Membrana citoplasmática -...
41
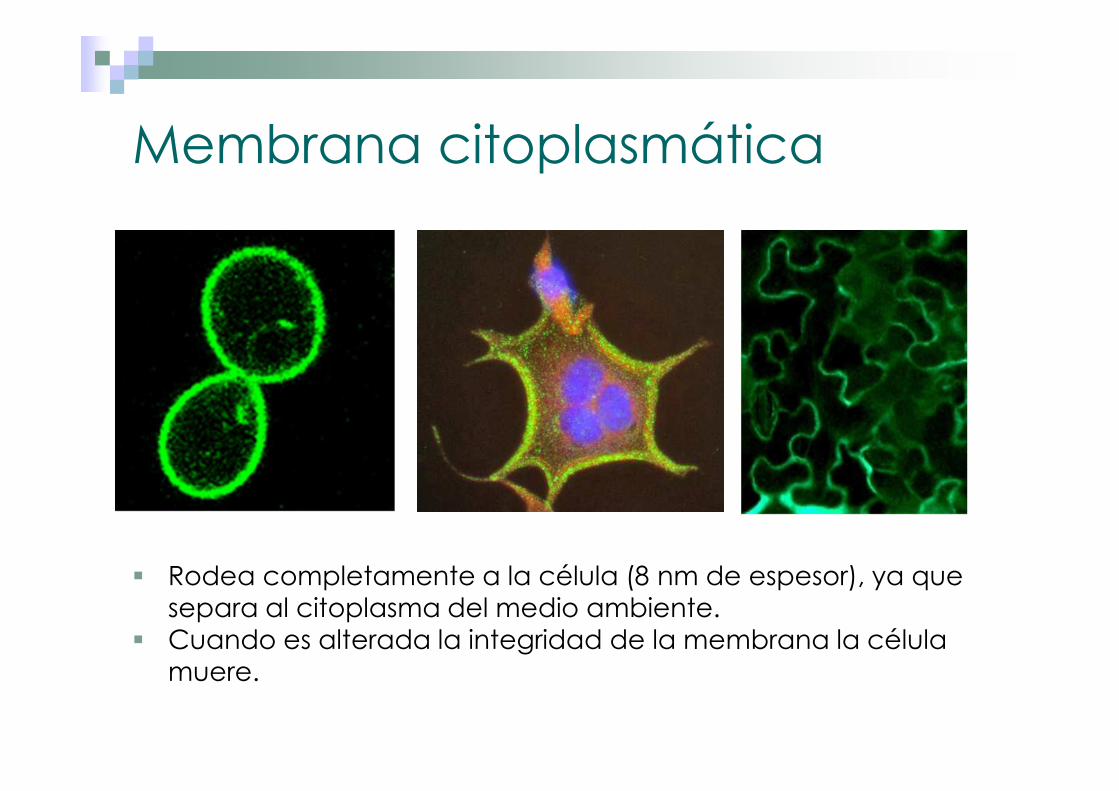
Membrana citoplasmática Rodea completamente a la célula (8 nm de espesor), ya que separa al citoplasma del medio ambiente. Cuando es alterada la integridad de la membrana la célula muere.
Transcript of Membrana citoplasmática -...
Membrana citoplasmática
� Rodea completamente a la célula (8 nm de espesor), ya que separa al citoplasma del medio ambiente.
� Cuando es alterada la integridad de la membrana la célula muere.

Membrana citoplasmática
� Membrana de alta selectividad, facilita la concentración de metabolitos específicos y la excreción de material de desecho.

Estructura
� Los fosfolípidos están formados por ácidos grasos (hidrofóbicos) y glicerol (relativamente hidrofílico) unidos por un enlace ester y un grupo PO4- que se encuentra unido al glicerol.

Estructura
�Existen diferentes formas químicas como resultado de la variación en a naturaleza de los ácidos grasos y los radicales unidos al grupo fosfato.
Estructura
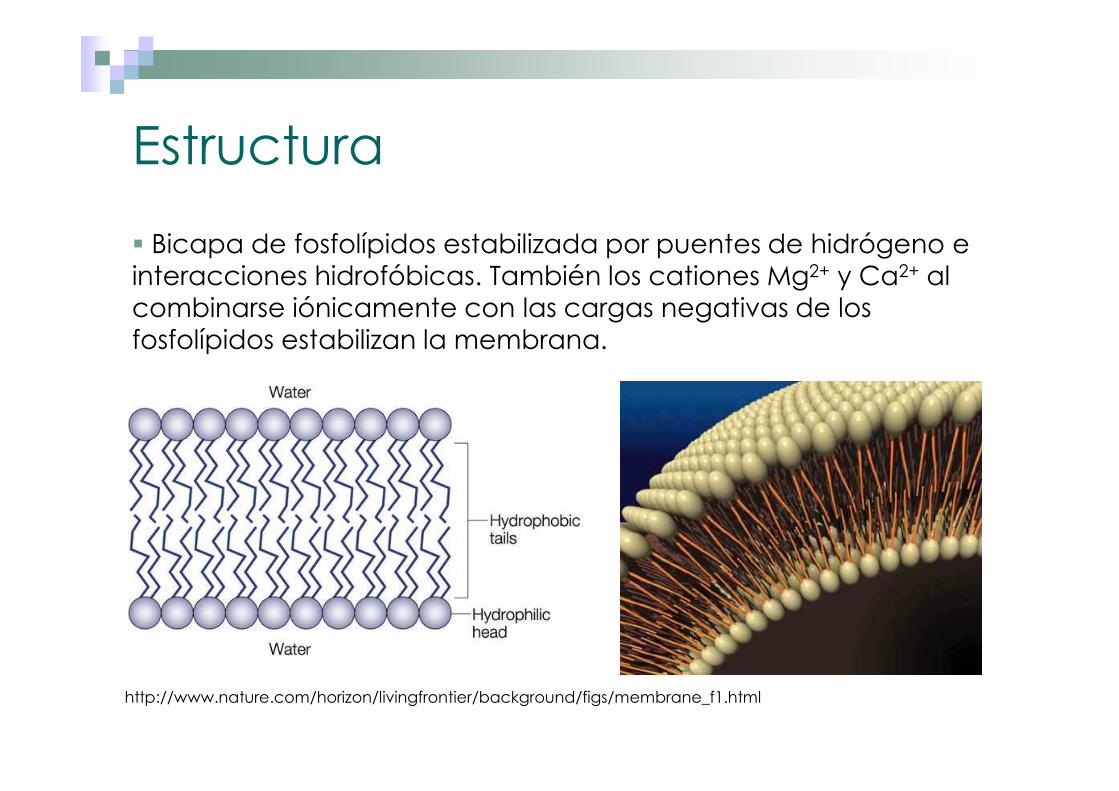
� Bicapa de fosfolípidos estabilizada por puentes de hidrógeno e interacciones hidrofóbicas. También los cationes Mg2+ y Ca2+ al combinarse iónicamente con las cargas negativas de los fosfolípidos estabilizan la membrana.
http://www.nature.com/horizon/livingfrontier/background/figs/membrane_f1.html

Estructura
A B
C D
A B
CCH3
CH3
CH3CH3
CH3CH3HO
A B
C D
HC
CH3
CH3
CH3CH3
CH2
CH2
CH2
HC CH3
Estructura general de los esteroles Colesterol Hopanoide
�En células eucariontes se encuentran también los esteroles y el porcentaje varía entre células. Estos esteroles dan rigidez y estabilizan la membrana. En bacterias, los Micoplasmas (bacterias sin pared celular) también contienen esteroles en su membrana para dar estabilidad.
�En algunas bacterias anaerobias (crecen en ausencia de oxígeno) se encuentran presentes moléculas similares a los esteroles. Los hopanoides no requieren de oxidación durante sus biosíntesis y tienen la misma función que los esteroles.
�Los hopanoides se encuentran en las membranas de muchas bacterias y tienen un papel similar a los esteroles en células eucarióticas.
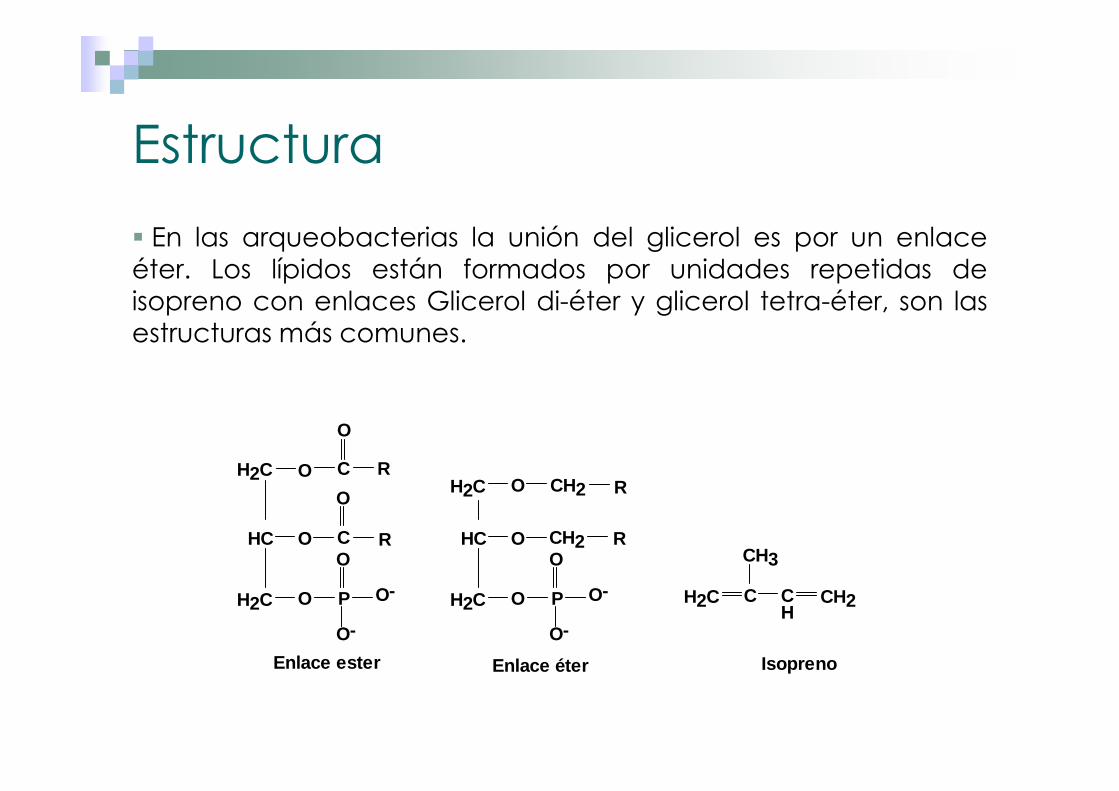
Estructura
� En las arqueobacterias la unión del glicerol es por un enlaceéter. Los lípidos están formados por unidades repetidas deisopreno con enlaces Glicerol di-éter y glicerol tetra-éter, son lasestructuras más comunes.
Enlace ester Enlace éter Isopreno
H2C P
H2C
HC
O
O-
O-
C
CO
O
OO
R
R
O
H2C P
H2C
HC
O
O-
O-
O
OO
R
R
CH2
CH2
H
CH3
H2C C C CH2

Bicapa lipídica en Arqueobacterias
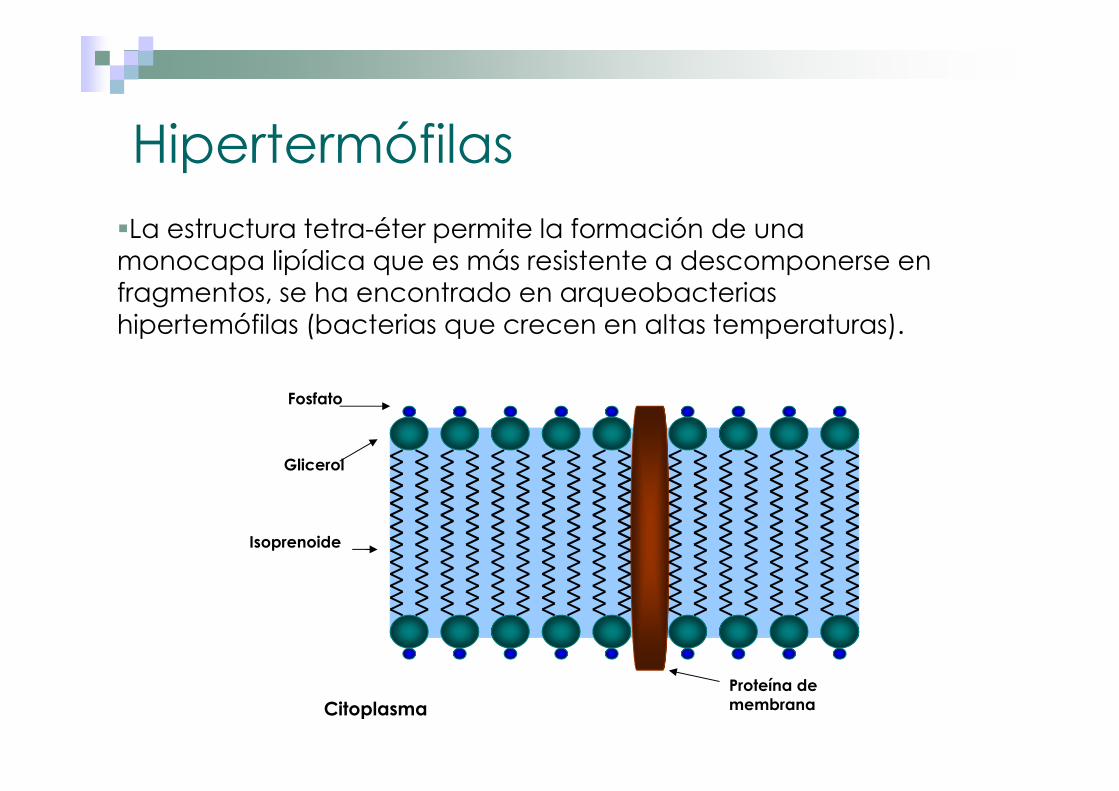
Hipertermófilas�La estructura tetra-éter permite la formación de una monocapa lipídica que es más resistente a descomponerse en fragmentos, se ha encontrado en arqueobacterias hipertemófilas (bacterias que crecen en altas temperaturas).
Isoprenoide
Fosfato
Glicerol
Citoplasma
Proteína de membrana

Proteínas de la membrana
�También las proteínas forman parte de la membrana citoplasmática. Las proteínas periplásmicas o integrales se encuentran embebidas en la membrana y las proteínas membranales periféricas algunas de estás se mantienen en la superficie de la membrana unidas por lípidos. Todas estas tienen gran importancia y dan a la membrana propiedades funcionales.
http://www.youtube.com/watch?v=ULR79TiUj80

Proteínas de la membrana
�La membrana no es una estructura rígida, la presencia de proteínas y proporciona movilidad aunada a que en condiciones normales de crecimiento la membrana presenta un aspecto fluido, por lo que al conjunto se le ha llamado modelo del mosaico fluido.
http://www.nature.com/horizon/livingfrontier/background/figs/membrane_f2.html
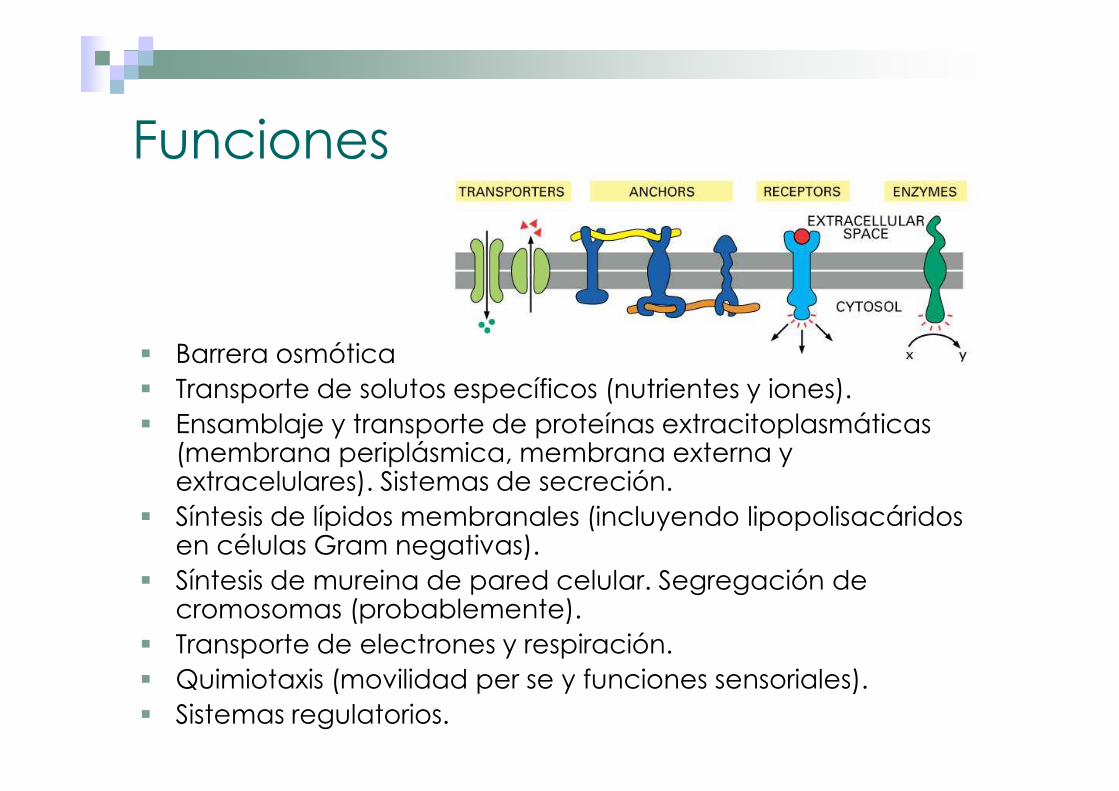
Funciones
� Barrera osmótica� Transporte de solutos específicos (nutrientes y iones).� Ensamblaje y transporte de proteínas extracitoplasmáticas
(membrana periplásmica, membrana externa y extracelulares). Sistemas de secreción.
� Síntesis de lípidos membranales (incluyendo lipopolisacáridos en células Gram negativas).
� Síntesis de mureina de pared celular. Segregación de cromosomas (probablemente).
� Transporte de electrones y respiración.� Quimiotaxis (movilidad per se y funciones sensoriales).� Sistemas regulatorios.
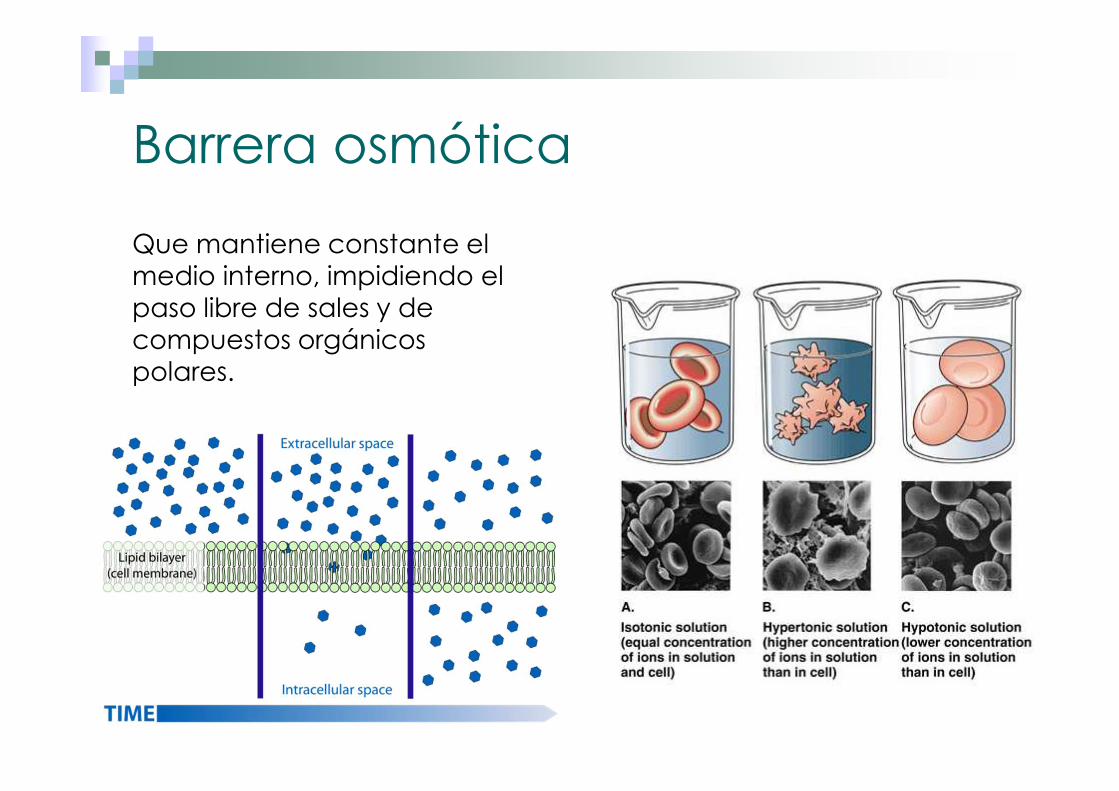
Barrera osmótica
Que mantiene constante el medio interno, impidiendo el paso libre de sales y de compuestos orgánicos polares.

Transporte de solutos específicos
1. Transporte pasivo inespecífico (Difusión simple). Consiste en la difusión pasiva de ciertas sustancias para las que la membrana es impermeable, debido a la diferencia de concentración (∆C) a ambos lados de dicha membrana (la sustancia tiene mayor concentración fuera que dentro de la célula). Aparte de la diferencia de concentración, influyen:
El grado de permeabilidad (Costante P) de la membrana a la sustancia en cuestión y el área o superficie total (A) a través de la que se produce el transporte. La difusión simple se produce por el paso de estas sustancias a través de poros inespecíficos de la membrana citoplásmica.

Transporte de solutos específicos2. Transporte pasivo específico (Difusión facilitada). Permite el
paso de compuestos por difusión a través de una proteína integral de membrana (permeasa o facilitador), cuya conformación determina un canal interior, y por el cual un sustrato específico puede alcanzar el interior, sin gasto de energía. Las proteínas transportadoras, que pueden ser de dos tipos:
�Proteínas de canal. Pequeños poros que permiten el paso en especial de iones inorgánicos (Na+, K+, Cl-, Ca2+), por lo que también se denominan canales iónicos, y se abren o se cierran en respuesta a estímulos eléctricos o a moléculas transmisoras.
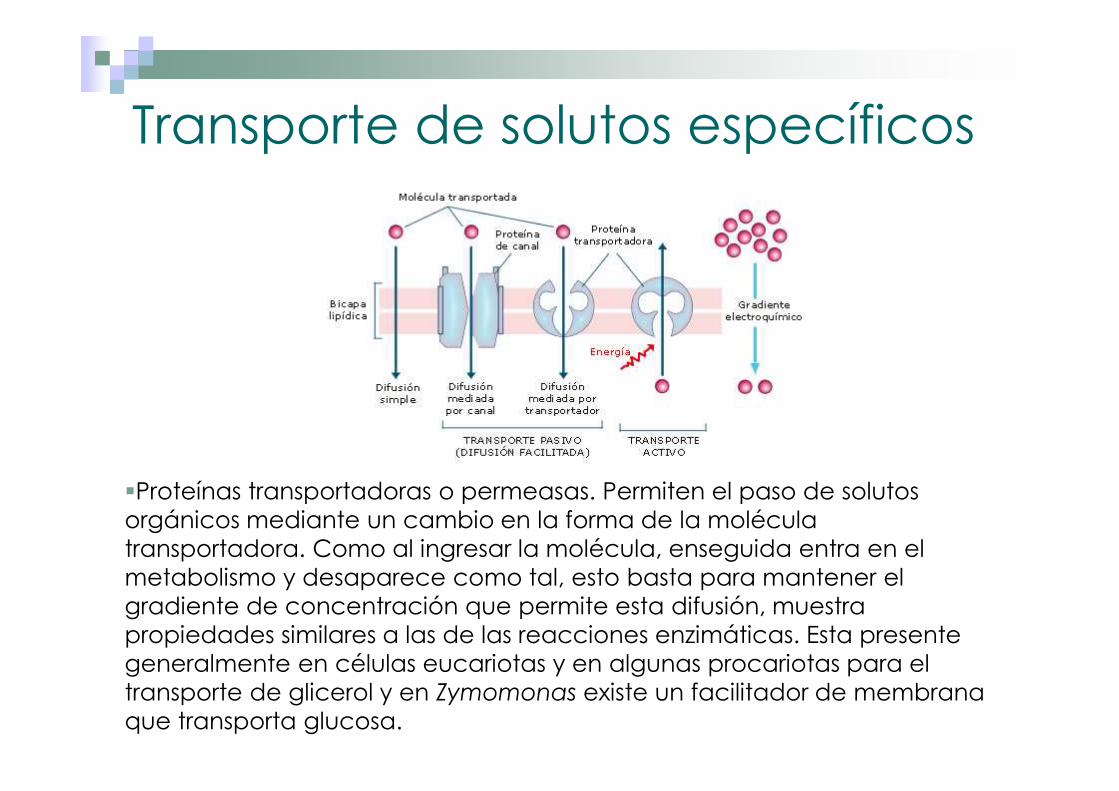
Transporte de solutos específicos
�Proteínas transportadoras o permeasas. Permiten el paso de solutos orgánicos mediante un cambio en la forma de la molécula transportadora. Como al ingresar la molécula, enseguida entra en el metabolismo y desaparece como tal, esto basta para mantener el gradiente de concentración que permite esta difusión, muestra propiedades similares a las de las reacciones enzimáticas. Esta presente generalmente en células eucariotas y en algunas procariotas para el transporte de glicerol y en Zymomonas existe un facilitador de membrana que transporta glucosa.

Transporte de solutos específicos
3. Transporte activo. Requiere un aporte energético, ya que se lleva a cabo en contra del gradiente electroquímico. Siempre ocurre mediante proteínas transportadoras especiales acopladas a una fuente de energía, como la hidrólisis del ATP. Así bombean sustancias de un lugar a otro de la membrana. Estos sistemas son comunes en procariotas debido a que generalmente crecen en concentraciones bajas de nutrientes.
Generalmente las proteínas transportadoras son similares y forman 12 α-helices. Existen tres clases de transportes: Uniporte Simporte y Antiporte.

Transporte de solutos específicos
En la mayoría de los casos del transporte activo, que supone un trabajo osmótico, se realiza a expensas de un gradiente de H+, por hidrólisis de ATP o por otro componente rico en energía:
�Transporte simple
Tipo Moléculas transportadas
Uniporte K+, lisina
Simporte de protones β-galactósido, Glutamato, HPO42-, HSO4
-
Simporte de sodio Glutamato
Antiporte de sodio/protones Reciclaje de sodio
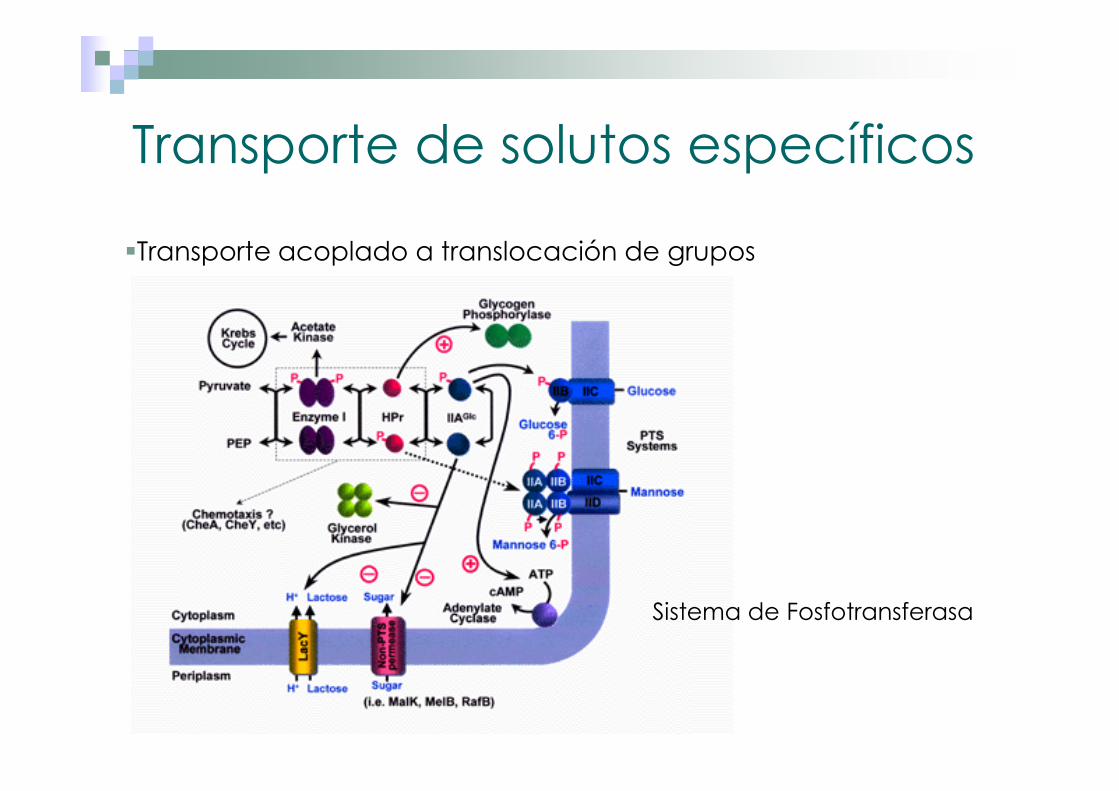
Transporte de solutos específicos
�Transporte acoplado a translocación de grupos
Sistema de Fosfotransferasa

Transporte de solutos específicos
�Transporte activo dirigido por ATP
Se denomina de transportadores ABC o ATPasas de tráfico, y se conocen muchos ejemplos en eubacterias y arqueas. Se trata de un sistema de varios componentes, en el que existen proteínas periplásmicas que captan el sustrato con gran afinidad, y lo llevan hasta unas proteínas de membrana, las cuales acoplan el paso de dicho sustrato hasta el citoplasma (sin alterarlo químicamente) con la hidrólisis de ATP.

Transporte de solutos específicos
�Porinas u otras proteínas de membrana externa para lograr la difusión del sustrato desde el medio hasta el espacio periplásmico, por ejemplo, en enterobacterias OmpF y OmpC (BtuB).�Proteína(s) solubles de espacio periplásmico que se unen al sustrato con gran afinidad (BtuF).�Un heterodímero formado por dos proteínas integrales de membrana por donde pasa el sustrato (BtuC).�Dos proteínas periféricas de membrana citoplásmica, en el lado citoplásmico, que acopla la hidrólisis de ATP con el transporte unidireccional del sustrato a través de la membrana (BtuD).Transporte de Vitamina B12
en Escherichia colihttp://moose.bio.ucalgary.ca/index.php?page=CK_Projects

Ensamblaje y transporte de proteínas extracitoplasmáticas�Translocación. Transporte de proteínas intra o a través de la membrana.�Exportación. Cuando la proteína es translocada al periplasma.�Secreción. Cuando la proteína es transportada al medio extracelular, dentro de otra célula o a la superficie celular.�Excreción. Transporte extracelular de moléculas que nos son de origen proteico (por ejemplo productos de la fermentación).

Sistemas de secreción.� Sistema Sec (General Secretory Pathway “GSP”). Sistema de
translocación y exportación de proteínas no plegadas.� Sistema Tat (Twin arginine translocation). Sistema de
translocación y exportación de proteínas plegadas.� Translocasa YidC. Sistema de translocación de proteínas de
Membrana Interna.
http://www.nature.com/nature/journal/v406/n6796/fig_tab/406575a0_F1.html

Sistema Sec�Péptido líder
�Proteína chaperona (SecB)
�Complejo de proteínas de unión (SecYEG)
�ATPasa citoplasmática (SecA)
�SecYEG + Sec A= translocasa. Es la responsable del movimiento de la proteína a través de la membrana citoplasmática.
�El complejo SecYE forma un canal conocido como translocón o canal conductor de proteínas (CCP).
�SecG, estimula el transporte y SecD, SecF y yajC son regulatorias.
�Péptido líder (secuencia líder o secuencia señal).
�Peptidasa del péptido señal (SPaseI).
General Secretory Pathway (GSP). Sistema de translocación y exportación de proteínas no plegadas.

Mecanismo del sistema Sec
1) La proteína chaperona SecB se une a una preproteína naciente en el citosol, estabilizando su conformación no plegada.2a) El complejo SecB-preproteína es dirigido hacia la translocasa SecYEG unida a SecA (3a).2b) El complejo SecB-preproteína se asocia a la proteína SecA y se dirige a SecYEG en la MI (3b). 4) Se une la secuencia señal a SecA, lo cual estabiliza la interacción SecB-SecA.
5) La preproteína se transfiere a SecA. La unión de ATP por SecA promueve el inicio de la translocación y la liberación de SecB del complejo ternario.La hidrólisis del ATP permite el paso de la proteína a través del translocón.
MENSAJE BIOQUÍMICO, Vol. XXVII (2003)
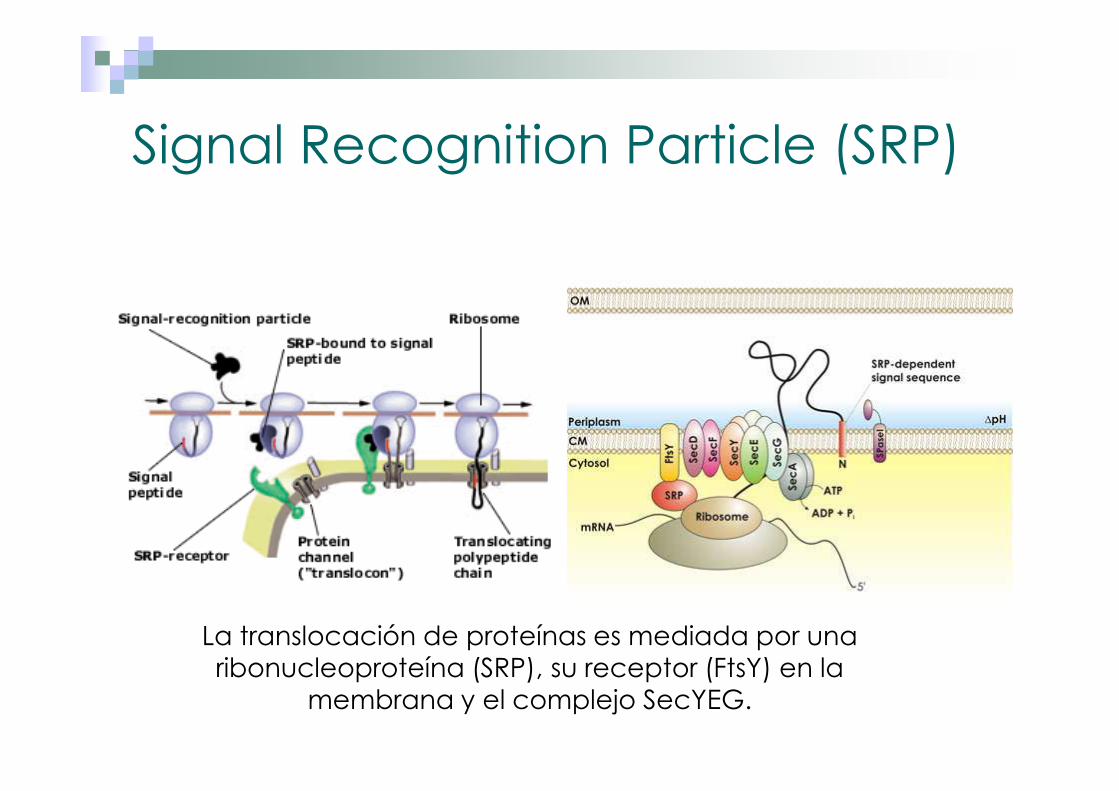
Signal Recognition Particle (SRP)
La translocación de proteínas es mediada por una ribonucleoproteína (SRP), su receptor (FtsY) en la
membrana y el complejo SecYEG.

Sistema Tat
Nature Reviews Molecular Cell Biology 2, 350-356, 2001.
Twin arginine translocation (Tat). Sistema de translocación y exportación de proteínas plegadas.
Se ha encontrado en:
�La membrana citoplasmática en bacterias y arqueas.
�Membranas tilakoides de los cloroplastos en plantas.
�Posiblemente en la membrana interna de la mitocondria de las plantas.

Sistema Tat
�Peptido señal (SRRXFLK) en el extremo amino terminal.�Tat translocasa: Proteínas membranales (Tat A, Tat B y Tat C).
Biochemical Society Transactions (2007) Volume 35, part 5

Aminoácido Código de tres letras Código de una letra
Alanina Ala A
Arginina Arg R
Asparagina Asn N
Ácido aspártico Asp D
Cisteína Cys C
Glutamina Gln Q
Ácido glutámico Glu E
Glicina Gly G
Histidina His H
Isoleucina Ile I
Leucina Leu L
Lisina Lys K
Metionina Met M
Fenilalanina Phe F
Prolina Pro P
Serina Ser S
Treonina Thr T
Triptófano Trp W
Tirosina Tyr Y
Valina Val V
Nomenclatura de
aminoácidos
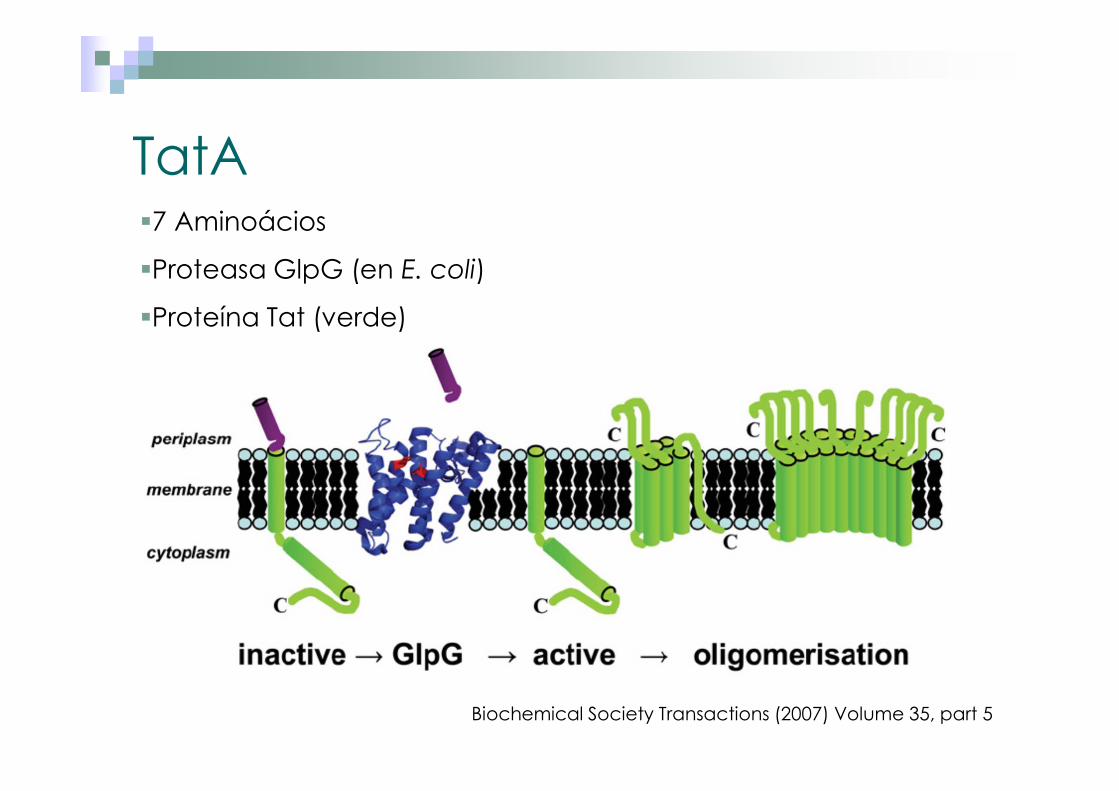
TatA�7 Aminoácios
�Proteasa GlpG (en E. coli)
�Proteína Tat (verde)
Biochemical Society Transactions (2007) Volume 35, part 5
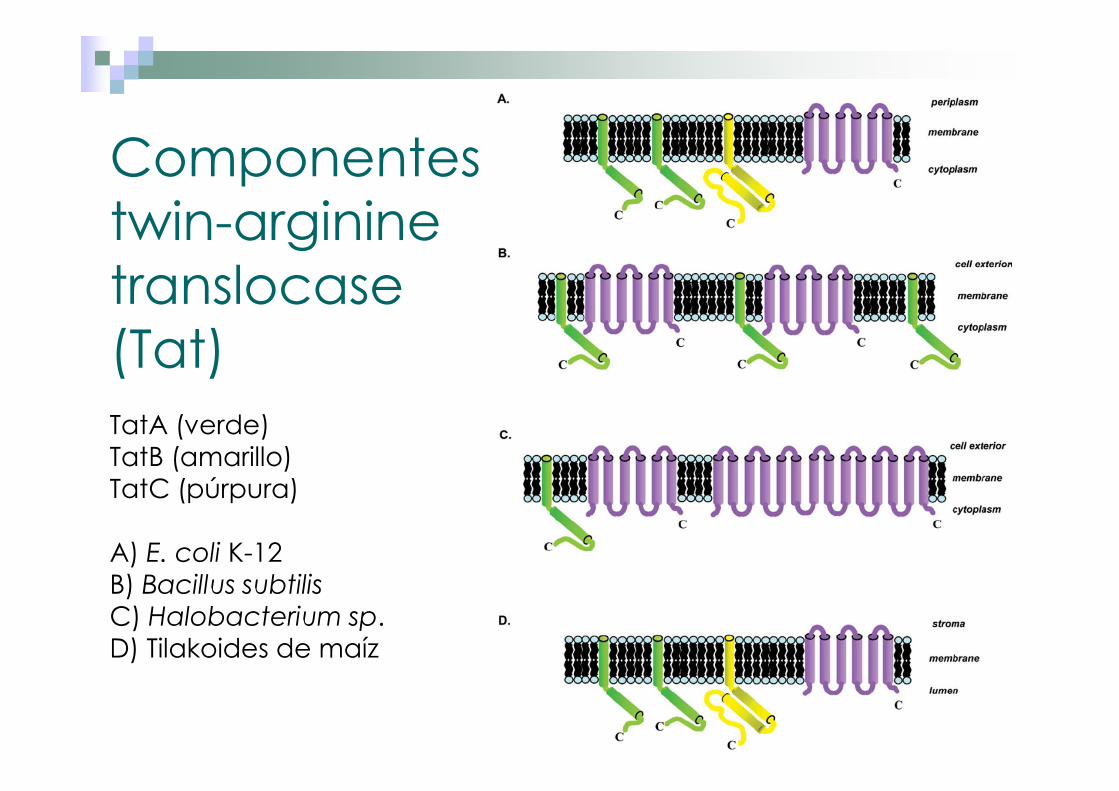
Componentes twin-argininetranslocase(Tat)TatA (verde)TatB (amarillo)TatC (púrpura)
A) E. coli K-12B) Bacillus subtilisC) Halobacterium sp. D) Tilakoides de maíz

Modelo Tat
Annu. Rev. Microbiol. 2006. 60:373–95

Translocasa YidC
EMBO reports 4, 10, 939–943 (2003)
Las proteínas de membrana interna (IMP) pueden translocarse a través de tres formas:
� Ruta de YidC.� Ruta SRP-YidC.� Ruta SRP-Sec translocón-
YidC.
Translocación de proteínas de Membrana Interna.

YidC insertasa dependiente e independiente de Sec
Nature Reviews Microbiology 6, 234-244 (March 2008)
La insertasa YidC tienen la función de promover la inserción de proteínas de membrana en conjunción con la translocasa Sec (o Sec-SRP) o independiente de ella.
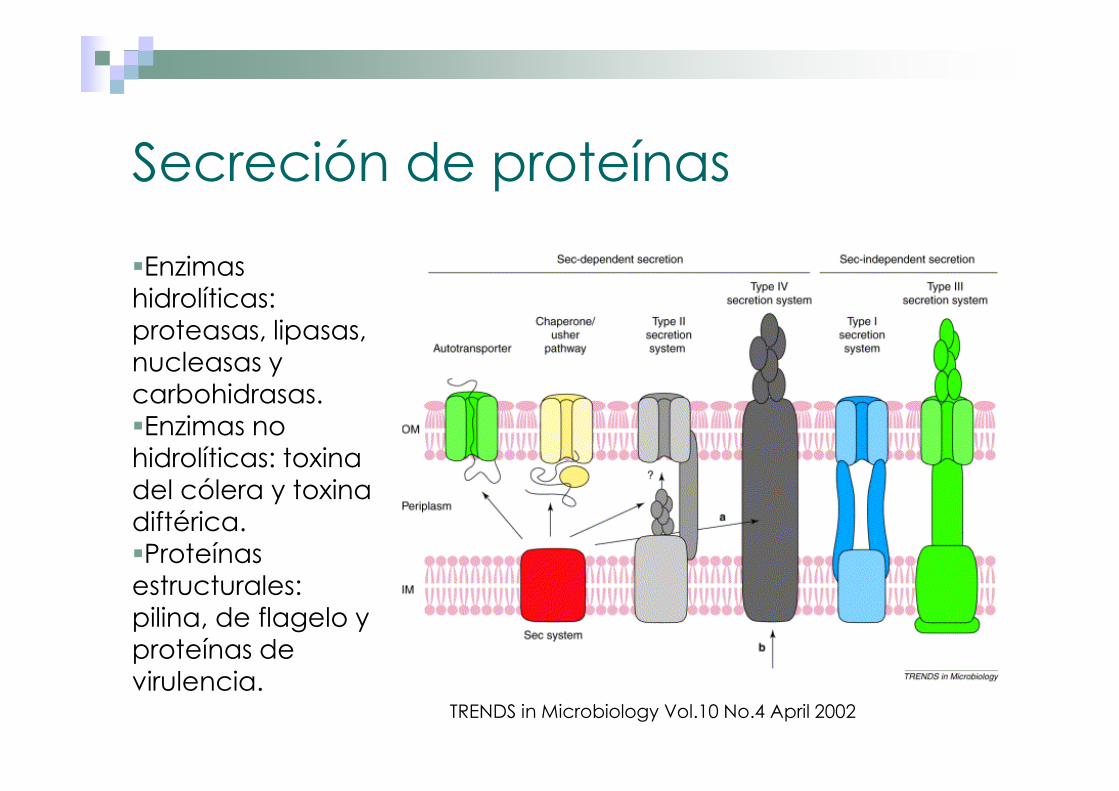
Secreción de proteínas
�Enzimas hidrolíticas: proteasas, lipasas, nucleasas y carbohidrasas.�Enzimas no hidrolíticas: toxina del cólera y toxina diftérica.�Proteínas estructurales: pilina, de flagelo y proteínas de virulencia.
TRENDS in Microbiology Vol.10 No.4 April 2002
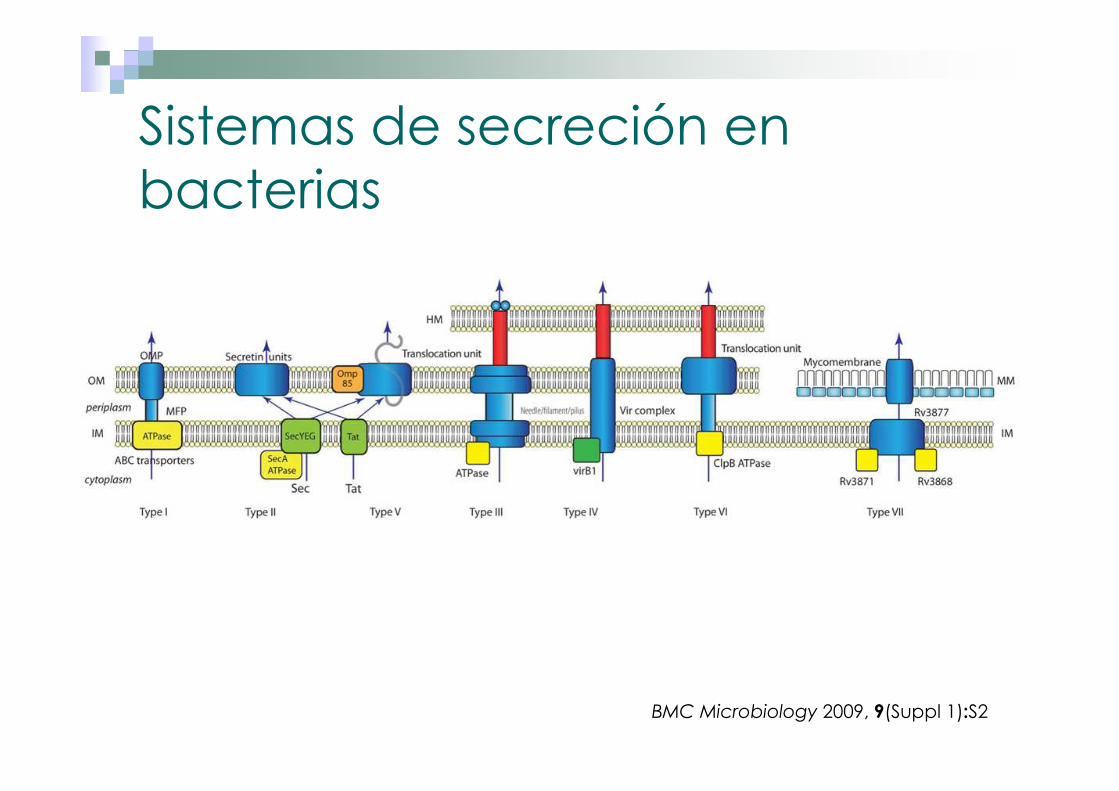
Sistemas de secreción en bacterias
BMC Microbiology 2009, 9(Suppl 1):S2

Segregación de cromosomas (probablemente).

Transporte de electrones y respiración.
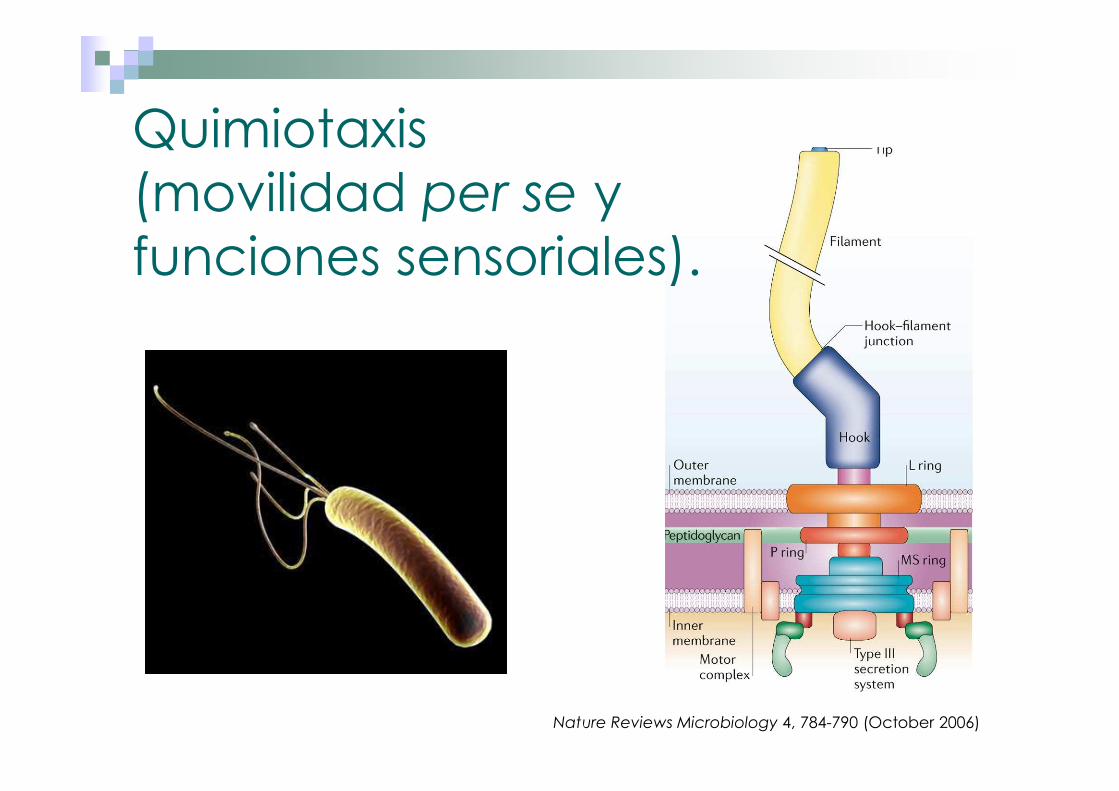
Quimiotaxis (movilidad per se y funciones sensoriales).
Nature Reviews Microbiology 4, 784-790 (October 2006)
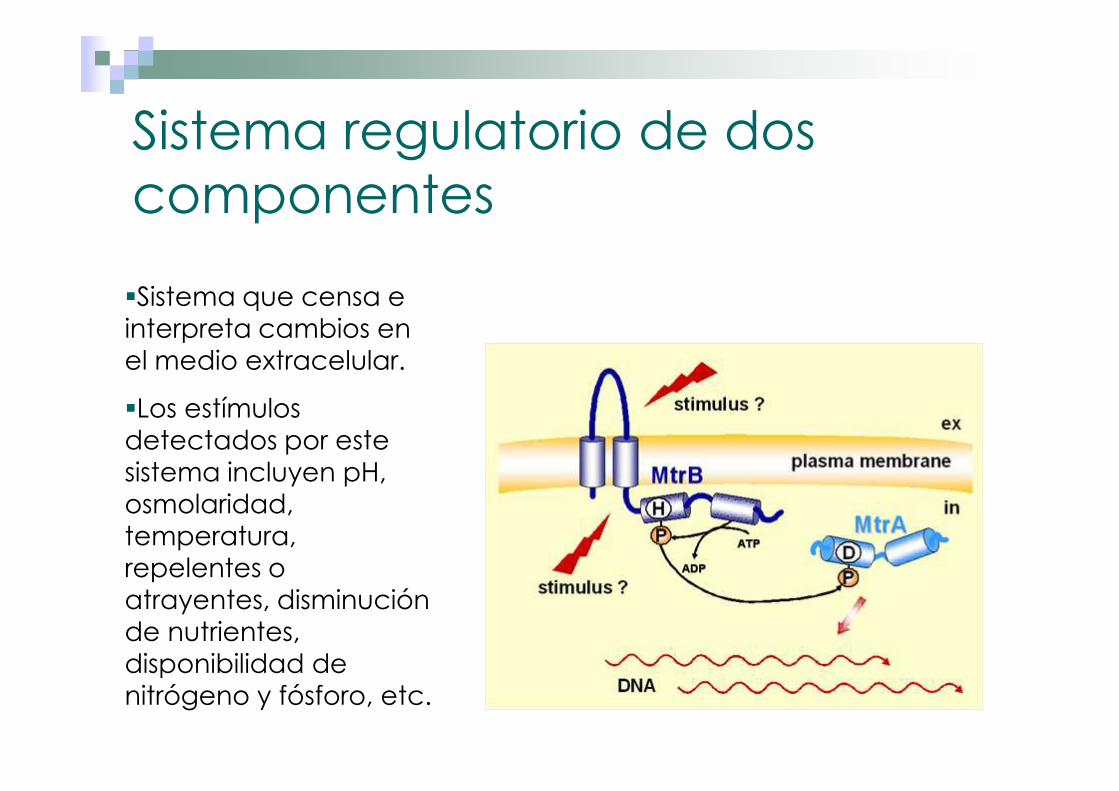
Sistema regulatorio de dos componentes
�Sistema que censa e interpreta cambios en el medio extracelular.
�Los estímulos detectados por este sistema incluyen pH, osmolaridad, temperatura, repelentes o atrayentes, disminución de nutrientes, disponibilidad de nitrógeno y fósforo, etc.

Modelo general del sistema de dos componentes
El estímulo no entra a la célula, sino que es detectada por un sensor a nivel de membrana, el cual transduce el estímulo hacia una proteína citoplásmica, regulador, la cual a su vez interacciona con secuencias determinadas al comienzo del operón, para regularlo, generando así la respuesta adaptativa correspondiente a la señal ambiental.
Moat, Microbial Physiology 2002