
evaluación de la capacidad biodegradadora de materia orgánica de ...
102
EVALUACIÓN DE LA CAPACIDAD BIODEGRADADORA DE MATERIA ORGÁNICA DE BACTERIAS AISLADAS DE LA RIZOSFERA DE DOS PLANTAS DE LA RIBERA DEL RIO COMBEIMA-TRAMO UT JUAN PABLO LEAL GUARNIZO Trabajado de Grado presentado como requisito parcial para optar al título de Biólogo Director ELIZABETH MURILLO PEREA Magister en Química UNIVERSIDAD DEL TOLIMA FACULTAD DE CIENCIAS BASICAS PROGRAMA DE BIOLOGÍA IBAGUE 2015
Transcript of evaluación de la capacidad biodegradadora de materia orgánica de ...

EVALUACIÓN DE LA CAPACIDAD BIODEGRADADORA DE MATERIA ORGÁNICA DE BACTERIAS AISLADAS DE LA RIZOSFERA DE DOS PLANTAS DE LA
RIBERA DEL RIO COMBEIMA-TRAMO UT
JUAN PABLO LEAL GUARNIZO
Trabajado de Grado presentado como requisito parcial para optar al título de Biólogo
Director ELIZABETH MURILLO PEREA
Magister en Química
UNIVERSIDAD DEL TOLIMA
FACULTAD DE CIENCIAS BASICAS PROGRAMA DE BIOLOGÍA
IBAGUE 2015


3
AGRADECIMIENTOS
Expreso mis más sinceros agradecimientos y aprecio a la profesora Elizabeth murillo
Perea, quien pacientemente guío mi labor, impulsó, motivó y contribuyo con
orientaciones, correcciones y dedicación incansable en mi formación académica y en la
construcción de este documento.
Agradezco también a cada una de las personas que hicieran su aporte e influyeron en mí
para mejorar y cumplir con los objetivos.
Detrás de todo el esfuerzo y el trabajo dedicado en conseguir este fruto y logro de vida,
siempre es necesario agradecer infinitamente a mi familia, por su apoyo incondicional
especialmente a mi madre y hermano.

4
CONTENIDO
Pág.
INTRODUCCIÓN ........................................................................................................ 10 1. OBJETIVOS ............................................................................................................ 13 1.1 OBJETIVOS GENERALES ................................................................................... 13
1.2 OBJETIVOS ESPECÍFICOS ................................................................................. 13
2. MARCO REFERENCIAL ........................................................................................ 15 2.1 MARCO TEÓRICO ................................................................................................ 15 2.1.1 El bosque ripario y su funcionalidad en las dinámicas del ecosistema acuático…
.................................................................................................................................... 15 2.1.2 Efecto de la vegetación riparia en la contaminación de los ecosistemas
Acuáticos ..................................................................................................................... 17
2.2 ESTADO DEL ARTE ............................................................................................. 20 3. MATERIALES Y MÉTODOS .................................................................................. 24 3.1 ZONA DE ESTUDIO ............................................................................................. 24 3.2 MATERIALES........................................................................................................ 25 3.3 METODOLOGIA .................................................................................................... 25 3.3.1 Evaluación de la composición florística del BRC-UT .......................................... 25
3.3.2 Selección de las especies vegetales de mayor importancia ecológica en el
BRC-UT ....................................................................................................................... 26
3.3.3 Caracterización fisicoquímica del suelo ribereño y del agua del rio ................... 26 3.3.4 Aislamiento de las rizobacterias asociadas a las plantas seleccionadas ........... 26 3.3.5 Aislamiento de bacterias endófitas de la raíz asociadas a las plantas
seleccionadas ............................................................................................................. 27 3.3.6 Selección de las bacterias solubilizadoras de fosfatos ....................................... 28 3.3.7 Determinación de la capacidad bacteriana para solubilizar el fósforo ............... 28

5
3.3.8 Selección de las cepas bacterianas con capacidad de transformar el amonio .. 29 3.3.9 Selección de cepas con capacidad de transformación del nitrito ....................... 29 3.3.10 Ensayo de verificación de biorremediación en un agua residual sintética ........ 30 3.3.11 Análisis estadístico ........................................................................................... 30
4. RESULTADOS Y DISCUSIÓN ............................................................................... 32 4.1 COMPOSICIÓN FLORÍSTICA DEL BRC-UT ........................................................ 32 4.1.1 Clasificación taxonómica del material vegetal encontrado en la zona
de estudio .................................................................................................................... 32
4.2 SELECCIÓN DE LA DOS ESPECIES VEGETALES CON MAYOR IMPORTANCIA
ECOLÓGICA EN LA ZONA DE ESTUDIO .................................................................. 35
4.2.1 Determinación del índice de valor de importancia (IVI) en las especies vegetales
.................................................................................................................................... 40
4.3 CARACTERIZACIÓN FISICOQUÍMICA DEL SUELO RIBEREÑO
Y EL AGUA DEL RIO .................................................................................................. 45 4.4 AISLAMIENTO Y CARACTERIZACIÓN DE LAS BACTERIAS ASOCIADAS A LA
RAÍZ DEL CHILCO Y PASTO ELEFANTE……………………………………………… . 57 4.4.1 Aislamiento de rizobacterias en las especies seleccionadas ............................. 57 4.4.2 Aislamiento de bacterias endófitas ..................................................................... 62 4.4.3 Selección de las bacterias solubilizadoras de fósforo ........................................ 64 4.4.4 Prueba cualitativa de solubilización en las cepas seleccionadas ....................... 67 4.4.5 Determinación de la capacidad bacteriana para solubilizar el fósforo ................ 69 4.4.6 Selección de cepas bacterianas con capacidad de transformación del amonio ....
.................................................................................................................................... 73
4.4.7 Selección de cepas con capacidad de transformación del nitrito ...................... 75 4.4.8 Ensayo de verificación de biorremediación en un agua residual sintética .......... 79
CONCLUSIONES ....................................................................................................... 80
RECOMENDACIONES ............................................................................................... 82
REFERENCIAS BIBLIOGRAFÍCAS ........................................................................... 83

6
LISTA DE TABLAS
Pág.
Tabla 1. Composición del agua residual sintética ....................................................... 30 Tabla 2. Clasificación taxonómica de los individuos colectados en el BRC-UT
organizados por familia, especie y número de individuos por especie ....................... 33
Tabla 3. Distribución de las especies encontradas en cada uno de los transeptos .... 34
Tabla 4. Medición de los parámetros ecológicos (diámetro de la base (DB), cobertura
foliar (CF) altura y biomasa (peso seco)) en las especies vegetales del BRC-UT ..... 36
Tabla 5. Especies de mayor importancia en los 5 transeptos evaluados.................... 42
Tabla 6. Caracterización fisicoquímica del suelo ribereño bajo estudio ..................... 45
Tabla 7. Caracterización fisicoquímica del agua del rio Combeima en época de lluvias
en dos puntos de la ribera del rio ............................................................................... 49 Tabla 8. Efecto de la temperatura sobre la solubilidad del oxígeno .......................... 50 Tabla 9. Rizobacterias aisladas de la raíz del Chilco y caracterizadas por su morfología
colonial ...................................................................................................................... 60 Tabla 10. Rizobacterias aisladas de la raíz del Pasto elefante y caracterizadas por su
morfología colonial ..................................................................................................... 61 Tabla 11. Bacterias endófitas aisladas y caracterizadas por su morfología colonial de la
raíz del Chilco ........................................................................................................... 62
Tabla 12. Bacterias endófitas aisladas y caracterizadas por su morfología colonial de la
raíz del Pasto elefante ............................................................................................... 62 Tabla 13. Aislados solubilizadores de fósforo inorgánico caracterizados según su
morfología colonial .................................................................................................... 65
Tabla 14. Medida del halo de solubilización en medio SRS con fosfato tricálcico ...... 68
Tabla 15. Principales procesos microbianos involucrados en la solubilización de
fosfatos ....................................................................................................................... 72 Tabla 16. Ensayo de detección de la transformación de nitrito en el medio de cultivo,
utilizando el reactivo de Griess y Zn en polvo ............................................................. 76

7
LISTA DE FIGURAS
Pág.
Figura 1. Relación funcional entre el bosque de ribera y el ecosistema acuático ...... 16
Figura 2. Ilustración de la zona de estudio ............................................................... 24 Figura 3. Parámetros ecológicos evaluados en las especies encontradas en cada uno
de los transeptos ........................................................................................................ 37 Figura 4. Índice de valor de importancia (IVI) de las especies de cada uno de los
transeptos ................................................................................................................... 41
Figura 5. Cepa Relef16 con acidificación del medio y halo de hidrólisis en medio SRS
................................................................................................................................... 64 Figura 6. Medición del halo de solubilización en medio SRS con fosfato tricálcico a las
24 y 48 h de la cepa Relef16. ...................................................................................... 67 Figura 7. Determinación del fosfato disponible después de la acción solubilizadora de
las 3 cepas seleccionadas .......................................................................................... 69 Figura 8. Resultados del ANOVA (GLM) para la determinación del fósforo disponible . .
.................................................................................................................................... 70
Figura 9. Resultados que evidencian la normalidad de los datos y el ajuste del modelo
................................................................................................................................... 71
Figura 10. Registro del comportamiento de la biomasa durante el periodo de
incubación de la bacteria en el caldo amonio .............................................................. 73 Figura 11. Detección de (NH4)2 SO4 con el reactivo de Nessler, en cada uno de los
tubos incubados con las cepas bacterianas .............................................................. 74

8
RESUMEN El rio Combeima, principal arteria fluvial de la ciudad de Ibagué, brinda gran cantidad
de servicios ambientales a sus habitantes, no obstante, durante todo su trayecto por la
ciudad el rio recibe gran diversidad de residuos sólidos y líquidos que afectan la
calidad del cuerpo de agua. Sin embargo, la acción conjunta de la vegetación riparia y
microorganismos permite realizar procesos de regulación natural que ayudan a mitigar
efectos negativos provocados al recurso hídrico. Este trabajo se interesó en seleccionar
las dos especies de mayor importancia ecológica sobre el bosque ripario del rio
Combeima-tramo UT, aislar y caracterizar las bacterias presentes en la rizósfera de las
especies vegetales seleccionadas, evaluar su habilidad para disminuir el exceso de
nutrientes y determinar algunos parámetros ecológicos. Pseudelephantopus spiralis y
Pennisetum purpureum son los organismos vegetales de mayor importancia ecológica
en el BRC-UT. Se obtuvieron 17 rizobacterias y 8 endófitas de P. spiralis, mientras que
18 rizobacterias y 14 endófitas provinieron de P. purpureum. Un total de 11 cepas con
capacidad para solubilizar el fósforo inorgánico fueron aisladas y caracterizadas, se
destacan las cepas RCH1, RCH2 y RCH10. Se encontraron 24 cepas con capacidad
de trasformación del nitrito a nitrato, entre las que se destacan por su mejor habilidad
ENDOCH1, ENDOCH5 y ENDOELEF10. El estudio demostró que P. spiralis y P.
purpureum pueden ser tenidas en cuenta en un proceso de fitorremediación de aguas
con exceso de materia orgánica. Es necesario realizar ensayos sobre agua
contaminada para verificar la acción fitorremediadora de estos organismos.
Palabras clave: vegetación riparia, rio Combeima, bacterias solubilizadoras de fósforo,
biorremediación, rizobacterias.

9
ABSTRACT
The Combeima river, main fluvial artery of the city of Ibagué, provides large amount of
environmental services to its inhabitants, however, throughout its journey around the
city the river receives great diversity and amount of solid waste and liquid that affect not
only the quality of the body of water. However, the joint action of the riparian vegetation
and microorganisms allows you to perform processes of natural regulation that help
mitigate negative effects caused to the water resource. This work are interested, select
the two species of greatest ecological importance on the riparian forest of the river
Combeima UT, isolate and characterize the bacterium present in the rhizosphere of
selected plant species, to assess their ability to reduce excess nutrients, and some
ecological parameters were determined. Pseudelephantopus spiralis and Pennisetum
purpureum plant organisms are of greater importance in the ecological (BRC-UT). Were
obtained 17 rhizobacteria and 8 endophytes of P. spiralis, while 18 rhizobacteria and 14
endophytes came from P. purpureum. A total of 11 strains with the ability to solubilize
inorganic phosphorous were isolated and characterized, highlights the RCH1 strains,
RCH2 and RCH10. We found 24 strains with the ability to transform the nitrite, among
which are by their best skill ENDOCH1, ENDOCH5 and ENDOELEF10. The study
showed that Pseudelephantopus spiralis and Pennisetum purpureum can be taken into
account in a phytoremediation process water with excess organic matter. It is necessary
to carry out tests on contaminated water to verify the action phythoremediating of these
organisms.
Key Words: riparian vegetation, Combeima River, phosphorus solubilizing bacterium,
bioremediation, rhizobacteria.

10
INTRODUCCIÓN La importancia de los ríos como ecosistemas estratégicos para el desarrollo de la vida
en sus diferentes niveles es conocido desde la antigüedad. Grandes civilizaciones
asentadas en la riberas de ríos tuvieron gran desarrollo (comunicación, suministro de
agua y alimento, agricultura, etc.), gracias al entendimiento de sus dinámicas y
aprovechamiento de los recursos que ofrecían estos ecosistemas; ejemplos evidentes
son la población Mesopotámica (rio Tigris y Éufrates), la china (ríos Hoang-Ho y Yang-
Tse-Kiang), la egipcia (rio Nilo) y la india (rio Indo) (Hughes, 1981).
En la actualidad aún se mantiene ese vínculo cultural, social y biológico con los ríos,
que impulsa el crecimiento de algunas de las ciudades modernas. Es el caso particular
del rio Combeima que en sus riberas acoge a la ciudad de Ibagué y brinda gran
cantidad de servicios ambientales a sus habitantes. Este rio nace en el nevado del
Tolima y en su recorrido, de 57.7 km hasta su desembocadura en la cuenca mayor del
rio Coello, drena una extensión de 27.421 ha correspondientes al 18.2 % del área de
la ciudad de Ibagué. En su viaje, cumple funciones básicas como la de abastecer a la
ciudad del 80% del agua requerida por las redes del acueducto municipal, además de
19 satelitales.
De sus aguas son extraídos anualmente 214 millones de m3 del líquido para la
irrigación de 7.000 ha de cultivos altamente tecnificados de arroz y sorgo, localizados
en la meseta de Ibagué (CONPES 3570, 2009; Vanegas, M. 2002). Del rio Combeima
también se derivan algunas actividades económicas como la extracción de materiales
rocosos; otras relacionadas con su valor paisajístico y turístico como en las zonas de
Pastales, Villa restrepo y Juntas, área rural de Ibagué; estéticas y ecológicas como en
el jardín botánico Alejandro Von Humboldt-UT.
No obstante, durante todo su trayecto por la ciudad, el rio recibe gran diversidad y
cantidad de residuos sólidos (plásticos, papel, piedra, animales muertos, recipientes de

11
diferente naturaleza, etc.) y líquidos (solubles e insolubles) que afectan no sólo la
calidad del cuerpo de agua sino también el soporte boscoso que constituye su ribera y
ponen en riesgo la disponibilidad de recursos y servicios ambientales en este
ecosistema.
El material vegetal conocido como bosque ripario, representa la última línea de defensa
para la protección de la calidad del agua y del ecosistema acuático (Robins y Cain
2002), mantiene las condiciones naturales del caudal del rio, evita la degradación del
suelo y perturbaciones en el lecho acuífero, amortigua el ingreso de contaminantes,
regula la temperatura y la entrada de luz. Sin mencionar que constituye una zona con
una altísima biodiversidad de plantas y animales en comparación con otras zonas, es el
refugio de especies vulnerables tanto de plantas, como de animales (Robins y Cain
2002), provee de hábitat a gran cantidad de especies silvestres; aves, reptiles, insectos
y anfibios, entre otros. En general, se trata de un ecosistema muy fértil y productivo,
con un valor ecológico importante en la salud y protección del rio frente a la amenaza
de la deposición de residuos, que podría redundar en una pérdida de los recursos y
servicios ambientales que de él se derivan.
De esta manera, los bosques riparios juegan un papel importantísimo como filtros
naturales, que por medio de su vegetación, sujeta de manera natural a una fuerte
dinámica ocasionada por la influencia del caudal del rio o por la aguas que escurren
superficialmente, ayudan a mitigar la problemática del vertimiento de residuos a estos
ecosistemas (Corbacho et al. 2003). La calidad de las aguas del rio puede verse
mejorada con esta dinámica natural, impidiendo la incorporación de nutrientes a las
aguas y retrasando procesos de eutrofización. Así mismo, en las riberas queda
atrapado un porcentaje muy elevado de sedimentos y con ello se reducen los sólidos
en suspensión de las aguas (Karssies y Prosser, 1999).
Otras contribuciones ecológicas de la dinámica entre el bosque ripario y el cuerpo de
agua, que benefician la salud del rio, es la disminución de la temperatura de las aguas,
que mejora el contenido en oxígeno disuelto en ellas. Gracias al efecto de sombreado

12
que también regula la entrada de luz, favoreciendo el crecimiento de las algas. De
hecho, la vegetación riparia se encuentra más conectada a la vida que sucede dentro
del río que a la que sucede fuera de él; la vegetación nativa que crece en los bancos es
la principal fuente que da vida a los organismos acuáticos de pequeñas quebradas
(Corbacho et al. 2003).
Conscientes de la importancia ecológica que manifiesta el bosque ripario para la salud
de todo el ecosistema del rio, y de las dinámicas naturales que mantiene las
disponibilidad de recursos y servicios ambientales derivados de la buena salud del
mismo, en este trabajo se buscó conocer el comportamiento de la vegetación riparia y
el entorno ecológico de esta frente a la contaminación presente en las aguas del rio
Combeima. Para ello, el trabajo evaluó la capacidad biodegradadora de las bacterias
aisladas de la rizósfera de dos plantas de la ribera del Rio Combeima -Tramo UT para
degradar el N y el P contenido las aguas del rio de interés; buscando con esto
contribuir al mejoramiento de la calidad del más importante acuífero de la ciudad de
Ibagué, además de establecer la fundamentación para un método natural de
recuperación de aguas y mantenimiento del bosque de ribera.

13
1. OBJETIVOS 1.1 OBJETIVOS GENERALES
Seleccionar las dos especies vegetales de mayor importancia ecológica desarrolladas
en la ribera del rio Combeima-tramo UT.
Evaluar la capacidad para solubilizar el fósforo inorgánico Ca3(PO4)2 y transformar el
amonio (NH4+) contenido en el sulfato de amonio (NH4)2SO4 y el nitrito (NO2-) por parte
de las bacterias aisladas de la rizósfera de las dos especies vegetales de mayor
importancia ecológica encontradas en la zona ribereña del rio Combeima-tramo UT.
1.2 OBJETIVOS ESPECÍFICOS
Seleccionar las dos especies de mayor importancia ecológica sobre el bosque ripario
del rio Combeima-tramo UT, mediante la determinación de la cobertura vegetal, la
densidad de especies y la biomasa.
Aislar e identificar las bacterias presentes en la rizósfera de las especies vegetales
seleccionadas.
Seleccionar y evaluar las bacterias con capacidad para oxidar al amonio a nitrito
(nitrosomonas), y así mismo, determinar aquellas con capacidad de transformar el
nitrito a nitrato (nitrobacter).
Establecer la habilidad de los aislados para solubilizar el fosfato inorgánico
suministrado como fosfato tricálcico.
Valorar aspectos fisicoquímicos del agua y del suelo ribereño.

14
Verificar la acción natural de descontaminación de aguas de las bacterias asociadas a
la raíz de las plantas de ribera, con el fin de implementarlo como parte de un sistema
de recuperación de aguas residuales.

15
2. MARCO REFERENCIAL 2.1 MARCO TEÒRICO
2.1.1 El bosque ripario y su funcionalidad en las dinámicas del ecosistema acuático
Un área riparia (perteneciente al banco de un río), es un área encontrada junto o
directamente influenciada por un cuerpo de agua. Se refiere a comunidades bióticas
que viven a ambos lados de los ríos, quebradas, lagos e incluso algunos humedales
(Robins y Cain 2002). Son ejemplos: los ecosistemas a ambos lados de quebradas y
ríos como bancos aluviales y humedales, terrazas de inundación, las cuales interactúan
con el río en tiempos de crecidas o inundaciones; la vegetación dependiente del
suministro de agua en el suelo, la cual es proveída por un río adyacente; ecosistemas
adyacentes a drenajes y canales que desembocan en quebradas ríos o humedales, o
simplemente áreas que rodean lagos (Olson, 2000).
La vegetación riparia o bosque ripario en zonas más conservadas, cubre las márgenes
de los ríos, se desarrollan en suelos donde el nivel freático está influenciado por la
dinámica fluvial, representan una vegetación azonal que corresponde a un ecotono
entre el ecosistema terrestre y acuático. A menudo hay un contraste marcado entre las
especies riparias y las que crecen en suelos zonales y no relacionados
hidrológicamente con los ríos: son especies adaptadas a suelos fértiles y capaces de
resistir la inundación. De esta manera los bosques riparios se convierten en
componente integral de los ríos y tienen una importancia ecológica clave en la
funcionalidad del ecosistema acuático.
El rio y el bosque ripario constituyen una unidad funcional que intercambia materia y
energía en el contexto de su cuenca (Dodds, 2002; Naiman et al., 2005); representa un
ecosistema dinámico del que se derivan recursos y servicios ambientales importantes
como la regulación en los flujos hidrológicos, reducción del impacto de deslaves e

16
inundaciones, disminución de la erosión del suelo, baja de la sedimentación en los
cursos de agua, mantenimiento o mejoramiento de la calidad del agua (filtración de
contaminantes potenciales), de la recarga de acuíferos y de hábitats acuáticos, y
conservación de suelos (CCAD-PNUD/GEF, 2002; Izko y Burneo, 2003; Angelsen y
Kaimowitz, 2001; Landell-Mills y Porras, 2002; Robertson y Wunder, 2005). La figura 1
ilustra la relación entre el bosque de ribera y el ecosistema acuático.
Figura 1 Relación funcional entre el bosque de ribera y el ecosistema acuático
Fuente: Elosegi & Sabater (2009). Conceptos y técnicas en ecología fluvial. FBBVA
La existencia de amplias bandas de bosque ripario con suelos permeables y bajo
coeficiente de escorrentía, debido a la frondosidad natural de la vegetación y la
presencia de residuos orgánicos en sus suelos, retrasa la formación de avenidas y
disminuye considerablemente el porcentaje de agua de lluvia que llegan al cauce del
rio. Favoreciendo de esta manera la recarga de acuíferos y ejerciendo un control de los
sedimentos hacia los cuerpos de agua y de los que son transportados por el cuerpo de
agua. La vegetación riparia tiene la capacidad de retener y aprovechar el fósforo
proveniente de los sedimentos, absorbe gran parte de los nitratos disueltos en las
escorrentías, funcionando como un filtro verde que retiene partículas y nutrientes que
llegan por escorrentía o por vía subsuperficial. Proceso importantísimo que tiene un
efecto directo en la protección y calidad de las aguas (Karssies y Prosser, 1999;
Mander y Hayakawa, 2005).

17
2.1.2 Efecto de la vegetación riparia en la contaminación de los ecosistemas acuáticos
El principal servicio de los ecosistemas fluviales, se enfoca hacia el abastecimiento de
agua para consumo en actividades domésticas y productivas (agricultura, industria y
generación de energía). Sin embargo, estos mismos sectores beneficiados del
ecosistema fluvial (la agricultura, la industria y actividades municipales) generan un
problema de contaminación en los cuerpos de agua y afectación del ecosistema,
debido principalmente a la descarga de gran cantidad de aguas residuales derivadas
de sus actividades (Baker, 2002; Baker et al., 2003,2004; Zhuo et al.,2010). Estas
aguas residuales provocan pérdida de la calidad del agua y en consecuencia un
detrimento de los servicios ecosistémicos.
En zonas urbanas el suministro de materia orgánica, en gran medida por el vertimiento
de aguas residuales, son una pesada entrada de nitrógeno y fósforo, así como una
fuente de bacterias heterotróficas, cuya actividad causa un agotamiento significativo
del oxígeno presente en las aguas que están en contacto con el punto de descarga
(Servais y Garnier, 1993; Garnier et al, 1992 a, b); así por ejemplo: la descarga de
efluentes que contienen altas concentraciones de nitrógeno, causa excesiva demanda
de oxígeno en las aguas receptoras, tiene un efecto tóxico en peces y otros
organismos, generan la eutrofización de ríos y lagos y aumenta la formación de
nitrosaminas que son cancerígenas (Dapena et al., 2004,2006).
Durante mucho tiempo el fenómeno de la eutrofización, se había considerado como un
proceso natural, un resultado de la descarga normal de nutrientes, sedimentos y otros
materiales alóctonos (externos) en los sistemas acuáticos. Actualmente se habla de
“Eutrofización Cultural”, la cual está asociada a la intervención del ser humano (Silvério
2006) con su principal causa la descarga de aguas servidas ricas en nutrientes, que
contribuyen al cambio trófico del cuerpo de agua receptor. Sus efectos son un aumento
de la biomasa y un empobrecimiento de la diversidad, con la proliferación de algas
unicelulares, cianobacterias y macrofitas en exceso en la superficie que impiden que
la luz penetre hasta las profundidades del ecosistema, lo que provoca la imposibilidad

18
de llevar a cabo la fotosíntesis en el fondo de dicho cuerpo de agua y por lo tanto la no
producción de oxígeno libre, al mismo tiempo aumenta la actividad metabólica
consumidora de oxígeno de los organismos descomponedores, que empiezan a recibir
excedentes de materia orgánica generados en la superficie.
El fenómeno de la eutrofización en los ecosistemas acuáticos es debido entonces al
incremento en el suministro y disponibilidad de nutrientes, que se origina de fuentes
externas a las naturales, principalmente de nitrógeno y fosforo; aunque también en
ocasiones, otros minerales como sílice, potasio, calcio, hierro o manganeso tiene
efectos en los procesos biogeofísicos y biológicos que naturalmente ocurren en el
ecosistema acuático. Causando un estrés en el ecosistema que lleva un estado de
equilibrio alterado, una modificación en su funcionamiento y acelerar procesos
indeseables (Chalar 2006).
Se sabe además que la eutrofización en aguas superficiales originada por grandes
descargas de fósforo y nitrógeno, contribuye al mal funcionamiento de los ecosistemas
acuáticos dificultando su utilización adecuada por parte de la sociedad desde el punto
de vista económico, para el consumo humano o simplemente paisajístico. Los suelos
aledaños a aguas con problemas de eutrofización tienen igualmente pocas
posibilidades de ser utilizados en la agricultura y la ganadería por lo que se requiere
implementar medidas tendientes a la reducción de la carga de nutrientes.
Sin embargo, la carga contaminante de materia orgánica que suministran estas aguas
residuales puede regularse mediante un proceso de autodepuración propio de los
cuerpos de agua relacionada con la actividad de los microorganismos, la flora y la
fauna acuática natural que habita en el bosque ripario.
La presencia de vegetación riparia contribuye a la estabilidad de las orillas, con ayuda
del sistema radicular aumenta la cohesión del suelo y su resistencia, a la vez que disipa
la energía y velocidad de las aguas, disminuyendo el riesgo de erosión por la acción
de la corriente. El bosque ripario proporciona además un efecto de sombrío sobre el

19
cuerpo de agua, ayudando a regular la temperatura del agua y manteniéndola bien
oxigenada. Representa una fuente de materia orgánica particulada (hojas, frutos, flores,
ramas) que sirven como recursos alimentarios para organismos loticos. Sirve como
hábitat natural para diferentes especies, que encuentran en él refugio y alimento. Las
raíces de las plantas típicas de los bancos proveen un sitio de refugio para las
especies acuáticas en momentos de crecidas, así como también de depredadores
(Chará, 2003). Los suelos de las zonas riparias proveen de sitios con condiciones
ideales para mamíferos que viven o se refugian en cuevas, así también como para
otros organismos que van desde insectos hasta aves (Robins y Cain 2002).
Por su parte, las bacterias heterotróficas bajo condiciones aerobias son capaces de
oxidar bioquímicamente el material orgánico biodegradable, produciendo dióxido de
carbono, agua, amonio y nueva biomasa (Madigan et al. 2003); pueden asimilar
compuestos directamente o actuar sobre ellos hidrolizándolos, como paso previo a su
incorporación. En el proceso de hidrolisis microbiana las macromoléculas como por
ejemplo aminoácidos, péptidos, monosacáridos y largas cadenas de ácidos grasos son
hidrolizadas a oligómeros y monómeros por la actividad microbiana antes de ser
posteriormente degradadas. La hidrólisis microbiana, por tanto, es un paso clave para
la degradación de la materia orgánica en los procesos de depuración de las aguas
(Dueholm et al. 2001, Morgenroth et al. 2002). Esta hidrólisis es realizada por
exoenzimas excretadas por microorganismos hidrolizantes (Gessesse et al. 2003).
Existen también bacterias quimioautótrofas que obtienen su energía para el crecimiento
y desarrollo de elementos constituyentes de la materia orgánica (nitrógeno, azufre,
hierro o hidrógeno). Algunas de estas bacterias (nitrosomonas y nitrobacterias) son
importantes para el proceso de la nitrificación en el cual el amonio se convierte en
nitratos y en una etapa posterior, la denitrificación del nitrato se produce óxido nitroso y
gas nitrógeno proceso importante para la degradación de este contaminante. (Edwards,
2000)

20
Algunas bacterias descomponen el material orgánico por acción mutualista con las
plantas, obtienen los nutrientes necesarios (compuestos del carbono) y el oxígeno de la
planta hospedera; a su vez ésta recibe compuestos del proceso de mineralización
microbiana. Estas bacterias se encuentran asociadas principalmente en la raíz de las
plantas y se denominan rizobacterias. Estas, particularmente tienen una importante
contribución en la degradación y remoción de contaminantes (Chaudhry et al. 2005);
han sido reportadas numerosas bacterias aisladas de la rizosfera de plantas por sus
capacidades degradativas (Atterby et al. 2002).
2.2 ESTADO DEL ARTE
La contaminación de las aguas de los ríos, quebradas, arroyos y aguas subterráneas,
constituye un grave problema global en el contexto actual, especialmente porque los
residuos de las actividades humanas son vertidos directamente a estos ecosistemas
sin tratar o con tratamientos deficientes. Una de las principales descargas a estos
ecosistemas son las aguas municipales compuestas de materia orgánica, nutrientes
(fundamentalmente nitrógeno y fósforo), cantidades traza de compuestos orgánicos
recalcitrantes y metales (Bitton, 2005), que crean graves problemas ambientales como
procesos de eutrofización, perdida de la diversidad en estos ecosistemas y lo más
importante la perdida de utilización del recurso.
La capacidad de distinguir los efectos individuales y acumulativos de los controles
bióticos (microorganismos nativos y vegetación riparía), físicos y químicos de la
excesiva productividad en los ecosistemas acuáticos eutrofizados por la abundancia
de materia orgánica antropogénica, es clave para entender, predecir, y gestionar este
problema.
La posibilidad de mejoramiento de los ecosistemas acuáticos mediante el empleo de
microorganismos de ambientes contaminados como herramienta biotecnológica que
pueda ser aplicada en la rectificación de aguas contaminadas es ampliamente utilizada
(Wagner y Loy, 2002; Gentry et al., 2004). Las posibilidades de uso de estos

21
organismos son derivadas de su capacidad adaptativa, la que, a su vez, está asociada
a la habilidad para degradar una gran variedad de compuestos orgánicos, inorgánicos
y sintéticos (Fabiano et al. 2003; Llagas y Gómez 2006). Aunque para algunos autores
la implementación de estos métodos biológicos tendientes a disminuir la carga orgánica
en cuerpos de agua no es fácilmente predecible (Bitton 2005).
Lo anterior podría explicarse, al menos parcialmente, la utilización de tratamientos
fisicoquímicos, ya sea como método alternativo o complementario a las biotecnologías,
ayudan a la remoción parcial de la carga orgánica; pero el costo de los reactivos y la
alta demanda química de oxígeno resultante con estos procesos, hace necesario el
empleo de otros métodos (Vidal et al., 2000). Así por ejemplo, se habla de que el
tratamiento biológico para remoción del fósforo representa una opción con una relación
costo-efectividad mejor que los tratamientos fisicoquímicos de precipitación de la carga
orgánica (Reardon, 1994). En el tratamiento de un agua residual utilizando radiación
gamma los resultados mostraron que ésta es una buena opción, pero acoplado a un
tratamiento físico previo y su aplicación se da a los contaminantes remanentes del
proceso. Se señala además que la utilización de los equipos que emiten este tipo de
radiación son costosos (Medina y Gómez 2009).
Se insiste entonces en las técnicas biorremediadoras con resultados y perspectivas
prometedoras; así por ejemplo, Rodríguez et al (2011), evidenciaron la remoción de
materia orgánica y de nitrógeno amoniacal por acción de bacterias nitrificantes y
desnitrificantes en un reactor tipo batch. Los resultados revelaron una densidad
microbial de aproximadamente 2.35×107 NMP/100 ml para las bacterias nitrificantes y
de 2.25×107 NMP/100 ml para las desnitrificantes; una eficiencia en el proceso del
99%, 98% y 71% en el DQO, DBO5 y N–NH+ 4, respectivamente.
Moreno, Naranjo y Koch (2010), evaluaron la capacidad de cianobacterias mediante
dos tipos de tratamiento (suspensión e inmovilización en alginato de calcio) para la
remoción de nitrógeno amoniacal, fósforo total y disminución de la demanda química de

22
oxígeno (DQO) en agua residual semisintetica y sintética, donde se demostró la
eficiencia del cultivo inmovilizado de Nostoc sp. para la reducción nitrógeno y fósforo.
Salgado et al (2012), comprobaron la capacidad metabólica de rizobacterias aisladas
de Typha dominguensis un planta hidrófita, para degradar distintos componentes de
una agua contaminada por materia orgánica y disminuir la DQO de un agua residual
sintética compleja. Lo que demuestra el potencial de este tipo de microorganismos para
su incorporación en un sistema de tratamiento de aguas y contribuir a la remediación
de efluentes contaminados por materia orgánica.
Huang et al. (2013) al igual que Ho et al. (2013), realizaron ensayos en los que
demostraron la efectividad y aplicabilidad de las técnicas de fitorremediacion, llegando
a evidenciarse su alto potencial para reducir la polución por materia orgánica en
ambientes contaminados. Los investigadores concluyeron que los microorganismos de
la rizósfera juegan papel importante en los procesos fitorremediadores.
Además, recientes estudios evidencian el uso potencial de bacterias endófitas aisladas
de ambientes contaminados, por ser viables en la degradación de distintos
contaminantes y por otorgar resistencia a la planta a estos contaminantes, en los
sistemas de fitorremediacion (Porteus-Moore et al., 2006; Van der lelie, 2005;
Lodewyckx et al., 2002).
En el caso particular de Colombia cabe mencionar que el desarrollo de tratamientos
para la recuperación de aguas residuales abarca un gran número de investigaciones
enfocadas fundamentalmente hacia dos alternativas de aplicación: el tratamiento
fisicoquímico y el biológico; la integración de ambos procesos parece reportar una
mejor eficiencia en la remoción de contaminantes.
Ejemplos del avance en materia de investigación en estos campos son el trabajo
realizado por Acelas et al. (2014), quienes crearon un medio hibrido con una resina
microporosa de intercambio anionico con óxidos hidratados de Fe, Zr y Cu

23
inmovilizado para la remoción de fosfatos en aguas residuales. Bustillo, Mehrvar y
Quiñones (2013), evaluaron el rendimiento y capacidad de un tratamiento combinado
de un reactor anaeróbico, un reactor de lodos activados con aeración y un proceso de
UV/H2O2 para la remoción de COT en aguas residuales. El proceso integrado de ABR-
AS-UV/H2O2 alcanzan un rango óptimo de eliminación de COT de 92,46% a un TRH
de 41 h. Mendoza, Osorio y Benítez (2010), combinaron un sistema solar photo-fento y
biológico, que logró la mineralización y degradación de pesticidas (ácido 2,4-
diclorofenoxiacético y Diuron) en aguas residuales del cultivo de caña, con una
mineralización de 79.8% en el agua residual preparada y 82,5% en las aguas
residuales industriales.
La biorremediación también ha despertado el interés en algunos estudiosos del tema
de descontaminación de un agua residual en nuestro país, destacando el uso de
microalgas (Chlorellavulgaris, Scenedesmusdimorphus) y macrofitos (Lemna
minúscula) dada la eficiencia en la reducción de amonio, fósforo y DQO en aguas
residuales con sustancias recalcitrantes (Valderrama et al., 2002; González, Cañizares
y Baena, 1997).
No obstante, en el caso particular del Departamento del Tolima son limitadas las
investigaciones en la temática en cuestión. Una revisión de literatura relacionada con
en el tema que nos ocupa dejó ver algunos trabajos, en su mayoría desarrollados en la
Universidad del Tolima, en los que se aplican métodos de biorremediación en aguas
residuales industriales (Flórez y Cotes, 2006; Medina, 2013).
Estas investigaciones, y otras no menos importantes, inducen a pensar si es posible
lograr remover parte de la materia orgánica de las aguas del rio Combeima, mediante
la utilización de bacterias aisladas de las raíces de algunas de las especies vegetales
que se desarrollan en la ribera del cuerpo hídrico más importante para la ciudad de
Ibagué.

24
3. MATERIALES Y METODOS 3.1 ZONA DE ESTUDIO
La zona de trabajo corresponde a un área de bosque ripario que se extiende por una
de las orillas del rio Combeima en su paso por el Jardín botánico Alexander von
Humboldt-Universidad del Tolima (4º 25´49.5´´ N y 75º 12´58.6´´ W, 1170 y 1070
m.s.m, 22.5 °C) y que acompaña el recorrido del rio e influye de manera directa en sus
dinámicas. En este trabajo se determinó llamar esta área del bosque “Bosque ripario rio
Combeima-tramo UT” (BRC-UT). La figura 2 ilustra la ubicación del área de muestreo y
algunas particularidades de la misma.
Figura 2. Ilustración de la zona de estudio
Fuente: Pagina web universidad del Tolima (2015)
Zona de estudio (BRC-UT)
Jardín botánico AVH
Rio Combeima

25
3.2 MATERIALES
Se trabajó con la vegetación riparia típica de la zona anteriormente descrita, utilizando
principalmente arbustos y arvenses desarrollados a lo largo de 50 m de la ribera y 5 m
de profundidad del cauce del rio hacia adentro. Adicionalmente, se trabajó con el suelo
ribereño y el agua del rio, como una manera de complementar la información obtenida
a través del material vegetal.
Para todos los ensayos se trabajó con reactivos analíticos de alto grado de pureza. Los
medios de cultivo fueron selectivos para las pruebas aplicadas.
3.3 METODOLOGIA
3.3.1 Evaluación de la composición florística del BRC-UT:
La selección de las especies vegetales se inició con una visita inspeccional a fin de
determinar la composición florística de la zona. Se realizó un muestreo de las especies
vegetales presentes en 50 m2 del BRC-UT, dividiendo el área de interés en 5
transeptos de 10 m x 1 m con el fin de entender la distribución de las especies a lo
largo de la ribera del rio. La selección de los transeptos se hizo considerando lugares
en los que las especies se encontraran influenciadas por el cauce del rio y
desarrolladas en el suelo particular de este; se tuvieron en cuenta puntos en los que la
diversidad era evidente y en los que prevalecían tanto arvenses como arbustos. En
cada uno de los transeptos se colectó un individuo de cada especie y se llevó a cabo
un conteo del número de individuos para cada caso, se tomaron medidas de cobertura
vegetal, área basal, altura y también se determinó la biomasa de cada una de la
especies. La clasificación taxonómica de los individuos colectados se realizó en el
herbario Toli de la Universidad del Tolima.

26
3.3.2 Selección de las especies vegetales de mayor importancia ecológica en el BRC-
UT:
Para seleccionar las dos especies de mayor importancia ecológica del BRC-UT, se
evaluó en el material vegetal encontrado en la zona parámetros ecológicos que
argumentaran un mejor desarrollo y adaptación a las condiciones particulares de esa
zona de transición. Se determinaron en cada especie colectada en los 5 transeptos los
parámetros de: cobertura vegetal, representada por la cobertura foliar y el área de
suelo ocupado, así como la biomasa y densidad de la especie. Cada parámetro
ecológico se expresó mediante un porcentaje relativo al área total de muestreo, la
sumatoria de estos valores relativos permitió establecer la importancia ecológica para
cada especie, lo cual se define como Índice de Valor de Importancia (IVI).
3.3.3 Caracterización fisicoquímica del suelo ribereño y del agua del rio:
El suelo ribereño y el agua del rio se caracterizaron mediante la determinación de
algunos parámetros fisicoquímicos (% materia orgánica, % de nitrógeno, mineralogía,
textura y solidos totales, DQO, nitrógeno amoniacal, fosfatos y fosforo total
respectivamente) siguiendo las metodologías aplicadas en el laboratorio LASEREX-
Departamento de Química-UT, las cuales se apoyan en los métodos analíticos del
laboratorio de suelos IGAC (Instituto geográfico Agustín Codazzi, 2006) y los métodos
normalizados para el análisis de aguas potables y residuales (Standard Methods for the
Examination of Water and Wastewater, 1989), respectivamente.
3.3.4 Aislamiento de las rizobacterias asociadas a las plantas seleccionadas: Para el aislamiento de las rizobacterias, se colectaron 3 especímenes de cada especie
seleccionada que estuvieran en relativa influencia con el agua del rio, cuidando no
remover el suelo adherido a sus raíces. Las muestras fueron guardadas en bolsas
debidamente etiquetadas para su traslado al laboratorio en donde se sometieron a un
proceso para la separación de las bacterias, el cual se puede resumir así:
Se removió el suelo débilmente adherido a las raíces, y el fuertemente unido a la raíz
se retiró utilizando 100 ml de agua destilada, la suspensión agua-suelo se sometió a
un proceso de extracción por agitación manual durante 30 min, después de lo cual se

27
dejó sedimentar. Del extracto obtenido se tomaron 100µL y se llevaron a 1000 µL (10-1
µL de extracto/mL solución) con agua destilada, esta se constituyó en la solución
madre, a partir de la cual se prepararon diluciones seriadas desde 10-3 a 10-5 µL/mL.
De cada una de estas diluciones se tomaron 100 µL que se sembraron en cada caja
de Petri, realizando 3 réplicas de siembra en placa en agar nutriente y se incubaron por
48 h (30 °C).Se seleccionaron colonias con características visibles diferentes de cada
siembra; los aislados se caracterizaron teniendo en cuenta su morfología y respuesta
fisiológica a la tinción de Gram y la observación al microscopio.
3.3.5 Aislamiento de bacterias endófitas de la raíz asociadas a las plantas
seleccionadas:
Para el aislamiento de las bacterias endófitas, se trabajó con la raíz tratada
previamente para el aislamiento de la rizobacterias como lo describió Pérez et al.
(2010). La raíz limpia se sometió a un proceso de desinfección consistente en lavar la
raíz con agua destilada autoclavada por lo menos 3 veces, se llevó a inmersión en
alcohol al 70% durante 10 min con agitación manual, seguidamente se hizo inmersión
en solución de hipoclorito de sodio (NaOCl) al 5% con agitación, lavando nuevamente
la raíz por lo menos 3 veces con agua destilada autoclavada, finalmente la raíz se
expuso 10 min a la luz ultravioleta (λ 256 nm) para concluir de esta forma con el
proceso de desinfección.
La raíz ya desinfectada se cortó en pequeños fragmentos y se maceró en un mortero
estéril adicionándole un volumen de 2 ml de buffer fosfato. El extracto obtenido,
producto del macerado, se pasó a un frasco estéril; se tomaron luego 100 µl del
extracto y se llevó a 1000 µL (10-1 µL/mL) con agua destilada, obteniendo una
solución madre, a partir de la cual se prepararon diluciones seriadas hasta 10-5. µL/mL.
Finalmente se sembraron sobre cajas de Petri conteniendo agar nutriente, 100 µl del
extracto concentrado y de las disoluciones 10-1 hasta la 10-5 µL/mL. Una vez absorbido
el volumen sembrado por la superficie del medio se sellaron las cajas con papel vinipel
y se llevaron a la incubadora a 30 °C para el crecimiento.

28
3.3.6. Selección de las bacterias solubilizadoras de fosfatos: Para evidenciar la capacidad de solubilización de fosfatos inorgánicos se realizó una
prueba cualitativa que inició con la repicación de las bacterias aisladas en medio
Sundara, Rao y Sinha (SRS) (1963), utilizando cajas de Petri que contenían medio de
cultivo sólido SRS con fosfato tricálcico, las cuales se sometieron a incubación (30°C,
5-7 días). Finalizado el tiempo se seleccionaron las cepas bacterianas que mostraron
crecimiento con acidificación del medio de cultivo y formación de halo de hidrólisis
(cambio de color de morado a amarillo).
La capacidad solubilizadora del fósforo se corroboró mediante otro ensayo cualitativo
que además permitió escoger las cepas más funcionales dentro del grupo que dejó ver
la bioactividad en el ensayo anterior.
El ensayo de verificación consistió en realizar dos pozos de 6.6 mm de diámetro c/u en
una caja de Petri que contenía medio SRS con fosfato tricálcico; se adicionaron 25 μl
de suspensión de bacterias a una concentración 0.5 McFarland (1.5 x 10-8 UFC/ml) en
cada pozo. Las cajas sembradas, se llevaron a incubación (30°C), al término del tiempo
se evaluó el diámetro, en mm, del halo de hidrólisis formado (cambio de color de
morado a amarillo). Las mediciones se realizaron a las 24 y 48 h después de la
siembra, teniendo en cuenta la medida del diámetro del pozo para descontarlo;
aquellas que presentaron mayor diámetro fueron seleccionadas para continuar con
ellas las restantes pruebas (Martínez y García, 2002).
3.3.7 Determinación de la capacidad bacteriana para solubilizar el fósforo: Se cuantificó la habilidad bacteriana para convertir el fosfato tricálcico insoluble en
fósforo soluble a través del método del fosfomolibdeno, utilizando el Test de fosfatos
(PMB) MERCK. Se trabajó con las 3 cepas consideradas como las de mayor capacidad
solubilizadora. El inóculo bacteriano se ajustó a una concentración de 0.5 McFarland
(1.5 x 10-8 UFC/ml). 1ml del inóculo se sembró en tubos con 24 ml de medio SRS
líquido que contenía la sal de fósforo sin indicador de pH. Los tubos con su contenido
se incubaron con agitación (30°C, 150 rpm, 5 horas/día). Simultáneamente se

29
prepararon controles de cada uno los tratamientos con 25 ml de medio sin inóculo y se
incubaron bajo las mismas condiciones que las muestras. Las lecturas del fósforo
disponible en el medio de cultivo se realizaron a los 4, 8 y 12 días de incubación;
además, se realizó una lectura del medio liquido SRS a los 0 (cero) días, en tanto que
las muestras fueron centrifugadas a 10.000 rpm (10 min), y con el sobrenadante se
hizo la medición. Los valores obtenidos son la media de dos determinaciones.
3.3.8 Selección de las cepas bacterianas con capacidad de transformar el amonio:
Para determinar la capacidad de nitrificación de las cepas aisladas de la raíz de las
plantas seleccionadas, se sembró una unidad formadora de colonia en tubos de ensayo
con 3 ml de caldo amonio, se garantizó la presencia de oxígeno para el proceso de
nitrificación, utilizando un tapón de algodón que permitiera solamente el flujo de este y
evitara la contaminación del medio. Los tubos inoculados se incubaron a 28°C durante
dos semanas. Transcurrido este periodo de tiempo, se evaluó el crecimiento de las
bacterias para trasformar el nitrógeno contenido en (NH4)2SO4 en las formas oxidadas
del nitrógeno (NO2- y NO3-). Para la detección de nitritos (NO2-), se adicionaron 2
gotas del reactivo de Griess, que desarrolla una coloración roja después de 5 minutos
en los tubos de respuesta positiva. Los tubos que no presentan cambio de coloración
(negativos), se les adiciona zinc en polvo para detectar la presencia de nitrato en el
medio, los tubos positivos toman coloración rojiza naranja; los que no cambian fueron
tomados como negativos, a los cuales se les adicionó el reactivo de Nessler para
confirmar la presencia de amonio, indicativo de que no existió proceso de nitrificación.
3.3.9 Selección de cepas con capacidad de trasformación del nitrito: Para determinar la capacidad de utilización del nitrito como fuente de energía por parte
de las bacterias aisladas, se realizó un repique de cada una de las cepas en cajas de
Petri con medio autotrófico para Nitrobacter 756c (DSMZ). Las cajas se incubaron a
27°C por 14 días. Después del periodo de incubación se evidenció el crecimiento típico
en forma de gotas de agua.

30
3.3.10 Ensayo de verificación de biorremediación en un agua residual sintética:
Para verificar la acción biorremediadora de las cepas seleccionadas en las anteriores
pruebas que mostraron capacidad de solubilización del fósforo inorgánico y
trasformación del nitrito (nitrificantes) se realizó un bioensayo, utilizando un agua
residual de concentración media, cuya composición aparece relacionada en la tabla 1.
Tabla 1. Composición del agua residual sintética
Compuesto Concentración (mg/L)
Peptona 160 mg/L
NaNO2 30 mg/L
KH2PO4 28 mg/L
MgSO4.7H2O 2 mg/L
CaCl2.H2O 4 mg/L
NaCl 7 mg/L
Para el bioensayo se trabajó con 200 ml de agua residual sintética contenida un frasco
de vidrio estéril, a los cuales se inoculó 1 ml de suspensión bacteriana (1.5 x 10-8
UFC/ml, equivalente a 0.5 McFarland). Se utilizó como control, un frasco con agua
residual sintética sin inóculo de bacteria. Se utilizaron 3 cepas bacterianas que
mostraron la mejor capacidad de trasformación del nitrito a nitrato y 3 cepas con la
mayor habilidad de solubilización del fósforo inorgánico. Los frascos utilizados en la
prueba de las bacterias nitrificantes se cubrieron con papel aluminio para impedir la
entrada de luz; el flujo de oxigeno se garantizó tapando la boca del frasco con gaza
estéril. Después de un periodo de 15 días se verificó la disminución de los parámetros
DQO, fosfatos y nitritos, al contrastar la lectura inicial y final en cada caso. Cada
ensayo se realizó por triplicado.
3.3.11 Análisis estadístico: Con los resultados de la determinación del fósforo disponible se realizó un ANOVA
(GLM) utilizando el paquete estadístico Minitab 15, buscando comprobar el efecto de

31
diferentes factores; tiempo del ensayo, tratamientos, réplica y sus interacciones, sobre
la cuantificación del fósforo disponible (PO4) durante el ensayo.
Se construyó el modelo estadístico:
𝑦𝑦 = 𝜇𝜇 + 𝜏𝜏𝑖𝑖 + 𝛽𝛽𝑗𝑗 + 𝛾𝛾𝑘𝑘 + (𝜏𝜏𝑖𝑖𝛽𝛽𝑗𝑗)𝑖𝑖𝑗𝑗 + (𝜏𝜏𝑖𝑖 𝛾𝛾𝑘𝑘)𝑖𝑖𝑘𝑘 + �𝛽𝛽𝑗𝑗𝛾𝛾𝑘𝑘�𝑗𝑗𝑘𝑘 + 𝜀𝜀𝑖𝑖𝑗𝑗𝑘𝑘
Donde,
Y = cantidad de fosfato
μ = cantidad de fosfato promedio
Τi = efecto producido por el tiempo
βj = efecto producido por el tratamiento
γk= efecto producido por la replica
(Τi βj)ij = interacción tiempo-tratamiento
(Τi γk)ik = interacción tiempo-replica
(βj γk)jk = interacción tratamiento-replica
εijk = error aleatorio
Que mostró ser el de mejor normalidad de los datos y el más adecuado para demostrar
el efecto de los diferentes factores.

32
4. RESULTADOS Y DISCUSION 4.1 COMPOSICIÓN FLORÍSTICA DEL BRC-UT
4.1.1 Clasificación taxonómica del material vegetal encontrado en la zona de estudio: La tabla 2 muestra el listado de las especies vegetales encontradas en el BRC-UT,
agrupadas por familias, nombre científico de la entidad botánica, número de individuos
contabilizados para cada caso y el código de referencia para la determinación
taxonómica, dado por el Herbario Toli-UT.
Tal como muestra la tabla 2, en los 5 transeptos muestreados del BRC-UT (50 m2) se
encontraron 29 especies vegetales, en su mayoría arbustos y arvenses, distribuidas en
17 familias. Su clasificación taxonómica se realizó mediante comparación con
especímenes tipo encontrados en la colección del herbario Toli. Se destacan por la
abundancia de especies las familias Asteraceae (4 especies en su mayoría herbáceas)
y Poaceae (5 especies entre arbustos y herbáceas), las restantes muestran diversidad
entre uno y dos especies. Dos especímenes colectados, “especie 4 y 13”, no pudieron
ser clasificados correctamente, logrando sólo determinar la familia a la que
pertenecen.
Pseudelephantopus spiralis resultó ser la especie de mayor representatividad dentro de
las Asteráceas con 180 individuos inventariados, desarrollados muy cercanos al lecho
del rio y formando un tapete de vegetación que cubría parte del lecho del rio. En su
mayoría estas herbáceas se encontraban en estado de floración.
Por su parte, de Pennisetum purpureum (familia Poaceae) se inventariaron 328
individuos, en su mayoría arbustos; a diferencia de las Asteráceas encontradas, éstos
se extendían más allá de la ribera del rio logrando distribuirse de una mejor manera y
evidenciado un domino en la zona de estudio.

33
Tabla 2. Clasificación taxonómica de los individuos colectados en el BRC-UT organizados por familia, especie y número de individuos por especie
Familia Especie N° individuos
Cód. individuo comparado
(Herbario Toli)
Asteraceae
Pseudelephantopus spiralis (Less.) Crong. 180 6686
Baccharis trinervis Persoon 6 6463 Ageratum conyzoides L. 7 01 0305 Synedrella sp. 50 01 0305
Euphorbiaceae
Acalypha diversifolia Jacq. 10 000 602 Especie 4 1
Malvaceae Sida rhombifolia L. 1 0104 58
Poaceae
Pennisetum purpureum Schumach. 328 0104 70 Pseudochinolaena polystachya (H.B.K). Stapf 74 000 570
Sporobolus pyramidalis P. Beauv. 1 000 547 Gynerium sagitattum (Aubl.) Beaun. 23 00 6052 Especie 13 50 Amaranthacea
e Cyathula achyrantoides (Kunth) Moq 13 00 9139
Zingiberaceae Renealmia sp. 26 00 8926 Piperaceae
Piper aduncum L. 7 00 9831 Piper angustifolium Lam. 5 00 8785
subfami. Faboideade Arachis pintoi Krapov & W.C Greg 9 010 520
subfami. Mimosoide
Zygia longifolia (H. & B.) Britton & Rose 5 00 9070
Acanthaceae Justicia sp. 19 07 943 Commelinacea
e
Tripogandra serrulata (Vahl) Handlos 60 00 8247
Commelina diffusa Burm. F. 6 01 0293
Sapindaceae Cupania cinérea Poepp. 1 000 927 Costaceae costus sp. 4 Lauaracea Persea sp. 1 00 3128 Lythraceae Adenaria floribunda H.B.K 2 000 940 Solanaceae Solanum sp. 2
Passifloraceae Pssiflora sp. 2 07 661 - Blechnum sp. 24 - Pteris sp. 6 Número total de individuos (N) 923 Número total de especies (S) 29 Fuente: autores

34
La distribución de las entidades botánicas colectadas en los 5 transeptos muestreados,
aparece consignada en la tabla 3.
Tabla 3. Distribución de las especies encontradas en cada uno de los transeptos.
Especie N° de individuos/ transepto T1 T2 T3 T4 T5
Pseudelephantopus spiralis (Less.) Crong. 30 144 6 - - Acalypha diversifoliaJacq. 4 - 6 - -

35
T1 a T5: Transeptos Fuente: autores
Aunque la tabla deja ver heterogeneidad en la distribución florística, algunas especies
fueron encontradas en más de 3 transeptos, es el caso de Pseudelephantopus
spiralis, Pennisetum purpureum, Pseudochinolaena polystachya, Baccharis trinervis,
Piper angustifolium, Tripogandra serrulata.
La tabla también muestra que en los transeptos T2, T3 y T4 aparece un número de
especies semejante (13 a 14), en tanto que T1 fue el transepto con casi la mitad de
Sida rhombifolia L. 1 - - - - Pennisetum purpureum Schumach. 64 25 39 - 200 Pseudochinolaena polystachya (H.B.K). Stapf 37 1 30 6 - Cyathula achyrantoides (Kunth) Moq 1 - - 12 - Renealmia sp. 16 - - 10 - Sporobolus pyramidalis P.Beauv. 1 - - - - Baccharis trinervis Persoon - 1 3 2 - Blechnum sp. - 24 - - - Pteris sp. - 6 - - - Piper aduncum L. - 2 - 5 - Piper angustifolium Lam. - 3 1 1 - Arachis pintoi Krapov& W.C Greg - 9 - - Zygia longifolia (H. & B.) Britton & Rose - 4 1 - - Justicia sp. - 4 - 15 - Gynerium sagitattum (Aubl.) Beaun. - 23 - - Tripogandra serrulata (Vahl) Handlos - 7 25 28 - Especie 4 - - 1 - - Cupania cinérea Poepp. - - 1 - - Commelina diffusaBurm. F. - - 6 - - costus sp. - - 4 - - Persea sp. - - 1 - - Adenaria floribunda H.B.K - - 1 1 - Solanum sp. - - - 2 - Ageratum conyzoides L. - - - 7 - Pssiflora sp. - - - 2 - Especie 13 - - - 50 - Synedrella sp. - - - 50 - Número total de individuos (N) 154 253 125 191 200 Número total de especies (S) 8 13 14 14 1

36
especies de los anteriores y, por su parte, T5 presentó uniformidad total ya que sólo fue
hallada una sola especie (Pennisetum purpureum Schumach).
4.2 SELECCIÓN DE LAS DOS ESPECIES VEGETALES CON MAYOR IMPORTANCIA ECOLÓGICA EN LA ZONA DE ESTUDIO
La escogencia de las dos especies vegetales de mayor representatividad en la zona
riparia muestreada, se realizó con base en la determinación de algunos parámetros
ecológicos como lo muestra la tabla 4; los valores consignados corresponden a la
media de 3 determinaciones para cada caso.
Importa aclarar que como consecuencia de las notorias diferencias observadas en el
tamaño de los individuos que conformaban la población vegetal, se determinó registrar
la altura de los 3 especímenes tomados al azar de cada especie, para determinar la
biomasa representativa de ella, expresada como peso seco. De igual manera, el
diámetro promedio de la base (DB) para cada especie da a entender un valor de
espacio de suelo ocupado por la especie, expresado como porcentaje.
La abundante información que deja ver la tabla obligó a agruparla, así:
• El valor promedio de cada parámetro ecológico, por ejemplo DB, se multiplicó
por el número de individuos por especie en el transepto, el dato obtenido
correspondió a un valor de área de suelo cubierto.
• Aplicando esta misma metodología se calcularon valores para el área foliar y la
biomasa
• Cada valor obtenido se expresó en porcentaje con respecto al área total
estudiada
• Adicionalmente se tuvo en cuenta, la densidad de cada especie, expresada en
porcentaje.
Tabla 4. Medición de los parámetros ecológicos (diámetro de la base (DB), cobertura foliar (CF) altura y biomasa (peso seco)) en las especies vegetales del BRC-UT.
Especie
Cobertura vegetal (diámetro cm)
Altura (cm)
W
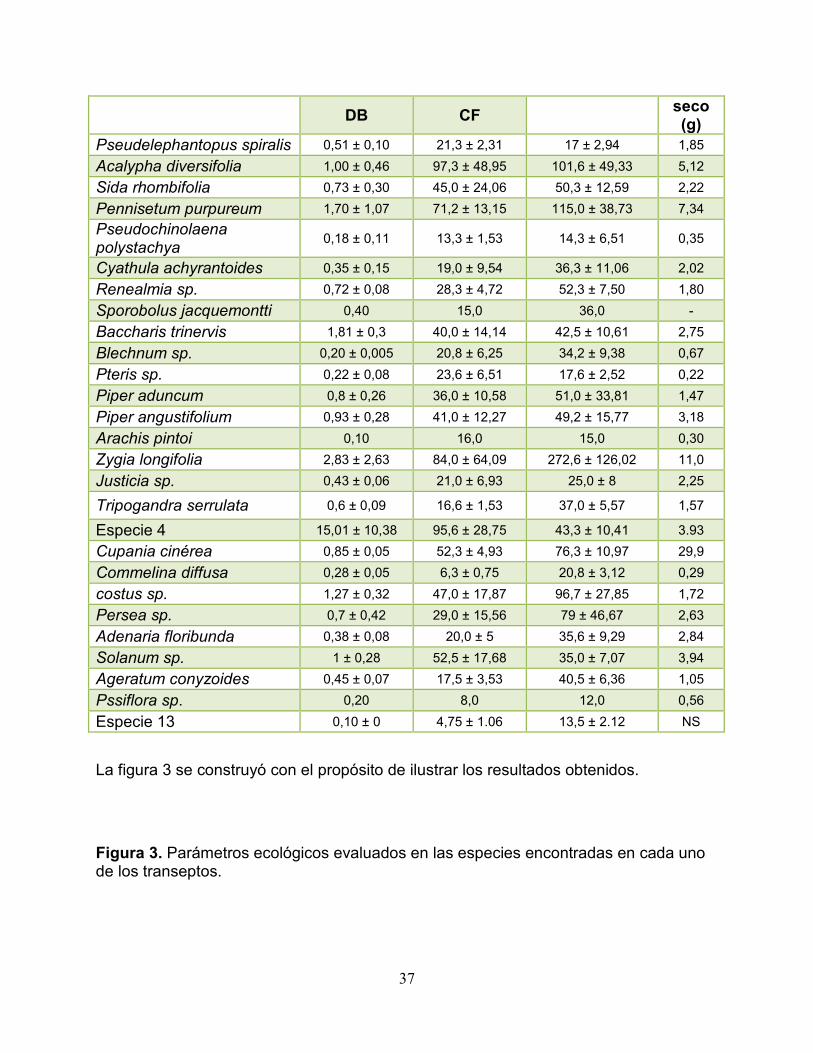
37
La figura 3 se construyó con el propósito de ilustrar los resultados obtenidos.
Figura 3. Parámetros ecológicos evaluados en las especies encontradas en cada uno de los transeptos.
DB CF seco (g)
Pseudelephantopus spiralis 0,51 ± 0,10 21,3 ± 2,31 17 ± 2,94 1,85 Acalypha diversifolia 1,00 ± 0,46 97,3 ± 48,95 101,6 ± 49,33 5,12 Sida rhombifolia 0,73 ± 0,30 45,0 ± 24,06 50,3 ± 12,59 2,22 Pennisetum purpureum 1,70 ± 1,07 71,2 ± 13,15 115,0 ± 38,73 7,34 Pseudochinolaena polystachya 0,18 ± 0,11 13,3 ± 1,53 14,3 ± 6,51 0,35
Cyathula achyrantoides 0,35 ± 0,15 19,0 ± 9,54 36,3 ± 11,06 2,02 Renealmia sp. 0,72 ± 0,08 28,3 ± 4,72 52,3 ± 7,50 1,80 Sporobolus jacquemontti 0,40 15,0 36,0 - Baccharis trinervis 1,81 ± 0,3 40,0 ± 14,14 42,5 ± 10,61 2,75 Blechnum sp. 0,20 ± 0,005 20,8 ± 6,25 34,2 ± 9,38 0,67 Pteris sp. 0,22 ± 0,08 23,6 ± 6,51 17,6 ± 2,52 0,22 Piper aduncum 0,8 ± 0,26 36,0 ± 10,58 51,0 ± 33,81 1,47 Piper angustifolium 0,93 ± 0,28 41,0 ± 12,27 49,2 ± 15,77 3,18 Arachis pintoi 0,10 16,0 15,0 0,30 Zygia longifolia 2,83 ± 2,63 84,0 ± 64,09 272,6 ± 126,02 11,0 Justicia sp. 0,43 ± 0,06 21,0 ± 6,93 25,0 ± 8 2,25
Tripogandra serrulata 0,6 ± 0,09 16,6 ± 1,53 37,0 ± 5,57 1,57
Especie 4 15,01 ± 10,38 95,6 ± 28,75 43,3 ± 10,41 3.93 Cupania cinérea 0,85 ± 0,05 52,3 ± 4,93 76,3 ± 10,97 29,9 Commelina diffusa 0,28 ± 0,05 6,3 ± 0,75 20,8 ± 3,12 0,29 costus sp. 1,27 ± 0,32 47,0 ± 17,87 96,7 ± 27,85 1,72 Persea sp. 0,7 ± 0,42 29,0 ± 15,56 79 ± 46,67 2,63 Adenaria floribunda 0,38 ± 0,08 20,0 ± 5 35,6 ± 9,29 2,84 Solanum sp. 1 ± 0,28 52,5 ± 17,68 35,0 ± 7,07 3,94 Ageratum conyzoides 0,45 ± 0,07 17,5 ± 3,53 40,5 ± 6,36 1,05 Pssiflora sp. 0,20 8,0 12,0 0,56 Especie 13 0,10 ± 0 4,75 ± 1.06 13,5 ± 2.12 NS

38
A1) A2) A1. sp1 (Pseudelephantopus spiralis), sp2 (Acalypha diversifolia), sp3 (Sida rhombifolia), sp4 (Pennisetum purpureum), sp5
(Pseudochinolaena polystachya), sp6 (Cyathula achyrantoides), sp7 (Renealmia sp.), sp8 (Sporobolus jacquemontti)
A2. sp1 (Baccharis trinervis), sp2 (Pseudelephantopus spiralis), sp3 (Blechnum sp.), sp4 (Pteris sp.), sp5 (Piper aduncum), sp6
(Piper angustifolium), sp7 (Pennisetum purpureum), sp8 (Pseudochinolaena polystachya), sp9 (Arachis pintoi), sp10 (Zygia
longifolia), sp11 (Justicia sp.), sp12 (Tripogandra cumanensis)
A3) A4) A3. sp1 (Tripogandra cumanensis), sp2 (Acalypha diversifolia), sp3 (Piper angustifolium), sp 4 (Especie 4), sp5 (Cupania cinérea),
sp6 (Pseudochinolaena polystachya), sp7 (Commelina diffusa), sp8 (costus sp.), sp9 (Pennisetum purpureum), sp10
(Pseudelephantopus spiralis), sp11 (Persea sp.), sp12 (Baccharis trinervis), sp13 (Zygia longifolia), sp14 (Adenaria floribunda)
A4. sp1 (Piper angustifolium), sp2 (Solanum sp.), sp3 (Cyathula achyrantoides), sp4 (Justicia sp.), sp5 (Renealmia sp.), sp6
(Adenaria floribunda), sp7 (Tripogandra cumanensis), sp8 (Piper aduncum), sp9 (Pseudochinolaena polystachya), sp10 (Ageratum
conyzoides), sp11 (Pssiflora sp.), sp12 (Baccharis trinervis), sp13 (Especie 13)
Tal como se observa, ninguna de las variables muestra una tendencia clara en su
comportamiento sino más bien, se nota irregularidad en cada una, e inexistencia de
0
10
20
30
40
50
60
70
80
sp1 sp2 sp3 sp4 sp5 sp6 sp7 sp8
Transepto 1. % parametros ecologicos
Densidadrelativa
% area desuelo ocupado
% Biomasa
0
10
20
30
40
50
60
70
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9
sp10
sp11
sp12
Transepto 2. % parametros ecologicos
Densidadrelativa % area desuelo ocupado % biomasa
0
10
20
30
40
50
60
70
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9
sp10
sp11
sp12
sp13
sp14
Transepto 3. % parametros ecologicos
Densidadrelativa
% area desueloocupado % biomasa
0
5
10
15
20
25
30
35
40
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9
sp10
sp11
sp12
sp13
Transepto 4. % parametros ecologicos
Densidadrelativa
% area desuelo ocupado
% biomasa

39
una correlación entre ellas. No obstante, es posible anotar algunas particularidades en
cada uno de los transeptos muestreados, así:
El transepto 1 (figura A1), se tipifica porque Pseudelephantopus spiralis (sp1) y
Pseudochinolaena polystachya (sp5), muestran una densidad relativa que supera
considerablemente (2 veces el valor) superior al área de suelo ocupado y al porcentaje
de biomasa desarrollado; se entiende entonces que estas especies las constituyen
individuos de porte pequeño, sin embargo su abundante cantidad en la zona no
representa una buena cobertura de suelo y biomasa. Contrariamente, Pennisetum
purpureum (sp4) se distribuye de mejor forma ocupando una mayor área de suelo, tal
vez por la necesidad de extender de mejor manera su mayor biomasa, disposición que
la convierte en la especie más importante en esta zona. Las restantes especies (sp2,
sp3 sp6, sp7 y sp8) dejan ver un balance moderado entre el área ocupada y la
biomasa que poseen, aunque entre ellas se destaca sp7 (Renealmia sp.) por los
mayores valores de los parámetros evaluados.
La figura A2, por su parte, ilustra el comportamiento de las especies desarrolladas en el
transepto 2; en este caso Blechnum sp (sp3), Pteris sp (sp4) y Arachis pintoi (sp9),
conforman un grupo en el que la biomasa y el área de suelo ocupado es
considerablemente más baja (por debajo de la mitad del valor de la densidad relativa)
que la densidad relativa, dando a entender que los individuos se dispersan a lo largo y
ancho de la zona, sin ocupar un gran espacio ni tener una biomasa significativa,
comportamiento asociado, como se mencionó en el transepto 1, al tipo de crecimiento
vegetativo que poseen (herbáceas de porte pequeño). Pennisetum purpureum (sp7) y
Zygia longifolia (sp10), evidencian una dinámica contraria a las anteriores: los pocos
individuos encontrados en la zona, tienen mejor desarrollo vegetativo (mayor biomasa y
área de suelo ocupado), lo que las convierte en dos especies de moderada importancia
en el transepto, derivado posiblemente de su mejor adaptabilidad a las condiciones de
la zona. Pseudelephantopus spiralis (sp2), se muestra como la especie de mayor
importancia biológica en el transepto que nos ocupa, dado que más del 50% de la

40
zona está ocupada por ella, deja ver los valores ecológicos más altos y muestra un
buen balance entre ellos.
En el transepto 3 (figura A3) se nota claramente que más del 70% de los individuos
pertenecen a las especies Tripogandra cumanensis (sp1), Pseudochinolaena
polystachya (sp6) y Pennisetum purpureum (sp9) dominando la zona. No obstante,
Pennisetum purpureum se destaca por su mayor biomasa y área de suelo ocupado,
consecuentemente debe considerársele como la de mayor representatividad biológica
en este transepto.
La zona 4 se diferenció claramente de las anteriores, tal como lo deja ver la figura A4.
De manera general se ve que en la zona hay un equilibrio evidente entre las especies,
ya que los parámetros ecológicos en cada una de ellas muestran valores de
significativa importancia y dignos de ser tenidos en cuenta. Se nota también que el
material biológico encontrado en la zona está distribuido entre 7 especies (sp2, sp3,
sp4, sp5, sp7, sp8 y sp10) que muestran un 5% o más de biomasa, se destacan entre
ellas: sp4 y sp7 con una biomasa superior al 20%.
Importa hacer ver que sp4, sp5, sp7, sp8 y sp10 evidencian una mayor área de suelo
ocupado en relación al número de individuos desarrollados. Por su parte, sp13
sobresale por su mayor densidad relativa en la zona.
Commelina diffusa (sp7) se revela como la de mayor importancia ecológica por tener
los valores ecológicos más altos
Debe aclararse que en el transepto 5 se evidenció un dominio total de Pennisetum
purpureu, esta especie se encontraba distribuida más allá del área muestreada y
abarcando una gran extensión de la ribera del rio.
De forma resumida puede anotarse que es evidente el dominio de algunas especies
vegetales en los diferentes transeptos muestreados, esta condición se deriva de una
mejor adaptación biológica a las condiciones particulares del suelo de la ribera del rio,

41
así como también a las dinámicas hidrológicas y características fisicoquímicas y
microbiológicas del agua rio Combeima; expresándose de alguna manera un mayor
número de individuos en estas especies (mayor densidad), área de suelo cubierto,
cobertura foliar y biomasa.
4.2.1 Determinación del Índice de valor de Importancia (IVI) en las especies vegetales:
Debe reconocerse que los parámetros ecológicos determinados dan una información
valiosa acerca del nivel de adaptabilidad de las especies vegetales encontradas bajo
las condiciones de suelo y clima del área de interés del rio Combeima, no obstante el
alto volumen de información obtenido demanda la aplicación de un tratamiento que
conjuntamente agrupe los datos obtenidos, genere un conocimiento más global de la
naturaleza de las especies relacionada con su hábitat, y así mismo se pueda reducir la
información a una expresión sencilla. Con este criterio en mente, en este trabajo se
consideró pertinente calcular el “Índice de valor de importancia” (IVI), el cual mide el
valor ecológico relativo de cada especie en la zona muestreada. Para tal efecto se
aunó la información expresada por tres parámetros: densidad relativa, área de suelo
cubierto y biomasa.
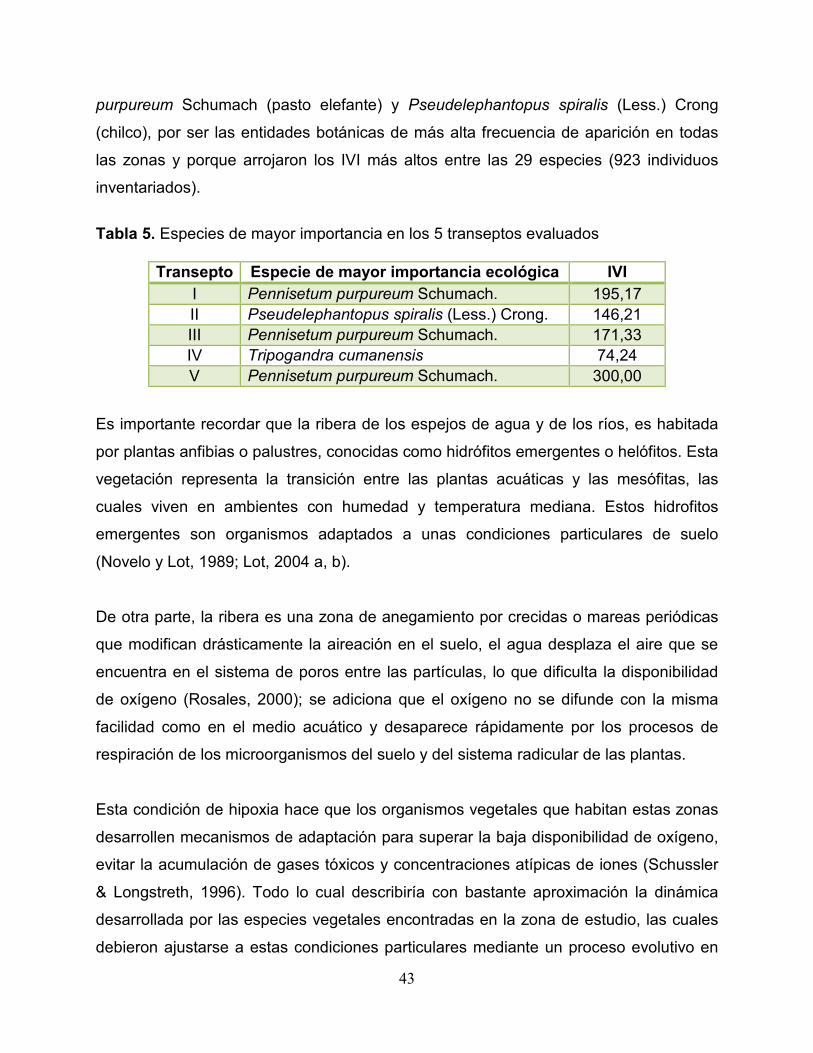
La figura 4 ilustra los valores encontrados para cada una de las especies en los
diferentes transeptos. Lo que se observa en la figura 4 confirma la discusión anterior,
evidenciándose claramente cuáles son las especies vegetales dominantes en cada
área muestreada. Se nota además que en los transeptos 1, 2 y 3 existe poca
diversificación y equilibrio ecológico en los organismos vegetales; caso contrario del
transepto 4 donde se observa un número abundante de individuos, alta diversidad de
especies, y equilibrio en el valor ecológico de cada especie.
Figura 4. Índice de valor de importancia (IVI) de las especies de cada uno de los transeptos.

42
A1) A2) A1. sp1 (Pseudelephantopus spiralis), sp2 (Acalypha diversifolia), sp3 (Sida rhombifolia), sp4 (Pennisetum purpureum), sp5
(Pseudochinolaena polystachya), sp6 (Cyathula achyrantoides), sp7 (Renealmia sp.), sp8 (Sporobolus jacquemontti) A2. sp1 (Baccharis trinervis), sp2 (Pseudelephantopus spiralis), sp3 (Blechnum sp.), sp4 (Pteris sp.), sp5 (Piper aduncum), sp6
(Piper angustifolium), sp7 (Pennisetum purpureum), sp8 (Pseudochinolaena polystachya), sp9 (Arachis pintoi), sp10 (Zygia
longifolia), sp11 (Justicia sp.), sp12 (Tripogandra cumanensis)
A3) A4) A3. sp1 (Tripogandra cumanensis), sp2 (Acalypha diversifolia), sp3 (Piper angustifolium), sp 4 (Especie 4), sp5 (Cupania cinérea),
sp6 (Pseudochinolaena polystachya), sp7 (Commelina diffusa), sp8 (costus sp.), sp9 (Pennisetum purpureum), sp10
(Pseudelephantopus spiralis), sp11 (Persea sp.), sp12 (Baccharis trinervis), sp13 (Zygia longifolia), sp14 (Adenaria floribunda)
A4. sp1 (Piper angustifolium), sp2 (Solanum sp.), sp3 (Cyathula achyrantoides), sp4 (Justicia sp.), sp5 (Renealmia sp.), sp6
(Adenaria floribunda), sp7 (Tripogandra cumanensis), sp8 (Piper aduncum), sp9 (Pseudochinolaena polystachya), sp10 (Ageratum
conyzoides), sp11 (Pssiflora sp.), sp12 (Baccharis trinervis), sp13 (Especie 13)
Sobre la base de lo observado (figura 4) y los mayores IVI encontrados (tabla 5) en las
especies de cada transepto se determinó continuar el estudio con Pennisetum
0
50
100
150
200
sp1 sp2 sp3 sp4 sp5 sp6 sp7 sp8
Transepto 1. IVI
IVI
0
50
100
150
200
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9
sp10
sp11
sp12
Transepto 2. IVI
IVI
020406080
100120140160
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9
sp10
sp11
sp12
sp13
sp14
Transepto 3. IVI
IVI
01020304050607080
sp1
sp2
sp3
sp4
sp5
sp6
sp7
sp8
sp9 sp…
sp…
sp…
sp…
Transepto 4. IVI
IVI
43
purpureum Schumach (pasto elefante) y Pseudelephantopus spiralis (Less.) Crong
(chilco), por ser las entidades botánicas de más alta frecuencia de aparición en todas
las zonas y porque arrojaron los IVI más altos entre las 29 especies (923 individuos
inventariados).
Tabla 5. Especies de mayor importancia en los 5 transeptos evaluados
Transepto Especie de mayor importancia ecológica IVI I Pennisetum purpureum Schumach. 195,17 II Pseudelephantopus spiralis (Less.) Crong. 146,21 III Pennisetum purpureum Schumach. 171,33 IV Tripogandra cumanensis 74,24 V Pennisetum purpureum Schumach. 300,00
Es importante recordar que la ribera de los espejos de agua y de los ríos, es habitada
por plantas anfibias o palustres, conocidas como hidrófitos emergentes o helófitos. Esta
vegetación representa la transición entre las plantas acuáticas y las mesófitas, las
cuales viven en ambientes con humedad y temperatura mediana. Estos hidrofitos
emergentes son organismos adaptados a unas condiciones particulares de suelo
(Novelo y Lot, 1989; Lot, 2004 a, b).
De otra parte, la ribera es una zona de anegamiento por crecidas o mareas periódicas
que modifican drásticamente la aireación en el suelo, el agua desplaza el aire que se
encuentra en el sistema de poros entre las partículas, lo que dificulta la disponibilidad
de oxígeno (Rosales, 2000); se adiciona que el oxígeno no se difunde con la misma
facilidad como en el medio acuático y desaparece rápidamente por los procesos de
respiración de los microorganismos del suelo y del sistema radicular de las plantas.
Esta condición de hipoxia hace que los organismos vegetales que habitan estas zonas
desarrollen mecanismos de adaptación para superar la baja disponibilidad de oxígeno,
evitar la acumulación de gases tóxicos y concentraciones atípicas de iones (Schussler
& Longstreth, 1996). Todo lo cual describiría con bastante aproximación la dinámica
desarrollada por las especies vegetales encontradas en la zona de estudio, las cuales
debieron ajustarse a estas condiciones particulares mediante un proceso evolutivo en

44
su sistema radicular y fisiológico que dio como resultado la generación de los
mecanismos necesarios para desarrollarse bajo estas condiciones, entre otros:
• Desarrollo de raíces y rizomas capaces de desplegarse muy bien en el hábitat.
• Presencia de órganos emergentes capaces de generar un tejido parenquimático
con espacios intercelulares particularmente grandes (aerénquima), que actúa
como un reservorio de gas que oxigena a los órganos sumergidos.
• Disponer también de tallos huecos y de grandes cámaras de aire para facilitar la
oxigenación de los órganos sumergidos: vástagos y raíces (Teal & Kanwisher,
1966).
Es indiscutible que las cuatro especies vegetales que relaciona la tabla 5 se
constituyen en modelo de esta adaptación, logrando destacarse muy bien Pennisetum
purpureum Schumach (IVI 195,1707) y Pseudelephantopus spiralis (Less.) Crong (IVI
171,33), como las más representativas de la zona bajo estudio.
Complementario a los mecanismos anteriormente descritos, algunas hidrófitas agrupan
sus hojas a manera de tallo, aunque hueco y esponjoso, para lograr superar la
condición de baja disponibilidad de oxígeno en el suelo, es el caso de Pennisetum
purpureum, Gynerium sagitattum, y Renealmia sp, las cuales presentan además raíces
bien desarrolladas de tipo adventicias.
La alta distribución en una gran porción del suelo ribereño bajo estudio por parte de
Pseudelephantopus spiralis, Pseudochinolaena polystachya, Sporobolus pyramidalis,
Tripogandra serrulata, Commelina diffusa, podría atribuirse a la ramificación de sus
rizomas, además de evidenciar la macromorfología que describe el párrafo que
antecede. Es importante resaltar que Pseudelephantopus spiralis mostró la mejor
condición de distribución entre estas especies, con raíces de longitud variada, rizomas
extendidos firmemente arraigados al sustrato mediante el desarrollo extensivo de
raíces adventicias que impiden que la planta sea arrastrada por las aguas en los

45
períodos de inundación (Arbo et al, 2000), a lo que se adiciona la generación de brotes
de hojas en formas de roseta en cada rizoma, que facilitan su propagación.
Probablemente otras especies (por ejemplo Pennisetum purpureum), logran su
adaptación a la baja disponibilidad de oxígeno y nutrientes, mediante el desarrollo de
un tipo de raíces especializadas, los neumatóforos, para suplir la escasez de oxígeno
en estos ambientes. Se trata de raíces adventicias que nacen en cada uno de los
estolones, con geotropismo negativo (Arbo, et al, 2000).
Debe recordarse que, en general, los sistemas ribereños están conformados por la
asociación de especies vegetales anfibias o palustres (sólo la raíz mantiene contacto
directo con el agua), acuáticas arraigadas o con hojas flotantes y acompañadas de
plantas terrestres de lugares muy húmedos. Los vegetales que logran adaptarse al
estrés hídrico por excesiva humedad tienen sus raíces envueltas en fango y suelo,
debajo de él puede haber 1 metro de agua o más. Bajo estas condiciones logran
desarrollarse desde herbáceas hasta algunas arbóreas (Lot y Novelo, 1978;
Sculthorpe, 1967).
Cabe resaltar que en este trabajo se encontraron organismos de baja y mediana talla
de desarrollo tales como: Pseudelephantopus spiralis, Pseudochinolaena polystachya,
Sporobolus pyramidalis, Justicia sp., Adenaria floribunda, Tripogandra serrulata,
Commelina diffusa y Pennisetum purpureum, Gynerium sagitattum, Renealmia sp.,
Solanum sp., encontrándose muy pocas de naturaleza arbórea: Cupania cinérea,
Persea sp., Zygia longifolia, Piper aduncum, Piper angustifolium; sin embargo las de
mayor dominancia fueron las de menor porte principalmente, Pseudelephantopus
spiralis y Pennisetum purpureum, dando a entender que el aportante de nutrientes no
es sólo el suelo sino también el rio, que por sus dinámicas mantiene un flujo de
nutrientes hacia los suelos aledaños, depositando diferentes elementos transportados
por el cuerpo de agua que permiten el desarrollo de estas especies.

46
4.3 CARACTERIZACIÓN FISICOQUÍMICA DEL SUELO RIBEREÑO Y EL AGUA DEL
RIO
Para evaluar y conocer la condición del suelo ribereño en que se desarrollan las
especies vegetales seleccionadas, se tomó una porción de suelo en influencia de las
plantas seleccionadas para continuar con ellas el trabajo (Pennisetum purpureum y
Pseudelephantopus spiralis). En el momento en que fueron colectados los
especímenes para los aislamientos. Los resultados de esta caracterización
fisicoquímica se ven en la tabla 6. Tabla 6. Caracterización fisicoquímica del suelo ribereño bajo estudio
Parámetro
Unidad
Resultados
Suelo Chilco Suelo Pasto elefante
pH 6,8 6,8 Nitrógeno (orgánico total) % 0,117 0,111
Materia orgánica (M.O) % 0,7 0,6 C.I.C (capacidad de intercambio catiónico) meq.100g-1 21 20
Fósforo (P) (soluble) mg.Kg -1 93 95 Calcio (Ca) meq.100g-1 6,2 6,4
Magnesio (Mg) meq.100g-1 3,2 3,7 Sodio (Na) meq.100g-1 0,2 0,2 Potasio (K) meq.100g-1 0,18 0,6 Hierro (Fe) mg.Kg -1 95 95 Cobre (Cu) mg.Kg -1 1,1 1 Zinc (Zn) mg.Kg -1 7,9 6,9 Manganeso (Mn) mg.Kg -1 44 44 Boro (B) mg.Kg -1 0,3 0,35 Azufre (S) mg.Kg -1 28 31 Aluminio (Al) meq.100g-1 ND ND Saturación de Aluminio % ND ND Saturación de Bases % 46,6 54,5 Relación Ca/Mg 1,9 1,7 Relación (Ca+Mg)/K 52,2 16,8

47
Relación Mg/K 17,8 6,2
Textura
Arcilla % 6,5 6,5 Limo % 18 18 Arena % 75,5 75,5 Clave Franco arenoso Franco arenoso
Los resultados que muestra la tabla 6 dejan ver algunas deficiencias en el aporte de
nutrientes que puede hacer el suelo a la vegetación riparia que alberga. Es el caso del
nitrógeno, donde se nota una baja disponibilidad (0.1 %). La materia orgánica, que a
su vez sirve como fuente de nitrógeno (0.7- 0.6 %). y otros nutrientes, es igualmente de
limitada aportación nutricional. Esta condición de deficiencia se relaciona con la textura
y mineralogía particular que presenta el suelo (Church, 2002). En este caso el suelo
ribereño, resultó ser un sistema constituido por partículas gruesas (arenoso), que
evidencia todas las características de este tipo de suelo: capacidad de drenaje alta,
que permite la pérdida por percolación de la materia orgánica y del nitrógeno presente;
por su bajo contenido de material fino (arcillas y limos); baja retención de las partículas
minerales e incorporación por formación de agregados estables de la materia orgánica
y el nitrógeno. El suelo arenoso es un suelo de tipo ácido (tabla 6), y aunque es
preferido por muchas plantas, en general carece de los nutrientes que lo hacen ideal
como un medio fértil por sí mismo.
Todo lo anterior permite inferir que la vegetación desarrollada en los suelos ribereños
del rio Combeima-tramo UT, en cierta medida, no sólo depende del aporte de
nutrientes que hace el suelo (por sí mismo carente de ellos) sino también de la entrada
de nutrientes y minerales que hace el rio, el cual debe suplir las falencias presentadas
por el suelo. Esta suplementación puede llegar a través de la deposición de sedimentos
por las inundaciones periódicas en la ribera que suministra la materia orgánica y el
nitrógeno necesarios para el desarrollo de la vegetación.
De esta forma se puede afirmar que el suelo ribereño del rio Combeima-tramo UT se
caracteriza por presentar:

48
Pérdida de nutrientes por lavado
Deficiencias en materia orgánica y nitrógeno
Ser una zona de acumulación de sedimentos y materiales trasportados por el rio
A las dificultades por nutrientes que deben afrontar las especies riparias que habitan la
fuente hídrica de interés en este trabajo, se suman los altos niveles de fósforo
registrados en el suelo ribereño (93 - 95 mg.Kg-1), que tendría su origen en la frecuente
deposición de sedimentos y materiales presentes en el cuerpo de agua que llegan a las
orillas cuando ocurren las inundaciones. Es importante explicar que el rio Combeima
sufre en gran parte de su recorrido por la descarga de aguas servidas de origen
doméstico, cargadas de detergentes y desechos organofosforados que finalmente se
acumulan en la ribera.
Los demás componentes minerales del suelo (K, Mg, Ca, Na, Fe, Cu, Zn, Mn, B, S) se
encuentran en buena proporción, comparado con estudios de mineralogía de suelos
con vocación forestal protectora y pasturas (Echeverri et al., 2014; Gonzáles et al.,
2009), y se explica por la descarga de sedimentos que hace el rio de rocas
meteorizadas de diferente naturaleza que suministra diversos minerales y mejoran la
composición del suelo.
En general, las dos muestras de suelo analizadas, tomadas en diferentes puntos de la
ribera del rio y en el lugar de desarrollo de las especies vegetales de interés, presentan
la misma condición fisicoquímica. Ver tabla 6. Se entiende de esta manera que ambas
especies encuentran un suelo cuyas características fisicoquímicas a lo largo de la
ribera favorece su desarrollo, lo que justifica su dominancia en la zona bajo estudio y,
por ende, los mayores IVI encontrados.
Algunos autores (Mcclain and Richey, 1996; Naiman et al., 2005), sostiene que la
presencia de vegetación riparia en este tipo de suelo es de particular importancia, ya
que ella mejora las condiciones fisicoquímicas y el dinamismo en el flujo de nutrientes
y materiales, además ayuda a compactar el suelo. La materia orgánica que llega al

49
suelo, proveniente de estas plantas sirve de colchón para evitar la pérdida de
nutrientes por el lavado. El sistema radicular de estas plantas, por su parte, juega un
papel importante en el mantenimiento de unas condiciones favorables para los
procesos de biodegradación con microorganismos, retención, remoción y asimilación
de los nutrientes y materiales evitando su incorporación nuevamente al cuerpo de
agua.
Principalmente, la vegetación que se desarrolla en estos suelos con exceso de fósforo,
contribuye al manejo y reducción de este elemento, por su incorporación en forma de
biomasa a través de los procesos de biodegradación y bioasimilacion en asociación
con los microorganismos; lo que contribuiría, al menos en parte, el riesgo de alcanzar
concentraciones exageradamente altas de este elemento en el suelo y ejerciendo un
control de la contaminación por este elemento en el cuerpo del agua (Dorioz et al.,
2006).
En este contexto, se consideró importante realizar un análisis fisicoquímico al cuerpo
de agua, cuyos resultados contribuyen a enriquecer la información suministrada por el
suelo en cuanto a las condiciones de calidad y de cantidad de los parámetros químicos
incorporados en el cuerpo de agua, independientemente de si se trata de un agua
continental (superficial y subterránea), marina o costera, durante un tiempo indefinido o
definido en un área específica.
Sobre esta base se quiso conocer además, las condiciones en las que se encontraba el
agua el rio Combeima en la zona de interés, para la época en la que se realizó el
muestreo y recolección del material vegetal. Tratando de entender un poco las
dinámicas hidrológicas del rio, que afectan la calidad de sus aguas y su entorno
ecológico, en particular las especies vegetales asociadas en sus orillas que de manera
directa reciben y realizan un manejo de los materiales, sedimentos y nutrientes
trasportados por el cuerpo de agua. Se evaluaron entonces algunos parámetros de
importancia sobre la calidad del agua del rio: sólidos totales, nitrógeno amoniacal,
fosfatos, fósforo total y DQO, en dos puntos del rio Combeima: al inicio de una zona en

50
la que la vegetación riparia acompañaba el cauce del rio y otro al final. La toma de la
muestra de agua se realizó en periodo de aguas altas, después de una evidente
creciente del rio. Los resultados de la evaluación de estos parámetros se observan en
la tabla 7.
Tabla 7. Caracterización fisicoquímica del agua del rio Combeima en época de lluvias
en dos puntos de la ribera del rio.
Parámetro Unidad Resultado
Punto inicial Punto final pH 0-14 7,55 7,5 Temperatura °C 20,3 20,1 Solidos Totales mg/L 2114 1914 DQO mg/L 56,3 104,3 Nitrógeno amoniacal
mg/L 3 2,8
Fósforo (Fosfatos) mg PO4-3/L 1,4 0,9 Fósforo total mg P/L 0,73 0,67
Los resultados que muestra la tabla 7 dejan ver un agua con pH entre los rangos de
neutralidad, no obstante el pH de un cuerpo de agua puede variar a lo largo de un
amplio rango de valores, dependiendo de factores intrínsecos (Capacidad
amortiguadora del sistema de alcalinidad carbonato-bicarbonato, evaporación,
intensidad de procesos biológicos tales como fotosíntesis, respiración y actividades de
descomposición de materia orgánica, entre otros) y extrínsecos (composición de suelos
adyacentes, depósitos superficiales y lecho rocoso, presión parcial de CO2 en la
atmósfera, temperatura, etc) del ambiente acuático (Roldan, 1992)
A su vez, la temperatura es un factor determinante en la solubilidad del oxígeno en el
agua. La tabla 8 deja ver una relación entre las dos variables. Podría entonces
esperarse una cantidad relativamente alta de O2 disuelto y con ello una alta actividad
fotosintética.

51
Tabla 8. Efecto de la temperatura sobre la solubilidad del oxígeno
Sin embargo la tabla 7 muestra un cuerpo de agua afectado por los altos niveles de sólidos (haciendo alusión a materia suspendida o disuelta), que pueden afectar
adversamente la calidad de un cuerpo de agua o un efluente volviéndolas de mal sabor
para el paladar e inducir una reacción fisiológica adversa en el consumidor. En general
se acepta como un límite de 500 mg/L de sólidos disueltos para el agua potable
(Fuentes y Massol-Deyá, 2002). En la tabla 7 se puede observar que las muestras
analizadas tienen un nivel 4 veces superior a los valores aceptados (2114-1914 mg/L).
El promedio de sólidos disueltos totales para los ríos de todo el mundo ha sido
estimado en alrededor de 120 ppm (Livingston, 1963). Cabe advertir que la zona del rio
Combeima bajo estudio se encuentra influenciada por las dinámicas de la ciudad, que
claramente intervienen en sus componentes naturales: reducción de la franja de
vegetación riparia, desestabilización de sus orillas, modificación del lecho del rio y
pérdida de la calidad de sus aguas, con asentamientos de población en parte de su
recorrido, que suministran una carga importante de contaminación y cierto grado de
perturbación al ecosistema acuático, que redunda en una baja productividad y
transformación de la materia orgánica y materiales en exceso en este punto del rio,
asociado principalmente a la descarga de aguas servidas directamente al rio sin ningún
tratamiento por la población aledaña. Se adiciona que el rio es considerado como un
depósito de basuras, desperdicios y materiales de todo tipo que generan problemas de
contaminación a lo largo de su recorrido.
Temperatura (°C)
Oxígeno disuelto (mg/L)
10 11.3
20 9.2
30 7.6

52
Un parámetro que arroja información importante a nivel ecológico es la “Demanda
Química de Oxígeno” (DQO). Puede considerársele como una medida de la necesidad
de oxígeno requerido para la oxidación total de los constituyentes orgánicos a
productos inorgánicos, excluyendo el oxígeno que convierte el nitrógeno reducido a
nitrato (EPA, 2000). Para los sitios de muestreo llamados “inicial” y “final” se obtuvieron
valores de 56,3 mg O2 /L y 104,3 mg O2 /L, respectivamente (tabla 7); notándose
claramente que en su recorrido el agua del rio sufre un deterioro importante, logrando
casi doblar los requerimientos de O2 para degradar la materia orgánica que se acumula
en una longitud de apenas 50 m; pudiendo entonces pensarse en una alta variabilidad
de contaminantes de tipo orgánico e inorgánico en donde se incluirían desde nutrientes
(N, P, etc.) hasta sustancias tóxicas, pero que en cualquier caso otorgan al agua poca
capacidad de depuración, debido a la pérdida de la biodiversidad acuática y al bajo
contenido de oxígeno disuelto.
De otra parte, aunque el nitrógeno es un nutriente esencial para los organismos vivos,
su presencia en gran exceso puede transformarlo en una sustancia tóxica (Rozic et al.,
2000). Niveles por encima de 0.2 mg L-1 de amoniaco libre disuelto en agua son
extremadamente perjudiciales para la vida acuática, provocando una aguda
disminución del O2 disuelto en el agua (Dapena et al., 2004,2006). La eliminación de
nitrógeno de las aguas residuales no es algo deseable sino que es exigido por
entidades como la Directiva Europea 91/271/EEC, en el tratamiento de aguas
residuales urbanas.
Los valores de nitrógeno amoniacal hallados en este trabajo (3 - 2.8 mg/L), son un
indicativo más del grado de contaminación en el que se encuentra el agua del rio por el
exceso de materia orgánica y, especialmente, por un exceso de compuestos
nitrogenados. El origen del nitrógeno amoniacal en aguas superficiales es diverso: la
industria del petróleo, la farmacéutica, los fertilizantes o la industria alimenticia
(Gangbazo et al., 1995; Hooda et al., 2000), son sólo unos pocos ejemplos de cómo la

53
actividad antrópica afecta la calidad del agua; el N es, tal vez, el mayor contribuyente a
la degradación de un cuerpo de agua, especialmente por materia orgánica.
Se debe reconocer entonces que el vertido incontrolado de los efluentes industriales y
domésticos produce un alto impacto en el medio acuático que los recibe. En el caso
que nos ocupa, la presencia del xenófobo estaría dado principalmente por la descarga
de aguas cloacales sin ningún tratamiento y control, originado por la población
asentada a lo largo de la ribera del rio Combeima (Servais y Garnier, 1993; Garnier et
al, 1992 a, b); además del aporte que hacen los terrenos agrícolas cargados de
fertilizantes (compuestos de nitrógeno y fósforo) que por escorrentía y lavado
suministran una importante cantidad de nutrientes a las aguas superficiales y
subterráneas del rio, resultando en un agua degradada en su calidad en las partes
bajas del rio.
Esta contaminación evidente por nitrógeno es causante en el medio acuático de
procesos de eutrofización (ocasionado por la presencia de fosfatos y nitratos,
especialmente) y reducción del oxígeno disuelto, además de ser desencadenante de
problemas de salud a la población humana y animal. Algunas formas de nitrógeno,
como los nitratos, en un alto contenido en el agua, ocasionan problemas de
metahemoglobinemia en niños (Fernández et al, 2011). En la eutrofización se
desencadena, el florecimiento de algas que al morir, dejan un exceso de material
orgánico en el fondo del rio, que es descompuesto por bacterias y microorganismos
que consumen gran cantidad de oxígeno y generan un agotamiento de éste en el
medio acuático, lo que a su vez se traduce en una pérdida de la vida que depende este
elemento (Mitchell et al. 1991).
Debe admitirse que el fósforo después del nitrógeno, es el nutriente inorgánico más
requerido por plantas y microorganismos, por lo que se considera como un elemento
esencial para la vida; no obstante cuando aparece en cantidades excesivas en un
cuerpo de agua pasa a constituirse, al igual que el nitrógeno, en indicador de
contaminación. En las aguas superficiales, un exceso de fósforo, en combinación con
temperaturas altas y luz solar, estimula el crecimiento de algas (Redfield, 1958).

54
Los resultados que muestra la tabla 7 para los fosfatos en la zona del rio bajo estudio
(1.4 – 0.9 mg PO4-3 /L, para el punto inicial y final respectivamente; y 0.73- 0.67mg
Ptotal/L, para el punto inicial y final respectivamente); mientras que en aguas no
contaminadas superficiales se manejan datos entre 10 a 50 µg P/L y entre 0.005 –
0.02 mg PO4-3 /L; para aguas de excelente calidad los valores aceptados son menores
a 0.001 mg/l. UNESCO (1996). Los valores obtenidos para el rio Combeima
claramente superan a los típicamente aceptados para aguas de buena calidad, dando a
entender claramente la contaminación por fosfatos, fundamentalmente provenientes de
los detergentes, los cuales abundan en cantidad en las aguas residuales domésticas
que llegan al rio durante todo su recorrido por la ciudad.
Las plantas como el Pasto Elefante y el Chilco dispondrían, según los resultados
obtenidos, de cantidad suficiente de fósforo a través de las aguas para efectuar sus
procesos metabólicos; no obstante, a la luz de las actividades que se producen en un
cuerpo de agua, el contenido de fósforo total está representado en: fósforo en
suspensión en materia particulada (la mayor cantidad) y fósforo en forma disuelta que
es bastante bajo. El incremento de la cantidad de fosfatos en el agua del rio, se origina
a partir de fertilizantes y detergentes que llegan al rio por escurrimientos agrícolas,
desechos industriales y aguas negras. Bajo la óptica anteriormente descrita es posible
afirmar que la vegetación riparia está sometida a estrés excesivo por cuanto el medio,
suelo y agua, contiene fósforo en abundancia aunque no necesariamente disponible
para las plantas puesto que una buena parte de estos nutrientes estaría asociada a la
materia orgánica convirtiéndolo en fósforo inmovilizado.
No debe perderse de vista que las plantas necesitan fósforo para crecer y desarrollar
su potencial genético y formar la base de gran número de compuestos, de los cuales
los más importantes son los fosfatos. Las especies deben absorberlo del suelo, donde
se encuentra en muy baja concentración, normalmente en niveles que varían entre 5 y
30 mg.kg-1, debido a que el fósforo soluble reacciona con Ca, Fe o Al (fosfatos de Al-P,
de Fe-P y de Ca-P), provocando su precipitación o fijación y disminuyendo su

55
disponibilidad para los vegetales. La materia orgánica que constituye hasta el 50 % del
suelo es un importante reservorio de P inmovilizado.
Nuestros resultados concuerdan con algunos estudios previos realizados por
CORTOLIMA, en los que también se evidencia la contaminación y pérdida de la calidad
de las aguas del rio Combeima hacia las partes bajas de su cuenca, lo que comprende
la entrada del rio por la ciudad y el recorrido por ella (área que coincide con la zona de
estudio de este trabajo). La entidad controladora medioambiental en el Tolima halló que
en las estaciones de monitoreo Rio Combeima- Chapetón (encuentro del rio con la
ciudad) y Rio Combeima- Totumo (salida del rio de la ciudad), los índices de calidad
alcanzan niveles que permiten calificar el agua como de “mala calidad”. Por su parte,
los índices de contaminación ICOMO e ICOSUS (afectación por niveles altos de
materia orgánica y material suspendido, respectivamente) fueron particularmente
elevados en el punto Rio Combeima- Totumo. El estudio atribuye la disminución de la
calidad del recurso hídrico en estos tramos del rio a las descargas industriales
(Carlima, Fruver, Fatextol, Toliplásticos, entre otras) y a descargas de aguas residuales
domésticas de la parte sur, centro y occidente de la ciudad de Ibagué (CORTOLIMA,
2003).
Es preciso hacer ver que la biodegradabilidad de las aguas residuales no es una
propiedad intrínseca de éstas, sino que depende en gran medida de la población
microbiológica a la que esta se enfrente. Una estrategia que puede ser aplicada para el
manejo de la contaminación tan evidente en este tramo del rio, que favorece procesos
de mejoramiento de la calidad del agua y la recuperación del ecosistema acuático, es el
uso de bandas protectoras de vegetación riparia (buffer- strips). Un método que logra la
restauración ecológica del rio y contribuye en el manejo de la contaminación difusa,
proveniente de zonas agrícolas y por la población aledaña, que realiza descargas de
aguas servidas a lo largo de la ribera del rio Combeima.
Esta estrategia resalta el concepto de Bio-asimilacion y Bio-transformacion del exceso
de nutrientes y contaminantes que hace la vegetación natural desarrollada en estas

56
zonas contaminadas, a través de la incorporación de procesos naturales físicos y
biológicos, logrando reducir, transformar y acumular contaminantes en el suelo
ribereño e impedir su retorno al medio acuático. (Osborne y Kovacic, 1993).
Estas bandas protectoras de vegetación que se establecen a lo largo de la ribera del
rio, ayudan a resistir el impacto de la erosión y los focos de contaminación (EPA, 1984).
Cuando permanece en estado natural, protegen o tampona el cuerpo de agua frente a
los impactos procedentes de un exceso de nutrientes de zonas altas del rio, que
pueden depositarse en la ribera y de la contaminación proveniente del uso de suelo en
terrenos adyacentes. De esta manera esta vegetación se constituye en un ecotono
entre el medio acuático y el medio terrestre colindante.
Entre las funciones de la vegetación en estas zonas se destaca que:
- Controlan la entrada de la radiación solar, disminuyen la llegada de luz y las
fluctuaciones de temperatura de las aguas, evitando así la pérdida del oxígeno
disuelto por aumento de la temperatura. El efecto regulador depende de la
dimensión de la banda arbolada, 10 a 30 metros de ancho es recomendable,
(Gray y Leiser, 1989).
- Reducen la entrada de sedimentos y nutrientes al rio actuando como filtro de
las escorrentías superficiales o del propio cuerpo de agua. La presencia de
vegetación, unida con el aporte de residuos orgánicos que hacen al suelo,
determina mayor rigurosidad y resistencia al paso de las aguas, disminuyendo
su velocidad, favoreciendo la infiltración y la retención de sedimentos en la
llanura de inundación (Gray y Leiser, 1989).
- La vegetación riparia es capaz de retener y aprovechar el fósforo que llega
absorbido a los sedimentos, además de absorber gran parte de los nitratos
disueltos en las escorrentías con lo cual se reduce la entrada de estos nutrientes
en elevados porcentajes al agua del rio, según sea la dimensión de la banda

57
riparia, su pendiente y la carga de nutrientes proveniente de la ladera y del rio.
(US Department of the Army, 1991).
- Las bandas riparias generan también efectos positivos en la recuperación de la
composición del suelo y resistencia a la erosión (Peterson et al. 1992).
- Ante la baja disponibilidad de nutrientes las plantas que habitan en ecosistemas
riparios aumentan el crecimiento de las raíces o los pelos radicales o su
densidad, con el objetivo de lograr la exploración de una mayor superficie y
volumen de suelo. Este comportamiento sustenta la tendencia de este tipo de
vegetación a extender sus raíces, ocupar la mayor cobertura posible del suelo e
intentar proveerse de nutrientes a través de las aguas, pero aunque en un agua
contaminada muchos de ellos (N y P, por ejemplo) se encuentran en cantidades
relativamente altas no está disponible para suplir las necesidades de la planta.
(Peterson et al. 1992).
Importa recordar el papel que desarrollan los microorganismos benéficos del suelo, que
pueden promover el crecimiento de las plantas y también evitar la infección del tejido
vegetal por patógenos, estos son denominados PGPR (plant growth promoting
rhizobacteria; rizobacterias promotoras del crecimiento vegetal). Estos
microorganismos pueden encontrarse en asociaciones simbióticas o de vida libre.
Estos últimos están asociados a las partículas del suelo generando interacciones con
las raíces de las plantas, en la zona de la rizósfera (Peña y Reyes, 2007). Las PGPR
pueden promover el crecimiento vegetal mejorando la disponibilidad de nutrientes en el
suelo mediante la solubilización de fósforo, fijación de nitrógenos y producción de
sideróforos (Bobadilla y Rincón, 2008). De igual manera, se ha demostrado que las
rizobacterias ayudan a disminuir la resistencia a la conductividad hidráulica, lo cual le
da a las plantas una mayor tolerancia a periodos de sequía (Rivieros, 2008).
Los objetivos del trabajo nos condujeron hasta las rizobacterias y particularmente las
endófitas que habitan dentro de los tejidos de las plantas seleccionadas
(Pseudelephantopus spiralis y Pennisetum purpureum), al menos durante una parte de

58
su ciclo de vida logran establecer una asociación simbiótica, sin causarles daño alguno,
y producen grandes beneficios a las plantas. Las bacterias endófitas cumplen una gran
diversidad de funciones, tales como promotoras de crecimiento vegetal, control
biológico sobre una diversidad de fitopatógenos, mejoramiento de la eficiencia de los
procesos de fitorremediación de compuestos tóxicos en la rizósfera (Porteus-Moore et
al., 2006; Van der lelie, 2005; Lodewyckx et al., 2002).
Con este criterio en mente, en este trabajo se consideró de vital importancia entender
la dinámica de las interacciones benéficas que ocurren en la raíz de las plantas
seleccionadas, con microorganismos específicamente funcionales como bacterias
nitrificantes, solubilizadores de fosfatos, como una forma de entender, al menos en
parte, las interacciones “suelo-planta-microorganismos-ambiente”. De esta manera, se
complementó la información hasta ahora obtenida mediante el aislamiento de los
microorganismos asociados a la rizósfera de Pseudelephantopus spiralis y Pennisetum
purpureum, especies que se muestran como las mejor adaptadas a la zona bajo
estudio.
4.4 AISLAMIENTO Y CARACTERIZACIÓN DE LAS BACTERIAS ASOCIADAS A LA
RAÍZ DEL CHILCO Y PASTO ELEFANTE.
4.4.1 Aislamiento de rizobacterias en las especies seleccionadas: Las tablas 9 y 10 muestran las rizobacterias aisladas y purificadas, desarrolladas en
Agar nutriente, después de seleccionar las colonias bacterianas con características
morfológicas diferenciadas.
Un total de 35 cepas bacterianas desarrolladas bajo influencia de la raíz o sobre su
superficie (rizobacterias) fueron aisladas a partir de las especies vegetales bajo
estudio. Se caracterizaron de acuerdo a su morfología colonial y purificaron en agar
nutriente; de éstas se aislaron 17 cepas de la raíz del Chilco (Pseudelephantopus
spiralis), con la pérdida de una cepa por contaminación (crecimiento de otras bacterias

59
en el medio de cultivo), y 18 aislados corresponden al pasto elefante (Pennisetum
purpureum), con contaminación de 3 aislados.
La rizósfera es la zona biológicamente activa del suelo alrededor de la raíz en donde se
llevan a cabo interacciones con microorganismos, entre los que se incluyen bacterias y
hongos (Singh et al., 2004). Las bacterias que interaccionan con la raíz de la planta, se
denominan rizobacterias y particularmente están sujetas a un ambiente de
suplementación de agua, oxígeno y nutrientes por la actividad de las plantas (Cardon y
Whitbeck, 2007).
La presencia y cantidad de rizobacterias halladas en Pennisetum purpureum (18 cepas)
y Pseudelephantopus spiralis (17 cepas) es un indicativo más, del papel que
desempeña la raíz de la vegetación riparia (como P. purpureum y P. spiralis) en los
procesos de adaptación a las condiciones de la zona de estudio. En un trabajo
armonioso y de mutua ayuda con la población microbiana asociada a ellas, permiten
que, a diferencia de otros vegetales, se adapten a las condiciones poco favorables para
el desarrollo y supervivencia de este ecosistema por demás agresivo y de difícil
adaptación para un ser vivo. La asociación planta-microorganismos explica además el
manejo de la contaminación por parte de esta vegetación: la raíz exuda una gran
cantidad y gama de compuestos orgánicos que estimula la actividad microbiana, ésta a
su vez colabora en la solubilización, degradación y transformación de diferentes
compuestos como potenciales contaminantes y los hacen disponibles a la planta para
así proveerla de nutrientes que sólo de esta forma podrían obtener o generar
resistencia a estos.
No obstante, cabe admitir que estas interacciones bióticas se dan en base a la
atracción de organismos tanto benéficos como patógenos, haciendo que el recuento de
bacterias sea mayor que en otras regiones del suelo donde no hay raíces (Cardon y
Whitbeck, 2007), y que la efectividad de las bacterias solubilizadoras de fósforo
depende de la capacidad de los aislados para colonizar la rizósfera y mantener su

60
actividad biológica; su adecuado desempeño está influenciado por factores tales como
el pH, la salinidad y la temperatura (Madigan et al., 2006).
Sin embargo, la diversidad de especies de bacterias que colonizan este hábitat es
significativamente menor que la encontrada en otras regiones del suelo, lo que indica
que existe una fuerte presión de selección en la rizósfera. Pese a ésto, gran cantidad
de rizobacterias se encuentran interaccionando activamente junto con los demás
componentes del ecosistema de la rizósfera. Esta población de bacterias tiene
diferentes niveles tróficos y hábitos de vida. Los saprofitos benéficos, de una diversidad
de grupos microbianos, son capaces de promover el crecimiento vegetal y la salud de
las plantas, incluyen: descomponedores de detritus orgánicos, rizobacterias promotoras
del crecimiento vegetal (PGPR) y los antagonistas de hongos y bacterias patógenos de
la raíz (Marilley y Arango, 1999). Particularmente, las PGPR participan en muchos
procesos importantes de la planta, resultando Pseudomonas y Bacillus como los
géneros más comúnmente descritos para el control bilógico de patógenos, ciclos de
nutrientes y desarrollo de semillas (Perssello-Cartieaux et al., 2003; Barea et al., 2004;
Zahir et al., 2003; Barea et al., 2005).
Esta población de rizobacterias sustentaría el mayor IVI encontrado para Pennisetum
purpureum y Pseudelephantopus spiralis, ya que estas contribuyen en los procesos de
adaptación de las especies a las condiciones adversas en la ribera del rio Combeima:
escasez de nutrientes en el suelo y la columna de agua, bajo contenido de oxígeno en
el suelo, además de la contaminación evidente del agua del rio por factores bióticos y
abióticos.

61
Tabla 9. Rizobacterias aisladas de la raíz del Chilco y caracterizadas por su morfología colonial
ID Forma Tamaño Superficie Elevación Borde Estructura interna Consistencia Cromogénesis
(AN) Tinción de
Gram
Rch1 Irregular 5 mm Rugosa Plana Lobulado Traslucida Cremosa Blancuzca Bacilos (-) Rch2 Redonda 5 mm Lisa Plana Entero Opaca Mucosa Blancuzca Bacilos (-)
Rch3 Extendida por todo el medio - - - - - Membranosa-
mucosa Blancuzca Bacilos (-)
Rch4 Puntiforme Rugosa Plana Entero Traslucida Mucosa Naranja Cocos (-) Rch5 Redonda 8 mm Lisa Plana Entero Opaca Mucosa Amarillento Bacilos (+)
Rch6 Irregular Rugosa Convexa Entero Traslucida Grasosa Blancuzca Cocos (-)
Rch7 Redonda 6 mm Rugosa Convexa Entero Traslucida Mucosa Amarillento Bacilos (-) Rch8 Redonda 7 mm Lisa Plana Entero Traslucida Grasosa Amarillento Bacilos (+)
Rch9 Redonda 5 mm Pliegues Plana Entero Traslucida Cremosa- grasosa Blancuzca Bacilos (-)
Rch10 Redonda 5 mm Plana Convexa Entero Traslucida Grasosa Blancuzca Cocos (+)
Rch11 Redonda 4 mm Plana Convexa Entero Traslucida Cremosa Amarillento Cocos (+) Bacilos (-)
Rch12 Contaminado - - - - - - - -
Rch13 Puntiforme - Rugosa Convexa Entero Opaca Mucosa Rosado Cocos (+)
Rch14 Redonda 2mm Lisa Plana Entero Traslucida Grasosa Blanco Bacilos (-) Rch15 Irregular 4 mm Lisa Plana Irregular Traslucida Grasosa Blanco Cocos (+)
Rch16 Puntos irregulares - Rugosa Plana Lobulado Opaca Cremosa Blancuzca Cocos (-)
Rch17 Puntos irregulares - Rugosa Convexa Lobulado Traslucida Cremosa Amarillento Cocos (+)

62
Tabla 10. Rizobacterias aisladas de la raíz del Pasto elefante y caracterizadas por su morfología colonial
ID Forma Tamaño Superficie Elevación Borde Estructura interna Consistencia Cromogenesis
(AN) Tinción de
Gram
Relef1 Redonda 3 mm Lisa Plana Entero Traslucida Mucosa Amarillo Cocos (+)
Relef2 Contaminado - - - - - - - - Relef3 Contaminado - - - - - - - -
Relef4 Puntos irregulares - Rugosa Convexa Lobulado Traslucida Mucosa Blancuzca Cocos (+)
Relef5 Irregular 10 mm Lisa Plana Irregular Traslucida Mucosa Amarillenta Cocos (+)
Relef6 Puntos irregulares Rugosa Convexa Irregular Traslucida Mucosa Blancuzca Bacilos (+)
Relef7 Fusiforme - Rugosa Elevada Filamentoso Traslucida Mucosa Blancuzca Cocos (+) Bacilos (-)
Relef8 Redonda 5 mm Rugosa Convexa Entero Traslucida Mucosa Amarillo Cocos (+)
Relef9 Redonda 20 mm Lisa Plana Irregular Traslucida Cremosa-grasosa Blancuzca Cocos (+)
Relef10 Redonda 50 mm Lisa Plana Lobulado Traslucida Cremosa Blancuzca cocos (+)
Relef11 Contaminado - - - - - - - -
Relef12 Irregular - Rugosa Elevada Lobulado Traslucida Cremosa-grasosa Blancuzca Cocos (+)
Bacilos (-)
Relef13 Irregular - Rugosa Convexa Lobulado Traslucida Cremosa Blancuzca Cocos (+)
Relef14 Irregular - Rugosa Convexa Irregular Traslucida Mucosa- viscosa Blancuzca Bacilos (-)
Relef15 Irregular - Lisa Plana Entero Traslucida Grasosa Amarillo Cocos (+)
Relef16 Puntiforme - Rugosa Convexa Entero Opaca Cremosa amarillo claro Cocos (+)
Relef17 Extendida por una parte del
medio - Rugosa Convexa Entero Opaca Mucosa Rojizo Bacilos (-)
Relef18 Redonda - Rugosa Elevada Irregular Traslucida Membranosa Amarillo Cocos (+)

63
4.4.2 Asilamiento de bacterias endófitas: En las tablas 11 y 12 se muestran las bacterias endófitas de la raíz de las
plantas seleccionadas, aisladas y purificadas después de seleccionar las colonias bacterianas con características
morfológicas diferenciadas que se desarrollaron en Agar nutriente.
Tabla 11. Bacterias endófitas aisladas y caracterizadas por su morfología colonial de la raíz del Chilco ID Forma Tamaño Superficie Elevación Borde Estructura interna Consistencia Cromogenesis en AN Tinción de Gram
Endoch1 Redonda 8 mm Rugosa Plana Ondulado Opaca Cremosa Amarillo claro Cocos(+) Endoch2 Redonda 3 mm Lisa Plana Entero Traslucida Cremosa Blancuzca Cocos(+) Endoch3 Irregular 1,5 cm Hongo - Entero - - Blancuzca - Endoch4 Irregular 1,5 cm Rugosa Elevada Entero Traslucida Cremosa Blancuzca Bacilos(-) Endoch5 irregular 3 cm Rugosa Plana Lobulado Traslucida Grasosa Blancuzca Cocos (-) Endoch6 Redonda 5 mm Superficie Entero Amarillo claro Cocos (+) Endoch7 Redonda 3mm Lisa Plana Entero Traslucida Viscosa Amarillo naranja Cocos (+) Endoch8 irregular 3 mm Lisa Plana Entero Trasparente Grasosa Trasparente Bacilos (-)
Tabla 12. Bacterias endófitas aisladas y caracterizadas por su morfología colonial de la raíz del Pasto elefante
ID Forma Tamaño Superficie Elevación Borde Estructura Interna Consistencia Cromogenesis (AN) Tinción de Gram
Endoelef1 Redonda 1 cm Lisa Plana Entero Trasparente Grasosa Amarillo-traslucido Cocos (+) Endoelef2 Redonda 3 mm Lisa Plana Entero Traslucida Grasosa Blancuzca Bacilos (-) Endoelef3 Redonda 1 cm Rugosa Plana Filamentoso Traslucida Grasosa Blancuzca Cocos (-) Endoelef4 Redonda 1 cm Rugosa Plana Radiada Trasparente Grasosa Trasparente Bacilos (+) Endoelef5 Estriada 3 cm Lisa Plana Filamentoso Trasparente Grasosa Amarillo traslucido Bacilos (+) Cocos (+) Endoelef6 Irregular 2 cm Rugosa Plana Filamentoso Opaca Cremosa Blancuzca Cocos (-) Endoelef7 Rizoide 5 mm Rugosa Plana Filamentoso Opaca Cremosa Blanco amarilloso Cocos (+) Endoelef8 Contaminado - - - - - - - - Endoelef9 Irregular 5 mm Rugosa Plana Entero Opaca Membranosa Blancuzca Cocos (+)
Endoelef10 Redonda 1 cm Rugosa Plana Lobulado Traslucida Grasosa Trasparente Cocos (-) Endoelef11 Contaminado -- - - - - - - - Endoelef12 Redonda 7 mm Lisa Plana Entero Opaca Cremosa Blancuzca Cocos (+) Endoelef13 Redonda 5 mm Lisa Plana Entero Trasparente Grasosa Amarillo traslucido Cocos (+) Endoelef14 Redonda 5 mm Lisa Plana Radiada Traslucida Blancuzca Cocos (+)

64
Un total de 21 cepas bacterianas endófitas fueron aisladas de la raíz de las 2 plantas
seleccionadas, las cuales fueron caracterizadas y purificadas. 7 cepas se aislaron de la
raíz del Chilco y 14 de la raíz del Pasto elefante, con un aislado contaminado.
Aunque se desconoce la participación directa de las bacterias endofíticas, se ha
demostrado que promueven el crecimiento de la planta, solubilizan nutrientes
indispensables como fósforo o contribuyen a asimilar el nitrógeno en las plantas. Las
bacterias calificadas como “endófitas” residen en tejidos de la planta, principalmente en
espacios intercelulares. La mayor distribución se encuentra en la raíz y está
influenciada por la disponibilidad de nutrientes y por la competencia con las bacterias
de la rizósfera, raramente se les encuentra en espacios intracelulares y dentro de
tejidos vasculares (Bacon y White, 2000). Su presencia en la raíz de las especies que
cohabitan con ellas otorga beneficios a la planta, por el grado de mutualismo que tiene
este sistema planta-microorganismo.
La presencia de bacterias endófitas en la raíz del Chilco y del Pasto elefante abre
posibilidades sobre su potencial aplicación en procesos de fitorremediacion, sobre la
base de que muchas de estas bacterias tienen potencial para transformar una
diversidad de compuestos químicos y mejorar la capacidad de adaptación de la planta en zonas con excesos de nutrientes o algún tipo de contaminante (Araújo et al., 2002
Rosenblueth et al., 2006; Li et al., 2007).
Existe evidencia de la presencia de bacterias endófitas en pastos y de sus beneficios.
Hurek y Reinhold-Hurek (2003), por ejemplo, aislaron e identificaron una bacteria
endófita, Azoarcus BH72, de tejidos de raíces de la especie de pasto Leptochloa fusca
L. Kunth, la cual mejora la capacidad de fijar nitrógeno biológicamente en la planta.
Scott (2001) aisló y encontró un hongo endófito, Epichloë, asociado con varias
especies de pastos y demostró que este microorganismo es un posible agente de
control biológico en estas especies de plantas.

65
Estudios moleculares recientes (Germain et al. 2006; Franks et al. (2006); Ryan et al.
2008) sobre diversidad de bacterias endófitas han revelado una alta riqueza de
filotipos, que promueven el crecimiento de las plantas, suprimen fitopatógenos, ayudan
a remover contaminantes, solubilizan fosfatos y contribuyen a la asimilación biológica
de nitrógeno. Así las cosas, el estudio prosiguió en realizar pruebas, cualitativas y
cuantitativas, que se constituyeran en un punto de apoyo sólido y en la evidencia de la
capacidad fitorremediadora del Chilco y del Pasto Elefante.
4.4.3 Selección de las bacterias solubilizadoras de fósforo: Los organismos con
capacidad de solubilizar el fosfato insoluble, se seleccionaron según su habilidad para
crecer acidificando el medio y formando un halo que indica el proceso de hidrólisis
realizado después de haberlas sometido a incubación en medio SRS con fosfato
tricálcico (figura 5), según la metodología propuesta por Beltrán et al, 2005. Los
resultados aparecen consignados en la tabla 13.
Figura 5. Cepa Relef16 con acidificación del medio y halo de hidrólisis en medio SRS
Fuente: autores
En total 10 rizobacterias aisladas de las plantas seleccionadas, mostraron capacidad
para solubilizar el fósforo aportado por el fosfato tricálcico insoluble, 5 de estos
aislados provenían de la raíz del Chilco y 5 aislados de la raíz del Pasto Elefante,
solamente una bacteria endófita, aislada de la raíz de este último, mostró esta
funcionalidad.

66
Tabla 13. Aislados solubilizadores de fósforo inorgánico caracterizados según su morfología colonial
ID Forma Tamaño Superficie Elevación Borde Estructura
interna Consistencia Cromogénesis (AN) Tinción Gram
Rch1 Irregular 5 mm Rugosa Plana Lobulado Traslucida Cremosa Blancuzco Bacilos (-)
Rch2 Redonda 5 mm Lisa Plana Entero Opaca Mucosa Blancuzco Bacilos (-)
Rch3 Irregular - - - - - Membranosa-
mucosa Blancuzco Bacilos (-)
Rch10 Redonda 5 mm Plana Convexa Entero Traslucida Grasosa Blancuzco Cocos (+)
Rch17 Puntos
irregulares - Rugosa Convexa Lobulado Traslucida Cremosa Amarillento Cocos (+)
Relef9 Redonda 20 mm Lisa Plana Irregular Traslucida Cremosa-grasosa Blancuzco Cocos (+)
Relef10 Redonda 50 mm Lisa Plana Lobulado Traslucida Cremosa Blancuzco Cocos (+)
Relef13 Irregular - Rugosa Convexa Lobulado Traslucida Cremosa Blancuzco Cocos (+)
Relef14 Irregular - Rugosa Convexa Irregular Traslucida Mucosa- viscosa Blancuzco Bacilos (-)
Relef16 Puntiforme - Rugosa Convexa Entero Opaca Cremosa Amarillo claro Cocos (+)
Endoelef10 Redonda 1 cm Rugosa Plana Lobulado Traslucida Grasosa Trasparente Cocos (-)

67
Se han sugerido diferentes mecanismos por los cuales las bacterias transforman los
fosfatos insolubles a formas solubles, tanto directos como indirectos. Entre ellos se
destacan: i) la acción de ácidos orgánicos producidos por estos microorganismos como
el oxálico, cítrico, butírico, malónico, láctico, succínico, málico, glucónico, acético,
glicónico, fumárico, adípico, indolacético y 2-cetoglucónico (Watanabe F. 1965;
Paredes y Espinosa. 2010) ii) quelación de los elementos responsables de la
insolubilidad de los fosfatos presentes. iii) asimilación directa de fosfatos insolubles por
microorganismos que lo acumulan en sus células y los liberan posteriormente (Puente
et al, 2009).
Otros posibles mecanismos aceptados por la comunidad científica y que, igualmente,
pueden sustentar la capacidad solubilizadora de fósforo son la formación de sideróforos
(Aguado et al. 2012) y la producción de enzimas fosfatasas (Nannipieri et al, 2011).
Dado que el ion hierro Fe+3 tiene muy poca solubilidad a pH neutro y por ende no
puede ser utilizado por los organismos, los microorganismos (incluidos bacterias,
principalmente Gram negativas, hongos y plantas) forman pequeñas moléculas
llamadas “sideróforos” (del griego “transportador de hierro”), muchas de ellas de
naturaleza peptídica (de 0.5 a 1.0 KDa) no ribosomales, de alta afinidad por el hierro
que actúan de manera específica como agentes quelantes para secuestrar hierro en
presencia de otros metales (particularmente el fósforo y liberar así el fósforo ligado a
los minerales del suelo) y reducirlo a Fe+2, una forma mucho más soluble que pueden
ser asimilados por mecanismos de transporte activo y aprovechables para la nutrición
de los vegetales (Salibury y Ross 1992).
En cualquier caso es relevante mencionar que la solubilización de fósforo es un
proceso intervenido por enzimas, entre ellas: fosfatasas, que participan en la
desfosforilación de los grupos fosfoéster unidos a la materia orgánica; fitasas, que
catalizan el proceso de hidrólisis del ácido fítico liberando de forma secuencial hasta
seis grupos ortofosfatos libres y para producir penta, tetra, di y monofosfatos y
finalmente inositol libre; las fosfonatasas y C-P liasas, enzimas que participan en la

68
ruptura del enlace carbono- fósforo de los organofosfonatos. (Vincent, et al, 1992; Oh
et al, 2004).
4.4.4 Prueba cualitativa de solubilización en las cepas seleccionadas: se utilizaron 11
cepas bacterianas para desarrollar la prueba cualitativa de solubilización, la cual
permitió definir, de acuerdo a la medida del halo de hidrólisis formado en cada caso,
aquellas que presentaron mejor capacidad de solubilizar el fosfato. Ver figura 6.
Figura 6. Medición del halo de solubilización en medio SRS con fosfato tricálcico a las
24 y 48 h de la cepa Relef16.
Fuente: autores
La tabla 14 deja ver las cepas seleccionadas de mayor funcionalidad para convertir el
fosfato insoluble y no disponible en fósforo soluble y asimilable por la planta; de igual
manera la tabla muestra los halos de solubilización formados a las 24 y 48 h.
Los resultados de la prueba muestran el valor promedio y la desviación estándar de
dos mediciones del halo de solubilización, observándose que el tamaño de éste
aumentó para todas la cepas de un periodo de tiempo al otro, aunque en algunos casos
el aumento fue mayor: RCH10 (Δ 1.65 mm) > RCH2 (Δ 1.1 mm) > RCH1 (Δ 0.25 mm).

69
Tabla 14. Medida del halo de solubilización en medio SRS con fosfato tricálcico
ID cepas Diámetro del halo (mm ± DE)
24h 48h RCH1 8,1 ± 0,9 8,3 ± 0,5 RCH2 7,8 ± 0,5 8,9 ± 0,3 RCH3 6,4 ± 0 7,4 ± 0,6 RCH10 8,4 ± 0 10,0 ± 2,3 RCH17 2,1 ± 0,4 5,1 ± 0,21 RELEF9 3,2 ± 1,1 4,9 ± 0,3 RELEF10 3,1 ± 0,5 4,25 ± 0,2 RELEF13 4,1 ± 0,5 5,0 ± 0,3 RELF14 6,6 ± 0,2 7,1 ± 0,1 RELEF16 6,0 ± 0,8 8,1 ± 1 ENDOELEF10 5,8 ± 0,8 7,9 ± 0,2
En un ensayo realizado por Faccini y Garzón (1997), obtuvieron halos entre 2.4 a 3.5
mm trabajando con medio Pikovskaya con fosfato tricálcico como fuente de fosfato,
Martínez y Martínez (1996), encontraron halos de solubilización de las cepas probadas
entre 0.5 a 3.5 mm, al igual que Beltrán et al (2005); en su estudio reportaron medidas
de halos entre 2 y 20 mm. De otra parte, Barragán y Zambrano (2003), trabajando en
medio SRS observaron a las 48 h halos de solubilización de 11.83 y 18.16 mm para
Azotobacter chrococcum y Burkholderia cepacia, respectivamente. Rodriguez y Urrego
(2003) por su parte, obtuvieron halos entre 5 y 7 mm en el mismo medio.
En nuestro trabajo los halos de solubilización de las 11 cepas probadas oscilaron entre
2.1 ± 0.4 y 8,05 ± 0,9 mm a las 24 h de incubación, y entre 4,25 ± 0,2 a 10,05 ± 2,3
mm a las 48 h; con lo que podría afirmarse que estas cepas bacterianas, tienen una
buena capacidad de solubilización del fosfato comparada con lo revelado por otros
autores en estudios realizados con propósitos similares.
De acuerdo al criterio aplicado (tamaño del halo) para seleccionar las cepas con mejor
capacidad de solubilización, se escogieron a RCH1, RCH2, RCH10 como las de mejor

70
comportamiento en la prueba. Con estas 3 cepas bacterianas se determinó realizar
algunos ensayos adicionales que confirmaran su capacidad para ser utilizadas en un
proceso de biorremediación.
4.4.5 Determinación de la capacidad bacteriana para solubilizar el fósforo:
El contenido de fosfato disponible, se puede observar en la figura 7.
Figura 7. Determinación del fosfato disponible después de la acción solubilizadora de
las 3 cepas seleccionadas
Fuente: autores
No obstante a que la figura 7 muestra básicamente el fósforo disponible en función del
tiempo de experimentación, indirectamente deja ver la habilidad metabólica de las
cepas bacterianas utilizadas en el ensayo para solubilizar el fosfato y permitir su
disponibilidad. Con esta óptica podrían hacerse algunas observaciones:
• Se nota una clara diferencia entre el control y los tratamientos, dejando ver
entre ellos un comportamiento semejante en los 4 primeros días del ensayo
con una actividad acelerada en este período de tiempo en las tres cepas de
bacterias.
0
20
40
60
80
100
120
140
160
180
0 5 10 15
PO4
(mg/
l)
Días
CONTROL
RCH1
RCH2
RCH10

71
• Las tres diferentes cepas disminuyen su actividad a partir del cuarto día, no
obstante RCH1 mantiene una mayor actividad que las otras dos.
• Entre el día cuatro y el octavo la disponibilidad del fósforo decae, quizá porque
es utilizado por las bacterias en la medida que se produce; RCH2 y RCH10
evidencian mayor actividad.
• A partir del octavo día, RCH1 parece verse afectada en gran medida por la
acidez del medio ya que su funcionalidad decrece en forma evidente; en tanto
que RCH2 y RCH10 entran en franca recuperación al reanudar la actividad
enzimática o quizá fueron sucedidas por bacterias oportunistas que logran
adaptarse mejor a los pHs ácidos del medio, volviéndose a notar un incremento
en los niveles del fósforo disponible.
De esta forma, puede inferirse que las tres cepas bacterianas podrían actuar
conjuntamente en un proceso de biorremediación de aguas, concretamente sobre
fosfatos provenientes de detergentes de uso común en la población y que llegan al rio a
través de las aguas servidas. No obstante, el proceso de depuración podría iniciarse
con cepas RCH1 y continuar con RCH2 y RCH10 cuando el pH del medio haya
descendido.
Figura 8. Resultados del ANOVA (GLM) para la determinación del fósforo disponible

72
A partir del ANOVA (GLM) realizado a los resultados del ensayo (figura 8), se demostró
que existe una diferencia significativa entre la cantidad de fosfato disponible
cuantificado en el control y el cuantificado con las 3 cepas utilizadas en la prueba, es
decir, se puede afirmar con 95% de probabilidad que las cepas RCH1, RCH2 y RCH10
se diferencian del control y evidencian capacidad de solubilizacion del fosforo. El factor
tiempo, estadísticamente no evidencio un efecto significativo sobre la cantidad de PO43,
posiblemente porque los valores de PO4-3 cuantificados en los diferentes tiempos del
ensayo, están influenciados por el consumo de fósforo que hacen las bacterias.
Además no hubo ningún efecto de las interacciones entre factores.
Figura 9. Resultados que evidencian la normalidad de los datos y el ajuste del modelo

73
En los últimos años, se ha avanzado en la comprensión de los procesos bioquímicos
involucrados en la solubilización de fósforo inorgánico mediada por ácidos orgánicos
(He Z et al, 2006; Ramachandran et al 2006). Un resumen de los principales
mecanismos involucrados en la solubilización de fosfatos por bacterias se muestra en
la tabla 15, alguno(s) de los cuales podría (n) explicar, al menos en parte, la
bioactividad observada por las cepas objeto del ensayo en nuestro trabajo.
Tabla 15. Principales procesos microbianos involucrados en la solubilización de
fosfatos
TIPO DE PROCESO
PRINCIPAL CAUSA DE DISOLUCIÓN MINERAL
PRINCIPAL REACCIÓN QUE LLEVA A LA
DISOLUCIÓN MINERAL
APLICABILIDAD A FOSFATOS MINERALES
REFERENCIAS
Acidificación del medio
Liberación de protones (H+) o producción de ácidos inorgánicos fácilmente disociables
Disminución del pH del medio, formación de hidrofosfatos de mayor solubilidad
Fosfatos de Ca+2
(Rodríguez & Fraga, 1999; Richardson & Simpson, 2011)
Formación de complejos metálicos
Liberación de ácidos orgánicos o complejos (quelantes)
Formación de complejos metálicos (incluyendo quelatos en el caso de ácidos di-tricarboxílicos o hidrocarboxílicos)
Fosfatos de Ca, Al+3 y Fe+3
(Paredes & Espinosa , 2010; Illmer & Schinner, 1995; Chen et al, 2006)
Reducción de metales
Actividad redox de bacterias o sus exudados (metabolitos secundarios)
Reducción de metales con estados de oxidación variable (ligados a fosfatos) a un bajo estado de oxidación (resultando en un fosfato más soluble)
Fe+3 fosfato
(Arcand & Schneider, 2006; Gerretsen, 1948)
Disolución de fosfatos mediada por enzimas
Liberación extracelular de enzimas específicas (Fosfatasas)
Hidrólisis enzimática de esteres de fosfato orgánicos pobremente solubles liberando fosfatos inorgánicos
Varios ésteres de fosfatos orgánicos (fitatos, fosfolípidos)
(Rodríguez et al, 2006; Schwab, 1989; Nannipieri et al 2011)
Disolución indirecta de fosfatos
Estimulación microbiana de exudación de ácidos orgánicos a la planta
El mismo mecanismo para el tipo 2 (formación de complejos metálicos) pero liberados por la interacción planta-microorganismo
Fosfatos de Ca+2, Al+3 y Fe+3 (Chen et al, 2006)

74
4.4.6 Selección de cepas bacterianas con capacidad de transformación del amonio:
Se quiso comprobar la capacidad de la totalidad de las cepas bacterianas aisladas de
transformar el (NH4)2SO4 en nitritos y nitratos para obtener energía a partir de estas
reacciones, habilidad propia de las bacterias nitrificantes. En el ensayo se utiliza caldo
de amonio como única fuente de energía. La bacteria debe ser capaz de transformar el
nitrógeno suministrado a partir de sulfato de amonio, (NH4)2SO4, para su desarrollo. Sin embargo, en este trabajo, una vez transcurrido el período de incubación de las
bacterias, no se evidenció un incremento significativo en la biomasa bacteriana (figura
10). De igual manera, al realizar las pruebas de detección de nitritos y nitratos (reactivo
Griess y Zinc en polvo, respectivamente) no se observaron cambios de coloración en
ninguno de los tubos que permitiera sacar conclusiones en relación a la bioactividad
bacteriana; por el contrario, el reactivo de Nessler permitió detectar en todos los tubos
la presencia de amonio. Ver figura 11
Figura 10. Registro del comportamiento de la biomasa durante el periodo de
incubación de la bacteria en el caldo amonio
Cepa Relef 5 a los 7 de incubación Fuente: autores

75
Figura 11. Detección de (NH4)2 SO4 con el reactivo de Nessler, en cada uno de los
tubos incubados con las cepas bacterianas.
Fuente: autores
Los resultados obtenidos indican que ninguna de las cepas aisladas tiene la capacidad
de oxidar el amonio, NH4+, del sulfato de amonio, (NH4)2SO4, a nitrito (NO2–), y a partir
de éste último transformarlo a nitrato (NO3-); es decir, no hubo oxidación. Se entiende
entonces que las cepas objeto del ensayo no son microorganismos aerobios con
habilidad para utilizar el oxígeno molecular (O2) como receptor de electrones.
El proceso sigue unos pasos en los que el átomo de nitrógeno se encuentra sucesivamente bajo las siguientes formas:
Nitrificación Oxidación Desnitrificaciòn Amonio Nitrito Nitrato NH4
+ (nitrosomonas) NO2– (nitrobacter) NO3
- (Pseudomonas) NO2 NO N2O N2 Dióxido de nitrógeno óxido nítrico óxido nitroso
No obstante, las plantas, ya sean terrestres, acuáticas o de la frontera entre estos dos
sistemas, tal como la vegetación riparia, toman el nutriente en forma de nitrato, NO3 -.
La concentración de nitratos en aguas superficiales normalmente es baja (0-18 mg/L),
pero puede llegar a alcanzar elevados niveles como consecuencia de las prácticas
agrícolas o residuos urbanos y ganaderos; la presencia de especies riparias en aguas

76
contaminadas se constituye entonces en un controlador por los excesos de este
componente químico.
Como ya se dijo, en el proceso de nitrificación biológica ocurre la oxidación secuencial
aeróbica del amonio a nitrito y luego a nitrato. Estos dos pasos están catalizados por
organismos procariotas quimilitótrofos denominados: bacterias amonio oxidantes
(AOB), Archaea amonio oxidantes (AOA) y bacterias nitrito oxidantes (NOB). De esta
manera cada paso del proceso es realizado por un tipo de bacterias distinto. Hasta el
momento no existen organismos capaces de realizar ambos pasos por si mismos
(Daims et al, 2009).
Debido a que no se encontraron bacterias amonio oxidantes (AOB), se buscó entonces
conocer si estas cepas podrían tener la habilidad de utilizar el nitrito y oxidarlo a nitrato,
es decir de comportarse como bacterias nitrito oxidantes (NOB).
4.4.7 Selección de cepas con capacidad de trasformación del nitrito: En esta prueba se
trabajó con la totalidad de los aislados (rizobacterias y bacterias endófitas). Pasado el
periodo de incubación de 14 días para el crecimiento de bacterias nitrito oxidantes en
medio autotrófico para Nitrobacter 756c (DSMZ), se evidenció crecimiento bacteriano
típico en forma de gotas de agua por parte de algunas de las cepas probadas. Se
seleccionaron aquellas que crecieron con esta característica y se realizó un ensayo
cualitativo de comprobación de la utilización y transformación de nitrito por parte de
estas. Los resultados de este ensayo se muestran la tabla 16.

77
Tabla 16. Ensayo de detección de la transformación de nitrito en el medio de cultivo, utilizando el reactivo de Griess y Zn en polvo.
A B C D
MEDIO MEDIO + BACTERIA T° 24 h
RCH4 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLO
RCH6 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLORCH11 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLORCH13 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLORCH14 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLORCH16 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA TENUE AMARILLORCH17 ROJIZO NARANJADO AMARILLO NARANJADO NARANJA AMARILLORELEF4 AMARILLO NARANJADO AMARILLO NARANJA TENUE AMARILLO PALIDORELEF6 AMARILLO NARANJADO AMARILLO NARANJADO TENUE NARANJA INTENSO AMARILLO PALIDORELEF9 AMARILLO AMARILLO TENUE NARANJA AMARILLO MUY TENUE
RELEF10 AMARILLO INTENSO AMARILLO INTENSO NARANJA AMARILLO MUY TENUERELEF13 AMARILLO AMARILLO INTENSO NARANJA FUERTE AMARILLO TENUERELEF14 AMARILLO AMARILLO INTENSO NARANJA FUERTE AMARILLO TENUERELEF18 ROJIZO AMARILLO NARANJA FUERTE AMARILLO TENUE
ENDOCH1 AMARILLO NARANJADO AMARILLO TENUE NARANJA FUERTE NARANJA TENUEENDOCH2 AMARILLO INTENSO AMARILLO INTENSO NARANJA FUERTE AMARILLO MUY TENUEENDOCH5 AMARILLO NARANJADO AMARILLO NARANJA MUY FUERTE NARANJA TENUEENDOCH6 AMARILLO INTENSO AMARILLO NARANJA FUERTE AMARILLO TENUE
ENDOELEF3 AMARILLO INTENSO AMARILLO NARANJA AMARILLO TENUEENDOELEF4 AMARILLO AMARILLO NARANJADO NARANJA AMARILLO TENUEENDOELEF7 AMARILLO NARANJADO AMARILLO NARANJA INTENSO AMARILLO TENUE
ENDOELEF10 AMARILLO INTENSO AMARILLO NARANJA FUERTE NARANJA TENUEENDOELEF12 AMARILLO AMARILLO TENUE NARANJA AMARILLO MUY TENUE
CONTROL DETECCION DE NITRITOCONTROL REACTIVO DE GRIESSID CEPA
NARANJA AMARILLO
MEDIO + BACTERIA+ RG+ Zn polvo
RCH1 ROJIZO NARANJADO AMARILLO NARANJADO
REACTIVO DE GRIESS (RG)
INCOLORO AMARILLO NARANJADO
0

Como control se utilizó una porción del medio sin bacteria, teniendo la precaución de
tomar la muestra lo más alejada de la población microbiana. Se adicionó RG (reactivo
de Griess) para detectar la presencia de nitritos (NO2-) en esta porción medio. La
columna A muestra las coloraciones observadas (fundamentados en que el RG en
presencia del ion NO2- se torna de color rojizo). Los resultados observados en la
columna A permiten hacer 2 grupos de bacterias, dado que en algunos casos se nota
disminución de la tonalidad rojiza (< cantidad de nitrito detectado), llegando incluso a
coloraciones amarillas de baja intensidad. La prueba se repitió tomando como muestra
una porción del medio + bacteria. Lo observado se muestra en la columna B,
claramente se nota que en todos los casos, hay una decoloración, en relación con el
control. Unas horas después se evidenció que las coloraciones se tornaban rojizas,
indicando que aún persiste una porción relativa mente baja de NO2- .
El método sugiere que si no hay cambio de color (rojizo), se adicione polvo de Zn, no
obstante en nuestro experimento se decidió agregar en todo los casos este reactivo,
que detecta la presencia de NO3- . Las tonalidades observadas en un tiempo inmediato
y después de 24h, se muestra en las columnas C y D, respectivamente. Es evidente
que la columna C muestra que el grupo de bacterias transformó el NO2- a NO3-
(coloración naranja) al adicionar el Zn (polvo), no obstante algunos de los organismos
muestran mayor actividad que los demás, esto se aprecia al observar que la coloración
naranja persiste en poco casos, después de las 24h de haber agregado el Zn, mientras
los demás lo pierden y se ven tonalidades hacia el amarillo. La persistencia de esta
coloración naranja puede ser debida a que en estos casos la cantidad de NO3- que
reaccionó con el Zn es mayor, evidenciando una mayor transformación del NO2- a
NO3-. En los demás casos es muy poco y se pierde (tonalidades amarillos). De acuerdo
a esta observación se decidió seleccionar las cepas que mostraron este
comportamiento en mayor intensidad.
De esta manera se evidenció que un total de 24 cepas bacterianas tienen la capacidad
de transformar el nitrito a nitrato, 12 de estas cepas se asilaron de la raíz del Chilco (8
rizobacterias y 4 endófitas), las restantes 12 se hallaron en el Pasto Elefante (7

rizobacterias y 5 endófitas), entre este grupo de bacterias seleccionaron a ENDOCH1,
ENDOCH5, ENDOELEF10, como las de mejor comportamiento en la transformación
del nitrito. Estas 3 cepas seleccionadas muestran potencial para su utilización en un
proceso de biorremediacion de exceso de nitrito y posibilitan disponibilidad de nitrato a
las plantas que allì habitan, por ejemplo el Chilco y el Pasto Elefante donde fueron
halladas.
Las bacterias nitrito oxidantes (NOB) forman cuatro grupos filogenéticos distintos:
Nitrobacter, Nitrococcus, Nitrospina y Nitrospira (Mota et al, 2005). Estas bacterias
tienen un papel importante en la regulación y eliminación del exceso de nitrógeno, ya
que realizan la conversión del nitrito a nitrato, paso final del proceso de nitrificación. El
siguiente proceso, comúnmente conocido “desnitrificación” devuelve el nitrógeno fijado
a la atmósfera por procesos de respiración microbiana, a través de la reducción
desasimilatoria de nitratos (NO3-) y nitritos (NO2-) a N2O y N2, respectivamente, como
se ve este mecanismo contribuye a mantener el equilibrio del nitrógeno en el suelo.
Las plantas y diferentes microorganismos - arqueas, bacterias y hongos son capaces
de incorporar los nitratos producto del proceso mediado por los NOB en su biomasa, lo
que reduce la perdida por lixiviación y desnitrificación (Gorfer et al., 2011). La presencia
de NOB en la raíz del chilco y pasto elefante, garantizan una disponibilidad de
nitrógeno en forma de nitratos para que estas especies lo asimilen en su metabolismo,
mejorando su condición de adaptación a la escasez de nutrientes y explican el porqué
de la relación simbiótica con este tipo de bacterias, que se encuentran en gran número
en la raíz de esta plantas (Chilco, 12 cepas halladas, 8 rizobacterias y 4 endófitas y en
Pasto Elefante: 12 cepas, 7 rizobacterias y 5 endófitas).
Quizá una razón que explica el no haber encontrado bacterias oxidantes de amonio
(AOB) en la raíz de estas especies es porque éstas no tienen una funcionalidad tan
determinante para la planta como las NOB, las cuales permiten la disponibilidad del
nitrato, y están más relacionadas con los niveles de nitrógeno altos en el suelo.

4.4.8 Ensayo de verificación de biorremediación en un agua residual sintética: La
prueba de laboratorio realizada para verificar la capacidad de biorremediación de un
agua residual por parte de las bacterias seleccionadas como de mejor comportamiento
(3 rizobacterias y 3 endófitas) se efectuó, como ya fue dicho en la metodología,
preparando un agua elaborada mediante la mezcla de componentes químicos que
simularan las condiciones naturales de un agua en alto grado de contaminación. No
obstante, los resultados obtenidos en más de 5 repeticiones realizadas arrojaron
resultados que revelaban la baja capacidad de los organismos escogidos para reducir
el nivel de degradación de la muestra sintética. Este resultado da entender que es todo
el grupo de organismos, rizobacterias y endófitas, las que actúan conjuntamente y no
unas pocas de ellas como se pretendió hacer en este trabajo aunque éstas fueran las
más activas. Importa recordar que de las dos especies seleccionadas se aislaron un
total de 52 aislados y es preciso hacer ver que no es fácil recrear los factores bióticos y
abióticos igualmente importantes para el desarrollo de este ensayo.
De otra parte, se presentaron problemas de contaminación del control que impidieron
insistentemente realizar un análisis comparativo entre la muestra y éste.

CONCLUSIONES
A través del presente estudio se definió la composición florística del bosque
ripario rio Combeima-UT (BRC-UT) que alberga en 50 m2, 29 especies
vegetales, distribuidas en 17 familias, destacándose las familias Asteraceae y
Poaceae como las de mayor diversidad por el número de especies y por tener
las especies más abundantes en la zona.
Los parámetros ecológicos (cobertura vegetal, biomasa y densidad de especie),
indicaron que las dos especies de mayor importancia ecológica en el BRC-UT
son Pseudelephantopus spiralis (Less.) Crong (180 individuos) y Pennisetum
purpureum Schumach (326 individuos).
De la raíz de estas dos especies vegetales, se aisló un total de 35 rizobacterias;
17 cepas de la raíz del Chilco (Pseudelephantopus spiralis) y 18 aislados del
Pasto Elefante (Pennisetum purpureum). Se aislaron además, 21 bacterias
endófitas, 7 de la raíz del Chilco y 14 del Pasto elefante, que a su vez
comprueban la importancia de la raíz de estas especies en los procesos de
adaptación a las condiciones de la ribera y a los procesos de transformación de
los nutrientes en esta zona particularmente dinámica.
A partir de la raíz de ambas especies vegetales, se aislaron 11 cepas
bacterianas con capacidad solubilizadora de fósforo inorgánico, una sola de ellas
(ENDOELEF10) resultó ser una bacteria endófita de P. purpureum (Pasto
Elefante); todo lo cual demuestra la funcionalidad de la rizósfera en la
transformación de este nutriente.
Las cepas RCH1 RCH2 y RCH10 mostraron la mejor capacidad para solubilizar
el fósforo y convertirlo en fosfato disponible, lo que demuestra el potencial para
ser usadas en un sistema de fitorremediación de aguas cargadas de
detergentes. Por su parte ENDOCH1, ENDOCH5, ENDOELEF10 se revelaron

como las de mejor capacidad de oxidación de NO2- (NO2- a NO3-), lo cual
sustenta su aplicación en procesos de la biorremediacion para aguas con
exceso de nitritos.
Pennisetum purpureum y Pseudelephantopus spiralis pueden ser tenidas en
cuenta en un proceso de fitorremediación de aguas con alta carga de nitrógeno
y fósforo
Para nuestro conocimiento este es uno de los pocos trabajos realizados que
plantea la recuperación de las aguas del rio Combeima utilizando los propios
recursos que el rio ofrece, como lo es la vegetación riparia y los
microorganismos de la rizosfera de estas plantas.

RECOMENDACIONES
Es necesario realizar ensayos biológicos donde se compruebe la actividad
fitorremediadora de estas especies vegetales en aguas residuales domésticas.
Puesto que es evidente que el rio Combeima sufre por la contaminación de sus
aguas y su entorno ecológico, se hace necesario implementar estrategias para la
recuperación de este ecosistema acuático tan importante para las actividades de
desarrollo de Ibagué y de su población. Una podría ser utilizar la vegetación
riparia, como es el caso de Pseudelephantopus spiralis (Less.) Crong y
Pennisetum purpureum Schumach, como barrera de protección o como medida
de restauración ecológica en zonas donde se haya perdido este tipo de
vegetación que sirve de mecanismo natural de autodepuración.

REFERENCIAS BIBLIOGRÁFICAS
- Acelas, N.; Martin, B.; López, D. & Jefferson, B. (2014). Selective removal of
phosphate from wastewater using hydrated metal oxides dispersed within anionic
exchange media. Chemosphere, 119, 1353–1360.
- Aguado, G.; Moreno, B.; Jiménez, B.; García, E. & Preciado, R. (2012). Impacto
de los sideróforos microbianos y fitosidéforos en la asimilación de hierro por las
plantas: una síntesis. Revista fitotecnia Mexicana, 35 (1) online. ene/mar.
- American Public Health Association. (1984). Standard Methods For the
Examination of Water and Wastewater. American Public Health Association.
Washington, D.C.
- American Public Health Association (APHA), American Water Works Association
(AWWA), Water Environment Federation (WEF). (2005). In: Eaton, A.D, Clesceri,
L.S., Rice, E.W., Greenberg, A.E. (Eds.) 21st edition, Standard Methods for the
Examination of Water and Wastewater. APHA, AWWA, and WEF Publishing,
Washington DC.
- Angelsen, A. & Kaimowitz, D. (2001). Agricultural technologies and tropical
deforestation. CABI Publishing. Wallingford, UK; New York, NY.
- Araújo, W.L.; Marcon, J.; Maccheroni, W.Jr.; van Elsa, J.D.; van Vuurde J.W.L. &
Azevedo J.L. (2002). Diversity on endophytic bacterial populations and their
interaction with Xylella fastidiosa in Citrus plants. Appl Environ Microbiol, 68,
4906-4914.

- Arbo, M.; López, M.; Schinini, A. & Pieszko, G. (2000). Plantas palustres del
Macrosistema Iberá. Instituto de Botánica del Nordeste (IBONE) - Facultad de
Cs. Agrarias - UNNE.Sargento Cabral 2131 - (3400) Corrientes - Argentina.
- Arcand, M. & Schneider, K. (2006). Plant and microbial-based mechanisms to
improve the agronomic effectiveness of phosphate rock: a review. Annals of the
Brazilian Academy of Sciences, 78, 791-807.
- Atterby, H.; Smith, N.; Chaudhry, Q. & Stead, D. (2002). Exploiting microbes and
plants to clean up pesticide contaminated environments. Pesticide Outlook.
February, 9–13.
- Bacon, C.W., & White, J.F. (2000). Microbial endophytes. Marcel Dekker Inc.,
New York, N.Y.
- Baker, A. (2002). Fluorescence excitation–emission matrix characterization of
river waters impacted by a tissue mill effluent. Environ.Sci.Technol, 36, 189–195.
- Baker, A.; Inverarity, R.; Charlton, M. & Richmond, S. (2003). Detecting river
pollution using fluorescence spectrophotometry: case studies from the Ouseburn,
NE England. Environmental Pollution, 124 , 57–70
- Baker, A.; Ward, D.; Lieten, S.H.; Periera, R.; Simpson, E.C. & Slater, M. (2004).
Measurement of protein-like fluorescence in river and waste water using a
handheld spectrophotometer. Water Research, 38, 2934–2938.
- Barea, J. M., Azcón, R. y Azcón-Aguilar, C. 2004. Mycorrhizal fungi and plant
growth promoting rhizobacteria. In: Varma A, Abbott L, Werner D, Hampp, R.
eds. Plant surface microbiology. Heardelberg, Germaniy. Springer-Verlag. 351-
371.

- Barea, J. M., Pozo, M. J., Azcón, R. y Azcón-Aguilar, C. 2005. Microbial co-
operation in the rhizosphere. Journal of Experimental Botany, 56, 1761-1778.
- Barragan, C. & Zambrano, D. (2003). Produccion de bacterias fosfato
solubilizadoras mediante fermentacion discontinua en caldo Pikovskaya
modificado. Trabajo de Grado. Carrera de Microbiologia Industrail. Pontificia
Universidad Javeriana. Colombia. 122 p.
- Beltran, S.; Torrado, Y.; Martinez, M & Matiz, A. (2005). Aislamiento de bacterias
con actividad fosfato solubilizadora y produccion de un inoculo mixto en
fermentacion discontinua. Trabajo de Grado. Carrera de Microbiologia Industrial.
Pontificia Universidad Javeriana. Colombia. 105 p.
- Beltrán, S.; Torrado, Y.; Martínez, M. & Matiz, A. (2005). Asilamiento de
bacterias con actividad fosfato solubilizadora y producción de un inoculo mixto
en fermentación discontinua. (Tesis de pregrado). Facultad de ciencias:
Pontificia Universidad Javeriana. Colombia.
- Bitton, G. (2005). “Wastewater Microbiology” Third Ed., Wiley-Liss, New York.
- Bobadilla, C. & Rincón, S.C. (2008). Aislamiento y producción de bacterias
fosfatos solubilizadoras a partir de compost obtenido de residuos de plaza.
(Tesis pregrado). Facultad de Ciencias: Pontificia Universidad Javeriana.
Colombia.
- Bustillo, C.; Mehrvar, M. & Quiñones E. (2013). Combined anaerobic-aerobic and
UV/H2O2 processes for the treatment of synthetic slaughterhouse wastewater.
Journal of environmental science and health. Part A, Toxic/hazardous
substances & environmental engineering, 48 (9), 1122-35.
- Cardon, Z.G. & Whitbeck, J.L. (2007). The Rhizosphere An Ecological
Perspective. Primera edición. Academic Press. USA. pp. 212.

- CCAD-PNUD/GEF. (2002). El Corredor Biológico Mesoamericano: una
plataforma para el desarrollo sostenible regional/ Proyecto para la consolidación
del Corredor Biológico Mesoamericano. Managua, Nicaragua. 1, 24.
- Chalar, G. (2006). Dinámica de la Eutrofización a Diferentes Escalas
Temporales: Embalse Salto Grande (Argentina-Uruguay). En: Eutrofizaçãona
América do Sul: causas, consequências e tecnología para gerenciamento e
control. Editado. J. GaliziaTundisi, T. MatsumuraTundisi & C. SidagisGalli. São
Carlos, SP, Brasil. 87-101.
- Chará, J.O. (2003). Manual para la evaluación biológica de ambientes acuáticos
en microcuencas ganaderas. Fundación centro para la investigación en sistemas
sostenibles de producción agropecuaria. Colombia. CIPAV.
- Chaudhry, Q.; Blom–Zandstra, M.; Gupta, S. & Joner, E.J. (2005). Utilising the
synergy between plants and rhizosphere microorganisms to enhance breakdown
of organic pollutants in the environment. Environ. Sci. Pollut. Res., 12, 34–48.
- Chen, Y.; Rekha, P.; Arum, A.; Shen, F.; LA, W. & Young, C. (2006). Phosphate
solubilizing bacteria from subtropical soil and their tricalcium phosphate
solubilizing abilities. Applied Soil Ecology, 34, 33-41.
- Church, M. (2002). Geomorphic thresholds in riverine landscapes. Freshwater
Biol, 47, 541–557.
- Comisión Europea. Aplicación de la Directiva 91/271/CEE del Consejo. (1991),
sobre el tratamiento de las aguas residuales urbanas. Luxemburgo: Oficina de
Publicaciones Oficiales de las Comunidades Europeas.

- Corbacho, C.; Sánchez, J.M. & Costillo, E. (2003). Patterns of structural
complexity and human disturbance of riparian vegetation in agriculture
landscapes of a Mediterranean area. Agriculture, Ecosystems and Environment,
13, 495-507.
- Daims, H.; Maixner, F. & Schmid, M.C. (2009). The Nitrifying microbes: Ammonia
oxidizers, nitrite oxidizers, and anaerobic ammonium oxidizers. En: Nielsen, P.,
Daims, H y Lemmer, H. 2009. FISH Handbook for Biological Wastewater
Treatment: Identification and quatification of microorganisms in activated sludge
and biofilms by FISH. p 9-17. Ed. IWA Publishing. Londres, Reino Unido.
- Dapena, A.; Campos, J.L.; Mosquera, A. & Mendez, R. (2006). Anammox
process for nitrogen removal from anaerobically digested fish canning effluents. Water Science & Technology, 53 (12), 265–274.
- Dapena, A.; Campos, J.L.; Mosquera, A.; Jetten, M. & Mendez, R. (2004).
Stability of the AMAMMOX process in a gas-lift reactor and a SBR. Journal of
Biotechnology, 110, 159–170.
- Dodds, W.K. (2002). Freshwater Ecology. Concepts and environmental
applications. Academic Press. San Diego.
- Dorioz, J.; Wang, D.; Poulenard, J. & Trevisan, D. (2006). The effect of grass
buffer strips on phosphorus dynamics-A critical review and synthesis as a basis
for application in agricultural landscapes in France. Agriculture, Ecosystems &
Environment, 117, 4-21.
- Dueholm, T.E.; Andreasen, K.H. & Nielsen, P.H. (2001). Transformation of lipids
in activated sludge. Water Sci. Technol. 43, 165–172.

- Echeverri, L.; Estévez, J.V. & Bedoya, J.G. (2014). Caracterización física,
química y mineralógica de suelos con vocación forestal protectora, región Andina
Central Colombiana. Rev.Fac.Nal.Agr.Medellín, 67(2), 7335-7343.
- Edwards, C.A. (2000). Earthworms. Chapter 8 in: Soil Biology Primer. Soil and
Water Conservation Society. Rev. Edition. Ankeny Iowa.
- Fabiano, M.; Marrale, D. & Misic, C. (2003). Bacteria and organic matter
dynamics during a bioremediation treatment of organic-rich harbour sediments.
Marine Pollution Bulletin, 46 (9), 1164-1173.
- Faccini, G. & Garzon, S. (1997). Evaluacion del efecto de un inoculo dual de
bacterias solubilizadoras de fosfato y Azotobacter chroococcum en el cultivo de
papa criolla var yema de huevo (Solanum phureja). Trabajo de Grado. Carrera
de microbiologia Industrial. Pontificia Universidad Javeriana. Bogota. pg 57-65
- Fernández, A.; Martínez, M.; Nerín de la Puerta, C.; Pla Martínez, A. & López,
R. (2011). Informe del Comité Científico de la Agencia Española de Seguridad
Alimentaria y Nutrición (AESAN) en relación a la evaluación del riesgo de la
exposición de lactantes y niños de corta edad a nitratos por consumo de acelgas
en España. Ministerio de sanidad, servicios sociales e igualdad, España.
- Flórez, Y. & Cotes, A. (2006). Bioabsorción de metales pesados por salvinia
natans de los lixividados del relleno sanitario Combeima de Ibagué. Revista
Tumbaga, 1, 89-100
- Franks, A.; Ryan, P.R.; Abbas, A.; Mark, G.L. & O’Gara, F. (2006). Molecular
tools for studying plant growth-promoting Rhizobacteria (PGPR): Molecular
techniques nfor soil and rhizosphere microorganisms. CABI Publishing,
Wallingford, Oxfordshire, UK.

- Fuentes, F. & Massol-Deyá, A. (2002). Manual de laboratorios ecología de
microorganismos. Departamento de Biologia. Universidad de Puerto Rico.
- Gangbazo, G.; Pesant, A.; Barnett, G.; Charuest, J. & Cluis, D. (1995). Water
contamination by ammonium nitrogen following the spreading of hog manure and
mineral fertilizers. Journal of Environmental Quality, 24 (3), 420-425.
- Garnier, J.; Billen, G. & Servais, P. (1992b). Physiological characteristics and
ecological role of small and large sized bacteria in a polluted river (Seine River,
France). Archiv für Hydrobiologie–Beiheft Ergebnisse der Limnologie, 37, 83-94.
- Garnier, J.; Servais, P. & Billen, G. (1992a). Dynamics of bacterioplankton in the
river Seine (France): impact of parisian effluents. Canadian Journal of
Microbiology, 38, 56-64.
- Gentry, T.J.; Rensing, C. & Pepper, I.L. (2004). New approaches for
bioaugmentation as a remediation technology. Critical Reviews in Environmental
Science and Technology, 34, 447-494.
- Germain, K.; Lui, X.; Cabellos, G.; Hogan, J.; Ryan, D. & Dowling, D.N. (2006).
Bacterial endophyte-enchanced phyto-remedation of the organochorine herbicide
2,4 dichlorophenoxyacetic acid. FEMS Microbiol Ecol, 57, 302-310.
- Gerretsen, F.C. (1948). The influence of microorganisms on the phosphate
intake by the plant. Plant and Soil I, 1, 51-81.
- Gessesse, A.; Dueholm, T.; Petersen, S.B. & Nielsen, P.H. (2003). Lipase and
protease extraction from activated sludge. Water Res. 37, 3652–3657.
- González, E.M.; Pedraza, A. & Pérez, M.M. (2009). Caracterización agrológica
del suelo y diagnóstico de su fertilidad en la estación experimental del campus

nueva granada, Cajicá (Cundinamarca, Colombia). Revista Facultad de ciencias
básicas, 4 (1), 82-104.
- González, L.E.; Cañizares, R.O.; & Baenaa, S. (1997). Efficiency of ammonia
and phosphorus removal from a colombian agroindustrial wastewater by the
microalgae chlorella vulgaris and scenedesmus dimorphus. Bioresource
Technology, 60, 259-262.
- Gorfer, M.; Blumhoff, M.; Klaubauf, S.; Urban, A.; Inselsbacher, E.; Bandian,
D…Strauss, J. (2011). Community profiling and gene expression of fungal
assimilatory nitrate reductases in agricultural soil. Multidisciplinary Journal of
Microbial Ecology, 5 (11), 1771-1783. Recuperate of: doi:10.1038/ismej.2011.53
- Gray, D.H. & Leiser, A.T. (1989). Biotechnical Slope Protection and Erosion
Control. Robert E. Krieger Publ. Co., Malabar, Florida.
- He Z, O.T.; Cade-menun, B.J.; Erich, M.S & Honeycutt C.W. (2006). Spectral
and chemical characterization of phosphates associated with humic substances.
Soil Science Society of America Journal, 70, 1741-51.
- Ho, Y.; Hsieh, J. & Huang, C. (2013). Construction of a plant–microbe
phytoremediation system: Combination of vetiver grass with a functional
endophytic bacterium, Achromobacter xylosoxidans F3B, for aromatic pollutants
removal. Bioresource Technology,145, 43-47.
- Hooda, P.; Edwards, A.; Anderson, H. & Miller, A. (2000). A review of water
quality concems in livestock farming areas. Science of The Total Environment.
250, 143-167.
- Huang, L.; Zhuo, J.; Guo, W.; Spencer, R.; Zhang, Z. & Xu, J. (2013). Tracing
organic matter removal in polluted coastal waters via floating bed

phytoremediation. Marine Pollution Bulletin. Recuperate of
:http://dx.doi.org/10.1016/j.marpolbu l.2013.03.0 32
- Hughes, D. (1981). Ecología de las civilizaciones antiguas. México: Fondo de
Cultura Económica.
- Hurek, T. & Reinhold-Hurek, B. (2003). Azoarcus sp. strain BH72 as a model for
nitrogen-fixing grass endophytes. Journal of Biotechnology, 106, 169-178.
- Illmer, P. & Schinner, F. (1995). Solubilization of inorganic calcium phosphates-
solubilization mechanisms. Soil Biology and Biochemistry, 27, 257-63.
- Izko, X. & Burneo, D. (2003). Herramientas para la valoración y gestión forestal
sostenible de los bosques sudamericanos. Unión Mundial para la Naturaleza,
Oficina Regional para Suramérica (UICN-Sur). Imprenta Mariscal. Quito,
Ecuador.
- Karssies, L.E. & Prosser, I.P. (1999). Guidelines for riparian filter trips for
Queensland irrigators. CSIRO Land and Water Technical Report32/99.
Canberra, Australia, Organización de Investigación Científica e Industrial de
Commonwealth (CSIRO).
- Landell-Mills, N. & Porras, T. I. (2002). “Silver bullet or fools’ gold? A global
review of markets for forest environmental services and their impact on the poor”.
Instruments for sustainable private sector forestry series. International Institute
for Environment and Development, London.
- Li, J.H.; Wang, E.T.; Chen, W.F. & Chen, W.X. (2008). Genetic diversity and
potential promotion of plant growth detected in nodule endophytic bacteria of
soybean grown in Heilongjiang province of China. Soil Biol Biochem, 40, 238 –
246.

- Livingston, D. (1963). Chemical composition of rivers and lakes. Professional
paper 440G. U.S. Geol. Surv., Washington, DC.
- Llagas, W.A. & Gómez E.G. (2006). Diseño de humedales artificiales para el
tratamiento de aguas residuales en la UNMSM. Revista del Instituto de
Investigaciones FIGMMG, 15, 85-96.
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E. R. B.; Taghavi, S.;
Mergeay, M. & van der Lelie, D. (2002). Endophytic bacteria and their potential
applications. Critical Reviews in Plant Sciences, 21, 583-606.
- Lot, A. & Novelo, A. (1978). Laguna de Tecocomulco, Hgo. Guías botánicas de
excursiones de México. Sociedad Botánica de México A.C. México, D.F.
- Lot, A. (2004a). Flora y vegetación de los humedales de agua dulce en la zona
costera del Golfo de México. In: Caso, M., I. Pisanty y E. Ezcurra (comps.).
Diagnóstico ambiental del Golfo de México. Instituto Nacional de Ecología-
Secretaría de Medio Ambiente y Recursos Naturales. México, D.F., México. 521-
553.
- Lot, A. (2004b). Fanerógamas acuáticas. In: García-Mendoza, A. J., M. J.
Ordóñez y M. Briones-Salas (eds.). Biodiversidad de Oaxaca. Instituto de
Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la
Conservación de la Naturaleza-World Wildlife Found. México, D.F. 237-248.
- Madigan, M.T.; Martinko, J.M. & Parker, J. (2003). Brock Biología de los
microorganismos. 10a ed. Prentice Hall. New Jersey. 1011pp.

- Madigan, M.T.; Martinko, J.M. & Parker, J. (2006). Biología de los
Microorganismos. Decima edición. Editoral Pearson Prentice Hall. España. pp.
986.
- Mander, U. & Hayakawa, Y. (2005). Purification processes, ecological functions,
planning and design of buffer zones in agricultural watersheds. Ecological
Engineering, 24, 421–432.
- Marilley, L. & Aragno, M. (1999). Phylogenetic diversity of bacterial communities
differing in degree of proximity of Lolium perenne and Trifolium repens roots.
Applied Soil Ecology, 13, 127–136.
- Martínez, M. & Martínez, A. (1996). Effect of phosphate solubilizing bacteria
during the rooting period of sugar cane (Saccharum offinarum) Venezuela
variety, on the growers oasis sustrate. Trabajo de Grado. Carrera de
bacteriologia. Pontificia Universidad Javeriana. Colombia. pp 83-92.
- Martínez, P. & García, D. (2002). Aislamiento y evaluación preliminar de
bacterias rizosféricas, diazotróficas y solubilizadores de fósforo de las especies
forestales altoandinas Weinmannia tomentosa y Escallonia mytilloides. Revista
Intropica, 5, 63-76.
- Mcclain, M. & Richey, J. (1996). Regional-scale linkages of terrestrial and lotic
ecosystems in the Amazon Basin: A conceptual model for organic matter. Archiv
für Hydrobiologie. 113 (Supplement) 111-125.
- Medina, E. & Gómez, A. (2009). Tratamiento de aguas residuales mediante
irradiación gamma. Quivera, 11(1), 12-21.
- Medina, J.P. (2013). Eliminación de vertimiento de aguas residuales industriales
(ARI) en la (CTO) de (ECP) mediante la instalación de un sistema de tratamiento
e inyección. (Tesis de Especialización). Facultad de ingeniería forestal.

Universidad del Tolima. Ibagué
- Mendoza, M.; Osorio, P. & Benítez, N. (2010). Decontamination of industrial
wastewater from sugarcane crops by combining solar photo-Fenton and
biological treatments. Journal of Hazardous Materials, 177, 851-855.
- Ministerio de Ambiente, Vivienda y Desarrollo Territorial (MAVDT), Ministerio
del Interior y de Justicia: Dirección de Prevención y Atención de Desastres
(DPAD), Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM),
Instituto Colombiano de Geología y Minería (INGEOMINAS), Departamento
Nacional de Planeación (DNP), Dirección de Desarrollo Urbano y Política
Ambiental (DDUPA), Corporación Autónoma Regional del Tolima
(CORTOLIMA). (2009). Estrategias de mitigación del riesgo en la cuenca del río
Combeima para garantizar el abastecimiento de agua en la ciudad de Ibagué,
CONPES 3570. Bogotá, D.C.
- Mitchell, M.; Stapp, W. & Bixby, K. (1991). Manual de campo de Proyecto del
Río: una guía para monitorear la calidad del agua en el Río Bravo. Segunda
edición. Proyecto del Río. New Mexico, USA.
- Moreno, M.; Naranjo, B. & Koch, A. (2010). Evaluación de dos métodos para la
reducción de nitrógeno, fósforo y DQO de aguas residuales, mediante un cultivo
axénico de cianobacterias y un consorcio microbiano, inmovilizados y en
suspensión. Revista Ciencia, 13, 55-61.
- Morgenroth, E.; Kommedal, R. & Harremoes P. (2002). Processes and modeling
of hydrolysis of particulate organic matter in aerobic wastewater treatment – a
review. Water Sci. Technol. 45, 25–40.

- Mota, C.; Ridenoure, J. & Cheng y De los Reyes, F. (2005). High levels of
nitrifying bacteria in intermittently aerated reactors treating high ammonia
wastewater. FEMS Microbiology Ecology, 54, 391-400.
- Naiman, R.; Decamps, H. & Mc clain, M. (2005). Riparia: Ecology, Conservation
and Management of Streamside Communities, Elsevier/Academic Press, San
Diego, CA, USA. 430 p.
- Nannipieri, P.; Giagnoni, L.; Landi, L. & Renella, G. (2011). Role of phosphatase
enzymes in soil. In: Bunemann EK, Oberson A, Frossard E, editors. Phosphorus
in action Soil biology. 26. Berlin: Springer; p. 215-41.
- Novelo, A. & Lot, A. (1989). Importancia de la vegetación acuática en los
ecosistemas naturales. In: Memorias del simposio internacional sobre la ecología
y conservación del delta de los ríos Grijalva y Usumacinta. Instituto Nacional de
Investigaciones sobre Recursos Bióticos. División Regional Tabasco y Gobierno
del Estado de Tabasco. Villahermosa. pp.: 5-14.
- Oh, B.; Choi, W.; Park, S.; Kim Y. & Oh, T. (2004). Biochemical properties and
substrate specificities of alkaline and histidine acid phytase. Applied Microbiology
and Biotechnology, 63, 362-372.
- Olson, H.; Chan, S.; Weaver, G.; Cunningham, P.; Moldenke, A.; Progar,
R.…Peterson, E. (2000). Characterizing stream riparian upslope habitats and
species in Oregon managed headwater forests. International conference on
riparian ecology and management in multi-land use watersheds. U.S. American
Water Resources Association. Parks and Wildlife Commission of the Northern
Territory. Australia. Journal of Biogeography, 27, 843–868.
- Osborne, L. & Kovacic, D. (1993). Riparian vegetated buffer strips in water-
quality restoration and stream management. Freshwater Biology, 29, 243-258.

- Paredes, M. & Espinosa, D. (2010). Ácidos orgánicos producidos por
rizobacterias que solubilizan fosfato: una revisión crítica. Terra Latinoamericana,
28(1), 61-70.
- Peña, H. & Reyes, I. (2007). Aislamiento y evaluación de bacterias fijadoras de
nitrógeno y disolventes de fosfatos en la promoción del crecimiento de la
lechuga (Lactuca sativa l.). Interciencia, 32 (8), 560-565.
- Perez, A.; Rojas, J. & Fuentes, J. (2010). Diversidad de bacterias endófitas
asociadas a raíces del pasto Colosuana (Bothriochloa pertusa) en tres
localidades del Departamento de Sucre, Colombia. Acta Biológica Colombiana,
15 (2), 219-228.
- Persello-Cartieaux, F.; Nussaume, L. & Robaglia, C. (2003). Tales from the
underground: molecular plant-rhizobacteria interactions. Plant, Cell and
Environment, 26 (2), 189-199
- Peterson, R.C.; Petersen, L.B.M. & Lacuorsiere, J. (1992). “A Buil-ding-block
Model for Stream Restoration”, en River Conservation and Management, P.J.
Boon, P. Calow y G.E. Petts (eds.). 293-310, Jhon Wiley & sons, Chichester.
- Porteous-Moore, F.; Barac, T.; Borremans, B.; Oeyen, L.; Vangronsveld, J.; Van
Der Lelie, D… Moore E.R.B. (2006). Endophytic bacterial diversity in poplar trees
growing on a BTEX-contaminated site: the characterisation of isolates with
potential to enhance phytoremediation. Systematic and Applied Microbiology, 29,
539- 556.
- Prato, T. & Shi, H. (1990). A comparison of erosion and water pollution control
strategies for an agricultural watershed. Water Resources Research, 26, 199-
205.

- Puente, M.; LI, C. & Bashan, Y. (2009). Rock-degrading endophytic bacteria in
cacti. Environmental and Experimental Botany, 66, 389-401.
- Ramachandran, S.; Fontanille, P.; Pandey, A. & Larroche, C. (2006). Gluconic
Acid: properties, applications and microbial production. Food Technology and
Biotechnology, 44(2), 185-95.
- Reardon, K. (1994). An immobilized-cell fluidized-bed bioreactor for 2,4-
dinitroluene biodegradation. Air Force Office of Scientific Research Report.
- Reardon, R.D. (1994). Cost Implication of Design for Nutrient Removal. En R.D.
Reardon. Water Environment Federation 67th Annual Conference and Exposition
(pp. 551-562) Chicago, IL
- Redfield, A. (1958). The biological control of chemical factors in the environment.
Am. Sci. 46: 206-221.
- Richardson, A.E. & Simpson, R.J. (2011). Soil Microorganisms Mediating
Phosphorus Availability. Plant Physiology. July 156:989-96.
- Rivieros, J.A. (2008). Evaluación de microorganismos con potencial en
biofertilizacion para un cultivo de uchuva (Physalis peruviana L.) en etapas de
semillero y vivero. Universidad Nacional de Colombia. Facultad de Agronomía.
Página 11 – 12.
- Robertson, N. & Wunder, S. (2005). Fresh tracks in the forest: Assessing
incipient payments for environmental services initiatives in Bolivia (draft). In
CIFOR. Bogor.
- Robins, J.D. & Cain, J.R. (2002). The past and present condition of the Marsh
Creek watershed. Berkeley, CA: Natural Heritage Institute. 71 p.

- Rodriguez, D. & Urrego, L. (2003). Evaluacion preliminary de dos matrices para
inmovilizacion de bacterias diazotroficas y solubilizadoras de fósforo aisladas
del bosque alto andino Cundinamarqués. Trabajo de Grado. Carrera de
Microbiologia Pontificia Universidad Javeriana. Bogota. 165 p.
- Rodríguez, D.; Ramírez, O. & Peñuela, G. (2011). Behavior of nitrifying and
denitrifying bacteria in a sequencing batch reactor for the removal of ammoniacal
nitrogen and organic matter. Desalination. 273 447–452.
- Rodríguez, H. & Fraga, R. (1999). Phosphate solubilizing bacteria and their role
in plant growth promotion. Biotechnology Advances. 17: 319-39.
- Rodríguez, H.; Fraga, R.; Gonzalez, T. & Bashan, Y. (2006). Genetics of
phosphate solubilization and its potential applications for improving plant growth-
promoting bacteria. Plant and Soil. 287: 15-21.
- Roldan, (1992). Fundamentos de Limnologia Neotropical. Editorial Universidad
de Antioquia, Medellin.
- Rosales, J. (2000). An ecohydrological approach for riparian forest biodiversity
conservation in large tropical rivers. Ph.D. Thesis. Universidad of Birmingham,
Birmingham, UK.
- Rosenblueth, M. & Martínez-Romero, E. (2006). Bacterial endophytes and their
Interactions with hosts. Ame Phylopathol Soci, 19, 827- 837.
- Rozic, M.; Cerjan-Stefanovic. S.; Kurajica, S.; Vancina, V. & Hodzic, E. (2000)
Ammoniacal nitrogen removal from water by treatment with clays and zeolites,
Water Research, 34(14), 3675-3681.

- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J. & Dowling D.N. (2008).
Bacterial endophytes: recent developments and applications. FEMS Microbiol
Lett. 278 (1), 1-9.
- Salgado, I.; Duran, C.; Cruz, M.; Carballo, M. & Martinez, A. (2012). Bacterias
rizosfericas con potencialidades fisiologicas para eliminar materia organica de
aguas residuales. Rev. Int. Contam. Ambie. 28 (1) 17-26.
- Salibury, F. & Ross, C. (1992). Plant Physiology. USA,California. Wadsworth,
Publishing Co. Belmont
- Schussler, E. & Longstreth, D. (1996). Aerenchyma develops by cell lysis in roots
and cell separation in leaf petioles in Sagittaria lancifolia (Alismataceae). Amer.
J. Bot. 83: 1266-1273.
- Schwab, A.P. (1989). Manganese-phosphate solubility relationships in an acid
soil. Soil Sci Soc Am J. 53: 1654-60.
- Scott, B. (2001). Epichloë endophytes: fungal symbionts of grasses. Current
Opinion in Microbiology, 4(4), 393–398.
- Sculthorpe, C. (1967). The biology of aquatic plants. Edward Arnold Ltd.
Londres. 610 pp.
- Servais, P. & Garnier, J. (1993). Contribution of heterotrophic bacterial
production to the carbon budget of the river Seine (France). Microbiol. Ecol. 25,
19-33.
- Silvério, P. (2006). O Proceso de Regulamentação do Uso de Fósforo em
detergente emPó no Brasil. En: Eutrofizaçãona América do Sul: causas,
consequências e tecnologías para gerenciamento e control. J. Galizia Tundisi, T.
MatsumuraTundisi& C. SidagisGalli (eds.). São Carlos, SP, Brasil. p. 507-520.

- Singh, B.K.; Millard, P.; Whiteley, A.S. & Murell, J.C. (2004). Unravelling
rhizosphere–microbial interactions: opportunities and limitations. Trends in
Microbiology, 12, 386-393.
- Sundara, R. & Sinha, W. (1963). Phosphate dissolving microorganisms in the soil
and rhizosfere. Indian J Agric Sci. 33: 272-278
- Teal, J. & Kanwisher, J. (1966). Gas transport in the marsh grass, Spartina
alterniflora. J. Exp. Bot. 17: 355-361.
- UNESCO (1996). Water Quality Assessments. Londres. Reino Unido.
- U.S. Department of the Army (1991). Buffer strips for riparian zones
management (a literature Review). Department of the Army, Crops of Engineer,
for State of Vermont.
- U.S. EPA (2000). EPA/625/R-99/010. Manual Constructed Wetlands Treatment
of Municipal Wastewaters.
- Valderrama, L.; Del Campo, C.; Rodríguez, C.; De-Bashan, L. & Bashan, Y.
(2002). Treatment of recalcitrant wastewater from ethanol and citric acid
production using the microalga Chlorella vulgaris and the macrophyte Lemna
minuscule. Water Res. 36: 4185 - 4192.
- Van der Lelie, D.; Barac, T. & Taghavi, S. (2005). Response to Newman: new
uses of endophytic bacteria to improve phytoremediation. Trends Biotechnol
23:8-9.
- Vanegas, M. (2002). Estudio complementario del caso Ibagué, Colombia.
Convenio IDRC – OPS/HEP/CEPIS 2000 – 2002.

- Vidal, G.; Carvalho, A.; Mendez, R. & Lema, J. (2000). Influence of the content in
fats and proteins on the anaerobic biodegradability of dairy wastewaters.
Bioresource Technol. 74, 231-239.
- Vincent, J.; Crowder, M. & Averill, B. (1992). Hydrolysis of phosphate
monoesters: a biological problem with multiple chemical solution. Trends.
Biochem Sci 17: pp.105-110.
- Wagner, M. & Loy, A. (2002). Bacterial community composition and function in
sewage treatment systems. Current Opinion in Biotechnology 2002, 13:218–227.
- Watanabe, F. (1965). Test of an ascorbic acid method for determining
phosphorus in water and NaHCO3 extracts from soils. Soil Sci Soc Am J; 29:
677-8.
- Zahir, Z.A.; Arshad, M. & Frankenberger, W.T. (2003). Plant growth promoting
rhizobacteria: applications and perspectives in agriculture. Advances in
Agronomy, 81, 97-168.
- Zhuo, J.F.; Guo, W.D.; Deng, X.; Zhang, Z.Y.; Xu, J. & Huang, L.F., (2010).
Fluorescence excitation–emission matrix spectroscopy of CDOM from Yundang
Lagoon and its indication for organic pollution. Spectrosc. Spec. Anal. 30, 1539–
1544.


















