
Phenotypic and Molecular Characterization of Brazilian ...
15
agronomy Article Phenotypic and Molecular Characterization of Brazilian Capsicum Germplasm Bruna Dias Gomes Brilhante 1 , Talles de Oliveira Santos 2 , Pedro Henrique Araújo Diniz Santos 2 , Samuel Henrique Kamphorst 2, * , José Dias Souza Neto 3 , Leandro Heitor Rangel 4 , Fernanda Vargas Valadares 4 , Rafael Nunes de Almeida 4 , Rosana Rodrigues 2 , Alexandre Cristiano Santos Júnior 3 and Monique Moreira Moulin 3, * Citation: Brilhante, B.D.G.; Santos, T.d.O.; Santos, P.H.A.D.; Kamphorst, S.H.; Neto, J.D.S.; Rangel, L.H.; Valadares, F.V.; de Almeida, R.N.; Rodrigues, R.; Júnior, A.C.S.; et al. Phenotypic and Molecular Characterization of Brazilian Capsicum Germplasm. Agronomy 2021, 11, 854. https://doi.org/10.3390/ agronomy11050854 Academic Editor: Monica Boscaiu Received: 26 March 2021 Accepted: 24 April 2021 Published: 27 April 2021 Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affil- iations. Copyright: © 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https:// creativecommons.org/licenses/by/ 4.0/). 1 Laboratório de Melhoramento Genético Vegetal, Universidade Federal do Espírito Santo (UFES), Alegre, ES 29500-000, Brazil; [email protected] 2 Laboratory of Genetics and Plant Breeding, Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF), Campos dos Goytacazes, RJ 28013-602, Brazil; [email protected] (T.d.O.S.); [email protected] (P.H.A.D.S.); [email protected] (R.R.) 3 Laboratory of Genetics and Molecular Biology, Instituto Federal do Espírito Santo—Campus de Alegre (IFES), Alegre, ES 29500-000, Brazil; [email protected] (J.D.S.N.); [email protected] (A.C.S.J.) 4 Laboratory of Agricultural Engineering, Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF), Campos dos Goytacazes, RJ 28013-602, Brazil; [email protected] (L.H.R.); [email protected] (F.V.V.); [email protected] (R.N.d.A.) * Correspondence: [email protected] (S.H.K.); [email protected] (M.M.M.) Abstract: Population growth and the high demand for food production has caused environmental degradation losses in biodiversity. The conservation of genetic resources is the most appropriate strategy for maintaining the variability of species of great importance, such as Capsicum. In this study, we describe the characterization of 69 pepper accessions of four Capsicum species from different regions of Brazil on the basis of qualitative and quantitative descriptors and ISSR markers. The 11 quantitative descriptors grouped the 69 populations into five clades. The fruit traits had the greater discrimination power. A strong correlation was detected between some pairs of quantitative descriptors: petiole and fruit length, fruit diameter and fresh fruit mass, leaf length and leaf width, canopy and plant height, stem diameter and plant height, stem diameter and canopy diameter. A negative correlation was also observed between plant height and pericarp thickness. Moreover, the molecular analysis grouped accessions into five clades. The morpho-agronomic and molecular characterization were efficient to estimate the genetic diversity between accessions, being important tools for the knowledge and use of accessions. This study is of great importance to preserve pepper germplasm and to provide data to facilitate the process of the selection in breeding programs. Keywords: peppers; genetic resources; germplasm bank; molecular markers 1. Introduction The genus Capsicum (Solanaceae) is home to peppers, a culture extremely relevant to Brazil and grown all over the world. The genus has about 42 described species [1]. Of these, five species are classified as domesticated: Capsicum annuum L., Capsicum baccatum L. var. pendulum, Capsicum pubescens Ruiz and Pav., Capsicum frutescens L. and Capsicum chinense Jacq. The other species are considered semi-domesticated and wild. Pepper is considered to be one of the most important vegetables in the world. This is attributed to its versatility, since its use is used in the culinary, pharmaceutical, and cosmetics industry [2]. The most pungent varieties have great appeal in the markets of Southeast Asia, South America, and also in Africa, while sweet peppers and non-pungent varieties are more consumed in North America and Europe [3]. According to the latest data from FAO (Food and Agriculture Organization) [4], in 2019, around 38 million and 4.3 million tons of green and dry pepper were produced worldwide, Agronomy 2021, 11, 854. https://doi.org/10.3390/agronomy11050854 https://www.mdpi.com/journal/agronomy
Transcript of Phenotypic and Molecular Characterization of Brazilian ...

agronomy
Article
Phenotypic and Molecular Characterization of BrazilianCapsicum Germplasm
Bruna Dias Gomes Brilhante 1, Talles de Oliveira Santos 2 , Pedro Henrique Araújo Diniz Santos 2 ,Samuel Henrique Kamphorst 2,* , José Dias Souza Neto 3, Leandro Heitor Rangel 4, Fernanda Vargas Valadares 4,Rafael Nunes de Almeida 4 , Rosana Rodrigues 2 , Alexandre Cristiano Santos Júnior 3
and Monique Moreira Moulin 3,*
�����������������
Citation: Brilhante, B.D.G.; Santos,
T.d.O.; Santos, P.H.A.D.; Kamphorst,
S.H.; Neto, J.D.S.; Rangel, L.H.;
Valadares, F.V.; de Almeida, R.N.;
Rodrigues, R.; Júnior, A.C.S.; et al.
Phenotypic and Molecular
Characterization of Brazilian
Capsicum Germplasm. Agronomy 2021,
11, 854. https://doi.org/10.3390/
agronomy11050854
Academic Editor: Monica Boscaiu
Received: 26 March 2021
Accepted: 24 April 2021
Published: 27 April 2021
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2021 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
1 Laboratório de Melhoramento Genético Vegetal, Universidade Federal do Espírito Santo (UFES),Alegre, ES 29500-000, Brazil; [email protected]
2 Laboratory of Genetics and Plant Breeding, Universidade Estadual do Norte Fluminense DarcyRibeiro (UENF), Campos dos Goytacazes, RJ 28013-602, Brazil; [email protected] (T.d.O.S.);[email protected] (P.H.A.D.S.); [email protected] (R.R.)
3 Laboratory of Genetics and Molecular Biology, Instituto Federal do Espírito Santo—Campus de Alegre (IFES),Alegre, ES 29500-000, Brazil; [email protected] (J.D.S.N.); [email protected] (A.C.S.J.)
4 Laboratory of Agricultural Engineering, Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF),Campos dos Goytacazes, RJ 28013-602, Brazil; [email protected] (L.H.R.);[email protected] (F.V.V.); [email protected] (R.N.d.A.)
* Correspondence: [email protected] (S.H.K.); [email protected] (M.M.M.)
Abstract: Population growth and the high demand for food production has caused environmentaldegradation losses in biodiversity. The conservation of genetic resources is the most appropriatestrategy for maintaining the variability of species of great importance, such as Capsicum. In this study,we describe the characterization of 69 pepper accessions of four Capsicum species from differentregions of Brazil on the basis of qualitative and quantitative descriptors and ISSR markers. The11 quantitative descriptors grouped the 69 populations into five clades. The fruit traits had thegreater discrimination power. A strong correlation was detected between some pairs of quantitativedescriptors: petiole and fruit length, fruit diameter and fresh fruit mass, leaf length and leaf width,canopy and plant height, stem diameter and plant height, stem diameter and canopy diameter. Anegative correlation was also observed between plant height and pericarp thickness. Moreover,the molecular analysis grouped accessions into five clades. The morpho-agronomic and molecularcharacterization were efficient to estimate the genetic diversity between accessions, being importanttools for the knowledge and use of accessions. This study is of great importance to preserve peppergermplasm and to provide data to facilitate the process of the selection in breeding programs.
Keywords: peppers; genetic resources; germplasm bank; molecular markers
1. Introduction
The genus Capsicum (Solanaceae) is home to peppers, a culture extremely relevantto Brazil and grown all over the world. The genus has about 42 described species [1]. Ofthese, five species are classified as domesticated: Capsicum annuum L., Capsicum baccatumL. var. pendulum, Capsicum pubescens Ruiz and Pav., Capsicum frutescens L. and Capsicumchinense Jacq. The other species are considered semi-domesticated and wild.
Pepper is considered to be one of the most important vegetables in the world. Thisis attributed to its versatility, since its use is used in the culinary, pharmaceutical, andcosmetics industry [2]. The most pungent varieties have great appeal in the markets ofSoutheast Asia, South America, and also in Africa, while sweet peppers and non-pungentvarieties are more consumed in North America and Europe [3].
According to the latest data from FAO (Food and Agriculture Organization) [4], in 2019,around 38 million and 4.3 million tons of green and dry pepper were produced worldwide,
Agronomy 2021, 11, 854. https://doi.org/10.3390/agronomy11050854 https://www.mdpi.com/journal/agronomy

Agronomy 2021, 11, 854 2 of 15
respectively, in a cultivated area of about 3.7 million hectares. Pepper consumption andproduction data in Brazil are still scarce, mainly because most peppers are produced bysmall producers and sold in popular markets and fairs. However, even though little isknown about the pepper market in Brazil, they are part of the country’s wealth and cultureand are a valuable asset of biodiversity [5]. A wide variety of types, names, size, colors,and flavor pungencies are found throughout Brazilian territory [6,7]. It is estimated thatthe country has the largest number of wild species, in addition to being considered animportant center of diversity of domesticated species of the genus Capsicum [8,9].
With the growing global demand for food—a consequence of the increase inpopulation—there is also an increase in the cultivated area and inadequate exploitationof the available genetic resources [10–12]. As a result, there is a great loss of biodiversityfor many species, resulting from the process of genetic erosion. These anthropic actionsrequire immediate actions to maintain the resources available for human survival. In thiscontext, the United Stations 2030 Agenda [13] provides several guidelines for more sus-tainable production models that guarantee food security and the preservation of availablegenetic resources.
From this perspective, accessing existing genetic variability and creating measuresto preserve existing genetic resources is one of the most important strategies for plantbreeding programs [14,15]. This is because the success in improving a variety lies in thegenetic diversity available in the studied germplasm. Therefore, in order to meet thedemand of the consumer market with regard to the productivity and quality of Capsicum—an extremely important culture for Brazil—it is essential that breeders have access to thegenetic diversity sheltered by the genus Capsicum. That is the reason why it is so importantto implement measures to preserve local varieties, as well as studies on the collection andcharacterization of germplasm.
In order for one to make the accessions of a germplasm bank available for use inimproving characteristics and launching new cultivars, it is essential that they be properlydescribed [16,17]. In this sense, Capsicum germplasm characterization studies have beenbased on predictive methods that take into account agronomic, physiological, morpho-logical, bromatological, and molecular descriptors [18–21]. The study of morphologicalcharacters associated with molecular markers has been used by many studies because theyprovide more accurate genetic information of the genotypes studied [22].
Many molecular markers are used to study Capsicum diversity, namely, RAPD (randomamplification polymorphism DNA) [23–25], AFLP (amplified fragment length polymor-phism) [26–28], SSR (simple sequence repeat) or microsatellites [29–32], and ISSR (intersimple sequence repeats) [8,33–36]. As the new generation sequencing techniques havebecome more accessible, new studies were carried out and were able to access the variationsexisting in Capsicum in greater depth. As an example, the study by D’Agostino et al. [37]determined the complete plastome nucleotide sequence of eight Capsicum species, allowingfor the identification of sequence variations that enable the development of extremelyuseful and informative PCR-based markers to discriminate Capsicum species. More recently,Colonna et al. [38], in a study sequencing 1.8% of the genome of 11 commercially importantCapsicum species from 51 countries, described the genomic variation in addition to findingloci associated with fruit-related phenotypes, which guarantees future studies with greaterprecision in the association of characteristics of interest for breeding programs.
The associated use of phenotypic characteristics to molecular studies of accessionsgenerates more information about the diversity existing in a germplasm collection, whichallows for significant advances in the identification of accessions with potential for breedingprograms. In addition, it provides an overview of the species’ conservation status, ensuringthat strategic measures are taken to conserve the species’ genetic resources. In this study,we describe the characterization of 69 Capsicum accessions from different regions of Brazil,performed using qualitative and quantitative characters and ISSR molecular markers.ISSR is one of the most widely used molecular markers that are reproducible, highlypolymorphic, independent of environmental influence, cost-effective, and do not require

Agronomy 2021, 11, 854 3 of 15
prior sequence knowledge [39]. The results obtained contribute to the knowledge of thepotential and use of accessions in breeding programs, as well as to the conservation ofpepper species, a culture of great importance in the country.
2. Materials and Methods2.1. Plant Material, Experimental Conditions, and Phenotyping
Sixty-nine Capsicum accessions from the germplasm bank of the Instituto Federal doEspírito Santo (IFES)—Campus de Alegre, located in Alegre, ES, Brazil, were evaluated.Sixty-four accessions were from four Brazilian regions (North, Northeast, Midwest, andSoutheast) and five were commercial varieties (IFES 09, IFES 51, IFES 99, IFES 100, andIFES 101) from Alegre, ES (Southeast). These accessions were characterized into fourspecies (Capsicum chinense, Capsicum baccatum var. pendulum, Capsicum frutescens, andCapsicum annuum) according to the description of Moscone et al. [40] (SupplementaryMaterials—Table S1).
Three seeds of each accession were sown in a 128-cell polystyrene trays containingcommercial substrate. Germination and seedling development were monitored daily, andirrigation was performed once a day, usually in the morning. There was no humidity andtemperature control. The seedlings were kept in greenhouse conditions until transplantingto the field, at the stage of four to six pairs of definitive leaves. In the field, the plants werearranged at a spacing of 1.0 m between rows and 0.5 m between plants. By the occurrenceof thinning, only one seedling of each accession was kept.
The experiment was carried out in a complete randomized block design with fourreplications, totaling 276 experimental plots in the experimental area of the same institution.Physico-chemical and granulometric analysis of the soil in the experimental area wasperformed to calculate the need for correctives and fertilizers. Covering fertilization wasperformed 30 days after planting; 3 g of the NPK 25-00-20 (Heringer S.A., Manhuaçu, MG,Brasil) formulation was used. All cultural treatments used followed the recommendationsfor the pepper culture. The irrigation shifts vary according to the plants needing at differentstages of development.
The accessions were characterized on the basis of 33 specific morpho-agronomictraits (Tables S2 and S3) for Capsicum following Bioversity International descriptors forthe genus [41]. Eleven descriptors were based on quantitative traits and 22 were based onqualitative traits. Among qualitative descriptors, the presence of capsaicin in fruits wasevaluated on the basis of Derera’s [42] method, where a fruit placenta sample (approxi-mately 1 cm2) was immersed in a solution of 3 mL of ammonium vanadate for 15 h. Afterthis period, the presence of brown spots in the placenta indicated the presence of capsaicin.
2.2. Genotyping
Samples of young leaves from the accessions were collected in bulk for extraction ofgenomic DNA. The extraction followed the procedure described by Doyle and Doyle [43],with modifications proposed by Daher [44]. The integrity and quantification of the genomicDNA were verified via 0.8% agarose gel using the High DNA Mass Ladder marker (100 pb)(Invitrogen, Waltham, MA, USA).
On the basis of some studies published in the literature [45–48], we selected and tested50 ISSR primers to determine optimal amplification reaction conditions. Of this amount,17 ISSR primers were selected for the analysis of DNA amplification reactions in thisstudy once they resulted in profiles of well-separated fragments. The ideal amplificationtemperature (Tm) and cycling time and discarded ISSR markers can be find at Table S4. ThePCR reactions were carried out in a thermocycler (Veriti 384-well Thermal Cycler AppliedBiosystems, Invitrogen, Waltham, MA, USA). The PCR reaction was carried out in a final25 µL, containing 30 ng of template DNA (3 µL), 13.3 µL of ultrapure water, 2.5 µL ofbuffer + 1 × PCR MgCl2 (1.9 mM), 2 µL of dNTP mix (0.4 mM), 0.2 µL of Taq polymerase(1 U), and 4 µL of each primer (0.08 µM) (Cellco Biotec). The PCR reactions were conductedas follows: 5 min at 94 ◦C for initial denaturation, followed by 45 cycles of 5 min at 94 ◦C,

Agronomy 2021, 11, 854 4 of 15
30 s at 46.6–52 ◦C, 3 min at 72 ◦C, and a final extension of 7 min at 72 ◦C. The amplifiedfragments were separated on a 2% agarose gel, stained with 4 µL of the TIV dye, andsubjected to UV light from the Minibis Pro photocomputer (Bio-imaging System, Israel,Jerusalem), evaluating the most consistent bands (Figure S1).
2.3. Data Analysis
Qualitative descriptors were subjected to descriptive statistical analysis, while quan-titative descriptors were subjected to analysis of variance (ANOVA) and means valueswith significant differences were grouped using the Scott-Knott test (α = 0.05). The relativecontribution of quantitative morpho-agronomic traits for diversity between accessions wasobtained through the Singh [49] method. The estimates of Pearson’s correlation coefficientswere obtained on the basis of the average of repetitions between the traits combined two bytwo. The divergence between accessions was obtained by the average Euclidean distanceon the basis of the 33 morpho-agronomic descriptors. The values of the morpho-agronomictraits were used for UPGMA (unweighted pair group method with arithmetic mean) clus-tering analysis in RStudio software [50]. To evaluate the consistency of the clusters, wecalculated the co-phenetic correlation coefficient [51]. The significance of this coefficientwas verified using the Student’s t-test at 5% probability. Statistical analyses were performedwith the software GENES [52].
ISSR results were interpreted as presence or absence of bands and expressed in abinary matrix. The dissimilarity matrix between accessions based on molecular markerswas obtained using the Jaccard index and grouping performed using the UPGMA method.The correlation between the dissimilarity matrices of the morpho-agronomic descriptorsand ISSR markers was obtained with the aid of the Dendextend package [53] in the RStudiosoftware (version 4.0.4) [50].
3. Results3.1. Morpho-Agronomic Characterization
The qualitative traits stem shape, stem pubescence, and calyx pigmentation traits wereconsidered monomorphic and were classified as angular, sparse, and present, respectively(Table S5). The intermediate growth habit was found in 82.6% of the accessions, while14.5% had an erect growth habit and 2.9% prostrate habit.
The shape of the leaves varied between ovate (49.3%), deltoid (44.9%), and lanceolate(5.8%). The nodal anthocyanin grouped the accessions into four classes: light purple(55.1%), green (24.7%), purple (14.5%), and dark purple (5.7%).
Analyzing flowers, 53.6% had an intermediate position, 43.5% erect, and 2.9% pendant.The number of flowers per axillary varied from one (36.2%), two and three (10.2%), or more(53.6%). The anthers showed a violet color for 66.7% of the accessions and yellow with alight blue spot for 33.3%. The corolla color grouped the accessions into four distinct classes:greenish white (55.1%), white (34.8%), greenish yellow (8.7%), and purple (1.4%). Mostof the accessions (68.1%) did not present spots on the corolla. In contrast, the remaining31.9%, referring to accessions of C. baccatum var. pendulum exhibited a greenish yellowcorolla spot. The filament color varied between white (73.9%), yellow (23.3%), green (1.4%),and violet (1.4%). Analyzing the calyx margin, we found that 75.3% of accessions hadintermediate margin, 18.9% dentate margin, and 5.8% entire margin. Analyzing the calyxconstriction, we found it was present only in C. chinense, which represents 53.6% of all theevaluated accessions.
The fruits presented wide phenotypic variation (Figure 1). Regarding to the color ofthe fruits, we observed that, in the intermediate stage, the fruits presented color rangingfrom orange (78.2%), yellow (10.3%), green (7.2%), dark purple (2.9%), and red (1.4%). Inthe mature stage, 43.5% of accessions had medium red fruits, 34.8% dark red, 5.8% orange,4.3% pale orange, 4.3% light red, 2.9% lemon yellow, and 2.9% orange-yellow (Figure 1).There was also variation in the shape of the fruits—the triangular and elongated shapes

Agronomy 2021, 11, 854 5 of 15
being the ones that occurred most at 39.1% and 36.2%, respectively, followed by blocky(11.6%), round shape (8.7%), and campanulate (4.4%).
Agronomy 2021, 11, x FOR PEER REVIEW 5 of 15
The fruits presented wide phenotypic variation (Figure 1). Regarding to the color of the fruits, we observed that, in the intermediate stage, the fruits presented color ranging from orange (78.2%), yellow (10.3%), green (7.2%), dark purple (2.9%), and red (1.4%). In the mature stage, 43.5% of accessions had medium red fruits, 34.8% dark red, 5.8% orange, 4.3% pale orange, 4.3% light red, 2.9% lemon yellow, and 2.9% orange-yellow (Figure 1). There was also variation in the shape of the fruits—the triangular and elongated shapes being the ones that occurred most at 39.1% and 36.2%, respectively, followed by blocky (11.6%), round shape (8.7%), and campanulate (4.4%).
Most fruits showed a smooth surface (56.6%), followed by semi wrinkled (42%) and wrinkled (1.4%). For the fruit shape at blossom end, we found that 52.2% were pointed, 27.5% sunken, 17.4% blunt, and 2.9% pointed and sunken. In the analysis of fruit shape at pedicel attachment, we observed that the obtuse shape was the one that most occurred (55.1%), followed by the truncate (20.3%), acute (10.1%), cordate (11.6%), and lobate (2.9%). Regarding the corrugation of the fruit, 38 accessions—55.1%—were grouped into the slightly corrugated class, followed by very corrugated (23.2%) and intermediate (21.7%). Capsaicin was present in 75.4% of accessions and absent in the remaining 24.6%.
Figure 1. Phenotypic representation of fruits of 69 Brazilian Capsicum spp. accessions studied. *—commercial varieties.
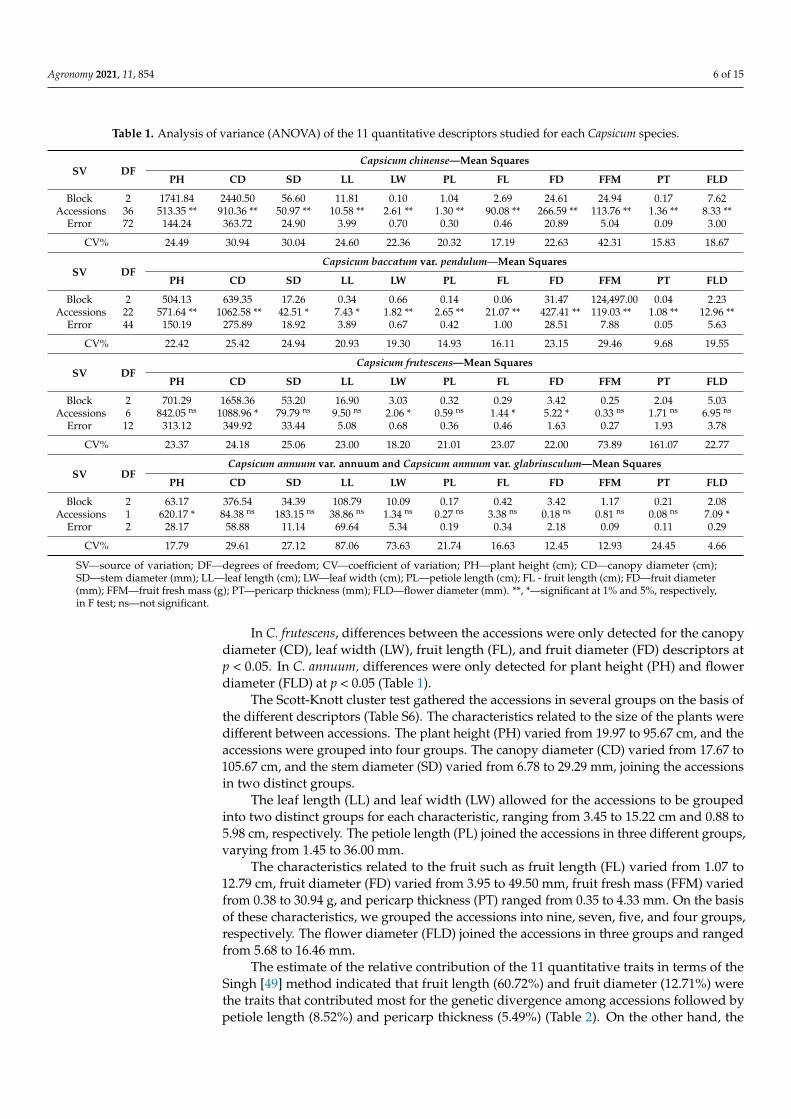
Concerning quantitative traits, analysis of variance (ANOVA) resulted in the detec-tion of significant phenotypic variation between the 69 accessions. In particular, great var-iability was observed in Capsicum chinense and C. baccatum var. pendulum accessions in terms of the 11 quantitative descriptors, with a significant difference between genotypes being detected by F test (α = 0.01), except for the stem diameter (SD) and leaf length (LL), which exhibited differences at p < 0.05 (Table 1).
In C. frutescens, differences between the accessions were only detected for the canopy diameter (CD), leaf width (LW), fruit length (FL), and fruit diameter (FD) descriptors at p
*
*
*
*
*
Figure 1. Phenotypic representation of fruits of 69 Brazilian Capsicum spp. accessions studied. *—commercial varieties.
Most fruits showed a smooth surface (56.6%), followed by semi wrinkled (42%) andwrinkled (1.4%). For the fruit shape at blossom end, we found that 52.2% were pointed,27.5% sunken, 17.4% blunt, and 2.9% pointed and sunken. In the analysis of fruit shapeat pedicel attachment, we observed that the obtuse shape was the one that most occurred(55.1%), followed by the truncate (20.3%), acute (10.1%), cordate (11.6%), and lobate (2.9%).Regarding the corrugation of the fruit, 38 accessions—55.1%—were grouped into theslightly corrugated class, followed by very corrugated (23.2%) and intermediate (21.7%).Capsaicin was present in 75.4% of accessions and absent in the remaining 24.6%.
Concerning quantitative traits, analysis of variance (ANOVA) resulted in the detectionof significant phenotypic variation between the 69 accessions. In particular, great variabilitywas observed in Capsicum chinense and C. baccatum var. pendulum accessions in terms of the11 quantitative descriptors, with a significant difference between genotypes being detectedby F test (α = 0.01), except for the stem diameter (SD) and leaf length (LL), which exhibiteddifferences at p < 0.05 (Table 1).
Agronomy 2021, 11, 854 6 of 15
Table 1. Analysis of variance (ANOVA) of the 11 quantitative descriptors studied for each Capsicum species.
SV DFCapsicum chinense—Mean Squares
PH CD SD LL LW PL FL FD FFM PT FLD
Block 2 1741.84 2440.50 56.60 11.81 0.10 1.04 2.69 24.61 24.94 0.17 7.62Accessions 36 513.35 ** 910.36 ** 50.97 ** 10.58 ** 2.61 ** 1.30 ** 90.08 ** 266.59 ** 113.76 ** 1.36 ** 8.33 **
Error 72 144.24 363.72 24.90 3.99 0.70 0.30 0.46 20.89 5.04 0.09 3.00
CV% 24.49 30.94 30.04 24.60 22.36 20.32 17.19 22.63 42.31 15.83 18.67
SV DFCapsicum baccatum var. pendulum—Mean Squares
PH CD SD LL LW PL FL FD FFM PT FLD
Block 2 504.13 639.35 17.26 0.34 0.66 0.14 0.06 31.47 124,497.00 0.04 2.23Accessions 22 571.64 ** 1062.58 ** 42.51 * 7.43 * 1.82 ** 2.65 ** 21.07 ** 427.41 ** 119.03 ** 1.08 ** 12.96 **
Error 44 150.19 275.89 18.92 3.89 0.67 0.42 1.00 28.51 7.88 0.05 5.63
CV% 22.42 25.42 24.94 20.93 19.30 14.93 16.11 23.15 29.46 9.68 19.55
SV DFCapsicum frutescens—Mean Squares
PH CD SD LL LW PL FL FD FFM PT FLD
Block 2 701.29 1658.36 53.20 16.90 3.03 0.32 0.29 3.42 0.25 2.04 5.03Accessions 6 842.05 ns 1088.96 * 79.79 ns 9.50 ns 2.06 * 0.59 ns 1.44 * 5.22 * 0.33 ns 1.71 ns 6.95 ns
Error 12 313.12 349.92 33.44 5.08 0.68 0.36 0.46 1.63 0.27 1.93 3.78
CV% 23.37 24.18 25.06 23.00 18.20 21.01 23.07 22.00 73.89 161.07 22.77
SV DFCapsicum annuum var. annuum and Capsicum annuum var. glabriusculum—Mean Squares
PH CD SD LL LW PL FL FD FFM PT FLD
Block 2 63.17 376.54 34.39 108.79 10.09 0.17 0.42 3.42 1.17 0.21 2.08Accessions 1 620.17 * 84.38 ns 183.15 ns 38.86 ns 1.34 ns 0.27 ns 3.38 ns 0.18 ns 0.81 ns 0.08 ns 7.09 *
Error 2 28.17 58.88 11.14 69.64 5.34 0.19 0.34 2.18 0.09 0.11 0.29
CV% 17.79 29.61 27.12 87.06 73.63 21.74 16.63 12.45 12.93 24.45 4.66
SV—source of variation; DF—degrees of freedom; CV—coefficient of variation; PH—plant height (cm); CD—canopy diameter (cm);SD—stem diameter (mm); LL—leaf length (cm); LW—leaf width (cm); PL—petiole length (cm); FL - fruit length (cm); FD—fruit diameter(mm); FFM—fruit fresh mass (g); PT—pericarp thickness (mm); FLD—flower diameter (mm). **, *—significant at 1% and 5%, respectively,in F test; ns—not significant.
In C. frutescens, differences between the accessions were only detected for the canopydiameter (CD), leaf width (LW), fruit length (FL), and fruit diameter (FD) descriptors atp < 0.05. In C. annuum, differences were only detected for plant height (PH) and flowerdiameter (FLD) at p < 0.05 (Table 1).
The Scott-Knott cluster test gathered the accessions in several groups on the basis ofthe different descriptors (Table S6). The characteristics related to the size of the plants weredifferent between accessions. The plant height (PH) varied from 19.97 to 95.67 cm, and theaccessions were grouped into four groups. The canopy diameter (CD) varied from 17.67 to105.67 cm, and the stem diameter (SD) varied from 6.78 to 29.29 mm, joining the accessionsin two distinct groups.
The leaf length (LL) and leaf width (LW) allowed for the accessions to be groupedinto two distinct groups for each characteristic, ranging from 3.45 to 15.22 cm and 0.88 to5.98 cm, respectively. The petiole length (PL) joined the accessions in three different groups,varying from 1.45 to 36.00 mm.
The characteristics related to the fruit such as fruit length (FL) varied from 1.07 to12.79 cm, fruit diameter (FD) varied from 3.95 to 49.50 mm, fruit fresh mass (FFM) variedfrom 0.38 to 30.94 g, and pericarp thickness (PT) ranged from 0.35 to 4.33 mm. On the basisof these characteristics, we grouped the accessions into nine, seven, five, and four groups,respectively. The flower diameter (FLD) joined the accessions in three groups and rangedfrom 5.68 to 16.46 mm.
The estimate of the relative contribution of the 11 quantitative traits in terms of theSingh [49] method indicated that fruit length (60.72%) and fruit diameter (12.71%) werethe traits that contributed most for the genetic divergence among accessions followed bypetiole length (8.52%) and pericarp thickness (5.49%) (Table 2). On the other hand, the

Agronomy 2021, 11, 854 7 of 15
traits canopy diameter and leaf width were the ones that contributed less to the divergencebetween accessions, with a relative contribution of 0.75 and 0.17%, respectively.
Table 2. Relative contribution of 11 quantitative traits to genetic divergence among 69 Capsicumaccessions by the Singh method.
Trait Relative Contribution (%)
Fruit length (cm) 60.72%Fruit diameter (mm) 12.71%Petiole length (cm) 8.52%
Pericarp thickness (mm) 5.49%Fruit fresh mass (g) 3.26%Plant height (cm) 3.07%
Flower diameter (mm) 2.31%Leaf length (cm) 1.92%
Stem diameter (mm) 1.09%Canopy diameter (cm) 0.75%
Leaf width (cm) 0.17%
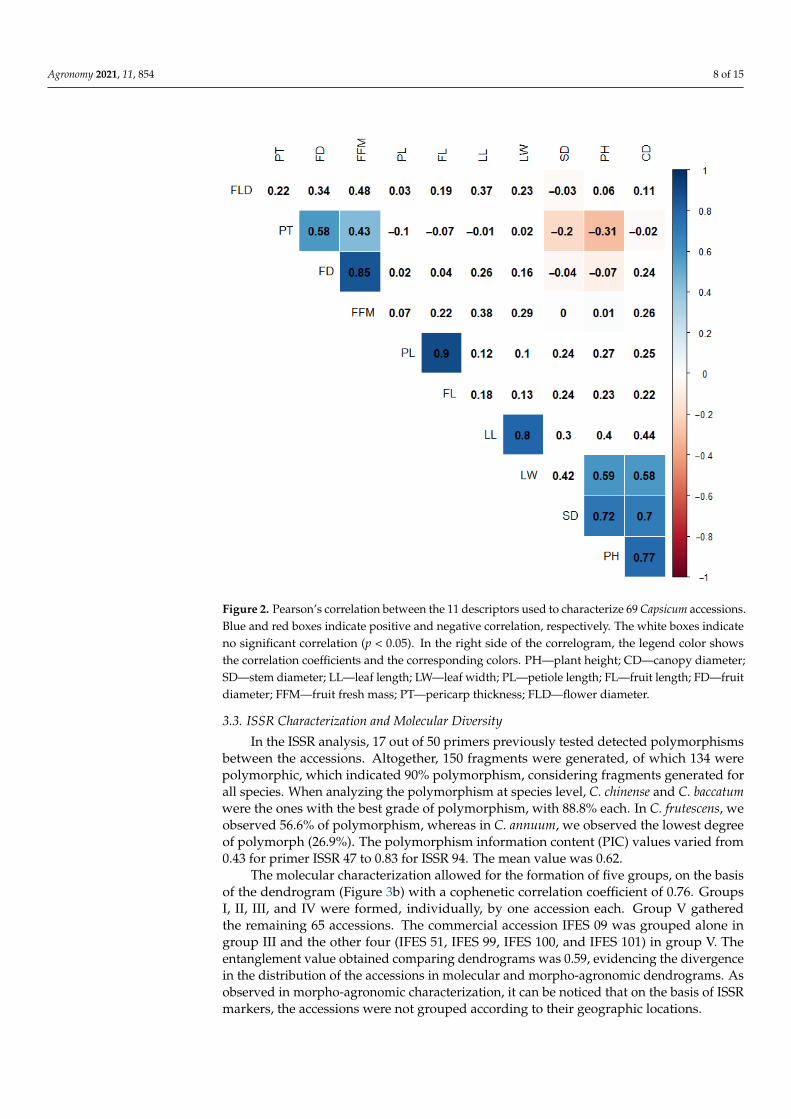
The existence of correlation between the quantitative traits was verified using Pear-son’s correlation coefficient (Figure 2). A very strong correlation was found between petiolelength (PL) and fruit length (FL) (0.90). Strong correlation was found between fruit diam-eter (FD) and fresh fruit mass (FFM) (0.85), leaf length (LL) and leaf width (LW) (0.80),plant height (PH) and canopy diameter (0.77), stem diameter and plant height (0.72), andstem diameter (SD) and canopy diameter (CD) (0.70). A moderate negative correlation wasdetected between plant height (PH) and pericarp thickness (PT) (−0.31).
3.2. Diversity Among Capsicum Accessions and Correlated Traits
To group accessions, we calculated dissimilarities on the basis of the squared Eu-clidean distances. The dendrogram obtained in the grouping of accessions allowed for theformation of five distinct groups (Figure 3a), and the cophenetic correlation coefficient was0.82. Groups I and II allocated only one accession each. The accession allocated in group Ipresented the highest values for plant height, canopy diameter, and stem diameter, havingelongated fruits of orange color in the intermediate stage and red when ripe.
The accession allocated in group II presented the lowest values for the traits of plantheight, canopy diameter, stem diameter, and leaf length and width. Its triangular fruits wereyellow in the intermediate stage and red when ripe. Group III gathered nine accessions,with the highest values for leaf length and width. In addition, they had an intermediategrowth habit; filament colored in white—except for the IFES 02 accession, which hadyellow filament—pigmented calyx; semi-wrinkled fruit surface; and absence of capsaicin,except for the IFES 67 and IFES 91 accessions, which showed pungent fruits.
Group IV was formed by four accessions in which the erect growth habit descriptorspredominated as well as the position of the flowers. The number of flowers per node inthe group’s accessions was two or three and violet in the anthers and greenish yellowcorolla without the presence of spots. The filament of these accessions was yellow in colorand the calyx had the presence of pigment. Its pointed fruits had a long shape with asemi-wrinkled surface and an acute base. In addition, its fruits were slightly corrugatedand with the presence of capsaicin. Group V gathered the remaining 54 accessions, which,in general, had smaller plants; short petioles; and smaller and thinner fruits and with lesspericarp thickness and, consequently, with less fresh mass. The growth habit of plants wasintermediate, and its fruits had capsaicin.
Agronomy 2021, 11, 854 8 of 15
Agronomy 2021, 11, x FOR PEER REVIEW 7 of 15
petiole length (8.52%) and pericarp thickness (5.49%) (Table 2). On the other hand, the traits canopy diameter and leaf width were the ones that contributed less to the divergence between accessions, with a relative contribution of 0.75 and 0.17%, respectively.
Table 2. Relative contribution of 11 quantitative traits to genetic divergence among 69 Capsicum accessions by the Singh method.
Trait Relative Contribution (%) Fruit length (cm) 60.72%
Fruit diameter (mm) 12.71% Petiole length (cm) 8.52%
Pericarp thickness (mm) 5.49% Fruit fresh mass (g) 3.26% Plant height (cm) 3.07%
Flower diameter (mm) 2.31% Leaf length (cm) 1.92%
Stem diameter (mm) 1.09% Canopy diameter (cm) 0.75%
Leaf width (cm) 0.17%
The existence of correlation between the quantitative traits was verified using Pear-son’s correlation coefficient (Figure 2). A very strong correlation was found between pet-iole length (PL) and fruit length (FL) (0.90). Strong correlation was found between fruit diameter (FD) and fresh fruit mass (FFM) (0.85), leaf length (LL) and leaf width (LW) (0.80), plant height (PH) and canopy diameter (0.77), stem diameter and plant height (0.72), and stem diameter (SD) and canopy diameter (CD) (0.70). A moderate negative correlation was detected between plant height (PH) and pericarp thickness (PT) (−0.31).
Figure 2. Pearson’s correlation between the 11 descriptors used to characterize 69 Capsicum acces-sions. Blue and red boxes indicate positive and negative correlation, respectively. The white boxes Figure 2. Pearson’s correlation between the 11 descriptors used to characterize 69 Capsicum accessions.Blue and red boxes indicate positive and negative correlation, respectively. The white boxes indicateno significant correlation (p < 0.05). In the right side of the correlogram, the legend color showsthe correlation coefficients and the corresponding colors. PH—plant height; CD—canopy diameter;SD—stem diameter; LL—leaf length; LW—leaf width; PL—petiole length; FL—fruit length; FD—fruitdiameter; FFM—fruit fresh mass; PT—pericarp thickness; FLD—flower diameter.
3.3. ISSR Characterization and Molecular Diversity
In the ISSR analysis, 17 out of 50 primers previously tested detected polymorphismsbetween the accessions. Altogether, 150 fragments were generated, of which 134 werepolymorphic, which indicated 90% polymorphism, considering fragments generated forall species. When analyzing the polymorphism at species level, C. chinense and C. baccatumwere the ones with the best grade of polymorphism, with 88.8% each. In C. frutescens, weobserved 56.6% of polymorphism, whereas in C. annuum, we observed the lowest degreeof polymorph (26.9%). The polymorphism information content (PIC) values varied from0.43 for primer ISSR 47 to 0.83 for ISSR 94. The mean value was 0.62.
The molecular characterization allowed for the formation of five groups, on the basisof the dendrogram (Figure 3b) with a cophenetic correlation coefficient of 0.76. GroupsI, II, III, and IV were formed, individually, by one accession each. Group V gatheredthe remaining 65 accessions. The commercial accession IFES 09 was grouped alone ingroup III and the other four (IFES 51, IFES 99, IFES 100, and IFES 101) in group V. Theentanglement value obtained comparing dendrograms was 0.59, evidencing the divergencein the distribution of the accessions in molecular and morpho-agronomic dendrograms. Asobserved in morpho-agronomic characterization, it can be noticed that on the basis of ISSRmarkers, the accessions were not grouped according to their geographic locations.

Agronomy 2021, 11, 854 9 of 15
Agronomy 2021, 11, x FOR PEER REVIEW 8 of 15
indicate no significant correlation (p < 0.05). In the right side of the correlogram, the legend color shows the correlation coefficients and the corresponding colors. PH—plant height; CD—canopy diameter; SD—stem diameter; LL—leaf length; LW—leaf width; PL—petiole length; FL—fruit length; FD—fruit diameter; FFM—fruit fresh mass; PT—pericarp thickness; FLD—flower diame-ter.
3.2. Diversity Among Capsicum Accessions and Correlated Traits To group accessions, we calculated dissimilarities on the basis of the squared Euclid-
ean distances. The dendrogram obtained in the grouping of accessions allowed for the formation of five distinct groups (Figure 3a), and the cophenetic correlation coefficient was 0.82. Groups I and II allocated only one accession each. The accession allocated in group I presented the highest values for plant height, canopy diameter, and stem diame-ter, having elongated fruits of orange color in the intermediate stage and red when ripe.
The accession allocated in group II presented the lowest values for the traits of plant height, canopy diameter, stem diameter, and leaf length and width. Its triangular fruits were yellow in the intermediate stage and red when ripe. Group III gathered nine acces-sions, with the highest values for leaf length and width. In addition, they had an interme-diate growth habit; filament colored in white—except for the IFES 02 accession, which had yellow filament—pigmented calyx; semi-wrinkled fruit surface; and absence of capsaicin, except for the IFES 67 and IFES 91 accessions, which showed pungent fruits.
Group IV was formed by four accessions in which the erect growth habit descriptors predominated as well as the position of the flowers. The number of flowers per node in the group’s accessions was two or three and violet in the anthers and greenish yellow corolla without the presence of spots. The filament of these accessions was yellow in color and the calyx had the presence of pigment. Its pointed fruits had a long shape with a semi-wrinkled surface and an acute base. In addition, its fruits were slightly corrugated and with the presence of capsaicin. Group V gathered the remaining 54 accessions, which, in general, had smaller plants; short petioles; and smaller and thinner fruits and with less pericarp thickness and, consequently, with less fresh mass. The growth habit of plants was intermediate, and its fruits had capsaicin.
Jaccard Index Average Euclidean Distance
Entanglement = 0.59
Figure 3. UPGMA dendrograms based on (a) morpho-agronomic traits and (b) genetic diversity of 69 pepper accessions.
4. Discussion
Studies related to genetic resources—such as the collection, characterization, andconservation of germplasm—of Capsicum are of great relevance for the preservation ofthe genetic diversity of the genus and for its use in breeding programs [37,38,54–57]. Theacquisition and maintenance of germplasm banks are strategies that allow us to explore andexpand the genetic variability of the acquired accessions and are fundamental to mitigatethe effects of the loss of genetic variability caused by the exploitation of domesticatedspecies [5]. Therefore, it is only possible to develop more productive varieties, resistant todiseases and reduce the impacts of genetic erosion when there is prior knowledge of theexisting variability [5,58]. For these, this study aimed to characterize 69 Capsicum accessionsfrom different regions of Brazil on the basis of qualitative and quantitative descriptors andISSR markers.
Among the qualitative descriptors studied, three were monomorphic (stem shape,stem pubescence, and calyx pigmentation), i.e., none of them were effective in discrimi-nating the studied accessions, and thus they can be eliminated in future studies, reducingthe time consumed for the evaluation of these characteristics. Carvalho et al. [59], whenstudying the diversity between accessions of Capsicum frutescens, also listed these traitsas monomorphic and inefficient in discriminating the accessions studied. However, ourresults report a great phenotypic variation for those descriptors related to flowers, andaccording to Baral and Bosland [60], to differentiate between the species of Capsicum, thedescriptors related to inflorescence are essential, such as the position of the flower and thepresence of annular constriction of the calyx, which are used to distinguish the accessionsof C. frutescens and C. chinense.
Wide variation between accessions was also detected for quantitative descriptors,revealing a great diversity in characteristics related to the plant and fruits. In Brazil,peppers are widely accepted, and their use is quite diversified. Peppers are eaten fresh,

Agronomy 2021, 11, 854 10 of 15
used in the preparation of sauces and condiments. In this sense, the consumer market hasacceptance for the different sizes, colors, and pungency levels of the Capsicum fruits.
For the fresh mass, the values varied widely—between 0.38 (IFES 100) and 30.94 g(IFES 33). In general, the fruits of the accessions of C. baccatum var. pendulum presented thehighest values for the characteristic, a common fact for this species that includes the sweetpepper variety with fruits appreciated for in natura consumption. Accessions belonging tothe species C. frutescens were the ones that presented the lowest values for this characteristic.This species includes the ‘chilli-pepper’ morphotype, in Brazil ‘pimenta malagueta’, which,due to its small size and high pungency, is widely used in the production of sauces andcanned peppers. The attributes of length and diameter of the fruit also showed a highvariation, with values between 1.07 and 12.79 cm, and 3.95 and 49.50 mm, respectively.Similar results for fruit length and diameter have been reported by Cardoso et al. [17] who,studying the diversity between 116 Capsicum accessions, found values between 0.93 and13.64 cm, and 4 and 59 mm, respectively. According to Leite et al. [61], longer fruits arevery important for genetic improvement programs of Capsicum, mainly when they aredestined for the food industry, as in the production of paprika or for use in cosmetics andpharmaceuticals.
Regarding pericarp thickness, the accessions showed values between 0.35 and 4.33 mm.According to Abud et al. [62], accessions with thicker pericarp are more important forthe fresh consumer market, since thicker pericarps gives greater resistance to fruits inpost-harvest treatments and guarantees an appearance more appreciated by consumers,in addition to conferring a higher degree of resistance to pathogens and parasites [22].In general, significant differences were found for all quantitative morpho-agronomiccharacteristics, an indicating the presence of great genetic variation between accessions(Table S6).
By estimating the relative contribution of the descriptors to the variability betweenaccessions on the basis of the method of Singh [38], we noticed that characteristics inherentto the fruit are the ones that most contribute to study the degree of diversity among the69 accessions. The fruit length (60.72%) and fruit diameter (12.71%) stood out. These resultscorroborate previous studies, which indicated that characteristics associated with fruits hada great discriminatory power [63]. A possible explanation for the greater contribution of thecharacteristics related to the fruit to the diversity in the different studies of Capsicum wasbecause the fruit is the portion consumed by humans and also by birds, being responsiblefor the dissemination of Capsicum seeds. This leads to the hypothesis that selection anddomestication were based on this plant organ, which has a diverse range of colors, shapes,sizes, degree of pungency, and aromas [64].
When it comes to Capsicum improvement, depending on the characteristics to beimproved, the use of traits correlated with the trait of interest facilitates the process ofindirect gain in the selection. In this perspective, Pearson’s correlation coefficient was usedin this study. It is a measure of linear association between two quantitative variables, inwhich one characteristic can be selected on the basis of another.
The results (Figure 2) showed that concomitant gains in fresh fruit mass (FFM) andpericarp thickness (PT) were possible when selecting genotypes on the basis of fruitdiameter (FD), since there is a positive correlation between fruit diameter and the first twotraits (FFM and PT). Studying 40 accessions of C. baccatum, Rêgo et al. [63] also found apositive correlation between pericarp thickness and fresh fruit mass, which corroboratedthe results also reported by Leite et al. [61] and, more recently, Bianchi et al. [22]. Studies ofthe association between easy-to-measure characteristics and those related to productivityand quality are crucial for breeding programs, since they contribute to the selection ofsuperior plants with little time consumption [58,60,65,66].
The associations between the characteristics related to the plant’s architecture alsohelp in improving Capsicum for ornamental purposes. A major factor for the use ofpepper trees in the floriculture market is the plant’s growth capacity in pots [67]. For this,characteristics such as short stature and leaf density are essential. Our results show a strong

Agronomy 2021, 11, 854 11 of 15
positive correlation between leaf length and width (0.80), between plant height and canopydiameter (0.77) and stem diameter (0.72); therefore, the selection of smaller plants or withless leaf length/width would result in reductions in the other aspects related to the goodarchitecture of the plant in the pots.
The description of the diversity between the accessions based on the morpho-agronomicdescriptors allowed for the formation of five groups. It is possible to infer that the dis-similarity matrix revealed data adjusted to the dendrograms. Such adjustment can beconfirmed by the high value of the cophenetic correlation coefficient, which according toRohlf and Fisher [68] must present values above 0.8. Thus, the results were satisfactoryand revealed the conformity between the graphic representations of the distances andtheir original matrices. A good adjustment between the graphical representation of thedistances and their original matrix makes it possible to make inferences through visualassessment [67].
Accessions could not be grouped on the basis of their geographic origin since geno-types from the same location were gathered in different groups. Other studies also reportedthe lack of correlation between geographic location and accession grouping [16,17,58,69].As in this study, Finger et al. [70], when evaluating the diversity between 49 accessions ofC. chinense of Brazil, did not show a relationship between the geographic distance and theestimated diversity. The authors reported that this may have been a reflection of geneticdrift and the selection of plants in different environments, rather than geographic location.Another possibility was the occurrence of cross-pollination at a rate that varies in Capsicumfrom 0.5 to 70%, contributing to the genetic contamination of the seeds.
Characterization through molecular markers is an effective strategy when associatedwith phenotyping studies based on multi-categorical descriptors. However, when it comesto SSR primers, these DNA markers are species-specific, and in the case of our work, fourpepper species of Capsicum (Capsicum annuum, Capsicum baccatum var. pendulum, Capsicumchinense Jacq., and Capsicum frutescens) were used and only for the species Capsicum annuumwe have a good number of SSR primers already described. In this sense, countless tests oftransferability of SSR primers to other species are necessary, which requires a lot of time,manpower, and financial resources. We reiterate that for the species Capsicum baccatum var.pendulum, we do not have any SSR primers described in the literature yet. In many studies,ISSR primers have been used to study genetic diversity [71–73], being considered quiteefficient to discriminate diversity in germplasm of unique or many species. The efficacyof the markers quantified by PIC values showed that the ISSR markers were effective inassessing the genetic relatedness in the accessions. Generally, the PIC values were greaterthan 0.5, a value considered satisfactory according to Mandal [71].
Our results show that the molecular markers indicated the presence of 90% of polymor-phism considering all accessions studied. Thul et al. [24], in a molecular characterizationof six Capsicum species with ISSR markers, found 98.7% of polymorphism. In contrast,Olatunji et al. [35], studying the diversity among four Capsicum species with ISSR mark-ers, found 14 polymorphic loci out of the 75 amplified, which represented only 18.6% ofpolymorphism. These results show the efficiency of the markers used in our work to detectdiversity in the germplasm collection evaluated. When analyzing each species, we foundthe fragments generated by ISSR markers in C. chinense and C. baccatum showed a greatlevel of polymorphism. More recently, Bianchi et al. [22], studying 55 accessions of C.chinense, found 47% polymorphism for the 97 fragments produced by the ISSR markersused in their study with C. chinense accessions.
The correlation between the groups formed by the morphological and molecularcharacterization was considered low, with an agreement rate of only 41. Results like thisare very common in studies of Capsicum diversity when molecular markers and phenotypicdescription of germplasm are associated [22,66,74]. One possible reason to this is the factthat the markers used in this study may be not associated with the evaluated characteristicsand because they also amplify coding and non-coding regions of the genome. Since thephenotype is the result of coding regions, there is a low correlation with the bands generated

Agronomy 2021, 11, 854 12 of 15
by the molecular markers. Similar to phenotypic characterization, ISSR markers did notgroup accessions according to their geographic location. The absence of an associationbetween geographic origin and molecular markers is probably caused by the exchange ofseeds between producers in nearby regions as well as unrestricted transport of fruits andseeds between different regions of Brazil [16,17].
However, even though the morpho-agronomic and molecular characterization didnot allow us to separate the accessions by region and both did not present similarity inthe formed clusters, we assumed the importance of these two methods to discriminatethe diversity of a germplasm collection, guaranteeing access to the wide variability of thegenetic resources to plant breeding programs.
5. Conclusions
A high level of variation was detected for the qualitative and quantitative descriptors.The characteristics associated with the fruit were those that showed greater relevance inthe discrimination of accessions and correlation among some important traits related tofruits, and plant architecture is an important tool to facilitate the next studies for pepperbreeding for many branches of the pepper market.
The morpho-agronomic and molecular characterization were efficient to estimate thegenetic diversity between accessions, showing significant divergence, being importanttools for the knowledge and use of accessions in breeding programs.
This study is going to allow for the establishment of a collection of great importanceto preserve pepper germplasm and to provide morpho-agronomical data to facilitate theprocess of the selection in the breeding programs. The shown characterization would beuseful for a possible next genome-wide association study.
Supplementary Materials: The following are available online at https://www.mdpi.com/article/10.3390/agronomy11050854/s1, Figure S1: Representative sample of 2% agarose gel. DNA amplifiedfragments using ISSR marker 106. M—molecular weight marker. Table S1: Accession identification of69 Capsicum accessions from Instituto Federal do Espírito Santo—Campus de Alegre germplasm collection.Table S2: Twenty-two qualitative descriptors used to phenotype 69 Capsicum accessions according toIPGRI (1995). Table S3: Eleven quantitative descriptors used to phenotype 69 Capsicum accessionsaccording to IPGRI (1995). Table S4: Information about 17 ISSR primers used in the genotyping of69 Capsicum accessions from Instituto Federal do Espírito Santo—Campus de Alegre germplasm collectionand 33 discarded primers. Table S5: Descriptive analysis of 22 qualitative descriptor of 69 Capsicumpopulations from Instituto Federal do Espírito Santo—Campus de Alegre germplasm collection. Table S6:Grouping of 11 quantitative means of 69 capsicum accessions by Scott-Knott method at 5% probability.
Author Contributions: Conceptualization, B.D.G.B., A.C.S.J. and M.M.M.; methodology, B.D.G.B.,T.d.O.S., P.H.A.D.S., S.H.K., J.D.S.N., L.H.R., F.V.V., R.N.d.A., R.R., A.C.S.J. and M.M.M.; software,B.D.G.B., T.d.O.S., P.H.A.D.S., S.H.K., J.D.S.N. and M.M.M.; validation, B.D.G.B., T.d.O.S., P.H.A.D.S.,S.H.K., J.D.S.N. and M.M.M.; formal analysis, B.D.G.B., T.d.O.S., P.H.A.D.S., S.H.K., J.D.S.N., L.H.R.,F.V.V., R.N.d.A. and M.M.M.; investigation, B.D.G.B., R.R. and M.M.M.; resources, B.D.G.B., R.R. andM.M.M.; data curation, B.D.G.B., R.R. and M.M.M.; writing—original draft preparation, B.D.G.B.,T.d.O.S., P.H.A.D.S., R.R., A.C.S.J. and M.M.M.; writing—review and editing, B.D.G.B., T.d.O.S.,P.H.A.D.S., S.H.K., R.R., A.C.S.J. and M.M.M.; visualization, B.D.G.B., R.R. and M.M.M.; supervision,B.D.G.B., A.C.S.J. and M.M.M.; funding acquisition, B.D.G.B. and M.M.M. All authors have read andagreed to the published version of the manuscript.
Funding: This research was financed in part by Instituto Federal de Educação, Ciência e Tecnologiado Espírito Santo—Campus de Alegre and Coordenação de Aperfeiçoamento de Pessoal de NívelSuperior (CAPES/Brasil) with the scholarship of the first author—finance code 001.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: Not applicable.
Conflicts of Interest: The authors declare no conflict of interest.

Agronomy 2021, 11, 854 13 of 15
References1. Barboza, G.E.; Carrizo García, C.; Leiva González, S.; Scaldaferro, M.; Reyes, X. Four new species of Capsicum (Solanaceae) from
the tropical Andes and an update on the phylogeny of the genus. PLoS ONE 2019, 14, e0209792. [CrossRef] [PubMed]2. Van Zonneveld, M.; Ramirez, M.; Williams, D.E.; Petz, M.; Meckelmann, S.; Avila, T.; Bejarano, C.; Ríos, L.; Peña, K.; Jäger, M.; et al.
Screening Genetic Resources of Capsicum Peppers in Their Primary Center of Diversity in Bolivia and Peru. PLoS ONE 2015, 10,e0134663. [CrossRef] [PubMed]
3. Scossa, F.; Roda, F.; Tohge, T.; Georgiev, M.I.; Fernie, A.R. The Hot and the Colorful: Understanding the Metabolism, Geneticsand Evolution of Consumer Preferred Metabolic Traits in Pepper and Related Species. CRC. Crit. Rev. Plant Sci. 2019, 38,339–381. [CrossRef]
4. Food and Agriculture Organization of the United Nations. FAOSTAT—Food and Agriculture Database. Available online:http://www.fao.org/faostat (accessed on 19 March 2021).
5. Neitzke, R.S.; Barbieri, R.L.; Vasconcelos, C.S.; Priori, D.; Fischer, S.Z.; Götzke, M.L.D.; Romano, C.M.; Heiden, G. Diversity inCapsicum Landraces Cultivated in Brazil. Acta Hortic. 2011, 531–536. [CrossRef]
6. Da Silva Neto, J.J.; do Rêgo, E.R.; Nascimento, M.F.; Silva Filho, V.A.L.; de Almeida Neto, J.X.; do Rêgo, M.M. Variabilidade empopulação base de pimenteiras ornamentais (Capsicum annuum L.). Rev. Ceres 2014, 61, 84–89. [CrossRef]
7. Fayos, O.; de Aguiar, A.; Jiménez-Cantizano, A.; Ferreiro-González, M.; Garcés-Claver, A.; Martínez, J.; Mallor, C.; Ruiz-Rodríguez,A.; Palma, M.; Barroso, C.; et al. Ontogenetic Variation of Individual and Total Capsaicinoids in Malagueta Peppers (Capsicumfrutescens) during Fruit Maturation. Molecules 2017, 22, 736. [CrossRef]
8. Rodrigues, R.; Gonçalves, L.S.; dos Bento, C.S.; Sudré, C.P.; Robaina, R.R.; do Amaral Júnior, A.T. Combining ability and heterosisfor agronomic traits in chili pepper. Hortic. Bras. 2012, 30, 226–233. [CrossRef]
9. De Santos, T.O.; Moulin, M.M.; Rangel, L.H.; Pirovani, R.O.L.; Valadares, F.V.; de Almeida, R.N.; Silva, L.O.E. Characterizationand Diversity of Peppers (Capsicum spp.) Genotypes Based on Morphological Traits Using Multivariate Analysis. J. Exp. Agric.Int. 2019, 1–10. [CrossRef]
10. Grafton, R.Q.; Daugbjerg, C.; Qureshi, M.E. Towards food security by 2050. Food Secur. 2015, 7, 179–183. [CrossRef]11. Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff,
B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [CrossRef]12. Pereira-Dias, L.; Gil-Villar, D.; Castell-Zeising, V.; Quiñones, A.; Calatayud, Á.; Rodríguez-Burruezo, A.; Fita, A. Main Root
Adaptations in Pepper Germplasm (Capsicum spp.) to Phosphorus Low-Input Conditions. Agronomy 2020, 10, 637. [CrossRef]13. United Nations Transforming Our World: The 2030 Agenda for Sustainable Development; ONU: New York, NY, USA, 2015.14. Moulin, M.M.; Rodrigues, R.; Ramos, H.C.C.; Bento, C.S.; Sudré, C.P.; Gonçalves, L.S.A.; Viana, A.P. Construction of an integrated
genetic map for Capsicum baccatum L. Genet. Mol. Res. 2015, 14, 6683–6694. [CrossRef]15. Moulin, M.M.; Rodrigues, R.; Bento, C.S.; Gonçalves, L.S.A.; Santos, J.O.; Sudré, C.P.; Viana, A.P. Genetic dissection of agronomic
traits in Capsicum baccatum var. pendulum. Genet. Mol. Res. 2015, 14, 2122–2132. [CrossRef]16. Baba, V.Y.; Rocha, K.R.; Gomes, G.P.; de Fátima Ruas, C.; Ruas, P.M.; Rodrigues, R.; Gonçalves, L.S.A. Genetic diversity of
Capsicum chinense accessions based on fruit morphological characterization and AFLP markers. Genet. Resour. Crop Evol. 2016, 63,1371–1381. [CrossRef]
17. Cardoso, R.; Ruas, C.F.; Giacomin, R.M.; Ruas, P.M.; Ruas, E.A.; Barbieri, R.L.; Rodrigues, R.; Gonçalves, L.S.A. Genetic variabilityin Brazilian Capsicum baccatum germplasm collection assessed by morphological fruit traits and AFLP markers. PLoS ONE 2018,13, e0196468. [CrossRef]
18. Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.;Wong, Z.-Z.; et al. Vitamin Variation in Capsicum Spp. Provides Opportunities to Improve Nutritional Value of Human Diets.PLoS ONE 2016, 11, e0161464. [CrossRef] [PubMed]
19. Deka, S.D. Taxonomic diversity of cultivated Capsicum genotypes of India. Electron. J. Plant Breed. 2017, 8, 660. [CrossRef]20. dos Pessoa, A.M.S.; do Rêgo, E.R.; dos Santos, C.A.P.; de Carvalho, M.G.; de Mesquita, J.C.P.; do Rêgo, M.M. Inheritance of flower
traits in ornamental pepper. Agropecuária Técnica 2018, 39, 50. [CrossRef]21. Sharmin, A.; Hoque, M.E.; Haque, M.M.; Khatun, F. Molecular Diversity Analysis of Some Chilli (Capsicum spp.) Genotypes
Using SSR Markers. Am. J. Plant Sci. 2018, 9, 368–379. [CrossRef]22. Bianchi, P.A.; Silva, L.R.A.; Alencar, A.A.S.; Santos, P.H.A.D.; Pimenta, S.; Sudré, C.P.; Corte, L.E.D.; Gonçalves, L.S.A.; Rodrigues,
R. Biomorphological Characterization of Brazilian Capsicum Chinense Jacq. Germplasm. Agronomy 2020, 10, 447. [CrossRef]23. Costa, F.R.; Pereira, T.N.S.; Vitória, A.P.; Campos, K.P.; Rodrigues, R.; Silva, D.H.; Pereira, M.G. Genetic diversity among Capsicum
accessions using RAPD markers. Crop. Breed. Appl. Biotechnol. 2006, 6, 18–23. [CrossRef]24. Thul, S.T.; Darokar, M.P.; Shasany, A.K.; Khanuja, S.P.S. Molecular Profiling for Genetic Variability in Capsicum Species Based on
ISSR and RAPD Markers. Mol. Biotechnol. 2012, 51, 137–147. [CrossRef]25. Soares de Souza, C.; Schuelter, A.R.; Finger, F.L.; Dias Casali, V.W. Variabilidade genética em acessos de Capsicum chinense por
meio de marcadores isoenzimáticos e rapd. Sci. Agrar. 2013, 14. [CrossRef]26. Thul, S.T.; Shasany, A.K.; Darokar, M.P.; Khanuja, S.P.S. AFLP Analysis for Genetic Diversity in Capsicum Annuum and Related
Species. Nat. Prod. Commun. 2006, 1, 1934578X0600100309. [CrossRef]27. Islam, M.A.; Sinha, P.; Sharma, S.S.; Negi, M.S.; Neog, B.; Tripathi, S.B. Analysis of genetic diversity and population structure in
Capsicum landraces from North Eastern India using TE-AFLP markers. Plant Mol. Biol. Report. 2016, 34, 869–875. [CrossRef]

Agronomy 2021, 11, 854 14 of 15
28. Moreira, A.F.P.; Ruas, P.M.; de Ruas, C.F.; Baba, V.Y.; Giordani, W.; Arruda, I.M.; Rodrigues, R.; Gonçalves, L.S.A. Genetic diversity,population structure and genetic parameters of fruit traits in Capsicum chinense. Sci. Hortic. 2018, 236, 1–9. [CrossRef]
29. Shirasawa, K.; Ishii, K.; Kim, C.; Ban, T.; Suzuki, M.; Ito, T.; Muranaka, T.; Kobayashi, M.; Nagata, N.; Isobe, S.; et al. Developmentof Capsicum EST–SSR markers for species identification and in silico mapping onto the tomato genome sequence. Mol. Breed.2013, 31, 101–110. [CrossRef] [PubMed]
30. Meng, C.; Wei, X.; Zhao, Y.; Yuan, Y.; Yang, S.; Wang, Z.; Zhang, X.; Sun, J.; Zheng, X.; Yao, Q.; et al. Genetic Diversity Analysis ofCapsicum Genus by SSR Markers. Mol. Plant Breed. 2017. [CrossRef]
31. Azevedo, C.D.O.; Rodrigues, R.; Sudré, C.P. Microsatellites for detecting inconsistencies in Capsicum cultivars registration inBrazilian database: More than meets the eye. Hortic. Bras. 2019, 37, 285–293. [CrossRef]
32. Guzmán, F.A.; Moore, S.; de Vicente, M.C.; Jahn, M.M. Microsatellites to enhance characterization, conservation and breedingvalue of Capsicum germplasm. Genet. Resour. Crop Evol. 2020, 67, 569–585. [CrossRef]
33. Rodrigues, R.; da Costa Batista, F.R.; Moulin, M.M. Molecular Markers in Capsicum spp. Breeding. In Production and Breeding ofChilli Peppers (Capsicum spp.); Springer International Publishing: Cham, Switzerland, 2016; pp. 81–95. [CrossRef]
34. Ou, L.; Zou, X. Inter simple sequence repeat analysis of genetic diversity of five cultivated pepper species. Afr. J. Biotechnol. 2012,11, 752–757. [CrossRef]
35. Olantunji, T.L.; Afolayan, A.J. Contributions to the Classification of Capsicum annuum L. and Capsicum frutescens L. in WestAfrica Using Morphological Traits. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 47, 135–142. [CrossRef]
36. Rana, M.; Sharma, R.; Sharma, P.; Bhardwaj, S.V.; Sharma, M. Estimation of Genetic Diversity in Capsicum annuum L. GermplasmUsing PCR-Based Molecular Markers. Natl. Acad. Sci. Lett. 2014, 37, 295–301. [CrossRef]
37. D’Agostino, N.; Tamburino, R.; Cantarella, C.; De Carluccio, V.; Sannino, L.; Cozzolino, S.; Cardi, T.; Scotti, N. The CompletePlastome Sequences of Eleven Capsicum Genotypes: Insights into DNA Variation and Molecular Evolution. Genes 2018, 9, 503.[CrossRef] [PubMed]
38. Colonna, V.; D’Agostino, N.; Garrison, E.; Albrechtsen, A.; Meisner, J.; Facchiano, A.; Cardi, T.; Tripodi, P. Genomic diver-sity and novel genome-wide association with fruit morphology in Capsicum, from 746k polymorphic sites. Sci. Rep. 2019,9, 10067. [CrossRef]
39. Olatunji, T.L.; Afolayan, A.J. Evaluation of genetic relationship among varieties of Capsicum annuum L. and Capsicum frutescensL. in West Africa using ISSR markers. Heliyon 2019, 5, e01700. [CrossRef] [PubMed]
40. Moscone, E.A.; Scaldaferro, M.A.; Grabiele, M.; Cecchini, N.M.; Sánchez García, Y.; Jarret, R.; Daviña, J.R.; Ducasse, D.A.;Barboza, G.E.; Ehrendorfer, F. The evolution of chili peppers (Capsicum—Solanaceae): A cytogenetic perspective. Acta Hortic.2007, 137–170. [CrossRef]
41. IPGRI. Descriptors for Capsicum (Capsicum spp.); International Plant Genetic Resources Institute: Rome, Italy, 1995,ISBN 978-92-9043-216-6.
42. Derera, F. Condiment Paprika: Breeding, Harvesting and Commercialisation: A Report for the Rural Industries Research and DevelopmentCorporation; RIRDC: New South Wales, Australia, 2000; p. 33.
43. Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1987, 12, 39–40.44. Daher, R.F.; Pereira, M.G.; Tupinambá, E.A.; Amaral Junior, A.T.; Aragão, W.M.; Ribeiro, F.E.; Oliveira, L.O.; Sakiyama, N.S.
Assessment of coconut tree genetic divergence by compound sample RAPD marker analysis. Crop. Breed. Appl. Biotechnol. 2002, 2,431–438. [CrossRef]
45. Souza, D.C.L.; Gomes, L.J.; Blank, A.F.; Gois, I.B.; Pereira, G.S.; Alves, P.B.; Silva-Mann, R. Characterization of wild genotypes ofAroeira: Subsidy for plant breeding. J. Agric. Biotechnol. Sustain. Dev. 2014, 6, 39–49. [CrossRef]
46. jiang, Y.; Liu, J.-P. Analysis of genetic diversity of Piper spp. in Hainan Island (China) using inter-simple sequence repeat ISSRmarkers. AFRICAN J. Biotechnol. 2011, 10, 16763–16768. [CrossRef]
47. Yao, H.; Zhao, Y.; Chen, D.F.; Chen, J.K.; Zhou, T.S. ISSR primer screening and preliminary evaluation of genetic diversity in wildpopulations of Gycyrrhiza uralensis. Biol. Plant. 2008, 52, 117–120. [CrossRef]
48. Refaat, M.H.; Elgarhy, H.A.S. Relationship between hybrid performance and genetic diversity based on ISSR-PCR markers inPepper (Capsicum annuum. L.). Ann. Agric. Sci. 2007, 45, 1565–1579.
49. Singh, D. The relative importance of characters affecting genetic divergence. Indian J. Genet. Plant Breed. 1981, 41, 237–245.50. R Core Team. R: A Language and ENVIRONMENT for Statistical Computing. R Foundation for Statistical Computing: Vienna,
Austria, 2021. Available online: http://www.R-project.org/ (accessed on 26 April 2021).51. Cruz, C.D..; Regazzi, I.A.J.; Carneiro, P.C.S. Modelos Biométricos Aplicados ao Melhoramento Genético; Editora UFV: Viçosa, MG,
Brasil, 2012; Volume 1.52. Cruz, C.D. GENES—A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013,
35, 271–276. [CrossRef]53. Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015,
31, 3718–3720. [CrossRef]54. Lima, M.F.; Carvalho, S.I.C.; Ragassi, C.F.; Bianchetti, L.B.; Faleiro, F.G.; Reifschneider, F.J.B. Characterization of a pepper
collection (Capsicum frutescens L.) from Brazil. Genet. Mol. Res. 2017, 16. [CrossRef]55. Martinez, A.L.A.; Araújo, J.S.P.; Ragassi, C.F.; Buso, G.S.C.; Reifschneider, F.J.B. Variability among Capsicum baccatum accessions
from Goiás, Brazil, assessed by morphological traits and molecular markers. Genet. Mol. Res. 2017, 16. [CrossRef]

Agronomy 2021, 11, 854 15 of 15
56. Nankar, A.N.; Todorova, V.; Tringovska, I.; Pasev, G.; Radeva-Ivanova, V.; Ivanova, V.; Kostova, D. A step towards BalkanCapsicum annuum L. core collection: Phenotypic and biochemical characterization of 180 accessions for agronomic, fruit quality,and virus resistance traits. PLoS ONE 2020, 15, e0237741. [CrossRef]
57. Bianchi, P.A.; Dutra, I.P.; Moulin, M.M.; Santos, J.O.; Santos Júnior, A.C. Morphological characterization and analysis of geneticvariability among pepper accessions. Ciência Rural 2016, 46, 7. [CrossRef]
58. González-López, J.; Rodríguez-Moar, S.; Silvar, C. Correlation Analysis of High-Throughput Fruit Phenomics and BiochemicalProfiles in Native Peppers (Capsicum spp.) from the Primary Center of Diversification. Agronomy 2021, 11, 262. [CrossRef]
59. Carvalho, S.I.C.; Bianchetti, L.B.; Ragassi, C.F.; Ribeiro, C.S.C.; Reifschneider, F.J.B.; Buso, G.S.C.; Faleiro, F.G. Genetic variabilityof a Brazilian Capsicum frutescens germplasm collection using morphological characteristics and SSR markers. Genet. Mol. Res.2017, 16. [CrossRef] [PubMed]
60. Baral, J.B.; Bosland, P.W. Unraveling the Species Dilemma in Capsicum frutescens and C. chinense (Solanaceae): A Multiple EvidenceApproach Using Morphology, Molecular Analysis, and Sexual Compatibility. J. Am. Soc. Hortic. Sci. 2004, 129, 826–832. [CrossRef]
61. Leite, W.S.; Pavan, B.E.; Alcântara Neto, F.; Matos Filho, C.H.A.; Feitosa, F.S.; Oliveira, C.B. Multivariate Exploratory Approachand Influence of Six Agronomic Traits on Soybean Genotypes Selection. Nativa 2016, 4, 206–210. [CrossRef]
62. Abud, H.F.; Araujo, R.F.; Pinto, C.M.F.; Araujo, E.F.; Araujo, A.V.; Santos, J.A. Caraterização morfométrica dos frutos de pimentasmalagueta e biquinho. Rev. Bras. Agropecuária Sustentável 2018, 8. [CrossRef]
63. Rêgo, M.M.; Sapucay, M.J.L.C.; Rêgo, E.R.; Araújo, E.R. Analysis of divergence and correlation of quantitative traits in ornamentalpepper (Capsicum SPP.). Acta Hortic. 2015, 389–394. [CrossRef]
64. Paran, I.; van der Knaap, E. Genetic and molecular regulation of fruit and plant domestication traits in tomato and pepper. J. Exp.Bot. 2007, 58, 3841–3852. [CrossRef]
65. Naegele, R.P.; Mitchell, J.; Hausbeck, M.K. Genetic Diversity, Population Structure, and Heritability of Fruit Traits in Capsicumannuum. PLoS ONE 2016, 11, e0156969. [CrossRef] [PubMed]
66. Leite, P.S.S.; Rodrigues, R.; Silva, R.N.O.; Pimenta, S.; Medeiros, A.M.; Bento, C.S.; Gonçalves, L.S.A. Molecular and agronomicanalysis of intraspecific variability in Capsicum baccatum var. pendulum accessions. Genet. Mol. Res. 2016, 15. [CrossRef]
67. Fortunato, F.L.G.; Do Rêgo, E.R.; De Carvalho, M.G.; Dos Santos, C.A.P.; Do Rêgo, M.M. Genetic diversity in ornamental pepperplants. Comun. Sci. 2019, 10, 364–375. [CrossRef]
68. Rohlf, F.J.; Fisher, D.R. Tests for Hierarchical Structure in Random Data Sets. Syst. Zool. 1968, 17, 407. [CrossRef]69. Sokal, R.R.; Rohlf, F.J. The comparison of dendrograms by objective methods. Taxon 1962, 11, 33–40. [CrossRef]70. Finger, F.L.; Lannes, S.D.; Schuelter, A.R.; Doege, J.; Comerlatto, A.P.; Gonçalves, L.S.A.; Ferreira, F.R.A.; Clovis, L.R.; Scapim, C.A.
Genetic diversity of Capsicum chinensis (Solanaceae) accessions based on molecular markers and morphological and agronomictraits. Genet. Mol. Res. 2010, 9, 1852–1864. [CrossRef] [PubMed]
71. Mandal, A.; Datta, A.K.; Datta, S.; Gupta, S. Genetic assessment of eight Corchorus spp. (Tiliaceae) using RAPD and ISSR markers.Nucleus 2013, 56, 23–30. [CrossRef]
72. Jia, X.-D.; Wang, T.; Zhai, M.; Li, Y.-R.; Guo, Z.-R. Genetic Diversity and Identification of Chinese-Grown Pecan Using ISSR andSSR Markers. Molecules 2011, 16, 10078–10092. [CrossRef]
73. Sunar, S.; Yildirim, N.; Sengul, M.; Agar, G. Genetic diversity and relationships detected by ISSR and RAPD analysis amongAethionema species growing in Eastern Anatolia (Turkey). C. R. Biol. 2016, 339, 147–151. [CrossRef] [PubMed]
74. Da Costa, F.R.; Pereira, T.N.S.; Sudré, C.P.; Rodrigues, R. Marcadores RAPD e caracteres morfoagronômicos na determinação dadiversidade genética entre acessos de pimentas e pimentões. Ciência Rural 2008, 39, 696–704. [CrossRef]


















