
Contribución al conocimiento de los Catopidae (excepto...
23
Contribución al conocimiento de los Catopidae (excepto subfamilia Bathyxiinae) de la Península Ibérica (Coleoptera staphylinoidea) Marina Blas Esteban ADVERTIMENT. La consulta d’aquesta tesi queda condicionada a l’acceptació de les següents condicions d'ús: La difusió d’aquesta tesi per mitjà del servei TDX (www.tesisenxarxa.net ) ha estat autoritzada pels titulars dels drets de propietat intel·lectual únicament per a usos privats emmarcats en activitats d’investigació i docència. No s’autoritza la seva reproducció amb finalitats de lucre ni la seva difusió i posada a disposició des d’un lloc aliè al servei TDX. No s’autoritza la presentació del seu contingut en una finestra o marc aliè a TDX (framing). Aquesta reserva de drets afecta tant al resum de presentació de la tesi com als seus continguts. En la utilització o cita de parts de la tesi és obligat indicar el nom de la persona autora. ADVERTENCIA. La consulta de esta tesis queda condicionada a la aceptación de las siguientes condiciones de uso: La difusión de esta tesis por medio del servicio TDR (www.tesisenred.net ) ha sido autorizada por los titulares de los derechos de propiedad intelectual únicamente para usos privados enmarcados en actividades de investigación y docencia. No se autoriza su reproducción con finalidades de lucro ni su difusión y puesta a disposición desde un sitio ajeno al servicio TDR. No se autoriza la presentación de su contenido en una ventana o marco ajeno a TDR (framing). Esta reserva de derechos afecta tanto al resumen de presentación de la tesis como a sus contenidos. En la utilización o cita de partes de la tesis es obligado indicar el nombre de la persona autora. WARNING. On having consulted this thesis you’re accepting the following use conditions: Spreading this thesis by the TDX (www.tesisenxarxa.net ) service has been authorized by the titular of the intellectual property rights only for private uses placed in investigation and teaching activities. Reproduction with lucrative aims is not authorized neither its spreading and availability from a site foreign to the TDX service. Introducing its content in a window or frame foreign to the TDX service is not authorized (framing). This rights affect to the presentation summary of the thesis as well as to its contents. In the using or citation of parts of the thesis it’s obliged to indicate the name of the author.
Transcript of Contribución al conocimiento de los Catopidae (excepto...

Contribución al conocimiento de los Catopidae (excepto subfamilia Bathyxiinae) de la Península
Ibérica (Coleoptera staphylinoidea)
Marina Blas Esteban
ADVERTIMENT. La consulta d’aquesta tesi queda condicionada a l’acceptació de les següents condicions d'ús: La difusió d’aquesta tesi per mitjà del servei TDX (www.tesisenxarxa.net) ha estat autoritzada pels titulars dels drets de propietat intel·lectual únicament per a usos privats emmarcats en activitats d’investigació i docència. No s’autoritza la seva reproducció amb finalitats de lucre ni la seva difusió i posada a disposició des d’un lloc aliè al servei TDX. No s’autoritza la presentació del seu contingut en una finestra o marc aliè a TDX (framing). Aquesta reserva de drets afecta tant al resum de presentació de la tesi com als seus continguts. En la utilització o cita de parts de la tesi és obligat indicar el nom de la persona autora. ADVERTENCIA. La consulta de esta tesis queda condicionada a la aceptación de las siguientes condiciones de uso: La difusión de esta tesis por medio del servicio TDR (www.tesisenred.net) ha sido autorizada por los titulares de los derechos de propiedad intelectual únicamente para usos privados enmarcados en actividades de investigación y docencia. No se autoriza su reproducción con finalidades de lucro ni su difusión y puesta a disposición desde un sitio ajeno al servicio TDR. No se autoriza la presentación de su contenido en una ventana o marco ajeno a TDR (framing). Esta reserva de derechos afecta tanto al resumen de presentación de la tesis como a sus contenidos. En la utilización o cita de partes de la tesis es obligado indicar el nombre de la persona autora. WARNING. On having consulted this thesis you’re accepting the following use conditions: Spreading this thesis by the TDX (www.tesisenxarxa.net) service has been authorized by the titular of the intellectual property rights only for private uses placed in investigation and teaching activities. Reproduction with lucrative aims is not authorized neither its spreading and availability from a site foreign to the TDX service. Introducing its content in a window or frame foreign to the TDX service is not authorized (framing). This rights affect to the presentation summary of the thesis as well as to its contents. In the using or citation of parts of the thesis it’s obliged to indicate the name of the author.

DEPARTAMENTO DE ZOOLOGÍA FACULTAD DE BIOLOGÍA
UNIVERSIDAD DE BARCELONA
"Contribución a l conocimiento de los Catopidae (excepto la subfam.Bathyscilnae) de l a Península Ibérica. (Coleóptera Sta-phylinoidea
Memoria redactada para optar a l grado de Doctor en Ciencias Biológicas presentada por l a Licenciada Marina Blas Esteban.
VQB2 del Director de la Memoria Enrique Gadea Buisán, Catedrático - Director del Departamento de Zoología de l a Facultad de Biología de l a Universidad de Barcelona.
Barcelona, a de de

46
El palpo maxilar está formado por tres artejos, e l último mucho más corto que e l penúltimo (subfam.Anemadinae) o bien aproximadamente de l a misma longitud (subf ams. Ptomaphag inae y Catopinae).
3.- TÓRAX
A.- FORMA del protórax variable, en general transverso. En algunos casos l a variabilidad intraespecíflea es superior a l a existente entre especies diferentes, aunque por lo común es bastante uniforme dentro de cada grupo (género),
PIEZAS ESTERNALES. La familia Catopidae se caracteriza y diferencia de las más próximas por presentar las cavidades coxales anteriores cerradas por detrás, separadas del borde posterior del prostemón; por los epimeritos p-rotorácicos cuadrangulares; por m ser visibles los estigmas protorácicos; por las cavidades coxales intermedias no transversas; y por e l epistemito mesotorácico sin alcanzar e l borde posterior de dicha cavidad.
Las tres subfamilias presentes en nuestra fauna se separan con relativa facilidad por l a siguiente combinación de caracteres esternales.
a) Subfam.Ptomaphaginae : mesosternón con una marcada q u i l l a longitudinal media; epistemitos me so torácicos separados de los epimeritos pos una sutura; cavidades coxales intermedias separadas por tma apófisis mesostemal soldada a l mesosternón; epistemitos metatorácicos anchos, triangulares o trapezoidales; cavidades coxales posteriores contiguas.

47
b) Subfam. Anemadinae : mesosternón con qu i l l a longitudinal media (tribu Anemadini) o sin e l l a (tribu Pa-racatopini), cuando existe poco marcada; apófisis mesos-temal libre, no soldada a l mesosternón; cavidades coxales intermedias confluentes; epistemitos metatorácicos lineares, no prolongados hacia detrás; cavidades coxales posteriores confluentes.
c) Subfam.Catopinae : mesosternón plano, sin quil l a longitudinal media; epimeritos mesotorácicos transversos, separados de los epistemitos por una sutura; apófisis meso sternal libre, no soldada a l metastemón; cavidades coxales intermedias confluentes; epistemitos metatorácicos l i neares; cavidades coxales posteriores contiguas.
C. - ÉLITROS , dé forma variable, generalmente más o menos ovalados y alargactos, convexos, con l a estria sutural siempre manifiesta, las discales por e l contrario, faltan en muchos casos, formadas tanto una como otras generalmente por series de puntos alineados longitudinalmente. En algunos casos existen también estriólas transversas (subfams.Ptomapha-ginae y Anemadinae). El ángulo sutural variable en algunos casos según los sexos.
D. - PATAS , en unos casos cortas y gruesas y en otros más o menos largas y finas.
Coxas sin caracteres morfológicos especiales, muy desarrolladas en e l gen.Choleva L a t r e i l l e ; trocánteres muy modificados en los d'd' de algunos grupos, presentando una seri e de características constantes a nivel especifico; fémures de estmctura bastante f i j a , solamente los Ó6 de Speo-

48
nemadus Jeannel presentan sobre ellos un surco en e l que se encaja l a q u i l l a t i b i a l durante la flexión y los t/d* de a l gunos Catops Paykull presentan sobre la cara ventral un tubérculo ; tibias de forma muy variable, constante a nivel específico, en algunos casos muy modificadas en los <^<f , s i nuosas, torcidas o con tubérculos, simples en las , aima-dura apical de las tibias muy constante a nivel de subfamil i a y de tribu, constituida por un peine apical de espinas cortas e iguales (subfam.Ptomaphag inae), por dos espolones internos, dos extemos y por una serie de espinas irregulares alineadas sobre dicho borde sin formar una corona (subfam. A£iemadinae y tribu Catopini) en los tres pares de patas¿ con esta misma estructura en las patas anteriores en los representantes de l a tribu Cholevini, mientras que en las intermedias y posteriores está constituida pos dos espolones internos y por una serie de espinas cortas e iguales formando una corona regular apical; tarsos de cinco artejos, los del primer par de patas siempre dilatados en los 66 , e l primero de los intermedios dilatado en los 66 de l a tribu Catopini y los dos primeros artejos de dicho par dilatados en los dcf de l a subfam.Anemadinae.
4.- ABDOMEN .
A.- MORFOLOGlA ABDOMINAL. La morfología abdominal nos sirve para establecer las grandes divisiones del orden, así como para la ordenación natural de las diferentes familias que lo integran e incluso a nivel específico.
La importancia de dicho carácter, fue puesta de ma-

49
n i f i e s t o por JEANNEL & PAULIAN (1944), l o s cuales de acuerdo con él, modlficaxon l a sistemática general d e l grupo. Por o t r a parte se puede s e g u i r muy b i e n l a evolución de l a est r u c t u r a abdominal, por l o que es un carácter válido para e s t a b l e c e r l a f i l o g e n i a d e l grupo, aunque ésta, se ha de apoy a r también en o t r o s c a r a c t e r e s t a l e s como l a e s t r u c t u r a l a r v a l , e s t r u c t u r a a l a r , t i p o de c i c l o biológico, e t c . .
En l o s Catopidae, e l abdomen es de t i p o haplogas-t r o , con s e i s e s t e m l t o s v i s i b l e s , salvo en l o s cíd* de l o s Anemadinae en l o s que son v i s i b l e s s i e t e , ya que e l segmento g e n i t a l no está invaginacki y con octo t e r g u l t o s m a n i f i e s t o s , de e l l o s l o s s e i s primeros fusionados y muy poco q u l t i -n lzados, aunque se mantiene en gene r a l l a separación entre e l l o s , l o s dos t e r g u l t o s r e s t a n t e s más. e s c l e r i f i c a d o s f o r man p a r t e de l o s dos últimos segmentos abdominales. E l t e r -g u l t o correspondiente a l segmento g e n i t a l (92), sólo es v i s i b l e en l o s dcf de l o s Anemadinae por no e s t a r invaginado dicho segmento, a l c o n t r a r i o de l o que sucede en l o s r e s -tantes grupos.
B«- SEGMENTO GENITAL Y A P m P U M GENITAL. Estos c a r a c t e r e s han. de ser estudiados por separado en e l cf , ya que l a a r madura g e n i t a l o edeago no parece ser de o r i g e n segmentario, correspondiendo a una esclerificación de l a p a r t e t e r m i n a l d e l c a n a l eyaculador, no teniendo por e l l o ninguna relación con e l segmento g e n i t a l . No sucede así en l a ̂ , en l a que l a araiadura g e n i t a l es de o r i g e n segmentario, correspondiendo a l o s apéndices d e l 92 segmento abdominal o segmento gen i t a l , por e l l o en este caso, ambos c a r a c t e r e s se han de est u d i a r simultáneamente.

50
* SEGMENTO GENITAL MASCULINO. Corresponde a l igual que en l a ̂ a l 92 segmento abdominal, estando muy modificado en l a mayor parte de los éé , menos en las , dependiendo también del grupo de que se trate. En general está invaginado,
a) Subfam,Ptomaphaginae. Segmento genital muy modificado (fig,l7 d), reducido a un anillo en gran parte membranoso, el terguito muy poco quitinizado y e l estemito reducido a una especie de manubrio con e l que se articulan los dos pleuritos que tienen forma de paleta,
b) Subfam,Anemadinae, Segmento genital poco modificado (figs. 21 d y 24 b), formado por un terguito pubescente en e l ápice, dos pleuritos también pubescentes en el ápice y un estemito bastante modificado, a modo de manubrio, ocupando la linea media. Este segmento no está invaginado, por lo que siempre es visib l e ,
c) Subfam,Catopinae, Segmento genital muy modifi-«
cado, reducido a un anillo en gran parte membranoso (figs, 30 e, 45 e y 38 b), formado por un terguito y un pleuroes-temito pubescente en e l ápice,
** ARMADURA GENITAL MASCULINA. En e l cf dicha armadura parece no ser de origen segmentario, correspondiendo a una esclerificación de l a región d i s t a l del canal eyaculador. Es de tipo articulado, diferenciando dos partes, e l lóbulo medio, en cuyo interior se sitúa e l saco interno que está en conexión con e l canal eyaculador y e l tegmen, formado por los estilos y por l a lámina basal; los estilos se articulan sobre la cara dorsal del lóbulo medio, aunque secundariamente pueden cambiar de posición.

51
La e s t r u c t u r a d e l edeago es muy constante a n i v e l e s p e c i f i c o , siendo por tanto vn carácter válido para separar especies, géneros y grupos su p e r i o r e s . Por o t r a p a r t e su única función es i n t e r v e n i r en e l momento de l a cópula, por l o que no está afectada por fenómenos ada p t a t i v o s y de convergencia.
'^*~ Lóbulo medio, de forma v a r i a b l e , en ge n e r a l b i e n d e s a r r o l l a d o , simétrico y más o menos arqueado h a c i a l a cara v e n t r a l ; e l o r i f i c i o a p i c a l de dicho lóbulo v e n t r a l , aunque secundariamente puede pasar a ser d o r s a l en algunos grupos; a menudo presenta una prolongación v e n t r a l denominada lámina v e n t r a l , que rodea l a base d e l saco i n t e r n o , f a l tando únicamente en l o s Ptomaphag inae.
a ) Subfam.Ptomaphaginae, en e l l a e l lóbulo medio presenta una gran v a r i a b i l i d a d , dándose todos l o s t i p o s pos i b l e s . En l a mayor parte de l a s e s p e c i e s es simétrico y en l a s más p r i m i t i v a s e l o r i f i c i o a p i c a l se mantiene v e n t r a l , mientras que en l a s más evolucionadas, dicho o r i f i c i o ha pasado a ser d o r s a l , desplazándose a l borde derecho o a l i z quierdo según l o s géneros; este desplazamiento produce una c i e r t a asimetría que en alg m o s - c a s o s es muy marcada ( a l g u -
A d e l o p s i s P o r t e v i n ) y m o d i f i c a c i o n e s importantes d e l edeago.
En l o s representantes ibéricos de e s t a s u b f a m i l i a , e l lóbulo medio es simétrico, p r o v i s t o de unos p e l o s sensor i a l e s en e l ápice y e l o r i f i c i o a p i c a l se h a l l a desplazado a l borde i z q u i e r d o de l a c a r a d o r s a l . Carecen de lámina vent r a l .
b) Subfam.Anemadinae, en sus rep r e s e n t a n t e s e l ló-

5?.
bulo medio es simétrico, recto o moderadamente curvado hacia la cara ventral, el o r i f i c i o apical de mismo ventral, sin lígulas y con lámina ventral.
En el gen. Speonemadus Jeannel, e l tegmento de la región apical del lóbulo medio presenta unas pequeñas rugosidades en las que se insertan pelos (foto 28).
c) Subfam.Catopinae. En este grupo, e l lóbulo medio es de forma variable, en general tubular, simétrico y muy curvado hacia la cara ventral, en algunos casos muy comprimido lateralmente; e l o r i f i c i o apical de éste ventral, con lígulas pubescentes o no; ápice del lóbulo medio normalmente entero, pero en ocasiones los lóbulos laterales se mantienen independientes, por lo que en estos casos parece bifido o trífido según el desarrollo de dichos lóbulos y frecuentemente presenta sobre la cara dorsal surcos más o menos proftmdos. Siempre con lámina ventral.
Tegmen, diferencia dos zonas, los estilos y la lámina basal, su desarrollo depende del grupo de que se trate.
a) Subfam.Ptomaphaginae. Estilos insertos lateralmente en la base del lóbulo medio, poco desarrollados, muy gráciles y unidos a dicho lóbulo, armados de varias sedas en el ápice. Lámina basal totalmente atrofiada.
b) Subfam.Anemadinae. Estilos insertos dorsalmen-te en l a base del lóbulo medio, robustos, sobrepasando en algunos casos la longitud de dicho lóbulo, armados de varios dientes y sedas en e l ápice; e l número y situación de tonos y otros es constante a nivel específico. En e l gen. Speonemadus Jeannel los estilos poseen unos orgánulos eva-
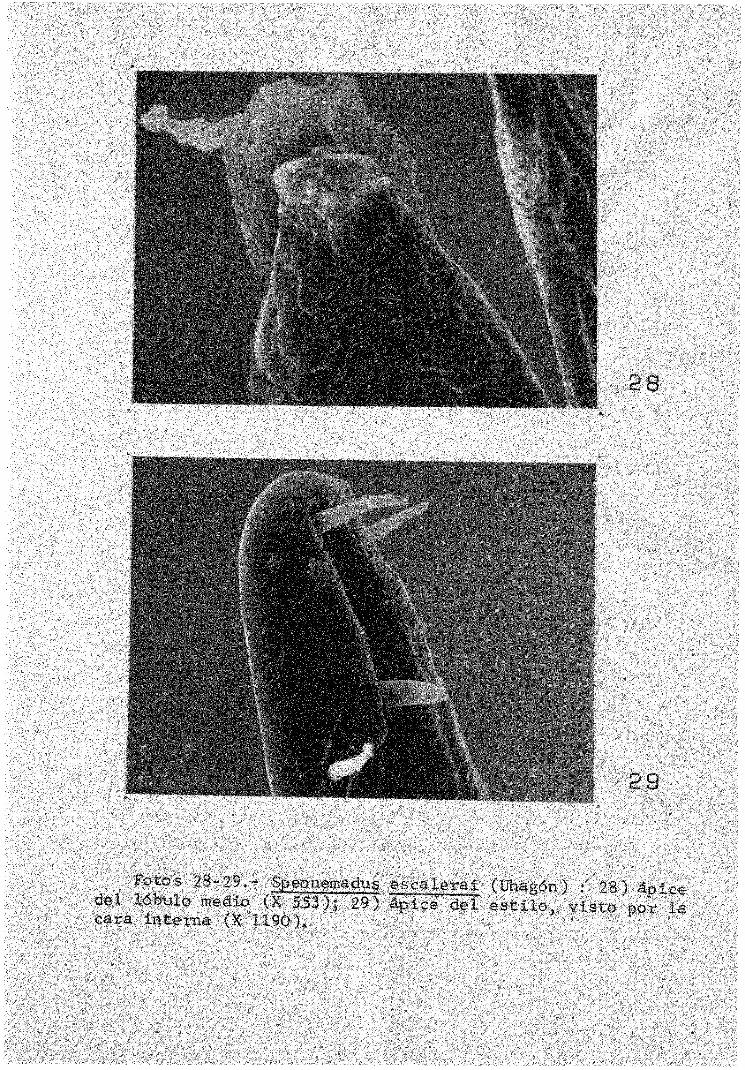
Fotos 28-29.- Speonemadus e s c a l e r a ! (Uhagón) : 28) ápice d e l lóbulo medio (X 553); 29) ápice d e l e s t i l o , v i s t o por l a cara i n t e r n a (X 1190),

54
ginables, posiblemente con función sensorial (foto 29). Lámina basal reducida a xjna estrecha banda quitinosa en los representantes Ibéricos,
c) Subfam.Catopinae, Estilos de forma variable, en la tribu Cholevini bien desarrollados, robustos, no afilados en e l ápice y armados en él de varias sedas, cuyo número y situación es constante a nivel especifico y en la tribu Catop i n i menos desarrollados, gráciles, y en general muy a f i l a dos en e l ápice, excepcionalmente engrosados (géneros Depros-cia Erichson y los Catops Paykull del grupo mor lo); en gener a l armado s de dos sedas en e l ápice. Lámina basal bien desarrollada.
En todos los casos e l número de sedas y espinas asi como su situación en los estilos es constante a nivel específico.
Y• ~ Saco interno 2. ̂ troadura copulatriz. E l saco in- . temo está en conexión directa con e l canal eyacula do r, siendo en'realidad l a parte terminal de éste. Su estructura es muy constante a nivel específico y en los casos más sencillos su superficie interna está revestida por una serie de escamas o dentículos quitinosos, que se levantan en todas las d i recciones cuando se evagina en e l momento de l a cópula, ya que en posición de reposo está invaginado' a modo de dedo de guante en e l interior del lóbulo medio. Sin embargo en mucho s casos, esta armadura copulatriz sencilla evo luciona, originándose a expensas de estas formaciones primitivas dientes, faneras, espinas, estiletes, etc., de forma y situación constante a nivel específico.
Por otra parte, se observa el mismo tipo de piezas

55
copulatrices del saco interno^en especies diversamente adaptadas, pero que poseen caracteres de filiación comunes, por lo que de acuerdo con este carácter se puede saber e l grado de parentesco y evolución de las diferentes especies de un miaño grupo o género. Estudios de este tipo se han realizado en e l gen.Choleva L a t r e i l l e con excelentes resultados.
En e l presente trabajo, se ha intentado hacer un estudio de la armadura copulatriz en las diferentes subfamil i a s con ayuda del "scanning", obteniéndose imos resultá
is
dos favorables, aunque debido a las pocas especies estudiadas, no se pueden dar resultados definitivos.
a) Subfam.Ptomaphagínae. Es en este grupo donde el saco interno está más modificado, pues está armado de un gran estilete evaginable muy quitinizado, sin piezas qui t i -nosas accesorias. Este carácter junto con l a forma del lóbulo medio y situación del o r i f i c i o apical de éste, nos da idea del alto grado de evolución del grupo. Por otra parte jmto a- estos caracteres aparecen otros muy primitivos, lo que también nos indica, se trata de un grupo muy antiguo.
b) Subfam.Anemadinae. En las especies ibéricas de esta subfamilia, la armadura copulatriz está constituida por dos cadenas longitudinales de dientes quitino sos engranados por l a base y sirviendo de sostén articulado a l saco interno cuando se evagina; además algunas especies poseen un diente ventral accesorio. Ademas de las estructuras mencionadas, toda l a cara interna está tapizada de pequeñas espinas articuladas.
En las fotos 30 y 31 correspondiente s a Hormo sacus clathratus (Perris) y a Speonemadus escalerai (Uhagón) res-

Fotos 30-31.- Armadura d e l saco i n t e r n o de : 30) Hormosa-cus c l a t h r a t u s ( P e r r i s ) (X 1011); 31) Speonemadus e s c a l e r a ! (Uhagón) (X 559), ^

57
pectivamente, se pueden observar l a s dos cadenas de di e n t e s q u i t i n o s o s y l a s espinas que t a p i z a n e l saco i n t e r n o .
c ) Sub fam.Catop inae. Este grupo es e l que presenta m a mayor v a r i e d a d de formas en l a amadura d e l saco i n t e r n o . En p r i n c i p i o d i c h a armadura está c o n s t i t u i d a por una s e r i e de d i e n t e s a r t i c u l a d o s que t a p i z a n e l i n t e r i o r d e l saco i n terno; secundariamente en algunos géneros y especies estos d i e n t e s se han modificado y transformado en o t r a s e s t r u c t u ras.más complejas, que en algunos casos son u t i l i z a d a s como un carácter más para d e f i n i r grupos. Por e l l o se analizará su e s t r u c t u r a a n i v e l genérico en l o s grupos en que ha sido estudiada.
Gen,Nargus Thomson, Armadura d e l saco i n t e r n o const i t u i d a en g e n e r a l por un diente a p i c a l y v e n t r a l de forma v a r i a b l e y por una s e r i e de espinas y formaciones q u i t i n o s a s agrupadas en fascículos. En algunos casos e x i s t e un die n t e a d i c i o n a l .
Gen.Choleva L a t r e i l l e , Es en e s t e género donde se ha estudiado con mayor d e t a l l e l a e s t r u c t u r a y evolución de l a armadura c o p u l a t r i z . Por o t r a p a r t e es e l grupo en e l que.presenta una mayor complejidad y v a r i e d a d de formas.
En e l saco'interno de.-los. Choleva L a t r e i l l e , es nec e s a r i o d i s t i n g u i r dos regiones, l a a p i c a l y l a media y b a s a l , que han evolucionado independientemente; e s t a s r e g i o nes corresponden respectivamente a zona d e l saco i n t e r n o que se -evaglna durante l a cópula y a l a que queda permanentemente en e l i n t e r i o r d e l lóbulo medio por r e p r e s e n t a r l a c o n t i nuación d e l c a n a l eyaculador..
En l a s formas más p r i m i t i v a s , l a región a p i c a l es-

58
tá foxtnada por cuatro o cinco f i l a s longitudinales articuladas, dirigidas hacia e l ápice, pero en las más evolucionadas, estás espinas se han modificado, originando una serie de formaciones diferentes, siendo la más interesante de todas ellas e l diente ventral que existe en l a mayor parte de especies, todas las de Cho leva (s. str. ), que se ha formado a expensas de dos f i l a s de espinas convergentes hacia e l ápice. Esta transformación así como la posterior evolución de la forma del diente, se debe a la presión ejercida por dicha región del saco interno contra e l terguito del segmento gen i t a l femenino, ya que es la pieza responsable de l a abertura de las vias genitales femeninas, mediante e l levantamiento det terguito, por ello l a evolución del diente apical y del terguito del segmento genital femenino es correlativa por las presiones recíprocas (JEANNEL,1923).
En las especies en que f a l t a e l diente ventral, está sustituido por \ino dorsal, propio del subgén. Cholevop-sis Jeannel. En ocasiones esta región presenta también pequeñas agrupaciones de espinas articuladas que se erizan durante la cópula y que se pueden transformar incluso en dientes compuestos.
Por lo que se refiere a la zona media y basal, l a armadura primitiva del saco interno, a l igual que en la región apical, está constituida por bandas longitudinales de espinas dirigidas hacia dicha región, que se modifican y transforman en las formas más evolucionadas, en las que se especializan en órganos propulsores del esperma.
Parece ser, que la evolución de las piezas de l a región basal es anterior a l a de l a región apical y que d i cha evolución se ha realizado en pocas direcciones, obser-

59
vándose e l mismo t i p o de p i e z a s , en todas l a s e s p e c i e s de tana misma e s t i r p e , s i r v i e n d o por tanto para c a r a c t e r i z a r l a s , aunque no es éste, e l tkiico carácter u t i l i z a d o para e s t a b l e c e r l a filiación de l o s d i v e r s o s grupos de especies.
G e n .Sciodrepoides Hatch, La armadura c o p u l a t r i z está c a r a c t e r i z a d a por pre s e n t a r tina p i e z a q u i t i n p s a en f o r ma de Y en l a región b a s a l y fascículos de espinas y pequeños dientes en l a región media y a p i c a l .
Gen.Catops P a y k u l l . En este género se han r e a l i z a do hasta e l presente muy pocos e s t u d i o s sobre l a e s t r u c t u r a de l a armadura c o p u l a t r i z (ZWICK,1968), es por e l l o y por l a gran s i m i l i t u d morfológica de algunas especies muy estrechamente emparentadas/que se ha tratado de d e s c i f r a r y v a l o r a r d i c h a e s t r u c t u r a .
Aparentemente e l saco i n t e r n o , a l menos e l de l a s especies ibéricas, carece de p i e z a s q u i t i n o s a s complicadas, ya que no se ven por t r a n s p a r e n c i a a través d e l lóbulo medio como ocurre por ejemplo en e l gen. Choleva L a t r e i l l e . S i n embargo xma. vez extraído e l saco intern o y a b i e r t o l o n g i t u dinalmente para v e r con mayor d e t a l l e l a s formaciones que l o t a p i z a n , su e s t r u c t u r a deja de ser simple, presentando tana gran v a r i e d a d de formas.
Este e s t u d i o se ha r e a l i z a d o con e l "scanning" en v a r i a s especies d e l grupo d e l C.fuscus, observándose, que i n c l u s o dentro de una misma es p e c i e , l a forma de l a s e s p i nas varía según l a asna d e l saco i n t e r n o de que se t r a t e .
En todas l a s especies estudiadas se observa en l a región a p i c a l una formación q u i t i n o s a en forma de V i n v e r t i d a , que es doble ( f o t o s 32 y 33), correspondientes a Catops n i g r i c a n s (Spence), en l a f o t o 32 se pueden o b s e r v a r
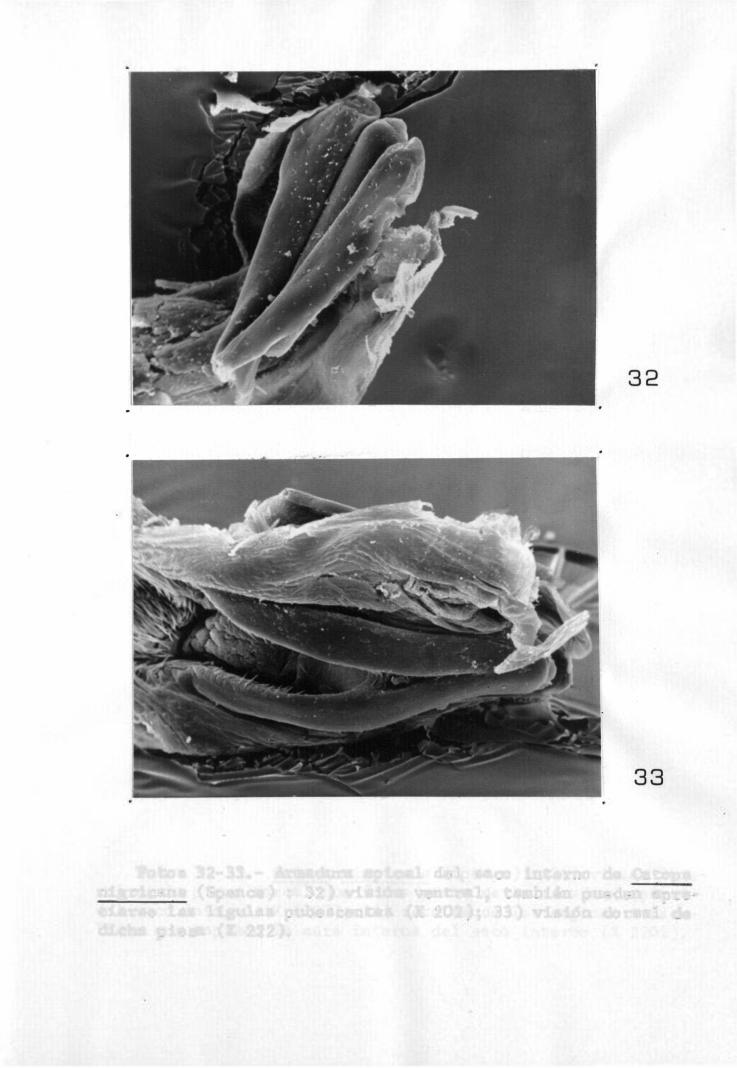
Fotos 32-33.- Armadura apical del saco interno de Catops nigrícans (Spence) : 32) visión ventral, también pueden apreciarse las lígulas pubescentes (X 202); 33) visión dorsal de dicha pieza (X 222).

Fotos 34-35.- Catops nígrlcans (Spence) : 34) visión l a t e r a l de l a p i e z a d o r s a l que forma p a r t e de l a armadura a p i c a l d e l saco i n t e r n o (X 1100); 35) d e t a l l e de l a s espinas a r t i c u ladas que t a p i z a n l a cara i n t e r n a d e l saco i n t e r n o (X 2201),
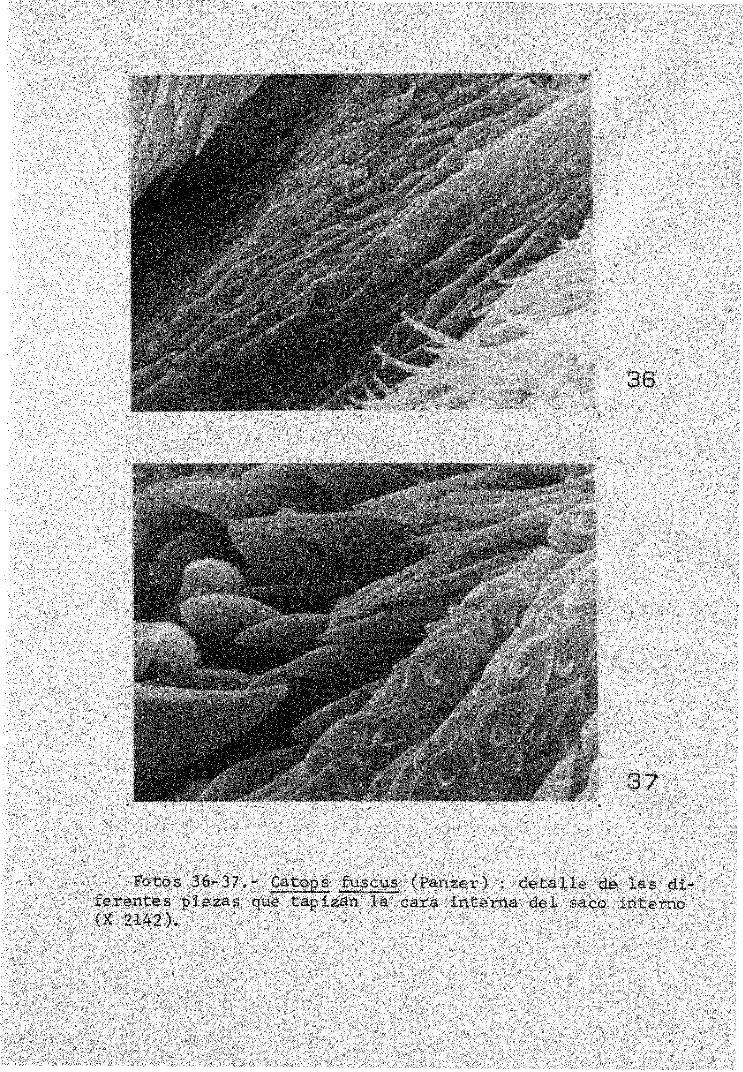
Fotos 36-37.- Catops fuscus (Panzer) : d e t a l l e de l a s d i f e r e n t e s p i e z a s que t a p i z a n l a cara i n t e r n a d e l saco i n t e r n o (X 2142).

Foto 38.- Catops fuscus (Panzer), detalle de las piezas que tapizan la cara interna del saco interno (X 5355).
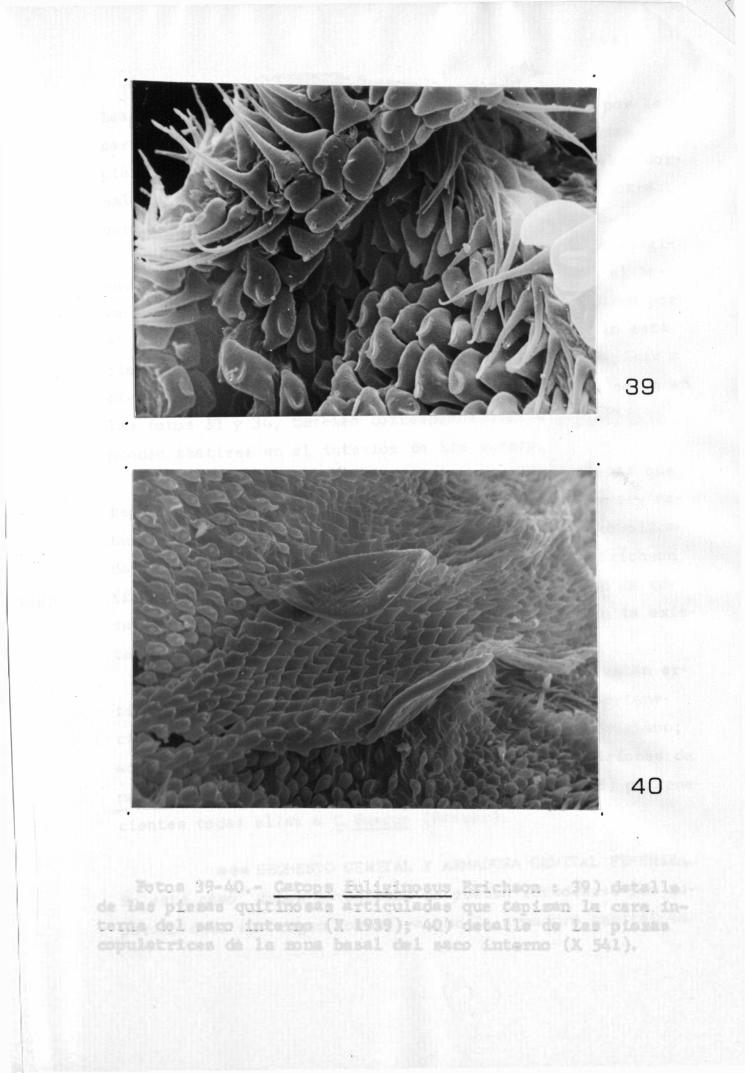
Fotos 39-40.- Catops fullglnosus Erichson : 39) detalle de las piezas quitinosas articuladas que tapizan la cara interna del saco interno (X 1939); 40) detalle de las piezas copulatrices de l a zona basal del saco interno (X 541).
I 1

65
las lígulas pubescentes y la pieza mencionada vista por la cara ventral y en la foto 33 se puede observar la misma pieza vista por l a cara dorsal. Ambas piezas, ventral y dorsal están unidas por e l ángulo de la V, zona que por otra parte está en contacto con e l ápice del lóbulo medio.
Esta pieza juega un papel importante en l a evagl-nación del saco interno, ya que se realiza mediante e l levantamiento de l a pieza dorsal, pasando e l saco interno por el espacio que queda entre los dos brazos de la V, En esta pieza se. observa por la cara interna una serie de espinas o dientes articulados de función desconocida que como se ve en las fotos 33 y 34, también correspondientes a C,nigricans pueden abatirse en el interior de los surcos.
Por lo que se refiere a las restantes piezas que tapizan la cara interna del saco interno son de forma y naturaleza variada, no observándose formaciones muy complicadas, s i se exceptúa l a existente en C.fuligiñosus Erichson (foto 40) en l a región basal, aunque la complejidad de esta fotmaclón no es comparable en ningún momento con la existente en las especies de otros géneros (Choleva).
La mayor parte de las piezas observadas están articuladas como se puede ver en las fotos 35 y 39 pertenecientes a C,nigricans (Spence) y a C,fuligiñosus Erichson; además de las espinas típicas, existen otras formaciones de naturaleza y misión desconocida (fotos 36, 37 y 38) pertenecientes todas ellas a C,fuscus (Panzer).
*** SEGMENTO GENITAL Y ARMADURA GENITAL FEMENINA. En este caso, ambos caracteres no pueden estudiarse por separado dado su origen común, ya que la armadura genital es -

66
de origen segmentario, correspondiendo a los apéndices modificados del segmento genital.
a) Subfam.Ptomaphaginae. Segmento genital bien desarrollado, formado por un terguito pubescente en e l ápice, dos pleuritos también pubescentes y un esternito bastante modificado (fig.17 e), constituido por una membrana translúcida y por un par de apéndices, las gonapófisis fuertemente quitinizadas, compuestas de dos piezas, una lateral poco quitinizada y otra dorsal dirigida hacia atrás; esta última, el estilo, posee en e l ápice una larga seda sensorial, que juega un papel importante en e l determinismo de l a puesta (DELAY,1978) y varias sedas más cortas.
b) Subfam.Anemadinae. Segmanto genital bien desarrollado, fotmado por un terguito, dos pleuritos y un esternito. La estructura del esternito es muy similar a la del grupo anterior, aunque la fotma de las gonapófisis es diferente .
c) Sub fam.Ca top inae. Segmento genital bien desarrollado, diferenciando a l iguial que las subfamilias anteriores un terguito, dos pleuritos y un esternito; la estructura de este último, también es s'imllar a l a de los grupos precedentes. E l terguito en algunos casos (géns.Choleva L a t r e i l l e y Sciodrepoides Hatch) está muy quitinizado, siendo su forma muy constante a nivel específico, por lo que resulta un carácter válido para determinar las (figs.35 c, 36 b, 37 b, 38 c, 40 b, 45 f y 46 d) de las diferentes especies. En los restantes géneros ibéricos dicho terguito está muy poco quitinizado, siendo su forma muy parecida en las diferentes especies, por lo que no se usa en su sistemática.


















