
Detección molecular de Helicobacter pylori como ...
74
Detección molecular de Helicobacter pylori como prospectiva para su identificación a partir de cuerpos de agua MARÍA DEL MAR ÁLVAREZ QUINTERO VALERIA CEBALLOS TORO Trabajo de investigación como requisito para optar al título de Microbiólogo ASESOR Dra. Adalucy Álvarez Aldana CO-ASESOR Dr. Fernando Rafael Siller López Universidad Libre Seccional Pereira Facultad de Ciencias de la Salud Programa de Microbiología 2018
Transcript of Detección molecular de Helicobacter pylori como ...

Detección molecular de Helicobacter pylori como prospectiva para su
identificación a partir de cuerpos de agua
MARÍA DEL MAR ÁLVAREZ QUINTERO
VALERIA CEBALLOS TORO
Trabajo de investigación como requisito para optar al título de Microbiólogo
ASESOR
Dra. Adalucy Álvarez Aldana
CO-ASESOR
Dr. Fernando Rafael Siller López
Universidad Libre Seccional Pereira
Facultad de Ciencias de la Salud
Programa de Microbiología
2018

TABLA DE CONTENIDO
Página
1. PLANTEAMIENTO DEL PROBLEMA Y PREGUNTA DE
INVESTIGACIÓN
6
2. JUSTIFICACIÓN 7
3. MARCO TEÓRICO
3.1. Reseña histórica 8
3.2. Microbiología del microorganismo
3.2.1. Taxonomía 8
3.2.2. Morfología 9
3.2.3. Genoma 9
3.2.4. Características bioquímicas 10
3.2.5. Factores de virulencia 11
3.2.5.1. Flagelina 11
3.2.5.2. Ureasa 11
3.2.5.3. Adhesinas 12
3.2.5.4. VacA y CagA 12
3.3. Epidemiología 13
3.4. Enfermedades asociadas a la infección
3.4.1. Dispepsia 13
3.4.2. Gastritis 14
3.4.3. Úlcera péptica 14
3.4.4. Cáncer gástrico 14
3.4.5. Linfoma gástrico tipo MALT 15
3.5. Ruta de transmisión
3.5.1. Ruta iatrogénica 15
3.5.2. Ruta fecal-oral 15
3.5.3. Ruta gastro-oral 16
3.5.4. Ruta oral-oral 16
3.6. Mecanismos de supervivencia en el agua
3.6.1. Forma viable no cultivable (VBNC) 16
3.6.2. Formación de biopelículas 17
3.6.3. Asociación con amebas de vida libre (FLA) 18
3.7. Dosis infectiva 19
3.8. Métodos de detección 19
3.8.1. Cultivo 20
3.8.2. Histología 20
3.8.3. Test rápido de ureasa 20
3.8.4. Detección de antígeno en heces 20
3.8.5. Test de aliento con urea 20
3.8.6. Serología 21
3.8.7. Métodos de reacción en cadena de la polimerasa 22
3.8.7.1. PCR de punto final 22
3.8.7.2. PCR en tiempo real 24

4. OBJETIVOS
4.1. Objetivo general 25
4.2. Objetivos específicos 25
5. METODOLOGÍA
5.1. Elaboración de un manuscrito de revisión de tema sobre el papel del
agua en la ruta de transmisión de H. pylori
26
5.2. Análisis de los cebadores usados para la detección molecular de H. pylori 26
5.3. Detección del gen ureA de H. pylori por PCR en tiempo real 27
5.4.Detección del gen ureA de H. pylori por PCR de punto final 27
5.5. Preparación de diluciones a partir de ADN de cepas de referencia 28
5.6. Determinación de la concentración de las diluciones 28
5.7. Límite de detección de ADN de H. pylori 29
6. RESULTADOS Y DISCUSIÓN
6.1. Manuscrito de revisión sobre el papel del agua en la ruta de
transmisión de H. pylori
30
6.2. Análisis de los cebadores usados para la detección molecular de H. pylori 31
6.3. Comparación de dos metodologías moleculares para la detección de H.
pylori
32
6.4. Diluciones obtenidas a partir de ADN de cepas de referencia
36
6.5. Determinación de la concentración de las diluciones 36
6.6. Límite de detección de ADN de H. pylori 37
7. CONCLUSIONES 42
8. PERSPECTIVAS A FUTURO 43
9. AGRADECIMIENTOS 44
10. REFERENCIAS BIBLIOGRÁFICAS 45

ÍNDICE DE TABLAS
Página
Tabla 1. Métodos de detección de H. pylori 19
Tabla 2. Cebadores utilizados para la detección molecular de H. pylori mediante
PCR.
26
Tabla 3. Componentes de la mezcla de reacción para la PCR en tiempo real 27
Tabla 4. Componentes de la mezcla de reacción para la PCR de punto final 27
Tabla 5. Resumen de algunos artículos relacionados con el papel del agua en la
transmisión de H. pylori.
30
Tabla 6. Resultado cuantificación de la PCR en tiempo real para detectar el
número límite de copias de ADN por μl de H. pylori.
34
Tabla 7. Concentración esperada de las diluciones realizadas a partir del ADN de
la cepa de referencia 12908 frente a las concentraciones obtenidas mediante
fluorómetro.
36
Tabla 8. Resultado cuantificación de la PCR en tiempo real para detectar el
número límite de nanogramos de ADN por μl de H. pylori.
39

ÍNDICE DE FIGURAS
Página
Figura 1. Patrón de crecimiento exponencial de las copias de ADN mediante PCR 23
Figura 2. Fragmento de la secuencia del gen ureA de H. pylori con la secuencia
de los cebadores ureA-F/ureA-R
31
Figura 3. Características de los cebadores ureA-F/ureA-R específicos para el gen
ureA de H. pylori.
32
Figura 4. Electroforesis en gel de agarosa al 3% y el producto amplificado por
PCR de punto final.
33
Figura 5. Resultado PCR en tiempo real para detección de cepa de referencia
12908 y el límite de detección de copias de ADN por μl de H. pylori.
33
Figura 6. Electroforesis en gel de agarosa al 3% y el producto amplificado por
PCR en tiempo real.
35
Figura 7. Resultado PCR en tiempo real para detectar el límite de nanogramos de
ADN por μl de H. pylori.
39
Figura 8. Electroforesis en gel de agarosa al 3% y los productos amplificado por
PCR de punto final y PCR en tiempo real.
40

1. PLANTEAMIENTO DEL PROBLEMA Y PREGUNTA DE INVESTIGACIÓN
La infección por Helicobacter pylori puede generar ciertas enfermedades, entre ellas gastritis
crónica, enfermedad ulcero-péptica y puede ser asociado a carcinoma gástrico y a linfoma del
tejido linfoide asociado a mucosas (MALT, del inglés mucus-associated lymphoid tissue
lymphoma). Es importante mencionar que la infección por este microorganismo presenta una alta
tasa de morbilidad, pero una baja tasa de mortalidad y es curable con terapia antimicrobiana.
Diversos estudios han intentado identificar otros reservorios aparte de la mucosa gástrica humana
y otra posibles rutas de transmisión ya que las más comunes son oral-oral, gastro-oral y fecal oral.
Por esta razón se ha estudiado el agua como posible vehículo para la diseminación de la infección
dado que, en los últimos años, se ha detectado la presencia de H. pylori en distintos tipos de aguas
como en pozos, ríos, biofilms asociados a tuberías y a redes de distribución. Esta posible ruta
aumenta la probabilidad de contraer la infección por parte de la comunidad, por lo que se ha
aumentado la realización de estudios epidemiológicos que pretenden determinar si la prevalencia
de H. pylori se asocia con el tipo de fuente de agua, apoyándose fuertemente la idea de que esta
puede actuar como ruta de transmisión tal como se mencionó anteriormente.
En los últimos años se ha evidenciado un alto interés por parte de la comunidad científica en la
identificación y análisis de H. pylori a través de muestras distintas a su hábitat gástrico normal, sin
embargo la elevada sensibilidad de H. pylori a las condiciones ambientales y de cultivo hacen
difícil su recuperación a partir de muestras no gástricas, como es el caso del agua. Todo esto genera
una gran incertidumbre en científicos e investigadores ya que el limitado conocimiento sobre el
agua como ruta de trasmisión de la bacteria y su dosis infectiva en esa matriz hacen que no pueda
haber una difusión certera de la información respecto a este tema, lo cual resulta determinante en
el consiguiente desarrollo de la enfermedad, ya que si estos hallazgos no son transmitidos a la
comunidad no se adoptarán medidas de precaución al respecto. Siendo así, es necesario la
realización de más estudios al respecto.
En el marco del desarrollo del proyecto “Estandarización de un método molecular para la detección
de Helicobacter pylori a partir de agua” financiado en la convocatoria 08, se planteó la
investigación denominada “Detección molecular de Helicobacter pylori como prospectiva para ser
utilizada en cuerpos de agua”, para contestar las siguientes preguntas de investigación: ¿Cuál es el
límite de detección de ADN de H. pylori mediante un método molecular?, ¿Existen diferencias en
el límite de detección según el tipo de método molecular (PCR en tiempo real y PCR de punto
final)?

2. JUSTIFICACIÓN
Gran parte de las investigaciones en H. pylori se realizan a partir de muestras directas de pacientes
humanos. No obstante, en los últimos años se han realizado trabajos que muestran hallazgos de la
bacteria en fuentes no gástricas, sin embargo la información respecto a estas investigaciones es
muy limitada.
La alta tasa de infección por H. pylori en las naciones en desarrollo, está estrechamente relacionado
con factores socioeconómicos, como la pobreza, malas condiciones sanitarias y el hacinamiento
en hogares lo que indica que la transmisión de la infección se puede reducir mediante la mejora
de las condiciones higiénico sanitarias de la población y el uso regular de agua potable.
La vía de transmisión de H. pylori no es bien conocida, y al parecer el agua juega un papel
importante. Se ha observado la presencia de H. pylori en aguas subterráneas, depósitos de
almacenamiento de agua potable, muestras de aguas residuales y en agua contaminada que también
puede ser un foco de contaminación de este microorganismo, debido a la contaminación fecal de
lugares en donde el agua no es tratada y los individuos la consumen así. Estas poblaciones
presentan una tasa mucho más alta de infección por este patógeno proporcionando sospechas de
que el agua si tiene un papel importante transmisión de la infección. Se dice que H. pylori puede
ser inactivada por cloro, pero otros estudios demuestran que este patógeno puede sobrevivir en
bajas concentraciones de cloro en su estado viable no cultivable y puede formar biofilms para
volverse más resistentes al tolerar el efecto biocida del cloro en sistemas de distribución de agua.
Esta situación se agrava aún más sabiendo que H. pylori suspendido en agua tiene muy bajo poder
de cultivo comparado con otros patógenos de origen acuático. Motivo por el cual el uso de técnicas
moleculares permite detectar la presencia de ADN de H. pylori sin importar su viabilidad, siendo
entonces el método de detección más adecuado. Este estudio es de gran importancia a nivel de
salud pública y ecología de H. pylori en cuanto su distribución en el ambiente, además, aporta en
el conocimiento, el estudio y entendimiento sobre la detección, transmisión e infección con H.
pylori.

3. MARCO TEÓRICO
3.1. Reseña histórica
Los primeros aportes sobre microorganismos en la mucosa gástrica fueron hechos a finales del
siglo XIX y las primeras observaciones relacionadas con Helicobacter pylori fueron hechas por
Bizzozero en 1893, quien comprobó la presencia de bacterias Gram-negativas en las células
parietales y glándulas gástricas de perros (1); a principios del siglo XX anatomistas y patólogos
reportaron la presencia de microorganismos en forma espiral en el estómago de animales, y poco
después, bacterias similares fueron observadas en el estómago de humanos con enfermedad ulcero
péptica y cáncer gástrico (2). En 1979 el patólogo Robin Warren observó por primera vez la
bacteria en una muestra de mucosa gástrica con gastritis crónica activa. En el año 1981, el
gastroenterólogo Barry Marshall colaboró con los microbiólogos para encontrar una técnica de
cultivo adecuado que favoreciera el crecimiento de la bacteria (3). Warren y Marshall aislaron por
primera vez, la bacteria desde la superficie del epitelio gástrico en el año 1983. Un año después,
Marshall demostró que esta bacteria podía colonizar el estómago humano e inducir inflamación de
la mucosa gástrica al ingerir un cultivo bacteriano que le produjo gastritis, la cual curó con sales
de bismuto y metronidazol (4).
Esto dio pie para debatir respecto a que los factores como el estrés y el estilo de vida eran las
principales causas de la enfermedad ulcero-péptica y se estableció que la bacteria causaba más del
90% de las úlceras duodenales y el 80% de las úlceras gástricas. Se continuaron realizando estudios
al respecto, algunos de los cuales tenían el objetivo de promover información sobre este
microorganismo a nivel del continente europeo, sus técnicas de diagnóstico y métodos de
investigación clínica y básica. Finalmente, Warren y Marshall fueron galardonados con el Premio
Nobel de Medicina y Fisiología del 2005 por el “Descubrimiento de la bacteria Helicobacter pylori
y su papel en la gastritis y enfermedad ulcero péptica” (3), revolucionando la gastroenterología.
3.2. Microbiología del microorganismo
3.2.1. Taxonomía
Inicialmente H. pylori fue llamada Campylobacter pyloridis (5), pero tras un análisis de las
secuencias de ADNr 16S se comprobó su divergencia con otras cepas de Campylobacter y se
confirmó el hallazgo de un nuevo género “Helicobacter”, para el año de 1989 se renombró como
Helicobacter pylori (6).
H. pylori pertenece al dominio Bacteria, filo Proteobacteria, clase Epsilonproteobacteria en la
subdivisión Thiobacteria, familia Helicobacteraceae y género Helicobacter. Esta familia también
incluye los géneros: Wolinella, Flexispira, Sulfurimonas, Thiomicrospira y Thiovuluml (7).

Las especies del género Helicobacter pueden ser subdivididas, en gástricas y entero-hepáticas.
Ambos grupos demuestran un alto nivel de especificidad de órgano. Las especies gástricas son
incapaces de colonizar el tracto gastrointestinal inferior o el hígado y las entero-hepáticas la
superficie de la mucosa gástrica. H. pylori hace parte de las especies gástricas, que se han adaptado
a condiciones inhóspitas como la encontrada en la superficie de la mucosa gástrica (8).
3.2.2. Morfología
H. pylori es un bacilo Gram negativo, con morfología espiral cuando se encuentra en la mucosa
gástrica dado que dicha forma facilita su movilidad en la viscosidad del moco gástrico; es menos
espiral cuando crece en medios artificiales y puede pasar de la forma bacilar a la forma cocoide en
cultivos prolongados o sometidos a situaciones no favorables para su crecimiento como exposición
a O2, antibióticos y cuando se encuentra en el agua (9), se ha asumido que estas formas son
indicativas de un estado latente y ayudan a la supervivencia del microorganismo en el ambiente.
La estructura cocoide es considerada la forma persistente de H. pylori y es denominada VBNC
(estado viable pero no cultivable) (10). La forma vegetativa oscila entre 0,5 a 1,0 µm de ancho y
de 3 µm de largo (11), no forma endosporas y posee de 2 a 6 flagelos unipolares de
aproximadamente 3 µm de longitud (12).
Varios estudios han señalado que los cambios en la morfología de H. pylori están estrechamente
relacionada con los genes implicados en la síntesis de peptidoglucano. Iniciando por la división de
los enlaces peptídicos por acción de Csd1, Csd2 y Csd3.S eguido de esto, los enlaces intrapéptidos
también se escinden por acción secuencial de Csd3, de una DL-carboxipeptidasa desconocida y
Csd4, que permite una relajación inicial del peptidoglicano en la zona de torsión, perdiendo la
forma helicoidal. A continuación, los monómeros que han sido sometidos a la acción enzimática,
son reconocidos por diferentes transglicosilasas líticas, como Slt y MltD, que se eliminan para la
posterior inserción de nuevos monómeros. Finalmente, la inserción de nuevos monómeros en el
peptidoglicano y la elongación, se realiza mediante PBP1, PBP2 y proteínas de membrana (MreC).
Esto aumenta la longitud de las cadenas de glucanos y los enlaces cruzados (13).
3.2.3. Genoma
La primera secuencia del genoma completo de H. pylori se publicó en 1997, esta secuencia
correspondió a la cepa 26695 aislada de un paciente de Reino Unido que presentaba gastritis. El
genoma de la cepa 26695 está incluido en un cromosoma circular de 1’667.867 pb el cual contiene
1.587 genes (14, 15). Las propiedades del genoma son constantes, con un tamaño de genoma
aproximadamente de 1,6 kbp, un contenido promedio de GC del 39% y aproximadamente 1.500–
1.600 secuencias de codificación predichas por cepa. Los datos de los análisis de transcriptoma,
metiloma y proteoma han proporcionado información importante sobre la regulación de los genes
y las propiedades de virulencia de H. pylori. El genoma de H. pylori es altamente metilado y exhibe

una variación específica de la cepa. El análisis del transcriptoma primario reveló una complejidad
inesperada en la regulación génica de H. pylori. Muchos sitios de inicio de la transcripción se
ubicaron dentro de los operones y opuestos a los genes anotados que significan desacoplamiento
de policistrones y transcripción antisentido. H. pylori codifica más de 60 ARN pequeños, incluido
el ARN 6S que antes se consideraba ausente. Los ARN pequeños son mediadores importantes de
la regulación postranscripcional bajo estrés y durante la virulencia (16).
La diversidad genética de H. pylori y la presencia de genes específicos en cada cepa ha hecho
difícil identificar el orden de los genes en el cromosoma; algunos estudios han demostrado que la
diversidad nucleotídica de genes ortólogos entre aislamientos independientes de H. pylori es muy
alta y que es muy raro encontrar dos cepas con la misma secuencia de genes, se ha demostrado que
la recombinación entre alelos de genes dentro de la población de la bacteria es extremadamente
común; sin embargo debido a la redundancia en el código genético, es más alta la divergencia en
los genes que en las respectivas proteínas codificadas (7).
Otros estudios basados en el análisis de microarreglos ADN, reportaron la existencia de genomas
centrales con 1.091 o 1.281 genes (17, 18), se ha descrito que este núcleo de genes es
indispensables para la colonización de la mucosa gástrica del huésped y que 33 de ellos son
indispensable para la viabilidad de la bacteria; una de las mayores diferencias entre el contenido
de genes de la cepas de H. pylori es la presencia o ausencia de una región de 40 Kb de ADN
cromosomal conocido como isla de patogenicidad (Cag -PAI) (19).
3.2.4. Características bioquímicas
Las especies de Helicobacter son quimioorganótrofas y tienen un metabolismo respiratorio. La vía
de la glucolisis-gluconeogénesis probablemente compromete los principales medios de producción
de energía, como también los puntos de inicio de muchas vías de biosíntesis. Presentan
parcialmente las vías metabólicas Entner-Doudoroff, de pentosas fosfato, y el ciclo de ácidos
tricarboxílicos, sin embargo la vía del glioxilato está ausente. Esta bacteria oxida glucosa (20), no
hidroliza gelatina, almidón, caseína o tirosina. Produce gran cantidad de enzimas como oxidasa,
ureasa y catalasa las cuales son muy útiles para su identificación microbiológica (7); fosfolipasas
y alcohol deshidrogenasas le permiten modificar las barreras de la mucosa y acceder a través del
epitelio gástrico (21), fosfatasas y ATPasas esenciales para la generación de energía, síntesis y
transporte de iones y productos celulares (22).
3.2.5. Factores de virulencia
H. pylori genera una fuerte respuesta inmune, humoral y celular en toda la mucosa gástrica,
principalmente actúan los neutrófilos y luego se amplía la respuesta inflamatoria por la interacción
de linfocitos, células mastoides, macrófagos y células no inmunes las cuales liberan gran

cantidad de mediadores químicos. Frente a esto, H. pylori posee factores de virulencia que la
ayudan a continuar la colonización del epitelio gástrico superficial, la profundidad de las criptas y
el espacio entre las células epiteliales (9).
3.2.5.1. Flagelina
La movilidad para H. pylori es fundamental para colonizar la mucosa gástrica, posee entre dos y
seis flagelos monopolares, característica inusual que es distinta del resto de proteínas flagelares,
las cuales son homo poliméricas. Cada flagelo está compuesto por dos flagelinas (FlaA y FlaB).
FlaB se localiza en la base del flagelo, mientras que FlaA, se encuentra en el exterior. Además, la
morfología espiral o helicoidal de la bacteria facilita su movilidad en la viscosidad del moco
gástrico, y produce una proteasa que digiere el moco facilitando su avance (9).
3.2.5.2. Ureasa
El jugo gástrico normal posee un pH inferior a 4 el cual le confiere un carácter bactericida y por
tanto capaz de eliminar a muchas de las bacterias que llegan al estómago con la ingesta de
alimentos; conociéndose al estómago como la “barrera ácida”. Siendo así, la colonización de la
mucosa gástrica por H. pylori, implica su capacidad para sobrevivir en ese ambiente ácido. La
clave para la adaptación al pH gástrico reside en la producción de ureasa. Esta enzima “urea
amidohidrolasa” cataliza la hidrólisis de urea para proporcionar amonio y carbamato, el cual se
descompone para producir otra molécula de amonio y ácido carbónico, con lo cual neutraliza el
ambiente de las células gástricas colonizadas. La ureasa representa alrededor de un 5% del total
de las proteínas celulares de H. pylori, se localiza en el citosol y en su superficie, aunque esta
última puede tratarse de un mecanismo no específico para la exportación de la enzima (23).
La ureasa es producida por numerosas especies de bacterias. Además se ha demostrado como un
potente factor de virulencia para algunas especies, como Proteus mirabilis, Staphylococcus
saprophyticus y Helicobacter pylori. La ureasa es fundamental para la colonizaicon de mucosa
gástrica por parte de H. pylori además es un potente inmunógeno que provoca una respuesta
inmune vigorosa. Esta enzima se utiliza para la identificación taxonómica y para el diagnóstico y
seguimiento después del tratamiento. (24). El amoníaco producido por la ureólisis aumenta el pH
gástrico, proporcionando así un ambiente permisivo para la colonización del estómago, la nueva
estructura dodecamérica de la enzima le permite que permanezca activa en la superficie celular a
un pH gástrico ácido. Dentro de esta estructura los 12 sitios activos se agrupan dentro del interior
de la cubierta proteica. Esto puede permitir una alta concentración local de amoníaco dentro de la
enzima que puede proteger a los grupos quelantes de níquel de la protonación. Esta proteína
participa en el paso 1 de la ruta secundaria que sintetiza CO2 y NH a partir de la urea. Las proteínas
que se sabe que están involucradas en esta ruta son: ureA (subunidad alfa), ureB (subunidad beta)
(25).

3.2.5.3. Adhesinas
La colonización de la mucosa lleva implícito como paso previo la capacidad de la bacteria para
adherirse al epitelio gástrico, lo cual es esencial para la inducción de gastritis. Esta adherencia
ocurre mediante la interacción entre las adhesinas de la bacteria y los receptores del hospedero que
están representados por algunas proteínas de la matriz extracelular. Generando lesiones que se
caracterizan por la pérdida de las microvellosidades del epitelio gástrico en el sitio de adhesión, lo
que conduce a la formación de una estructura similar a un pedestal de unos 5 nm de diámetro, con
uniones estrechas entre la bacteria y la superficie celular. La adhesión a la mucosa gástrica es
necesaria para la inducción y liberación de citocinas proinflamatorias (23).
3.2.5.4. VacA y CagA
VacA (Vacuolization Associated Gen) fue nombrado asi por su capacidad para inducir la
formación de vacuolas en células eucariotas. La diferencia en las capacidades de vacuolización
está determinada por las variaciones en las tres regiones del gen vacA: la señal (s1 y s2), las
regiones intermedias (i1 e i2) y las regiones medias (m1 y m2). Una combinación de diferentes
secuencias en las 3 regiones conduce a múltiples alelos y determina la actividad de vacuolización.
La actividad de vacuolización es alta en genotipos s1/m1, intermedia en genotipos s1/m2 y ausente
en genotipos s2/m2.
CagA (Cytotoxin Associated Gen) es una oncoproteína que se inyecta en las células huésped a
través de una estructura de pilus llamada sistema de secreción tipo IV (T4SS). El locus del gen
que codifica CagA y T4SS se denomina isla de patogenicidad Cag (Cag PAI). Después de ser
inyectado en las células huésped, este gen altera las vías de transducción de señales intracelulares
que facilitan la transformación maligna de las células epiteliales gástricas. Existe evidencia directa
de que CagA es una proteína que necesita de fosforilación para conferir la oncogénesis. Además,
CagA y T4SS aumentan la inflamación gástrica a través de la señalización NFκB y el aumento de
la secreción de IL-8 que predisponen a la inestabilidad genética y carcinogénesis. CagA también
puede causar cambios epigenéticos, como la hipermetilación de histonas, que conducen a la
regulación negativa de los genes supresores de tumores o microRNAs (26).
3.3. Epidemiología
H. pylori afecta aproximadamente el 50% de la población mundial (27, 28), es adquirida en la
niñez y persiste durante mucho tiempo si no es tratada con terapia antimicrobiana (29), las
condiciones por las que se presenta la infección no están establecidas, se cree que esta está
influenciada por las variaciones geográficas, factores genéticos, edad, sexo, condiciones socio-
económicas, bajo nivel sociocultural, precarias condiciones higiénico –sanitarias, entre otras (30).
En el mundo se observa mayor prevalencia en los países en desarrollo que en los desarrollados

(80-90% vs 30-50%) (31, 4), los que se encuentran en desarrollo se caracterizan por altas tasas de
infección durante la niñez a diferencia de países desarrollados, donde la tasa de infección infantil
es baja con tendencia de aumento en función de la edad (32).
La prevalencia de la infección puede variar entre países de un mismo continente, incluso entre
regiones de un mismo país y se ha relacionado con el nivel de virulencia de la cepas y factores v
asociados s al huésped, es por esto, que la infección puede iniciar desde edades muy tempranas sin
presentar sintomatología aparente y puede pasar toda la vida sin tratamiento (33) o generar
rápidamente enfermedades asociadas. Por ejemplo, hay reportes en la literatura de mayor
prevalencia de las cepas citotóxicas (cagA+, cagE+ y vacA s1/m1,) en pacientes con cáncer,
metaplasia y ulcera en comparación con la observada en pacientes con gastritis no atrófica (34), lo
que ha caracterizado los vacA s1/m1 y cagA como marcadores de virulencia (35, 36).
3.4. Enfermedades asociadas a la infección
La infección por H. pylori es muy variable ya que puede estar influenciado por factores
dependientes tanto del microorganismo como del huésped, dichos factores determinan la evolución
de la infección hacia un estado crónico asintomático o el desarrollo de manifestaciones clínicas
(37).
La colonización por H. pylori no supone una enfermedad en sí misma, pero si ha sido asociado a
enfermedades como dispepsia, gastritis crónica, ulcera péptica, linfoma gástrico tipo MALT, y
cáncer gástrico. Los síntomas más comunes son alteraciones dispépticas transitorias (náuseas,
vómitos o sensación de plenitud), inflamación de la mucosa e hipoclorhidria. La inflación de la
mucosa y la gastritis se correlaciona con la excreción de ácido durante la colonización; cuando se
presenta hipoclorhidria se puede observar pangastritis con inflamación crónica, atrofia de la
mucosa, metaplasia intestinal y acaba desencadenándose un cáncer gástrico (2).
3.4.1. Dispepsia
La dispepsia se ha definido como dolor abdominal persistente o molestias centradas en la parte
superior del abdomen en pacientes que no tienen una clara explicación de los síntomas, la
patogenia es desconocida, pero se le han atribuido alteraciones de la percepción visceral, secreción
de ácido gástrico, y la motilidad, además puede indicar la presencia de enfermedades graves, como
úlcera péptica o cáncer gástrico (38). Esta enfermedad se ha considerado un enorme problema de
salud pública por la prevalencia y el elevado consumo de recursos que provoca (39). En la
actualidad se ha descrito que el 1% de pacientes infectados tienen riesgo de contraer la enfermedad
(40).

3.4.2. Gastritis
La gastritis se presenta como un dolor epigástrico, náuseas y vómito (9), se caracteriza por una
inflamación crónica activa de la mucosa gástrica guiada por monocitos, linfocitos y neutrófilos;
su severidad está determinada por la densidad de las células inflamatorias que infiltran la lámina
de la mucosa y sub-mucosa. Una de las características de esta enfermedad en la edad pediátrica
es la mayor presencia de linfocitos y células plasmáticas y una afectación más leve que la que tiene
lugar en el adulto (41). H. pylori puede causar gastritis antral predominante (asociada con una
mayor producción de ácido estimulada por la comida), lo que predispone a la úlcera duodenal o
pangastritis (asociado a una baja producción de ácido gástrico), lo que predispone a la ulceración
gástrica y adenocarcinoma gástrico (42). Tras la evolución de la gastritis crónica se puede generar
gastritis crónica atrófica; la mucosa gástrica se deteriora presentándose alteración de las células
epiteliales, reducción en la secreción de pepsinógeno I (PGI) y II (PGII) y una mala absorción de
los metabolitos esenciales como la vitamina B12 y finalmente un desencadenamiento en
metaplasia intestinal (43).
La gastritis crónica atrófica se ha relacionado con la presencia de H. pylori, porque las células
foveolares poseen receptores para esta bacteria. Se ha descrito que entre el 1- 10% de paciente
infectados tienen riesgo de desarrollar gastritis crónica con úlceras gastroduodenales (40).
3.4.3. Ulcera péptica
La ulcera péptica es una brecha en la mucosa gástrica o duodenal que va hasta la submucosa, las
infracciones poco profundas son llamadas erosiones y aunque a veces son insignificantes pueden
ser precursores de ulceras, las dos principales causas de esta enfermedad son la infección por H.
pylori y los medicamentos anti-inflamatorios no esteroideos. La relación de la enfermedad con H.
pylori se explica a través de algunos genes de virulencia como vacA que esta presenten en casi
todas las cepas de la bacteria, sin embargo, la actividad de la esta citotoxina es positiva en un 40-
60% de los pacientes infectados con esta bacteria y solo alrededor del 15 % de los individuos
infectados desarrollan la enfermedad (42).
3.4.4. Cáncer gástrico
El cáncer gástrico se ha descrito como un proceso de varios pasos el cual empieza con gastritis
crónica a través de la atrofia glandular, metaplasia y displasia, atrofia gástrica y metaplasia
intestinal. H. pylori ha sido clasificado por la organización mundial de la salud como carcinogénico
tipo 1 por su asociación con cáncer gástrico. Se cree que las interacciones de la bacteria con la
célula madre gástrica inducen a daños moleculares en la célula (44). Este microorganismo posee
una relación asociativa con el adenocarcinoma gástrico no cardial y el 0,1-3% de los pacientes se
encuentran en riesgo. La enfermedad también ha sido asociado con la edad, esta hipótesis se

explica por la acumulación de lesiones en la células madres, evadiendo el control homeostático y
dando como resultando cáncer (40).
3.4.5. Linfoma gástrico tipo MALT
El tracto gastrointestinal es el sitio más frecuente de los linfomas primarios extra nodales no
Hodgkin y en el estómago está localizado más de 75% de este tipo de tumores (45). Los linfomas
gástricos tipo MALT son tumores derivados de la zona marginal de los folículos linfoides, su
asociación con H. pylori es tan grade que la erradicación de la bacteria se convirtió en un estándar
para el manejo de estos tumores (46). La erradicación de la infección causa la regresión de la
mayoría de los linfomas MALT gástricos localizados (45). El riesgo de contraer la enfermedad es
2,8 veces mayor en personas infectadas con H. pylori ; Se dice que entre el 72 y 98% de los
pacientes con Linfoma gástrico tipo MALT están infectados con la bacteria; sin embargo, la
erradicación de la bacteria solo cura la enfermedad en un 70 a 80% de los casos (4).
3.5. Rutas de transmisión
Las rutas de transmisión de H. pylori aún son poco conocidas pero las más aceptadas son las vías
iatrogénica, fecal-oral, gastro-oral y oral-oral; aunque también se ha descrito que la bacteria puede
ser transmitida a través de algunos alimentos, animales y agua (48).
3.5.1. Ruta iatrogénica
La transmisión iatrogénica constituye una ruta por la cual personas pueden ser infectadas por
medio de elementos que han estado en contacto con heces, secreciones orales y jugos gástricos
contaminados por H. pylori (49). Esta ruta se ve protagonizada principalmente por material médico
utilizado en la detección del patógeno, como es el caso de los endoscopios, los cuales son puestos
en contacto con la mucosa gástrica de una persona infectada para luego ser utilizado con otra (50).
3.5.2. Ruta fecal- oral
Esta ruta de transmisión hace referencia a las enfermedades adquiridas cuando las partículas
fecales de un hospedero se introducen en la boca de un huésped, las principales causan son : agua
contaminada con heces y no tratada antes de consumir, alcantarillado deficiente, los alimentos
manipulados con heces, o cualquier objeto que haya estado en contacto con estas. La evidencia
acerca de la transmisión del patógeno por esta ruta se encuentra basada en estudio realizados donde
se ha podido cultivar H. pylori de heces humanas. Esta ha sido aislada a partir de heces de niños,
en adultos su detección es menos frecuente (49).

3.5.3. Ruta gastro-oral
Esta ruta hace referencia a la incidencia de H. pylori en los jugos gástricos, los cuales por medio
del vomito tienen contacto con otras personas las cuales pueden infectarse. Existen altas tasas de
infección en niños, entre los cuales los vómitos y el reflujo gastro-esofágico son comunes y más
frecuentes que en los adultos, además del contacto con otros objetos que son llevados a la boca
(23). Esta es una ruta no posee mucha relevancia en comparación de las fecal- oral y oral- oral, sin
embargo, el aislamiento de la bacteria en el jugo gástrico y vomito de personas infectadas, hace
que la ruta gastro- oral sea una fuente posible de transmisión de H. pylori (51).
3.5.4. Ruta oral-oral
Muchos científicos tienen la hipótesis de que la ruta oral-oral de transmisión de H. pylori es la más
probable, sobre todo en los países desarrollados. Esta se da a través del contacto con saliva
infectada por el patógeno, se encuentra presencia de H. pylori en lesiones de la cavidad oral y en
la placa dental. Esta transmisión se logra a través de contacto íntimo, al compartir alimentos y
utensilios contaminados y contacto con la placa dental, la cual es un importante reservorio,
perpetuando el ciclo infeccioso de la bacteria (52).
Hace algunos años pocas investigaciones implicaban el agua como vehículo de transmisión de H.
pylori, pero hoy en día hay suficiente evidencia de la presencia de esta bacteria en el agua a nivel
mundial, por lo que se puede implicar al agua tratada y no tratada como posible vehículo de
transmisión (53).
La hipótesis de que el agua es una vía de transmisión de H. pylori está respaldada por dos tipos de
estudios; los epidemiológicos que han demostrado una mayor prevalencia de la infección en países
en vía de desarrollo que padecen problemas relacionados con la distribución sanitaria del agua a
la población y los estudios que han detectado o aislado H. pylori de diversas fuentes de agua. Por
este motivo, son varios los estudios que se han centrado en determinar el potencial del agua como
fuente de transmisión de H. pylori y su capacidad de producir la infección clínica.
3.6. Mecanismos de supervivencia en el agua
3.6.1. Forma viable no cultivable
Muchas especies de bacterias entran en el estado viable no cultivable (VBNC) cuando están
expuestas a condiciones estresantes como inanición, bajas temperaturas y en general condiciones
ambientales desfavorables, siendo esta una estrategia de adaptación para la supervivencia a largo
plazo. A diferencia de las células normales que son cultivables en medios adecuados y se
convierten en colonias, las células VBNC son células a pesar de estar vivas que pierden su
capacidad de crecer en medios de cultivo en los que normalmente crecen. A pesar de esto, las

células VBNC no se consideran células muertas debido a que tienen su membrana intacta que
contiene la información genética en perfecto estado, además de ser metabólicamente activas (54).
La capacidad de ingresar al estado VBNC puede ser ventajosa para la bacteria, pero poco favorable
para la salud humana ya que bajo los métodos convencionales de cultivo no es posible su
detección, por lo que si en una muestra todas o algunas bacterias se encuentran en estado VBNC,
la cantidad total de bacterias viables será subestimada o no se detectará (55) y tratándose de un
patógeno humano, podrá infectar a la persona que consuma el agua en la cual no pudo ser
detectado.
Se ha demostrado la viabilidad y la capacidad de cultivo de H. pylori en agua hasta por 75 horas a
10°C. Así mismo, se determinó que el total de células no disminuía por períodos mucho más largos
de tiempo en condiciones adecuadas (2 años a 4°C) (56).
Varios estudios han señalado que los cambios en la morfología de H. pylori están estrechamente
relacionada a los genes implicados en la síntesis de peptidoglucano. En primer lugar, ocurre la
división de los enlaces peptídicos por acción de Csd1, Csd2 y Csd3. Luego, los enlaces
intrapéptidos también se escinden por acción secuencial de Csd3, de una DL-carboxipeptidasa
desconocida y Csd4, que permite una relajación inicial del peptidoglicano en la zona de torsión,
perdiendo la forma helicoidal. A continuación, los monómeros que han sido sometidos a la acción
enzimática, son reconocidos por diferentes transglicosilasas líticas, como Slt y MltD, que se
eliminan para la posterior inserción de nuevos monómeros. Finalmente, la inserción de nuevos
monómeros en el peptidoglicano y la elongación, se realiza mediante PBP1, PBP2 y proteínas de
membrana (MreC). Esto aumenta la longitud de las cadenas de glucanos y los enlaces cruzados
(13).
La forma espiral se ha asociado con la capacidad de moverse en un medio viscoso, como el moco
gástrico. Esto significa que la forma helicoidal puede ser innecesaria en ambientes acuáticos ya
que puede implicar un costo energético innecesario para la bacteria (13), siendo este otro posible
motivo que justifique el cambio de su morfología
3.6.2. Formación de Biopelículas
La formación de biopelículas en el ambiente ha sido propuesto como una estrategia de H. pylori
para sobrevivir en los sistemas de distribución de agua (56).
Las biopelículas son comunidades microbianas embebidas dentro de una matriz orgánica
polimérica autoproducida y adherida a superficies vivas o inertes, son sólidas y presentan canales
por donde fluye el agua y los nutrientes a las zonas más profundas. Los primeros análisis de la
producción de biofilms por parte de bacterias, como H. pylori, se llevaron, mediante estudios de
microscopía electrónica, observándose la presencia de formas espirales, cocoides y degenerativas.
La formación de la biopelícula conlleva un proceso de cinco fases: 1) Adsorción reversible de la

bacteria a la superficie; 2) Unión irreversible a la superficie; 3) Fase inicial de maduración con
crecimiento y desarrollo microbiano; 4) Producción del exopolímero y, 5) Desarrollo final de la
colonia con dispersión de células colonizadoras (57).
Cuando H. pylori se adhiere a una superficie, comienza la activación del quorum sensing así como
los mecanismos de formación de los biofilms. Se ha trabajado con biofilms en sistemas de tuberías
y tanques de almacenamiento y algunos autores creen que así es como H. pylori puede ser viable
en el agua (58) debido a que, al ubicarse en la parte interna de estos tanques y tuberías, las hace
inaccesibles y difícil de alcance por los desinfectantes (59).
En Colombia se evaluó la presencia de H. pylori en muestras de agua y biopelículas de los grifos
de instituciones educativas oficiales de la ciudad de Medellín a través de una técnica de reacción
en cadena de la polimerasa (PCR) de punto final, identificando la bacteria por cultivo en 11.2% y
PCR en 2.1% de las muestras (53). En este caso, el cultivo tuvo mejor rendimiento que la PCR
probablemente porque los componentes de las biopelículas actuaron como inhibidores de la PCR
(60), por los genes utilizados para la amplificación o por la técnica de procesamiento de muestras
(53).
La capacidad de H. pylori para crecer y formar biopelículas in vitro e in vivo podría ser una ventaja
para la especie para evitar lesiones causadas por factores estresantes químicos, como la terapia
antimicrobiana in vivo, o el estrés inducido por la privación de nutrientes en el medio ambiente
(56).
La presencia de las biopelículas de H. pylori en agua se ha verificado mediante PCR, sondas de
ADN e hibridación fluorescente in situ (FISH). Así mismo, se ha presentado evidencia in vitro que
sugiere que las interacciones con bacterias específicas (por ejemplo, Mycobacterium chelonae)
dentro de una biopelícula de múltiples especies puede contribuir a la viabilidad de H. pylori (61).
Es necesario tener en cuenta que aunque una población disponga de aguas municipales potables y
de grifos adecuados, la calidad del agua de la potabilización no es la misma en todas partes, además
el estado del acueducto como daños previos que exponen al agua potable a contaminación con
materia fecal, la antigüedad del acueducto y la presencia de biopelículas en las tuberías, deben
estudiarse como factores predictores de contaminación del agua (53).
3.6.3. Asociación con amebas de vida libre (FLA)
Se ha demostrado previamente que las amebas de vida libre (FLA), como Acanthamoeba,
Naegleria, Vermamoeba o Balamuthia, pueden actuar como huéspedes de algunos patógenos
bacterianos capaces de resistir la digestión amebiana. Dentro de ellas, las bacterias sobreviven y
son más resistentes a las condiciones ambientales que normalmente las matarían. Por lo tanto, FLA

actúa como "caballo de Troya" para ARB, permitiendo la supervivencia bacteriana y/o la
transmisión a susceptibles hospedadores (62, 63).
Recientemente, un estudio in vitro ha indicado que H. pylori puede sobrevivir dentro de
Acanthamoeba castellanii después de resistir un tratamiento de desinfección por cloración. FLA
proporciona refugio para H. pylori, lo que le permite sobrevivir bajo diferentes tratamientos y
hacer posible su transmisión a humanos. El análisis de qPCR de las muestras mostró la presencia
de ADN de H. pylori intracelular en 28 (50,9%) de las muestras de aguas residuales y en 11
(91,7%) de las muestras de agua potable. Así mismo, se indicó que no se pudieron recuperar
colonias por cultivo de las muestras de FLA positivas, debido a que la bacteria podría haber
adquirido su estado VBNC, ya que mediante DVC-FISH se observó su viabilidad después del
tratamiento con hipoclorito (62, 64).
3.7. Dosis infectiva
La dosis infectiva estimada para el ser humano se encuentra entre 104 - 1010 UFC/L (65, 66).
Al realizar una evaluación cuantitativa del riesgo microbiano (QMRA), se determinó un nivel
máximo de H. pylori en agua potable de un organismo por litro (<1 organismo/L). Igualmente se
determinó la mediana de H. pylori en agua potable para el rango anual de riesgo de infección en
6.84 x 10-02/L para humanos (67).
3.8. Métodos de detección
En la Tabla 1 se presentan los métodos de detección más comunes para H. pylori, los cuales son
descritos a continuación:
Invasivos No invasivos
Directos Cultivo En diferentes muestras (Jugo
gástrico, saliva, heces) Histología
Técnicas moleculares Antígeno en heces
Indirectos Ureasa rápida Prueba de aliento
Serología
Tabla 1. Métodos de detección de Helicobacter pylori (8)

3.8.1. Cultivo
Habitualmente se le considera una técnica difícil sin embargo bajo ciertas precauciones la mayoría
de los laboratorios consiguen el crecimiento del microorganismo de una manera eficiente. El
cultivo, además de método diagnóstico, tiene la ventaja de tipificar el organismo y determinar su
sensibilidad frente a agentes antibacterianos, lo que tiene importancia desde el punto de vista
epidemiológico para conocer posibles resistencias frente a regímenes terapéuticos. La
especificidad del cultivo es del 100%, no obstante, su sensibilidad es menor a la de otras técnicas
diagnósticas (68).
3.8.2. Histología
Esta técnica consiste en la observación de microorganismos en forma espiral en los cortes
histológicos de las biopsias. La localización de H. pylori es característica en plena barrera mucosa.
Es un método de diagnóstico bastante sencillo y tiene la gran ventaja, de proporcionar
simultáneamente información precisa sobre los cambios morfológicos de la mucosa gástrica (68).
3.8.3. Test rápido de ureasa
La capacidad de H. pylori para producir grandes cantidades de ureasa ha servido para desarrollar
un método rápido y sencillo de diagnóstico. Para su realización se coloca una muestra gástrica en
un tubo con urea y un indicador que cambia la coloración del medio al variar el pH. Si la muestra
contiene ureasa (y por tanto H. pylori) se hidroliza la urea y se forman iones amonio, que aumentan
el pH de la solución, generándose un cambio de color. La rapidez del resultado depende del
número de bacterias presentes en la muestra, siendo suficiente una mínima cantidad de
microorganismos para que la prueba sea positiva (69).
3.8.4. Detección de antígenos en heces
La eficacia diagnóstica de esta prueba de antígeno en heces es alta tanto para el diagnóstico
primario de infección por H. pylori como para el monitoreo de erradicación (sensibilidad y
especificidad del 80-95%). La prueba de antígeno en heces es simple y fácil de usar en la práctica
clínica, su única limitación es la manipulación fecal. Su sencillez, rapidez y economía hacen de
esta técnica una prometedora alternativa para el diagnóstico «no invasor» de la infección (70).
3.8.5. Test de aliento con urea
La prueba del aliento se basa en la capacidad de la ureasa producida por H. pylori para hidrolizar
una solución de urea previamente marcada, bien con 13C o con 14C. El CO2 marcado se absorbe,

se difunde a la sangre, es transportado a los pulmones y de allí excretado a través del aliento
espirado. Tras un ayuno nocturno se recoge una muestra basal de aire espirado, después se
administra una solución de ácido cítrico con el fin de retrasar el vaciamiento gástrico, seguida de
una solución acuosa con la urea marcada (8).
3.8.6. Serología
Las pruebas serológicas para el diagnóstico de la infección por H. pylori se basan en estudiar la
presencia de anticuerpos específicos frente a antígenos de este microorganismo que aparecen como
consecuencia de la respuesta inmunitaria tanto local como sistémica, que se produce tras la
infección. Por ello, estas técnicas indican únicamente una exposición previa al microorganismo,
pero no discriminan entre personas con infección activa y enfermedad e individuos sanos
previamente expuestos a la infección (69).
Uno de los principales problemas para aceptar el agua como posible reservorio de H. pylori es la
incapacidad de aislar y detectar la especie de las muestras de agua mediante técnicas
convencionales. Por lo tanto, la aplicación de este tipo de métodos es vital para la detección rápida,
sensible y específica de H. pylori en entornos acuáticos es de suma importancia.
El principal desafío cuando se realiza un muestreo ambiental es la incapacidad de demostrar la
existencia de H. pylori viable en el agua, que potencialmente puede colonizar el estómago o el
duodeno porque los métodos moleculares utilizados para detectar las bacterias en el medio
ambiente son incapaces de distinguir entre bacterias muertas; sin embargo, han surgido técnicas
como la DVC-FISH (Hibridación fluorescente in situ en combinación con la incubación de conteo
viable directo) la cual que permite la detección específica de células viables y tiene la ventaja de
no ser inactivado por inhibidores de muestras (71).
Ensayos de anticuerpos fluorescentes indirectos (IFA) utilizan anticuerpos específicos que
permiten la enumeración de células tanto cultivables como no cultivables y las formas espiral y
cocoide, pero no puede detectar genes marcadores de virulencia como cagA lo cual no lo hace muy
útil al momento de querer confirmar la presencia de H. pylori a través de estos genes de virulencia
(72). Además de las anteriores técnicas, existen otras como la amplificación isotérmica medida
por bucle (LAMP por sus siglas en inglés) que es un mecanismo más rápido y fácil. Esta técnica
es una reacción de amplificación de una etapa, que puede producir una gran cantidad de copias en
menos de una hora en condiciones isotermas. La ventaja más importante de este método es que no
necesita desnaturalización del ADN (73). Además, se puede realizar sin un termociclador y los
resultados pueden observarse a simple vista. También se utiliza la citometría de flujo, la cual es
una técnica analítica que tiene el potencial de hacer una distinción entre los cuatro estados
fisiológicos de las bacterias: reproductivamente viable, metabólicamente activa, intacta y
permeabilizada. Puede determinar las proporciones de los estados VBNC y VC y las células
muertas, en función de la integridad de la membrana de la bacteria (74).

3.8.7. Métodos de reacción en cadena de la polimerasa
3.8.7.1. PCR de punto final
En 1987, Kary Mullis y Fred Faloona publicaron el artículo titulado “Síntesis específica de ADN
in vitro mediante una reacción en cadena catalizada por polimerasa” donde describen un método
por el cual una secuencia de ácido nucleico puede amplificarse exponencialmente in vitro, siendo
posible alterar la secuencia amplificada o agregarle nueva información. Para lograr la
amplificación se hace necesario conocer con detalle los extremos de la secuencia para sintetizar
oligonucleótidos que hibriden con ellos, y que esté disponible una pequeña cantidad de la
secuencia para iniciar la reacción. El producto de dicha reacción es una molécula de ADN de doble
cadena (dsADN) con los extremos correspondientes a los extremos 5' de los oligómeros empleados
(75). Después de la publicación de este artículo, en 1993 el Dr. Mullis ganó el Premio Nobel de
Química por su descubrimiento de la metodología de Reacción en Cadena de la Polimerasa (PCR)
(76).
La PCR es una técnica simple para copiar un fragmento de ADN en el laboratorio con reactivos
fácilmente disponibles. Debido a que el número de copias aumenta exponencialmente, se pueden
hacer más de 100 mil millones en unas cuantas horas. Se trata de un proceso cíclico donde, con
cada ciclo, el número de fragmentos de ADN se duplica (77).
Esta técnica, como su nombre lo indica, se fundamenta en la polimerasa, una enzima termoestable
obtenida a partir de Thermus aquaticus (Taq). El método utiliza un par de cebadores
oligonucleotídicos sintéticos, cada uno hibridando a una hebra de un fragmento de interés de ADN
de doble cadena (dsADN), con el par abarcando una región que se amplificará exponencialmente.
Los cebadores recocidos actúan como un sustrato para la ADN polimerasa (Taq), creando una
hebra de ADN complementaria a través de la adición secuencial de deoxinucleótidos (dNTP) que
han sido agregados a la mezcla de reacción. Experimentalmente, el proceso consiste en tres pasos
básicos:
I. Desnaturalización por calentamiento a 94 o 95°C.
II. Alineamiento del cebador a las hebras de ADN monocatenario (ADNss) de 55 a 65°C.
III. Extensión de imprimación a 72ºC (78).
En el primer paso (desnaturalización) la doble hélice de ADN se separa formando dos hebras. Para
ello se realiza una incubación de la muestra a altas temperaturas (93-97ºC). La renaturalización se
producirá cuando la temperatura disminuya; en el segundo paso (hibridación), los cebadores se
unen a las zonas 3´ complementarias que flanquean el fragmento que se quiere amplificar, esto se
realiza gracias a la bajada de la temperatura (50-65ºC). Finalmente en la tercera etapa (elongación)
se produce la síntesis de una cadena sencilla (produciéndose un fragmento de doble cadena por la
complementariedad) en la dirección 5´→ 3´ mediante la enzima DNA polimerasa, la cual incorpora
los deoxinucleótidos fosfato presentes en el medio siguiendo la cadena molde. Es importante
mencionar que los productos obtenidos tras la tercera etapa son de dos tipos: un producto corto y

un producto largo. El producto corto tiene una longitud definida por los extremos 5´ de los
cebadores y contiene exactamente la secuencia que se desea amplificar; es el fragmento que se
almacena de manera exponencial durante la reacción. Por otra parte, el producto largo es aquel
que incorpora las cadenas de ADN originales de la muestra y cuyos extremos 3´ no están definidos.
Sin embargo, al final de la PCR, la cantidad de producto corto sintetizado es muy superior en
comparación con el producto largo, por lo que generalmente para posteriores estudios a partir del
producto de la PCR, se desecha (79).
Al repetir estos pasos varias veces, generalmente de 30 a 40 ciclos, la secuencia objetivo del ADN
resultante se amplifica exponencialmente (Figura 1), dando como resultado miles de millones de
copias del llamado "amplicon".
Figura 2. Patrón de crecimiento exponencial de las copias de ADN mediante PCR.
Esta PCR de punto final clásica es una reacción de ausencia/presencia porque mide la formación
de productos de ADN después de un número de ciclos determinado, lo que arroja información
cualitativa sobre la presencia o ausencia de un determinado gen, pero no revela información sobre
la cantidad existente de ADN. Esta técnica de PCR se ha convertido en una de las herramientas
más influyentes en las ciencias biológicas y médicas (78).
Para poder evidenciar el resultado de la PCR de punto final es necesario realizar un corrido
electroforético, mediante el cual es posible separar fragmentos de ADN en función de su tamaño,
visualizarlos mediante una sencilla tinción y determinar así el contenido de ácidos nucleicos de
una muestra, teniendo una estimación de su concentración y grado de entereza. Pudiendo confirmar
así que el amplicón sea del tamaño esperado (80).
A lo largo de los años se han descrito numerosas adaptaciones y aplicaciones de la PCR de punto
final clásica, incluida la PCR múltiple, PCR o cuantitativa (qPCR), PCR con transcriptasa inversa
(RT-PCR), PCR semicuantitativa, PCR competitiva cuantitativa, entre otros.

3.8.2.2. PCR Tiempo real
En la PCR a tiempo real, los procesos de amplificación y detección se producen de manera
simultánea. Además, mediante detección por fluorescencia se puede medir durante la
amplificación la cantidad de ADN sintetizado en cada momento, debido a que la emisión de
fluorescencia en la reacción es proporcional a la cantidad de ADN formado. Esto permite conocer
y registrar en todo momento la cinética de la reacción de amplificación. Los termocicladores para
llevar a cabo la PCR a tiempo real incorporan un lector de fluorescencia, el cual puede ser de dos
tipos: agentes intercalantes y sondas específicas marcadas con fluorocromos
- Agentes intercalantes: Son fluorocromos que aumentan la emisión de fluorescencia
cuando se unen a ADN de doble hélice. El incremento de ADN en cada ciclo se refleja en
un aumento proporcional de la fluorescencia emitida. El más empleado es el SYBR Green.
- Sondas de hibridación específicas: Son sondas marcadas con dos tipos de fluorocromos,
un donador y un aceptor. El proceso se basa en la transferencia de energía fluorescente
mediante resonancia (FRET) entre las dos moléculas. Las más utilizadas son las sondas
TaqMan, beacons y FRET (81).
Entre las ventajas de la PCR en tiempo real está el uso de la fluorescencia para detectar los
productos de la PCR; para diferenciar no solo un resultado positivo de uno negativo, sino también
para poder cuantificar los organismos durante la curva de crecimiento. Además de esto; una ventaja
revolucionaria es que el tubo con los productos de PCR no tiene que abrirse para realizar un corrido
electroforético y observar el amplicón ya que el análisis se realiza en el instrumento durante la
PCR (76).

4. OBJETIVOS
4.1. Objetivo general
Detectar Helicobacter pylori por métodos moleculares como prospectiva para su identificación a
partir de cuerpos de agua.
4.2. Objetivos específicos
• Elaborar un manuscrito de revisión sobre el papel del agua en la ruta de transmisión de H.
pylori.
• Comparar dos metodos moleculares para la detección de H. pylori.
• Establecer el límite de detección de ADN de H. pylori.

5. METODOLOGÍA
5.1. Elaboración de un manuscrito de revisión de tema sobre el papel del agua en la ruta de
transmisión de H. pylori
Se realizó una búsqueda amplia y sistemática de la literatura en las bases de datos PubMed y
Science Direct utilizando los términos Helicobacter pylori, water, detection methods, VBNC y
cocoid form, para lo cual se empleó el conector booleano AND y se restringió la bibliografía a
artículos originales y algunos de revisión publicados en los últimos 5 años.
5.2. Análisis de los cebadores para la detección molecular de H. pylori
Para la detección molecular de H. pylori se utilizaron los cebadores ureA-F y ureA-R debido a que
estos presentan una homología del 100% con el gen ureA específico de H. pylori. Por tal motivo
y mediante el Software GeneRunner versión 6.5.51., se buscó confirmar la especificidad de los
cebadores para amplificar el gen ureA y verificar tanto el tamaño del amplicón a obtener como la
homología señalada anteriormente.
En la tabla 2 se describen los cebadores, sus secuencias respectivas y la fuente bibliográfica de los
mismos.
Tabla 2. Cebadores utilizados para la detección molecular de H. pylori mediante PCR.
Gen Cebador Secuencia Artículo
ureA ureA- F 5’- CGTGGCAAGCATGATCCAT -3’ Schabereiter-Gurtner C,
Hirschl AM, Dragosics B,
Hufnagl P, Puz S, Kova´ch
Z, et al. Novel Real-Time
PCR Assay for Detection of
Helicobacter pylori
Infection and Simultaneous
Clarithromycin
Susceptibility Testing of
Stool and Biopsy
Specimens. Journal of
Clinical Microbiology.
2004; 42(10): 4512–4518.
ureA-R 5’- GGGTATGCACGGTTACGAGTTT -3’
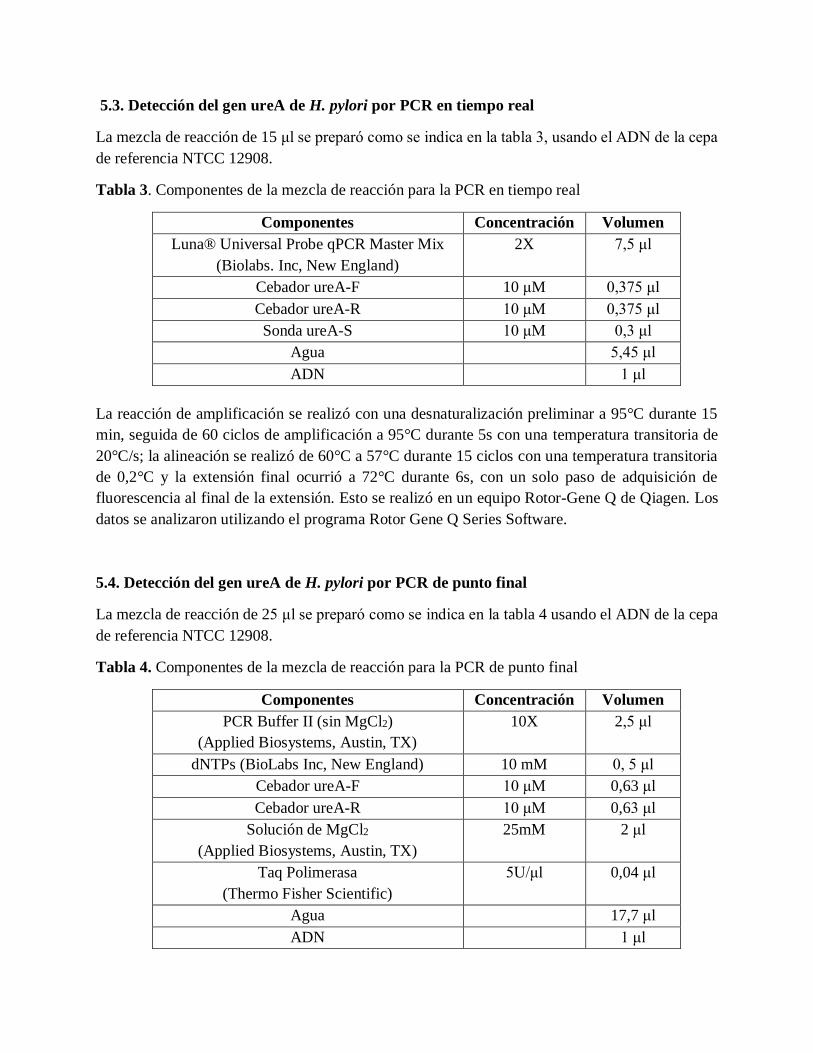
5.3. Detección del gen ureA de H. pylori por PCR en tiempo real
La mezcla de reacción de 15 μl se preparó como se indica en la tabla 3, usando el ADN de la cepa
de referencia NTCC 12908.
Tabla 3. Componentes de la mezcla de reacción para la PCR en tiempo real
Componentes Concentración Volumen
Luna® Universal Probe qPCR Master Mix
(Biolabs. Inc, New England)
2X 7,5 μl
Cebador ureA-F 10 μM 0,375 μl
Cebador ureA-R 10 μM 0,375 μl
Sonda ureA-S 10 μM 0,3 μl
Agua 5,45 μl
ADN 1 μl
La reacción de amplificación se realizó con una desnaturalización preliminar a 95°C durante 15
min, seguida de 60 ciclos de amplificación a 95°C durante 5s con una temperatura transitoria de
20°C/s; la alineación se realizó de 60°C a 57°C durante 15 ciclos con una temperatura transitoria
de 0,2°C y la extensión final ocurrió a 72°C durante 6s, con un solo paso de adquisición de
fluorescencia al final de la extensión. Esto se realizó en un equipo Rotor-Gene Q de Qiagen. Los
datos se analizaron utilizando el programa Rotor Gene Q Series Software.
5.4. Detección del gen ureA de H. pylori por PCR de punto final
La mezcla de reacción de 25 μl se preparó como se indica en la tabla 4 usando el ADN de la cepa
de referencia NTCC 12908.
Tabla 4. Componentes de la mezcla de reacción para la PCR de punto final
Componentes Concentración Volumen
PCR Buffer II (sin MgCl2)
(Applied Biosystems, Austin, TX)
10X 2,5 μl
dNTPs (BioLabs Inc, New England) 10 mM 0, 5 μl
Cebador ureA-F 10 μM 0,63 μl
Cebador ureA-R 10 μM 0,63 μl
Solución de MgCl2
(Applied Biosystems, Austin, TX)
25mM 2 μl
Taq Polimerasa
(Thermo Fisher Scientific)
5U/μl 0,04 μl
Agua 17,7 μl
ADN 1 μl

La reacción de amplificación se realizó con una desnaturalización preliminar a 95°C durante 4
min, seguida de 30 ciclos de amplificación a 95°C durante 30s, alineación a 55°C durante 30 s y
elongación a 72° durante 30 s. Se realizó una extensión final a 70°C durante 7 min y una etapa
extra de enfriamiento a 4°C durante 59 min. Esto se realizó en un Termociclador convencional
Swift™ MiniPro, ESCO.
Luego se realizó separación electroforética de los productos amplificados a 100 voltios por 120
minutos, en un gel de agarosa al 3% en buffer de corrido TAE 1X. El marcador de peso molecular
usado fue Hyperladder™ 50 pb (Bioline). El gel fue teñido con SYBR safe y las bandas se
evidenciaron en transiluminador bajo luz ultravioleta.
5.5. Preparación de diluciones a partir de ADN de cepas de referencia
Se utilizó un ADN control: AMPLIRUN® Helicobacter pylori DNA CONTROL (Vircell,
14MBC049001) obtenido a partir de la cepa de referencia es la ATCC 26695, el cual tenía una
concentración de 18.000 copias/µl. A partir del ADN control se realizaron diluciones 1:10 hasta
alcanzar una concentración teórica de 1.8 copias/µl.
Por otro lado, se utilizó ADN de la cepa de referencia NTCC 12908, el cual se encontraba a una
concentración de 16 ng/µl. A partir de este ADN se realizaron diluciones hasta alcanzar una
concentración teórica de 1 ng/µl.
5.6. Determinación de la concentración de las diluciones
Las diluciones realizadas a partir de las dos cepas de referencia (ATCC 26695 y NTCC 12908)
fueron cuantificadas mediante dos metodologías: espectrofotometría y fluorometría.
Para la cuantificación mediante espectrofotometría, se utilizó un Espectrofotómetro Multiskan Go,
usando la placa μDrop™ (Thermo Scientific™) y se analizó mediante Soft Skant Software 5.0 for
microplate readers. El procedimiento consistió en colocar 2 µl de cada dilución por duplicado en
la placa μDrop y un blanco, que podía ser TE 1X o agua; una vez depositadas todas las muestras,
se introdujo la placa en el espectrofotómetro, el cual realizó la lectura y arrojó los datos obtenidos.
Para la cuantificación mediante fluorometría, se utilizó el Fluorometro QubitTM 4 Fluorometer de
Invitrogen (Thermo Fisher Scientific, Singapore, 739256). En este caso se inició preparando la
solución de trabajo, para la cual se diluyó 1:200 el reactivo QubitTM dsDNA BR Reagent
concentrado en DMSO (Life Technologies Corporation, Eugene, 97402) en el Buffer Qubit TM
dsDNA BR Buffer (Life Technologies Corporation, Eugene, 97402). Igualmente se prepararon las
dos soluciones estándar Qubit TM dsDNA BR Standard #1 (Life Technologies Corporation,
Eugene, 97402) y Qubit TM dsDNA BR Standard #2, (Life Technologies Corporation, Eugene,
97402). Estas soluciones al igual que las muestras se prepararon a un volumen final de 200 μl,

agregando, para el caso de los estándares 10 μl de cada uno y 190 μl de la solución de trabajo y
para el caso de las muestras, 2 μl de cada una y 198 μl de la solución de trabajo. Luego cada tubo
fue insertado de manera individual (primero los estándares), se tomó la lectura y se registraron los
datos obtenidos.
5.7. Límite de detección de ADN de H. pylori
El límite de detección se abordó por ambas metodologías moleculares (PCR en tiempo real y PCR
de punto final); así mismo, se abordó mediante dos estrategias, las cuales fueron el límite de
detección de copias de ADN/µl y de nanogramos de ADN/µl de H. pylori.
Para abordar estas dos estrategias se utilizaron las diluciones realizadas a partir de las dos cepas
de referencia (ATCC 26695 y NTCC 12908).
Una vez confirmadas mediante cuantificación las concentraciones de las anteriores diluciones, se
procedió a realizar las reacciones en cadena de la polimerasa, utilizando para ambas los cebadores
ureA-F/ureA-R bajo las condiciones descritas en los numerales 5.3 y 5.4. Todos los ensayos se
realizaron por triplicado, para los dos métodos moleculares y para las dos unidades de medición
utilizadas.
Posterior a esto se utilizó la siguiente fórmula (82) mediante la cual fue posible obtener tanto el
número de copias de ADN a partir de la cantidad de ADN en nanogramos, como la cantidad de
nanogramos de ADN a partir del número de copias; esto último al despejar la fórmula como se
muestra a continuación:
Número de copias de ADN =cantidad de ADN (ng) ∗ 6.022x10ˆ23
longitud del ADN ∗ 1x10ˆ9ngml
∗ 650 Daltons
Despeje de la fórmula de tal manera que el valor a obtener sea la cantidad de ADN en nanogramos,
así:
Cantidad de ADN (ng) =Número de copias de ADN ∗ longitud del ADN ∗ 1x10ˆ9
ngml
∗ 650 Daltons
6.022x10ˆ23

6. RESULTADOS Y DISCUSIÓN
6.1. Manuscrito de revisión sobre el papel del agua en la ruta de transmisión de H. pylori
En la tabla 5 se describen algunos de los artículos consultados y un breve resumen de los hallazgos.
Diferentes trabajos han logrado detectar H. pylori en cuerpos de agua, siendo capaz de resistir y
tolerar las prácticas de desinfección normalmente empleadas en el tratamiento del agua potable.
Igualmente se han descrito algunos mecanismos que le permiten sobrevivir a H. pylori en
ambientes adverso para ella, entre ellos está la forma viable no cultivable a la cual pasan cuando
están expuestas a condiciones estresantes o diferentes a las que están acostumbradas, siendo esto
una estrategia de adaptación para la supervivencia a largo plazo. Este mecanismo de supervivencia
es el más estudiado ya que se ha demostrado que H. pylori en su forma cocoide presente en agua
conserva una serie de factores de virulencia encargados de la colonización e invasión de la mucosa
gástrica; por ejemplo, mantiene detectable los niveles de ureasa, continúa la síntesis de proteínas
en pequeñas cantidades y produce también pequeñas cantidades de ADN; demostrando que estas
células no cultivables son formas infecciosas. En el tejido gástrico, por ejemplo, las formas
cocoides (VBNC) pueden permanecer latentes durante mucho tiempo y retener los factores de
virulencia, por lo que también pueden ayudar a los fracasos del tratamiento y a la recurrencia de
la infección y de las enfermedades gastroduodenales. Otra característica importante que le permite
a la bacteria sobrevivir en agua es su capacidad de adherirse a diferentes materiales y formar
biopelículas.
Se encuentra que uno de los principales problemas para aceptar el agua como posible reservorio
de H. pylori es la incapacidad de aislar rutinariamente la especie de las muestras de agua mediante
técnicas convencionales de cultivo microbiológico. Por lo tanto, la aplicación de métodos
moleculares para la detección rápida, sensible y específica de H. pylori en entornos acuáticos es
de suma importancia. El artículo de revisión se presenta como Anexo 1.

6.2. Análisis de los cebadores usados para la detección molecular de H. pylori
GeneRunner mostró un alineamiento específico de los cebadores ureA-F/ureA-R con la secuencia
del gen Ureasa (Figura 2) de H. pylori, así mismo arrojó el tamaño en pares de bases (pb) del
amplicón, corroborando la longitud de 77pb.
Tabla 5. Resumen de artículos analizados en el manuscrito generado de revisión del tema sobre
el papel del agua en la transmisión de H. pylori.
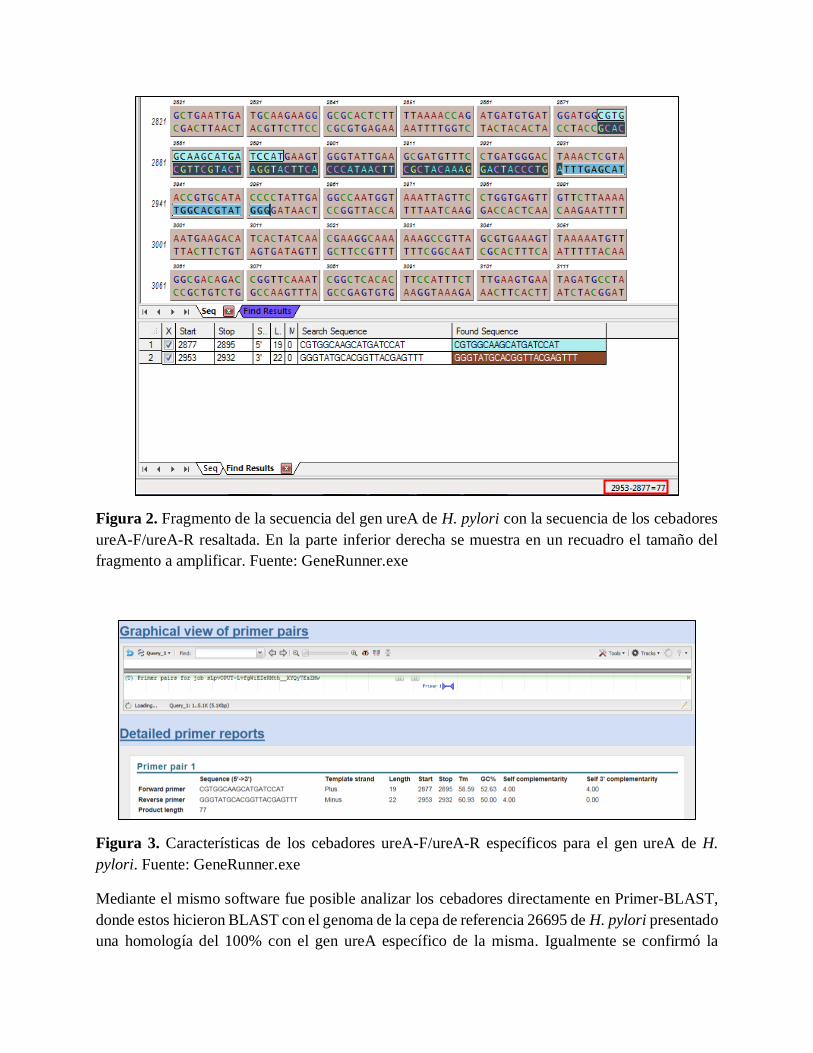
Figura 2. Fragmento de la secuencia del gen ureA de H. pylori con la secuencia de los cebadores
ureA-F/ureA-R resaltada. En la parte inferior derecha se muestra en un recuadro el tamaño del
fragmento a amplificar. Fuente: GeneRunner.exe
Figura 3. Características de los cebadores ureA-F/ureA-R específicos para el gen ureA de H.
pylori. Fuente: GeneRunner.exe
Mediante el mismo software fue posible analizar los cebadores directamente en Primer-BLAST,
donde estos hicieron BLAST con el genoma de la cepa de referencia 26695 de H. pylori presentado
una homología del 100% con el gen ureA específico de la misma. Igualmente se confirmó la

información obtenida en GeneRunner como la longitud del amplicón y las posiciones de inicio y
fin de cada uno de ellos; además arrojó información adicional como la temperatura de melting
(Tm), el porcentaje Guanina/Citosina (GC%) y la complementariedad. En la Figura 3 se muestran
en detalle los cebadores ureA-F/ureA-R
La PCR es una herramienta de suma importancia utilizada en muchas investigaciones. Es necesario
conocer la reacción y adecuar sus condiciones antes de hacer uso extensivo de ella; uno de los
pasos fundamentales es contar con los cebadores o primers y con las características específicas de
cada uno de ellos. Esto permitirá sortear exitosamente uno de los pasos primordiales para obtener
la mejor calidad y cantidad del producto a amplificar. El diseño cuidadoso de cebadores es uno de
los aspectos más importantes para obtener los resultados esperados; cebadores mal diseñados
pueden amplificar otros fragmentos de ADN distintos a los buscados (amplificación inespecífica).
Por este motivo, el uso de GeneRunner, un programa de utilidad en genética molecular que permite
introducir una secuencia de ADN, observar su cadena complementaria, encontrar una subsecuencia
dentro de esa secuencia, ver las enzimas de restricción y más (83), fue de gran ayuda para verificar
la homología, la especificidad en el alineamiento y la longitud del fragmento obtenido usando los
primers ureA-F/ureA-R como se puede observar n la Figura 3.
6.3. Comparación de dos metodologías moleculares para la detección de H. pylori
Al observar los resultados obtenidos para la PCR de punto final (figura 4) y PCR tiempo real
(figura 5) se demostró que fue posible amplificar la cepa de referencia NTCC 12908 bajo estas dos
metodologías. El resultado obtenido para PCR en tiempo real se evidenció igualmente mediante
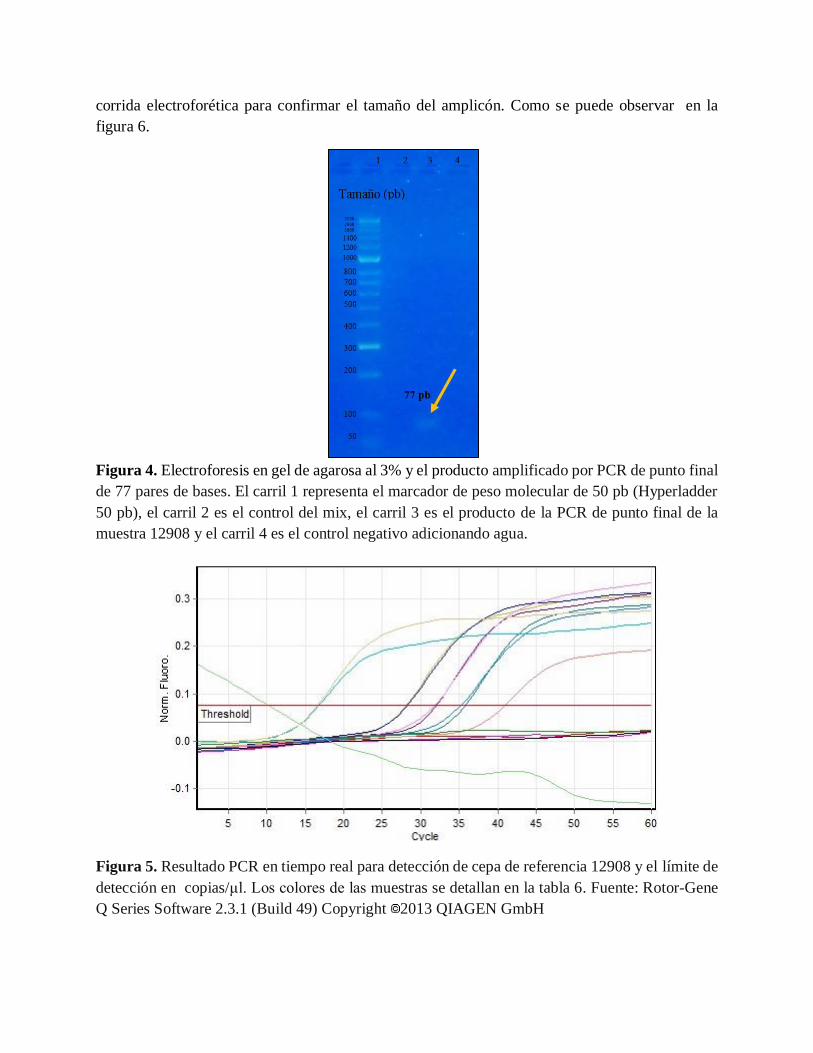
corrida electroforética para confirmar el tamaño del amplicón. Como se puede observar en la
figura 6.
Figura 4. Electroforesis en gel de agarosa al 3% y el producto amplificado por PCR de punto final
de 77 pares de bases. El carril 1 representa el marcador de peso molecular de 50 pb (Hyperladder
50 pb), el carril 2 es el control del mix, el carril 3 es el producto de la PCR de punto final de la
muestra 12908 y el carril 4 es el control negativo adicionando agua.
Figura 5. Resultado PCR en tiempo real para detección de cepa de referencia 12908 y el límite de
detección en copias/μl. Los colores de las muestras se detallan en la tabla 6. Fuente: Rotor-Gene
Q Series Software 2.3.1 (Build 49) Copyright 2013 QIAGEN GmbH
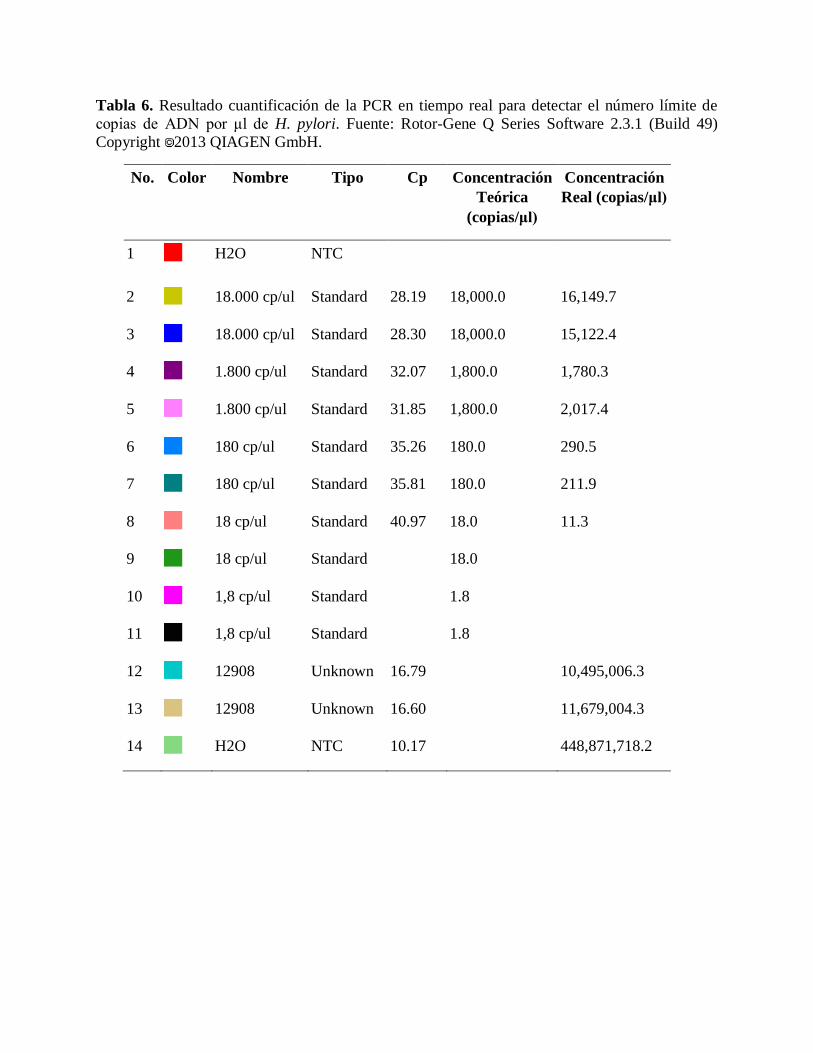
Tabla 6. Resultado cuantificación de la PCR en tiempo real para detectar el número límite de
copias de ADN por μl de H. pylori. Fuente: Rotor-Gene Q Series Software 2.3.1 (Build 49)
Copyright 2013 QIAGEN GmbH.
No. Color Nombre Tipo Cp Concentración
Teórica
(copias/μl)
Concentración
Real (copias/μl)
1 H2O NTC
2
18.000 cp/ul Standard 28.19 18,000.0 16,149.7
3
18.000 cp/ul Standard 28.30 18,000.0 15,122.4
4
1.800 cp/ul Standard 32.07 1,800.0 1,780.3
5
1.800 cp/ul Standard 31.85 1,800.0 2,017.4
6
180 cp/ul Standard 35.26 180.0 290.5
7
180 cp/ul Standard 35.81 180.0 211.9
8
18 cp/ul Standard 40.97 18.0 11.3
9
18 cp/ul Standard
18.0
10
1,8 cp/ul Standard
1.8
11
1,8 cp/ul Standard
1.8
12
12908 Unknown 16.79
10,495,006.3
13
12908 Unknown 16.60
11,679,004.3
14
H2O NTC 10.17
448,871,718.2

Figura 6. Electroforesis en gel de agarosa al 3% y el producto amplificado por PCR en tiempo
real de 77 pares de bases. El carril 1 representa el producto de la PCR en tiempo real de la muestra
12908, el carril 2 es el control del Mater Mix, el carril 3 es el control negativo adicionando agua y
el carril 4 es el marcador de peso molecular de 50 pb (Hyperladder 50 pb).
El diagnóstico por PCR de H. pylori tiene una sensibilidad y especificidad de 95% y su principal
ventaja es que se puede detectar el microorganismo sin importar la viabilidad de la bacteria en las
muestras. La especificidad de esta técnica viene dada por el uso de oligonucleótidos sintéticos,
específicos para determinado gen y que facilitan la amplificación de una secuencia nucleotídica,
que a su vez es específica para H. pylori; es por esta razón que en los últimos años se ha diseñado
una variedad de pruebas diagnósticas para esta bacteria por medio de PCR, basándose en:
oligonucleótidos provenientes de diferentes genes, entre ellos el gen de la ureasa A (ureA) (84).
Al utilizar los primers ureA-F y ureA-R para amplificar un fragmento específico del gen ureA se
obtuvieron resultados óptimos como se puede ver en las figuras 4 y 5, ya que si fue posible
amplificar el ADN la cepa de referencia NTCC 12908 por las dos técnicas, confirmando que se
trataba del amplicón de 77pb en el gel de agarosa al 3%. Aunque los resultados fueron los
esperados para las dos metodologías es importante realizar una comparación que permita
identificar los pros y contra de cada técnica para elegir la más adecuada; la PCR de punto final
mide el producto de PCR acumulado al final de los ciclos; sin embargo, esta no cuantifica, pero la
comparación de la intensidad de la banda amplificada en un gel con los estándares de una
concentración conocida puede proporcionarle resultados semicuantitativos. Por otro lado, la PCR
en tiempo real mide el producto amplificado a medida que se produce; esta técnica cuantifica al
recopilar los datos obtenidos durante la fase de crecimiento exponencial (log) cuando la cantidad

del producto de la PCR es directamente proporcional a la cantidad de ácido nucleico de la plantilla
(85).
Entre las desventajas de la PCR de punto final se encuentran su baja precisión, baja sensibilidad,
baja resolución, no es automatizada, solo discrimina basándose en el tamaño, los resultados no se
expresan como números y se debe realizar un tratamiento post-PCR. Entre las ventajas de la PCR
en tiempo real se encuentra un aumento en el rango de detección, no se necesita un procesamiento
posterior a la PCR, recopila datos en la fase de crecimiento exponencial, un aumento en la señal
de fluorescencia es directamente proporcional a la cantidad de amplicones generados. Por lo
anterior, la PCR de punto final es recomendada para amplificación de ADN en secuenciación,
genotipado y clonación y la PCR en tiempo real se recomienda para cuantificación de la expresión
génica, verificación de micromatrices, control de calidad y validación de ensayos, detección de
patógenos, genotipado de polimorfismos de un solo nucleótico (SNP), variación del número de
copias, análisis de microARN, cuantificación viral, experimentos de ARN de interferencia (85).
6.4. Diluciones obtenidas a partir de ADN de cepas de referencia
A partir de la cepa de ATCC 26695 que se encontraba a una concentración 18.000 copias/µl, se
obtuvieron las siguientes diluciones: 1.800 copias/µl, 180 copias/µl, 18 copias/µl y 1.8 copias/µl.
Por otro lado, usando ADN de la cepa de referencia NTCC 12908, la cual se encontraba a una
concentración de 16 ng/µl se obtuvieron las siguientes diluciones 16 ng/µl, 13 ng/µl, 10 ng/µl, 7
ng/µl, 4 ng/µl y 1 ng/µl.
6.5. Determinación de la concentración de las diluciones
Tabla 7. Concentración esperada de las diluciones realizadas a partir del ADN de la cepa de
referencia 12908 frente a las concentraciones obtenidas mediante fluorómetro.
La cuantificación del ADN usando el método de fluorometría no permitió detectar la concentración
de las copias/μl ya que el kit con el que se prepararon los estándares de detección, permitía una
cuantificación entre 2 y 1000 ng y las copias/μl se encontraban en un rango no detectable. Por el
Concentración esperada Concentración calculada
16 ng/µl 15.3 ng/µl
13 ng/µl 13.0 ng/µl
10 ng/µl 9.8 ng/µl
7 ng/µl 7.6 ng/µl
4 ng/µl 5.3 ng/µl
1 ng/µl 2.2 ng/µl

contrario, para el caso de las diluciones realizadas a partir del ADN de la cepa patrón 12908, se
pudo medir la concentración y así compararla con la concentración teórica a la que se pretendía
llegar en nanogramos/μl. Estos resultados se muestran en la tabla 7. Por otro lado la cuantificación
del ADN usando el método de espectrofotometría arrojó unos resultados muy inconsistentes, ya
que las muestras con mayor concentración de ADN, arrojaban un resultado negativo e inferior a
aquellas muestras de menor concentración,
Existen diversos métodos para estimar la concentración de ácidos nucleicos en una preparación.
Si la muestra es pura (no contiene cantidades significativas de contaminantes como proteínas,
fenol, agarosa o incluso otros ácidos nucleicos), su medición mediante espectrofotometría es la
más adecuada. Por el contrario, si la cantidad de ADN o ARN es muy pequeña, o si la muestra
contiene grandes cantidades de impurezas, la cuantificación puede llevar a cabo por la intensidad
de fluorescencia. Debido a su rapidez, simplicidad y ausencia de daño, la cuantificación de ADN
por espectrofotometría ha sido ampliamente utilizada. Sin embargo, esta técnica es poco sensible,
y para la mayoría de los espectrofotómetros se requiere una concentración de ADN de al menos 1
µg/mL (86). El método de cuantificación por fluorometría se basa en tintes de base de
fluorescencia que se unen específicamente al ADN, ARN o proteína y el método de cuantificación
por espectrofotometría se basa en mediciones de absorbancia UV (mide la absorbancia a 260 nm
y la relación 260nm/280nm. Un aspecto importante es que la fluorometría mide con precisión tanto
el ADN como el ARN de una misma muestra, en cambio la espectrofotometría no es capaz de
distinguir una de la otra (87).
Las diferencias entre la técnica de fluorometría y espectrofotometría se basan en que el fluorómetro
cuantifica con precisión el ADN en muestras con concentraciones tan bajas como 10
picogramos/μl; claro está que esto depende del equipo utilizado, ya que el utilizado en esta
investigación media en un rango entre 2 y 1.000 ng/μl. Por el contrario, el espectrofotómetro no
se recomienda para concentraciones por debajo de 2 nanogramos/μl, ya que cuando se miden
concentraciones inferiores, presenta una alta variación en los resultados (87).
6.6. Límite de detección de ADN de H. pylori
Límite de detección de copias/μl de H. pylori por PCR en tiempo real
El límite de detección de copias de ADN por μl de H. pylori obtenido mediante PCR en tiempo
real fue de 11.3 copias/μl como se señala en la figura 5 y se describe en la tabla 6. Se esperaría que
el límite de detección fuera de 18 copias/μl en base a las diluciones hechas; sin embargo como ya
se sabe que la PCR en tiempo real cuantifica, esta arrojo el valor real de la concentración mínima
que se detectó el cual fue de 11.3 copia/μl como se mencionó anteriormente. En la tabla 6 se
presenta el nombre de cada muestra y el color al que corresponde en la gráfica, el ciclo en el que
empezó a emitir fluorescencia la concentración teórica y la concentración real o calculada por el
equipo. En este límite de detección de copias de ADN por μl de H. pylori obtenido mediante PCR
en tiempo real, el valor de Cp (ciclo del proceso de amplificación en el que se supera un umbral

establecido de fluorescencia o valor “threshold”) (88) fue de 40.97 para la muestra correspondiente
a la concentración de 11.3 copia/μl como se observa en la figura 5 señalado con una flecha. Este
valor concuerda con los valor de Cp obtenidos en otro estudio que detectó el gen ureA para H.
pylori, los cuales variaron entre 40.0 y 44.4 (89).
En los últimos años se han realizado estudios que determinan el límite de detección de H. pylori
en agua bajo la concentración de UFC/ml, observándose en uno de los casos que los cebadores
usados lograron detectar desde 3.0 x 103 UFC en un litro de agua (90) sin embargo, las UFC se
limitan a unidades de mayor tamaño, lo cual puede ser perjudicial al momento de detectar la
bacteria en agua, ya que esta puede encontrase en concentraciones muy pequeñas, inferiores
incluso a una célula, por lo cual la detección de la bacteria mediante copias de ADN amplia el
rango detección y así evita pasar por alto las concentraciones mínimas de la bacteria en agua, lo
cual podría evitar problemas de salud pública a los que conlleva la infección por H. pylori.
Barrera et al., obtuvieron en su investigación que el límite de detección de la técnica de PCR en
tiempo real para Granulovirus en muestras de larva, suelo y en un bioplaguicida fue de 1,25 x 105
copias del gen (125.000 copias del gen), lo que equivale a 6,4 x 10-4 ng (0,00064 nanogramos) de
ADN (91), siendo este un valor superior al límite de detección obtenido en el presente proyecto.
Por otro lado, en un estudio basado en la detección molecular de Legionella spp a partir de muestras
ambientales mediante qPCR (PCR cuantitativa en tiempo real), se obtuvo que el límite de
detección fue de 25 copias por 10 μl de la muestra, lo que equivale a 2,5 copias/μl (0,0000094 ng)
(92), siendo este valor un poco más similar, pero sin embargo inferior al límite de detección que
obtuvimos para H. pylori mediante la misma metodología.
Límite de detección de copias de ADN por μl de H. pylori por PCR de punto final
La PCR de punto final no pudo detectar ninguna de las muestras estándar de copias por μl debido
a que esta técnica es poco sensible y no amplifica valores tan pequeños. Por este motivo, otros
estudios han implementado y recomiendan la PCR en tiempo real como la técnica más adecuada,
sensible y específica para determinar concentraciones dadas en cantidad de copias de ADN.
Aunque se dice que la PCR de punto final o convencional presenta un límite de detección más alto
de lo deseable, como se pudo comprobar en este estudio; el estudio realizado por Moreno y
Agudelo, presentan unos resultados que divergen por completo, ya que mediante una PCR
convencional simple obtuvieron un límite de detección para Leptospira interrogans de 0,012 ng,
lo que equivale a 2.38×103 copias (2.380 copias de ADN) (93), siendo estos valores mucho
menores a los que se obtuvieron mediante esta técnica
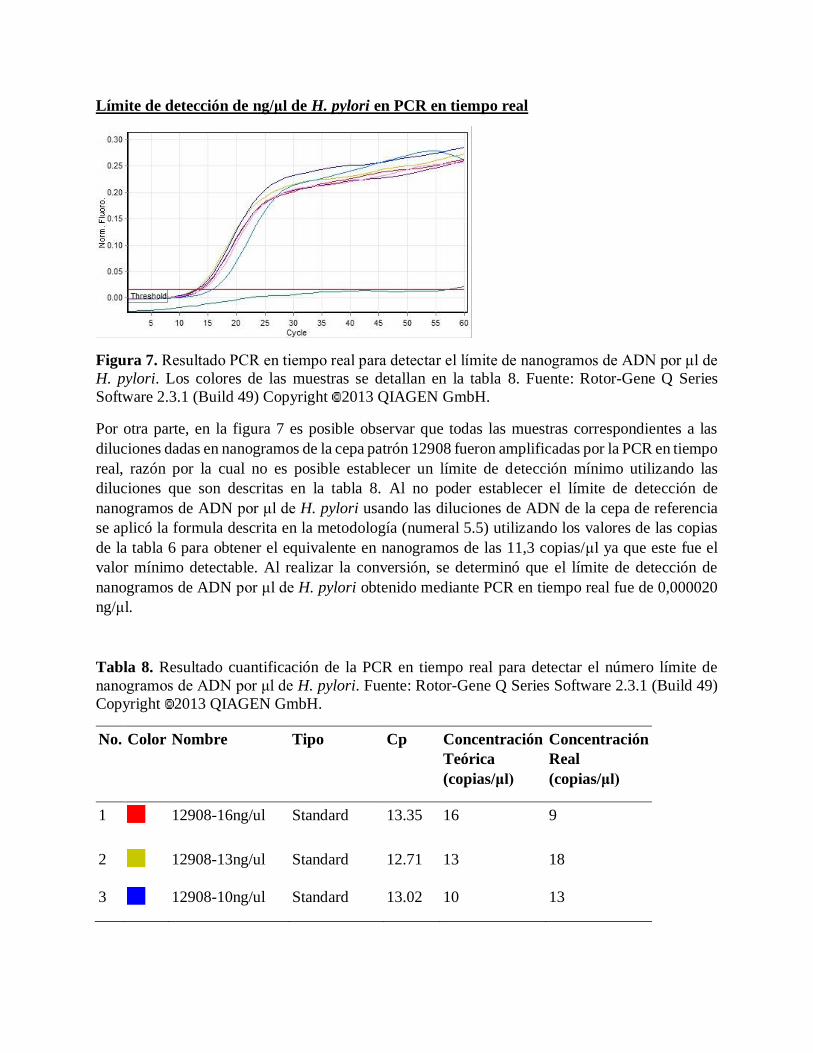
Límite de detección de ng/μl de H. pylori en PCR en tiempo real
Figura 7. Resultado PCR en tiempo real para detectar el límite de nanogramos de ADN por μl de
H. pylori. Los colores de las muestras se detallan en la tabla 8. Fuente: Rotor-Gene Q Series
Software 2.3.1 (Build 49) Copyright 2013 QIAGEN GmbH.
Por otra parte, en la figura 7 es posible observar que todas las muestras correspondientes a las
diluciones dadas en nanogramos de la cepa patrón 12908 fueron amplificadas por la PCR en tiempo
real, razón por la cual no es posible establecer un límite de detección mínimo utilizando las
diluciones que son descritas en la tabla 8. Al no poder establecer el límite de detección de
nanogramos de ADN por μl de H. pylori usando las diluciones de ADN de la cepa de referencia
se aplicó la formula descrita en la metodología (numeral 5.5) utilizando los valores de las copias
de la tabla 6 para obtener el equivalente en nanogramos de las 11,3 copias/μl ya que este fue el
valor mínimo detectable. Al realizar la conversión, se determinó que el límite de detección de
nanogramos de ADN por μl de H. pylori obtenido mediante PCR en tiempo real fue de 0,000020
ng/μl.
Tabla 8. Resultado cuantificación de la PCR en tiempo real para detectar el número límite de
nanogramos de ADN por μl de H. pylori. Fuente: Rotor-Gene Q Series Software 2.3.1 (Build 49)
Copyright 2013 QIAGEN GmbH.
No. Color Nombre Tipo Cp Concentración
Teórica
(copias/μl)
Concentración
Real
(copias/μl)
1 12908-16ng/ul Standard 13.35 16 9
2
12908-13ng/ul Standard 12.71 13 18
3
12908-10ng/ul Standard 13.02 10 13

4
12908-7ng/ul Standard 13.76 7 6
5
12908-4ng/ul Standard 14.00 4 5
6
12908-1ng/ul Standard 15.74 1 1
7
MMix NTC 56.73
Límite de detección de nanogramos de ADN por μl de H. pylori por PCR de punto final
Figura 8. Electroforesis en gel de agarosa al 3% y los productos amplificado por PCR de punto
final y PCR en tiempo real de 77 pares de bases. Los carriles 1, 2, 3, 4, 5 y 6 representan los
productos de la PCR de punto final, los cuales equivalen a 16 ng/μl, 13 ng/μl, 10 ng/μl, 7 ng/μl, 4
ng/μl y 1 ng/μl respectivamente, el carril 7 es el control negativo adicionando agua y el carril 8 es
el marcador de peso molecular de 50 pb (Hyperladder 50 pb). Los carriles 9, 10, 11, 12, 13 y 14
representan los productos de la PCR en tiempo real, los cuales equivalen a 16 ng/μl, 13 ng/μl, 10
ng/μl, 7 ng/μl, 4 ng/μl y 1 ng/μl respectivamente
Como se puede ver en la figura 8 el límite de detección de nanogramos de ADN por μl de H. pylori
obtenido mediante PCR de punto final fue de 5 ng/μl , debido a que fue la cantidad más baja de
ADN que se logró amplificar mediante la mencionada metodología. Se esperaría que el límite de

detección fuera de 4 ng/μl en base a las diluciones hechas; sin embargo, cuando se realizó la PCR
en tiempo real utilizando las diluciones en nanogramos/μl, este arrojo el valor real de las
concentraciones, obteniendo que la concentración mínima que se detectó fue de 5 ng/μl (tabla 6).
En esta figura también se evidencian los tamaños de los amplicones obtenidos mediante PCR en
tiempo real (Figura 5) los cuales se encuentran ubicados al lado derecho después del marcador de
peso molecular. Aunque para esta metodología no es necesario realiza corrido electroforético, se
realizó para verificar el tamaño de los amplicones.
Se puede notar que en la figura 8 es difícil visualizar las bandas amplificadas; sin embargo, al
momento de observar directamente el gel en el transiluminador, fue posible visualizar las 5 bandas,
donde la última indicaba la mínima cantidad que se detectó y se señala con una flecha.
Este fenómeno de que las bandas de ADN no se ven nítidamente en el gel puede ser causado
porque el ADN se ha degradado, porque la cantidad o la concentración de ADN en las muestras
cargadas al gel eran insuficientes; porque las moléculas de ADN al ser de un tamaño pequeño se
difundieron durante la tinción o debido a un gel de agarosa muy grueso, por lo cual se recomienda
evitar la contaminación con nucleasas, aumentar la cantidad de ADN, y preparar geles más
delgados (94, 95).
Por otra parte, la PCR de punto final no amplificó ninguna de las muestras estándar de copias por
μl, por lo cual se calculó a través de la página scienceprimer.com (se fundamenta en la misma
fórmula descrita en la metodología, numeral 5.5), el equivalente en copias de los 5 nanogramos
(concentración real), debido a que esa fue la cantidad mínima detectable mediante PCR de punto
final como puede confirmarse a través de la electroforesis en la figura 7. Fue así como se determinó
que el límite de detección de copias de DNA por μl de H. pylori obtenido mediante PCR de punto
final fue de 2,7 x 106 lo que corresponde 2.700.000 copias/μl.
Finalmente se puede decir que, aunque las técnicas de PCR en tiempo real y PCR de punto final
tienen una alta sensibilidad (mayor en el caso de la PCR en tiempo real), presentan la desventaja
de no poder discriminar entre células viables y no viables. Por este motivo y para lograr la
detección de células viables mediante estas dos metodologías, se han desarrollado diferentes
alternativas, como el uso del agente intercalante de ADN fluoróforo propono monoazida (PMA),
el cual es un colorante fotorreactivo de alta afinidad que se une preferentemente a dsDNA. Es
impermeable a las membranas celulares intactas y, por lo tanto, penetra solo a través de las
membranas dañadas de las células muertas para intercalarse en el ADN, evitando su amplificación
(96). El uso de este agente intercalante le confiere un valor agregado a la técnica, en especial
cuando se trata de detectar H. pylori en agua, debido a que, además de buscar detectar la presencia
de la bacteria, lo ideal sería detectar la presencia de células viables, ya que al estar metabólicamente
activas y con todos sus factores de virulencia, son capaz de causar enfermedad en las personas que
consuman el agua. Al poder confirmar la presencia de células viables en una cantidad tal que sea
infecciosa para el humano, se podría confirmar definitivamente el agua como mecanismo de
transmisión para H. pylori.

7. CONCLUSIONES
Se establecieron las condiciones de la PCR tiempo real y de punto final para la detección del gen
ureA utilizando los cebadores ureA-F y ureA-R de Helicobacter pylori y se comprobó su
especificidad.
Se logró determinar el límite de detección en nanogramos de ADN por μl de H. pylori para PCR
en tiempo real y para PCR de punto final.
Se logró determinar el límite de detección en copias de ADN por μl de H. pylori para PCR en
tiempo real y para PCR de punto final.
Se determinaron las diferencias entre cuantificación por método espectrofotométrico y
fluorométrico.
Se determinaron las diferencias entre PCR en tiempo real y PCR de punto final.

8. PERSPECTIVAS A FUTURO
En esta investigación se abordó una primera fase que consistió en la escogencia del método
molecular de reacción en cadena de la polimerasa y el límite de detección; se propone una segunda
fase mediante la cual se contamine agua con la bacteria bajo condiciones controladas de laboratorio
y una tercera fase para investigar directamente en cuerpos de agua la detección de H pylori.
Probar nuevas condiciones para la PCR con el fin de optimizar el límite de detección para poder
hallar concentraciones más pequeñas de la bacteria.
Estandarizar una metodología molecular que pueda ser utilizada por entidades públicas que
suministren agua potable a la población para la detección de H. pylori con el fin de disminuir los
casos de incidencia de la infección que puedan ser adquiridos a través del agua.

9. AGRADECIMIENTOS
Agradecemos a Dios, quien nos ha llenado de fortaleza y sabiduría en cada momento de este largo
trayecto.
A nuestras madres, Martha Liliana Ceballos y Nancy Quintero, quienes han velado por nuestro
bienestar y educación, siendo nuestro apoyo en todo momento.
A nuestras familias, por ser acompañantes en tiempos duros de trabajo y ahora de satisfacción en
el cumplimiento de una nueva etapa.
A la Universidad Libre seccional Pereira y a cada uno de los docentes que hicieron parte de nuestro
proceso educativo; gracias a ellos, la formación que hemos tenido en los últimos 5 años de nuestras
vidas ha sido de suma importancia.
Agradecemos especialmente a la Doctora Adalucy Álvarez Aldana y al Doctor Fernando Siller
López por su paciencia, dedicación y apoyo, por dedicar gran parte de su tiempo y compartir su
valioso conocimiento académico, profesional y personal; porque siempre nos motivaron a seguir
adelante con este proceso y nos facilitaron el acceso a herramientas fundamentales en la
elaboración del proyecto.
A nuestros compañeros, los cuales conocimos y nos han acompañado a lo largo de estos años,
convirtiéndose en personas muy importantes para nosotras.
A esas personas que aunque hoy no nos acompañan, se sienten orgullosos de ver lo que hemos
logrado a través de todo este recorrido.
Nuestra gratitud para todos ellos y la recompensa que, de aquí en adelante, día a día, pondremos
nuestro mayor empeño para ser unas excelentes profesionales y unas personas que siempre darán
lo mejor de sí mismas.

10. REFERENCIAS BIBLIOGRÁFICAS
1. Marshall BJ. One Hundred Years of Discovery and Rediscovery of Helicobacter pylori and
Its Association with Peptic Ulcer Disease. En: Mobley H, Mendz G & Hazell S, editors.
Helicobacter pylori: Physiology and Genetics. Washington (D.C.): ASM Press; 2001.
Disponible en: https://www.ncbi.nlm.nih.gov/books/NBK2432/
2. Kusters JG, van Vliet AH, Kuipers EJ. Pathogenesis of Helicobacter pylori infection. Clin
Microbiol Rev [Internet] 2006 [Consultado 20 oct 2018]; 19(3): 449-490. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1539101/
3. Pajares JM, Gisbert JP. Helicobacter pylori: its discovery and relevance for medicine. Rev
Esp Enf Dig [Internet] 2006 [Consultado 20 oct 2018]; 98(3): 770-785. Disponible en:
http://scielo.isciii.es/pdf/diges/v98n10/punto.pdf
4. McColl KE. Clinical practice. Helicobacter pylori infection. N Engl J Med [Internet] 2010
[Consultado 20 oct 2018]; 362(17): 1597-604. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/20427808
5. Proença-Modena JL, Olszanski Acrani G, Brocchi M. Helicobacter pylori: phenotypes,
genotypes and virulence genes. Future Microbiol [Internet] 2009 [Consultado 20 oct 2018];
4(2): 223-240. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/19257848
6. Brown LM. Helicobacter Pylori: Epidemiology and Routes of Transmission. Epidemiol
Rev [Internet] 2000 [Consultado 20 oct 2018]; 22(2): 283-297. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/11218379
7. Mobley HLT, Mendz GL, Hazell SL. Helicobacter pylori: Physiology and Genetics.
[Internet]. Washington (D.C.): American Society for Microbiology (AMS Press); 2001.
[Consultado 20 oct 2018]. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/21290711
8. Rivera M, Contreras F, Terán A, Fouillioux C. Helicobacter Pylori: Enteropatógeno
frecuente del ser humano. AVFT. [Internet] 2004 [Consultado 21 oct 2018]; 23(2).
Disponible en: http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0798-
02642004000200003
9. Agudo Pena S. Estudio molecular de los factores de virulencia y de la resistencia a
claritromicina en la infección por Helicobacter pylori. Tesis Doctoral. [Internet] 2010
[Consultado 21 oct 2018]. Madrid: Universidad Complutense de Madrid. Disponible en:

https://www.researchgate.net/publication/47761657_Estudio_molecular_de_los_factores
_de_virulencia_y_de_la_resistencia_a_claritromicina_en_la_infeccion_por_Helicobacter
_pylori
10. Cellini L. Helicobacter pylori: A chameleon-like approach to life. World J Gastroenterol.
[Internet] 2014 [Consultado 21 oct 2018]; 20(19): 5575-5582. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4024766/
11. Alarcón T, Baquero M, Domingo D, López-Brea M, Royo G. Diagnóstico microbiológico
de la infección por Helicobacter pylori. [Internet]. Madrid: SEIMC; 2004 [Consultado 21
oct 2018]. Disponible en:
https://www.seimc.org/contenidos/documentoscientificos/procedimientosmicrobiologia/s
eimc-procedimientomicrobiologia17.pdf
12. Croxen MA, Sisson G, Melano R, Hoffman PS. The Helicobacter pylori Chemotaxis
Receptor TlpB (HP0103) Is Required for pH Taxis and for Colonization of the Gastric
Mucosa. J Bacteriol. [Internet] 2006 [Consultado 21 oct 2018]; 188(7): 2656-2665.
Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/16547053
13. Fernandes RM, Silva H, Oliveira R, Almeida C, Azevedo NF, Vieira MJ. Morphological
transition of Helicobacter pylori adapted to water. Future Microbiol [Internet]. 2017
[Consultado 21 oct 2018]; 12:1167-1179. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/28879777
14. Jung DO, Helene KB, Marios G, Jian X, Robert SF, Lucinda AF, et al. The complete
genome sequence of a chronic atrophic gastritis Helicobacter pylori strain: Evolution
during disease progression. Proc Natl Acad Sci U S.A. [Internet] 2006 [Consultado 21 oct
2018]; 103(26): 9999-10004. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1480403/
15. Tomb JF, White O, Kerlavage AR, Clayton RA, Sutton GG, Fleischmann RD, et al. The
complete genome sequence of the gastric pathogen Helicobacter pylori. Nature [Internet]
1997 [Consultado 21 oct 2018]; 388(6642: 539-547. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/9252185
16. Kienesberg S, Zechner EL. Helicobacter pylori. Reference Module in Biomedical
Sciences. [Internet] 2017 [Consultado 21 oct 2018]. Elsevier Inc.
17. Salama N, Guillemin K, McDaniel TK, Sherlock G, Tompkins L, Falkow S. A whole-
genome microarray reveals genetic diversity among Helicobacter pylori strains. Proc Natl
https://www.ncbi.nlm.nih.gov/pubmed/?term=Oliveira%20R%5BAuthor%5D&cauthor=true&cauthor_uid=28879777

Acad Sci U S A [Internet] 2000 [Consultado 21 oct 2018]; 97(26), 14668-73. Disponible
en: https://www.ncbi.nlm.nih.gov/pubmed/11121067
18. Han YH, Liu WZ, Shi YZ, Lu LQ, Xiao S, Zhang QH, et al. Comparative genomics
profiling of clinical isolates of Helicobacter pylori in Chinese populations using DNA
microarray. J Microbiol [Internet] 2007 [Consultado 21 oct 2018] 45(1), 21-28. Disponible
en:
https://www.researchgate.net/publication/6464852_Comparative_Genomics_profiling_of
_clinical_isolates_of_Helicobacter_pyloli_in_Chinese_populations_using_DNA_microar
ray
19. McClain M, Shaffer C, Israel D, Peek R, Cover T. Genome sequence analysis of
Helicobacter pylori strains associated with gastric ulceration and gastric cancer. BMC
Genomics. [Internet] 2009 [Consultado 21 oct 2018]; 10: 3. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2627912/
20. Chalk PA, Roberts AD, Blows WM. Metabolism of pyruvate and glucose by intact cells of
Helicobacter pylori studied by 13C NMR spectroscopy. Microbiology. [Internet] 1994
[Consultado 21 oct 2018]; 140: 2085-92. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/7921258
21. Cesareo SD, Langton SR. Kinetic properties of Helicobacter pylori urease compared with
jack bean urease. FEMS Microbiol Lett. [Internet] 1992 [Consultado 21 oct 2018]; 78(1),
15-21. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/1468614
22. Suerbaum S, Josenhans C. Helicobacter pylori evolution and phenotypic diversification in
a changing host. Nat Rev Microbiol. [Internet] 2007 [Consultado 21 oct 2018]; 5(6), 441-
452. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/17505524
23. Rivas F, Hernández F. Helicobacter pylori: Factores de virulencia, patología y
diagnóstico. Rev Biomed. [Internet] 2000 [Consultado 24 oct 2018]; 11:187-205.
Disponible en: http://www.cirbiomedicas.uady.mx/revbiomed/pdf/rb001136.pdf
24. Eaton KA, Brooks CL, Morgan DR, Krakowka S. Essential role of urease in pathogenesis
of gastritis induced by Helicobacter pylori in gnotobiotic piglets.
Infect Immun. [Internet] 1991 [Consultado 6 nov 2018]; 59 (7): 2470–2475. Disponible
en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC258033/
25. UniProt. UniProtKB P14916 (URE23_HELPY). [Internet] Disponible en:
https://www.uniprot.org/uniprot/P14916

26. Chang WL, Yeh YC, Sheu BS. The impacts of H. pylori virulence factors on the
development of gastroduodenal diseases. J Biomed Sci. [Internet] 2018 [Consultado 20
nov 2018]; 25(1): 68. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6131906/pdf/12929_2018_Article_466.p
df
27. Archampong TNA, Asmah RH, Wiredu EK, Gyasi RK, Nkrumah KN, Rajakumar K.
Epidemiology of Helicobacter pylori infection in dyspeptic Ghanaian patients. The Pan
African Medical Journal [Internet] 2015 [Consultado 21 oct 2018]; 20: 178. Disponible en:
http://www.panafrican-med-journal.com/content/article/20/178/full/
28. Ghasemi-Kebria F, Ghaemi E, Azadfar S, Roshandel G. Epidemiology of Helicobacter
pylori infection among Iranian children. Arab Journal of Gastroenterology. [Internet] 2013
[Consultado 21 oct 2018]; 14(4): 169-172. Disponible en:
http://europepmc.org/abstract/MED/24433647
29. Figueroa M, Cortés A, Pazos Á, Bravo LE. Sensibilidad in vitro a amoxicilina y
claritromicina de Helicobacter pylori obtenido de biopsias gástricas de pacientes en zona
de bajo riesgo para cáncer gástrico. Biomédica. [Internet] 2012 [Consultado 21 oct 2018];
32(1): 32-42. Disponible en:
https://www.revistabiomedica.org/index.php/biomedica/article/view/454
30. Eusebi LH, Zagari RM, Bazzoli F. Epidemiology of Helicobacter pylori infection.
Helicobacter. [Internet] 2014 [Consultado 21 oct 2018]; 1: 1-5. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/25167938
31. Windsor HM, Abioye-Kuteyi EA, Leber JM, Morrow SD, Bulsara MK, Marshall BJ.
Prevalence of Helicobacter pylori in Indigenous Western Australians: comparison
between urban and remote rural populations. Med J Aust. [Internet] 2005 [Consultado 24
oct 2018]; 182(5): 210-213. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/15748129
32. Bruce MG, Maaroos HI.Epidemiology of Helicobacter pylori infection. Helicobacter.
[Internet] 2008 [Consultado 24 oct 2018]; 1: 1-6. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/18783514
33. Choi SS, Chivers PT, Berg DE. Point Mutations in Helicobacter pylori's fur Regulatory
Gene that Alter Resistance to Metronidazole, a Prodrug Activated by Chemical Reduction.
PLoS ONE. [Internet] 2011 [Consultado 24 oct 2018]; 6: 18236. Disponible en:
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0018236

34. Quiroga AJ, Cittelly DM, Bravo MM. Frecuencia de los genotipos babA2, oipA y cagE de
Helicobacter pylori en pacientes colombianos con enfermedades gastroduodenales.
Biomédica. [Internet] 2005 [Consultado 24 oct 2018]; 25: 325-334. Disponible en:
https://www.revistabiomedica.org/index.php/biomedica/article/download/1357/1472
35. Cittelly DM, Huertas GM, Martínez JD, Oliveros R, Posso H, Bravo MM, et al. Los
genotipos de Helicobacter Pylori en gastritis no atrófica difieren de los encontrados en
úlcera Péptica, lesiones premalignas y cáncer gástrico. Rev Med Chile. [Internet] 2002
[Consultado 24 oct 2018]; 130(2): 1-12. Disponible en:
https://scielo.conicyt.cl/scielo.php?script=sci_arttext&pid=S0034-98872002000200003
36. Galvis Arevalo A. Prevalencia de los genotipos de virulencia de Helicobacter pylori CagA,
VacA, BabA2 e IceA en pacientes colombianos con dispepsia funcional. [Tesis de
Pregrado]. Bogotá: Pontificia Universidad Javeriana; 2010. Disponible en:
https://repository.javeriana.edu.co/handle/10554/818
37. Macenlle García RM. Prevalencia de la infección por Helicobacter pylori en la población
general adulta de la provincia de Ourense y estudio de factores de riesgo asociados. [Tesis
de Pregrado]. Madrid: Universidad de Santiago de Compostela. Disponible en:
https://minerva.usc.es/xmlui/bitstream/handle/10347/2375/9788497509657_content.pdf?s
equence=1
38. Blum AL, Talley NJ, O'Moráin C, van Zanten SV, Labenz J, Stolte M, et al. Lack of Effect
of Treating Helicobacter pylori Infection in Patients with Nonulcer Dyspepsia. N Engl J
Med. [Internet] 1998 [Consultado 24 oct 2018]; 339: 1875-1881. Disponible en:
https://www.nejm.org/doi/full/10.1056/NEJM199812243392602
39. Gisbert JP. Enfermedades relacionadas con Helicobacter pylori: dispepsia, úlcera y
cáncer gástrico. Gastroenterología y Hepatología. [Internet] 2011 [Consultado 24 oct
2018]; 33(S2): 1-92. Disponible en: http://www.elsevier.es/es-revista-gastroenterologia-
hepatologia-14-articulo-enfermedades-relacionadas-con-helicobacter-pylori-
S0210570511700176
40. Gómez M, Ruíz O, Páramo Hernández D, Albis R, Sabbagh LC. Erradicación del
Helicobacter pylori: encuesta realizada por la Asociación Colombiana de
Gastroenterología. Revista Colombiana de Gastroenterologia. Rev Colom Gastro.
[Internet] 2015 [Consultado 24 oct 2018]; 30(1), 25-31. Disponible en:
http://www.redalyc.org/pdf/3377/337738642005.pdf

41. Di L. Subcomittee on chronic abdominal pain. Chronic Abdominal pain in children.
Pediatrics. [Internet] 2005 [Consultado 24 oct 2018]; 115: 370-38.
42. Majumdar D, Bebb J, Atherton J. Helicobacter pylori infection and peptic ulcers.
Medicine. [Internet] 2011 [Consultado 24 oct 2018]; 39(3): 154- 161. Disponible en:
https://www.medicinejournal.co.uk/article/S1357-3039(10)00309-9/fulltext?mobileUi=0
43. Schöttker B, Adamu MA, Weck MN, Müller H, Brenner H. Helicobacter pylori infection,
chronic atrophic gastritis and major cardiovascular events: A population-based cohort
study. Atherosclerosis. [Internet] 2012 [Consultado 24 oct 2018]; 220(2): 569-574.
Disponible en: https://www.atherosclerosis-journal.com/article/S0021-9150(11)01100-
2/fulltext?mobileUi=0
44. Ding S. Helicobacter pylori infection, oncogenic pathways and epigenetic mechanisms in
gastric carcinogenesis. Future Oncol. [Internet] 2010 [Consultado 24 oct 2018]; 6(5): 851-
862. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/20465395
45. Andriani A, Miedico A, Tedeschi L, Patti C, Di Raimondo F, Leone M, et al. Management
and long-term follow-up of early stage H. pylori-associated gastric MALT-lymphoma in
clinical practice: An Italian, multicentre study. Dig Liver Dis. [Internet] 2009 [Consultado
24 oct 2018]; 41(7): 467-473. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/18945654
46. Parsonnet J, Hansen S, Rodriguez L, Gelb AB, Warnke RA, Jellum E, et al. Helicobacter
pylori Infection and Gastric Lymphoma. N Engl J Med. [Internet] 1994 [Consultado 24 oct
2018]; 330(18): 1267-1271. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/8145781
47. Fischbach W, Goebeler-Kolve ME, Dragosics B, Greiner A, Stolte M. Long term outcome
of patients with gastric marginal zone B cell lymphoma of mucosa associated lymphoid
tissue (MALT) following exclusive Helicobacter pylori eradication therapy: experience
from a large prospective series. Gut. [Internet] 2004 [Consultado 24 oct 2018]; 53(1), 34-
37. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/14684573
48. Breckan RK, Paulssen EJ, Asfeldt AM, Kvamme JM, Straume B, Florholmen J. The All-
Age Prevalence of Helicobacter pylori Infection and Potential Transmission Routes. A
Population-Based Study. Helicobacter [Internet]. 2016 [Consultado 24 oct 2018]; 21(6):
586-595. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/27172105

49. Palomino Camargo C, Tomé Boschian E. Helicobacter pylori: Rol del agua y los alimentos
en su transmisión. An Venez Nutr. [Internet] 2012 [Consultado 24 oct 2018]; 25(2): 85-
93. Disponible en: http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0798-
07522012000200005
50. Cava F, Cobas G. Dos décadas de Helicobacter pylori. Vaccimonitor. [Internet] 2003
[Consultado 24 oct 2018]; 12(1): 1-10. Disponible en:
http://www.redalyc.org/pdf/2034/203414596001.pdf
51. Ramírez A, Mendoza D, Leey J, Guerra J. Estudio del Helicobacter pylori en el Perú. Rev
Perú Med Exp Sal Púb. [Internet] 2002 [Consultado 24 oct 2018]; 19(4): 209-214.
Disponible en: http://www.scielo.org.pe/scielo.php?pid=S1726-
46342002000400009&script=sci_abstract
52. Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clin Microbiol Rev. [Internet] 1997
[Consultado 25 oct 2018]; 10(4): 720-41. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/9336670
53. Cuervo CM, Gaviria AM. Detección de Helicobacter pylori en muestras de agua y
biopelícula de los grifos de las instituciones educativas oficiales en la ciudad de Medellín.
Acta Med Colomb [Internet]. 2017 [Consultado 25 oct 2018]; 42(2): 121-128. Disponible
en: http://www.scielo.org.co/pdf/amc/v42n2/0120-2448-amc-42-02-00121.pdf
54. Laam L, Mendis N, Trigui H, Oliver JD, Faucher SP. The importance of the viable but non-
culturable state in human bacterial pathogens. Front Microbiol [Internet] 2014
[Consultado 25 oct 2018]; 5: 258. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040921/
55. Mali S, Vrushi L, Hili N. The impact of drinking water on the prevalence of H. pylori in
humans in Elbasan, Albania. Eur Water [Internet] 2017 [Consultado 25 oct 2018]; 58: 263-
266. Disponible en:
https://www.researchgate.net/publication/321184977_The_impact_of_drinking_water_on
_the_prevalence_of_H_Pylori_in_humans_in_Elbasan_Albania
56. García A, Salas Jara MJ, Herrera C, González C. Biofilm and Helicobacter pylori: From
environment to human host. World J Gastroenterol [Internet] 2014 [Consultado 25 oct
2018]; 20(19): 5632-5638. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4024771/

57. Bayona Rojas MA, Gutiérrez Escobar AJ. Biopelícula: un mecanismo de supervivencia de
Helicobacter pylori. Rev U.D.C.A Act & Div Cient [Internet] 2013 [Consultado 25 oct
2018]; 16(2): 335-342. Disponible en:
http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0123-
42262013000200007&lng
58. Montero Campos V, Hernández Soto A, Camacho Sandoval J. Culture and Molecular
Identification of Helicobacter pylori in Drinking Water from Areas of High and Low
Incidence of Gastric Cancer in Costa Rica. Op J Med Microbiol [Internet] 2014
[Consultado 25 oct 2018]; 4(4): 261-269. Disponible en:
https://www.scirp.org/journal/PaperInformation.aspx?PaperID=52896
59. Bahrami A. Rahimi E. Detection of Helicobacter pylori in City Water, Dental Units Water,
and Bottled Mineral Water in Isfahan, Iran. Scientific World Journal [Internet] 2013
[Consultado 25 oct 2018]; 280510. Disponible
en: https://www.ncbi.nlm.nih.gov/pubmed/23606812
60. Otero R W. Helicobacter pylori en agua potable ¿Es la ruta de la infección? Acta Med
Colomb [Internet] 2017 [Consultado 25 oct 2018]; 42(2): 87-89. Disponible en:
http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-
24482017000200087&lng=en
61. Hathroubi S, Servetas SL, Windham I, Merrell DS, Ottemann KM. Helicobacter pylori
Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol Mol Biol Rev
[Internet] 2018 [Consultado 25 oct 2018]; 82(2) pii: e00001-18. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/29743338
62. Moreno-Mesonero L, Moreno Y, Alonso JL, Ferrús MA. Detection of viable Helicobacter
pylori inside free-living amoebae in wastewater and drinking water samples from Eastern
Spain. Environ Microbiol [Internet] 2017 [Consultado 25 oct 2018]; 19(10): 4103-4112.
Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/28707344
63. Amirhooshang A, Ramin A, Ehsan A, Mansour R, Shahram B. High frequency of
Helicobacter pylori DNA in drinking water in Kermanshah, Iran, during June–November
2012. J Water Health. [Internet] 2014 [Consultado 25 oct 2018]; 12(3): 504-512.
Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/25252354
64. Bai X, Xi C, Wu J. Survival of Helicobacter pylori in the wastewater treatment process
and the receiving river in Michigan, USA. J Water Health [Internet] 2016 [Consultado 25
oct 2018]; 14(4): 692-8. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/27441864

65. Graham DY, Opekun AR, Osato MS, El-Zimaity HM, Lee CK, Yamaoka Y, Monath TP.
Challenge model for Helicobacter pylori infection in human volunteers. Gut 2004; 53(9):
1235-43. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/15306577
66. Valdivieso M, Bussalleu A, Sexton R, Boehnke K, Osorio S, Novoa I, et al. Clinical,
Epidemiologic, and Genomic Studies (SWOG S1119) of Helicobacter Pylori in Lima, Peru:
Role of Contaminated Water. J Cancerol [Internet] 2016 [Consultado 25 oct 2018]; 3: 52-
63. Disponible en: http://www.journalofcancerology.com/pdf/JCancer2016_02_52-63.pdf
67. Ryan M, Hamilton K, Hamilton M, Haas CN. Evaluating the Potential for a Helicobacter
pylori Drinking Water Guideline. Risk Anal [Internet] 2014 [Consultado 25 oct 2018];
34(9): 1651-62. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/24660760
68. Gisbert JP, Critical review of diagnostic methods of Helicobacter pylori infection.
Gastroenterologia y Hepatologia. [Internet]. 2000 [citado 2018 Nov 28]; 23(3): 109-152.
Disponible en: http://www.elsevier.es/es-revista-gastroenterologia-hepatologia-14-
articulo-revision-critica-los-metodos-diagnosticos-9806
69. Ordoñez Frías JS, Otero Regino W, Aspectos prácticos en métodos diagnósticos para la
infección por Helicobacter pylori: una revisión narrativa. Revista de Gastroenterología del
Perú. 20017; 37(3): 246-253. Disponible en:
http://www.scielo.org.pe/pdf/rgp/v37n3/a09v37n3.pdf
70. Mones J, Gisbert JP, Borda F, Domínguez-Muñoz E. Indicaciones, métodos diagnósticos
y tratamiento erradicador de Helicobacter pylori. Rev Esp Enferm Dig 2005; 97: 348-74.
Disponible en: http://scielo.isciii.es/scielo.php?script=sci_arttext&pid=S1130-
01082005000500007
71. Santiago P, Moreno Y, Ferrús MA. Identification of Viable Helicobacter pylori in Drinking
Water Supplies by Cultural and Molecular Techniques. Helicobacter [Internet]. 2015
[Consultado 1 nov 2018]; 20(4): 252-9. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/25655472
72. Chomvarin C, Wongboot W, Tirapattanun A, Kanthawong S, Wongwajana S, Tongpim S,
et al. Detection of Helicobacter pylori in Aquati Environments and Drinking Waters in
Northeastern Thailand. Chiang Mai J Sci. [Internet]. 2017 [Consultado 1 nov 2018]; 44(3):
731-741. Disponible en:
https://www.researchgate.net/publication/318786035_Detection_of_Helicobacter_pylori_
in_aquatic_environments_and_drinking_waters_in_northeastern_Thailand.

73. Chamanrokh P, Shahhosseiny MH, Assadi MM, Nejadsattari T, Esmaili D. Three Tests
Used to Identify Non-Culturable Form of Helicobacter pylori in Water Sample.
Jundishapur J Microbiol [Internet]. 2015 [Consultado 1 nov 2018]; 8(4): e16811.
Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4449853.
74. Assadi MM, Chamanrokh P, Whitehouse CA, Huq A. Methods for Detecting the
Environmental Coccoid Form of Helicobacter pylori. Front Public Health [Internet]. 2015
[Consultado 1 nov 2018]; 3: 147. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4446911/
75. Mullis KB, Faloona FA. Specific synthesis of DNA in vitro via a polymerase-catalyzed
chain reaction. Methods Enzymol. [Internet] 1987 [Consultado 8 nov 2018]; 155: 335-50.
Disponible en: https://www.sciencedirect.com/science/article/pii/0076687987550236
76. Barton Rogers B. The Evolution of the Polymerase Chain Reaction to Diagnose Childhood
Infections. Pediatric and Developmental Pathology. [Internet] 2015 [Consultado 8 nov
2018]; 18(6): 495–503. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/26701384
77. Mullis KB. The unusual origin of the polymerase chain reaction. Sci Am. [Internet] 1990
[Consultado 8 nov 2018]; 262(4): 56-65. Disponible en:
https://pdfs.semanticscholar.org/14bf/9cc89c9885d7ad46a384d0e5661ac07c752f.pdf
78. Overbergh L, Vig S, Coun F, Mathieu C. Quantitative Polymerase Chain Reaction. En:
Patrinos GP. Molecular Diagnostics. Ed. 3. Academic Press; 2016. p. 41 -58. Disponible
en: http://dx.doi.org/10.1016/B978-0-12-802971-8.00004-3
79. Mas E, Poza J, Ciriza J, Zaragoza P, Osta R, Rodellar C. Fundamento de la Reacción en
Cadena de la Polimerasa (PCR). AquaTIC. [Internet] 2001 [Consultado 8 nov 2018]; (15).
Disponible en: www.revistaaquatic.com/ojs/index.php/aquatic/article/download/139/128
80. Fierro Fierro F. Electroforesis de ADN. En: Cornejo Romero A, Serrato Díaz A, Rendón
Aguilar B, Rocha Muníve MG. Herramientas Moleculares aplicadas en Ecología: Aspectos
Teóricos y Prácticos. Ed. 1. 2014. p. 27-51. Disponible en:
https://micrositios.inecc.gob.mx/publicaciones/libros/710/electroforesis.pdf
81. Costa J. Reacción en Cadena de la Polimerasa (PCR) a tiempo real. Enferm Infecc
Microbiol Clin [Internet] 2004 [Consultado 6 nov 2018]; 22(5): 299-305. Disponible en:
https://www.sciencedirect.com/science/article/pii/S0213005X0473092X

82. Lorenz TC. Polymerase Chain Reaction: Basic Protocol Plus Troubleshooting and
Optimization Strategies. J Vis Exp [Internet]. 2012 [Consultado 20 nov 2018]; 68: e3998.
Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4846334/
83. Jimenez. A. Manual de neurogenética. [Internet] España: Díaz de Santos S.A.; 2003.
[Consultado 20 nov 2018]. Disponible en: https://es.scribd.com/document/312077394/A-
Jimenez-Scrig-Manual-de-Neurogentica
84. Premoli Gloria, González Anajulia, Millán-Mendoza Beatriz, Percoco Tiziana, Vielma
Amilcar. Diagnóstico de Helicobacter pylori mediante la reacción en cadena de la
polimerasa. Rev Cubana Med Trop [Internet]. 2004 [Consultado 20 nov 2018]; 56(2):
85-90. Disponible en: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0375-
07602004000200001&lng=es
85. ThermoFisher Scientific. Real-Time vs. Digital PCR vs. Traditional PCR. [Internet].
ThermoFisher Scientific. [Consultado 20 nov 2018]. Disponible en:
https://www.thermofisher.com/co/en/home/life-science/pcr/real-time-pcr/real-time-pcr-
learning-center/real-time-pcr-basics/real-time-vs-digital-vs-traditional-pcr.html
86. Guevara P, Campelo R, Clark R, Guariglia V, Moronta F, Rizzolo K. Manual de
Laboratorio [Internet]. Venezuela: Universidad Central de Venezuela; 2009. [Consultado
22 nov 2018]. Disponible en:
http://www.ciens.ucv.ve:8080/generador/sites/geneticamolecular/archivos/Manual%20La
b%20Fund%20Gen%20Mol%20Vers%2020%20mayo%202009.pdf
87. Whittaker J. Nanodrop vs. Qubit [Internet]. Personalized genes; 2015. [Consultado
22 nov 2018]. Disponible en: http://www.personalizedgenes.com/nanodrop-or-qubit/
88. Catalán Cuenca V. La técnica de PCR: ¿en qué fase de desarrollo técnico se encuentra?
Rev Salud Ambiental. [Internet]. 2006 [Consultado 24 nov 2018]; 6(1-2): 85-88.
Disponible en: http://ojs.diffundit.com/index.php/rsa/article/viewFile/301/260
89. Schabereiter-Gurtner C, Hirschl AM, Dragosics B, Hufnagl P, Puz S, Kova´ch Z, et al.
Novel Real-Time PCR Assay for Detection of Helicobacter pylori Infection and
Simultaneous Clarithromycin Susceptibility Testing of Stool and Biopsy Specimens.
Journal of Clinical Microbiology [Internet]. 2004 [Consultado 23 nov 2018]; 42(10):
4512–4518. Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC522301/
90. Valverde Osorio CA. Estandarización de un método de detección molecular de
Helicobacter pylori a partir de agua. [Tesis de pregrado]. Armenia: Universidad del
Quindío, 2010.

91. Barrera G, Murcia J, Cerón J, Cuartas P, Guzmán C, Villamizar L. Real Time PCR: a useful
methodology for detection and quantitation of Granulovirus [Internet] 2016 [Consultado
25 nov 2018]; 18(2): 24-31. Disponible en:
http://www.scielo.org.co/pdf/biote/v18n2/v18n2a04.pdf
92. Ferrando Monllor N. Desarrollo y validación de un nuevo sistema de detección molecular
basado en qPCR de Legionella spp. en muestras ambientales. [Tesis de maestría]. Alicante:
Instituto Universitario del Agua y de las Ciencias Ambientales; 2013. Disponible en:
https://iuaca.ua.es/es/master-agua/documentos/-gestadm/trabajos-fin-de-master/tfm-2013-
nuria-ferrando.pdf
93. Moreno N, Agudelo Flórez P. Aplicación de las pruebas de PCR convencional simple y
múltiple para la identificación de aislamientos de Leptospira spp. en Colombia. Rev Peru
Med Exp Salud Pública. [Internet]. 2010 [Consultado 25 nov 2018]; 27(4): 548-56.
Disponible en: http://www.scielo.org.pe/pdf/rins/v27n4/a09v27n4.pdf
94. Yábar Varas CA. Manual de procedimientos de electroforesis para proteínas y ADN
[Internet]. Lima: Ministerio de Salud, Instituto Nacional de Salud; 2008. [Consultado 20
nov 2018]. Disponible en:
http://www.ins.gob.pe/insvirtual/images/otrpubs/pdf/Manual%20Electroforesis%2038.pd
f
95. Herráez A. Plásmidos en electroforesis. [Internet]. Universidad de Alcalá. 2011.
[Consultado 20 nov 2018]. Disponible en: http://biomodel.uah.es/an/plasmido/inicio.htm
96. Moreno Y, Ferrus MA. Specific detection of culturable Helicobacter pylori cells from
wastewater treatment plants. Helicobacter [Internet]. 2012 [Consultado 25 nov 2018];
17(5): 327-32. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/22967115.

ANEXO 1
Manuscrito de Revisión
El agua como ruta de transmisión de Helicobacter pylori
Valeria Ceballos-Toro*, María del Mar Álvarez-Quintero*, Adalucy Alvarez-Aldana!
Resumen: Helicobacter pylori es un bacilo móvil Gram negativo cuya morfología es espiral. Se
considera catalasa, oxidasa y ureasa positiva. Es una bacteria de distribución mundial cuya prevalencia
varía de acuerdo a las características epidemiológicas de la población. Las rutas de transmisión
descritas son: oral-oral, gastro-oral y fecal-oral; aunque también se ha descrito su transmisión a través
de alimentos, animales y agua. Para la realización del siguiente artículo de revisión se realizó una
búsqueda en las bases de datos PubMed y Science Direct utilizando los términos Helicobacter pylori,
water, detection methods, VBNC y cocoid form. Obteniendo como resultado que existe una alta
prevalencia de esta bacteria en países en vía de desarrollo que cuentan con condiciones deficientes de
saneamiento, por lo que el agua se convierte en uno de los canales por el cual viaja la bacteria, se
mantiene con vida, llega al estómago del ser humano y lo coloniza. H. pylori sobrevive en agua gracias
a varios mecanismos como es su cambio a forma cocoide (VBNC), su capacidad para formar biofilms
y de asociarse con amebas de vida libre. Debido a su presencia en forma viable pero no cultivable es
difícil de detectar en las muestras de agua, por lo que es necesaria la utilización de métodos moleculares
como PCR, FISH y LAMP, sin embargo, estas técnicas deben mejorarse para poder detectar la
viabilidad de las células para que sean métodos 100% efectivos, lo cual beneficiará la salud pública.
Palabras clave: Helicobacter pylori, agua, detección, riesgo sanitario.
Abstract: Helicobacter pylori is a Gram negative mobile bacillus. Its morphology is spiral and is
considered catalase, oxidase and urease positive. It is a bacterium of worldwide distribution whose
prevalence varies according to geographical distribution, ethnicity, race, socioeconomic factors,
hygiene and sanitation. There are several distribution routes but the best known are oral-oral, gastro-
oral and fecal-oral; it has also been described that the bacteria can be transmitted through food, animals
and water. For the realization of the following review article, a search was made in the databases
PubMed, PubMed Central and Science Direct using the terms Helicobacter pylori, water, detection
methods, VBNC y cocoid form. Obtaining as a result that there is a high prevalence of this bacterium
in developing countries that have poor sanitation conditions, so that water becomes one of the channels
through which the bacteria travels, stays alive, arrives to the stomach of the human being and colonizes
it. H. pylori survives in water thanks to several mechanisms such as its change to coccoid form
(VBNC), its ability to form biofilms and to associate with free-living amoebas. Due to its presence in
a viable but not cultivable way, it is difficult to detect in water samples, so it is necessary to use
molecular methods such as PCR, FISH and LAMP, however, these techniques must be improved in
order to detect the viability of the cells to be 100% effective methods, which will benefit public health.
Key words: Helicobacter pylori, water, detection, health risk
*Estudiantes de séptimo semestre. Semillero Obvio-Microbio. Programa de Microbiología. Universidad
Libre Seccional Pereira !Docente investigador Programa de Microbiología. Universidad Libre Seccional Pereira

Introducción
Helicobacter pylori es un bacilo móvil Gram negativo, microaerófilo, su morfología típica
es en espiral de aproximadamente 3.5 x 0.5 µm. Bioquímicamente es ureasa1, oxidasa y catalasa
positivo; se encuentra colonizando el estómago de aproximadamente la mitad de la población
mundial y es la causa de gastritis, úlceras pépticas, cáncer gástrico y linfoma de células B2.H.
pylori fue la primera especie bacteriana probada para causar cáncer y está clasificado como un
carcinógeno del grupo I por el Agencia Internacional para la Investigación sobre el Cáncer
(IARC)3.
Es una bacteria de distribución mundial cuya prevalencia varía de acuerdo a la distribución
geográfica, etnia, raza, factores socioeconómicos, de higiene y saneamiento. La infección puede
presentarse a cualquier edad pero se presenta con mayor frecuencia en edades tempranas, donde
puede permanecer asintomática o hacerse sintomática en la adultez; no produce inmunidad de
memoria y suele darse la reinfección4, 5.
Las rutas de transmisión de H. pylori aún son poco conocidas pero las más aceptadas son
las vías oral-oral, gastro-oral y fecal-oral; aunque también se ha descrito que la bacteria puede ser
transmitida a través de alimentos, animales y agua6.
Hace algunos años pocas investigaciones implicaban el agua como vehículo de transmisión
de H. pylori, pero hoy en día hay suficiente evidencia de la presencia de esta bacteria en el agua a
nivel mundial, por lo que se puede implicar al agua tratada y no tratada como posible vehículo de
transmisión7.
La hipótesis de que el agua es una vía de transmisión de H. pylori está respaldada por dos
tipos de estudios; los epidemiológicos que han demostrado una mayor prevalencia de la infección
en países en vía de desarrollo que padecen problemas relacionados con la distribución sanitaria del
agua a la población y los estudios que han detectado o aislado H. pylori de diversas fuentes de
agua8, 9. Por este motivo, son varios los estudios que se han centrado en determinar el potencial del
agua como fuente de transmisión de H. pylori y su capacidad de producir la infección clínica10.
La presente revisión tiene como objetivo compilar la información actual con relación a la
presencia de Helicobacter pylori en aguas; las metodologías de detección y su cambio morfológico
de forma espiral a forma cocoide en el agua así como su patogenicidad al encontrarse en este
estado.
Metodología
Se realizó una búsqueda amplia y sistemática de la literatura en las bases de datos PubMed
y Science Direct utilizando los términos Helicobacter pylori, water, detection methods, VBNC y
cocoid form, para lo cual se empleó el conector booleano AND. La búsqueda arrojó como resultado
797 y 12.614 artículos respectivamente, de los cuales 136 y 3.360 correspondían a trabajos no
mayores a 5 años de publicación, el cual fue un criterio de aceptación, ya que se restringió la
bibliografía a artículos originales publicados en los últimos 5 años y algunos artículos de revisión.

2
Se seleccionaron artículos sobre los métodos de detección de H. pylori en agua, sobre el cambio
de forma espiral a forma cocoide (viable no cultivable) que sufre la bacteria en el agua y en general
sobre evidencias del agua como mecanismo de transmisión de la bacteria H. pylori.
Discusión
La primera asociación entre el agua y la infección por H. pylori fue sugerida en Perú en
1991 cuando se observó que los niños peruanos que consumían agua no tratada, tenían mayor
prevalencia de H. pylori9, 11 y aun hoy en día Perú tiene una prevalencia
Resultados
En la tabla 1 se describen algunos de los artículos consultados y un corto resumen de los
hallazgos.
Tabla 2. Breve resumen de algunos artículos relacionados con la transmisión de H. pylori a través
del agua

3
estimada del 45,5%; encontrándose en agua potable la mayor cantidad de la bacteria, equivalente
a 1.6 x 106 copias del genoma/L. Un estudio publicado en este año arrojó que el agua potable en
Lima está contaminada con H. pylori el 20.3% del tiempo, resultando positivas 49 de 241
muestras13.
Diferentes estudios han mostrado la prevalencia de esta bacteria en países en vía de
desarrollo, esto relacionado con las circunstancias socioeconómicas y la calidad de vida llevan a
la falta de acceso a agua potable de calidad, a un saneamiento adecuado, a medidas higiénicas
estrictas y a una buena atención en salud, permitiendo estas situaciones que el agua contaminada
sea uno de los canales por el cual viaja la bacteria, se mantiene con vida, llega al estómago del ser
humano y lo coloniza6, 13, 14.
En las poblaciones del Norte de Europa y América del Norte, alrededor de un tercio de los
adultos se encuentran infectados por la bacteria, mientras que en Europa del Sur y Este, América
del Sur y Asia, la prevalencia de H. pylori a menudo es superior al 50%, siendo estos últimos
países en desarrollo que aun cuentan con servicios de saneamiento deficientes15.
En este punto aparece otro posible factor detrás de las diferencias geográficas y económicas
en la incidencia de la infección y son las fuentes mejoradas que permiten el acceso al agua de estas
poblaciones, las cuales consisten en métodos artesanales que aplican las personas que no tienen
suministro continuo de agua, como lo son el agua entubada en el hogar, pozos de sondeo,
manantiales de arenas de cavado bien protegidos, agua de lluvia y fuentes de agua públicas o
externas, las cuales pueden llegar a ser fuentes directas de H. pylori como arroyos y aguas
superficiales contaminadas por contaminación fecal, industrial y doméstica16, 17, 18.
H. pylori ha sido detectada en varias fuentes de agua, incluyendo lagos, ríos, agua del grifo, agua
de pozo, aguas de riego, agua de mar, agua mineral, agua de unidades dentales, agua potable y sus
sistemas de distribución como lo han demostrado diferentes estudios realizados en Colombia, Perú,
Estados Unidos, España, Costa Rica, Irán, Pakistán, entre otros19.
Aunque algunos estudios han encontrado que el agua clorada carece de la bacteria, se dice
que arrojaron esta conclusión debido a que se basaron en la falta de recuperación utilizando
métodos de cultivo estándar, ya que muchos otros autores han demostrado que H. pylori es capaz
de resistir y tolerar las prácticas de desinfección normalmente empleadas en el tratamiento del
agua potable20, 21. Se dice que el cambio de morfología permite a las células evadir el tratamiento
con cloro del agua utilizada para el consumo humano22.
En un estudio realizado en España se expuso el patógeno a una concentración de 0.96 mg/L
de cloro libre durante varios tiempos control, observando que H. pylori perdía su capacidad de
crecimiento sobre medios de cultivo pasados 5 minutos. Sin embargo, mediante las técnicas
moleculares FISH y PCR observó que un porcentaje elevado de las células pasaban a forma viable
no cultivable, demostrando así que la bacteria puede sobrevivir a concentraciones elevadas de
cloro libre y que los tratamientos de desinfección en aguas potables podrían ser ineficientes para
la eliminación de éste patógeno de las redes de distribución23.

4
Aunque varios estudios epidemiológicos proponen el agua como un intermediario en la
transmisión fecal-oral de H. pylori20; se ha encontrado que la detección de esta bacteria en muestras
de agua no siempre se correlaciona con la presencia de bacterias fecales indicadoras (E. coli o
Enterococcus). Es más, se ha demostrado que H. pylori es más resistente a la cloración que E. coli.
Lo cual puede constituir un grave problema de salud pública, ya que a menudo se intenta predecir
la seguridad de los cuerpos de agua en función de los niveles de bacterias indicadoras fecales
cuando el agua tratada no está necesariamente exenta de H. pylori24, 25.
Aparte de ser resistente al cloro, H. pylori es resistente al ozono19 y a la radiación UV como
tratamiento terciario, hecho que pudo deberse al alto caudal de agua tratado en el estudio, el cual
no permitió una penetración total de la radiación ultravioleta durante un tiempo suficiente, o bien
que el sistema de radiación UV no se encontraba en buenas condiciones de mantenimiento23.
Se han descrito algunos mecanismos que le permiten sobrevivir a Helicobacter pylori en
un ambiente adverso para ella como es el agua. Entre ellos encontramos la conversión a su forma
cocoide (forma viable no cultivable) (Fig 1), su capacidad para adherirse a diferentes materiales y
agregarse junto con otras bacterias para forma estructuras complejas en tuberías u otras superficies
en contacto con agua (biopelículas) y también para sobrevivir dentro de amebas de vida libre
(FLA).
Fig 1. Cambios morfológicos en Helicobacter pylori. Forma espiral (flecha negra larga), forma V (flecha
negra corta), forma U (flecha blanca corta) y forma cocoide o viable no cultivable (flecha blanca larga) 26.
Forma viable no cultivable - VBNC
Muchas especies de bacterias entran en el estado viable no cultivable (VBNC) cuando están
expuestas a condiciones estresantes como inanición, bajas temperaturas y en general condiciones
ambientales desfavorables, siendo esta una estrategia de adaptación para la supervivencia a largo
plazo. A diferencia de las células normales que son cultivables en medios adecuados y se
convierten en colonias, las células VBNC son células vivas que pierden su capacidad de crecer en
medios de cultivo en los que normalmente crecen. A pesar de esto, las células VBNC no se
consideran células muertas debido a que tienen una membrana intacta que contiene la información
genética en perfecto estado, además de ser metabólicamente activas27.

5
La capacidad de ingresar al estado VBNC puede ser ventajosa para la bacteria, pero
representa un riesgo para la salud humana ya que bajo los métodos convencionales de cultivo no
es posible su detección, por lo que si en una muestra todas o algunas bacterias se encuentran en
estado VBNC, la cantidad total de bacterias viables será subestimada o no se detectará28 y
tratándose de un patógeno humano, podrá infectar a la persona que consuma el agua en la cual no
pudo ser detectado.
Se ha demostrado la viabilidad y la capacidad de cultivo de H. pylori en agua hasta por 75
horas a 10 °C. Así mismo se determinó que el total de células no disminuía por períodos mucho
más largos de tiempo en condiciones adecuadas (2 años a 4 °C) 29.
Varios estudios han señalado que los cambios en la morfología de H. pylori están
estrechamente asociada a los genes implicados en la síntesis de peptidoglucano. En primer lugar,
ocurre la división de los enlaces peptídicos por acción de Csd1, Csd2 y Csd3. Luego, los enlaces
intrapéptidos también se escinden por acción secuencial de Csd3, de una DL-carboxipeptidasa
desconocida y Csd4, que permite una relajación inicial del peptidoglicano en la zona de torsión,
perdiendo la forma helicoidal. A continuación, los monómeros que han sido sometidos a la acción
enzimática, son reconocidos por diferentes transglicosilasas líticas, como Slt y MltD, que se
eliminan para la posterior inserción de nuevos monómeros. Finalmente, la inserción de nuevos
monómeros en el peptidoglicano y la elongación, se realiza mediante PBP1, PBP2 y proteínas de
membrana (MreC). Esto aumenta la longitud de las cadenas de glucanos y los enlaces cruzados30.
La forma espiral se ha asociado con la capacidad de moverse en un medio viscoso, como
el moco gástrico. Esto significa que la forma helicoidal puede ser innecesaria en ambientes
acuáticos y también puede implicar un costo energético innecesario para la bacteria30, siendo este
otro posible motivo que justifique el cambio de forma.
Por mucho tiempo se consideró que muchos de los problemas que se han encontrado en la
recuperación de formas viables de H. pylori en el agua, se debe a que estas pasan a formas cocoides
o VBNC. Desde un punto de vista más amplio, ahora se acepta que lo que ocurre es que estas
bacterias no se logran cultivar bajo métodos rutinarios y convencionales de laboratorio29 sino que
son necesarios métodos de detección molecular.
Formación de Biopelículas
La formación de biopelículas en el ambiente ha sido propuesto como una estrategia de H.
pylori para sobrevivir en los sistemas de distribución de agua22.
Las biopelículas son comunidades microbianas embebidas dentro de una matriz orgánica
polimérica autoproducida y adherida a una superficie viva o inerte; son sólidas y presentan canales
por donde fluyen el agua y los nutrientes a las zonas más profundas. Los primeros análisis de la
producción de biofilm por parte de bacterias, como H. pylori, se llevaron, mediante estudios de
microscopía electrónica, observando formas espirales, cocoides y degenerativas. La formación de
la biopelícula conlleva un proceso de cinco fases: 1) Adsorción reversible de la bacteria a la
superficie; 2) Unión irreversible a la superficie; 3) Fase inicial de maduración con crecimiento y

6
desarrollo microbiano; 4) Producción del exopolímero y, 5) Desarrollo final de la colonia con
dispersión de células colonizadoras31.
Cuando H. pylori se adhiere a una superficie, comienza la activación del quorum sensing
así como los mecanismos de formación de los biofilms. Se ha trabajado con biofilms en sistemas
de tuberías y tanques de almacenamiento y algunos autores creen que así es como H. pylori puede
ser viable en el agua25 debido a que al ubicarse en la parte interna de estos tanques y tuberías, las
hace bastante inaccesibles y difícil de alcanzar por los desinfectantes32.
En Colombia se evaluó la presencia de H. pylori en muestras de agua y biopelículas de los
grifos de instituciones educativas oficiales de la ciudad de Medellín a través de una técnica de
reacción en cadena de la polimerasa (PCR) directa convencional, identificando la bacteria por
cultivo en 11.2% y PCR en 2.1% de las muestras7. En este caso, el cultivo tuvo mejor rendimiento
que la PCR probablemente porque los componentes de las biopelículas actuaron como inhibidores
de la PCR11, por los genes utilizados para la amplificación o por la técnica de procesamiento de
muestras7.
La capacidad de H. pylori para crecer y formar biopelículas in vitro e in vivo podría ser
una ventaja para la especie para evitar lesiones causadas por factores estresantes químicos, como
la terapia antimicrobiana in vivo, o el estrés inducido por la privación de nutrientes en el medio
ambiente22.
La presencia de las biopelículas de H. pylori en agua se ha verificado mediante PCR,
sondas de ADN e hibridación fluorescente in situ (FISH). Así mismo, se ha presentado evidencia
in vitro que sugiere que las interacciones con bacterias específicas (por ejemplo, Mycobacterium
chelonae) dentro de una biopelícula de múltiples especies puede contribuir a la viabilidad de H.
pylori33.
Es necesario tener en cuenta que aunque una población disponga de aguas municipales
potables y de grifos adecuados, la calidad del agua de la potabilización no es la misma en todas
partes, además el estado del acueducto como daños previos que exponen al agua potable a
contaminación con materia fecal, la antigüedad del acueducto y la presencia de biopelículas en las
tuberías, deben estudiarse como factores predictores de contaminación del agua7.
Asociación con amebas de vida libre (FLA)
Se ha demostrado previamente que las amebas de vida libre (FLA), como Acanthamoeba,
Naegleria, Vermamoeba o Balamuthia, pueden actuar como huéspedes de algunos patógenos
bacterianos capaces de resistir la digestión amebiana (bacterias resistentes a amebas, ARB). Dentro
de ellas, las bacterias sobreviven y son más resistentes a las condiciones ambientales que
normalmente las matarían. Por lo tanto, FLA actúa como "caballo de Troya" para ARB,
permitiendo la supervivencia bacteriana y/o la transmisión a susceptibles hospedadores34, 35.
Recientemente, un estudio in vitro ha indicado que H. pylori puede sobrevivir dentro de
Acanthamoeba castellanii después de resistir un tratamiento de desinfección por cloración. FLA
proporciona refugio para H. pylori, lo que le permite sobrevivir bajo diferentes tratamientos y

7
hacer posible su transmisión a humanos. El análisis de qPCR de las muestras mostró la presencia
de ADN de H. pylori intracelular en 28 (50,9%) de las muestras de aguas residuales y en 11
(91,7%) de las muestras de agua potable. Así mismo, se indicó que no se pudieron recuperar
colonias por cultivo de las muestras de FLA positivas, debido a que la bacteria podría haber
adquirido su estado VBNC, ya que mediante DVC-FISH se observó su viabilidad después del
tratamiento con hipoclorito34, 36.
Virulencia de las formas cocoides de H. pylori
Para determinar si H. pylori representaba un riesgo significativo para la salud pública, era
necesario realizar una evaluación cuantitativa del riesgo microbiano (QMRA), la cual fue realizada
por Ryan et al., quienes sugirieron un nivel máximo del contaminante en agua potable de un
organismo por litro (<1 organismo/L). Igualmente se determinó la mediana de H. pylori en agua
potable para el rango anual de riesgo de infección en 6.84 x 10-02/L para humanos37.
Históricamente el cambio en la morfología (de bacilo espiral a cocoide) ha planteado dudas
sobre si H. pylori es realmente viable e infeccioso en el agua, sin embargo se han realizado pocos
estudios al respecto en humanos, pero si se ha demostrado la viabilidad infecciosa de esta forma
de H. pylori en ratones, tanto a través de agua como de sonda nasogástrica38.
En el estudio realizado por Boehnke, se suministraron a ratones SS1 concentraciones
estáticas variables de H. pylori en agua de: 1.29 x 105, 106, 107, 108 y 109 UFC/L, estableciendo
como concentración mínima infecciosa de H. pylori en agua para esta cepa de ratones 106 UFC/L.
Además, se sugirió que la dosis infecciosa en humanos puede ser menor que para los ratones
porque H. pylori no es un habitante normal del ratón39, coincidiendo con los resultados de la
QMRA mencionada previamente y con otros estudios que establecieron la dosis infectante
estimada para el ser humano en 104 - 1010 UFC/L 11, 38.
Por otra parte, Guimarães et al, realizaron un estudio que pretendía evaluar el efecto de la
exposición al agua en varios mecanismos de H. pylori que inducen una respuesta en las células del
huésped, para lo cual se utilizó la línea celular epitelial gástrica humana AGS y se obtuvieron
importantes resultados, como que después de 24h de exposición al agua H. pylori pierde su
capacidad de cultivo (Fig 2) al cambiar su morfología a forma cocoide la cual es viable pero no
cultivable; así como la capacidad de inducir la producción de IL-8 por las células AGS; su adhesión
a las células se ve disminuida en un 40% y se limita la influencia de la bacteria sobre la apoptosis
de la célula huésped40. Sin embargo, es importante aclarar que en otros estudios, la bacteria ha
perdido su capacidad de cultivo a las 48 h e incluso a las 72 h30.
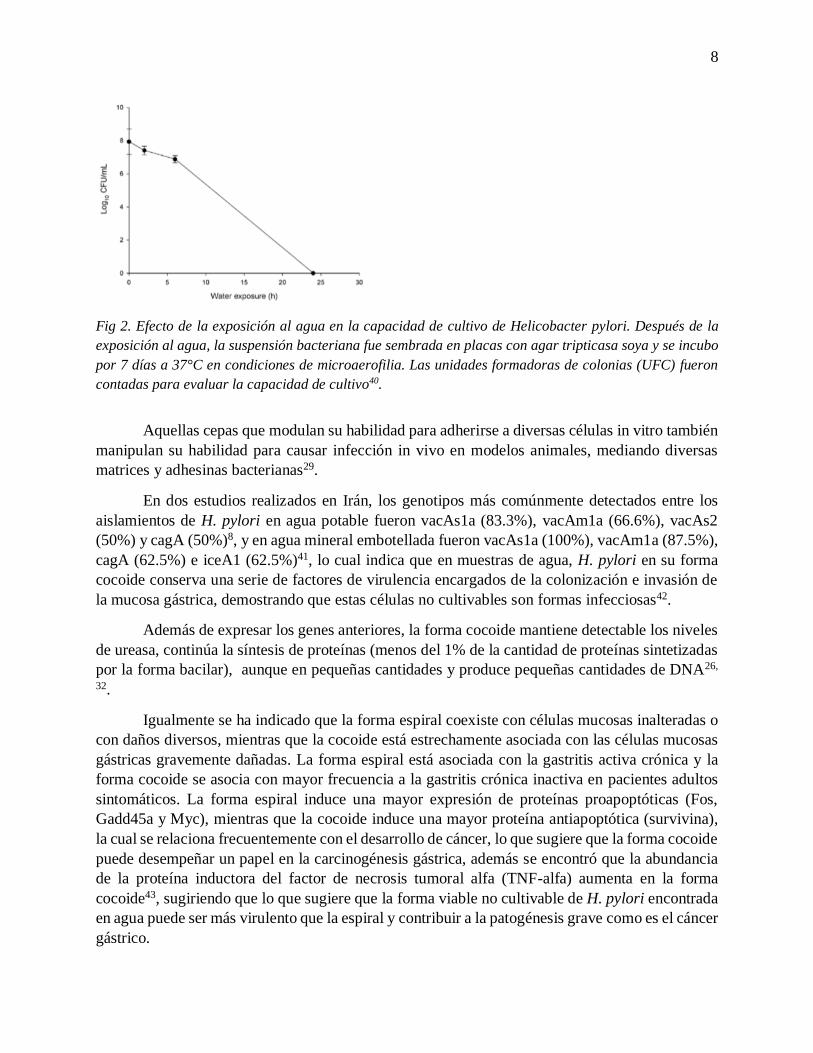
8
Fig 2. Efecto de la exposición al agua en la capacidad de cultivo de Helicobacter pylori. Después de la
exposición al agua, la suspensión bacteriana fue sembrada en placas con agar tripticasa soya y se incubo
por 7 días a 37°C en condiciones de microaerofilia. Las unidades formadoras de colonias (UFC) fueron
contadas para evaluar la capacidad de cultivo40.
Aquellas cepas que modulan su habilidad para adherirse a diversas células in vitro también
manipulan su habilidad para causar infección in vivo en modelos animales, mediando diversas
matrices y adhesinas bacterianas29.
En dos estudios realizados en Irán, los genotipos más comúnmente detectados entre los
aislamientos de H. pylori en agua potable fueron vacAs1a (83.3%), vacAm1a (66.6%), vacAs2
(50%) y cagA (50%)8, y en agua mineral embotellada fueron vacAs1a (100%), vacAm1a (87.5%),
cagA (62.5%) e iceA1 (62.5%)41, lo cual indica que en muestras de agua, H. pylori en su forma
cocoide conserva una serie de factores de virulencia encargados de la colonización e invasión de
la mucosa gástrica, demostrando que estas células no cultivables son formas infecciosas42.
Además de expresar los genes anteriores, la forma cocoide mantiene detectable los niveles
de ureasa, continúa la síntesis de proteínas (menos del 1% de la cantidad de proteínas sintetizadas
por la forma bacilar), aunque en pequeñas cantidades y produce pequeñas cantidades de DNA26,
32.
Igualmente se ha indicado que la forma espiral coexiste con células mucosas inalteradas o
con daños diversos, mientras que la cocoide está estrechamente asociada con las células mucosas
gástricas gravemente dañadas. La forma espiral está asociada con la gastritis activa crónica y la
forma cocoide se asocia con mayor frecuencia a la gastritis crónica inactiva en pacientes adultos
sintomáticos. La forma espiral induce una mayor expresión de proteínas proapoptóticas (Fos,
Gadd45a y Myc), mientras que la cocoide induce una mayor proteína antiapoptótica (survivina),
la cual se relaciona frecuentemente con el desarrollo de cáncer, lo que sugiere que la forma cocoide
puede desempeñar un papel en la carcinogénesis gástrica, además se encontró que la abundancia
de la proteína inductora del factor de necrosis tumoral alfa (TNF-alfa) aumenta en la forma
cocoide43, sugiriendo que lo que sugiere que la forma viable no cultivable de H. pylori encontrada
en agua puede ser más virulento que la espiral y contribuir a la patogénesis grave como es el cáncer
gástrico.

9
En el tejido gástrico, las formas cocoides (VBNC) pueden permanecer latentes durante
mucho tiempo y retener los factores de virulencia, por lo que también pueden ayudar a los fracasos
del tratamiento y la recurrencia de la infección y de las enfermedades gastroduodenales26. La
recurrencia de la infección es posible debido a la ocurrencia de recombinación genética con otras
cepas de H. pylori que podrían estar presentes en el mismo huésped, lo que da como resultado una
mayor diversificación genética la cual puede ayudar a la bacteria a adaptarse a un nuevo huésped
después de la transmisión40.
Así como existen múltiples estudios que demuestran la presencia y la capacidad infectiva
de H. pylori en agua; en otro estudio realizado por Boehnke et al con la cepa de ratones SS1 y H.
pylori, ninguno de ellos mostró signos de infección, lo cual puede ser explicado debido a que la
variabilidad genética de las cepas de H. pylori es amplia, por lo que es posible que algunas cepas
carezcan de la capacidad de persistir en el agua y por el contrario se transmitan únicamente a través
de otras rutas, como las vías de persona a persona o vía fecal-oral44.
Métodos de detección
Uno de los principales problemas para aceptar el agua como posible reservorio de H. pylori
es la incapacidad de aislar rutinariamente la especie de las muestras de agua mediante técnicas
convencionales de cultivo microbiológico. Por lo tanto, la aplicación de métodos moleculares para
la detección rápida, sensible y específica de H. pylori en entornos acuáticos es de suma
importancia.
Usualmente cuando se logra cultivar H. pylori a partir de muestras ambientales como el
agua, no es posible aislarse completamente debido al crecimiento de la microbiota competitiva en
un agar selectivo, demostrando que los medios de cultivo actualmente disponible no son lo
necesariamente útiles para el aislamiento de las bacterias del medio ambiente en especial H. pylori.
Cuando muestras de la presunta colonias se analizan, células Gram-negativas exhiben una
morfología típica de la bacteria pero se mezclan con otros bacilares que no son H. pylori45. Para
favorecer la eliminación de esta microbiota competitiva se han desarrollado técnicas alternativas
como son la separación inmunomagnética (IMS) y el tratamiento ácido23.
El principal desafío cuando se realiza un muestreo ambiental es la incapacidad de demostrar
la existencia de H. pylori viable en el agua, que potencialmente puede colonizar el estómago o el
duodeno porque los métodos moleculares utilizados para detectar las bacterias en el medio
ambiente son incapaces de distinguir entre bacterias muertas; sin embargo, han surgido técnicas
como la DVC-FISH (Hibridación fluorescente in situ en combinación con la incubación de conteo
viable directo) que permite la detección específica de células viables y tiene la ventaja de no ser
inactivado por inhibidores de muestras, así como la implementación del agente intercalante PMA
a la técnica qPCR10. Esta última sigue siendo un desafío, ya que solo demuestra la integridad de la
membrana y puede conducir a una sobreestimación de la población de bacterias viables en algunas
condiciones de inactivación42.
La prueba de qPCR para H. pylori presenta por si sola varias ventajas: no se basa en el
cultivo; muchas muestras pueden analizarse rápidamente (hasta 384 muestras en 2 h); se eliminan

10
muchas de las fuentes de error humano del proceso de análisis y disminuye el potencial de
contaminación46. Sin embargo, debe tenerse en cuenta que la mayoría de los resultados positivos
de qPCR podrían deberse al ADN de células no viables, por lo que este método no es adecuado
para determinar el potencial infeccioso de una muestra10.
Por su parte, la PCR convencional es el método más utilizado hasta el momento, siendo
más sensible y específico, pero tiene algunas limitaciones, como numerosos ciclos térmicos,
costoso termociclador y un método de identificación y manifestación del producto lento y difícil2.
Cuando se utiliza la PCR como método de detección, es necesario realizar un tratamiento
previo o enriquecimiento que aumente la sensibilidad de la prueba mejorando así las tasas de
detección debido a que existen inhibidores de la enzima Taq polimerasa presentes en el agua (por
ejemplo, en las aguas residuales los ácidos húmicos) que pueden producir resultados falsos
negativos45.
La viabilidad de H. pylori en muestras de agua puede evaluarse mediante PCR de
transcripción inversa (RT-PCR), y ensayos de anticuerpos fluorescentes indirectos (IFA). La
prueba IFA utiliza anticuerpos específicos que permiten la enumeración de células tanto
cultivables como no cultivables y las formas espiral y cocoide, pero no puede detectar genes
marcadores de virulencia como cagA lo cual no lo hace muy útil al momento de querer confirmar
la presencia de H. pylori a través de estos genes de virulencia47.
Además de las anteriores técnicas, existen otras como la amplificación isotérmica medida
por bucle (LAMP por sus siglas en inglés) que es un mecanismo más rápido y fácil. Esta técnica
es una reacción de amplificación de una etapa, que puede producir una gran cantidad de copias en
menos de una hora en condiciones isotermas. La ventaja más importante de este método es que no
necesita desnaturalización del ADN2. Además, se puede realizar sin un termociclador y los
resultados pueden observarse a simple vista46. También se utiliza la citometría de flujo, la cual es
una técnica analítica que tiene el potencial de hacer una distinción entre los cuatro estados
fisiológicos de las bacterias: reproductivamente viable, metabólicamente activa, intacta y
permeabilizada. Puede determinar las proporciones de los estados VBNC y VC y las células
muertas, en función de la integridad de la membrana de la bacteria46.
Es importante resaltar que la transmisión por agua también ha sido sugerida para otras
especies de Helicobacter como H. mustelae, H. muridarum, H. felis, H. canadensis, H. pullorum,
H. canis y H. cetorum. Utilizando para su detección la técnica PCR rRNA 16S y 23S, descubriendo
que el uso de 16S era más específico para los resultados esperados48.
Para la detección molecular se han utilizado diversos genes como ureC32 que se ha
demostrado que es único y esencial para el crecimiento de H. pylori21 rRNA 16S (cebadores Hp1,
Hp2 y Hp3)49, glmM25, ureA, cagA, vacA19, iceA, oipA, cagE and baba241.
A medida que los instrumentos para la secuenciación de ADN se vuelven más extendidos
y convenientemente portátiles, y mientras que el costo de la secuenciación continue disminuyendo,
la metagenómica probablemente se convertirá en la tecnología principal utilizada en microbiología

11
ambiental y ecología microbiana para la detección de H. pylori y otras bacterias en muestras
ambientales46. No obstante, por el momento sigue siendo necesario adaptar algunos los métodos
moleculares para detectar la viabilidad de H. pylori en los suministros de agua50.
Referencias
1. Matsumoto H, Shiotani A, Nishibayashi H, KamadaT, Kimura T, Fujimura Y, et al.
Molecular Detection of H. pylori Using Adherent Gastric Mucous to Biopsy Forceps.
Helicobacter [Internet]. 2016 Dec [cited 2018 May 23]; 21(6): 548-553. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/27061611
2. Chamanrokh P, Shahhosseiny MH, Assadi MM, Nejadsattari T, Esmaili D. Three Tests Used
to Identify Non-Culturable Form of Helicobacter pylori in Water Sample. Jundishapur J
Microbiol [Internet]. 2015 April [cited 2018 May 23]; 8(4): e16811. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4449853/
3. Testerman TL, Morris J. Beyond the stomach: An updated view of Helicobacter pylori
pathogenesis, diagnosis, and treatment. World J Gastroenterol [Internet]. 2014 Sep. [cited
2018 May 26]; 20(36): 12781–12808. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4177463/
4. De Pardo Ghetti EM. Helicobacter Pylori: un problema actual. Gac Med Bol [Internet]. 2013
Dic [citado 2018 May 31]; 36(2): 108-111. Disponible en:
http://www.scielo.org.bo/scielo.php?script=sci_arttext&pid=S1012-
29662013000200013&lng=es
5. Suárez Rivera JJ, Almaguer Betancourt YM, Martínez Garrido R. Comportamiento
higiénico-sanitario de pacientes con diagnóstico de úlcera gastroduodenal por Helicobacter
pylori. Rev Cubana Med Gen Integr [Internet]. 2013 Dic [cited 2018 Jun 01]; 29(3): 328-
335. Disponible en: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-
21252013000400006&lng=es
6. Breckan RK, Paulssen EJ, Asfeldt AM, Kvamme JM, Straume B, Florholmen J. The All-
Age Prevalence of Helicobacter pylori Infection and Potential Transmission Routes. A
Population-Based Study. Helicobacter [Internet]. 2016 Dec [cited 2018 May 23]; 21(6): 586-
595. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/27172105
7. Cuervo CM, Gaviria AM. Detección de Helicobacter pylori en muestras de agua y
biopelícula de los grifos de las instituciones educativas oficiales en la ciudad de Medellín.
Acta Med Colomb [Internet]. 2017 June [cited 2018 Apr 28]; 42(2): 121-128. Disponible en:
http://www.scielo.org.co/pdf/amc/v42n2/0120-2448-amc-42-02-00121.pdf
8. Ranjbar R, Khamesipour F, Jonaidi-Jafari N, Rahimi E. Helicobacter pylori isolated from
Iranian drinking water: vacA, cagA, iceA, oipA and babA2 genotype status and
antimicrobial resistance properties. FEBS Open Bio [Internet]. 2016 Apr [cited 2018 May
23]; 6(5): 433-41. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/27419049.

12
9. Aziz RK. Khalifa MM, Sharaf RR. Contaminated water as a source of Helicobacter pylori
infection: A review. J Adv Res [Internet]. 2015 Jul. [cited 2018 May 31]; 6(4), 539–547.
Disponible en: https://www.sciencedirect.com/science/article/pii/S2090123213000970
10. Santiago P, Moreno Y, Ferrús MA. Identification of Viable Helicobacter pylori in Drinking
Water Supplies by Cultural and Molecular Techniques. Helicobacter [Internet]. 2015 Aug
[cited 2018 May 23]; 20(4): 252-9. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/25655472
11. Otero R W. Helicobacter pylori en agua potable ¿Es la ruta de la infección? Acta Med
Colomb [Internet]. 2017 June [cited 2018 Apr 28]; 42(2): 87-89. Disponible en:
http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-
24482017000200087&lng=en
12. Boehnke KF, Brewster RK, Sánchez BN, Valdivieso M, Bussalleu A, Guevara M. et al. An
assessment of drinking water contamination with Helicobacter pylori in Lima, Peru.
Helicobacter [Internet]. 2018 Apr [cited 2018 May 27]; 23(2): e12462. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/29316052
13. Duarte Pastor LD. Análisis de la problemática de salud ambiental provocada por
Helicobacter pylori presente en fuentes hídricas contaminadas. [Tesis de Especialización].
Bogotá: Universidad Militar Nueva Granada; 2017. Disponible en:
https://repository.unimilitar.edu.co/bitstream/10654/16565/3/DuartePastorDanielaLizbeth2
017.pdf
14. Mentis A, Lehours P, Mégraud F. Epidemiology and Diagnosis of Helicobacter pylori
infection. Helicobacter [Internet]. 2015 Sep [cited 2018 may 29]; 20(1): 1-7. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/26372818
15. Eusebi LH, Zagari RM. Bazzoli F. Epidemiology of Helicobacter pylori Infection.
Helicobacter [Internet]. 2014 Sep. [cited 2018 May 25]; 19(1): 1-5. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/25167938
16. Eichelberger L, Murphy G, Etemadi A, Abnet CC, Islami F, Shakeri R, et al. Risk of Gastric
Cancer by Water Source: Evidence from the Golestan Case-Control Study. PLoS One
[Internet]. 2015 May [cited 2018 May 26]; 10(5): e0128491. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/26023788
17. Porras C, Nodora J, Sexton R, Ferreccio C, Jimenez S, Dominguez RL, et al. Epidemiology
of Helicobacter pylori infection in six Latin American countries (SWOG Trial S0701).
Cancer Causes Control [Internet]. 2013 Feb. [cited 2018 May 29]; 24(2): 209–15. Disponible
en: https://www.ncbi.nlm.nih.gov/pubmed/23263777
18. Baingana RK, Kiboko Enyaru J, Davidsson L. Helicobacter pylori infection in pregnant
women in four districts of Uganda: role of geographic location, education and water sources.
BMC Public Health [Internet]. 2014 Sep [cited 2018 May 27]; 14: 915. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/25190150
19. Castillo Canón AP. Estandarización de la técnica de PCR en tiempo real para la detección
de Helicobacter pylori en aguas de consumo. [Tesis de Maestría]. Bogotá: Universidad

13
Nacional de Colombia; 2014. Disponible en:
http://bdigital.unal.edu.co/46636/1/05599317.2014.pdf
20. Bayona Rojas MA, Gutiérrez Escobar AJ. Helicobacter Pylori: Vías de transmisión.
Medicina [Internet]. 2017 Sep [cited 2018 May 31]; 39(3), 210-220. Disponible en:
http://revistamedicina.net/ojsanm/index.php/Medicina/article/view/118-4.
21. El-Sharouny E, El-Shazli H, Olama Z. Detection of Helicobacter pylori DNA in Some
Egyptian Water Systems and Its Incidence of Transmission to Individuals. Iran J Public
Health [Internet]. 2015 Feb [cited 2018 May 26]; 44(2): 203–210. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4401878/
22. García A, Salas Jara MJ, Herrera C, González C. Biofilm and Helicobacter pylori: From
environment to human host. World J Gastroenterol [Internet]. 2014 may [cited 2018 may
28]; 20(19): 5632-5638. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4024771/
23. Cuellar P. Transmisión de Helicobacter pylori a través del agua: estudio de la presencia del
patógeno e identificación de formas viables mediante técnicas moleculares. [Tesis Doctoral].
Valencia: Universidad Politécnica de Valencia; 2016. Available from:
https://riunet.upv.es/bitstream/handle/10251/75086/SANTIAGO%20-
%20TRANSMISI%C3%93N%20DE%20HELICOBACTER%20PYLORI%20A%20TRA
V%C3%89S%20DEL%20AGUA%3A%20ESTUDIO%20DE%20LA%20PRESENCIA%
20DEL%20PAT%C3%93....pdf?sequence=1
24. Holman CB, Bachoon DS, Otero E , Ramsubhag A. Detection of Helicobacter pylori in the
coastal waters of Georgia, Puerto Rico and Trinidad. Mar Pollut Bull [Internet] 2014 Feb
[cited 2018 may 29]; 79(1-2): 354-8. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/24332757
25. Montero Campos V, Hernández Soto A, Camacho Sandoval J. Culture and Molecular
Identification of Helicobacter pylori in Drinking Water from Areas of High and Low
Incidence of Gastric Cancer in Costa Rica. Open Journal of Medical Microbiology [Internet].
2014 Dec [cited 2018 May 28]; 4(4): 261-269. Disponible en:
https://www.scirp.org/journal/PaperInformation.aspx?PaperID=52896
26. Sarema M, Corti R. Role of Helicobacter pylori coccoid forms in infection and
recrudescence. Gastroenterol Hepatol [Internet]. 2016 Jan [cited 2018 May 26]; 39(1): 28-
35. Disponible en: https://www.sciencedirect.com/science/article/pii/S2444382415000073
27. Laam L, Mendis N, Trigui H, Oliver JD, Faucher SP. The importance of the viable but non-
culturable state in human bacterial pathogens. Front Microbiol [Internet]. 2014 Jun. [cited
2018 May 31]; 5: 258. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040921/
28. Mali S, Vrushi L, Hili N. The impact of drinking water on the prevalence of H. pylori in
humans in Elbasan, Albania. Eur Water [Internet]. 2017 Jun. [cite 2018 May 31]; 58: 263-
266. Disponible en:

14
https://www.researchgate.net/publication/321184977_The_impact_of_drinking_water_on_
the_prevalence_of_H_Pylori_in_humans_in_Elbasan_Albania
29. Monsalve LA. Prevalencia de Helicobacter pylori en el agua de consumo humano de
pacientes diagnosticados con cáncer gástrico Helicobacter pylori positivo en el Instituto
Nacional de Enfermedades Neoplásicas 2015 – 2016. [Tesis de Pregrado]. Lima:
Universidad Nacional Mayor de San Marcos; 2017. Disponible en:
http://cybertesis.unmsm.edu.pe/handle/cybertesis/7195
30. Fernandes RM, Silva H, Oliveira R, Almeida C, Azevedo NF, Vieira MJ. Morphological
transition of Helicobacter pylori adapted to water. Future Microbiol [Internet]. 2017 Oct
[cited 2018 May 24]; 12:1167-1179. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/28879777
31. Bayona Rojas MA, Gutiérrez Escobar AJ. Biopelícula: un mecanismo de supervivencia de
Helicobacter pylori. Rev U.D.C.A Act & Div Cient [Internet]. 2013 Dec [cited 2018 May
31]; 16(2): 335-342. Disponible en:
http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0123-
42262013000200007&lng
32. Bahrami A. Rahimi E. Detection of Helicobacter pylori in City Water, Dental Units Water,
and Bottled Mineral Water in Isfahan, Iran. Scientific World Journal [Internet]. 2013 Mar.
[cited 2018 May 29]; 280510. Disponible
en: https://www.ncbi.nlm.nih.gov/pubmed/23606812
33. Hathroubi S, Servetas SL, Windham I, Merrell DS, Ottemann KM. Helicobacter pylori
Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol Mol Biol Rev
[Internet]. 2018 May. [citado 2018 May 31]; 82(2) pii: e00001-18. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/29743338.
34. Moreno-Mesonero L, Moreno Y, Alonso JL, Ferrús MA. Detection of viable Helicobacter
pylori inside free-living amoebae in wastewater and drinking water samples from Eastern
Spain. Environ Microbiol [Internet]. 2017 Oct [cited 2018 May 28]; 19(10): 4103-4112.
Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/28707344
35. Amirhooshang A, Ramin A, Ehsan A, Mansour R, Shahram B. High frequency of
Helicobacter pylori DNA in drinking water in Kermanshah, Iran, during June–November
2012. J Water Health. [Internet]. 2014 Sep. [citado 2018 May 31]; 12(3): 504-512.
Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/25252354
36. Bai X, Xi C, Wu J. Survival of Helicobacter pylori in the wastewater treatment process and
the receiving river in Michigan, USA. J Water Health [Internet]. 2016 Aug [cited 2018 May
28]; 14(4): 692-8. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/27441864
37. Ryan M, Hamilton K, Hamilton M, Haas CN. Evaluating the Potential for a Helicobacter
pylori Drinking Water Guideline. Risk Anal [Internet]. 2014 Sep [cited 2018 May 27];
34(9): 1651-62. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/24660760
38. Valdivieso M, Bussalleu A, Sexton R, Boehnke K, Osorio S, Novoa I, et al. Clinical,
Epidemiologic, and Genomic Studies (SWOG S1119) of Helicobacter Pylori in Lima, Peru:
https://www.ncbi.nlm.nih.gov/pubmed/?term=Oliveira%20R%5BAuthor%5D&cauthor=true&cauthor_uid=28879777
https://www.ncbi.nlm.nih.gov/pubmed/?term=Azevedo%20NF%5BAuthor%5D&cauthor=true&cauthor_uid=28879777
https://www.ncbi.nlm.nih.gov/pubmed/?term=Hamilton%20K%5BAuthor%5D&cauthor=true&cauthor_uid=24660760

15
Role of Contaminated Water. J Cancerol [internet]. 2016 Feb. [cited 2018 May 31]; 3: 52-
63. Disponible en: http://www.journalofcancerology.com/pdf/JCancer2016_02_52-63.pdf
39. Boehnke KF, Eaton KA, Valdivieso M, Baker LH, Xi C. Animal Model Reveals Potential
Waterborne Transmission of Helicobacter pylori Infection. Helicobacter [Internet]. 2015 Oct
[cited 2018 May 26]; 20(5): 326-33. Disponible
en: https://www.ncbi.nlm.nih.gov/pubmed/25664781
40. Guimarães Nuno M, Azevedo Nuno F, Vieira Maria J, Figueiredo Ceu. Water-induced
modulation of Helicobacter pylori virulence properties. Mem. Inst. Oswaldo Cruz [Internet].
2014 July [cited 2018 Abr 30]; 109(4): 414-419. Disponible en:
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0074-
02762014000400414&lng=en. http://dx.doi.org/10.1590/0074-0276140024
41. Ranjbar R, Khamesipour F, Jonaidi-Jafari N, Rahimi E. Helicobacter pylori in bottled
mineral water: genotyping and antimicrobial resistance properties. BMC
Microbiol [Internet]. 2016 Mar [cited 2018 May 26]; 16: 40. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/26970903
42. Vesga FJ, Moreno Y, Ferrús MA, Campos C, Trespalacios AA. Detection of Helicobacter
pylori in drinking water treatment plants in Bogotá, Colombia, using cultural and molecular
techniques. Int J Hyg Environ Health [Internet]. 2018 May [cited 2018 May 26]; 221(4):
595-601. Disponible en:
https://www.sciencedirect.com/science/article/pii/S1438463918301378?via%3Dihub
43. Loke MF, Ng CG, Vilashni Y, Lim J, Ho B. Understanding the dimorphic lifestyles of
human gastric pathogen Helicobacter pylori using the SWATH-based proteomics approach.
Sci Rep [Internet]. 2016 May [cited 2018 May 28]; 6: 26784. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/27222005
44. Boehnke KF, Eaton KA, Fontaine C, Brewster R, Wu J, Eisenberg JNS, et al. Reduced
infectivity of waterborne viable but nonculturable Helicobacter pylori strain SS1 in mice.
Helicobacter [Internet]. 2017 Aug [citado 2018 May 23]; 22(4): e12391. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/28436616
45. Moreno Y, Ferrus MA. Specific detection of cultivable Helicobacter pylori cells from
wastewater treatment plants. Helicobacter [Internet]. 2012 Oct [citado 2018 may 26]; 17(5):
327-32. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/22967115
46. Assadi MM, Chamanrokh P, Whitehouse CA, Huq A. Methods for Detecting the
Environmental Coccoid Form of Helicobacter pylori. Front Public Health [Internet]. 2015
May [cited 2018 May 23]; 3: 147. Disponible en:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4446911/
47. Chomvarin C, Wongboot W, Tirapattanun A, Kanthawong S, Wongwajana S, Tongpim S,
et al. Detection of Helicobacter pylori in Aquati Environments and Drinking Waters in
Northeastern Thailand. Chiang Mai J Sci. [Internet]. 2017 Jul [cited 2018 May 29]; 44(3):
731-741. Disponible en:
https://www.ncbi.nlm.nih.gov/pubmed/?term=Boehnke%20KF%5BAuthor%5D&cauthor=true&cauthor_uid=25664781

16
https://www.researchgate.net/publication/318786035_Detection_of_Helicobacter_pylori_i
n_aquatic_environments_and_drinking_waters_in_northeastern_Thailand
48. Cunachi AM, Fernández M, Delgado P, Suárez M. Detection of Helicobacter DNA in
different water source and penguin feces from Greenwich, Dee and Barrientos Islands
Antarctica. Polar Biol [Internet]. 2016 Sep. [citado 2018 May 23]; 39(9): 1539-1546.
Disponible en: https://link.springer.com/article/10.1007/s00300-015-1879-5#citeas.
49. Flores-Encarnación M, Meza-de la Rosa JL, Aguilar-Gutiérrez GR, Cabrera-Maldonado C,
Aquino-Bonilla AG, Enríquez-Guerra MA, et al. Presence of Helicobacter pylori into
municipal water in common use. Bas Res J Med Clin Sci [Internet]. 2015 Jun. [citado 2018
May 23]; 4(6): 180-185. Disponible en:
http://basicresearchjournals.org/medicine/pdf/Flores-Encarnacion.pdf
50. Khan A. Farooqui A, Kazmi SU. Presence of Helicobacter pylori in drinking water of
Karachi, Pakistan. J Infect Dev Ctries [Internet]. 2011 Sep. [citado 2018 May 31]; 6(3): 251-
5. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/22421606


















