Organelos celulares
37
Organelos celulares Estructura y función APARATO DE GOLGI El aparato de Golgi, es también llamado complejo o cuerpo de Golgi, fue descrito por primera vez por Camilo Golgi en 1898 y su existencia fue cuestionada durante décadas. La técnica del reactivo de Schiff-ácido peryódico marca los glúcidos del Golgi y esto hace que se puede ver con el microscopio óptico. Su estructura membranosa fue observada por primera vez al microscopio electrónico por Dalto y Felix en 1954. Se encarga de la distribución y el envío de los productos químicos de la célula. En las células animales es un orgánulo que se localiza próximo al centrosoma, el cual suele estar en las cercanías del núcleo. Como el centrosoma es el principal centro organizador de microtúbulos, la posición central del aparato de Golgi en las células animales depende de la organización del sistema de microtúbulos. Por tanto la existencia de este orgánulo en posición central en las células animales depende de la organización del sistema de microtúbulos Modifica proteínas y lípidos (grasas) que han sido construidos en el retículo endoplasmático y los prepara para expulsarlos fuera de la célula. Estructura
-
Upload
luisalberg -
Category
Documents
-
view
22 -
download
0
description
Estructuras principales
Transcript of Organelos celulares
Organelos celularesEstructura y funcinAPARATO DE GOLGIEl aparato de Golgi, es tambin llamado complejo o cuerpo de Golgi, fue descrito por primera vez por Camilo Golgi en 1898 y su existencia fue cuestionada durante dcadas. La tcnica del reactivo de Schiff-cido perydico marca los glcidos del Golgi y esto hace que se puede ver con el microscopio ptico. Su estructura membranosa fue observada por primera vez al microscopio electrnico por Dalto y Felix en 1954. Se encarga de la distribucin y el envo de los productos qumicos de la clula.En las clulas animales es un orgnulo que se localiza prximo al centrosoma, el cual suele estar en las cercanas del ncleo. Como el centrosoma es el principal centro organizador de microtbulos, la posicin central del aparato de Golgi en las clulas animales depende de la organizacin del sistema de microtbulos. Por tanto la existencia de este orgnulo en posicin central en las clulas animales depende de la organizacin del sistema de microtbulosModifica protenas y lpidos (grasas) que han sido construidos en el retculo endoplasmtico y los prepara para expulsarlos fuera de la clula.EstructuraEl aparato de Golgi est formado por una serie de grupos de cisternas aplanadas o dictiosomas que se disponen regularmente formando pilas. En una clula puede haber varios de estos apilamientos y algunas cisternas localizadas en dictiosomas prximos estn conectadas lateralmente. A todo el conjunto de pilas y sus conexiones es a lo que se denomina complejo del Golgi. El nmero (normalmente de 3 a 8) y el tamao de las cisternas en cada dictiosoma es variable y depende del tipo celular, as como del estado fisiolgico de la clula.
Entre las cisternas, dentro de cada pila, existen numerosas protenas fibrosas en las que se encuentran embebidas las cisternas. Este entramado, denominado matriz, podra ayudar en el mantenimiento de la estructura del orgnulo. Sin embargo, se ha demostrado que la integridad del aparato de Golgi depende principalmente de la organizacin del sistema de microtbulos, por ejemplo, durante la mitosis desaparece, y tambin del trfico vesicular desde el retculo endoplasmtico, si ste se detiene el Golgi tambin desaparece.En las clulas vegetales, que no tienen centrosoma, hay numerosas estructuras similares a aparatos de Golgi poco desarrollados dispersas por el citoplasma. En las clulas vegetales las cisternas del aparato de Golgi son ms pequeas que en las clulas animales y no estn unidas formando apilamientos, aunque el nmero de cisternas dispersas puede variar entre decenas y ms de cien.El aparato de Golgi es un orgnulo polarizado. En un dictiosoma se diferencian dos caras diferentes, una cara cis o de entrada y una cara trans o de salida. La cara cis est relacionada con el retculo endoplasmtico y de la cara trans surgen diferentes vesculas de transporte que se dirigen a sus destinos finales: la membrana plasmtica, los lisosomas y las vesculas de secrecin.En el lado cis existe un proceso continuo de formacin de cisternas con material procedente de la fusin de compartimentos tbulo vesiculares denominados ERGIC (endoplasmic reticulum Golgi intermediate compartment), que se forman con material proveniente del retculo endoplasmtico. El lado trans tambin posee una organizacin tbulo-vesicular denominada TGN (entramado trans del aparato de Golgi o trans Golgi network), donde las cisternas con las molculas procesadas se deshacen en vesculas que se dirigen a otros compartimentos celulares. Por tanto se da un trasiego constante de molculas desde el lado cis al trans, pero tambin hay un flujo de reciclado en sentido contrario mediado por vesculas recubiertas con protenas COP I que parten de las zonas laterales de las cisternas y se dirigen hacia otras cisternas ms prximas al lado cis. Es un orgnulo en constante renovacin y el flujo de molculas afecta a su organizacin y a su tamao. Este orgnulo est especialmente desarrollado en clulas con fuerte secrecin.FuncionesEl aparato de Golgi desempea funciones esenciales para la clula:Interviene en procesos de secrecin. Las protenas destinadas a ser secretadas al exterior son sintetizadas en el RE rugoso y posteriormente transferidas al complejo de Golgi, donde son empaquetadas en las vesculas de secrecin. Dichas vesculas se fusionan con la membrana plasmtica y vierten su contenido al exterior por exocitosis. Puede tratarse de una secrecin constitutiva, en la que las protenas que abandonan la clula no estn sealizadas y es la que se produce por defecto en todas las clulas; las vesculas estn recubiertas por protenas de tipo COP I. Puede ser tambin una secrecin regulada, realizada por clulas especializadas; en ellas las protenas quedan almacenadas en vesculas recubiertas por clatrina y como respuesta a una seal extracelular se fusionan con la membrana plasmtica y las protenas abandonan la clula.Reciclaje de la membrana plasmtica. Durante la exocitosis, la membrana de las vesculas secretoras pasa a formar parte de la membrana plasmtica. Esto permite reponer los componentes de la membrana que se incorporan a la clula por endocitosis, lo que consitituye un reciclaje de la membrana plasmtica.Glicosilacin. El complejo de Golgi contiene numerosas enzimas glicosil-transferasas que realizan reacciones de glicosilacin, tanto en molculas de lpidos como de protenas, a medida que estas van pasando a travs del complejo de Golgi. En estas reacciones se eliminan algunos de los azcares del oligosacrido aadido en el RE y se adicionan otros. La glucosidacin en las protenas puede ser de dos tipos: a) Los sacridos ricos en manosa aadidos en el retculo endoplasmctico (glucosidacin tipo N) son posteriormente modificados en el aparato de Golgi mediante la adicin de nuevos azcares. b) En el aparato de Golgi se aade oligosacridos con unin tipo O a los grupos hidroxilos de aminocidos como la serina, la treonina y la hidroxilisina. Este tipo de glucosilacin ocurre en los proteoglucanos. Tambin se aaden los grupos sulfatos a los glucosaminoglucanos. Formacin de lisosomas.Formacin de vacuolas en las clulas vegetales.Interviene adems en otras funciones como son la sntesis de los componentes de la matriz extracelular en las clulas animales, la sntesis de los componentes de la matriz de la pared celular, tales como pectinas y hemipectinas, en las clulas vegetales, la formacin del tabique telofsico en las clulas vegetales,...LISOSOMAS Los lisosomas son vesculas rodeadas de membrana que contienen enzimas hidrolticas encargadas de las digestiones intracelulares. Metchnikoff y sus colaboradores articularon a finales del siglo XIX la idea de que el material fagocitado era digerido en compartimentos intracelulares acidificados. Estos compartimentos fueron denominados lisosomas y aparecen en todas las clulas eucariotas. Son orgnulos muy heterogneos en cuanto a su forma y tamao. Su diversidad refleja la amplia variedad de materiales que estn digiriendo.Se han encontrado aproximadamente 40 tipos de enzimas lisosomales, todas ellas hidrolasas cidas, cuyo pH ptimo es prximo a 5. Entre ellas hay proteasas, nucleasas, glicosidasas, lipasas, fosfatasas, sulfatasas y fosfolipasas. No todos los lisosomas son iguales y pueden contener juegos diferentes de enzimas. La membrana de los lisosomas protege al resto de la clula de esta actividad destructora, pero si sta se rompiese el pH citoplasmtico, prximo a 7.2, sera un obstculo para la actividad de estas enzimas; esta membrana es por tanto resistente a la accin de las enzimas hidrolticas debido a que la mayora de sus protenas estn muy glicosiladas. Dicha membrana contiene una bomba de protones que mantiene el pH cido en el interior del lisosoma y protenas de transporte que permiten que los productos resultantes de la digestin pasen al citosol.Los lisosomas se forman a partir de vesculas que se desprenden del aparato de Golgi y reciben distintos nombres segn el estado de degradacin de las molculas que contienen: primarios, secundarios y cuerpos residuales. Los cuerpos residuales contienen material que ya no puede ser degradado y quedan almacenados en el interior celular o, como veremos ms adelante, se fusionan con la membrana plasmtica expulsando dicho material el medio extracelular.FuncionesLa funcin principal de los lisosomas es intervenir en la digestin intracelular de macromolculas.Dependiendo de la procedencia del material implicado en la digestin se pueden distinguir tres procesos diferentes:Los lisosomas son considerados como la estacin final de la va endoctica. La mayora de las molculas que van a ser degradadas por esta va tienen que pasar previamente por los endosomas. Las protenas que no se reciclan de nuevo a la membrana plasmtica o al TGN del aparato de Golgi desde los compartimentos endosomales son degradas en los lisosomas. La formacin de los lisosomas es un asunto controvertido. Unos autores proponen que se forman por gemacin o maduracin a partir de los endosomas tardos que ya contienen todas las enzimas degradativas necesarias as como las molculas a degradar. Otros autores proponen que los lisosomas son orgnulos independientes de los endosomas y que reciben vesculas desde los endosomas o se producen fusiones entre endosomas tardos y lisosomas.Para que las protenas integrales de la membrana plasmtica sean dirigidas a los lisosomas se ha de producir una ubiquitinacin de su parte citoslica, es decir, la adicin de una molcula denominada ubiquitina. Ello es necesario para que las protenas de membrana interaccionen con la maquinaria de reparto que se encuentran en los endosomas y no vuelvan a la membrana citoplasmtica en vesculas de reciclado. Las interacciones con diversos complejos proteicos mantienen a las protenas ubiquitinadas en zonas limitadas de la membrana endosomal, que poseen una cubierta en la que est presente la clatrina. Todo ello hace que sean retenidas en los endosomas tempranos y depus transportadas a los cuerpos multivesiculares, a los endosomas tardos, y de ah a los lisosomas, donde se degradan. Este mecanismo afecta a receptores, transportadores, canales, etctera. Los receptores que no son ubiquitinados pero s endocitados, cuando llegan a los endosomas tempranos suelen reciclarse hacia la membrana celular.Las partculas obtenidas por fagocitosis siguen una va propia. Las partculas como bacterias o restos celulares quedan en el interior celular englobadas por membrana formando un compartimento que madurar y se convertir en el denominado fagosoma. La degradacin de estas partculas se produce cuando se fusionan los fagosomas con los lisosomas.Una tercera va de llegada de molculas a los lisosomas es la autofagia. Consise en la digestin de material de origen endgeno, es decir, de partes de la propia clula. Para ello, un orgnulo defectuoso se rodea de membranas procedentes del retculo endoplasmtico, formndose un autofagosoma, el cual se fusiona con un lisosoma y se inicia la digestin. La autofagia est relacionada con el recambio de los componentes celulares: destruye los componentes celulares defectuosos o que ya no son necesarios para la clula; interviene en la destruccin de los tejidos y orgnulos durante la metamorfosis de los insectos y anfibios. Asegura la nutricin en condiciones desfavorables (ayuno), viviendo la clula de sus propios materiales.Pero adems a los lisosomas han de llegar las hidrolasas cidas encargadas de la digestin. stas se empaquetan en vesculas en el TGN del aparato de Golgi, las cuales se fusionarn con los endosomas tardos y desde ah llegan a los lisosomas. En el aparato de Golgi se aade a las enzimas lisosomales un grupo glucdico fosfatado, la manosa-6-fosfato, que es reconocido por un receptor en el TGN del aparato de Golgi. La interaccin del dominio citoslico de este receptor con la cubierta de clatrina permite englobar al receptor ms la hidrolasa en vesculas que se dirigirn hacia los endosomas tardos, y desde ah hasta los lisosomas. Aunque ste sea el mecanimso principal existen otras protenas que no requieren la fosforilacin de las manosas para ir a los lisosomas. Estas protenas son las integrales de la membrana. Estas protenas contienen una secuencia de aminocidos de destino que se encuentra en la cara citoslica de la protena.Capacidad de participar en una exocitosis regulada. Por ejemplo, en el hgado se secretan enzimas lisosmicas a la bilis. Tambin se ha observado la exocitosis de orgnulos con caractersticas similares a los lisosomas como es el caso de los melanocitos (los grnulos de melanina que pasarn a los queratinocitos que darn el color moreno a la piel). El acrosoma de los espermatozoides, una vescula cargada de numerosas enzimas hidrolticas, se libera durante la fecundacin. Se ha propuesto desde hace tiempo que las clulas eucariotas son capaces de eliminar las sustancias que no pueden degradas ms y esto sera posible si los lisosomas terminan por expulsar su material cuando se fusionan con la membrana plasmtica. En las clulas de mamferos donde slo se produce secrecin constitutiva se ha visto que bajo ciertas condiciones pueden realizar exocitosis regulada, por ejemplo, por una elevacin de la concentracin de calcio intracelular, lo cual ocurre, por ejemplo, durante las pequeas roturas de la membrana citoplasmtica, como vimos en el apartado dedicado a las membranas. VACUOLAS Junto con los cloroplastos y la pared celular, las vacuolas son componentes caractersticos de las clulas vegetales.Una vacuola es una vescula muy grande, llena de lquido y rodeada por una membrana, denominada tonoplasto; esta membrana es selectivamente permeable, e interviene especialmente en el mantenimiento de la turgencia celular y en el crecimiento. Se forman por fusin de vesculas procedentes del aparato de Golgi.El nmero y tamao de las vacuolas vara segn el tipo de clula y las distintas fases del desarrollo. La clula vegetal inmadura contiene gran cantidad de pequeas vacuolas, que aumentan de tamao y se fusionan entre s cuando la clula crece. En la clula madura, una sola vacuola puede llegar a ocupar hasta el 90% del volumen celular, quedando el citoplasma reducido a una fina capa alrededor de la pared celular.EstructuraLa composicin de las vacuolas vara de acuerdo con el tipo de planta y estado fisiolgico. El principal componen del lquido vacuolar es el agua, que puede contener sales, azcares y protenas en disolucin. A veces, algunas sales estn lo suficientemente concentradas en la vacuola como para formar cristales; son frecuentes los cristales de oxalato clcico que pueden adoptar distintas formas; pueden aparecer con forma de arena cristalina, de agujas en los rafidios, columnas en los estiloides (Eichhornia crassipes), prismtica en los cristales prismticos simples o compuestos: las drusa. Otras vacuolas contienen aceites y esencias.FuncionesLas vacuolas realizan, entre otras, las siguientes funciones:Almacenan gran cantidad de sustancias:Nutritivas, como las protenas de reserva de muchas semillas.Productos de deshecho txicos, que aislan del resto del citoplasma, como la nicotina o el opio. Estas molculas nocivas se liberan de las vacuolas cuando una planta es comida o daada, proporcionando un sistema de defensa contra los depredadores.La vacuola, es a menudo un lugar de concentracin de pigmentos. Los colores azul, violeta, prpura, rojo de las clulas vegetales se deben, usualmente, a un grupo de pigmentos llamados antocianinas (responsables de las coloraciones de frutas y verduras).Poseen actividad digestiva, ya que pueden contener gran variedad de enzimas hidrolticas. Estn implicadas en la degradacin de macromolculas y en el reciclaje de los componentes celulares.Regulan la presin de turgencia. La concentracin de materiales disueltos dentro de la vacuola es muy alta, superior a la del citosol. La entrada de agua en la vacuola por osmosis desde el citosol hace que esta se expanda y cree una presin hidrosttica o turgencia dentro de la clula. Esto est equilibrado por la resistencia mecnica que presenta la pared celular.El aumento de tamao de las clulas vegetales se debe, en gran parte, a la acumulacin de agua en las vacuolas, lo que supone un sistema muy econmico para el crecimiento de las clulas vegetales.En una misma clula puede haber diferentes vacuolas con distintas funciones. Por ejemplo, con funcin lisosmica y con funcin de almacenamiento. Los centrolos.Son corpsculos cilndricos formados por 9 tbulos perifricos simples o tabicados y dos tubos centrales, rigen el movimiento de los cromosomas en las divisiones celulares. Estructura y composicin qumicaCentrolo es cada una de l as dos estructuras de forma cilndrica que se encuentran en el centro de un orgnulo de las clulas eucariticas denominado centrosoma. Al par de centrolos se conoce con el nombre de diplosoma; stos se disponen perpendicularmente entre s.El centrosoma est formado por el diplosoma, el material periocentriolar (material de aspecto amorfo) y las fibras del ster (microtbulos que se organizan en forma de radios). La estructura del centrolo es similar a la parte denominada cuerpo basal o cinetosoma de un cilio o de un flagelo. Consiste en un cilindro abierto, de unos 0,2 por 0,5 micrmetros, cuyas paredes estn formadas por 9 tripletes (grupos de tres) de microtbulos, los cuales se mantienen unidos mediante conexiones, las principales sustancias qumicas que los componen son actina, dineina , nexina, ADN y monmeros alfa y beta de tubulina. No poseen microtbulos centrales como en el caso del axonema o tallo del cilio. Formados por 9 conjuntos de 3 microtbulos, y tienen su propio ADN.Ubicacin Estn cerca del ncleo, hay 2 centrolos por clula y estn presentes solo en clulas animales, aunque pueden existir en vegetales inferiores. Las clulas de los vegetales superiores (angiospermas) carecen de centrolos, por lo que la formacin del huso acromtico se realiza a partir del material periocentolar del centrosoma. En cuanto a su clasificacin solo se puede decir que al conjunto de centrolos se le llama diplosoma.
FuncinLa funcin principal de los centrolos es la formacin y organizacin del os microtbulos que constituyen el huso acromtico en la divisin del ncleo celular. Durante el proceso de divisin de la clula, los centrolos se desplazan hasta colocarse a lados opuestos de la clula, es entonces cuando de cada uno surge un racimo de filamentos radiales al que se le denomina ster. Posteriormente, se forma un huso entre ambos centrolos por medio de los filamentos. Estos filamentos estn compuestos de protena y por cantidades mnimas de cido ribonucleico. Los cromosomas se adhieren a estos filamentos por el centrmero y entonces son empujadas unas a un lado de la clula, y otras al lado contrario.La funcin principal de los centrolos es la formacin y organizacin de los filamentos que constituyen el huso acromtico cuando ocurre la divisin del ncleo celular.Por ejemplo en la espermiognesis las espermtidas se convierten en espermatozoides. Para ello, se reduce el citoplasma, el ncleo se alarga y queda en la cabeza del espermatozoide, las mitocondrias se colocan en el cuello y los centrolos originan un flagelo.
Cloroplastos
Cloroplasto, es el orgnulo citoplasmtico, con forma de disco, de entre 4 y 6 micrmetros de dimetro que se encuentra en las clulas vegetales y en las de las algas, donde se lleva a cabo la fotosntesis (proceso que permite la transformacin de energa luminosa en energa qumica), una cadena de reacciones que utiliza la energa de la luz solar para combinar agua y dixido de carbono en presencia de clorofila y producir oxgeno y azcares tiles para los animales. Sin fotosntesis, la atmsfera no tendra oxgeno suficiente para mantener la vida animal.Estructura y composicin qumica
Como las mitocondrias tambin tienen una membrana interna y una externa, pero a diferencia de las mitocondrias en su matriz cuentan con un sistema interconectado de vesculas planas con forma de disco llamadas tilacoides. Las vesculas tilacoidales se agrupan una sobre otra para formar las estructuras denominadas grana. Dentro de los tilacoides se encuentra la clorofila y dems enzimas que participan en la fotosntesis El estroma (el espacio citoslico que rodea a los tilacoides) es dnde se lleva a cabo la fijacin del carbono (en forma de CO2) en compuestos orgnicos. Al igual que las mitocondrias, los cloroplastos poseen su propio DNA. Los plstidos verdes, llamados cloroplastos, de las plantas y algas verdes son mayores e incluso ms parecidos a bacterias que las mitocondrias. Esto es debido a que: Tienen tambin su propio DNA y mRNA. Sus ribosomas tambin son del mismo tamao que los de las bacterias. Como en el caso de las mitocondrias, los plstidos se encuentran aislados del resto de la clula por una membrana. Su DNA, al igual que el DNA bacteriano, carece de histonas. Tambin se dividen directamente en dos. El DNA, RNA y protenas que producen estos orgnulos son extraordinariamente semejantes a los de las bacterias, especialmente a los de las cianobacterias. Contienen clorofila a y clorofila b.
UbicacinAparecen en mayor cantidad en las clulas de las hojas, lugar en el cual parece que pueden orientarse hacia la luz. En una clula puede haber entre 40 y 50 cloroplastos, y en cada milmetro cuadrado de la superficie de la hoja hay 500.000 cloroplastos.
ClasificacinLa mayora, pero no todos, han conservado los sistemas fotosintticos con pigmentos verdes, rojos o verde azulados. Si bien es cierto que todas las clulas vegetales contienen algn tipo de plstido, los hay tambin incoloros (leucoplastos), no fotosintticos aunque aun no se conoce su funcin. Los plastos evolucionaron a partir de simbiontes fotosintticos ms de una vez; por tanto, son considerados polifilticos, como demuestran los diferentes tipos de plastos: plstidos verdes corrientes de los vegetales (clorofila a y b procedentes de Prochloron), plastos verde azulados y rodoplastos de algas rojas (clorofila a y ficobiliprotenas, procedentes de varios tipos de cianobacterias), y diatomeas y algas pardas (clorofilas a y c).
Funcin
En los cloroplastos se lleva a cabo la funcin cloroflica, de la que depende en la actualidad toda la vida del planeta. Es frecuente encontrar en muchos de ellos cmulos de almidn, formados al polimerizarse la glucosa obtenida durante los procesos de asimilacin fotosinttica del anhdrido carbnico. El proceso ms comn es la fotosntesis. Este proceso se realiza en estructuras especializadas llamadas cloroplastos y que se encuentran principalmente en hojas y partes verdes, de los vegetales.Estos cloroplastos tienen una estructura compleja formada a grandes rasgos por: estroma y tilacoides o granum. Los tilacoides son estructuras vesiculares (en forma de bolsas o discos) que contienen en su interior la molcula de clorofila, responsable de "captar" la energa luminosa (en forma de fotones), excitarse y, mediante este proceso, liberar energa que es captada por algunas molculas como el ADP y el NADP ( que reaccionan formando ATP y NADPH2). Esta reacciones, que dependen directamente de la luz pueden ser de dos tipos: foto fosforilacin cclica y foto fosforilacin acclica. La molcula de clorofila es grande y formada por C, O, H y Mg. En general se presenta en dos formas: clorofila A o alfa y clorofila B o beta. Estos pigmentos varan en su composicin qumica (C55H72N4O5Mg y C55H70N4O6Mg respectivamente), se encuentran en proporciones desiguales dentro de las plantas y reaccionan a diferentes longitudes de onda luminosa (420nm la alfa y 663 nm la beta). Con la energa que se almacena en las reacciones luminosas de la fotosntesis y las molculas de NADPH2 que se forman ah, la clula vegetal puede "fijar" el CO2 atmosfrico y formar molculas complejas como los azcares. Estas reacciones no requieren ya de la intervencin de la luz y se realizan en el estroma del cloroplasto, formando un ciclo conocido como Ciclo de Calvin-Benson. Las molculas de clorofila, que absorben luz para llevar a cabo la fotosntesis, estn unidas a las lamelas. La energa luminosa capturada por la clorofila es convertida en adenosin-trifosfato (ATP) y molculas reductoras (NADPH) mediante una serie de reacciones qumicas que tienen lugar en los grana. Los cloroplastos tambin contienen grnulos pequeos de almidn donde se almacenan los productos de la fotosntesis de forma temporal. En las plantas, los cloroplastos se desarrollan en presencia de luz, a partir de unos orgnulos pequeos e incoloros que se llaman proplastos. A medida que las clulas se dividen en las zonas en que la planta est creciendo, los proplastos que estn en su interior tambin se dividen por fisin. De este modo, las clulas hijas tienen la capacidad de producir cloroplastos.
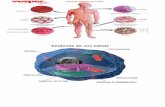
ConclusinClula eucariota; animal las estructuras internas de la clula animal estn separadas por membranas. Destacan las mitocondrias, orgnulos productores de energa, as como las membranas apiladas del retculo endoplasmtico liso (productor de lpidos) y rugoso (productor de protenas). El aparato de Golgi agrupa las protenas para exportarlas a travs de la membrana plasmtica, mientras que los lisosomas contienen enzimas que descomponen algunas de las molculas que penetran en la clula. La membrana nuclear envuelve el material gentico celular y los centriolos que participan en la reproduccin celular.Eucariota, vegetal Las clulas vegetales, as como las animales, presentan un alto grado de organizacin, con numerosas estructuras internas delimitadas por membranas. La membrana nuclear establece una barrera entre la cromatina (material gentico) y el citoplasma. Las mitocondrias, de interior sinuoso, convierten los nutrientes en energa que utiliza la planta. A diferencia de la clula animal, la vegetal contiene cloroplastos, unos orgnulos capaces de sintetizar azcares a partir de dixido de carbono, agua y luz solar. Otro rasgo diferenciador es la pared celular, formada por celulosa rgida, y la vacuola nica y llena de lquido, muy grande en la clula vegetal.CENTRIOLOS Son dos pequeos cilindros localizados en el interior del centrosoma, exclusivos de clulas animales. Con el microscopio electrnico se observa que la parte externa de los centrolos est formada por nueve tripletes de microtbulos. Los centrolos se cruzan formando un ngulo de 90.
Los cloroplastosLos cloroplastos son orgnulos celulares exclusivos de las clulas vegetales. Tienen un tamao variable de unas plantas a otras, pero en las plantas superiores es de alrededor de cinco micras de dimetro. Al igual que las mitocondrias, los cloroplastos tienen la capacidad de multiplicarse por divisin.
El nmero de cloroplastos vara de unas especies a otras, desde las que tienen una sola por clula, que se divide sincrnicamente con el ncleo, hasta las que tienen cincuenta o ms. Los cloroplastos tienen tambin una doble membrana limitante. La membrana interna emite prolongaciones al espacio interior. Estas prolongaciones son tubulares, pero de trecho en trecho se ensanchan y aplanan formando discos. Los discos, a su vez, pueden apilarse para formar una estructura llamada grana, en las que son muy abundantes sustancias tales como las clorofilas y los carotenoides. Dentro de los tilacoides se encuentra la clorofila y dems enzimas que participan en la fotosntesis (proceso que implica la conversin de energa luminosa en energa qumica en forma de ATP). El estroma (el espacio citoslico que rodea a los tilacoides) es dnde se lleva a cabo la fijacin del carbono (en forma de CO2) en compuestos orgnicos. Al igual que las mitocondrias, los cloroplastos poseen su propio DNA. Su estructura es an ms compleja que la mitocondria: adems de las dos membranas de la envoltura, que no se repliegan formando crestas, los cloroplastos tienen numerosos sacos internos en forma de disco (denominados tilacoides), interconectados entre s, que estn formados por una membrana que encierra el pigmento verde llamado clorofila. Desde el punto de vista de la vida terrestre, los cloroplastos desempean una funcin an ms esencial que la de las mitocondrias: en ellos ocurre la fotosntesis; esta funcin consiste en utilizar la energa de la luz solar para activar la sntesis de molculas de carbono pequeas y ricas en energa, y va acompaada de liberacin de oxgeno. Los cloroplastos producen tanto las molculas nutritivas como el oxgeno que utilizan las mitocondrias. La fotosntesis es un proceso fundamental para los seres vivos, ya que permite la transformacin de la energa luminosa procedente del Sol en energa qumica, que es almacenada para ser utilizada segn las necesidades del organismo. A partir de elementos inorgnicos sencillos, el agua y el dixido de carbono, se sintetiza la glucosa, que a su vez puede ser empleada para la produccin de almidn, sacarosa y polisacridos, asumiendo por tanto una funcin estructural; o bien puede servir de sustrato en las reacciones de respiracin celular, con las que se obtiene energa (bajo la forma de molculas de ATP); en ese caso desempea una funcin energtica. En la fotosntesis tambin se produce, mediante un proceso denominado foto fosforilacin, cierta cantidad de ATP. No todos los seres vivos estn en condiciones de realizar este proceso fundamental: esta capacidad slo la poseen algunas bacterias, las cianobacterias, las algas y las plantas, que poseen las molculas de clorofila necesarias para la absorcin de la luz solar y las enzimas indispensables para realizar las reacciones fotosintticas.
MITOCONDRIASEstructuraMembranasLa mitocondria est rodeada por dos membranas diferentes en lo que corresponde a funcin y actividad enzimtica. stas separan tres espacios: el citosol, el espacio intermembrana y la matriz mitocondrial.-Membrana externa: bicapa lipdica exterior la cual es permeable a iones, metabolitos y otras macromolculas. Esto ltimo se debe a que contiene unas protenas llamadas porinas, las cuales forman poros. Esta membrana realiza relativamente pocas funciones enzimticas o de transporte.-Membrana interna: sta membrana contiene ms protenas, carece de poros y es ms selectiva en comparacin con la membrana externa. Contiene muchos complejos enzimticos y sistemas de transporte transmembrana; stos estn implicados en la translocacin de molculas.La composicin de la membrana mitocondrial interna hay una gran cantidad de protenas (cerca de un 80%), y son adems exclusivas en este organelo:1.Cadena trasportadora de electrones, compuesta por cuatro complejos enzimticos fijos y dos transportadores de electrnos mviles.2.Un complejo, el canal de protones ATP-sintasa que cataliza la sintetisis ATP (fosforilacin oxidativa).3. Protenas transportadoras que permiten el paso de iones y molculas (cidos grasos, cido pirvicos, ADP, ATP, O2 y agua). Espacio intermembranoso Es un lquido similar al hialoplasma, este se localiza entre las dos membranas. Posee una alta concentracin de protones debido al bombeo de los mismos por los complejos enzimticos de la cadena respiratoria. En este espacio se encuentran diversas enzimas que intervienen en la trasferencia del enlace de alta energa del ATP.
Matriz mitocondrialContiene menos molculas que el citosol, aunque tiene iones, metabolitos, ADN circular bicatenario, ribosomas tipo 70S y contiene ARNm. Se podra decir que tiene todos los organelos que tendra una clula procariota. En sta parte de la mitocondria hay diversas rutas metablicas esenciales para la vida, cmo el cclo de Krebs y la beta-oxidacin de cidos grasos, adems de la oxidacin de aminocidos y algunas reacciones de sntesis de urea y grupos hem.Funcin.La principal funcin de la mitocondria en la oxidacin de metabolitos (cclo de Krebs, beta-oxidacin y oxidacin de cidos grasos). A partir de esto ltimo la mitocondria puede ser el principal productor de energa mediante la fosforilacin oxidativa, que es dependiente de la cadena transportadora de electrones. Tambin sirve como almacn de sustancias como iones, agua y otros compuestos. Adems sta organela aporta en la transcripcin de informacin gentica a partir del ARN mitocondrial (ARNm)RETICULO ENDOPLASMATICOEl retculo endoplsmico es un sistema de comunicacin interna de las clulas. Se compone de varias membranas, la creacin de canales que se extienden desde el citoplasma a la membrana nuclear (membrana que rodea el ncleo de la clula). Dentro de la red, diversas sustancias son transportadas de un punto a otro, dependiendo de la necesidad. Por ejemplo, las vesculas en el aparato de Golgi, que contienen las enzimas que son transportados desde el retculo endoplsmico a la membrana celular. Hay dos tipos de retculo endoplsmico: retculo endoplsmico liso y retculo endoplsmico rugoso. Retculo endoplsmico rugoso. Miles de ribosomas estn pegados a este tipo de red. Se encuentra en la mayora de las clulas secretoras, como el pncreas. Tambin hace que el transporte de protenas producidas en polirribosomas sean agregadas a varias partes de la clula: el aparato de Golgi, el ncleo, mitocondrias, etc. Retculo endoplsmico liso. El retculo endoplsmico liso (REL) carece de ribosomas, y su principal funcin es desintoxicar el cuerpo. Es el que hace el metabolismo del etanol (alcohol) en las clulas hepticas, y otras sustancias extraas. Tambin es responsable de producir algunos lpidos como el colesterol. Las clulas musculares, que tiene la ATP, una molcula que almacena energa que se utilizar en el movimiento.RIBOSOMAS
La estructura primaria se corresponde con una molcula flexible de gran longitud con una disposicin asimtrica en las bases nitrogenadas y conteniendo una elevada proporcin de guanina y escasa de citosina, en lo que respecta a organismos inferiores y vegetales; a medida que avanzamos en la escala evolutiva aumenta la citosina y va siendo menor la diferencia entre nucletidos. Se pudo demostrar la existencia de una estructura secundaria con la espectroscopa viendo que la molcula de ARN se puede plegar debido a su flexibilidad. Si las bases que estn en oposicin son complementarias se unen y da el aspecto de que es una helice de ADN, hay otras zonas donde no hay oposicin de bases complementarias y no se produce unin quedando como asas, a veces seguidos denominndose estructura de trbol que contiene tambin un ARN transferente.En cuanto a la estructura terciaria se han propuesto varios modelos: segn un modelo se producira por un amontonamiento de cadenas de cidos y dara lugar a estructuras en varilla. Segn otro modelo propuesto por Attardi y Amaldi en el cual aparecen hlices de ARN ribosmico irradiando de un eje comn. La funcin de los ribosomas es la sntesis de protenas. Este es el proceso mediante el cual el mensaje contenido en el ADN nuclear, que ha sido previamente transcrito en un ARN mensajero, es traducido en el citoplasma, juntamente con los ribosomas y los ARN de transferencia que transportan a los aminocidos, para formar las protenas celulares y de secrecin.El ribosoma lee el ARN mensajero y ensambla la protena con los aminocidos suministrados por los ARN de transferencia, este proceso se denomina sntesis de protenas.Los polisomas se encargan de sintetizar protenas de localizacin celular, mientras que los ribosomas del RER se encargan de sintetizar protenas de exportacin, o sea que se irn de la clula hacia otro lugar donde se necesite.Para la sntesis de protenas se requiere la unin de una subunidad mayor y otra menor. Al terminar de fabricar las protenas, se separan. Cuando vuelven a sintetizar se unen dos diferentes.Para la sntesis de protenas los ribosomas se asocian en grupos mediante un filamento de unos 2 nm. de espesor. Estos ribosomas asociados se denominan polisomas, y suelen adoptar una figura en espiral. La subunidad menor queda hacia el interior de la espiral.Los ribosomas forman polisomas para realizar cualquier tipo de sntesis proteicas: tanto la efectuada por los ribosomas libres como la realizada por los asociados a membranas (RER). En el RER la subunidad mayor es la que se adosa a la membrana.El nmero de ribosomas que forman un polisoma y la longitud de ARN-m que los une varan segn el peso molecular de las protenas que se va a sintetizar.Para la sntesis de protenas, los ribosomas recorren el ARNm desde un extremo a otro. Por cada tres nucletidos recorridos, incorporan un aminocido a la cadena de protenas que estn sintetizando; aminocidos que les proporciona el ARNt .Cuando han completado el recorrido, los ribosomas se liberan del ARNm y suelta la protena ya terminada. Mientras se est sintetizando protenas, por cada ribosoma que abandona el polisoma en el extremo final, otro se incorpora en el inicial, de modo que el ritmo de trabajo del polisoma siempre es complejo.El ARNt y la cadena de aminocidos que se est formando se encuentran en un canal situado en la subunidad mayor.PEROXISOMASLos peroxisomas, caracterizados por De Duve en 1960, llevan a cabo gran variedad de funciones, como son: catabolismo de cidos grasos de cadena muy larga, de cido pipeclico, de cidos dicarboxlicos, y del cido fitnico, y biosntesis de plasmalgenos y de cidos biliares. El peroxisoma es una organela celular que est presente en todos los tejidos excepto en el eritrocito maduro. Es particularmente prominente en el hgado, donde puede ocupar del 1,5 al 2% del volumen celular parenquimatoso, y en el rin. En estos tejidos los peroxisomas aparecen en el microscopio electrnico como una organela redonda u oval, con un dimetro promedio de aproximadamente 500 manmetros, rodeados por una membrana simple. Los peroxisomas cerebrales son ms pequeos, con un dimetro promedio de 140 manmetros. Los peroxisomas contienen una matriz ms o menos finamente granulada, en la que se encuentra una masa llamativamente densa denominada nucleoide. Este tiene una estructura muy regular semejante a un cristal con aspecto muy variable. El anlisis bioqumico muestra un contenido de peroxidasa, catalasa, uratooxidasa y aminoacidooxidasa, es decir, enzimas totalmente diferentes de las que se encuentran en los lisosomas. Es necesaria una tincin para las catalasas para identificarlos en las neuronas cerebrales de los roedores, tanto del cerebro como del cerebelo, que contiene peroxisomas durante las primeras dos semanas posnatales; algunos estn presentes en los cuerpos neuronales despus de la tercera semana. Son prominentes en los oligodendrocitos durante el perodo de mielinizacin activa. Esta organela fue la ltima organela subcelular descrita. Fue identificada por primera vez en 1954 en el rin del ratn y debido a que su funcin era desconocida se le asign el nombre de microcuerpo (microbody). El trmino de peroxisoma fue asignado a esta organela por De Duve y Baudhin debido a su papel en la formacin y oxidacin del perxido de hidrgeno. En 1976, Lazarow y De Duve mostraron que la organela tambin desempeaba un papel en la oxidacin de los cidos grasos y este hallazgo fue seguido por el descubrimiento, que en la actualidad an contina expandindose, de que es el sitio de una amplia variedad de reacciones. Muchas de estas reacciones afectan al metabolismo lipdico. El sistema de b-oxidacin peroxisomal de los cidos grasos utiliza reacciones enzimticas completamente distintas de las de la mitocondria. La oxidacin peroxisomal de los cidos grasos no est ligada a la produccin de energa. La oxidacin de los cidos grasos no ramificados con una longitud de cadena de hasta 18 o 20 carbonos, una fuente clave de energa, se lleva a cabo tanto en la mitocondria como en el peroxisoma, teniendo lugar en la mitocondria del 70 al 95%. El peroxisoma se usa exclusivamente para la oxidacin de un conjunto distinto de cidos grasos, como los cidos grasos de cadena muy larga, con una longitud de cadena de 24 o ms carbonos y, como se ha demostrado muy recientemente, cidos grasos de cadena ramificada, como el cido fitnico y pristnico. Los peroxisomas tambin contienen una va paralela para la biosntesis del colesterol. Todava no est clara su relacin con la va bien conocida del retculo endoplsmico. Al menos 50 reacciones enzimticas se han localizado en el peroxisoma, y el nmero contina aumentando.