
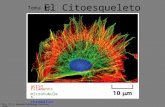
Citoesqueleto y Motilidad Celular
33
-
Upload
francisco-hidalgo -
Category
Documents
-
view
37 -
download
0
description
clase de citoesqueleto
Transcript of Citoesqueleto y Motilidad Celular


Universidad del ZuliaFacultad de MedicinaEscuela de MedicinaCátedra: BiologíaProf.: Mgs. Patricia de Moreno
EL CITOESQUELETOEL CITOESQUELETOY LA MOTILIDAD CELULARY LA MOTILIDAD CELULAR

CITOESQUELETOCITOESQUELETO
1. Francisco Hiidalgo
Es el sistema esquelético de las células eucariotas que tiene funciones análogas. Esta compuesto por tres estructuras filamentosas bien definidas que en conjunto constituyen una red interactiva, estos son:
Microtúbulos Microfilamentos Filamentos Intermedios
Cada uno de estos tres tipos de filamentos citoesqueleticos es un polímero de subunidades proteicas unidas mediante enlaces débiles no covalentes. Este tipo de construcción se presta a un ensamble y un desensamble rápidos, que dependen de una regulación celular compleja.

CITOESQUELETOCITOESQUELETO
1. Francisco Hiidalgo
Funciones Principales
Propiedades de los Microtubulos, los filamentos Intermedios y los filamentos de Actina

ESTUDIO DEL CITOESQUELETOESTUDIO DEL CITOESQUELETO
2. Adriana Hornes
El uso del microscopio con fluorescencia:El microscopio con fluorescencia, tiene una participación prepoderante en esta revolución al permitir a los investigadores observar en forma directa los procesos moleculares en las células vivas.En la técnica más usual se sintetizan proteínas con marcas fluorescentes dentro de la célula como una proteína fusionada que contiene proteína verde fluorescente(PVF).
La miscroscopia de fluorescencia también puede usarse para revelar la localización de una proteína en concentraciones muy bajas dentro de la célula.

MICROTUBULOSMICROTUBULOS
3. Franchesca
Son estructuras tabulares huecas que se encuentran en casi todas las células eucariotas. Forman parte de los cilios y flagelos así como del huso mitotico, los Microtúbulos tienen un diámetro externo de 25nm y puede extenderse a lo largo y ancho de la célula; su pared esta formada por proteínas globulares dispuestas en hileras longitudinales protofilamentos.

MICROTUBULOSMICROTUBULOS
3. Franchesca
Proteínas relacionadas con los Microtúbulos:
Además de las proteínas que están compuestas por Microtúbulos esta la componen proteínas adicionales llamadas proteínas relacionadas con Microtúbulos (MAP) las cuales tienen un dominio que se une al lado de un microtúbulo y otro que sobresale hacia fuera como un filamento de la superficie del microtúbulo. Estas pueden incrementar la estabilidad de los Microtúbulos y promover su ensamble.

FUNCIONES DE LOS MICROTUBULOSFUNCIONES DE LOS MICROTUBULOS
4. Georgina Isea
Como soportes y Organizadores estructurales:
Los microtúbulos son lo bastante rígidos para resistir fuerzas que pudieran comprimir o doblar la fibra.
Esta propiedad les permite brindar soporte
mecánico, tal como vigas de acero que
sostienen a un edificio.
En las células Animales, los microtúbulos se extienden
con un patrón radical.

Como agentes de Motilidad Intracelular:
Las células vivas están repletas de actividad. El estudio de la Motilidad Intracelular se iniciara con las células nerviosas, cuyos movimientos intracelulares dependen de un conjunto muy organizado de microtúbulos y otros filamentos del citoesqueleto.
FUNCIONES DE LOS MICROTUBULOSFUNCIONES DE LOS MICROTUBULOS
4. Georgina Isea
Trasporte Axónico
Transporte Retrógrado
Transporte Anterógrado
Cuando se inyecta aminoácidos marcados en el cuerpo celular se incorporan proteínas marcadas que se mueven hacia el axón y viajan x toda su extensión.

Las Proteínas motoras de una Célula son aquellas que convierten la energía química en energía mecánica.
PROTEÍNAS MOTORAS QUE SE CRUZAN EN EL PROTEÍNAS MOTORAS QUE SE CRUZAN EN EL CITOESQUELETO MICROTUBULARCITOESQUELETO MICROTUBULAR
5. Ana Inciarte
Estas se dividen en tres grandes familias:
Cinesinas Miosinas
Dineína citoplásmica

PROTEÍNAS MOTORAS QUE SE CRUZAN EN EL PROTEÍNAS MOTORAS QUE SE CRUZAN EN EL CITOESQUELETO MICROTUBULARCITOESQUELETO MICROTUBULAR
5. Ana Inciarte
a
_Estructura de una molécula de cinesina que consta principalmente de dos cadenas pesadas que se entrelazan en la región del tallo. Las cabezas generadoras de fuerzas se unen almicrotúbulo, y la cola a la carga que se transporta.
Tallo
Cadenas pesadas
Bisagra flexible
Cabeza Cuello Cola
Cadenas Ligeras
b
_Diagrama esquemático de una molécula de cinesina moviendo una vesícula a lo largo de una vía microtubular.
_Estructura de una molécula de dineína citoplásmica que consta de dos cabezas globulares grandes generadoras de fuerza, cada una compuesta de una cadena pesada de dineína, y un tallo, un número de subunidades más pequeñas en la base de la molécula, que se presume median la unión de laproteína motora a la carga que se transporta.
aCadenas pesadas Cadenas
ligeras
CabezaPie Tallo
_Diagrama esquemático de dos vesículas moviéndose en dirección opuesta a lo largo del mismo microtúbulo, una impulsada por cinesina moviéndose en dirección anterógrada y la otra por dineína citoplásmica en dirección retrógrada.
bCuerpo Celular
Cinesina
Dineína
Terminación del Axón
Complejo de Dineína
Cinesina:
Dineina Citoplasmatica:

CENTROS ORGANIZADORES DE LOS CENTROS ORGANIZADORES DE LOS MICROTUBULOSMICROTUBULOS
6. Anlin Leguisimo
Los centros de organizaciones de microtúbulo son estructuras especializadas que intervienen en diversos fenómenos de la célula.La unión de un microtúbulo depende de su localización y de su orientación.
Los Centrosomas :Son estructuras que contienen dos centriolos con forma de barril rodeados por material peric3ntriolar.

CENTROS ORGANIZADORES DE LOS CENTROS ORGANIZADORES DE LOS MICROTUBULOSMICROTUBULOS
6. Anlin Leguisimo
Estos se encuentran en la base del cilio o flagelo, tienen estructura idéntica a la de los centriolos. De hecho los cuerpos basales y centriolo pueden dar origen uno al otro.
Los Centrosomas, Cuerpos basales y otros MTOC:
El centrosoma es el principal sitio de inicio de los Microtúbulos en las células animales y por lo general permanecen en el centro de la red microtubular.

CENTROS ORGANIZADORES DE LOS CENTROS ORGANIZADORES DE LOS MICROTUBULOSMICROTUBULOS
6. Anlin Leguisimo
Nucleación del microtúbulo
Nucleacion de un micro túbulo en el centrosoma
En la nucleación de microtúbulos se demuestra la participación del centrosoma en la organización del cito esqueleto microtubular.
Participación de la tubulina gamma en la función del centrosoma

CILIOS Y FLAGELOS: ESTRUCTURA Y FUNCIÓNCILIOS Y FLAGELOS: ESTRUCTURA Y FUNCIÓN
Naivelen Hernandez
Los cilios y flagelos son organelos móviles similares a vellosidades que sobresalen de la superficie de diversas células eucariotas. En esencia, son la misma estructura.
Desempeñan una función sensorial al vigilar las propiedades de los líquidos extracelulares.

Brazos de Dineína:
ESTRUCTURA DE LOS CILIOS Y FLAGELOSESTRUCTURA DE LOS CILIOS Y FLAGELOS
El cambio de conformación de la Dineína sirve como fuerza básica de impulso para el movimiento ciliar o flagelar.
La Dineína que se encuentra en el axonema de cilios y flagelos recibe el nombre de Dineína axonémica, una proteína de gran tamaño encargada de la conversión de energía química del ATP en energía mecánica de la locomoción ciliar. Esta molécula de Dineína consiste tres cadenas pesadas y varis cadenas intermedias y ligeras, cada cadena pesada de Dineína está formada por un pie largo, una cabeza en forma de rueda, y un tallo.
9. Nicola Hindle

ESTRUCTURA DE LOS CILIOS Y FLAGELOSESTRUCTURA DE LOS CILIOS Y FLAGELOS
9. Nicola Hindle
En un axonema el tallo de dineína se encuentra anclado a la superficie del túbulo A , con las cabezas orientadas al Túbulo B. El mecanismo de locomoción se puede resumir en 4 pasos:
Paso1: Los brazos de dineína anclados en el TA de adhieren a los sitios de unión del TB superior.Paso2: La dineína experimenta un cambio en su conformación o golpe de poder que ocasiona el deslizamiento.Paso3:Los brazos de dineína de desprendieron del túbulo B dela pareja superior.Paso4: los brazos de unen de nuevo a la pareja superior para comenzar el ciclo.

MECANISMO DE LOCOMOCIÓN CILIAR Y MECANISMO DE LOCOMOCIÓN CILIAR Y FLAGELARFLAGELAR
La regulación de la locomoción ciliar y flagelar:
Empieza con la regulación de los brazos de dineína. Se cree que el par central de microtúbulos y los rayos radiales determinan que brazo de dineína se activan en un instante determinado, conforme este rota, el par central barre periódicamente cada rayo radial, con lo que parece enviar una señal del rayo al brazo de dineína en el túbulo adyacente, lo que activa el brazo para que se efectúe su movimiento de balanceo. La activación o desactivación de los brazos de dineína se realiza mediante la eliminación o adición de grupos Pi de unos o más polipéptido motores.
9. Nicola Hindle

FUNCIÓN DE LOS CILIOS EN EL FUNCIÓN DE LOS CILIOS EN EL DESARROLLO Y LA ENFERMEDADDESARROLLO Y LA ENFERMEDAD
10. Suendy Huerta
Situs inversus
Síndrome de Kartagener
Estructura anormaldel axonema

FUNCIÓN DE LOS CILIOS EN EL FUNCIÓN DE LOS CILIOS EN EL DESARROLLO Y LA ENFERMEDADDESARROLLO Y LA ENFERMEDAD
9. Suendy Huerta
Nodo embrionario
CilioMicrotubulos centrales
La rotación de los cilios nodales mueve el liquido circulante al lado izquierdo de la linea media del embrión, desarrollando al azar la asimetría entre izquierda y derecha.

FILAMENTOS INTERMEDIOS FILAMENTOS INTERMEDIOS
11. Annielis Hernandez
Componentes del citoesqueleto formados por agrupaciones de proteínas
fibrosas.Estructura
Función
Rigidez a la célula.
Rigidez al núcleo.
Regulación de transcripción.

FILAMENTOS INTERMEDIOS FILAMENTOS INTERMEDIOS
11. Annielis Hernandez
Principales proteínas Filamentosas
Propiedades y Distribución de las principales proteínas de filamentos intermedios en los mamíferos

FILAMENTOS INTERMEDIOS FILAMENTOS INTERMEDIOS
12. María Isabel Hernández
Ensamble y desensamble de filamentos intermedios:
La unidad básica del ensamble de los IF es un tetrámero formado por dos dímeros que se alinean lado a lado en forma intercalado Los pares de monómeros se organizan en orientación paralela, con sus extremos alineados para formar dímeros, según el tipo de filamento intermedio los dímeros pueden formarse por monómeros idénticos (homodimero) o no idénticos (heterodimeros) que a su vez se organizan en forma intercalada antiparalela para formar tetrámeros.

MICROFILAMENTOSMICROFILAMENTOS
14. Gabriel higuera
Ensamble y desensamble de los microfilamentos:Pasos:
1-Adicion filamentos preformados de actina, a una solucion de actina en presencia de ATP
2-Concentración alta: continua la adicion de sub-unidades en ambos extremos.
3-Decae la concentración de ATP-actina y la adicion continua en el extremo mas (+). Pero cesa en el extremo menos (-).
4-Se agregan monomeros al extremo “+” y no al extremo “-”. Se consumen mas monomeros con la elongacion del filamento. Y se alcanza un nivel de equilibrio.
5- En estado de equilibrio las subunidades añadidas en el extremo ‘+’ se retiran del extremo ‘-’.Las subunidades individuales en realidad se mueven a lo largo del filamento (“Efecto noria”)

MIOSINAMIOSINA
Es una proteína con una cola y una cabeza que se une con otra formando un dimero y a su vez estos se unen formando un gran polímero de miosina, asemejándolo a ramitas de árbol. Todas estas miosinas conforman la fibra de miosina gruesa y central en el sarcomero.
Es una enzima mecano - química, es decir, convierte la energía química en mecánica por eso es también llamada de proteína motora. Entonces, en los movimientos generados por esos elementos, la miosina es el motor, los filamentos de actina son los raíles y el ATP, el combustible.
15. Adriana Hernández

MIOSINAMIOSINA
Estructura: La miosina I tiene una (monomérica); en esta, tiene casa de campo de conexión para la membrana plasmática a la cual se prende; por la cabeza ella se prende a la actina. La miosina II y V son diméricas.
Fisiología: En las células musculares acanaladas, los filamentos de actina y de miosina están arreglados de manera definida y estable en el citoplasma, para que puedan promover la contracción muscular.
15. Adriana Hernández

PROTEÍNAS DE UNIÓN CON LA ACTINAPROTEÍNAS DE UNIÓN CON LA ACTINA
Kiaved Hidalgo
De los filamentos de Actina se han aislado casi 100 diferentes proteínas enlazadas a esta, pertenecientes a muchas familias de uno u otro tipo de célula.
El tema es muy complejo y todavía no muy bien comprendido, las proteínas enlazadas a la actina se pueden dividir en varias categorías según sus presuntas funciones en las células:
1.-Proteinas que secuestran monómeros: Secuestran o toman los monómeros de actina. Los cambios en la concentración o actividad de proteínas que secuestran actina, pueden variar el equilibrio monómero – polímero en cierta región de la célula y provocar que en ese momento se favorezca la polimerización o la despolimerización.
2.-Proteinas que bloquean extremos o (casquetes): Estas pueden regular la longitud de los filamentos de actina al enlazarse a uno u otro extremo del mismo, y así formar un casquete. Ciertas proteínas bloqueadoras de extremos pueden promover la formación de nuevos filamentos (nucleación) y al mismo tiempo inhibir el alargamiento de los filamentos existentes. Lo que da como resultado filamentos mas pequeños.
3.-Proteinas de enlace transversal: Existe una gran variedad de proteínas capaces de alterar la organización tridimensional de una población de filamentos de actina. Cada proteína tiene dos o mas sitios de enlace para actina y por lo tanto pueden enlazar dos o mas filamentos de actina a la vez.

PROTEÍNAS DE UNIÓN CON LA ACTINAPROTEÍNAS DE UNIÓN CON LA ACTINA
Kiaved Hidalgo
4.-Proteinas que cortan filamentos: Este tipo de proteínas son capaces de enlazarse a un filamento ya existente y partirlo en dos reduciendo su longitud.
5.-Proteinas enlazadas a la membrana: Son proteínas que forman enlaces entre filamentos de actina y la membrana plasmática. Ya sea de forma directa por medio de fijación a una proteína integral de membrana o indirectamente por fijación a una proteína periférica de la membrana.

PROTEÍNAS DE UNIÓN CON LA ACTINAPROTEÍNAS DE UNIÓN CON LA ACTINA
Kiaved Hidalgo
Proteínas Enlazadas a la Actina

Interacciones célula-sustrato y papel de las fibras de esfuerzo: Las fibras de esfuerzo son muy parecidas a las miofibrillas del tejido del músculo esquelético, más que cualquier otra estructura en una célula no muscular. Además de acuna, las fibras de esfuerzo contienen algunas otras proteínas típicas de células musculares, incluyendo miosina, tropomiosina y a-actinina.
EJEMPLOS DE CONTRACTILIDAD Y MOTIVILIDAD NO EJEMPLOS DE CONTRACTILIDAD Y MOTIVILIDAD NO MUSCULARMUSCULAR
20. Liliangel Hernandez
Locomoción celular: La locomoción celular, como ocurre en un organismo multicelular, de ordinario es imposible de observar, ya que tiene lugar en tejidos opacos y células indistinguibles de otras células que las rodean.

CRECIMIENTO AXONICOCRECIMIENTO AXONICO
Los axones comienzan a brotar de las neuronas poco después que estas han terminado de dividirse y han alcanzado su posición correcta en el cerebro. Para las primeras porciones del cerebro que se desarrollan, esto sucede cuando el embrión es sólo de algunos milímetros de longitud, continuando el proceso hasta alrededor del nacimiento.
En el extremo de cada axón en crecimiento hay una estructura llamada cono de crecimiento, que tiene tres partes principales:
21. María Del Valle Hernández
Núcleo central, rico en microtúbulos, mitocondrias y otros orgánulos.
Cuerpo, del que salen los filopodios.
Filopodios, prolongaciones finas en la expansión terminal del extremo axónico, que se forman y desaparecen rápidamente y entre los cuales se encuentran los lamelipodios, que son expansiones laminares.

CRECIMIENTO AXONICOCRECIMIENTO AXONICO
Una de las teorías que explican el avance del cono axónico propone:
21. María Del Valle Hernández
En el frente de avance ocurre la polimerización de la Actina
y empuja hacia adelante.
Actina polimerizada se retira hacia el centro del cono, con ayuda
de miosina.
Los monómeros generados por la despolimerización en la parte trasera se dirigen hacia la parte
delantera polimerizando de nuevo en el frente.
La estructura del cono de crecimiento se aplana y extiende filopodios, lo que
sugiere una búsqueda de señales apropiadas para dirigir el
crecimiento.
Los receptores de señales suelen ser el primer eslabón de una
cadena de segundos mensajeros intracelulares.
El segundo mensajero más
importante es el calcio.
Otros segundo mensajero importantes son
los nucleótidos cíclicos.

GRACIASGRACIASPOR SU ATENCION…














![Motilidad Celular[1]](https://static.fdocuments.es/doc/165x107/55cf9976550346d0339d872c/motilidad-celular1.jpg)