
CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE
83
CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE Pseudomonas putida por TRANSPOSON mini Tn5 y EVALUACIÓN DE LOS GENES AFECTADOS LAIDY JOHANA CORTÉS ORDOÑEZ PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS CARRERA DE MICROBIOLOGIA INDUSTRIAL São Paulo, Brasil. Julio 2008
Transcript of CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE

CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE
Pseudomonas putida por TRANSPOSON mini Tn5 y EVALUACIÓN DE LOS GENES
AFECTADOS
LAIDY JOHANA CORTÉS ORDOÑEZ
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
CARRERA DE MICROBIOLOGIA INDUSTRIAL
São Paulo, Brasil.
Julio 2008

CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE
Pseudomonas putida por TRANSPOSON mini Tn5 y EVALUACIÓN DE LOS GENES
AFECTADOS
LAIDY JOHANA CORTÉS ORDOÑEZ
TRABAJO DE GRADO Presentado como requisito parcial
Para optar al titulo de
MICROBIOLOGA INDUSTRIAL
DIRECTOR:
Dr. José Gregorio Cabrera Gomez Biólogo, Ph.D.
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
CARRERA DE MICROBIOLOGIA INDUSTRIAL
São Paulo, Brasil.
Julio 2008

NOTA DE ADVERTENCIA Artículo 23 de la Resolución N° 13 de Julio de 1946
“La Universidad no se hace responsable por los conceptos emitidos por
sus alumnos en sus trabajos de tesis. Solo velará por que no se publique
nada contrario al dogma y a la moral católica y por que las tesis no
contengan ataques personales contra persona alguna, antes bien se vea
en ellas el anhelo de buscar la verdad y la justicia”.

CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE
Pseudomonas putida por TRANSPOSON mini Tn5 y EVALUACIÓN DE LOS GENES
AFECTADOS
LAIDY JOHANA CORTÉS ORDOÑEZ
APROBADO
Dr. José Gregorio Cabrera Gomez Biólogo, Ph.D.
Director
Rafael Costa Santos Rocha Sonia Regina da Silva Queiroz Biólogo, Ph.D. Ingeniera Química Ph.D. Jurado Jurado

CONSTRUCCIÓN DE UNA BIBLIOTECA DE MUTANTES PHA- NEGATIVO DE
Pseudomonas putida por TRANSPOSON mini Tn5 y EVALUACIÓN DE LOS GENES
AFECTADOS
LAIDY JOHANA CORTÉS ORDOÑEZ
APROBADO
Ingrid Schuler, Ph.D Janeth Arias MSc Decana Académica Directora de Carrera

Dedicatoria: Dedicatoria: Dedicatoria: Dedicatoria:
A mi mama, A mi mama, A mi mama, A mi mama,
Yolanda Ordoñez Yolanda Ordoñez Yolanda Ordoñez Yolanda Ordoñez
por todo por todo por todo por todo
El El El El amor , ded amor , ded amor , ded amor , dedicación y comprensiónicación y comprensiónicación y comprensiónicación y comprensión
Durante toda mi vida, Durante toda mi vida, Durante toda mi vida, Durante toda mi vida,
Por apoyarme Por apoyarme Por apoyarme Por apoyarme
Cuando nadie mas lo hacia…Cuando nadie mas lo hacia…Cuando nadie mas lo hacia…Cuando nadie mas lo hacia…

VI
Agradecimientos:
Al Dr. José Gregorio Gomez por haberme permitido trabajar em su grupo
de investigación; por su orientación, colaboración y apoyo durante la
realización de este trabajo.
A la Dra. Luiziana Ferreira da Silva por su importante papel en el enlace
entre dos instituciones, para el intercambio de conocimientos.
Al Dr. Rafael costa, por su apoyo constante e incondicional en las etapas
claves de este trabajo.
A la Dra. Sonia Regina Queiroz, por su orientación y enseñanzas en el
área de biología molecular.
A todo el equipo de trabajo de laboratorio, compañeros como Dani,
Marco, Nuri, Tatiana, Sayuri, Rogerio, Camila y Francy, por su
colaboración en repetidas ocasiones, que hicieron de cada día de trabajo
un día mas de alegría en Brasil.
A Margarita, Oswaldo, Daniel y Andrea, por brindarme un hogar en Brasil
y permitirme ser un miembro más de la familia, por su apoyo, consejos y
momentos compartidos, sin los cuales no hubiera sido posible la
realización de este trabajo.
A Andrea Murillo, por su apoyo, complicidad, amistad y discusiones,
durante todo este tiempo.

VII
A Margarita Barreneche, por sus constantes consejos y jalones de orejas
que no me dejaron decaer y me hicieron reaccionar en los momentos
más difíciles.
A mi Tía Gloria, Nata, Miguel, y demás familia por brindarme su amor y
apoyo durante todos estos años.
A mis amigos de toda la vida, Lady, Yulieth, Ingrid, Dieguito , Carlos y
Winston, por los momentos de alegría y lagrimas compartidos en mas de
cinco años.
A mi papa, por su apoyo en los momentos finales de este trabajo.
Y Finalmente al ser mas importante en mi vida, mi mama, por confiar en
mi en cada momento, por brindarme su amor y apoyo incondicional en
cada instante.

VIII
TABLA DE CONTENIDOS
Página 1. Introducción
15
2. Marco teórico
17
2.1 Bioplasticos
17
2.2 PHAs
18
2.3 Síntesis de polihidroxialcanoatos
20
2.4 Biosíntesis de PHA a partir de sustratos relacionados
y no relacionados.
21
2.5 Biosíntesis de Polihidroxialcanoatos, Catabolismo y Regulación.
22
2.6 Biosíntesis de PHAMCL y metabolismo de ácidos grasos.
26
2.7 Arquitectura macromolecular de las Inclusiones.
28
2.8 Aplicaciones Biotecnológicas de los Polihidroxialcanoatos
29
2.9 Polihidroxialcanoatos en el área medica.
30
2.10 Producción Industrial
33
3. Planteamiento del problema y justificación.
34
4. Objetivos.
36
4.1 Objetivo general.
36
4.2 Objetivos específicos.
36
5. Materiales y métodos.
37
5.1 Microorganismos y plásmidos
37
5.2 Condiciones de cultivo
38
5.3 Preservación de las cepas bacterianas
38

IX
5.4 Construcción de la biblioteca y aislamiento de mutantes deficiente en el acumulo de PHAMCL utilizando transposon mini Tn5
39
5.4.1 Selección y aislamiento de mutantes deficientes en el acumulo de PHAs.
40
5.5 Evaluación de la producción de PHAsMCL
41
5.6 Métodos analíticos
41
5.6.1 pH
41
5.6.2 Masa seca celular
41
5.6.3 Cantidad y composición de PHA
42
5.7 Manipulación de DNA
43
5.7.1 Extracción de DNA plasmidial y genómico
43
5.7.2 Digestión de DNA plasmidial y genómico
43
5.7.3 Electroforesis en gel de agarosa
44
5.7.4 Ligación de DNA
44
5.8 Inserción de DNA
44
5.8.1 Transformación por choque térmico
44
5.8.2 Transformación por electroporación
45
5.9 Recolección de la información
46
6. Resultados y discusión
47
6.1 Construcción de la biblioteca y aislamiento de mutantes
deficientes en el acumulo de PHAMCL utilizando
transposon mini Tn5
47
6.2 Selección y aislamiento de mutantes deficientes en el acumulo de PHAs
47
6.3 Evaluación de la producción de PHAsMCL
53
6.4 Identificación de los Genes afectados en los mutantes 66

X
7. Conclusiones
68
8. Recomendaciones
69
9. Bibliografía
71
10. Anexos
79
10.1 Anexo 1: Composición medio mineral y de acumulo
79
10.2 Anexo 2: Composición caldo nutritivo
80
10.3 Anexo 3: Composición medio Luria Bertani
81
10.4 Anexo 4: Composición Buffer de transformación
82

XI
LISTA DE TABLAS
TABLA
Página
Tabla 1 Cepas bacterianas y plásmidos utilizados 37
Tabla 2 Perfil de acumulo de los mutantes parciales de P.
putida.
49
Tabla 3 Producción de PHAMCL por mutantes de Pseudomonas putida a partir de glucosa.
53
Tabla 4 Producción de PHAMCL por mutantes de
Pseudomonas putida a partir de acido octanóico.
55
Tabla 5 Producción de PHAs por mutantes parciales de
Pseudomonas putida a partir de glucosa
60
Tabla 6 Producción de PHAs por mutantes parciales de
Pseudomonas putida a partir de acido octanóico
63

XII
LISTA DE FIGURAS
Página
Figura 1 Fórmula general de monómeros detectados en PHA.
20
Figura 2 Genes de biosíntesis de PHAMCL en P. oleovorans
23
Figura 3 Organización estructural del granulo de PHA e Interconexiones metabólicas entre las diferentes vías involucradas en la biosíntesis y catabolismo de PHBs y PHAs.
25
Figura 4 Fotografía microscopia electrónica de Pseudomonas putida, evidenciando presencia de gránulos de acumulo.
28
Figura 5 Esquema de inserción del transposon mini Tn5
39
Figura 6 Foto mutante Lfm 814 deficiente en el acumulo de PHAMCL a partir de glucosa en MMGLU.
51
Figura 7 Foto mutante Lfm 814 deficiente en el acumulo de PHAMCL a partir de glucosa en MMOCT.
51
Figura 8 Foto mutante Lfm 822 deficiente en el acumulo de PHAs a partir de Glucosa.
52
Figura 9 Foto mutante Lfm 824 deficiente en el acumulo de PHAMCL a partir de glucosa y octanóato.
52
Figura 10 Producción de PHA por mutantes de Pseudomonas putida a partir de glucosa.
54
Figura 11 Producción de PHA por mutantes de Pseudomonas putida a partir de acido octanóico.
55
Figura 12 Comparación de producción de PHA por mutantes parciales de Pseudomonas putida en diferentes tiempos a partir de glucosa.
61
Figura 13 Comparación de monómeros de PHA producidos por mutantes parciales de Pseudomonas putida en diferentes tiempos a partir de glucosa.
62
Figura 14 Comparación de producción de PHA por mutantes parciales de Pseudomonas putida en diferentes tiempos a partir de acido octanóico.
64
Figura 15 Comparación de monómeros de PHA producidos por mutantes parciales de Pseudomonas putida en diferentes tiempos a partir de acido octanóico.
65

RESUMEN
Los polihidroxialcanoatos (PHA) son poliésteres producidos por diversas
bacterias y acumulados bajo la forma de gránulos intracelulares de
reserva de carbono, energía o equivalentes reductores, pudiendo
representar hasta un 80% de la masa seca celular; estos polímeros son
acumulados de forma mas expresiva bajo condiciones desbalanceadas de
crecimiento, cuando hay exceso de fuente de carbono disponible y
limitación de por lo menos un nutriente esencial como el nitrógeno. La
biosíntesis de polihidroxialcanoatos depende principalmente del
microorganismo y de la fuente de carbono proporcionada, diferentes vías
metabólicas están relacionadas con la formación de moléculas de
hidroxiacil-CoA, principal precursor de los PHA; La mayoría de los
intermediarios de PHAMCL son obtenidos por el metabolismo de ácidos
grasos, aunque otros monómeros (sintetizados a partir de diferentes
fuentes de carbono) pueden ser obtenidos por medio de diferentes vías
bioquímicas. Pseudomonas putida ha sido descrita en diversos trabajos,
por su amplia capacidad de sintetizar PHAMCL, entre los genes que están
involucrados en la síntesis de este biopolímero se destacan el gen phaC
que codifica para la PHA sintasas, el gen phaZ que codifica para las
despolimerasas, el gen phaG codifica a la enzima que enlaza la
biosíntesis de ácidos grasos de novo y la síntesis de PHA y finalmente los
genes phaJ, FabG, entre otros, responsables de la transferencia de
intermediarios de la β- oxidación de ácidos grasos a la síntesis de PHA.
En este trabajo se obtuvieron mutantes de Pseudomonas putida
deficientes en el acumulo de PHAMCL a través de inserción del transposon
miniTn5; Se obtuvo un mutante deficiente en el acumulo de PHAs a partir
de glucosa y acido octanóico, estando posiblemente afectado en el gen
phaC, un segundo mutante afectado solamente en el acumulo a partir de
carbohidratos, indicando que posiblemente fue afectado en el gen phaG y
una serie de mutantes parciales posiblemente afectados en algún gen
involucrado en la síntesis de enzimas responsables de la transferencia de
intermediarios de la β – oxidación de ácidos grasos a la síntesis de PHA.

ABSTRACT
Polyhydroxyalkanoates (PHA) are polyesters produced by several
bacterias they are accumulated under the form of intracellular granules as
carbon, energy or equivalent reducers storage, being able to represent
until 80% of the cellular dry mass; these polymers are accumulated in a
more expressive way under unbalance growth conditions, when there is
an excess of source of available carbon and limitation of at least one of
essential nutrient as nitrogen. Polyhydroxyalkanoates biosynthesis
depends mainly on the microorganism and of the source of carbon
proportionate, different metabolic pathways are related with the formation
of hidroxiacil-CoA molecules, main precursor of the PHA. Most of the
intermediates of PHAMCL are obtained by the metabolism of fatty acids,
although other monomers (synthesized from different carbon sources) can
be obtained from different biochemical pathways. Pseudomonas putida
has been described in diverse works, for its wide capacity to synthesize
PHAMCL, among the genes that are involved in the synthesis of this
biopolymer the gene phaC that codes for the PHA synthases, the gene
phaZ that codes for the depolymerases, the gene phaG that codes to the
enzyme that connects the biosynthesis of fatty acids of novo and the
synthesis of PHA and finally the genes phaJ, FabG stand out among
others, responsible for the transfer of intermediates of the β - oxidation of
fatty acids to the synthesis of PHA. In this work mutants of Pseudomonas
putida affected in the accumulation of PHAMCL, through insert of the
transposon miniTn5 were obtained. A mutant affected in the accumulation
of PHAs from glucose and octanoic acid, being possibly affected in the
gene phaC, a second mutant only affected in the accumulation from
carbohydrates, indicating that it was possibly affected in the gene phaG
and a series of partial mutants possibly affected in some gene involved in
the synthesis of enzymes responsible for the transfer of intermediates of
β-oxidation of fatty acids to the synthesis of PHA.

15
1. INTRODUCCIÓN
En la actualidad el uso de plásticos se ha convertido en una necesidad
del día a día del hombre moderno, llegando a tener múltiples y variadas
aplicaciones en diferentes ámbitos de la sociedad. Esto sumado al
crecimiento desmesurado de la población mundial y los altos niveles de
consumo que se llega a tener de este producto junto con la falta de
conciencia y conocimiento del daño que puede llegar a causar; ha
generado un gran deterioro en el planeta debido a la contaminación
ambiental causada por el acumulo de grandes cantidades de este
plástico no biodegradable, en forma de basura alrededor de todo el
mundo.
Es ahí cuando surge el interés por parte del gobierno, industria y grupos
ambientalistas, en hallar una solución viable para detener el problema. En
respuesta a esto nace la necesidad de buscar y/o desarrollar materiales
biodegradables que cumplan las exigencias del mercado así como las
especificaciones y funciones que cumplen los tradicionales plásticos sin
causarle daño al medio ambiente.
Los polihidroxialcanoatos (PHAs) han demostrado ser una excelente
alternativa para ello; siendo polímeros de fácil biodegradabilidad,
sintetizados comúnmente en la naturaleza por microorganismos y con
muchos tipos de aplicaciones en la industria de embalajes, medicina,
farmacia y agricultura, debido a la gran variedad de monómeros que
puede contener en sus cadenas, los cuales le confieren grandes
propiedades físicas y termoplásticas. Además de esto pueden ser
sintetizados a partir de sustratos renovables y de bajo costo, como
sustratos provenientes de material de desecho de la industria agrícola,
tales como bagazo de caña de azúcar, aceites vegetales, lactosa
hidrolizada, entre otros.

16
Debido a ello en las ultimas cinco décadas los PHAs se han convertido en
objeto de estudio de varias industrias y grupos de investigación alrededor
del mundo; interesados en adquirir el conocimiento suficiente y desarrollar
estrategias útiles para su producción. Actualmente se tiene conocimiento
de la composición de los monómeros que forman de estos polímeros,
los microorganismos capaces de producirlos, las vías bioquímicas de
biosíntesis, los sustratos adecuados a proporcionar al microorganismo
para obtener el polímero deseado y las enzimas involucradas en la
biosíntesis a partir de diferentes sustratos así como también se conocen
algunos de los genes implicados en la conformación de la maquinaria
metabólica necesaria para la síntesis de PHAs en varios
microorganismos.
Este estudio pretende contribuir aportando conocimientos acerca de los
genes involucrados en las vías de biosíntesis de PHA MCL a partir de
carbohidratos y ácidos grasos como fuentes de carbono; encontrando los
genes que intervienen en la síntesis de enzimas que participan en la
trasferencia de intermediarios de la ruta β-oxidativa de ácidos grasos a la
vía de síntesis de PHAs, o revelando y confirmando la presencia de
algunos genes involucrados en las vías biosinteticas, que han sido citados
en estudios previos a este. Esto mediante la inducción de mutaciones en
el genoma de Pseudomonas putida, conocida como una bacteria capaz
de producir PHAs MCL eficazmente, buscando la inactivación de los
genes anteriormente nombrados en los mutantes obtenidos, incapaces
ahora de producir polihidroxialcanoatos a partir de las fuentes de carbono
testadas; los cuales servirán como recurso de análisis para cumplir
nuestro objetivo.

17
2. MARCO TEÓRICO 2.1 Bioplásticos Los biomateriales son productos naturales que son sintetizados y
catabolizados por diferentes organismos a los cuales se les ha
encontrado amplias aplicaciones biotecnológicas. Ellos pueden ser
asimilados por muchas especies (biodegradables) y no causan efectos
tóxicos en el huésped (biocompatible), confiriéndoles una considerable
ventaja con respecto a otros productos sintéticos convencionales (Luengo
et al,. 2003).
Los bioplasticos son un tipo especial de biomaterial. Ellos son poliésteres,
producidos por una serie de microorganismos, cultivados bajo diferentes
condiciones ambientales y nutricionales (Madison & Huisman, 1999).
Estos polímeros, que son usualmente producidos en la naturaleza, son
acumulados como materiales de reserva (en forma móvil, Amorfa,
gránulos líquidos), permitiendo a los microorganismos sobrevivir bajo
condiciones de estrés (Sudesh et al., 2000).
El número y tamaño de los gránulos, la composición del monómero, la
estructura macromolecular y las propiedades físico – químicas varían
dependiendo del organismo productor (Anderson & Dawes, 1990).
La microscopia de fuerza atómica y la espectroscopia confocal Ramal son
técnicas actualmente usadas para el análisis de los gránulos de PHA.
Los bioplasticos pueden ser aislados por centrifugación o por extracción
por medio de solventes con cloroformo, trifluoroetanol, dicloroetano,
propileno carbonato, clorídeo de metileno o por acido di cloroacético. Sus
pesos moleculares (alcanzando de 50 000 a 1 000 000 Da) han sido
establecidos por dispersión de luz, cromatografía en gel, análisis de
sedimentación y por medidas de viscosidad intrínseca, y la composición
de los monómeros ha sido determinada por cromatografía de gases,
espectroscopia de masas y resonancia magnética nuclear (Di Lorenzo &
Silvestre, 1999).
La mayoría de los bioplasticos conocidos contienen, como monómeros,
diferentes intermediarios de la β – Oxidación [ (S) – 3- hidroxiacil – CoAs]

18
, los cuales son enzimaticamente polimerizados por la condensación del
grupo carboxilo, presente en el thioester CoA monomérico con el grupo 3
– Hidroxi (o el grupo tiol) del siguiente ( Zinn et al., 2001).
Otros bioplasticos, contienen monómeros inusuales (ácidos 4,5,6-
hidroxialcanoicos), los cuales son sintetizados a través de diferentes vías
metabólicas, sugiriendo esto que las enzimas biosinteticas están
ampliamente distribuidas, y que la producción de un tipo particular de
poliéster es un evento especifico de cada cepa (Di Lorenzo & Silvestre,
1999)
Lemoigne fue el primero en describir un bioplastico - poli(3-
hidroxibutirato) (PHB) en Bacillus megaterium; esta observación inicial fue
casi olvidada hasta la mitad de la década de 1970, cuando debido a la
crisis de el petróleo, un movimiento científico trato de descubrir fuentes
alternativas al combustible fosil; hoy en día ya han sido establecidas las
estructuras, vías de biosíntesis y aplicaciones de muchos de estos
bioplasticos (Luengo et al., 2003).
2.2 PHAs
Los polihidroxialcanoatos (PHA) son poliésteres producidos por diversas
bacterias y acumulados bajo la forma de gránulos intracelulares de
reserva de carbono, energía o equivalentes reductores, pudiendo
representar hasta un 80% de la masa seca celular (Anderson & Dawes,
1990).
Los polihidroxialcanoatos (PHA) representan una compleja clase de
poliésteres que son sintetizados por la mayoría de géneros bacterianos y
miembros de la familia Halobacteriaceae de Archea. Muchos de estos
procariotas sintetizan poli(3- hidroxibutirato), poli (3HB), y otros PHAs
como componentes de reserva y depositan estos poliesteres como
inclusiones insolubles en el citoplasma (Rehm et al., 1999).
Después de haber sido detectado poli(3HB) en un amplio rango de
especies bacterianas, todo lo referente a el y a la biosíntesis de
poliésteres ha experimentado un dramático desarrollo durante la última

19
década (Steinbüchel et al.,1996). Los poli(3HB) fueron detectados
también en organismos eucariotes e incluso en humanos; mas de 150 o
sea diferentes ácidos hidroxialcanoicos son ahora conocidos como
constituyentes de PHA bacteriano, esta variedad de constituyentes y la
posibilidad de tener diferentes combinaciones permite la biosíntesis de un
gran numero de diferentes poliésteres. Las rutas de biosíntesis para
muchos de estos poliésteres a partir de diferentes sustratos relacionados
y no relacionados han sido investigadas detalladamente. Así mismo los
genes involucrados de la biosíntesis de PHA , provenientes de muchas
bacterias han sido clonados y analizados a nivel molecular con el fin de
conocer cual es su papel (Steinbüchel et al.,1996).
Además de los ácidos nucleicos, proteínas, polisacáridos, polifosfatos,
polisoprenoides, los PHA han sido referidos como otra clase de
biopolímeros con relevancia fisiológica; junto con la lignina, el cual
representa uno de los biopolímeros más abundantes en la naturaleza, los
PHA representan la séptima clase de biopolímeros con relevancia
fisiológica (Müller & Seebach, 1993).
Estos polímeros son acumulados de forma mas expresiva bajo
condiciones desbalanceadas de crecimiento, o sea cuando hay exceso de
fuente de carbono disponible y limitación de por lo menos un nutriente
esencial para la multiplicación celular (Brandl et al., 1990).
Actualmente cerca de 150 monómeros diferentes han sido identificados
como constituyentes de PHA producidos por bacterias a partir de diversas
fuentes de carbono (Rehm & Steinbüchel, 1999, Rehm, 2003). Los
monómeros constituyentes de PHA son clasificados en dos grandes
grupos (Steinbüchel & Valentin, 1995): (1) Aquellos de cadena corta
(HASCL – “HydroxyAcids of Short-Chain-Length”), conteniendo de 3 a 5
atómos de carbono en la cadena principial, y (2) aquellos de cadena
mediana (HAMCL – “HydroxyAcids of Medium-Chain-Length”), conteniendo
de 6 a 16 átomos de carbono en la cadena principal. En base a esto los
PHA pueden ser entonces clasificados como : (1) PHASCL, compuestos
por HASCL y (2) PHAMCL , formados por HAMCL.

20
En cuanto se refiere a la capacidad de síntesis microbiana, mientras los
PHASCL son encontrados en bacterias pertenecientes a los mas diversos
grupos taxonómicos (Steinbüchel, 1991), los PHAMCL solo han sido
detectados en Pseudomonas de el grupo I de homologia de RNA
ribosomal (Pseudomonas sensu stricto) (Huisman et al., 1989).
Una formula general para los diversos monómeros observados como
constituyentes de PHA es representada en la figura 1.1 a continuación :
Figura 1. Fórmula general de monómeros detectados en PHA.
2.3 Síntesis de Polihidroxialcanoatos Las vías metabólicas de síntesis de los polihidroxialcanoatos de cadena
corta y de cadena media han sido bastante estudiadas (Steinbuchel &
Eversloh, 2003)
El principal intermediario para la biosíntesis de polihidroxialcanoatos de
cadena corta (PHASCL) es el acetil-CoA, proveniente principalmente de la
oxidación de diferentes fuentes de carbono como carbohidratos, ácidos
orgánicos y alcoholes; la producción de polihidroxialcanoatos a partir de
carbohidratos (sacarosa, glucosa, y fructosa) esta compuesta por tres
importantes enzimas clave (Rehm, B. 2003)
La primera enzima perteneciente a la vía biosintetica de los
polihidroxialcanoatos es la β-cetotiolasa, la cual cataliza la condensación
de dos moléculas de acetil-CoA para formar acetoacetil-CoA; Esta enzima
es codificada por el gen phaA.
El siguiente paso es la reducción del acetoacetil-CoA a (R)-3-
Hidroxibutiril-CoA catalizada por la acetoacetil-CoA reductasa
dependiente de NADPH. La expresión de esta enzima es codificada por
el gen phaB (Suriyamongkol et al., 2007).

21
La principal enzima señalada en la síntesis de PHA es denominada PHA
sintasa, responsable por la polimerización de unidades de (R)-3-
hidroxiacil-CoA, formando el polihidroxialcanoato (Steinbuchel, 1996). Ella
es absolutamente estereoespecifica para los esteroisomeros (R) y su
especificidad por el sustrato definirá la naturaleza del polímero final. Esta
enzima puede ser encontrada en los gránulos de PHA durante el
acumulo; las moléculas de hidroxiacil CoA servirán de sustrato para esta
enzima, codificada por el gen phaC (Rehm et al., 1999).
Las PHA sintasas son ampliamente estudiadas y pueden ser clasificadas
a grandes rasgos en cuatro tipos. Las PHA sintasa de tipo I, son
especificas para monómeros de ácido hidroxialcanoico de cadena corta
(C3 a C5), mientras que las de tipo II son especificas para la
polimerización de monómeros de ácido hidroxialcanoico de cadena
media (C6 a C13). El gen que codifica estas dos clases es denominado
phaC. Las PHA sintasas de tipo III son caracterizadas por la presencia
de dos subunidades (I) la PhaC, que presenta semejanza en la
secuencia de aminoácidos igual a 28% con las PhaC de las clases I y II;,
y (II) PhaE. Las PHA sintasas de tipo IV son semejantes a las de tipo III,
con la diferencia que el PhaE es sustituido por PhaR. El tamaño de la
cadena polimérica a ser formada es dependiente del tipo de PHA sintasa
en cada microorganismo productor (Rehm, B. 2003).
2.4 Biosíntesis de PHA a partir de sustratos relacionados y no
relacionados
La incorporación de diferentes monómeros depende principalmente del
microorganismo y de la fuente de carbono utilizada. Diferentes vías
metabólicas, presentes en los microorganismos son responsables por la
formación de diferentes moléculas de hidroxiacil-CoA (Gomez et
al.,1996).
Existen dos grandes grupos de sustratos utilizados en la biosíntesis
de estos biopolímeros: los sustratos relacionados y los sustratos no

22
relacionados. (Steinbuchel, 1996). La síntesis a partir de estratos
relacionados se denomina de esta forma ya que la composición del
polímero producido refleja claramente el sustrato proporcionado; este tipo
de síntesis es la que permite la incorporación de mas diversidad de
monómeros al polímero. En la síntesis a partir de sustratos no
relacionados, como es el caso de diversas Pseudomonas sp, cuando son
cultivadas en presencia de glucosa, fructosa, glicolato, acetato, glicerol,
etc., producen un polimero conteniendo 3- hidroxidecanoato (3HD) como
principal constituyente, además del 3-hidroxioctanoato (3HO), 3-
hidroxihexanoato (3HHx), 3-hidroxidodecanoato (3HDd), 3-
hidroxitetradecanoato (3HTd), 3-hidroxi-5-cis-dodecenoato (3HDd∆ 5) y 3-
hidroxi-7-cis-tetradecenoato (3HTd∆7) como constituyentes secundarios.
Las síntesis de PHAMCL a partir de sustratos relacionados y no
relacionados pueden ser claramente distinguidas por las vías metabólicas
que proporcionan los intermediarios. Mientras que en la síntesis a partir
de sustratos relacionados la β-oxidación de ácidos grasos es la principal
vía para suministrar intermediarios, en la síntesis a partir de sustratos no
relacionados la biosíntesis de ácidos grasos suple los intermediarios
(Gomez, 2000)
Comparados con la biosíntesis a partir de sustratos relacionados, no
existen muchos trabajos que utilicen los sustratos no relacionados para la
producción de polihidroxialcanoatos; aunque en los últimos años se ha
observado un aumento significativo en el empleo de fuentes de carbono
simples y de bajo costo para la producción de polímeros biodegradables
con estructura diferente de la fuente de carbono empleada (Rocha, 2007).
2.5 Biosíntesis de Polihidroxialcanoatos, Catabolismo y Regulación La organización de los genes de biosíntesis de PHAMCL en Pseudomonas
oleovorans se muestra en la figura 2. El operon phaC1ZCD codifica dos

23
polimerasas (PhaC1 y PhaC2), la despolimerasa (PhaZ) y la proteína
PhaD (Steinbuchel, 2001).
Fuente: (Luengo et al., 2003).
Figura 2. Genes de biosíntesis de PHAMCL en P. oleovorans.
Las dos polimerasas son miembros de la subfamilia de las α \ β
hidrolasas, catalizan la condensación en PHAS de varios derivados (R)-3-
hidroxi-acil-CoA (saturados, insaturados, lineales, cíclicos, ramificados o
sustituidos con diferentes grupos funcionales como átomos alógenos,
hidróxidos, ciano, carboxi, o grupos fenilo) cuyas cadenas alcanzan a
tener entre C5 y C14 átomos. Ambas enzimas son completamente
similares en su secuencia de aminoácidos (cerca del 50%) y en
especificidad de sustrato (derivados de 3-OH-acyl-CoA y 4-,5- o 6-
OH-acyl-CoAs), aunque cuando se expresa en un huésped ajeno, ellas
son capaces de polimerizar otros monómeros. La identificación de los
residuos de aminoácidos requeridos para la catálisis ha permitido la
modificación de la velocidad de catálisis en varias PHA sintasas.
La mayoría de los intermediarios de PHAMCL son obtenidos por β –
oxidación de ácidos grasos, aunque otros monómeros (sintetizados a
partir de diferentes fuentes de carbono) pueden ser obtenidos por medio
de diferentes vías bioquímicas (Figura 3). El uso de una u otra parece ser
característico de cada cepa y del sustrato proporcionado (Garcia et al.,
1999).
Un diferente tipo de polimerasa (tipo III) existe en Cromatium vinosum,
Thiocystis violacea, Thiocapsa pfennigii y Synechocystis sp. PCC 6803
(Sudesh et al., 2000). Esta es formada por PhaC y PhaE, los cuales
principalmente sintetizan PHAS SCL pero también polimerizan monómeros
SCL y MCL.

24
La existencia de dos polimerasas en el mismo microorganismo
(probablemente consecuencia de la duplicación de genes) representa un
evento interesante en la evolución que podría haber contribuido a la
transición bioquímica de PHBS (el cual solo requiere una sola polimerasa)
a PHAMCL (donde dos enzimas están involucradas). Aunque se necesitan
mas estudios para confirmar esta hipótesis (Luengo et al., 2003).
Los genes phaC1 y phaC2 están separados por un tercer gen que codifica
el producto PhaZ (figura 2). Esta proteína, contiene la convencional
región lipasa box, lo cual muestra cierta homologia con las PHA
despolimerasas (enzimas involucradas en la movilización de PHA SCL) y
con un gran número de enzimas hidroliticas, sugiriendo que estas
participan en la liberación de derivados hidroxiacil – CoA provenientes de
PHAS. La localización topológica de PhaZ (superficie del granulo), y la
incapacidad de ciertas bacterias para movilizar PHAS MCL cuando phaZ es
mutado apoya firmemente su función fisiológica.

25
Figura 3. Fuente: (Luengo et al., 2003).
Organización estructural del granulo de PHA e interconexiones metabólicas entre las diferentes vías involucradas en la biosíntesis y catabolismo de PHBs y PHAs. (a) via de oxidación de alcanos. (1) Alcano 1- monooxigenasa. (2) Alcohol deshidrogenasa. (3) aldehido deshidrogenasa. (b) ß- oxidación de acidos grasos (4) Acil – CoA ligasa (5) Acil-CoA deshidrogenasa, (6) Enoil-CoA hidratasa, (7) 3- hidroxiacil- CoA deshidrogenasa, (8) 3- cetiolasa, (9) (R)- enoil-CoA hidratasa, (10) 3- cetoacil- CoA reductasa. (c) Biosintesis a partir de carbohidratos. (11) ß- cetiolasa, (12) NADPH- dependiente acetoacil-CoA reductasa. (d) sintesis de acidos grasos de novo. (13) acetil- CoA carboxilasa, (14) ACP- maloniltransferasa, (15) 3- cetoacil-ACP sintasa, (16) 3- cetoacil-ACP reductasa, (17) 3- hidroxiacil-ACP reductasa, (18) enoil-ACP reductasa, (19) 3- hidroxiacil-ACP-CoA transcilasa.

26
2.6 Biosíntesis de PHAMCL y Metabolismo de ácidos grasos El metabolismo de ácidos grasos representa una de las vías metabólicas
más comunes para suplir monómeros de hidroxialcanoato (HA) a la
síntesis de PHA (Sudesh et al., 2000). Los intermediaros generados
mediante la degradación de ácidos grasos vía β- Oxidación pueden
proveer hidroxialcanoil- CoA (HA-CoA) como sustrato para mcl-PHAs.
Los intermediarios de la β- Oxidación de ácidos grasos incluyen el enoil-
CoA, 3-cetoacil-CoA y el (S)-3-hidroxiacil-CoA, pueden servir como
precursores de mcl-(R)-3-hidroxiacil-CoA, el cual es usado directamente
en la síntesis de PHAMCL (Suriyamongkol et al., 2007).
El gen que codifica para la PHA sintasa (phaC) en Aeromonas caviae,
está flanqueado por phaJ, el cual codifica la enzima hidratasa enoil-CoA,
que cataliza una hidratación (R)- especifica del 2-enoil-CoA
proporcionando unidades monómero de (R)-3-Hidroxiacil-CoA para la
síntesis de PHA a través de la β-oxidación de ácidos grasos (Fukui & Doi,
1997).
Una segunda ruta para la síntesis de PHAMCL en bacterias es através del
uso de intermediarios de la biosíntesis de ácidos grasos de novo, la cual
provee monómeros HA-ACP. En contraste a P. oleovorans y P. fragii ( los
cuales usan intermediarios de la β- Oxidación), P. aeruginosa y P.putida
producen PHAMCL cuando crecen a partir de sustratos no relacionados,
tales como la glucosa. Esto es porque P. oleovorans y P. fragii usan los
ácidos grasos como fuente de carbono mediante la β- Oxidación para
producir 3-Hidrociacil-CoA, sustrato de PHAMCL sintasa (Suriyamongkol et
al., 2007).
Por otro lado la biosíntesis de ácidos grasos es la principal ruta para la
síntesis de 3-Hidroxiacil-CoA en P. aeruginosa y Pseudomonas putida
durante su crecimiento a partir de fuentes de carbono que son
metabolizadas a acetil-CoA, como carbohidratos, acetato o etanol. El gen
phaG codifica a la enzima que enlaza la biosíntesis de ácidos grasos de
novo y la síntesis de PHA; esta ha sido clonado de P. putida (Rehm et al.,
1998; Hoffmann et al., 2000a,b).

27
El producto de este gen cataliza la conversión de (R)-3-hidroxiacil-ACP,
intermediario de la vía de biosíntesis de ácidos grasos a su
correspondiente derivado CoA.
Se ha demostrado según estudios, que la expresión de phaG en E. coli da
a la bacteria recombinante la nueva capacidad de producir ácidos grasos-
mcl libres como precursores 3(HD) (Zheng et al., 2004) y su expresión en
P. oleovorans y P. fragii produce PHAMCL (Fiedler et al., 2000; Hoffmann
et al., 2000b), a partir de fuentes de carbono no relacionadas a la
estructura 3HD, tales como la glucosa y la fructosa.

28
2.7 Arquitectura macromolecular de las Inclusiones
PHAs son acumulados intracelularmente (como polímeros amorfos o
móviles) en gránulos de diferentes tamaños (Figura 4), ellos están
rodeados por una monocapa fosfolipídica, que contiene PHAsinas PhaF y
PhaI, polimerasas, despolimerasas y proteínas citosolicas no especificas
ligadas a el granulo (Figura 3). PhaI participa en la formación y
estabilización de los gránulos, mientras que PhaF está involucrada en la
estabilización de los gránulos y actúa como un regulador (Prieto et al.,
1999). La función de la monocapa fosfolipídica de envoltura aún no ha
sido bien establecida, aunque se cree que es necesaria para el contacto
de PHAS con el agua (previniendo la transición del poliéster del estado
liquido amorfo a una forma cristalina mas estable), y también desempeña
un papel como barrera protectora (evitando daño celular causado por la
interacción de PHAS con las estructuras internas celulares o con las
proteínas citosolicas). Si la monocapa fosfolipídica es en realidad
requerida para proteger las células desde el comienzo de la formación del
granulo, puede ser asumido que esta envoltura puede ser extendida
alrededor del granulo tan larga como se incremente el tamaño del
granulo. Por ello deben existir enzimas específicamente involucradas en
la síntesis de la monocapa fosfolipídica, asociadas con el poliéster
(Luengo et al., 2003).
Figura 4. Fotografía microscopia electrónica de Pseudomonas putida evidenciando presencia de gránulos de acumulo .Fuente: (Luengo et al., 2003).

29
2.8 Aplicaciones Biotecnológicas de los Polihidroxialcanoatos
Durante la década de 1980s la compañía británica Imperial Chemicals
Industries (ICI), desarrollo un proceso comercial para producir Poly(3HB),
y un copolímero relacionado conocido como poli-3-hidroxibutirato-co-R-3-
hidroxivalerato, poli(3HB-co-3HV). Estos polímeros fueron vendidos bajo
el nombre comercial Biopol®, y fueron desarrollados principalmente como
remplazo renovable y biodegradable para plásticos derivados del petróleo
(Williams & Martin, 2008).
En sus inicios, este tipo de material fue usado para la fabricación de
botellas, fibras, látex y varios productos de interés agrícola y comercial.
En 1990, fue lanzado comercialmente a partir de PHAs (P3HB-co-3HB),
una botella biodegradable para shampoo de una compañía alemana de
cosméticos (Preusting,1992). Desde entonces diversas aplicaciones a
pequeña escala han sido desarrolladas para P3HB o P3HB-co-3HV.
Aunque algunas aún no se encuentran disponibles comercialmente, una
serie de aplicaciones potenciales han sido vislumbradas para PHAMCL.
Babu et al., 1997, evaluaron el uso de PHAsMCL como componentes de
adhesivos sensibles a la presión. Formulaciones conteniendo PHAs
producidos por P. oleovorans a partir de octanoato (P3HHx-co-3HO) no
presentaron las propiedades esperadas. En cambio, formulaciones
basadas en PHAs producidos por P. oleovorans a partir de nonanoato
o a partir de mezclas de nonanoato o 10-undecenoato con octanoato
presentaron un buen desempeño en características especificas para este
tipo de aplicación, con valores dentro del rango presentado por adhesivos
sensibles a la presión en uso comercial (Babu et al., 1997).
PHAs producidos por P. putida a partir de ácidos grasos derivados de
aceite de linaza fueron utilizados como agentes ligantes en formulaciones
de tintas, que permitieron la sustitución de solventes orgánicos por agua,
posibilitando el desarrollo de tintas no agresivas en el medio ambiente.
Las tintas producidas presentaron un buen desempeño cuando fueron

30
evaluadas con respecto a las características específicas para estos
materiales (Van der Walle et al., 1999; Buisman et al., 2000)
En los últimos años el interés por los polihidroxialcanoatos ha venido
creciendo notablemente debido a sus diversas propiedades físicas,
químicas y mecánicas.
Actualmente, en Japón, la empresa Mitsubishi produce P3HB a partir de
metanol con el nombre de Biogreen®, en Brasil la empresa PHB Industrial
produce y comercializa este mismo producto a partir de sacarosa extraída
de caña de azúcar. Ambas empresas lo producen a pequeña escala,
aproximadamente 100 toneladas por año (Rocha, 2007).
Los polihidroxialcanoatos también pueden serempleados en diferentes
áreas; tales como medicina, industrias químicas y farmacéuticas.
(Hrabak,1992) .
Alrededor del mundo 14 compañías trabajaron o trabajan en PHA. En los
últimos años, 10 empresas chinas comenzaron a prestar atención a los
PHA. Entre ellos ZheJiang TianAnha estado produciendo un copoliester
de 3- hidroxibutirato y 3- hidroxivalerato, denominado PHBV, alcanzando
a producir cerca de 1000 toneladas al año; Otras compañías están
desarrollando procesos de producción a escala piloto de varios PHAs,
buscándole dar un alto valor agregado a las diferentes aplicaciones que
se le pueden lograr dar con el debido proceso de producción a estos
polímeros (Chen, 2008).
2.9 Polihidroxialcanoatos en el área medica
Por muchos años la investigación en el área farmacéutica fue focalizada
en el descubrimiento y síntesis de nuevas sustancias con actividad
biológica. En tanto, en las dos últimas décadas, creció el interés por el
desarrollo de nuevos métodos de administración de drogas. La aparición
de nuevos grupos de agentes terapéuticos como los ácidos nucleicos, los
polipéptidos y hormonas de crecimiento ha impulsado la investigación en
sistemas de liberación controlada por tratarse de medicamentos con alto

31
valor agregado, aplicadas en bajas concentraciones. Los primeros
polímeros usados en el desarrollo de microencapsulados para la
liberación de drogas fueron las gomas de silicona y polietileno; aunque,
a pesar de ser biocompatibles, estos materiales no eran biodegradables y
las microcápsulas eran utilizadas con más frecuencia como implantes.
Debido al hecho de que estos materiales pueden dejar residuos tóxicos
en el organismo, era necesario removerlos quirúrgicamente (Desai et al.
1965).
En el área medica los polímeros biodegradables han sido utilizados son el
Homo y el copolímero de lactato (PLA) y glicolato (PGA) , desarrollados
por la Du Pont, y que han sido empleados por la Johnson & Johnson en la
fabricación de hilos quirúrgicos para suturas reabsorbibles por el
organismo (Chulia et al., 1994).
Steinbüchel et.at. 1996 informo en su estudio que el acido 3-
Hidroxibutírico puede ser encontrado en el plasma humano por tratarse
de un polímero biocompatible, lo que aumento el interés por este material
en el área medica.
Los polihidroxialcanoatos se presentan entonces como candidatos
alternativos para este tipo de aplicación. Dentro de estos el homopolímero
P3HB y su copolímero P3HB -co- 3HV son los mas citados para uso
farmacéutico, con la ventaja de ser biocompatibles y degradados por
microorganismos y no sintetizados por vía química, presentando así
menos problemas toxicológicos. Como desventaja, en comparación a
los PLA y PGA, presentan menos degradabilidad debido al alto grado de
cristalización (Pouton et al.,1996).
Como resultado de todos los trabajos y productos desarrollados, estos
polímeros llegaron a ser ampliamente usados, lo cual se convirtió
oportunidades para su evolución como biomateriales médicos. Estos
esfuerzos dieron como resultado una serie de pruebas clínicas muy
promisorias, continuando hasta el día de hoy, la búsqueda por desarrollar

32
constantemente productos conteniendo este tipo de materiales para ser
aprovechados en usos médicos in vivo (Williams & Martin, 2008).
En los últimos años los polihidroxialcanoatos, han emergido como una
clase de polímeros biodegradables con distintas aplicaciones en
ingeniería de tejidos. Los PHAs han sido evaluados para la producción de
“scaffolds”, que son piezas diseñadas para proveer una estructura soporte
para las células que están siendo implantadas. Algunas propiedades que
son fundamentales para el uso de un determinado material en la
fabricación de estas piezas, incluyen: (I) Biocompatibilidad, (II) capacidad
para permitir el crecimiento de las células del tejido (III) capacidad para
guiar y organizar el desarrollo de los tejidos de una determinada manera
(IV) difusión adecuada de nutrientes y metabolitos (V) biodegradación,
cuando es necesario, generando productos no tóxicos. Los resultados de
las evaluaciones preliminares de estas propiedades indican que PHAs
tendrán un importante papel en la ingeniería de tejidos y en el desarrollo
de productos de tejidos vivos para aplicaciones terapéuticas (Gomez,
2000).
En la ultima década estos polímeros comenzaron a ser testados para su
empleao como suturas, implantes, ingeniería de tejidos neuro y
cardiovascular, fijación de fracturas, tratamiento de narcolepsia y adicción
al alcohol, vehículos de administración de drogas, microencapsulación
celular, analgésicos, radiopotenciador, preservativos químicos, algunos
presentan propiedades antihelmínticas, antitumorales (aquellos que
contienen monómeros aromáticos o están ligados a nucleósidos)
(Angelova & Hunheler, 1999).
Gracias a todos estos avances, los PHAs demostraron ser un atractivo
material para aplicaciones biomédicas de interés comercial. Diversos
productos de PHA, han sido puestos en venta por algunas compañías,
como TEPHA. (USA), Metabolix. (USA) y P & G. (USA), (Misra et al.,
2006) entre ellos se destacan las suturas quirurgicas, vendajes, soportes
para tejidos, películas, dispositivos de reparación de tejidos, sistemas de

33
placas, mallas quirúrgicas, sistemas de liberación controlada de
compuestos activos, curativos, pos – lubrificantes, usos ortopédicos,
como sustrato pericárdico, implantes e injertos (Chen & Wu, 2005).
2.10 Producción Industrial
Existen principalmente dos limitaciones en la producción en masa de los
bioplasticos: primero, la dificultad implicada en la síntesis del polímero a
partir de precursores de bajo costo y segundo, el alto costo de su
recuperación (Anderson & Dawes , 1990).
Sin embargo, actualmente el conocimiento acerca de las vías de
biosíntesis y su regulación ha permitido la construcción de organismos
recombinantes (otros microorganismos, hongos, plantas) capaces de
sintetizar bioplasticos de fuentes de carbono de bajo costo (molazas,
sucrosa, lactosa, glicerol, aceites y metano). Sin embargo, los procesos
fermentativos que son llevados a cabo con bacterias recombinantes y
plantas transgénicas no son competitivos con los procesos industriales
convencionales de síntesis de plásticos (Steinbüchel et al., 1998).
Una estrategia diferente de producción es síntesis enzimática de
bioplasticos. Aunque su producción en el laboratorio es económicamente
no aconsejable, la caracterización de las enzimas de biosíntesis, tal y
como el conocimiento acerca de los requerimientos energéticos para tales
procesos, podría facilitar su paso a gran escala y por lo tanto la
producción de bioplasticos nuevos o modificados en biorreactores (Lee &
Choi, 1999).

34
3. PLANTEAMIENTO DEL PROBLEMA Y JUSTIFICACIÓN
La búsqueda de productos que puedan remplazar las funciones
comúnmente realizadas por los tan contaminantes plásticos, con la misma
o mayor eficacia y sobre todo sin causarle daño al medio ambiente, ha
despertado el interés por los biomateriales, caracterizados por su fácil
biodegradabilidad al poder ser asimilados por muchas especies cuando
son eliminados y por su biocompatibilidad al no causar efectos tóxicos
sobre la naturaleza y seres vivos.
Los polihidroxialcanoatos son un grupo bastante diversificado de
poliésteres que han despertado el interés científico, tecnológico e
industrial en diversos países, pues además de ser termoplásticos
biodegradables, pueden ser sintetizados por fermentación a partir de
materias primas renovables. Pudiendo ser así, los PHAs excelentes
substitutos de los plásticos convencionales en aplicaciones de descarte
muy rápidas, pues podrían ser depositados en rellenos sanitarios sin
causar dificultades para la degradación de otros materiales constituyentes
de la basura; en los casos de recolecta selectiva de basura, podrían ser
reunidos al restante de la materia prima orgánica y utilizados en la
producción de fertilizantes. Como en la producción de PHAs son utilizadas
materias primas derivadas de la agricultura, por lo tanto rápidamente
renovables, se tendría un sistema cíclico y equilibrado, semejante a los
sistemas naturales (Gomez, 2000).
Gran variedad de bacterias son capaces de acumular poli(3-
hidroxialcanoatos) (PHAs) como reservas de carbono y energía. Debido a
su potencial los PHAs han sido ampliamente estudiados por grupos
académicos e industriales. Pseudomonas sp sintetiza en su mayoría
polihidroxialcanoatos de cadena media (PHAsMCL), los cuales consisten
en monómeros que contienen de 6 a 8 átomos de carbono en su cadena.
Aunque algunos de los PHAs han sido desarrollados, comercializados y
vendidos, en general el uso de estos polímeros ha sido dificultado por los
altos costos de producción. La reducción de estos costos podría ser

35
alcanzada de varios modos, incluyendo el incremento de la productividad
o usando plantas transgénicas para la producción de PHA, propiciando
que los niveles de PHA lleguen de un 20% a 40% del peso seco de la
planta (Klinke et al., 2000).
Por esta razón es de gran importancia realizar estudios que contribuyan al
desarrollo este tipo de polímeros como alternativa económica y
ecológicamente viable a los plásticos comunes.
Este trabajo pretende contribuir aportando conocimientos y herramientas
para el modelaje y producción de PHA, mediante la construcción de una
biblioteca de mutantes de Pseudomonas putida incapaces de acumular
polihidroxialcanoatos a partir de carbohidratos y/o ácidos grasos; los
genes afectados tras la mutación, mas adelante servirán como
herramientas para cumplir tal fin.

36
4. OBJETIVOS
4.1 Objetivo General
• Construir una biblioteca de mutantes por transposon, obtenida a
partir de una cepa salvaje de Pseudomonas putida , afectados en
la biosíntesis de polihidroxialcanoatos a partir de glucosa y/o
octanoato como únicas fuentes de carbono.
4.2 Objetivos Específicos
• Utilizar el transposon miniTn5 para generar una mutación en el
genoma de Pseudomonas putida que inactive algunos de los
genes involucrados en la biosíntesis de PHAs a partir de glucosa y
octanoato como fuentes de carbono.
• Evaluar metabolitamente las cepas de mutantes PHA-negativo
obtenidas, en cuanto a crecimiento, degradación de sustrato y
producción de PHAs.
• Identificar los genes involucrados en las vías metabólicas
responsables de la transferencia de intermediarios del
metabolismo de ácidos grasos para la síntesis de PHAs.

37
5. MATERIALES Y MÉTODOS
5.1 Microorganismos y plásmidos
Las características de cada una de las cepas bacterianas que fueron
utilizadas en este trabajo, se muestran a continuación en la tabla 1.
Tabla 1. Cepas bacterianas y plásmidos utilizados
Bacterias Características Referencia
Pseudomonas putida IPT
046
Cepa salvaje aislada de una
muestra de suelo, capaz de
acumular PHAMCL a partir de
carbohidratos y/o oleos
vegetales.
Gomez, 2000.
Pseudomonas putida IPT
461
Cepa mutante deficiente en
el acumulo de PHAMCL a
partir de Glucosa y
octanoato. Utilizada como
control.
Pseudomonas putida IPT
463
Cepa mutante deficiente en
el acumulo de PHAMCL a
partir de Glucosa.
Utilizada como control.
E. coli S17 -1 λpir
pUTKm – IPT624
Cepa que carga el plásmido
que contiene el transposon
mini Tn5
Lorenzo et al., 1990
E. coli DH5α
E. coli XL1- Blue
SupE44 lacU169(80
lacZM15) hsdR 17 recA 1
endA 11 gyrA 96 thi-1 relA1
Hanahan, 1983
pUC BM20 Vetor de clonación,
Replicon ColE1, Ampr
Böehringer Mannheim
GMBH
pUT Km2 Plasmido suicida que carga
El transposon mini Tn-5
conteniendo el gen
reportero lacZ sin promotor.
Lorenzo et al., 1990

38
5.2 Condiciones de cultivo
Las cepas de Pseudomonas putida y sus mutantes fueron cultivadas a 30º
C en caldo nutriente (Peptona 5 g/L y extracto de carne 3 g/L) o medio
mineral (Anexo 1) descrito por (Ramsay et al., 1990), conteniendo
glucosa o octanoato como fuentes de carbono y kanamicina (50 µg/ml)
en el cultivo de los mutantes. Las cepas de Escherichia coli fueron
cultivadas en medio Luria-Bertani (Sambrook, 1989) suplementado con
kanamicina (50 µg/ml).
5.3 Preservación de las cepas bacterianas
Cada una de las cepas bacterianas a ser preservada fue cultivada en
caldo Luria-Bertani (suplementado con kanamicina para el cultivo de
E.coli) por 24 horas, luego se tomaron 5ml de cada cultivo para ser
diluidos en 5ml de una solución acuosa de Glicerol al 20%. La
suspensión de células en la solución de glicerol fue distribuida en tubos
eppendorf de 1ml con (500µL por tubo), los cuales fueron mantenidos en
un congelador de refrigerador domestico por 2 horas para finalmente ser
congelados en freezer a -80ºC.
Para la preservación de las cepas bacterianas por liofilización, 40 ml de
cultivo en caldo nutriente (24 horas) se llevaron a centrifugar, luego las
celulas resultantes fueron resuspendidas en 2 ml de solución acuosa de
leche desnatada (10%) y glutamato de sodio (5%). La suspensión de
células en el lioprotector fue distribuida en ampollas de vidrio (0,2 ml por
ampolla), congelada lentamente hasta -35 °C y luego liofilizada en el
equipo de liofilización Labconco, modelo 5-75050 (Silva et al., 1992). Las
ampollas fueron cerradas al vacío y almacenadas en refrigerados
domestico a 4°C.

39
5.4 Construcción de la biblioteca y aislamiento de mutantes
deficientes en el acumulo de PHAMCL utilizando transposon Mini- Tn5
La inserción del transposon fue realizada através de conjugación de la
cepa de Pseudomonas putida IPT 046 con una E. coli S17-1 donadora,
que carga el plásmido suicida pUT Km2 el cual contiene el transposon
mini Tn5 (Lorenzo et al., 1990) el gen reportero lacZ sin promotor y el gen
de resistencia a la kanamicina (Figura 5).
Figura 5. Esquema de inserción del transposon mini Tn5. Fuente: Braz,
2006.
Para ello cada cepa fue estriada e incubada deacuerdo a sus condiciones
óptimas de crecimiento durante 48 horas, P. putida se cultivo en agar LB
a 30 °C y E.coli S17-1 conteniendo el transposon, fue cultivada en LBK a
37°C.
Posteriormente fue realizado un inoculo de cada cepa en 25 ml de medio
liquido de LB y LBK respectivamente durante 24 horas; transcurrido el
tiempo de incubación se procedió a realizar filtración por membrana
utilizando 10 ml de cada cultivo, colocando 5 ml de cada uno
escalonadamente para ser filtrados juntos en una membrana de 0,45 µm
de poro. Esta membrana que contiene las células fue colocada en una
placa de medio LB e incubada a 30°C por 24 horas para así propiciar la
conjugación entre las dos cepas. Pasadas las 24 horas, la membrana fue
lavada para obtener una suspensión celular en solución salina 0,85%,
luego se realizaron diluciones seriadas hasta 10-2 , estas junto con la
suspensión celular concentrada fueron plaqueadas en medio mineral
glucosa mas kanamicina (MMGK) y en medio mineral octanoato mas

40
kanamicina (MMOCTK), colocando 100 µl de suspensión por superficie de
medio.; las placas se dejaron incubando durante 72 horas a 30°C.
A través de la selección con el antibiótico kanamicina, solo crecieron en
los medios aquellas colonias de P. putida que contenían el transposon
insertado en su genoma, ya que E.coli no es capaz de crecer en medio
mineral y Pseudomonas putida no es resistente a la kanamicina. Debido
a esto las colonias que crecieron en MMGK y MMOCTK se consideraron
como células de P. putida que habían insertado en su genoma el
transposon que carga consigo el gen de resistencia a la kanamicina.
5.4.1 Selección y aislamiento de mutantes deficientes en el acumulo
de PHAs
A partir de clones obtenidos en la biblioteca de mutantes, se
seleccionaron por características fenotípicas aquellos incapaces de
acumular PHAs a partir de Glucosa y Octanoato. Fueron tomadas
aquellas colonias que presentaron tonalidad clara medio transparente en
comparación a las otras colonias de las bacterias que acumulaban de
color blanco opaco.
Estas colonias seleccionadas se transfirieron a MMGK y MMOCTK con el
fin de ser reaisladas, posteriormente se transfirió cada mutante aislado a
medio MMGLU y MMOCT para confirmar si efectivamente eran
deficientes en el acumulo de PHAs , para ello se colocaron tres cepas
control junto con el mutante en cada medio, la cepa salvaje de
Pseudomonas putida , como control positivo de acumulo, P. putida IPT
463 deficiente en el acumulo a partir de carbohidratos y P. putida IPT 461
deficiente en el acumulo tanto a partir de carbohidratos como de ácidos
grasos. Confirmado cada mutante se procedió a preservar cada cepa,
realizar ensayo cuantitativo de producción de PHA y a evaluar por ultimo
los genes afectados por la mutación.

41
5.5 Evaluación de la producción de PHAsMCL
Cada uno de los mutantes obtenidos junto con las cepas control, fueron
estriadas en agar nutriente y cultivadas por 72 horas. A partir de estas
placas se hizo inoculo en 25 ml de caldo nutritivo (Anexo2) , el cual se
llevo a incubación por 24 horas a 150 rpm. Después de transcurrido el
tiempo de incubación se tomó un volumen de 1ml del cultivo para inocular
25 ml de medio mineral (anexo1) conteniendo exceso de fuente de
carbono (5g/L de Glucosa, 3g/L de Octanoato) y limitando la fuente de
nitrógeno (0,5 g/L de (NH4)2SO4), el cultivo se realizó por 72 horas , 30°C,
150 rpm; Transcurrido el tiempo de cultivo se tomaron muestras para
determinar: pH, masa seca celular, cantidad y composición de PHAs.
5.6 Métodos Analíticos
5.6.1 pH
El pH fue determinado en el sobrenadante, después de la centrifugación
de el cultivo, en potenciómetro (Mettler – modelo Delta 350) utilizando
patrones de pH: 4,0; 7,0; y 9,2(Ingold).
5.6.2 Masa seca celular
La masa seca celular fue determinada gravimetricamente después de
centrifugar 10 ml del cultivo a 8500 rpm/ 10 min / 10ºC; posteriormente se
realizó filtración y lavado de las células con agua destilada en membrana
de poro 0,45 µm (Millipore). La membrana conteniendo las células fue
secada a 100 ºC por 4 horas.

42
Después del tiempo de secado a 100ºC fueron colocadas por 20 minutos
en un desecador a temperatura ambiente, para luego ser determinada la
masa seca celular (MS) por la siguiente ecuación:
MS= ((MMC – MM + HM)/VOL) X 1000
Donde:
MMC= masa de la membrana y células después del secado (g)
MM= masa de la membrana (g)
HM= Humedad média del lote de membranas (g)
VOL= volumen de la suspensión centrifugada (L)
5.6.3 Cantidad y composición de PHA
La cantidad y composición de PHA fueron determinadas a través de
cromatografía de gases de propil – esteres (Riis & Mai, 1988). Entre 10 y
15 mg de células liofilizadas fueron transferidas para tubos, a los cuales
se les adicionó 1,5 ml de una solución de acido clorhídrico en
propanol(1:4 v/v), 1,5 ml de 1,2-dicloroetano y 100 µl de una solución de
ácido benzóico (40g/l) en propanol. Los tubos fueron cerrados
fuertemente, agitados y sometidos a propanólisis por 3 horas a 100 ºC ,
con agitación después de los primeros 30 minutos de propanólisis.
Después de enfriarse, se adicionó a los tubos 4 ml de agua destilada,
agitándolos vigorosamente por 30 segundos. Después de separar la fase
acuosa (superior) de la fase orgánica (inferior), se descartó la fase
acuosa y se tomó la fase orgánica para luego ser analizada. Para ello se
tomó 1 µl de la fase orgánica la cual fue analizada después del
fraccionamiento de la muestra (splits 1:20) en cromatógrafo de gases
HP6890 Series GC System equipado con una columna HP-5 (5% fenil-
metil- siloxane, 30m de alto ; 0,25 mm de diámetro; 0,25 µm de espesor
del filme). El análisis fue llevado a cabo bajo las siguientes condiciones:

43
• Gas de arrastre: Hélio (0,8 ml/min)
• Temperatura del inyector: 250 ºC
• Temperatura del detector: 300 ºC
• Sistema de detección : Ionización de llam (FID)
• Programa de temperatura: 100ºC por 1 minuto, elevación de la
temperatura hasta 185ºC a 8 ºc/min y 185 ºC por 15 minutos.
Se utilizó acido benzoico como patrón interno. Los polímeros producidos
por P. putida a partir de diferentes fuentes de carbono o P3HB y P3HB-
co-3HV (Aldrich) fuerón utilizados como patrones para la generación de
las curvas de calibración. El PHA total fue calculado sumándose las
cantidades de los constituyentes 3HB, 3HV, 3HHx, 3HHp, 3HO, 3HN,
3HD, 3HDd y 3 HDd∆5.
5.7 Manipulación de DNA
5.7.1 Extracción de DNA plasmidial y genómico
Se utilizaron los siguientes Kits para la extracción y purificación deL DNA
y según los protocolos e instrucciones de los fabricantes:
• DNA genómico: PurelinkTM Genomic DNA Purification (Invitrogen
Corp.)
• DNA plasmidial: Quick Plasmid Miniprep (Invitrogen Corp).
5.7.2 Digestión del DNA plasmidial y genómico
Se tomaron aproximadamente de 100 ng a 300 ng del DNA plasmidial y
genómico extraído para realizar la digestión; la cual se realizo usando
endonucleasas de restricción en volumen final de 30 µl a 50 µl junto con el
buffer correspondiente a cada enzima utilizada según las

44
recomendaciones del fabricante (Invitrogen Corp.). Las reacciones de
digestión se llevaron a cabo a 37°C overnight. Trascurrido el tiempo
necesario para la digestión se inactivo la enzima a 65°C por 10 minutos.
5.7.3 Electroforesis en gel de agarosa
Las reacciones de digestión de los DNAs plasmidial y genómico fueron
sometidos a electroforesis en el de agarosa (0,8 %) en tampon de corrida
TBE1X (Tris 89 mM; ácido bórico 89 mM; EDTA 2 mM pH 8,0). Las
electroforesis fueron realizadas a 90 V, 80mA, 80W por 1 hora, utilizando
como marcador de peso molecular el DNA Ladder y λ Hindii de 1 Kb
(Invitrogen). Después de la electroforesis el gel fue coloreado con
brometo de etidio 0,5 µg/ml, y el corrido de los DNAs fue observado bajo
luz UV.
5.7.4 Ligación de DNA
Las reacciones de ligación fueron realizadas utilizando la enzima T4 DNA
ligaza (Invitrogen) junto con el buffer suministrado con la enzima por el
fabricante. Las reacciones de ligación se llevaron a cabo a 19°C
overnight.
Con el fin de conseguir una buena ligación entre el DNA del vector y el
DNA del fragmento aislado, se testaron varias proporciones 50:1, 100:1,
200:1, 400:1 para así saber cual de ellas era más eficaz.
5.8 Inserción del DNA
5.8.1 Transformación por choque térmico :
Se realizo un inoculo de E. coli XLI Blue en en 25 ml de medio LB
adicionando 250 µl de MgCl2 10 mM y 250 µl de MgSO4 10 mM; este se
incubo a 37°C , 150 rpm, hasta alcanzar un valor de densidad óptica a

45
650 nm DO650 = 0,3 – 0,5. Posteriormente se centrifugo 8 ml del cultivo
por 15 min / 4000 rpm/ 4°C en tubos falcón estériles. El pellet resultante
fue resuspendido en 4 ml de buffer de transformación (Composición
anexo 4), estas células fueron colocadas en hielo por 15 minutos y
centrifugadas de nuevo bajo las mismas condiciones de operación. Para
la obtención de las células competentes, se resuspendió de nuevo el
pellet en 0,8 ml de buffer de transformación, luego fue dividido en
alícuotas de 200 µl, estas se mantuvieron en baño de gelo mientras eran
utilizadas.
Para cada 200 µl de células competentes, se adicionaron 5 µl de DNA
(Aproximadamente 500 ng), la mezcla fue colocada en baño de hielo
durante 30 minutos y luego sometida a choque térmico a 42°C por 90
segundos, inmediatamente se coloco en hielo para ser enfriada.
Posteriormente se adicionaron a cada mezcla 600 µl de medio LB y
llevadas a incubar por 1 hora a 37°C. Los clones transformantes fueron
seleccionados en agar LB conteniendo el antibiótico apropiado (Ampicilina
o Kanamicina); IPTG (isopropil-tio-β-D-galactosídeo) y X-Gal (5-bromo-4-
cloro-3-indolil-β-D-galactosídeo) también fueron adicionados para
diferenciar los clones recombinantes que contenían el inserto de DNA
(Queiroz, 2008).
5.8.2 Transformación por electroporación
Las células electrocompetentes de la cepa de E. coli DH5α fueron
preparadas deacuerdo al protocolo descrito por Ausubel y colaboradores
(1992). Para realizar las transformaciones por electroporación fueron
utilizados 40 µl de las células competentes de E. coli DH5α y 1 µl de
las ligaciones del vector con el inserto. Esta mezcla fue transferida a
celdas de electroporación de 0,1 cm. Las células fueron sometidas a
pulsos de 1,8 kV/cm, capacitancia de 24 µF y resistencia de 200 Ω, e
inmediatamente incubadas en 1 ml de medio LB por 1 hora a 37 °C.
Finalmente se adicionaron 250 µl de la suspensión en placas de LB, a las

46
cuales fueron adicionados previamente 40 µl de X-gal solución 20 mg/ml
(5-bromo-4-cloro-3-indolil-β-D-galactosídeo) y 7 µl de IPTG solución al
20% (isopropil-tio-β-D-galactosídeo) mas Ampicilina o Kanamicina; las
células fueron distribuidas uniformemente sobre toda la superficie del
medio con ayuda del asa de vidrio, las placas fueron llevadas a incubar a
37 °C por 16 horas (Braz, 2006).
5.9 Recolección de la Información
La recolección de la información se realizó IN SITU, en el laboratorio de
Fisiología de los microorganismos del instituto de ciencias biomédicas de
la Universidad de São Paulo, Brasil. Se llevó un diario de laboratorio el
cual se utilizó como guía y registro de evolución de la investigación.

47
6. RESULTADOS Y DISCUSIÓN
6.1 Construcción de la biblioteca y aislamiento de mutantes
deficientes en el acumulo de PHAMCL utilizando transposon Mini- Tn5
El tamaño del genoma de P. putida corresponde a 6.181.863 Kb
conteniendo 5420 ORFs; siendo necesario obtener 7000 clones como
mínimo para representar la biblioteca de mutantes de esta bacteria; Con
el fin de cumplir este objetivo se construyó una biblioteca de mutantes de
Pseudomonas putida utilizando el transposon mini Tn5 (Lorenzo et al.,
1990). Evaluando cerca de 15000 clones en este trabajo.
La biblioteca fue utilizada para buscar mutantes afectados en la síntesis
de PHAs a partir de glucosa y/o acido octanoico como fuentes de
carbono. Uno de los principales objetivos de la construcción de esta
biblioteca fue identificar y tener un instrumento para el análisis de los
genes involucrados en la síntesis de PHAMCL a partir de carbohidratos y
ácidos grasos en Pseudomonas putida.
La mutación fue realizada através de la inserción del transposon mini Tn5,
el cual se inserto de forma aleatoria en el genoma de P. putida, dando
como resultado una única inserción del material genético por clon. La
transposición ocurrió tras realizarse el proceso de conjugación entre la
cepa de Pseudomonas putida y una E. coli S17-1 donadora del
plásmido suicida
pUT-Km2 el cual contiene el transposon mini Tn5 (Lorenzo et al., 1990) y
el gen de resistencia a la kanamicina. Mediante esta técnica se
obtuvieron cerca de 15000 mutantes los cuales fueron evaluados en
busca de mutantes afectados en la biosíntesis de PHA.
6.2 Selección y aislamiento de mutantes deficientes en el acumulo
de PHAs
Los mutantes fueron seleccionados en medio mineral glucosa mas
kanamicina (MMGK) y medio mineral octanoato mas kanamicina

48
(MMOCTK) con exceso de fuente de carbono (Glucosa 10 g/L –
Octanoato 3 g/L) respectivamente, y limitación de fuente de nitrógeno
((NH4)2SO4 0,2 g/L) para propiciar el acumulo de PHA; la kanamicina fue
usada en los medios como agente de selección para que solo pudiesen
crecer aquellos mutantes que tenían el inserto del transposon en su
genoma.
La selección se realizó entre los mutantes que crecieron en los medios
por características fenotípicas, tomando como referencia el color blanco
opaco característico de las colonias cuando las bacterias se encuentran
acumulando y la tonalidad clara medio transparente cuando no lo hacen.
Obedeciendo a las características anteriormente mencionadas fueron
seleccionados 25 mutantes en total, considerados como deficientes en el
acumulo de PHA a partir de una o ambas fuentes de carbono probadas.
Estos fueron reaislados en un medio con las mismas condiciones de
donde fueron tomados para certificar su pureza, posteriormente fueron
testados en medio mineral con glucosa en exceso 10 g/L (MMGLU) y
medio mineral con octanoato 3 g/L (MMOCT) junto con las cepas control
IPT O46 (cepa salvaje que acumula a partir de ambos sustratos), IPT 463
( cepa control mutante que acumula a partir de octanoato mas no a partir
de glucosa) y IPT 461 ( cepa control mutante que no acumula a partir de
ninguno de los dos sustratos probados); con el fin de probar
cualitativamente si efectivamente se trataban de mutantes de
Pseudomonas putida afectados en el acumulo de PHAs a partir de uno o
de ambos sustratos.
A partir de esta prueba se encontró que 5 de los mutantes no acumulaban
PHA a partir de glucosa pero si lo hacían a partir de octanoato, 1 mutante
no acumulaba ni a partir de glucosa ni a partir de octanoato y los otros 19
mutantes acumulaban PHA a partir de ambos sustratos pero en menor
proporción en comparación a la cepa salvaje, considerándose así
mutantes parciales. Como se muestra en la Tabla 2, se realizó un perfil
comparativo de acumulo en placa a las 24 horas de los mutantes

49
parciales junto con la cepa salvaje en glucosa y octanóato, identificando
en ella los mutantes clave para análisis posteriores.
Tabla 2. Perfil de acumulo de los mutantes parciales de P. putida.
Cepa bacteriana MMGLU MMOCT
IPT 046 ++++ ++++
M11 + ++
M15 ++ +++
M18 + ++
M31 ++ ++
M30 ++ ++
M29 ++ +++
M28 ++ +++
M27 +++ +++
M26 +++ +++
M25 ++ ++
M24 ++ -
M23 ++ ++
M22 ++ ++
M37 +/- +/-
M36 + +
M34 +/- +/-
M35 +++ +++
M33 +++ ++
M32 +++ +++
- : no presento crecimiento; +/- , +, ++, +++, ++++; intensidad creciente de crecimiento.

50
A partir de estos resultados, se observo preliminarmente según la
intensidad y crecimiento de los mutantes en placa, que efectivamente se
encontraban acumulando a partir de ambos sustratos pero en menor
proporción, presentando colonias menos opacas en comparación a la
cepa utilizada como control de Pseudomonas putida, acumulando a una
velocidad menor que la cepa salvaje, posiblemente debido a que pueden
estar afectados en algunos de los genes involucrados en la transferencia
de intermediarios de la β- Oxidación de ácidos grasos para la síntesis de
PHAMCL, como el gen phaJ, el cual codifica la enzima hidratasa enoil-CoA,
que cataliza una hidratación (R)- especifica del 2-enoil-CoA
proporcionando unidades monómero de (R)-3-Hidroxiacil-CoA para la
síntesis de PHA a través de la β-oxidación de ácidos grasos
(Suriyamongkol et al., 2007); el FabG el cual codifica la 3-cetoacil-CoA
reductasa que cataliza la reducción del 3-cetoacil-CoA para obtener el
(R)-3-Hidroxiacil-CoA o el gen que codifica en la conversión del (S)-3-
hidroxiacil-CoA. Estando afectados en alguno de estos genes,
efectivamente pueden estar acumulando pero en menor proporción
debido a que si se ven afectados en el suministro de algún precursor, la
bacteria tendría dos opciones más para transferir intermediarios de la β-
Oxidación a la síntesis de PHAMCL , lo cual disminuye la capacidad de
acumulo mas no la inactiva.
Así mismo también se observo ausencia o disminución de crecimiento de
algunos de los mutantes, como en el caso del mutante M24, lo cual pudo
ser debido a que fueron afectados en algún gen involucrado con el
crecimiento de la bacteria. Pasando a no ser de nuestro interés.
Por otro lado se confirmó en las pruebas en placa de acumulo que 5 de
los 25 mutantes iniciales estaba afectados en la biosíntesis de PHAMCL a
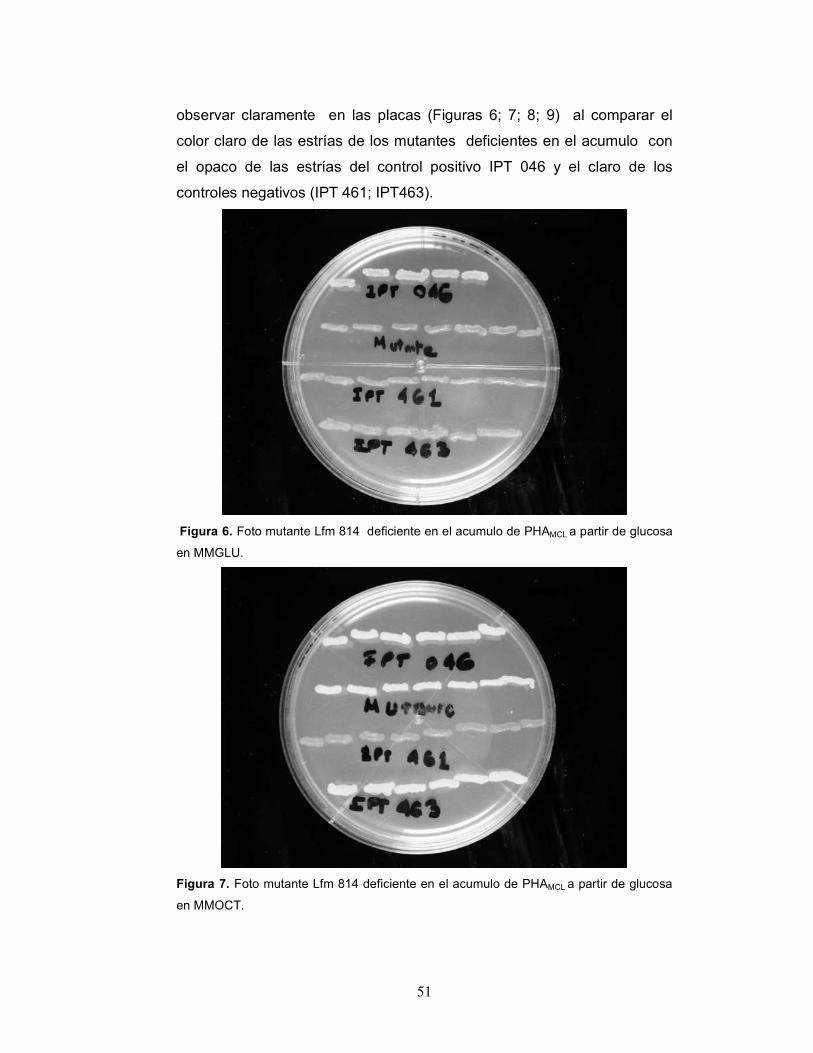
partir de glucosa, (Figuras 6; 7; 8) pasando a ser denominados Lfm 814,
Lfm 820, Lfm 821, Lfm 822 y Lfm 823, y que uno se encontraba
afectado en la síntesis del polímero tanto a partir de glucosa como de
octanoato, pasando a ser denominado Lfm 824 (Figura 9). Esto se puede
51
observar claramente en las placas (Figuras 6; 7; 8; 9) al comparar el
color claro de las estrías de los mutantes deficientes en el acumulo con
el opaco de las estrías del control positivo IPT 046 y el claro de los
controles negativos (IPT 461; IPT463).
Figura 6. Foto mutante Lfm 814 deficiente en el acumulo de PHAMCL a partir de glucosa
en MMGLU.
Figura 7. Foto mutante Lfm 814 deficiente en el acumulo de PHAMCL a partir de glucosa
en MMOCT.
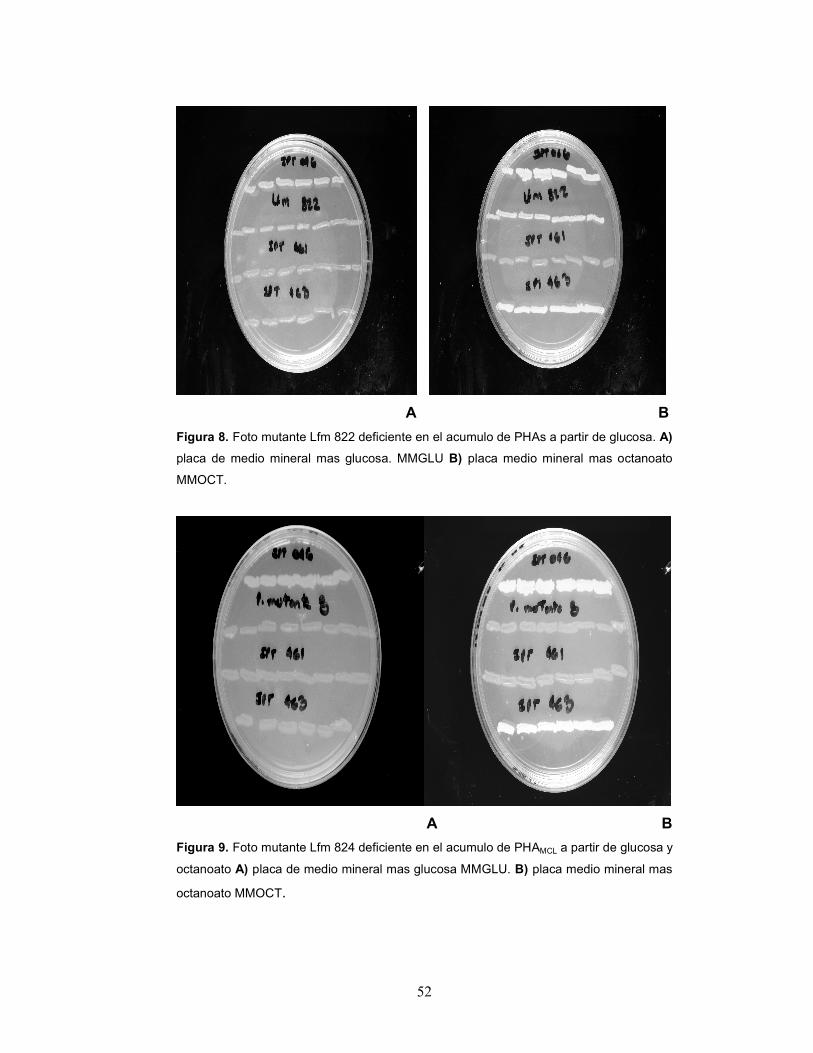
52
A B
Figura 8. Foto mutante Lfm 822 deficiente en el acumulo de PHAs a partir de glucosa. A)
placa de medio mineral mas glucosa. MMGLU B) placa medio mineral mas octanoato
MMOCT.
A B
Figura 9. Foto mutante Lfm 824 deficiente en el acumulo de PHAMCL a partir de glucosa y
octanoato A) placa de medio mineral mas glucosa MMGLU. B) placa medio mineral mas
octanoato MMOCT.

53
6.3 Evaluación de la producción de PHAsMCL
Con el fin de confirmar cuantitativamente la capacidad o incapacidad de
acumular PHAMCL de los mutantes obtenidos a partir de glucosa y/o acido
octanóico, se procedió a evaluar la producción del biopolímero (Tablas 3;
4 y Figuras 10; 11).
Tabla 3. Producción de PHAMCL por mutantes de Pseudomonas
putida a partir de Glucosa
Cepa X pH 3 HHx 3HO 3HD 3HDD 3HDd∆∆∆∆5 PHA PHA
(g/L) (mol %) (mol%) (mol%) (mol%) (mol%) (%MSC) (g/L) IPT 046 1,58 6,9 2,68 30,03 63,40 0,00 3,89 42,31 0,67 Lfm 814 0,85 6,6 0,00 0,00 0,00 0,00 0,00 0,00 0,00 Lfm 824 0,83 6,5 0,00 0,00 0,00 0,00 0,00 0,00 0,00 Lfm 821 0,32 6,7 0,00 0,00 0,00 0,00 0,00 0,00 0,00 Lfm 822 1,33 7,1 0,00 31,27 68,73 0,00 0,00 43,33 0,58 Lfm 820 1,59 6,8 0,00 30,19 69,81 0,00 0,00 45,22 0,72 IPT 463 0,98 6,4 0,00 0,00 0,00 0,00 0,00 0,00 0,00 IPT 461 0,89 7,0 0,00 0,00 0,00 0,00 0,00 0,00 0,00

54
IPT 046 Lfm 814 Lfm 824 Lfm 821 Lfm 822 Lfm 820 IPT 463 IPT 461
S1
0
5
10
15
20
25
30
35
40
45
50
CepasPHA (%MSC)
Producción de PHA por mutantes a partir de glucosa
Figura 10. Producción de PHA por mutantes de Pseudomonas putida
a partir de glucosa
55
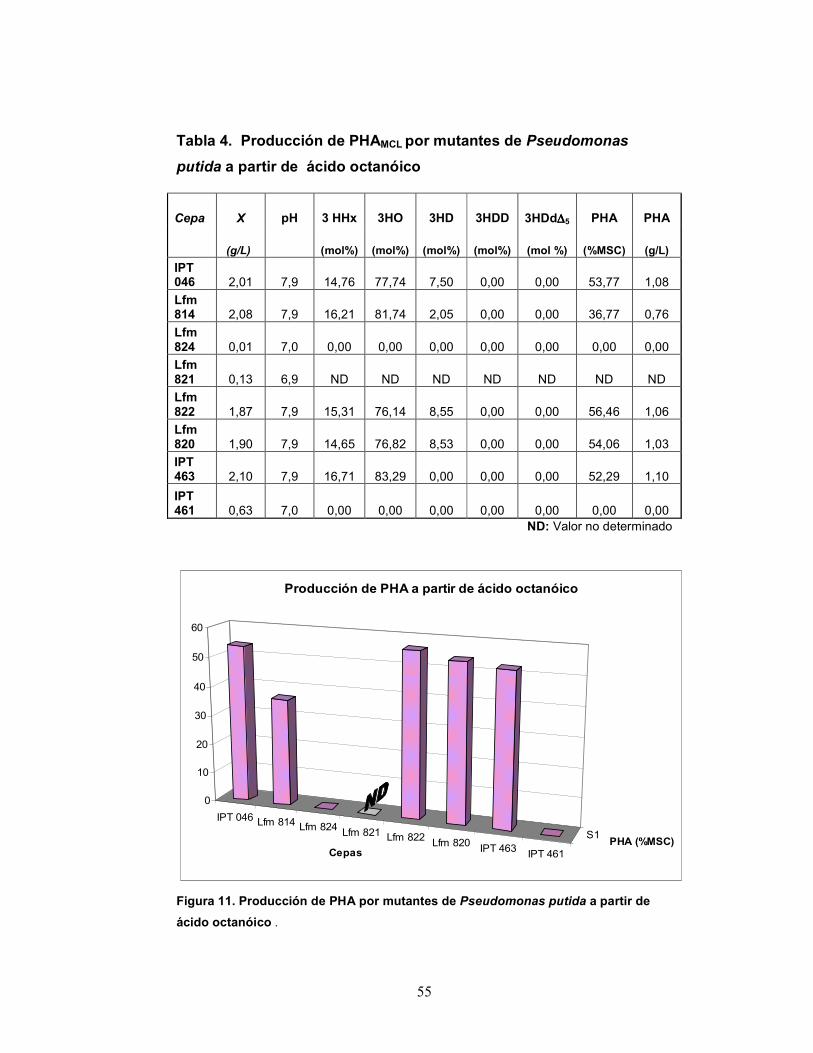
Tabla 4. Producción de PHAMCL por mutantes de Pseudomonas
putida a partir de ácido octanóico
ND: Valor no determinado
IPT 046 Lfm 814 Lfm 824 Lfm 821 Lfm 822 Lfm 820 IPT 463 IPT 461
S1
0
10
20
30
40
50
60
CepasPHA (%MSC)
Producción de PHA a partir de ácido octanóico
Figura 11. Producción de PHA por mutantes de Pseudomonas putida a partir de
ácido octanóico .
Cepa X pH 3 HHx 3HO 3HD 3HDD 3HDd∆∆∆∆5 PHA PHA
(g/L) (mol%) (mol%) (mol%) (mol%) (mol %) (%MSC) (g/L)
IPT 046 2,01 7,9 14,76 77,74 7,50 0,00 0,00 53,77 1,08
Lfm 814 2,08 7,9 16,21 81,74 2,05 0,00 0,00 36,77 0,76
Lfm 824 0,01 7,0 0,00 0,00 0,00 0,00 0,00 0,00 0,00
Lfm 821 0,13 6,9 ND ND ND ND ND ND ND
Lfm 822 1,87 7,9 15,31 76,14 8,55 0,00 0,00 56,46 1,06
Lfm 820 1,90 7,9 14,65 76,82 8,53 0,00 0,00 54,06 1,03
IPT 463 2,10 7,9 16,71 83,29 0,00 0,00 0,00 52,29 1,10
IPT 461 0,63 7,0 0,00 0,00 0,00 0,00 0,00 0,00 0,00

56
Como se puede observar en los datos presentados en las tabla 3; 4; y las
figuras 10 y 11, de los cinco presuntos mutantes deficientes en el
acumulo de PHA a partir de glucosa que se creían tener, solo tres fueron
confirmados en los ensayos de producción de PHA.
La cepa Lfm 814 mostró ser incapaz de acumular PHAMCL a partir de
carbohidratos, y capaz de seguir acumulando a partir de acido octanóico.
Mientras que la cepa salvaje mostró acumular cerca de un 45 % de su
masa seca celular en PHA a partir de carbohidratos y alcanzar un índice
de biomasa cercano a los 1,6 g/L a las 72 horas de cultivo, la cepa
mutante no acumuló ningún tipo de PHA y no alcanzó a tener 1 g/L de
biomasa. Mostrando un perfil similar al del control IPT 463 utilizado en
los ensayos, el cual no acumula tampoco a partir de glucosa mas lo
continua haciendo a partir de acido octanóico, e igualmente que el
mutante obtenido en este estudio no alcanzo a tener mas de 1 g/L de
masa seca celular.
En el estudio realizado por (Gomez, 2000) de obtención de mutantes de
Pseudomonas putida por radiación UV, encontró este mismo tipo de
mutantes, para lo cual le dio la explicación de que se trataban de
mutantes que debían estar afectados en algún gen involucrado en la
codificación de una enzima responsable de un paso especifico de la
síntesis de PHA a partir de estos sustrato.
Pseudomonas putida utiliza como principal ruta para la síntesis de
PHAMCL a partir de sustratos no relacionados tales como la glucosa, la
biosíntesis de ácidos grasos de novo para la síntesis de monomeros de
3-Hidroxiacil-CoA.
El gen phaG codifica a la enzima que enlaza la biosíntesis de ácidos
grasos de novo y la síntesis de PHA; esta fue clonado de P. putida
(Rehm et al., 1998; Hoffmann et al., 2000a,b). El producto de este gen
cataliza la conversión de (R)-3-hidroxiacil-ACP, intermediario de la vía de
biosíntesis de ácidos grasos a su correspondiente derivado CoA.
En contraste P. oleovorans y P. fragii son incapaces de acumular
PHAMCL a partir de carbohidratos, para su síntesis usan los ácidos grasos

57
como fuente de carbono mediante la β- Oxidación para producir 3-
Hidrociacil-CoA, sustrato de PHAMCL sintasa (Suriyamongkol et al., 2007).
Se ha demostrado según estudios, que la expresión de phaG en E. coli da
a la bacteria recombinante la nueva capacidad de producir ácidos grasos-
mcl libres como precursores 3(HD) (Zheng et al., 2004) y su expresión en
P. oleovorans y P. fragii produce PHAMCL (Fiedler et al., 2000; Hoffmann
et al., 2000b), a partir de fuentes de carbono no relacionadas a la
estructura 3HD, tales como la glucosa y la fructosa.
Por ello se cree que el mutante Lfm 814 puede estar afectado en el gen
phaG, el cual al ser inactivado por la inserción del transposon dejo de
codificar para la enzima 3-Hidroxiacil-ACP-CoA transcilasa, paso clave en
la síntesis de PHAMCL a partir de carbohidratos como se menciono
anteriormente.
Por otro lado se obtuvo un mutante total, deficiente en el acumulo tanto a
partir de glucosa como a partir de octanoato, la cepa mutante Lfm 824,
confirmo en los ensayos de producción de PHA no encontrarse
acumulando a partir de ninguno de los dos sustratos, presentando un
perfil similar al de la cepa control IPT 461, la cual también es deficiente en
el acumulo a partir de carbohidratos y ácidos grasos.
Este mutante muy posiblemente debe estar afectado en algún gen phaC,
el cual codifica para la principal enzima señalada en la síntesis de
polihidroxialcanoatos, necesaria para la síntesis del biopolímero a partir
de cualquier sustrato; la PHA sintasa, responsable por la polimerización
de unidades de (R)-3-hidroxiacil-CoA, formando el polihidroxialcanoato
(Steinbuchel, 1996), su especificidad por el sustrato define la naturaleza
del polímero final.
Adicional a esto, se obtuvo un mutante deficiente en el acumulo de
PHAMCL a partir de glucosa, el cual fue denominado Lfm 821, sin embargo
no se pudo confirmar en los ensayos de producción de PHA si
efectivamente acumulaba o dejaba de acumular a partir de acido
octanóico, por lo cual no se puede aun plantear una hipótesis clara acerca

58
de los genes que pudieron ser afectados en este mutante, en tanto no se
repitan de nuevo las pruebas de producción de PHA para esta cepa.
Paralelamente, se realizó un ensayo para evaluar cuantitativamente la
producción de PHAs a través del tiempo de algunos mutantes que se
consideraban parciales, (definiendo como mutantes parciales en este
trabajo a aquellos que aún se encontraban acumulando PHAMCL pero en
menor proporción, designados con la letra M, Tabla 2.) Ya que en las
pruebas en placa a pesar de seguir acumulando PHA, mostraban hacerlo
en menor proporción al ser comparados con la cepa salvaje.
En los ensayos de producción e PHA con los posibles mutantes parciales,
se observaron diferentes proporciones de monómeros; a partir de Glucosa
como fuente de carbono se encontró presencia de 3 HDd∆5 en menor
proporción, seguido por 3 HDD, 3HHx, 3HO y con una buena proporción
3 HD (Tabla 5; Figura 13). Esto tanto en la cepa salvaje como en los
mutantes, siendo estos monómeros usuales en PHAs a partir de
carbohidratos, como es el caso del HDd∆5.
Al evaluar cuantitativamente la producción de PHAMCL a partir de glucosa,
las cepas mostraron un comportamiento similar al de la cepa salvaje, sin
embargo cabe destacar que la cepa M18 acumulo un poco más que la
cepa salvaje a partir de glucosa (Tabla 5; Figura 12) y la cepa M11
mostró acumular PHA en menor proporción a partir del mismo sustrato
comparándola con la cepa salvaje a las 24 horas (Tabla 5; Figura 12).
Así mismo también se observo una disminución en la concentración de
PHAMCL, en todas las cepas a las 72 horas de cultivo, esto talvez debido al
posible reconsumo del polímero que puede estar realizando la bacteria
En los ensayos a partir de acido octanóico como fuente de carbono, se
encontró presencia de 3HHx, 3HD y 3 HO en mayor proporción, tal y
como se esperaba cuando se proporciona este sustrato para la
producción de PHAMCL (Tabla 6; Figura 15); adicional a esto al calcular la

59
producción de PHA en (g/L) se observó que efectivamente la cepa
mutante M18 estaba acumulando menos que la cepa salvaje (Tabla 6;
Figura 14), siendo este un mutante parcial, el cual puede estar
posiblemente afectado en uno de los genes que codifican para las
enzimas responsables de la transferencia de intermediarios de la β-
oxidación de ácidos grasos para la síntesis de PHA, tales como el gen
phaJ que codifica para la (R)- enoil-CoA hidratasa y el gen FabG que
codifica para la 3-cetoacil-CoA reductasa; de esta forma al tener uno de
los genes inactivados, el otro puede seguir funcionando igual y producir
una enzima que se encargue de la transferencia de uno de los
intermediarios, produciendo finalmente PHA pero en menor proporción ya
que cuenta con menos elementos para la síntesis.
Por ello, seria muy interesante en estudios futuros, seguir buscando
mutantes de este tipo e intentar identificar cual de los genes fue afectado
en este mutante.

60
Tab
la 5. Producción de PHAs por mutantes parciales de Pseudomonas putida a partir de gluco
sa
Cep
a Hora
X (g/L)
pH
3 HHx
3HO
3HD
3HDD
3HDd
∆ ∆∆∆5
PHA
PHA
(mol %)
(mol %)
(mol %)
(mol %)
(mol %)
(%MSC)
(g/L)
IPT 046
24
0,98
6,
30
0,00
33
,05
66,9
5
0,00
0
,00
34
,23
0,34
48
1,
46
6,80
0,
00
35,3
6 66
,64
0,
00
0,0
0
42,7
2 0,
62
72
1,
38
6,50
2,
89
32,4
1 61
,10
0,
00
3,6
0
39,6
1 0,
55
M36
24
1,22
7,
00
0,00
33
,07
66,9
3
0,00
0
,00
32
,71
0,40
48
1,
93
6,90
2,
67
30,9
1 59
,22
3,
45
3,7
5
43,7
6 0,
84
72
1,
46
6,70
3,
12
34,4
3 62
,45
0,
00
0,0
0
35,4
7 0,
52
M18
24
0,87
6,
60
0,00
28
,37
71,6
3
0,00
0
,00
34
,07
0,30
48
2,
51
6,40
2,
71
26,8
8 65
,65
0,
00
4,7
7
48,4
7 1,
22
72
1,
53
6,80
2,
58
26,3
8 62
,86
3,
53
4,6
6
47,0
1 0,
72
M11
24
1,34
7,
00
0,00
65
,52
34,4
8
0,00
0
,00
15
,84
0,21
48
1,
64
6,50
2,
98
30,3
7 66
,65
0,
00
0,0
0
46,
2 0,
76
72
1,
66
6,70
0,
00
28,4
8 62
,88
3,
99
4,6
5
44,9
2 0,
75

61
Prod
ucció
n de
PHA
a pa
rtir d
e Gluco
sa
0102030405060
2448
7224
4872
2448
7224
4872
IPT
046
M36
M18
M11
Tiem
po (h
) / Cep
as
PHA (%MSC)
Prod
ucción
PHA
en (g/L) a
patir de
Gluco
sa
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
2448
7224
4872
2448
7224
4872
IPT
046
M36
M18
M11
Tiem
po (h
)/ Ce
pas
PHA (g/L)
Figura 12. C
omparación de Producción de PHA por mutantes parciales de Pseudomonas putida en diferen
tes tiem
pos a
partir de gluco
sa

62
IPT 046
0,00
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
2448
72
Tiempo (h)
mol (%)
3 H
Hx
3HO
3HD
3HD
D3H
DdD
5
M 36
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
24
48
72
Tiempo (h)
mol (%)
3 H
Hx
3 H
O3 H
D3 H
DD
3 H
Dd5
M18
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
24
4872
Tiempo (h)
mol (%)
3 H
Hx
3 H
O3 H
D3 H
DD
3H
Dd5
M11
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
24
48
72
Tiempo (h)
mol (%)
3 H
Hx
3 H
O3 H
D3 H
DD
3 H
Dd5
Figura 13. Comparación de monómeros de PHA producidos por mutantes parciales de Pseudomonas putida en diferentes tiempos a
partir d
e gluco
sa

63
Tab
la 6. Producción de PHAs por mutantes parciales de Pseudomonas putida a partir de ácido octan
óico
Cep
a
Hora
X (g/L)
pH
3 HHx
3HO
3HD
3HDD
3HDd
∆ ∆∆∆5
PHA
PHA
(mol %)
(mol %)
(mol %)
(mol %)
(mol %)
(%MSC)
(g/L)
IPT 046
24
1,02
7,
8 10
0,00
0,
00
0,0
0 0,
00
0,00
5,
08
0,05
48
2,
01
7,9
14,4
3
76,
88
8,6
9 0,
00
0,00
5
3,0
3 1,
07
72
1,
62
7,8
13,7
5
76,
64
9,6
1 0,
00
0,00
5
1,1
6 0,
83
M36
24
0,47
7,
9 15
,05
8
4,9
5 0
,00
0,00
0,
00
27,
31
0,13
48
1,
93
7,8
ND
N
D
ND
N
D
ND
N
D
ND
72
1,
73
8,0
12,8
7
74,
47
12,6
6 0,
00
0,00
4
7,9
4 0,
83
M18
24
0,31
7,
5 0,
00
0,00
10
0,0
0 0,
00
0,00
2,
97
0,01
48
0,
98
7,8
16,8
6
72,
49
10,6
5 0,
00
0,00
5
1,5
7 0,
51
72
1,
74
7,7
16,3
8
71,
50
12,1
2 0,
00
0,00
4
6,5
3 0,
81
M11
24
0,30
7,
4 15
,96
8
4,0
4 0
,00
0,00
0,
00
15,
54
0,05
48
1,
93
7,8
13,6
7
75,
14
11,2
0 0,
00
0,00
5
2,9
9 1,
02
72
1,
57
7,8
13,2
2
74,
40
12,3
8 0,
00
0,00
4
3,7
2 0,
69
ND:
Va
lor
no
det
erm
inad
o
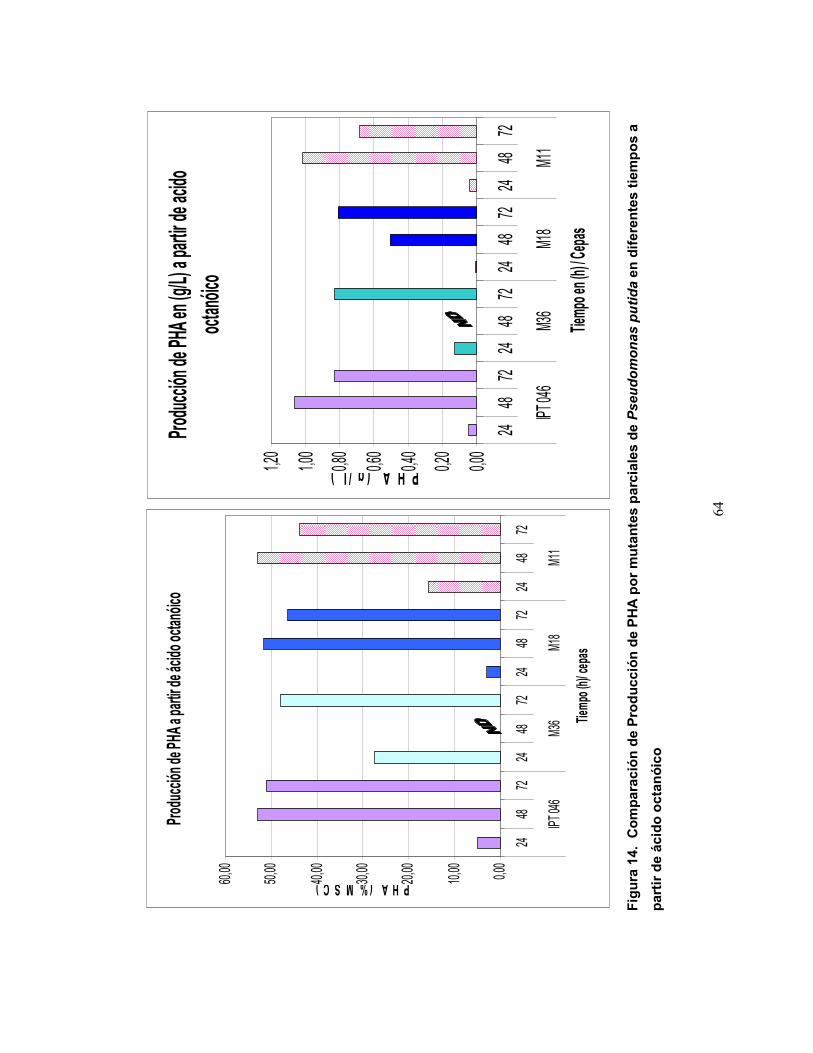
64
Prod
ucció
n de P
HA a pa
rtir de
ácido
octan
óico
0,00
10,00
20,00
30,00
40,00
50,00
60,00
2448
7224
4872
2448
7224
4872
IPT 04
6M3
6M1
8M1
1
Tiemp
o (h)/
cepa
s
PHA (%MSC)
Produ
cción
de PH
A en (
g/L) a
partir
de ac
ido
octan
óico
0,00
0,20
0,40
0,60
0,80
1,00
1,20
2448
7224
4872
2448
7224
4872
IPT 04
6M3
6M1
8M1
1
Tiemp
o en (
h) / C
epas
PHA (g/L)
Figura 14. C
omparación de Producción de PHA por mutantes parciales de Pseudomonas putida en diferen
tes tiem
pos a
partir de ácido octan
óico

65
IPT 046
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
90,0
0
100,0
0
24
48
72
Tiempo (h)
(mol%)
3 H
Hx
3 H
O3 H
D
M36
0,0
0
5,0
0
10,0
0
15,0
0
20,0
0
25,0
0
30,0
0
35,0
0
40,0
0
45,0
0
24
48
72
Tiempo (h)
(mol%)
3 H
Hx
3 H
O3 H
D
M18
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
90,0
0
100,
00
2448
72
Tiempo (h)
(mol%)
3 H
Hx
3 H
O3
HD
M 11
0,0
0
10,0
0
20,0
0
30,0
0
40,0
0
50,0
0
60,0
0
70,0
0
80,0
0
90,0
0
100
,00
24
4872
Tiempo (h)
(mol%)
3 H
Hx
3 H
O3
HD
Figura 15. Comparación de monómeros de PHA producidos por mutantes parciales de Pseudomonas putida en diferentes tiempos a
partir de ác
ido octan
óico

Por otro lado se esperaba encontrar mutantes de Pseudomonas putida
deficientes en el acumulo de PHAMCL solo a partir de acido octanóico, para
así encontrar un gen directamente relacionado con la transferencia de
intermediarios de la β-Oxidación de ácidos grasos para síntesis de PHA,
mas no fue posible tras varios intentos y ensayos, al igual que en otros
estudios anteriores como en el de Gómez,(2000), donde tampoco se
obtuvieron mutantes de este tipo, esto talvez puede ser debido a que a
través de la β-Oxidación se obtiene mas de un intermediario, incluyendo
el enoil-CoA, 3-cetoacil-CoA y el (S)-3-hidroxiacil-CoA, los cuales pueden
servir como precursores de mcl-(R)-3-hidroxiacil-CoA, usado
directamente en la síntesis de PHAMCL (Suriyamongkol et al., 2007).
Teniendo que bloquear de esta manera mas de un gen para incapacitar
totalmente a la bacteria en la producción de PHA a partir de ácidos
grasos.
6.4 Identificación de los Genes afectados en los mutantes
Con el fin de identificar los genes de los mutantes de Pseudomonas
putida afectados por la inserción del transposon mini Tn5, el DNA
genómico de los mutantes Lfm 814 y Lfm 824 fue digerido con la enzima
de restricción Pst1. Paralelamente a esto se realizo la digestión y
posterior defosforilación del vector de clonación pUCBM20, con el fin de
impedir que los extremos del vector del volvieran a unir y cerrar, de esta
manera se aumentan las posibilidades de inserción del fragmento de DNA
de interés dentro del vector.
Una vez realizado esto, se procedió a realizar la ligación entre el DNA
digerido de los mutantes y el vector de clonación, posteriormente se
realizaron dos tipos de procedimientos con el vector ya ligado; el primero
tratamiento fue transferir el vector a una E. coli XL-Blue por
transformación por choque térmico en presencia de CaCl2.
El segundo tratamiento fue transferir el vector a una E.coli DH5α por
medio de transformación por electroporación.

67
Posteriormente se evalúo la eficiencia de cada método seleccionando las
colonias que recibieron el fragmento en LBA y LBK, para así saber cual
era el mas adecuado y eficaz para encontrar el fragmento de DNA con el
transposon.
Tras varios tratamientos, se encontró una eficiencia similar de
transformación con ambos métodos, solo aumentando cuando se
utilizaba pUCBM20 defosforilado en la ligación.
La diferencia básicamente radico en la rapidez y practicidad que presento
el uso de electroporación, ya que en unos pocos minutos se podía tratar
una mayor cantidad de muestras, obteniendo así un mayor número de
ensayos y consecuentemente mayor posibilidad de hallar el fragmento de
DNA de interés.
Aunque se realizaron varios intentos no fue posible encontrar un clon de
interés. Para lograrlo es recomendable seguir estableciendo las mejores
condiciones de ligación y tratamiento.
Lo que se esperaba o se espera encontrar a futuro es el fragmento de
DNA con el transposon inserto en un plásmido adentro de una E. coli
transformada, que logre crecer en LBK, ya que si se encontraba una
colonia en este medio era clara evidencia de que se trataba de la E.coli
con el vector, el gen y el trasnposon miniTn5 que le conferiría resistencia
a la kanamicina. Para después poder proseguir a realizar la extracción y
secuenciación, obteniendo finalmente la identidad de los genes afectados
en los mutantes.
La obtención de estos mutantes puede ser empleada en un futuro para la
identificación y clonación de los genes involucrados en la síntesis de PHA
a partir de carbohidratos y ácidos grasos, convirtiéndose en una
herramienta para su posible modelaje o producción en organismos mas
eficientes .

68
7. CONCLUSIONES
• Se obtuvieron mutantes de Pseudomonas putida deficientes en el
acumulo de PHAMCL a través de inserción del transposon Tn5,
mostrando ser una herramienta eficaz en la generación de nuevos
mutantes deficientes en el acumulo de PHAMCL.
• Fue posible obtener un mutante incapaz de producir PHAs a partir
de carbohidratos, indicando que posiblemente fue afectado en el
gen phaG, relacionado con la producción de PHAMCL a partir de
sustratos no relacionados metabolizados a Acetil-CoA.
• Se obtuvo un mutante deficiente en el acumulo de PHAs a partir de
Glucosa y acido octanóico, estando posiblemente afectado en el
gen phaC, que codifica para la PHA sintasa necesaria en la
síntesis de PHA a partir de cualquier sustrato.
• Fue obtenido un tercer mutante afectado en el acumulo a partir de
carbohidratos, sin embargo es necesario verificar si es capaz o no
de acumular a partir de ácido octanóico.
• Tras varios intentos, no fue posible identificar mutantes afectados
específicamente en el acumulo de PHA a partir de acido octanóico,
sugiriendo que el producto de mas de un gen debe contribuir para
el direccionamiento de intermediarios de la β-oxidación para la
síntesis de PHA.
• Aunque aún no fue posible identificar los genes afectados en los
mutantes obtenidos, mediante los ensayos se logro demostrar que
era más eficiente el uso del vector pUCBM20 defosforilado y el
empleo de electroporación, para obtener mayores eficiencias de
transformación.

69
8. RECOMENDACIONES
• El uso del transposon mini Tn5 para generar mutaciones aleatorias
en el genoma de la bacteria, mostró ser eficaz, además de ser una
herramienta para la obtención de mutantes, también representa
una ventaja en la identificación de los genes afectados e
involucrados en una vía o proceso de interés, ya que su inserción
demarca mas fácilmente el sitio del genoma afectado en
comparación a otros métodos usados comúnmente en la obtención
de mutantes como la radiación Ultravioleta, en la cual no se
distinguen fácilmente los genes afectados por ella.
• Por tratarse de una búsqueda de mutantes que solamente sean
incapaces de acumular a partir de glucosa y acido octanóico, se
recomienda sembrar directamente la suspensión después de la
conjugación en medio mineral, con el sustrato de prueba en
condiciones de exceso y limitación de nitrógeno mas kanamicina,
para la selección de los mutantes deficientes, ya que tras varios
ensayos e intentos de obtener mutantes se demostró que de esta
manera era posible obtener y seleccionar los mutantes con mayor
facilidad y rapidez.
• Tras la obtención de los posibles mutantes parciales en este
trabajo se recomienda hacer nuevos ensayos de producción de
PHAs en función del tiempo con cada uno de ellos, con el fin de
confirmar cuantitativamente si se trata de mutantes parciales en el
acumulo de PHA.
• Para la futura identificación de los genes afectados en los
mutantes, se recomienda el uso del vector de clonación pUCBM20
defosforilado, ya que mostró mayor eficiencia de ligación y

70
transformación; así como también se recomienda el uso de
electroporación como técnica de transformación por su eficiencia y
practicidad.

71
9. BIBLIOGRAFIA
• Anderson AJ, Dawes EA. 1990. Occurrence, metabolism, metabolic
role, and industrial uses of bacterial polyhydroxyalkanoates.
Microbiol. Rev, 54:450-472.
• Angelova N, Hunkeler D. 1999. Rationalizing the design of
polymeric biomaterials. Trends Biotechnol. 17:409-421.
• Ausubel, FM.; Brent, R. Kingston, RE.; Moore, D.D.; Seidmam, J.G;
Smith, J.A.; Struhl K. 1992. Short protocols in molecular Biology
. 3nd edition. Greene Publishing Associates and John Wiley & Sens.
• Babu, G.N. Hammar, W.J.; Rutherford, D.R. 1997. Poly-3-
hydroxyalkanoates as pressure sensitive adhesive. In: Steinbüchel,
A.; Poirier, Y. & Witholt, B. (eds). 1996 International Symposium
on Bacterial Polyhydroxyalkanoates., NRC Research Press,
Ottawa. P. 48-55.
• Brandl, H., Gross, R. A, Lenz, R. W., Fuller, R. C. 1990. Plastic
from bacteria and for bacteria: poly (β-Hydroxyalkanoates) as
natural , biocompatible and biodegradable polyesters. Adv.
Biochem. Eng. and Biotechnol., 41: 77-93.
• Braz, V.S. 2006. Caracterização de genes envolvidos na resposta a
metais em Caulobacter crescentus. Tese Doutorado em
Microbiologia. Instituto de Ciências Biomédicas, Universidade
de São Paulo, São Paulo, Brasil. 145 pag.
• Buisman, G.J.H.; Cuperus, F.P. Weusthuis, R.A. 2000. Poly(3-
hydroxyalkanoate) paint and method for the preparation thereof. US
Patent 6.024.784.

72
• Chen G. Qiang, Wu Qiong. 2005. The applications of
polyhydroxyalkanoates as tissue engineering materials.
Biomaterials. 26: 6565 – 6578.
• Chen G. Qiang. 2008. Recent Biodegradable Plastics Development
In China. International Centre for Science and High
Technology.<http://www.ics.trieste.it/Portal/ActivityDocument.aspx?i
d=5397 > [ Consulta: 24 Jun. 2008].
• Chulia, D. Delail, M, Pourcelot, Y. 1994. Powder Technology and
Pharmaceutical Processes, Amsterdam: Elservier Science.
• Desai SJ, Simonelli, A.P, Higuchi W.I. 1965. Investigation of factors
influencing release rateo f solid drug dispersal in inertet matrices.
J,Pharm. Sci. V.54, p. 1459 – 1464.
• Di Lorenzo ML, Silvestre C. 1999. Non-isothermal crystallization of
polymers. Prog Polym Sci , 24:917-950.
• Fiedler S, Steinbüchel A, Rehm B. 2000. PhaG-mediated synthesis
of poly(3-hydroxyalkanoates) consisting of medium – chain –length
constituents from non-related carbon sources in recombinant
Pseudomonas fragi . Appl. Environ. Microbiol. 66: 2117-24.
• Fukui T, Doi Y. 1997. Cloning and analysis of the poly(3-
hydroxybutyrateco-3-hydroxyhexanoato) biosynthesis genes of
Aeromonas caviae. J. Bacteriol. 179: 4821-30
• García B, Olivera ER, Miñambres B, Fernández-Valverde M,
Cañedo LM, Prieto MA, García JL, Martínez M, Luengo JM. 1999.

73
Novel biodegradable aromatic plastics from a bacterial source. J.
Biol. Chem . 274:29228-29241
• Gomez,J.G. C.; Rodrigues, M.F. A.; Torres, B. B.; Netto, C. L. B.;
Oliveira, M.S; Silva, L.F. 1996. Evaluation of soil gram- negative
bactéria yielding polyhydroxyalkanoic acids from carbohidratos and
propiónico acid. Appl. Microbiol. Biotech., v. 45, p. 785 – 791.
• Gomez J.G. C. 2000. Produção por Pseudomonas sp de
polihidroxialcanoatos contendo monômeros de cadeia média a
partir de carboidratos: Avaliação da eficiência, modificação da
composição e obtenção de mutantes. Tese (Doutorado) –
Instituto de Ciências Biomédicas, Universidade de São Paulo,
São Paulo, Brasil. 155 pag.
• Hanahan, D. 1983. Studies on transtransformation of Escherichia
coli with plasmids. J. Mol. Biol. 166: 557.
• Hoffmann N, Steinbüchel A, Rehm BHA. 2000a. The Pseudomonas
aeruginosa phaG gene product is envolved in the synthesis of
polyhydroxyalkanoic acid consisting of medium-chain-length
constituents from non-related carbon sources. FEMS Microbiol.
Biotechnol. 184:253-9.
• Hoffmann N, Steinbüchel A, Rehm BHA. 2000b. Homologous
functional expression of cryptic phaG from Pseudomonas
oleovorans establishes the transacylase-mediated
polyhydroxyalkanoate biosynthetic pathway. Appl. Microbiol.
Biotechnol. 54: 665-70.
• Holmes ,P.A. 1985 Applications of PHB, Microbially produced
biodegradable thermoplastic. Phys. Tecnhol., v.16 p.32-36.

74
• Hrabak, O. 1992 Industrial production of Poly - β- Hidrpxibutirato.
FEMS Microbiol. V. 103, p.251 -256.
• Huisman G.W., De Leeuw O, Eggink G. 1989. Synthesis of
polyhydroxyalkanoates is a common feature of fluorescent
pseudomonads. Appl. Environ. Microbiol. 55: 1949 -1954.
• Klinke S, Duner M, Scott G, Kessler B, Witholt B. 2000. Inactivation
of Isocitrate Lyase leads to increased production of medium- chain-
length Poly(3-Hydroxyalkanoates) in Pseudomonas putida .
American Society for Microbiology. 66: 3: 909-913.
• Lee SY, Choi J. 1999. Production and degradation of
polyhydroxyalkanoates in waste environment. Waste Management
. 19:133-139.
• Lorenzo D.V., Herrero M, Jakubzik U, Timmis K. 1990. Mini-Tn5
transposon derivates fon insertion mutagénesis, promoter probing,
and chromosomal insertion of cloned DNA in Gram-negative
Eubacteria. Journal of Bacteriology. 172: 6568-6572.
• Luengo J. M, García B., Sandoval A, Naharroy G, Olivera E. L.
2003. Bioplastics from microorganisms. Current Opinion in
Microbiology, 6:251–260.
• Madison LL, Huisman GW. 1999. Metabolic engineering of poly(3-
hydroxyalkanoates): from DNA to plastic. Microbiol. Mol. Biol.
Rev. 63:21-53.

75
• Müller, H. M., Seebach, D. 1993. Poly(hidroxyfettsäureester), eine
fünfte Hlasse von physiologisch bedeutsamen organischen
Bioplymeren. Angew. Chem. 105: 483-509.
• Murray RGE, Doetsch RD, Robinow CF. 1994. Determinative and
cytological light microscopy. In Manual of Methods for General
Bacteriology. Edited by Gerhardt P, Murray RGE, Wood WA, Krieg
NR. Washington DC, ASM; vol 10: 21-41.
• Pouton, C. W Akthar,S. 1996. Biosynthetic polyhydroxyalkanoates
and theirpotential in drug delivery. Adv. Drug Delivery Rev. V.18,
P. 133 – 162.
• Preusting, H.R. 1992. Pseudomonas oleovorans as a source of
bioplastics: production and characterization of poly(3-
hydroxyalkanoates). The Netherlands. PhD Thesis – University of
Gröningen.
• Prieto MA, Buhler B, Jung K, Witholt B, Kessler B. 1999. PhaF, a
polyhydroxyalkanoate-granule-associated –protein of
Pseudomonas oleovorans GPo1 involved in the regulatory
expression system for pha genes. J. Bacteriol . 181: 858-68.
• Queiroz S.R. 2008. Estudo do metabolismo de ácidos graxos em
Pseudomonas putida visando à modulação da composição
monomérica de elastômero biodegradável. Tese (Doutorado),
Interunidades em Biotecnologia USP, Instituto Butantã, IPT.
São Paulo, Brasil. 143 pag.
• Ramsay,B. A., Lomaliza, K. Chavarie, C. 1990. Production of poly-
β-hydroxybutyric-co- β- hidroxyvaleric acids. Appl. Environ.
Microbiol. 56: 2093- 2098.

76
• Rehm B. H. A, Steinbüchel A. 1999. Biochemical and genetic
analysis of PHA synthases and other proteins required for PHA
synthesis. International Journal of Biological Macromolecules.
25: 3- 19.
• Rehm, B. H. A. 2003. Polyester synthases: natural catalysis for
plastics. Biochem. J., V.376, p. 15 – 33.
• Rehm, B. H. A.; Steinbuchel, A. 1999. Biochemical and genetic
analysis of PHA synthases and other proteins required por PHA
synthesis. Int. J. Biol. Macromol., V.25, p. 3-19.
• Rehm, B. H. A.; Kruger N.; Steinbüchel, A. 1998, A new metabolic
link between fatty acid de novo synthesis and polyhydroxyalkanoic
acid synthesis. J. Biol. Chem. 273: 24044-51.
• Riis, V & Mai, W. 1988. Gas chromatographic determination of poly-
β-hydroxybutiric acid in microbial biomass after hydrochloric acid
propanolysis. J. Chromatogr. 445: 285- 289.
• Rocha,R. C. S. 2007. Estudo da biossíntese de poli-3-hidroxi-4-
pentenoato em Burkholderia cepacia. Tese (Doutorado) –
Instituto de Ciências Biomédicas, Universidade de São Paulo,
São Paulo, Brasil.
• Sambrook, J, Fritsch, E,. Maniatis, T. 1989. Molecular cloning: a
laboratory manual. 2ª Ed. New York : Cold Spring Harbor Lab.
Press.
• Silva, L.F.; Gomez, J.G.C.; Oliveira, M.S. 1992. Freeze-drying of
industrial yeast strains: influence of growth conditions, cooling rates

77
and suspending media on the viability of recovered cells. Rev.
Microbiol. 23: 117- 122.
• Steinbüchel A, Füchstenbusch B. 1998. Bacterial and other
biological systems for polyester production. Trends Biotechnol
.16:419-427
• Steinbüchel A. 2001. Perspectives for biotechnological production
and utilization of biopolymers: metabolic engineering of
polyhydroxyalkanoate biosynthesis pathways as a successful
example. Macromol. Biosci. 1:1-24.
• Steinbuchel A. Lutke – Eversloh T. 2003. Metabolic engineering
and pathway construction for biotechnological production of
relevant polyhydroxyalcanoates in microorganisms. Biochem. Eng.
J. V. 16 p. 81 – 96.
• Steinbüchel, A. & Valentin, H.E. 1995. Diversity of bacterial
polyhydroxyalkanoic acids. FEMS Microbiology letters. 128: 219-
228.
• Steinbüchel, A. & Valentin, H.E. 1995. Diversity of bacterial
polyhydroxyalkanoic acids. FEMS Microbiology letters. 128: 219-
228.
• Steinbuchel, A. 1996. PHB and other polyhydroxyalkanoic acids. In:
Rehm, H.J., Reed, G., Roehr, M. (Ed.) Products of primary
metabolinm, John & Sons, p. 405 - 464.
• Sudesh K, Abe H, Doi Y. 2000 Synthesis, structure and properties
of polyhydroxyalkanoates: biological polyesters. Prog. Polym. Sci.
25:1503-1555.

78
• Suriyamongkol P, Weselake R, Narine S, Moloney M, Shah S.
2007. Biotechnological approaches for the production of
polyhydroxyalkanoates in microorganisms and plants- A review.
Biotechnology Advances. 25. 148-175.
• Misra K. Superb, Sabeel P. Valappil, Ipsita Roy, Aldo R.
Boccannini. 2006. Polyhydroxyalkanoate (PHA)/ Inorganic Phase
Composites for Tissue Engineering Applications. American
Chemical Society. 7:8: 2249-2258.
• Van der Walle, G.A.M.; Buisman, G.J.H.; Weusthuis, R.A. 1999.
Development og environmentally friendly coatings and paints using
medium-chain- length poly(3-hydroxyalkanoates) as the polymer
binder. Int. J. Biol. Macrool. 25: 123-128.
• Williams S. Martin D. 2008. Applications of PHAs in Medicine and
Pharmacy. Tepha, Inc. 91-103.
• Zheng Z, Zhang MJ, Zhang G, Chen GQ. 2004. Production of 3-
hydroxydecanoic acid by recombinant Escherichia coli HB101
harboring phaG gene. Antonie van Leeuwnhoek. 85: 93- 101.
• Zinn M, Witholt B, Egli T. 2001. Occurrence, synthesis and medical
application of bacterial polyhydroxyalkanoate. Adv. Drug Rev.
53:5-21.

79
10. ANEXOS
ANEXO 1
• MEDIO MINERAL
ELEMENTO CONCENTRACIÓN
Na2HPO4 3,5 g/L
KH2PO4 1,5 g/L
MgSO4 . 7H2O 0,20 g/L
CaCl2 . 2H2O 0,01 g/L
Citrato férrico amoniacal 0,06g
Solución de Elementos Trazos 1ml
(NH4)2SO4 0,2 g/L (para medio sólido)
(NH4)2SO4 0,5 g/L (para medio liquido)
• Fuentes de carbono para medio mineral de acumulo
ELEMENTO CONCENTRACIÓN
Glucosa 10 g/L
Acido Octanóico 3 g/L

80
ANEXO 2
• CALDO NUTRITIVO
ELEMENTO CONCENTRACIÓN
Peptona 5 g/L
Extracto de carne 3 g/L

81
ANEXO 3
• MEDIO LURIA BERTANI (LB)
ELEMENTO CONCENTRACIÓN
Triptona 10 g/L
Extracto de levadura 5 g/L
NaCl 5 g/L
• ANTIBIOTICOS Y CONCENTRACIONES UTILIZADAS EN LOS
MEDIOS LBA Y LBK
Antibiótico Solución Stock
(mg/ml)
Concentración final
(µg/mL)
Ampicilina (A) sal
sodico
100 (en H2O) 100
Canamicina (K) 50 (en H2O) 50

82
ANEXO 4
• Buffer de transformación:
Tris/HCl (Hidrocloreto de hidroximetil aminometano) 10 mM CaCl2 50 mM MgCl2 10 mM MgSO4 10 mM
Água destilada qsp 1000 mL pH 8,0


















