LOS GENES · moleculares de la herencia y ... te se puede ver la organización típica de los genes...
16
LOS GENES CIENCIAS NATURALES Introducción. ¿Qué dicen los genes? | ADN: el material genético | El genoma eucariota | ¿Cómo se expresa la información contenida en el ADN? | El proceso de transcripción | Topología de un gen eucariota típico | ¿Cómo se traduce el mensaje? | Regulación de la expresión de genes eucariotas | Transcripción diferencial de genes | Procesamiento alternativo de intrones | Traducción selectiva de ARNm | Modificación diferencial de proteínas Autora: Dra. Nora B. Calcaterra (UNR y CONICET) | Coordinación Autoral: Dr. Alberto Kornblihtt (UBA y CONICET) EXPLORA LAS CIENCIAS EN EL MUNDO CONTEMPORÁNEO PROGRAMA DE CAPACITACIÓN MULTIMEDIAL
Transcript of LOS GENES · moleculares de la herencia y ... te se puede ver la organización típica de los genes...
LOS GENESCIENCIAS NATURALES
Introducción. ¿Qué dicen los genes? | ADN: el material genético | El genoma eucariota | ¿Cómo se expresa lainformación contenida en el ADN? | El proceso de transcripción | Topología de un gen eucariota típico | ¿Cómo se traduce elmensaje? | Regulación de la expresión de genes eucariotas | Transcripción diferencial de genes | Procesamientoalternativo de intrones | Traducción selectiva de ARNm | Modificación diferencial de proteínas
Autora: Dra. Nora B. Calcaterra (UNR y CONICET) | Coordinación Autoral: Dr. Alberto Kornblihtt (UBA y CONICET)
EXPLORALAS CIENCIAS EN EL MUNDO CONTEMPORÁNEO
PROGRAMA
DE CAPACITACIÓN
MULTIMEDIAL

2 EXPLORA CIENCIAS NATURALES
INTRODUCCIÓN. ¿QUÉ DICEN LOS GENES?
ADN: EL MATERIAL GENÉTICO
A partir de los experimentos deGregor Mendel fue posible pos-
tular la existencia de factores here-dables, responsables de transmitir lascaracterísticas de una especie de gene-ración en generación. Estos factoresheredables fueron posteriormenteidentificados como genes, secuenciasespecíficas de nucleótidos de ADN ubi-cadas en los cromosomas de las célu-las. Pero ¿cuál es la naturaleza de laconexión entre los cromosomas y loscaracteres heredables? ¿Cuáles sonlos mecanismos que una célula utiliza
para especificar cómo será su descen-dencia? ¿Qué dicen los genes real-mente? ¿Cómo es su mensaje tradu-cido por la célula en característicasespecíficas, tales como el color deojos o la altura de una persona, laforma de las hojas, el tamaño, elcolor y el sabor de los frutos?
En este fascículo analizaremos algu-nos mecanismos moleculares que lle-van a cabo las células eucariotas yque permiten responder las pregun-tas planteadas.
U na vez demostrado que el mate-rial genético es el ADN, fue
necesario conocer la estructura tridi-mensional de esta molécula parapoder responder a dos preguntas:cómo se replica y de qué manera diri-ge la síntesis de proteínas. En 1953,los científicos James Watson y FrancisCrick propusieron un modelo queestableció las principales característi-cas de la estructura del ADN y permi-tió comenzar a entender las basesmoleculares de la herencia y la evolu-ción. El modelo estructural proponeque el ADN es una hélice de doblecadena de desoxirribonucleótidos, dediámetro uniforme, que se tuercehacia la derecha, antiparalela (las doscadenas corren en sentido contrario).
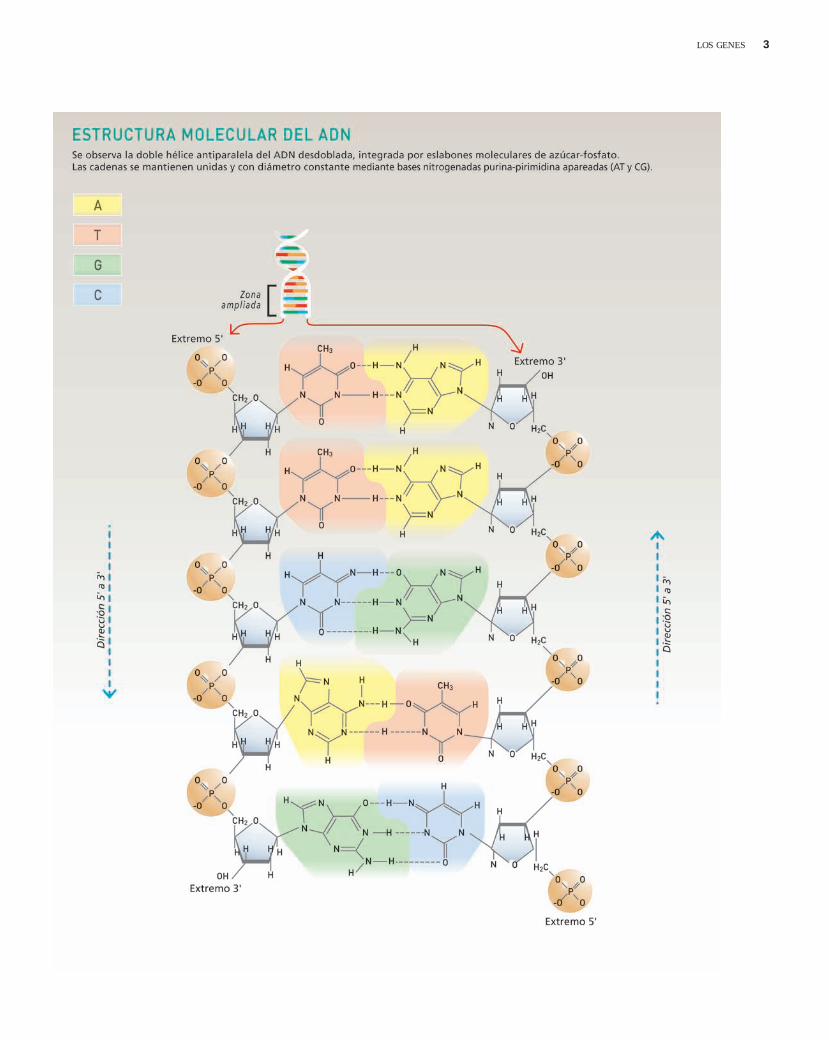
¿Qué quiere decir que las dos cade-nas son antiparalelas? La dirección deun polinucleótido puede definirsemirando los enlaces fosfodiéster entrenucleótidos adyacentes. En el esquele-to de azúcar-fosfato, un grupo fosfatounido al carbono 5' de una desoxirri-bosa se conecta al grupo hidroxilo del
Cromosoma de una célula de insectovisto con microscopio óptico.
carbono 3' de otra molécula de desoxi-rribosa, uniendo azúcares sucesivosentre sí. Por ello, los extremos de unpolinucleótido son distintos: en uno deellos el carbono 5' está libre (extremo5') mientras que en el otro lo está elcarbono 3' (extremo 3'). Las dos cade-nas de polinucleótidos de una molécu-la de ADN corren en sentidos opuestosdado que mientras una de ellas lo haceen sentido 5'3', la cadena comple-mentaria corre en sentido 3'5'.
En la molécula de ADN, los esquele-tos de azúcar-fosfato de las cadenasde polinucleótidos se enrollan alrede-dor de la parte externa de la hélice ylas bases nitrogenadas se ubican en elcentro. Las cadenas se mantienenunidas por apareamiento de las basesnitrogenadas purinas (A o G) o pirimi-dinas (T o C). En esta estructura,siempre una A (adenina) se enfrenta auna T (timina), y una C (citosina) seenfrenta a una G (guanina). Esta com-plementariedad de bases A-T y C-Ghace que se mantenga constante eldiámetro de la doble hélice.
En este fascículo no analizaremos endetalle los mecanismos molecularesmediante los que se replica una molé-cula de ADN. Sólo mencionaremosalgunas de las principales característi-cas de este proceso. La bioquímica dela replicación es semejante en célulasprocariotas y eucariotas. En todos loscasos ocurre de manera finamente re-gulada y con la participación de unagran cantidad de proteínas. In vivo, lareplicación ocurre una sola vez duran-te el ciclo de vida de una célula, duran-te la fase S de la interfase. Sin embar-go, in vitro es posible lograr que unamolécula de ADN se replique variasveces en un tubo de ensayo, sin la pre-sencia de células. Para ello se requiereADN, una enzima (ADN polimerasa) yuna mezcla de los cuatro desoxinucleó-tidos trifosfato. Esto se debe a que elADN sirve de molde para su propia sín-tesis, actuando como guía para la ubi-cación exacta de los nucleótidos com-plementarios en la nueva cadena. Laposibilidad de replicación del ADN invitro ha resultado de importancia fun-
Inst
itu
t Pa
steu
r /
Fran
cia
3LOS GENES

4 EXPLORA CIENCIAS NATURALES
E l genoma de un organismo sedefine como la totalidad del ADN
de sus cromosomas y los genes queeste contiene. Las organelas de loseucariotas (mitocondrias y plastidios)también contienen ADN. Por lo tanto,se puede hablar de genoma mitocon-drial o genoma plastídico para dife-renciarlos del genoma nuclear.
Todos los individuos de una mismaespecie tienen distribuido su genomaen un número característico de cromo-somas. Por ejemplo, las células somáti-cas de los humanos tienen 46 cromo-somas. Sin embargo, no somos la úni-ca especie en tener esa cantidad decromosomas; algunas especies deanimales y vegetales también tienen46 cromosomas. Por otro lado, el ta-maño del genoma no presenta unarelación directa con la complejidaddel organismo. Las células humanas,por ejemplo, tienen un genoma unas700 veces más grande que el de labacteria E. coli, pero las células de
algunos anfibios y plantas poseen ungenoma 30 veces mayor que el de lashumanas. Todo esto indica que no esel tamaño del genoma o el númerode cromosomas lo que hace única acada especie, sino la informaciónespecificada en los genes de esos cro-mosomas. Es el orden particular de lasbases purinas o pirimidinas, es decir,la secuencia de ADN, la que especifi-ca la exacta instrucción para generarun organismo particular.
Como se analizará en detalle luego,no todo el genoma de un organismolleva información para sintetizar unamolécula de ARN funcional. Así, en lamayoría de los genomas de los verte-brados, los genes no se encuentranyuxtapuestos, sino que están separa-dos por largos segmentos de ADN queconstituyen las regiones intergénicas.Estas regiones representan alrededorde un 70 % del total del genoma. Porotro lado, las secuencias de geneseucariotas que codifican proteínas ha-
bitualmente no son continuas, sinoque están interrumpidas por secuen-cias no codificantes. Las secuencias nocodificantes se denominan intrones ylas secuencias codificantes, exones. Enconsecuencia, los intrones, presentesen el genoma, están ausentes en elARNm y, por lo tanto, no se traducenen proteínas. Los exones son los seg-mentos que están presentes en elgenoma, en el ARNm precursor nuclear(pre-ARNm), permanecen en el ARNmmaduro citoplasmático y son traduci-dos en proteínas. En la página siguien-te se puede ver la organización típicade los genes en un cromosoma de ver-tebrado.
El conocimiento de la organizacióngeneral del genoma de varias espe-cies demostró las similitudes y dife-rencias estructurales existentes entreellas, pero también planteó el desafíode conocer la secuencia de nucleóti-dos completa del genoma de unaespecie en particular. Estos emprendi-
EL GENOMA EUCARIOTA
damental para el desarrollo de la bio-tecnología.
El ADN tiene toda la informaciónnecesaria para el funcionamiento ce-lular y el establecimiento de los ca-racteres específicos de una especie oindividuo. Por ello, es importante quepueda ser copiado con fidelidad paraser repartido entre la progenie, ya seancélulas que se regeneran o nuevosindividuos de una especie. Se han pro-puesto muchas hipótesis sobre cómose replica el ADN. En 1953, Watson yCrick formularon la hipótesis semicon-servativa que fue posteriormentedemostrada por Matthew Meselson yFranklin Stahl en 1957. En la replica-ción semiconservativa, la doble héli-ce se separa y cada una de las cade-nas sirve de molde para la síntesis deuna nueva cadena complementaria.
El resultado final son dos moléculasidénticas a la original, cada una con-teniendo una cadena original y otrasintetizada de novo.
El reconocimiento de que el ADNpodía copiarse de esta forma sugiriócómo esta molécula podía poseeruna de las características necesariasdel material genético: la capacidad demutar. Sin mutaciones no habría varia-bilidad genética ni diversidad de espe-cies. Las mutaciones representan cam-bios en la secuencia de bases en elADN. Si el ADN es copiado por unmecanismo que implica apareamientode bases complementarias, todo cam-bio en una cadena será reflejado enuna nueva secuencia durante un nue-vo ciclo de replicación, y la secuenciamutada pasará a las moléculas hijasmediante el mismo mecanismo.

5LOS GENES
mientos científicos son conocidoscomo Proyectos Genoma y sus aplica-ciones tienden a resolver problemasde la medicina, la biotecnología, lasfuentes de energía, etc. En la actuali-dad se han completado los ProyectosGenoma de varias especies, entre ellasel de la mostaza silvestre (Arabidopsisthaliana), el de la mosca de la fruta(Drosophila melanogaster), el del gu-sano (Caenorhabditis elegans), el delratón (Mus musculus) y el del humano(Homo sapiens).
¿CÓMO SE EXPRESA LA INFORMACIÓN CONTENIDA EN EL ADN?
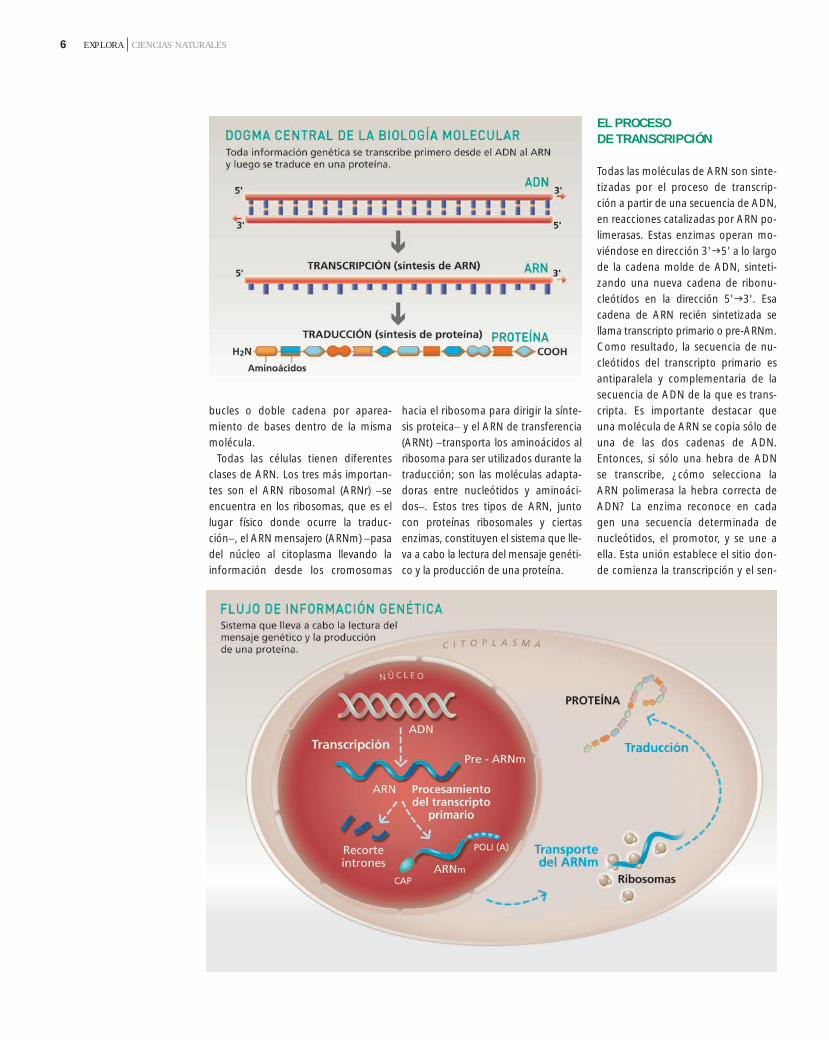
Unos años después de haber propues-to el modelo estructural del ADN,Francis Crick propuso "el dogma cen-tral de la biología molecular", quedice que la información fluye en elsentido: ADNARN proteína. Segúnsus palabras, "una vez que la informa-
ción pasó a una proteína no puedesalir nuevamente".
Este flujo de la información constade dos etapas: primero, en la trans-cripción, se copia la información de lasecuencia de ADN en informacióncorrespondiente a una secuencia deARN. Luego, durante la traducción,esta información se traduce en unasecuencia de aminoácidos, es decir,una proteína.
Si bien el flujo de la información esen un solo sentido, en algunos casos latranscripción puede ser invertida:ARNADN. Esta reacción ocurre me-diante enzimas específicas llamadastranscriptasas reversas, presentes, porejemplo, en los retrovirus como el VIH.Sin embargo, la traducción nunca pue-de ser invertida. Es decir, la informa-ción contenida en una proteína nopuede pasar al ADN. Esto explica porqué no es posible heredar los caracte-res adquiridos de los progenitores.
¿Cómo pasa la información del nú-
cleo (donde está el ADN) a las proteí-nas (que son sintetizadas en el cito-plasma)? ¿Cuál es la relación entre unasecuencia de nucleótidos específica yla secuencia de aminoácidos en unaproteína? Para responder a estos inte-rrogantes, Crick postuló la existenciade moléculas mensajeras y moléculasadaptadoras. Hoy sabemos que esasmoléculas son distintas clases de ARN.El ARN es un ácido nucleico muy seme-jante al ADN, pero entre ambas molé-culas hay diferencias: El azúcar que compone el ARN esribosa, en lugar de desoxirribosa. En lugar de timina, el ARN contie-ne una pirimidina relacionada, el ura-cilo (U). El ARN se encuentra como cadenasimple y no forma una estructura heli-coidal regular como el ADN.
A pesar de esto último, algunas mo-léculas de ARN presentan una estruc-tura secundaria, es decir que ciertaszonas de la molécula pueden formar
6 EXPLORA CIENCIAS NATURALES
bucles o doble cadena por aparea-miento de bases dentro de la mismamolécula.
Todas las células tienen diferentesclases de ARN. Los tres más importan-tes son el ARN ribosomal (ARNr) −seencuentra en los ribosomas, que es ellugar físico donde ocurre la traduc-ción−, el ARN mensajero (ARNm) −pasadel núcleo al citoplasma llevando lainformación desde los cromosomas
hacia el ribosoma para dirigir la sínte-sis proteica− y el ARN de transferencia(ARNt) −transporta los aminoácidos alribosoma para ser utilizados durante latraducción; son las moléculas adapta-doras entre nucleótidos y aminoáci-dos−. Estos tres tipos de ARN, juntocon proteínas ribosomales y ciertasenzimas, constituyen el sistema que lle-va a cabo la lectura del mensaje genéti-co y la producción de una proteína.
EL PROCESO DE TRANSCRIPCIÓN
Todas las moléculas de ARN son sinte-tizadas por el proceso de transcrip-ción a partir de una secuencia de ADN,en reacciones catalizadas por ARN po-limerasas. Estas enzimas operan mo-viéndose en dirección 3'5' a lo largode la cadena molde de ADN, sinteti-zando una nueva cadena de ribonu-cleótidos en la dirección 5'3'. Esacadena de ARN recién sintetizada sellama transcripto primario o pre-ARNm.Como resultado, la secuencia de nu-cleótidos del transcripto primario esantiparalela y complementaria de lasecuencia de ADN de la que es trans-cripta. Es importante destacar queuna molécula de ARN se copia sólo deuna de las dos cadenas de ADN.Entonces, si sólo una hebra de ADNse transcribe, ¿cómo selecciona laARN polimerasa la hebra correcta deADN? La enzima reconoce en cadagen una secuencia determinada denucleótidos, el promotor, y se une aella. Esta unión establece el sitio don-de comienza la transcripción y el sen-
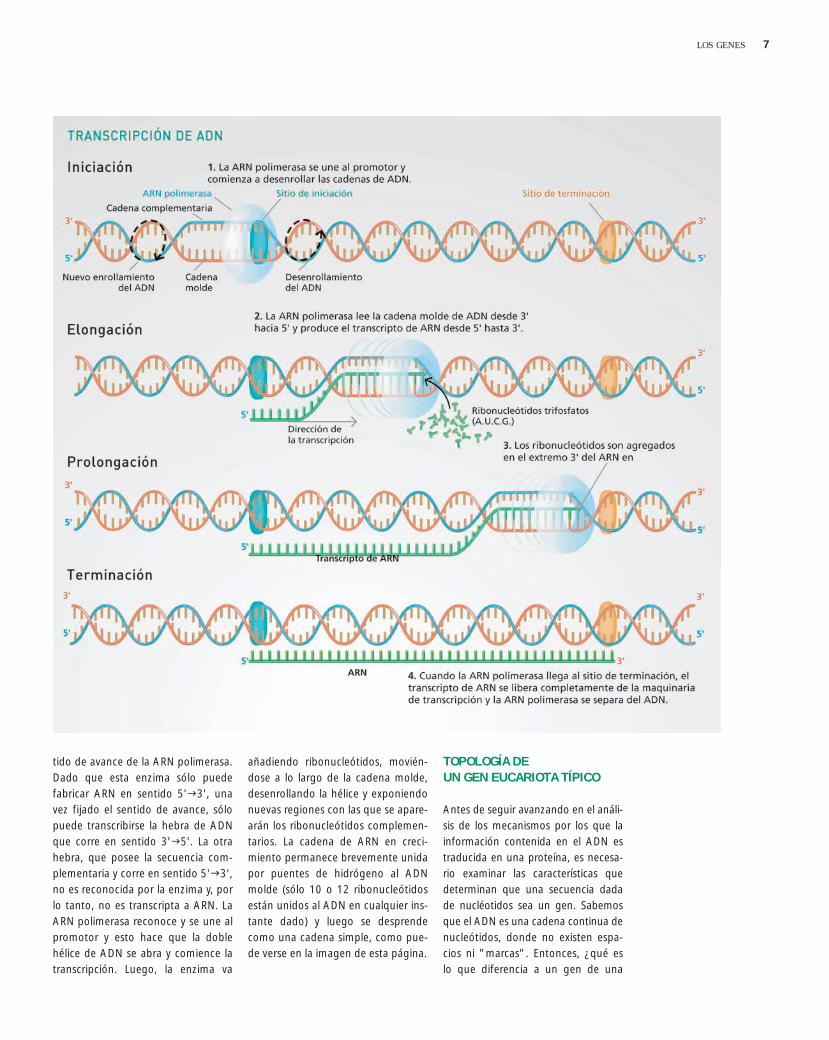
7LOS GENES
tido de avance de la ARN polimerasa.Dado que esta enzima sólo puedefabricar ARN en sentido 5'3', unavez fijado el sentido de avance, sólopuede transcribirse la hebra de ADNque corre en sentido 3'5'. La otrahebra, que posee la secuencia com-plementaria y corre en sentido 5'3',no es reconocida por la enzima y, porlo tanto, no es transcripta a ARN. LaARN polimerasa reconoce y se une alpromotor y esto hace que la doblehélice de ADN se abra y comience latranscripción. Luego, la enzima va
añadiendo ribonucleótidos, movién-dose a lo largo de la cadena molde,desenrollando la hélice y exponiendonuevas regiones con las que se apare-arán los ribonucleótidos complemen-tarios. La cadena de ARN en creci-miento permanece brevemente unidapor puentes de hidrógeno al ADNmolde (sólo 10 o 12 ribonucleótidosestán unidos al ADN en cualquier ins-tante dado) y luego se desprendecomo una cadena simple, como pue-de verse en la imagen de esta página.
TOPOLOGÍA DE UN GEN EUCARIOTA TÍPICO
Antes de seguir avanzando en el análi-sis de los mecanismos por los que lainformación contenida en el ADN estraducida en una proteína, es necesa-rio examinar las características quedeterminan que una secuencia dadade nucléotidos sea un gen. Sabemosque el ADN es una cadena continua denucleótidos, donde no existen espa-cios ni "marcas". Entonces, ¿qué eslo que diferencia a un gen de una

8 EXPLORA CIENCIAS NATURALES
secuencia intergénica? Típicamente,un gen eucariota presenta los siguien-tes elementos. El promotor y las regiones regula-doras (potenciadores y silenciadores).Como se verá más adelante, proteí-nas llamadas factores de transcripciónse unen a ellas y regulan la actividadde un gen. El sitio de iniciación de la transcrip-ción. Aquí se determina qué nucleótidoserá el extremo 5' del ARN. En el pre-ARNm, este extremo se modifica por elagregado de un nucleótido de 7-metil-guanosina, llamado caperuza o cap,tan pronto ha sido transcripto. Estamodificación del extremo 5' del pre-ARNm se llama capping. La caperuzaes necesaria para la unión del ARNm alribosoma y la subsiguiente traducción. El sitio de iniciación de la traducción(ATG). Esta secuencia (que se convier-
te en AUG en el ARN) se localiza a dis-tancias variables del sitio de iniciaciónde la transcripción, según el gen deque se trate. Entre el sitio de iniciaciónde la transcripción y el triplete ATGqueda comprendida una secuenciaque no se traduce, llamada región notraducida del extremo 5' o 5'UTR (delinglés 5' untranslated region). Exones e intrones. Secuencias pre-sentes en el gen y en el pre-ARNmque serán conservadas o eliminadas,respectivamente, durante el procesa-miento del transcripto. Los genes trans-criben pre-ARNm, que son más largosque los ARNm maduros porque contie-nen intrones que deben ser eliminadosen el núcleo antes de pasar al citoplas-ma. Este mecanismo se conoce como"procesamiento por corte y empalme"o splicing. El procesamiento del pre-ARNm es mediado por un complejo
de proteínas y pequeños ARN nuclea-res (small nuclear RNA o snRNA). Estecomplejo, llamado espliceosoma, seensambla reconociendo secuencias denucleótidos específicas en los intrones.Una vez ensamblado el espliceosoma,el pre-ARNm es cortado, se liberan lassecuencias reconocidas como intronesy se empalman los extremos de lassecuencias reconocidas como exones,para producir un ARNm maduro. El codón de terminación. Puede te-ner tres diferentes conformaciones:TAA, TGA y TAG. La región 3'UTR que, aunque estranscripta, no es traducida a proteí-nas. Esta región incluye la secuenciaAATAAA, necesaria para la poliadeni-lación, una modificación del ARN queconsiste en el agregado de una "cola"de unos 200 o 300 nucleótidos de ade-nina. Estas A son agregadas enzimáti-

9LOS GENES
camente al transcripto; no son parte dela secuencia génica. La cola de poli(A)confiere estabilidad al ARNm, permiteque este salga del núcleo y su ausenciainhibe la traducción.
A partir de lo explicado sobre loselementos característicos de un geneucariota se desprende que sólo po-demos hablar de ARNm una vez queel transcripto de ARN (pre-ARNm) hasufrido las modificaciones en susextremos 5' (capping) y 3' (poliadeni-lación) y hayan sido procesadas sussecuencias intrónicas (splicing).
¿CÓMO SE TRADUCE EL MENSAJE?
Finalizadas la transcripción, las modifi-caciones de los extremos 5' y 3' y elsplicing, el ARNm es exportado al ci-toplasma donde se traduce el mensa-je. Durante la traducción, el orden denucleótidos del ARNm especifica elorden de aminoácidos en un poli-péptido. De acuerdo con el códigogenético, cada triplete o codón enel ARNm codifica un aminoácido enparticular.
El código genético está compuestopor 64 codones: 61 de ellos especifi-can aminoácidos y los otros 3 son
señales de finalización del procesode traducción. Teniendo en cuentaque las proteínas están compuestaspor 20 aminoácidos diferentes yexisten 61 codones para especificar-los, es obvio que más de un tripletepuede codificar el mismo aminoáci-do. Por eso decimos que el códigogenético es redundante o degenera-do. Otra característica del códigogenético es su universalidad: el mis-mo código es utilizado por todos losorganismos.
La síntesis de proteínas ocurre en losribosomas, que consisten en dos subu-nidades, una grande y otra pequeña,cada una formada por ARNr y proteí-nas. La subunidad grande contieneuna depresión en una de sus superfi-cies, en la que se ajusta la subunidadpequeña. El ARNm se inserta en unsurco formado entre las superficies decontacto de las dos subunidades.
Para la traducción también se requie-ren los ARNt, que están plegados enuna estructura tridimensional que seasemeja a la letra "L". Tienen un triple-te de bases, el anticodón, en el plega-miento central de la estructura secun-daria. Una enzima específica, la ami-noacil ARNt sintetasa, agrega un ami-noácido determinado en el extremo3' de cada ARNt, convirtiéndolos enaminoacil-ARNt. El apareamiento debases complementarias codón-antico-dón permite trasladar la informaciónespecificada en el ARNm a un aminoá-cido específico.
La traducción se realiza en variasetapas. La iniciación ocurre cuando lasubunidad ribosómica más pequeñase une al extremo 5' de una moléculade ARNm y se mueve en sentido 5'3'hasta encontrar el codón de iniciaciónAUG. La primera molécula de amino-acil-ARNt, que lleva el aminoácidometionina (Met), se acopla con elcodón de iniciación. El complejo de
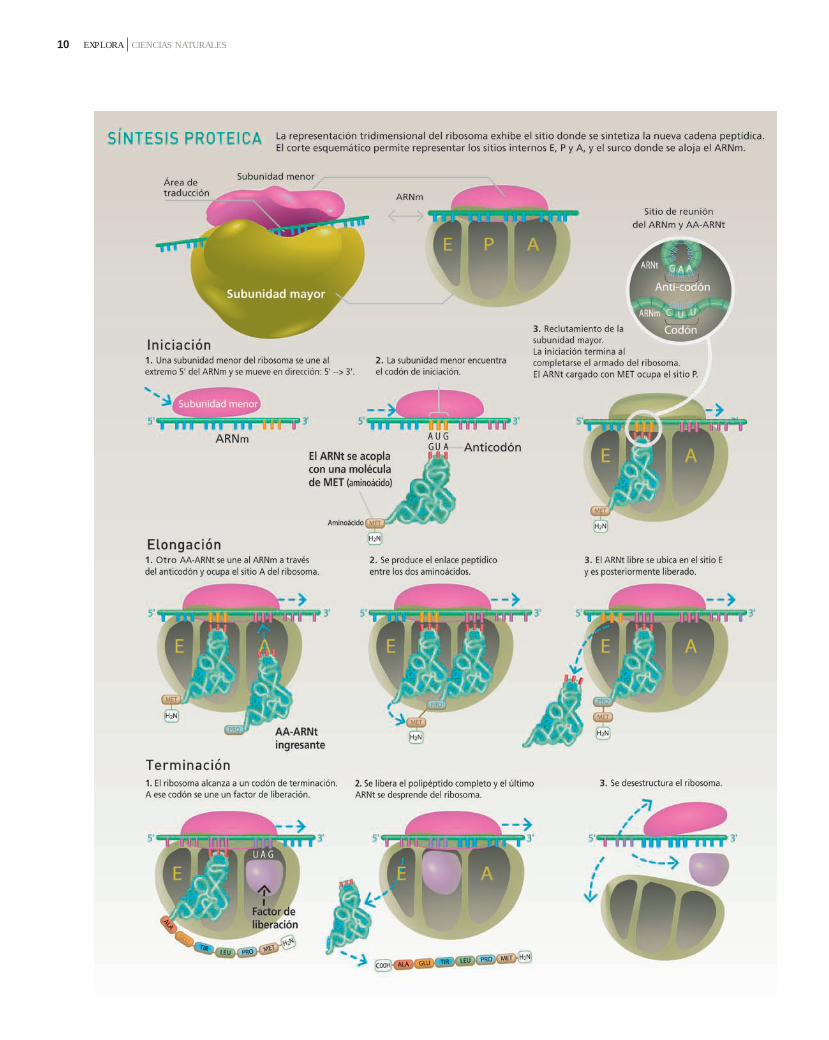
10 EXPLORA CIENCIAS NATURALES

11LOS GENES
REGULACIÓN DE LA EXPRESIÓN DE GENES EUCARIOTAS
primer aminoácido y su ARNt. El ribo-soma se mueve a lo largo de la cade-na de ARNm en una dirección 5'3',y el segundo ARNt, con el dipéptidounido (peptidil-ARNt), se mueve des-de el sitio A al sitio P. El primer ARNtse mueve hacia el sitio E y se despren-de del ribosoma, dejando el aminoá-cido que transportaba en el péptidoen formación. En suma, durante laelongación del péptido, el sitio A uneel aminoacil-ARNt correspondiente alaminoácido que deberá incorporarseal péptido en formación, el sitio P uneel peptidil-ARNt y el sitio E es el sitiode salida para el ARNt que estuvo pre-viamente unido al sitio P. Un terceraminoacil-ARNt se coloca en el sitio Ay se forma otro enlace peptídico. La
iniciación se completa cuando lasubunidad ribosómica grande se unea estas estructuras. El ribosoma arma-do contiene ahora 3 sitios: A, P y E; eneste momento, el complejo ARNt-METocupa el sitio P y los sitios A y E que-dan vacantes.
La elongación comienza cuando unsegundo aminoacil-ARNt se coloca enel sitio A y su anticodón se acopla conel ARNm. Se forma un enlace peptídi-co entre los dos aminoácidos. Lo inte-resante es que esta reacción es catali-zada por un pequeño ARN presente enla subunidad mayor del ribosoma. Aestos ARN con actividad enzimáticase los denomina ribozimas. Al mismotiempo que ocurre la unión de dos ami-noácidos se rompe el enlace entre el
L a finalización de varios ProyectosGenoma mostró que el número de
regiones que codifican proteínas nopresenta una correlación con la com-plejidad del organismo ni con su origenevolutivo. C. elegans, D. melanogastery Homo sapiens contienen aproxi-madamente 19.000, 14.000 y 25.000genes, respectivamente. ¿Cómo pue-de el genoma de la mosca de la frutacontener menos genes que el delindudablemente más simple gusano?El conocimiento de la secuencia com-pleta de todos los genes de un indivi-duo no alcanza para responder estapregunta. "La secuencia no es el finaldel día. Es sólo el comienzo", diceSydney Brenner, un líder de estos pro-yectos. La respuesta está en los dife-rentes proteomas, es decir, en el con-junto de proteínas que cada uno deellos expresa. No basta con conocerla secuencia del genoma completo,hay que caracterizar las proteínas quelos genes ordenan elaborar, por eso elproteoma es el actual desafío de labiología.
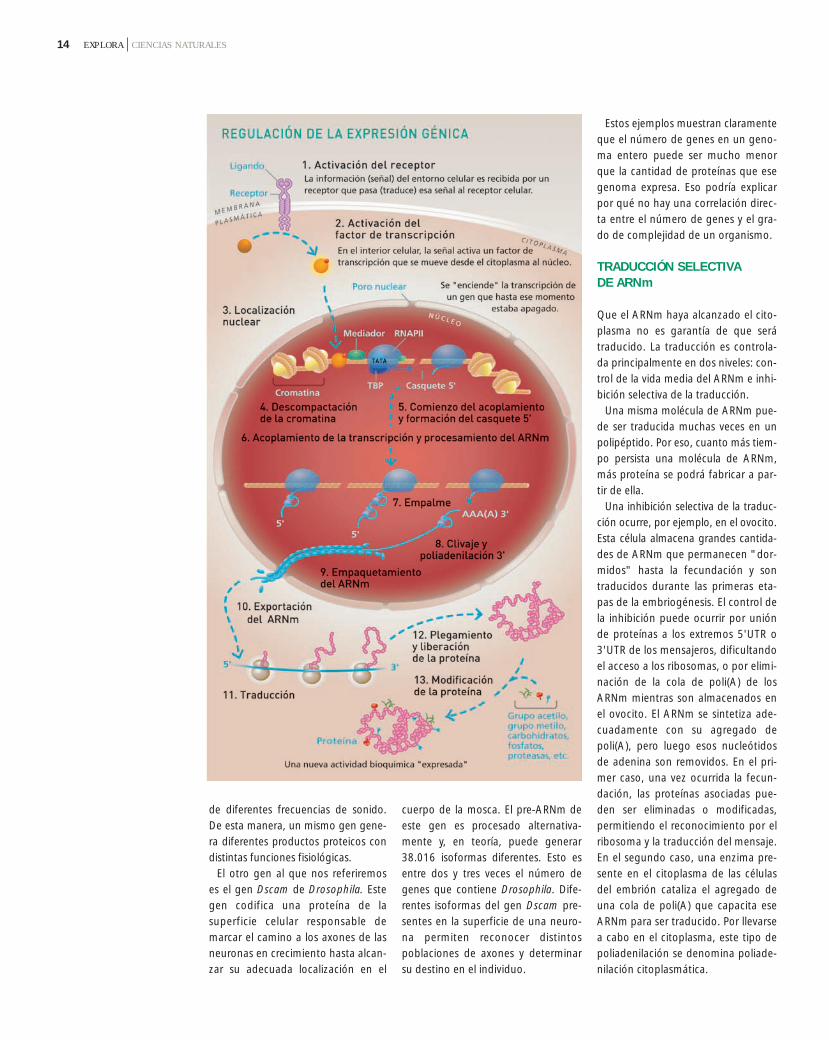
¿Qué determina los tipos y cantida-des de las diversas proteínas que carac-terizan un organismo o una célula enparticular? Todas las células de un or-ganismo multicelular con tejidos espe-cializados poseen los mismos genes.Entonces, ¿cómo es que en un mismoorganismo una célula puede diferen-ciarse en un glóbulo blanco y otra encélula de la piel? Lo que ocurre es que,aunque los genes están siempre pre-sentes en un determinado genoma, noestán siempre “encendidos”. Decimosque un gen se expresa (se enciende)cuando se transcribe y el ARNm se tra-duce a proteína. El control de cualquie-ra de los pasos de este proceso puedeconducir a una expresión génica dife-rencial en tipos celulares específicos.Por lo tanto, los glóbulos blancos sediferencian de las células de la piel noporque tengan diferentes genes, sinoporque expresan diferentes proteínas.
La regulación de la expresión génicapuede ocurrir en distintos niveles: Transcripción diferencial de genes, re-gulando cuáles de los genes son trans-
criptos a ARNm en un momento o tipocelular determinado. Procesamiento alternativo de intro-nes, regulando cuál de los ARN trans-criptos (o qué parte de ese ARNnuclear) será transportado al citoplas-ma para ser traducido a proteína. Selección en el núcleo de los ARNmque deben ser exportados al citoplasma. Traducción selectiva del ARNm,regulando cuál de los ARNm será tra-ducido a proteína en el citoplasma. Desestabilización selectiva de ARNmen el citoplasma. Modificación diferencial de proteí-nas, regulando qué proteínas seránfuncionales en la célula.
TRANSCRIPCIÓN DIFERENCIAL DE GENES
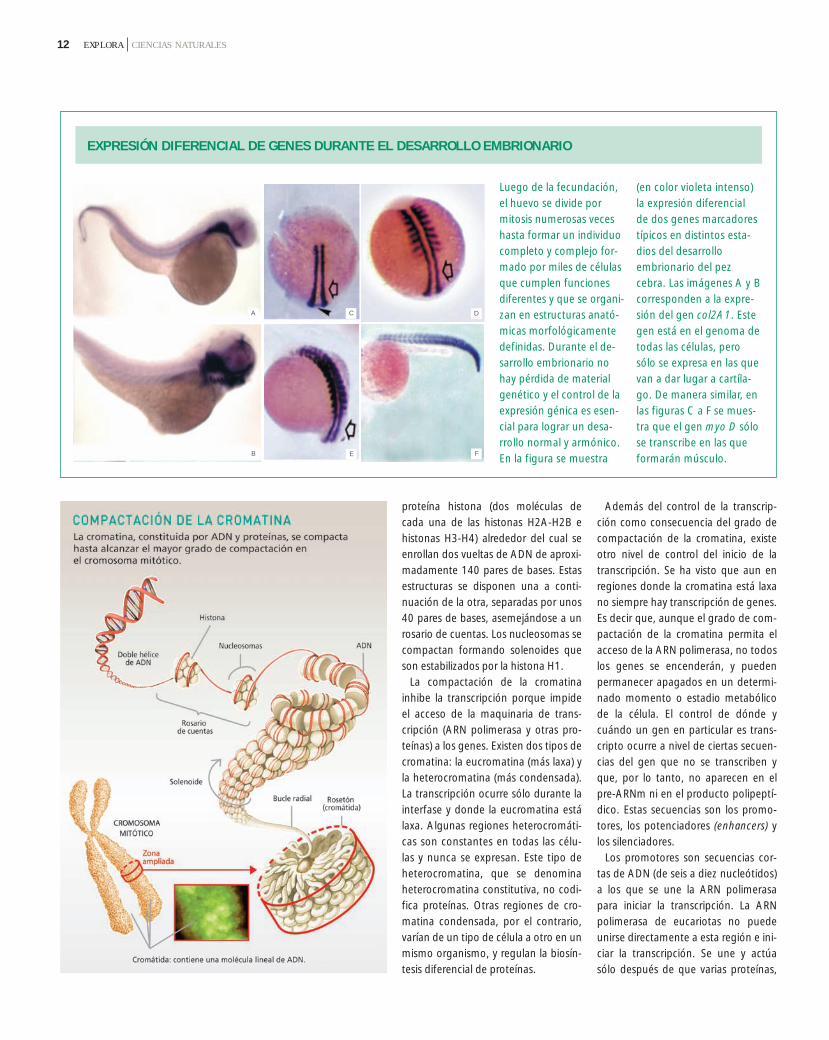
Los genes eucariotas están conteni-dos en un complejo de ADN y proteí-nas llamado cromatina. La unidadbásica de la cromatina es el nucleoso-ma, que está compuesto por un cora-zón proteico de ocho moléculas de la
cadena peptídica naciente siempreestá unida al ARNt que se está mo-viendo del sitio A al sitio P, y el amino-acil-ARNt entrante siempre ocupa elsitio A. Este paso se repite una y otravez incorporando los distintos ami-noácidos hasta que se completa elpolipéptido.
La terminación ocurre cuando elribosoma alcanza un codón de termi-nación, el polipéptido se escinde delúltimo ARNt y el ARNt se desprendedel sitio E. El sitio A es ocupado por unfactor de liberación que produce ladisociación de las dos subunidades delribosoma.
A modo de resumen, la imagen dela página anterior ilustra los procesosdescriptos.
12 EXPLORA CIENCIAS NATURALES
proteína histona (dos moléculas decada una de las histonas H2A-H2B ehistonas H3-H4) alrededor del cual seenrollan dos vueltas de ADN de aproxi-madamente 140 pares de bases. Estasestructuras se disponen una a conti-nuación de la otra, separadas por unos40 pares de bases, asemejándose a unrosario de cuentas. Los nucleosomas secompactan formando solenoides queson estabilizados por la histona H1.
La compactación de la cromatinainhibe la transcripción porque impideel acceso de la maquinaria de trans-cripción (ARN polimerasa y otras pro-teínas) a los genes. Existen dos tipos decromatina: la eucromatina (más laxa) yla heterocromatina (más condensada).La transcripción ocurre sólo durante lainterfase y donde la eucromatina estálaxa. Algunas regiones heterocromáti-cas son constantes en todas las célu-las y nunca se expresan. Este tipo deheterocromatina, que se denominaheterocromatina constitutiva, no codi-fica proteínas. Otras regiones de cro-matina condensada, por el contrario,varían de un tipo de célula a otro en unmismo organismo, y regulan la biosín-tesis diferencial de proteínas.
Además del control de la transcrip-ción como consecuencia del grado decompactación de la cromatina, existeotro nivel de control del inicio de latranscripción. Se ha visto que aun enregiones donde la cromatina está laxano siempre hay transcripción de genes.Es decir que, aunque el grado de com-pactación de la cromatina permita elacceso de la ARN polimerasa, no todoslos genes se encenderán, y puedenpermanecer apagados en un determi-nado momento o estadio metabólicode la célula. El control de dónde ycuándo un gen en particular es trans-cripto ocurre a nivel de ciertas secuen-cias del gen que no se transcriben yque, por lo tanto, no aparecen en elpre-ARNm ni en el producto polipeptí-dico. Estas secuencias son los promo-tores, los potenciadores (enhancers) ylos silenciadores.
Los promotores son secuencias cor-tas de ADN (de seis a diez nucleótidos)a los que se une la ARN polimerasapara iniciar la transcripción. La ARNpolimerasa de eucariotas no puedeunirse directamente a esta región e ini-ciar la transcripción. Se une y actúasólo después de que varias proteínas,
EXPRESIÓN DIFERENCIAL DE GENES DURANTE EL DESARROLLO EMBRIONARIO
Luego de la fecundación,el huevo se divide pormitosis numerosas veceshasta formar un individuocompleto y complejo for-mado por miles de célulasque cumplen funcionesdiferentes y que se organi-zan en estructuras anató-micas morfológicamentedefinidas. Durante el de-sarrollo embrionario nohay pérdida de materialgenético y el control de laexpresión génica es esen-cial para lograr un desa-rrollo normal y armónico.En la figura se muestra
(en color violeta intenso)la expresión diferencial de dos genes marcadorestípicos en distintos esta-dios del desarrolloembrionario del pezcebra. Las imágenes A y Bcorresponden a la expre-sión del gen col2A1. Estegen está en el genoma detodas las células, perosólo se expresa en las quevan a dar lugar a cartíla-go. De manera similar, enlas figuras C a F se mues-tra que el gen myo D sólose transcribe en las queformarán músculo.
A
B
C D
FE

13LOS GENES
llamadas factores de transcripciónbasales, se han ensamblado sobre elpromotor del gen.
Existen otros factores de transcrip-ción, llamados reguladores, que seunen a las regiones potenciadoras osilenciadoras. Los potenciadores sonsecuencias de ADN a las que se unenfactores de transcripción reguladores yactivan la utilización de un promotor,controlando la eficiencia y la velocidadde la transcripción. De manera similar,las secuencias silenciadoras puedenbloquear específicamente la transcrip-ción de un gen en un grupo de célulaso regular el momento en que un gense expresa. Estas secuencias puedenestar incluso a distancias muy lejanasdel promotor. Son muy variadas y espe-cíficas de cada gen, regulan su expre-sión dependiendo del tipo celular, deseñales provenientes de otras células,del entorno, etc.
Una vez que una célula se ha dife-renciado ocurre una serie de procesoscelulares para mantener estable supatrón de transcripción y asegurar quesu progenie conserve las mismas carac-terísticas y propiedades.
PROCESAMIENTO ALTERNATIVO DE INTRONES
Originalmente se pensó que cualquie-ra fuera la composición de intrones yexones de un pre-ARNm determinado,este daría lugar al mismo ARNm. Sinembargo, se ha demostrado que dife-rentes tipos celulares que transcribenel mismo pre-ARNm, pero lo procesande manera diferencial, generan dife-rentes ARNm y, por lo tanto, distintasproteínas.
En la ilustración del procesamientodiferencial del pre-ARNm se muestracómo es posible generar familias deproteínas a partir del procesamientodiferencial de un único pre-ARNm. Laeliminación de ciertos exones poten-ciales en una célula pero no en otrasconduce a la formación de proteínasestrechamente relacionadas, pero dis-tintas. Estas proteínas diferentes codi-ficadas por el mismo gen se denomi-nan isoformas.
El procesamiento alternativo delpre-ARNm determina qué secuenciapresente en el pre-ARNm será elimi-nada como intrón. Lo que es un intrónen el núcleo de una célula puede no
serlo en el núcleo de otra. En los extre-mos de los intrones existen secuenciasconservadas en todos los genes quepueden ser reconocidas o no por elespliceosoma de forma regulada. Enconsecuencia, la maduración por cor-te y empalme es un proceso regulado.
La existencia de un procesamientoalternativo de un pre-ARNm implicaque genes con docenas de intronespueden originar miles de proteínas re-lacionadas pero diferentes. A modode ejemplo, describiremos dos genespara los que el procesamiento alterna-tivo de intrones permite generar unaextraordinaria diversidad molecular.Uno de ellos es el gen slo, que codificauna proteína de membrana presenteen las células del oído interno. Estaproteína es, al menos en parte, la res-ponsable de la percepción del ampliorango de frecuencias que el oído pue-de percibir. Existen al menos ocho si-tios de procesamiento alternativo en elpre-ARNm de slo, lo que posibilita quemás de 500 ARNm diferentes puedanser finalmente generados. Este proce-samiento alternativo es funcionalmen-te relevante ya que distintas isoformasdel gen slo participan en la percepción
14 EXPLORA CIENCIAS NATURALES
de diferentes frecuencias de sonido.De esta manera, un mismo gen gene-ra diferentes productos proteicos condistintas funciones fisiológicas.
El otro gen al que nos referiremoses el gen Dscam de Drosophila. Estegen codifica una proteína de lasuperficie celular responsable demarcar el camino a los axones de lasneuronas en crecimiento hasta alcan-zar su adecuada localización en el
cuerpo de la mosca. El pre-ARNm deeste gen es procesado alternativa-mente y, en teoría, puede generar38.016 isoformas diferentes. Esto esentre dos y tres veces el número degenes que contiene Drosophila. Dife-rentes isoformas del gen Dscam pre-sentes en la superficie de una neuro-na permiten reconocer distintospoblaciones de axones y determinarsu destino en el individuo.
Estos ejemplos muestran claramenteque el número de genes en un geno-ma entero puede ser mucho menorque la cantidad de proteínas que esegenoma expresa. Eso podría explicarpor qué no hay una correlación direc-ta entre el número de genes y el gra-do de complejidad de un organismo.
TRADUCCIÓN SELECTIVA DE ARNm
Que el ARNm haya alcanzado el cito-plasma no es garantía de que serátraducido. La traducción es controla-da principalmente en dos niveles: con-trol de la vida media del ARNm e inhi-bición selectiva de la traducción.
Una misma molécula de ARNm pue-de ser traducida muchas veces en unpolipéptido. Por eso, cuanto más tiem-po persista una molécula de ARNm,más proteína se podrá fabricar a par-tir de ella.
Una inhibición selectiva de la traduc-ción ocurre, por ejemplo, en el ovocito.Esta célula almacena grandes cantida-des de ARNm que permanecen "dor-midos" hasta la fecundación y sontraducidos durante las primeras eta-pas de la embriogénesis. El control dela inhibición puede ocurrir por uniónde proteínas a los extremos 5'UTR o3'UTR de los mensajeros, dificultandoel acceso a los ribosomas, o por elimi-nación de la cola de poli(A) de losARNm mientras son almacenados enel ovocito. El ARNm se sintetiza ade-cuadamente con su agregado depoli(A), pero luego esos nucleótidosde adenina son removidos. En el pri-mer caso, una vez ocurrida la fecun-dación, las proteínas asociadas pue-den ser eliminadas o modificadas,permitiendo el reconocimiento por elribosoma y la traducción del mensaje.En el segundo caso, una enzima pre-sente en el citoplasma de las célulasdel embrión cataliza el agregado deuna cola de poli(A) que capacita eseARNm para ser traducido. Por llevarsea cabo en el citoplasma, este tipo depoliadenilación se denomina poliade-nilación citoplasmática.

15LOS GENES
MODIFICACIÓN DIFERENCIAL DE PROTEÍNAS
Una vez que un polipéptido ha sidosintetizado pueden ocurrir diferentescambios para determinar si será bioló-gicamente activo o no. Algunas prote-ínas no son activas a menos que seanmodificadas por agregado covalentede un grupo fosfato (fosforilación) oacetato (acetilación), y también poragregado de azúcares (glicosilación).Otras deben unir iones o incorporarmoléculas orgánicas pequeñas llama-das grupos prostéticos o cofactores.Por ejemplo, la hemoglobina, ademásde la correcta síntesis de los cuatropolipéptidos que la componen, debeincorporar a su estructura un grupoprostético "hemo" para poder cum-plir con su función biológica de trans-portar el oxigeno a través de la san-gre. Todos estos procesos ocurrenluego de que la traducción finalizó, ypor ello se conocen como "modifica-ciones postraduccionales": son regu-ladas y sólo en el caso de llevarse acabo adecuadamente la proteína seráfuncional y la expresión del gen sehará evidente.
¿QUÉ ES UN GEN?
Según el contexto en que se anali-za, un gen puede ser definido dediferentes maneras. Desde el puntode vista mendeliano, un gen es unaunidad de herencia que determinauna característica fenotípica. Deacuerdo con la teoría cromosómicade la herencia, es posible asignarcada uno de los genes de un orga-nismo a un locus específico en loscromosomas, por lo que algunasveces se usa el termino locus comosinónimo de gen. La biología celulardefine gen como una unidad deinformación determinada por unasecuencia específica de nucleótidosque se halla en un sitio particular deun cromosoma. Todas estas defini-
ciones son útiles, pero no abordanel aspecto funcional. En 1909, el físico británicoArchibald Garrod fue el primero ensugerir que los genes dictan losfenotipos por medio de enzimasque catalizan procesos químicosespecíficos en una célula. Poste-riormente, en 1940, Tatum sugirióentonces la hipótesis un gen: unaenzima. Para explicar el origen delas proteínas que no son enzimas ylas formadas por más de un tipo deunidad, fue necesario adoptar unaexpresión más exacta y global de loque es un gen y así se propuso lahipótesis un gen: un polipéptido.Aun esta definición debe ser refina-da y ampliada. Por un lado, lamayoría de los genes eucariotascontienen intrones que pueden ser
procesados de manera reguladamediante splicing alternativo; porotro lado, existen largas porcionesde esos genes que no poseen unsegmento correspondiente a un poli-péptido −por ejemplo, las regiones5' y 3' UTR, la cola de poli(A)−.También es aceptado que las regio-nes promotoras y las secuenciasreguladoras, que no son transcriptasa ARN, son parte de un gen funcio-nal ya que deben estar presentespara que ocurra la transcripción.Además, una definición moleculardebería ser lo suficientemente ampliacomo para incluir el ADN que lleva lainformación para sintetizar ribozi-mas, ARNr, ARNt y snARN. Una defi-nición más adecuada de gen sería"una región del ADN requerida parala síntesis de un ARN funcional".

16 EXPLORA CIENCIAS NATURALES
Al inicio del fascículo se mencionandos enunciados de la biologíamolecular de diferente naturaleza:la hipótesis semiconservativa y eldogma central. En primer lugartenemos la hipótesis, que es, desdesu sentido más literal, una suposi-ción o conjetura: un enunciadoprovisorio que tiene la capacidadde ser sometido a pruebas empíri-cas mediante la observación, laexperimentación u otro tipo deintervenciones sobre el mundo.
El conjunto de pruebas empíricasque atraviesa una hipótesis seconoce como contrastación; la con-trastación de una hipótesis admitedos resultados extremos y muchos"grises" intermedios. Esos resulta-dos extremos son la validación, quesignifica que la hipótesis pasa laprueba con éxito y es aceptada pro-visionalmente para seguir avanzan-do en la investigación científica, y larefutación, que refiere a que la
hipótesis no soporta las pruebas yes descartada totalmente comoerrónea. Los puntos intermediosentre estas dos posturas, que sonmuy frecuentes en las cienciasnaturales, incluyen intervencionesno totalmente concluyentes, inter-pretaciones provisionales de losdatos o validaciones parciales.
Antiguamente se hablaba de"verificación" o de "comproba-ción" de una hipótesis, queriendosignificar que los datos empíricospueden proporcionar una basesegura e incuestionable para losenunciados de la ciencia, y que deellos cabe decir que son "verda-deros". Pero la historia de la cien-cia nos ha mostrado que esta esuna mirada epistemológica inge-nua. Los resultados de la ciencia,por su naturaleza, son siempreprovisorios, perfectibles y sujetosa revisión y crítica.
Buena parte de la provisoriedadde la ciencia se debe a la impor-tante carga teórica que posee laexploración del mundo natural;esta está mediada por las ideas,
las finalidades, los métodos y losvalores de los científicos. Con uncambio en cualquiera de todosesos elementos se pueden intro-ducir modificaciones en el conoci-miento científico "establecido".Esta idea queda ilustrada en laplaqueta final que recorre losdiferentes sentidos que se dio alconcepto de gen a lo largo de lahistoria de la biología.
En segundo lugar, se mencionaen el fascículo un dogma, quetambién podríamos llamar princi-pio o axioma. Este tipo de enun-ciado es mucho más fundamental,abstracto y estructurante que lashipótesis. No significa esto que nopueda ser reemplazado en la histo-ria de la ciencia, sino que un prin-cipio participa del núcleo de unateoría, dando sentido a muchísi-mos modelos enlazados entre sí.Por tanto, un cuestionamiento alos principios científicos asume laforma de una auténtica "revolu-ción", en la cual muchos elemen-tos de la ciencia son alterados pro-fundamente. Se puede producir,
incluso, lo que llamaríamos un"cambio de paradigma", es decir,una nueva elección de presupues-tos teóricos y metodológicos.
Ahora bien, todos los enuncia-dos de la ciencia (hipótesis, dog-mas y muchos otros a los que nodaremos nombres específicos),independientemente de las dife-rencias en su grado de validación,son luego puestos en marcha paraseguir avanzando y, al mismo tiem-po, quedan sujetos a constanterefinamiento, revisión y crítica. Estose puede ver muy bien en el fascículo, en donde enunciadosmás establecidos coexisten connuevos hallazgos que son hoyobjeto de investigación "puntera".
El estilo del texto científico,como hemos comentado en otrofascículo, parece dar estatuto de"descripción verdadera" a todassus afirmaciones. Esta es una carac-terística retórica de las ciencias, quecae lejos de la propia metodologíaautocorrectiva y revisionista queellas tienen, basada en el uso de losmodelos y las inferencias.
EPISTEMOLOGÍA Agustín Adúriz-Bravo
Bibliografía Alberts, B., D. Brays, J. Lewis, K. Raff, K. Roberts y J. D. Watson: Biología
molecular de la célula, Barcelona, Omega, 1996; 3ª ed.Alberts, B., A. Johnson, J. Lewis, M. Raff, K. Roberts y P. Walter: Biología
molecular de la célula, Barcelona, Omega, 2002; 4ª ed.Curtis, H., N. S. Barnes, A. Schnek y G. Flores: Biología, Buenos Aires,
Panamericana, 2000.De Robertis, E., J. Hib y R. Ponzio: Biología celular y molecular,
Buenos Aires, El Ateneo, 1996.Lordish, H., A. Berk, S. L. Zipurskey, P. Matsudaira, D. Baltimore y J.
Darnell: Biología celular y molecular, Buenos Aires, Panamericana, 2002; 4ª ed.
Purves, W. K., D. Sadava, G. H. Orians y H. C. Heller: Vida. La ciencia de la biología, Buenos Aires, Panamericana, 2002.
AgradecimientosLa Dirección Nacional de Gestión Educativa agradece a las siguientes ins-tituciones y personas por permitirnos reproducir material fotográfico ycolaborar en la documentación de imágenes: Institut Pasteur (Francia);Chantal Policieux.
Coordinadora del Áreas Curriculares,Lic. Cecilia CrestaCoordinadora del Área de CienciasNaturales, Lic. Nora BahamondeCoordinadores del Área deCapacitación, Lic. Carlos Ruiz y Lic. Margarita MarturetCoordinadoras del Programa deCapacitación. Explora, Lic. V. Nespereira,Lic. M. D. Urizar y Lic. A. Vendrov
Coordinadora de Edición,Lic. Raquel FrancoCoordinación y documentación, Lic. Rafael BlancoEdición, Lic. Gonzalo BlancoDiseño y diagramación, DG María Eugenia MásCorrección, Norma A. Sosa Pereyra
http://explora.educ.ar
Ministro de Educación, Prof. Alberto Estanislao Sileoni
Secretario de Educación, Lic. Jaime Perczyk
Jefe de Gabinete, A.S. Pablo Urquiza
Subsecretario de Equidad y Calidad Educativa, Lic. Eduardo
Aragundi
Directora Nacional de Gestión Educativa, Lic. Delia Méndez
Directora de Educación Secundaria, Lic. Virginia Vázquez Gamboa