MODULACION DEL CALCIO INTRACELULAR POR INSULINA EN EL CARDIOMIOCITO DE...
70
Universidad de Chile Facultad de Ciencias Químicas y Farmacéuticas Departamento de Bioquímica y Biología Molecular Laboratorio de Transducción de Señales MODULACION DEL CALCIO INTRACELULAR POR INSULINA EN EL CARDIOMIOCITO DE RATA ADULTA Memoria para optar al título de Química Farmacéutica DEISY CAROLINA PIVET SILVA Profesor Patrocinante: Directores de Memoria: Dr. Sergio Lavandero G. Dr. Sergio Lavandero G. Dr. Ariel Contreras F. Santiago de Chile, 2010
Transcript of MODULACION DEL CALCIO INTRACELULAR POR INSULINA EN EL CARDIOMIOCITO DE...

Universidad de Chile
Facultad de Ciencias Químicas y Farmacéuticas
Departamento de Bioquímica y Biología Molecular
Laboratorio de Transducción de Señales
MODULACION DEL CALCIO INTRACELULAR POR
INSULINA EN EL CARDIOMIOCITO DE RATA ADULTA
Memoria para optar al título de Química Farmacéutica
DEISY CAROLINA PIVET SILVA
Profesor Patrocinante: Directores de Memoria:
Dr. Sergio Lavandero G. Dr. Sergio Lavandero G.
Dr. Ariel Contreras F.
Santiago de Chile, 2010

ii
AGRADECIMIENTOS
Primero quisiera agradecer a mis padres Mary y Eduardo,
quienes me han apoyado toda mi vida y sin duda han sido
pilares fundamentales en mi formación académica, gracias a
ustedes por el cariño, la preocupación y por confiar en mí
siempre. También a mi hermana Evelyn y su familia, mi cuñado
Ariel y mis adorables sobrinos Catalina y Alonso por ser parte
de mi vida y a la vez motivos de felicidad.
A Pedro por su continuo soporte desde el momento en que
nos conocimos cuando era yo mechona y su permanente
entrega de amor, indiscutiblemente eres muy importante en mi
vida. A su familia, en especial a sus padres Carmen y Rubén
por su preocupación y por integrarme como una más de
ustedes.
Al Profesor Sergio Lavandero por darme esta oportunidad de
conocer un mundo nuevo para mí y guiarme en este proceso y
por supuesto a todos los miembros del Laboratorio tanto de
Cardios como de Fibros por ayudarme en este arduo camino.
Particularmente quisiera agradecer a Fidel, Ruth, mi compañera
Coni y mi infaltable compañero de confocal David por hacer
más feliz la estadía, así como también a mi tutor Ariel quien me
enseñó gran parte de lo que aprendí. Gracias también al
Profesor Guillermo Díaz por su preocupación y consejos.
Muchas gracias a todos ustedes quienes formaron parte de
esta hermosa etapa de mi vida ya que sin vosotros todo
hubiese sido distinto.

iii
Esta memoria se realizó en el Laboratorio de Transducción de Señales del Centro
FONDAP Estudios Moleculares de la Célula y Departamento de Bioquímica y Biología
Molecular, Facultad Ciencias Químicas y Farmacéuticas de la Universidad de Chile y
contó con el financiamiento de los Proyectos FONDECYT 1080436 y FONDAP
1501006 (Dr. Sergio Lavandero).

iv
INDICE GENERAL
INDICE GENERAL ....................................................................................................... iv
INDICE DE FIGURAS ................................................................................................... vi
ABREVIATURAS ......................................................................................................... viii
RESUMEN ..................................................................................................................... x
SUMMARY ................................................................................................................... xii
1. INTRODUCCION ........................................................................................................... 1
1.1 Corazón y diabetes mellitus ............................................................................. 1
1.2 Características estructurales y funcionales de los cardiomiocitos adultos ........ 2
1.3 Insulina y su sistema transduccional ................................................................ 4
1.4 IGF-1 y su sistema transduccional ................................................................... 6
1.5 Acciones de Insulina en el corazón .................................................................. 7
1.6 Calcio ............................................................................................................... 9
1.7 Insulina y calcio .............................................................................................. 10
2. HIPOTESIS ................................................................................................................. 12
3. OBJETIVO GENERAL ................................................................................................. 12
4. OBJETIVOS ESPECIFICOS ........................................................................................ 12
5. MATERIALES Y METODOS ........................................................................................ 13
5.1 Reactivos ....................................................................................................... 13
5.2 Animales ........................................................................................................ 13
5.3 Obtención de cardiomiocitos de rata adulta .................................................... 14
5.4 Obtención de extractos proteicos para electroforesis ..................................... 15
5.5 Determinación de proteínas ........................................................................... 15
5.5.1 Método de Lowry ........................................................................................ 15
5.5.2 Método de Bradford .................................................................................... 16
5.6 Electroforesis en geles de poliacrilamida ........................................................ 16
5.7 Electrotransferencia de proteínas ................................................................... 16
5.8 Inmunowestern blot ........................................................................................ 16
5.9 Determinación de los niveles intracelulares de calcio ..................................... 17
5.10 Inmunofluorescencia indirecta ........................................................................ 18
5.11 Caracterización de cardiomiocitos adultos ..................................................... 18

v
5.11.1 Cardiomiocitos adultos para inmunowestern blot e Inmunofluorescencia ... 18
5.11.2 Cardiomiocitos adultos para medición de calcio por microscopía confocal . 19
5.12 Expresión de resultados y análisis estadístico ............................................... 21
6. RESULTADOS ............................................................................................................ 22
6.1 Activación de la vía de señalización canónica de
insulina en cardiomiocitos adultos .................................................................. 22
6.1.1 Efecto de insulina sobre la fosforilación del receptor de insulina ................ 22
6.1.2 Efecto de insulina sobre la fosforilación de Akt ........................................... 23
6.1.3 Efecto de insulina sobre la fosforilación de ERK 1/2 ................................... 23
6.2 Efecto de insulina en los niveles intracelulares de
calcio en el cardiomiocito adulto ..................................................................... 25
6.2.1 Efecto de insulina en los niveles de calcio intracelular de
cardiomiocitos adultos en ausencia de calcio externo ................................ 26
6.2.2 Efecto de insulina en los niveles de calcio intracelular y frecuencia de
contracción de cardiomiocitos adultos en presencia de calcio externo ....... 31
6.3 Efecto de IGF-1 sobre los niveles de calcio intracelular de
cardiomiocitos adultos .................................................................................... 37
6.3.1 Efecto de IGF-1 en los niveles de calcio intracelular de cardiomiocitos
adultos en ausencia de calcio externo ........................................................ 37
6.3.2 Efecto de IGF-1 en los niveles de calcio intracelular y frecuencia de
contracción de cardiomiocitos adultos en presencia de calcio externo ....... 37
6.3.3 Respuesta diferencial de cardiomiocitos adultos frente a IGF-1 1 µM ....... 41
6.4 Participación del calcio en la activación de la vía de
señalización del receptor de insulina .............................................................. 42
6.4.1 Participación del calcio en la fosforilación del receptor de insulina ............. 42
6.4.2 Participación del calcio en la fosforilación de Akt ........................................ 42
7. DISCUSION ................................................................................................................. 44
8. CONCLUSIONES ........................................................................................................ 49
9. BIBLIOGRAFIA ............................................................................................................ 50

vi
INDICE DE FIGURAS
Figura 1. Caracterización de los cardiomiocitos adultos.............................................. 20
Figura 2. Insulina estimula la fosforilación del receptor de insulina (RI) y
de las proteínas kinasas Akt y ERK2 en el cardiomiocito adulto .................. 24
Figura 3. Identificación de zonas de análisis (ROI) en el cardiomiocito adulto ............ 25
Figura 4. Efecto de insulina sobre la liberación de calcio desde reservorios
intracelulares en el cardiomiocito adulto evaluado en el citoplasma celular . 28
Figura 5. Efecto de insulina sobre la liberación de calcio desde reservorios
intracelulares en el cardiomiocito adulto evaluado en el núcleo celular ....... 29
Figura 6. Efecto de insulina sobre la liberación de calcio desde reservorios
intracelulares en el cardiomiocito adulto evaluado en la membrana celular . 30
Figura 7. Efecto de insulina sobre los niveles de calcio intracelular en
presencia de calcio externo evaluado en el citoplasma celular .................... 32
Figura 8. Efecto de insulina sobre los niveles de calcio intracelular en
presencia de calcio externo evaluado en el núcleo celular .......................... 33
Figura 9. Efecto de insulina sobre los niveles de calcio intracelular en
presencia de calcio externo evaluado en la membrana celular .................... 34
Figura 10. Efecto de insulina sobre la frecuencia de contracción de
cardiomiocitos de rata adulta ....................................................................... 36
Figura 11. Experimento que resume el efecto de insulina sobre los niveles
de calcio intracelular .................................................................................... 36

vii
Figura 12. Efecto de IGF-1 sobre la liberación de calcio desde reservorios
intracelulares en el cardiomiocito adulto ...................................................... 39
Figura 13. Efecto de IGF-1 sobre el calcio intracelular en presencia de
calcio externo y la frecuencia de contracción............................................... 40
Figura 14. Respuesta diferencial de cardiomiocitos adultos frente a IGF-1 1 µM ......... 41
Figura 15. Participación del calcio en la fosforilación del receptor de insulina
y proteína kinasa Akt ................................................................................... 43

viii
ABREVIATURAS
µg = Microgramo
µL = Microlitro
µm = Micrómetro
µM = Micromolar
ADP = Adenosina difosfato
ATP = Adenosina trifosfato
BAPTA-AM = BAPTA-acetometil éster
BDM = 2,3 butanodiona monoxima
BSA = Seroalbúmina de bovino
Ca2+ = Calcio
CICR = Liberación de calcio inducida por calcio
DHP = Dihidropiridina
DTT = Ditiotreitol
ECC = Acoplamiento excitación contracción
EDTA = Ácido etilendiaminotetraacético
EGTA = Ácido etilenglicoltetraacético
ERK = Proteína kinasa regulada extracelularmente
Fluo-3 AM = Fluo-3 acetometil éster
g = Gramo
GH = Hormona de crecimiento
h = Hora
HEPES = ÁcidoN-(2-hidroxietil)piperazina-N’-(2-etanosulfónico)
IGF-1 = Factor de crecimiento análogo a insulina tipo 1
IGFBP = Proteínas transportadoras de IGF-1
IGF-1 R = Receptor de IGF-1
Ins = Insulina
Iono = Ionomicina
IP3 = inositol-1,4,5-trifosfato
IP3R = Receptor de inositol 1,4,5-trifosfato
IRS = Primer sustrato del receptor de insulina
Iso = Isoproterenol
kDa = Kilo Dalton

ix
Kd = Constante de disociación
M199 = Medio 199
mA = Miliamperes
MAPK = Proteína kinasa activada por mitógenos
MEC = Matriz extra celular
mg = Milígramo
min = Minuto
mL = Mililitro
mM = Milimolar
NADH = Nicotinamida adenina dinucleótido reducido
nm = Nanómetro
nM = Nanomolar
N.S. = No significativo
PBS = Tampón fosfato salino
PDK = Kinasa de la piruvato deshidrogenasa
PI3K = Fosfatidil inositol 3-Kinasa
PIP2 = Fosfatidil inositol 4,5- bifosfato
PIP3 = Fosfatidil inositol 3,4,5- trifosfato
PKA = Proteína kinasa A
PKB = Proteína kinasa B (Akt)
aPKC = Proteína kinasa C atípica
PLC = Fosfolipasa C
PMSF = Fenilmetanosulfonil fluoruro
RI = Receptor de Insulina
ROI = Región óptica de interés
rpm = Revoluciones por minuto
RyR = Receptor de ryanodina
SDS = Dodecil sulfato de sodio
SEM = Error estándar de la media
Túbulo T = Túbulo transverso
TIRF = Fluorescencia de reflexión total interna
V = Volt
VLDL = Lipoproteina de muy baja densidad
Zn2+ = ión Zinc

x
RESUMEN
El cardiomiocito adulto es una célula terminalmente diferenciada con escasa
capacidad replicativa y responsable de la contracción del miocardio. El ión calcio en
conjunto con el AMP cíclico son los segundos mensajeros que regulan la contracción
del cardiomiocito. Por otra parte la diabetes mellitus tipo II se asocia a un mayor riesgo
cardiovascular (“cardiomiopatía diabética”), siendo muy frecuentes alteraciones
sistólicas y diastólicas en el corazón de un paciente diabético. Dado que insulina es la
principal hormona asociada a la diabetes es importante determinar si existe alguna
relación entre ella y el calcio en el cardiomiocito adulto. En una investigación previa de
nuestro Laboratorio se describió que insulina aumenta el calcio intracelular en cultivos
primarios de cardiomiocitos neonatos, presentando esta respuesta dos componentes,
uno rápido dependiente del calcio externo y el canal tipo L y receptor de ryanodina y
otro lento dependiente de la vía transduccional IP3/receptor IP3. Interesantemente, este
último componente media los efectos de insulina a nivel de la translocación del GLUT4
a la membrana plasmática y el posterior ingreso de glucosa a estas células. En base a
estos antecedentes se propuso en esta memoria como hipótesis que insulina también
aumenta los niveles intracelulares de Ca2+ en el cardiomiocito adulto a través de un
mecanismo dependiente del Ca2+ extracelular y del almacenado en reservorios
intracelulares.
Nuestro objetivo consistió en estudiar la activación del sistema de señalización río
abajo del receptor de insulina y su dependencia del calcio y el efecto tanto de insulina
como IGF-1 en los niveles intracelulares de calcio en el cardiomiocito adulto de rata.
Las células se expusieron a insulina 100 nM por distintos períodos de tiempo y se
evaluó la activación del receptor mediante su autofosforilación en tirosina y de las
proteínas kinasas Akt (Ser 473) y Erk 1/2 mediante Western blot. La presencia del
receptor de insulina en el cardiomiocito de rata adulta se detectó por
inmunofluorescencia en tyr1150/1151. Las tres proteínas evaluadas alcanzaron un
máximo de activación significativo con respecto a los controles a los 5 min post-
estímulo con insulina. Por otra parte, las células preincubadas con la sonda
fluorescente para calcio Fluo3- AM se estudiaron por microscopía confocal para
determinar cambios en los niveles de calcio intracelulares inducidos por insulina e IGF-

xi
1. Ambos péptidos no produjeron aumentos significativos en los niveles de calcio
intracelulares tanto en medio de reposo sin calcio como en medio de reposo con calcio
y evaluando distintas zonas celulares y concentraciones de los compuestos, mientras
que conocidos estímulos (ionomicina y KCl), empleados como controles positivos,
aumentaron el calcio intracelular. Asimismo, insulina e IGF-1 no modificaron la
frecuencia de contracción del cardiomiocito adulto. Sin embargo, ensayos de
fosforilación de la proteína Akt (Ser 473) en ausencia de calcio intracelular mediante el
uso de BAPTA-AM mostraron niveles inferiores a los observados en presencia de este
catión. En cambio, la fosforilación del receptor de insulina no se modificó al quelar el
calcio intracelular en el cardiomiocito adulto.
Estos resultados sugieren que probablemente las diferencias en cuanto a fenotipo y
genotipo, así como las diferencias en las preferencias de sustratos energéticos de
cardiomiocitos neonatos y adultos podrían constituir razones por las cuales ambos
tipos celulares se comportan de manera distinta en cuanto a la relación insulina y
calcio.
En conclusión, insulina activa su vía transduccional en el cardiomiocito adulto. No
obstante, ni insulina ni IGF-1 inducen cambios en los niveles intracelulares de calcio en
el cardiomiocito adulto ya sea afectando la entrada desde el extracelular o la salida
desde reservorios intracelulares de manera detectable con Fluo3- AM. Sin embargo,
dado que este catión moduló la activación de la vía de señalización de insulina a nivel
de la proteína kinasa Akt, futuras investigaciones deberán aclarar estas discrepancias.

xii
SUMMARY
Intracellular calcium modulation by insulin in adult rat cardiomyocytes
Cardiomyocytes are fully differentiated cells with limited proliferation capacity and
responsible for myocardial contraction. Ca2+ and cyclic AMP are second messengers
regulating cardiomyocyte contraction. On the other hand type II diabetes mellitus has
been associated with increased cardiovascular risk (diabetic cardiomyopathy), being
frequent systolic and diastolic alterations in the diabetic heart. Because insulin is the
main hormone linked to diabetes, it is important to determine whether there is any
relationship between this peptide hormone and calcium in the adult cardiomyocyte. In a
previous work in our laboratory, we reported that insulin increased intracellular Ca2+ in
primary cultured neonatal cardiomyocytes. This response has two components, a rapid
one characterized by its dependence by external Ca2+ and the L-type Ca2+
channel/ryanodine receptor, and a slow component depending on IP3/IP3 receptor
signaling pathway. Interestingly the latter component mediates insulin effects on GLUT4
translocation to the plasma membrane and subsequent glucose uptake. Based on this
evidence, we propose that insulin also increases intracellular Ca2+ levels in isolated
adult rat cardiomyocytes through a mechanism involving extracellular and intracellular
Ca2+.
The aim of this work was to study the activation of insulin receptor downstream
signaling pathway and its regulation by Ca2+ as well as the effects of insulin and IGF-1
on the intracellular Ca2+ levels in isolated adult rat cardiomyocytes.
To this end, cells were exposed to 100 nM insulin for different time periods and the
activation of the insulin receptor was assessed by its tyrosine autophosphorylation and
the phosphorylation of protein kinases Akt (Ser 473) and Erk 1/2 by Western Blot. The
presence of tyr1150/1151 insulin receptor in the adult rat cardiomyocyte was
investigated by indirect immunofluorescence. These three signaling proteins reached a
maximum activation at 5 min after insulin stimulation. On the other hand, cells
preincubated with the fluorescent probe for calcium Fluo3-AM were studied by confocal
microscopy to determine changes on intracellular Ca2+ levels induced by insulin and
IGF-1. Both peptides did not produce any significant increases in intracellular calcium

xiii
levels both in presence and absence of extracellular calcium and evaluating different
cell areas and concentrations of the compounds. Stimuli like ionomycin and KCl, used
as positive controls, increased intracellular Ca2+ levels in cardiac cells. Furthermore,
insulin and IGF-1 did not affect the frequency of adult cardiomyocyte contraction.
However, Akt phosphorylation on Ser 473 was lower in cells cultured with the
intracellular calcium chelator BAPTA-AM that those cultured with medium containing
this cation. In contrast, insulin receptor phosphorylation was not modified by BAPTA-
AM on isolated adult rat cardiomyocytes.
These results suggest that probably the differences in phenotype and genotype, as
well as differences in energy substrate preferences by neonatal and adult
cardiomyocytes could explain why both cell types behave differently with regard to the
relationship between insulin and calcium.
In summary, insulin activates its canonical signaling pathway in adult
cardiomyocytes. However, neither insulin nor IGF-1 changed intracellular Ca2+ levels in
isolated rat adult cardiomyocytes. However Ca2+ modulated the activation of the
canonical insulin receptor signaling pathway at the level of Akt and future research
should clarify these discrepancies.

1
1. INTRODUCCION
1.1. Corazón y diabetes mellitus
El sistema cardiovascular posee dos principales componentes: el sistema
circulatorio compuesto por vasos sanguíneos que distribuyen la sangre a través de
todo el cuerpo y tejidos y el corazón como órgano central. Este último está situado
entre los pulmones en el centro del pecho y es una bomba muscular que expulsa la
sangre hacia las arterias con el fin de entregar a las células oxígeno y nutrientes y a su
vez retirar los desechos producidos en el metabolismo (1).
Por otro lado, en Chile la prevalencia global de diabetes de acuerdo a la Encuesta
Nacional de Salud de 2003 fue de 4,2%, 4,8% en hombres y 3,8% en mujeres. Sin
embargo, este valor se ve aumentado en el rango etario de 45-64 años a 9,4% y a
15,2% en mayores de 65 años (2). En consecuencia, esta enfermedad se ha
posicionado en la sociedad con una alta prevalencia contra la cual la lucha cada día se
hace más ardua.
La diabetes mellitus es un factor de riesgo conocido para la insuficiencia cardiaca
congestiva. Se piensa que este riesgo está asociado a enfermedades en arterias
coronarias y sus complicaciones, especialmente aterosclerosis (3). Al momento del
diagnóstico de diabetes tipo 2, es común que los pacientes presenten manifestaciones
de aterosclerosis. La principal causa de insuficiencia cardiaca congestiva es la
enfermedad cardiaca isquémica a pesar de los avances en la prevención y en el
tratamiento; por lo tanto diabetes e insuficiencia cardiaca están relacionadas (4). La
diabetes mellitus también puede afectar la función y estructura cardiaca en ausencia de
cambios en la presión sanguínea y de enfermedad de arterias coronarias, esta
condición se denomina cardiomiopatía diabética (5).
La cardiomiopatía diabética es una condición clínica diagnosticada cuando se
desarrolla disfunción ventricular diastólica en pacientes con diabetes y en ausencia de
aterosclerosis e hipertensión arterial (6). Algunas alteraciones del corazón diabético
incluyen hipertrofia de cardiomiocitos, fibrosis intersticial y microangiopatía
intramiocardica (4). A su vez, en pacientes con diabetes mellitus se observa una

2
disminución en la contractilidad cardiaca, esta alteración puede ser explicada por
diversos mecanismos, entre ellos están daño en la homeostasis del calcio,
desregulación del sistema renina-angiotensina, aumento del estrés oxidativo,
metabolismo alterado y disfunción mitocondrial (5).
1.2. Características estructurales y funcionales de los cardiomiocitos adultos
Dos tercios de corazón adulto está compuesto por células no musculares
principalmente fibroblastos. Estos fibroblastos cardiacos están inmersos dentro de la
matriz extracelular (MEC) del tejido conectivo y son responsables de su síntesis. Bajo
condiciones fisiológicas, los fibroblastos proveen un soporte mecánico para los
cardiomiocitos y participan en la coordinación de la función contráctil del corazón (7).
Además de secretar componentes de la matriz extracelular, los fibroblastos cardiacos
producen una serie de factores de crecimiento que probablemente median una
interacción entre fibroblastos y cardiomiocitos (8). Estrés externo, como por ejemplo
una herida, produce un cambio en el fenotipo de los fibroblastos cardiacos a otro tipo
celular conocido como miofibroblasto (9).
Aproximadamente un tercio del corazón está compuesto por células musculares
responsables de la contracción de este órgano, los cardiomiocitos. No obstante,
existen numerosas diferencias entre los tipos celulares de cardiomiocitos adultos y
neonatos o fetales que apuntan a satisfacer las necesidades del corazón propias de la
etapa de desarrollo en que se encuentra. Es así como los cardiomiocitos adultos de
mamíferos han experimentados cambios para llegar a un fenotipo adulto diferenciado.
Esto incluye disminución de la expresión de genes fetales y aumento de la expresión
de genes responsables del fenotipo adulto. Estos cambios permiten conseguir una
diferenciación de la estructura del cardiomiocito para suplir las necesidades del
corazón que avanzan hacia una mayor presión sistólica luego del nacimiento. Por lo
tanto, se produce un cambio en las isoformas y tipos de proteínas relacionadas con la
estructura y la contractilidad cardiaca, originándose una sarcomerización más
desarrollada y compacta que en la edad neonata. Junto con esta diferenciación se
produce una diferenciación terminal cuando además los cardiomiocitos pierden su
capacidad de dividirse (10).
Otra de las diferencias dice relación con el acoplamiento excitación- contracción
(ECC) de los cardiomiocitos. El acoplamiento excitación- contracción es la base de la

3
función cardiaca a nivel celular. Este proceso involucra varios componentes que
comunican la excitación eléctrica de la membrana celular a la generación de señales
de calcio citosólicas que gatillan la contracción celular (11).
En ambos casos, cardiomiocitos adultos y neonatos, se ha visto que al remover el
calcio del medio extracelular de los cardiomiocitos cesa la contracción de los mismos,
por lo tanto el influjo de calcio desde el extracelular sería indispensable para la
contracción del músculo cardiaco. El calcio entra a los cardiomiocitos principalmente a
través de los canales de calcio tipo L sensibles a dihidropiridinas (DHP). Sin embargo,
en células cardiacas adultas, la mayor parte del calcio responsable de la contracción
proviene del almacenamiento interno en el retículo sarcoplásmico (12). Al parecer, a
diferencia de los cardiomiocitos adultos, los neonatos carecerían de sistema de túbulos
T el cual juega un rol central en el acoplamiento excitación- contracción de
cardiomiocitos ventriculares adultos y por lo tanto las células cardiacas neonatas
tendrían la capacidad de mantener la señalización de calcio citosólico sin liberación de
calcio desde el retículo sarcoplásmico produciéndose señalizaciones más
heterogéneas (11).
En cardiomiocitos, la contracción se inicia por la apertura de los canales tipo L
sensibles a voltaje, los cuales son más abundantes en los túbulos transversos (túbulos
T). El influjo de calcio producido a partir de esta apertura provoca que se abra el
receptor de Ryanodina (RyR) que a su vez es canal de calcio, liberándose el calcio
desde el interior del retículo sarcoplásmico hacia el citoplasma celular. Este proceso es
conocido como liberación de calcio inducida por calcio (del inglés: “calcium induced-
calcium release”, CICR). Posteriormente este calcio se adosa a la maquinaria contráctil
generándose la contracción celular. La interacción entre el canal de calcio tipo L y el
receptor de Ryanodina estaría dada por la existencia de los túbulos T (13).
Posterior al nacimiento, el corazón requiere aumentar el gasto cardíaco para ser
capaz de operar contra una resistencia vascular aumentada y de adaptarse a
variaciones en la tasa metabólica total y al suministro de sus requerimientos de
sustratos energéticos vitales (14). Es así como el corazón de mamíferos posterior al
nacimiento utiliza ácidos grasos de cadena larga como principal sustrato para la
producción de ATP, mientras que el corazón fetal, que funciona en un ambiente
relativamente hipóxico, obtiene su energía a partir del catabolismo de glucosa y lactato
dado que la producción glicolítica de ATP es más eficiente en cuanto al oxígeno que la

4
oxidación de ácidos grasos. Los cambios en el suministro de sustratos energéticos al
corazón, los cambios en el control hormonal del metabolismo energético en el corazón
y los cambios subcelulares dentro del miocardio son los responsables de las
diferencias en las preferencias energéticas del corazón después del nacimiento (15).
1.3. Insulina y su sistema transduccional
Insulina es un péptido de 51 aminoácidos secretado por las células β del páncreas y
la principal hormona que participa en la captación de glucosa y la homeostasis de la
misma. El monómero de insulina maduro está compuesto por dos cadenas, una
cadena A (de 21 aminoácidos) y una cadena B (de 30 aminoácidos) unidas por 3
puentes disulfuro (16). El proceso de liberación de insulina comienza con las células β
pancreáticas sensando los niveles de glucosa sanguínea elevados e introduciéndola al
interior de las mismas a través de un transportador de glucosa de alta capacidad y baja
afinidad, GLUT-2. Una vez al interior de las células la glucosa es fosforilada a glucosa-
6-fosfato por una glucokinasa y posteriormente es metabolizada a través de la
glicólisis obteniéndose como producto piruvato, NADH y ATP (17). Los procesos
oxidativos derivados del ingreso de glucosa a las células β-pancreáticas se traducen en
mayor producción de ATP en la mitocondria y por lo tanto una razón ATP/ADP mayor
en el citoplasma. El aumento de esta razón cierra los canales de potasio sensibles a
ATP produciéndose una disminución en el flujo de salida hiperpolarizante de potasio
conduciendo a una despolarización de la membrana plasmática, un influjo de calcio
extracelular a través de los canales de calcio dependientes de voltaje y en
consecuencia a un gran aumento en los niveles de calcio intracelular que gatilla la
exocitosis de las vesículas que contienen la insulina almacenada (18) en forma de
hexámeros. Estos hexámeros están compuestos por 3 dímeros unidos entre sí a través
de dos iones de Zn2+ que se encuentran en el centro y a su vez, los monómeros que
conforman los dímeros están unidos a través de puentes de hidrógeno en sus cadenas
principales. Sin embargo, la insulina debe disociarse a monómero para actuar sobre su
receptor (16).
El receptor de insulina (RI) pertenece a la superfamilia de los receptores con
actividad tirosina kinasa intrínseca. A diferencia de otros integrantes de esta familia, el
RI completo es una glicoproteína heterotetramérica compuesto por dos subunidades α
y dos subunidades β unidas entre sí por puentes disulfuro de la forma (αβ)2. La
subunidad α del RI es de 135 kDa y de ubicación extracelular, mientras que la

5
subunidad β pesa 95 kDa y contiene una porción extracelular, una secuencia
transmembrana y un dominio tirosina kinasa intracelular (19). Insulina se une a su
receptor a través de la subunidad extracelular α del mismo. Esta unión produce un
acercamiento de las dos subunidades α con lo cual se produce un cambio
conformacional que permite que el ATP se una al dominio intracelular de la subunidad
β. Esta unión activa la autofosforilación del receptor que a su vez activa la actividad
kinasa del mismo hacia sustratos proteicos intracelulares (20). Los primeros sustratos
proteicos en ser fosforilados son las proteínas sustratos del receptor de insulina (IRSs)
incluyendo IRS-1, IRS-2, IRS-3, IRS-4, Gab-1 y Shc, que están ligadas a la activación
de dos principales vías de señalización: la vía de la fosfatidil inositol 3- kinasa (PI3K) –
Akt/ proteína kinasa B (PKB), la cual es responsable de la mayoría de las acciones
metabólicas de insulina, y la vía de las MAPK (Ras- mitogen activated protein kinase),
que regula la expresión de algunos genes y junto con la vía PI3K controlan el
crecimiento celular y la diferenciación (21).
La unión de IRSs a la subunidad regulatoria de PI3K resulta en su activación, la cual
fosforila al fosfatidil inositol 4,5- bifosfato (PIP2) en la posición tres convirtiéndolo en
fosfatidil inositol 3,4,5- trifosfato (PIP3). Este fosfolípido activa las proteínas kinasas
PDK1 y PDK2, resultando en la activación de la proteína Akt/PKB y una proteína
kinasa C atípica (PKC λ y PKC ζ). La proteína kinasa B (Akt) activada fosforila al
sustrato de 160 kDa (AS160), el cual participa en la translocación de vesículas de
GLUT4 mediada por insulina hacia la membrana plasmática. La activación de aPKC
también está involucrada en esta translocación en respuesta a insulina (22).
Además de la vía de señalización de insulina dependiente de PI3K, otra rama
importante de esta vía compromete a la proteína Shc. Shc se une al receptor de
insulina activado y es fosforilada por un mecanismo similar a IRS-1, luego une una
proteína adaptadora, Grb2 (23). Esta unión resulta en la activación del factor de
intercambio de GTP, Sos que a su vez activa la GTPasa pequeña Ras, que recluta la
kinasa serina/treonina Raf a la membrana donde es completamente activada.
Posteriormente la señal es amplificada a través de dos kinasas río abajo: MAP kinasa
kinasa, MAPKK (MEK o ERK kinasa) y las kinasas reguladas extracelularmente, ERK
1/2, las que son únicamente reguladas por fosforilación. MEK es fosforilada en dos
residuos de serina por Raf y luego las ERK son fosforiladas en un residuo de tirosina y

6
en otro de treonina por MEK (24). Esta rama dependiente de MAPK de la señalización
de insulina modula acciones tales como crecimiento, sobrevida y diferenciación (25).
También se ha visto que la proteína c-Cbl participa de manera relevante en la
acción de insulina. Esta función estaría regulada por dos proteínas adaptadoras, APS
(adapter containing PH and SH2 domains) y CAP (Cbl-associated protein). CAP
interactúa constitutivamente con Cbl a través de su dominio SH3 C- terminal. Luego de
la estimulación, el receptor de insulina cataliza la fosforilación de APS en tirosina. Una
vez fosforilada, APS recluta a c-Cbl hacia el receptor de insulina para posteriores
fosforilaciones en tirosina. Luego de la fosforilación de Cbl, el complejo CAP/Cbl migra
hacia la membrana plasmática, esto conduce al reclutamiento del complejo Crk/C3G
hacia la membrana donde C3G, un factor de intercambio de GTP activa a la proteína G
pequeña TC10. La activación de TC10 ocurre de manera independiente de la vía PI3K
y parece colaborar en la translocación de GLUT4 hacia la membrana estimulada por
insulina (26).
1.4. IGF-1 y su sistema transduccional
IGF-1 es un péptido de cadena simple de 70 aminoácidos estructuralmente
homólogo a la pro-insulina. Es sintetizado principalmente en el hígado y el riñón
aunque la producción de manera local en otros tejidos pareciera ser importante para
mediar, a través de mecanismos paracrinos o autocrinos, los efectos anabólicos y de
promoción del crecimiento de la hormona del crecimiento (GH). IGF-1 circula unido a
proteínas transportadoras (IGFBPs), las cuales sirven además para prolongar su vida
media, modular la especificidad de tejidos y potenciar o neutralizar sus acciones
biológicas (27). El receptor de IGF-1 (IGF-1 R) y el receptor de insulina (RI) comparten
un alto grado de identidad en sus estructuras. Al igual que el receptor de insulina, el
receptor de IGF-1 está compuesto de dos subunidades α extracelulares donde se une
el ligando y dos subunidades β transmembrana que poseen actividad tirosina kinasa
intrínseca. El mayor grado de homología entre el receptor de insulina y el receptor de
IGF-1 se encuentra en el dominio transmembrana con actividad tirosina kinasa
intrínseca (subunidades β) con alrededor de 84% de homología, mientras que el mayor
grado de diferencia se encuentra en los dominios C- terminales, donde comparten
alrededor de un 44 % de identidad (28).

7
La unión de IGF-1 a su receptor promueve la dimerización del receptor y la
autofosforilación en residuos de tirosina, conduciendo a la posterior fosforilación de
proteínas tales como las proteínas sustratos del receptor de insulina (IRSs) y el
acoplamiento de proteínas que contienen homología Src (Shc) (29), transduciendo
señales a efectores río abajo. A través de estos efectores, IGF-1 activa dos cascadas
de señalización principales, la vía Ras- Raf- MEK- ERK y la vía PI3K- Akt/PKB (30).
1.5 Acciones de insulina en el corazón
Los principales blancos de insulina son el músculo esquelético, el tejido adiposo y el
hígado, aquí la insulina participa en la regulación de etapas críticas en el metabolismo
intermediario. Sin embargo, muchos otros tejidos expresan receptores de insulina y por
lo tanto sus funciones podrían estar reguladas por este péptido. Entre las acciones de
insulina en el corazón podemos distinguir algunas indirectas y otras directas. Insulina
aumenta la síntesis de proteínas y suprime su degradación en hígado y músculo
esquelético contribuyendo a la economía de carbohidratos y nitrógeno. A su vez afecta
la degradación de triglicéridos en el tejido adiposo, aumentando la disponibilidad de
ácidos grasos que pueden ser sustratos de la β-oxidación (31). Estos efectos
constituyen acciones indirectas.
Insulina también posee acciones directas sobre el metabolismo cardíaco como por
ejemplo en la captación de glucosa, el metabolismo de ácidos grasos, y la glicólisis. En
cuanto a la captación de glucosa se sabe que se requiere insulina para promover la
translocación del transportador de glucosa más distribuido en el corazón, GLUT4 hacia
la membrana celular desde compartimientos intracelulares a través de tráfico de
vesículas (32). No obstante, insulina no es el único estímulo que induce captación de
glucosa sino también lo hacen la contracción del tejido cardíaco (33) y el ejercicio
aunque de distintas maneras (34). Más aún, insulina estimula la captación de glucosa a
través de la promoción de la expresión génica de GLUT (32).
En ausencia de estímulo, los transportadores GLUT-4 permanecen en
compartimientos intracelulares con una combinación de exocitosis lenta y endocitosis
rápida (35). Insulina promueve la translocación de GLUT-4 a la membrana celular a
través de la estimulación de la exocitosis y también disminuye la endocitosis. El efecto

8
de estimulación de insulina sobre la captación de glucosa alcanza 2 a 14 veces en los
cardiomiocitos y ha sido atribuida principalmente a la translocación de GLUT-4 (36).
Por otro lado, se sabe que los ácidos grasos contribuyen en un 70% al ATP total
generado por un corazón aeróbico. Insulina controla los niveles circulantes de ácidos
grasos y lipoproteínas actuando sobre el tejido adiposo y sobre el hígado, pero también
regula el metabolismo de ácidos grasos en el corazón. Sin embargo, el corazón tiene
baja capacidad de sintetizar y almacenar ácidos grasos, por lo cual recibe ácidos
grasos exógenos como ácidos grasos unidos a albúmina y lipoproteínas ricas en
triglicéridos tales como la lipoproteína de muy baja densidad (VLDL) y los
quilomicrones (37). Además, insulina regula la síntesis de lipoproteina lipasa en
cardiomiocitos, enzima responsable de la hidrólisis de triglicéridos plasmáticos en la
superficie del lumen de las células vasculares endoteliales (31), liberando ácidos
grasos a la circulación y permitiendo su disponibilidad para el corazón.
Otro proceso metabólico afectado por insulina es la glicólisis. La vía glicolítica
permite producir dos moléculas de piruvato a partir de una molécula de glucosa- 6-
fosfato y tiene la ventaja de producir ATP sin requerir de oxígeno (38). En este proceso
hay varios pasos involucrados, pero la fosfofructokinasa-1 es considerada una de los
participantes más importantes de esta vía, más específicamente, la enzima regulatoria
clave. Esta enzima cataliza la transformación de fructosa 6-fosfato a fructosa 1,6-
bifosfato, utilizando un ATP.
La enzima fosfofructokinasa es inhibida por altos niveles de ATP o citrato y el
control de esta enzima es crucial para regular la glicólisis. Es conocido que la forma
cardiaca de fosfofructokinasa- 2, la cual es una enzima bifuncional que forma fructosa
2,6 bifosfato, un activador alostérico potente de la proteína limitante de la glicólisis,
fosfofructokinasa-1 (39), tiene una función importante en la activación de la glicólisis
estimulada por insulina. Este rol está dado tal vez por el control que ejerce la fructosa
2,6-bifosfato sobre la enzima fosfofructokinasa- 1 debido a que es capaz de superar los
efectos inhibitorios del ATP (40).
La hexokinasa también es afectada por insulina de manera indirecta. La hexokinasa
puede ser inhibida por la glucosa 6-fosfato, por lo tanto cuando los niveles de insulina
disminuyen, hay una acumulación de fructosa 6-fosfato y de glucosa 6-fosfato que es
una consecuencia de la carencia de estimulación de la fosfofructokinasa-1. Sin

9
embargo, altos niveles de insulina estimulan la fosfofructokinasa- 1 y no hay inhibición
de la hexokinasa. Hay algunos trabajos que declaran que la estimulación de la glicólisis
por insulina en el corazón está dada por el reclutamiento de GLUT-4 y también por el
aumento de fructosa- 2,6- bifosfato (41) y que esta activación de la fosfofructokinasa-2,
así como el transporte de glucosa, está mediada por la PI3K, uno de los componentes
de la vía de señalización de insulina (42).
1.6. Calcio
El calcio es un mensajero muy ubicuo y de mucha importancia para la señal
intracelular en las células vivas que ha demostrado regular diversas actividades como
por ejemplo la transcripción de genes, la proliferación celular y la contracción muscular.
Las células modulan la duración, la amplitud, la frecuencia y la extensión espacial de
los eventos de señalización de calcio, razón por la cual el calcio regula procesos tan
variados como los antes mencionados (43). Las señales de calcio pueden ocurrir del
orden de los microsegundos para desencadenar procesos como la exocitosis, del
orden de minutos u horas para controlar el crecimiento celular y la transcripción génica
y también pueden ocurrir en distintos espacios celulares, pueden llevarse a cabo en
dominios microscópicos, entrar a organelos o permanecer en el citoplasma para
producir una señal en toda la célula, así como también puede coordinar las actividades
entre células vecinas, ya sea difundiendo a través de uniones “gap” o por liberación de
mensajeros paracrinos (44).
Es así como cada tipo celular expresa un conjunto único de componentes de la
señalización de calcio para crear sistemas de señalización de calcio con diferentes
propiedades espaciales y temporales. Casi todos los sistemas de señalización de
calcio funcionan generando pulsos breves de calcio que son creados por variaciones
en las llamadas reacciones de encendido que son aquellas que introducen calcio al
citoplasma y las de apagado a través de las cuales las señales de calcio son
removidas ya sea por acción de tampones, bombas e intercambiadores (45).
Específicamente en el corazón, el calcio además contribuye a alcanzar la
despolarización celular de manera sincronizada y posteriormente a activar proteínas
contráctiles a través del mecanismo de ECC. Como se describió anteriormente, este

10
mecanismo consiste en la ocurrencia de la liberación de calcio desde reservorios
intracelulares, principalmente el retículo sarcoplásmico, en respuesta a la
despolarización de la membrana plasmática a través de los canales- receptores de
Ryanodina (RyR), proceso conocido como liberación de calcio inducida por calcio
(CICR) (46). Sin embargo, el calcio no sólo es importante para la contracción y
relajación normal del músculo, sino también es un importante segundo mensajero de
varias vías de señalización en el corazón. Se ha observado que la homeostasis del
calcio, así como también las vías de señalización dependientes de calcio son
importantes en el desarrollo de hipertrofia, insuficiencia y arritmias cardiacas (47).
1.7. Insulina y calcio
La regulación del calcio intracelular en el miocardio es determinante de la función
contráctil. Es por esta razón que en diversos estudios se ha planteado la posibilidad de
que la homeostasis del calcio en las células tenga alguna implicancia en la
cardiomiopatía diabética. Se ha observado que en animales diabéticos hay defectos en
la contracción que incluyen disminuciones en las cinéticas de acortamiento y relajación,
es decir, menor velocidad de contracción, prolongación de la contracción y retraso de
la relajación y también disminución en la presión, menor gasto cardiaco y menor
frecuencia cardiaca en reposo (48). Asimismo, se postula que los contenidos de calcio
de corazones diabéticos pueden estar alterados debido a que defectos en la
contractilidad cardiaca pueden deberse a cambios en la capacidad del miocardio para
regular el calcio durante la diabetes (49).
Los efectos de insulina sobre los niveles de calcio intracelular y sobre las corrientes
de calcio provenientes del exterior han sido objeto de algunos estudios en distintos
tipos celulares. En hepatocitos de rata, insulina aumenta las oscilaciones de calcio en
el núcleo de estas células y este aumento es independiente del calcio extracelular,
pero cuya señalización involucraría a la proteína fosfolipasa C (PLC) y al inositol
trifosfato (IP3) atribuyéndole un rol a insulina en el crecimiento del hígado y su
regeneración (50).
Acercándonos más a nuestro modelo celular, en músculo esquelético de ratas
neonatas se observó que insulina estimula las corrientes de calcio de canales tipo L

11
produciéndose un aumento en el calcio intracelular debido a la apertura posterior de los
canales- receptores de Ryanodina y que no involucraría según los resultados la vía
PLC-IP3. (29). Más aún, existen estudios previos en este mismo tipo celular que
especifican aumentos en el calcio intracelular estimulado por insulina particularmente
localizados en la zona próxima a la membrana celular y no en el calcio citosólico global
(51).
Por otro lado, se ha visto en ratones diabéticos tipo 1 que poseen un defecto en la
contractilidad cardiaca y una disminución de las corrientes de calcio de canales tipo L
debido a una menor actividad de señalización de la vía PI3K en el corazón (52). A su
vez, de manera concordante con estos datos, se ha demostrado en cardiomiocitos
auriculares de humanos y en cardiomiocitos ventriculares de ratas adultas a través de
técnicas electrofisiológicas que insulina induce un aumento en las corrientes de calcio
tipo L en un 86 ± 11% y un 140 ± 12 % respectivamente sobre los niveles basales
(53,54).
Estudios previos en nuestro laboratorio utilizando cardiomiocitos de rata neonata
como modelo, han logrado establecer que insulina produce un aumento transitorio en
el calcio intracelular compuesto por dos señales diferentes. La primera provendría de la
apertura de los canales de calcio tipo L y la posterior apertura de los canales-
receptores de Ryanodina y la segunda de la liberación de calcio desde el retículo
sarcoplásmico mediada por el receptor de IP3 (55).
Como se ha planteado anteriormente, existen diversos estudios que avalan la
participación de insulina en la homeostasis del calcio intracelular en distintos modelos
celulares. Particularmente en el corazón, se han evaluado los efectos de insulina sobre
los movimientos de calcio llegando a establecerse que insulina induce un aumento en
las corrientes de calcio mediadas por los canales tipo L y un aumento en el calcio
intracelular a través de los receptores de Ryanodina y de IP3 en el retículo
sarcoplásmico. No obstante, a la fecha no se ha establecido el rol de insulina en la
regulación del calcio en el cardiomiocito adulto que corresponde a un modelo que se
asemeja más al fisiopatológico real y que posee diferencias marcadas en el fenotipo y
genotipo con el cardiomiocito neonato. Estas variaciones nos conducen a pensar en
posibles disimilitudes en su comportamiento frente a insulina, por lo tanto nos parece
fundamental estudiar la relación insulina y calcio en el cardiomiocito adulto.

12
2. HIPOTESIS
Insulina induce un aumento en los niveles intracelulares de Ca2+ en el cardiomiocito
adulto a través de un mecanismo dependiente del Ca2+ extracelular y del almacenado
en reservorios intracelulares.
3. OBJETIVO GENERAL
Estudiar el papel del Ca2+ como segundo mensajero de la acción de insulina en
cardiomiocitos adultos aislados.
4. OBJETIVOS ESPECIFICOS
Estudiar la activación canónica del sistema transduccional de insulina en
cardiomiocitos aislados de corazón de rata adulta.
Caracterizar las cinéticas de Ca2+ inducida por diferentes concentraciones de
insulina en medio de reposo con y sin calcio externo.
Caracterizar las cinéticas de Ca2+ inducida por diferentes concentraciones de
IGF-1 en medio de reposo con y sin calcio externo.
Estudiar la participación del Ca2+ en la activación de la vía de señalización
canónica de insulina.

13
5. MATERIALES Y MÉTODOS
5.1. Reactivos
Los siguientes reactivos se adquirieron en Sigma Chemical Co. (St. Louis, MO,
EEUU), medio M199, SDS, NaVO4, PMSF, glicerol, DTT, Tween-20, ácido pirúvico
(Cat. No. 501568), reactivo de Folin Ciocalteau, 2,3-butanodiona monoxima (Cat. No.
B0753-100G), anticuerpo monoclonal anti-beta-actina (Cat. No. A5316) y anticuerpo
monoclonal anti-beta-tubulina (Cat. No. T0198). De Merck (Darmstadt, Alemania) se
obtuvieron: NaOH, HCl, Na2CO3, Na2HPO4, CuSO4•5H2O, NaF, pirofosfato de sodio,
Tritón X-100, MgCl2, 2-propanol. De Amresco (Solo, OH, EEUU): NaCl, leupeptina,
aprotinina, HEPES sal sódica, Tris base, acrilamida, bisacrilamida, EGTA. De
Invitrogen (Carlsbad, CA, EEUU) se adquirió: laminina (natural mouse, Cat. No. 23017-
015). De Molecular Probes (actualmente subsidiaria de Invitrogen) se obtuvieron:
fluo3-acetoximetilester (Fluo3-AM), BAPTA-AM, ácido plurónico y Alexa fluor 488 anti-
rabbit. De Winkler (Santiago, Chile) se obtuvieron: KCl, 2-mercaptoetanol, TEMED,
BSA. De Calbiochem (La Jolla, CA, EEUU) se adquirieron: anti-IgG conejo, anti-IgG
ratón. Los anticuerpos primarios anti p-Akt, Akt, p-Erk, p-RI se adquirieron en Cell
Signaling Technology. El anticuerpo primario anti Erk1/2 (K-23) se obtuvo de Santa
Cruz Biotechnology Inc. (Santa Cruz, CA, EEUU). Otros reactivos: reactivo de Bradford
(BioRad, EEUU), Dako (Dako Cytomation, CA, EEUU). El reactivo quimioluminiscente
para Western blot (Western Lightning) se adquirió en PerKinElmer Life Sciences, Inc.
(Boston, MA, EEUU). Ketamina y xilazina utilizadas fueron de uso veterinario.
Colagenasa A (Cat. No. 11088793001) se adquirió de Roche (Mannheim, Alemania).
5.2. Animales
Se utilizaron ratas macho Sprague- Dawley adultas de aproximadamente 2 meses
de edad con un peso entre 200- 250 g, provenientes del bioterio de la Facultad de
Ciencias Químicas y Farmacéuticas, Universidad de Chile. Todos los estudios con
animales contaron con la aprobación del Comité de Bioética de nuestra institución.

14
Esta investigación se realizó acorde a la guía “Guide for the care and use of laboratory
animals” publicada por Instituto de Salud de Estados Unidos, (NIH publicación N°85-23,
revisada en 1985).
5.3. Obtención de cardiomiocitos de rata adulta
Las ratas se anestesiaron con una mezcla ketamina: xilacina (2:1). Luego se extrajo
el corazón desde la cavidad torácica y se puso en un recipiente que contenía buffer
Gerard (0,19 mM NaH2PO
4, 1,01 mM Na
2HPO
4, 10 mM HEPES, 128 mM NaCl, 4 mM
KCl, 1,39 mM MgSO4, 5,5 mM glucosa, 2 mM ácido pirúvico, pH 7,4) frío.
Inmediatamente el corazón se canuló a través de la arteria aorta en una bomba
peristáltica para proceder a una retroperfusión. Se hizo pasar por el corazón 3
soluciones en medio Gerard consecutivamente: CaCl2 2,6 mM por 1 min o hasta que el
corazón no contuviera sangre, EGTA 2,5 mM por 2 min o hasta que dejase de
contraerse el corazón y colagenasa A 0,1- 0,13% por 30 min en recirculación para
digerir el corazón. Una vez finalizado este proceso el corazón se retiró de la bomba, se
cortó longitudinalmente y se disgregó mecánicamente con ayuda de un par de pinzas.
Posteriormente se introdujo en una botella plástica, se agregó solución de colagenasa
A y se llevó a baño termorregulado (37°C) por 15 min y en constante agitación. Luego
se tomó el sobrenadante y se traspasó a un tubo plástico de 15 mL y se centrifugó por
2 min a 500 rpm para obtener los cardiomiocitos adultos separados de otros tipos
celulares. El tejido remanente se volvió a poner en agitación con nueva solución de
digestión en el baño termorregulado. Una vez obtenidos los pellets celulares de al
menos 2 digestiones en el baño termorregulado, las células se resuspendieron en
M199 (HEPES modificado) suplementado con 2,3-butanodiona monoxima (BDM) 5 mM
(para evitar la contracción y favorecer el pegado de las células) y se procedió al
sembrado de los cardiomiocitos en las placas de trabajo pre tratadas durante 24 h. con
laminina 5 µg/mL en PBS estéril. Después de 15 min se cambió el medio de las células
por M199 fresco con/sin BDM dependiendo del experimento que se realizaría.
Finalmente las células se incubaron (5% CO2 y 95% aire a 37°C) 2-3 h antes de
realizar los ensayos.

15
5.4 Obtención de extractos proteicos para electroforesis
Luego de los tiempos de estímulos necesarios para realizar los experimentos las
células se lavaron con PBS (NaCl 80%, KCl 2%, Na2HPO4 *7H2O 19,5%, KH2PO4 0,2%
(p/v)) frío (4°C) 3 veces. Posteriormente, las células se trataron con buffer de lisis (10
mM Tris-HCl pH 7.2, 150 mM NaCl, 0,1% SDS, 1% Tritón X-100, 1% deoxicolato, 5 mM
EDTA) suplementado con 20 mM NaF, 1 mM pirofosfato de Na, 1 mM NaVO4, y 1 mM
PMSF, 1 µg/mL leupeptina, 1 µg/mL aprotinina. Una vez obtenido el lisado celular, este
se centrifugó a 15.000 rpm por 10 min a 4°C y posteriormente se separó el pellet
(desecho) del sobrenadante puesto que esta última es la fracción utilizada.
La concentración de proteínas de las muestras se determinó a través de 2 métodos:
Bradford (Bio-Rad protein assay) y Lowry. Posterior a la cuantificación, las muestras se
prepararon utilizando un 25% de tampón SDS-PAGE 4x (25 mM Tris-HCl pH 7,5, 32%
v/v glicerol, 20% 2-mercaptoetanol, 9,2% SDS, 0,02% azul de bromofenol), fracciones
solubles de las muestras (c.s.p.) y H2O (c.s.p.) y se incubaron por 7 min a 95ºC. Luego
se guardaron las muestras a -20°C.
5.5. Determinación de proteínas
5.5.1. Método de Lowry
Para este método primero se preparó el reactivo A mezclando partes iguales de una
solución de cobre-tartrato-carbonato (preparado por mezcla de 50 mL de 0,2% p/v
CuSO4•5H2O y 0,4% p/v tartrato dipotásico con 50 mL de 20% p/v Na2CO3), SDS 10%
y NaOH 0,8N. Luego se preparó el reactivo B diluyendo 1 volumen del reactivo de Folin
Ciocalteau con 5 volúmenes de agua nanopura. Posteriormente en tubos Eppendorf se
mezcló 5 µL de muestra de proteínas con 395 µL de agua nanopura y 400 µL de
reactivo A, se dejó reposar por 10 min a temperatura ambiente y luego se agregó 200
µL de reactivo B (Folin). Esta mezcla se incubó a 40ºC por 30 min y se midió la
absorbancia a 750 nm. Para realizar la curva de calibración se utilizó albúmina de
suero bovino (BSA).

16
5.5.2. Método de Bradford
Se mezclaron 5 µL de muestra con 200 µL de reactivo de Bradford (Bio-Rad protein
assay) y 795 µL de agua. Se midió absorbancia a 595 nm. Como estándar para realizar
la curva de calibración se utilizó albúmina de suero bovino (BSA).
5.6. Electroforesis en geles de poliacrilamida
La separación de las proteínas de acuerdo a su masa molecular se realizó mediante
electroforesis en geles de poliacrilamida entre el 8% y 12%. Para la detección se
cargaron entre 25 y 40 µg de extracto proteico. La electroforesis se realizó a un voltaje
constante de 100 Vs por 2,5 h. en tampón de electroforesis 1x que se obtuvo a partir
de una dilución de buffer de electroforesis 10x (tris base 30,25 g, glicina 144 g, SDS 10
g, agua 1000 mL).
5.7. Electrotransferencia de proteínas
Una vez realizada la electroforesis, las proteínas se electrotransfirieron a una
membrana de nitrocelulosa de 0,2 µm (BioRad) a amperaje constante (400 mA)
durante 90 min en tampón de transferencia 1x que se obtuvo mezclando 100 mL de
tampón de transferencia 10x (glicina 144 g, tris base 30,25 g, agua 1000 mL), 200 mL
de metanol y 700 mL de agua destilada.
5.8. Inmunowestern blot
Una vez transferidas, las membranas se bloquearon con tampón de bloqueo (TBS
1x; Tween-20 0,1%; BSA 3%) durante 60 min a temperatura ambiente y posteriormente
se incubaron con los anticuerpos primarios en solución de bloqueo toda la noche a las
diluciones recomendadas por los fabricantes: pAkt, Akt, pERK, pRI 1:1000; ERK 1/2
(K23), tubulina, actina 1:2000 y a 4°C.
Posterior a la incubación con los anticuerpos primarios, las membranas se lavaron 3
veces por 5 min con TBS 1X / Tween-20 al 0,1%, y se incubaron por 2 h a temperatura

17
ambiente con anti-IgG de ratón o conejo, según correspondiera, conjugados con
peroxidasa, en solución de bloqueo a un título de 1:5.000.
Una vez pasado el tiempo de incubación con el anticuerpo secundario las
membranas fueron lavadas 3 veces por 5 min con TBS 1x - Tween-20 al 0,1% y se
procedió a la detección de la unión específica de los anticuerpos a las proteínas. Para
esto, las membranas se incubaron por 1 min en solución de ECL (enhanced
chemiluminescence) y luego se expusieron a la película de fotografía Kodak-Biomax.
Las películas obtenidas se digitalizaron y las imágenes se analizaron por densitometría
mediante el programa ImageJ 1.42q.
Para detectar otras proteínas en las mismas membranas, éstas se sometieron a un
proceso que permite quitar los anticuerpos adheridos a la membrana en corto tiempo
(“mild stripping”: glicina 1,5% p/v, SDS 0,1% p/v, tween 20 1% v/v, pH 2,2). Posterior a
este proceso, las membranas se lavaron 3 veces por 5 min con TBS 1x- Tween-20 al
0,1% y luego se procedió a bloquear nuevamente las membranas y de esta manera se
continuó con el proceso anteriormente descrito.
5.9. Determinación de los niveles intracelulares de calcio
Para realizar estos ensayos se trabajó con células adheridas en cubreobjetos de
25 mm (pre-tratados con laminina 5 µg/mL) que a su vez estaban contenidos en placas
de 6 pocillos o 35 mm. Los cardiomiocitos adultos se lavaron utilizando medio Krebs
con calcio (140 mM NaCl, 5 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 10 mM HEPES, 1
g/mL glucosa, pH 7,4), se incubaron con la sonda fluorescente para calcio Fluo3-AM
(5,5 µM) durante 25 min a temperatura ambiente y luego se lavaron nuevamente con
medio Krebs con calcio o sin calcio (145 mM NaCl, 5 mM KCl, 1,0 mM EGTA, 1 mM
MgCl2, 10 mM HEPES-Na, 5,6 mM glucosa, pH 7,4) dependiendo del experimento que
se llevaría a cabo. Luego el cubreobjetos que contenía las células fue montado en una
cámara especial de 1 mL de capacidad y esta cámara se posicionó en un microscopio
confocal (Carl Zeiss Axiovert 135 M-LSM Microsystems) a través del cual se realizaron
los registros. Las imágenes fluorescentes se recolectaron cada 0.983 seg con el lente
63x, con una configuración para Fluo3-AM, excitación 488 nm; emisión 526 nm. y en
modo de campo completo o escaneo en línea según correspondiese.

18
Una vez obtenidos los registros, éstos se analizaron con el programa ImageJ 1.42q
midiendo la fluorescencia en una región óptica de interés (ROI) al interior de las células
y en todas las imágenes adquiridas. La intensidad de fluorescencia es proporcional al
calcio que se encuentra al interior de las células. Esta intensidad se expresó como
Porcentaje de fluorescencia relativa a la fluorescencia basal de cada célula.
5.10. Inmunofluorescencia indirecta
Los cardiomiocitos adultos se sembraron en cubreobjetos de vidrio pre-tratados
(18 mm) como ya se mencionó anteriormente, en placas de 12 pocillos. Transcurrido
el periodo de estimulación, las células se fijaron con paraformaldehído (4% p/v en PBS,
20 min), se permeabilizaron (PBS-0,3% v/v Tritón X-100, 30 min) y se bloquearon (2%
p/v BSA en PBS, 1 h). Este procedimiento se realizó sobre hielo a 4°C con lavados
entre cada paso, utilizando PBS frío (4°C). Posteriormente, las células se incubaron
con el anticuerpo primario, anti fosfo-Receptor de Insulina (RI) 1:100 diluido en PBS-
2% BSA toda la noche, se lavaron y se incubaron con el anticuerpo secundario IgG de
conejo conjugado a Alexa 488, (1:400) diluido en PBS-2% BSA, con Rodamina-
Faloidina (1:500 para marcar la actina sarcomérica) y Hoescht (1:1.000 para marcar los
núcleos celulares) por 2 h y protegido de la luz. Finalmente, a los cubreobjetos se les
agregó 10 µL de DAKO para extender el tiempo de decaimiento del fluoróforo y fueron
montados en portaobjetos. Las muestras así tratadas se observaron en microscopio
confocal.
5.11. Caracterización de cardiomiocitos adultos
Los cardiomiocitos adultos que se utilizaron para realizar los experimentos
cumplían con ciertas exigencias ya sea analizando el cultivo en general o las células
individuales dependiendo de los experimentos que se llevarían a cabo.
5.11.1. Cardiomiocitos adultos para inmunowestern blot e inmunofluorescencia
Para realizar estos experimentos lo más importante fue evaluar la confluencia
de células lograda luego de la extracción en las placas de cultivo. Se consideró
suficiente una confluencia superior a 50% e insuficiente bajo este porcentaje. En el

19
caso de que las placas no alcanzaran la confluencia establecida, estas eran
desechadas ya que el número de células existentes no sería el necesario para llevar a
cabo estos experimentos de manera exitosa, representando un alto riesgo de fracaso
del experimento. La Figura 1A muestra dos cultivos representativos de ambos casos.
5.11.2. Cardiomiocitos adultos para medición de calcio por microscopía confocal
Para las mediciones de los niveles de calcio intracelulares la cantidad de
células no necesariamente debe ser muy alta, sin embargo, debido a que se registra
sólo un campo por cubreobjetos y en muchos casos este campo posee entre 1 a 3
cardiomiocitos adultos, es imperativo elegir muy bien las células que se van a registrar
basándose en características morfológicas de viabilidad. Lo primero que observamos
es la forma de la célula, esta debe ser tipo bastón, también conocida como “rod-shape”
y no células en forma esférica (“round-shape”) ya que esas son células apoptóticas que
se caracterizan por la compactación de su volumen celular. Es muy importante también
diferenciar las células viables de aquellas que a pesar de tener forma de bastón
corresponden a las llamadas células necróticas, que al igual que las células
apoptóticas no son viables y que se caracterizan por su estructura cilíndrica que solo
es mantenida por el citoesqueleto y en las cuales existe pérdida de la integridad de la
membrana celular junto con un aumento del volumen de la célula. En el caso de los
registros de calcio realizados en presencia de calcio externo se consideró además,
como muestra de viabilidad de los cardiomiocitos adultos, la presencia de contracción
celular, es decir, en lo posible, se registró sólo células que mantuvieran una
contracción basal constante. No obstante, debido a razones evidentes, para la elección
de células en las mediciones de calcio en ausencia de calcio extracelular solo nos
basamos en las características morfológicas de viabilidad mencionadas anteriormente.

20

21
Figura 1. Caracterización de los cardiomiocitos adultos. (A) Esta imagen (izquierda)
muestra el porcentaje de confluencia aceptable para realizar experimentos de Western blot e
inmunofluorescencia, mientras que a la derecha vemos una cantidad de células insuficientes
para este tipo de experimentos. (B) Viabilidad de cardiomiocitos adultos de acuerdo a su
morfología. (C) Cardiomiocito en su estado basal (izquierda) y en contracción (derecha).
5.12. Expresión de resultados y análisis estadístico
Los resultados mostrados corresponden al promedio ± SEM de, al menos, tres
experimentos independientes. Los datos se analizaron con el programa GraphPad
Prism5 y la significancia estadística se evaluó mediante test de Student o ANOVA de
una vía. Los post-test ANOVA utilizados fueron Tuckey´s o Dunnett según
correspondiera. Se consideró como límite de significancia estadística, valores de
p<0,05.
La fluorescencia se expresó como porcentaje de fluorescencia relativa a la
fluorescencia basal de cada célula,[(F-F0)/F0] x100, donde F representa el valor de
fluorescencia en cada punto y F0 representa el valor basal de fluorescencia.

22
6. RESULTADOS
6.1. Activación de la vía de señalización canónica de insulina en cardiomiocitos
adultos
Lo primero que se realizó a nivel experimental fue asegurarnos de que el sistema de
señalización de insulina funcionaba en nuestro modelo celular. Para este fin se
utilizaron dos técnicas previamente descritas en Materiales y Métodos, el
inmunowestern blot y la inmunofluorescencia indirecta. Los cardiomiocitos de rata
adulta se trataron con insulina 100 nM por distintos periodos entre 5 a 60 min.
Posteriormente, se evaluó la fosforilación de proteínas que canónicamente se activan
al ser estimulado el receptor de insulina por su ligando.
6.1.1. Efecto de insulina sobre la fosforilación del receptor de insulina
La Figura 2A muestra el resultado del Western blot para el receptor de insulina
fosforilado (p-RI) en cardiomiocitos adultos estimulados con insulina. Se observa que la
fosforilación del receptor en los residuos Tyr1150/1151 pasa de un basal prácticamente
inexistente a una fosforilación máxima al ser estimulado con insulina 100 nM por 5 min
(p<0,01). A medida que el tiempo de estímulo con insulina aumenta, la fosforilación del
receptor tiende a disminuir, sin embargo, esta tendencia no posee significancia
estadística.
La presencia del receptor de insulina (RI) fosforilado a los 5 min post-estimulación
con insulina se corroboró por inmunofluorescencia indirecta como se muestra en la
Figura 2B, utilizando un anticuerpo monoclonal anti-fosfo receptor de insulina
(Tyr1150/1151), obteniéndose como resultado un aumento de la marca del p-RI
respecto al control y una localización subcelular más definida. A su vez, se puede
apreciar un pequeño desplazamiento de la colocalización de la marca del p-RI con
respecto a la actina sarcomérica en las células estimuladas con insulina en relación al
control.

23
6.1.2. Efecto de insulina sobre la fosforilación de Akt
Continuando río debajo de la vía de señalización del receptor de insulina se
investigó qué sucedía con la fosforilación de la proteína Akt en presencia de insulina
100 nM y a los tiempos anteriormente mencionados. La Figura 2C muestra que la
fosforilación de Akt alcanzó su máximo a los 5 min de estímulo (p<0,001). Este nivel de
fosforilación se mantuvo a los 10 min de estímulo y comenzó a decaer ya a los 20 min.
Esta fosforilación decayó drásticamente a los 60 min posteriores al pulso de insulina.
6.1.3. Efecto de insulina sobre la fosforilación de ERK 1/2
En el caso de las proteínas kinasas ERK 1/2 (Figura 2D) se observó un
comportamiento diferencial de la isoforma ERK 2 (p42) respecto a ERK 1 (p44),
reflejándose esta diferencia en que la fosforilación de ERK 1 casi no se modificó por la
acción de insulina en nuestro modelo; sin embargo, ERK 2 aumentó significativamente
su fosforilación (p<0,001), alcanzando su nivel máximo a los 5 min al igual que las
otras proteínas analizadas.

24
Figura 2. Insulina estimula la fosforilación del receptor de insulina (RI) y de las proteínas
kinasas Akt y ERK2 en el cardiomiocito adulto. La fosforilación del RI (A), Akt (C) y ERK2
(D) se evaluó mediante Western blot, observándose la máxima fosforilación para las 3 proteínas
a los 5 min de estímulo con insulina 100 nM. (B) Inmunofluorescencia contra el receptor de
insulina fosforilado. Los datos mostrados son representativos de un mínimo de 3 experimentos
independientes.

25
6.2. Efecto de insulina en los niveles intracelulares de calcio en el cardiomiocito
adulto
El efecto de insulina sobre los niveles intracelulares de calcio se evaluó utilizando
microscopía confocal y la sonda fluorescente Fluo3-AM como se describió en
Materiales y Métodos. Las mediciones se realizaron en ausencia o presencia de calcio
externo para estudiar la procedencia del calcio detectado por la sonda al interior de las
células.
Los análisis de los registros de mediciones de los niveles de calcio en presencia de
insulina ya sea utilizando medio externo con o sin calcio se efectuaron en distintas
zonas celulares: citoplasma, núcleo y membrana celular. La Figura 3 muestra una
representación gráfica de las zonas evaluadas en los cardiomiocitos adultos.
Figura 3. Identificación de zonas de análisis (ROI) en el cardiomiocito adulto. En la imagen
se señalan las 3 zonas analizadas para determinar si se producían movimientos de calcio
estimulados por insulina.

26
6.2.1. Efecto de insulina en los niveles de calcio intracelular de cardiomiocitos
adultos en ausencia de calcio externo
Para estudiar los cambios que pudiese producir insulina en el calcio, los
cardiomiocitos adultos se expusieron a distintas concentraciones de insulina (Ins) entre
10 nM y 10 µM, manteniéndolos en una primera instancia sin calcio externo con el fin
de investigar si el calcio que pudiera movilizar insulina en el cardiomiocito adulto
proviene de reservorios intracelulares. A partir de estos experimentos se obtuvo las
cinéticas de calcio que se observan en las figuras sucesivas. Estas mediciones se
efectuaron en el modo campo completo.
La primera zona evaluada fue el citoplasma celular. La Figura 4A muestra las
cinéticas de calcio desprendidas del análisis de una región de interés (del inglés
“region of interest”, ROI) ubicada en el citoplasma del cardiomiocito representativas de
cada una de las concentraciones de insulina ensayada.
Debido a que a simple vista, basándose en las cinéticas de calcio, pareciera no
haber liberaciones de este catión desde reservorios intracelulares estimuladas por
insulina, se expresó las mediciones en variables objetivas como fluorescencia relativa
promedio y fluorescencia relativa máxima posterior al estímulo con respecto a los
valores basales y a su vez con respecto a los controles (cinéticas no mostradas), dado
que en la mayoría de las cinéticas pareciera haber una pendiente en la fluorescencia
que a medida que transcurre el tiempo se mantiene, pero cuya fluorescencia cada vez
se hace mayor. La Figura 4B muestra los gráficos correspondientes a fluorescencia
relativa promedio y fluorescencia relativa máxima evaluadas en el citoplasma celular.
Como se observa en estos gráficos no hay diferencias significativas en ninguno de
estos dos parámetros.
Como se mencionó anteriormente, además del citoplasma celular, se analizó la
zona nuclear y aquella próxima a la membrana plasmática del cardiomiocito adulto. La
Figura 5A muestra las cinéticas de calcio representativas correspondientes al análisis
de la región de interés (ROI) ubicada en alguno de los dos núcleos celulares del
cardiomiocito adulto para las concentraciones de 10 nM, 100 nM, 1 M y 10 M de
insulina (Ins). En la figura 5B se muestran los gráficos de fluorescencia relativa
promedio y fluorescencia relativa máxima que se realizaron a partir de las cinéticas de
calcio. A su vez, en la Figura 6A se muestran las cinéticas de calcio correspondientes

27
al análisis de la región de la membrana de los cardiomiocitos adultos a las distintas
concentraciones antes mencionadas de insulina y en la Figura 6B se presenta la
fluorescencia relativa promedio y fluorescencia relativa máxima para la misma zona. Al
igual que en la región citoplasmática, en las regiones nucleares y de membrana los
niveles de calcio intracelular no se ven afectados al estimular las células con insulina.
La mayoría de los registros de movimientos de calcio se acompañaron de
controles positivos que permiten demostrar que las células responden a otros
estímulos distintos de insulina y que el método utilizado es adecuado y que, por lo
tanto, en nuestro modelo celular no hay liberación de calcio desde reservorios
intracelulares dependiente de insulina de manera detectable con Fluo3-AM.
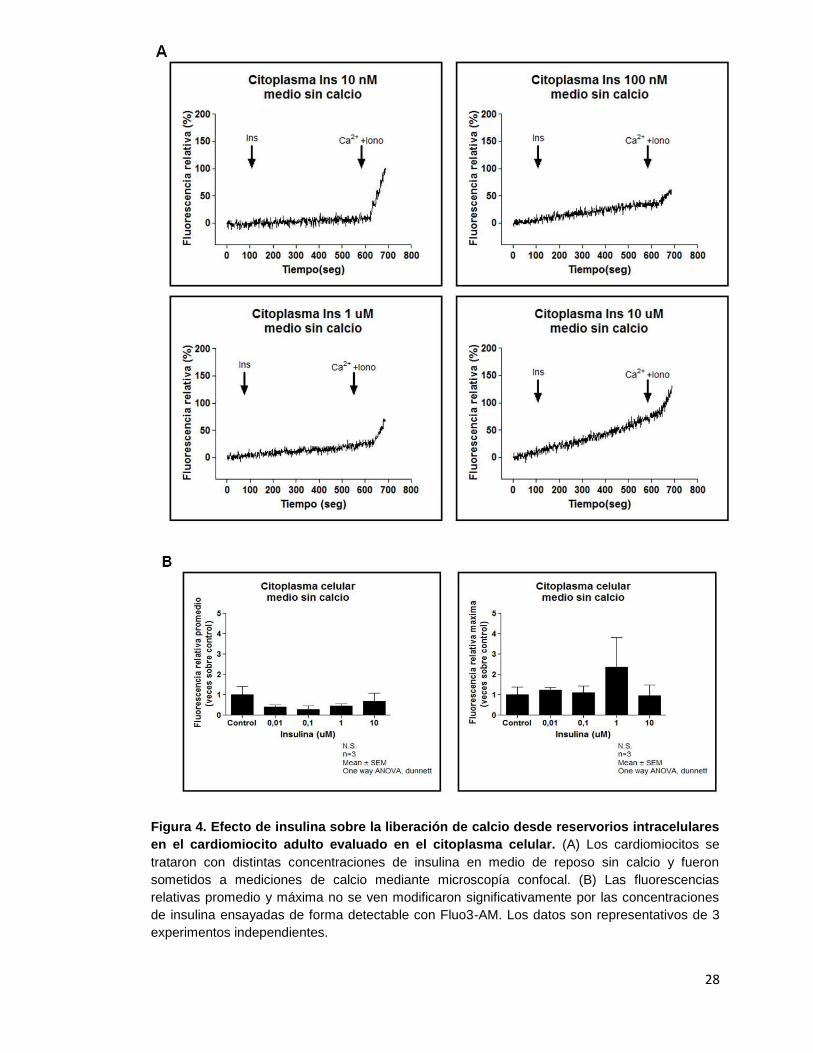
28
Figura 4. Efecto de insulina sobre la liberación de calcio desde reservorios intracelulares
en el cardiomiocito adulto evaluado en el citoplasma celular. (A) Los cardiomiocitos se
trataron con distintas concentraciones de insulina en medio de reposo sin calcio y fueron
sometidos a mediciones de calcio mediante microscopía confocal. (B) Las fluorescencias
relativas promedio y máxima no se ven modificaron significativamente por las concentraciones
de insulina ensayadas de forma detectable con Fluo3-AM. Los datos son representativos de 3
experimentos independientes.
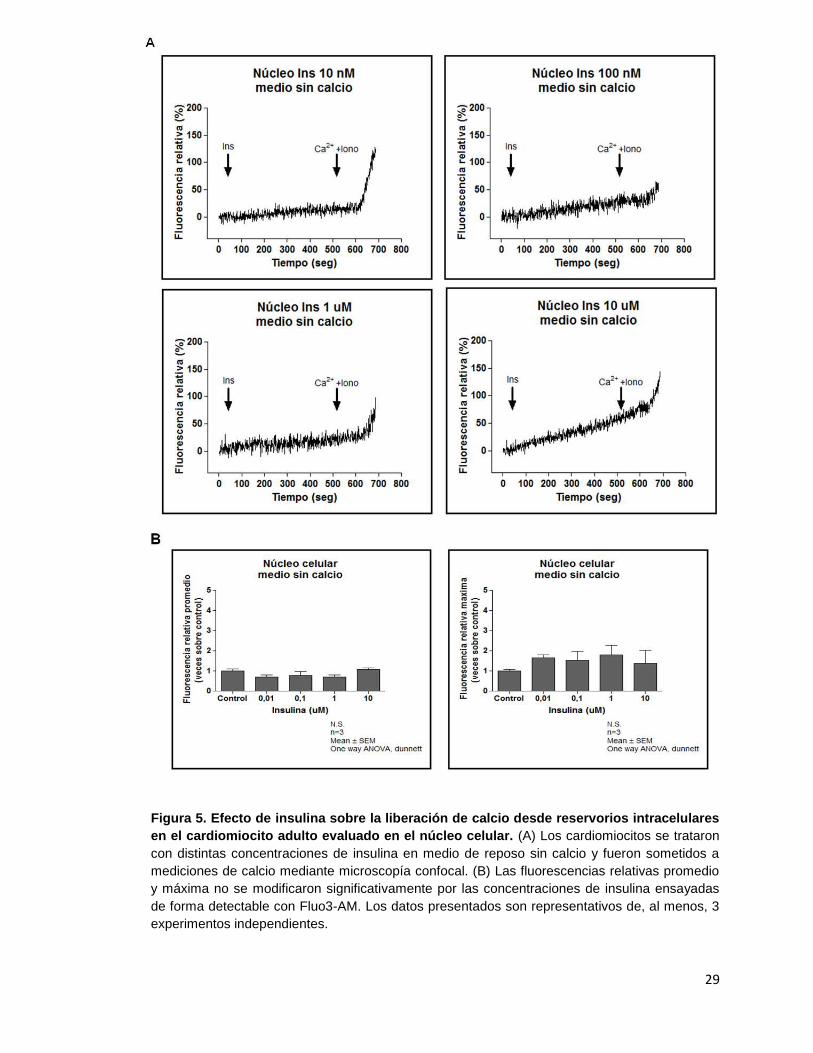
29
Figura 5. Efecto de insulina sobre la liberación de calcio desde reservorios intracelulares
en el cardiomiocito adulto evaluado en el núcleo celular. (A) Los cardiomiocitos se trataron
con distintas concentraciones de insulina en medio de reposo sin calcio y fueron sometidos a
mediciones de calcio mediante microscopía confocal. (B) Las fluorescencias relativas promedio
y máxima no se modificaron significativamente por las concentraciones de insulina ensayadas
de forma detectable con Fluo3-AM. Los datos presentados son representativos de, al menos, 3
experimentos independientes.

30
Figura 6. Efecto de insulina sobre la liberación de calcio desde reservorios intracelulares
en el cardiomiocito adulto evaluado en la membrana celular. (A) Los cardiomiocitos fueron
tratados con distintas concentraciones de insulina en medio de reposo sin calcio y sometidos a
mediciones de calcio mediante microscopía confocal. (B) Las fluorescencias relativas promedio
y máxima no cambiaron significativamente. Los resultados son representativos de, al menos, 3
experimentos independientes.

31
6.2.2. Efecto de insulina en los niveles de calcio intracelular y frecuencia de
contracción de cardiomiocitos adultos en presencia de calcio externo
Posterior a las mediciones realizadas en ausencia de calcio, se estudió si insulina
inducía un aumento de calcio en el interior de los cardiomiocitos adultos dependiente
de una entrada de calcio desde el extracelular, ya que según lo observado
anteriormente insulina no produce liberaciones de calcio detectables con Fluo3-AM
desde reservorios intracelulares del cardiomiocito adulto.
Con este fin, los cardiomiocitos se cultivaron en presencia de calcio externo durante
las mediciones de calcio y expusieron a distintas concentraciones de insulina: 10 nM,
100 nM, 1 µM y 10 µM. La Figura 7A muestra las cinéticas representativas derivadas
del análisis del citoplasma celular obtenidas de estos ensayos. En la Figura 7B se
muestran los datos cuantitativos del efecto de insulina en sus distintas concentraciones
sobre el calcio intracelular de cardiomiocitos en presencia de calcio externo medido en
una región de interés ubicada en el citoplasma celular. Estos gráficos muestran que no
hay diferencias significativas en las fluorescencias relativas promedio y máxima
posterior al estímulo con respecto a la fluorescencia previa al estímulo y comparado
con los controles.
Al igual que para las mediciones de cinéticas de calcio en ausencia de calcio
externo, también se evaluó el comportamiento de otras zonas del cardiomiocito adulto
en cuanto a los movimientos de los niveles de calcio intracelular. Cabe destacar que
como se mencionó previamente en Materiales y Métodos, las células elegidas para
estos registros presentaban un nivel de contracción basal y se usó como prueba de su
vialidad. En la Figura 8A se muestran las cinéticas de calcio correspondientes a la
evaluación de los niveles de calcio en un núcleo. La Figura 8B muestra los gráficos de
fluorescencia relativa promedio y fluorescencia relativa máxima de la misma zona. De
manera similar, la Figuras 9A y B presentan los movimientos de calcio evaluados en la
membrana celular y los correspondientes gráficos de fluorescencia relativa promedio y
fluorescencia relativa máxima desprendidos de estas cinéticas, respectivamente. El
análisis estadístico indicó que no hay diferencias significativas entre las fluorescencias
relativas promedio y máxima en células estimuladas con insulina y sus controles.
Estos resultados permiten concluir que insulina tampoco induce entrada de calcio
desde el medio extracelular a los cardiomiocitos adultos de manera detectable con
Fluo3-AM.
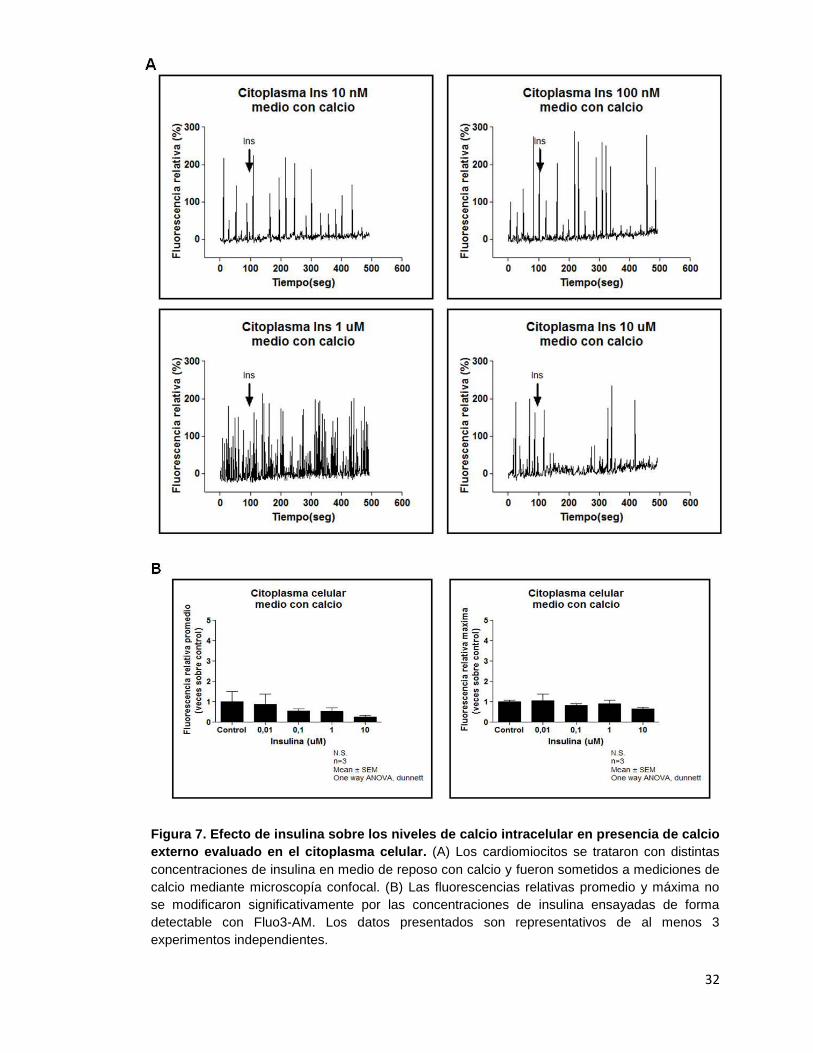
32
Figura 7. Efecto de insulina sobre los niveles de calcio intracelular en presencia de calcio
externo evaluado en el citoplasma celular. (A) Los cardiomiocitos se trataron con distintas
concentraciones de insulina en medio de reposo con calcio y fueron sometidos a mediciones de
calcio mediante microscopía confocal. (B) Las fluorescencias relativas promedio y máxima no
se modificaron significativamente por las concentraciones de insulina ensayadas de forma
detectable con Fluo3-AM. Los datos presentados son representativos de al menos 3
experimentos independientes.

33
Figura 8. Efecto de insulina sobre los niveles de calcio intracelular en presencia de calcio
externo evaluado en el núcleo celular. (A) Los cardiomiocitos se trataron con distintas
concentraciones de insulina en medio de reposo con calcio y fueron sometidos a mediciones de
calcio mediante microscopía confocal. (B) Las fluorescencias relativas promedio y máxima no
cambiaron significativamente por las concentraciones de insulina ensayadas de forma
detectable con Fluo3-AM. Los datos presentados son representativos de al menos 3
experimentos independientes.
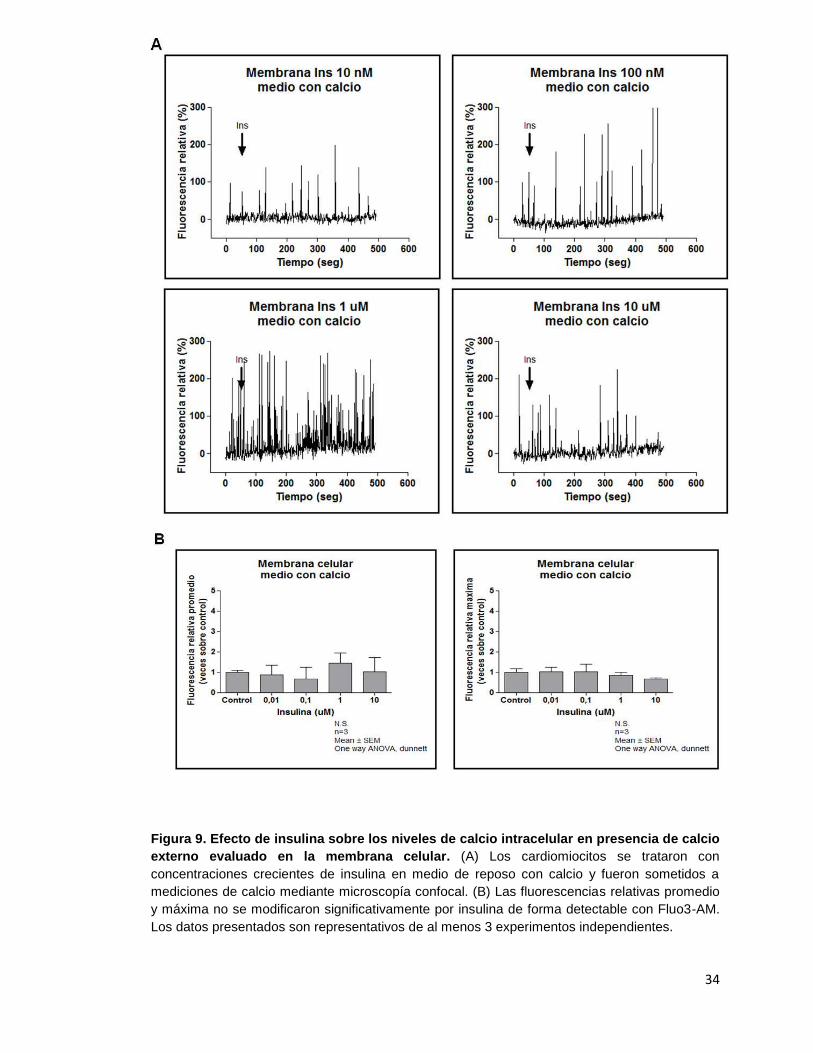
34
Figura 9. Efecto de insulina sobre los niveles de calcio intracelular en presencia de calcio
externo evaluado en la membrana celular. (A) Los cardiomiocitos se trataron con
concentraciones crecientes de insulina en medio de reposo con calcio y fueron sometidos a
mediciones de calcio mediante microscopía confocal. (B) Las fluorescencias relativas promedio
y máxima no se modificaron significativamente por insulina de forma detectable con Fluo3-AM.
Los datos presentados son representativos de al menos 3 experimentos independientes.

35
A continuación se evaluó si insulina tenía algún efecto sobre la frecuencia de
contracción de los cardiomiocitos adultos. Con este fin se realizaron experimentos con
cardiomiocitos adultos en presencia de calcio extracelular y registro en el modo
escaneo en línea (“line scan”). La Figura 10 muestra que insulina no cambia la
frecuencia de contracción de cardiomiocitos adultos a ninguna concentración de las
ensayadas. En esta serie de experimentos, la presencia de contractilidad sirvió como
medida cualitativa de viabilidad, además de las características morfológicas
comentadas en Materiales y Métodos.
A modo de resumen, la Figura 11A muestra un gráfico que presenta el
comportamiento del calcio intracelular del cardiomiocito adulto en presencia de insulina
100 nM, isoproterenol 10 µM e ionomicina 1 µM. Insulina no afecta los niveles de calcio
intracelular mientras que isoproterenol aumenta notoriamente la frecuencia de
contracción e ionomicina induce un drástico aumento en los niveles de calcio interno al
generar poros en la membrana plasmática. La Figura 11B muestra el aumento agudo
que produce insulina en los niveles de calcio intracelular en los cardiomiocitos
neonatos, acción previamente descrita por nuestro Laboratorio.

36
Figura 10. Efecto de insulina sobre la frecuencia de contracción de cardiomiocitos de rata
adulta. La frecuencia de contracción no se modificó significativamente a las concentraciones de
insulina ensayadas de forma detectable con Fluo3-AM.
Figura 11. Experimento que resume el efecto de insulina sobre los niveles de calcio
intracelular. Efecto secuencial de insulina (100 nM), isoproterenol (10 µM) e ionomicina (1 µM)
en los niveles intracelulares de calcio en cardiomiocitos adultos (A) o de insulina (100 nM) y KCl
(50 mM) en cardiomiocitos neonatos (B).

37
6.3 Efecto de IGF-1 sobre los niveles de calcio intracelular de cardiomiocitos
adultos
En la siguiente serie de experimentos se buscó investigar si IGF-1, un análogo
estructural de insulina, aumentaba los niveles de calcio intracelular en cardiomiocitos
adultos. Estos experimentos, de igual manera que los anteriores, se realizaron por
microscopía confocal utilizando la sonda Fluo3-AM para detectar los cambios del calcio
al interior de las células tanto en medio de reposo con y sin calcio externo. En estos
experimentos sólo se registraron los cambios en el citoplasma celular.
6.3.1. Efecto de IGF-1 en los niveles de calcio intracelular de cardiomiocitos
adultos en ausencia de calcio externo
Para estudiar el efecto de IGF-1 sobre el calcio intracelular de cardiomiocitos
adultos se expusieron las células a un medio externo en ausencia de calcio a fin de
dilucidar si IGF-1 estimulaba liberación de calcio desde los reservorios intracelulares.
Luego las células se trataron con dos concentraciones de IGF-1: 100 nM y 1 µM. Los
registros de las cinéticas y datos cuantitativos se muestran en las Figuras 12A-B,
respectivamente. Las fluorescencias relativas promedio y máxima muestran que no hay
diferencias significativas en los parámetros medidos a ninguna de las dos
concentraciones ensayadas (Figura 12B) Sin embargo, a 1 µM hubo una tendencia al
aumento de la fluorescencia relativa máxima, sin que llegara a ser significativa. En las
cinéticas representativas también se muestran controles positivos para descartar
problemas de viabilidad o fallas de la metodología empleada.
6.3.2. Efecto de IGF-1 en los niveles de calcio intracelular y frecuencia de
contracción de cardiomiocitos adultos en presencia de calcio externo
A fin de estudiar si existía un componente externo de calcio que pudiese ser
movilizado por IGF-1 se realizaron experimentos en presencia de calcio externo. A
similitud de los experimentos anteriores se utilizaron concentraciones de IGF-1: 100 nM
y 1 µM. La Figura 13A muestra las cinéticas de calcio representativas con los
respectivos controles positivos. La Figura 13B muestra que las fluorescencias relativas
máxima y promedio no se modificaron significativamente con IGF-1 respecto al
comportamiento basal de las células. También se apreció una tendencia a aumentar la

38
fluorescencia relativa promedio a 1 µM de IGF-1, pero que no alcanzó a ser
estadísticamente significativa.
Finalmente mediante el análisis de escaneo en línea se investigó si IGF-1
modificaba la frecuencia de contracción del cardiomiocito adulto. Los registros de
frecuencia de contracción de cardiomiocitos muestran que IGF-1 100 nM ó 1 µM no
modificaron significativamente este parámetro respecto al control (Figura 13C).

39
Figura 12. Efecto de IGF-1 sobre la liberación de calcio desde reservorios intracelulares
en el cardiomiocito adulto. (A) Los cardiomiocitos se incubaron con distintas concentraciones
de IGF-1 en medio de reposo sin calcio y fueron sometidos a mediciones de calcio mediante
microscopía confocal. (B) Las fluorescencias relativas promedio y máxima no se modificaron
significativamente a las concentraciones de IGF-1 ensayadas de forma detectable con Fluo3-
AM. Los datos presentados son representativos de 3 experimentos independientes.

40
Figura 13. Efecto de IGF-1 sobre el calcio intracelular en presencia de calcio externo y la
frecuencia de contracción. (A) Los cardiomiocitos se trataron con IGF-1 a las concentraciones
indicadas en medio de reposo con calcio y fueron sometidos a mediciones de calcio mediante
microscopía confocal. (B) Las fluorescencias relativas promedio y máxima y (C) la frecuencia
de contracción no cambiaron significativamente por acción del IGF-1 en forma detectable con
Fluo3-AM. Los datos presentados son representativos de al menos 3 experimentos
independientes.
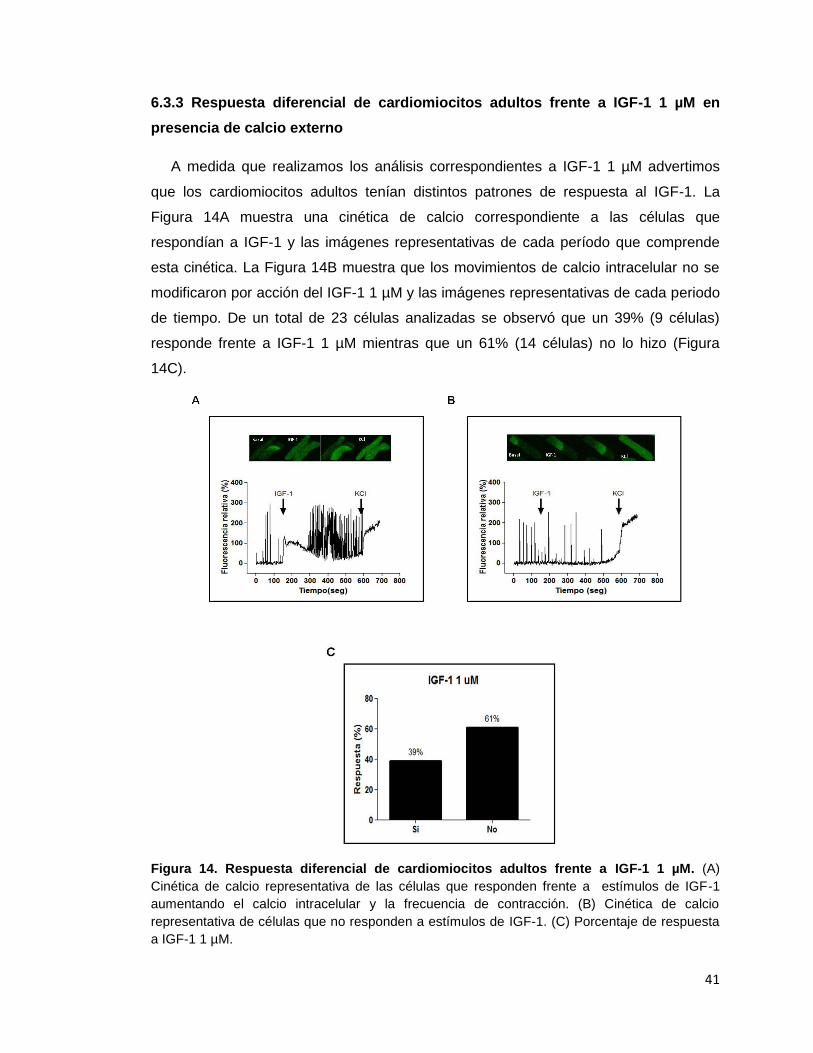
41
6.3.3 Respuesta diferencial de cardiomiocitos adultos frente a IGF-1 1 µM en
presencia de calcio externo
A medida que realizamos los análisis correspondientes a IGF-1 1 µM advertimos
que los cardiomiocitos adultos tenían distintos patrones de respuesta al IGF-1. La
Figura 14A muestra una cinética de calcio correspondiente a las células que
respondían a IGF-1 y las imágenes representativas de cada período que comprende
esta cinética. La Figura 14B muestra que los movimientos de calcio intracelular no se
modificaron por acción del IGF-1 1 µM y las imágenes representativas de cada periodo
de tiempo. De un total de 23 células analizadas se observó que un 39% (9 células)
responde frente a IGF-1 1 µM mientras que un 61% (14 células) no lo hizo (Figura
14C).
Figura 14. Respuesta diferencial de cardiomiocitos adultos frente a IGF-1 1 µM. (A)
Cinética de calcio representativa de las células que responden frente a estímulos de IGF-1
aumentando el calcio intracelular y la frecuencia de contracción. (B) Cinética de calcio
representativa de células que no responden a estímulos de IGF-1. (C) Porcentaje de respuesta
a IGF-1 1 µM.

42
6.4. Participación del calcio en la activación de la vía de señalización del receptor
de insulina
A continuación se estudió a través de otra técnica distinta de la microscopía
confocal, si insulina participaba de alguna manera en movimientos de calcio al interior
de las células. Para este fin se investigó la fosforilación de dos proteínas blanco de
insulina: su propio receptor RI y la proteína kinasa Akt mediante Western blot tal como
se describe en Materiales y Métodos. A su vez, las células se preincubaron con
BAPTA-AM, un quelante de calcio que ingresa a la célula y/o EGTA, un quelante de
calcio que no atraviesa la membrana plasmática y que por lo tanto actúa sobre el
calcio extracelular.
6.4.1. Participación del calcio en la fosforilación del receptor de insulina
La Figura 15A muestra que el receptor de insulina se fosforila por acción de insulina
100 nM a los 5 min. La preincubación de los cardiomiocitos adultos con BAPTA- AM
(50 µM) y/o EGTA (2 mM) por 30 min previos al pulso de Insulina de 5 min mostró que
ninguno de los dos quelantes modificó la fosforilación del RI dependiente de insulina.
Es decir, el calcio externo y el interno no participan en la fosforilación del receptor de
insulina.
6.4.2. Participación del calcio en la fosforilación de Akt
La Figura 15B muestra que la proteína quinasa Akt se fosforila a los 5 min post-
exposición a insulina 100 nM. Los cardiomiocitos adultos, al igual que en la figura
anterior, se preincubaron con BAPTA- AM (50 µM) y/o EGTA (2 mM) por 30 min
previos al pulso de insulina. Los análisis muestran que la fosforilación de Akt aumentó
100 veces sobre el basal en presencia de insulina 100 nM por 5 min (p<0,001) y que
en presencia de BAPTA- AM (50 µM) esta fosforilación decayó a niveles cercanos al
basal con una diferencia estadísticamente significativa (p<0,001) con respecto a
insulina.
En el caso de la participación del calcio extracelular, EGTA 2 mM no afectó la
fosforilación de Akt y su magnitud es similar a la observada en presencia sólo de

43
insulina (p<0,001 vs control). En presencia de insulina, BAPTA- AM y EGTA, la
fosforilación de Akt fue menor en comparación a las células tratadas sólo con insulina
(p<0,01) y por lo tanto no habría participación del calcio extracelular en la fosforilación
de Akt.
Figura 15. Participación del calcio en la fosforilación del receptor de insulina y proteína
kinasa Akt. Las células se preincubaron con BAPTA- AM (50 µM) y/o EGTA (2 mM) por 30 min
previos a la estimulación con insulina 100 nM por 5 min. (A) La fosforilación del receptor de
insulina no se modificó por la presencia/ausencia de calcio intracelular ni extracelular. (B) La
fosforilación de la proteína Akt inducida por insulina disminuye en presencia del quelante
intracelular de calcio pero no se modifica por la presencia de EGTA. Los datos mostrados son
representativos de un mínimo de 3 experimentos independientes.

44
7. DISCUSION
Nuestro Laboratorio ha estado estudiando la acción de la insulina en el
cardiomiocito, su sistema de transducción y su vinculación con el metabolismo
energético, específicamente a nivel del transporte de glucosa y la regulación de la
dinámica mitocondrial. En un trabajo previo realizado en cultivos primarios de
cardiomiocitos neonatos se encontró que el calcio intracelular media los efectos de
insulina a nivel de la translocación del GLUT4 a la membrana plasmática y el posterior
ingreso de glucosa a estas células. El cardiomiocito neonato es el principal modelo
utilizado en la actualidad, caracterizándose por obtener ATP a través del metabolismo
de la glucosa. Por el contrario, el cardiomiocito adulto normal genera energía a través
de la utilización de ácidos grasos. Sin embargo, bajo condiciones de estrés se produce
un switch metabólico en el cardiomiocito adulto, retornando al uso de glucosa como
principal combustible energético. Dado estos antecedentes, en esta tesis se investigó
si esta nueva vía transduccional del receptor de insulina dependiente de calcio también
opera en el cardiomiocito adulto. En primer término en el presente trabajo se corroboró
la presencia del receptor de insulina en el cardiomiocito de rata adulta y que su
activación produce la autofosforilación del receptor y posterior aumento de los niveles
de las formas fosforiladadas de Akt y Erk ya a los 5 minutos de estimulación con
insulina tal como se ha observado en estudios anteriores (56). Estos hallazgos
permiten establecer que insulina activa la vía canónica de señalización del receptor de
insulina en el cardiomiocito adulto.
Por otra parte, utilizando las mismas herramientas experimentales que permitieron
estudiar los cambios en los niveles intracelulares de calcio en el cardiomiocito neonato,
no fue posible detectar movimientos de calcio intracelulares inducidos por insulina en el
cardiomiocito adulto. Insulina no estimuló el ingreso de calcio extracelular ni su
movilización desde reservorios intracelulares, pues células incubadas en ausencia de
calcio externo y otras en presencia de calcio externo no mostraron cambios en los
niveles de calcio al ser estimuladas con insulina de manera detectable con la sonda
utilizada. Más aún, los análisis realizados en las distintas zonas celulares del
cardiomiocito adulto arrojaron los mismos resultados. En ninguna de las zonas
analizadas, es decir, ni en el citoplasma, núcleo ni en la membrana celular se logró

45
detectar variaciones de los niveles de calcio inducidos por insulina. Estos datos difieren
en parte con algunos estudios previos donde se logró detectar aumentos de calcio
inducidos por insulina en núcleos de hepatocitos de rata adulta con la sonda Fluo4- AM
(50) y en los cuales a su vez en músculo esquelético se evaluaron los niveles de calcio
utilizando una sonda lipofílica derivada de Indo-1, conocida como FIP18, que permite
evaluar las señales de calcio en la zona cercana a la membrana celular (57), llegando
a la conclusión de que insulina producía aumentos en los niveles de calcio sólo en la
zona cercana a la membrana celular pero no en el calcio citosólico global detectado
utilizando la sonda Indo-1 que es de ubicación citoplasmática (51). No obstante, se
puede estar seguro de la bioactividad de insulina a partir de los resultados obtenidos
de los estudios de inmunofluorescencia y Western blot.
Para eliminar la posibilidad de la ausencia de resultados positivos debido a perdida
de la viabilidad celular, además de evaluar la morfología celular, en cada uno de los
registros de movimientos de calcio se realizaron controles positivos que permitieron
establecer que la célula seleccionada podría responder a estímulos que aumentan los
niveles de calcio intracelular. Con este fin, se utilizó KCl para despolarizar la membrana
celular (58) o ionomicina, un ionóforo para calcio utilizado para investigar procesos que
requieren movimientos de calcio (59). Las células estudiadas respondieron a estos
estímulos aumentando sus niveles intracelulares de calcio comprobando que nuestra
sonda detectaba niveles de calcio elevados y que las células respondían a estímulos
que producen aumentos del catión en su interior.
Como se mencionó anteriormente, nuestro laboratorio determinó la presencia de
transientes de calcio estimuladas por insulina en cardiomiocitos neonatos logrando
establecer que estas transientes estaban compuestas por dos señales claramente
identificadas. La primera dada por el ingreso de calcio desde el extracelular a través de
los canales dependientes de voltaje de tipo L y una posterior apertura de los canales
de calcio receptores de Ryanodina permitiendo un aumento brusco y agudo en el calcio
citosólico. La segunda señal está compuesta por la salida de calcio desde el interior del
retículo sarcoplásmico pero esta vez mediada por el receptor de IP3 (55). En base a
estos antecedentes se evaluó movimientos de calcio en cardiomiocitos neonatos con el
propósito de determinar si éramos capaces de detectar movimientos de calcio
inducidos por insulina en este modelo celular y así poder descartar la posibilidad de un
mal manejo del protocolo de medición de calcio. Es así como a partir de estos análisis

46
se logró reproducir los resultados obtenidos previamente en el laboratorio en
cardiomiocitos neonatos, lo cual indica que el procedimiento efectuado para concretar
las mediciones y los análisis de las mediciones de calcio se ejecutó adecuadamente.
También se estudió por microscopía confocal en el formato “line scan” si insulina
modifica la frecuencia de las espigas de calcio asociadas a la contracción. Los
resultados mostraron que insulina no modificó la frecuencia de contracción de
cardiomiocitos adultos a ninguna de las concentraciones ensayadas de manera
detectable con Fluo3- AM.
Debido a la similitud entre insulina e IGF-1 y a otros trabajos efectuados en nuestro
Laboratorio, se evaluó si IGF-1 produciría movimientos de calcio en el cardiomiocito
adulto. Como se indicó antes, datos previos señalan que IGF-1 aumenta los niveles de
calcio en cardiomiocitos de rata neonata a través de una vía de señalización que
involucra la participación de PI3K- PLC- IP3 (30). En base a estos antecedentes
efectuamos mediciones de calcio a través de microscopía confocal similares a los
realizados con insulina. IGF-1 no aumentó los niveles intracelulares de calcio ya sea
por su movilización desde el extracelular o del almacenado en reservorios
intracelulares. Usando los controles positivos respectivos y realizando evaluaciones
morfológicas de las células estudiadas, se estableció que las células eran viables. Sin
embargo, se observó que el 39% de las células estudiadas respondían a
concentraciones muy altas de IGF-1 (1 µM). Tampoco IGF-1 logró cambios en las
espigas contráctiles del cardiomiocito adulto, pudiendo establecer que IGF-1, al igual
que insulina, no modificó la frecuencia de contracción.
Finalmente se evaluó la participación del calcio en la activación de la vía de
señalización canónica de insulina en nuestro modelo celular. Los niveles del receptor
de insulina fosforilado no se modificaron por la ausencia de calcio intracelular lograda
con la presencia de BAPTA- AM ni en ausencia de calcio extracelular lograda con la
presencia de EGTA. Sin embargo, la fosforilación de la proteína kinasa Akt
dependiente de insulina disminuyó en presencia de BAPTA- AM, pero no en presencia
de EGTA. Estos resultados sugieren que el calcio intracelular claramente tiene un rol
en la activación de la vía de señalización de insulina en el cardiomiocito adulto ya que
la fosforilación de Akt en ausencia de este catión cae a niveles cercanos al basal. Por
lo tanto, este último dato invita a meditar acerca de la posibilidad de que tal vez la
sonda utilizada para medir los niveles de calcio intracelular no es la más adecuada,

47
aunque Fluo3- AM posee una Kd de 390 nM y en consecuencia tiene una sensibilidad
adecuada.
Los cardiomiocitos adultos son células bastante grandes que poseen túbulos T en
su membrana en estrecha relación con el retículo sarcoplásmico. Por lo tanto, en caso
de producirse liberaciones de calcio desde el retículo sarcoplásmico inducidos por
insulina, estas señales podrían estar localizadas en la proximidad de la membrana
plasmática del cardiomiocito adulto. Es decir, podría existir una compartamentalización
de las señales de calcio. En nuestros análisis intentamos medir calcio en la cercanía de
la membrana plasmática, sin embargo esta es una aproximación gruesa ya que esa
zona claramente puede estar siendo enmascarada por el calcio que permanece en el
citoplasma celular. Para evitar este problema y como una manera de descartar
definitivamente la posibilidad de movimientos de calcio inducidos por insulina en el
cardiomiocito adulto a futuro se sugiere usar una sonda que permita localizar las
mediciones en la región próxima a la membrana celular que es donde se encuentran
los canales de calcio tipo L en los túbulos T en estrecha relación con el retículo
sarcoplásmico de las células. Otra posibilidad sería utilizar microscopía TIRF (Total
internal reflection fluorescence), la cual permite excitar sólo los fluoróforos ubicados en
los primeros 200 nM desde la superficie a estudiar.
En resumen, existen tareas pendientes que permitirían comprobar o rechazar
nuestra hipótesis de trabajo ya que con los antecedentes actuales sólo podemos decir
que insulina no induce movimientos de calcio en el cardiomiocito adulto de manera
detectable con la sonda Fluo3- AM, pero si podemos afirmar que el calcio participa en
la activación de la vía de señalización de insulina de una forma aún desconocida. No
obstante, un gran aporte de este trabajo a nuestra experiencia como investigadores es
el hecho de que trabajar con un modelo celular u otro, en este caso con cardiomiocitos
neonatos o adultos, marca una gran diferencia en los resultados que podrían
obtenerse. Como se sabe, existen distintas preferencias en cuanto a los sustratos
metabólicos utilizados por cardiomiocitos neonatos y adultos. Los primeros utilizan
principalmente glucosa para obtener energía y los segundos ácidos grasos. Esta
diferencia podría ser de gran relevancia a la hora de mirar objetivamente los resultados
obtenidos en el presente trabajo y constituir una razón por la cual los cardiomiocitos en
sus distintas etapas de desarrollo se comportarían de manera disímil.

48
Cabe destacar que la presente investigación es una primera aproximación al estudio
de enfermedades cardiovasculares vinculadas a la diabetes, como por ejemplo
cardiomiopatía diabética, en el modelo adulto que es comúnmente el afectado por
estas patologías. El conocimiento del comportamiento normal de los cardiomiocitos
adultos frente a insulina es el primer paso para posteriormente estudiar y entender las
modificaciones celulares en el estado patológico, específicamente en condiciones de
hipertrofia celular, característica principal de la cardiomiopatía diabética.

49
8. CONCLUSIONES
El receptor de insulina está presente en el cardiomiocito de rata adulta de
manera relativamente homogénea en el citoplasma celular y en la membrana
celular.
Insulina activa su cascada de señalización canónica en el cardiomiocito adulto,
estimulando la fosforilación de su receptor y las proteínas kinasas Akt y Erk1/2.
Insulina e IGF-1, a las distintas concentraciones ensayadas, no aumentaron los
niveles de calcio, evaluados por microscopía confocal y la sonda sensora
Fluo3-AM, en los compartimentos citosólico, nuclear y en la membrana celular
del cardiomiocito adulto.
La fosforilación de la proteína kinasa Akt dependiente de insulina es regulada
por el calcio intracelular en el cardiomiocito adulto.
En base a estos antecedentes, el calcio forma parte del sistema transduccional
del receptor de insulina en el cardiomiocito neonato pero no en el cardiomiocito
adulto.

50
9. BIBLIOGRAFIA
(1) Heart book, Yale University, Pag: 3-10; 133-148.
(2) Encuesta nacional de salud (2003), ministerio de salud, Chile, 2004.
(3) Alain G. Bertoni, Arthur Tsai, Edward K. Kasper, Frederick L. Brancati Diabetes
And Idiopathic Cardiomyopathy. Diabetes Care, 26:2791–2795, 2003.
(4) Lars Ryde´n, Karolinska Sjukhuset, Diabetes mellitus and congestive heart
failure. European Heart Journal, 20, 789–795, 1999.
(5) Sihem Boudina, E. Dale Abel. Diabetic Cardiomyopathy Revisited. Circulation,
115;3213-3223, 2007.
(6) Indu G. Poornima, Pratik Parikh, Richard P. Shannon. Diabetic Cardiomyopathy
The Search for a Unifying Hypothesis. Circ Res. ;98:596-605, 2006.
(7) Masaki Ieda, Takatoshi Tsuchihashi, Kathryn N. Ivey, Robert S. Ross, Ting-
Ting Hong, Robin M. Shaw, and Deepak Srivastava. Cardiac Fibroblasts
Regulate Myocardial Proliferation through β1Integrin Signaling. Dev Cell; 16(2):
233–244, 2009.
(8) Norifumi Takeda, Ichiro Manabe, Yuichi Uchino, Kosei Eguchi, Sahohime
Matsumoto, Simon J. Conway, Ryozo Nagai. Cardiac fibroblasts are essential
for the adaptive response of the murine heart to pressure overload. J. Clin.
Invest. 120:254–265, 2010.
(9) Ichiro Manabe, Takayuki Shindo, Ryozo Nagai. Gene Expression in Fibroblasts
and Fibrosis Involvement in Cardiac Hypertrophy. Circ Res.;91:1103- 1113,
2002.

51
(10) Nawazish Naqvi, Ming Li, Eiji Yahiro, Robert M. Graham, Ahsan Husain.
Insights into the Characteristics of Mammalian Cardiomyocyte Terminal
Differentiation Shown Through the Study of Mice with a Dysfunctional c-Kit.
Pediatr Cardiol.; 30(5): 651–658, 2009.
(11) Topi Korhonen, Sandra L. Hanninen, Pasi Tavi. Model of Excitation-Contraction
Coupling of Rat Neonatal Ventricular Myocytes. Biophysical Journal, vol 96,
1189–1209 2009.
(12) Noritsugu Tohse, Sumihiko Seki, Takeshi Kobayashi, Masaaki Tsutsuura,
Masato Nagashima, Yoichi Yamada. Development of Excitation- Contraction
Coupling in Cardiomyocytes. Japanese Journal of Physiology, 54, 1-6, 2004.
(13) Michael Ibrahim, Abeer Al Masri, Manoraj Navaratnarajah, Urszula Siedlecka,
Magdi H. Yacoub, Cesare M. N. Terracciano. Prolonged mechanical unloading
affects cardiomyocyte excitation-contraction coupling, transverse-tubule
structure, and the cell surface. The FASEB Journal. Vol 24, (3321- 3329), 2010.
(14) A-Olufemi Makinde, Paul F. Kantor and Gary D. Lopaschuk. Maturation of fatty
acid and carbohydrate metabolism in the newborn heart. Molecular and Cellular
Biochemistry 188: 49–56, 1998.
(15) John J. Lehman, MD, and Daniel P. Kelly, MD. Gene Regulatory Mechanisms
Governing Energy Metabolism during Cardiac Hypertrophic Growth. Heart
Failure Reviews, 7, 175–185, 2002.
(16) Mattia Falconi, Maria Teresa Cambria, Antonio Cambria and Alessandro
Desideri. Structure and Stability of the Insulin Dimer Investigated by Molecular
Dynamics Simulation. Journal of Biomolecular Structure &Dynamics.18 (5): 761-
772, 2001.
(17) Patrick E MacDonald, Jamie W Joseph and Patrik Rorsman. Glucose-sensing
mechanisms in pancreatic b-cells. Phil. Trans. R. Soc. B. 360, 2211-2225, 2005.

52
(18) Leonid E Fridlyand and Louis H Philipson. Glucose sensing in the pancreatic
beta cell: a computational systems analysis. Theoretical Biology and Medical
Modelling. 7:15, 2010.
(19) Cecil C. Yip and Peter Ottensmeyer. Three-dimensional Structural Interactions
of Insulin and Its Receptor. The Journal of Biological Chemistry vol. 278, No. 30,
Issue of July 25, pp. 27329–27332, 2003.
(20) Yoshiaki Kido, Jun Nakae, and Domenico Accili. The Insulin Receptor and Its
Cellular Targets. The Journal of Clinical Endocrinology & Metabolism. vol. 86,
No. 3, 2001.
(21) Cullen M. Taniguchi, Brice Emanuelli and C. Ronald Kahn. Critical nodes in
signalling pathways: insights into insulin action. Nature Reviews, Molecular Cell
Biology. vol 7, Febrero 2006.
(22) Kangduk Choi and Young-Bum Kim. Molecular Mechanism of Insulin
Resistance in Obesity and Type 2 Diabetes. The Korean Journal of Internal
Medicine, vol. 25, No. 2, Junio 2010.
(23) Antti Virkamäki, Kohjiro Ueki, and C. Ronald Kahn. Protein–protein interaction
in insulin signaling and the molecular mechanisms of insulin resistance. The
Journal of Clinical Investigation. Vol 103, No 7, Abril 1999.
(24) Jacques Pouysse´ gur and Philippe Lenormand. Fidelity and spatio-temporal
control in MAP kinase (ERKs) signalling. Eur. J. Biochem. 270, 3291–3299,
2003.
(25) Brian J. DeBosch, Anthony J. Muslin. Insulin signaling pathways and cardiac
growth. Journal of Molecular and Cellular Cardiology, 44; 855–864, 2008.
(26) Jun Liu, Stephanie M. DeYoung, Joseph B. Hwang, Erin E. O’Leary, and Alan
R. Saltiel. The Roles of Cbl-b and c-Cbl in Insulin-stimulated Glucose Transport.

53
The Journal Of Biological Chemistry. Vol. 278, No. 38, 36754–36762,
Septiembre 2003.
(27) Graziella Castellano, Flora Affuso, Pasquale Di Conza and Serafino Fazio. The
GH/IGF-1 Axis and Heart Failure. Current Cardiology Reviews, 5, 203-215,
2009.
(28) Joelle Dupont, Javed Khan, Bao-He Qu, Paul Metzler, Lee Helman, And Derek
Leroith. Insulin and IGF-1 Induce Different Patterns of Gene Expression in
Mouse Fibroblast NIH-3T3 Cells: Identification by cDNA Microarray Analysis.
Endocrinology 142(11):4969–4975, 2001.
(29) A Espinosa, M Estrada and E Jaimovich. IGF-I and insulin induce different
intracellular calcium signals in skeletal muscle cells. Journal of Endocrinology
182, 339–351, 2004.
(30) Cristian Ibarra, Manuel Estrada, Loreto Carrasco, Mario Chiong, José L.
Liberona, Cesar Cárdenas, Guillermo Díaz-Araya, Enrique Jaimovich, and
Sergio Lavandero. Insulin-like Growth Factor-1 Induces an Inositol 1,4,5-
Trisphosphate-dependent Increase in Nuclear and Cytosolic Calcium in Cultured
Rat Cardiac Myocytes. The Journal Of Biological Chemistry. Vol. 279, No. 9,
7554–7565, 2004.
(31) Roger W. Brownsey , Adrienne N. Boone , Michael F. Allard. Actions of insulin
on the mammalian heart: metabolism, pathology and biochemical mechanisms.
Cardiovascular Research ,34; 3–24, 1997.
(32) Ding An and Brian Rodrigues. Role of changes in cardiac metabolism in
development of diabetic cardiomyopathy. Am J Physiol Heart Circ Physiol 291:
H1489–H1506, 2006.
(33) Joost J. F. P. Luiken, Susan L. M. Coort, Debby P. Y. Koonen, Dick J. van der
Horst . Arend Bonen . Antonio Zorzano . Jan F. C. Glatz. Regulation of cardiac

54
long-chain fatty acid and glucose uptake by translocation of substrate
transporters. Eur J Physiol, 448: 1–15, 2004.
(34) Johanna T Lanner, Joseph D Bruton, Abram Katz and Hakan Westerblad.
Ca2+ and insulin-mediated glucose uptake. Current Opinion in Pharmacology,
8:339–345, 2008.
(35) Martin OJ, Lee A & McGraw TE. GLUT4 distribution between the plasma
membrane and the intracellular compartments is maintained by an insulin-
modulated bipartite dynamic mechanism. Journal of Biological Chemistry, 281
484–490, 2006.
(36) Stagsted J, Olsson L, Holman GD, Cushman SW, Satoh S. Inhibition of
internalization of glucose transporters and IGF-II receptors. Mechanism of action
of MHC class I-derived peptides which augment the insulin response in rat
adipose Cells. J Biol Chem, 268:22809–22813, 1993.
(37) Izaäk Frederik Kodde, Johan van der Stok, Ryszard T. Smolenski, Jan Willem
de Jong. Metabolic and genetic regulation of cardiac energy substrate
preference. Comparative Biochemistry and Physiology, Part A 146, 26–39,
2007.
(38) Cristiana Vitale, Peter Collins. Optimization of Cardiac Metabolism in Diabetes
Mellitus. Current Pharmaceutical Design, 14, 2537-2550, 2008.
(39) Joanne S. Ingwall. Energy metabolism in heart failure and remodelling.
Cardiovascular Research, 81, 412–419, 2009.
(40) Kurland IJ, Pilkis SJ. Covalent control of 6-phosphofructo-2- kinaserfructose-
2,6-bisphosphatase: insights into autoregulation of a bifunctional enzyme.
Protein Sci; 4:1023–1037, 1995.
(41) Louis Hue, Christophe Beauloye, Anne-Sophie Marsin, Luc Bertrand, Sandrine
Horman and Mark H. Rider. Insulin and Ischemia Stimulate Glycolysis by Acting

55
on the Same Targets Through Different and Opposing Signaling Pathways.
Journal of Molecular and Cellular Cardiology, 34, 1091- 1097, 2002.
(42) Christophe Depre, Mark H. Rider And Louis Hue. Mechanisms of control of
heart glycolysis. Eur. J. Biochem, 258, 277-290, 1998.
(43) Martin D. Bootman, Peter Lipp and Michael J. Berridge. The organisation and
functions of local Ca2+ signals. Journal of Cell Science, 114, 2213-2222, 2001.
(44) Martin D. Bootman, Daniel R. Higazi, Stephen Coombes, H. Llewelyn Roderick.
Calcium signalling during excitation-contraction coupling in mammalian atrial
myocytes. Journal of Cell Science 119, 3915-3925, 2006.
(45) Michael J. Berridge,Martin D. Bootman and H. Llewelyn Roderick. Calcium
Signalling: Dynamics, Homeostasis And Remodelling. Molecular Cell Biology, 4,
517-529, 2003.
(46) Min Fu, Meng Wu, Ji-Feng Wang, Yan-Jiang Qiao, Zhao Wang. Disruption of
the intracellular Ca2+ homeostasis in the cardiac excitation–contraction coupling
is a crucial mechanism of arrhythmic toxicity in aconitine-induced
cardiomyocytes. Biochemical and Biophysical Research Communications, 354,
929–936, 2007.
(47) Susumu Minamisawa, Yoji Sato, Myeong-Chan Cho. Calcium cycling proteins
in heart failure, cardiomyopathy and arrhythmias. Experimental and Molecular
Medicine, Vol. 36, No. 3, 193-203, 2004.
(48) Nazmi Yaras, Mehmet Ugur, Semir Ozdemir, Hakan Gurdal, Nuhan Purali,
Alain Lacampagne, Guy Vassort, and Belma Turan. Effects of Diabetes on
Ryanodine Receptor Ca Release Channel (RyR2) and Ca2 Homeostasis in Rat
Heart. Diabetes 54,3082- 3088 ,2005.
(49) Grant N. Pierce, James C. Russell. Regulation of intracellular Ca2q in the heart
during diabetes. Cardiovascular Research 34, 41–47, 1997.

56
(50) Michele A. Rodrigues, Dawidson A. Gomes, Viviane A. Andrade, M. Fatima
Leite, Michael H. Nathanson. Insulin Induces Calcium Signals in the Nucleus of
Rat Hepatocytes. Hepatology 48(5):1621–1631, 2008.
(51) Joseph D. Bruton, Abram Katz, Håkan Westerblad. Insulin increases near-
membrane but not global Ca21 in isolated skeletal muscle. Proc. Natl. Acad.
Sci. USA 96, 3281–3286, 1999.
(52) Zhongju Lu, Ya-Ping Jiang, Xin-Hua Xu, Lisa M. Ballou, Ira S. Cohen, Richard
Z. Lin. Decreased L-Type Ca2_ Current in Cardiac Myocytes of Type 1 Diabetic
Akita Mice Due to Reduced Phosphatidylinositol 3-Kinase Signaling. Diabetes
56, 2780-2789, 2007.
(53) Sebastian Maier, Friedbert Aulbach, Andreas Simm, Volkmar Langec, Heiner
Langenfeld,Holger Behre , Ulrich Kersting , Ulrich Walter , Michael Kirstein.
Stimulation of L-type Ca current in human atrial myocytes by insulin.
Cardiovascular Research 44, 390–397 1999.
(54) Friedbert Aulbach , Andreas Simm , Sebastian Maier , Heiner Langenfeld ,
Ulrich Walter,Ulrich Kersting, Michael Kirstein. Insulin stimulates the L-type Ca
current in rat cardiac myocytes. Cardiovascular Research 42, 113–120, 1999.
(55) A. E. Contreras-Ferrat, B. Toro, R. Bravo, V. Parra, C. Vásquez, C. Ibarra, D.
Mears, M. Chiong, E. Jaimovich, A. Klip, and S. Lavandero. An Inositol 1,4,5-
Triphosphate (IP3)-IP3 Receptor Pathway Is Required for Insulin-Stimulated
Glucose Transporter 4 Translocation and Glucose Uptake in Cardiomyocytes.
Endocrinology, 151(10):0000–0000, 2010.
(56) Christophe Montessuit, Irène Papageorgiou, and René Lerch. Nuclear
Receptor Agonists Improve Insulin Responsiveness in Cultured Cardiomyocytes
through Enhanced Signaling and Preserved Cytoskeletal Architecture.
Endocrinology 149(3):1064–1074, 2008.

57
(57) Lothar A. Blatter’, Ernst Niggli. Confocal nearmembrane detection of calcium in
cardiac myocytes. Cell Calcium, 23(5), 269-279, 1998.
(58) Paul H. Ratz1,2, Amy S. Miner, and Suzanne E. Barbour. Calcium-independent
phospholipase A2 participates in KClinduced calcium sensitization of vascular
smooth muscle. Cell Calcium. 2009 July; 46(1): 65–72, 2009.
(59) Zhanghua Gao, Yingfa Li, John P. Cooksey, Thomas N. Snaddon, Stefan
Schunk, Eddy M. E. Viseux, Stephen M. McAteer, and Philip J. Kocienski. A
Synthesis of an Ionomycin Calcium Complex. Angew. Chem. Int. Ed. 48, 5022 –
5025, 2009.