Evaluación de patrones para lima tahití (Citrus latifolia ...
67
Evaluación de patrones para lima tahití (Citrus latifolia Tanaka) frente a Citrus tristeza virus (CTV) Johnny Camilo Beltrán Montoya Universidad Nacional de Colombia Facultad, Agronomía (Escuela de posgrados) Bogotá, Colombia 2012
Transcript of Evaluación de patrones para lima tahití (Citrus latifolia ...

Evaluación de patrones para lima tahití (Citrus latifolia Tanaka) frente a
Citrus tristeza virus (CTV)
Johnny Camilo Beltrán Montoya
Universidad Nacional de Colombia
Facultad, Agronomía (Escuela de posgrados)
Bogotá, Colombia
2012


Evaluación de patrones para lima tahití (Citrus latifolia Tanaka) frente a
Citrus tristeza virus (CTV)
Johnny Camilo Beltrán Montoya
Trabajo de investigación presentado como requisito parcial para optar al título de:
Magister en Ciencias agrarias con énfasis en fitoprotección integrada
Director:
Ph.D., Oscar Arturo Oliveros Garay
Línea de Investigación:
Virus de plantas
Universidad Nacional de Colombia
Facultad, Agronomía (Escuela de posgrados)
Bogotá, Colombia
2012


¡Estudiemos y capitalicemos conocimiento!
Lo que aprendemos a hacer y lo que
tenemos en la cabeza es lo único que nos
queda hasta el día de nuestra muerte. Las
cosas materiales son polvo que arrastra el
viento.
A mi mamá, esposa e hijo.
Agradezco el apoyo y tiempo para llevar a
cabo mis estudios.
¡La felicidad es tan tangible que podemos ser
felices cuando queramos y nos decidamos a
serlo!


Agradecimientos
Al grupo del laboratorio de biotecnología vegetal de la facultad de agronomía de la
Universidad Nacional de Colombia.
Al profesor Oscar Oliveros agradezco brindarme la posibilidad de trabajar en este
proyecto y por su aporte intelectual.
Al doctor Javier Orduz y su grupo de trabajo por el apoyo y oportuna colaboración
A Walter Turizo por su oportuna colaboración y la amistad gestada en el transcurso de
este trabajo.
A mis compañeros de laboratorio por hacer un sano equipo de trabajo.
A Corpoica por poner a disposición sus instalaciones, enfocadas en la investigación
especializada de cítricos y por la colaboración de las personas que hicieron posible este
trabajo.


Resumen y Abstract IX
Resumen
En el presente trabajo se evaluó la respuesta de patrones para lima Tahití frente a
infecciones de Citrus tristeza virus. El documento se divide en tres capítulos. El primer
capítulo es una revisión general del estado del arte de CTV, que abarca generalidades
sobre sus hospederos, distribución, genoma viral, síntomas que generan las diferentes
cepas y antecedentes de lima Tahití en Colombia. El segundo capítulo abarca la
identificación de cepas de CTV que infectan en campo mediante el uso de primers
específicos de los genes de la polimerasa y p23, y secuenciación de los productos de
PCR. Los resultados muestran que las cepas de genotipos VT son las de mayor
incidencia, solas o de manera mixta con cepas T30 y T36. El tercer capítulo presenta los
resultados de la respuesta de expresión de “stemp pitting” de lima Tahití injertada sobre
seis patrones diferentes y su relación con los genotipos de CTV que infectan en campo.
La relación de dependencia entre patrones, respuesta sintomática y genotipos de CTV
se realizó mediante test de independencia de chi-cuadrado. Se identificó que la
respuesta de stem pitting es dependiente del patrón y de la localidad geográfica, pero
independiente de la mezcla de cepas que infectaron los cítricos.
Palabras clave: Lima Tahití, CTV, PCR, patrón.
Abstract
This study evaluated the response Tahitian lime grafted in different citrus rootstocks
against Citrus Tristeza Virus infections. The document is divided into three chapters. The
first chapter is an overview of the state of the art of CTV covering their hosts, distribution,
viral genome, symptoms induced by different strains and Tahitian lime characteristics.
The second chapter deals with the identification of CTV isolates that infect field using
PCR and specific primers for polymerase and p23 genes, and sequencing of the PCR
products. Results reveal VT genotypes strains exhibited the highest incidence, alone or

mixed with strains such T30 and T36. The third chapter deals with the response of stem
pitting expression in Tahitian lime grafted on six different rootstocks and their relationship
with CTV genotypes found in the Colombian field. A dependency relationship between
citrus patterns, symptomatic response and CTV genotypes was performed using chi-
square independence. It was found dependency response between stem pitting
rootstocks and geographical locality, but independency relationship of the mixture of
genotypes infecting a specific citrus.
.
Key words: Tahitian lime, CTV, PCR, rootstock.

Contenido XI
Contenido
Tabla de contenido
Resumen ......................................................................................................................... IX
Introducción .................................................................................................................. 15
Objetivo General ............................................................................................................ 17
Objetivos Específicos ................................................................................................... 17
1. Estado del arte........................................................................................................ 18 1.1 Tristeza de los cítricos................................................................................... 18 1.2 Agente causal: Citrus Tristeza Virus .............................................................. 19 1.2.1 Genoma Viral ............................................................................................. 20 1.2.2 Síntomas causados por CTV ...................................................................... 22 1.2.3 Transmisión ............................................................................................... 25 1.3 Hospedero: Citrus latifolia Tanaka (Lima Tahití) ............................................ 25 1.4 Bibliografía .................................................................................................... 28
2. Identificación de cepas de Citrus Tristeza Virus mediante marcadores moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka). ........... 34
Resumen ............................................................................................................ 34 2.1 Introducción .............................................................................................. 35 2.2 Materiales y métodos .................................................................................... 37 2.2.1Extracción de ARN, transcripción reversa (RT) y reacción en cadena de la polimerasa (PCR). ............................................................................................... 39 2.3 Resultados .................................................................................................... 41 2.4 Discusión ...................................................................................................... 46 2.5 Bibliografía .................................................................................................... 47
3. Patrones para lima Tahití C. latifolia (Tanaka) frente a la severidad de stem pitting ocasionado por Citrus Tristeza Virus ............................................................... 51
3.1 Introducción .................................................................................................. 52 3.2 Materiales y métodos. ................................................................................... 54 3.3 Resultados .................................................................................................... 58 3.4 Discusión ...................................................................................................... 61 3.5 Bibliografia .................................................................................................... 63
Conclusiones ................................................................................................................. 67

Contenido XII
Lista de figuras
Figura 1-1: Distribución mundial del virus de la tristeza de los cítricos……………………………………………..….………….4
Figura 1-2: Micrografia electrónica de cepas purificadas de CTV………………………………………………………………….6
Figura 1-3: Organización del genoma viral de CTV. Tomado de Karasev et al, 1995…………………………………………7
Figura 1-4: Clasificación de asilamientos de CTV propuesta por Hilf et al., 2005………………………………………………9
Figura 1- 5: Clasificación de asilamientos de CTV propuesta por Sambade et al., 2003…………………………………….9
Figura 1 6: Síntomas causados por CTV. Tomado de Moreno et al., 2008……………………………………………………...11
Figura 2-1: Esquema de toma de muestras y flujo de decisión para seleccionar plantas a evaluar …………………………22
Figura 2-2: Electroforesis en gel de agarosa para visualizar amplicones de VT-POL deCTV (695 pb) de las muestras de lima Tahití colectadas en el huerto Corpoica La Libertad ……………………………………………………………………...…26
Figura 2-3: Electroforesis en gel de agarosa para visualizar amplicones T30 POL de CTV (696 pb) de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La ibertad……………………………………………………………………26
Figura 2-4: Electroforesis en gel de agarosa para visualizar amplicones T36 POL de CTV (714 pb) de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La Libertad…………………………………………………………………………………………………………………………………27
Figura 2-5: Electroforesis en gel de agarosa para visualizar amplicones del gen P23 de CTV de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La Libertad………………………………………………………………………………………………………………………………...28
Figura 2-6: Electroforesis en gel de agarosa para visualizar amplicones de CTV (p23) de las muestras de lima tahti colectadas en el huerto ubicado en Corpoica Nataima……………………………………………………………………………..29
Figura 3-1: Patrones micro injertados con lima Tahití y establecidos en campo……………………………….…………..39
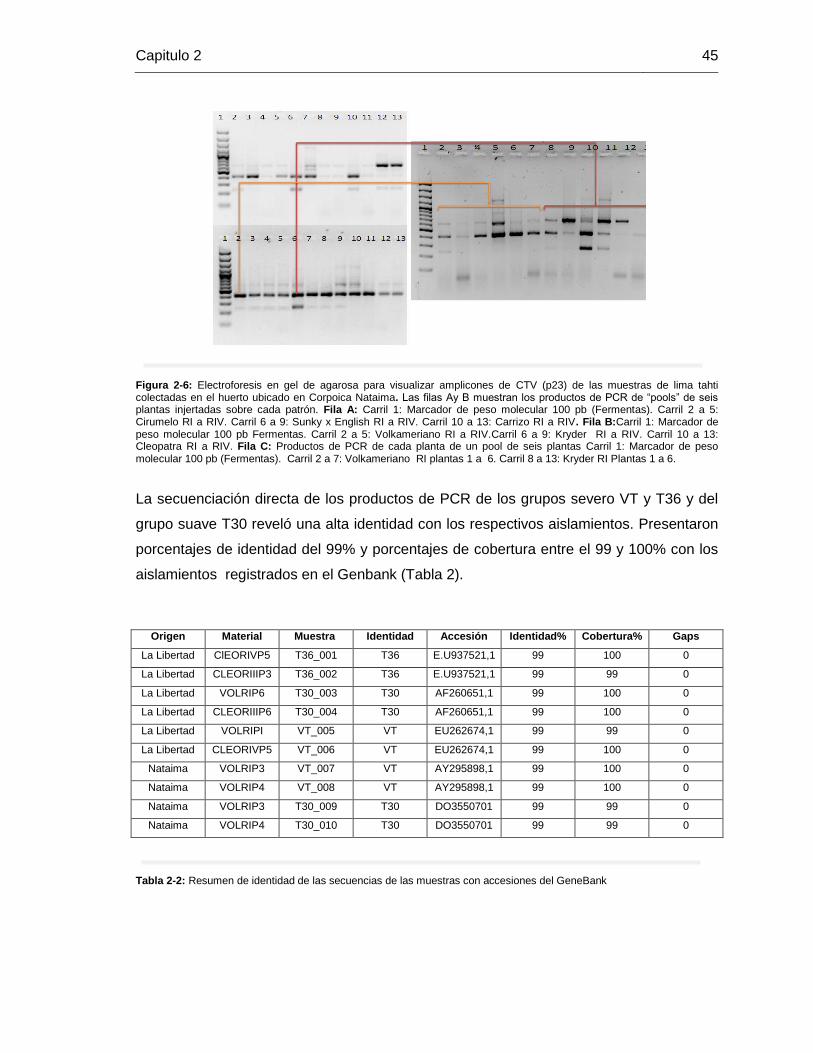
Figura 3-2: Escala diagramática para la evaluar la intensidad de acanaladuras de tallo. Tomada de Meissner-Filho et al. (2002)…………………………………………………………………………………………………………………………………..40
Figura 3-3: Distribución de las cepas de CTV que infectaron de manera natural en campo…………………………………..41
Figura 3-4: Severidad de stem pitting (dimensión 2) vs patrón y localidad (dimensión 1)……………………………………..44
Figura 3-5: Severidad de stem pitting vs asilamientos de CTV……………………………………………………………………45

Contenido XIII
Lista de tablas
Tabla 2-1: Primers utilizados para detección de CTV. Tomado de Hilf et al., 2005 y Sambade et al., 2003…………….25
Tabla 2-2: Resumen de identidad de las secuencias de las muestras con accesiones del GeneBank……………….………29
Tabla 3-1 Tabla de frecuencia de la severidad en stem pittin en diferentes patrones y dos localidades
diferentes………………………………………………………………………………………………………………………………..43

Contenido XIV
Abreviaturas Abreviatura Término
CTV Citrus tristeza virus SP Stem pitting QD Quick decline

Introducción
A nivel mundial los cítricos son cultivados en el subtropico y en el trópico, sin embargo,
las principales zonas productoras del mundo se encuentran ubicadas en el subtrópico
entre los 25 y 40 grados de latitud en ambos hemisferios y se denomina como el cinturón
citrícola conformado por California, Florida, China, Japón, Brasil, Argentina, Australia y
Suráfrica. Según la encuesta Nacional Agropecuaria de 2010, en Colombia hay plantadas
62.409 ha de cítricos, de las cuales 36.943 ha están sembradas en naranja y 25.466 ha
plantadas entre mandarina, tangelo y limas ácidas. El grupo de las limas ácidas está
compuesto por la lima Tahití (Citrus latifolia) y el limón pajarito (Citrus aurantifolia), estas
son originarias del archipiélago malayo, no toleran bajas temperaturas y alcanzan la
mejor calidad interna y coloración en las regiones tropicales cálidas (Orduz-Rodriguez,
2012). El cultivo de lima Tahití en Colombia tiene altas potencialidades en cuanto a
diversificación de cultivos y exportación, el país tiene tierras aptas para el establecimiento
de cultivos, la demanda interna no está satisfecha y en los momentos de
desabastecimiento es necesario importar limón verdadero, sin embargo, el modelo de
producción no cuenta con paquetes tecnológicos que garanticen la sanidad vegetal del y
presenta diversas limitaciones fitosanitarias. En Colombia la tristeza de los cítricos tiene
alto impacto sobre lima tahiti que es uno de los hospederos más susceptibles. En campo
hay alta presión de áfidos vectores, las prácticas de vivero no garantizan material vegetal
libre de virus o como mínimo infectado con cepas suaves y se constituyen como una
fuente de diseminación regional de enfermedades. Las estrategias de manejo de tristeza
son (1) material vegetal libre de virus, (2) selección de patrones para conferir tolerancia,
(3) erradicación de plantas infectadas con cepas severas, (4) esquemas de protección
cruzada (5) movilización de material vegetal sano, (6) regulación oficial sobre los
productores de material vegetal. Mediante estudios previos del centro de investigación
C.I Corpoica La libertad, se han reconocido patrones para lima tahiti con buen potencial
de adaptación a condiciones agroecológicas de las zonas de producción.

16 Estado del arte.
La limitaciones tecnológicas del cultivo de lima Tahití esta constituidas principalmente
por la falta de material de propagación con características de calidad, resistencia a
enfermedades y adaptación a condiciones edafoclimáticas. El patrón volkameriana es el
más usado en la citricultura colombiana, tiene ventajas debido a su precocidad, pero el
color de los frutos generados es verde claro, lo que disminuye el interés en el mercado
de exportación. Lima Tahití en uno de los hospedantes más sensibles al virus de la
tristeza de los cítricos y esta es una de las principales limitaciones de la longevidad de
los cultivos. En los llanos orientales se reporta tiempo de vida útil de los huertos llega
hasta los ocho diez años máximo causa de la severidad de CTV. En este sentido, el
fortalecimiento tecnológico para la producción de limas ácidas en Colombia requiere la
evaluación de patrones y programas de selección de materiales promisorios de alta
productividad que generen garantías para aumentar el área sembrada y la reconversión
de los materiales de producción que actualmente son utilizados.
En la citricultura moderna el esquema de propagación se basa en el uso de injertos con
el fin de generar precocidad en la producción, resistencia a la planta frente a
enfermedades y adaptación a suelos. El injerto cítrico se compone de un patrón y una
copa. El patrón confiere varias características de producción y repuesta a enfermedades
sistémicas a la planta, debe presentar alta compatibilidad con la copa, buena adaptación
a clima y suelos, confiere más de 20 características a la planta cítrica, entre ellas el vigor,
tolerancia a factores abióticos como salinidad, acidez, resistencia o tolerancia frente a
enfermedades como Phytophtora. sp, Citrus Tristeza Virus, Citrus psorosis virus, Citrus
exocortis viroid, entre otros. La copa debe provenir de plantas madres sanas (libres de
virus), con características de alto rendimiento y calidad de frutos. En el presente trabajo
se evaluó la respuesta de patrones para copas de lima Tahití, obtenidas a partir de
yemas libres de virus, respecto al carácter de conferir tolerancia frente a infección de
Citrus Tristeza Virus. En trabajos previos realizados por el banco de germoplasma del C.I
Corpoica Palmira se obtuvieron micro-injertos de lima Tahití libres de tristeza de los
cítricos con patrones promisorios en citricultura, el comportamiento de los cuales fue
subsecuentemente evaluado en campo.
La principal fuente de diseminación de CTV es la movilización de material infectado de
cítricos. Los viveros de propagación en Colombia no cuentan con un programa de
certificación de yemas libres de virus. La falta de regulación en el transporte de cítricos
puede generar un flujo de CTV a nivel regional. La diseminación local es mediada por

Capitulo 1 17
áfidos vectores presentes en campo como Aphis gosippy y Toxoptera citricida. Las
medidas de manejo de CTV deben incluir cuarentena de algunas cepas severas,
programas de certificación de yemas, el uso de patrones tolerantes y protección cruzada
con cepas suaves dependiendo de la incidencia y las cepas predominantes de cada zona
(Moreno et al., 2009). El uso de patrones tolerantes a CTV constituye una estrategia de
importancia para el manejo de la tristeza de los cítricos y la modernización de la
citricultura del país. Para generar programas de protección cruzada, es necesaria la
identificación de cepas suaves y la evaluación de su potencial para la protección frente a
cepas severas locales.
En el marco de este trabajo se espera que el material plantado en campo se infecte con
las cepas locales asociados a la región geográfica, razón por la cual la evaluación de
este material se realizó a los dos años de establecido el cultivo en campo. Durante este
periodo, el material en campo estaría expuesto a infecciones por las cepas locales de
CTV que son inoculadas por áfidos vectores. Estos árboles probablemente expresan
síntomas de tristeza asociados a la especificidad del genotipo viral, el patrón y la zona de
evaluación.
El presente trabajo se compone de tres capítulos principales. El primero presenta una
revisión general del estado del arte, el segundo es referente a un articulo enfocado en la
identificación de cepas de CTV en campo infectando lima Tahití, el tercer capítulo es un
artículo que trata de la respuesta de las plantas de lima Tahití injertadas sobre varios
patrones frente a la infección de campo de cepas de CTV.
Objetivo General
Evaluar la respuesta de patrones para lima Tahití (c. latifolia Tanaka) frente a Citrus
Tristeza Virus (CTV)
Objetivos Específicos
Identificar las cepas de CTV que infectan lima Tahití (C. latifolia) en campo mediante RT-
PCR y secuenciación.

18 Estado del arte.
Analizar la relación entre las cepas de CTV detectadas en campo y la respuesta
sintomática inducida en C. latifolia injertado sobre diferentes patrones.
1. Estado del arte
1.1 Tristeza de los cítricos
La tristeza de los cítricos es una enfermad de origen viral causada por Citrus Tristeza
Virus (CTV). Las sinonimias de esta enfermedad son Citrus quick decline virus (Fawcett
& Wallace, 1946); Citrus seedling yellows virus (Fraser, 1952); Grapefruit stem pitting
virus (Oberholzer et al., 1949) y Lime die-back virus (Hughes & Lister, 1949). Infecta
todas las especies e híbridos de cítricos, incluso hay reportes de infección en “parientes”
de los cítricos, tales como Aeglopsis, Afraegle, Fortunella y Pamburus, Poncirus. Fuera
de las Rutáceas, el único hospedero conocido de CTV, son algunas especies de
Passiflora (Bar Joseph et al., 1989)

Capitulo 1 19
Figura 1-1: Distribución mundial del virus de la tristeza de los cítricos. CMI (1978)
El primer determinante de dispersión de la tristeza de los cítricos data del siglo XIX
cuando se empezó a utilizar Citrus auranti (naranjo agrio) para conferir resistencia frente
a la pudrición de raíz causada por Phytophthora sp. Esta experiencia ocasionó la
diseminación de CTV desde zonas endémicas a zonas libres por la movilización de
material infectado (Bar Joseph et al., 1989). Las epidemias más importantes de tristeza
se reportaron en Argentina en 1930, en Brasil en 1937, en California en 1939, en Florida
en 1951, en España 1957, en Israel en 1970 y en Venezuela en 1980, (Webber, 1943;
Davino et al., 2003; Garnsey et al., 2000). En la actualidad CTV se encuentra en la
mayoría de las regiones de producción de cítricos del mundo, este virus se encuentra en
países de Europa, África, Asia, Norte América, Oceanía y Sur América.
1.2 Agente causal: Citrus Tristeza Virus
Citrus Trizteza Virus pertenece al género Closterovirus, familia Closteroviridae. Las
partículas de este virus son descritas como filamentos flexuosos de 10 a 11 nm de
diámetro y 1900 a 2000 nm de longitud. El genoma de CTV está constituido por RNA de
cadena sencilla, de sentido positivo, sin cola de poli-A en el extremo 3' (Bar Joseph y
Lee, 1989). Posee dos proteínas de cápside: una proteína mayor de 25 kDa que cubre el
95% de la partícula viral y una proteína menor (proteína divergente) de 27 kDa que cubre
el 5% de la partícula al extremo amino terminal (Granoff & Webster, 1999)
Figura 1-2: Micrografia electrónica de cepas purificadas de CTV. Tomada de:Bar Joseph y Lee, 1989AAB.

20 Estado del arte.
1.2.1 Genoma Viral
El RNA genómico (gRNA) de CTV se encuentra compuesto por 12 marcos de lectura
abiertos (ORFs) y regiones no traducibles (UTRs) en los extremos 5' y 3'. Los ORFs 1a y
1b codifican proteínas asociadas a la replicación. El ORF 1 codifica para metiltransferasa,
dos proteasas y helicasa. 1b codifica RNA polimerasa dependiente de RNA. Estas dos
abarcan la mitad 5' del genoma y son traducidos directamente por el gRNA. Los 10 ORF
adicionales se localizan en el extremo 3' y codifican las proteínas p33 (ORF2), p6 (ORF
3), p65 (ORF 4), p61 (ORF 5), p27 (ORF 6), p25 (ORF 7), p18 (ORF 8), p13 (ORF 9), p20
(ORF 10) y p23 (ORF 11). p65 es homologa a proteínas de choque térmico y junto con
las proteínas de la capside p25 y p27 están involucradas con el ensamblaje del virión.
p20 está asociada con la acumulación de cuerpos de inclusión en las células infectadas.
La función de p33, p18 y p13 se relaciona con la capacidad de inducir infección sistémica
en el rango de cítricos hospederos (Tatineni et al., 2011). Los ORFs del extremo 3´ de
CTV se expresan a través de un set de RNAs subgenómicos 3´ coterminales los cuales
tienen sus versiones correspondientes de RNA negativo (Hilf et al., 1995).
Figura 1-3: Organización del genoma viral de CTV. Tomado de Karasev et al, 1995
La cubierta del virión de CTV se encuentra formada por la proteína mayor CP de 23000
Dalton y una proteína menor CPm de 21000 Dalton que se encuentran presentes en un
proporción de 5 a 1(Bar Joseph y Lee, 1989).
CTV tiene el genoma más largo entre los virus que infectan plantas, presenta variabilidad
diferencial, el extremo 5’ tiene mayor variabilidad que extremo 3’. Este último es
relativamente conservado (aproximadamente 90% de identidad nucleotídica entre
aislamientos), mientras que la identidad encontrada en el extremo 5’ es menor al 70%
(Hilf et al. 2000). Los genomas de CTV secuenciados en su totalidad son: T36 y T30 de
Florida (Karasev et al.,1995; Albiach-Marti et al., 2000), VT de Israel (Mawassi et al.,
1996), T385 de España (Vives et al. 1999), SY568 de California (Yang et al. 1999),

Capitulo 1 21
NUagA de Japón (Suastika et al., 2001), Qaha de Egyto y un aislamiento mexicano. El
genoma de CTV como mínimo tiene tres genes que codifican supresores de
silenciamiento. CP es supresor de silenciamiento intercelular, p23 supresor de
silenciamiento intracelular y P20 supresor de silenciamiento en ambos niveles (Lu et al
2004)
La clasificación de genotipos de CTV ha identificado dos grupos (Hilf et al., 2005). El
primer grupo denominado VT contiene aislamientos VT, B14, T30 y T3, el segundo grupo
denominado T36, contiene aislamientos B83, B33 y T36. El grupo VT esta compuesto por
determinados subgrupos de aislamientos VT (B370, B152, B1, B370, B59, B79, B219),
B14 (B425, B211, B41), T30 (B213, B348, B270 – I, B2) y un subgrupo de aislamientos
T3. El grupo T36 tiene aislamientos B83, B33, T36 (B183, B359, B358). Esta clasificación
fue propuesta con base en los patrones compartidos de los marcadores genéticos
utilizados. Los aislamientos asignados al genotipo T30 tuvieron una mínima variabilidad
en la secuencia de nucleótidos entre la secuencia de los marcadores K17 y POL. La
variabilidad entre la secuencia de nucleótidos de los marcadores fue mayor para
genotipos asignados al grupo T36.
El gen p23 codifica una proteína de 23 KDa, se localiza adyacente al extremo 3' del
genoma de CTV. Este gen no tiene genes homólogos en otros closterovirus. P23 se
acumula en niveles moderados respecto a otras proteínas virales (Pappu et al., 1997),
sin embargo el RNA subgenomico de p23 es el segundo más abúndate en el RNA
mensajero viral de los tejidos infectados (Hilf et al., 1995; Navas – Castillo et al., 1997) y
se acumula más rápido que otros RNA sub genómicos en protoplastos infectados, lo que
sugiere que p23 está involucrado en los primeros pasos para la replicación o
transcripción (Navas – Castillo et al., 1997). Cítricos transgénicos que expresan p23 de
CTV exhiben sintomatología similar al de plantas infectadas (Ghorebel et al., 2001) lo que
demuestra que p23 se encuentra involucrado en el desarrollo de síntomas y
probablemente juega un papel determinante en la patogenicidad de CTV.
La función del gen p23 de CTV es asociada al control de la acumulación asimétrica del
número de hebras de RNA durante la replicación y es potencial determinante de la
patogenicidad de CTV. La región 78 – 80 de p23 se muestra como un marcador útil para
diferenciar grupos de aislamientos suaves, severos y atípicos. Los aislamientos suaves

22 Estado del arte.
tienen en la posición 78 – 80 de p23 Leu79 y Lys80; los aislamientos severos tienen Ser79 y
Arg80 y los atípicos tienen Gly78 (Sambade et al., 2003)
Figura 1-4: Clasificación de asilamientos de CTV propuesta por Hilf et al., 2005
Figura 1-5: Clasificación de asilamientos de CTV propuesta por Sambade et al., 2003.
1.2.2 Síntomas causados por CTV
Los virus que infectan plantas inducen la formación de diversas estructuras ó
alteraciones en la célula que están en función de la replicación del genoma o del
movimiento intracelular de estos. Se pueden formar esferulas, vesículas o cuerpos
multivesiculares, que pueden estar conectados o no con el citoplasma. La especificidad
en los organelos objetivo de los virus varía de acuerdo a la familia y genero (Laliberté et

Capitulo 1 23
al., 2010). Las proteínas virales de CTV forman inclusiones citoplasmáticas en las células
del floema, estás inclusiones están asociadas con el aumento de numero y tamaño de
vacuolas, invaginación de la membrana nuclear y degradación de cloroplastos que
ocasionan un desarrollo anormal y el colapso en general de las células (Rodriguez et al,
2009; Ecale Zhou et al., 2002).
La infección de CTV, se localiza en los conductos vasculares del floema de la planta,
impidiendo el transporte de asimilados a través de estos (Bar Joseph, 1970). Al interior
de las células acompañantes del floema, la cantidad de cuerpos de inclusión varía de
acuerdo a la cepa del virus (Bar Joseph y Lee, 1989). Las infecciones por CTV descritas
en las células del floema generan invaginaciones de la membrana nuclear y degradación
de los cloroplastos (Ecale Zhou et al., 2002).
La expresión sintomática de las plantas infectadas con CTV es variable y esta difiere de
acuerdo a la interacción patrón – copa y al tipo de cepa viral que causé la infección (Bar
Joseph, 1989). Sin embargo, los síntomas esenciales reportados en el mundo están
asociados a decaimiento rápido (Quick decline - QD) de las especies cítricas injertadas
sobre el patrón C. aurantium en donde se produce necrosis del floema en la unión del
patrón y la copa. El segundo síntoma de importancia es la acanaladura de tallo (stem
pitting - SP), en tallo y ramas de varias especies de cítricos, disminuyendo el vigor de la
planta, el tamaño de los frutos y el rendimiento en producción sin tener en cuenta el
patrón utilizado. El tercer síntoma está asociado a crecimiento retardado (stunting) y
clorosis en la hojas reportado en patrones de C. aurantium, C. limón y C. paradisi Macf
(Garnsey et al., 1987; Sambade et al., 2003). Los determinantes patogénicos de CTV aún
no están estrictamente definidos, no es claro si los síntomas son inducidos por una
variante genómica predominante o por una combinación de variantes genómicas de la
población.

24 Estado del arte.
Figura 16: Síntomas causados por CTV. Tomado de Moreno et al., 2008.a) Sintoma de decaimiento rápido
en naranja dulce propagada sobre naranjo agrio. B) Arboles en diferente estado de declinamiento. c) Sintoma severo de stem pitting en el tronco de árbol de toronja injertado sobre Poncirius trifoliata. d)Fruto de toronja pequeños de árbol de toronja injertado sobre Poncirius trifoliata. e) Plantula con crecimiento retardado y hojas con clorosis. f) Clorosis en las nervaduras de hojas de Citrus macrophylla.g) Stem pitting en el tronco de plántulas de lima mejicana. h) Lima mejica transgénica expresando p23 (izquierda) y hoja de lima mejicana no transgénica infectada con CTV.

Capitulo 1 25
1.2.3 Transmisión
La evolución de los géneros de la familia closteroviridae está asociada con fuerzas de
selección ejercidas por sus insectos vectores, resultando en tres linajes principales
asociados a la transmisión de closterovirus por áfidos, crinivirus por moscablanca y
ampeñovirus por cochinillas (Karasev, 2000). La trasmisión de CTV es semipersistente y
en plantas infectadas está dirigida a células asociadas al floema (Martelli et al.2002).
CTV tiene dos vías principales de transmisión. (1) por yemas para propagación
infectadas con CTV, siendo este el responsable de la dispersión regional. (2) Vectores
del genero Aphis y del genero Toxoptera, siendo los más reconocidos Toxoptera citricida
y Aphis gossypii (Rocha-Peña et al. 1995). Los factores que influencian la dispersión en
campo están relacionados con la densidad de población de áfidos, las condiciones
ambientales que influencian la dinámica en las poblaciones de los vectores, los
aislamientos virales predominantes en campo y la eficiencia de transmisión por cepas
específicas como en el caso T. citricida que es el vector más eficiente de aislamientos
severos (Bar Joseph y Lobestein 1972; Marroquin et al, 2004; Hermoso de Mendoza et
al., 1984).
1.3 Hospedero: Citrus latifolia Tanaka (Lima Tahití)
El centro de origen de Citrus latifolia es desconocido, sin embargo se ha llegado a pensar
que este es un hibrido entre lima mejicana (Citrus aurantifolia) y limón (Citrus limón). Este
cítrico es de porte alto con hojas lanceoladas y pecíolos alados; sus frutos son ovales,
oblongos o elípticos de piel verde y lisa que al madurar se torna amarilla, generalmente
sin semilla, sus flores son ligeramente violetas y el polen es inviable. C. latifolia es
altamente susceptible a CTV, a los aislamientos que inducen decaimiento rápido (QD) y
los que inducen acanaladuras de tallo y ramas (SP). En los llanos orientales de Colombia
los huertos de producción de lima Tahití reportan que estos no superan los ocho años de
producción a consecuencia de la severas epidemias de tristeza de los cítricos (Mateus et
al., 2010).
Históricamente los patrones utilizados para lima Tahití han presentado dificultades por su
susceptibilidad a la enfermedad tristeza de los cítricos y otros patógenos. En Colombia el
patrón más utilizado para lima Tahití es Volkameriana (C. volkameriana Ten. Y Pasq.) no
es susceptible a Phytophtora. sp y se reporta con niveles de tolerancia a CTV. Las

26 Estado del arte.
plantas injertadas sobre este patrón son vigorosas, uno de sus atractivos en lima Tahití
es su precocidad, sin embargo una de las limitantes es el color verde pálido de sus
frutos, que afecta la demanda de exportación. El patrón naranjo amargo (C. aurantium
L.) y C. macrophyla son altamente productivos, pero altamente susceptibles a CTV, por
tanto no son promisorios para zonas endémicas de CTV. En Brasil el limón rangpur (C.
limonia Osbeck. (pro sp.)) es ampliamente usado como patrón para lima Tahití por su alta
resistencia a la sequia, precocidad y alta productividad. El limón rugoso (C. jambhirii
Lush), es susceptible a Phytophthora. sp y no es recomendable como patrón para lima
Tahití. Los patrones tradicionales como limón rugoso (C. jambhirii Lush), naranjo agrio
(C.aurantium L.) y naranjo dulce (C. sinensis [L.] Osb.) han sido poco utilizados en la
citricultura colombiana y hoy día no tienen ningún uso debido a su susceptibilidad a
problemas fitosanitarios, en especial a CTV y a gomosis. El patrón Carrizo (C. sinensis x
P. trifoliata (L.)) es un hibrido intergenérico entre naranja dulce por naranja trifoliada,
tolerante a enfermedades sistémicas, susceptible a Citrus Exocortis Virod, ha mostrado
gran potencial productivo en la zona cafetera de Colombia, no obstante, su
comportamiento como patrón para lima Tahití no está bien determinado. Citrumelo
Swingle (C. paradisi x P. trifoliata (L.) Raf.) es un hibrido intergenérico entre toronja por
naranja trifoliada, tiene buen comportamiento en suelos ácidos para lima Tahití, en Brasil
induce buenas productividades para lima Tahití y en Colombia supera en producción a
algunos patrones injertados con madarina “Arrayana”. El patrón cleopatra (C. reticulata
Blanco.) es susceptible a deficiencias de micronutrientes, produce frutos pequeños y
crece mejor en zonas tropicales que subtropicales, no es prometedor como patrón para
lima Tahití (Campbell, 1979; Campbell 1991, Figueiredo et al 2001) En Colombia
cleopatra ha sido un patrón evaluado para lima Tahtií, los huertos evaluados por
Hernandez y Parrado, 2008, se establecieron en 1997 y han alcanzado de manera
gradual el 100% de incidencia. El síntoma más frecuente encontrado en campo es “stem
pitting”, no obstante en esta evaluación no fue posible corroborar el tipo de cepa presente
en campo. Las cepas tipo VT están asociadas a este tipo síntomas y de acuerdo al
contexto es una cepa ampliamente distribuida y establecida en las zonas de producción
de cítricos en Colombia. No obstante, algunos árboles del huerto empiezan a colapsar a
partir del año 8 de establecimiento que podría ocurrir por los eventos de re-inoculación
del virus por los vectores presentes en campo y el titulo viral, por la interacción entre
varios tipos de cepas de CTV o por la compatibilidad presente entre el patrón cleopatra y
lima Tahití.

Capitulo 1 27
En Venezuela los patrones cleopatra carrizo, volkameriano y citrumelo preliminarmente
fueron catalogados como promisorios para injertarlos sobre Naranja valencia (C. sinensis
(L.) Osbeck.) por su buen comportamiento frente a CTV (Mendt et al., 1988). Aunque
estas respuestas se dieron en una especie cítrica diferente a lima Tahití, las condiciones
de suelos de Venezuela pueden ser homologables a los de algunas regiones de
Colombia, como es el caso de los llanos orientales. Evaluaciones de patrones como
carrizo, limón volkameriano y cleopatra no presentan efectos sobre la productividad y
calidad de los frutos, sin embargo si hay disminución de la longevidad; de las plantas
injertadas sobre limón rugoso colapsan alrededor del año 5 debido a pudriciones de raíz;
las plantas de lima Tahití injertadas sobre mandarina cleopatra colapsan entre los años 5
y 8 probablemente debido a una baja afinidad entre el patrón cleopatra y lima Tahití
(Stenzel y Neves 2004; Figueiredo et al. 2007). No obstante en este último no se tuvo en
cuenta la influencia de CTV sobre la baja longevidad de estos materiales. Una situación
similar es reportada para lima Tahití injertada sobre el patrón cleopatra, en la cual se
reporta que la longevidad se limita hasta el año 8 y se atribuye esta situación a
problemas fitosanitarios como CTV (Quiroga et al., 2010). Las selecciones de Poncirus
trifoliata dentro de las cuales se encuentra el patrón kryder son tolerantes a
enfermedades sistémicas con Citrus Tristeza Virus, Citrus psorosis virus y
Phythophthora; presenta problemas de adaptación a suelos salinos y susceptible a
exocortis, en lima Tahití presenta porte bajo y menor volumen de la copa. Una de sus
seleccines fly dragon (P. trifoliata var monstrosa) es un patrón enanizante que induce
porte bajo en general y se referencia como tolerante a Phytophtora, SP-CTV y
nematodos, pero susceptible a Citrus exocortis viroid (Sanches-Stuchi, 2007). El patrón
Sunki x English es un hibrido entre la naranja trifoliada y la mandarina “Sunki” (P.trifoliata
English x C. sunki), es el patrón con mayor resistencia Phythophthora de 90 patrones
evaluados en el Valle del Cauca en la década de 1960, en Colombia ha alcanzado un
alto reconocimiento por alcnzar altas producciones injertado sobre naranja “Garcia
Valencia” en la zona cafetera y en mandarina “Oneco”; forma plantas de porte medio, alta
calidad de frutos y sanidad de la planta (Orduz-Rodriguez, 2012). Mandarina cleopatra
(Citrus reshi Hort. Ex Tan.) es el patrón cítrico más utilizado e nivel mundial, es poco
precoz para entrada en producción, produce frutos de buena calidad, es tolerante a
algunas cepas de Citrus Tristeza Virus, Citrus psorosis virus, Citrus exocortis viroid.

28 Estado del arte.
1.4 Bibliografía
Albiach-Marti, M R., Mawassi, M., Gowda, S., Satyaranayana, T., Shanker, M.E., Almira,
E.C., Vives. M.C., López, C., Guerri, J., Florez, R., Moreno. P., Garnsey, S., M.,
Dawson, W.O. 2000. Sequences of Citrus Tristeza Virus separated in time and space are
essentially identical. J. Virol. 74, 6856- 6865.
Bar – Joseph y Lee, R. Descriptions of plant viruses. 1989. AAB.
Bar-Joseph, M. Lobestein, G. Cohen, J. (1970) Partial purification of viruslike particles
assiated with the citrus trsiteza virus. Phytopathology 60: 75-78
Bar Joseph.M y Loebenstein. G .1972. Effects os strain, source plant, and temperatura
on the transmissibility of Citrus Tristeza Virus by melón aphid. Phytopathology 63: 716 –
720.
Bar-Joseph, M.; Marcus, R.; Lee, R.F. (1989) The continuous challenge of Citrus Tristeza
Virus control.Annual Review of Phytopathology 27, 291-316.
Campbell, C.W. 1979.Tahití lime production in Florida.University of Florida. Florida
Cooperative. Service,EVA.
Campbell, C.W. 1991. Production of the lime (Citrus latifolia, Tanaka).In florida. University
of Florida. Tropical Region 2:184-192.
CMI (1978) Distribution Maps of Plant Diseases No. 289 (edition 5). CAB International,
Wallingford, UK.
Davino, S., Davino, M., Sambade, A., Guardo, M. and Caruso, A.(2003) The first Citrus
Tristeza Virus outbreak found in a relevant citrus producing area of Sicily, Italy. Plant Dis.
87, 314.
Ecale Zhou, C.L., Ammar, E. D., Sheta, H., Kelley, S., Polek, M., Ullman, D.E. 2002
Citrus Tristeza Virus ultraestructure and associated cytopathology in Citrus sinensis and
Citrus aurantifolia Canadian Journal of Botany 80:512-525
Fraser, L. (1952) Seedling yellows, an unreported virus disease of citrus. Agricultural
Gazette of New South Wales 63, 125-131.
Figueiredo, J.O., Stuchi, E.S., Laranjeira, F.F., Donadio, l.C., Teófilo-Sobrinho, J.,
Sempionato, O.R. y Muller, G.W. 2001. Porta-enxertos para lima ácida ‘Tahití ’ em duas
regiões do Estado de São Paulo. Laranja, v.22, n.1, p.203- 213.

Capitulo 1 29
Garnsey, S. M., D. J. Gumpf, C. N. Roistacher, E. L. Civerolo, R. F. Lee, R. K. Yokomi,
and M. BarJoseph 1987. Toward a standardized evaluation of the biological properties of
Citrus Tristeza Virus.Phytophylactica 19: 151-157.
Garnsey, S.M., Gottwald, T.R., Hilf, M.E., Matos, L. and Borbón, J. (2000) Emergence
and spread of severe strains of Citrus Tristeza Virus isolates in the Dominican Republic.
In: Proceedings of the 14th Conference of the International Organization of Citrus
Virologists (da Graça, J.V., Lee, R.F. and Yokomi, R. K., eds), pp. 57–68. Riverside, CA:
IOCV.
Granof, A., y Webster, R. 1999. Encyclopedia of Virology. Second Edition, Vol
2.Academic Press.
Ghorbel, R., Lopez, C., Fagoaga, C., Moreno, P., Navarro, L., Flores, R., y Peña, L. 2001.
Transgenic citris expressing the citris tristeza virus p23 protein exhibit viral-like symptons.
Molecular plant pathology (2001) 2(1), 27 – 36
Hermoso de Mendoza, A., Ballester-Olmos, J.F. and Pina, J.A. (1984) Transmission of Citrus Tristeza Virus by aphids (Homoptera, Aphididae) in Spain. In: Proceedings of the 9th Conference of the International Organization of Citrus Virologists (Garnsey, S.M., Timmer, L.W. and Dodds, J.A., eds), pp. 23–27. Riverside, CA: IOCV. Hernandez, F., y Quiroga, J. 2008. Análisis de la influencia del virus de la tristeza de los
citricos CTV, en copas de lima tahití (citrus latifolia Tanaka), injertadas sobre patrón de m
andarina cleopatra (citrus reshni horth. ex tan), en Corpoica, La Libertad. Tesis. Escuela
de Ingeniaría Agronómica. Universidad de los llanos. Villavicencio – Colombia.
Hilf, M. E and Garnsey, S. M. 2000.Characterization and Classification of Citrus Tristeza
Virus Isolates by Amplification of Multiple Molecular Markers. Fourteenth IOCV
Conference, 2000—Citrus Tristeza Virus
Hilf, M.E., Karasev, A.V., Pappu, H.R., Gumpf, D.J., Niblett, C.L. and Garnsey, S.M.
1995. Characterization of citrus tristeza virus subgenomic RNAs in infected tissue.
Virology, 208, 576–582.
Hilf, M. E., Mavrodieva, V. A., and Garnsey, S. M. 2005. Genetic marker analysis of a
global collection of isolates of Citrus risteza virus:Characterization and distribution of CTV
genotypes and association withsymptoms. Phytopathology 95:909-917.
Hughes, W.A. y Lister, C.A. (1949) Lime disease in the Gold Coast. Nature 164, 880
Karasev AV, Boyko VP, Gowda S, Nikolaeva OV, Hilf ME, Koonin EV, Nibblet CL, Cline
K,Gumpf DJ, Lee RF, Garnsey SM and Dawson WO (1995) Complete sequence of the
Citrus Tristeza Virus RNA genome. Virology 208, 511–520.
Karasev, A.V. (2000). Genetic diversity and evolution of closteroviruses. Annu. Rev.

30 Estado del arte.
Phytopathol.38, 293-324.
Laliberté, J.F., Sanfacon, H. 2010 Cellular remoldeling during plant virus infection. Annu.
Rev. Phytopathol. 48:69-9.
Lu, R., Folimonov.. M., Shinatahu. M., Li, W., Dawson., Ding .S. 2004. Proc. Natl. Acad.
Mateus, D., Pulido, X., Gutierrez, A., y Orduz-Rodriguez, J.O. 2010. Evaluación
económica de la producción de cítricos cultivados en el Piedemonte del Departamento
del Meta durante 12 años. Orinoquia [online]. 2010, vol.14, n.1.
Marroquin, C., Olmos, A., Gorris, M.T., Bertolini, E., Martinez, C., Carbonell, E.A.,
Hermoso de Mendoza, A., and Cambra, M. (2004). Estimation of the number of aphids
carryng Citrus Tristeza Virus that visit adult citrus trees. Virus Research.
Mawassi, M., Mietkiewska, E., Gofman, R., Yang, G., and Bar-Joseph, M. 1996.Unusual sequence relationships between two isolates of Citrus Tristeza Virus. J. Gen. Virol. 77:2359-2364.
Mendt, R., F. Ochoa, L. Vilalba, H. Uhlig, G. Perez, A. Cedeno, and T. Barreto 1988.
Evaluation of citrus tristeza virus tolerant rootstocks grafted with Valencia orange in
Venezuela, p. 107-112. In: Proc. 10th Conf. IOCV., IOCV, Riverside.
Moreno, P., Ambros, S., Albiach-Marti, M.R., Guerri, J., Pena, L., 2008. Citrus
tristezavirus: a pathogen that changed the course of the citrus industry. Mol. Plant
Pathol.9, 251–268.
Navas – Castillo, J., Albiach-Marti, M.R., Godwa, S., Hilf, M.E., Garsney, SM., y Dawson,
W.O. (1997) Kinetics of accumulation of citrus trsiteza virus RNAs. Virology , 199, 25 –
46.
Oberholzer, P.C.J.; Mattews, I.; Stiemie, S.F. (1949) The decline of grapefruit trees in
South Africa. A preliminary report on so-called "stem pitting". Science Bulletin of the
Department of Agriculture South Africa No. 297.
Orduz-Rodriguez, J.O. 2012. Manual para el Cultivo de Frutales en el Trópico. Capitulo
cítricos. Editorial Produmedios
Pappu, S.S., Febres, V.V., Pappu, H.R., Lee, R.F. and Niblett, C.L. (1997)
Characterization of the 3 proximal gene of the citrus tristeza closterovirus genome. Virus
Res. 47, 51–57.
Quiroga - Cardona, Julio., Hernández- Parrado, F. L., Silva- Herrera, M., Orduz-
Rodríguez, J.O. 2010. Comportamiento de la producción de lima Tahití (Citrus latifolia
Tanaka), injertada sobre el patrón de Mandarina Cleopatra (Citrus reticulata Blanco) y la

Capitulo 1 31
influencia del virus de la tristeza (CTV) en condiciones del piedemonte del Meta, 1997-
2008. Orinoquia, vol. 14, núm. 1, junio, 2010, pp. 5-15
Rodriguez, P., De Perez, G., Guzman, M., 2009. Detección del virus de la tristeza de los
cítricos por serología, microscopía e hibridación in situ. Revista Colombiana De
Biotecnología ISSN: 0123-3475 ed: Instituto De BiotecnologiaIbunv.11 fasc.1 p.94 –
106.
Rocha-Peña, M.A., Lee, R.F., Lastra, R., Niblett, C.L., Ochoa, F.M., Garnsey, S.M., and
Yokomi, R.K. (1995). Citrus Tristeza Virus and its aphid vector Toxóptera citricida:
Threats to citrus production. Plant Disease 79, 437-445.
Ruiz-Ruiz, S., Moreno, P., Guerri, J., and Ambrós, S. 2009. Discrimination between mild and severe Citrus Tristeza Virus isolates with a rapid and highly specific real-time reverse transcription-polymerase chain reaction method using TaqMan LNA probes. Phytopathology 99:307-315.
Sambade, A., Lopez, C., Rubio, L., Flores, R., Guerri, J., Moreno, G. 2003.
Polymorphism of a specific región in gen p23 of Citrus Tristeza Virus allows
discrimination between mild ans severe isolates. Arch Virol 148: 2325 – 2340.
Sanches-Stuchi, E. 2007. Trifoliata Flying Dragon: Um excelente porta-enxerto =ara limão
Tahit. Em memória Congreso Embrapa Mandioca e fruticultura Tropipcal-Estacao
experimental de citricultura de Bebedouro. :8-10.
Stenzel, Neusa. M.C., y Neves. C. (2004) Rootstocks for 'Tahití ' lime. Sci. agric.
(Piracicaba, Braz.) [online]. 2004, vol.61, n.2, pp. 151-155. ISSN 0103-
9016. http://dx.doi.org/10.1590/S0103-90162004000200005
Suastika, G., Natsuaki, T., Terui, H., Kano, T., Ieki, H. and Okuda, S.(2001) Nucleotide
sequence of Citrus Tristeza Virus seedling yellows isolate. J. Gen. Plant Pathol. 67, 73–
77.
Tatineni, S., Robertson, C., Garnsey, S., y Dawson, W. 2011. A plant virus evolved by
acquiring multiple nonconserved genes to extend its host range. Proc Natl Acad Sci U S
A. 2011 Oct 18;108(42):17366-71. Epub 2011 Oct 10.
Vives, M.C., Rubio, L., López,C., Navas-Castillo, J., Albiach-Marti, M.R., Dawson, W.O.
Guerri, J., Florez , R., Moreno, P. (1999). The complete genome sequence of the major
component of a mild Citrus Tristeza Virus isolate.J. Gen. Virol. 80, 811-816.
Webber, H.J. 1943.The Tristeza Disease of Sour-Orange-Rootstock. Proc.
Am.Soc.Hort.Sci. 43:160-168.

32 Estado del arte.
Yang, Z,N., Mathews, D.M., Dodds, J.A., Mirkov, T.E. (1999). Molecular characterization
of an isolate of Citrus Tristeza Virus that causes severe symptoms in sweet orange.Virus
Genes 19, 131-142.


2. Identificación de cepas de Citrus Tristeza Virus mediante marcadores moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
Resumen
Citrus Tristeza Virus (CTV) es causa de la enfermedad viral más deletérea para la
producción de cítricos a nivel mundial, y usualmente está presente en campo como un
complejo de asilamientos. En este trabajo se identificaron cepas de CTV en dos parcelas
experimentales de lima Tahití generadas por micro-injerto de ápices caulinares in vitro
sobre seis patrones diferentes. Las parcelas se establecieron en los centros de
investigación de Corpoica Nataima (Espinal – Tolima - Colombia) y La Libertad
(Villavicencio – Meta – Colombia). El material se evaluó después de dos años de
crecimiento en campo. Se empleó RT-PCR mediante primers del gen de la polimerasa
que permiten la identificación de los genotipos específicos VT, T30 y T36, y primers del
gen p23 para discriminar cepas de los grupos suaves y severos. El genotipo VT fue de
mayor incidencia en las dos localidades, mientras que el genotipo T30 se encontró en las
dos localidades pero con menor incidencia en La Libertad. El genotipo T36 se encontró
únicamente en la Libertad. Se identificaron plantas con infecciones mixtas VT/·T30 y
VT/T30/T36. Las secuencias nucleotídicas del gen POL corroboraron los resultados
obtenidos por RT-PCR, así los aislamientos colombianos T36 presentaron 99% de
identidad con la secuencia del genoma de T36 registrada en el GeneBank. Sin embargo,
se sugiere corroborar la presencia de este genotipo mediante el uso de marcadores
moleculares de otros genes de CTV.
Abstract
Citrus Tristeza Virus (CTV) is the most damaging virus of citrus in the worldwide. Usually,
it is present in field trees as a complex of isolates. In this study, CTV isolates were

Capitulo 2 35
identified in two experimental tahitían lime plots produced by micro-graffting of caulinar
budwood in vitro on six different rootstocks. Plots were established in research centers of
Nataima (Espinal – Tolima – Colombia) and La Libertad (Villavicencio – Meta –
Colombia).The material was evaluated after two years of established crops. RT-PCR was
done using gene polymerase specific molecular markers to identify genotypes VT, T30
and T36 and gene p23 markers to identify mild and severe isolates. Isolates of severe
genotype VT exhibited de higher incidence in the two localities, mean while the isolates of
the mild genotype T30 were found in two localities with less incidence in La Libertad.
Severe genotype T36 was found in La Libertad. Mixed infections VT/T30 and VT/T30/T36
were identified. Nucleotide sequences of gene polymerase of isolates identified as
genotypes VT, T30 and T36 corroborated the obtained results by RT-PCR. Isolates with
genotype T36 showed 99% identity with the genome sequence T36. However, this result
must be confirmed using molecular markers for other genes of CTV.
2.1 Introducción
El agente causal de la tristeza de los cítricos es Citrus Tristeza Virus (CTV), se encuentra
establecido en todas las áreas de producción y es considerado como el virus más
limitante en la producción mundial de cítricos.CTV pertenece al género Closterovirus de
la familia Closteroviridae. Las partículas virales son filamentos flexuosos de 10 a 11 nm
de diámetro y 1900 a 2000 nm de longitud, y exhiben una arquitectura particular
“rattlesnake” por el ensamblaje de dos proteínas de capside, CP de 25kD y CPm de 27kD
(Febres et al., 1996). El genoma de CTV es RNA de cadena sencilla con sentido positivo
(Bar Joseph et al1972; Bar Joseph y Lee, 1989).Existen una amplia gama de técnicas
para detección de CTV, siendo las más frecuentes ELISA, RT–PCR y RT–qPCR.
Mediante ELISA se utilizan anticuerpos monoclónales y policlónales para discriminar
entre cepas suaves y severas (Garnsey et al., 1978; Gonsalves, et al., 1978; Peñaranda
et al., 1996). Las técnicas moleculares emplean el uso de marcadores específicos para
discriminar el genotipo de las diferentes cepas de CTV. Algunos trabajos han conjugado
la aplicación de técnicas de imnunoimpresión, serológicas y moleculares para el rápido
diagnóstico de CTV (Rodriguez et al, 2009).
La variabilidad en la expresión de síntomas es utilizada para diferenciar cepas de CTV.
Las cepas suaves hacen referencia a que solo causan síntomas suaves o son

36 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
asintomáticos en el hospedero cítrico indicador. Las cepas severas pueden causar
síntomas de “quick decline”, “stem pitting” o ambos, estos pueden variar en la intensidad
(Garnesey et al., 1987).
El RNA genómico (gRNA) de CTV se encuentra compuesto por 12 marcos de lectura
abiertos (ORFs) y regiones no traducibles (UTRs) en las terminaciones 5' y 3'. El primer
ORF se encuentra divido por dos regiones 1a y 1b. 1a codifica replicasas. 1b codifica
RNA polimerasa dependiente de RNA. Estas dos ORFs abarcan la mitad 5' del genoma y
son traducidos directamente por el gRNA. En la mitad 3' se encuentran ubicados 10 ORF
que codifican las proteínas p33 (ORF2), p6 (ORF 3), p65– hHSP70 (ORF 4), p61 (ORF
5), p27- CPm (ORF 6), p25 – CP (ORF 7), p18 (ORF 8), p13 (ORF 9), p20 (ORF 10) y
p23 (ORF 11). p65 y p62 son homologas a proteínas de choque térmico y junto con las
proteínas de la capside p25 y p27 están involucradas con el ensamblaje del virion. p20 es
asociada con la acumulación de cuerpos de inclusión en las células infectadas. La
función de p33, p18 y p13 se relaciona con la capacidad de inducir infección sistémica en
el rango de cítricos hospederos (Tatineni et al., 2011) y se ha demostrado que no son
necesarias para la replicación y ensamblaje de los viriones (Satyanarayana et al., 1999,
2000). Los ORFs del extremo 3´ de CTV se expresan a través de un set de RNAs
mensajeros 3´- coterminales llamados RNAs subgenómicos (Hilf et al., 1995). Una alta
variabilidad ha sido encontrada en CTV, debido a que la RNA polimerasa dependiente de
RNA - dRpR no posee actividad exonucleasa para corrección de errores en la replicación
y a eventos de recombinación (Bar-joseph et al., 1893; Moreno et a., 2008; Cerni et al.,
2007). Análisis de secuencias de nucleótidos muestran que el genoma de CTV es
conservado en los ORF cercanos al extremo 3′ con una identidad del 90% entre
aislamientos y con mayor divergencia en los ORFs 1a y 1b cercanos al extremo 5 ′con
una identidad del 40% entre aislamientos (Albiachi et al., 2000; López et al., 1998;
Mawassi et al.,1996).
La detección de un genotipo especifico de CTV es posible mediante PCR con primers
diseñados a partir de variantes nucleotídicas del gen RNA polimerasa (Hilf et al., 2005). A
partir de este método de detección, CTV ha sido clasificados en tres grupos principales:
VT (B370, B152, B1, B79, B59, B219) que generan síntomas de acanaladuras de tallo y
ramas, T30 (B213, B348, B271, B270-1) que generan síntomas suaves, y T36 (B83, B33,

Capitulo 2 37
T36) que generan decaimiento rápido (Hilf et al., 1999; Hilf y Garnsney 2000; Hilf et al.,
2005). Por otro lado marcadores específicos diseñados a partir del polimorfismo del gen
p23 permiten discriminar genotipos severos (VT, Barão B, T 305, T388), suaves (T300,
T312, T32, entre otros) y atípicos (T36, Galego 50, K, entre otros) (Sambade et al.,
2003).
Trabajos de caracterización de cepas colombianas de CTV reportan la presencia de
suaves y severas. Las cepas B126 y B165 de Colombia están relacionadas con
aislamientos que generan acanaladuras de tallo y en análisis filogenéticos muestran
relación con aislamientos de India y Japón. Las cepas B272 y B274 están asociadas con
aislamientos suaves T4 y T30 de Florida. La comparación de secuencias del gen CP de
algunas cepas colombianas con la secuencia del aislamiento T36 indicó rangos de
identidad de 93% a 97% (Peñaranda et al., 1996). Este estudio preliminar sugirió la
presencia de cepas T36 en cultivos colombianos, sin embargo esta región del genoma de
CTV es altamente conservada y es necesario evaluar otros marcadores moleculares para
CTV que proporcionen mayor información. Los trabajos de detección en Colombia
indican una amplia distribución y una alta incidencia de CTV en general. El uso de
técnicas serológicas ó moleculares para discriminar cepas suaves de severas, han
determinado una amplia distribución de cepas severas que generan acanaladuras de
tallo (Peñaranda et al., 1996; Oliveros et al., 2001; Delgado et at., 2004; Martínez y
Guzmán 2007), la distribución de cepas suaves es menos amplia y se reportan
principalmente en Mompóx (Peñaranda et al., 1996). El objetivo del presente trabajo fue
identificar las cepas de CTV que infectan dos poblaciones de lima Tahití ubicadas en dos
posiciones geográficas diferentes.
2.2 Materiales y métodos
La muestras se colectaron del centro de investigación Corpoica Nataima, en el Espinal
(Tolima – Colombia) ubicado en las coordenadas 4° 12’ 56’’ N y 72° 56’ 3’’ O, a 430
msnm, con una temperatura media de 28 ºC y una humedad relativa del 70% y del centro
de investigación Corpoica La Libertad a 2 Km de Villavicencio (Meta –Colombia) ubicado
en las coordenadas 9º 6' N, 73º 34' O, a 320 msnm, con una temperatura media de 27 ºC
y humedad relativa entre el 70 y el 90%. Cada una de las poblaciones de lima Tahití se
encontraba injertada sobre patrones de seis especies o cultivares híbridos: volkameriano

38 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
(C. volkameriana Ten. Y Pasq.), citrumelo swingle (C. paradisi x P. trifoliata (L.) Raf.),
mandarina cleopatra (C. reticulata Blanco.), sunky x english (Citrus sunki Hort. ex Tan. x
Poncirus trifoliata (L.)) , kryder (Poncirius trifoliata L Raf), y carrizo (C. sinensis x P.
trifoliata (L.)). En campo, cada especie de injerto se distribuyó en grupos de seis plantas
y este arreglo se repitió cuatro veces. Las muestras se rotularon de acuerdo al patrón,
numero de repetición y numero de planta. Se procesaron de la siguiente manera: i) Se
tomaron nervaduras de hojas jóvenes y corteza de ramas con tejido verde en los cuatro
puntos cardinales de cada una de las plantas. ii) Se homogenizó en una sola muestra
cada grupo de seis plantas injertadas sobre un mismo patrón.iii) El material vegetal se
macero en nitrógeno liquido iv) Cada grupo de plantas se proceso por RT – PCR. iv) En
los grupos de plantas en los que se detectaron cepas de interés ó varias cepas, se
procesó independientemente cada planta (ver figura 1)
Figura 2-1: Esquema de toma de muestras y flujo de decisión para seleccionar plantas a evaluar
El genotipo de cada uno de las cepas de CTV se determinó por RT – PCR mediante el
uso de parejas primers genotipo específicos para VT, T30 y T36. Los productos RT-PCR
se visualizaron por electroforesis en gel de agarosa al 2% teñido con bromuro de etidio.
Los productos de RT – PCR fueron secuenciados y las secuencias obtenidas se
compararon con las registradas en el GeneBank. La RT – PCR se realizó en un
termociclador BIORAD C1000.

Capitulo 2 39
2.2.1Extracción de ARN, transcripción reversa (RT) y reacción en cadena de la polimerasa (PCR).
La extracción de ARN se realizó mediante el protocolo Trizol® (Invitrogen). Este consistió
en tres fases: la primera rompimiento de paredes celulares con Trizol®, separación con
cloroformo y gradientes de densidad por centrifugación. La segunda fase fue de
precipitación de ácidos nucleícos mediante el uso de alcohol isopropílico y gradientes de
densidad por centrifugación. La tercera fase consistió en el lavado del pellet de ARN con
etanol al 75%. Posteriormente se re suspendió el pellet en 50μl de agua grado molecular
tratada con dietil pirocarbonato (DEPC).
Transcripción reversa
Se realizó en un volumen de 25μl que contenía 5μl de RNA, 1μl del primer ramdom
hexamers (Invitrogen), 1μl de dNTPs 10mM (Invitrogen) y 8μl de agua DEPC, 2μl de
Buffer first strand M-MLV 10X (Invitrogen), 2μl de DTT 100mM (Invitrogen), 1μl RNAasa
Out 40U (Invitrogen), 1μl de M-MLV-RT (Invitrogen) y 4μl de MgCl2 25mM (Invitrogen). El
RNA se denaturó a 65 ºC por 5 minutos. La reacción se incubó 10 minutos a 25 ºC, se
sintetizó por 90 minutos a 37 ºC, y se finalizó 10 minutos a 72 ºC.
PCR Convencional
La PCR convencional se realizó con los primers de la región polimerasa en reacciones de
25μl. Cada una contenía 2.5μl de Buffer taq polimerasa 10X sin MgCl2 (Invitrogen), 1μl de
dNTPs 10 mM (Invitrogen), 0.25μl 10 mM primer sentido, 0.25μl 10 mM anti – sentido de
los primers CP, VTPOL, T30POL y T36POL según el caso; 1.5μl de MgCl2 50mM
(Invitrogen), 5μl de producto de RT. 0.4μl de Taq polimerasa 5U/μl (Invitrogen) y 14.1μl
de agua DEPC. El programa de amplificación consistió en una temperatura de
denaturación inicial por 5 minutos a 94 ºC, posteriormente 35 ciclos a 94 ºC por 30
segundos, 65 ºC por 30 segundos y 72 ºC por 60 segundos. Para finalizar una
temperatura de extensión a 72 ºC por 10 minutos.
La amplificación con primers CP indica la presencia general de CTV y es un control
interno de la transcripción reversa. Los primers VT-POL producen un amplicon de 695pb
e indica la presencia de genotipos severos VT que ocasionan acanaladuras de tallo en
lima Tahití. Los primers T30-POL producen un amplicon de 696pb e indica la presencia

40 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
de genotipos que ocasionan síntomas suaves en lima Tahití. Mientras que los primers
T36-POL permiten la amplificación de un amplicon de 714pb e indica la presencia de
cepas T36 que ocasionan decaimiento rápido en cítricos injertados sobre naranjo agrio y
lima Tahití (Hilf et al. 2005)
PCR bidireccional
Esta PCR se efectuó mediante el uso de primers del ORF p23. Se utilizaron dos parejas
de primers; una pareja externa y una interna. La reacción se realizó a 25μl. Cada una
contenía 2.5μl de Buffer taq polimerasa 10X sin MgCl2 (Invitrogen), 1μl de dNTPs 10 mM
(Invitrogen), Los primers externos (0.25μl 10 mM primer pm85 (+), 0.25μl 10 mM primer
pm86 (-)), los primers internos (0.25μl 10 mM primer pm82 (+), 0.25μl 10 mM primer
pm83 (-)), 1.5μl de MgCl2 50mM (Invitrogen), 5μl de producto de RT. 0.4μl de taq
polimerasa 5U/μl (Invitrogen) y 14.1μl de agua DEPC. . El programa de amplificación
consistió en una temperatura de denaturación inicial por 94 ºC por 120 segundos,
posteriormente 35 ciclos de 94 ºC por 30 segundos, 50 ºC por 30 segundos y 72 ºC por
60 segundos. Para finalizar una temperatura de extensión a 72 ºC por 120 segundos.
Los primer externos están basados en secuencias conservadas y los internos son
derivados de secuencias de grupos severos, suaves y atípicos. Los grupos suaves (T32,
T35, T300, T312, T346 y T385) producen amplicones de 612 y 239 a la vez. Los grupos
severos ( VT y Barão B) producen amplicones de 612 y 450 pb a la vez; otras cepas del
grupo severo (T305, T388, C269-6, Cald-CB y Val-CB) producen adicionalmente un
fragmento de 239 pb. Los grupos atípicos (T36 y K ) producen un solo amplicon de 612
pb. (Sambade et al. 2003)
Primer Secuencia
VTPOL (+) gacgctagcgatggtcaagc
VTPOL (-) ctcggctcgctttcttacgt
T36POL (+) atggacgacgaaacaaagaaattg
T36POL (-) tcaacgtgtgttgaatttccca
T30POL (+) gatgctagcgatggtcaaat
T30POL (-) ctcagctcgctttctcacat
P23_PM82 aaacacgataaggcatcgag
P23_PM83 cacttacgttcagtcttgagcg
P23_PM85 ggacaaactttiitttctgtgaacctttc
P23_PM86 gatgaagtggtgttcacggagaactc
Tabla 2-1: Primers de utilizados para detección de CTV. Tomado de Hilf et al., 2005 y Sambade et al., 2003

Capitulo 2 41
Secuenciación
Los productos de PCR previamente purificados fueron secuenciados en un secuenciador
ABI3730 XL por el método Byg Dye Terminator. Dos productos de PCR obtenidos con los
primers utilizados de la región de polimerasa para cada una de las poblaciones
evaluadas, fueron seleccionados para su secuenciación.
2.3 Resultados
Todos los grupos de muestras evaluados mediante PCR con primers del gen CP
indicaron infección con CTV. En el 100% de las muestras se observaron amplicones
672pb que corresponden al tamaño esperado para este amplicon. Lo que indicó que por
lo menos una de las seis plantas que conforman cada grupo, estaba infectada con CTV.
En la población del huerto de cítrios de La Libertad (Villavicencio – Meta – Colombia) se
encontró una alta incidencia de cepas VT respecto a las cepas T30 y T36. El 100% de los
grupos de plantas evaluadas estaban infectadas con cepas VT. El 8 % de los grupos de
plantas evaluadas con el marcador estaban infectadas con cepas T30. El 13 % de los
grupos de plantas evaluadas con cepas T36. Lo anterior constituye evidencia de
infecciones mixtas VT/T30/T36 y VT/T30, las cuales fueron detectadas en los grupos de
platas injertadas sobre los patrones Volkameriano RI, Cleopatra RIII, cleopatra RIV y
carrizo RIV. Este resultado, llevó a evaluar la infección de estos grupos planta a planta.
En la revaluación planta a planta se encontró que todas las plantas estaban infectadas
con CTV, y de igual manera se encontró alta incidencia de cepas VT respecto a cepas
T30 y T36. El 100% de las plantas estaban infectadas con cepas VT. Se identificaron
plantas con infecciones mixtas, el 27% (6 plantas) tenían infección mixta entre cepas
VT/T30, el 34% (8 plantas) de las plantas se encontraban infectadas con mezcla VT/T36
y el 4% (1 planta) tenía infección mixta entre cepas VT/T30/T36. El 35% de plantas
restantes estaba infectado únicamente con cepas VT.

42 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
Figura 2-2: Electroforesis en gel de agarosa para visualizar amplicones de VT-POL deCTV (695 pb) de las muestras de lima Tahití colectadas en el huerto Corpoica La Libertad. Las filas A y B muestran los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón. Fila A: Carril 1: Marcador de peso molecular 100 bp (Fermentas). Carril 2 a 5: Kryder RI a RIV. Carril 6 a 9: Volkameriano RI a RIV. Carril 10 a 13: Sunky x English RI a RIV. Fila B: Carril 1: Marcador de peso molecular 100 bp (Fermentas). Carril 2 a 5: Citrumelo RI a RIV. Carril 6 a 9: Cleopatra RI a RIV. Carril 10 a 13: Carrizo RI a RIV. Fila C: Productos de PCR de cada planta de un pool de seis plantas. Carril 1: Marcador de peso molecular 100 bp (Fermentas). Carril 2: Control positivo Carril 3: Blanco de reacción.Carril 4 a 8: Cleopatra RIV plantas 1 a 5. Carril 9 a 14: Cleopatra RIII plantas 1 a 6. Carril 15 a 20: Volkameriano RI plantas 1 a 6. Carril 21 a 24: Carrizo RIV plantas 1 a 4.
Figura 2-3: Electroforesis en gel de agarosa para visualizar amplicones T30 POL de CTV (696 pb) de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La Libertad. Las filas Ay B muestran los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón. Fila A: Carril 1: Marcador de peso molecular Fermentas 100pb. Carril 2 a 5: Kryder RI a RIV. Carril 6 a 9: Volkameriano RI a RIV. Carril 10 a 13: Sunky x English RI a RIV. Fila B: Carril 1: Marcador de peso molecular Fermentas. Carril 2 a 5: CitrumeloRI a RIV. Carril 6 a 9: Cleopatra RI a RIV. Carril 10 a 13: Carrizo RI a RIV.Fila C: Las filas Ay B muestran los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón Carril 1: Marcador de peso molecular Fermentas 100pb. Carril 2: Control positivo Carril 3: Blanco de reacción .Carril 4 a 9: Cleopatra RIII platas 1 a 6. Carril 10 a 15:Volkameriano RI Plantas 1 a 6

Capitulo 2 43
Figura 2-4: Electroforesis en gel de agarosa para visualizar amplicones T36 POL de CTV (714 pb) de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La Libertad. La fila A muestra los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón. Fila A: Carril 1: Marcador de peso molecular 100 pb (Fermentas). Carril 2 a 5: Citrumelo RI a RIV. Carril 6 a 9: Cleopatra RI a RIV. Carril 10 a 13:Cariizo RI a RIV. Carril 14 a 17: Sunky x English RI a RIV. Carril 18 a 19:Cleopatra RI a RII. Las filas B y C muestran productos de PCR de cada planta de un pool de seis plantas Fila B. Carril 1: Marcador de peso molecular 100pb (Fermentas). Carril 2: Control positivo Carril 3: Blanco de reacción Carril 4 a 9: Cleopatra RIII plantas 1 a 6. Carril 10 a 14: Cleopatra RIV plantas 1 a 5. Fila C. Carril 1: Marcador de peso molecular Fermentas. Carril 2 a 7: Carrizo RIV plata 1 a 6
Cada grupo de muestras se evaluó nuevamente con los primers de la región p23. Se
visualizaron amplicones de 612pb y 412pb en el 96% de las muestras, indicando que
estos grupos de plantas estaban infectados con cepas severas que podrían ser tipo VT.
El 4% de las muestras restantes produjo adicionalmente una banda de 239pb, lo que
indicaría que estos grupos de muestras podrían estar infectados en mezcla de cepas
severas y suaves. Estas dos bandas adicionales se identificaron en los grupos de plantas
volkameriano RI y cleopatra RIII. Se revaluaron estos grupos planta a planta más los
grupos cleopatra RIV y carrizo RIV en donde no se había identificado cepas suaves con
los primers de la región de polimerasa.
En la revaluación planta a planta en el 73% de las muestras se visualizaron únicamente
amplicones de 612pb y 412pb, lo que indica que estas plantas estaban infectadas con
asilamientos severos presumiblemente VT. En el 27% de las muestras se observó un
amplicon adicional de 239pb, lo que indicó que estas plantas se encontraban infectadas
con mezcla de cepas suaves y severas, esto en correspondencia a lo encontrado con los
primers de la región polimerasa.

44 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
Figura 2-5: Electroforesis en gel de agarosa para visualizar amplicones del gen P23 de CTV de las muestras de lima Tahití colectadas en el huerto ubicado en Corpoica La Libertad. Las filas Ay B muestran los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón. Fila A: Carril 1: Marcador de peso molecular 100 pb (Fermentas) Carril 2 a 5: Kryder RI a RIV. Carril 6 a 9: Volkameriano RI a RIV. Carril 10 a 13: Sunky x English RI a RIV. Carril 14 a 15: Citrumelo RI a RII. Fila B: Carril 1: Marcador de peso molecular 100 pb (Fermentas). Carril 2 a 3: Citrumelo RIII a RIV. Carril 4 a 7: Cleopatra RI a RIV. Carril 8 a 11: Cariizo RI a RIV. Fila C: Productos de PCR de cada planta de un pool de seis plantas. Carril 1: Marcador de peso molecular 100pb (Fermentas). Carril 2 a 7: Cleopatra RIII Planta 1 a 6. Carril 8 a 12: Cleopatra RIV Planta 1 a 5. Carril 13 a 18: Volkameriano RI Planta 1 a 6. Carril 19 a 24: Carrizo RIV planta 1 a 6.
En la población del huerto de cítricos de Nataima (Espinal –Tolima – Colombia), todos los
grupos de injertos evaluados presentaron infección por CTV y se detectaron únicamente
cepas VT y T30. La incidencia de cepas VT fue mayor respecto a la incidencia de cepas
T30. El 50% de los grupos de plantas estaban infectadas únicamente con cepas VT y el
50% estaba infectado con mezcla de cepas VT/T30. Se identificaron cuatro grupos de
plantas con mezcla de cepas VT y T30, se escogieron los grupos de plantas
volkameriano RI y kryder RI para evaluar planta a planta. En esta evaluación de cada
planta se encontró que el 66% de las plantas se encontraban infectadas únicamente con
cepas VT. El 34% de la plantas presentó infecciones mixtas entre cepas VT/T30. Al
evaluar el grupo de muestras mediante PCR del gen p23, se encontró que el 54%
amplificaron únicamente bandas de 612pb y 412pb indicando la presencia de cepas VT,
el 46 % amplifico una banda adicional de 239 pb que indica la mezcla de cepas suaves y
severas en estos grupos de plantas. Se escogieron dos grupos para evaluarlos planta a
planta (kryder RI y Volkameriano RI), se identificó que el 33% de las plantas tenían
infección mixta entre cepas suaves y severas, frente al 67% restante que indico infección
únicamente con aislamientos suaves.
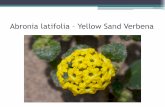
Capitulo 2 45
Figura 2-6: Electroforesis en gel de agarosa para visualizar amplicones de CTV (p23) de las muestras de lima tahti colectadas en el huerto ubicado en Corpoica Nataima. Las filas Ay B muestran los productos de PCR de “pools” de seis plantas injertadas sobre cada patrón. Fila A: Carril 1: Marcador de peso molecular 100 pb (Fermentas). Carril 2 a 5: Cirumelo RI a RIV. Carril 6 a 9: Sunky x English RI a RIV. Carril 10 a 13: Carrizo RI a RIV. Fila B:Carril 1: Marcador de peso molecular 100 pb Fermentas. Carril 2 a 5: Volkameriano RI a RIV.Carril 6 a 9: Kryder RI a RIV. Carril 10 a 13: Cleopatra RI a RIV. Fila C: Productos de PCR de cada planta de un pool de seis plantas Carril 1: Marcador de peso molecular 100 pb (Fermentas). Carril 2 a 7: Volkameriano RI plantas 1 a 6. Carril 8 a 13: Kryder RI Plantas 1 a 6.
La secuenciación directa de los productos de PCR de los grupos severo VT y T36 y del
grupo suave T30 reveló una alta identidad con los respectivos aislamientos. Presentaron
porcentajes de identidad del 99% y porcentajes de cobertura entre el 99 y 100% con los
aislamientos registrados en el Genbank (Tabla 2).
Origen
Origen
Material Muestra Identidad
con:
:genotipo
Accesión Identidad%
(%)
Cobertura%
(%)
Gaps
La Libertad ClEORIVP5 T36_001 T36 E.U937521,1 99 100 0
La Libertad CLEORIIIP3 T36_002 T36 E.U937521,1 99 99 0
La Libertad VOLRIP6 T30_003 T30 AF260651,1 99 100 0
La Libertad CLEORIIIP6 T30_004 T30 AF260651,1 99 100 0
La Libertad VOLRIPI VT_005 VT EU262674,1 99 99 0
La Libertad CLEORIVP5 VT_006 VT EU262674,1 99 100 0
Nataima VOLRIP3 VT_007 VT AY295898,1 99 100 0
Nataima VOLRIP4 VT_008 VT AY295898,1 99 100 0
Nataima VOLRIP3 T30_009 T30 DO3550701 99 99 0
Nataima VOLRIP4 T30_010 T30 DO3550701 99 99 0
Tabla 2-2: Resumen de identidad de las secuencias de las muestras con accesiones del GeneBank

46 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
2.4 Discusión
Los resultados indican que las plantas de los huertos de lima Tahití de La Libertad tienen
infección de un complejo de cepas T30, VT y T36, frente a la población de Nataima cuyo
complejo de infección se restringe a cepas T30 y VT. La incidencia de infección por
cepas del grupo severo VT fue alta en los huertos evaluados, acorde con lo reportado
previamente acerca de su amplia distribución e incidencia en Colombia (Peñaranda et al.,
1996; Oliveros et al., 2010; Delgado et at., 2004; Martínez y Guzmán 2007). Se
identificaron cepas suaves T30 en las dos poblaciones de lima Tahití, y la proporción de
infección fue mayor en Nataima respecto a La Libertad. Esto podría indicar que las cepas
de este tipo posiblemente se han dispersado por el país, encontrándose inicialmente en
Mompóx (Bolivar) y luego en Cundinamarca (Peñaranda et al., 1996; Oliveros et al.,
2010). En el presente trabajo, las cepas T30 fueron detectadas en Villavicencio (Meta) y
Espinal (Tolima). Se identificaron cepas T36 en el huerto La Libertad (Villavicencio –
Meta) a través de los marcadores específicos del gen de la polimerasa, esta detección
corrobora estudios preliminares que sugieren la presencia de este tipo de cepas
mediante secuencias del gen CP (Peñaranda et al., 1996). Las secuencias del gen de la
polimerasa indican identidades entre el 99 y el 100% de las muestras positivas para las
cepas VT, T30 y T36. La detección de las cepas T36 es preliminar y debe ser
corroborada con marcadores de otros genes que sean informativos para discriminar este
tipo de cepas. Estudios de infección mixta presentes en campo han mostrado que es
frecuente encontrar mezcla de cepas VT/T30, y en casos excepcionales T30/T36 (Roy y
Brlansky, 2004).
Los resultados obtenidos muestran plantas con infecciones mixtas en varias
combinaciones: plantas infectadas con cepas VT/T30, VT/T36 y VT/T30/T36. Se encontró
alta concordancia entre los primers del gen de la polimerasa y los primers del gen p23
para identificar genotipos de CTV. Muestras que indicaron infección únicamente con VT
con los primers del gen de la polimerasa, con p23 indicaron que pertenecían al grupo
severo que podrá ser VT ó Barao B.
La notable predominancia en campo de cepas VT respecto a T30 y T36 genera
interrogantes de la forma en que se dio la dinámica de infección de CTV. El material fue
plantado al mismo tiempo, sin embargo las proporciones de infección son diferentes.

Capitulo 2 47
¿Hay alguna adaptación de las diferentes cepas de CTV a los vectores locales? En los
campos de cítricos de Colombia es predominante el vector Toxoptera citricida y se sabe
que este es vector mas eficiente de las cepas de CTV tipo VT, de manera tal que podría
haber una relación lógica con la dinámica de infección, ¿Fue efectiva la producción de
yemas libres de virus? Realmente no se logro llevar una trazabilidad del material desde la
liberación en vivero hasta su establecimiento en campo, no obstante se piensa que
probablemente cepas T36 encontradas únicamente en el huerto de La Libertad son
procedentes de campo, ya que de proceder del vivero habría mayor probabilidad de
encontrarla en Nataima y como se mostro este resultado fue negativo. ¿Por qué se da
una delta de incidencia tan alta en tan solo dos años? El vector Toxoptera citricida tiente
la capacidad de infectar el 95% de una población de cítricos en 2 a 4 años (Gottwald et
al., 1996), es predominante en campo y probablemente se una de las causas de dicho
comportamiento. No obstante, no tenemos evidencia de la actividad del vector en las
parcelas de estudio para explicar este comportamiento, o sí puede tener mayor afinidad
por la transmisión de cepas VT.
Con las secuencias obtenidas fue posible confirmar los resultados obtenidos por RT-PCR
del gen de la polimerasa. Se encontraron valores de incidencia diferentes entre cepas VT
y T30 en las dos áreas geográficas evaluadas. Las plantas de lima Tahití de Nataima
presentaron masivamente infecciones mixtas entre cepas suaves T30 y severos VT. Los
primers del gen p23 mostraron las muestras con infecciones mixtas VT/T30, lo cual se
evidencia por la amplificación de bandas de 612, 412 y 239 simultáneamente(Giampan et
al., 2010).
En la Libertad no fue posible identificar plantas con infección exclusiva de cepas del
grupo severo T36 mediante RT-PCR bidireccional del gen p23. Esto debido a que la
totalidad de los cítricos tenían infección con cepas del grupo VT en mezcla con otras
cepas. Por lo tanto, la banda de 612pb que indica la infección con la cepa T36 nunca la
encontraríamos sola a menos de que el cítrico presente únicamente infección con
estacepa.
2.5 Bibliografía
Albiach-Marti, M R., Mawassi,M., Gowda, S., Satyaranayana, T., M.E., Shanker, S.,
Almira, E.C., Vives, M.C., López, C.,Guerri, J.,Florez,R., Moreno, P., Garnsey, S., M.,

48 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
Dawson, W.O. 2000. Sequences of Citrus Tristeza Virus separated in time and space are
essentially identical. J. Virol. 74, 6856- 6865.
Bar-Joseph, M., Loebenstein, G. and Cohen, J. (1972) Further purificationand
characterization of threadlike particles associated with thecitrus tristeza disease. Virology,
50, 821–828.
Bar-Joseph, M., Marcus, R., Lee, R.F. (1989) The continuous challenge of Citrus Tristeza
Virus control.Annual Review of Phytopathology 27, 291-316.
Cerni, S., Ruscic, J., Nolasco., G, Zivko., G, Krjacic., M, skoric, D. 2007. Stem pittin and
seedling yellows symptoms of Citrus Tristeza Virus infection may be determinate by minor
sequences variants. Virus Genes 36:241-249.
Delgado, J., Guzmán, M., Caicedo, A., Ordúz, J. 2004. CTV screening of Colombian germplasm banks kept in Palmira (Valle del Cauca) and Villavicencio (Meta) and preliminary estimation of CTV variants by RFLP and SSCP 44 Meeting of the American Phytopathological Society Caribbean Division La Habana, mayo 24-28. Memorias.
Febres, V.J., Ashoulin, L., Mawassi, M., Frank, A., Bar-Joseph, M., Manjunath, K.L., Lee,
R.F. and Niblett, C.L. 1996.The p27 protein is present at one end of Citrus Tristeza Virus
particles. Phytopathology, 86, 1331–1335.
Garnsey, S.M., Gonsalves, D., and Purcifull, D. E. 1978. Rapid diagnosis of citrus
Tristeza virus infections by sodium dodecyl ulfate-immunodiffusion procedures.
Phytopathology 68:88-95.
Garnsey, S. M., D. J. Gumpf, C. N. Roistacher, E. L. Civerolo, R. F. Lee, R. K. Yokomi,
and M. BarJoseph 1987. Toward a standardized evaluation of the biological properties of
Citrus Tristeza Virus.Phytophylactica 19: 151-157.
Gonsalves, D., Purcifull, D., Garnsey, S.M. 1978. Purification and serology of Citrus
Tristeza Virus.Phytopathology 68, 553-559.
Hilf, M.E., Karasev, A.V., Pappu, H.R., Gumpf, D.J., Niblett, C.L. and Garnsey, S.M. 1995.
Characterization of citrus tristeza virus subgenomic RNAs in infected tissue. Virology,
208, 576–582.
Hilf, M. E y Garnsey, S.M. 2000. Characterization and Classification of Citrus Tristeza Virus Isolates by Amplification of Multiple Molecular Markers. Fourteenth IOCV Conference, 2000—Citrus Tristeza Virus
Hilf, M.E., Mavrodieva, V. A., and Garnsey, S. M. 2005. Genetic marker analysis of a
global collection of isolates of Citrus Tristeza Virus:Characterization and distribution of
CTV genotypes and association withsymptoms. Phytopathology 95:909-917.

Capitulo 2 49
López, C., Ayllón, M. A., Navas-Castillo, J., Guerri, J., Moreno, P., and Flores, R. 1998.
Molecular variability of the 5′- and 3′-terminal regions of Citrus Tristeza Virus
RNA.Phytopathology 88:685-691.
Martínez, S , Guzmán, M 2007 Serological analysis of Colombian Citrus Tristeza Virus
isolates Phytopathology 97: S175
Mawassi, M., Mietkiewska, E., Gofman, R., Yang, G., and Bar-Joseph, M. 1996.Unusual
sequence relationships between two isolates of Citrus Tristeza Virus.J. Gen. Virol.
77:2359-2364.
Oliveros, O , Torres, J , Morales, G , Guzmán, M , Acosta, O , Peñaranda J 2001 Two
common haplotypes of 106 Rev. Colomb. Biotecnol. Vol. XI No. 1 Julio 2009 94-106.
The CPm gene(p27) in Colombian field isolates of the Citrus Tristeza Virus In:
proccedings of the 15th Conference of the International Organization of Citrus virologists
Riverside: IOCV University of California, Abstracts
Oliveros-Garay, O.A, Martinez-Salazar, N-, Torres-Ruiz, Y., Acosta, O. CPm gene
diversity infield isolates of Citrus Tristeza Virus from Colombia.Arch
Virol. 2009;154(12):1933-7
Peñaranda, J. Acosta, O. Guzman, M. Barney, A. Pappu, H, Pappu, S. Manjunath, L. Febre, J. Niblett, L. 1996. Incidence and characterization of mild and severe isolates of Citrus Tristeza Virus from Colombia. Thirteenth IOCV Conference. Rodriguez, P., De Perez, G., Guzman, M., 2009. Detección del virus de la tristeza de los
cítricos por serología, microscopía e hibridación in situ.
Revista Colombiana De Biotecnología ISSN: 0123-3475 ed: Instituto De
BiotecnologiaIbunv.11 fasc.1 p.94 – 106.
Roy, A. and Brlansky, R.H. 2004. Genotype classification and molecular evidence for the presence of mixed infections in Indian Citrus Tristeza Virus isolates. Arch. Virol. 149 (10), 1911-1929 Ruiz-Ruiz, S., Moreno, P., Guerri, J., and Ambrós, S. 2009. Discrimination between mild and severe Citrus Tristeza Virus isolates with a rapid and highly specific real-time reverse transcription-polymerase chain reaction method using TaqMan LNA probes. Phytopathology 99:307-315.
Sambade. A., Lopez, C., Rubio, L., Flores, R., Guerri, J., Moreno, G. 2003.
Polymorphism of a specific región in gen p23 of Citrus Tristeza Virus allows
discrimination between mild ans severe isolates. Arch Virol 148: 2325 – 2340.
Satyanayanana, T., Gowda, S., Boyko, V.P., Albiach-Martí, M.R., Mawassi, M., Navas-
Castillo, J., Karasev, A.V., Dolja, V., Hilf, M.E., Lewandowsky, D.J., Moreno, P., Bar-
Joseph, M., Garnsey, S.M. and Dawson, W.O. 1999 An engineered closterovirus RNA

50 Identificación de aislamientos de Citrus Tristeza Virus mediante marcadores
moleculares de genotipo especifico en plantas de Citrus latifolia (Tanaka).
replicon and analysis of heterologous terminal sequences for replication. Proc. Natl Acad.
Sci. USA, 96, 7433–7438.
Satyanayanana, T., Gowda, S., Mawassi, M., Albiach-Martí, M.R., Ayllón, M.A.,
Robertson, C., Garnsey, S.M. and Dawson, W.O. 2000 Closterovirus encoded HSP70
homolog and p61 in addition to both coat proteins function in efficient virion assembly.
Virology, 278, 253–265.
Tatineni, S., Robertson. C, Garnsey, S., y Dawson, W. 2011. A plant virus evolved by
acquiring multiple nonconserved genes to extend its host range. Proc Natl Acad Sci U S
A. 2011 Oct 18;108(42):17366-71. Epub 2011 Oct 10.

3. Patrones para lima Tahití C. latifolia (Tanaka) frente a la severidad de stem pitting ocasionado por Citrus Tristeza Virus
Resumen
Citrus Tristeza Virus induce diversos síntomas dependiendo de tipo de aislamiento ó el
complejo de infecciones mixtas y de la especie y variedad del hospedero cítrico. En el
presente trabajo se evaluó la severidad de “stem pittin” en lima Tahití injertada sobre seis
patrones diferentes. En trabajos preliminares se identificaron las cepas de CTV que
infectaron en campo el material objeto de evaluación. Se identificaron cepas de VT
predominantes que generan “stem pitting” solas y en infecciones mixtas con cepas T30 y
T36. La relación entre la severidad de “stem pitting” con la localidad, los diferentes
patrones y cepas de CTV se analizaron mediante análisis de correspondencias múltiples,
tablas de contingencia y test de independencia chi cuadrado. Se encontró que la
severidad de “stem pitting” está asociada con la localidad geográfica y el patrón, pero no
con la mezcla de cepas virales. El patrón Kryder injertado con lima Tahití se muestra
prometedor frente a infección por CTV para los materiales cultivados en las localidades
de La Libertad y Nataima
Abstract
Citrus Tristeza Virus induces different symptoms, which are related with virus genotype or
mixed infections and type of host specie and variety. This study evaluated the severity of
“stem pittin” in Tahitian lime grafted on six different patterns. In preliminary work identified
CTV strains that infected field material evaluated. Isolates classified as genotype VT were
the most frequent and they generate stem pitting alone or in mixed infections with isolates
with genotypes T30 and T36. The relationship between the severity of stem pitting with
the geographical locality, the different rootstocks and CTV genotype were analyzed by
multiple correspondence analysis, contingency tables and chi square test of

52 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
independence. The severity of stem pitting depends on the location and rootstocks, but
not the mixture of viral strains. The rootstock Kryder show promise against CTV for La
Libertad and Nataima
3.1 Introducción
La enfermedad tristeza de los cítricos es causada por Citrus Tristeza Virus (CTV), es un
agente viral limitante para la producción de cítricos a nivel mundial. Las estrategias de
manejo dependen de las condiciones específicas de cada región, dado por las cepas
predominantes y la incidencia de CTV. En este sentido las medidas de manejo generales
son: i). Programas de certificación de yemas libres de virus y cuarentena para evitar la
dispersión de cepas ii) Erradicación de plantas infectadas cuando las incidencias son
bajas. iii) Uso de patrones tolerantes a cepas de CTV. iv) Esquemas de protección
cruzada con aislamientos suaves frente asilamientos severos. (Gottwald et al., 2002;
Moreno et al., 2008; Navarroet al., 2002). Las estrategias de manejo de infecciones por
CTV en la citricultura colombiana son limitadas. La incidencia de CTV en Colombia es
muy alta y no está concentrada en una zona específica de manera tal que la erradicación
de plantas no es una opción. Las cepas severas de CTV son predominantes en el país
(Peñaranda et al., 1996; Oliveros et al., 2001; Delgado et at., 2004; Martínez y Guzmán
2007) y no hay legislación restrictiva para el flujo de material entre zonas. Las
plataformas de protección cruzada exitosas en otros países no son homologables a la
citricultura del país. En este orden de ideas las estrategias de manejo que están al
alcance son la producción yemas libres de virus y uso de patrones tolerantes a CTV con
buena adaptación al trópico bajo para el caso específico de lima Tahití.
La lima Tahití (Citrus latifoliaL.) puede ser originaria del archipiélago Malayo, no tiene
semillas y tiene buena adaptación a regiones tropicales (Castle y Gmitter, 1999), sin
embargo exhibe alta susceptibilidad a la enfermedad tristeza de los cítricos (Campbell,
1979). Al igual que otros cítricos su entrada en producción es tardía cuando esta
injertada sobre patrones como mandarina cleopatra (Orduz et al., 2006). La producción
promedio en parcelas experimentales se estima en 15 ton/ha. Sin embargo, a parir del
quinto año hay disminución en los rendimientos a causa de problemas fitosanitarios como
CTV (Orduz-Rodriguez et al., 2007; Quiroga et al., 2010).

Capitulo 3 53
Los cítricos se propagan mediante el uso de injertos para conservar las características de
las plantas madre. El éxito de esta forma de propagación depende de la selección del
patrón y del injerto (Agrios, 2005). El uso de patrón confiere varias ventajas como
precocidad en la producción, mayor vigor y uniformidad de la plantación, cierto control
sobre la calidad y cantidad de la cosecha para una misma variedad, adaptación a
problemas físico-químicos del suelo y tolerancia o resistencia a plagas y enfermedades
(Orduz y Baquero, 2003). En el presente trabajo se emplearon patrones promisorios
para la citricultura en general, como son Volkameriano, Kryder, Sunky x English,
Cleopatra, Carrizo y Citrumelo (Orduz, 2003; Orduz et al., 2006). Los antecedentes de
comportamiento frente a CTV se describen a continuación. El patrón Kryder (Poncirius
trifoliata L Raf) ha mostrado tolerancia a la mayoría de aislamientos de CTV y Fortunella
crasiflora Swing tolerancia a aislamientos específicos de CTV (Mestre et al., 1997; Rai,
2006). En los protoplastos de materiales de Fortunella hay replicación del virus, lo que
indica que el movimiento de CTV es lo que se restringe (Albiach-Martí et al., 2004) y se
presenta es tolerancia (Moreira et al., 1949; Yoshida, 1993; Mestreet al., 1997). El patrón
Swingle Citrumelo (Citrus paradisi Macf. x Poncirus trifoliata (L.) Raf) inicialmente fue
reportado como tolerante a CTV (Hutchison., 1974), sin embargo se detectó
susceptibilidad de este material frente a los aislamientos severos B3 y B28, y
adicionalmente tiene alta afinidad productiva con lima Tahití (Campbell 1991). El patrón
Carrizo Citrange (Citrus sinensis Osb. x Poncirus trifoliata (L.)) ha reportado niveles de
tolerancia heredados de Poncirus trifoliata (Garnesey et al., 1996) y ha presentado buen
comportamiento frente a CTV injertado con naranja valencia (Mendtet al., 1988). Sunki x
english (Citrus sunkiHort. ex Tan. x Poncirus trifoliata (L.)) podría tener buenos niveles de
tolerancia heredados de Poncirus trifoliata (L.). El limón Volkameriano (Citrus
Volkameriana Ten. y Pasq.) también ha presentado buen comportamiento frente a CTV
cuando se injerta con naranja valencia, no obstante hay que tener en cuenta que naranja
valencia es menos susceptible a CTV y es posible que su comportamiento no sea
homologo en lima Tahití. Mandarina Cleopatra (Citrus reshni Horth. Ex Tan)) es
reconocido por niveles de tolerancia a CTV, sin embargo, no presenta alta afinidad con
lima Tahití y la longevidad productiva de las plantas se limita hasta el año ocho (Stenzel y
Neves 2004; Figueiredo et al. 2007, Quiroga et al., 2010). Cleopatra es uno de los
patrones utilizados con más frecuencia en Colombia, es un material de precocidad media
en producción, porte alto que dificulta la cosecha y baja longevidad en campo atribuida a
CTV (Orduz-Rodriguez y Avella 2008).

54 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
Las cepas severas VT son predominantes en campo, generan el sintoma “stem pitting”
que hacen referencia a acanaladuras en el tallo. Estas se dan por la interrupción de la
actividad meristematica en áreas limitadas del cambium, lo que resulta en un crecimiento
radial irregular (Moreno et al., 2008). Los síntomas que generan estas cepas no colapsan
el árbol, pero afecta su crecimiento y desarrollo, producción y calidad de los frutos, y las
pérdidas ocasionadas por este tipo de cepas alcanzan hasta el 48% en huertos
comerciales de cítricos en Sur Africa (Marais et al., 1996). Las limas ácidas son
altamente sensibles a las acanaladuras de tallo. Es posible identificar acanaladuras en el
tallo principal en plantas maduras y en las ramas del tercio medio (McClean y Van Der
Plank, 1955). Las cepas T30 se consideran suaves y en algunos casos se presentan
como asintomáticos. Las cepas del genotipo T36 son severas “quick decline”, que indica
decaimiento rápido sobre todos los cítricos injertados sobre naranjo agrio, es decir un
colapso de la planta. El objetivo de este trabajo fue evaluar la severidad de “stem pitting”
producida por cepas de CTV infectadas naturalmente en campo sobre seis patrones para
lima Tahití.
3.2 Materiales y métodos.
Para el desarrollo de este trabajo se establecieron dos parcelas experimentales en
campo. La primera en el centro de investigación Corpoica Nataima, en el Espinal Tolima
ubicado en las coordenadas 4° 12’ 56’’ N y 72° 56’ 3’’ O, a 430 msnm, con una
temperatura media de 28 ºC y una humedad relativa del 70%. La segunda en el centro
de investigación Corpoica La Libertad a 2 Km de Villavicencio, ubicado en las
coordenadas 9º 6' N, 73º 34' O, a 320 msnm, con una temperatura media de 27 ºC y
humedad relativa entre el 70 y el 90%. La fase de laboratorio del presente estudio se
desarrolló en el laboratorio de Biotecnología vegetal, de la facultad de agronomía, de la
Universidad Nacional de Colombia.
Material vegetal y colecta de muestras

Capitulo 3 55
.
Figura 3-1: Patrones micro injertados con lima Tahití y establecidos en campo
El material vegetal se obtuvo del banco de germoplasma de cítricos ubicado en el centro
de investigación Corpoica Palmira. Seis patrones cítricos (Volkameriano, Kryder, Sunky x
English, Cleopatra, Carrizo y Citrumelo) fueron micro-injertados con lima Tahití y llevadas
a vivero. En campo se establecieron seis plantas por patrón. Las seis plantas se ubicaron
en el mismo surco. Cada grupo de seis plantas se estableció de forma aleatoria y se
ubicaron cuatro repeticiones
La sintomatología de tristeza se evaluó a través de la intensidad de “stem pitting”
presente en las ramas del tercio medio de los cítricos. Se tomaron ramas con corteza
verde de 20 cm de largo en cada uno de los ejes cardinales del dosel de cada planta. Las
ramas fueron auto clavadas para facilitar el removimiento de la corteza. Se asigno el
valor de intensidad de acanaladuras de la siguiente manera: 1 = ausencia de
acanaladuras, 2 =escaso o menor número de acanaladuras, 3 = moderado numero de
acanaladuras, 4 = muchas acanaladuras superficiales y pocas profundas, 5 = muchas
acanaladuras superficiales y profundas. La escala categórica se utilizó de acuerdo a lo
establecido por Meissner-Filho et al., (2002). La representación grafica de cada una de
las categorías se presenta en la figura 3-2.
.
Citrumelo Swingle (Citrus paradisi Macf. x Poncirus
trifoliata (L.) Raf.)
Sunki x english (Citrus sunkiHort. ex Tan. x Poncirus
trifoliata (L.))
Carrizo (Citrus sinensis Osb. x Poncirus trifoliata (L.)
Kryder (Poncirus trifoliata(L.) Raf.)
Mandarina cleopatra (Citrus reticulata Blanco).
Volkameriana (Citrus Volkameriana Ten. y Pasq.)
Injerto Portan injerto
Lima Tahiti(Citrus latifolia
Tanaka)
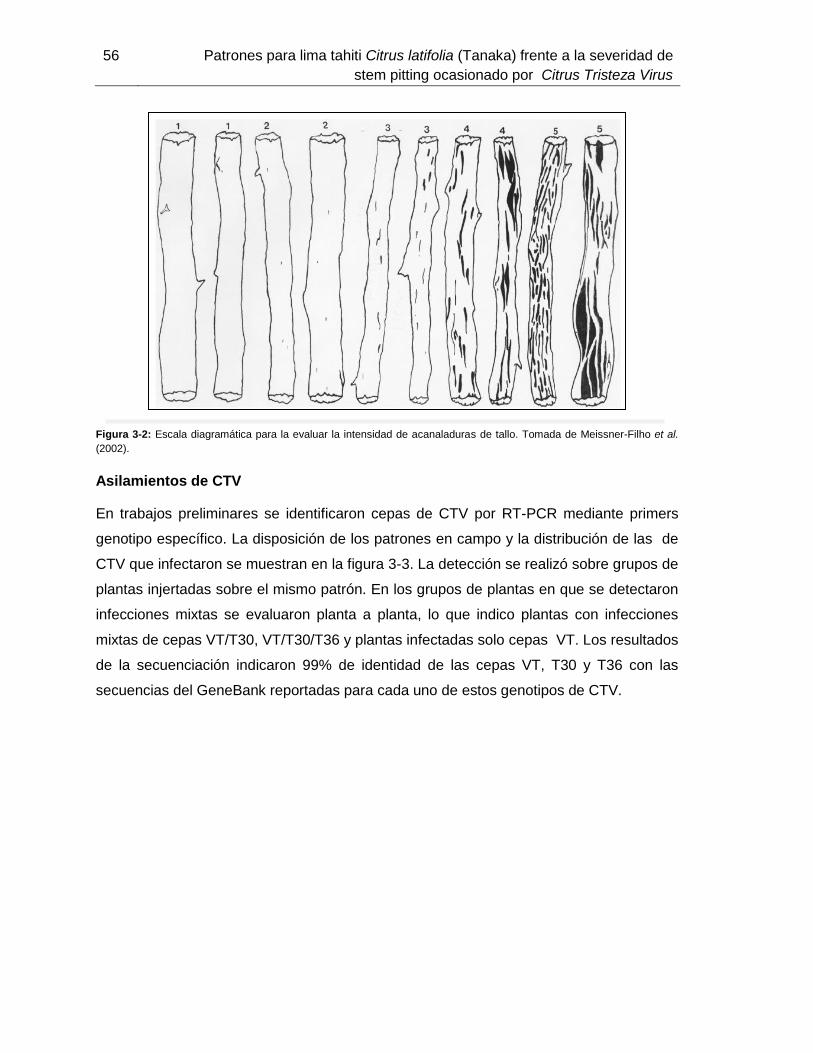
56 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
Figura 3-2: Escala diagramática para la evaluar la intensidad de acanaladuras de tallo. Tomada de Meissner-Filho et al.
(2002).
Asilamientos de CTV
En trabajos preliminares se identificaron cepas de CTV por RT-PCR mediante primers
genotipo específico. La disposición de los patrones en campo y la distribución de las de
CTV que infectaron se muestran en la figura 3-3. La detección se realizó sobre grupos de
plantas injertadas sobre el mismo patrón. En los grupos de plantas en que se detectaron
infecciones mixtas se evaluaron planta a planta, lo que indico plantas con infecciones
mixtas de cepas VT/T30, VT/T30/T36 y plantas infectadas solo cepas VT. Los resultados
de la secuenciación indicaron 99% de identidad de las cepas VT, T30 y T36 con las
secuencias del GeneBank reportadas para cada uno de estos genotipos de CTV.
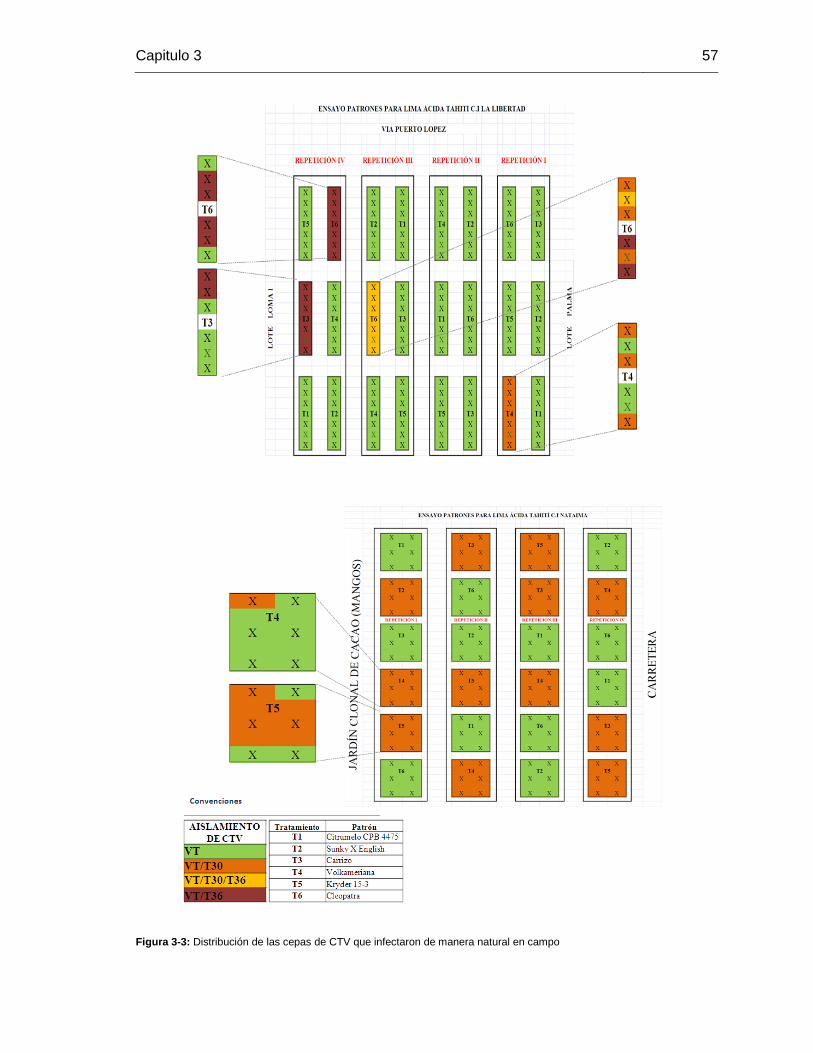
Capitulo 3 57
Figura 3-3: Distribución de las cepas de CTV que infectaron de manera natural en campo
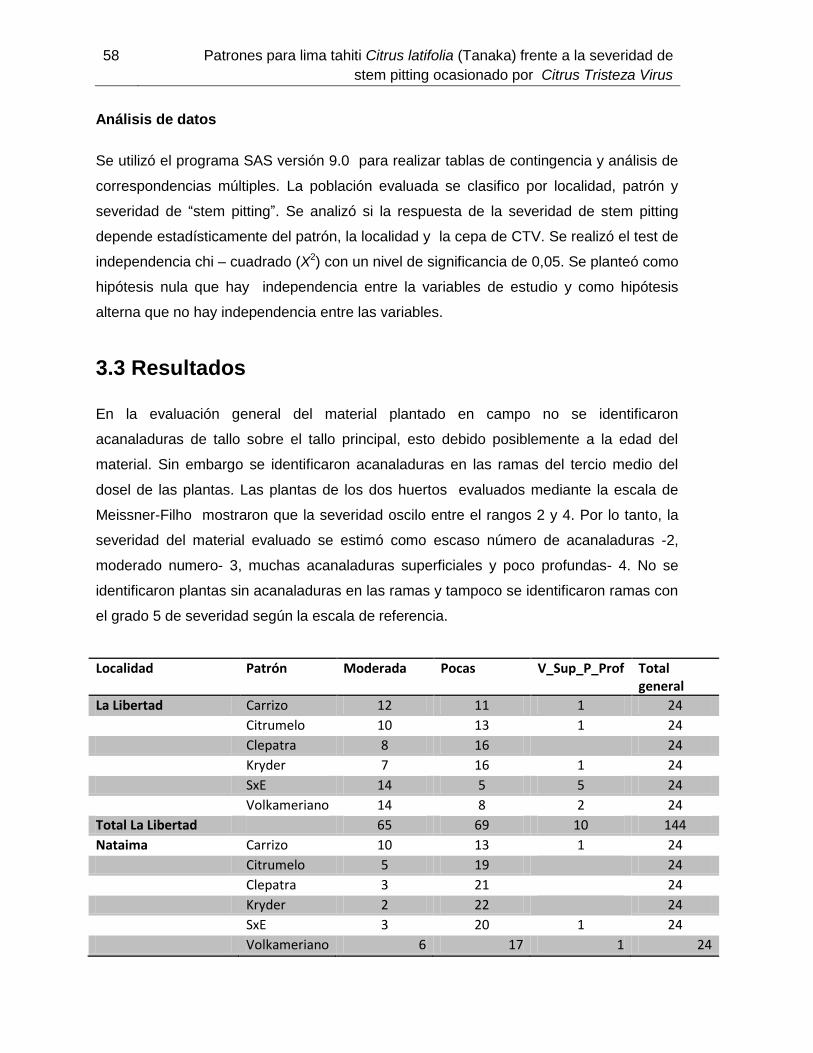
58 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
Análisis de datos
Se utilizó el programa SAS versión 9.0 para realizar tablas de contingencia y análisis de
correspondencias múltiples. La población evaluada se clasifico por localidad, patrón y
severidad de “stem pitting”. Se analizó si la respuesta de la severidad de stem pitting
depende estadísticamente del patrón, la localidad y la cepa de CTV. Se realizó el test de
independencia chi – cuadrado (X2) con un nivel de significancia de 0,05. Se planteó como
hipótesis nula que hay independencia entre la variables de estudio y como hipótesis
alterna que no hay independencia entre las variables.
3.3 Resultados
En la evaluación general del material plantado en campo no se identificaron
acanaladuras de tallo sobre el tallo principal, esto debido posiblemente a la edad del
material. Sin embargo se identificaron acanaladuras en las ramas del tercio medio del
dosel de las plantas. Las plantas de los dos huertos evaluados mediante la escala de
Meissner-Filho mostraron que la severidad oscilo entre el rangos 2 y 4. Por lo tanto, la
severidad del material evaluado se estimó como escaso número de acanaladuras -2,
moderado numero- 3, muchas acanaladuras superficiales y poco profundas- 4. No se
identificaron plantas sin acanaladuras en las ramas y tampoco se identificaron ramas con
el grado 5 de severidad según la escala de referencia.
Localidad Patrón Moderada Pocas V_Sup_P_Prof Total general
La Libertad Carrizo 12 11 1 24
Citrumelo 10 13 1 24
Clepatra 8 16 24
Kryder 7 16 1 24
SxE 14 5 5 24
Volkameriano 14 8 2 24
Total La Libertad 65 69 10 144
Nataima Carrizo 10 13 1 24
Citrumelo 5 19 24
Clepatra 3 21 24
Kryder 2 22 24
SxE 3 20 1 24
Volkameriano 6 17 1 24

Capitulo 3 59
Total Nataima 29 112 3 144
Total general 94 181 13 288
Tabla 3-1 Tabla de frecuencia de la severidad en stem pittin en diferentes patrones y dos
localidades diferentes. Pocas= (2) escaso número de acanaladuras, Intermedio= (3) moderado numero de
acanaladuras, V_Sup_P_Prof = (4) muchas acanaladuras superficiales y pocas profunda
En la Tabla 3-1 se presentan los valores de severidad “stem pitting” de las plantas en los
dos huertos evaluados. La relación entre estas variables se describió y analizó mediante
análisis de correspondencias múltiples. La frecuencia de plantas con severidad grado 2
fue de 78% en Nataima frente a 48% de la Libertad. La frecuencia de severidad grado 3
fue de 45% en La Libertad frente a 20% de Nataima. La frecuencia de severidad grado 4
fue de 7% en La Libertad frente a 2% de Nataima. Respecto a la variedad de patrón, se
observó que el 21% de las plantas injertadas sobre el patrón Sunky x English de La
Libertad presentaron severidad grado 4 frente al 58% que presentaron grado 3 de
severidad. Las plantas injertadas sobre el patrón carrizo presentaron frecuencias
similares de plantas con severidad en grados 2 y 3 en las dos localidades. Los patrones
cleopatra y kryder presentaron las altas frecuencias de severidad grado 2, así: cleopatra
tuvo frecuencia de 88% en Nataima y 67% en La Libertad. Kryder tuvo frecuencia de 92%
en Nataima y 67% en La Libertad.
En la figura 3-3 se muestra la validación de los datos de la tabla de frecuencias mediante
un grafico de correspondencias múltiples: De acuerdo al aporte de inercia, la dimensión 1
está conformada por las variables localidad y patrón, y la dimensión 2 está conformada
por la variable severidad. La severidad de stem pittin sobre carrizo y volkameriana fue
similar en la Libertad, en el grafico se ubican equidistantes de los puntos de severidad
intermedio y menor. Los patrones cleopatra y kryder muestran cercanía con la variable
menor severidad. El patrón Volkameriano está más relacionado con grado moderado de
severidad en La Libertad y grado menor. En la Tabla 3-1 se presentan los valores de
severidad “stem pitting” de las plantas en los dos huertos evaluados. de severidad en
Nataima.
La severidad de stem pittin se comporta de manera dependiente del patrón (test de
independencia chi cuadrado pvalor= 0.0195<0.05) y se comporta de manera dependiente
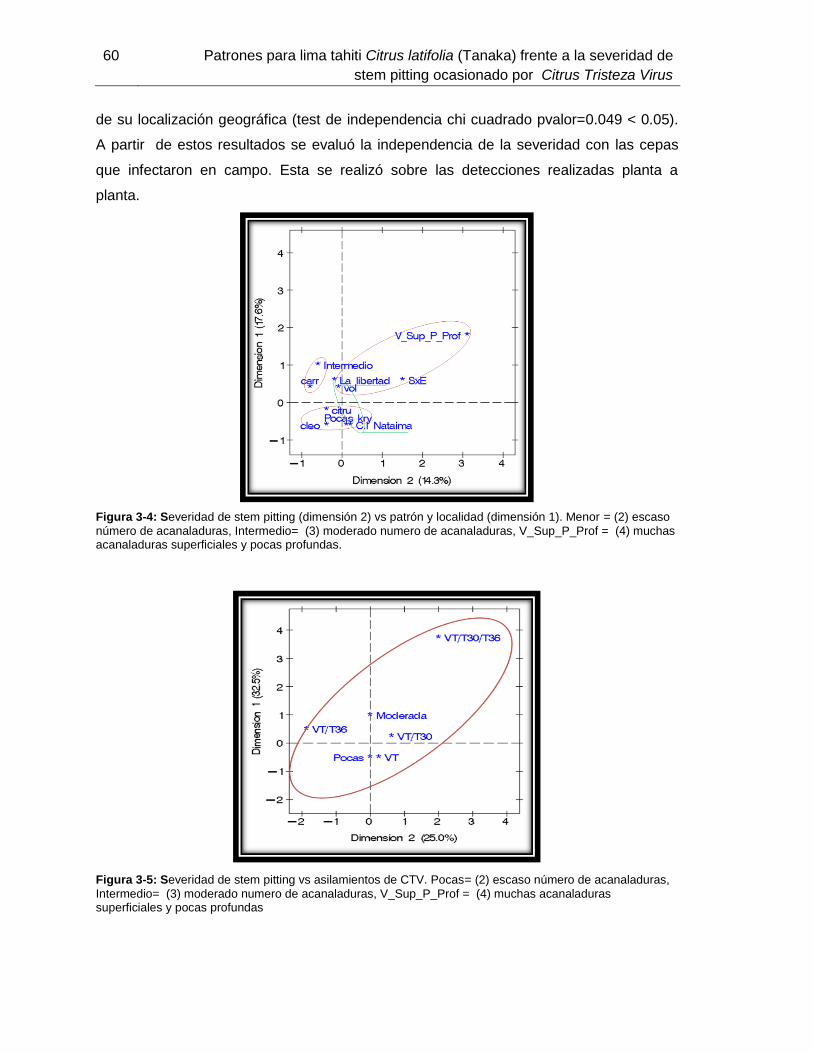
60 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
de su localización geográfica (test de independencia chi cuadrado pvalor=0.049 < 0.05).
A partir de estos resultados se evaluó la independencia de la severidad con las cepas
que infectaron en campo. Esta se realizó sobre las detecciones realizadas planta a
planta.
Figura 3-4: Severidad de stem pitting (dimensión 2) vs patrón y localidad (dimensión 1). Menor = (2) escaso
número de acanaladuras, Intermedio= (3) moderado numero de acanaladuras, V_Sup_P_Prof = (4) muchas acanaladuras superficiales y pocas profundas.
Figura 3-5: Severidad de stem pitting vs asilamientos de CTV. Pocas= (2) escaso número de acanaladuras,
Intermedio= (3) moderado numero de acanaladuras, V_Sup_P_Prof = (4) muchas acanaladuras superficiales y pocas profundas

Capitulo 3 61
La expresión de severidad de stem pitiing se comportó de manera independiente del la
mezcla de cepas que infectaron(test de independencia chi cuadrado pvalor= 0.3672>
0.05), de manera tal se muestra en grafico 3-4 que las plantas infectadas con cepas VT,
VT/T30 y VT/T36 expresan grados de severidad similares. Una sola planta presentó
infección mixta de cepas VT/T30/T36, sin embargo, su expresión de severidad se ubico
dentro de los mismos rangos que plantas infectadas con otras cepas.
3.4 Discusión
La eliminación de la corteza a nivel de ramas jóvenes, permite evaluar estrías o
acanaladuras longitudinales a nivel del tejido vascular acorde a sintomatología descrita
como síndrome de acanaladura de tallos “stem pitting” (Meissner-Filho et al.,2002; Magri
et al., 2011). Se encontró que la severidad de CTV es variable según el patrón y la
localización, pero no según el complejo de cepas virales. La cepa viral predominante en
campo fue VT para las dos localidades. Las plantas en que se identificaron infecciones
con la cepa VT o infecciones mixtas VT/T30, VT/T36 ó VT/T30/T36 no tuvieron un efecto
sobre la severidad de CTV de acuerdo a la escala de referencia. En el estado fenológico
en el que se evalúan estas plantas, no es posible hacer inferencias para explicar
modulación de expresión de síntomas mediante procesos de protección cruzada.
Tampoco es posible establecer cuál de los genotipos virales presentes en una infección
mixta, es predominante en las plantas a fin de explicar el comportamiento de severidad
obtenido. El uso de otras escalas como Ndongo ó Horsfall y Barrat no es aplicable a la
edad de las plantas, ni al estado sanitario, pues con cualquiera de estas es muy posible
confundir otros síntomas generados por CTV con los síntomas causados por otros
disturbios como clorosis en las nervaduras. La escala de Ndongo sería aplicable en
estados más avanzados de la enfermedad en la planta y fenológicamente más
desarrollada. Acorde con lo anterior la escala propuesta por Meissner-Filho et al (2002)
permitió tener una alta resolución para medir la severidad de la tristeza de los cítricos en
lima Tahití .
El test de chi cuadrado indicó que la severidad de “stem ptiting” se comporta de manera
dependiente del patrón y de la localidad. La dependencia de la localidad se puede
explicar a partir de la diferencia de las condiciones ambientales, aunque no hay un
estadístico certero de la relación entre las variables ambiente vs severidad. La

62 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
comparación de los valores de severidad entre las dos localidades, muestra que grupos
de plantas con un mismo patrón infectadas con cepas virales del mismo genotipo
presentan un contraste en la severidad, como lo es el caso del patrón Sunky x English
cuya severidad tiende al mayor grado en La Libertad, situación que ocurre de manera
general con otros patrones. No obstante no conocemos como ha sido la actividad del
vector en campo y como afectan los eventos de re-inoculación (Cambra et al., 2002)
sobre la respuesta sintomática de la planta.
La dependencia del patrón se podría explicar por la tolerancia específica de la especie de
cada patrón. Se observa que hay consistencia entre las proporciones de las frecuencias
de severidad para algunos patrones entre localidades. La severidad del patrón carrizo fue
muy similar en las dos localidades, se esperaban grados de severidad bajos teniendo en
cuenta que este patrón procede de Poncirus trifoliata (Garnesey et al., 1996), sin
embargo los valores de severidad fueron muy proporcionales entre los grados 2 y 3 para
las dos localidades. El patrón citrumelo que también tiene herencia de P. trifoliata
presentó valores de severidad similares a carrizo en La Libertad y una severidad grado 2
en Nataima. El patrón kryder presenta un alto grado de consistencia con las referencias
de tolerancia (Mestre et al., 1997; Rai, 2006), la severidad grado 2 fue las más frecuente
entre las plantas de las dos localidades. El número de plantas con severidad grado 2 de
los patrones citrumelo, cleopatra, y carrizo fue mayor que el número de árboles con
grado 3 de severidad, de manera consistente en las dos localidades. Sin embargo la
frecuencia de severidad grado 2 siempre fue mayor en Nataima, se puede decir que la
severidad de estos patrones es intermedia entre kryder y sunky x english. El patrón limón
volkamieriano se comporto de manera inversa en las dos localidades, en la Libertad se
presentó un mayor número de plantas con severidad grado 3 respecto a las grado 2 y en
Nataima se presentó una distribución inversa. El patrón mandarina cleopatra ha sido
caracterizado por exhibir baja afinidad con lima Tahití (Stenzel y Neves 2004;
Figueiredoet al. 2007, Quiroga et al., 2010) y en algunos casos se limita longevidad
atribuida a problemas fitosanitarios como CTV (Orduz-Rodriguez y Avella 2008). Hasta el
momento cleopatra se ubica en un punto intermedio de severidad de CTV y no se hacen
evidentes incompatibilidades.

Capitulo 3 63
En La Libertad se identificaron plantas infectadas con cepas T36 en infección mixta con
cepas VT y T30. Sin embargo, no se observó variación en la expresión de síntomas
asociados a “stem pitting”. Durante el período de realización de este estudio, estos
árboles no han desarrollado síntomas de declinación rápida - quick deckine -. Varios
factores podrían estar modulando esta respuesta como son el titulo viral, el estado
fenológico de las plantas y variables climáticas, lo que quiere decir que en un futuro
podrían generar la sintomatología asociada a plantas infectadas con este tipo de cepas y
colapsar.
Al momento podemos inferir que hay patrones prometedores en términos de baja
severidad frente a CTV, sin embargo el estado fenológico del material plantado no
permite determinar un mejor patrón. Es necesario que este material de campo continúe
con los procesos infección de cepas de CTV y se haga un seguimiento en el tiempo tanto
de la dinámica de infección como de la respuesta de severidad.
3.5 Bibliografia
Agrios, G.N. 2005. Plant Patology. Fifth Edition .Elsevier American Press. Department of
Plant Pathology.University of Florida. U.S.A. p.922.
Albiach-Martí, M.R., Grosser, J.W., Gowda, S., Mawassi, M., Satyanarayana, T.,
Garnsey, S.M. and Dawson, W.O. 2004 Citrus Tristeza Virus replicates and forms
infectious virions in protoplast of resistant citrus relatives. Mol. Breeding, 14, 117–128.
Cambra, M.C., Martínez, M., Marroquín, C., Gorris, M.T., Zaragoza, I.S., López, A.,
Olmos, A., y Hermoso de Mendoza, A. 2002. Epidemiology of Citrus Tristeza Virus
(CTV) in Citrus Varieties Cultivated Under Plastic Net Covers.Fifteenth IOCV Conference,
2002—Short Communications.
Campbell, C.W., 1979.Tahití lime production in Florida University of Florida. Florida
Cooperative. Service, EVA.
Campbell, C.W. 1991. Production of the lime (Citrus latifolia, Tanaka). In florida.
University of Florida. Tropical Region 2:184-192.
Castle, W.S. y Gmitter, F.G. 1999.Rootstock and scion selection. En: Citrus health
management. Timmer, L.W. y Duncan, L.W. (eds.). University of Florida, APS Press, pp.
21-35.

64 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
Delgado, J , Guzmán, M , Caicedo, A , Ordúz, J 2004 CTV screening of Colombian
germplasm banks kept in Palmira (Valle del Cauca) and Villavicencio (Meta) and
preliminary estimation of CTV variants by RFLP and SSCP 44 Meeting of the American
Phytopathological Society Caribbean Division La Habana, mayo 24-28 Memorias
Garnsey, S.M., Su, H.J. and Tsai, M.C. (1996) Differential susceptibility of pummelo and
Swingle citrumelo to isolates of Citrus Tristeza Virus. In: Proceedings of the 13th
Conference of the International Organization of Citrus Virologists (da Graça, J.V., Moreno,
P. and Yokomi., R.K., eds), pp. 138–146. Riverside, CA: IOCV.
Gottwald, T.R., Polek, M. and Riley, K. (2002) History, present incidence, and spatial
distribution of Citrus Tristeza Virus in the California Central Valley.In Proceedings of the
15th Conference of the International Organization of Citrus Virologists (Duran-Vila, N.,
Milne, R.G. and da Graça, J.V., eds), pp. 83–94. Riverside, CA: IOCV.
Hutchison, D. J. 1974. Swingle citrumelo-A promising rootstock hybrid. Proc. Fla. State
Hort. Soc. 87:89-91.
Figueiredo, .J.O., Stuchi, E.S., Laranjeira, F.F., Donadio, l.C., Sobrinho, J., Sempionato,
O.R. y Muller, G.W. 2001. Porta-enxertos para lima ácida ‘Tahití ’ em duas regiões do
Estado de São Paulo. Laranja, v.22, n.1, p.203- 213.
Martínez, S., y Guzmán, M 2007 Serological analysis of Colombian Citrus Tristeza Virus
isolates Phytopathology 97: S175
McClean, A.P.D., y Van Der Plank, J.E. 1955. The role of seedling yellows and stem
pitting in tristeza of citrus.Phytopathology 45:222-224.
Magri, W., Corazza, M ., Zanutto, C., Carvalho-Nunes, W., Müller, G. 2011. SSCP
analysis of Citrus Tristeza Virus protectives isolates in Pêra sweet orange clones under
northern Paraná state, Brazil conditions. Citrus Research & Technology, Cordeirópolis,
v.32, n.1, p.9-16, 2011
Marais, L. J., Marais, M. L., y Rea, M. 1996. Effect of Citrus Tristeza Stem Pitting on Fruit
Size and Yield of Marsh Grapefruit in Southern Africa. Thirteenth IOCV Conference,
1996- Citrus Tristeza Virus
Meissner-Filho, P.E., Soares-Filho, W., Dos. S., Velame, K.V.C., Diamantino, E.P., y
Diamantino, M.S.A.S. 2002. Reação de porta-enxertos híbridos ao Citrus Tristeza Virus.
Fitopatologia Brasileira 27:312-315. 2002.
Moreira, S, Costa, A.S and Grant, T.J. 1949. Conhecimentos atuais sobre a tristeza dos
citrus. Revista de Agricultura 24:335- 345.
Moreno, P., Ambros, S., Albiach-Marti, M.R., Guerri, J., Pena, L., 2008. Citrus
tristezavirus: a pathogen that changed the course of the citrus industry. Mol. Plant
Pathol.9, 251–268.

Capitulo 3 65
Mestre, P.F., Asins, M.I., Carbonell, E.A and Navarro, L. (1997) New gene(s) involved in
the resistance of Poncirus trifoliata (L.) Raf. to Citrus Tristeza Virus. Theor Appl Genet
95:691-695.
Navarro, L., Pina, J.A., Juárez, J., Ballester-Olmos, J.F., Arregui, J.M., Ortega, C.,
Navarro, A., Duran-Vila, N., Guerri, J., Moreno, P., Cambra, M., Medina, A. and
Zaragoza, S. (2002) The Citrus Variety Improvement program in Spain in the period
1975–2001. In: Proceedings of the 15th Conference of the International Organization of
Citrus Virologists (Duran-Vila, N., Milne, R.G. and da Graça, J.V., eds), pp. 306–316.
Riverside, CA: IOCV.
Ndongo, B, Ambang Z, Belibi Messanga L, Ngodo Melingui J, B, Ongono Y. Vigour and
Behaviour of fifteen citrus varieties against tristeza in the forest zone of Cameroon.
Department of Plant Biology Faculty of Sciences, University of Yaoundé I. Afr. J.
Biotechnology.2007;6(12):1403-1409
Oliveros, O., Torres, J., Morales, G., Guzmán, M., Acosta, O., Peñaranda, J. 2001 Two
common haplotypes of 106 Rev. Colomb. Biotecnol. Vol. XI No. 1 Julio 2009 94-106.
The CPm gene(p27) in Colombian field isolates of the Citrus Tristeza Virus In:
proccedings of the 15th Conference of the International Organization of Citrus irologists
Riverside: IOCV University of California, Abstracts
Orduz, J.O, Baquero, J.E. 2003. Aspectos básicos para el cultivo de los cítricos en el
piedemonte llanero. Revista Achagua 2003; 7(9):7-19
Orduz, J.O. 2003. Evaluación de patrones en cítricos en suelos ácidos en condiciones de
vivero en el Trópico bajo de Colombia. Revista Achagua. 2003; 7(9) 24-27.
Orduz, J.O., Arango, l., Monroy, H., Fischer, G. 2006 Comportamiento de la mandarina.
Arrayana en seis patrones en suelos ácidos del piedemonte Llanero de Colombia. Agr.
Col. 2006;24(2):266-273.
Orduz-Rodríguez, J. O., Chacón-Díaz, A., Linares-Briceño, V.M. 2007. Evaluación del
potencial de rendimiento de tres especies y un híbrido de cítricos en la región del Ariari
del departamento del Meta (Colombia) durante doce años, 1991- 2003. Revista
ORINOQUIA - Universidad de los Llanos - Villavicencio, Meta. Colombia. Volumen 11 -
Nº 2.
Orduz-Rodriguez, J., Avella, F. 2008.Comportamiento de 26 cultivares de naranja en
condiciones del piedemonte del Meta, Colombia. Revista Colombiana De Ciencias
Hortícolas - Vol. 2 - No.2 - pp. 157-172.
Peñaranda, J., Acosta, O., Guzman-Barney, M., Pappu, H., Pappu, S., Manjunath, L.,
Febre, J., Niblett, L. 1996. Incidence and characterization of mild and severe isolates of
Citrus Tristeza Virus from Colombia. Thirteenth IOCV Conference.

66 Patrones para lima tahiti Citrus latifolia (Tanaka) frente a la severidad de
stem pitting ocasionado por Citrus Tristeza Virus
Quiroga-Cardona, J., Hernández- Parrado, F.L., Silva- Herrera, M., Orduz-Rodríguez,
J.O. 2010. Comportamiento de la producción de lima Tahití (Citrus latifolia Tanaka),
injertada sobre el patrón de Mandarina Cleopatra (Citrus reticulata Blanco) y la influencia
delvirus de la tristeza (CTV) en condiciones del piedemonte del Meta, 1997-2008.
Orinoquia, vol. 14, núm. 1, junio, 2010, pp. 5-15
Rai, M. (2006) Refinement of the Citrus Tristeza Virus resistance gene (CTV positional
map in Poncirus trifoliata and eneration of transgenic grapefruit(Citrus paradisi ) plant
lines with candidate resistance genes in this region. Plant Mol. Biol. 61, 399–414.
Stenzel, Neusa. M.C., y Neves. C. (2004) Rootstocks for 'Tahití ' lime. Sci. agric.
(Piracicaba, Braz.) [online]. 2004, vol.61, n.2, pp. 151-155. ISSN 0103-
9016. http://dx.doi.org/10.1590/S0103-90162004000200005
Yoshida, T. (1993) Inheritance of immunity to Citrus Tristeza Virus of trifoliate orange in
some citrus intergeneric hybrids.Bull Fruit Tree Res Station 25:33-43

Conclusiones
En campo se identificaron cepas correspondientes a los genotipos VT, T30 y T36. La
cepa VT fue la de mayor incidencia en las dos localidades. La cepa T30 se encontró en
las dos localidades, pero con menor incidencia en La Libertad. La cepa T36 se encontró
únicamente en la Libertad. Se identificaron plantas con infecciones mixtas VT/·T30 y
VT/T30/T36. Las secuencias de los productos de PCR corroboraron los resultados
obtenidos. Todas las secuencias presentaron 99% de identidad con las secuencias
registradas en el GeneBank, sin embargo en trabajos posteriores se sugiere corroborar la
presencia de cepas T36 en Colombia mediante otros marcadores moleculares. Se
recomienda realizar seguimiento a los síntomas de los arboles en los que se identificó
cepas T36 y T30.
El estudio de la relación entre el patrón, las cepas de CTV y la severidad de stem pitting
indicó que la severidad de stem pitting depende de la localidad y el patrón, pero no de la
mezcla de cepas virales. En estudios posteriores se requiere estudiar los factores que
modulan la respuesta sintomática de arboles de lima tahti infectados con diferentes
cepas. No obstante esta respuesta podría ser variable en función del tiempo y trayectoria
del los arboles en campo. Preliminarmente el patrón Kryder injertado con lima Tahití se
muestra prometedor frente a CTV para las localidades de La Libertad y Nataima, de
acuerdo a la severidad de stem pitting presentada y al movimiento viral restringido de
CTV (Albiach-Martí et al., 2004).