Organos linfoides
20
Inmunología Órganos linfoides Síntesis realizada por: Q.C. Miguel Ángel Ortiz Gil www.quimicaclinicauv.blogspot.com Julio 2009
-
Upload
miguel-angel -
Category
Documents
-
view
100.118 -
download
2
Transcript of Organos linfoides

Inmunología
Órganos linfoides
Síntesis realizada por:
Q.C. Miguel Ángel Ortiz Gil
www.quimicaclinicauv.blogspot.com
Julio 2009

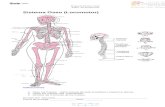
Órganos linfoides
Las células que participan en las respuestas inmunitarias se encuentran
organizadas formando tejidos y órganos, para llevar a cabo sus funciones con la
máxima eficacia. El conjunto de estas estructuras se denomina sistema linfoide. El
sistema linfoide está formado por linfocitos, células accesorias (macrófagos y
células presentadoras de antígeno) y, en algunos tejidos, células epiteliales. Los
principales órganos y tejidos linfoides se clasifican en primarios (centrales) y
secundarios (periféricos) (Fig. No. 1).
Figura No. 1. Principales órganos y tejidos linfoides.
Órganos linfoides primarios
Los órganos linfoides primarios son donde las células del sistema inmune se
diferencian a partir de células madre, proliferan y maduran hacia células con
capacidad efectora. En estos órganos linfoides adquieren sus receptores
antigénicos específicos, y también aprenden a discriminar entre autoantígenos,
que serán tolerados y antígenos extraños que serán atacados (Fig. No. 2).

Los órganos linfoides primarios son los lugares en los que se produce
mayoritariamente la linfopoyesis. En ellos, los linfocitos se diferencian a partir de
las células madre linfoides, proliferan y dan lugar, finalmente, a células maduras
funcionales. En los mamíferos, las células T maduran en el timo, mientras que las
células B maduran en el hígado del feto y en la médula ósea.
Figura No. 2. Órganos linfoides primarios.
Las células T se desarrollan en el timo
El timo de los mamíferos es un órgano bilobulado, situado en la cavidad torácica,
por encima del corazón y los grandes vasos sanguíneos. Cada uno de sus dos
lóbulos principales está compuesto a su vez de lobulillos, separados entre sí por
trabéculas de tejido conjuntivo. En cada uno de estos lobulillos, las células
linfoides (timocitos) se encuentran repartidos entre una zona cortical externa y una
médula interna. En la zona cortical se agrupan densamente la mayoría de los
timocitos relativamente inmaduros en fase de proliferación; las células que se
encuentran en la médula son más maduras, lo que implica que existe un gradiente
de diferenciación desde la corteza hacia la médula. Los timocitos medulares
maduros expresan CD44, mientras que los timocitos corticales no. En el seno de

los lobulillos existe una red de células epiteliales, que es necesaria para que se
produzca el proceso de diferenciación en el que las células pretímicas
procedentes de la médula ósea se transforman en linfocitos T maduros (Fig. No.
3).
Figura No. 3. Desarrollo de linfocitos T.
Los lobulillos del timo contienen tres tipos de células epiteliales
En los lobulillos del timo se pueden distinguir al menos tres tipos diferentes de
células epiteliales, de acuerdo con su distribución, estructura, función y fenotipo.
Son las células epiteliales nodriza de la corteza externa, las células epiteliales
corticales, que forman la red epitelial, y las células epiteliales medulares, que se
suelen disponer en forma de agregados. En los lobulillos del timo también se
encuentran células dendríticas interdigitantes (CDI) y macrófagos (ambos
procedentes de la médula ósea) especialmente en la zona limítrofe entre la
corteza y la médula.
La circulación de las células hacia y desde el timo se produce a través de las
vénulas de endotelio alto (VEA) regionales. Las células epiteliales, las CDI y los

macrófagos expresan moléculas CPH, que son esenciales para el desarrollo y
selección de las células T. En la médula del timo se suelen encontrar corpúsculos
de Hassall. No se conoce su función, pero parece que contienen células epiteliales
degeneradas, ricas en citoqueratinas de elevado peso molecular.
Los linfocitos T más inmaduros llegan a la corteza del timo a través de los vasos
sanguíneos. Los precursores de los Linfocitos T llegan por vía arterial llegan a la
corteza y a través de los capilares pasan a la médula, de la médula salen por los
capilares venosos. Los linfocitos se diferencian en el trayecto de la corteza a la
médula. La diferenciación consiste en la presentación por parte de las células
epiteliales de sus proteínas HLA sucediendo la llamada selección positiva.
Después las células dendríticas y los macrófagos enseñan a los timocitos los
antígenos HLA con péptidos propios en su hendidura (selección negativa).Con
esta selección se eliminan el 95 % de los posibles linfocitos T. La selección
positiva (elimina linfocitos T con receptores poco apropiados) se realiza en la
corteza y en la selección negativa (médula) se eliminan los linfocitos que
reconocen elementos propios del organismo.
Linfopoyesis T comienza con la llegada de los precursores de los linfocitos T, que
durante el proceso de maduración intratímica, reciben el nombre de timocitos.
Durante esta fase mueren muchos timocitos, aproximadamente el 95 por 100 de
ellos, debido a que se eliminan aquellos que reconocen los antígenos propios del
organismo. El resto de las células abandonan el timo, vía sanguínea, como
linfocitos T maduros.
Estos linfocitos colonizan los órganos linfoideos secundarios, situándose en la
zona paracortical de los ganglios linfáticos y vainas paracorticales linfocíticas del
bazo. Se han identificado algunos factores de transcripción que son
imprescindibles para la diferenciación de los linfocitos a lo largo de la linfopoyesis.
Entre estos destacan PU.1 e IKAROS que controlan el desarrollo de células T y B

mientras que GATA-3 solo afecta el compromiso de las células T y E2A, EBF y
Pax controlan el compromiso B.
En el timo se han identificado células precursoras que poseen capacidad de
generar células T, NK, B y células dendríticas del timo, y a lo largo de su
diferenciación los precursores más evolucionados van perdiendo paulatinamente
la capacidad de generar células B, NK y células dendríticas en este orden.
Durante el proceso de maduración intratímico, los timocitos adquieren una serie de
moléculas nuevas en su superficie. Estas moléculas van apareciendo
secuencialmente en los diferentes estadíos de maduración intratímica así como,
en general, en todos los procesos de maduración y diferenciación
hematopoyéticos.
Posteriormente estas células, ya en el timo, maduran distinguiéndose varios
estados diferenciativos con la presencia de diferentes marcadores de superficie.
Así en los timocitos inmaduros aparecen los marcadores CD7 y CD2, añadiéndose
en un estadío posterior de maduración ( timocito común), el marcador CD1. Ya en
el timo va a ocurrir una especialización funcional, distinguiéndose dos
subpoblaciones de timocitos maduros: Una es aquella que expresa en su
superficie el marcador CD4 y que será el precursor inmediato de los linfocitos T
colaboradores y los linfocitos T reguladores que aparecen en sangre periférica.
La otra expresa en la superficie el marcador CD8 y dará origen a los linfocitos T
citotóxicos/supresores circulantes. En ambas subpoblaciones se pierde la
expresión de la molécula CD1. Los timocitos más inmaduros no expresan CD3,
CD4 ni CD8, por lo que son conocidos como células triples negativas. A medida
que van madurando, en estas células se produce la reorganización del TCR, la
expresión del complejo CD3 y de las moléculas CD4 y CD8 conjuntamente
(células dobles positivas), para después perder una u otra quedando bien como
CD4-CD8+ o como CD4+CD8- (Fig. No. 4).

Figura No. 4. Linfopoyesis T.
El timo de los mamíferos experimenta una involución a lo largo del tiempo. En los
seres humanos, la atrofia comienza en el momento de la pubertad, y prosigue a lo
largo de toda la vida del individuo. La involución timica se inicia en la zona cortical,
que puede llegar a desaparecer totalmente, aunque sigan persistiendo restos
medulares. La atrofia cortical está relacionada con la sensibilidad de los timocitos
corticales a los esteroides. Sin embargo, es posible que durante la vida adulta se
sigan produciendo linfocitos T en el timo, aunque en pequeñas cantidades.
Lugares de desarrollo de las células B de los mamíferos
Los linfocitos B se desarrollan directamente en los islotes de células
hematopoyéticas que se encuentran en el hígado del feto y en la médula ósea del
feto y del adulto. La médula ósea de los seres humanos adultos, además de
constituir el lugar en el que se desarrollan las células B, contiene células T
maduras y abundantes células plasmáticas. Por tanto, la médula ósea de los seres
humanos también es un importante órgano linfoide secundario.

Durante el proceso de maduración de los linfocitos B, a partir de la célula
progenitora (CFU-B), se distinguen varios estadíos de diferenciación, que incluyen
las células pre-pre-B, las células pre-B, células B inmaduras y linfocitos B maduros.
En cada uno de estos estadíos de maduración las células expresan distintas
moléculas en la superficie, utilizadas como marcadores para conocer el estado de
diferenciación. Ya en las células pre-B se detecta la presencia de cadena pesada
mu intracitoplasmática, adquiriéndose en la siguiente fase madurativa la capacidad
de sintetizar las cadenas ligeras y pesadas de las inmunoglobulinas IgM e IgD,
detectables en la superficie celular. En consecuencia, la mayoría de los linfocitos B
expresan estos dos tipos de inmunoglobulinas en su superficie. Posteriormente
estos linfocitos, mediante un proceso de reordenamiento génico, se especializarán
en la producción de una sola clase de las inmunoglobulinas IgG, IgA, IgM, IgD e IgE
(Fig. No. 5).
Figura No. 5. Desarrollo de Linfocitos B.
Órganos linfoides secundarios
Los órganos linfoides secundarios son el bazo, los ganglios linfáticos y los tejidos
asociados a mucosas (TLAM), entre los que se encuentran las amígdalas y las
placas de Peyer del íleon. Los órganos linfoides secundarios proporcionan a los
linfocitos un entorno en el que éstos pueden interaccionar entre sí, con las células
accesorias y con los antígenos. Una vez concluido el desarrollo de los linfocitos

en los órganos linfoides primarios, dichas células migran hacia los tejidos
periféricos secundarios (Fig. No. 6).
Figura No. 6. Órganos linfoides secundarios.
El bazo responde ante los antígenos transportados por la sangre, mientras que los
ganglios linfáticos protegen al organismo frente a los antígenos que transporta el
sistema linfático, procedentes de la piel o de superficies internas. En ambos casos,
las respuestas frente a los antígenos consisten en la secreción de anticuerpos
hacia la circulación y en respuestas locales mediadas por células.
El sistema de mucosas ejerce una protección frente a los antígenos que penetran
directamente en el organismo a través de los epitelios mucosos y en él se produce
el primer encuentro (iniciación) entre el antígeno que penetra por las superficies
mucosas y las células inmunes. Así, se suelen encontrar tejidos linfoides
asociados a las superficies que recubren el tracto intestinal (tejido linfoide
asociado al intestino, o TLAI), el tracto respiratorio (tejido linfoide asociado a los
bronquios, o TLAB) o el tracto genitourinario. En estos casos, el principal

mecanismo efector es la secreción directa de IgA (IgAs) sobre la superficie del
epitelio mucoso en cuestión.
El bazo
El bazo se encuentra situado en el cuadrante superior izquierdo del abdomen,
detrás del estómago y próximo al diafragma, mide 13x8cm y pesa 180 y 250 g en
el adulto. Está rodeado exteriormente por una cápsula formada por fibras de
colágeno, que penetran en el parénquima del órgano en forma de trabéculas
cortas. Estas últimas, junto con una red reticular, constituyen el soporte en el que
se asientan las diversas células que contiene el órgano. El bazo contiene dos tipos
principales de tejidos, la pulpa roja y la pulpa blanca (Fig. No.7).
Figura No. 7. Estructura del bazo.
La pulpa blanca está formada por tejido linfoide, la mayor parte del cual está
dispuesto alrededor de una arteriola central, y que se denomina capa linfoide
periarteriolar. La CLP contiene zonas de células T y zonas de células B; las

células T se encuentran situadas alrededor de la arteriola central; las células T se
encuentran situadas alrededor de la arteriola central; las células B se pueden
encontrar organizadas en forma de folículos primarios «no estimulados»
(agregados de células B vírgenes) o folículos secundarios «estimulados» (en los
que aparece un centro germinal con células de memoria).
Los centros germinales contienen también células dendríticas foliculares y
macrófagos fagocíticos. En la región marginal (la zona que recubre al manto de los
folículos secundarios) se encuentran macrófagos especializados y una
subpoblación especial de células B, que responden a antígenos independientes
del timo de tipo II (polisacáridos). Los macrófagos y las células dendríticas
foliculares presentan los antígenos a las células B del bazo. Las células B y otros
linfocitos entran y salen libremente de la CLP a través de los capilares
procedentes de las arteriolas centrales que irrigan la región marginal. Algunos
linfocitos, especialmente los plasmoblastos en fase de maduración, pueden
atravesar la región marginal y alcanzar la pulpa roja a través de ciertos puentes.
La pulpa roja, es un tejido está formado por sinusoides y cordones celulares, que
contienen macrófagos residentes, eritrocitos, plaquetas, granulocitos, linfocitos y
numerosas células plasmáticas. Además de las funciones inmunitarias que
desempeña, el bazo constituye también un reservorio de plaquetas, eritrocitos y
granulocitos. El bazo es el lugar en que son destruidos las plaquetas y los
eritrocitos envejecidos, en un proceso que se lleva a cabo en la pulpa roja y se
denomina «hemocatéresis» (Fig. No. 8).
Figura No. 8. Pulpa roja.

Todo esto es posible gracias a la organización vascular del bazo. Las arterias
centrales se encuentran rodeadas por la CLP y dan lugar a capilares arteriales que
vierten su contenido en los cordones de la pulpa roja. De esta manera, las células
circulantes llegan a estos cordones y quedan atrapadas en ellos. Los macrófagos
reconocen y fagocitan a las plaquetas y eritrocitos envejecidos; las células
sanguíneas que no son ingeridas y destruidas pueden retornar a la circulación
sanguínea atravesando los orificios que determina los endotelios discontinuos en
las paredes de las sinusoides venosos, mientras que el plasma fluye libremente a
través de dichas paredes (Fig. No. 9).
Figura No.9. Estructura de la pulpa blanca del bazo. La pulpa blanca se separa por la
vaina periarteriolar linfoide (PALS) rodeada de células B. Dentro de cada folículo primario
existe un grupo de células foliculares dendríticas (FDC). El (BC) se cree que representan
las zonas por las que linfocitos entran y salen de la pulpa blanca.
Ganglios linfáticos y sistema linfático
Los ganglios linfáticos forman parte de una red que filtra los antígenos contenidos
en el líquido intersticial de los tejidos y en la linfa durante el transporte de estos
líquidos desde la periferia hasta el conducto torácico y los restantes conductos
colectores principales. Los ganglios linfáticos se suelen encontrar situados en los
puntos de ramificación de los vasos linfáticos.

Existen determinadas zonas que drenan regiones superficiales y profundas del
organismo, como el cuello, las axilas, las ingles, el mediastino y la cavidad
abdominal, en las que se observa una acumulación estratégica de ganglios
linfáticos. Los ganglios linfáticos que protegen a la piel son superficiales, y se
denominan ganglios subcutáneos. Los ganglios linfáticos profundos, que protegen
a las mucosas respiratoria, digestiva y genitourinaria, se denominan ganglios
viscerales o profundos.
Los ganglios linfáticos humanos presentan un diámetro de 2-10 mm, son
redondeados o con forma de riñón y presentan una depresión, denominada hilio,
en la zona de entrada y salida de los vasos sanguíneos. La linfa llega hasta el
ganglio linfático a través de varios vasos linfáticos aferentes, y sale del mismo a
través de un único vaso linfático eferente, situado en el hilio. Los ganglios linfáticos
típicos están rodeados por una cápsula de colágeno. Los diversos componentes
celulares se encuentran dispuestos sobre un armazón de trabéculas radiales y
fibras de reticulina. Un ganglio linfático contiene una región de células B (córtex),
una región de células T (paracórtex) y una médula central, en la que aparecen
cordones celulares que contienen células T, células B, células plasmáticas y
macrófagos (Fig. No. 10).
Figura No. 10. Estructura de un ganglio linfático.

La región paracortical contiene gran cantidad de CPA (células interdigitantes), que
expresan activamente antígenos de superficie CPH de clase II. Estas CPA
proceden de la piel (células de Langerhans) o de las mucosas (células
dendríticas), transportando hasta el ganglio los antígenos procedentes de la
superficie interna o externa del organismo. La mayor parte del tejido linfoide se
encuentra en las regiones corticales y paracortical. La médula también contiene
algo de tejido linfoide, que forma cordones separados por senos linfáticos
(medulares) que desembocan en el seno terminal, a partir del cual surge el vaso
linfático eferente. Los senos linfáticos se encuentran recubiertos de células
fagociticas, especialmente en la región medular.
Cuando la linfa fluye desde los vasos aferentes hasta los eferentes, estas células
fagociticas capturan las partículas antigénicas contenidas en las misma y las
transportan hasta el tejido linfoide del ganglio linfático. El córtex contiene
agregados de células B, formando folículos primarios o secundarios, mientras que
las células T se encuentran situadas principalmente en el paracórtex. Por tanto,
cuando una zona de la piel o de una mucosa se ve expuesta a un antígeno
dependiente de células T, las células T del paracórtex de los ganglios linfáticos
que drenan dicha zona proliferan activamente.
Los folículos secundarios de los ganglios linfáticos estimulados por antígenos
poseen centros germinales. Son parecidos a los centros germinales de las
regiones de células B de las CLP esplénicas y del TLAM. Las células del centro
germinal pueden ser grandes o pequeñas, y se denominan centroblastos y
centrocitos, respectivamente. Las células B en fase de proliferación de los centros
germinales presentan una forma nuclear bien definida.
Los centros germinales están rodeados por un manto de linfocitos. Las células B
de esta capa contienen grandes cantidades de IgM e IgD de superficie. En la
mayor parte de los folículos secundarios, este manto o corona se encuentra
engrosada en dirección a la cápsula del ganglio. Los folículos secundarios

contienen, CPA dendríticas foliculares, algunos macrófagos y escasos linfocitos T
CD4 que interaccionan con las células dendríticas del centro germinal. Parece que
todas estas células, junto con los macrófagos especializados de los senos
marginales, desempeñan un papel en las respuestas de las células B y,
especialmente, en el desarrollo de las células B de memoria, que probablemente
es la función principal de los centros germinales.
Tejido linfoide asociado a mucosas (TLAM)
Los agregados de tejido linfoide no encapsulado son especialmente abundantes
en la lámina propia y en la submucosa de los tractos gastrointestinal (Fig. No. 11),
respiratorio y genitourinario. Las células linfoides se pueden encontrar como
agregados difusos o estructuradas en forma de ganglios aislados o agrupados,
que contienen centros germinales (folículos secundarios).
Las amígdalas de los seres humanos contienen una cantidad considerable de
tejido linfoide, que en muchas ocasiones presentan folículos secundarios con
zonas T interpuestas con vénulas de endotelios altos. Hay tres tipos
fundamentales de amígdalas: las palatinas, las faríngeas (adenoides) y las
linguales, que constituyen el anillo de Waldeyer. En los bronquios y en el tracto
genitourinario también se encuentran acumulaciones parecidas de tejido linfoide.
Los epitelios respiratorios, genitourinarios y digestivos contienen células
dendríticas, que tienen la misión de ingerir, transportar y procesar los antígenos
hasta los ganglios linfáticos de drenaje.
En la lámina propia de la pared intestinal se encuentran acumulaciones de tejido
linfoide, que suelen alcanzar la submucosa. Estas acumulaciones pueden adoptar
forma de nódulos solitarios o nodulos agregados, como en el apéndice. Las placas
de Peyer se localizan en el íleon distal. El epitelio intestinal (epitelio asociado a los
folículos, EAF) que recubre las placas de Peyer es un epitelio especializado, que
permite el transporte de los antígenos hacia el tejido linfoide. Esta misión es

llevada a cabo por ciertas células epiteliales, denominadas células «M» debido a
la presencia de numerosas microvellosidades en su superficie luminal y que se
localizan entre los enterocitos. Estas células contienen profundas invaginaciones
de la membrana plasmática basolateral, que forman invaginaciones en las que se
alojan linfocitos B y T, células dendríticas y macrófagos. Los antígenos y los
microorganismos entran por estas invaginaciones hacía el tejido linfoide mucoso
organizado subyacente al epitelio.
Figura No. 11. Mucosa del tracto gastrointestinal.
Las respuestas inmunitarias humorales a nivel de la mucosa son principalmente
del isotipo IgA. Las IgA de secreción son unos anticuerpos capaces de atravesar
las membranas mucosas, impidiendo de esa forma la penetración de los
microorganismos infecciosos.
Linfocitos de mucosas
Estos linfocitos están situados en el tejido conjuntivo que forma la lámina propia y
en el seno del tejido epitelial. Los linfocitos de la lámina propia (LLP) son
principalmente células T activadas, pero también se observa la presencia de
numerosas células B activadas y células plasmáticas. Estas células plasmáticas
secretan principalmente IgA, que atraviesa las células epiteliales y es liberada a la
luz del conducto en cuestión.

Los linfocitos intraepiteliales (LIE) son principalmente células T, que presentan
características fenotípicas diferentes de las de los LLP. La mayoría de las células
T LLP y LIE pertenecen a la subpoblación CD45RO de células de memoria. Se
sabe que los LIE liberan citocinas, entre las que se encuentran el IFNy e IL-5. Se
ha sugerido que los LIE pueden estar encargados de la detección de células
propias mutantes o infectadas por virus.
CIRCULACIÓN DE LOS LINFOCITOS
Una vez que alcanzan los tejidos secundarios, los linfocitos no se limitan a
permanecer allí; muchos de ellos se desplazan de un órgano linfoide a otro a
través de la sangre y de la linfa.
Aunque algunos linfocitos abandonan la circulación sanguínea a través de vénulas
no especializadas, en la mayoría de los mamíferos este proceso ocurre a través
de una zona especializada de las vénulas poscapilares, denominada vénula de
endotelio alto o VEA. Estas vénulas se encuentran principalmente en el paracórtex
de los ganglios linfáticos, existiendo también algunas en el córtex, pero no en la
médula. Algunos linfocitos, fundamentalmente células T, penetran en los ganglios
linfáticos a través de los vasos linfáticos aferentes, no a través de las VEA; ésta es
la vía principal de llegada del antígeno a los ganglios linfáticos. Además de en los
ganglios linfáticos, este tipo de vena se localiza en el TLAM y en el timo.
Las VEA controlan la circulación de los linfocítos
Las VEA están recubiertas de células endoteliales cúbicas. Estas células se
encuentran activadas y expresan diversas moléculas de adherencia, que no
aparecen en las células endoteliales planas y en reposo propias de las vénulas
normales. Uno de los mecanismos de activación de las células endoteliales es la
producción a nivel local de citocinas, como el IFNy, la IL-1 y el TNF.

Las células endoteliales se pueden activar y dar lugar a VEA en los lugares en los
que existe un proceso inflamatorio crónico, como por ejemplo en la piel o en la
membrana sinovial, en las que normalmente no existe este tipo de vénulas. A su
vez, esto puede provocar la acumulación de subpoblaciones específicas de
linfocitos T en la zona en que se han formado las VEA. Las moléculas que
expresan las células endoteliales activadas pertenecen a la superfamilia de las
inmunoglobulinas, como en el caso de ICAM-1 (CD54), ICAM-2 (CD102) o VCAM-
1 (CD106), o a la familia de las selectinas, como por ejemplo la selectina E
(ELAM-1: CD62E) y la selectina P (CD62P).
Se cree que entre los linfocitos y las células endoteliales se establecen varias
interacciones receptor-ligando diferentes, que sirven para dirigir los linfocitos hacia
los órganos diana. Estas interacciones se producen mediante unas «adresinas»
expresadas por las células endoteliales, como MadCAM-1 presente en los
endotelios en el tejido intestinal o VCAM-1 de otras células endoteliales.
Las células linfoides de los ganglios linfáticos retornan a la circulación a través de
los vasos linfáticos eferentes, cuyo contenido se vierte finalmente en la vena
subclavia izquierda a través del conducto torácico. Este proceso permite que un
gran número de linfocitos específicos de un antígeno determinado entre en
contacto con el mismo en el microentorno de los órganos linfoides periféricos. Los
linfocitos circulan continuamente de uno a otro ganglio, pero cuando un antígeno
penetra en los ganglios linfáticos de un animal sensibilizado previamente frente a
dicho antígeno, la circulación queda interrumpida transitoriamente,
aproximadamente durante 24 horas.
Una de las diferencias entre el sistema TLAM y los órganos linfoides sistémicos es
que las células linfoides asociadas a mucosas circulan principalmente a través del
sistema linfoide de mucosas. Así, las células linfoides estimuladas de las placas
de Peyer atraviesan los ganglios linfáticos regionales y son vertidas al torrente
circulatorio, pero retornan rápidamente a la lámina propia del intestino. Esta

recirculación específica es posible debido a que las células linfoides reconocen
moléculas de adherencia de tipo «adresinas» expresadas específicamente por las
células endoteliales de las vénulas poscapilares de la mucosa, y que no se
encuentran presentes en las VEA de los ganglios linfáticos. Por ello, la
estimulación antigénica en una zona de la mucosa induce una respuesta de
anticuerpos que afecta, predominantemente al TLAM.

Bibliografía:
ü Yang-Xin Fu and David D. Chaplin. Development and maturation ofsecondary lymphoid tissues. Annu. Rev. Immunol. 1999. 17:399 433.
ü Thomas Boehm and Conrad C Bleul. The evolutionary history of lymphoidorgans. Nature Immunology, Vol. 8 No. 2, February 2007.
ü Reina E. Mebius and Georg Kraal. Strucrure and function of the spleen.Nature Reviews Immunology, Vol, 5, August 2005.
ü Reinhard Pabst. Plasticity and heterogeneity of lymphoid organs what arethe criteria to call a lymphoid organ primary, secondary or tertiary?Immunology Letters 112 (2007) 1 8.
ü Ulrich H. von Andrian and Thorsten R.Mempel. Homing and cellular traffic inlymph nodes. Nature Reviews Immunology. Vol,3, November 2003.
ü Troy D Randall, Damian M Carragher, and Javier Rangel-Moreno.Development of secondary lymphoid organs. Annu Rev Immunol. 2008; 26:627 650.
ü Gerd Muller and Martin Lipp. Concerted action of the chemokine andlymphotoxin system in secondary lymphoid-organ development. CurrentOpinion in Immunology 2003, 15:217 224.
ü David D. Chaplin and Yang-Xin Fu. Cytokine regulation of secondarylymphoid organ development. Current Opinion in Immunology 1998, 10:289-297