
Intralmetabolismo1 2010
47
Introducción al Metabolismo y Bioenergética Introducción a las oxidaciones biológicas y al ciclo de Krebs Prof. Vanessa Miguel @bioquitips
-
Upload
vanessa-miguel -
Category
Health & Medicine
-
view
466 -
download
0
Transcript of Intralmetabolismo1 2010

Introducción al Metabolismo y Bioenergética
Introducción a las oxidaciones biológicas y al ciclo de Krebs
Prof. Vanessa Miguel@bioquitips

Metabolismo
Conjunto de todas las reacciones enzimáticas de la célula mediante las cuales adquieren y utilizan la energía que necesitan para realizar sus funciones
Representa la suma de todos los cambios químicos que convierten los nutrientes en energía y productos celulares químicamente complejos
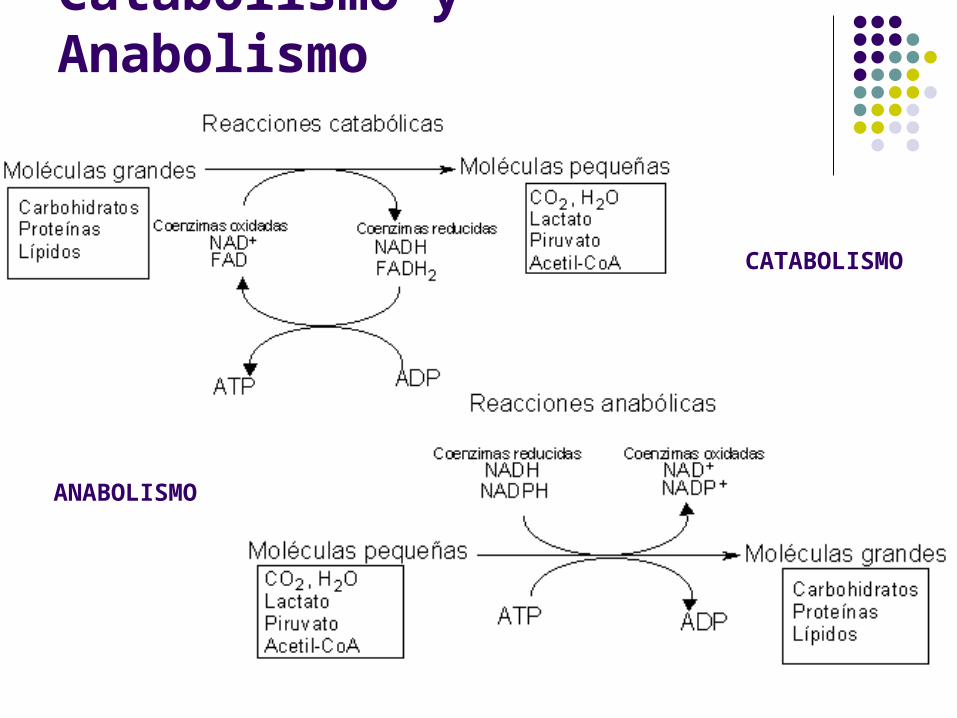
Catabolismo y Anabolismo
CATABOLISMO
ANABOLISMO

Control de la actividad de las enzimas en respuesta a la concentración de metabolitos a. Control por la concentración del producto de una reacciónb. Regulación alostérica
Control por modificación de la enzima (modificación covalente)
Isoenzimas Compartamentalización Complejos multienzimáticos Especialización metabólica Control de la cantidad de enzima
Mecanismos de regulación del metabolismo (enzimas)

Termodinámica Ley Cero (principio del equilibrio
termodinámico). Si dos sistemas A y B están en equilibrio termodinámico, y B está en equilibrio termodinámico con un tercer sistema C, entonces A y C están a su vez en equilibrio termodinámico.
Primera Ley (principio de conservación de la energía): si se realiza trabajo sobre un sistema o bien éste intercambia calor con otro, la energía interna del sistema cambiará.
Segunda Ley: el calor no puede pasar de forma espontánea de una fuente fría a otra más caliente.
Tercera Ley: que es imposible alcanzar una temperatura igual al cero absoluto
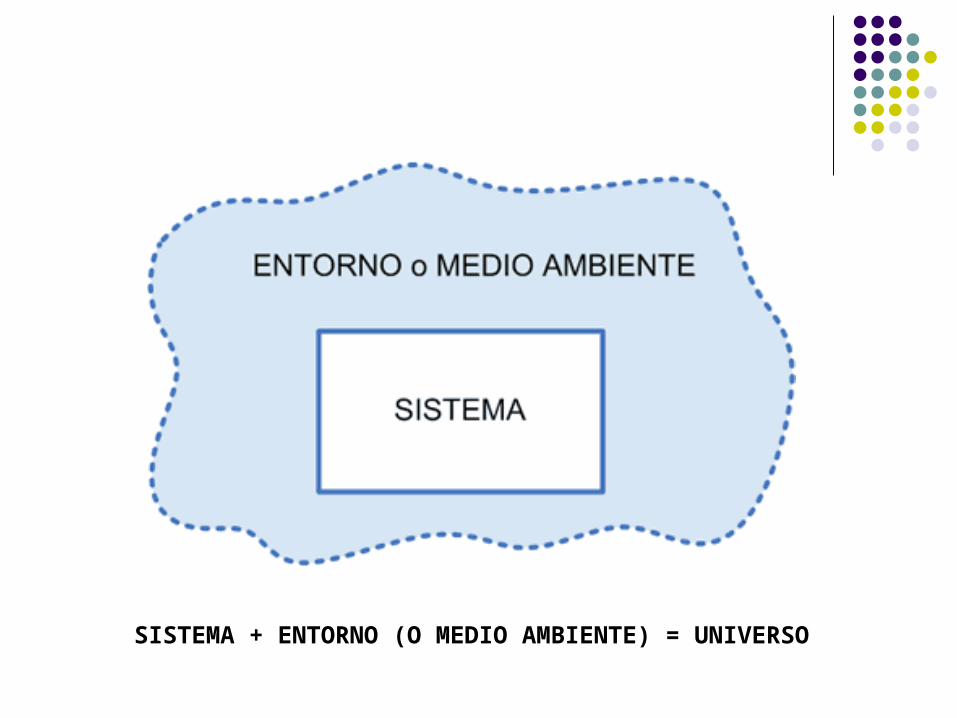
SISTEMA + ENTORNO (O MEDIO AMBIENTE) = UNIVERSO
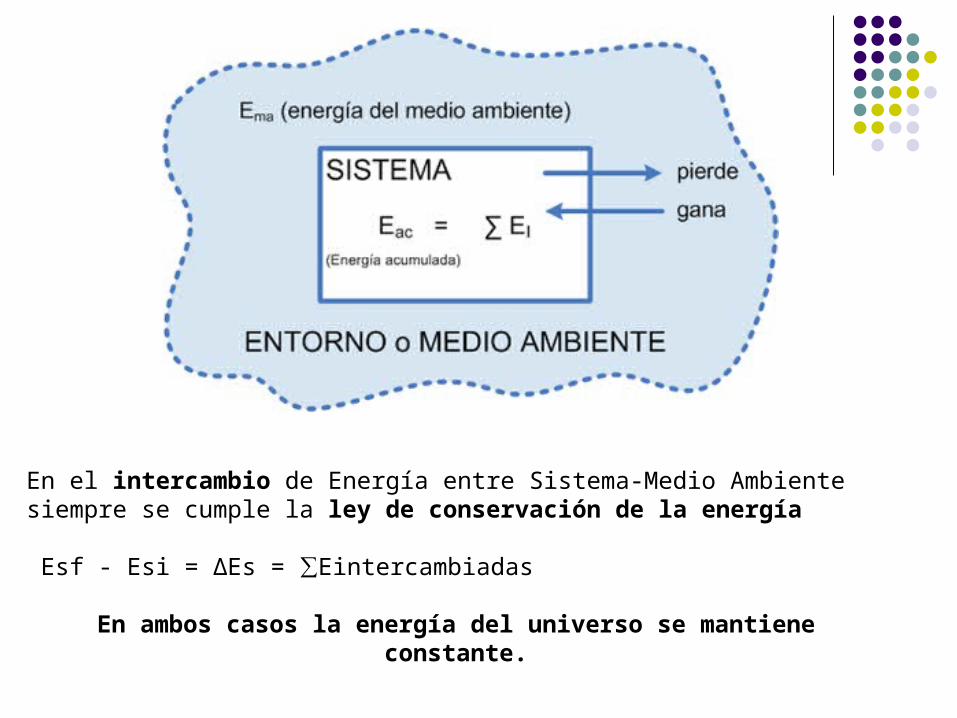
En el intercambio de Energía entre Sistema-Medio Ambiente siempre se cumple la ley de conservación de la energía
Esf - Esi = ∆Es = ∑Eintercambiadas
En ambos casos la energía del universo se mantiene constante.

Tipos de Energía (ej.) 1 mol de sustancia pura en ciertas condiciones de P y T.
Energías intramoleculares Nuclear Electrónica Traslacional Vibracional Rotacional(3, 4, y 5 son las Energías Térmicas)Energías intermoleculares Fuerzas de atracción interiónicas (sales) Fuerzas de atracción Ión-dipolo Unión por puente de hidrógeno Fuerzas de atracción dipolo-dipolo Fuerzas de atracción entre iones o dipolos-moléculas polarizables Fuerzas entre dipolos instantáneos-dipolos inducidos Fuerzas hidrofóbicas Fuerzas de repulsión

Antes que seguir ... ¿qué es entropía?

Si fueramos ingenieros..

Entropía fácil…

El flujo de energía es la esencia de la vida, se comprende mejor a una célula como un complejo sistema de transformación de energía

Espontaneidad de las reacciones
Las reacciones químicas que tienen lugar espontáneamente suelen ser exotérmicas, es decir, con desprendimiento de calor.
El calor que se desprende o absorbe en una reacción a presión constante recibe el nombre de Entalpía ( H). Las reacciones en las que se desprende calor son exotérmicas: H < 0
Sin embargo, hay reacciones endotérmicas que cursan espontáneamente, H > 0; por ejemplo, la disolución de sulfato amónico en agua.

Espontaneidad de las reacciones
La disolución de sulfato amónico en agua es un proceso por el cual se pasa de un estado cristalino altamente ordenado, a otro de mucho mayor desorden molecular.
La función termodinámica que mide el desorden de un sistema recibe el nombre de Entropía ( S). Puede observarse que muchas reacciones que cursan espontáneamente lo hacen con incremento positivo de entropía: S > 0.
Sin embargo, hay procesos espontáneos que cursan con disminución de entropía. Ej: la solidificación del agua a 0 ºC.

Gibbs fue el primero en proponer que un sistema químico busca, no solamente el estado de mínima energía o entalpía (H), sino también el estado de máximo desorden o entropía (S).
Definió una nueva función de estado, la energía libre, G, que relaciona la entalpía y la entropía: G = H - TS
Pueden cursar espontáneamente aquellos procesos en los que se desprende energía libre ( G < 0); pero no aquellos procesos en los que se absorbe energía libre ( G > 0)
Energía Libre de Gibbs

Situaciones:
Si H < 0 (exotérmico) y S > 0 (aumenta el "desorden") para cualquier temperatura, el G < 0 siempre.
Cuando H < 0 (exotérmico) y S < 0 (disminuye el desorden) el proceso ocurrirá espontáneamente si es suficientemente exotérmico para sobrepasar la disminución en entropía
Si H > 0 (endotérmico) y S > 0; G < 0 solamente a T bien altas.
Energía Libre de Gibbs

En procesos irreversibles (espontáneos) G < 0; esto representa una tendencia hacia el estado final. En una reacción química indica que la formación de productos está favorecida.
Por el contrario, un valor de G > 0 representa una tendencia de espontaneidad hacia los reactivos a partir de los productos.
En procesos reversibles (de equilibrio) G = 0; esto representa la condición de un sistema estable en que la velocidad de formación de productos y sustratos es igual.
Energía Libre de Gibbs

Dada una reacción A B, ésta cursará de izquierda a derecha cuando las concentraciones de A y B sean tales que la G <0.La energía libre del sistema viene dada por:
Donde G0 es la Energía Libre Standard de la reacción (reactivos a concentración 1M, en condiciones STP) . Conociendo G0 podemos calcular si un proceso puede tener lugar espontáneamente o no en unas determinadas condiciones.
G = G0 + RT ln [A]
[B]
Energía Libre de Gibbs

En el equilibrio, G = 0; por tanto,
= G0 + RT ln [Aeq][Beq]
Pero[Aeq][Beq] = Keq Por tanto, G0 = -RT ln Keq
Energía Libre de Gibbs

Condiciones intracelulares:Temperatura: 37 ºC, equivalentes a 310 ºK[Glucosa-6-fosfato], 1 mM[Glucosa], 0.01 mM[fosfato], 10 mM
Glucosa 6P + H2O Glucosa + Pi
G0 = -3250 cal/mol
G =G0 + RT ln[Glucosa] [Fosfato]
[Glucosa-6-fosfato]
Sustituyendo, obtenemos:
G = -3250 + 1.98*310*2.303* log 10-5*10-3
10-2 = -8900 cal/mol

Estructuras complejas
Estructuras simples
Catabolismo AnabolismoG
Acoplamiento catabolismo-anabolismo

En general, podemos decir:
Reacciones catabólicas: G < 0
Reacciones anabólicas: G > 0
Supongamos una reacción anabólica mediante la cual se formaun enlace químico entre A y B; esquemáticamente,
A + B A-B (G > 0)
La reacción no puede tener lugar espontáneamente. ¿Cómo puede entonces tener lugar en el metabolismo?

Lo que ocurre en el metabolismo es que las reaccionesendergónicas (G > 0) se acoplan a reacciones exergónicas (G < 0) de manera que :
1. La energía liberada en una de las reacciones es utilizada por la otra.
2. La suma total de energías libres de una y otra reacción da un G < 0, por lo que el proceso en conjunto tiene lugar espontáneamente.
Acoplamiento catabolismo-anabolismo

Así, la reacción
A + B A-B (G1 > 0)
Se acopla a
X-Y + H2O X + Y (G2 < 0)
Dando lugar a una reacción global
A + B + X-Y + H2O A-B + X + Y
Siendo |G2 | > |G1|
(G < 0)

Acoplamiento catabolismo-anabolismo
La reacción que se acopla en los seres vivos a procesos endergónicos es, en la mayoría de los casos, la hidrólisis de polifosfatos, principalmente la hidrólisis de ATP
O-POPO
OO
O-O-
R + H2O O-P
O
O-
HOPO
O
O-
R OH +
OCH2 N
N
N
N
NH2
OHOH
OPOPOP-O
O O O
O-O-O-
ATP
Hidrólisis de polifosfatos
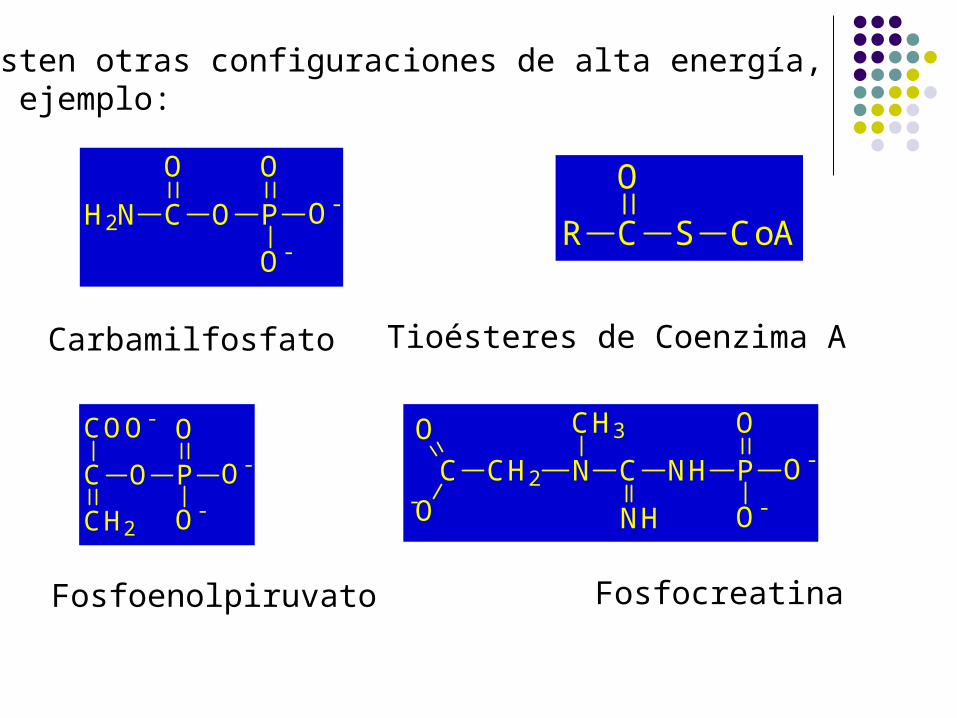
Existen otras configuraciones de alta energía,por ejemplo:
Fosfoenolpiruvato
O-PO
O
O-
C
CH2
COO-
O-P
O
O-
NHC
NH
N
CH3
CH2C
O
-O
Fosfocreatina
O-P
O
O-
OC
O
H2N
Carbamilfosfato
R C
O
S CoA
Tioésteres de Coenzima A
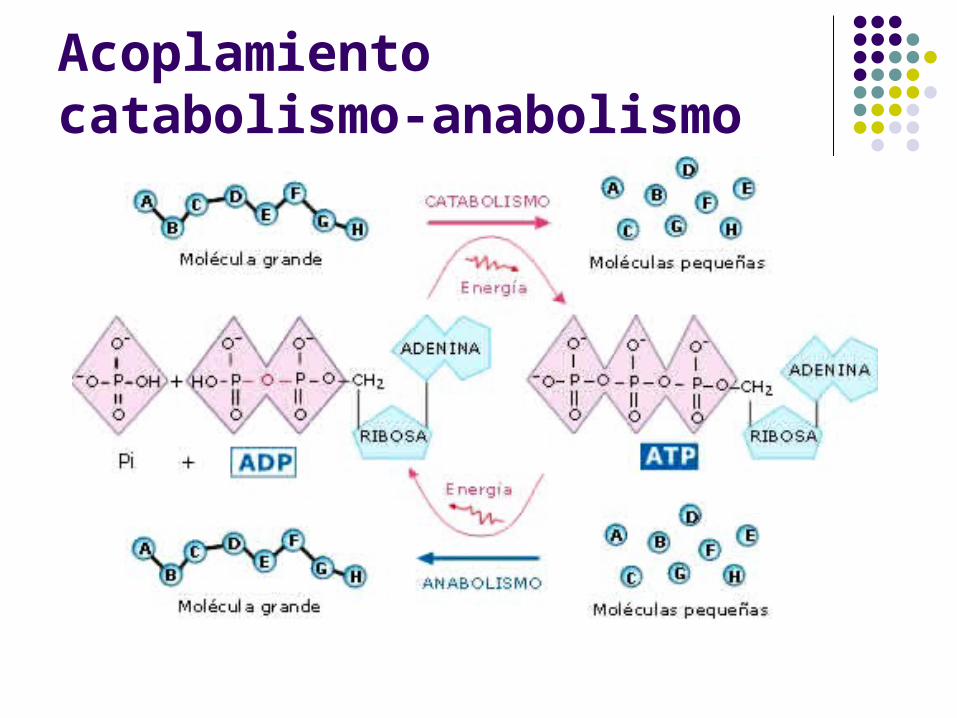
Acoplamiento catabolismo-anabolismo

Acoplamiento catabolismo-anabolismo

Los procesos catabólicos productores de energía generan ATP, que se empleará en todas aquellas reacciones endergónicas en las que sea requerido.
El ATP puede ser sintetizado mediante: Fosforilación a nivel de sustrato Fosforilación oxidativa
Acoplamiento catabolismo-anabolismo

Óxido-Reducción
Las reacciones de óxido reducción son reacciones de transferencia de electrones. En estas reacciones un elemento se oxida, es decir pierde electrones, y otro elemento gana electrones, es decir, se reduce.
El elemento que se oxida es el agente reductor porque dona los electrones, mientras que el elemento que se reduce es el agente oxidante.

Catabolismo y Anabolismo
CATABOLISMO
ANABOLISMO

Coenzimas óxido-reducción
Adenina
RibosaAdenina
Ribosa
NAD+ NADH + H+
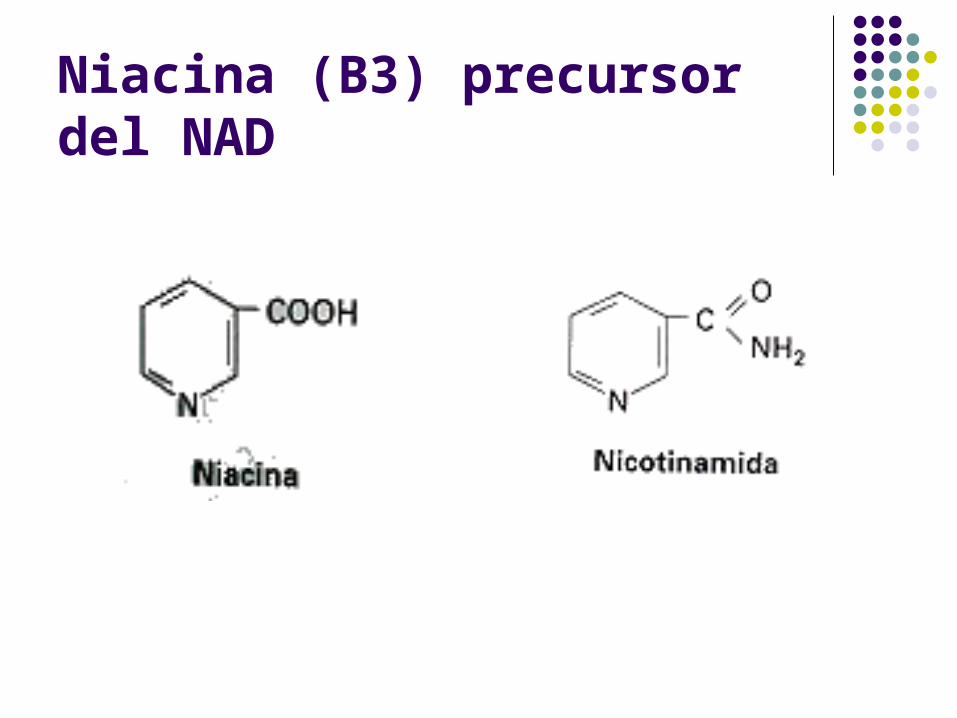
Niacina (B3) precursor del NAD

Coenzimas óxido-reducción
FAD
FADH2

Rivoflavina (B2) precursor FAD

MITOCONDRIA
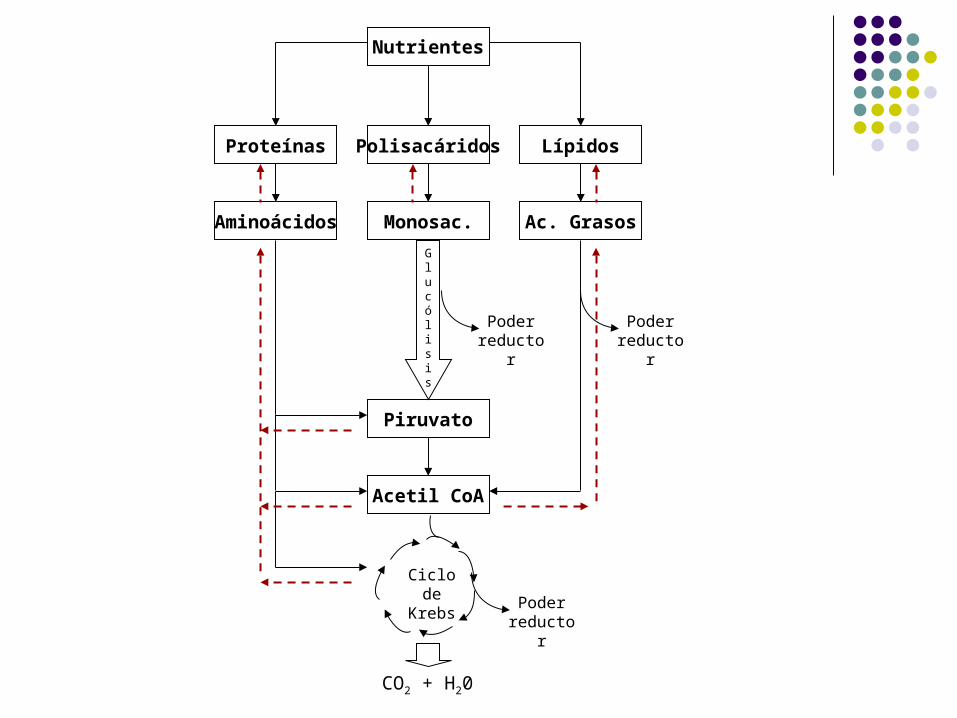
Ciclo de Krebs
Nutrientes
Proteínas Polisacáridos Lípidos
Aminoácidos Monosac. Ac. Grasos
Glucólisis
Piruvato
Acetil CoA
Poder reductor
Poder reductor
Poder reductor
CO2 + H20

Ciclo de Krebs
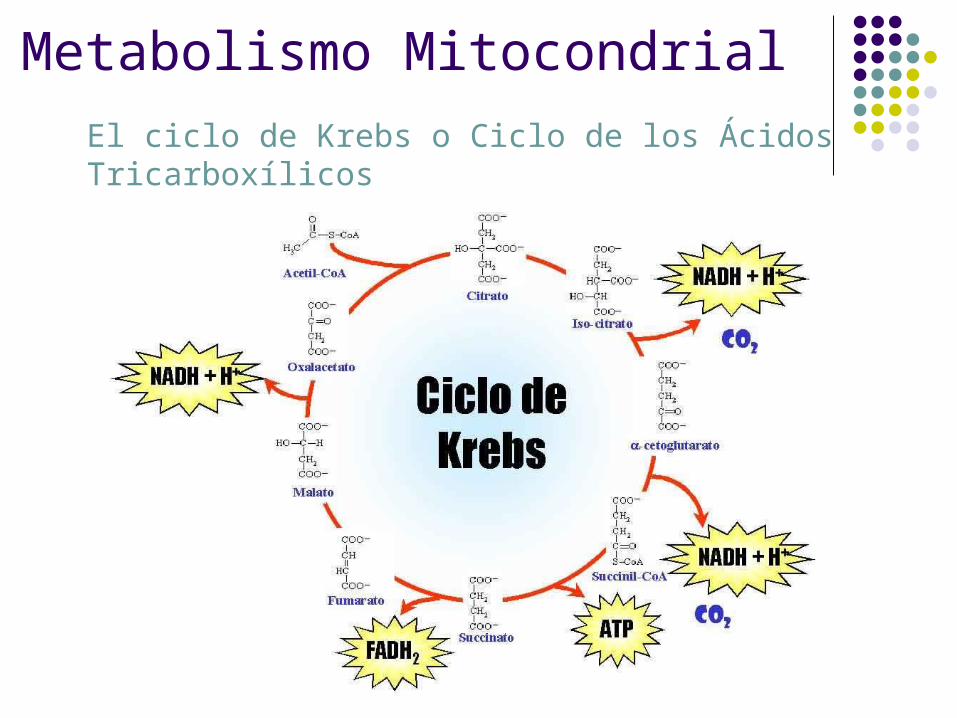
Metabolismo Mitocondrial
El ciclo de Krebs o Ciclo de los Ácidos Tricarboxílicos

Complejos multienzimáticos
Ejemplo: El complejo multienzimático Piruvato deshidrogenasa (PDH) está formado
por múltiples copias de tres enzimas:
Piruvato deshidrogenasa
(E1)
Dihidrolipoil transacetilasa
(E2)
Dihidrolipoil deshidrogenasa
(E3)

Complejos multienzimáticosPiruvato deshidrogenasa
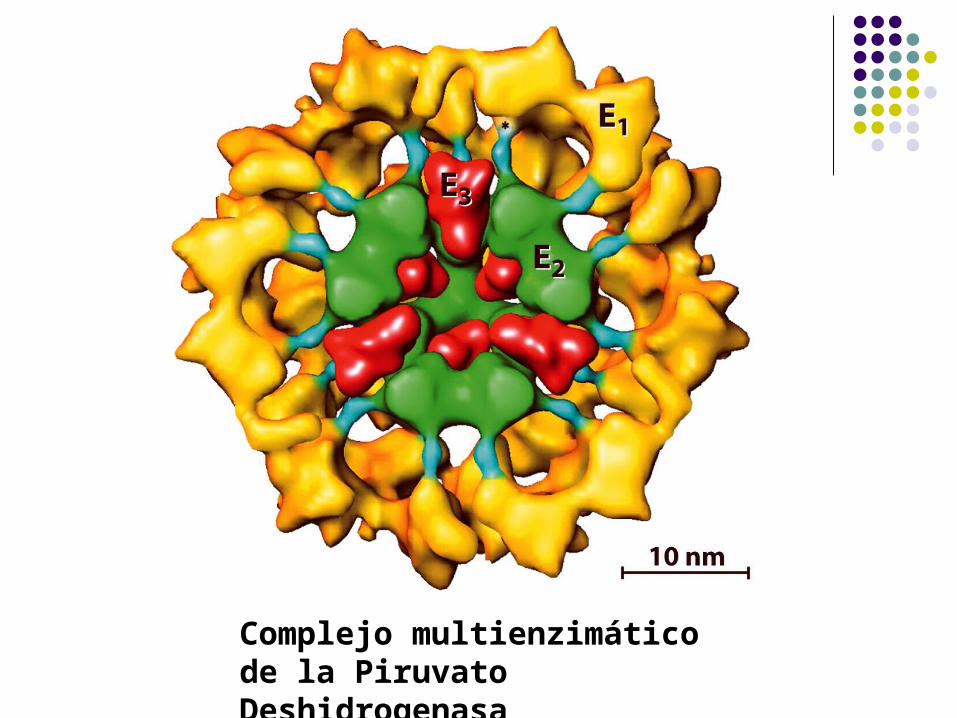
Complejo multienzimático de la Piruvato Deshidrogenasa

Complejo multienzimático de la Piruvato Deshidrogenasa

Complejo multienzimático de la Piruvato Deshidrogenasa

Regulación de la Piruvado Deshidrogensa
h

Regulación Piruvato Deshidrogenasa


















