
Histologia Parte 4 - Tejido Nervioso
25
Autor: Dra. Cecilia Koenig
-
Upload
janno59fifty -
Category
Documents
-
view
36 -
download
5
Transcript of Histologia Parte 4 - Tejido Nervioso

Autor: Dra. Cecilia Koenig

2
Características y funciones básicaso Neuronas
Tipos de neuronas Estructura celular básica del citoplasma de la neurona Estructura y funciones básicas del axón
Transporte axonal Conducción del impulso nervioso Sinapsis
Uniones intercelulares de las neuronaso Células de sostén
Astrocitos Oligodendrocitos Células ependimarias Microglía Células de Schwann Células satélite o capsulares
Estructura básica del SNPo Nervioso Ganglios
Estructura básica del SNCo Redes neuronales en la sustancia gris: cerebeloo Organización tisular propia de sustancia gris y sustancia blanca
Receptores sensorialeso Células de Merkelo Oídoo Fotoreceptores

3
Características y funciones básicas del tejido nervioso
Se origina desde el ectoderma y sus principales componentes son las células, rodeadas de escaso material intercelular. Las células son de dos clases diferentes: neuronas o células nerviosas y neuroglia o células de sostén. (Fig 1).
Es el tejido propio del Sistema Nervioso el cuál, mediante la acción coordinada de redes de células
nerviosas: (Fig 2)
recoge información procedente desde receptores sensoriales procesa esta información, proporcionando un sistema de memoria y genera señales apropiadas hacia las células efectoras.
Las células de sostén rodean a las neuronas y desempeñan funciones de soporte, defensa, nutrición y regulación de la composición del material intercelular
El Sistema Nervioso Central (SNC), se origina desde el epitelio del tubo neural y su tejido nervioso contiene neuronas, células de neuroglia y capilares sanguíneos que forman la barrera hemato-encefalica. (Fig 2)
El Sistema Nervioso Periférico (SNP), que conecta los receptores sensoriales con SNC. y a este con las células efectoras, se desarrolla a partir de la cresta neural y sus células se asocian a otros tejidos del organismo. Sin embargo, es una extensión del tejido nervioso del SNC ya que zonas de las neuronas sensitivas y efectoras y todas las interneuronas se encuentran en el SNC, mientras que los ganglios nerviosos y los nervios periféricos corresponde al tejido nervioso propio del SNP (Fig 3)
Figura 1 Figura 2
Figura 3
4
NeuronasSon las células funcionales del tejido nervioso. Ellas se interconectan formando redes de comunicación que transmiten señales por zonas definidas del sistema nervioso . Los funciones complejas del sistema nervioso son consecuencia de la interacción entre redes de neuronas, y no el resultado de las características específicas de cada neurona individual.
La forma y estructura de cada neurona se relaciona con su función específica, la que puede se:
recibir señales desde receptores sensoriales conducir estas señales como impulsos nerviosos, que consisten en cambios en la
polaridad eléctrica a nivel de su membrana celular transmitir las señales a otras neuronas o a células efectoras
En cada neurona existen cuatro zonas diferentes
el pericarion que es la zona de la célula donde se ubica el núcleo (Fig 1), y desde el cuál nacen dos tipos de prolongaciones (Fig 2)
las dendritas que son numerosas y aumentan el área de superficie celular disponible para recibir información desde los
terminales axónicos de otras neuronas (Fig 3 y 4)
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6

5
El axón que nace único y conduce el impulso nervioso de esa neurona hacia otras células (Figs 5 y 6) ramificándose en su porción terminal (telodendrón)
Uniones celulares especializadas llamadas sinapsis, ubicadas en sitios de vecindad estrecha entre los botones terminales de las ramificaciones del axón y la superficie de otras neuronas (Fig 4 y 5)
El tamaño de las células nerviosas es muy variable pero su cuerpo celular puede llegar a medir hasta 150 um y su axón más de 100 cm
Cada zona de las células nerviosas se localiza de preferencia en zonas especializadas del tejido nervioso.
Los cuerpos celulares , la mayor parte de las dendritas y la arborización terminal de una alta proporción de los axones se ubican en la sustancia gris del SNC (Fig 6) y en los
ganglios del SNP (Fig 7 y 8)
Los axones forman la parte funcional de las fibras nerviosas y se concentran en los haces de la sustancia blanca del SNC; y en los nervios del SNP
Tipos de neuronasSegún el número y la distribución de sus prolongaciones, las neuronas se clasifican en:
bipolares, que además del axón tienen sólo una dendrita; se las encuentra asociadas a receptores en la retina y en la mucosa olfatoria
Figura 7 Figura 8
Figura 1 Figura 2
Figura 3 Figura 4

6
seudo-unipolares, desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente cono un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal; es el caso de las neuronas sensitivas espinales (Fig 1)
multipolares desde las que, además del axón, nacen desde dos a más de mil dendritas lo que les permite recibir terminales axónicos desde múltiples neuronas distintas (Fig 2). La mayoría de las neuronas son de este tipo. Un caso extremo do lo constituye la célula de Purkinje que recibe más de 200.000 terminales nerviosos( Fig 3 y 4)
Estructura celular de la neuronaLas neuronas son células sintetizadoras de proteínas, con un alto gasto de energía metabólica, ya que se caracterizan por:
presentar formas complejas y una gran área de superficie de membrana celular, a nivel de la cuál debe mantener un gradiente electroquímico importante entre el intra y el extracelular
secretar distintos tipos de productos a nivel de sus terminales axónicos
requerir un recambio contante de sus distintos organelos y componentes moleculares ya que su vida suele ser muy larga (hasta los mismos años que el individuo al que pertenecen).
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6

7
Por estas razones:
El núcleo es grande y rico en eucromatina, con el nucléolo prominente. (Fig 1) El ergastoplasma que se dispone en agregados de cisternas paralelas entre las
cuales hay abundantes poliribosomas (Fig 1) Al microscopio de luz se observan como grumos basófilo o cuerpos de Nissl, (fig 2) los que se extienden hacia las ramas gruesas de las dendritas
El aparato de Golgi se dispone en forma perinuclear y da origen a vesículas membranosas, con contenidos diversos, que pueden desplazarse hacia las dendritas o hacia el axón.
Las mitocondrias son abundantes y se encuentran en el citoplasma de toda la neurona.
Los lisosomas son numerosos (fig 3) y originan cuerpos residuales cargados de lipofucsina que se acumulan de preferencia en el citoplasma del soma neuronal (fig 4)
El citoesqueleto aparece, al microscopio de luz, como las neurofibrilla (fig 5), que corresponden a manojos de neurofilamentos (filamentos intermedios), vecinos a los abundantes microtúbulos (neurptúbulos) (fig 6).
Estos últimos se asocian a proteinas específicas (MAPs: proteínas asociadas a microtúbulos) que determinan que el citoesqueleto de microtúbulos pueda:
definir compartimentos en el citoplasma neuronal: la MAP-2 se asocia a los microtúbulos del pericarion y dendritas
mientras que la proteína tau se asociada a los microtúbulos del axón. dirigir el movimiento de organelos a lo largo de los microtúbulos: la kinesina, se
desplaza hacia el extremo (+), mientras que la dineína, se desplazan hacia elextremo (-) de los microtúbulos
Las dendritas nacen como prolongaciones numerosas y ramificadas desde el cuerpo celular (fig 7):. sin embargo en las neuronas sensitivas espinales se interpone un largo axón entre las dendritas y el pericarion (fig 8). A lo largo de las dendritas existen las espinas dendríticas, pequeñas prolongaciones citoplasmáticas, que son sitios de sinapsis (figs 9 y 10). El citoplasma de las dendritas contiene mitocondrias, vesículas membranosas, microtúbulos y neurofilamentos.
Figura 7 Figura 8
Figura 9 Figura 10

8
El axón es de forma cilíndrica y nace desde el cono axónico que carece de ergastoplasma y ribosomas (fig 11). Elcitoplasma del axón (axoplasma) contiene mitocondrias, vesículas, neurofilamentos y microtúbulos paralelos. Su principal función es la conducción del impulso nervioso Se ramifica extensamente sólo en su región terminal
(telodendrón) la que actúa como la porción efectora de la neurona, ya que así cada terminal axónico puede hacer así sinapsis con varias neuronas (fig 12) o células efectoras. (fig 13)
Estructura y funciones básicas del axónSus principales funciones son:
el transporte de organelos y moléculas, por el axoplama, entre el pericarion y las ramas del telodendrón. Este es necesario para la mantención del axón y de las células asociadas a él, y para permitir la llegada al pericarion de factores reguladores que modulan su comportamiento.
la conducción del impulso nervioso, como el desplazamiento del potencial de acción generado por cambios en la permeabilidad a iones a lo largo de la membrana celular axonal (axolema) de las fibras nerviosas, en que el axón está rodeado por la vaina de células de sostén.
Figura 11 Figura 12
Figura 13

9
Transporte axónicoEl transporte de organelos, enzimas, agregados macromoleculares y metabolitos, es una función de axoplasma en la cuál intervienen directamente los microtúbulos (Fig 1).
Ocurre en dos direcciones:
anterógrado, desde el soma neuronal hacia el telodedrón y
retrógrado desde los botones terminales hacia el soma neuronal
La velocidad del transporte varía entre:
un flujo lento de 0,5 um/min, al cual se desplazan agregados moleculares como las sub-unidades proteicas que forman al citoesqueleto axonal.
el flujo rápido anterógrado al cuál los organelos membranosos se transladan a velocidades de unos 300 um/min. (Figs 2,3 y 4)
el flujo rápido retrógrado en el cuál vesículas membranosas provenientes de los botones terminales, son transportados hacia el pericarion a unos 200 um/min.
Los transportes axonales rápidos están mediados por la interacción molecular entre microtúbulos y los dos moléculas son capaces de desplazarse a lo largo de los microtúbulos.
En el flujo rápido anterógrado es la molécula de kinesina, unida a un receptor en la membrana del organelo transportado que se desplaza, a expensas de ATP, desde el extremo (-) del microtúbulo, ubicado en el pericarion hacia su extremo (+)
En el flujo rápido retrógrado es la molécula de dineína citoplasmática (MAP1C) unida a un receptor en la membrana del organelo transportado la que se desplaza interactuando con la tubulina a expensas de ATP, desde el extremo (+) del microtúbulo, ubicado en el terminal axónico hacia su extremo (-)
Figura 1 Figura 2
Figura 3 Figura 4

10
Conducción del impulso nervioso por el axón
En el SNC los axones están rodeados por la mielina de los oligodendrocitos (fibras nerviosas mielínicas del SNC), mientras que en el SNP pueden estar rodeados, ya sea, por
prolongaciones citoplasmáticas de las células de Schwann (fibras amielínicas)(Fig 1) o por la mielina las células se Schwann (fibras nerviosas mielínicas del SNP) (Fig 2)
Los impulsos nerviosos son ondas transitorias de inversión del voltaje que existe a nivel de la membrana celular, que se inician el sitio en que se aplica el estímulo. Cada una de estas ondas corresponde a un potencial de acción,
Este proceso es posible porque entre las macromoléculas que, como proteínas integrales, ocupan todo el espesor del axolema se encuentran.
la bomba de sodio-potasio, capaz de transportar activamente sodio hacia el extracelular intercambiandolo porpotasio.
canales para Na sensibles a voltaje, que determinan en la inversión del voltaje de la membrana ya que al abrirse y permitir la entrada de Na+ hacen que el interior de la membrana se vuelva positiva,
canales para K sensibles a voltaje, cuya activación contribuye al retorno a la polaridad inicial, por salida de iones K desde el interior del axoplasma.
En las fibras nerviosas amielínicas el impulso se conduce, como una onda continua de inversión de voltaje hasta los botones terminales de los axones en la forma indicada en el párrafo anterior. La velocidad que es proporcional al diámetro del axón y varía entre 1 a 100 m/s.
En las fibras nerviosas mielínicas, el axón está cubierto por una vaina de mielina formada por la aposición de
una serie de capas de membrana celular, que actúa como un aislante eléctrico del axón.
A lo largo del axón, la mielina es formadas por células sucesivas y en cada límite intercelular existe un anillo sin mielina que corresponde al nodo de Ranvier (Fig 3).
Es en este sitio donde puede ocurrir flujo de iones a través de la membrana axonal. A nivel de los nodos de Ranvier el axolema tiene una alta concentración de los canales de Na sensibles a voltaje, en.. La consecuencia es una conducción saltatoria del potencial de acción ya que la inversión del voltaje inducido a nivel de un nódulo de Ranvier se
Figura 1 Figura 2
Figura 3

11
continúa por propagación pasiva rápida de la corriente por el interior del axón y por el extracelular hasta el nódulo siguiente donde produce la inversión del voltaje. La consecuencia de esta estructura es que en los axones mielínicos la conducción del impulso nervioso es más rápida. La velocidad de conducción del impulso nervioso es proporcional al diámetro del axón y a la distancia entre los nodos de Ranvier.
SinapsisConducen el impulso nervioso sólo en una dirección. Desde el terminal pre-sináptico se envián señales que deben ser captadas por el terminal post-sináptico.
Existen dos tipos de sinapsis, eléctricas y químicas que difieren en su estructura y en la forma en que transmiten el impulso nervioso.
Sinapsis eléctricas: corresponden a uniones de comunicación entre las membranas plasmáticas de los terminales presináptico y postsinápticos . las que al adoptar la configuración abierta permiten el libre flujo de iones desde el citoplasma del terminal presinático hacia el citoplasma del terminal postsináptico..
Sinapsis química: se caracterizan porque las membranas de los terminales presináptico y postsináptico están engrosadas y las separada la hendidura sinátpica, espacio intercelular de 20-30 nm de ancho. El terminal presináptico se caracteriza por contener mitocondrias y abundantes vesículas sinápticas, que son organelos revestidos de membrana que contienen neurotransmisores (Fig 1)
Al fusionarse las vesículas sinápticas con la membrana se libera el neurotrasmisor que se une a receptores específicos localizados en la membrana post-sináptica, en la cuál se concentran canales para cationes activados por ligandos
Al llegar el impulso nervioso al terminal presináptico se induce: la apertura de los canales para calcio sensibles a voltaje, el subir el ncalcio intracelular se activa la exocitosis de las vesículas sinápticas que liberan al neurotransmisor hacia la hendidura sináptica. La unión del neurotrasmisor con su receptor induce en la membrana postsinática la apertura de los canales para cationes activados por ligandosdeterminando cambios en la permeabilidad de la membrana que pueden: inducir la depolarización de la membrana postsinática: sinápsis exhitatorias; o hiperpolarizar a la membrana postsináticas: sinapsis inhibitorias.
La sumatoria de los impulsos exitatorios e inhibitorios que llegan por todas las sinapsis que se relacionan con a cada neurona( 1000 a 200.000) determina si se produce o no la descarga del potencial de acción por el axón de esa neurona
Figura 1

12
Uniones intercelulares de las neuronasLas neuronas se asocian entre sí y con otras células en estructuras especializadas que permiten:
inducir la descarga del impulso nervioso de una neurona sensitiva por asociación de la ramificación terminal periférica de su axón con células epiteliales de los receptores sensoriales.
estimular la acción de células efectoras de otros tejidos, gracias a la capacidad del terminal axónico de asociarse a estas células. De hecho a este tipo de uniones se les considera un tipo especial de sinapsis química.
transmitir el impulso nervioso desde una neurona a otra mediante estructuras especializadas, las sinapsis, que se encuentran el sitio de contacto de cada terminal axónico con las dendritas o el pericarion de otra neurona.
Células de sosténEn el tejido nervioso del SNC, por cada neurona hay entre 10 a 50 células de neuroglia (Fig 1), y que a diferencia de las neuronas retienen su capacidad de proliferar
Existen 4 clases de células de neuroglia:
astrocitos (astroglia) oligodendrocitos (oligodendroglia) células ependimarias microglia
En el tejido nervioso del SNP, tanto las neuronas, en los ganglios, como los axones ubicados en las fibras nerviosas, están rodeadas por de células de sostén (Fig. 2).
Se distinguen dos tipos:
células de Schwann células satélites o capsulares
Figura 1 Figura 2

13
AstrocitosTienen formas estrelladas y presentan largas prolongaciones que se extienden hacia las neuronas y hacia los láminas basales que rodean a los capilares sanguíneos (pies terminales) (Fig 1), o que separan al tejido nervioso del conjuntivo laxo de la piamadre, constituyendo la glia limitante
Las prolongaciones de los astrocitos contienen manojos de
filamentos intermedios específicos formados por la proteína ácida fibrilar.
Se han identificado dos tipos de astroglia: astrocitos fibrosos que se asocian de preferencia a las fibras nerviosas de la sustancia blanca (Fig 2). y astrocitos protoplasmáticos que se concentran de preferencia asociados a los pericariones, dendritas, terminaciones axónicas en la sustancia gris (Fig 3)
OligodendrocitosSon más pequeños y con menos prolongaciones que la astroglía (Fig. 1).
Su núcleo es rico en heterocromatina y su citoplasma contiene ergastoplasma, polirribosomas libres, un aparato de Golgi desarrollado y un alto contenido n microtúbulos, tanto en el citoplasma que rodea al núcleo como en sus prolongaciones. Su función más notable es la formación de la mielina, que rodea a los
axones del SNC.
El proceso de mielinización del axón por el oligodendrocito es similar al de la célula de Schwann. Sin embargo una oligodendroglia puede formar mielina en cada una de sus prolongaciones que se adhieren inicialmente a un axón, de modo que internodos mielinizados de varios axones dependen un oligodendrocito.
Figura 1 Figura 2
Figura 3
Figura 1

14
Células ependimariasForman un tipo de epitelio monostratificado que reviste las cavidades internas del SNC que contienen al líquido cefalo raquídeo (ventrículos y conducto del epéndimo).
Se unen entre sí por comlejosde unión similares a los epiteliales pero carecen de zona de oclusión, de modo que el liquido cefaloraquídeo se comunica con los espacios intercelulares existentes entre las células nerviosas y las glías. Presentan además largas prolongaciones en su zona basal que se asocian a las prolongaciones de la astroglia y en su superficie apical presenta microvellocidades y cilios.
MicrogliaSe caracterizan por ser pequeñas, con un denso núcleo alargado y prolongaciones largas y ramificadas.
Contienen lisosomas y cuerpos residuales. Si bien se la clasifica generalmente como célula de la neuroglia ellas presentarnel antígeno común leucocítico y el antígeno de histocompatibilidad clase II, propio de las células presentadoras de antígeno.
Células de SchwannLas células de Schawnn se originan de la cresta neural y acompañan a los axones durante su crecimiento, formando la vaina que cubre a todos los axones del SNP desde su segmento inicial hasta sus terminaciones. Ellas son indispensables para la integridad estructural y funcional del axón.
Fibras nerviosas amielínicas: cuando el axón asociado a la célula de Schwann es de pequeño diámetro se aloja en una concavidad de la superficie de la célula de Schawnn, rodeado por espacio intercelular y conectado hacia el exterior mediante el mesaxón. Varios axones pueden estar alojados de esta forma en la misma célula (Fig 1).
Figura 1 Figura 2
Figura 3

15
Fibras nerviosas mielínicas: Los axones de mayor diámetro inducen el proceso de formación de la mielina por la célula de Schwann (Fig 2) En las fibras mielínicas cada célula de Schawnn rodea a solo un axón y su vaina de mielina se ubicada vecina al axón con el resto de su citoplasma en la zona externa. Por fuera, la célula de Schawnn se asocia mediante su lámina basal que al endoneuro.
El largo de cada célula de Schawnn varía entre 200 -2000 um. Entre las sucesivas células de Schwann existen zonas sin mielina llamadas los nodos de Ranvier. (Fig 3).
La mielina está compuesta por capas de membrana de la célula de Schwann las cuales se disponen así durante el proceso de mielinización , el cual comienza con lainvaginación de un axón superficie de la célula de Schwann, de manera que el axolema se adosa estrechamente a la membrana plasmática de la célula de Schwann por una parte, y las membranas de la
célula de Schwann que se enfrentan en el mesaxón (Fig 4). Se produce luego un crecimiento en espiral del citoplasma de la célula de Schwann que se traduce en un crecimiento del mesaxón en forma tal que se enfrentan las membranas plasmáticas de la célula de Schwann por sus caras extracelulares y por sus caras intracelulares (Fig 5) Al fusionare las caras extracelulares se genera la llamada línea interperiodica (línea densa menor) y al desplazarse el citoplasma y fusionarse las caras intracelulares de las membranas se originan las líneas periódicas ( líneas densas mayores) (Figs 6 y 7)
El citoplasma de la célula de Schwann permanece (Fig 8):
junto al axón; junto a la superficie externa de la célula entre las lamelas internodales de la mielina: en
las cisuras de Scmidt-Lantermann a nivel de los nodos de Ranvier, el citoplasma
en los extremos celulares de cada vuelta de mielina permanece y no ocurre la fusión de las membranas plasmáticas. La lengueta más externa de la célula de Schwann y su lámina basal cubren al axón en esta zona.
Figura 4 Figura 5
Figura 6 Figura 7
Figura 8

16
Células satélites o capsulares
Son células pequeñas localizadas en los ganglios, alrededor del pericaron (Figura 1), las dendritas y terminales axónicos (Figs. 2 y 3).
Están rodeadas por lámina basal y separan a las células nerviosas del estroma fibrocolagenoso presente en lel tejido propio del SNP (Figs. 3 y 4).
Estructura básica del SNPEn el SNP las células nerviosas se organizan formando dos tipos de estructuras:
NERVIOSque son haces de fibras nerviosas, mielínicas o amielínicas, mantenidas juntas por tejido conjuntivo organizado en forma específica
(Fig. 1).Los axones de las fibras nerviosas pueden pertenecer a neuronas motoras del SNC, neuronas sensitivas y neuronas pre o post-ganglionares del SNA
Figura 1 Figura 2
Figura 3 Figura 4
Figura 1 Figura 2
Figura 3
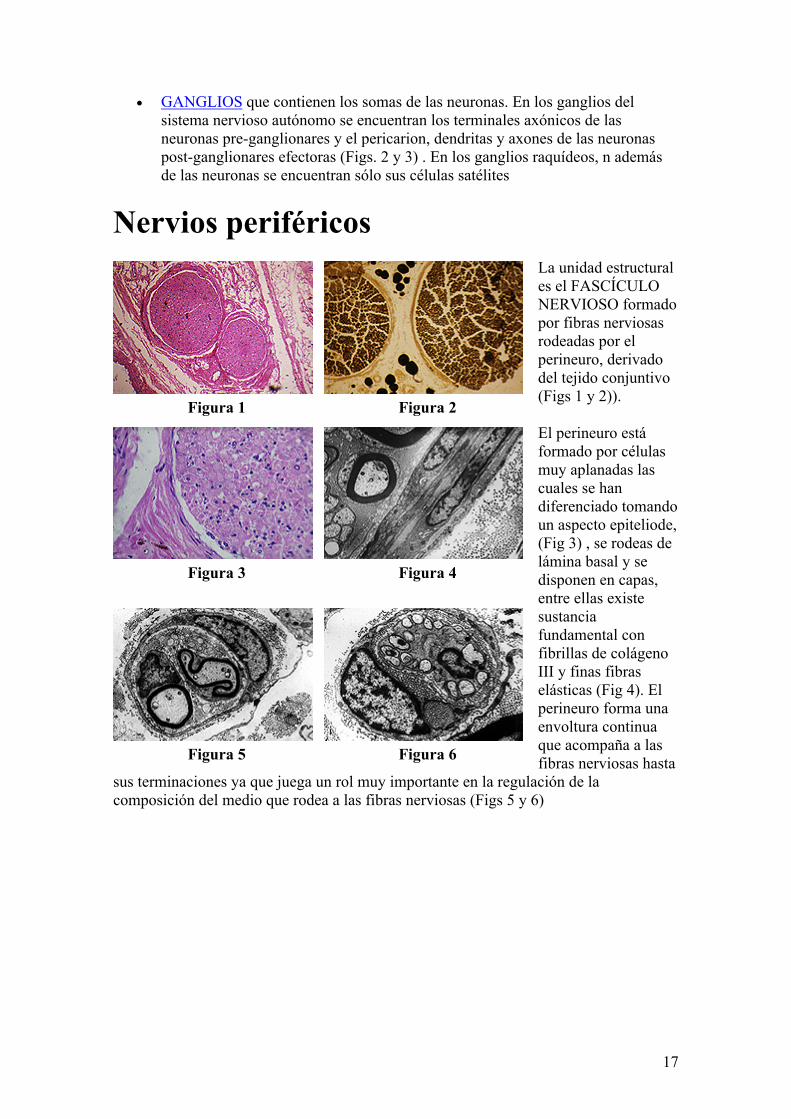
17
GANGLIOS que contienen los somas de las neuronas. En los ganglios del sistema nervioso autónomo se encuentran los terminales axónicos de las neuronas pre-ganglionares y el pericarion, dendritas y axones de las neuronas post-ganglionares efectoras (Figs. 2 y 3) . En los ganglios raquídeos, n además de las neuronas se encuentran sólo sus células satélites
Nervios periféricosLa unidad estructural es el FASCÍCULO NERVIOSO formado por fibras nerviosas rodeadas por el perineuro, derivado del tejido conjuntivo (Figs 1 y 2)).
El perineuro está formado por células muy aplanadas las cuales se han diferenciado tomando un aspecto epiteliode, (Fig 3) , se rodeas de lámina basal y se disponen en capas, entre ellas existe sustancia fundamental con fibrillas de colágeno III y finas fibras elásticas (Fig 4). El perineuro forma una envoltura continua que acompaña a las fibras nerviosas hasta
sus terminaciones ya que juega un rol muy importante en la regulación de la composición del medio que rodea a las fibras nerviosas (Figs 5 y 6)
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6
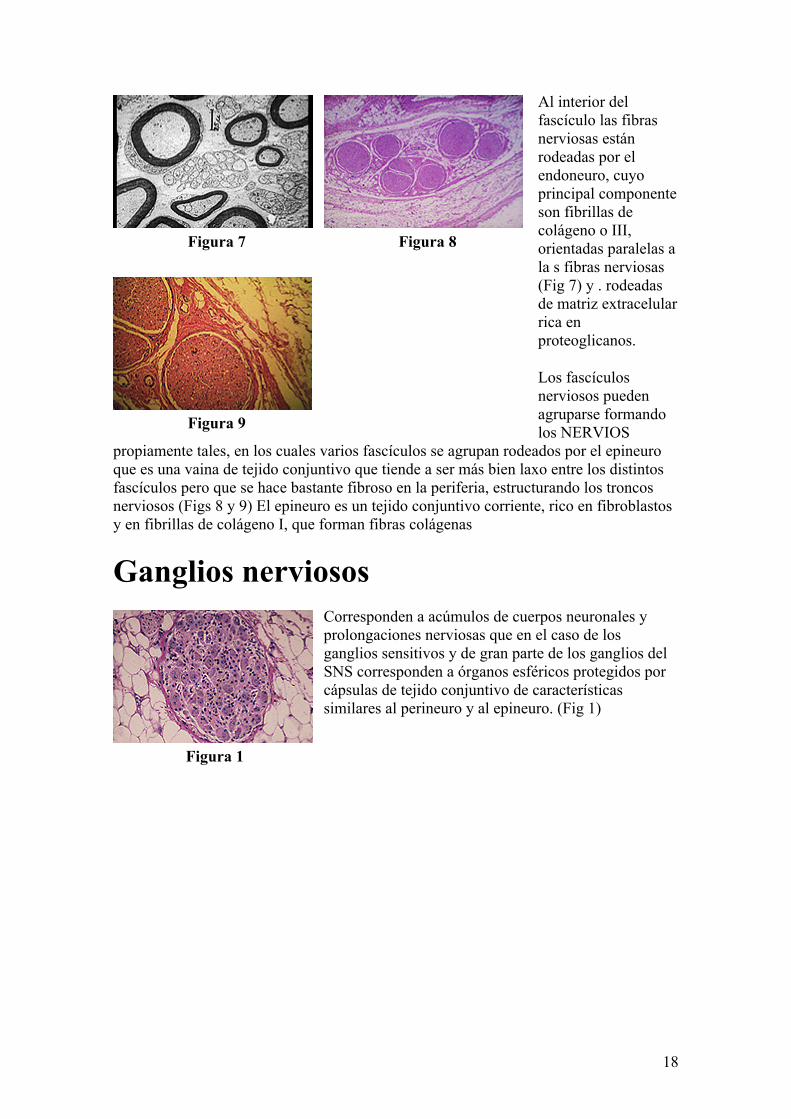
18
Al interior del fascículo las fibras nerviosas están rodeadas por el endoneuro, cuyo principal componente son fibrillas de colágeno o III, orientadas paralelas a la s fibras nerviosas (Fig 7) y . rodeadas de matriz extracelular rica en proteoglicanos.
Los fascículos nerviosos pueden agruparse formando los NERVIOS
propiamente tales, en los cuales varios fascículos se agrupan rodeados por el epineuro que es una vaina de tejido conjuntivo que tiende a ser más bien laxo entre los distintos fascículos pero que se hace bastante fibroso en la periferia, estructurando los troncos nerviosos (Figs 8 y 9) El epineuro es un tejido conjuntivo corriente, rico en fibroblastos y en fibrillas de colágeno I, que forman fibras colágenas
Ganglios nerviososCorresponden a acúmulos de cuerpos neuronales y prolongaciones nerviosas que en el caso de los ganglios sensitivos y de gran parte de los ganglios del SNS corresponden a órganos esféricos protegidos por cápsulas de tejido conjuntivo de características similares al perineuro y al epineuro. (Fig 1)
Figura 7 Figura 8
Figura 9
Figura 1
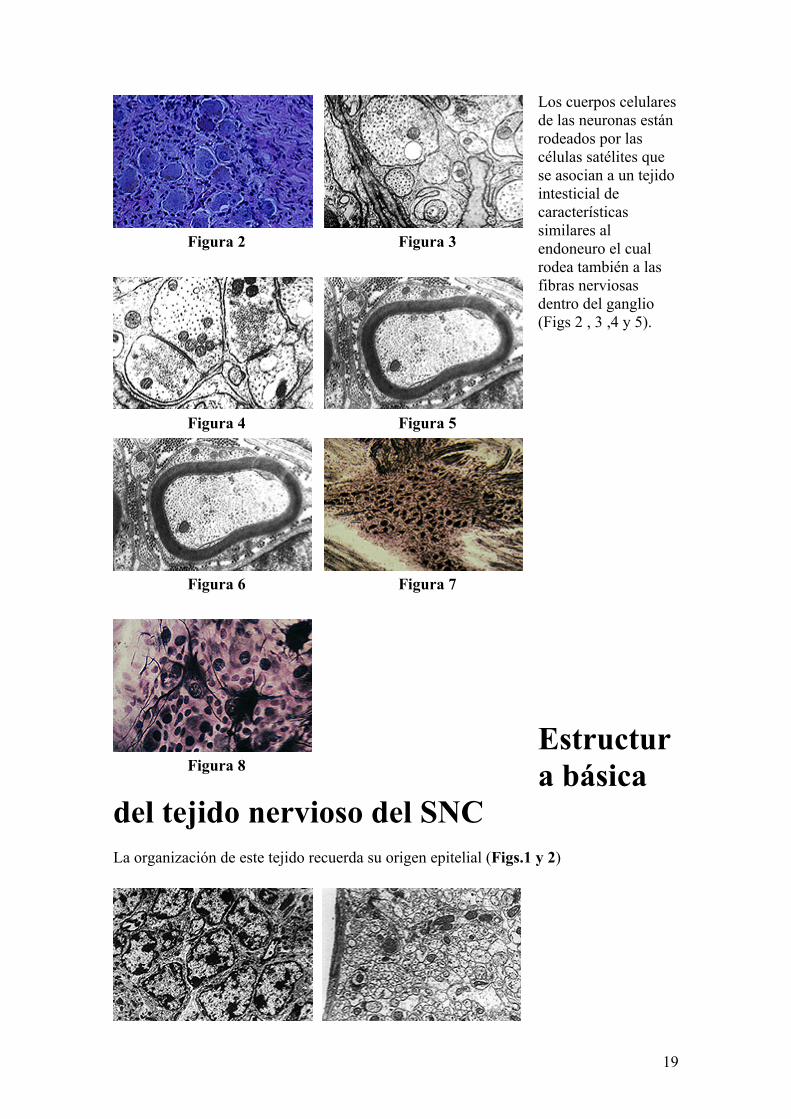
19
Los cuerpos celulares de las neuronas están rodeados por las células satélites que se asocian a un tejido intesticial de características similares al endoneuro el cual rodea también a las fibras nerviosas dentro del ganglio (Figs 2 , 3 ,4 y 5).
Estructura básica
del tejido nervioso del SNCLa organización de este tejido recuerda su origen epitelial (Figs.1 y 2)
Figura 2 Figura 3
Figura 4 Figura 5
Figura 6 Figura 7
Figura 8

20
Figura 1 Figura 2
Se extiende desde los conductos que contienen al líquido cefalo-raquídeo hasta la primera capa de tejido conjuntivo que lo rodea externamente: la píamadre de la cuál está separado por una lámina basal.
El tipo de organización estructural que define las propiedades del SNC son las complejas redes interneuronales que lo forman ubicándose la mayor parte del cuerpo de estas neuronas en la sustancia gris, proyectándose hacia los axones mielínizados hacia la sustancia blanca, para conectar entre sí a neuronas ubicadas en distintas zonas del encéfalo o la médula espinal.
Redes neuronalesComo ejemplo:
Corteza Cerebelosa
La sustancia gris, dispuesta vecina a la superficie externa de las laminillas cerebelosas (Fig.1) está formada por neuronas dispuestas en tres capas diferentes (Fig.2): la capa externa o molecular y la externa o glomerulosa, que contienen neuronas más bien pequeñas.
Figura 1 Figura 2
Entre ambas se ubica la capa de las células de Purkinje, grandes neuronas dispuestas en una sola capa, cuyas dendritas se proyectan hacia la capa molecular y su axón atraviesas la capa granulosa y se dirige hacia la sustancia blanca ubicada al centro de cada laminilla. La citoarquitectura de las neuronas cerebelosas y la posible interacción entre ellas las estableció el histólogo español Ramón y Cajal usando cortes histológicos teñidos con la técnica de Golgi, mediante la cuál en cada corte se tiñen sólo pocas neuronas pero cada una de ellas se tiñe entera. La observación de muchos cortes de cerebelo en cada uno de los cuales se teñían distintos grupos de neuronas le permitió a Ramón y Cajal realizar los esquemas, vigentes hasta hoy de la estructura de la corteza cerebelosa.

21
Células de Purkinje de frente (Fig.3).
Figura 3
Células en canasta (Figs. 4, 5, 6 y 7)
Figura 4 Figura 5
Figura 6 Figura 7
Células estrelladas (Fig. 8)
Figura 8

22
Células de la granulosa (Figs.9 y 10)
Figura 9 Figura 10
Dendritas células de Purkinje tangenciales y fibras paralelas (Figs.11, 12 y 13)
Figura 11
Figura 12 Figura 13
Organización tisular propia del tejido nervioso en sustancias gris y blancaTejido de la sustancia gris
La ultraestructura de las zonas ya descritas de la corteza cerebelosa permite observar su la organización tisular, en cortes de tejido:
Pericariones de células de la granulosa Sinapsis entre fibras musgosas y dendritas de células granulosas (Fig.2) Pericarones de células de Purkinje (Fig.3) Sinapsis entre axón (¿células en canasta?) y pericarones de células de Purkinje

23
Dendritas de células de Purkinje: espinas dendríticas : posible sinapsis con fibras trepadoras o con fibras paralelas
Capa molecular: Sinapsis entre dendritas de células de Purkinje y fibras paralelas: axones de células de la granulosa .
Los perfiles membranosos entre las células nerviosas corresponden a prolongaciones de células de neuroglia, entre las cuales especialmente la astroglia tiene una organización muy precisa.
Como características generales de la sustancia gris destaca:
estrecha vecindad entre las membranas de las células nerviosas y las membranas de las células gliales, quedando siempre un espacio delgado de grosor relativamente constante entre ellas, lleno de material extracelular
disposición de prolongaciones gliales alrededor de las sinapsis químicas, aislándolas del escaso espacio intercelular
concentración de células de astroglia protoplasmática en este tejido dendritas y somas neuronales rodeadas siempre de prolongaciones de
oligodendrocitos y astrocitos y separadas por ellas de otras prolongaciones nerviosas, excepto en los sitios de sinapsis.
capilares sanguíneos rodeados siempre por prolongaciones gliales ausencia de elementos ajenos al tejido derivado del tubo neural, con la única
excepción de las células de microglia y de la irrigación.
Tejido de la sustancia blanca
Corresponden a fibras nerviosas mielinicas en las cuales los axones muestran su vaina de mielina dependiente de los oligodendrocitos y rodeados por los astrocitos fibrosos, en forma tal que cortes a través de tractos de sustancia blanco muestran como principales componentes a axones con su vaina mielínica, a citoplasma y núcleos de oligodendrocitos y astrocitos fibrosos y a capilares sanguíneos.
Receptores sensorialesSon estructuras en las cuales existen células capaces de responder con una gran sensibilidad a señales específicas del entorno, y de transferir la información recibida a terminales nerviosos aferentes al SNC que corresponden a axones de neuronas sensitivas. El proceso que hace que el receptor sensorial responda de un modo útil al estímulo se denomina transducción sensorial.
En los quimioreceptores es la unión de un ligando específica a receptores presentes en la membrana celular de la célula sensorial la que determina un cambio en la permeabilidad a cationes de canales sensibles a ligando presentes en su membrana.
Esto induce a la célula receptora a generar una señal que modifica la permeabilidad a iones de la membrana de los terminales nerviosos adyacentes a ellas generando eventualmente en ellos un potencial de acción que se transmite hacia el SNC.

24
En los mecanoreceptores es la deformación que experimenta la membrana celular de la célula sensorial la que determina la apertura de canales para cationes, generándose una señal que modifica la permeabilidad a iones de la membrana de los terminales nerviosos adyacentes a ellas generando eventualmente en ellos un potencial de acción que se transmite hacia el SNC .
En los fotoreceporess es un cambio inducido por la luz en la estructura de moléculas específicas presentes en membranas de la célula sensorial lo que genera una señal que determina la apertura de canales para cationes en la membrana plasmática de la célula receptora, generándose una señal que modifica la permeabilidad a iones de la membrana de los terminales nerviosos adyacentes a ellas generando eventualmente en ellos un potencial de acción que se transmite hacia el SNC.
Corpúsculos de tacto de Merkel (Células de Merkel) Son células capaces de actuar como receptores sensitivos a la presión. Son células especializadas del epitelios estratificado de la epidermis, concentradas en la palma de la mano y la planta de los pies. Las células de Merkel se ubican entre las células de la capa germinativa y se asocian a las células epiteliales vecinas por medio de desmosomas y su citoplasma se caracteriza por su abundancia en filamentos intermedios de citoqueratina.
Su cara basal se asocia una terminal nerviosa que adopta una forma de disco y que corresponde a la terminal de una fibra aferente, que corresponde al axón de una neurona sensitiva.
El citoplasma de las células de Merkel es capaz de sintetizar y acumular vesículas membranosas que contienen un material denso a los electrones que contienen cromograninas asociadas a moléculas pequeñas parecidas a las catecolaminas .
La célula de Merkel al ser deformada por una compresión la epidermis sería estimulada a liberar sus vesículas que contienen una sustancia capaz de actuar como un neurotransmisor y podría inducir la depolarización del terminal nervioso asociada a ella, la cuál eventualmente generaría la descarga de un potencial de acción en el axón de la neurona sensitiva.
Por sus características de ser capaz de liberar una sustancia química capaz de ejercer su acción al unirse un ligando específico en la superficie de la membrana del terminal nervioso se considera que la célula de Merkel pertenece al sistema APUD (o sistema neuroendocrino difuso).
Organo de Corti (oído)Las células receptoras son las célula ciliadas capaces de traducir el cambios en la posición de sus estereocilios, inducidos desplazamientos de la densa capa de matriz extracelular, la membrana tectoria, en señales eléctricas hacia los termínales sensitivos asociados a ellas.

25
El sitio de transducción de la señal mecánica que determina el cambio en la posición de los estereocilios de modo que estas células epiteliales ricas en estereocilios actúan como mecanorceptores en los cuales la deformación generada por el cambio en la posición de los estereocilios regula el comportamiento de los canales para cationes presentes en la membrana de las células ciliadas y determina la depolarización de la membrana de la célula ciliada la cual se transmite a las terminales nerviosos aferentes al SNC (sensitivas).
Fotorreceptores (retina)Las células fotosensibles de la retina corresponden a células de características algo distintas: los conos y los bastones.
La especialización que les confiere el carácter de actuar como receptores de luz es sin embargo compartida por ellas y corresponde a la existencia del llamado segmento externo de las células.
Esta zona de la célula parece corresponder a un cilio altamente especializado que presenta una serie de laminillas membranosos en su interior, formadas por vesículas membranosas aplanadas que contienen como molécula integral de su membrana a la rodopsina.
Basta la llegada de un fotón a las moléculas de rodopsina para desencadenar una secuencia de reacciones que se traducen finalmente en una activación de los canales a cationes sensibles a ligandos en la membrana de conos o bastones la que determina una señal que induce la liberación de vesículas de mediadores químicos hacia los terminales axónicos sensitivos (axones de neuronas horizontales de la retina) de que hacen contacto con la base de las células sensitivas fotosensibles desencadenando en ellas un potencial de acción.









![Gartner, Leslie P. - Texto Atlas de Histologia, 2da Edición [9 Tejido Nervioso]](https://static.fdocuments.es/doc/165x107/563db835550346aa9a918e35/gartner-leslie-p-texto-atlas-de-histologia-2da-edicion-9-tejido-nervioso.jpg)








