. Tratamiento: daño mecánico
Los resultados obtenidos se muestran en la Figura 35. Los datos que aquí
se presentan representan una cuantificación indirecta de los niveles de capsidiol,
ya que debido a la conocida inestabilidad de este metabolito in planta, aunada a
los largos períodos de muestreo (≥28 d) solo pudo detectarse un producto de
degradación de este metabolito (ver Figs. 11 y 24-26), cuya identidad no pudo
ser determinada, pero cuyo espectro de masas resultó semejante al del cis-9,10-
dihidrocapsenona, un posible catabolito estable de capsidiol detectado en
cultivos celulares en suspensión de C. annuum (Ward et al., 1977; Whitehead et
al., 1987). Se considero, por lo tanto, que la dihidrocapsenona indica los niveles
de su precursor el capsidiol). Los niveles detectados en las plantas en estudio
resultaron de 3 a 4 órdenes de magnitud más altos a los reportados en la
literatura en tejido de N. tabacum inducido por patógenos o PMAP.

81
Figura 34. Comparación entre los niveles de ácido clorogénico en plantas WT y
transgénicas adultas medidos al cumplirse el ciclo de vida de Bemisia tabaci (28
días después de la oviposición). En hoja local (HL), hoja sistémica (HS) y raíz.
Los asteriscos indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤
0.01 **).
0
5
10
15
HL HS RAÍZug
clo
rogé
nic
o/m
g P
F
WTNt-LePS 08
B
0
5
10
15
HL HS RAÍZug
clor
ogén
ico
/mg
PF
WT
Nt-LeSYS 03
** ** *
C
0
5
10
15
HL HS RAÍZu
g cl
oro
gén
ico/
mg
PF
WT
WT X Nt-LeSYS 03
***
*
A

82
En plantas WT, los niveles basales de dihidrocapsenona en plantas
jóvenes fueron entre 6.5 (en raíz) y 7.5 (en la hoja distal superior) veces mayores
a los detectados en tejidos equivalentes de plantas adultas, en las cuales no se
detectó dihidrocapsenona l en las hojas C-F. Además, el daño mecánico abatió
los niveles de dihidrocapsenona en todos los tejidos analizados de plantas
jóvenes (la disminución fue altamente significativa a nivel local y sistémico en la
hoja dañada y en raíz, respectivamente, y marginal a nivel sistémico en la hoja
adyacente superior), mientras que lo contrario se observó en plantas adultas, en
las cuales el daño indujo una acumulación altamente significativa de
dihidrocapsenona en todos los tejidos (aunque sin alcanzar los niveles
detectados en las plantas jóvenes). Esto sugiere que la síntesis constitutiva e
inducida de capsidiol esta regulada por el estado de desarrollo de la planta.
El efecto del daño mecánico también se modificó en las tres líneas de
plantas transgénicas analizadas el cual fue, asimismo, influido por el estado de
desarrollo. De manera que en plantas jóvenes, el daño mecánico en plantas Nt-
LePS 08 y WT × Nt-LeSYS 03 indujo una acumulación de dihidrocapsenona en
todos los tejidos analizados (excepto en la respuesta local en la línea Nt-LePS
08), aunque los niveles basales en estas dos líneas de plantas fueron, en
promedio, aproximadamente 6 veces menores a los detectados en las plantas
WT. Sin embargo, los niveles de detectado dihidrocapsenona en plantas WT y
transgénicas (de estas dos líneas) dañadas mecánicamente fueron muy
similares. En plantas Nt-LeSYS 03, el efecto del daño fue mucho más notable,
particularmente en follaje, ya que al parecer indujo la síntesis de novo de este
metabolito, el cual no se detectó en plantas intactas, mientras que en raíz no se
detectaron cambios significativos. A pesar de ello, los niveles de
dihidrocapsenona en plantas Nt-LeSYS 03 dañadas fueron, en general, menores
a los detectados en el resto de las plantas dañadas analizadas.
Al igual que en plantas WT, el daño mecánico en plantas adultas de las
líneas plantas Nt-LePS 08 y WT × Nt-LeSYS 03 indujo una acumulación
generalizada y altamente significativa de dihidrocapsenona. La diferencia radicó
en el hecho de que los niveles de este metabolito acumulados en las plantas
transgénicas dañadas fueron muy elevados, entre 9 (respuesta sistémica en hoja
distal superior en las línea WT × Nt-LeSYS 03) y 80 (hoja dañada de ambas

83
líneas) veces más altos a los detectados en plantas WT dañadas. En plantas Nt-
LeSYS 03 adultas, que a diferencia de las plantas jóvenes tenían niveles basales
detectables de dihidrocapsenona en follaje, pero que en raíz fueron 1.8 veces
menores a los detectados en plantas jóvenes, sólo se observó un incremento
significativo en los niveles de dihidrocapsenona a nivel sistémico en la hoja distal
superior. En la hoja distal inferior el efecto del daño fue inhibitorio, mientras que
no se observaron cambios significativos en la hoja dañada ni en la raíz. En
plantas adultas de esta línea, la acumulación sistémica de dihidrocapsenona
inducida por daño no fue de la magnitud observada en las dos líneas restantes,
sino similar a la observada en plantas WT.
6.4.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas)
Los resultados se muestran en la Figura 36. En plantas WT adultas
sometidas a herbivoría por M. sexta, se observó un incremento con respecto a
los niveles basales de dihidrocapsenona, el cual fue más marcado en la hoja
dañada y en la raíz (8.3× más alta). El incremento en dihidrocapsenona en
ambas hojas distales fue menor, particularmente en la hoja superior. Los niveles
de dihidrocapsenona detectados en plantas adultas sometidas a herbivoría
también fueron superiores a los detectados en plantas adultas dañadas
mecánicamente.
En la línea Nt-LePS 08, la herbivoría causó una marcada reducción en los
niveles de dihidrocapsenona, hasta valores no detectables en ambas hojas
distales (respuesta sistémica acropetala y basipetala), mientras que en la hoja
dañada C-F (respuesta local), la dihidrocapsenona aumentó hasta niveles
detectables, en contraste con la ausencia de este metabolito en hojas C-F de
plantas intactas. A su vez, el contenido de dihidrocapsenona en raíces de
plantas de esta línea dañadas por M. sexta se redujo a niveles no detectables, lo
que contrastó con la acumulación significativa de dihidrocapsenona detectada en
raíces de plantas dañadas mecánicamente.

84
PLANTA JÓVEN PLANTA ADULTA
Figura 35. Niveles de capsidiol, cuantificado directamente en su posible
producto de degradación (dihidrocapsenona), en plantas jóvenes y adultas, WT y
transgénicas intactas 0 día y cinco días después de haber sido sometidas a daño
mecánico. En hoja local (HL), hoja sistémica (HS), hoja distal vieja (HDV), hoja
distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia estadísticamente
significativa (P ≤ 0.05*; P ≤ 0.01 **).
WT
0.0
0.5
1.0
1.5
HL HS RAÍZ
0 día
5 día
** **P=0.065
A
WT X Nt-LeSYS 03
0.0
0.5
1.0
1.5
HL HS RAÍZ
0 día
5 día
****
C
Nt-LePS 08
0.0
0.5
1.0
1.5
HL HS RAÍZ
0 día
5 día
** **
E
Nt-LeSYS 03
0.0
0.5
1.0
1.5
HL HS RAÍZ
0 día
5 día
** **
G
WT
0.0
0.1
0.2
0.3
0.4
0.5
HL HDV HDJ RAÍZ
0 día
5 día
******
**
B
WT X Nt-LeSYS 03
0
5
10
15
HL HDV HDJ RAÍZ
0 día
5 día**
**
** **
D
Nt-LePS 08
0
5
10
15
HL HDV HDJ RAÍZ
0 día
5 día
**
****
**
F
Nt-LeSYS 03
0.0
0.1
0.2
0.3
HL HDV HDJ RAÍZ
0 día
5 día
**
**
H

85
En la línea WT × Nt-LeSYS 03 se observó un patrón similar aunque
menos marcado, particularmente en la respuesta local a la herbivoría, que no fue
diferente a la detectada en plantas intactas de la misma edad. En raíces de esta
planta, los niveles de dihidrocapsenona en plantas dañadas por M. sexta fueron
3.7× más altos a los basales.
Los niveles de dihidrocapsenona detectados en plantas Nt-LePS 08 y WT
× Nt-LeSYS 03 dañadas por M. sexta fueron considerablemente inferiores a los
detectados en plantas WT dañadas. Asimismo, en ambas líneas se observó un
contraste muy marcado entre los niveles de dihidrocapsenona acumulados en
plantas dañadas mecánicamente (acumulación hasta niveles excepcionalmente
altos) o por larvas de M. sexta (disminución en los niveles o represión de su
síntesis).
Una vez más, la línea Nt-LeSYS 03 se comportó en forma diferente a las
otras dos. En todos los tejidos analizados, los niveles de dihidrocapsenona se
incrementaron en respuesta a herbivoría, con respecto a los niveles basales
detectados en plantas Nt-LeSYS 03 intactas de la misma edad. El efecto más
marcado fue en la hoja dañada y en la hoja distal inferior (niveles 5.3× más altos
en ambos casos). Los niveles detectados en plantas sometidas a folivoría por M.
sexta también fueron más altos a los detectados en plantas dañadas
mecánicamente. La acumulación de dihidrocapsenona en follaje en respuesta a
herbivoría en estas plantas fue marginal o significativamente (respuesta
sistémica en hoja distal inferior) más alta a la detectada en plantas WT. Sin
embargo, al igual que en las líneas Nt-LePS 08 y WT × Nt-LeSYS 03, los niveles
de dihidrocapsenona en la raíces de plantas dañadas por M. sexta fueron
significativamente inferiores a los detectados en plantas WT similarmente
tratadas.

86
Figura 36. Comparación entre los niveles de capsidiol, cuantificado
indirectamente en forma de un posible producto de degradación
(dihidrocapsenona) en plantas WT y transgénicas adultas, 5 días después de
haber sido expuestas a herbivoría por larvas de M. sexta durante 24 h. En hoja
local (HL), hoja distal vieja (HDV), hoja distal jóven (HDJ) y raíz. Los asteriscos
indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
0.0
0.5
1.0
1.5
2.0
HL HDV HDJ RAÍZ
ug c
apsi
diol
/ mg
peso
fre
sco.
WTNt-LePS 08
** ***
B
**
0.0
0.5
1.0
1.5
2.0
HL HDV HDJ RAÍZug c
apsi
diol
/ mg
peso
fres
co.
WTNt-LeSYS 03
**
**P = 0.22
P= 0.07
C
0.0
0.5
1.0
1.5
2.0
HL HDV HDJ RAÍZug
cap
sidi
ol/ m
g p
eso
fres
co.
WT
WT X Nt-LeSYS 03
**
***
A

87
6.4.3. Tratamiento: infestación por mosquita blanca (plantas adultas)
Los resultados obtenidos de este tratamiento se muestran en la Figura 37.
La infestación por B. tabaci causó un marcado e inusual incremento en los
niveles de dihidrocapsenona en todos los tejidos analizados de plantas WT,
particularmente en la hoja infestada. Los altos niveles detectados apoyan la
proposición de que el dihidrocapsenona inducido por la infestación por B. tabaci
se acumuló gradualmente en forma de su catabolito más estable durante los
relativamente largos tiempos de muestreo empleados en este experimento (28
días). La acumulación sistémica de dihidrocapsenona también fue elevada (11 y
13× mayor en la raíz y la hoja distal superior, respectivamente, a la detectada en
plantas WT intactas de la misma edad). Comparando todas las plantas en
estudio, fue evidente que la infestación por B. tabaci indujo la mayor
acumulación de dihidrocapsenona en plantas WT.
Excepto por la caída en los niveles de dihidrocapsenona en las raíces de
plantas WT × Nt-LeSYS 03 infestadas, el patrón de acumulación de
dihidrocapsenona en respuesta a MB en las Nt-LePS 08 y WT × Nt-LeSYS 03
fue similar, aunque de menor magnitud, al observado en las plantas WT. En la
línea Nt-LeSYS 03 la respuesta sistémica en hoja y raíz también fue superior o
similar a la de las demás plantas, mientras que dihidrocapsenona en la hoja
infestada disminuyó hasta niveles no detectables. En la línea Nt-LePS 08 la
acumulación local de dihidrocapsenona en plantas infestadas y dañadas
mecánicamente fue similar, mientras que la respuesta sistémica, tanto en la hoja
distal superior como en la raíz, fue menor en las plantas infestadas.
En las tres líneas de plantas transgénicas la acumulación local y sistémica
(hoja distal superior y raíz) de dihidrocapsenona en respuesta a B. tabaci fue
igual o significativamente menor a la detectada en tejido equivalente de plantas
WT infestadas, excepto por la respuesta sistémica en hoja en plantas Nt-LeSYS
03, en la cual los niveles de dihidrocapsenona detectados fueron
significativamente más altos que en plantas WT.
En síntesis, los cambios observados en los niveles basales de
dihidrocapsenona mayores en plantas WT jóvenes, y su incremento en plantas
adultas, o disminución (plantas jóvenes) en respuesta al daño mecánico indican

88
que los niveles constitutivos o inducidos de este metabolito en planta están
ontogénicamente determinados. La acumulación de dihidrocapsenona en
respuesta a daño mecánico en plantas adultas y a infestación por MB se
potenció marcadamente en la línea Nt-LePS 08 y en menor grado en la línea WT
× Nt-LeSYS 03, particularmente en la hoja dañada o infestada. El daño por
insectos chupadores y masticadores indujo una acumulación local y sistémica de
dihidrocapsenona en plantas WT, la cual fue más intensa en respuesta a B.
tabaci. En general, la inducción de dihidrocapsenona en respuesta al daño por
insectos se redujo significativamente en las plantas transgénicas.
6.5. Cuantificación de nornicotina en plantas jóven es y/o adultas
6.5.1. Tratamiento: daño mecánico
Los resultados obtenidos se muestran en las Figuras 38 A y B. Éstos
indicaron que la síntesis basal de nornicotina (NN) y la inducción de su
acumulación en respuesta a daño mecánico en N. tabacum dependen del estado
de desarrollo de la planta, del tipo de tejido analizado y de la filotaxia de la
planta. De manera que no se detectó este alcaloide en plantas WT jóvenes ni en
la hoja de transición C-F de plantas adultas intactas. En éstas, NN se detectó en
las dos hojas distales, superior (0.041 µg/mg PF) e inferior (0.031 µg/mg PF) en
relación a la hoja C-F y más claramente en raíz (0.17 µg/mg PF). La cantidad de
NN detectada resultó menor a lo esperado para N. tabacum, donde este
alcaloide alcanzó hasta 0.35 µg/mg PF, equivalentes al 3% del contenido total de
alcaloides en esta especie (Saitoh et al., 1985). Al igual que en el análisis de
CGA descrito con anterioridad, esta variación podría haberse debido a
diferencias genéticas y/o analíticas.
El daño mecánico en plantas WT adultas indujo una síntesis y
acumulación significativa de NN en la hoja dañada (de niveles no detectables a
0.045 µg/mg PF) y en la hoja distal superior (de 0.041 a 0.12 µg/mg PF),
mientras que redujo su concentración a niveles no detectables en la hoja distal
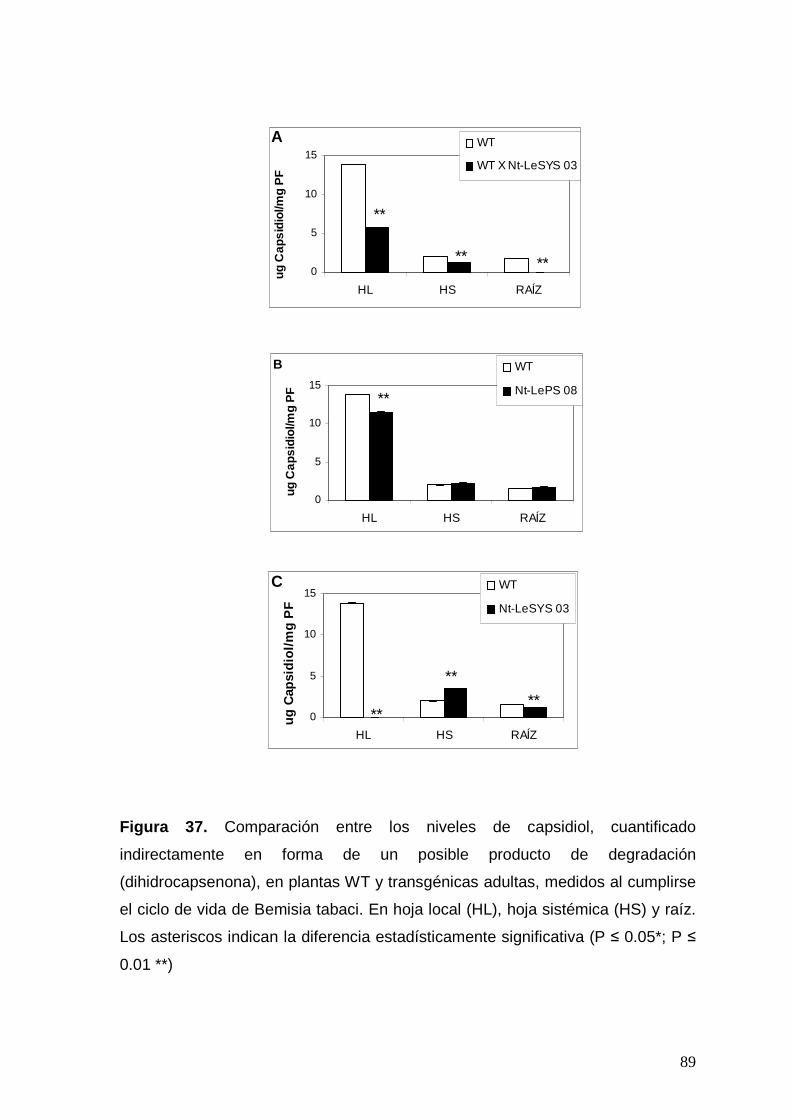
89
Figura 37. Comparación entre los niveles de capsidiol, cuantificado
indirectamente en forma de un posible producto de degradación
(dihidrocapsenona), en plantas WT y transgénicas adultas, medidos al cumplirse
el ciclo de vida de Bemisia tabaci. En hoja local (HL), hoja sistémica (HS) y raíz.
Los asteriscos indican la diferencia estadísticamente significativa (P ≤ 0.05*; P ≤
0.01 **)
0
5
10
15
HL HS RAÍZ
ug C
apsi
diol
/mg
PF
WT
Nt-LePS 08**
B
0
5
10
15
HL HS RAÍZ
ug C
apsi
diol
/mg
PF
WT
Nt-LeSYS 03
**
****
0
5
10
15
HL HS RAÍZug
Cap
sidi
ol/m
g P
F
WT
WT X Nt-LeSYS 03
****
**
A
C

90
inferior. También se observó una disminución significativa en el contenido de NN
en raíces de plantas adultas dañadas mecánicamente (de 0.17 a 0.04 µg/mg PF)
(28 días después de la oviposición). Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
La línea Nt-LeSYS 03 se caracterizó por ser la única línea en la cual se
detectó NN en forma constitutiva en follaje de plantas jóvenes. El daño mecánico
en estas plantas causó una reducción a niveles no detectables (ND) en la hoja
dañada (respuesta local inhibitoria), mientras que en la raíz indujo su síntesis de
novo (de ND a 0.05 µg/mg PF). En la hoja distal adyacente se detectó un
aumento marginal de NN (0.03 a 0.12 µg/mg PF). Un comportamiento muy
similar se observó en follaje de plantas adultas de la línea Nt-LeSYS 03 dañadas
mecánicamente, ya que se volvió a presentar una reducción significativa de NN
en hojas dañadas, mientras que el incremento detectado en hojas distales
superiores (7.6× mayor; de 0.09 a 0.69 µg/mg PF) fue ahora altamente
significativo. Los niveles de NN sintetizados en raíces de plantas jóvenes Nt-
LeSYS 03 dañadas mecánicamente resultaron muy similares a los niveles
constitutivos detectados en las raíces de plantas adultas, los cuales no variaron
significativamente en respuesta al daño mecánico.
El daño mecánico tuvo un efecto sistémico negativo sobre los niveles de
NN en las dos restantes líneas de plantas adultas transformadas (Nt-LePS 08 y
WT × Nt-LeSYS 03), observado en la hoja distal superior y raíz. Aunque en
ambas líneas el daño mecánico causó una disminución de NN a niveles no
detectables, el efecto observado fue particularmente severo en la raíz.
No se detectó NN en plantas de cualquiera de los genotipos analizados
sometidas a herbivoría por M. sexta o infestación por MB (resultados no
mostrados).

91
PLANTA JOVEN PLANTA ADULTA
Figura 38 . Niveles de nornicotina en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas a daño
mecánico (5 día). En hoja local (HL), hoja sistémica (HS), hoja distal vieja (HDV),
hoja distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
Nt-LeSYS 03
0.0
0.1
0.2
HL HS RAÍZug N
orni
cotin
a/m
g P
F
0 día
5 díaP=0.072
**
**
D
WT
0.0
0.1
0.2
0.3
HL HDV HDJ RAÍZug N
orni
cotin
a/m
g P
F
0 h
5 día
***
**
A
WT X Nt-LeSYS 03
0.0
0.1
0.2
0.3
HL HDV HDJ RAÍZug N
orni
cotin
a/m
g P
F
0 h
5 día
** **
B
Nt-LePS 08
0.0
0.1
0.2
0.3
HL HDV HDJ RAÍZug N
orni
cotin
a/m
g P
F
0 h
5 día
** **
C
Nt-LeSYS 03
0.0
0.5
1.0
HL HDV HDJ RAÍZug N
orni
cotin
a/m
g P
F
0 h
5 día
**
*
E

92
6.6. Cuantificación de fenoles totales en plantas j óvenes y/o adultas
6.6.1. Tratamiento: daño mecánico
Los resultados obtenidos se muestran en las Figuras 39 A y B. Los niveles
basales de fenoles totales en plantas WT adultas fueron superiores (entre 1.92 y
7.7× más altos) en las hojas distal inferior y de transición C-F, pero más bajos
(entre 1.7 y 1.1× menores) en raíz, que los observados en tejidos equivalentes
de plantas de las tres líneas transgénicas en estudio (Fig. 39 B)
Asimismo, los niveles basales de fenoles en la raíz de las tres líneas de
plantas transgénicas jóvenes fueron menores a los detectados en plantas WT de
la misma edad, particularmente en la línea WT × Nt-LeSYS 03 en la cual no se
detectaron. En follaje se repitió esta tendencia en las líneas Nt-LePS 08 (1.3 y 2
veces menores en las hojas inferiores y superiores, respectivamente) y WT × Nt-
LeSYS 03 (2.7 y 3.8 veces menores en hojas inferiores y superiores,
respectivamente), mientras que en la línea Nt-LeSYS 03, los niveles de fenoles
en follaje no variaron mucho en relación a los detectados en las plantas WT
(Fig.39 A).
En plantas WT jóvenes dañadas mecánicamente se observaron efectos
sistémicos contrastantes, ya que se detectó un incremento y disminución
significativos en la hoja distal adyacente superior y raíz, respectivamente. En las
plantas transgénicas jóvenes, el daño mecánico no indujo un cambio en los
niveles basales de fenoles en follaje (línea WT × Nt-LeSYS 03) o bien provocó
una reducción significativa de éstos (líneas Nt-LePS 08 y Nt-LeSYS 03). En raíz,
la disminución significativa de fenoles totales observada en plantas WT dañadas,
se repitió en la línea Nt-LePS 08, mientras que en las dos restantes se detectó
un incremento significativo (Figs. 39 C, G).
En plantas WT adultas dañadas mecánicamente, sólo se observó un
aumento significativo en fenoles totales a nivel sistémico en la hoja distal inferior.
En contraste con lo observado en las plantas Nt-LePS 08 jóvenes, el daño
mecánico aplicado a plantas adultas de esta línea indujo un incremento
significativo generalizado en fenoles totales, tanto en follaje como en raíz. En las
líneas Nt-LeSYS 03 y WT × Nt-LeSYS 03, se observó en general lo contrario, ya

93
que los niveles de fenoles totales detectados en la hoja dañada (respuesta local),
hojas distales inferiores y superiores y raíz (respuesta sistémica) resultaron
marginal o significativamente menores a los observados en plantas intactas
(excepto por el incremento significativo detectado en la hoja distal superior en la
línea WT × Nt-LeSYS 03) (Fig. 39 D).
6.6.2. Tratamiento: herbivoría por larvas de M. sexta (plantas adultas)
Los resultados obtenidos se muestran en la Figura 40. La herbivoría por
M. sexta causó, en general, una reducción en los niveles de fenoles totales (en
relación a los niveles basales detectados en plantas intactas de la misma edad)
en todos los tejidos analizados, tanto de plantas WT como transgénicas (excepto
en la línea Nt-LePS 08, en cuyas hojas distales superior e inferior,
respectivamente, se observó un ligero aumento). La disminución fue más
marcada en la hoja dañada, en la hoja distal inferior (es decir en las hojas más
viejas) y en raíz, mientras que en la hoja distal superior los niveles se
mantuvieron iguales o variaron levemente.
El nivel de fenoles detectado en follaje y raíz de las plantas transgénicas
dañadas por M. sexta resultó marginal o significativamente inferior al detectado
en plantas WT, excepto en la raíz de la línea Nt-LeSYS 03, cuyos niveles
resultaron significativamente superiores al de las plantas WT.
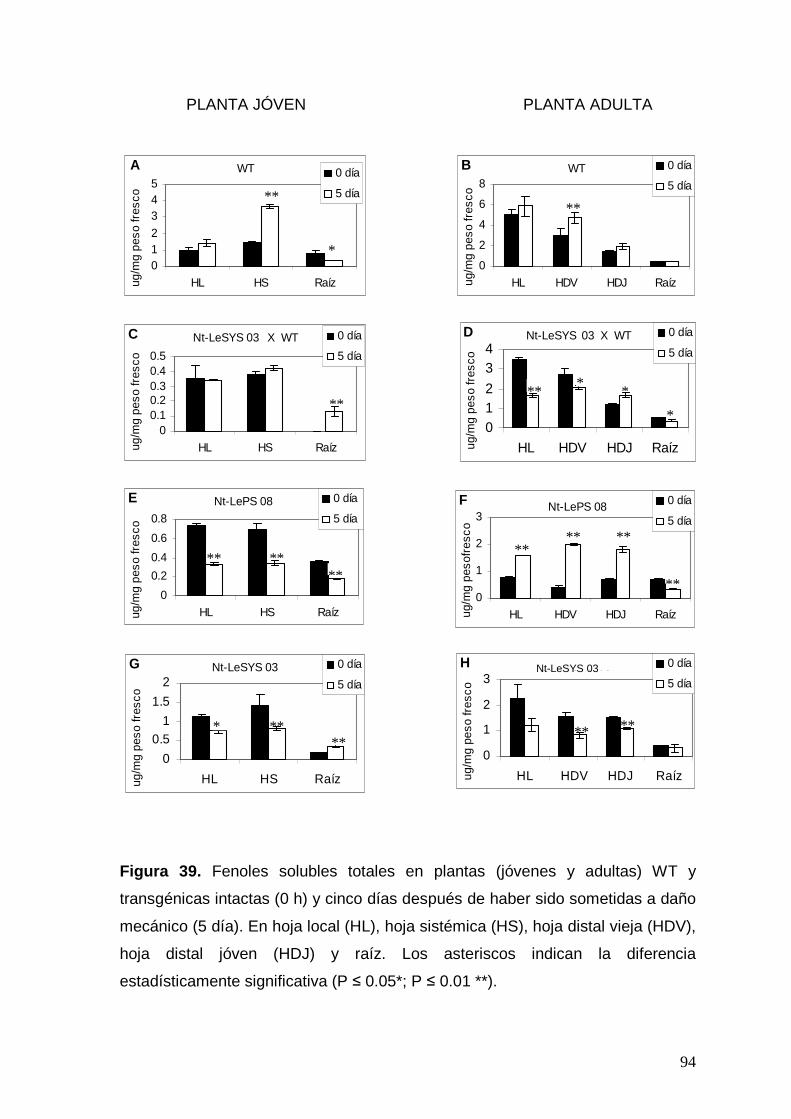
94
PLANTA JÓVEN PLANTA ADULTA
Figura 39. Fenoles solubles totales en plantas (jóvenes y adultas) WT y
transgénicas intactas (0 h) y cinco días después de haber sido sometidas a daño
mecánico (5 día). En hoja local (HL), hoja sistémica (HS), hoja distal vieja (HDV),
hoja distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
WT
012
345
HL HS Raízug/m
g pe
so fr
esco
0 día
5 día**
*
A
Nt-LePS 08
0
0.2
0.4
0.6
0.8
HL HS Raízug/m
g pe
so fr
esco
0 día
5 día
******
E
Nt-LeSYS 03
0
0.51
1.5
2
HL HS Raízug/m
g pe
so fr
esco
0 día
5 día
*** **
G
Nt-LeSYS 03 X WT
00.10.20.30.40.5
HL HS Raízug/m
g pe
so fr
esco
0 día
5 día
**
C
WT
0
2
4
6
8
HL HDV HDJ Raízug/m
g pe
so fr
esco
0 día
5 día
**
B
Nt-LePS 08
0
1
2
3
HL HDV HDJ Raízug/m
g pe
sofr
esco
0 día
5 día
** ** **
**
F
Nt-LeSYS 03 X WT
012
34
HL HDV HDJ Raízug/m
g pe
so fr
esco
0 día
5 día
**
*
**
D
Le-Nt'SYS 03
0
1
2
3
HL HDV HDJ Raízug/m
g pe
so fr
esco
0 día
5 día
** **
H
Nt-LeSYS 03

95
Figura 40. Comparación entre los niveles de fenoles totales solubles en plantas
WT y transgénicas adultas, 5 días después de haber sido expuestas a herbivoría
por larvas de M. sexta durante 24 h. En hoja local (HL), hoja distal vieja (HDV),
hoja distal jóven (HDJ) y raíz. Los asteriscos indican la diferencia
estadísticamente significativa (P ≤ 0.05*; P ≤ 0.01 **).
0
0.4
0.8
1.2
1.6
HL HDV HDJ Raíz
ug/m
g pe
so fr
esco
WT
Nt-LePS 08
** **
B
0
0.4
0.8
1.2
1.6
HL HDV HDJ Raíz
ug/m
g pe
so fr
esco
WT
WT X Nt-LeSYS 03
** **
*
**
A
0
0.4
0.8
1.2
1.6
HL HDV HDJ Raíz
ug/m
g pe
so fr
esco
WT
Nt-LeSYS 03
*
**
C

96
6.6.3. Tratamiento: infestación por mosquita blanca (plantas adultas)
Los resultados obtenidos se muestran en la Figura 41. En plantas WT, la
infestación por B. tabaci causó una disminución en los niveles de fenoles en
follaje, particularmente en la hoja infestada donde los niveles fueron 2.9 veces
menores a los detectados en plantas WT no infestadas de la misma edad. En la
raíz, los niveles de fenoles fueron similares en plantas WT infestadas y no
infestadas.
En las plantas transgénicas, el comportamiento en respuesta a la
infestación por B. tabaci fue variable: mientras que en la línea Nt-LePS 08 se
observó un aumento de fenoles totales tanto local (2× más alto) como sistémico
(3.3× más alto), con respecto a plantas no infestadas de la misma edad, en las
dos restantes la respuesta fue inhibitoria, excepto por el ligero incremento
detectado en la hoja distal superior de la línea WT × Nt-LeSYS 03. El efecto
inhibitorio fue más marcado en la hoja infestada (2.6× menor en ambas líneas).
En raíz de las tres líneas transformadas se detectó una disminución en los
niveles de fenoles con respecto a plantas no infestadas de la misma edad.
Los niveles de fenoles en todos los tejidos analizados de plantas
transgénicas infestadas fueron iguales (Nt-LePS 08) o significativamente
menores (Nt-LeSYS 03 y WT × Nt-LeSYS 03) a los detectados en plantas WT
infestadas. A su vez, el cambio en los niveles de fenoles totales detectado en
plantas transgénicas y WT en respuesta a herbivoría por M. sexta e infestación
por MB fue similar.
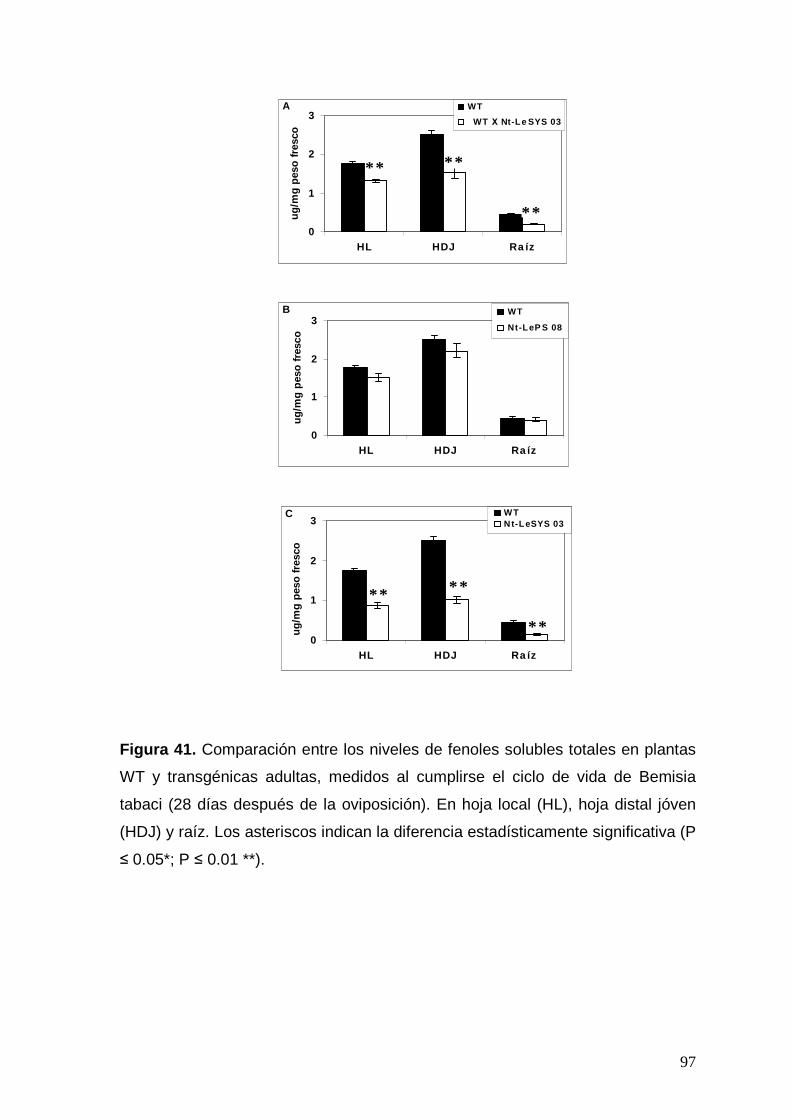
97
Figura 41. Comparación entre los niveles de fenoles solubles totales en plantas
WT y transgénicas adultas, medidos al cumplirse el ciclo de vida de Bemisia
tabaci (28 días después de la oviposición). En hoja local (HL), hoja distal jóven
(HDJ) y raíz. Los asteriscos indican la diferencia estadísticamente significativa (P
≤ 0.05*; P ≤ 0.01 **).
0
1
2
3
HL HDJ Ra íz
ug/m
g pe
so fr
esco
WT
Nt-LeP S 08
B
0
1
2
3
HL HDJ Ra íz
ug/m
g pe
so fr
esco
WT
WT Х Nt-Le SYS 03
** **
**
A
0
1
2
3
HL HDJ Ra íz
ug/m
g pe
so fr
esco
WTNt-LeSYS 03
** **
**
C

98
DISCUSIÓN DE RESULTADOS
Las plantas del género Nicotiana (Solanaceae) son capaces de sintetizar
una gran diversidad de metabolitos secundarios, principalmente alcaloides,
compuestos fenólicos y terpenoides, muchos de los cuales ejercen una influencia
sobre las interacciones biológicas que estas plantas tienen con su ambiente
circundante.
Este trabajo es parte de un estudio cuyo objetivo fue determinar el efecto
que la sobreexpresión de prosistemina (LsPS) y sistemina (LeSYS) de jitomate
en N. tabacum pudiera tener sobre la capacidad de responder a agobios
(a)bióticos. La inquietud de analizar los niveles constitutivos e inducibles de
metabolitos secundarios relacionados con defensa contra herbívoros y/o
patógenos (nicotina, nornicotina, ácido clorogénico, capsidiol y fenoles solubles
totales) en estas plantas surgió de la posibilidad de que la resistencia diferencial
contra herbivoría por larvas de M. sexta, así como las numerosas modificaciones
morfológicas y fisiológicas observadas en plantas transformadas con LePS y/o
LeSYS (Rocha-Granados, 2004 y resultados no publicados) podría deberse a
cambios en la síntesis y/o acumulación de estos metabolitos en follaje y raíz. En
apoyo a esta propuesta está el hecho de que la síntesis de novo de muchos de
estos compuestos, principalmente alcaloides, compuestos fenólicos y
terpenoides, se incrementa dramáticamente como respuesta al daño mecánico o
ataque por herbívoros y/o patógenos, y que este cambio es inducido en muchos
casos por ácido jasmónico, el cual funciona como una señal mediadora de
cambios en el metabolismo secundario en respuesta a muchos tipos de agobio
biótico y abiótico (Gundlach et al., 1992; Hildman et al., 1992; Reinbothe et al.,
1994). Es también bien conocido que la inducción de genes de defensa en
respuesta al daño en L. esculentum, parte de la cual es dependiente de
sistemina, ocurre a través de la intermediación de AJ (Schilmiller y Howe, 2005).
La nicotina es uno de los metabolitos secundarios de defensa más
estudiados. Es el alcaloide predominante en variedades comerciales de N.
tabacum, incluyendo la variedad Xanthi utilizada en esta investigación. La
nicotina constituye en promedio el 94.8% del total de alcaloides (ca. 11.5 mg/g
de peso seco) presentes en follaje de N. tabacum (subgénero Tabacum; sección

99
Genuinae); el resto está constituido por los alcaloides nornicotina (3.0%),
anabasina (0.3%) y anabatina (1.9%) (Saitoh et al., 1985). Se estima que la
nicotina contribuye a la resistencia contra insectos herbívoros gracias a su
capacidad de interactuar con receptores de acetilcolina del sistema nervioso de
animales (Steppuhn et al., 2004; Liu et al., 2005). Asimismo, se ha encontrado
que el daño al follaje, mecánico o por herbívoros, tanto en N. tabacum como en
las especies silvestres N. attenuata, N. sylvestris y otras, induce la síntesis de
novo de nicotina en raíces, la cual es posteriormente reubicada en el follaje (vía
el xilema) para contribuir a la defensa de la planta y reducir la pérdida de área
foliar por herbivoría. El ácido jasmónico, que es producido en las hojas dañadas
y rápidamente transportado a las raíces vía el floema, es un componente
esencial de la señalización entre hoja y raíz que facilita esta respuesta (Zhang y
Baldwin, 1997). De acuerdo a Ohnmeiss y Baldwin (2000), la distribución de la
nicotina proveniente de la raíz en el follaje estará determinada por los postulados
de la teoría de defensa óptima, que predice que los tejidos más valiosos para la
planta, o aquellos con una mayor probabilidad de ser atacados, serán los mejor
defendidos. Éstos son en general los tejidos jóvenes, con altas tasas
fotosintéticas y/o las estructuras reproductivas; sin embargo, se ha encontrado
que el valor del tejido varía de acuerdo a la ontogenia de la planta y puede estar
influido por su nivel nutricional. El ataque por herbívoros o patógenos también
puede alterar la cantidad y la distribución de metabolitos defensivos.
Como se observa en las Figuras 29 A y B, los niveles basales de nicotina
fueron similares en plantas WT jóvenes y adultas. Sin embargo, la acumulación
significativa de nicotina en respuesta a daño mecánico sólo se observó en los
tejidos analizados (predominantemente en la hoja dañada y en la hoja distal
joven) de las plantas adultas; en plantas jóvenes dañadas, se observó, en
contraste, una disminución generalizada en los niveles de nicotina, la cual fue
significativa a nivel sistémico. De acuerdo a la teoría de la defensa óptima,
descrita anteriormente, la diferencia entre plantas jóvenes y adultas podría
explicarse en parte, si se considera que el valor que tienen las hojas expandidas
en una planta joven es similar debido a que la diferencias en el estado de
desarrollo son mínimas, tal como se ha observado en plantas de N. sylvestris en
el estadio de roseta (Baldwin et al., 1997). Otros factores que podrían haber
contribuido a la débil respuesta en plantas jóvenes, en cuanto a la inducción de

100
la acumulación de nicotina en respuesta al daño mecánico, son los siguientes: i)
la alta disponibilidad de recursos en plantas jóvenes, los cuales al no estar
siéndo desviados hacia la generación de estructuras reproductivas o producción
de semillas, permiten la utilización de estrategias menos costosas desde el punto
de vista metabólico que la síntesis de nicotina, como el rebrote de tejidos
después del daño; ii) las plantas jóvenes son incapaces de fijar altas
concentraciones de nicotina; iii) la nicotina no es necesariamente el metabolito
de defensa más importante en esta etapa de desarrollo; iv) el similar valor que el
tejido foliar tiene en plantas jóvenes garantiza una distribución uniforme de
metabolitos de defensa, y/o v) la incapacidad de exportar suficiente JA a la raíz
para inducir la síntesis de nicotina, aún a pesar de que los tejidos jóvenes y
reproductivos son los que producen los mayores niveles de AJ, tanto
constitutivos como inducibles (Creelman y Mullet, 1997;Ohnmeiss y Baldwin,
2000). Esta última posibilidad podría haberse debido al hecho de que las hojas
inmaduras constituyen tejidos consumidores que importan fotosintatos desde las
hojas maduras y viejas, de manera que la exportación de AJ hacia el floema y
eventualmente hacia la raíz, podría estar comprometida en plantas jóvenes en
crecimiento activo y sin un aparato fotosintético eficiente.
En cuanto a las plantas WT adultas dañadas, la máxima acumulación de
nicotina se observó a nivel local y sistémico apical, lo que sugiere que estas
hojas tienen un mayor valor para la planta que la hoja distal basal. De manera
que la acumulación local de nicotina podría ser parte de una estrategia de
defensa para la defensa de tejidos valiosos (e. g., aquellos con las tasas
fotosintéticas más altas por área foliar, tal como se ha reportado para hojas
maduras en N. sylvestris) contra insectos, especialmente de aquellos que tienen
poca movilidad. También se ha especulado que la acumulación de defensas en
tejido dañado puede disuadir a los herbivoros a consumir tejido de menor valor e
inclusive, a migrar hacia plantas vecinas competidoras. En todo caso, los
resultados para nicotina obtenidos en este trabajo contrastaron con los
reportados para N. sylvestris, en las cuales se observó que la sensibilidad al
daño mecánico disminuía gradualmente a medida que las plantas avanzaban en
su estado de desarrollo (Ohnmeiss et al., 1997; van Dam et al. 2001).
En plantas transgénicas jóvenes, exceptuando la línea Nt-LeSYS 03, los
niveles basales de nicotina fueron menores a los detectados en las plantas WT.

101
El efecto negativo que el daño mecánico tuvo sobre el contenido de
nicotina en plantas WT jóvenes se hizo más marcado en la línea Nt-LePS 08
(disminución significativa generalizada), lo cual contrastó con la inducción local
(en Nt-LeSYS 03) y sistémica (en WT × Nt-LeSYS 03) de niveles
significativamente más altos de nicotina. En plantas adultas, el patrón de
acumulación de nicotina como respuesta al daño resultó muy similar entre
plantas WT y transgénicas, aunque estas últimas mostraron una respuesta más
intensa al daño, particularmente la respuesta sistémica en la hoja distal joven en
las líneas Nt-LePS 08 y Nt-LeSYS 03. Estos resultados sugieren que la
expresión de LePS y LeSYS en tabaco alteró el patrón de síntesis y distribución
de nicotina en la planta. Tomando en cuenta que la acumulación de metabolitos
defensivos en tabaco se produce de acuerdo al valor relativo que los tejidos
tienen para la planta atacada, es válido sugerir que el comportamiento
observado fue un indicio de que el valor de estas hojas en las plantas
transgénicas se incrementó. Las razones de este cambio se desconocen, pero
podrían estar relacionados con cambios fisiológicos causados por la
sobreexpresión de estos genes en tabaco, los cuales podrían haber alterado las
relaciones entre tejidos consumidores y tejidos fuente (debido a la síntesis
constitutiva de proteínas y metabolitos de defensa) o bien, podrían haber
alterado el desarrollo normal de N. tabacum. Este último argumento podría
aplicarse a la línea Nt-LeSYS 03, la cual presenta un fenotipo caracterizado por
alteraciones marcadas en su desarrollo, como enanismo, hojas modificadas,
floración precoz y senescencia acelerada (Rocha-Granados, 2004 y resultados
no publicados; ver Fig. 13). En plantas adultas de todos los genotipos analizados
se observó también un incremento significativo de nicotina en raíz en respuesta
al daño, lo que fue un indicio de que la síntesis de novo de AJ en las hojas y su
posterior transporte, vía floema, a la raíz, no se alteró en las plantas
transgénicas. Sin embargo, los niveles de nicotina en raíces de plantas Nt-
LeSYS 03 intactas y dañadas, fueron inferiores a los detectados en plantas WT y
demás plantas transgénicas. Es posible que la floración precoz y la senescencia
acelerada observada en esta línea hayan causado una reducción en la síntesis
de nicotina y/o un aumento en el catabolismo de nicotina como parte de una
estrategia para canalizar el nitrógeno hacia la producción de estructuras/tejidos

102
reproductivos. Esta propuesta se sustenta en cierta medida con lo observado en
plantas de N. attenuata y N. sylvestris, en las cuales se detectó una reducción en
el contenido de nicotina durante las etapas de elongación (generación de
estructuras reproductivas) y floración (Ohnmeiss y Baldwin, 2000; Baldwin,
2001).
La reducción en los niveles de nicotina, con respecto a plantas intactas,
observados en hojas de plantas WT y transgénicas sometidas a herbivoría por
larvas de M. sexta (Figuras 30 A y 30 B) estuvo de acuerdo a varios reportes en
la literatura en los que se ha observado que el daño causado por este insecto
tolerante a la nicotina en especies silvestres de Nicotiana, reprime su síntesis, a
pesar de un marcado aumento en los niveles de AJ, tanto en hoja como en raíz
(McCloud y Baldwin, 1997). Este efecto se ha atribuido a una síntesis de novo
sostenida de etileno, que al igual que en otras especies (e.g., Arabidopsis
thaliana, Griffonia simplicifolia), es antagónico al efecto inductivo local y/o
sistémico de AJ (Zhu-Salzman et al., 1998; Rojo et al., 1999; Kahl et al., 2000).
Además, se ha demostrado que está inhibición se debe a la supresión por etileno
de dos genes (putrescina-N-metil transferasas 1 y 2) que son esenciales para la
síntesis de nicotina en raíz (Winz y Baldwin, 2001). El contraste observado entre
daño mecánico (inducción) y herbivoría (represión), coincidió con estudios
previos que han demostrado que la síntesis de etileno, responsable a su vez de
la represión de la síntesis de nicotina, se produce exclusivamente en respuesta a
un grupo de amidas, sintetizadas a partir de la conjugación de ácidos grasos con
aminoácidos, presentes en las secreciones orales de M. sexta (Halitschke et al.,
2001; Schittko et al., 2001). Además de su efecto sobre la síntesis de nicotina, la
herbivoría por M. sexta también causa una reorganización transcripcional en
plantas de tabaco diferente a la observada en respuesta a daño mecánico, la
cual se caracteriza por la expresión diferencial de más de 500 genes
involucrados en fotosíntesis, transporte de electrones, síntesis y funcionamiento
del citoesqueleto, metabolismo de carbono y nitrógeno, y respuesta al daño o
invasión por patógenos (Hermsmeier et al., 2001).
La represión de la acumulación de nicotina en hojas atacadas por M.
sexta fue, sin embargo, mucho más marcada en las plantas transgénicas que en
las plantas WT. Estos resultados sugieren que la sobreexpresión de LePS y
LeSYS en tabaco potenció el efecto inhibitorio que la herbivoría por M. sexta

103
tiene sobre la síntesis de nicotina. Considerando que los ensayos de daño
mecánico no afectaron la inducción de la acumulación local y sistémica de
nicotina en las plantas transformadas, y que estudios previos han confirmado
que la síntesis de AJ no es afectada negativamente por herbivoría (McCloud y
Baldwin, 1997), es posible que la potenciación del efecto inhibitorio observado en
las plantas transgénicas se haya debido a una sobre-producción de etileno
endógeno en respuesta a las secreciones orales de M. sexta. Esta es una
posibilidad que deberá analizarse con mayor detalle, ya que no hay evidencia de
que la sobreexpresión de prosistemina, por lo menos en L. esculentum,
favorezca la síntesis de etileno en respuesta a herbivoría. El efecto sistémico en
raíz fue aún más marcado, ya que en las plantas transgénicas los niveles
disminuyeron marcadamente (1.67 veces en la línea Nt-LeSYS 03 y hasta
niveles no detectable en las otras dos líneas). Esto contrastó con las plantas WT,
en las cuales se detectó una acumulación de nicotina en raíz de plantas dañadas
por herbivoría que fue 6.8 veces mayor a la observada en plantas WT dañadas
mecánicamente. Este efecto se podría explicar en base a los argumentos de
Baldwin (2001), quién sugirió que la acumulación de nicotina en raíces de
plantas dañadas por herbívoros que consumen follaje constituye una estrategia
benéfica para la planta, ya que al localizar la síntesis y localización de este
metabolito en la raíz, se favorece el empleo de la nicotina para la defensa de
tejidos aéreos durante el subsiguiente período de recuperación, en la cual la
capacidad fotosintética de la planta (y por lo tanto la síntesis de novo de nicotina)
estará seriamente comprometida.
La infestación B. tabaci también tuvo un efecto negativo sobre la
acumulación de nicotina en plantas WT y transgénicas, particularmente a nivel
local. Es posible que el efecto observado haya sido debido, al igual a lo
observado por daño con M. sexta, a la síntesis facilitada de etileno, aunque esto
no ha sido demostrado aún. La única evidencia experimental a favor de esta
posibilidad, es la inducción de proteínas RP en jitomate en respuesta a la
infestación por este insecto (Mayer et al., 1996), las cuales dependen de etileno
para su expresión en patógenos (Kombrink y Somssich, 1997; Ohtsubo et al.,
1999). Sin embargo, parece más probable que la infestación por B. tabaci, que
es un insecto succionador que se alimenta directamente del floema, haya
cambiado la relación tejido fuente-tejido consumidor en los tejidos infestados, de

104
manera que la demanda de fotosintatos en la hoja infestada de las plantas WT
podría haber reducido la fuerza de la raíz como tejido consumidor, similarmente
a lo que ocurre cuando las plantas de tabaco entran en la fase de floración
(Shiroya et al., 1961), causando una disminución en el transporte de JA a la raíz,
con la consecuente supresión de la inducción de la síntesis de nicotina. Esta
posibilidad se apoya en los resultados obtenidos al analizar las raíces de las
plantas infestadas, excepto la línea Nt-LeSYS 03, en las cuales la nicotina se
redujo hasta niveles no detectables. Otra posibilidad es que los niveles de
nicotina regresaron a sus niveles basales, al menos en follaje, debido al largo
tiempo de muestreo, que se hizo 28 días después de haber infestado con
insectos adultos las hojas de plantas que, al inicio del experimento, tenían solo 4
hojas expandidas. De acuerdo a lo anterior, es probable que los bajos niveles de
nicotina detectados en hojas y raíces de plantas WT y transgénicas infestadas
por B. tabaci se hayan debido a una combinación de efectos ontogénicos y
fisiológicos, caracterizados estos últimos por cambios en el estatus fotosintético
del tejido fuente o consumidor de los tejidos analizados. Esta suposición
concuerda en cierto modo con evidencias experimentales previas obtenidas en
Nicotiana que indican que cualquier tratamiento capaz de incrementar la fuerza
de la raíz como tejido consumidor (e. g., fertilización con nitrógeno) facilita el
transporte de AJ y consecuente síntesis de nicotina, desde las hojas dañadas a
la raíz (Ohnmeiss y Baldwin, 2000) y con la noción de que las relaciones tejido
fuente-tejido consumidor ejercen una influencia muy marcada sobre la
resistencia de las plantas a agobios bióticos y abióticos (Coleman 1986;
Coleman y Leonard, 1995).
Después de la nicotina, la nornicotina es el alcaloide más abundante en N.
tabacum (3% y 6% del total de alcaloides en hojas y raíces, respectivamente)
(Saitoh et al., 1985). En ciertas especies como N. glauca, la nornicotina es
acilada mediante la formación de enlaces amida con ácidos grasos de 12 a 16
átomos de carbono en respuesta a daño mecánico y AJ exógeno; esta
transformación la hace más tóxica hacia herbívoros adaptados a la nicotina
como M. sexta (Laue et al., 2000). Los resultados obtenidos en plantas WT, en
las cuales solo se detectó nornicotina en las hojas distales superior e inferior y
en la raíz de plantas adultas intactas, coincidieron en cierto modo con los
reportados en plantas de N. sylvestris (cuyo contenido de nornicotina es superior

105
al reportado en N. tabacum) en las cuales la nornicotina y otros alcaloides
menores fueron cuantitativamente importantes sólo en plantas senescentes
(Baldwin, 1989). En forma similar a la acumulación de derivados acilados de
nornicotina en plantas de N. glauca dañadas y tratadas con AJ, la nornicotina se
acumuló en hojas dañadas y distales superiores de plantas WT. Por su parte, la
acumulación en respuesta al daño en plantas adultas se reprimió completamente
en las líneas Nt-LePS 08 y WT × Nt-LeSYS 03, mientras que en la línea Nt-
LeSYS 03 se observó una inducción sistémica apical relativamente intensa (ver
Figura 30). Además, esta línea fue la única en la que se detectó este alcaloide,
tanto en forma constitutiva como inducible, en hojas y raíces de plantas jóvenes.
Tampoco se detectó nornicotina en todos los genotipos sometidos a herbivoría
por M. sexta o a infestación por B. tabaci, lo que posiblemente fue un reflejo de
los niveles abatidos de nicotina detectados en respuesta a estos tratamientos.
Los resultados sugieren que la expresión de LeSYS en tabaco altera el flujo
biosintético de nitrógeno entre nicotina en raíz y nornicotina en follaje,
especialmente en plantas jóvenes. La presencia de nornicotina en plantas
jóvenes de esta línea podría haber sido consecuencia de la senescencia
acelerada que la caracteriza.
El ácido clorogénico (ACG) es un metabolito presente constitutivamente
en varias especies de plantas, particularmente en solanáceas, que funciona
como agente protector contra desafíos de tipo biótico. Por ejemplo, en plantas de
jitomate y tabaco con altos niveles de este metabolito (en tabaco se acumula en
los tricomas glandulares de la superficie de las hojas), se observó una
progresión más lenta y una reducción en los niveles de infección por
Pseudomonas syringae y patógenos fungosos, respectivamente (Maher et al.,
1994; Shedle et al., 2003). Asimismo, incrementos en el contenido de ácido
clorogénico fueron detectados en hojas de plantas infectadas con virus del
mosaico de tabaco, aunque éstos se asociaron más con la formación de
quinonas y productos poliméricos que con la generación de lesiones necróticas y
acumulación de compuestos fenólicos propias de la reacción hipersensible
(Tanguy y Martin, 1972). Su efecto se atribuye, al menos en parte, a la actividad
antioxidante de este metabolito. Esta cualidad favorece su oxidación, después
del daño causado por la infección microbiana o por insectos, por la enzima

106
polifenol oxidasa para generar quinonas reactivas, que tienen la propiedad de
limitar el crecimiento de microorganismos invasores y/o el daño tisular gracias a
su capacidad de formar entrecruzamientos con componentes de la pared celular
(Rober, 1989; Keinänen et al., 2001).
Los niveles basales de ACG resultaron mucho menor en hojas de plantas
jóvenes que en las de plantas adultas. Este resultado posiblemente fue un reflejo
del control estricto que sobre la biosíntesis de fenilpropanoides se presenta
durante el desarrollo de la planta (Bate et al., 1994; Whetten y Sederoff, 1995).
Al parecer, el nivel de estos metabolitos en hoja depende de un balance entre la
actividad de 3-desoxi-D-arabino-heptulosonato-7-fosfato-sintasa, DAHPS (ruta
del ácido shiquímico) y fenilalanina amonio liasa, FAL (ruta de los
fenilpropanoides de donde se produce finalmente el ACG). La actividad de FAL
también depende de señales (a)bióticas derivadas del medio ambiente, como
aquellas que se producen en respuesta al daño, lo que coincidió con la
acumulación sistémica y local de ACG (en plantas WT adultas). A su vez, la
expresión de LePS y de LeSYS en plantas transgénicas jóvenes causó una
disminución generalizada, en relación a las plantas WT, del contenido de ACG
en plantas intactas y dañadas. En plantas transgénicas adultas de la línea WT ×
Nt-LeSYS 03 y Nt-LeSYS 03 también se observó una marcada disminución en
los niveles de ACG locales y sistémicos de plantas dañadas, particularmente en
ésta última. La línea Nt-LePS 08 mostró un efecto altamente contrastante
caracterizado por niveles muy bajos de ACG en la hoja de transición tejido
fuente-consumidor, pero muy elevados en la hoja distal superior de plantas
intactas, situación que se invirtió en los mismos tejidos de plantas dañadas. A su
vez, los niveles de ACG disminuyeron drásticamente en plantas WT (reducción a
niveles no detectables en la respuesta sistémica en hoja y raíz) y transgénicas
(reducción generalizada a niveles no detectables en todos los tejidos analizados)
sometidas a herbivoría por M. sexta (Figs. 33 A, B y C). La infestación por B.
tabaci también provocó una disminución en los niveles de ACG, pero ésta fue de
mucho menor intensidad, excepto en hojas de la línea Nt-SYS 03, donde el ACG
se mantuvo por debajo de los límites de detección (ver Figura 34 B). Estos
resultados indican que al igual que en el caso de la nicotina, la herbivoría por
insectos, particularmente M. sexta, redujo los niveles de ACG en plantas WT y

107
que este efecto negativo se potenció en las plantas transgénicas, observándose
inclusive en plantas dañadas mecánicamente. Esto sugiere que el metabolismo
de fenilpropanoides se alteró, de un modo aún no determinado, por la expresión
de LePS y LeSYS en N. tabacum.
Sin embargo, los resultados obtenidos en plantas de la línea Nt-LeSYS
03, en las cuales se detectaron en general niveles significativamente reducidos
de ACG podrían ofrecer una posible pista sobre el mecanismo mediante la
expresión de LePS y LeSYS pues podrían estar afectando la ruta de los
fenilpropanoides, ya que el crecimiento y desarrollo anormal que se presenta en
estas plantas caracterizado por enanismo de las plantas, floración precoz,
morfología alterada de las hojas (lanceoladas, con bordes irregulares y lesiones
necróticas constitutivas) y flores (más pequeñas, con un desarrollo incompleto de
los estambres), lo que se reflejó en una producción de semillas reducida con
muy baja eficiencia de germinación (Rocha-Granados, 2004) se asemeja al
observado en plantas de tabaco en las cuales se encuentra alterado el
metabolismo de fenilpropanoides. Con estos antecedentes se podrían sugerir los
siguientes escenarios para explicar la reducción de ACG (y de fenoles totales) en
plantas de tabaco sobreexpresantes de LeSYS y LePS: i) alteración del balance
de aminoácidos aromáticos, tal como se observó en plantas transgénicas de
tabaco en las cuales la sobreexpresión de triptófano y tirosina descarboxilasas
resultó, curiosamente, en una reducción en los niveles de triptófano y tirosina.
Esto causó alteraciones fisiológicas y bioquímicas en las plantas transgénicas
debido a un giro compensatorio en el flujo de la ruta del ácido shiquímico para
favorecer la síntesis de estos dos aminoácidos y disminuir la de compuestos
fenilpropanoides; tales cambios se asociaron a la detección de niveles
anormales de metabolitos secundarios como nicotina y ácido clorogénico (Guillet
et al., 2000); ii) modificación del equilibrio entre citocininas y auxinas, similar a la
observada en plántulas de tabaco transformadas con variantes del gen 6b de
Agrobacterium tumefaciens, que se considera como un gen accesorio que
modula la acción de estas hormonas. Este cambio se atribuyó a una reducción
en el catabolismo de auxinas debido al efecto protector asociado con la
acumulación temprana de los fenilpropanoides escopolina y ACG (Gális et al.,
2002, 2004), y/o iii) alteración en la actividad de la enzima fenilalanina amonio
liasa (FAL), que es la enzima clave responsable de catalizar la reacción limitante

108
en la síntesis de ACG, que junto con la rutina, son los compuestos fenólicos
extraíbles más abundantes en hoja de tabaco. Esta última posibilidad se apoya
en el hecho de que plantas de tabaco en las cuales la expresión de FAL
endógeno se suprimió por la sobreexpresión de dos genes FAL exógenos,
provenientes de fríjol, presentaron niveles muy bajos de fenilpropanoides así
como un fenotipo caracterizado por alteraciones morfológicas severas como
enanismo, curvatura y cambio en la textura de las hojas, morfología y
pigmentación alterada de las flores, reducción de la viabilidad del polen, lesiones
necróticas y fluorescentes en hojas y lignificación reducida, particularmente en el
xilema (Elkind et al., 1990).
Una forma relativamente sencilla de comprobar que existe una relación
entre la reducción de ACG, un mayor catabolismo de auxinas y una consecuente
inhibición del crecimiento por citocininas en plantas Nt-LeSYS 03, podría hacerse
mediante la adición de niveles bajos (0.3 µM) de auxinas sintéticas exógenas
(ej., ácido naftalenacético) al medio de cultivo usado para la regeneración in vitro
de estas plantas, tal como fue demostrado por Gális et al., (2002).
Por lo menos siete sesquiterpenos bicíclicos (capsidiol, risitina, lubimina,
solavetivona, fituberina, fituberol y debneiol) se acumulan en respuesta a estrés
en N. tabacum (Guedes et al., 1982; Threlfall y Whitehead, 1988). De éstos, el
capsidiol es el más abundante, no sólo en N. tabacum sino en otras plantas
solanáceas como Capsicum annuum, Solanum tuberosum y Datura spp.
(Guedes et al., 1982). Usualmente, el capsidiol se encuentra ausente en tejidos
sanos y se sintetiza rápidamente en N. tabacum; la acumulación de esta
fitoalexina se detecta entre 12 y 24 h después de la infección de plantas intactas
por hongos, oomicetos, bacterias y virus o de la inducción de suspensiones
celulares con evocadores bíoticos (esporas fungosas, fragmentos de paredes
celulares de hongos, glicoproteínas derivadas de oomicetos como criptogeína,
celulasa, etc.) o abióticos (e. g., iones metálicos) (Threlfall y Whitehead, 1988;
Vögeli y Chappel, 1988, 1990; Yin et al., 1997; Keller et al., 1998). Sin embargo,
la acumulación de capsidiol en cultivos celulares inducidos de tabaco, papa y
chile es relativamente fugaz, ya que éste tiende a desaparecer con la misma
velocidad con la cual se acumuló, mediante un proceso catabólico que no ha
sido plenamente identificado (Threlfall y Whitehead, 1988). En contraste, la

109
síntesis de capsidiol está bien definida. Este metabolito, al igual que todas las
fitoalexinas sesquiterpénicas, se sintetiza a través de la ruta biosintética de los
isoprenoides. Su síntesis de novo es precedida por la activación de la enzima
clave 3-hidroxi-3-metilglutaril CoA reductasa (HMGR), de cuya reacción se
derivará eventualmente el farnesil pirofosfato (FPP). El FPP es el último
compuesto en común por el que compiten las rutas biosintéticas de los esteroles
y sesquiterpenos, respectivamente. En esta última, los esqueletos carbonados
son dirigidos hacia la síntesis de capsidiol y otras fitoalexinas similares mediante
una ruta cuya reacción determinante, catalizada por la enzima 5-epi-
aristoloqueno sintasa (EAS), es la conversión de FPP a 5-epi-aristoloqueno
(Dudley et al., 1986; Croteau et al., 1987; Threlfall y Whitehead, 1988; Bohlmann
et al., 2002).
En este estudio no se pudo detectar capsidiol sino un producto de
degradación (Ramírez E, comunicación personal) que bien podría ser similar a
aquellos reportados por otros investigadores, como la capsenona o la cis-9,10-
dihidrocapsenona (Zacharius et al., 1985; Whitehead et al., 1987). Es probable
que la ausencia de capsidiol en las muestras analizadas tomadas de las plantas
dañadas mecánicamente o por insectos se haya debido a los tiempos de
muestreo, los cuales rebasaron ampliamente la vida media del capsidiol (12-24
h). Resulta, sin embargo, difícil de explicar la detección de catabolitos del
capsidiol en tejidos de plantas intactas donde, debido a la naturaleza
estrictamente inducible de este metabolito, no debería estar presente (pero ver
los argumentos presentados más adelante).
Posiblemente, y tal como se ha observado con otros metabolitos
secundarios de N. tabacum analizados en este trabajo, la acumulación
constitutiva del capsidiol, la cual se asume por la presencia de su catabolito,
pudiera estar regulada también por la ontogenia de la planta, ya que los niveles
constitutivos de esta fitoalexina detectados tanto en hoja como en raíz, fueron de
9 a 30 veces más altos en plantas jóvenes. El efecto sobre los niveles de
capsidiol causado por daño mecánico, que se sabe induce levemente la
expresión de EAS (Yin et al., 1997), también varió con la edad de las plantas,
siendo inductivo en plantas adultas, e inhibitorio en plantas jóvenes (Fig. 35 A y
B). La detección de bajos niveles de dihicrocapsenona en plantas intactas,
aunque inesperada, tiene relación con resultados previos en los cuales se

110
detectó una expresión basal del gen GUS en hojas, tallos y raíces de un número
de plantas transgénicas de N. tabacum transformadas con el promotor de EAS4,
uno de las 12 a 15 copias del gen de esta sesquiterpeno ciclasa que se han
detectado en el genoma de N. tabacum (Fachini y Chappel, 1992). De manera
que la presencia de múltiples formas del gen EAS en tabaco sugiere que, en
esta especie, cada uno de los genes podría ser regulado independientemente, y
en forma diferencial, durante el desarrollo o en respuesta a diferentes estímulos
ambientales (Yin et al., 1997), lo que podría explicar los resultados contrastantes
entre plantas jóvenes y adultas obtenidos tanto con plantas intactas como
dañadas. En las raíces de plantas de N. tabacum intactas también se detectó
capsidiol cuantificado en forma directa como dihicrocapsenona cuyos niveles, al
igual que en follaje, fueron superiores en plantas jóvenes. Este resultado
coincidió con lo encontrado en raíces de plantas de tabaco silvestres (N.
attenuata y N. sylvestris), donde se detectó la expresión constitutiva del gen
EAS, a la cual se le atribuyó una leve acumulación de capsidiol (10 ng/g PF) en
raíces de plantas intactas (Bohlmann et al., 2002). De acuerdo a estos
investigadores, este hallazgo sugiere que el capsidiol no es solamente un
metabolito defensivo cuya acumulación se induce en respuesta a patógenos,
sino que constituye una defensa de tipo constitutivo que se expresa en forma
tejido específica en algunas especies de Nicotiana para impedir la invasión de
patógenos en la rizósfera, cuyas células epidérmicas, las cuales carecen de una
capa de cutícula protectora, son particularmente vulnerables debido a la abrasión
a la que continuamente están expuestas.
La sobreexpresión en tabaco de LePS y LeSYS redujo, en general, los
niveles constitutivos de capsidiol (dihidrocapsenona), en las plantas
transformadas, que en algunos casos (e. g., hojas de plantas jóvenes de la línea
Nt-LeSYS 03) estuvieron por debajo del límite de detección. El daño mecánico
tuvo un ligero efecto inductivo en plantas transgénicas jóvenes, que contrastó
con lo observado en plantas WT, mientras que en plantas transgénicas adultas
(excepto en la línea Nt-LeSYS 03) el daño mecánico causó una acumulación
extraordinaria de dihidrocapsenona, particularmente en plantas Nt-LePS 08 (Fig.
35 F). Estos resultados sugieren fuertemente que los mecanismos de regulación
que controlan la síntesis de este metabolito en N. tabacum se alteraron de tal
forma que permitieron la sobre acumulación de dihidrocapsenona en respuesta a

111
daño mecánico. Un posible blanco podría ser el gen EAS, cuya expresión está
estrechamente relacionada con la acumulación de capsidiol y otras fitoalexinas
sesquiterpénicas en N. tabacum. De modo que, al igual a lo que se ha observado
en la inducción del gen FAL (Kuhn et al., 1984; Bolwell et al., 1985; Lawton y
Lamb, 1987; Lois et al., 1989), que como se ha mencionado anteriormente es
una de las enzimas claves que regulan la ruta biosintética de los
fenilpropanoides, la síntesis elevada y estable de capsidiol en la plantas Nt-LePS
08 y WT × Nt-LeSYS 03 podría haberse debido a un incremento en: i) la tasa de
traducción de los ARNm que codifican para las diferentes formas del gen EAS y
la subsiguiente síntesis de novo de la enzima EAS; ii) la estabilidad de los
transcritos de EAS; y/o, iii) la velocidad de transcripción del gen EAS. Esta
hipótesis se apoya en los hallazgos de un estudio realizado por Vögeli y
Chappell (1990), quienes demostraron que la regulación de la actividad de la
enzima sesquiterpeno ciclasa en N. tabacum está mediada por un control, a nivel
transcripcional, de la expresión del gen EAS. Sin embargo, no puede descartarse
una posible contribución de otros genes reguladores de la síntesis de
isoprenoides, como el HMGR. Estas son posibilidades que quedan sujetas a
posterior investigación para ser verificadas o desmentidas.
La acumulación de capsidiol (dihidrocapsenona) en plantas WT sometidas a
herbivoría por larvas de M. sexta fue ligeramente superior a la detectada en
plantas dañadas mecánicamente (Figs. 35 y 36). Un efecto similar se observó en
plantas Nt-LeSYS 03 (Fig. 36 C), mientras que en las líneas Nt-LePS 08 y WT ×
Nt-LeSYS 03, la herbivoría fue claramente incapaz de inducir la acumulación de
los mismos niveles de capsidiol observados en plantas dañadas mecánicamente,
lo que sugiere que cualquiera que haya sido el factor generado por daño
mecánico que disparó la síntesis de capsidiol en estas plantas transgénicas, fue
neutralizado o suprimido por componentes presentes en las secreciones orales
de M. sexta, tal como se discutió anteriormente en el caso de la nicotina. A pesar
de ello, la leve inducción de capsidiol coincidió con los resultados reportados por
Bohlmann et al., (2002), quienes detectaron la acumulación de transcritos de
EAS y de capsidiol, aunque en niveles inferiores a los reportados en
suspensiones celulares de N. tabacum (Vögeli y Chappell, 1988), en tejido aéreo
de plantas de N. attenuata y N. sylvestris dañadas por larvas de M. sexta. Por el

112
contrario, se detectó una fuerte acumulación de capsidiol (dihidrocapsenona),
particularmente a nivel local (excepto en la línea Nt-LeSYS 03), en plantas
infestadas por mosquita blanca (Fig. 37 A a 37 C). Los altos niveles de capsidiol
detectados en estas plantas coinciden con la propuesta de que los insectos
chupadores que se alimentan directamente del floema son percibidos por la
planta como patógenos, debido a las similitudes que existen entre la manera en
que las hifas fungosas y el estilete de los insectos chupadores penetran la planta
y en menor medida, entre el tipo de enzimas hidrolíticas que se liberan durante la
invasión por hongos patógenos o alimentación de los insectos (Fidantsef et al.,
1999; Walling, 2000). La expresión de respuestas defensivas similares a las
inducidas por patógenos en plantas infestadas por insectos chupadores también
se ha atribuido a los virus y/o microorganismos, endo simbiontes que ellos
frecuentemente se encuentran (McKenzie et al. 2002). Es probable que la
inducción de capsidiol (dihidrocapsenona), por mosquita blanca detectada en las
plantas estudiadas, al igual a lo observado en plantas transgénicas de N.
tabacum transformadas con el promotor EAS4 fusionado al gen GUS (Yin et al.,
1997), haya sido independiente de ácido salicílico, ácido jasmónico y H2O2. Esta
propuesta se apoya, en cierta medida, en los resultados reportados por van de
Ven et al., (2000), quienes encontraron que la expresión del gen SLW3, inducida
en plantas de calabaza infestadas por B. argentifolii, fue independiente de estas
moléculas de señalización.

113
8. CONCLUSIONES
1. La extracción simple con metanol acético acuoso, resultó ser eficiente y
confiable para la obtención de metabolitos secundarios de interés (alcaloides,
fenoles y terpenos) con varias propiedades químicas, obviando la necesidad de
procesos de extracción más elaborados.
2. La técnica análitica por GC/MS en la cuantificación e identificación de
metabolitos de interés (nicotina, ác. clorogénico, capsidiol y nornicotina) no
modificados por la inserción de cadenas laterales (glicosilación, acetilación,
mutilación, etc.), resultó más confiable y precisa que HPLC.
3. La cantidad y distribución de metabolitos defensivos varió de acuerdo a la
ontogenia y tipo de desarrollo de la planta en Nicotina tabacum (WT y
transgénica), así como también por el tipo de tratamiento y tejido analizado.
4. La cantidad y distribución de metabolitos defensivos se modificó por la
expresión de LePS y LeSYS en N. tabacum, lo que podría tener relación con los
diferentes grados de resistencia contra herbivoría detectados entre plantas WT,
Nt-LePS y Nt-LeSYS y con el fenotipo anómalo de plantas Nt-LeSYS 03.

114
9. PERSPECTIVAS
Sería conveniente llevar a cabo un análisis tipo Tail-PCR para detectar
secuencias aledañas de la inserción del T-ADN en la línea Nt-LeSYS 03, con el
propósito de descartar la posibilidad de que el fenotipo alterado no se debió a la
interrupción de algún gen involucrado en procesos se desarrollo y crecimiento
de la planta, causante del fenotipo anormal.
Sabiéndo que el ácido jasmónico (AJ) induce la acumulación de nicotina
en plantas de N. attenuata, el cual se incrementa en las hojas dañadas y
rápidamente se transporta a las raíces vía floema, y que es un componente
esencial de la señalización entre hoja y raíz, sería interesante determinar los
niveles de AJ en plantas transgénicas de Nt-LePS y Nt-LeSYS en distintas
etapas de desarrollo para corroborar si existe una relación entre los niveles de
AJ in planta y los resultados obtenidos por daño mecánico y herviboría.
La inhibición de la síntesis de nicotina en hojas atacadas por M. sexta fue
mucho más marcada en las plantas transgénicas que en las plantas WT. Se
sugiere que la sobreexpresión de LePS y LeSYS en tabaco potencia el efecto
inhibitorio que la herbivoría por M. sexta tiene sobre la síntesis de nicotina,
posiblemene por una acumulación de etileno endógeno. De modo que sería
conveniente analizar esta posibilidad, pues no se conoce que la expresión de
sistemina y/o prosistemina incremente los niveles de Et en respuesta a
herbivoría por Manduca sexta.
Sería interesante cuantificar los niveles de nicotina el día de máxima
acumulación (5o día), en plantas infestadas con Bemisia tabaci, pues se sabe
que en plantas de Nicotiana los niveles de este metabolito se incrementan en
repuesta a daño, y verificar si la inducción de nicotina se reprime en este lapso al
igual como ocurre con herbivoría por Manduca sexta.
Sería interesante comprobar si existe algún desbalance o reducción de
algún componente de la ruta de los fenilpropanoides como FAL en la planta Nt-
LeSYS, pues su fenotipo anormal se asemeja al observado en plantas de tabaco
en las cuales se encuentra alterado el metabolismo de fenilpropanoides y si

115
existe una relación entre la reducción de ACG observado con un mayor
catabolismo de auxinas y una consecuente inhibición del crecimiento por
citocininas, debido a la expresión de LeSYS.
Considerando que el espectro de masas de la mayoría de los picos
obtenidos por GC/MS produjo señales que indicaban la presencia de
modificaciones por oligosacáridos y otros compuestos orgánicos, sería
conveniente realizar un análisis de los extractos después de su hidrólisis ácida
para tener una idea del grado de modificación que pudieran tener los metabolitos
secundarios de interés en tabaco. Resultaría interesante determinar si el grado
de modificación cambia en respuesta al daño mecánico y herbivoría, y si estos
cambios tienen relación con su actividad biológica.

116
10. BIBLIOGRAFÍA
Aerts RJ, Gisi D, De Carolis E, De Luca V, Baumann TW (1994) Methyl jasmonate vapor increases the developmentally controlled synthesis of alkaloids in Cantharanthus and Cinchona seedlings. Plant J 5: 635-643 Agrawal AA, Tuzun S Bent E (1999) Induced Plant Defenses Against Pathogens and Herbivores. Biochemistry, Ecology and Agriculture. St. Paul MN: American Phytopathological Society Apostol I, Heistein PF, Low PS (1989) Rapid stimulation of an oxidative burst during elicitation of cultured plant cell. Plant Physiol 90: 109-116 Baldwin IT (1989) The mechanism of damage-induced alkaloids in wild tobacco. J Chem Ecol 15: 1661-1680 Baldwin IT, Saedler R (2004) Virus-induced gene silencing of jasmonate-induced direct defences, nicotine and trypsin proteinase-inhibitors in Nicotiana attenuata. J Experimental Botany 55: 151-157 Baldwin IT, Schmelz EA, Ohnmeiss TE (1994) Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris. J Chem Ecol 20: 2139-2157 Baldwin IT, Zhang Z, Diab N, Ohnmeiss TE, McCloud E S, Lynds GY, Schemelz EA (1997) Quantification, correlations and manipulation of wound-induced changes in jasmonic acid and nicotine in Nicotiana sylvestris. Planta 201: 387-404 Baldwin IT, Gorham D, Schmelz EA, Lewandowski C, Ly nds GY (1998) Allocation of nitrogen to an inducible defense and seed production in Nicotiana attenuata. Oecologia 115: 541-552 Baldwin IT (1999) Inducible nicotine production in native Nicotiana as an example of adaptive phenotypic plasticity. J Chem Ecol 25: 3-30 Baldwin IT (2001) An ecologically motivated analysis of plant-herbivore interactions in native tobacco. Plant Physiol 127: 1449-1458 Bate Nj, Weiting JO, Meromi A, Meromi N-H, Doerner PW, Dixon RA, Lamb CJ, Elkind J (1994) Quantitative relationship between phenylalanine ammonia-lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate-determining step in natural product synthesis. Proc Natl Acad Sci USA 91: 7608-7612

117
Bergey DR, Howe GA, Ryan CA (1996) Polipeptide signaling for plant defensive genes exhibits analogies to defense signaling in animals. Proc Natl Acad Sci USA 93: 12053-12058 Birch PRJ, Avrova AO, Duncan JM, Lyon GD, Toth RL (1999) Isolation of potato genes that are induced during an early stage of the hypersensitive response to Phytophthora infestans. Mol Plant-Microbe Interactions 12: 356-361 Bohlmann J, Stauber EJ, Krock B, Oldham NJ, Gershen zon J, Baldwin IT (2002) Gene expression of 5-epi-aristolochene synthase and formation of capsidiol in roots of Nicotiana attenuata and N. sylvestris. Phytochemistry 60: 109-116 Bolwell GP, Bell JN, Cramer CL, Schuch W, Lamb CJ, Dixon RA (1985) L-Phenylalanine ammonia-lyase from Phaseolus vulgaris: characterization and differential induction of multiple forms from elicitor-treated cell suspension cultures. Eur J Biochem 149: 411-419 Botte M, Mabon F, LeMouillour M, Robins RJ (1997) Biosynthesis of nornicotine in root cultures of Nicotiana alata does not involve oxidation at C-5′ of nicotine. Phytochemistry 46: 117-122 Bowles, D. J. (1990) Defense-related proteins in higher plants. Annu Rev Biochem 59: 873-907 Brooks JW, Watson DG, Freer IM (1986) Elicitation of capsidiol accumulation in suspended callus cultures of Capsicum annuum. Phytochemistry 25: 1089-1092 Buchanan B, Gruissem W, Jones R (2000) Biochemistry & Molecular Biology of Plants. American Society of Plant Physiologists pp 1250-1311 Coleman JS (1986) Leaf development and leaf stress: increased susceptibility associated with sink-source transition. Tree Physiol 2: 289-299 Coleman JS, Leonard AS (1995) Why it matters where on a leaf a folivore feeds. Oecologia 101: 324-328 Creelman RA, Mullet JE (1997) Biosynthesis and action of jasmonates in plants. Annu Rev Plant Physiol Plant Mol Biol 48, 353-381 Croteau R, Gurkewitz MA, Johnson MA, Fish HJ (1987) Biochemistry of oleoresins, monoterpene and diterpene biosynthesis in lodgepole pine saplings infected with Ceratocystis clavigera or treated with carbohydrate elicitors. Plant Physiol 85: 1123-1128 Chappell J (1995) Biochemistry and molecular biology of the isoprenoid biosynthetic phathway in plants. Annu Rev Plant Mol Biol 46: 521-547 Corcuera LJ (1993) Biochemical basis for the resistance of barley to aphids. Phytochemistry 33: 741-747

118
Croft KPC, Voisey CR, Slusarenko AJ (1993) Volatile products of the lipooxigenase pathway evolved from Phaseolus vulgaris (L.) leaves inoculated with Pseudomonas syringae pv phaseolicola. Plant Physiol 101: 13-24 de Bruxelles GL, Roberts MR (2001) Signals regulating multiple responses to wounding and herbivores. Crit Rev Plant Sci 20: 487-521 Delaney TP, Ukness S, Vernooij B, Friedrich L, N, W ayman NK, Negrootto D, Gaffney T, Gutrella M, Kessman H, Ward E, Ryals J (1994) A central role of salicylic acid in plant disease resistance. Science 266: 1247-1250 Délano-Frier JP, Dowbrowski JE, Ryan CA (1999) The expression of tomato prosystemin in E. coli: A structural challenge. Protein Expression Purif 17: 74-82 Dempsey DMA, Shaun M, Klessig DF (1999) Salicylic acid and disease reistance in plants. Crit Rev Plant Sci 18: 547-575 Dicke M (1994) Local and systemic production of volatile herbivore-induced terpenoids: their role in plant-carnivore mutualism. J Plant Physiol 143: 465-472 Dietrich A, Mayer JE, Hahlbrock K (1990) Fungal elicitor trigger rapid, transient and specific protein phosphorylation in parsley cell suspension cultures. J Biol Chem 265: 6360-6368 Doares SH, Styrovets T, Weiler EW, Ryan CA (1995) Oligogalacturonides and chitosan activate plant defensive genes thrhough the octadecanoid pathway. Proc Natl Acad Sci USA 92: 4095-4098 Dombrowski JE, Pearce G, Ryan CA (1999) Proteinase inhibitor-inducing activity of the prohormone prosystemin resides exclusively in the C-terminal systemin domain. Proc Natl Acad Sci USA 96: 12947-12952 Dong X (1998) SA, JA, ethylene and disease resistance in plants. Curr Opin Plant Biol 1: 316-323 Dudley MW, Dueber R, MT, West CA (1986) Biosynthesis of the macrocyclic diterpene casbene in castor bean (Ricinus comunis L.) seedlings, changes in enzyme levels induced by fungal infection, and intracellular localization of the pathway. Plant Physiol 81: 335-342 Ebel J, Cosio EG (1994) Elicitors of plant defense responses. Int Rev Cytol 148: 1-36 Elkind, Y., Edwards, R., Mavanad, M., Hedrick, S.A. , Ribak, O., Dixon, R.A., Lamb, C.J. (1990) Abnormal plant development and down-regulation of phenylpropanoid biosynthesis in transgenic tobacco containing an heterologous phenylalanine ammonia-lyase gene. Proc Natl Acad Sci USA 87: 9057-9061

119
Fachini PJ, Chappel J (1992) Gene family for an elicitor-induced sesquiterpene cyclase in tobacco. Proc Natl Acad Sci USA 89: 11088-11092 Farmer EE, Ryan CA (1990) Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc Natl Acad Sci USA 87: 7713-7716 Farmer EE, Ryan CA (1992) Octadecanoid precursors of jasmonic acid activate the synthesis of wound-inducible proteinase inhibitors. Plant Cell 4: 129-134 Ferris PJ, Woessner JP, Waffenschmidt S, Kilz S, Dr ees J, Goodenough UW (2001) Glycosylated polyproline II rods with kinks as a structural motif in plant hydroxyproline-rich glycoproteins. Biochemistry 40: 2978-2987 Fidantsef, A.L., Stout, M.J., Thaler, J.S., Duffey, S.S., Bostock, R.M. (1999) Signal interactions in pathogen and insect attack: expression of lipoxygenase, proteinase inhibitor II, and pathogenesis-related protein P4 in the tomato Lycopersicon esculentum. Physiol Mol Plant Pathol 54: 97-114 Fiehn O, Kopka J, Dörman P, Altmann T, Trethewey RN , Willmitzer L (2000) Metabolite profiling for plant functional genomics. Nature Biotech. 18: 1157-1161 Freytag S, Arabatzis N, Hahlbrock K, Scmelzaer E (1994) Reversible cytoplasmic rearrangements precede wall aposition, hypersensitive cell death and defense-related gene activation in potato/Phytophthora infestans interactions. Planta 194: 123-135 Friedrich L, Lawton K, Ruess W, Masner P, Specker N, Rella MG, Meier B, Dichner S, Staub T, Uknes S, Metraux JP, Kessmann H , Ryals J (1996) A benzothiadiazole derivate induces systemic acquired resistance in tobacco. Plant J 10: 61-70 Gaffney T, Friedrich L,Vernooij B, Negrotto D, Nye G, Uknes, S, Ward E, Kessmann H, Ryals J (1993) Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261: 754-756 Gális I, Šimek P, Van Onckelen HA, Kakiuchi Y, Wabi ko H (2002) Resistance of transgenic tobacco seedlings expressing Agrobacterium tumefaciens C58-6b gene, to growth-inhibitory levels of cytokinin is associated with elevated IAA levels and activation of phenylpropanoid metabolism. Plant Cell Physiol 43: 939-950 Gális I, Kakiuchi Y, Šimek P, Wabiko H (2004) Agrobacterium tumefaciens Ak-6b gene modulates phenolic compound metabolism in tobacco. Phytochemistry 65: 169-179 Gatehouse JA (2002). Plant resistance towards insect herbivores: a dynamic interaction. New Phytol. 156: 145-169 Genoud T, Métraux JP (1999) Cross-talk in plant cell signaling: structure and function of the genetic network. Trends Plant Sci 4: 503-507

120
Geoffroy P, Legrand M, Fritig B (1990) Isolation and characterization of a proteinaceous inhibitor of microbial proteinases induced during the hypersensitive reaction of tobacco to tobacco mosaic virus. Mol Plant-Microbe Interactions 3: 327-333 Gershenzon J, Kreis W (1999) Biochemistry of terpenoids: monoterpenes, sesquiterpenes, diterpenes, sterols, cardiac glycosides and steroid saponins En M Wink (ed), Biochemistry of Plant Secondary Metabolism, Annual Plant Reviews, vol. 2, CRC Press LLC. pp. 222-299 Glawe GA, Zavala JA, Kessler A, van Dam N, Baldwin IT (2003) Ecological costs and benefits correlated with trypsin protease inhibitor production in Nicotiana attenuata. Ecology 84: 79-90 Goodspeed TH (1954) The Genus Nicotiana. Chronica Botanica, Waltham, MA. Gómez-Gómez L, Boller T (2002) Flagellin perception: a paradigm for innate immunity. Trends Plant Sci 7: 251-256 Görlach J, Volrath S, Knauf-Beiter G, Beckhove U, K ogel KH Oostendorp M, Staub T, Kessmann EWH, Ryals J (1996) Benzothiadiazole, a novel class of inducer of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell 8: 629-643 Graham JS, Hall G, Pearce G, Ryan CA (1986) Regulation of synthesis of proteinase inhibitors I and II mRNA in leaves of wounded tomato plants. Planta 169: 399-405 Grand J, Loake GJ (2000) Role of reactive oxygen intermediates and cognate redox signaling in disease resistance. Plant Physiol 124: 21-29 Guedes, M.E.M., Ku ć, J., Hammerschmidt, R., Bostock, R . (1982) Accumulation of six sesquitepenoid phytoalexins in tobacco leaves infiltrated with Pseudomonas lachrymans. Phytochemistry 21: 2987-2988 Guillet G, Poupart J, Basurco J, de Luca V (2000) Expression of tryptophan decarboxylase and tyrosine decarboxylase genes in tobacco results in altered biochemical and physiological phenoytpes. Plant Physiol 122: 933-943 Gundlach H, Muller MJ, Kutchan TM, Zenk MH (1992) Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc Natl Acad Sci USA 89: 2389-2393 Gundlanch H, Muller MJ, Kutchan TM, Zenk MH (1997) Biosynthesis and action of jasmonates in plants. Annu Rev Plant Physiol Plant Mol Biol 48: 355-381

121
Hain R, Reif H, Krause E, Langebartels R, Kindl H, Vornam B, Wiese W, Schreier PH, Stocker RH, Stenzel K (1993) Disease resistance results from foreign phytoalexin expression in a novel plant. Nature 361: 153-156 Halitschke R., Schittko U., Pohnert G., Boland W., Baldwin I.T. (2001) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. III. Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant Physiol 125: 711-717 Hallbrock K, Scheel D, Logemann E, Nürberger T, Par niske M, Reinold S, Sacks W, Schmelzer E (1995) Oligopepetide elicitor-mediated defense gene activation in cultured parsley cells. Proc Natl Acad Sci USA 92: 4150-4157 Hammond-Kosack KE, Jones JDG (1996) Resistance gene-dependent plant defense responses. Plant Cell 8: 1773-1791 Heo WD, Lee SH, Kim MC, Kim JC, Chung WS, Chun HJ, Lee KJ, Park HC, Choi JY, Cho MJ (1999) Involvement of specific calmodulin isoforms in salicylic acid-independent activation of plant disease resistance responses. Proc Natl Acad Sci USA 96: 766-771 Hermsmeier D, Schittko U, Baldwin IT (2001) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. I. Large-scale changes in the accumulation of growth- and defense-related plant mRNAs. Plant Physiol 125: 683-700 Hildmann T, Ebneth M, Peña-Cortés H, Sánchez-Serran o JJ, Willmitzer L, Prat S (1992) General roles of abscisic and jasmonic acids in gene activation as a result of mechanical wounding. Plant Cell 4: 1157-1170 Howles PA, Sewalt VJH, Paiva NL, Elkind Y, Bate NJ, Lamb C, Dixon RA (1996) Overexpression of L-phenylalanine ammonia-lyase in transgenic tobacco plants reveals control points for flux into phenylpropanoid synthesis. Plant Physiol 112: 1617-1624
Jacinto T, McGurl B, Franceschi V, Delano-Frier J, Ryan CA (1997) Tomato prosystemin promoter confers wound-inducible, vascular bundle-specific expression of the β-glucuronidase gene in transgenic tomato plants. Planta 203: 406-412 Jefferson RA, Kavanagh TA, Beven MW (1987) GUS fusions: β-glucuronidase as sensitive and versatile gene fusion marker in higher plants. EMBO J 6: 3901-3907 Johnson AW, Severson RF, Jackson DM (1992) Sources of resistance to insect pests of tobacco. CORESTA Inf Bull 3/4: 151-175 Jongsma MA, Bakker P, Peters J, Bosch D, Stiekema W J (1995) Adaptation of spodoptera exigua larvae to plant proteinase inhibitors by induction of gut

122
proteinase activity insensitive to inhibition. Proc Natl Acad Sci USA 92: 8041-8045 Jongsma, MA, Stiekema WJ, Bosch D (1996) Combatting inhibitor-insensitive proteases of insect pests. Trends Biotechol 14: 331-333 Kahl J, Siemens DH, Aerts RJ, Gäbler R, Kühnemann F , Preston CA Baldwin IT (2000) Herbivore-induced ethylene suppresses a direct defense but not a putative indirect defense against an adapted herbivore. Planta 210: 336-342 Keinänen M, Oldham NJ, Baldwin IT (2001) Rapid HPLC screening of jasmonate-induced increases in tobacco alkaloids, phenolics, and diterpene glycosides in Nicotiana attenuata. J Agric Food Chem 49: 3553-3558 Keller H, Czernic P, ponchet M, Ducrot PH, Back K, Chappell J, Ricci P, Marco Y (1998) Sesquiterpene cyclase is not a determining factor for elicitor- and pathogen-induced capsidiol accumulation in tobacco. Planta 205: 467-476 Keller H, Hohlfeld HV, Hahlbrock K, Schell D, Strac k D (1996) Changes in the accumulation of soluble and cell wall-bound phenolics in elicitor-treated cell suspension cultures and fungus-infected leaves of Solanum tuberosum. Phytochemistry 42: 389-396 Kessler A, Baldwin IT (2001) Defensive function of herbivore-induced plant volatile emissions in nature. Science 291: 2141-2144 Kleinig H (1989) The role of plastids in isoprenoid biosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:39-59 Knoesterm M, Van Loon LC, van den Hevel J, Henning J, Bol JF, Linthorst HJM (1998) Ethylene-insensitive tobacco lacks nonhost resistance against soil-borne fungi. Proc Natl Acad Sci USA 95: 1933-1937 Kombrink E., Somssich I.E. (1997) Pathogenesis related proteins and plant defense En Carroll CG, Tudzynski P (eds). Plant relationships. Berlin: Springer-Verlag, p 107-128 Kuhn DN, Chappell J, Boudet A, Hahlbrock K (1984) Induction of phenylalanine ammonia-lyase and 4-coumarate: CoA ligase mRNASs in cultured plant cells by UV light or fungal elicitor. Proc Natl Acad Sci USA 81: 1102-1106 Langebarterls C, Kerner K, Leonardi S, Scharaudner M, Trost M, Heller W, Sandermann H Jr (1991) Biochemical plant responses to ozone. 1. Differential induction of polyamine and ethylene biosynthesis in tobacco. Plant Physiol 95: 882-889 Laue G, Preston CA, Baldwin IT (2000) Fast track to the trichome: induction of N-acyl nornicotines precedes nicotine induction in Nicotiana repanda. Planta 210: 510-514

123
Lawton KA, Friedrich L, Hunt MH, Weymann K, Delaney T, Kessmann H, Staub T, Ryals J (1996) Benzothiadiazole induces disease resistance in Arabidopsis by activation of the systemic acquired resistance signal transduction pathway. Plant J 10:71-82 Lawton MA, Lamb CJ (1987) Transcriptional activation of plant defense genes by fungal elicitor, wounding, and infection. Mol Cell Biol 7: 335-341
Lee J, Vogt T, Scmidt J, Parthier B, Lobler M (1997) Methyl jasmonate-induced accumulation of coumaroyl conjugates in barley leaf segments. Phytochemistry 44: 589-592 Li L, Li C, Lee GI, Howe GA (2002a) Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato. Proc Natl Acad Sci USA 99: 6416-6421 Li C, Williams MM, Loh Y-T, Lee GI, Howe GA (2002b) Resistance of cultivated tomato to cell content-feeding herbivores is regulated by the octadecanoid-signaling pathway. Plant Physiol 130: 494-503
Linthorst HJ (1991). Pathogenesis-related proteins of plants. Crit Rev Plant Sci 10: 123-150. Liu Z, Williamson MS, Lansdell SJ, Denholm I, Milla r NS (2005) A nicotinic acetylcholine receptor mutation conferring target-site resistance to imidacloprid in Nilaparvata lugens (brown planthopper). Proc Natl Acad Sci USA 102: 8420-8425 Lois R, Dietrich A, Hahlbrock K, Schulz W (1989) A phenylalanine ammonia-lyase from parsley: structure, regulation, and identification of elicitor and light responsive cis-acting elements. EMBO J 8: 1631-1635 Lou Y, Baldwin IT (2003) Manduca sexta recognition and resistance among allopolyploid Nicotiana host plants. Proc Natl Acad Sci USA 100: 14581-14586 Lozoya Gloria E (2004) Molecular approaches to pepper phytoalexin production. CINVESTAV, Departamento de Ingeniería Genética. [email protected]. Maher EA, Bate NJ, Ni W, Elkind Y, Dixon RA, Lamb C J (1994) Increased disease susceptibility of transgenic tobacco plants with suppressed levels of preformed phenylpropanoid products. Proc Natl Acad Sci USA 91: 7802-7806 Masclaux C, Valadier M-H, Brugière N, Morot-Gaudry J-F, Hirel B (2000). Characterization of the sink/source transition in tobacco (Nicotiana tabacum L.) shoots in relation to nitrogen management and leaf senescence. Planta 211: 510-518 Mathieu Y, Kurkdjian A, Xia H, Guern J, Koller A, S piro MD, O’Neill M, Albersheim P, Darvill A (1991) Membrane responses induced by oligolacturonides in suspension-cultured tobacco cell. Plant J 1: 333-343

124
Mayer MJ, Narbad A, Parr AJ, Parker ML, Walton NJ, Mellon FA, Michael AJ (2001) Rerouting the plant phenylpropanoid pathway by expression of a novel bacterial enoyl-CoA hydratase/lyase enzyme function. Plant Cell 13: 1669-1682 Mayer RT, McCollum TG, McDonald RE, Polston JE, Doo stdar H (1996) Bemisia feeding induces pathogenesis-related proteins in tomato. En D Gerling, RT Mayer (eds.) Bemisia 1995: taxonomy, biology, damage control and management. Andover, Hants, UK: Intercept Ltd. P 179-188 McCloud ES, Baldwin IT (1997) Herbivory and caterpillar regurgitants amplify the wound-induced increases in jasmonic acid but not nicotine in Nicotiana sylvestris. Planta 203: 430-435 McGurl B, Pearce G, Orozco-Cárdenas M, Ryan CA (1992) Structure, expression, and antisense inhibition of the systemin precursor gene. Science 225:1570-1573 McKenzie C.L, Shatters R.G., Doostdar H., Lee S.D., Inbar M., Mayer R.T. (2002) Effect of geminivirus infection and Bemisia infestation on accumulation of pathogenesis-related proteins in tomato. Arch Insect Biochem Physiol 49: 203-214 Mehdy MC (1994) Active oxygen species in plant defense against pathogens. Plant Physiol 105: 467-472 Meindl T, Boller T, Felix G (1998) The plant wound hormone sistemin binds with the N-terminal to its receptor but needs the C-terminal part to activate it. Plant Cell 10: 1561-1570 Memelink J, Verpoorte R, kijne JW (2001) ORCAnization of jasmonate-reponsive gene expression in alkaloid metabolism. Trends Plant Sci 6: 212-219 Messiaen J, van Cutsem P (1994) Pectic signal transduction in carrot cell membrane: cytosolic and nuclear responses induced by oligogalacturonides. Plant Cell Physiol 35: 677-687 Murashige T, Skoog F (1962) A universal medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15: 473-479 Narváez-Vásquez J, Pearce G, Orozco-Cárdenas M, Fra nceschi VR, Ryan CA (1995) Autoradiographic and biochemical evidence for the systemic translocation of systemin in tomato plants. Planta 195: 593-600 Narváez-Vásquez J, Ryan CA (2004) The cellular localization of prosystemin: a functional role for phloem parenchyma in systemic wound signaling. Planta 218: 360-369

125
Narváez-Vásquez J, Pearce G, Ryan CA (2005) The plant cell matrix harbors a precursor of defense signaling peptides. Proc Natl Acad Sci USA 102: 12974-12977 Nelson CE, Walker-Simmons KM, Makus D, Zuroske G, G raham J, Ryan CA (1983) Regulation of syntesis and accumulation of proteinase inhibitors in leaves of wounded tomato plants En PA Hedin (ed) Plant Resistance in leaves. American Chemical Society, Washington, D.C. pp 103-122 Nicholson RL, Hammerschmidt R (1992) Phenolic compounds and their role in disease resistance. Annu Rev Phytopathology 30: 369-389 Niki T, Mitsuhara I, Seo S, Ohtsubo N, Oaci Y (1998) Antagonistic effect of salicylic acid and jasmonic acid on the expression of pathogenesis-related protein genes in wound mature tobacco leaves. Plant Cell Physiol 39: 500-507 Ohashi Y, Shigemi S, Hidehalu S, Hiroyuki K, Shigeo Y (2003) A diterpene as an endogenous signal for the activation of defense responses to infection with tobacco mosaic virus and wounding in tobacco. Plant Cell 15:863-873 Ohnmeiss TE, Baldwin IT (2000) Optimal defense theory predicts the ontogeny of an induced nicotine defense. Ecology 81: 1765-1783 Ohtsubo N, Mitsuhara I, Koga M, Seo S, Ohashi Y (1999) Ethylene promotes the necrotic lesion formation and basic PR gene expression in TMV-infected tobacco. Plant Cell Physiol 40: 808-817 Orozco-Cárdenas M, Ryan CA (1999) Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid path-way. Proc Natl Acad Sci USA 96: 6553-6557 Osbourn AE (1996) Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell 8: 1821-1831 O’Donnell PJ, Calvert C, Atzorn R, Wasternack C, Le yser HMO, Bowles DJ (1996) Ethylene as a signal mediating the wound response of tomato plants. Science 274: 1914-1917 Paré PW, Tumlinson JH (1997) Induced synthesis of plant volatiles. Nature 385: 30-31
Pallas JA, Paiva NL, Lamb C, Dixon RA (1996) Tobacco plants epigenetically suppressed in phenylalanine ammonia-lyase expression do not develop systemic acquired resistance in response to infection by tobacco mosaic virus. Plant J 10: 281-293 Parker JE (2003) Plant recognition of microbial patterns. Trends Plant Sci 8: 245-247

126
Pearce G, Strydom D, Johnson, S, Ryan CA (1991) A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science 253: 895-898 Pearce G, Johnson S, Ryan CA (1993) Structure-activity of deleted and substituted systemin, an 18-amino acid polypeptide inducer of plant defensive genes. J Biol Chem 268: 212-216 Pearce G, Moura DS, Stratmann J, Ryan CA (2001) Production of multiple plant hormones from a single polyprotein precursor. Nature 411: 817-820 Peña-Cortés H, Sánchez-Serrano JJ, Mertens R, Willm itzer L, Prat S (1989) Absicic acid is envolved in the wound-induced expresión of the proteinase inhibitor II gene in potato and tomato. Proc Natl Acad Sci USA 86: 9851-9855 Pieterse CMJ, van Loon LC (1999) Salicylic acid independent plant defence pathways. Trends Plant Sci 4: 52-58 Pieterse CMJ, van Wees SCM, van Pelt JA, Knoester M , Laan R, Gerrits N, Pelt JA, van Loon LC (1996) Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 8: 1225-1237 Qu N, Schittko U, Baldwin IT (2004) Consistency of Nicotiana attenautas’s herbivore- and jasmonate-induced transcriptional responses in the allotetraploid species Nicotiana quadrivalvis and Nicotiana clevelandii. Plant Physiol 135: 539-548 Reinbothe S, Mollenhauer B, Reinbothe C (1994) JIPs and RIPs: the regulation of plant gene expression by jasmonates in response to environmental cues and pathogens. Plant Cell 6: 1197-1209 Rhodes MJC (1994) Physiological roles for secondary metabolites in plants-some progress, many outstanding problems. Plant Mol Biol 24: 1-20 Richael C, Gilchrist D (1999) The hypersensitive response: a case of hold or fold? Physiol Mol Plant Pathol 55: 5-12 Richarsdson M (1991) Plant protein inhibitor of enzymes: their role in animal nutrition and plant defense. J Biol Educ 15: 178-182 Rickauer M, Brodschelm W, Bottin A, Véronesi C, Gri mal H, Esquerré-Tugayé MT (1997) The jasmonate pathway is involved differentially in the regulation of different responses in tobacco cells. Planta 202: 155-162 Rober KC (1989) Investigation on the synthesis of polyphenols and phytoalexins in rot-infected potato tubers. Biochem Physiol Pflanzen 184: 277-284

127
Rocha-Granados MC (2004) Sobre-expresión de la prosistemina y sistemina de jitomate (Lycopersicon esculentum) en plantas de tabaco (Nicotiana tabacum) y su influencia sobre la resistencia a insectos. Tesis de Doctorado. Cinvestav IPN-Unidad Irapuato Rocha-Granados MC, Sanchez-Hernández CV, Sanchez-He rnández C, Martinez-Gallardo NA, Ochoa-Alejo N, Delano-Frier J P (2005) The expresión of the hydroxyproline-rich glycopeptide systemin precursor A in response to (a) biotic stress and elicitors is indicative of its role in the regulation of the wound response in tobacco (Nicotiana tabacum L.). Planta (en prensa) Rodríguez-Concepción M, Gruissem W (1999) Arachidonic acid alters tomato HMGR expression and fruit growth and induces 3-hidroxy-3-methylglutaryl coenzyme A reductase-independent lycopene accumulation. Plant Physiol 119: 41-48 Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner HY, Hunt MD (1996) Systemic acquired resistance. Plant Cell 8: 1809-1819
Ryan CA (1981) Proteinase inhibitors. The Biochemistry of Plants. Academic Press Inc. 6: 351-369. Ryan CA (1990) Protease inhibitors in plants: genes for improving defenses against insects and pathogens. Annu Rev Phytopathol 28: 425-449 Ryan CA (2000) The systemin signaling pathway: differential activation of defensive genes. Biochem Biophys Acta 1477: 112-122 Ryan CA, Jagendorf A (1995) Self defense by plants. Proc Natl Acad Sci USA 92: 4075 Ryan CA, Moura DS (2002) Systemic signaling in plants: A new perception. Proc Natl Acad Sci USA 99: 6519-6520 Ryan CA, Pearce G (2001) Polypeptide hormones. Plant Physiol 125:65-68 Ryan CA, Pearce G (2003) Systemins: A functionally defined family of peptide signals that regulate defensive genes in Solanaceae species. Proc Natl Acad Sci USA 100: 14577-14580
Saitoh F, Norma M, Kawashima N (1985) The alkaloid contents of sixty Nicotiana species. Phytochemistry 24: 477-480 Sano H, Ohashi Y (1995) Involvement of small GTP-binding proteins in defense signal transduction pathways of higher plants. Proc Natl Acad Sci USA 92: 4138-4144 Saunders JW, Bush LP (1979) Nicotine biosynthetic enzyme activities in Nicotiana tabacum L. genotypes with different alkaloid levels. Plant Physiol 64: 236-240

128
Schaller A (1999) Oligopeptide signalling and the action of systemin. Plant Mol Biol 40: 763-769 Schaller A, Ryan CA (1995) Systemin: a polypeptide defense signal in plants. Bioassays 18: 27-33 Scheer JM, Ryan CA (1999) A 160-KD receptor on the surface of Lycopersicon peruvianum suspension-culture cell. Plant Cell 11: 1525-1535 Scheer JM, Ryan CA (2000) The systemin receptor SR160 from Lycopersicon peruvianum is a member of the LRR receptor kinase family. Plant Biol 99: 9585-9590 Scheer JM, Pearce G, Ryan CA (2003) Generation of systemin signaling in tobacco by transformation with the tomato systemin receptor kinase gene. Proc Natl Acad Sci USA 100: 10114-10117 Schilmiller AL, Howe GA (2005) Systemic signaling in the wound response. Curr Opin Plant Biol 8: 369-377 Schittko U, Baldwin IT (2003) Constraints to herbivore-induced systemic responses: bidirectional signaling along orthostichies in Nicotiana attenuata. J Chem Ecol 29: 763-770 Schittko U, Hermsmeier D, Baldwin IT (2001) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. II. Accumulation of plant mRNAs in response to insect-derived cues. Plant Physiol 125: 701-710 Seo S, Seto H, Koshino H, Yoshida S, Ohashi Y (2003) A diterpene as an endogenous signal for the activation of defense responses to infection with tobacco mosaic virus and wounding in tobacco. Plant Cell 15: 863-873 Shedle GL, Wesley SV, Korth KL, Chen F, Lamb C, Dix on R (2003) Phenylpropanoid compounds and disease resistance in transgenic tobacco with altered expression of L-phenylalanine ammonia-lyase. Phytochemistry 64: 153-161 Shiroya M, Lister GR, Nelson CD, Krotkov G (1961) Translocation of C14 in tobacco at different stages of development following assimilation of C14 by a single leaf. Can J Bot 39: 855-864 Shoji T, Yamada Y, Hashimoto T (2000) Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol 41: 831-839 Sinclair SJ, Johnson R, Hamill JD (2004) Analysis of wound-induced gene expression in Nicotiana species with contrasting alkaloid profiles. Func Plant Biol 31: 721-729

129
Sisson VA, Severson RF (1990) Alkaloid composition of the Nicotiana. Beitr Tabakforsch Int 14: 327-340 Snook ME, Mason PF, Sisson VA (1986) Polyphenols in the Nicotiana species. Tob Sci 30: 43-49 Snook ME, Johnson AW, Severson RF, Teng Q, White RA Jr, Jackson DM (1997) Hydroxygeranyllinalool glycosides from tobacco exhibit antibiosis in the tobacco budworm [Heliothis virescens (F.)]. J Agric Food Chem 45: 2299-2308 Staswick PE, Su W, Howell SH (1992) Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc Natl Acad Sci USA 89: 6837-6840 Steppuhn A, Gase K, Krock B, Halitscke R, Baldwin I T (2004) Nicotine’s defensive function in nature. PLoS Biol 2: 1074-1080 Swain T, Hillis WE (1959) The phenolic constituents of Prunus domesticus I: The quantitative analysis of phenolic constituents. J Agric Food Chem 10: 65-68 Stintzi A, Heitz T, Prasad V, Wiedemann-Merdinpglus S, Kauffmann S, Geoffroy P, Legrand M Friting B (1993) Plant pathogenesis-related proteins and their role in defense against pathogens. Biochimie 75: 687-706 Stout MJ, Duffey SS (1996) Characterization of induced resistance in tomato plants. Entomol Experimen Appl 79: 273-283 Stout MJ, Bostock RM (1999) Specificity of induced responses to arthropods and pathogen En Induced plant defenses against pathogen pathogens and herbivores: biochemistry, ecology and agriculture. APS press. 183: 210 Stout MJ, Findantsef AL, Duffey SS, Bostock RM (1999) Signal interactions in pathogens and herbivores of the tomato, Lycopersicum esculentm. Physiol Mol Plant Pathology 54: 115-130 Tanguy J, Martin C (1972) Phenolic compounds and the hypersensitivity reaction in Nicotiana tabacum infected with tobacco mosaic virus. Phytochemistry 11: 19-28 Thain JF, Doherty HM, Bowles DJ, Wildon DC (1990) Oligosaccharides that induce proteinase inhibitor activity in tomato plants cause depolarization of tomato leaf cells. Plant Cell Env 13: 569-574 Threlfall DR, Whitehead IM (1988) Co-ordinated inhibition of squalene synthetase and induction of enzymes of sesquiterpenoid phytoalexin biosynthesis in cultures of Nicotiana tabacum. Phytochemistry 27: 2567-2580

130
Tian M, Benedetti B, Kamoun S (2005) A second Kazal-like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis-related protease P69B of tomato. Plant Physiol 138: 1785-1783 Thordal-Christensen H (2003) Fresh insights into processes of nonhost resistance. Curr Opin Plant Biol 6: 351-357 Toumadje A, Johnson WC Jr (1995) Systemin has the characteristics of poly (L-proline) II type helix. J Am Chem Soc 117: 7023-7024 Tso TC (1972) Physiology and Biochemistry of Tobacco Plants. Dowden, Hutchinson and Ross; Stroudsburg, PA. van Dam NM, Horn M, Mareš M, Baldwin IT (2001) Ontogeny constrains systemic protease inhibitor response in Nicotiana attenuata. J Chem Ecol 27: 547-568 Van Loon LC, Pierpoint WA, Boller T, Conejero V (1994) Recommendations for naming plant pathogenesis-related proteins. Plant Mol Biol Rep 12: 245-264 van de Ven WTG, LeVesque CS, Perring TM, Walling LL (2000) Local and systemic changes in squash gene expression in response to silverleaf whitefly feeding. Plant Cell 12: 1409-1423 Vernooij B, Friedrich L, Morse A, Reist R, Kolditz- Jawhar R, Ward E, Uknes S, Kessmann H, Ryals J (1994) Salicylic acid is not the translocated signal responsible for inducing systemic acquired resistance but is required in signal transduction. Plant Cell 6: 959-965 Vögeli U, Chappel J (1988) Induction of sesquiterpene cyclase and suppression of squalene synthase activities in plant cell cultures treated with fungal elicitors. Plant Physiol. 88: 1291-1296 Vögeli U, Chappel J (1990) Regulation of sesquiterpene cyclase in cellulase-treated tobacco cell suspension cultures. Plant Physiol. 94: 1860-1866 Walling LL (2000) The myriad plant responses to herbivores. J Plant Growth Regul 19: 195-216 Ward EWB, Stoessl A, Stothers JB (1977) Metabolism of the sesquiterpenoid phytoalexins capsidiol and rishitin to their 13-hydroxy derivatives by plant cells. Phytochemistry 16: 2024-2025 Whetten R., Sederoff R. (1995) Lignin biosynthesis. Plant Cell 7: 1001-1013 Whitehead IM, Threlfall DR, Ewing DF (1987) Cis-9,10-dihyrocapsenone: a possible catabolite of capsidiol from cell suspension cultures of Capsicum annuum. Phytochemistry 26: 1367-1369

131
Whitehead IM, Threlfall DR, Ewing DF (1987) Cis-9,10-dihydrocapsenone: a possible catabolite of capsidiol from cell suspension cultures of Capsicum annuum. Phytochemistry 26: 1367-1369 Wink M (1998) Modes of action of alkaloids En Alkaloids MF Roberts, Wink M (eds) pp. 301-326. Plenum Press, New York Winz RA, Baldwin IT (2001) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. IV. Insect-induced ethylene reduces jasmonate-induced nicotine accumulation by regulating putrescine N-methyltransferase transcripts. Plant Physiol 125: 2189-2202 Wittstock U, Gershenzon J (2002) Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr Opin Plant Biol 5: 300-307 Yamamoto RT (1964) Mass rearing of the tobacco hornworm II. Larval rearing and pupation. J Econ Entomol 62: 1427-1431 Yin S, Mei L, Newmann J, Back K, Chappell J (1997) Regulation of sesquiterpene cyclase gene expression. Plant Physiol 115: 437-451 Zacharius RM, Kalan EB, Kimoto WI (1985) Biotransformation of potato stress metabolites rishitin, lubimin, and 15-dihydrolubimin by potato and soybean cultures. Plant Cell Rep 4: 1-3 Zavala, JA, Patankar AG, Gase K, Baldwin IT (2004a) Constitutive and inducible trypsin proteinase inhibitor production incurs large fitness costs in Nicotiana attenuata. Proc Natl Acad Sci USA 101: 1607-1612 Zavala JA, Patankar AG, Gase K, Hui D, Baldwin IT (2004b) Manipulation of endogenous trypsin proteinase inhibitor production in Nicotiana attenuata demonstrates their function as antiherbivore defenses. Plant Physiol 134: 1181-1190 Zhang Z-P, Baldwin IT (1997) Transport of [2-14C] jasmonic acid from leaves to roots mimics wound-induced changes in endogenous jasmonic acid pools in Nicotiana sylvestris. Planta 203: 436-441 Zhu-Salzman K., Salzman R.A, Koiwa H., Murdock L.L, Bressan R.A., HAsegawa P.M. (1998) Ethylene negatively regulates local expression of plant defense lectin genes. Plant Physiol 104: 365-372