DESARROLLO EMBRIONARIO TEMPRANO 2009
16
UNIVERSIDAD CENTRAL DE VENEZUELA FACULTAD DE CIENCIAS VETERINARIAS CÁTEDRA DE HISTOLOGÍA VETERINARIA GUÍA TEÓRICA DEL DESARROLLO EMBRIONARIO TEMPRANO DE LOS ANIMALES MATERIAL PREPARADO POR: PROFESOR FRANCISCO J. CABRERA A. Abril de 2009
-
Upload
estudiantes-de-veterinaria-primer-ano-2009 -
Category
Documents
-
view
3.034 -
download
3
Transcript of DESARROLLO EMBRIONARIO TEMPRANO 2009

UNIVERSIDAD CENTRAL DE VENEZUELA
FACULTAD DE CIENCIAS VETERINARIAS
CCÁÁTTEEDDRRAA DDEE HHIISSTTOOLLOOGGÍÍAA VVEETTEERRIINNAARRIIAA
GUÍA TEÓRICA DEL DESARROLLO EMBRIONARIO TEMPRANO DE LOS
ANIMALES
MATERIAL PREPARADO POR:
PROFESOR FRANCISCO J. CABRERA A.
Abril de 2009

IINNTTRROODDUUCCCCIIÓÓNN
Con este material se pretende facilitar la compresión de las diferentes etapas que
se suceden durante el desarrollo embriológico, los cuales muchas veces no están
bien desarrollados en algunos libros, en otros aparecen dispersos y/o con
diferentes términos, lo cual dificulta el aprendizaje, por ello se hace un resumen de
manera didáctica, clara y comprensible.
OOrriieennttaacciióónn
Este material de ninguna manera pretende sustituir a los libros, sino facilitar la
comprensión de los mismos, por lo cual este material no sustituye la bibliografía
recomendada.
Objetivos
Facilitar la comprensión, análisis y conocimiento de los procesos y etapas
fundamentales en el desarrollo embriológico, así como los diferentes métodos de
abordaje del diagnóstico.
Estos objetivos serán reforzados en el laboratorio de prácticas docentes, donde se
le demostrará material audiovisual, rotafolio, fotografías y proyección de videos
con este fin.
Se organiza el contenido partiendo de los conceptos básicos, pasando por las
ramas, procesos iniciales, diferenciación y sus procesos morfogenéticos, etc.
contando con la asistencia del profesor, pasantes, preparadores y técnicos. Se
contará con microscopios, láminas histológicas, rotafolio, proyector de diapositivas
o multimedia de acuerdo a la disponibilidad del recurso, se proyectará en
televisores imágenes del microscopio con salida a través de una cámara de video
adaptada al microscopio, y videos a través del VHS y televisores.
El estudiante se evaluará en teoría y práctica con preguntas claras, concisas y
objetivas que permitan evaluar la consecución de los objetivos.
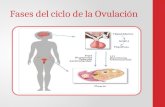
Conceptos Básicos

Cigoto : Es el óvulo fecundado o embrión unicelular, con la carga genética de su
especie restituida a su valor normal.
Tipos de cigoto
Según su tamaño
Microlecito Mesolecito Macrolecito
Según la cantidad de vitelo
Oligolecito Mesolecito Polilecito
Según la distribución del vitelo
Isolecito Vitelo distribuido por gradiente
Telolecito
Especies Invertebrados y mamíferos
Peces y anfibios
Reptiles y aves
Segmentación
Primer proceso del desarrollo embrionario, caracterizada por el clivaje 1 del cigoto.
MÓRULA : Embrión resultante de la segmentación, formado por una masa de 8 a
32 células (llamadas BLASTÓMERAS ).
Tipos de segmentación : depende de la cantidad de vitelo presente en el cigoto.
1. Holoblástica e igual o típica : da origen a la mórula de los
invertebrados y se caracteriza por un proceso de clivaje uniforme, en el
cual todas las blastómeras poseen el mismo tamaño.
2. Holoblástica desigual: da origen a la mórula de los peces y anfibios y
se caracteriza porque el clivaje de las blastómeras en el polo animal es
más veloz que en el polo vegetativo. Como consecuencia, las
blastómeras del polo animal son más pequeñas y numerosas
(micrómeras ) que las del polo opuesto, denominadas macrómeras . Este

evento ocurre por la acumulación de grandes gotas de vitelo en el polo
vegetativo.
3. Meroblástica o parcial: por este proceso se origina la mórula discoidal
de aves y reptiles , y se caracteriza por la división incompleta del
citoplasma del cigoto. En consecuencia, las blastómeras forman un
sincitio 2 temporal, el cual da paso a un embrión plano.
4. Holoblástica igual atípica: da origen a la mórula de los mamíferos , en
la cual el proceso de clivaje se realiza rotando en 90º el eje de división
celular en cada nueva mitosis. Morfológicamente, esta mórula es similar a
la de los invertebrados, pero su destino es diferente.
Repaso de conceptos :
1. Clivaje: Mitosis sucesivas, que no son sucedidas por crecimiento celular, dando
origen a células cada vez más pequeñas.
2. Sincitio: Masa de protoplasma multinucleado, obtenido por mitosis sin
separación (citodiéresis) del citoplasma de las células hijas (División Amitótica).

BLASTOGÉNESIS
Segundo proceso del desarrollo embrionario, caracterizado por la formación de
una cavidad (blastocele ) en el interior del embrión (ahora llamado blástula ), el
cual está formado por una capa (blastodermo ) de células.
a. BLASTOGÉNESIS EN
INVERTEBRADOS: el
proceso de
blastogénesis en los
invertebrados se
cumple por cavitación
de la masa celular de la mórula. Esto ocurre porque las blastómeras
segregan líquido, el cual se acumula en el centro del embrión. Esto produce
un embrión hueco, monodérmico ( es decir, formado por una capa de
células), llamado blástula típica de los invertebrados . La cavidad,
denominada blastocele , suele ser perfectamente simétrica y centrada.
b. BLASTOGÉNESIS EN PECES Y ANFIBIOS: debido a la asimetría
existente entre las micrómeras y las macrómeras, la cavitación de la mórula
ocurre por encima de la línea ecuatorial, es decir, a nivel del polo animal. El
nuevo embrión recibe el nombre de anfiblástula.

c. BLASTOGÉNESIS EN AVES Y REPTILES: en estas especies, el disco
embrionario descansa sobre el vitelo, separado de este por una cavidad
denominada cavidad sub-germinal. El propio disco embrionario es bastante
grueso, y sufre un proceso de delaminación que lo separa en dos capas,
unidas por los bordes. Ambas capas se encuentran separadas por el
blastocele y en lugar de tomar el nombre genérico de blastodermo, pasan a
denominarse epiblasto (la capa superior o superficial) e hipoblasto (la
capa inferior, más cercana al vitelo)
d. BLASTOGÉNESIS EN
MAMÍFEROS: ocurre
siguiendo el mismo proceso
que en los invertebrados,
con la única diferencia de
que se forma una masa
celular en el interior del
blastocele, adherida al polo
animal y que se conoce con
el nombre de nódulo
embrionario . Este nódulo
dará origen a las
estructuras propiamente embrionarias, mientras que el blastodermo dará
origen a las estructuras placentarias. Como paso final de este proceso, el
nódulo embrionario sufre una cavitación que formará a la cavidad
amniótica , separada del blastocele por un disco embrionario similar al de
las aves
6. Concepto de Gastrulación. Tercer proceso del desarrollo embrionario,
caracterizado por la formación de dos hojas germinales (ectodermo y
endodermo ) y por la aparición del primer eje longitudinal medial (línea
primitiva )

7. Mecanismos Fundamentales y Accesorios de la Gast rulación.
1. MECANISMOS FUNDAMENTALES DE LA GASTRULACIÓN:
1.1. Multiplicación celular
1.2. Crecimiento celular
1.3. Expansión celular
1.4. Delaminación
1.5. Invaginación
2. MECANISMOS ACCESORIOS DE LA GASTRULACIÓN:
2.1. Epibolia : consiste en la multiplicación y expansión de las micrómeras
(en peces y anfibios) o del hipoblasto de las aves, reptiles y mamíferos, con el
fin de encerrar la totalidad del vitelo dentro de un saco (saco vitelino )
2.2. Confluencia : por medio de los tres primeros mecanismos
fundamentales, algunas células del epiblasto o del ectodermo dorsal se
agrupan en la línea media dorsal del embrión, dando origen a la línea
primitiva .
8. Gastrulación en diferentes especies y formación de las hojas germinales
(ectodermo y endodermo).
a. EL PROCESO DE
GASTRULACIÓN EN LOS
INVERTEBRADOS se cumple
por invaginación de la pared
de la blástula a nivel de la
línea ecuatorial. Esto produce
un embrión didérmico, (es
decir, formado por dos capas

de células en lugar de una) llamado gástrula . La cavidad del blastocele es
reducida a un pequeño espacio entre la capa externa (ectodermo) y la Interna
(endodermo). El endodermo delimita una nueva cavidad, llamada intestino
primitivo o arquenterón (de la palabra griega αρθυεντερων), la cual se abre al
exterior por una abertura llamada blastoporo .
b. EL PROCESO DE GASTRULACIÓN EN PECES Y ANFIBIOS es similar al de
los invertebrados, con la salvedad de que la invaginación, iniciada a nivel
ecuatorial, se desvía dorsalmente para colocar el intestino primitivo por encima
y a los lados del vitelo, definiendo de este modo el saco vitelino.
c. EL PROCESO DE
GASTRULACIÓN EN AVES,
REPTILES Y MAMÍFEROS tiene
características propias por tratarse
de embriones discoidales. El
epiblasto y el hipoblasto no se

redefinen como ectodermo y mesodermo, respectivamente, sino después de
que se ha formado la línea primitiva. A partir de la línea primitiva, ubicado en el
epiblasto, ocurre un proceso de delaminación celular . El conjunto de células
desprendidas o delaminadas del epiblasto reemplaza al hipoblasto original y
crea una tercera capa, intermedia, la cual es denominada mesodermo. Solo en
este momento se puede decir que hay un ectodermo (antiguo epiblasto) y un
endodermo (el nuevo hipoblasto). El hipoblasto original es desplazado fuera
del embrión discoidal, como parte del mecanismo de epibolia y pasando a
formar parte del endodermo extraembrionario del saco vitelino. En definitiva, a
diferencia de los animales inferiores, la gástrula de aves, reptiles y
mamíferos es un embrión tridérmico (de tres capas).
9. Concepto de Implantación: Proceso mediante el cual, el embrión de los
mamíferos, se
introduce y anida en
la mucosa uterina.
Durante este
proceso, el
blastodermo
extraembrionario se
transforma en
trofoblasto , un tejido
compuesto por dos
tipos celulares, el
cual dará origen a las
estructuras placentarias de origen fetal. El trofoblasto posee una capa externa
de células sincitiales , es decir fusionadas en una gran masa celular
multinucleada (sincitiotrofoblasto ), que responsable por la invasión del
endometrio y el anclaje del embrión al tejido materno. La capa interna del
trofoblasto está formada por células que no se fusionan (citotrofoblasto ), y es
responsable por la formación de la membrana coriónica de la placenta.

10. Formación del mesodermo y aparición de estructu ras neurales.
Esta etapa es conocida con el nombre de Neurulación y se caracteriza por el
desarrollo del sistema nervioso y el comienzo del desarrollo del tubo digestivo y
corazón. También ocurre la segmentación del mesodermo paraxial, lo que da
origen a las somitas. El mesodermo se forma por medio de tres mecanismos,
según la especie:
Mecanismo enterocelómico: mecanismo típico de los invertebrados . Consiste
en la diferenciación simultánea de células mesodérmicas desde el endodermo y
del ectodermo, en 2 capas separadas que delimitan el celoma o cavidad corporal
(mesodermo lateral). Las dos capas celulares de mesodermo se fusionan en la
región dorsal del embrión, originando las regiones paraxial e intermedia del
mesodermo.
Delaminación: este mecanismo está presente en peces y anfibios , en los cuales
las futuras células mesodérmicas se desprenden tanto del ectodermo a lo largo de
la línea primitiva, como de regiones laterales del mismo.
Proliferación de células desplazadas y acumuladas a lo largo de la línea media
dorsal, formando la línea primitiva. Las células desprendidas se ubican entre el
ectodermo y el endodermo, para luego diferenciarse en células mesenquimatosas
(el mesodermo) ubicadas en un eje longitudinal denominado Notocorda. A partir
de la notocorda ocurre la proliferación lateral de las células mesodérmicas,
formando el mesodermo paraxial, intermedio y lateral, en ese orden. Este
mecanismo es el típico de reptiles, aves y mamíferos (ver la figura abajo)

Las células mesenquimatosas proliferan en zonas específicas, donde son más
abundantes: A lo largo de la línea media, bajo el ectodermo (Notocorda ); a ambos
lados de la notocorda, formando una masa continua (mesodermo paraxial ), que
posteriormente se fracciona, formando segmentos que rigen la división del cuerpo
en zonas (somitas ). El mesodermo paraxial, segmentado en somitas, es
responsable de la formación de una gran cantidad de estructuras de carácter
conectivo: dermis de la piel, tejido óseo vertebral, músculos asociados a la
columna vertebral, gónadas, corteza adrenal, etc.
Se forma además una zona externa de mesodermo, el cual no se segmenta
(Mesodermo Intermedio ) y que da origen a los túbulos del sistema genito-urinario
y al sistema cardiovascular. Finalmente, el mesodermo forma dos hojas exteriores
o mesodermo lateral , separadas por una cavidad (celoma ). La hoja externa o
superior se mantiene en contacto con la cara interna del ectodermo, formando la
somatopleura (el término designa por igual tanto a este mesodermo, como a la
suma de mesodermo y ectodermo), mientras que la hoja inferior da origen a la
esplacnopleura al relacionarse con la cara externa del endodermo.
El desarrollo del sistema nervioso se inicia cuando la notocorda, plenamente
formada, induce un engrosamiento por mitosis en el ectodermo ubicado
sobre ella . Este engrosamiento, llamado placa neural , es el primer esbozo del
sistema nervioso . La placa neural se invagina a lo largo, dando origen al surco
neural . Posteriormente, se inicia el cierre del surco neural desde su mitad hacia
los extremos, transformándose en un tubo neural con aberturas en cada extremo
(Neuroporo anterior y posterior). Los neuroporos se cierran en su momento,
siendo el cierre del neuroporo anterior el que más tarda. De hecho, la
transformación del surco neural en tubo neural es más lenta a nivel craneal, ya
que en esta área el neuroepitelio se desarrolla muy rápido. Esto permite que el
tubo neural sea muy ancho cranealmente, formando un tubo dilatado en tres
bolsas (Vesículas Cefálicas Primarias: Prosencéfalo, Mesenc éfalo y

Rombencéfalo ) que dan origen al cerebro, cerebelo y médula oblonga. El resto
del tubo neural mantiene un diámetro uniforme y dará origen a la médula espinal.
El sistema nervioso periférico
se origina de los bordes del
surco neural, doblados hacia
fuera y a los lados del tubo
neural, para luego separarse y
segmentarse: las crestas
neurales . Estas crestas dan
origen a las células de
neuroepitelio a partir de las
cuales se derivan los ganglios
y nervios del sistema nervioso
periférico, las células
secretoras del Sistema Entero-endocrino, médula adrenal, células parafoliculares
de la tiroides y los melanocitos, entre otras.
11. Desarrollo del celoma:
El celoma es la cavidad interna
del embrión, la cual da origen en
su momento a las cavidades
torácica y abdominal. Es un
espacio delimitado por un
mesotelio (epitelio plano simple
originado por el mesodermo de
la somatopleura y la
esplacnopleura ) que en su

momento tomará el nombre anatómico de lámina pleural y lámina peritoneal. El
celoma existe como espacio una vez que el desarrollo de la somatopleura y la
esplacnopleura encierran el antiguo espacio del blastocele. De aquí se desprende
una definición válida: El blastocele, al ser encerrado en mesodermo, se transforma
en el celoma.
En el caso de los anfibios, peces y invertebrados, la aparición del celoma no
implica ningún cambio especial, ya que la totalidad del embrión y su pequeño saco
vitelino han quedado encerrados por el ectodermo durante la gastrulación (Ver el
concepto de Epibolia). El caso es distinto para los animales amniotas (reptiles,
aves y mamíferos) Cuyos embriones son planos incluso tras haber empezado la
Neurulación. Se inicia, en estas especies, un movimiento de tejidos,
específicamente la somatopleura, que logra cerrarse sobre la zona dorsal del
endodermo, separando el tubo digestivo primitivo del saco vitelino. Se logra,
además, la transformación del embrión plano en uno tubular, con el saco vitelino
externo unido al cuerpo por medio de un cordón umbi lical.
12. Desarrollo de Los Anexos Fetales. Los llamados anexos o membranas
fetales son un conjunto de membranas asociadas a la estructura de la placenta,
originadas a partir de las porciones de las tres hojas blastodérmicas que nunca
quedaron dentro del embrión cuando este se cerró. Estas membranas, en el
orden en que se forman, son: corion (membrana coriónica), amnios (saco
amniótico), alantoides (vesícula alantoidea) y saco vitelino.
Mientras el epiblasto embrionario está sufriendo movimientos celulares similares a
aquéllos vistos en la Gastrulación del reptil o del ave, las células extraembrionarias
están formando los tejidos propios del mamífero que permiten al feto sobrevivir
dentro del útero. Aunque las células iniciales del trofoblasto de mamíferos se
dividen como la mayoría de las otras células del cuerpo, ellas dan lugar a una
población de células en las que la división nuclear ocurre en ausencia de
citocinesis. El tipo original de células trofoblásticas lo constituye una capa llamada
citotrofoblasto . El citotrofoblasto adhiere inicialmente al endometrio a través de

una serie de moléculas de adherencia. Es más, en humanos y roedores, estas
células también contienen enzimas proteolíticas que les permiten entrar en la
pared uterina y remodelar los vasos sanguíneos uterinos para que la sangre
maternal bañe los vasos sanguíneos fetales. Se piensa que el tejido del
sincitiotrofoblasto lleva más allá la progresión del embrión en la pared uterina
digiriendo el tejido uterino. El útero, a su vez, envía los vasos sanguíneos a esta
área, dónde ellos contactan con el sincitiotrofoblasto en el futuro. Poco después
de esto, el tejido mesodérmico se extiende afuera del embrión durante la
gastrulación.
Los estudios en embriones de
humanos y monos rhesus
han sugerido que el saco
vitelino y el hipoblasto son la
fuente de este mesodermo
extraembrionario. El
mesodermo extraembrionario
une las extensiones del
trofoblasto, dando lugar a los
vasos sanguíneos que llevan
los nutrientes de la madre al
embrión. El estrecho tallo de
mesodermo extraembrionario
que en el futuro une al
embrión con el trofoblasto
forma los vasos del cordón
umbilical. El órgano
extraembrionario formado
por tejido del trofoblasto y
el mesodermo rico en vasos
sanguíneos , se llama corion , y se funde con la pared uterina para crear la

placenta. Así, la placenta tiene tanto una porción materna (el endometrio que se
modifica durante el embarazo) y un componente fetal (el corion). Existen
diferentes tipos de corion que difieren según que tan estrechamente relacionados
estén con los tejidos maternos, pudiendo, o separarse con facilidad de ellos (como
en la placenta de contacto del cerdo), o pueden estar tan íntimamente integrados
con tejidos maternales que los dos no pueden separarse sin daño a la madre y el
feto en vías de desarrollo (como en la placenta caduca de la mayoría de los
mamíferos, incluso los humanos).
El saco amniótico se origina a partir de las células externas del nódulo
embrionario, que se quedan relacionadas al trofoblasto cuando el nódulo se
transforma en disco embrionario y se crea la cavidad amniótica. La función de esta
cavidad es llenarse de líquido (llamado líquido amniótico) el cual garantiza que el
embrión posea espacio suficiente para desarrollarse libremente y al mismo tiempo
proporciona un medio de protección hidráulica contra traumas físicos.
La vesícula alantoidea se
forma como una evaginación
temprana del piso de la zona
caudal del intestino primitivo y
sirve como una ruta de desvío
o de depósito de la orina. En
los mamíferos domésticos llega
a alcanzar un tamaño mayor al
de la cavidad amniótica. En los
humanos, se desarrolla muy
poco y queda incluido dentro
del cordón umbilical. El
alantoides aparece antes de

completarse el cierre del celoma, por lo que es una estructura bien diferenciada
antes de que la separación entre el intestino primitivo y el saco vitelino sea
completa.
Bibliografía recomendada
Carlson, b. M. (2000). Embriología Humana y Biología del Desarrollo . 2ª
Edición por Harcourt S. A., Madrid.
Hickman, F. M. y Hickman, C. P. (1991) ZOOLOGÍA, Manual de laboratorio . 7ª
Edición por Interamericana Mc.Graw-Hill, Nueva York.
Noden, C. y De Lahunta, A. (1990) Embriología de los Animales Domésticos .
Editorial Acribia, Barcelona
Sadler, T. W.(2002) Embriología Médica de Langman. 8ª Edición por Editorial
Médica Panamericana, Buenos Aires