
INST JOSE MARTI REGULACION ENZIMATICA BCM
28
TEMA 5: Otras biomoléculas de importancia biológica. BIOLOGÍA CELULAR Y MOLECULAR UNIDAD NO. 1. INTRODUCCIÓN AL ESTUDIO DE LA BIOLOGÍA CELULAR Y MOLECULAR. BASES MOLECULARES DE LA VIDA. Regulación enzimática. Concepto. Tipos. • Inducción y represión. • Modificación alostérica. • Modificación covalente. • Otros tipos de regulación: proteólisis e isomerización. Cofactores enzimáticos. • Generalidades. • Ejemplos
-
Upload
delgadilloas -
Category
Documents
-
view
2.706 -
download
1
description
INST JOSE MARTI REGULACION ENZIMATICA BCM
Transcript of INST JOSE MARTI REGULACION ENZIMATICA BCM

TEMA 5: Otras biomoléculas de importancia biológica.
BIOLOGÍA CELULAR Y MOLECULARUNIDAD NO. 1. INTRODUCCIÓN AL ESTUDIO DE LA BIOLOGÍA CELULAR Y
MOLECULAR. BASES MOLECULARES DE LA VIDA.
Regulación enzimática. Concepto. Tipos.
• Inducción y represión.
• Modificación alostérica.
• Modificación covalente.
• Otros tipos de regulación: proteólisis e isomerización.
Cofactores enzimáticos.
• Generalidades.
• Ejemplos

REGULACIÓN
Cap. 17: Págs. 299-300
Se da en sistemas o procesos
Capaz de variar su comportamiento en respuesta a
los cambios del entorno
La respuesta puede ser directa o indirecta
La respuesta tiende a modificar el estimulo
volviendo a la situación inicial.
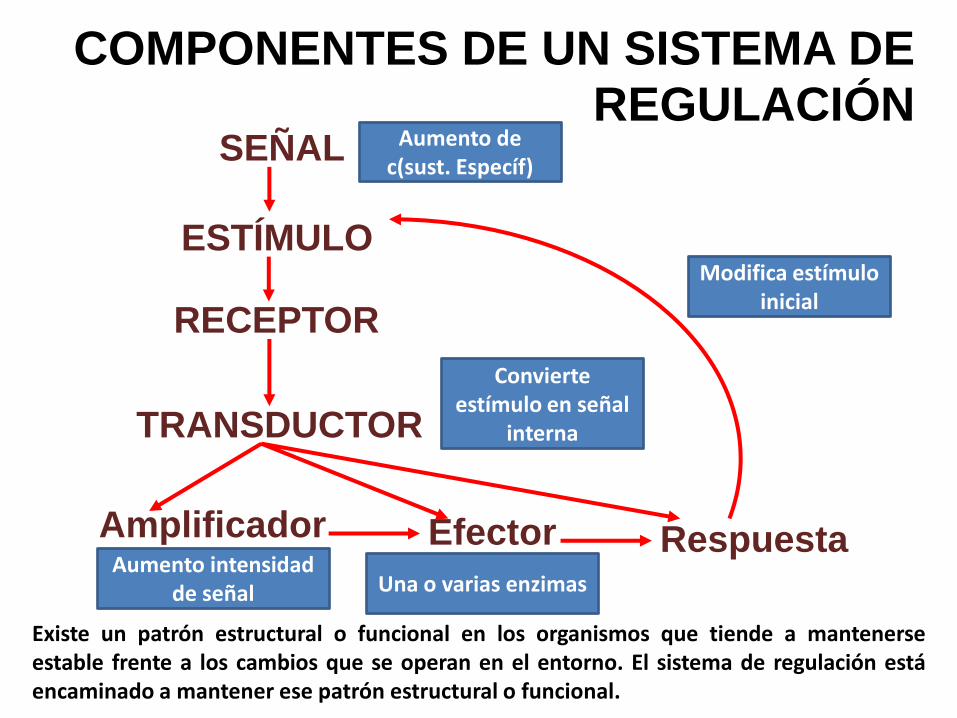
Existe un patrón estructural o funcional en los organismos que tiende a mantenerseestable frente a los cambios que se operan en el entorno. El sistema de regulación estáencaminado a mantener ese patrón estructural o funcional.
COMPONENTES DE UN SISTEMA DE REGULACIÓN
Convierte estímulo en señal
interna
SEÑAL
ESTÍMULO
RECEPTOR
TRANSDUCTOR
Amplificador Efector Respuesta
Aumento de c(sust. Específ)
Aumento intensidad de señal
Modifica estímulo inicial
Una o varias enzimas

MECANISMOS DE REGULACIÓNENZIMÁTICA
• Mecanismos que modifican la cantidad de enzimas: Inducción. Represión.
• Mecanismos que modifican la actividad enzimática: Modificación alostérica. Modificación covalente.

MECANISMOS QUE MODIFICAN LA [ENZIMAS]
• Inducción: la presencia de una sustancia en lacélula puede activar el proceso de síntesis de laenzima y por tanto aumentar su cantidad.
• Represión: el estímulo determina la disminuciónde la síntesis enzimática por lo cual la cantidadde enzima disminuye.
Ambos son mecanismos lentos porque implican síntesis o degradación de
enzimas involucradas y en ellos se da respuesta a una señal química
(metabolito)

MODIFICACIÓN ALOSTÉRICA
Mecanismo por el cual una sustancia denominada
efector alostérico se une a la enzima en un lugar llamado
sitio alostérico, mediante interacciones débiles y provoca
cambios conformacionales, que modifican la velocidad
de la reacción.
MECANISMOS QUE MODIFICAN LA ACTIVIDAD DE LAS ENZIMAS
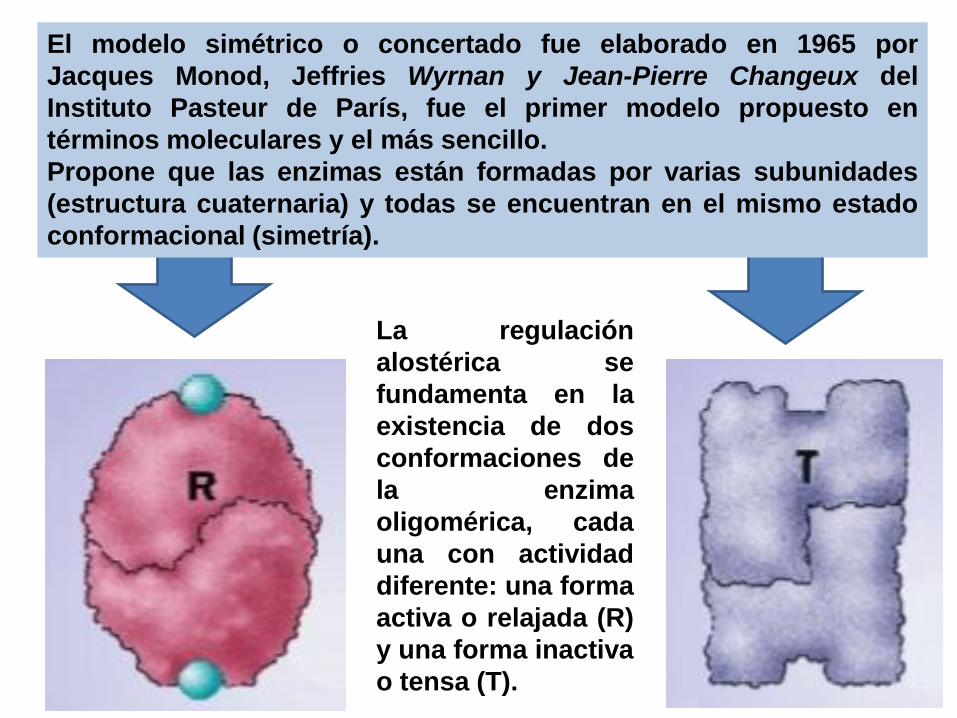
La regulación
alostérica se
fundamenta en la
existencia de dos
conformaciones de
la enzima
oligomérica, cada
una con actividad
diferente: una forma
activa o relajada (R)
y una forma inactiva
o tensa (T).
El modelo simétrico o concertado fue elaborado en 1965 por
Jacques Monod, Jeffries Wyrnan y Jean-Pierre Changeux del
Instituto Pasteur de París, fue el primer modelo propuesto en
términos moleculares y el más sencillo.
Propone que las enzimas están formadas por varias subunidades
(estructura cuaternaria) y todas se encuentran en el mismo estado
conformacional (simetría).
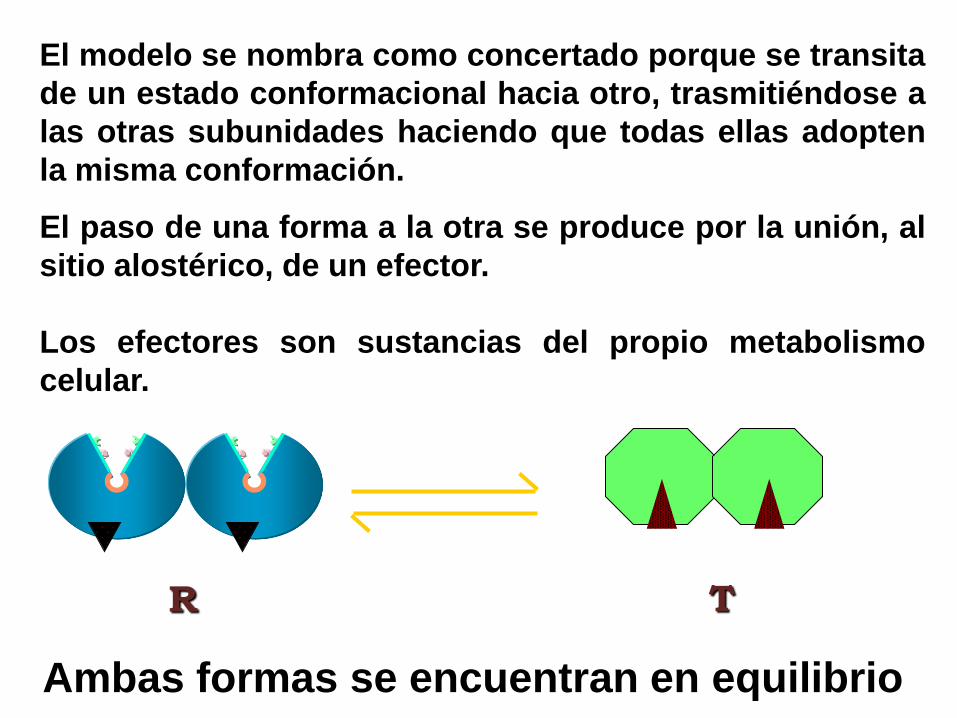
Ambas formas se encuentran en equilibrio
R T
El paso de una forma a la otra se produce por la unión, al
sitio alostérico, de un efector.
Los efectores son sustancias del propio metabolismo
celular.
El modelo se nombra como concertado porque se transita
de un estado conformacional hacia otro, trasmitiéndose a
las otras subunidades haciendo que todas ellas adopten
la misma conformación.
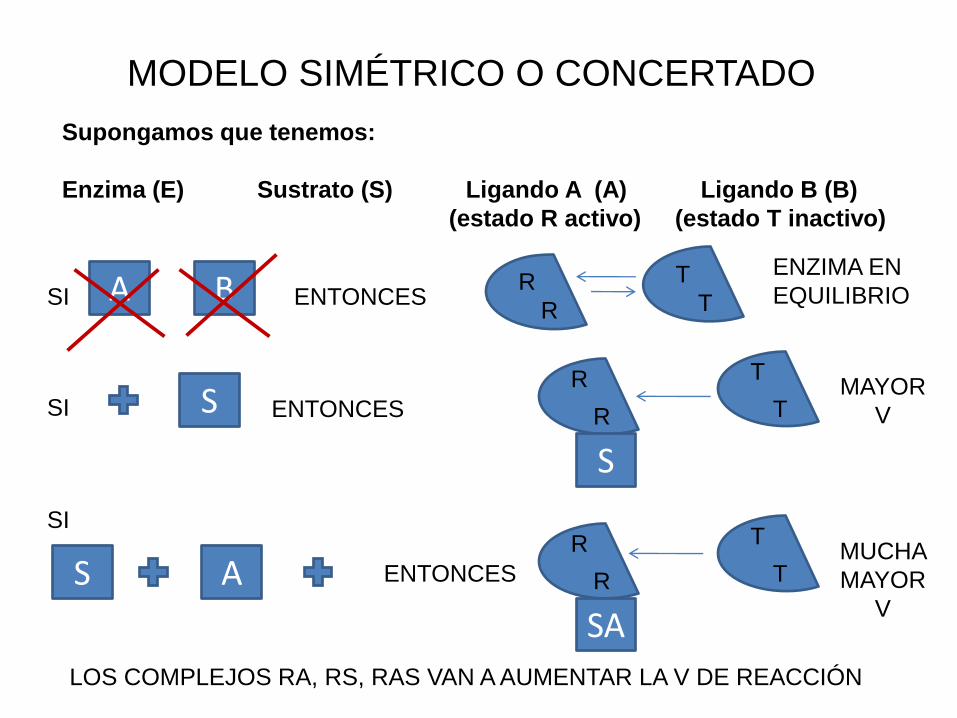
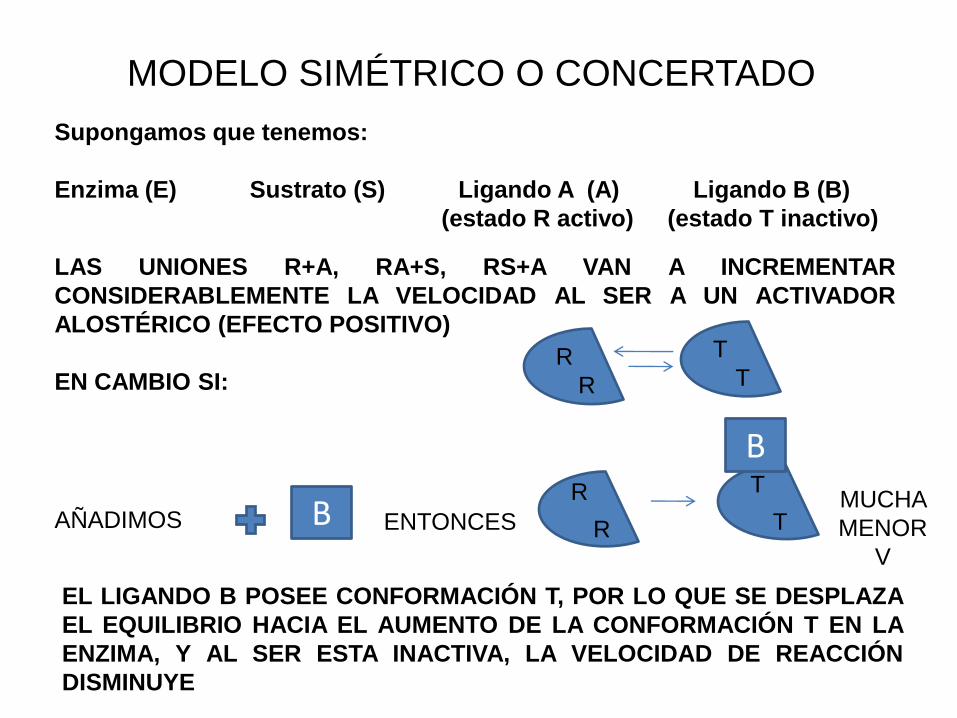
MODELO SIMÉTRICO O CONCERTADO
Supongamos que tenemos:
Enzima (E) Sustrato (S) Ligando A (A) Ligando B (B)
(estado R activo) (estado T inactivo)
A BSI ENTONCESR
R
ENZIMA EN
EQUILIBRIO
SI S ENTONCES
R
R
S
T
TMAYOR
V
SI
A ENTONCES
R
R
SA
T
TMUCHA
MAYOR
V
S
LOS COMPLEJOS RA, RS, RAS VAN A AUMENTAR LA V DE REACCIÓN
T
T
MODELO SIMÉTRICO O CONCERTADO
Supongamos que tenemos:
Enzima (E) Sustrato (S) Ligando A (A) Ligando B (B)
(estado R activo) (estado T inactivo)
B ENTONCES
R
R
T
T
B
MUCHA
MENOR
V
EL LIGANDO B POSEE CONFORMACIÓN T, POR LO QUE SE DESPLAZA
EL EQUILIBRIO HACIA EL AUMENTO DE LA CONFORMACIÓN T EN LA
ENZIMA, Y AL SER ESTA INACTIVA, LA VELOCIDAD DE REACCIÓN
DISMINUYE
LAS UNIONES R+A, RA+S, RS+A VAN A INCREMENTAR
CONSIDERABLEMENTE LA VELOCIDAD AL SER A UN ACTIVADOR
ALOSTÉRICO (EFECTO POSITIVO)
EN CAMBIO SI:R
R
T
T
AÑADIMOS

MODELO SECUENCIAL
Propone que:
• Existen dos conformaciones (R y T) posibles para
cada subunidad de la enzima.
• La unión del sustrato cambia la conformación de la
subunidad a la que se une, pero no altera de forma
apreciable la del resto de las subunidades.
• La transformación que ocurra en esa subunidad
puede determinar si la velocidad va a aumentar o
disminuir porque influye en la afinidad por el sustrato
del resto de las unidades (la incrementa o disminuye).
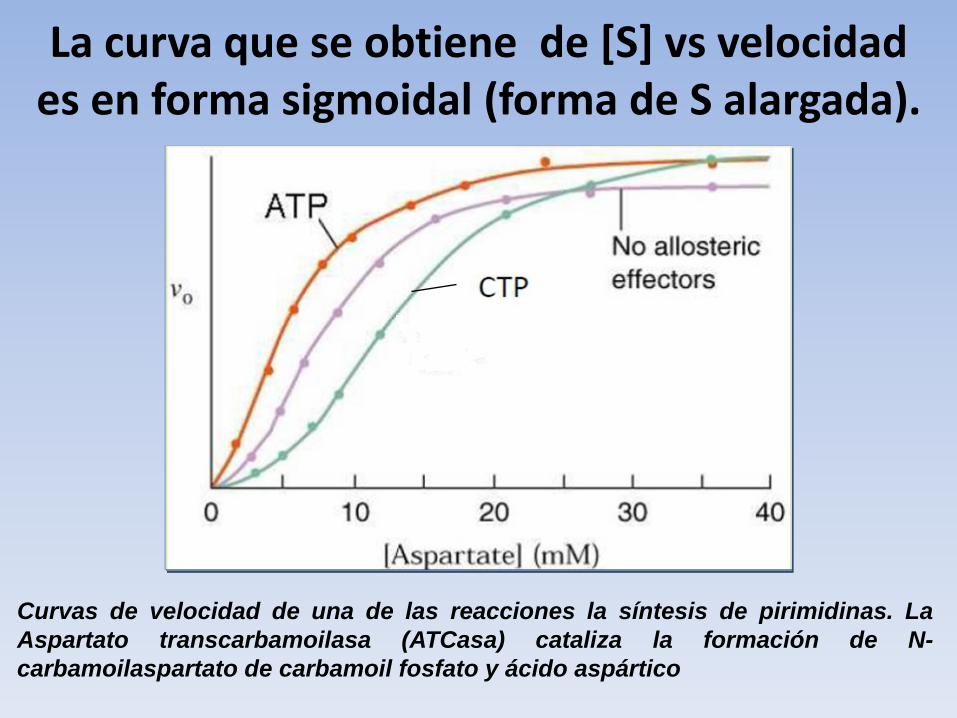
La curva que se obtiene de [S] vs velocidad es en forma sigmoidal (forma de S alargada).
Curvas de velocidad de una de las reacciones la síntesis de pirimidinas. La
Aspartato transcarbamoilasa (ATCasa) cataliza la formación de N-
carbamoilaspartato de carbamoil fosfato y ácido aspártico

CARACTERÍSTICAS DE LAS ENZIMAS ALOSTÉRICAS
• Son proteínas oligoméricas de elevado peso
molecular.
• Existen en varios estados conformacionales
interconvertibles y con afinidad diferente para cada
uno de sus ligandos.
• Los ligandos se unen a la enzima en sitios
específicos por fuerzas no covalentes y de forma
reversible, afectando el estado conformacional de
las enzimas.
• Los cambios conformacionales en una subunidad se
comunican en mayor o menor grado al resto de las
subunidades.

EJEMPLO DE REGULACIÓN ALOSTÉRICA
La pareja que con mayor frecuencia cumple estas funciones es la
formada por el ATP y el ADP, sus concentraciones relativas controlan
un gran número de actividades enzimáticas.
Formación de ATP:
ADP + Pi ATP + H2O
Formación de ADP:
ATP + H2O ADP + Pi
Al aumentar las concentraciones de ATP disminuyen las de ADP y
viceversa.
Ej: En la fosfofructoquinasa el ATP es un efector negativo (inhibidor) y
el ADP es positivo (activador), como las concentraciones de ambos
no pueden aumentar simultáneamente, en cada momento la enzima
sólo estará expuesta a elevadas concentraciones de uno de ellos y así
será su respuesta.

MODIFICACIÓN COVALENTE
Es el mecanismo mediante el cual la unión por enlace
covalente de un grupo químico a la enzima, le provoca
un cambio conformacional que produce una variación
de la velocidad de reacción.
Los tipos mis difundidos de modificación covalente son:
Fosforilación – desfosforilación (P),
Adenilación - desadenilación (AMP grupo adenilato); y
Intercambio de sulfihidrilos (SH) – disulfuros (SS).
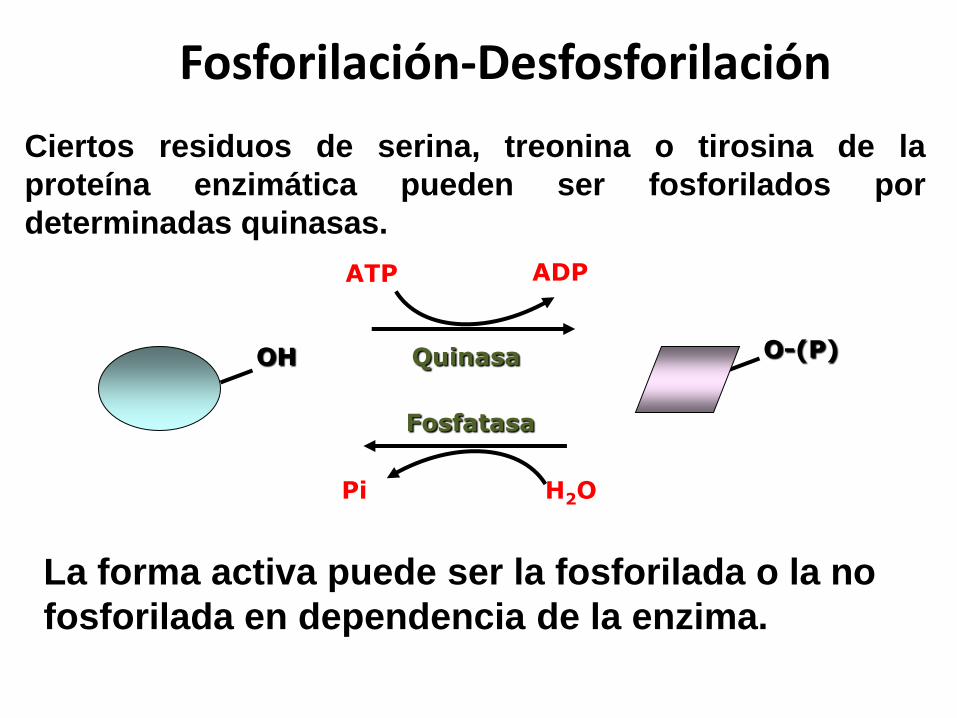
OH
ADPATP
Quinasa
H2OPi
La forma activa puede ser la fosforilada o la no
fosforilada en dependencia de la enzima.
O-(P)
Ciertos residuos de serina, treonina o tirosina de la
proteína enzimática pueden ser fosforilados por
determinadas quinasas.
Fosforilación-Desfosforilación
Fosfatasa
Cap. 17: Págs. 306-309 y 311

CARACTERÍSTICAS DE LA
MODIFICACIÓN COVALENTE
• Se modifica la composición de la
enzima, que conduce a un cambio
conformacional secundario y
modificación de la actividad.
• Menor rapidez que la modificación
alostérica.
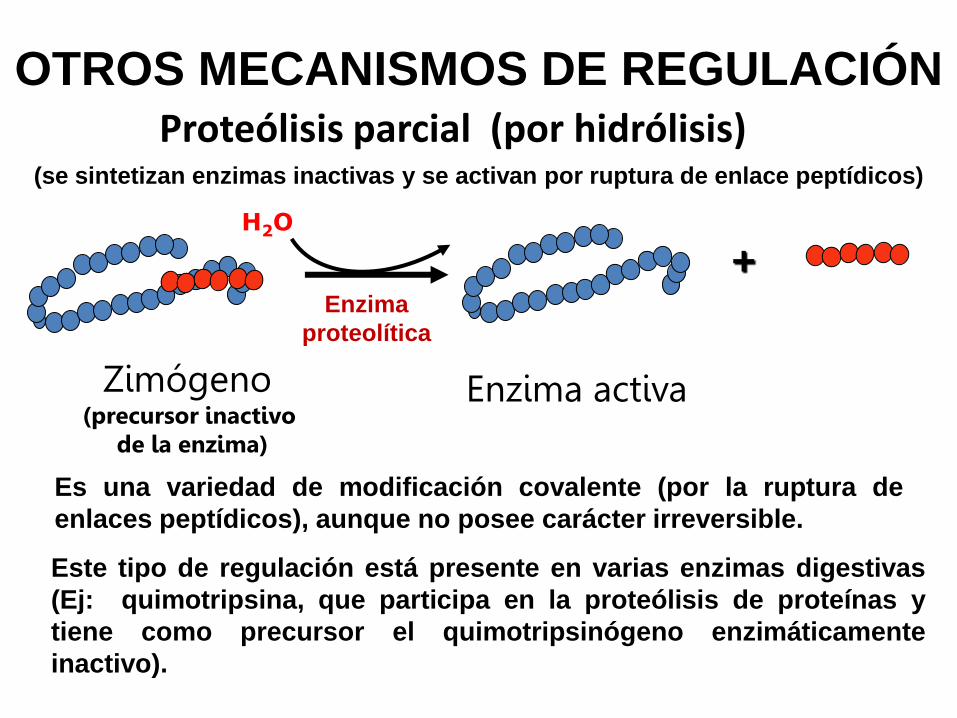
+
Zimógeno (precursor inactivo
de la enzima)
Enzima activa
Este tipo de regulación está presente en varias enzimas digestivas
(Ej: quimotripsina, que participa en la proteólisis de proteínas y
tiene como precursor el quimotripsinógeno enzimáticamente
inactivo).
Proteólisis parcial (por hidrólisis)
OTROS MECANISMOS DE REGULACIÓN
Es una variedad de modificación covalente (por la ruptura de
enlaces peptídicos), aunque no posee carácter irreversible.
H2O
Enzima
proteolítica
(se sintetizan enzimas inactivas y se activan por ruptura de enlace peptídicos)
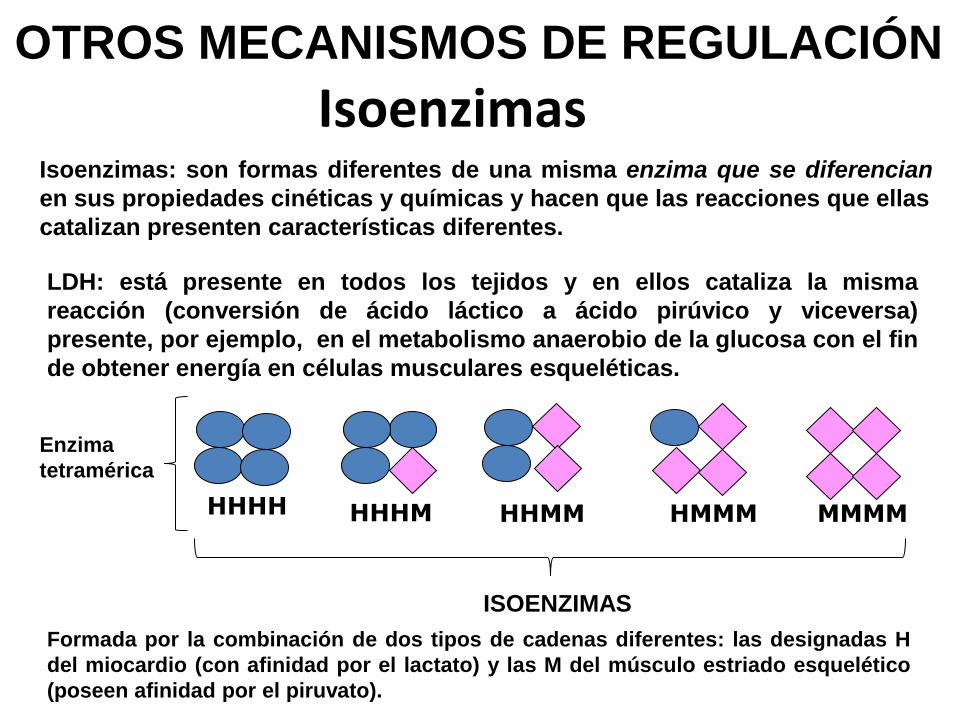
HHHH HHHM HHMM HMMM MMMM
IsoenzimasIsoenzimas: son formas diferentes de una misma enzima que se diferencian
en sus propiedades cinéticas y químicas y hacen que las reacciones que ellas
catalizan presenten características diferentes.
LDH: está presente en todos los tejidos y en ellos cataliza la misma
reacción (conversión de ácido láctico a ácido pirúvico y viceversa)
presente, por ejemplo, en el metabolismo anaerobio de la glucosa con el fin
de obtener energía en células musculares esqueléticas.
Formada por la combinación de dos tipos de cadenas diferentes: las designadas H
del miocardio (con afinidad por el lactato) y las M del músculo estriado esquelético
(poseen afinidad por el piruvato).
Enzima
tetramérica
ISOENZIMAS
OTROS MECANISMOS DE REGULACIÓN
COFACTORES
Sustancias de carácter no proteico y
bajo peso molecular; son moléculas o
iones imprescindibles para la acción
catalítica de muchas enzimas.

• Contribuyen a la unión entre la enzima
y el sustrato.
• Estabilizan la enzima en su
conformación más activa.
• Constituyen frecuentemente parte del
grupo catalítico principal.
• Son transportadores intraenzimáticos
o interenzimáticos en la reacción
catalizada.
FORMAS DE ACTUACIÓN DE LOS
COFACTORES
Cap. 19: Págs. 334-335
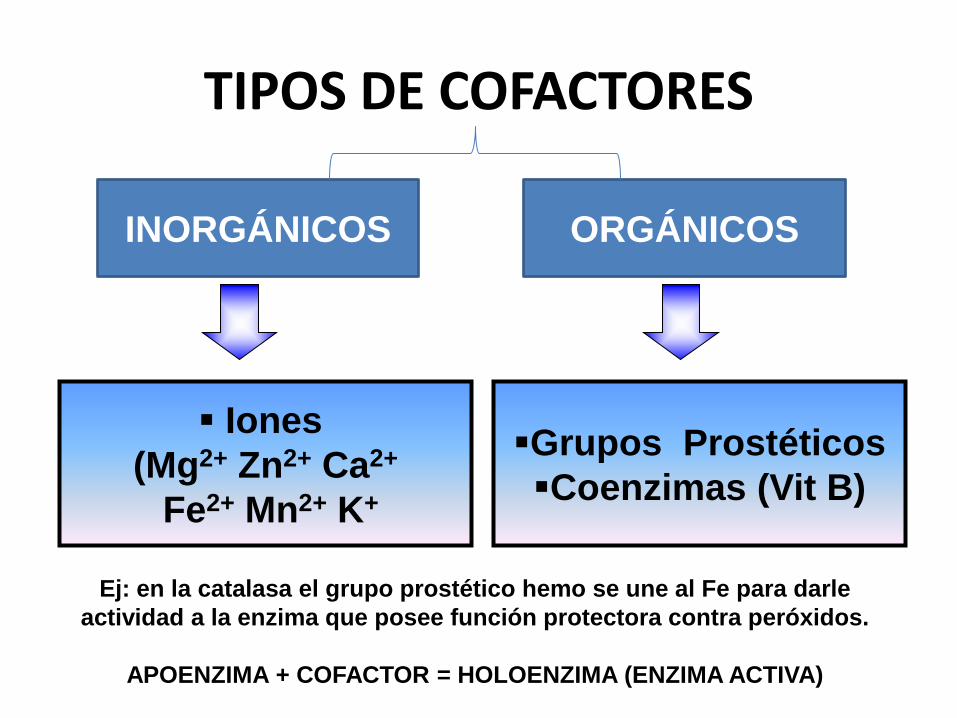
TIPOS DE COFACTORES
Grupos Prostéticos
Coenzimas (Vit B)
INORGÁNICOS ORGÁNICOS
Iones
(Mg2+ Zn2+ Ca2+
Fe2+ Mn2+ K+
Ej: en la catalasa el grupo prostético hemo se une al Fe para darle
actividad a la enzima que posee función protectora contra peróxidos.
APOENZIMA + COFACTOR = HOLOENZIMA (ENZIMA ACTIVA)
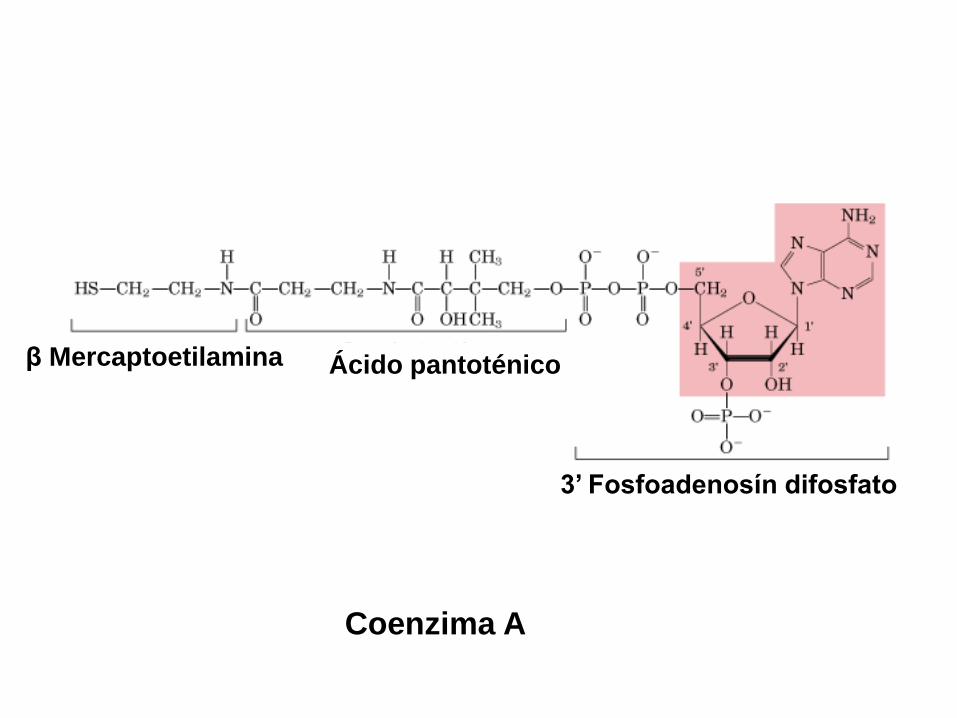
3’ Fosfoadenosín difosfato
Ácido pantoténicoβ Mercaptoetilamina
Coenzima A
Cap. 19: Págs. 343-344
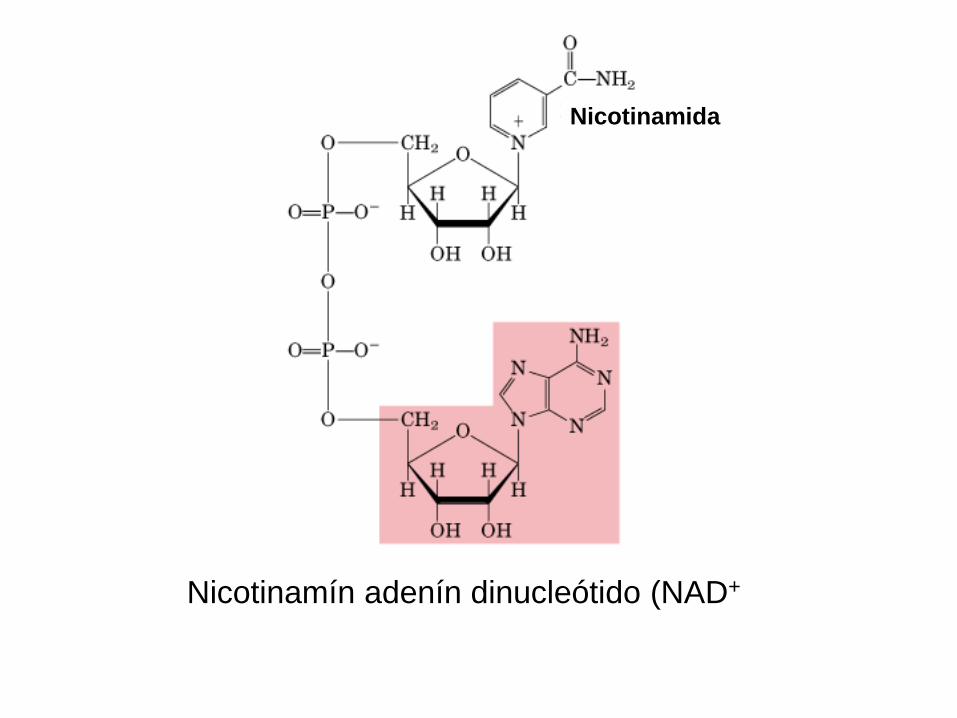
Nicotinamín adenín dinucleótido (NAD+
Nicotinamida
Cap. 19: Págs. 335-337
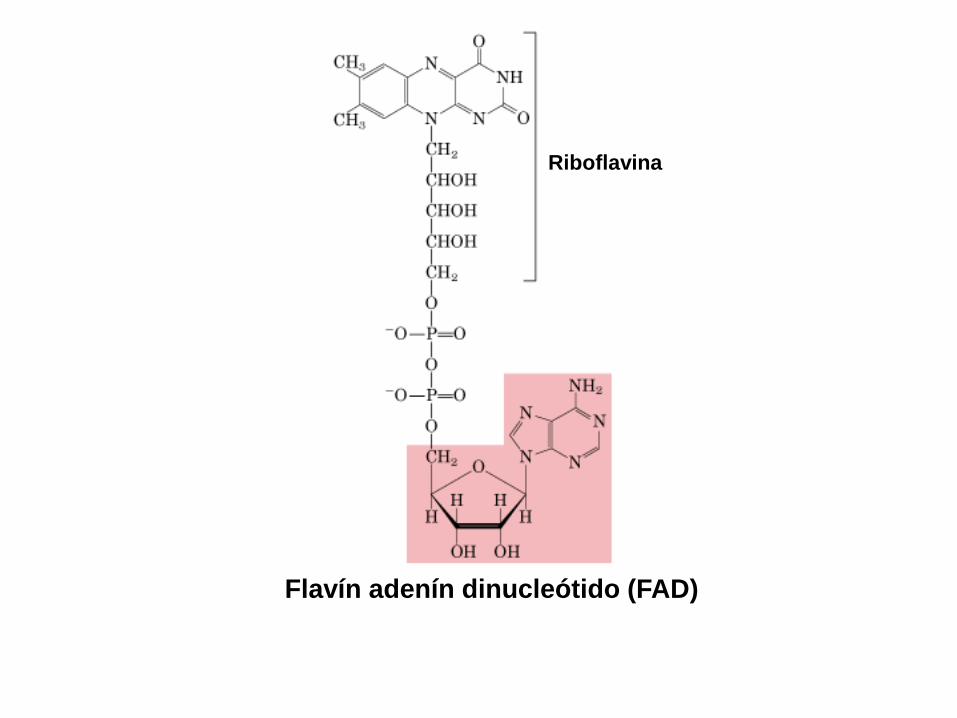
Flavín adenín dinucleótido (FAD)
Riboflavina
Cap. 19: Págs. 337-338
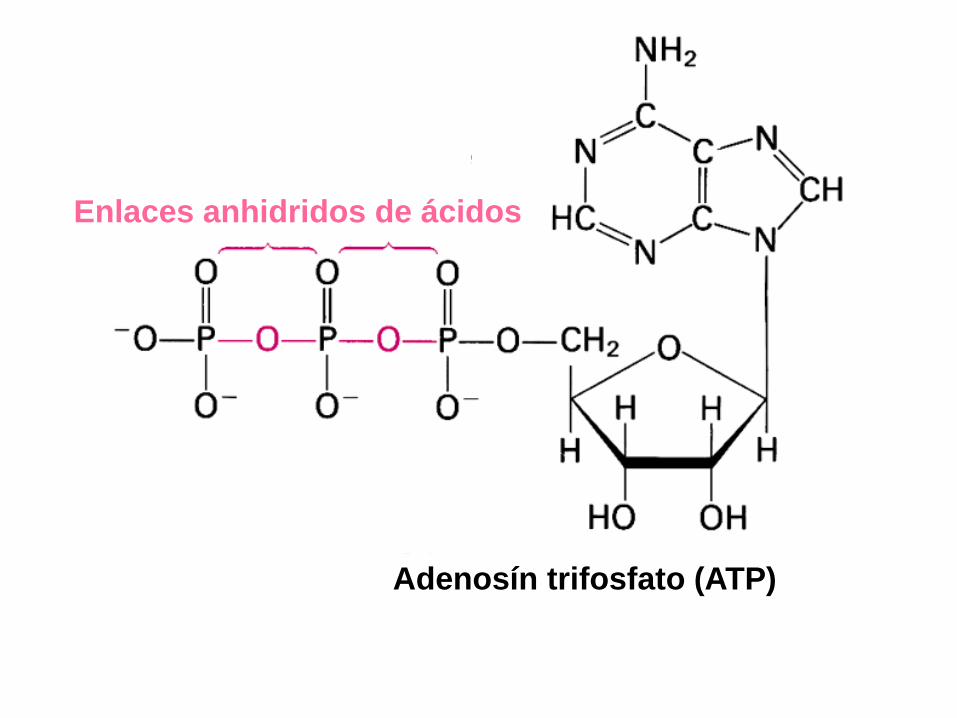
Enlaces anhidridos de ácidos
Adenosín trifosfato (ATP)
Cap. 19: Págs. 347-348

CONCLUSIONES• Las enzimas aceleran la velocidad de las reacciones disminuyendo
la energía de activación y su mecanismo básico de acción consta
de dos etapas, la de unión y la de transformación.
• La estructura tridimensional del centro activo y sus componentes
determinan la especificidad de sustrato y de acción de las
enzimas.
• Existen factores que influyen en la velocidad de la reacción
enzimática, modificando la estructura de la enzima y en particular
de su centro activo, aspecto de gran importancia en la práctica
médica.
• Las formas básicas de regulación enzimática se manifiestan por
variación en la cantidad, ya sea por inducción o represión y por
variación en su actividad, como la regulación alostérica y
covalente.
TAREA EVALUATIVA INDIVIDUAL
Explique porqué se realiza la determinación de
los niveles de enzimas séricas como la LDH
para detectar casos de infarto agudo del
miocardio.
ENTREGAR:
POR ESCRITO
FECHA DE ENTREGA:
VIERNES 12/10/2012
![Enzimatica [PLM]](https://static.fdocuments.es/doc/165x107/5571f22b49795947648c476e/enzimatica-plm.jpg)

















