
PARTE VIIciat-library.ciat.cgiar.org/Articulos_Ciat/tsbf/pdf/...clasificarse en función del grado...
32
PARTE VII Conclusiones Generales y Prioridades para la Investigación Futura
Transcript of PARTE VIIciat-library.ciat.cgiar.org/Articulos_Ciat/tsbf/pdf/...clasificarse en función del grado...

PARTE VII
Conclusiones Generales yPrioridades para la
Investigación Futura

Conclusiones Generales...
407
CAPÍTULO 24
Conclusiones Generales, ResultadosImportantes de la Investigación yNecesidades Futuras
J. J. Jiménez*, T. Decaëns**, R. J. Thomas*,L. Mariani*** y P. Lavelle***
Esta obra recopila los conocimientosrecientes sobre losmacroinvertebrados del suelo,haciendo énfasis especialmente enlas lombrices de tierra de lassabanas situadas sobre los suelosácidos de los Llanos Orientales deColombia. La mayor parte de estainvestigación se realizó en el CentroNacional de Investigaciones (CNI)Carimagua. La hipótesis básica queha servido de hilo conductor en estelibro es que las lombrices de tierrason un recurso natural que puedemanejarse para ayudar a lasostenibilidad de aquellosagroecosistemas cuyos recursosbiológicos se consideraríanindicadores potenciales de fertilidady de calidad del suelo.
En Carimagua se hizo uninventario de los recursos biológicos,es decir, de la abundancia y ladiversidad de las especies y de suecología particular, así como unaevaluación del impacto que causandichos recursos en el suelo y en las
plantas. En este libro se describe elimpacto de los ecosistemas, tantonaturales como antrópicos, sobre lascomunidades de la macrofauna delsuelo, y se estudian la biologíabásica, la dinámica de laspoblaciones, las estrategiasadaptativas y el ciclo de vida de laslombrices de tierra; asimismo, sedetallan los efectos que un grupofuncional, el de los ingenieros delecosistema, causa en diferentesaspectos físicos, químicos ybiológicos de estos suelos. Sepresentan, también, opciones paraconservar dichos recursos biológicosy para aprovecharlos en sistemas demanejo que integren diversasprácticas agrícolas. Finalmente, seresumen los resultados másrelevantes de la investigaciónobtenidos hasta la fecha y sediscuten las necesidades queenfrenta esta investigación en elfuturo.
La Fauna del Suelo comoRecurso Natural Valioso
La macrofauna del suelo en lasabana natural de Carimagua secaracteriza por su riquezataxonómica y su alta densidad depoblación. Las termitas (Isoptera)son el grupo más abundante (47% dela biomasa total) y lo siguen las
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
* Laboratoire d’Ecologie, UFR Sciences,Université de Rouen, F-76821 Mt SaintAignan Cedex, Francia.
*** Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.

408
El Arado Natural:…
lombrices (Oligochaeta) con un 31%del total. Entre los demás gruposrepresentados están las hormigas ylas avispas (Hymenoptera), losescarabajos (Coleoptera), las arañas(Arachnida), los miriápodos(Myriapoda), los nematodos(Nematoda), las cigarras(Homoptera), las chinches(Hemiptera), los isópodos (Isopoda) ylas moscas y mosquitos (Diptera).
Los dos grupos funcionales másimportantes de losmacroinvertebrados del suelo estánrepresentados en la sabana deCarimagua: son los“transformadores de la hojarasca”,p.ej., algunos coleópteros y laslombrices de hábito epígeo, y los“ingenieros del ecosistema”, como lastermitas, las hormigas, las lombricesendógeas y las anécicas. De modosimilar, y unida a la abundanteriqueza específica de la sabana, unagran diversidad funcional semanifiesta en cuatro tipos deestructuras biogénicas superficialesproducidas por 14 especies deingenieros del ecosistema; son lassiguientes: termiteros (3 tipos),placas de termitas (1), hormigueros(8) y heces de lombrices (2) (Decaënset al. 2001). Este descubrimientorefuerza la idea de que lacaracterización de la macrofauna encualquier ecosistema debe iracompañada de un inventario de lasestructuras biogénicas, las cualesson, además, el factor que une labiodiversidad de los organismos conla función que éstos desempeñan enel ecosistema (Lavelle 1997).
Cuando los ecosistemasnaturales son sustituidos poragroecosistemas, la intensidad de loscambios que experimenta lacomunidad de lombrices dependerá,en su mayor parte, de la amplitud delas modificaciones ambientalesinducidas por el uso agrícola delsuelo. Por ejemplo, la riqueza
específica disminuye drásticamentecuando la selva tropical o la sabanase convierten en monocultivosanuales (Dangerfield 1990; Decaënset al. 1994; Fragoso et al. 1997;Lavelle 1988; Lavelle y Pashanasi1989). De otra parte, pocos cambiosse observan cuando elagroecosistema es funcionalmentesimilar al ecosistema original, comoen el caso de un pastizal provenientede una sabana (Decaëns et al. 1994;Jiménez et al. 1998; Schneidmadl yDecaëns 1995) o el de un sistemaagroforestal que deriva del bosquetropical (Fragoso et al. 1997; Gilot-Villenave et al. 1995; Lavelle yPashanasi 1989).
La pérdida de la diversidad de lascomunidades de lombrices causauna alteración en el funcionamientodel ecosistema (Fragoso et al. 1997;Giller et al. 1997). En India, porejemplo, surgen problemas en loscultivos de té tras la desaparición delas lombrices de hábito anécicoporque la hojarasca, que sin ellas nopuede ser introducida rápidamenteen el suelo, se acumulaexcesivamente sobre éste (Senapatiet al. 1994). En América del Sur, laconversión del bosque amazónico enpastizales intensivos conduce a laeliminación de las especies nativasde lombrices en beneficio de otraespecie alóctona; ahora bien, laintensa actividad de ésta, unida a laausencia de la diversidad original,degradan el ecosistema (Barros et al.1998; Blanchart et al. 1999; Chauvelet al. 1999; Duboisset 1995).
La IntensificaciónAgrícola y los Ingenieros
del EcosistemaLas prácticas agrícolas puedenclasificarse, siguiendo un gradientecontinuo, desde las extensivas(utilización del fuego, baja carga

Conclusiones Generales...
409
animal) hasta las intensivas (usonormal de maquinaria agrícola,aplicación de grandes cantidades deinsumos y pastoreo intenso). Laintensificación agrícola y lasprácticas asociadas con ella, como laeliminación de la vegetación nativa,la mecanización del suelo y el uso depesticidas, conducen a ciertasmodificaciones ambientales(especialmente en el microclima y enel recurso trófico) que disminuyen lamayor parte de la biodiversidad delsuelo (Swift et al. 1996).
La biodiversidad de unagroecosistema se reduce no sólo enrelación con la macrofauna del suelosino también con las comunidadesvegetales y animales, encomparación con los ecosistemasnaturales (Giller et al. 1997; Swift yAnderson 1994; Swift et al. 1996).El concepto de intensificaciónagrícola incluye también cambios enla estructura de los agroecosistemas,por ejemplo cuando se pasa de unaagricultura tradicional a una másintensiva (Swift et al. 1996). Laintensificación agrícola se caracterizapor tres acciones (Giller et al. 1997;Swift y Anderson 1994):
• El uso más frecuente de lamisma parcela de suelo, es decir,la intensificación en el uso delrecurso.
• La especialización de las especiesproductivas, tanto animalescomo vegetales, es decir, elcomienzo de la pérdida de labiodiversidad de la cubiertavegetal inicial.
• La utilización de insumos, comofertilizantes y pesticidas, juntocon la mecanización del suelo.
Los diferentes agroecosistemasestudiados en Carimagua puedenclasificarse en función del grado deintensificación agrícola (Swift et al.1996), el cual puede expresarse
mediante un índice (adaptado deGiller et al. 1997) que permitevisualizar de manera precisa dichaintensificación en los LlanosOrientales (Decaëns y Jiménez2002). El orden ascendente deintensificación del gradiente sería elsiguiente (Figura 24-1):
Sabana nativa < Cultivos demarañón < Pastizalestradicionales <
Pastizales introducidos antiguos< Pastizales introducidosrecientes <
Monocultivos < Cultivos anuales
La riqueza taxonómica de lamacrofauna del suelo varía a lo largode este gradiente según dos factores:(a) la intensidad y la frecuencia de laperturbación y (b) la productividaddel ecosistema (cantidad y calidad delos recursos) (Huston 1996). Engeneral, se pueden distinguir dosgrandes agroecosistemas quecontrastan según los efectoscausados en las prácticas agrícolasempleadas: los pastizales y losmonocultivos anuales.
Los pastizales no sólo conservanla riqueza de especies de lombricessino que aumentan sus poblaciones(Jiménez et al. 1998), es decir, tienenuna capacidad de carga mayor. Larazón es la siguiente: los pastizales,que incorporan C al suelo mediantela fotosíntesis y las raíces,suministran además con losdesechos animales, en particular lasdeposiciones del ganado, recursosorgánicos de mayor calidad tantopara las lombrices como para lostransformadores de la hojarasca. Elestiércol del ganado es fragmentadoe incorporado al suelo rápidamente,junto con la hojarasca, por losingenieros del ecosistema, es decir,uno y otra son ingeridos por ellos yse mezclan con la parte mineral delsuelo en el tracto digestivo de laslombrices anécicas, o son mezclados
410
El Arado Natural:…
con suelo —en el que varían lacantidad de materia orgánica (MO) yde raíces— por las lombricesendógeas. En los pastizales hay unacantidad de raíces mucho mayor queen la sabana nativa (Rao 1998), unfactor que puede aumentar laspoblaciones de lombrices endógeas,como Andiodrilus n. sp., y tambiéninfluir en la acumulación decantidades considerables de C en elinterior del suelo (Fisher et al. 1994,1997).
En algunos casos, lacombinación de agroecosistemasfavorece los procesos antes descritos;por ejemplo, el sistema arroz-pastizal(Sanz et al. 1999), donde el pastizal,que queda establecido tras lasiembra del arroz, se beneficia de lafertilización inorgánica aplicada alcultivo de este cereal. Igualmente,en los pastizales de gramíneas con
leguminosas, la fertilización orgánicaaumenta la cantidad y la calidad dela MO. La presencia de laleguminosa en el sistema mejora elciclo de los nutrientes y favorece lafijación biológica del N (Thomas1992; Thomas et al. 1995).
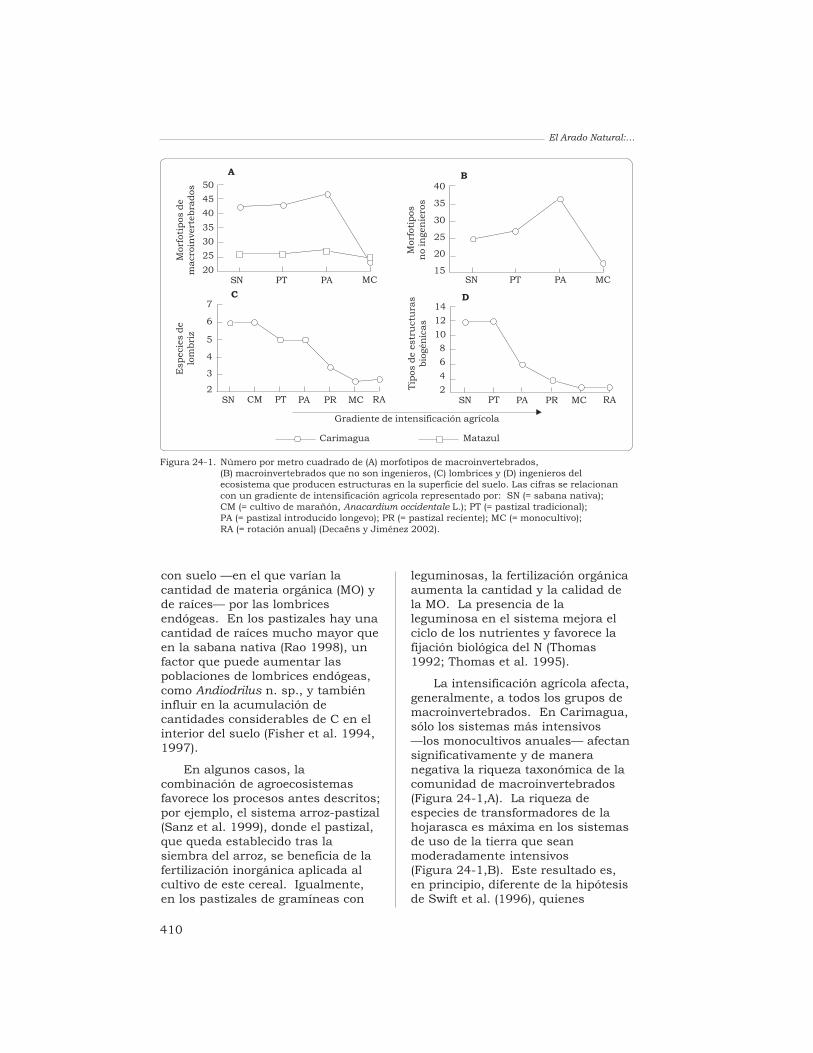
La intensificación agrícola afecta,generalmente, a todos los grupos demacroinvertebrados. En Carimagua,sólo los sistemas más intensivos—los monocultivos anuales— afectansignificativamente y de maneranegativa la riqueza taxonómica de lacomunidad de macroinvertebrados(Figura 24-1,A). La riqueza deespecies de transformadores de lahojarasca es máxima en los sistemasde uso de la tierra que seanmoderadamente intensivos(Figura 24-1,B). Este resultado es,en principio, diferente de la hipótesisde Swift et al. (1996), quienes
Figura 24-1. Número por metro cuadrado de (A) morfotipos de macroinvertebrados,(B) macroinvertebrados que no son ingenieros, (C) lombrices y (D) ingenieros delecosistema que producen estructuras en la superficie del suelo. Las cifras se relacionancon un gradiente de intensificación agrícola representado por: SN (= sabana nativa);CM (= cultivo de marañón, Anacardium occidentale L.); PT (= pastizal tradicional);PA (= pastizal introducido longevo); PR (= pastizal reciente); MC (= monocultivo);RA (= rotación anual) (Decaëns y Jiménez 2002).
A50454035302520
Carimagua Matazul
SN PT PA MC
7
6
5
4
3
2
C
SN PT PACM PR MC RA
40
35
30
25
20
15
1412108642
SN PT PA MC
SN PT PA PR MC RA
B
D
Mor
foti
pos
no
inge
nie
ros
Tipo
s de
est
ruct
ura
sbi
ogén
icas
Mor
foti
pos
dem
acro
inve
rteb
rado
sE
spec
ies
delo
mbr
iz
Gradiente de intensificación agrícola
Conclusiones Generales...
411
sostienen que la intensificaciónagrícola conduce sistemáticamente auna reducción de la diversidad. Sinembargo, esta hipótesis no fuerespaldada por el estudio realizadoen Matazul, cerca de Puerto López ya 100 km al este de Carimagua,donde no hubo una disminuciónsignificativa de la riqueza taxonómicaa lo largo del gradiente deintensificación.
En cuanto a las lombrices detierra, el número de especiesdisminuye progresivamente si separte de la sabana natural y seavanza a los monocultivos anuales(Figura 24-1,C). Por último, elnúmero de estructuras biogénicasidentificables sobre la superficie delsuelo, a menudo relacionadas con elnúmero de especies de ingenieros delecosistema que desarrollan parte desus actividades en la superficie delsuelo, también disminuyerápidamente cuando aumenta la
intensificación del sistema de uso dela tierra, excepto en las pasturasextensivas tradicionales (o sea, en elsistema menos intensivo)(Figura 24-1,D).
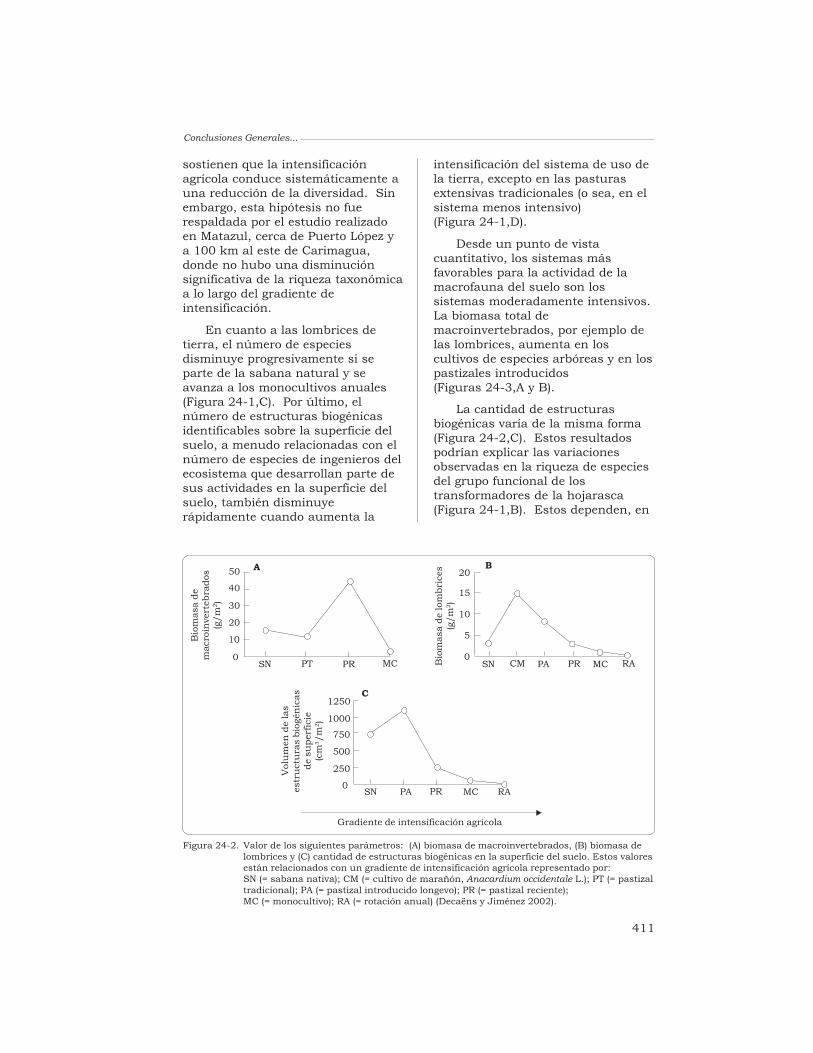
Desde un punto de vistacuantitativo, los sistemas másfavorables para la actividad de lamacrofauna del suelo son lossistemas moderadamente intensivos.La biomasa total demacroinvertebrados, por ejemplo delas lombrices, aumenta en loscultivos de especies arbóreas y en lospastizales introducidos(Figuras 24-3,A y B).
La cantidad de estructurasbiogénicas varía de la misma forma(Figura 24-2,C). Estos resultadospodrían explicar las variacionesobservadas en la riqueza de especiesdel grupo funcional de lostransformadores de la hojarasca(Figura 24-1,B). Estos dependen, en
Figura 24-2. Valor de los siguientes parámetros: (A) biomasa de macroinvertebrados, (B) biomasa delombrices y (C) cantidad de estructuras biogénicas en la superficie del suelo. Estos valoresestán relacionados con un gradiente de intensificación agrícola representado por:SN (= sabana nativa); CM (= cultivo de marañón, Anacardium occidentale L.); PT (= pastizaltradicional); PA (= pastizal introducido longevo); PR (= pastizal reciente);MC (= monocultivo); RA (= rotación anual) (Decaëns y Jiménez 2002).
Bio
mas
a de
mac
roin
vert
ebra
dos
(g/m
2 )
Bio
mas
a de
lom
bric
es(g
/m2 )
50
40
30
20
10
0
Gradiente de intensificación agrícola
20
15
10
5
0
1250
1000
750
500
250
0
SN CM PA PR MC RASN PT PR MC
A B
SN PA MC RA
C
Vol
um
en d
e la
ses
tru
ctu
ras
biog
énic
asde
su
perf
icie
(cm
3 /m
2 )
PR
412
El Arado Natural:…
gran medida, de tales estructuras.Por ejemplo, M. carimaguensis, uningeniero del ecosistema, modificalas condiciones de vida de otrosorganismos mediante la creación deestructuras biogénicas, es decir, deheces superficiales donde abundanciertos grupos de artrópodos(T. Decaëns 1999, sin publicar).
Las especies de lombrices deCarimagua no responden por igual ala intensificación agrícola. Engeneral, su densidad aumenta en lossistemas de uso de la tierramoderadamente intensivos ydisminuye en los sistemas másintensivos, aunque la amplitud deestas variaciones es grande ydepende de la especie (Decaëns yJiménez 2002). En los pastizalesintensivos, un género nuevoperteneciente a la familiaOcnerodrilidae aprovecha
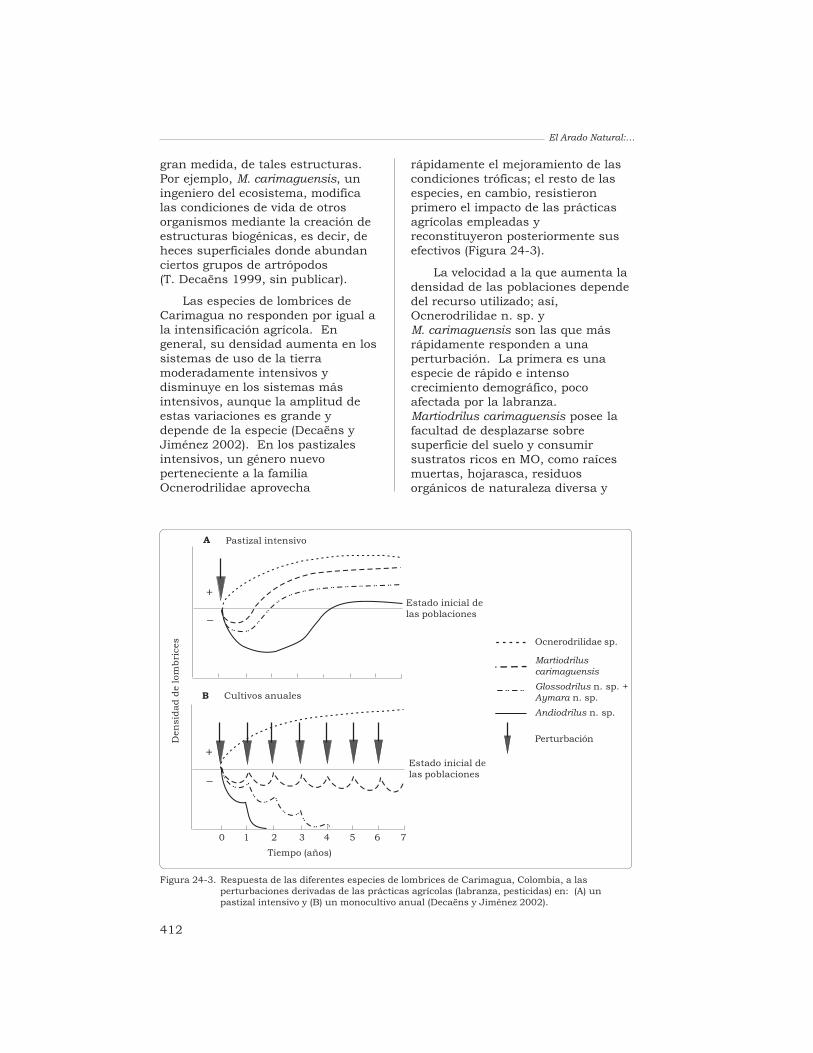
rápidamente el mejoramiento de lascondiciones tróficas; el resto de lasespecies, en cambio, resistieronprimero el impacto de las prácticasagrícolas empleadas yreconstituyeron posteriormente susefectivos (Figura 24-3).
La velocidad a la que aumenta ladensidad de las poblaciones dependedel recurso utilizado; así,Ocnerodrilidae n. sp. yM. carimaguensis son las que másrápidamente responden a unaperturbación. La primera es unaespecie de rápido e intensocrecimiento demográfico, pocoafectada por la labranza.Martiodrilus carimaguensis posee lafacultad de desplazarse sobresuperficie del suelo y consumirsustratos ricos en MO, como raícesmuertas, hojarasca, residuosorgánicos de naturaleza diversa y
Figura 24-3. Respuesta de las diferentes especies de lombrices de Carimagua, Colombia, a lasperturbaciones derivadas de las prácticas agrícolas (labranza, pesticidas) en: (A) unpastizal intensivo y (B) un monocultivo anual (Decaëns y Jiménez 2002).
A
B
Pastizal intensivo
Cultivos anuales
Estado inicial delas poblaciones
Estado inicial delas poblaciones
+
_
+
_
0 1 2 3 4 5 6 7
Den
sida
d de
lom
bric
es
Tiempo (años)
Ocnerodrilidae sp.
MartiodriluscarimaguensisGlossodrilus n. sp. +Aymara n. sp.Andiodrilus n. sp.
Perturbación

Conclusiones Generales...
413
heces de otras lombrices (Mariani etal. 2001). En los monocultivosanuales, las especies no tienen eltiempo suficiente para recuperar ladensidad que tenían antes de lasperturbaciones y la mayoría de ellasdesaparece de estos sistemas.
Ciclo de Vida, Estructurade la Comunidad y
PerturbaciónEl estudio de la diversidad y de losprocesos ecológicos asociados con esabiodiversidad y su función en elecosistema constituye la base de lacomprensión y del manejo de losecosistemas naturales y perturbados(Giller 1996). En general, lascomunidades de lombrices sonsensibles a los factores climáticos yedáficos, los cuales determinan ladisponibilidad de los recursosalimenticios y las condicionesmicroclimáticas. Existe, por tanto, elriesgo de que las alteraciones que elhombre causa en los ecosistemasnaturales alteren las comunidades demacroinvertebrados. Ahora bien, lacapacidad de respuesta de dichafauna puede emplearse para manejarsus actividades (Lavelle et al. 1994,1999).
En relación con la biología y laecología de las especies de la sabana,las hipótesis que se han probado sonlas siguientes (Jiménez 1999):
• La sustitución de la sabana nativapor el pastizal conduce a unareducción drástica de la riquezaespecífica y de la diversidad de laslombrices de tierra, que puedeconducir a la eliminación de lasespecies nativas.
• Aunque las especies nativasconsigan adaptarse al nuevosistema de uso de la tierra, laestructura de su comunidadcambiará.
• El manejo de la producciónprimaria en el pastizal y suaumento influyen en laabundancia y en la biomasa delas especies, bien incrementandoo bien reduciendo el número deindividuos.
El cambio que experimenta elecosistema natural, que es la sabananativa, influye en la estructurafuncional de la comunidad delombrices, pero poco o nada afecta lariqueza en especies de éstas. En lospastizales de Carimagua seencuentran las mismas especiesnativas que en la sabana natural yno se observa ninguna invasión deespecies exóticas. Sin embargo, laestructura de la comunidad hacambiado, es decir, la contribuciónrelativa de las diferentes categoríasecológicas y sus efectos en losagroecosistemas fueron alterados.
Dos pastizales introducidosconservan la fauna nativa enCarimagua: (a) Andropogon gayanusy Brachiaria decumbens, solos oasociados con la leguminosa Puerariaphaseoloides; (b) Brachiariahumidicola, sola o asociada conArachis pintoi u otras leguminosas.Estos resultados son excepcionalesporque, según la teoría generalmenteaceptada, las perturbaciones queexperimentan los ecosistemasnaturales traen consigo ladisminución o la desaparición de lasespecies nativas (Lee 1991). Lo máscomún es la aparición de especies dedistribución pantropical con unrango de tolerancia a las propiedadesfísico-químicas del medio muyamplio, como Pontoscolex corethrurusMüller (Cuadro 24-1) y Polypheretimaelongata Perrier (Barois et al. 1988;Barros et al. 1998; Fragoso et al.1999; Lavelle y Pashanasi 1989).
Es probable que el principalfactor que interviene en elmantenimiento de la fauna nativa

414
El Arado Natural:…
sea la similitud estructural yfuncional entre el ecosistema originaly el agroecosistema (Fragoso et al.1999). Una de las causas delestablecimiento de poblacionesexóticas de lombrices es lacolonización del nicho que dejanvacante las especies nativas tras unaalteración de su hábitat (Kalisz yWood 1995).
Del total de especies de la sabana,sólo dos crecieron significativamenteen abundancia en los pastizales deCarimagua (Jiménez 1999).
Estrategias AdaptativasLas sabanas tropicales sonecosistemas sometidos a una fuerteestacionalidad en la cual la épocaseca puede variar de 3 a 9 meses.Este patrón determina el tipo desabana y, por tanto, su régimen dehumedad y su vegetación (Solbrig etal. 1996). Una estacionalidad tanfuerte influye directamente en lahumedad del suelo, factor quedetermina los siguientes aspectos:
- la actividad, la dinámica y eltamaño de las poblaciones delas lombrices (Lavelle 1978,1983);
- las estrategias adaptativas ylos hábitos alimenticiosadoptados; y
- la localización en el suelodurante la época deactividad, ya que laspoblaciones de lombricesresponden a la severidadambiental con el fin de evitarsu extinción.
Las tres categorías ecológicasprincipales de lombrices (Bouché1977; Lavelle 1981) se basan en larespuesta que dan estos organismosa los factores limitantes del suelo, esdecir, a la escasa reserva denutrientes, al suelo compacto y a lascondiciones climáticas desfavorables.
Las especies se adaptan a talescondiciones de estrés mediantecambios en su actividad y en suestructura demográfica (en losparámetros de desarrollo comocrecimiento, fecundidad, mortalidad,etc.). Hay tres parámetros depoblación, todos ellos explicados porlas variaciones estacionales delambiente, que definen la estructurafuncional de la comunidad delombrices de Carimagua: laabundancia, el grado de actividad yla distribución vertical. Laestacionalidad aparece, por ejemplo,como el factor determinante de laactividad de las poblaciones y de lasdiversas estrategias adaptativasfrente a las condiciones adversas(Jiménez et al. 2000). Partiendo deestas observaciones, en Carimagua
Cuadro 24-1. Riqueza de especies de macroinvertebrados en dos ambientes: uno, un ecosistema debosque tropical en Manaos, Brasil, bajo el bosque primario y en un pastizal introducidode Brachiaria humidicola Rendle (Barros et al. 1998); y otro, un ecosistema de sabana enCarimagua, Colombia, en la sabana y en un pastizal introducido de B. humidicola +Pueraria phaseoloides (Decaëns et al. 1994).
Parámetro Bosque tropical Sabana(Manaos, Brasil) (Carimagua, Colombia)
En bosque En pastizal En sabana En pastizal
Morfoespecies (no.) 151 48 31-57 42-55
Especies en común (%) 15 54
Biomasa total (g PF/m2) 51.5 51.2 15.3 28.8-62.5
Biomasa de Pontoscolex corethrurus (%) 0 90 0 0

Conclusiones Generales...
415
se distinguen tres grupos deespecies, según la respuesta que denal estrés ambiental:
1. Especies de tamaño pequeño,localizadas superficialmente en elsuelo (en la época activa), condistribución vertical profundadurante la fase de quiescencia.Por ejemplo, Ocnerodrilidae n.sp., así como Aymara n. sp.(epígea) y Glossodrilus n. sp.(endógea), aunque la distribuciónvertical de las dos últimas no esmuy profunda.
2. Especies de tamaño intermedio,de localización superficialdurante la época lluviosa, y sinadaptación especial a la época desequía; por ejemplo, Andiodrilusn. sp. (endógea).
3. Especies de tamaño grande, condiapausa verdadera en las capasprofundas del suelo; por ejemplo,Andiorrhinus n. sp. (endo-anécica, aunque sin diapausa) yM. carimaguensis (anécica).
Ingenieros del Ecosistemadel Suelo
Los Capítulos del 11 al 23 estándedicados al estudio de los efectosproducidos por los ingenieros delecosistema en algunos procesosfísicos y químicos del suelo, y giranen torno a tres hipótesis (Decaëns1999):
• Los ingenieros del ecosistemaproducen estructuras biogénicasque son características de suespecie.
• Los ingenieros del ecosistemacausan, mediante las estructurasbiogénicas producidas, unimpacto significativo en losprocesos del suelo y en losorganismos que viven en él.
• La intensificación agrícola tieneefectos contrastantes en lascomunidades de ingenierosecológicos del suelo y altera, portanto, el impacto que éstosharían, ya que la composición dela comunidad de ingenieros y sufunción pueden variar.
La comprobación empírica yexperimental de estas hipótesis hapermitido establecer y describir elpapel funcional de los ingenieros delecosistema en los suelos de lassabanas colombianas, así comoevaluar el impacto que tienen lasprácticas agrícolas en laspoblaciones de tales ingenieros.
Martiodrilus carimaguensiscomo ingeniero del ecosistemaLos ingenieros ecológicos sonorganismos que tienen la facultad demodular la disponibilidad de losrecursos que serán usados por otrosorganismos vivos mediante lasestructuras biogénicas que producen(Jones et al. 1994, 1997; Lawton yJones 1995). Numerososmacroinvertebrados responden aesta definición (Anderson 1995;Jones et al. 1994; Lavelle 1997),incluyendo a M. carimaguensis. Estaespecie, en efecto, modifica suambiente haciendo pasar losmateriales (hojarasca, humus ysuelo) de un estado físico a otro, esdecir, abriendo macroporos en elsuelo, incorporando hojarasca alsuelo y formando agregados estables.Las estructuras producidas—galerías y turrículos— constituyenun recurso (microhábitat y recursotrófico, respectivamente) que losmacroinvertebrados y las raícesaprovechan directamente. Demanera relativamente indirecta,M. carimaguensis transforma elsuelo, llevándolo de un estado físicoinicial a uno bioturbado a través deuna serie de modificaciones en sus

416
El Arado Natural:…
propiedades físicas, en la dinámicade la MO y en la disponibilidad de losnutrientes, todo lo cual tiene unefecto positivo en el crecimiento delas plantas (Decaëns et al. 1999a,1999b).
En resumen, M. carimaguensisdesarrolla las siguientes acciones pormedio de las estructuras queproduce (Decaëns 1999):
• Suministra un recurso trófico, yaque Andiodrilus n. sp. yOcnerodrilidae n. sp. ingierenesas estructuras, y uno espacial,porque proporciona un espacio aciertos macroinvertebrados en elinterior de las galerías o bajo losturrículos.
• Modifica indirectamente ladisponibilidad y la calidad delrecurso trófico y del recursoespacial; es decir, al modificar laestructura del suelo y ladinámica de su MO, favorece ladisponibilidad de los nutrientesque requieren las plantas.
• Facilita el transporte de algunosorganismos, aproximándolos alos recursos.
Los atributos funcionales de unaespecie o de un grupo funcional seconsideran a diferentes niveles deorganización dentro de los sistemasjerárquicamente conformados. Encuanto aumenta la escala deobservación, de lo más próximo a lomás alejado, los niveles describen losmismos procesos (Allen y Starr1982). Una primera aproximaciónascendente (“bottom-up”, sensuAnderson 1995) ha mostrado que lasestructuras biogénicas producidaspor M. carimaguensis influyen en laestructura del suelo, en la dinámicade la MO del suelo, en la diversidadde la macrofauna y en el crecimientoradicular de las plantas. En una
aproximación descendente(“top-down”, sensu Anderson 1995),el impacto de M. carimaguensis en elecosistema sigue siendo apreciable,aunque está marcadoprogresivamente por los efectosproducidos por otras comunidadesde organismos y por los factoresambientales que regulan los procesosdel suelo dentro de la escalajerárquica (Decaëns et al. 1999a).
Tipología de las estructurasbiogénicasLos ingenieros del suelo delecosistema en Carimagua producenestructuras biogénicas decaracterísticas físicoquímicasparticulares de cada especie y,generalmente, diferentes del sueloque las circunda. Esto abre laposibilidad, mas allá de los criteriosclásicos de clasificación ecológica delas lombrices de tierra, de estableceruna clasificación nueva de losingenieros del ecosistema basada enlos atributos funcionales de susestructuras biogénicas y sinconsiderar necesariamente losaspectos taxonómicos (Decaëns et al.2001); por ejemplo, la ingeniería quehace M. carimaguensis en elecosistema disminuye lacompactación del suelo —un efectoque equilibra, en cierta forma, lacompactación proveniente de lasheces grandes y compactas queproduce la lombriz— y nos permitiríallamarla una especiedescompactadora. Hay queconsiderar también el efecto de laespecie perteneciente a la familiaOcnerodrilidae, pues su contribucióna la estabilidad estructural del suelopuede ser importante, aun cuandolos efectos que tenga en elcrecimiento de las plantas seanpequeños.

Conclusiones Generales...
417
Guía para el Manejo de laFauna Benéfica del Suelo
Tanto la investigación presentada enesta obra como las referenciasbibliográficas consultadas nos llevana ser cautos respecto a laformulación de una guía generalpara el manejo de la macrofauna delsuelo. Algunas prácticas, como lospastizales mejorados, puedengenerar, sin duda, un aumentosustancial de las poblaciones demacrofauna. Ahora bien, esteaumento puede ser benéfico, comoen el caso de M. carimaguensis en lassabanas colombianas, o perjudicial,como el efecto compactador deP. corethrurus en los pastizales de laAmazonia que, aunado a la pérdidade la fauna nativa del suelo, llevaríaa la degradación de los pastizales(Chauvel et al. 1999). La similitudentre el ecosistema original y elagroecosistema que de él provienesería el principal determinante de lasupervivencia, de la adaptación, dela capacidad de resistencia y de laestabilidad de las especies nativas delombrices, dentro de los límites delmanejo de los ecosistemas.
Antes de pensar en estableceruna de las guías indicadas, esnecesario conocer la biología y laecología básicas de la macrofauna decualquier ecosistema. Este procesoexige un activo trabajo deinvestigación, lo que explica laescasa presencia de estudiossimilares en la zona tropical. Elconocimiento mencionado debeenfocarse a los principales ingenierosdel ecosistema que se hanencontrado, puesto que los efectoscausados por estas especies tiendena dominar los que provienen de lasdemás poblaciones de la macrofauna(Lavelle 1997).
El caso de M. carimaguensis enlos Llanos Orientales de Colombia es,hasta ahora, una excepción, ya que
los efectos de esta especie en lospastizales mejorados tienden a serbeneficiosos cuando aumentan suspoblaciones (Parte III y Parte VI deesta obra). Estas lombrices son unejemplo excepcional de que lasactividades de la macrofauna nativacontribuyen a la conservación, a laproductividad y a la sostenibilidad deun agroecosistema, y logran mejorarestos parámetros.
La disposición espacial de unpastizal junto a un cultivo puedeacelerar la recuperación de laspoblaciones de macrofauna de esteúltimo. Esta práctica de manejonecesita ser investigada aún enprofundidad, especialmente si laespecie benéfica, que puedeestablecerse más rápidamente,ayuda a revertir los problemas dedegradación de la estructura delsuelo debidos a los cultivos, y haceasí innecesaria la solución de dichosproblemas mediante el empleo deestrategias intensivas y el uso demaquinaria costosa. Las lombricesde tierra se convierten entonces enun recurso que se podría utilizarpara mejorar la calidad delecosistema.
La disposición espacial deparcelas que conservan laspoblaciones de lombrices junto aaquéllas que tienden a reducirlas esun ejemplo de manejo indirecto(Figura 24-4). Lavelle et al. (1999)suministran mayor informaciónsobre este tema y lo discuten endetalle.
Tipología de una comunidadde macrofauna benéficaAunque es grande el número deespecies que pueden considerarseingenieros del ecosistema porquecausan cambios físicos en elambiente, no todos esos cambiostendrán consecuencias ecológicassignificativas (Jones et al. 1997). Sin

418
El Arado Natural:…
embargo, algunos ingenieros puedenmultiplicar estos efectos mediante laacción desarrollada por su dominiofuncional. Estos efectos dependende una serie de factores, entre losque se encuentran la densidadpoblacional, la distribución espaciallocal y regional, y el tipo deestructuras físicas producidas (Joneset al. 1997). Por tanto, es precisoidentificar las especies presentes enel suelo y las estructuras que ellasproducen, y describir con precisiónlos mecanismos mediante los cualeslos ingenieros del ecosistemainfluyen en el ambiente del suelo.Las interacciones tróficas eindirectas (interacciones deingeniería, sensu Jones et al. 1994)pueden incluirse en modelospredictivos del impacto quecausarían los ingenieros delecosistema. Estos modelos seránparticularmente útiles cuando seintente mejorar la sostenibilidad de
los agroecosistemas mediante unaumento de las actividades o de ladiversidad de la macrofauna delsuelo (o de ambos factores) (Giller etal. 1997). Los modelos seemplearían para probar las hipótesissobre la función de la biodiversidaden los agroecosistemas (Swift yAnderson 1994; Swift et al. 1996).
El estudio aquí presentadopermite sugerir que el manejo de ladiversidad de las lombrices de tierraen los agroecosistemas debe iniciarseestableciendo una comunidad quecontenga una o más especies epígeasy una o más especies anécicas —queconstruyan galerías verticales,produzcan estructuras biogénicas enla superficie y mezclen residuosvegetales con el sustrato mineral— yuna o más especies endógeas —quese alimenten de MO y de exudadosde raíces y horaden galeríashorizontales. La comunidad de
Figura 24-4. Alternativas de manejo en las tecnologías con lombrices (Senapati et al. 1999). Losrectángulos indican actividades humanas.
Vermicompost
Inoculación sola1. Potes2. Parcelas
Inoculación +manejo de
residuos orgánicos
Manejoorgánico
Rotación decultivos
Yurimaguas(Perú)
Tamil Nadu(India)
Carimagua(Colombia)
Yierpadi(India)
Producción masiva
Yurimaguas (Perú)Yierpadi (India)
Manejo directo(inoculación)
Manejo indirecto
En el suelo
Tecnologías con lombrices
Fuera del suelo

Conclusiones Generales...
419
lombrices de Carimagua respondeexactamente a este modelo, y haceposible la propuesta de un manejodeterminado de las comunidades delombrices en los agroecosistemasestudiados.
Las opciones de manejo de laactividad de las lombrices tendríanen cuenta, principalmente, lossiguientes aspectos relevantes:
• La comunidad de ingenieros delecosistema del suelo en elagroecosistema debería ser lamisma que la del ecosistemaoriginal, que es el natural.
• Los organismos ingenieros delsuelo y, en particular,M. carimaguensis, influyen endiferentes procesos relacionadoscon la fertilidad del suelo.
• Las actividades de los ingenierosdel suelo se han desarrolladobastante en los pastizales, perose reducen mucho en los cultivosanuales.
• Los efectos de los ingenieros delecosistema permanecen durantealgún tiempo porque lasestructuras biogénicasproducidas persisten en elentorno, aun después de ladesaparición del organismo quelas produjo.
La especie perteneciente a lafamilia Ocnerodrilidae yM. carimaguensis son las únicasespecies de lombriz que soportan laintensificación continua de losmonocultivos. Ocnerodrilidae n. sp.es una especie endógea polihúmicaque tolera la labranza porque susindividuos son de tamaño muypequeño. Los individuos deM. carimaguensis, aparte de la buenaestrategia con que, en la épocadesfavorable, ocupan los estratosprofundos del suelo (Jiménez yDecaëns 2000), migran también por
la superficie del suelo. Lacolonización que haríaM. carimaguensis de una parcelapobre en lombrices podría facilitarsesi se distribuyen adecuadamente lasparcelas. Además, las rotaciones enque un monocultivo sigue a unpastizal de varios años de serviciopueden ser favorables para eldesarrollo de poblacionesimportantes de lombrices.
Un sistema de producciónagrícola que preserve las lombricesdesarrollaría las siguientes prácticas:
• Mantener los pastizales nativos(la sabana) como reservorios debiodiversidad.
• Establecer cultivos forrajeroscomo bancos de proteína(Rippstein et al. 1996) quepermitan el desarrollo de unabiomasa de lombricesimportante.
• Sembrar cultivos anuales queroten con pastizales de variosaños (bancos de proteína) o concultivos forrajeros, de maneraque los fenómenos de migraciónse maximicen. En este sistema,las estructuras biogénicas queaparezcan serían indicadorescualitativos y cuantitativos delestado de las poblaciones de losingenieros del ecosistema.
Un desafío importante para elfuturo cercano es la identificación delos agroecosistemas sostenibles quemantienen niveles aceptables dediversidad y de biomasa de lamacrofauna del suelo, para optimizarlas actividades de ésta. Lainvestigación presentada en estaobra es un ejemplo de la decisión deabrir la “caja negra” de la biología delsuelo y del consiguiente desarrollo deuna guía de manejo de la faunaencontrada. Ahora bien, para quelos productores acepten y adoptenesta guía, los estudios mencionados

420
El Arado Natural:…
deben combinarse con un enfoquemultidisciplinario que incluya losconocimientos locales, laexperimentación en la finca delproductor y algunos estudiossocioeconómicos. De este modo, elsuelo se vería como un lugar másdiverso, productivo y saludable,gracias a un proceso que aumente elnúmero de opciones viables para elproductor.
Aceptación por losProductores
Cualquier práctica nueva de manejode los organismos del suelo debe seraceptada por el agricultor y requiere,a menudo, que éste la desarrolle y laadapte para que pueda incorporarlacon éxito a su sistema. Elconocimiento actual de la percepciónque tienen los agricultores de losorganismos del suelo es todavía muylimitado. Algunos estudios recientessobre esta visión del agricultor hanrevelado, por ejemplo, una granvariación tanto en los conocimientoslocales sobre las lombrices como enel uso que pueden hacer de ellas ensus sistemas agrícolas (Ortiz et al.1999). Los agricultores de Méxicoque fueron consultados estabanrelativamente bien informados sobrela importancia y los beneficios de laslombrices de tierra en la fertilidad delsuelo; los agricultores del Congo, encambio, tenían muy escasainformación al respecto. Sepercibieron también diferencias entrehombres y mujeres sobre este temaen el estudio de México (Ortiz et al.1999). Los conocimientos que tienenlos agricultores de una localidadsobre los organismos microscópicosson incluso menores que losrelacionados con los organismos másgrandes y visibles; los síntomas deenfermedades en las plantas serían,quizás, el único indicador obvio paraellos de la presencia de organismos
perjudiciales. No obstante, resultaestimulante anotar que losagricultores de México podíandistinguir entre tres o cuatro tipos delombrices según su tamaño, su colory el lugar donde se encontraban.Ortiz et al. (1999) publicaron elprimer informe, probablemente,sobre la percepción que tienen losagricultores de la macrofauna delsuelo.
Los agricultores y los científicosnecesitan desarrollar conjuntamenteun lenguaje común sobre la faunadel suelo, ya que así se aceptaríanmejor los conceptos sobre losorganismos del suelo y sobre lasprácticas de manejo de éstos, y seaumentaría, por ello,significativamente las poblaciones deestos organismos en los sistemas deproducción. Es necesario, también,desarrollar una guía del usuario,sencilla y amigable, para manejar lasactividades de los organismosbenéficos. Los agricultores necesitandistinguir, por ejemplo, entre lasespecies de lombrices que compactanel suelo y las que no lo compactan.Necesitan también métodos demuestreo simples y rápidos paraobtener, al menos, un cálculosemicuantitativo de la abundanciade la macrofauna. Los científicos,por su parte, necesitan establecer overificar los umbrales de abundanciade organismos o de poblaciones,desde los que se obtengan impactossignificativos en los procesos delsuelo y en la producción vegetal.
Estas herramientas y guías paratomar decisiones deben desarrollarseconjuntamente entre los agricultoresy los científicos, incorporando elconocimiento local existente y,cuando éste no exista,suministrando material educativo ydidáctico para los que puedan leer ypara los que no sepan hacerlo.Existen ejemplos de dicho material:uno de ellos es la serie de guías de

Conclusiones Generales...
421
manejo de los recursos naturales queha sido publicada para usarla en elmanejo de las cuencas (CIAT 1999).Estas guías indicarán cómoidentificar y desarrollar un enfoquesobre el manejo de los recursosnaturales y cómo desarrollar unmaterial atractivo para losagricultores y otros usuarios, comolos extensionistas y los integrantes deorganizaciones no gubernamentales(ONG). Actualmente se estánelaborando módulos de capacitaciónsobre biología del suelo.
Resultados Importantesde la Investigación
Capítulo 2Impacto del uso de la tierraen la macrofauna del suelode los Llanos Orientales deColombia1. Las comunidades de
macroinvertebradosresponden claramente a lasalteraciones derivadas delsistema de uso del suelo. Elpastoreo y la quemafavorecen la actividad de laslombrices, aunque suimportancia decrece cuandohay sobrepastoreo.
2. La asociación deB. decumbens y P.phaseoloides durante untiempo largo (15 años)conserva la calidad y labiodiversidad del suelo ymantiene también la riquezataxonómica de la sabana.
3. Los cultivos anuales tienenun impacto negativodramático en las poblacionesde lombrices y artrópodosporque reducen su biomasa,la densidad de suspoblaciones y su riquezataxonómica.
Capítulo 3Cuantificación de lamacrofauna del suelo enuna cuenca hidrográfica deColombia1. La diversidad, la abundancia
y la estructura funcional delas comunidades de losmacroinvertebrados del suelovariaban según el sistema deuso de la tierra; el bosquesecundario fue el ecosistemade mayor diversidad.
2. Varios organismos puedenser utilizados comoindicadores de calidad delsuelo en los ambientes deladera alterados de los Andescolombianos.
Capítulo 4Las comunidades delombrices en las sabanasnativas y en los pastizalesintroducidos de los LlanosOrientales de Colombia1. La riqueza específica de la
sabana nativa se mantuvo enlos pastizales, ya sean degramínea sola o asociada conleguminosas, y ningunaespecie exótica de lombrizcolonizó estosagroecosistemas.
2. La comunidad de lombricesde Carimagua estácompuesta por especiespertenecientes a lasprincipales categoríasecológicas. Estas especiesson de diverso tamaño yemplean estrategiasadaptativas diferentes que lespermiten sobrevivir a severascondiciones ambientales.
3. La conversión de la sabanaen pastizal produjodiferencias significativas entérminos de densidad y de

422
El Arado Natural:…
biomasa de las lombrices(10 veces mayores en elpastizal introducido), que sedebieron, principalmente, alespectacular aumento de laspoblaciones de la especieanécica M. carimaguensis.
4. En la sabana nativa, laslombrices endógeascontribuyeron con más del80% a la biomasa total delombrices, mientras que en elpastizal, la especie M.carimaguensis aportó el 88%de esa biomasa y lasendógeas sólo el 12%.
Capítulo 5Dinámica de poblaciones yestrategias adaptativas deMartiodrilus carimaguensis(Oligochaeta,Glossoscolecidae), unaespecie nativa de lassabanas bien drenadas deColombia1. La producción de heces
frescas o turrículos en lasuperficie del suelo fue 11veces superior en el pastizalintroducido que en la sabananativa, lo que indica que haydiferencias entre ambossistemas respecto a laabundancia de M.carimaguensis.
2. Martiodrilus carimaguensispresentó una verdaderadiapausa, que teníadiferencias si se trataba deun adulto o de un individuoinmaduro. Estos últimos sonactivos solamente 4 meses alaño, hasta la mitad delperíodo de lluvias. Estaslombrices construyencámaras de estivación al finalde sus galerías, y en ellashacen varios septos con lasheces.
3. La capacidad de entrar endiapausa antes de la llegadade la época seca y laconstrucción de galeríassemipermanentes son dosfactores que explican por quéla sequía afecta tan poco alos individuos de esta especiey por qué pueden mantenersu densidad poblacional.
4. Las diferencias en la calidadde la MO y el aporte que haceel estiércol del ganado, quees rápidamente incorporadoal suelo por especies dehábito anécico (M.carimaguensis y algunoscoleópteros) pueden ser losfactores que explican elaumento de la actividad deestas lombrices en lospastizales.
Capítulo 6Estrategias reproductorasde tres especies delombrices nativas de lassabanas de Carimagua(Colombia)1. La fuerte estacionalidad del
sitio restringe la deposiciónde los capullos a sólo8 meses del año. Seobservan dos estrategiasclaras: o bien deposicióncontinua, como enAndiodrilus n. sp., odeposición y diapausa, comoen M. carimaguensis; o bienmuerte de los adultos, comoen Glossodrilus n. sp.
2. Cuanto mayor es el tamañodel adulto, mayor es el delcapullo; M. carimaguensisinvierte el 16% del peso deladulto en la formación delcapullo, el porcentaje másalto registrado hasta la fecha.

Conclusiones Generales...
423
Capítulo 7Distribución vertical de laslombrices de tierra en lossuelos de los “Llanos” deColombia cubiertos porespecies forrajeras1. La sustitución de la sabana
por el pastizal no produceningún efecto en ladistribución vertical de laslombrices en el suelo.
2. Las especies más grandes nose encuentran a muchaprofundidad, ya que no huborelación entre el tamaño delas lombrices y laprofundidad a la que fueronencontradas. Sin embargo,una especie endógeapolihúmica muy pequeña,que se encuentrahabitualmente en las hecesde M. carimaguensis en elinterior del suelo, suelehallarse a una profundidadrelativa.
3. El principal factor en ladistribución vertical de laslombrices es la humedad delsuelo, aunque las diferenciasencontradas en este factorentre los adultos y losindividuos inmaduros de M.carimaguensis pueden sertambién de origen biótico. Elinicio temprano de ladiapausa de los individuosinmaduros estáfisiológicamente inducido; laseñal que desencadena elproceso necesita másinvestigación.
4. Durante la diapausa, tantolos adultos como lossubadultos de M.carimaguensis (los individuosmaduros) se encontraron amayor profundidad que losindividuos inmaduros.
Capítulo 8Distribución espacial de laslombrices en las sabanas desuelos ácidos de los LlanosOrientales de Colombia1. Las lombrices de la sabana y
de los pastizales, quepertenecían a categoríasecológicas diferentes y eran detamaño diverso, presentaronuna distribución espacialfuertemente agregada enparches de varias decenas demetros, en cualquiera de losdos sistemas de uso de latierra ensayados.
2. Existe una relación entre losíndices de agregación y eltamaño de las especies (larelación longitud/diámetropreclitelar, L/D). El análisisfactorial discrimina lasespecies grandes que viven acierta profundidad de las quese encuentran cerca de lasuperficie del suelo, y tambiénlas especies de agregaciónespacial fuerte pero conrelación L/D baja de lasespecies con valores altos deL/D, como en el caso deGlossodrilus sp., la cual tieneademás un índice deagregación cercano a 1. Elpatrón de distribuciónespacial de las lombrices seexplica por variablesbiológicas y no por el sistemade uso de la tierra.
Capítulo 9Diferencias en laprogramación de ladiapausa y en los patronesde estivación de algunasespecies de lombriz de tierradel trópico1. La actividad de las lombrices
estuvo determinada por la

424
El Arado Natural:…
fuerte estacionalidad delecosistema de las sabanas ycasi todas las poblaciones delombrices descendían a losestratos profundos del suelodurante la época seca.
2. Las especies que afrontan laépoca seca muestrandiferentes estrategiasadaptativas, por ejemplo ladiapausa facultativa uobligatoria, para evitar quesus tegumentos se sequen enel interior de las cámaras deestivación localizadas a ciertaprofundidad del suelo.
3. Los individuos inmaduros deM. carimaguensis comienzanla diapausa en la mitad de laépoca lluviosa; la fuerteestacionalidad de las sabanassería tal vez el factor queregula la densidadpoblacional de esta especie.
Capítulo 10La dieta de una lombrizanécica de las sabanascolombianas: Una cuestiónrelacionada con lascategorías ecológicas1. Se observaron diferencias
significativas mínimas alcomparar el contenido de lamolleja de los individuosprovenientes ya sea de lasabana o del pastizalintroducido. Los de lasabana tenían en la mollejauna cantidad relativamentegrande de complejos órgano-minerales. Se encontrótambién una gran cantidadde fragmentos vegetales< 500 µm (44.2% V/V) en losagregados órgano-minerales,de los cuales las raíces eranel componente principal.
2. El contenido de las mollejasanalizadas era el siguiente:
un 33% de fracción liviana deMO, un 6.6% de MO amorfay un 53% de partículasminerales.
3. Los individuos recolectadosen las dos parcelasensayadas compartían unrégimen alimenticio basadoen la necrofagia y en lacoprofagia, y a vecesadoptaban una dietatípicamente anécica, es decir,ingerían una mezcla dehojarasca y suelo.
4. Los estudios que seemprendan más tardedeberán considerar lahipótesis de la ingestiónselectiva de una masaórgano-mineral en lasuperficie del suelo, es decir,un comportamiento anécicotípico aunque con unafunción ecológica modificada.Este comportamiento sería laconsecuencia de unaadaptación de esta especie ala ausencia de hojarasca enla sabana que es sometida ala quema periódica.
Capítulo 11Propiedades de lasestructuras construidas porlos ingenieros delecosistema en la superficiedel suelo de una sabanacolombiana1. En las sabanas ácidas de
Carimagua, 14 especiesdiferentes demacroinvertebradosproducen estructurasbiogénicas sobre la superficiedel suelo, que son fácilmentedistinguibles entre sí:hormigueros (8 tipos),termiteros (3), placas determitas (1) y heces delombrices (2).

Conclusiones Generales...
425
2. Los turrículos producidos porAndiodrilus sp. (endógea) yM. carimaguensis (anécica)están formados por agregadosde gran tamaño que, en elcaso de esta última especie,son más estables que losagregados del suelo de tamañocomparable.
3. En conjunto, se hanidentificado tres tipos deestructuras biogénicas:a. compactas y ricas en MO
(turrículos);b. ligeramente compactas y
ricas en MO (termiteros); yc. granulares, no
compactadas y con bajoscontenidos de MO(hormigueros y placas determitas)
4. A partir de la tipología de lasestructuras biogénicas, sepuede establecer unaclasificación funcional de lafauna del suelo y valorar susefectos en el funcionamientodel ecosistema.
Capítulo 12Dinámica de la degradaciónde las heces superficiales dela lombriz de tierra en áreasde sabana y de pastizal enlos Llanos Orientales deColombia1. La formación de cámaras y
galerías en el interior de losturrículos se debió a lacolonización de éstos poralgunos macroorganismos(macroinvertebrados y raíces)y principalmente por laactividad de las termitaspertenecientes al géneroNasutitermes.
2. La persistencia de losturrículos deM. carimaguensis sobre lasuperficie del suelo parece
estar determinada por dosfactores antagónicos:a. la densidad de la
vegetación y la cantidadde hojarasca presenteque protege los turrículosdel impacto de la lluvia, y
b. el pisoteo del ganado queacelera su destrucción.
3. El papel que desempeñan losmacroinvertebradospequeños en lafragmentación de lasestructuras biogénicasproducidas por organismosde tamaño mayor apoya lahipótesis de que laestabilidad de losecosistemas depende de unadiversidad funcional alta.
4. Los individuos deM. carimaguensis soningenieros del ecosistema, yaque los turrículos queproducen son micrositiosdonde se regulan importantesprocesos físicos y químicosdel suelo. La producción degrandes cantidades deturrículos produce efectossignificativos en la estructuradel suelo, en sus propiedadesfísicas y en la dinámica de laMO en el perfil del suelo.
Capítulo 13Dinámica del nitrógeno enlas heces de las lombricesde tierra de la especieMartiodrilus carimaguensisJiménez y Moreno, en unOxisol de las sabanas deColombia1. La introducción de un
pastizal en un área desabana conduce al aumentode las reservas de MO, delcual dependería elincremento de la densidad delas lombrices de tierra.

426
El Arado Natural:…
2. Las lombrices de lospastizales, cuya densidad esalta, ingieren variastoneladas de suelo por año, yson por ello responsables delas grandes cantidades de Nmineralizado.
3. El manejo de lascomunidades de lombrices enlos pastizales introducidospuede convertirse en unmecanismo que aumente enel suelo la MO y el ciclo denutrientes, especialmente delN, reduciendo así el riesgo dedegradación del pastizal.
Capítulo 14Disponibilidad de fósforoen las heces de una especieanécica de lombriz de tierrade las sabanas de sueloOxisol en Colombia1. Martiodrilus carimaguensis
juega un papel importante enmejorar y en mantener ladisponibilidad del P en losOxisoles de baja fertilidad.
2. Todas las fracciones de Pencontradas en las heceseran mayores que las delsuelo que circundaba lasheces (aumento quecorrespondía al del P total),principalmente en lasfracciones inorgánicas lábilesde P.
3. Los individuos deMartiodrilus carimaguensis,en condiciones de campo,incorporan el P de lahojarasca o del materialvegetativo aéreo y radiculardescompuesto. Este P seincorpora en todas lasfracciones (orgánicas einorgánicas) de P, peroespecialmente en susfracciones inorgánicaslábiles.
Capítulo 15Materia orgánica del suelocontenida en las heces queproduce una comunidad delombrices anécico-endógeade un pastizal tropical, enColombia1. En las heces de lombriz se
encontraron contenidos deagregados estables más altosque en el suelo circundante,que correspondían, a su vez,a concentraciones altas de Corgánico.
2. La mezcla estrecha dematerial fresco con suelo(efecto anécico) llevó a mayorconcentración de recursosprimarios (carbohidratos ylignina) y a menorconcentración de compuestoshúmicos (C aromáticos ygrupos –COOH sustituidos enel C).
3. La actividad de las lombricestrajo consigo unaredistribución muy notoria dela MO en agregados detamaños diferentes.
4. La descomposición de la MOdel suelo asociada conpartículas de limo y arcilla sehalla, al parecer, en unestado más avanzado en elsuelo circundante que en lasheces.
Capítulo 16Dinámica del carbono y delnitrógeno en las heces dediferente edad depositadaspor lombrices en áreas desabana y de pastizal en losLlanos Orientales deColombia1. Las lombrices seleccionan
sustratos alimenticios cuyovalor de C orgánico se

Conclusiones Generales...
427
incrementa durante elenvejecimiento de losturrículos, lo cual se debe, enparte, a los procesos defijación del CO2, a lasraicillas muertas o a laactividad en las heces dealgunos grupos demacrofauna.
2. Durante el primer mes seobservan contenidos altos deNH4 en las heces de losindividuos deM. carimaguensis a causa,principalmente, de ladeposición continua dematerial fresco sobre elturrículo. Una vez que lalombriz abandona la galería,ese nivel desciende y elcontenido de N inorgánicoque queda provendría de laproducción de NO3
en hecesfrescas mediante procesos denitrificación.
3. En las heces frescas seobserva una rápidaproducción de NO3
que, mástarde, se difunde hacia elsuelo circundante, primeroen sentido vertical y despuésen sentido horizontal.
4. Entre 3 y 34 kg/ha al año deN inorgánico se liberan en lasheces frescas de losindividuos de M.carimaguensis en la sabana yen el pastizal,respectivamente, lo queequivale a casi un 22% del Ntotal anual asimilado por esteúltimo.
5. La gran cantidad de sueloprocesado por las lombriceshace extensivos,especialmente en el manejode la MO del suelo, losefectos que causaglobalmente dicho proceso enla fertilidad del suelo y en laproducción vegetal; éste esun paso fundamental en el
mejoramiento de lasostenibilidad de losagroecosistemas y en ladisminución de las emisionesde CO2 a la atmósfera.
Capítulo 17Características delintercambio iónico en lasheces de la lombriz anécicaMartiodrilus carimaguensisJiménez y Moreno, en unOxisol de una sabana deColombia1. La introducción de pastizales
en los suelos de sabana y elmanejo de éstos conducen acambios importantes en dosparámetros: la fertilidad delsuelo y el comportamientoelectroquímico de susnutrientes.
2. El tránsito del suelo por elintestino de la lombrizocasiona grandes cambios enel punto de carga cero,especialmente en la sabana,donde la calidad de la MO delsuelo y de la hojarasca esmuy baja.
3. Los cambios de laspropiedades del suelo quederivan de la actividad de laslombrices pueden llevar auna mejora en la capacidadde intercambio catiónico delos Oxisoles infértiles.
Capítulo 18Las comunidades de lamacrofauna de la superficiedel suelo asociadas con lasheces de las lombrices detierra en los LlanosOrientales de Colombia1. La densidad y la biomasa de
las tres categorías ecológicasde lombrices fueronfuertemente afectadas por la
-
-
+

428
El Arado Natural:…
cantidad de turrículosdepositados por M.carimaguensis sobre lasuperficie del suelo; además,las comunidades deorganismos encontradas enlos turrículos estabanrepresentadasprincipalmente (en sudensidad y en su biomasa)por especies epígeas en lasabana y por especiesanécicas en el pastizal.
2. Las heces de las lombricesparecen mejorar lascondiciones de vida de otrasespecies, puesto que lariqueza taxonómica y ladiversidad de los organismosaumentó cuando seincrementó el número dedeposiciones superficiales.
3. Los resultados obtenidosapoyan la hipótesis de lasbiodiversidades encajadas,según la cual los ingenierosdel ecosistema determinan laabundancia y la estructurade las comunidades deorganismos más pequeños.
Capítulo 19Efectos causados por laslombrices en el banco desemillas permanente delsuelo en sistemas desabana y de pastizal enColombia1. El porcentaje de germinación
de las semillas que sehallaban en las heces de laslombrices fue de 3 a 40 vecesmenor que en lasencontradas en el suelo, aconsecuencia de los dañosque sufren las semillas
cuando son ingeridas por laslombrices. En las hecesdepositadas sobre lasuperficie del suelo hay de18 a 878 semillasgerminables por m2 por año(hasta 8.7 millones desemillas por hectárea poraño), cantidad querepresenta del 1% al 13% deltotal de semillas del bancopermanente del suelo.
2. La diversidad, la riqueza deespecies y la equitatividad dela vegetación disminuyeronconsiderablemente en losagroecosistemas, si secomparan éstos con lasabana nativa.
3. Es débil la relación entre lacomposición específica delbanco de semillas del suelo yla vegetación que crece en lasuperficie del suelo,resultado que se explica porfactores ecológicos, como lareproducción vegetativa, laquema de la sabana, ladispersión de las semillas porel viento o los animales, y elsobrepastoreo.
4. La composición de lavegetación superficial estuvomás cerca de la que seobtuvo en el banco desemillas de las heces que dela obtenida en el banco desemillas del suelo.
5. Es posible considerar que losturrículos de M.carimaguensis son nichosimportantes de regeneraciónde la comunidad vegetal, yque la actividad de laslombrices es un factorrelevante de la diversidad y lasostenibilidad de losecosistemas.

Conclusiones Generales...
429
Capítulo 20Determinación, mediante laespectroscopía de RMN con31P, del nivel de fósforoorgánico de un Oxisol, enpastizales del trópicoderivados de la sabananativa1. El establecimiento de los
pastizales mejoradosaumentó, en más del 40%, elcontenido total de P en losácidos húmicos y fúlvicos, yestos valores fueron de 2 a4 veces superiores en lasheces que en el suelo que lascircundaba, lo que indica unenriquecimiento notable del Pextraíble.
2. El P del ácido húmico estuvosiempre dominado por lasespecies que contenían Porgánico, y la cantidad de Porgánico en el ácido húmicofue proporcionalmente másalta en las heces que en elsuelo.
3. La actividad de las lombricesmejora la disponibilidad del Pen los pastizales porque creauna reserva de P orgánicoque proviene de laestabilización física de ésteen el interior de las heces.
Capítulo 21Alteración de la lignina yde los carbohidratos en losagregados de un Oxisol, enpastizales tropicalesderivados de la sabananativa1. Los turrículos de las
lombrices se caracterizan porun notable enriquecimientoen C, en lignina y enazúcares de la fracción dearena, si se comparan conlos contenidos del suelo
alrededor de ellos, lo quesugiere una importanteincorporación de hojarasca ala parte mineral del suelo(efecto anécico).
2. La concentración decarbohidratos en la MOasociada a la fracción dearenas aumentó según elorden siguiente: sabananativa < B. decumbens < B.decumbens + P. phaseoloides< turrículos de lombrices deB. decumbens + P.phaseoloides.
3. La actividad de las lombricestiene un efecto importante enla redistribución de la MO delsuelo. En la fracción dearenas de los turrículos seencontraron valores altos deC, en comparación con elsuelo que los circundaba. Lahojarasca fresca fueincorporada a la partemineral del suelo, mientrasque la lignina se preservabaen los turrículos.
4. El enriquecimiento de lafracción de limos endetrimento de la de arcillasse produjo, probablemente,por la unión fuerte que existeentre las partículas de arcillay las de limos mediante elmucílago secretado por laslombrices.
Capítulo 22Efecto de la exclusión de lalombriz anécicaMartiodrilus carimaguensisJiménez y Moreno en laspropiedades del suelo y enel crecimiento de lasplantas, en pastizales delos Llanos Orientales deColombia1. Los efectos que causa la
ausencia de M. carimaguensis

430
El Arado Natural:…
en las propiedades del sueloy en el crecimiento de lasplantas están asociados auna compactación grande delsuelo, a un contenido bajo deC, a la saturación alta de Al,a una baja biomasa vegetal ya una alta biomasa demalezas; sin embargo, losprincipales factores quecontrolan las propiedades delsuelo fueron el tipo depastizal y la edad de éste.
2. La pérdida de alguna especie,principalmente si estáasociada a una reducciónimportante de la biomasa,conduce a pérdidassignificativas en elfuncionamiento delecosistema. Porconsiguiente, hay que prestaratención al manejo de lascomunidades de lombrices detierra en los agroecosistemas,con el fin de obtener, graciasa sus actividades, un efectofavorable en la fertilidad delsuelo.
Capítulo 23Estructura espacio-temporal de una comunidadde lombrices de tierra yheterogeneidad del suelo enun pastizal tropical enCarimagua, Colombia1. Mediante un estudio de
análisis espacial y unenfoque del nivel más alto almás bajo (“top-down”) sedemostró la existencia deuna estructura espacial en lacomunidad de lombrices delas sabanas de Carimagua.
2. Existe una relaciónsignificativa entre laestructura espacio-temporalde la comunidad delombrices y la heterogeneidad
espacial del suelo y de labiomasa radicular.
3. Los patrones de distribuciónespacial son estables ymuestran la oposiciónexistente entre Andiodrilussp. y Glossodrilus sp.,resultado que indicaría lapresencia de un fenómeno deexclusión competitiva.
4. Existe una oposición espacialentre las especies pequeñas ylas de mayor tamaño. Losparches de poblacióndominados por las especiesgrandes se oponen a otros enque las especies pequeñasson más abundantes, ycorresponden a áreascaracterizadas porpropiedades del suelo yvalores de biomasa radicularque son específicos.
Necesidades de laInvestigación Futura
Todo el conocimiento recopilado eneste estudio se está empleando en eldesarrollo de un modelo que permitael manejo indirecto de las actividadesde las lombrices de tierra, con el finde optimizar sus efectos benéficos enlos agroecosistemas (Figura 24-5;L. Mariani sin publicar, citado enLavelle 2000). El objetivo del modeloes establecer una acción permanentede la fauna del suelo en losagroecosistemas, con el fin de deobtener efectos favorables en laestructura física del suelo y en ladinámica de la MO del suelo. Elmodelo explica las interacciones queocurren entre tres entidades: elagroecosistema (o sea, las opcionesde manejo elegidas), las propiedadesdel suelo (la dinámica de la MO delsuelo y la estructura física de éste) yla macrofauna del suelo,especialmente las actividades de losingenieros invertebrados del suelo y

Conclusiones Generales...
431
los efectos que causan lasestructuras biogénicas producidaspor ellos (Lavelle 2000).
El seguimiento de las estructurasbiogénicas producidas por losingenieros del ecosistema del sueloes un primer paso para comprenderlas relaciones existentes entre labiodiversidad y la función que éstadesempeña. Además de este enfoquepreliminar, es necesario fortalecer eltrabajo orientado a entender losaspectos funcionales de estasestructuras, mediante el desarrollode estudios a diferentes escalasespacio-temporales y desde la escalade la estructura hasta la de laparcela o la población. Debenconsiderarse también la dinámica delas estructuras biogénicas durantesu envejecimiento y sus efectos
concomitantes en las diferentespropiedades del suelo. Esta es unalabor importante ya que, cuandodesaparecen de la parcela algunosgrupos de la fauna del suelo (porejemplo, las lombrices de tierra) susefectos pueden persistir dentro de laescala de su dominio funcional, esdecir, la drilosfera. Por lo tanto,gracias a los procesos bien sea físicoso químicos, como se ha mostrado enlos capítulos previos, que ocurren enlas estructuras biogénicas, lasostenibilidad del agroecosistemapuede mantenerse durante algúntiempo. Una prioridad de lainvestigación debería ser ladeterminación del período de vida deestas estructuras y de laspropiedades relacionadas con ellas,por ejemplo, la infiltración del agua.
Figura 24-5. Modelo propuesto para el manejo de las poblaciones de lombrices y de otros ingenieros delecosistema (L. Mariani, sin publicar; citado en Lavelle 2000).
Materia orgánica
Estructura
Disposición espacialde las parcelas
Prácticasagrícolas
AGROECOSISTEMA
SUELO
MigraciónTratamiento del suelo
Otros ingenierosdel ecosistema
Lombrices
Estructurasbiogénicas de
superficie
MACROFAUNA
Valor alimenticiodel suelo
Recursotrófico
Estimulación delcrecimiento vegetal
AgregaciónPorosidad
Indicador

432
El Arado Natural:…
Los ingenieros del ecosistema delsuelo deberían clasificarse tambiénsegún los efectos que ejerzan en laestructura física del suelo, en ladinámica de la MO del suelo, en labiodiversidad de otros organismos yen la producción primaria. Senecesitan más estudiospormenorizados para desarrollaropciones alternas de manejo, si sequiere prevenir o revertir ladegradación del suelo mediante lamanipulación de las actividades delos organismos ingenieros delecosistema que hagan impactopositivo y significativo en laestructura y en la fertilidad del suelo.Si se conserva una faunadiversificada en que haya especiesepígeas, endógeas y anécicas, sefavorecerán también aquellasespecies que no son ingenieras, esdecir, las epígeas o las quetransforman la hojarasca (Lavelle1996). Se puede, por tanto,optimizar e incrementar lasactividades de los organismosbenéficos del suelo a escala de laparcela o del agroecosistema, sindejar de atender los requisitos delproductor en términos deproductividad y de beneficios.
A escala de la parcela se podríaincrementar la capacidad de cargade algunas especies selectas de estosorganismos; sin embargo, y enprimer lugar, se debe conocer lasensibilidad de las diferentesespecies de la comunidad demacroinvertebrados a las diversasprácticas agrícolas empleadas, comola labranza, la fertilización y elcontrol de plagas, así como adiferentes regímenes alimenticios.
Es posible lograr un manejoindirecto de las actividades de lamacrofauna del suelo manteniendodiferentes sistemas de uso de latierra, naturales y no naturales, a laescala del paisaje. Los ecosistemasnaturales, en este caso la sabana
nativa, actuarían como el reservoriooriginal de la fauna del suelo, que esfuncionalmente diverso. Esnecesario estudiar, a la escala delagroecosistema o de la práctica demanejo, la capacidad que tienen lasespecies clave para migrar desde elpastizal mejorado y desde la sabananativa donde los niveles,respectivamente, de biomasa y dediversidad son altos, hacia sistemasde manejo de la tierra, como loscultivos, donde la biodiversidad esbaja.
Las estructuras biogénicas sepueden emplear como índice de lasactividades de los ingenieros delecosistema en los cultivos, yservirían de indicadores ensustitución de las mediciones muylaboriosas que requieren laspoblaciones de dichos ingenieros. Sise logra entender la dinámica (o sea,la producción y la degradación) delas estructuras biogénicas, seráposible establecer las densidadescríticas que sirvan como advertenciaoportuna de los efectos que podríacausar un cambio ocurrido en lasprácticas agrícolas.
Es difícil estudiar este tema auna escala que supere el tamaño delas estructuras biogénicas; por tanto,los trabajos descritos en la mayorparte de los capítulos de la obrasobre los efectos de los ingenieros delecosistema se han realizado en dichaescala. Se requieren aún estudiosque establezcan el papel funcional delos organismos del suelo y lasopciones de que se dispone paraoptimizar sus actividades medianteun modelo de manejo integrado enlos agroecosistemas. Esos estudioscaracterizarían primero lasestructuras biogénicas producidaspor otras especies de lombrices paraprobar la hipótesis de que los efectospercibidos en los procesos del sueloestán, de hecho, jerárquicamenteregulados. La dinámica de las

Conclusiones Generales...
433
malezas y la actividad de laslombrices son también un temadigno de un estudio futuro, lo mismoque el papel de las lombrices en ladispersión de los propágulos demicorriza y en la estimulación de lamicroflora.
Es necesario investigar tambiénel papel de la macrofauna en ladegradación de los pastizalestropicales, especialmente en elCerrado brasileño donde, a diferenciade lo que ocurre en Carimagua, sehan encontrado pruebas evidentes dedegradación física, química ybiológica. Las causas de ladegradación de los pastizales deltrópico húmedo no se hanesclarecido todavía (Fisher et al.1995). Hay que considerar la posiblecontribución de las especiesendémicas de lombriz de tierra —quese adaptan a los cambios ocurridosen el ecosistema original— al diseñode prácticas agrícolas cuyos efectosen el ecosistema sean menosdañinos. Este conocimientopermitirá mejorar la sostenibilidad,ya que el uso calculado de losrecursos biológicos podría mantenerla fertilidad de los ecosistemasnaturales y lograr un nivel alto deproductividad primaria en los suelospobres.
Los bioindicadores de ladiversidad y la calidad del suelo sontemas importantes que deberíanconsiderarse como una prioridad enla investigación futura. A partir de lapresencia y la abundancia de losgrupos de macrofauna mássensibles, se computa un índice y serelaciona con las prácticas agrícolas,para evaluar el valor de labiodiversidad en el mantenimiento delas propiedades físicas del suelo, enla conservación de su MO y en laproducción vegetal primaria. Esposible calcular un índice indirectomediante un análisis multivariableque establezca la relación que existe
entre la biodiversidad, la funciónestructural y las propiedades físicasy químicas del suelo. Estos estudios,tomados en conjunto, darían unmejor manejo del agroecosistemaporque promoverán la integración dela productividad, la sostenibilidad yla conservación de los recursosnaturales.
Finalmente, la adopción de lasprácticas de manejo que incrementeno estimulen, ya sea elestablecimiento o ya elmantenimiento de poblacionessignificativas de macrofauna,requiere de una labor adicional decapacitación y de motivación de losagricultores. Este proceso variará deun país a otro y de una región a otra.Es necesario establecer un lenguajecomún entre los agricultores y loscientíficos, de modo que losprincipios y los conceptos relativos almanejo de los organismos benéficosdel suelo ganen mayor aceptación ysean desarrollados más tarde por lospropios agricultores.
ReferenciasAllen TFH; Starr TB. 1982. Hierarchy:
perspectives for ecologicalcomplexity. University of ChicagoPress, Chicago, IL, Estados Unidos.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Londres. p 94-106.
Barois I; Cadet P; Albrecht A; Lavelle P.1988. Systèmes de culture et faunades sols: Quelques donnés. En:Feller C, ed. Fertilité des sols dansles agricultures paysannescaribéenes: effet des restitutionsorganiques. Institut Français deRecherche Scientifique pour leDéveloppement en Coopération(ORSTOM)–Martinique, París,Francia. p 85-96.

434
El Arado Natural:…
Barros E; Grimaldi M; Desjardins T;Sarrazin M; Chauvel A; Lavelle P.1998. Conversion of forest intopastures in Amazonia: effects on soilmacrofaunal diversity and soil waterdynamics. En: Proceedings of the16th World Congress of Soil Science,agosto 1998, Montpellier, Francia.ISSS, AISS, IBG, SICS and AFES,Montpellier, Francia. vol. 1, p 203(también en CD-ROM).
Blanchart E; Albrecht A; Alegre J;Duboisset A; Gilot-Villenave C;Pashanasi B; Lavelle P; Brussaard L.1999. Effects of earthworms on soilstructure and physical properties.En: Lavelle P; Brussaard L; HendrixPF, eds. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido. p 149-172.
Bouché MB. 1977. Strategieslombriciennes. En: Lohm U; PerssonT, eds. Soil organisms as componentof ecosystems. Ecol Bull 25:122-132.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm. Nature398:32-33.
CIAT. 1999. Decision making forsustainable natural resourcemanagement: nine tools that help.Cali, Colombia.
Dangerfield JM. 1990. Abundance,biomass and diversity of soilmacrofauna in savanna woodlandand associated managed habitats.Pedobiologia 34:141-150.
Decaëns T. 1999. Rôle fonctionnel etréponses aux pratiques agricoles desvers de terre et d’autres ingénieursécologiques dans les savanescolombiennes. Tesis (Ph.D.).Université Paris VI, París, Francia.412 p.
Decaëns T; Jiménez JJ. 2002. Earthwormcommunities under an agriculturalintensification gradient in Colombia.Plant Soil 240(1):133-143.
Decaëns T; Lavelle P; Jiménez JJ; EscobarG; Rippstein G. 1994. Impact of landmanagement on soil macrofauna inthe Oriental Llanos of Colombia. EurJ Soil Biol 30(4):157-168.
Decaëns T; Galvis JH; Amézquita E. 2001.Properties of some structurescreated by soil ecosystem engineersin a Colombian savanna. C R AcadSci, Ser III. 324(5):465-478.
Decaëns T; Jiménez JJ; Lavelle P. 1999a.Effects of exclusion of the anecicearthworm Martiodriluscarimaguensis Jiménez and Morenoon soil properties and plant growthin grasslands of the Eastern Plains ofColombia. Pedobiologia 43:835-841.
Decaëns T; Rangel AF; Asakawa N;Thomas R. 1999b. Carbon andnitrogen dynamics in earthwormcasts in grasslands of the EasternPlains of Colombia. Biol Fertil Soils30:20-28.
Duboisset A. 1995. Caractérisation etquantification par analyse d’imagedes modifications structuralesengendrées par Pontoscolexcorethrurus (Glossoscolecidae,Oligochaeta) sur un sol ferrallitiquecultivé (Pérou). DEA Pédologia,Université Nancy I, Francia.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Fisher MJ; Rao IM; Thomas RJ; LascanoCE. 1995. Grasslands in the well-watered tropical lowlands. En:Hodgson J; Illius AW, eds. Theecology and management of grazingsystems. CAB International,Wallingford, Reino Unido.p 393-425.

Conclusiones Generales...
435
Fisher MJ; Thomas RJ; Rao IM. 1997.Management of tropical pastures inacid-soil savannas of SouthAmerica for carbon sequestration inthe soil. En: Lal R; Kimble JM;Follet RF; Stewart BA, eds.Management of carbonsequestration in the soil. CRCPress, Boca Ratón, FL, EstadosUnidos. p 405-420.
Fragoso C; Brown GG; Patrón JC;Blanchart E; Lavelle P; PashanasiB; Senapati BK; Kumar T. 1997.Agricultural intensification, soilbiodiversity and ecosystem functionin the tropics: the role ofearthworms. Appl Soil Ecol 6:17-35.
Fragoso C; Lavelle P; Blanchart E;Senapati BK; Jiménez JJ; MartínezMA; Decaëns T; Tondoh J. 1999.Earthworm communities of tropicalagroecosystems: origin, structureand influence of managementpractices. En: Lavelle P; BrussaardL; Hendrix PF, eds. Earthwormmanagement in tropicalagroecosystems. CAB International,Wallingford, Reino Unido. p 27-55.
Giller PS. 1996. The diversity of soilcommunities, the “poor man’stropical rainforest”. BiodiversConserv 5:135-168.
Giller KE; Beare MH; Lavelle P; Izac A-MN; Swift MJ. 1997. Agriculturalintensification, soil biodiversity andagroecosystem function. Appl SoilEcol 6:3-16.
Gilot-Villenave C; Lavelle P; Keli J;Kouassi P; Guillaume G. 1995.Biological activity of soil underrubber plantations in Côte d’Ivoire.Acta Zool Fenn 196:186-189.
Huston MA. 1996. Biological diversity:the coexistence of species inchanging landscape. CambridgeUniversity Press, Reino Unido.
Jiménez JJ. 1999. Estructura de lascomunidades y dinámica de laspoblaciones de lombrices de tierraen sabanas naturales y perturbadasde Carimagua (Colombia). Tesis(Doctorado). UniversidadComplutense de Madrid, España.311 p.
Jiménez JJ; Decaëns T. 2000. Verticaldistribution of earthworms ingrassland soils from the ColombianLlanos. Biol Fertil Soils 32:463-473.
Jiménez JJ; Brown GG; Decaëns T; FeijooT; Lavelle P. 2000. Differences inthe timing of diapause and patternsof aestivation in some tropicalearthworms. Pedobiologia44:677-694.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78(7):1946-1957.
Kalisz PJ; Wood HB. 1995. Native andexotic earthworms in wildlandecosystems. En: Hendrix PF, ed.Earthworm ecology andbiogeography in North America.Lewis Publishers, Boca Raton, FL,Estados Unidos. p 117-126.
Lavelle P. 1978. Les vers de terre de lasavanne de Lamto (Côte d’Ivoire):Peuplements, populations etfonctions dans l’ecosystème. Tesis(Doctorado). Université Paris VI,Francia.
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecol 2(2):117-133.

436
El Arado Natural:…
Lavelle P. 1983. The soil fauna of tropicalsavannas; II: The earthworms. En:Bourlière F, ed. Tropical savannas.Elsevier Scientific Publishing,Amsterdam, Holanda. p 485-504.
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P. 2000. Ecological challenges forsoil science. Soil Sci 165:73-86.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B. 1994. Therelationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. Wiley andSons, Chichester, Reino Unido.p 137-169.
Lavelle P; Brussaard L; Hendrix PF, eds.1999. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido.
Lawton JH; Jones CG. 1995. Linkingspecies and ecosystems: organismsas ecosystem engineers. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Londres. p 141-150.
Lee KE. 1991. The diversity of soilorganisms. En: Hawksworth DL, ed.The biodiversity of microorganismsand invertebrates: its role insustainable agriculture. CABInternational, Wallingford, ReinoUnido. p 73-87.
Mariani L; Bernier N; Jiménez JJ; DecaënsT. 2001. Régime alimentaire d’un verde terre des savanes colombiennes:une remise en question des typesécologiques. C R Acad Sci, Ser III.324(8):733-742.
Ortiz B; Fragoso C; M’Boukou I; PashanasiB; Senapati BK; Contreras A. 1999.Perception and use of earthworms intropical farming systems. En: LavelleP; Brussaard L; Hendrix PF, eds.Earthworm management in tropicalagroecosystems. CAB International,Wallingford, Reino Unido.p 239-252.
Rao IM. 1998. Root distribution andproduction in native and introducedpastures in the South Americansavannas. En: Box, Jr. JE, ed. Rootdemographics and their efficienciesin sustainable agriculture,grasslands and ecosystems. KluwerAcademic Publishers, Amsterdam,Holanda. p 19-41.
Rippstein G; Lascano CE; Decaëns T.1996. La production fourragère dansles savanes d’Amerique de Sudintertropicale. Fourrages 145:33-52.
Sanz JI; Zeigler RS; Sarkarung S; MolinaDL; Rivera M. 1999. Sistemasmejorados arroz-pasturas parasabana nativa y pasturasdegradadas en suelos ácidos deAmérica del Sur. En: Guimarães EP;Sanz JI; Rao IM; Amézquita MC;Amézquita E, eds. Sistemasagropastoriles en sabanas tropicalesde América Latina. CIAT, Cali,Colombia. p 232-244.
Schneidmadl J; Decaëns T. 1995. Soilmacrofauna in managed savannas atMatazul farm, Eastern Plains,Colombia. En: CIAT annual report,1995. CIAT, Cali, Colombia.
Senapati BK; Panigrahi PK; Lavelle P.1994. Macrofaunal status andrestoration strategy in degraded soilunder tea cultivation in India. En:Etchevers JD, ed. Proceedings of the15th World Congress of Soil Science,Acapulco, México. p 64-75.

Conclusiones Generales...
437
Senapati BK; Lavelle P; Giri S; PashanasiB; Alegre J; Decaëns T; Jiménez JJ;Albrecht A; Blanchart E; MahieuxM; Rousseaux L; Thomas RJ;Panigrahi PK; Venkatachalam M.1999. In-soil earthwormtechnologies for tropicalagroecosystems. En: Lavelle P;Brussaard L; Hendrix PF, eds.Earthworm management in tropicalagroecosystems. CAB International,Wallingford, Reino Unido.p 199-237.
Solbrig OT; Medina E; Silva JF. 1996.Determinants of tropical savannas.En: Solbrig OT; Medina E; Silva JF,eds. Biodiversity and savannaecosystem processes. Springer-Verlag, Berlín, Alemania. p 31-41.
Swift MJ; Anderson JM. 1994.Biodiversity and ecosystem functionin agricultural systems. En: SchulzeED; Mooney HA, eds. Biodiversityand ecosystem function. Springer-Verlag, Berlín, Alemania. p 15-41.
Swift MJ; Vandermeer J; RamakrishnanPS; Anderson JM; Ong CK; HawkinsBA. 1996. Biodiversity andagroecosystem function. En:Mooney HA; Cushman JH; MedinaE; Sala OE; Schulze ED, eds.Functional roles of biodiversity: aglobal perspective. John Wiley andSons, Nueva York, Estados Unidos.p 261-298.
Thomas RJ. 1992. The role of legume inthe nitrogen cycle of productive andsustainable pastures. Grass ForageSci 47:133-142.
Thomas RJ; Lascano CE; Sanz JI; AraMA; Spain JM; Vera RR; Fisher MJ.1995. The role of forage grasses andlegumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science Series. LewisPublishers, Boca Ratón, FL,Estados Unidos. p 61-83.


















