
Dinámica temporal de las microalgas de un tratamiento ... · des de algas que intervienen en dicho...
10
Dinámica temporal de las microalgas de un tratamiento terciario de aguas residuales urbanas a escala de laboratorio MIGUEL ALVAREZ COBEL4S* y TOMÁS GALLARDO** * Instituto de la Grasa y sus Derivados (CSIC), Avda. Padre García Tejero. 4.. E-41012 Sevilla. ** Departamento de Biología Vegetal 1, Facultad de Biología, Universidad Complu- tense. E-28040 Madrid. Resumen. En un agua procedente de un tratamiento biológico secundario con lo- dos activos se inoculó un cultivo mixto de Ch/ore/lo saecharoph/la y Seeaedesmus acu- /us que creció en discontinuo en sendas cubetas a 25 ‘C y a 30 ‘C, respectivamente (ilu- minación II klux: fotoperiodo. [5/9), determinándose los siguientes parámetros: clo- rofila a. densidad óptica a 560 nm., número de células por especie, biomasa (peso seco y biovolúmenes). diversidad, producción primaria, respiración, capacidad foto- sintética y tasas de sustitución específica. Además, para todo el proceso se realizaron correlaciones lineales con los factores químicos cuantificados en un trabajo anterior ~ se determinaron las constantes de crecimiento. C.socchoroph/lo fue desplazada por Oso//la/oria amplí/bio, que apareció espontá- neamente en los cultivos, debido a una mejor adaptación a la luz y a los nutrientes por parte de la segunda. Todos los procesos ocurrieron más rápidamente a 30 ‘C que a 25 ‘(1? como era de esperar. El fósforo y el carbono controlaron el crecimiento de las algas en esas condiciones. La producción primaria presentó máximos en los minutos posteriores al encendido de la luz. Palabras clave: cultivo, aguas residuales, algas, Ch/ore/la, Seenedesmus, Oso/lía/o- na. Abstract. A mixed culture of Chlorel/a sac.chorophi/a and Seenedosmus acutus was inoculated in urban water produced by activated sludge in a secondary treatment. As- says were undertaken in batch cultures (T = 30 “C and 25 0C, light intensity = II klux, photoperiod = 15/9) and the following parameters were assessed: chlorophyll a, optical density at 560 nm., number of celís. biomass (biovolumes and dry weighfl, diversity, primarv productivity. respiration. assimilation numbers and species turnover rates. In addition, sorne correlations were tried among chemical factors found in a prevíous paper and species growth rates were calculated. Fo!, (o;ííp/u/ens/s N.~ 1465-74. EdiL. Universidad Complutense, 1989
Transcript of Dinámica temporal de las microalgas de un tratamiento ... · des de algas que intervienen en dicho...

Dinámica temporaldelas microalgasdeun tratamientoterciario deaguas
residualesurbanas a escaladelaboratorio
MIGUEL ALVAREZ COBEL4S* y TOMÁS GALLARDO**
* Instituto de la Grasa y sus Derivados (CSIC), Avda. Padre García Tejero. 4..
E-41012Sevilla.** Departamentode Biología Vegetal 1, Facultadde Biología,UniversidadComplu-
tense.E-28040Madrid.
Resumen.En un aguaprocedentede un tratamientobiológico secundariocon lo-dosactivosse inoculóun cultivo mixto de Ch/ore/lo saecharoph/la y Seeaedesmus acu-/us quecrecióen discontinuoen sendascubetasa 25 ‘C y a 30 ‘C, respectivamente(ilu-minación II klux: fotoperiodo. [5/9), determinándoselos siguientesparámetros:clo-rofila a. densidadóptica a 560 nm., número de célulaspor especie,biomasa(pesosecoy biovolúmenes).diversidad,producciónprimaria, respiración,capacidadfoto-sintéticay tasasde sustituciónespecífica.Además,paratodo el procesose realizaroncorrelacioneslinealescon los factoresquímicos cuantificadosen un trabajoanterior~ se determinaronlas constantesde crecimiento.
C.socchoroph/lo fue desplazadapor Oso//la/oria amplí/bio, que apareció espontá-neamenteen los cultivos, debidoa unamejor adaptacióna la luz y a los nutrientespor partede la segunda.Todoslos procesosocurrieronmásrápidamentea 30 ‘C quea 25 ‘(1? como era de esperar.El fósforo y el carbonocontrolaronel crecimientode lasalgasen esascondiciones.La producciónprimariapresentómáximosen los minutosposterioresal encendidode la luz.
Palabrasclave: cultivo, aguasresiduales,algas, Ch/ore/la, Seenedesmus,Oso/lía/o-na.
Abstract.A mixed cultureof Chlorel/a sac.chorophi/a andSeenedosmusacutus wasinoculatedin urbanwaterproducedby activatedsludge in a secondarytreatment.As-sayswereundertakenin batchcultures (T = 30 “C and25 0C, light intensity= II klux,photoperiod= 15/9)and the following parameterswereassessed:chlorophylla, opticaldensityat 560 nm., numberof celís.biomass(biovolumesanddry weighfl, diversity,primarv productivity. respiration. assimilationnumbersand speciesturnoverrates.In addition,sornecorrelationsweretried amongchemicalfactorsfound in a prevíouspaperandspeciesgrowth rateswere calculated.
Fo!, (o;ííp/u/ens/s N.~ 1465-74.EdiL. UniversidadComplutense,1989

66 Álvarez Cobelas, M. et al.
(7. saccharophila was outcompetedby Oscillatoria amph/bia (that spontaneouslyappearcdin the cultures)due to betier light ami nutrient adaptationsof te seconó.Ah te phenomenaproceededfasteraL 30 0C than at 25 0C, asexpected.Phosphorusand carboncontrolledalgal growth in such eonditions.Primaryproductivity showedmaximal values in theminutesthat followed the light turning-on.
Key words: culture,urbanwater, algae,Ch/arel/a, Scenedesmus, Oscillatoria.
INTRODUCCIÓN
Se ha recalcadoen un trabajoanterior (VELASCO & al., 1986) la im-portanciay el interésdel uso de las microalgasen el tratamientoterciariode la contaminaciónproducidapor aguasresiduales.En estapublicaciónnos referiremosestrictamentea la dinámica temporalde las comunida-desde algasque intervienenen dichoprocesoa escalade laboratorio.
El estudiode la competenciaentreespeciesde microalgasen el labo-ratorio no es nuevo (TILMAN, 1982), tanto paralas marinascomo paralas dulceacuicolas,pero rara vez se ha realizadocon taxonesmuy resis-tentesa la contaminaciónorgánica.Por otro lado, el númeromayor detrabajosse ha llevadoa cabocon diatomeas,resultandomuy reducidoelde experienciasconcianofíceasy clorofíceas(MUR & al., 1978; VINCENT& SILVESTER, 1979a,b).
El interésde éstasúltimas procededel hechode que,genéricamentehablando,constituyeel final de la sucesiónestivaldel fitoplanetonen eco-sistemaseutróficos.
MATERIAL Y METODOS
Las especiesempleadasen el cultivo mixto fueron: Oscillataria am-phibia Ag. (Cyanophyceae),(7/dore/la saccharop/’uila (Krtiger) Migula(Chlorophyceae),ScenedesmusacutusMeyen (Chlorophyceae).
Lasdosúltimasconstituyeronel inóculo mixto; laprimeraaparecióes-pontáneamenteen los cultivos, quizá a partir de estadosquiescentesenel aguaresidual.Lascondicionesde cultivo y los métodosde análisisquí-mico ya se han relatadoen el trabajoprecedente(VELASCO & al., 1986);sinembargo,nosgustaríaseñalaraquíel mediode cultivo ha sido el aguaresidual,lo cual es pocofrecuenteen estudiosde estaíndole.
Paralas algas, se estimó cl númerode célulasmedianterecuentosenmicroscopio-invcrtido.-A-partirde-dichascifras,-se averiguaronlas tasasde crecimientoy los tiemposde duplicación,suponiendouna dinámicaexponencial.Tambiénse estudióla concentraciónde clorofila a, por ex-tracción en metanol en caliente y espectrofotometría(MARKER & al.,1980). La producciónprimariay la respiraciónfueronseguidasmedianteun electrodode oxígenoYSI-57conagitadorincorporado,introducidoen

Dinámicatemporalde las microalgasde... 67
un vaso de precipitadolleno de cultivo y tapadocon unacubiertatrans-parente,sumergidoa su vez en el cultivo; las experienciasen faselumi-nosaeranprecedidasde cincominutosen laoscuridadparainhibir la fo-torrespiracián(1-LARRIS & PKÚCININ, 1977).Labiomasade las algasse cal-culó, ademásde mediantela clorofila a, porpesoseco(residuoa 100 OC),
biovolúmenesy densidadóptica(colorimetríaa 560 nm.). La fórmuladeLEwIS (1978) sirvió para conocerla tasa de sustituciónespecífica.Conlos datosdel númerode individuosse halló la diversidadmediantela fór-mula de Shannon-Weaver.
Por último, se realizaronalgunascorrelacioneslinealesentreparáme-tros químicosy biológicos.
RESULTADOS
En la tabla 1 se muestranlos datosqueno se hanincluido en lasgráfi-cas.
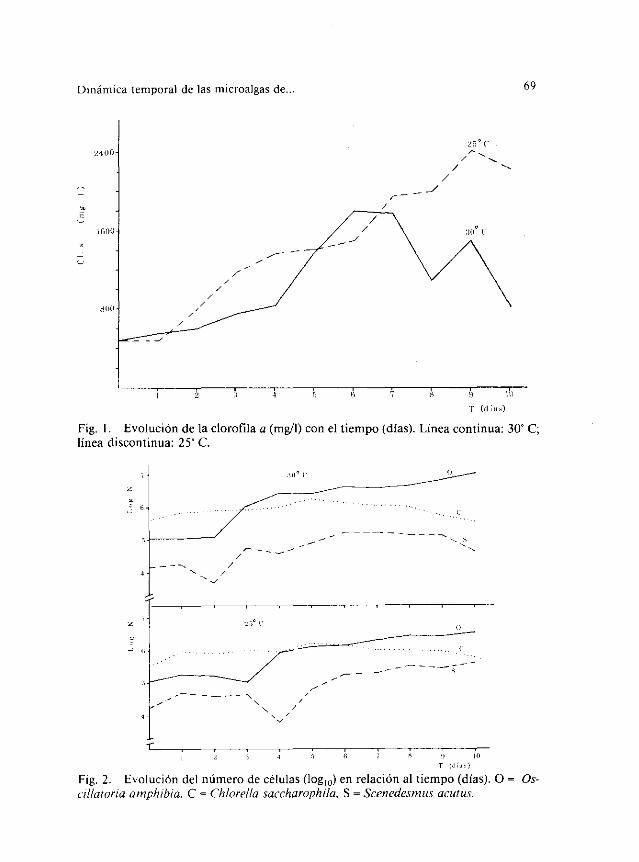
La biomasatotal de las algasofrecemáximosvariablessegúnel indi-cadorqueseconsidere.Pero,parala clorofila a, los picosse tienenen elsexto día (30 OC) y en el noveno(25 OC) (fig. 1). Lascorrelacionesde la ta-bla 2 nos señalanque es la densidadóptica (datos,en VELASCO & al.,1986) la que se liga significativamentea mayor númerode indicadoresde biomasa.En cualquiercaso,todaslas cifrasparecensermayoresa me-nor temperatura.
La clorofila a de los cultivos se encuentranegativamentecorrelacio-nadacon ortofosfatos,carbonoinorgánicoy amonio(tabla2), y positiva-mentecon los nitratos,lo cual podría sugerirqueel amonioes la princi-pal fuentenitrogenadaparalas especiesde algasestudiadas.La cantidadde clorofila a por célula, aun mostrandopautasalgo erráticas(tabla 1),apuntaunatendenciadecrecientecon el tiempo.
Seapreciaunacompetencia(ftg. 2) entre(7. saccharophilay O.amphi-bia, en la cual acabadominandola cianofíceaa pesarde la superiori-daddel algaverdeen los momentosiniciales. Dicho dominioes máspa-tentea 30’C quea25’C (tabla3), yaquelatasadecrecimientode O. amphi-bia es mayor a la temperaturamásalta. .5’. acutussin embargo,a pesardc presentaruna capacidadreproductorasuperiora (7. saccharophilanoconsigueaproximarsea lasotras algas.La cianofíceapresentacorrelacio-nessignificativasconel ortofosfatoy el carbonoinorgánico,mientrasqueS. saccharaphilano (tabla 3).
El florecimiento de O. amphibia en las experienciasse refleja en latasade sustituciónespecífica(fig. 3) y en la diversidad(tabla 1), aunqueno de la misma manera.Los cultivos acusanla entradamasivade dichotaxón (días3’, 30’ C y 4’, 25’ C, respectivamente),produciéndoseun au-mentosucesivode diversidad,quehacia el final declinapor efectode ladominanciadebidaa la cianoficea.

‘o ‘O e-— en~fl flO en a’ e--
oa e-- O 0 ‘t r~- Co=CM~O
‘00 — rl00
Coe-la’r~ 00
a’ 00 rl ‘tO 0—“no t-’,t CM’.’,en.n —CMCo ‘4
“o—~~’4 00Orn — ‘4 e--, -t e-— ej
Co en000 Co CM O~ 0—ao’t ‘otenCo
rl e-— 0c.O ‘o en -t 0~
-t —u~ -t
a’ rl te-- -t
Ú CM‘o
‘4(1CM ‘1. e-l-1-CM Co 00 “‘ CMeflen ,ttfla”4‘4en
0 <0 Co —o ‘o CM ‘00
-t ~r aa 000enCM enCM—CM0CM‘4CM
o Co en e-~ O’4en a’ 000CM ‘0 ‘0 ‘o
fl e-~- CM — a’ a’
CM -
Co=~na’o Co ‘o e-~ Co ~CM’N’OCO -
01~
0’ 00 ‘o ‘o -t ~t’4CMa’=enenCo~o-t’oa’e--
1 j—’r,r-~t’ooorlen
~‘,fl,tCo00tOOCo’oa’ (MO en -* ‘o ‘Oe-~Co’4~a.e-Co~De--00en’fl
e-- =0 ‘00 ‘4 0 CM =00 a’ 4 Co fl O ‘O
1 ——‘oso’o’n00’fl~t
(MCM ‘0CM ‘*0CM CM a’a’ 0CMenQnCoe-—’fl ‘t Coo ‘o ~ ~ -t 00 e-- en
en e-— CM ‘o rl ‘4 0CM a’ -*
—0 -t ‘4~ 00 ‘o -t
‘oCo CM ‘O Co 0 -t ‘00CM‘O O ej O -t CM ‘fl CM
1 dJWfe’<4do’o’n
0oOCo—~ ~o~oe-—lCo—t’OoOe--ie--lCo00’oenenCo ‘oCotr—a’13n’tCM0~n
— e-.. a’ O O a’ ‘113 Cl Cr~ O OenCMCMCM O O’0 ‘~nen O — Co e--a’ en
‘o ‘000 -tÚ
~¿ tta’-.a’.0.?. Co Co
--0CoCo0 0rw~CMen 00 1o oo a’ a’
QOUUQUUUQUUOUUQUQO
0~0.r0.r0~ 0.n0~n013n0’fl0~’en e-len (Men e--len e-lenCM en rl en rl en e-len CM
.0
c~o) ‘0%í0*’~3 elo
.0 ‘0--0’=0o ~, a
68 Alvarez Cobe/as, M. e! o/.
0
abae
O)
o
aueoeo
-0
00‘oo.0
o
‘cece
‘it-1
o) ‘ixE ~ZZ13,, -=~
69Dinámicatemporalde las mieroalgasde...
-1-
íj— a
Y
1~:1 -‘ 9 6 7 8 9 lO
T (dil)
Fig. 2. Evolucióndel númerode células(1og10)en relaciónal tiempo(días).O = Os-cí/lotoria amph/bia. C Chíarella saccharophila. 5 = Seenedesmus acutus.
~25 U
y
-‘-13
-Á
r
:10 U
1 td ja’)
Fig. 1. Evoluciónde laclorofila a (mg/l) conel tiempo(días). Líneacontinua 30 Clíneadiscontinua:25 C.
- y-13,

A/vare: (‘obelas. M. e! al.
TABLA 2. Correlaciones lineales entre diversos parámetros. * = significativa P<0,05;
(ns) = no significativas.
30’C 25’C
Cl. a-BiomasaCI. a-D.O.CI. a-PesosecoBiomasa-PesosecoPesoseco-DO.Biomasa-DO.Cl. a-PO~Cl. a-C. inorgánicoCl. a-NHCl. a-NO;POtChlorellaPO~OscillatoriaC. inorgánico-ChlorellaC. inorgánico-Oscilla¡oria
0,51 (ns)Q94*0,04 (ns)0,26 (ns)0,96*0,44 (ns)
~0,7l*-0,78k..Q74*0,66*
-0,49 (ns)~0,62*-0,40 (ns)~Q,75*
0,92*0,91*0,94*0,88*0,93*0,86*
~0,9l*..Q97*130,89*0 66’
-0,52 (ns)~0,84-0,43 (ns)~0,90*
La producciónprimariaes mayora25’ C quea 30’ C, aunquelos máxi-mos se alcancenantesaestaúltima temperatura.En cuantoa la respira-ción, el fenómenoes similar, pero no tan claro (tabla 1). El seguimientode la fotosíntesisen periodosde cinco minutos(fig. 4) muestra,en gene-ral, los picos de producciónde oxígenoinmediatamentedespuésdel en-cendidode la luz.
TABLA 3. Tasas de crecimiento, r (día-’) y tiemposde duplicación,t (horas).
30’Cr
25’Ctr t
CAcillo/oria aniph/biaCh/ore/la soe’charophi/aSeenedesmus acutus
0,2060,1190,133
356054
0,1620,11 50,140
446251
DISCUSIÓN
El desplazamientode (7. saccharophilapor O. amphibiaes un procesoquemerececiertasconsideraciones.Contrariamentea estudiosanterio-res,aquíse hancomparadoun taxónplanetónico(la clorofícea)y unoben-tónico (la cianofícea).Estaúltima forma comunidadescasi monoespecí-ficasde algasdurantela primaveray el veranoen arroyosmuy contami-nadospor sustanciasorgánicas.(7. saccharophilatendríala necesidaddemoverseen la columnavertical, de modo pasivo,paraevitar la fotoinhi-bición (HARRIS, 1978),mientrasqueO. amphibia,adaptadaavivir eneco-sistemassomeros,no; teniendoen cuentaquese hanempleadocondicio-
70

Dinámicatemporal de las microalgasde... 71
2-3’
1- ¡
Y
/Y>
2 :3 5 -y
1’ (djn~)
Fig. 3. Tasasdesustituciónespecífica(dí&’) en relaciónal tiempo(días).Líneacon-tinua: 30’ C; líneadiscontinua:25’ C.
131313~ 1’ fN‘a “ A ¡ Y.FC
‘-‘-13-”-
¡¡J NSI j3<>’ c ¡
3.2’ t II
fi
a
7, lO [5 20 25 50 o <0 ‘5 20 25 30
-r (mm 3 ‘U (ni ini
Fig. 4. Producciónprimaria(mg 02/1) frenteal tiempo(minutos)de un día represen-tativo. Las flechas señalanel comienzode la fase luminosapara la producción.P = producciónbruta; N = producciónneta; R = respiración.

72 res (‘obelas. Al. e’! al.
nes luminosasfotoinhibidas(11 klux = 200 gE.m.2.seg-’,HARRIS, 1978)parael plancton,la cianofíceatendríaventaja.Porotro lado, parecequeuna y otra presentandistintas respuestasfotorrespiratorias:mientrasenel cultivo mixto seapreciancaídasen la fotosintesistraslos primerosmi-nutos (fig. 4), en cultivos casi puros de O. arnphibia (datosinéditos)di-chasdisminucionesno se observan;de modo queestaúltima tendríaca-racterísticasde plantaC
4 y Csaccharophi/ade C3 (TOLBERT, 1974), se-gún los valoresbajos o elevadosde la excreciónde carbonoen la luz, lapequeñaexcreciónaumentaríala produccióny la ventajaadaptativa,portanto, de la cianofíceaen igualescondiciones.Se sabetambiénque mu-chas algasazulesposeenfosfatasaalcalina (HEATH & COOKE. 1975), lacual les permitemetabolizarpolifosfatoscuandodesapareceel ortofosfa-to; éstefenómenopodría ser otro elementodesfavorablepara(7. saecha-raphi/a. Además,VINCENT & SILVESTER(1 979b)señalanqueexistentoxi-nasexcretadaspor cianofíceasque inhiben el crecimientode (7. saceha-raphi/a. Todosestosfactorespodríanexplicarla dominanciade O. a¡np/ii-lila, que se refleja no sólo en el mayor númerode célulassino en la su-perior tasade crecimiento(tabla 3). Sin embargo,nuestrosdatosno nospermitensaberqué factoro factoresdesencadenanel florecimientode laetanofícea.
El crecimientode los cultivos persisteauncuandoalgunosde los nu-trientes(C y P) casise hanagotado.Estefenómenosedebea quesonmásimportantesparael crecimientolos contenidoscelularesde un nutrienteque su concentraciónambiental (RHEE. 1980), de modoque una reservanutritiva adecuadamantienea un cultivo en crecimientono habiendodi-chassustanciasen el ambiente;las ingestiones«suntuarias»(luxury upta-ke) estánbien documentadasen la literatura desdeKETCHUM (1939) yson inversamenteproporcionalesa la cantidadintracelulardel nutriente(BROWN & HARRIS, 1978, para el fósforo). La relación entrelas tasasdecrecimientoy los contenidosinternosde los nutrienteseslineal (DROOP.1973) por lo cual se hace másnecesarioestimaréstasque la concentra-ción en el medio. Se sabe,por otra parte,que la escasezde fósforoen elaguaestimulala ingestiónde carbono(LEAN & PICK, 1981); esto explica-ría los abruptosdescensosen el carbonoinorgánicoque sucedenal ago-tamiento del fósforo en los cultivos (datosno publicadosy tablas 1 y 2de VELASCO & al., 1986).
La limitación inicial parala nutrición es la del carbono,seguidaen eltiempopor la del fósforo;es bienconocidala inexistenciade limitacionesnutritivasmúltiples(RHEE, 1978;AHLGREN, 1980).Respectoalusodel ni-trógeno,no es fácil averiguarcuál es la forma preferidapor las algas; lanitrificación disminuyóa partir de los días7’ (30’ C) y 8’ (25’ C), aumen-tandoconcomitantementeel amonio,pero ignoramossi dicho fenómenose debea la muertede las bacteriasnitrificantesporel burbujeode la ai-reación(NAMEIAR & BOKIL, 1981) o al consumode las algas;y lo mismo

Dinámicatemporalde las mieroalgasde... 73
puededecirsedel amonio: ¿desaparecepor nitrificación o lo ingieren lasalgas?
Como se esperabapor ser un fenómenobienconocido,todoslos pro-cesosse acelerana mayor temperatura,con lo cual (al no haberentradade materia)los procesosdecrecimientoterminanantesa 30’ C quea25’ C.Por otro lado, las correlacionessignificativasson mayoresa menortem-peratura,quizáporqueéstaseaóptima paraestasalgas.
Las tasasde crecimientoson menoresa las observadaspor VINCENT& SILVESTER (1979a) para algasaisladasde agua residual,pero debete-nerseen cuentaquedichosautorestrabajabanconmediossintéticos,másequilibradosen nutrientes.
Los puntosde vista esbozadosanteriormentey las observacionesrea-lizadas ligan, a nuestrojuicio, los procesosqueocurrenen el ambienteque rodeaa las algasconlos que tienenlugaren el interiorde las células.La diferenciaentreunaaproximacióny otra es cadavez menoren el es-tudio de las algasmicroscópicas.
REFERENCIASBIBLIOGRÁFICAS
AHLGREN. c. 980. Effects on alga] growth ratesby multiple nutrient limitation .Arch.lIvárohiol? 89: 43-53.
BROWN, E. J. & R. F. HARRIS. 1978. Kinetics of algal transientphosphateuptakeand theccli quota concept.LI,no/i Oceanogí’. 23: 35-40.
DROOP. M. R. 1973.Somethoughtson nutrientlimitation in algae..1 Phyco/? 9:264-272.FIARRIS. o. p. 1978. Photosvdthcsis,productivityandgrowth: the physiologicalecology
of pbytoplankton.Ergeb. Limnologie 10: 1-171.IFARIZIS. 6. 1>. & u. u. PICCININ. 1977. Photosynthesisby naturalphytoplanktonpopula-
tions. .Arch. llvdrobio/i 80: 405-457.IIEATH. It. T. & ci. liv COOKL. 1975. The significance of alkaline phosphatasein an eu-
trophic lake. Verh. Ial. ¡‘en LimnoL 19: 959-965.KETCHUM, B. H. 1939. Re absorptionof phosphateandnitrateby illuminatedcultures
of NiI:schia e-los!erium. Am. J. Bol. 26: 339-407.LEAN. D. It. 5. & E. R. PICK. 1981. Photosynthetic responseof lake plankton to nutrient
enrichment:a testfor nutrient limitation. LimnoL Oceanogr.26: 1001-1019.LEWIS. W. M. 1978.Analysis of successionin a tropical phytoplankíon eommunityand
a newmeasureof successionrate.Am. No!. 112: 401-414.MARKER. A. E. U.. E. A. NUSCH, H. ¡tAl & B. RIEMANN. 1980. The measurement of pho-
tosyntheticpigmentsin freshwatersandstandardizationof methods:conelusionsandreeomendations.lúrgeb. Linínologie 14: 91-106.
M(JR.J. T.. U. J. C,0N5& E. ~an LIERE. 1978. Competition of the greenalgaScenedesmusand the blue-greenOscil/atoria. M/tt. In!. Leí; LimnoL 21: 473-479.
NAMBIAR. K. It. & 5. D. BOKIL. 1981. Luxury uptakeof nitrogen in flocculating algal-bac-
terial system. U’ater Rcsearch15: 667-669.RHEE. o. Y. 1978. Effects of N:P atomieratios andnitrate limitation on algalgrowth,
celí compositionandnitrate uptake.LimnoL Oceanogri 23: 10-25.

74 ÁlvarezCobelas, M. e! aL
Rl-lEE. o. c. 1980. Continuousculturein phytoplanktonecology. In: Advancesin Aqua-tic Microbiology 2: 151-203. AcademiePress. N. Y.
TILMAN. fi. 1982. Resource competition and community 5/ructure. Princeton Univ.Press.Princeton.
TOLBERT, N. E.1974. Photorespiration.In: W. D. P. STEWART (Ed.) A/gal Physiologyand Biochemistrv 474-504. Blackwell. Oxford.
VELASCO, J. L.. J. M. COLMENAREJO,M. ALVAREZ. A. BUSTOS& E. PARRA. 1986. Impactoam-biental de los venidos de aguasresidualesurbanas,1. Depuraciónintegralcon re-cuperaciónde nutrientes mediantetratamientoterciario fotosintético con mi-croalgas.Químicae lndus!r/a 32: 475-478.
VINCENT. W. E. & W. B. SILVESTER. 1979a. Growth of blue-greenalgaein the Manukau(New Zealand)oxidationponds.1. Growth potentialof oxidationpond waterandcomparativeoptima for blue-greenand green algal growth. VÍ-’ater Research13:711-716.
VINCENT. W E. & w. B. SILVESTER. 1 979b. Ibid. II. Experimentalsiudieson algal interac-tion. Water Research 13: 717-723.


















