Placa de marfil grabada de la estructura 10.049 del sector PP4 ... · de grandes construcciones...
30
Placa de marfil grabada de la estructura 10.049 del sector PP4-Montelirio de la Zona Arqueológica de Valencina de la Concepción-Castilleja de Guzmán (Sevilla). Foto: Miguel Ángel Blanco de la Rubia
Transcript of Placa de marfil grabada de la estructura 10.049 del sector PP4 ... · de grandes construcciones...

Placa de marfil grabada de la estructura 10.049 del sector PP4-Montelirio de la Zona Arqueológica de Valencina de la Concepción-Castilleja de Guzmán (Sevilla). Foto: Miguel Ángel Blanco de la Rubia

69MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN (SEVILLA). ECONOMÍA Y SIMBOLISMO DE LOS ANIMALES EN UNA COMUNIDAD DEL III MILENIOCorina Liesau von Lettow-Vorbeck1,3, María Teresa Aparicio Alonso2, Rafael Araujo Armero2, Laura Llorente Rodríguez1
Arturo Morales Muñiz1
DO
SSIE
R
THE FAUNA FROM THE PP4-MONTELIRIO SECTOR OF THE PREHISTORIC SITE OF VALENCINA DE LA CONCEPCIÓN (SEVILLE). ANIMAL ECONOMY AND SYMBOLISM ON A THIRD MILLENNIUM BC COMMUNITY FROM THE IBERIAN PENINSULA
1 Laboratorio de Arqueozoología, Universidad Autónoma de Madrid. [[email protected]]; [[email protected]]2 Museo Nacional de Ciencias Naturales. [[email protected]]; [[email protected]]3 Departamento de Prehistoria y Arqueología, Universidad Autónoma de Madrid. [[email protected]]
Recibido: 12/07/2013; Aceptado: 17/10/2013
69
Resumen:
Se estudian las faunas recuperadas en el sector PP4-Montelirio del yacimiento calcolítico de Valencina de la Concepción, distinguiendo aquellas recuperadas en tumbas con restos humanos de los depósitos carentes de éstos. En las segundas se documenta un espectro de mamíferos similar al del registro funerario lo que sin embargo difiere en las especies de moluscos y en determinadas faunas como las astas de ciervo, parte de un bucráneo de un carnero o el tritón (Charonia lampas) que apuntan al carácter simbólico de los depó-sitos funerarios. Un tercio de las 61 estructuras funerarias presenta la valva cóncava de la viera (género Pecten) y, en menor medida, lapas de la especie Cymbula nigra. Ambas especies, que interpretamos como ajuares, se sitúan siempre próximas a algún inhumado. La presencia recurrente de vieras pudiera asociarse con la identidad de determinados individuos. La totalidad de las especies documentadas en este estudio, a excepción del uro (Bos primigenius), se encuentran actualmente en la zona.
Palabras clave: Calcolítico, fauna, simbolismo, Arqueozoología funeraria, Península Ibérica, Tafonomía.
Abstract:
This paper reviews the faunas retrieved at the PP4-Montelirio sector of the Chalcolithic site of Valencina de la Concepción, setting apart remains found in association with or without human bones. Although the structures of non-funerary character exhibit a similar mammalian profile to those of funerary nature, the molluscs and some items such as red deer antlers, a ram’s bucranium or the triton (Charonia lampas) seem to have a clear symbolic nature. A third of the 61 funerary structures incorporate the concave valve of sca-llops (Genus Pecten), and, less often, limpet shells of the species Cymbula nigra. These two taxa, considered as funerary offers, are always placed next to some of the inhumations. The prevalence of scallops in the tombs may constitute an identity sign for the members of this community. All the species documented in our study, with the sole exception of the aurochs (Bos primigenius), are still to be found in the region.
Keywords: Chalcolithic, Fauna, Symbolism, Funerary Archaeozoology, Iberian Peninsula, Taphonomy.

70
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
1. INTRODUCCIÓN
Con una extensión de c. 468 ha (Vargas Jiménez, 2004: 141) y una elevada densidad de estructuras que, según sugieren recientes prospecciones geofí-sicas (Wheatley et al., 2012; Vargas Jiménez et al., 2012), se contarían por millares, el asentamiento de Valencina de la Concepción es el de mayor exten-sión, complejidad y variedad de estructuras hasta la fecha conocido para el Calcolítico peninsular. Loca-lizado sobre la zona norte de la plataforma elevada de la comarca de El Aljarafe, en la margen derecha del río Guadalquivir y próximo a la actual ciudad de Sevilla, se ubica en un medio muy favorable para el asentamiento humano. La variada disponibilidad de los recursos naturales (llanuras de inundación, el estuario, la cobertura forestal al interior, su cerca-nía a recursos mineros) queda manifiesta a través de una intensa ocupación (probablemente conti-nuada) desde finales del IV hasta bien entrado el I milenio ANE. El Aljarafe en su conjunto está ocu-pado de forma ininterrumpida a lo largo de la Anti-güedad, el Medievo y los siglos más recientes hasta su incorporación en el área metropolitana de Sevilla a finales del siglo XX.
Valencina es conocido desde el descubrimiento del tholos de La Pastora en 1860. Ulteriores hallazgos de grandes construcciones megalíticas como los tholoi de Matarubilla o de Montelirio, además de los varios centenares de estructuras de todo tipo reve-ladas por las más de 120 excavaciones arqueológi-cas realizadas desde los años 80 del pasado siglo, han dado lugar a un voluminoso y complejo regis-tro arqueológico, todavía mal conocido. La reciente publicación de las actas de una reunión monográfica llevada a cabo en 2010 coincidiendo con el 150 ani-versario del descubrimiento de La Pastora (García Sanjuán et al., 2013a) ha permitido poner al día la investigación del yacimiento, reuniendo numero-sos trabajos en torno a la naturaleza del territorio, aspectos paleoambientales, estudios antropológi-cos, secuencias ocupacionales, prácticas funerarias y votivas, además de aportar algunas contribucio-nes, aunque de manera bastante puntual, al estudio de las estrategias subsistenciales y productivas de este gran asentamiento prehistórico.
Son cinco los informes arqueozoológicos publicados hasta la fecha para el asentamiento calcolítico de Valencina:
• Sectores Cerro de la Cabeza, La Candelera y La Perrera (Hain, 1982), incluyendo 38.300 restos óseos (NMI = 558) encontrados en las excavacio-nes llevadas a cabo en 1975 y 1976.
• Sector Plan Parcial Matarrubilla, a partir de una muestra mucho más pequeña de 789 restos (NMI = 37) (Abril López et al., 2010).
• Sector La Gallega, basado en 9.025 restos faunís-ticos (NMI = 86) (Bernáldez Sánchez et al., 2013).
• Sector c/ Mariana de Pineda s/n, que incluye un total de 2.216 restos (NMI = 82) (Pajuelo Pando y López Aldana, 2013).
• Sector c/ Andalucía, con una muestra muy pequeña (NMI = 16) (Sardá Piñeiro, 2013).
A estos estudios cabría añadir otros más específicos sobre materias primas animales concretas u objetos específicos. Es el caso, por ejemplo, de los varios trabajos que se han publicado muy recientemente en relación con el abastecimiento, transformación, uso funerario y simbología del marfil en la Valencina del III milenio (Vargas Jiménez et al., 2012; Nocete Calvo et al., 2013; García Sanjuán et al., 2013b; Schuhmacher et al., 2013; Luciañez Triviño et al., 2014). Se trata de un tema que ha despertado un inusitado interés y que está posibilitando una lectura completamente nueva de los contactos a larga distancia de la Valencina del III milenio ANE, así como de las formas de expresión material del prestigio y del estatus social. También pueden citarse algunas primeras aproximaciones a la manufactura de “ídolos” o figurillas en materiales óseos faunísticos (Hurtado Pérez, 2013) o a las figu-rillas zoomorfas moldeadas o talladas en distintas materias primas animales (Valera et al., 2014).
Los resultados de estos estudios apuntan a que los animales tuvieron un papel capital, tanto en el plano económico como en el simbólico, en las sociedades que habitaron o frecuentaron el asen-tamiento Valencina durante el III milenio ANE (Gar-cía Sanjuán, 2013: 37-42). Sin embargo, la escasa cantidad de estudios arqueofaunísticos realizados en proporción a la gran cantidad de excavaciones realizadas recomienda prudencia a la hora de esta-blecer generalizaciones. Además, todavía no se ha realizado prácticamente ningún estudio sobre aspectos muy importantes como son los patrones de consumo, la diacronía y distribución espacial de

71
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
los restos, sus asociaciones con diferentes tipos de estructuras, las herramientas y objetos manufactu-rados con restos de animales, etc.
El objetivo principal de este artículo es precisamente contribuir a un mejor conocimiento del papel que tuvieron los animales en la economía y el mundo simbólico de las comunidades que ocuparon Valen-cina a lo largo del III milenio cal ANE, aportando nue-vos datos obtenidos de un estudio inédito realizado a partir del registro obtenido en las excavaciones lle-vadas a cabo en el sector PP4-Montelirio entre 2007 y 2008 (Mora Molina et al., 2013). De acuerdo con la síntesis preliminar realizada por el Grupo de Inves-tigación ATLAS de los resultados de esta excavación, se identificaron un total 134 estructuras prehistóri-cas (probablemente calcolíticas en la mayoría de los casos), además de otras 40 de cronología dudosa o indefinida (Mora Molina et al., 2013). Siguiendo los criterios de este estudio preliminar, se han iden-tificado un total de 61 funerarias divididas a su vez en tres tipos de acuerdo con su complejidad arqui-tectónica: estructuras de tipo megalítico, estructu-ras negativas con elementos pétreos1 y estructuras negativas sin elementos pétreos (Mora Molina et al., 2013: 266). En relación con el número de indivi-duos identificados en estas tumbas, predominan las estructuras con inhumaciones colectivas (43) frente a las individuales (17). Otras 73 estructuras son de carácter no funerario y también han sido clasificadas en tres tipos en función de la presencia de elementos pétreos o no, o de difícil asignación a falta de carac-terísticas arqueológicas definitorias.
La principal aportación de este trabajo es, por ello, el estudio de la fauna recuperada en las diferentes estructuras del sector PP4-Montelirio. Una vez cuan-tificada y valorada en relación con sus contextos, la muestra de mamíferos ha resultado demasiado exi-gua para realizar estudios comparados con otros análisis faunísticos realizados en Valencina, aunque se discuten someramente algunas cuestiones de interés. A pesar de que se desconoce su ubicación micro-espacial exacta, los numerosos moluscos
marinos encontrados en los contextos funerarios han ofrecido una valiosa información sobre su papel como ajuar funerario, aunque todavía requieren de un estudio más exhaustivo para conocer su naturaleza funcional o simbólica a lo largo del III milenio ANE.
2. MATERIAL Y MÉTODOS
Tras una inspección previa del material durante una visita al Museo Arqueológico de Sevilla realizada en octubre del 2013 (C.L.) pudimos constatar el acusado grado de alteración postdeposicional y de fractura-ción de los restos óseos y, sobre todo, la abundan-cia de especies de invertebrados, lo que suponía una importante dificultad añadida de cara a una identifi-cación in situ de la muestra. La necesidad de la con-sulta, no sólo de una colección de fauna comparada, sino también de diferentes especialistas con el fin de garantizar la identificación, recomendó el traslado de los materiales desde el Museo Arqueológico de Sevilla al Laboratorio de Arqueozoología la Universi-dad Autónoma de Madrid (LAZ-UAM) para el estudio, limpieza y toma de muestras2.
Además de las alteraciones postdeposicionales, un fac-tor que ha condicionado de forma considerable el estu-dio arqueofaunístico del PP4-Montelirio es que, salvo excepciones, la gran mayoría de las 134 estructuras prehistóricas documentadas no han podido ser todavía objeto de un estudio pormenorizado, un proceso que llevará años completar dado el volumen del material recuperado (cincuenta cajas de artefactos y otras tan-tas de material óseo humano). Ello es especialmente importante en los casos de las estructuras para las que el excavador cita vestigios de cronología no prehistó-rica (cerámica a torno, en algunos casos fragmentos de hierro, etc.), y en los que los registros de excavación no aclararan ni la exacta posición de dichos vestigios, ni su características. Como se explica más adelante, la inexistencia de estudios del registro material de las estructuras que presentan cerámica a torno obliga a tomar con cautela la asignación cronológica de ciertos
1 Las primeras son descritas como estructuras negativas con algún elemento de piedra, normalmente una o varias (escasas) lajas de piza-rra dispuestas horizontalmente o hincadas verticalmente, pero que no tienen papel estructural y parecen jugar más bien un papel simbólico en la definición del espacio, mientras que las segundas son estructuras negativas puras, sin lajas de ningún tipo.
2 Este traslado fue autorizado por la Secretaría de Cultura de la Consejería de Educación, Cultura y Deportes de la Junta de Andalucía mediante resolución con fecha de 29 de enero de 2014, aunque la entrega de los materiales en Madrid solo se verificaría el 29 de marzo de 2014, siendo el plazo de depósito posteriormente ampliado hasta el 17 de noviembre de 2014, fecha en que el material fue devuelto al Museo Arqueológico de Sevilla.

72
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
restos faunísticos3. Otro importante lote de restos no se puede estudiar en la actualidad, al desconocerse a qué estructuras se asignan las UUEE anotadas en las bolsas. Esperemos que en futuros trabajos puedan ser clasificados e incorporados en los diferentes horizon-tes de ocupación del sector PP4-Montelirio. Como el presente trabajo se centra en la ocupación calcolítica del yacimiento se adjunta en el Apéndice 1 una tabla con los restos prehistóricos indeterminados y roma-no-medievales, también descritos en el Apéndice 2.
Para el estudio arqueofaunístico se agrupan los res-tos siguiendo los criterios establecidos por Mora Molina et al. (2013: 266): para una mayor facilidad de expresión, y siguiendo el criterio del Grupo de Inves-tigación ATLAS de la Universidad de Sevilla que está estudiando el registro de estas excavaciones. Las estructuras que contenían restos humanos son desig-nadas como de carácter funerario, mientras que las que no los contenían son citadas como ‘no funerarias’ (sin entrar en un debate respecto a posibles significa-ciones ‘no funerarias’ de los restos óseos humanos).
El sedimento de esta intervención no fue cribado ni flo-tado durante o tras el proceso de intervención, aspecto que evidentemente ha influido negativamente en los resultados de este estudio y muy especialmente en la ausencia de piezas de reducido tamaño, caso de los microvertebrados y los caracoles terrestres de menor talla. Aunque también es cierto que se han recuperado esquirlas óseas y de moluscos en varios contextos que apenas superan los 3 mm de tamaño, los únicos hue-sos de ave (un tibiotarso de gallina, Gallus gallus) y de roedores (probablemente un metacarpo) son sin duda evidencia de lo mucho que se habría perdido durante el proceso de excavación. De ello es también testigo la gran escasez de restos de conejo, generalmente abundantes en este tipo de yacimientos. Sobre los problemas de este tipo de actuaciones se ha adver-tido ya en numerosas ocasiones (Payne, 1972; Gifford, 1981; Gilbert y Singer, 1982; Grayson, 1984).
La identificación osteológica se ha llevado a cabo gracias a la colección comparativa de AMM sita en el LAZ-UAM. Como apoyo bibliográfico se han con-
sultado puntualmente las obras de Schmid (1972) y Barone (1976), así como las de Boessneck (1980) y Prummel y Frisch (1986).
Los moluscos se identificaron según estructuras y UUEE, al ser los contextos clave para explicar su presencia. La identificación de moluscos se hizo con el concurso de la colección comparativa de AMM, así como con la ayuda de dos especialistas en malacofauna andaluza actual (Dr. Angel Luque del Villar, UAM) y fósil (Dr. José Luis Vera Peláez, Gaia Museum) y bibliografía relevante (Gofas et al., 2012). La terminología empleada en el estudio preliminar del asta de ciervo sigue los criterios definidos por Billamboz (1979) traducidos por Liesau (1988).
Tres han sido los estimadores de abundancia incor-porados al estudio de los restos de fauna: el Número de Restos (NR); el número mínimo de individuos (NMI) calculado para cada una de las UUEE en las respectivas estructuras, atendiendo criterios de lateralidad y edad (Clason, 1972) y el peso. El peso del material desglosado por taxones (exceptuando a los moluscos) se expresa en gramos para poder valorar adecuadamente la tanatomasa de cada taxón según estructuras y de forma global. La determina-ción de edad se llevó a cabo empleando la metodo-logía desarrollada por la Escuela de Múnich4. Para la correlación de cohortes de edades (en meses) se ha seguido la metodología propuesta por Morales (1976). El principal criterio utilizado para la deter-minación del sexo se ha basado en el dimorfismo sexual de algunas porciones esqueléticas (cuernos, caninos, pelvis, etc.), teniendo además en cuenta, otros criterios osteomorfológicos y métricos. La osteometría, realizada con calibres convencionales (error estimado ± 0.5 mm), sigue los criterios de von den Driesch (1976) con las siglas traducidas por Miguel y Morales (1984).
Para el estudio de las huellas de manipulación antró-pica hemos seguido las cinco secuencias expuestas por Rixson (1988: 49) para el descuartizamiento de las carcasas animales, así como la terminología desarrollada por Liesau (1998: 59-61).
3 La dificultad para establecer la cronología de las estructuras del sector PP4-Montelirio se ha visto incrementada por el efecto de las con-diciones edafoquímicas imperantes, que han ocasionado que numerosas muestras de hueso humano enviadas para datación radiocarbónica no hayan deparado fechas debido a los bajos niveles de colágeno que presentaban. Sin embargo, dos individuos procedentes de la estructura 10.042 se pueden asignar al Calcolítico Antiguo de acuerdo con dos fechas publicadas: CNA-1291: 4161±34 BP (2880-2629 cal BC 2σ) y CNA-1303: 4277±31 BP (3007-2779 cal BC 2σ) (García Sanjuán et al., 2013: 625).
4 Tablas de edades elaboradas por A. von den Driesch y J. Boessneck (inédito) a partir de la fusión epifisaria y el reemplazo o desgaste dentario.

73
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
Algunas valvas de vieras y de lapas procedentes de los registros funerarios 10.038 y 10.073 han sido seleccionadas para un estudio de huellas antrópi-cas, así como para proceder a una serie de análisis químicos destinados a conocer su composición. A tal efecto, en el laboratorio del Dpto. de Prehistoria y Arqueología de la UAM, Dña. Ana Isabel Pardo, res-tauradora de Patrimonio Arqueológico, ha procedido a la limpieza mecánica suave con agua-alcohol de las concreciones carbonatadas que dificultaban la inspección macroscópica de las piezas así como a la toma de muestra con vistas a estas analíticas que desarrollaremos en un futuro próximo.
3. COMENTARIOS SOBRE CUESTIONES TAFONÓMICAS
En general, el material recuperado de la excava-ción del PP4-Montelirio muestra un elevado grado de fracturación post-deposicional y causado por los procesos de extracción durante la excavación. Del casi medio millar de restos de mamíferos recupe-rados, el 46% del material no ha podido ser iden-tificado (Tab. 1 y Apéndice 2). Estas circunstancias desfavorables también limitan la calidad del regis-tro osteométrico, al no haberse recuperado ni un solo hueso largo completo, e incluso la mayoría de las piezas dentarias, generalmente muy resistentes a la acción de diferentes agentes tafonómicos, se encuentran fracturadas o muy alteradas en super-ficie (sobre todo en las estructuras 10.023, 10.042 y 10.088). Sin embargo, no podemos descartar que en otras estructuras funerarias el deficiente estado de conservación pueda también deberse a la mani-pulación antrópica relacionada con los rituales de enterramiento, al presentar numerosas piezas huellas de combustión (p.ej. estructura 10.088; ver apartado de Discusión).
El material arqueofaunístico destaca también por presentar abundantes concreciones de carbonatos y silicatos que recubren las superficies. Estas obser-vaciones apuntan a que, con independencia de otras condiciones edáficas, los rellenos de estas estructu-ras negativas estuvieron sometidos a condiciones de humedad con el nivel freático de forma recurrente y prolongada, cuestión que aceleró su descompo-sición. Posiblemente tal hecho explique el escaso colágeno disponible en los huesos humanos remiti-dos para datación. La única colección faunística que, como conjunto, presenta un estado de conservación
medianamente aceptable son los moluscos marinos y, en menor medida, los terrestres, muchos de los últimos aparecidos en el interior de algunos cráneos humanos (p.ej., estructuras 10.031 y 10.034).
Durante la identificación se han podido detectar una serie de huesos humanos que se mencionan breve-mente en el inventario realizado como complemento de este trabajo (ver Apéndice 2). Algunos de éstos se corresponden con tumbas conocidas (10.031, 10.049), mientras que otros están asociados a contextos que hasta ahora no se habían relacionado con prácticas funerarias (estructuras 10.024 y 10.111, UUEE 148 y 620 y restos de tres individuos en la UE 724). Estos huesos humanos tendrán que ser valorados en rela-ción con los artefactos del registro, dado que la dis-persión generalizada de restos humanos suele ser frecuente en yacimientos prehistóricos peninsulares de larga ocupación, donde sucesivas remociones de tierras son incorporadas de forma involuntaria a nuevas estructuras. La dispersión de restos huma-nos también se vería favorecida por la práctica de exposición previa de los cadáveres, aspecto señalado en varios yacimientos calcolíticos (Márquez Romero y Jiménez Jáimez, 2010; Gómez Pérez et al., 2011).
4. RESULTADOS Y DISCUSIÓN
En las tablas 1-7 se ofrece, desglosada por contextos y grupos faunísticos, la relación de restos estudiados (en el Apéndice 1 se refiere el desglose de faunas de los horizontes no calcolíticos). Tan sólo 29 estructu-ras (22% de las 134 estructuras calcolíticas detecta-das en este sector) ofrecen restos de fauna hasta el momento y los 467 restos aquí estudiados contras-tan con la abundancia de fauna registrada en otras zonas del yacimiento (Hain, 1982; Pajuelo Pando y López Aldana, 2013; Bernáldez Sánchez et al., 2013). Tan sólo el Sector Plan Parcial Matarrubilla, un área identificada con actividades metalúrgicas, exhibe una pobreza faunística similar a la aquí documentada (NR= 789; Abril López et al., 2010). Aunque probable-mente también influya el hecho de que en un yaci-miento del enorme tamaño de Valencina las condi-ciones edafoquímicas que afectan a la conservación del registro óseo pueden variar bastante de un sector a otro del yacimiento, las importantes diferencias en los recuentos de fauna entre sectores debe obede-cer a razones culturales más que postdeposicionales o tafonómicas, probablemente conectadas con los usos y posibles especializaciones que los distintos

74
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
sectores del mismo tuvieron a lo largo del tiempo, aunque esta cuestión necesita de un estudio deta-llado que todavía no se ha realizado.
Las diferencias que apreciamos entre las muestras procedentes de contextos funerarios y no funerarios dentro de lo que consideramos fauna depositada intencionalmente, se refieren a la abundancia dife-rencial de moluscos. En este apartado, los contextos funerarios duplican los valores que se aprecian en los contextos no funerarios (30% vs.17%). Frente a ello, dentro de los contextos funerarios, los mamí-feros suponen el 64%, lo que contrasta con el más de 80% de estos restos para los no funerarios. Se trata de diferencias que, aun siendo apreciables, tampoco nos permiten decir que ambos tipos de contextos sean particularmente diferentes en fun-ción de su fauna. Más significativas resultan las diferencias dentro de la industria ósea, por cuanto la de los depósitos funerarios quintuplica la de los depósitos no funerarios. Puede aquí, por tanto, exis-tir una diferencia que, sin duda, remite a la natura-leza de la actividad humana. Se trata, en cualquier caso, de una cuestión que habrá de ser abordada desde un contexto general y, a ser posible, una vez se disponga de muestras más apreciables que las aquí reseñadas. Por último, las diferencias faunísti-cas más apreciables entre los contextos funerarios y los no funerarios se refieren al conjunto de fauna que hemos considerado intrusiva. Centrada ésta fun-damentalmente sobre el grupo de los pulmonados (moluscos terrestres) la fauna intrusiva de los depó-
sitos funerarios quintuplica, tanto en número como en porcentaje, los valores registrados en los depósi-tos no funerarios (Tab. 1). Estamos ante un fenómeno que no podemos atribuir a la actividad humana y que nos está indicando que por alguna razón esta fauna intrusiva parece haber manifestado una predilección por los contextos funerarios (Fig. 1). En el siguiente apartado abundaremos sobre este extremo, pero antes de hacerlo resaltaremos que esta mayor fre-cuencia de fauna intrusiva en los depósitos funera-rios quizá nos esté indicando el auténtico carácter de los dos únicos restos de conejo detectados (Tab. 2).
Si el desglose de faunas lo realizamos a nivel de especies representadas, habremos de constatar algunas diferencias apreciables entre los contextos funerarios y no funerarios (Tabs. 2, 4 y 5). En el caso de los mamíferos, el ciervo es la especie dominante en los contextos funerarios, situándose un orden de magnitud por encima de los valores que exhibe en los depósitos no funerarios (Tab. 2). Tal patrón se nos antoja más significativo en la medida en que la muestra de mamíferos identificados en los depósitos funerarios es la mitad de la registrada en los depósi-tos no funerarios. De igual modo, los escasos restos de perro aparecen restringidos a contextos funera-rios, algo que no debe sorprendernos en la medida que el perro rara vez constituye un objeto regular de consumo en la Prehistoria. Las diferencias que pre-sentan las restantes especies entre contextos, mien-tras tanto, es posible que no podamos interpretarlas en clave bien de consumo bien ritual habida cuenta la
GRUPOCALCOLÍTICOFUNERARIO
CALCOLÍTICONO FUNERARIO TOTAL
NR NR% NR NR% NR NR%
MAMÍFEROS 46 33,6 99 46,3 145 41,3
MOLUSCOS MARINOS 41 29,9 23 10,7 64 18,2
MOLUSCOS DULCEACUICOLAS 14 6,5 14 4
MACROMAMÍFEROS SI 42 30,6 75 35 117 33,3
INDUSTRIA ÓSEA 8 5,8 3 1,4 11 3,1
SUBTOTAL ANTRÓPICO 137 100 214 100 351 100
AVES 1 6 1 0,8
MICROMAMÍFEROS SI 1 1 1 0,8
MOLUSCOS TERRESTRES 87 88,8 17 94 104 90
MOLUSCOS SI 10 10,2 10 8,2
SUBTOTAL INTRUSIVO 98 100 18 100 116 100
TOTAL ESTUDIADO 235 (41) 232 (7,8) 467 (24,8)
Tab. 1. Relación general del número de restos (NR) desglosados por grupos faunísticos (SI= restos sin identificar). Los valores porcentuales se calculan independientemente para las faunas que se consideran intrusivas y aquellas que se presumen depositadas intencionalmente, refiriéndose los valores entre paréntesis al porcentaje que supone la fauna intrusiva sobre el total estudiado.

75
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
similitud de los valores en ambos tipos de contextos. La única excepción podría ser la presencia del équido en los contextos no funerarios, pero lo minúsculo de la muestra impide aventurar hipótesis alguna de uso. Únicamente destacar que, aunque el asno se pos-tulaba introducido durante la Edad del Hierro en la Península Ibérica, el reciente hallazgo de la especie en niveles calcolíticos del yacimiento de Leceia (Por-tugal) (Cardoso et al., 2013) abre la posibilidad de que el resto que aquí consignamos como “Equus sp.” y que, en principio, debería corresponder a un caballo, pudiera pertenecer a un asno. Todo ello obliga a ser cautelosos con la asignación a especie de los restos de équidos peninsulares a partir del tercer milenio a.C. y no sólo a partir del primero.
Por lo que atañe a la representación esquelética de las muestras de mamíferos decir, en primer lugar, que al ser tan menguadas, habrá que tomar cual-quier aseveración con suma cautela. Centrándonos sobre los contextos funerarios, podríamos argumen-tar la existencia de una cierta preferencia por las porciones craneales dado que éstas suponen 2/3 del total (Tab. 3). Esta tendencia es muy discutible. Se
antoja inequívoca en el caso del ciervo donde la tota-lidad de la muestra procede de astas que apuntarían al carácter presumiblemente ritual de estas piezas al tiempo que explicarían la aparente abundancia de la especie en este tipo de contextos a la que antes nos hemos referido. Frente a ello, la abundancia relativa de porciones craneales en la vaca podría ser simplemente reflejo de la mayor robustez que exhiben los dientes frente a los huesos, ya que los primeros suponen el 90% de esta muestra craneal. Por último, la paupérrima representación de ovejas, perros y conejos impide interpretar la abundancia o la inexistencia de piezas craneales en clave de algún fenómeno determinista ya que tales presencias/ausencias pueden deberse exclusivamente a fenó-menos estocásticos más que de tipo aleatorio.
Por lo que se refiere a la representatividad esquelé-tica en contextos no funerarios, parece muy evidente que, a diferencia de los sesgos que apreciamos en los depósitos funerarios, las muestras más volumi-nosas (vaca y oveja/cabra) apuntan a la presencia de ejemplares completos en contextos no funera-rios, a diferencia de los sesgos que apreciamos en
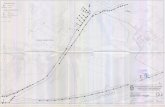
Fig. 1. Plano del sector PP4- Montelirio con la ubicación de las estructuras inalteradas frente a otras con faunas intrusivas por expolio o alteraciones antrópicas en períodos posteriores (10.024, 10.042 y 10.088), o por causas naturales (10.015; 10.032; 10.034 y 10.073). Diseño gráfico: José Manuel Durán Moreno.

76
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
los depósitos funerarios, (Tab. 4). Con todo y con ello es evidente que, una vez más, los dientes, que en conjunto suponen casi la cuarta parte del total, vuelven a presentar una marcada sobreabundancia que seguramente tiene mucho que ver con su mayor capacidad de pervivir en los depósitos al encontrarse más mineralizados que el hueso. Al tiempo que ello ocurre, resulta muy patente que no es posible reali-zar ningún tipo de inferencia sobre la representativi-dad esquelética en las muestras de équidos, suidos, ciervo y uro, habida cuenta lo paupérrimo de los con-tingentes recuperados.
En el caso de los moluscos, una diferencia apreciable entre las muestras de contextos funerarios y no fune-rarios remite al tamaño de las especies. En efecto, tanto Otala lactea como Theba pisana son los pulmo-nados de mayor tamaño y los únicos que aparecen en los contextos no funerarios (Tabs. 5 y 7). Frente a ello, la mayoría de las especies de caracoles terrestres de contextos funerarios son de pequeño, cuando no dimi-nuto, tamaño. Con todo, la diferencia más llamativa entre contextos referida a los moluscos habremos de encontrarla en la exclusiva presencia de las náya-des (moluscos dulceacuícolas) en los contextos no
TAXÓNCALCOLÍTICO FUNERARIO CALCOLÍTICO NO FUNERARIO
NR % NMI % PESO % NR % NMI % PESO %
Bos taurus 15 32,6 3 27,3 380 62,8 51 51,5 7 31,8 1266 54,1
Ovis aries 2 4,3 1 9,194 15,5
2 2 2 9,1192 8,2
Ovis/Capra 2 4,3 2 18,2 31 31,3 6 27,3
Sus sp. 2 4,3 1 9,1 6 1 10 10,1 3 13,6 86 3,7
Canis familiaris 3 6,5 1 9,1 12 2
Equus sp. 1 1 1 4,5 55 2,3
Bos primigenius 3 6,5 1 9,1 112 18,5 2 2 2 9,1 723 30,9
Cervus elaphus 17 37 1 9,1 (380) - 2 2 1 4,5 20 0,9
Oryctolagus cuniculus 2 4,3 1 9,1 1 0,2
TOTAL 46 100 11 100 605 100 99 100 22 100 2342 100
Tab. 2. Número de restos (NR), individuos (NMI) y peso de los mamíferos del PP4-Montelirio que se presumen acumulados por actividad antrópica, agrupados según contextos funerarios o no. Los restos de ciervo del Calcolítico funerario representan astas que, por desconocer si son piezas mudadas, no podemos atribuir a ejemplares cazados. Por esta razón, se indica su peso entre paréntesis no incluyéndose este valor en los cómputos globales.
Categoriaósea
Bostaurus
Ovisaries
Ovis/Capra Sus sp.
Canis fami-liaris
Cervus elaphus
Oryctolagus cuniculus
Bos primigenius TOTAL %
Asta/cuerno 2 17 19 41,3
Diente superior 5 1 6 13,0
Mandíbula 1 1 2,2
Diente inferior 2 1 3 6,5
Diente si 2 2 4,3
∑ Craneal 10 2 1 1 0 17 0 0 31 67,4
Vértebra 1 1 2,2
∑ Axial 1 0 0 0 0 0 0 0 1 2,2
Escápula 2 2 4,3
Húmero 1 1 1 1 4 8,7
Radio 1 2 3 6,5
Ulna 1 1 2,2
Metatarso 1 1 2,2
Pelvis 1 1 2,2
Tibia 2 2 4,3
∑ Apendicular 4 0 1 1 3 0 2 3 14 30,4
TOTAL 15 2 2 2 3 17 2 3 46 100
Tab. 3. Número de restos de los mamíferos desglosados por categorías esqueléticas en los contextos funerarios.

77
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
funerarios. Habida cuenta el doble carácter funcional (consumo, utensilios) y ritual que pueden implicar los restos de náyades, la desvinculación de estas conchas con los contextos funerarios apuntaría a usos exclusi-vamente utilitarios en Valencina (Hain, 1982: 144).
4.1. ESTRUCTURAS NO FUNERARIAS
Las estructuras no funerarias totalizan 73 (54% del total de las 134 detectadas de esta zona). De éstas, solo 9 (apenas el 12%) evidencian restos de fauna. Sin embargo, como bien podemos apreciar en la Tabla 5, la presencia de fauna en todas ellas es tes-timonial a excepción de la estructura 10.024 cuyos restos suponen más del 95% del total recuperado en estos 9 depósitos.
La fauna de la estructura 10.024 incluye un total de 16 taxones en donde aquellos que no constan como intrusivos con total seguridad (los pulmo-nados Otala lactea y Theba pisana y la gallina que obligatoriamente remite a una etapa post-calcolí-tica) suponen sólo el 8% del total estudiado (Tab. 5). La presencia de la gallina resulta de lo más relevante por cuanto indica una contaminación diacrónica del depósito que arroja dudas sobre si el resto de la fauna que cabe suponer originaria en la estructura 10.024 podría representar también fauna intrusiva5. En este sentido, el que ninguna de las restantes UUEE no funerarias -a las que cabría atribuir una condición como acumuladoras de res-tos alimentarios- presenten restos de mamíferos, que suponemos hubieron de representar el grueso de la alimentación de origen animal en el asenta-
Categoria ósea Equus sp.Bos
TaurusOvis aries
Ovis/Capra Sus sp.
Cervus elaphus
Bos primigenius TOTAL %
Asta/cuerno 1 1 1,0
Cráneo 3 1 4 8 8,1
Diente superior 1 2 3 3,0
Mandíbula 4 4 1 9 9,1
Diente inferior 2 3 5 5,1
Diente si 7 5 12 12,1
∑ Craneal 17 15 5 1 38 38,4
Vértebra 3 1 4 4,0
Costilla 12 1 13 13,1
∑ Axial 15 2 17 17,1
Escápula 1 1 1 2 5 5,1
Húmero 4 1 5 5,1
Radio 3 3 1 7 7,1
Ulna 1 1 1,0
Metacarpo 1 1 1 3 3,0
Pelvis 1 3 1 1 6 6,1
Fémur 1 1 1,0
Tibia 2 2 2,0
Metapodio 3 1 1 5 5,1
Astrágalo 1 1 2 2,0
Centrotarsal 1 1 1,0
FI 1 4 5 5,1
∑ Apendicular 1 18 2 14 5 1 2 43 43,6
TOTAL 1 51 2 31 10 2 2 99 100
Tab. 4. Número de restos de los mamíferos desglosados por categorías esqueléticas en los contextos no funerarios.
5 Una actuación de “prospección” realizada en 1998 con una pala excavadora causó daños a diferentes estructuras al excavar numerosas zanjas de gran longitud en todo el sector del PP4. Como han evidenciado posteriores excavaciones de José Peinado Cucarella en 2007 y 2008, dichas zanjas rompieron y desmocharon numerosas estructuras funerarias y su posterior relleno con tierras ya alteradas, tanto de los estratos superficiales, como de los depósitos prehistóricos, ha podido favorecer la intrusión de materiales recientes en varios contextos calcolíticos.

78
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
miento, resulta de lo más llamativo. En realidad, el que los moluscos de todas las restantes uni-dades hubieran sido potencialmente consumibles no apoya ni refuta su condición de piezas intru-sivas dado el paupérrimo número de restos que suponen.
Así las cosas, podemos asegurar que:
1) La única estructura que presenta un volumen de fauna que permitiría una valoración cuantitativa parece incluir cierta cantidad de fauna intrusiva, lo que deja en suspenso la posibilidad de hacer inferencias sobre el uso de los animales en época calcolítica hasta que se obtengan dataciones radiocarbónicas que confirmen la cronología cal-colítica de los restos.
2) La ausencia de estos taxones intrusivos en los restantes depósitos no funerarios puede ser simplemente resultado de la práctica ausencia de evidencias faunísticas en las mismas y,
3) Esta pobre representatividad de restos faunísti-cos en depósitos que podríamos pensar incorpo-ran vestigios alimentarios no parece que apoye tal hipótesis. La confirmación de ‘basureros’ quedaría por tanto supeditada a la valoración de otros registros complementarios al de la fauna.
Si se asume, en todo caso, que la fauna presente de la estructura 10.024 puede considerarse repre-sentativa de la economía animal del asentamiento calcolítico, lo que parece fuera de toda duda es que la fauna silvestre y, por ello, la caza y el marisqueo,
TAXON/UES 10.024 627 631 656 663 703 704 908 944 ∑
Bos taurus 51 51
Ovis aries 2 2
Ovis/Capra 31 31
Sus sp. 10 10
Bos primigenius 2 2
Cervus elaphus 2 2
Equus sp. 1 1
∑ MAMÍFEROS 99 99
Gallus gallus 1 1
Spisula cf. elliptica 1 1
Glycymeris sp. 1 1
Pecten sp. 1 1
Cerastoderma edule 1 1
Ostrea sp. 2 2
Ruditapes cf. decussatus 9 1 1 1 1 2 15
Antalis sp. 1 1
Charonia lampas 1 1
∑ MOLUSCOS MARINOS 12 1 3 1 1 1 1 1 2 23
Otala lactea 7 7
Theba pisana 10 10
∑ MOLUSCOS TERRESTRES 17 17
Unio delphinus 4 1 5
Potomida littoralis 7 7
Unionidae 2 2
∑ MOLUSCOS AGUA DULCE 13 1 14
MOLUSCO Indet. 1 1
MAMÍFEROS Indet. 75 75
INDUSTRIA ÓSEA 3 3
TOTAL 221 1 3 1 1 1 1 2 2 233
Tab. 5. Número de restos recuperados en las estructuras no funerarias, desglosados por taxones.

79
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
serían actividades secundarias en relación con la ganadería. Así, suponiendo que tanto los restos de los suidos como los de mamíferos indetermina-dos pudiesen añadirse al contingente doméstico, las cabañas ganaderas supondrían casi el 77% de la fauna recuperada (si restringimos los cálculos a las especies determinadas sin ningún género de dudas, los 84 restos de vacuno y ovicaprinos conti-nuarían suponiendo el 64% de los 130 restos inclui-dos en esta categoría). Si a ello le añadimos el prác-ticamente nulo aporte cárnico que habían supuesto los 23 restos de moluscos potencialmente consu-mibles (el diente de elefante o Antalis cabe inter-pretarlo como elemento ornamental), la importan-cia de la ganadería, “dominada” por el vacuno, se vería incrementada, resultado también avalado por el parámetro del peso, excluido de momento de la discusión, al no disponer hasta la fecha de ningún estudio del contexto y de dataciones radiocarbóni-cas de la estructura 10.024 que aporte datos sobre la funcionalidad de este recinto (ver Apéndice 2). A pesar de ello, también aquí parece confirmarse la importancia del ganado vacuno como principal proveedor cárnico, al igual que ha quedado ates-tiguado para otras áreas de este yacimiento (Hain, 1982: 141; Abril López et al., 2010: Pajuelo Pando y López Aldana, 2013: Tabla 5). No obstante, aquí no se confirma una significativa presencia de los suidos en cuanto al NR, resultado que merece dis-cutirse con más detalle en futuros trabajos habida cuenta lo referido en estudios previos (Hain, 1982: 143; Abril López et al., 2010:94; Pajuelo Pando y López Aldana, 2013: Tabla 5; Bernáldez Sánchez et al., 2013: 427).
4.2. ESTRUCTURAS FUNERARIAS
De las 61 estructuras funerarias detectadas en el PP4-Montelirio, escasamente la tercera parte (32,8%) incorporan fauna, lo cual, sin querer aven-turar demasiado, dada la escasez general de datos, podría sugerir rituales funerarios “poco comprome-tidos” con las ofrendas animales. Sea como fuere, se trata de un porcentaje muy superior de depósitos con fauna al registrado en las estructuras no funerarias. A ello se añaden dos hechos destacados (Tab. 6):
1) Los mamíferos no quedan circunscritos a un solo depósito, sino que aparecen en 6 de las 20 estructuras funerarias (30%).
2) Los mamíferos silvestres en general, y el ciervo en particular, representan una fracción aprecia-ble tanto del total de mamíferos (26% y 19% en el caso del ciervo) como, sobre todo, de los res-tos de mamíferos identificados taxonómicamente (e.d., 53,5% y 39,5% respectivamente).
Del mismo modo que ocurría con las estructuras no funerarias, en estos depósitos se constata una pre-sencia recurrente de fauna intrusiva. Así, aunque los pulmonados solamente se recuperan en una ter-cera parte de las estructuras (7), representan sobre el total de restos faunísticos el 38% de lo estudiado (casi el 61% de los moluscos) lo que prácticamente quintuplica la aportación de esta fauna en el caso de los depósitos no funerarios. De hecho, entre los mamíferos, constatamos otro caso adicional de fauna intrusiva (el roedor recuperado en la estruc-tura 10.031), al que quizás podríamos añadir, por su calidad de fauna bioturbadora, el contingente de conejo recuperado en la estructura 10.042.
ESPECIE/ ESTRUCTURA 10.023 10.031 10.032 10.042 10.042-49 10.088 TOTAL
Bos taurus 7 5 3 15
Ovis aries 2 2
Ovis/Capra 2 2
Sus sp. 2 2
Canis familiaris 3 3
Bos primigenius 3 3
Cervus elaphus 17 17
Oryctolagus cuniculus 2 2
Rodentia indet. 1 1
MAMÍFEROS SI 1 1 8 20 12 43
TOTAL MAMÍFEROS 8 1 1 29 27 23 89
Tab. 6. Número de restos de mamíferos recuperados en las estructuras funerarias desglosados por taxones.

80
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
Un grupo recurrente de faunas asociadas a los regis-tros funerarios son los moluscos terrestres. Los registros funerarios, especialmente los que ofrecen cavidades y espacios inicialmente no colmatados, permiten el cobijo o la hibernación de este tipo de fauna. Por ello, la mayor abundancia de caracoles intrusivos en las estructuras funerarias no debe ser casual y esto podemos apreciarlo, en primer lugar, en la composición faunística del grupo. Así, si entre las estructuras no funerarias solo constatábamos la presencia de dos especies de apreciable tamaño como son Otala lactea y Theba pisana, en el conjunto de los pulmonados de estructuras funerarias, estos grandes caracoles no suponen ni la quinta parte del total (19,5%), estando aquí las muestras dominadas por siete especies de pequeño o diminuto tamaño. Las razones pueden tener mucho que ver con los hábitats carroñeros, cuando no directamente carní-voros, de estos pequeños caracoles. En este sentido destacar que son especies que suelen congregarse
en torno a cadáveres de todo tipo, siendo en nuestro caso los humanos los candidatos prioritarios. Pero, sin desdeñar la primera hipótesis, tampoco es des-cartable que las razones de esta mayor presencia de pulmonados en las tumbas haya que buscarlas en las condiciones creadas por la descomposición de los cadáveres. En efecto, como casi todos los pulmona-dos de nuestras latitudes, los caracoles detectados en Valencina pertenecen a especies conocidas por sus hábitos hibernantes colectivos. Como quiera que tanto el proceso de descomposición de un cadáver como el cierre que suponemos tuvieron las tumbas, sin duda contribuyeron a incrementar la tempera-tura del depósito funerario, tal fenómeno hubo de haber atraído a muchos caracoles durante su fase de agregación previa a la hibernación. En el PP4-Monte-lirio un excelente ejemplo es la estructura megalítica 10.034, en la que en todos los cráneos de los siete individuos inhumados se han recuperado distintas especies de pulmonados totalizando 59 individuos,
ESPECIE/NMI/AJUARES 10.012
10.015
10.016
10.022
10.023
10.031
10.032
10.034
10.036
10.037
10.038
10.040
10.042
10.042-49
10.049
10.067
10.073
10.074
10.081
10.083
10.088 ∑
Glycymeris sp. 1 1
Pecten maximus 1 1 1 1 1 1 3 9
Pecten jacobeus 1 1
Pecten sp. 1 1 1 1 1 2 7
Cymbula nigra 1 1 1 1 4
Eastonia rugosa 1 1
Ruditapes cf. decussatus 17 17
Charonia lampas 1 1
∑ MOLUSCOS MARINOS 1 1 1 1 2 1 1 2 1 2 5 1 3 17 41
Xerotricha madritensis 13 3 16
Xerotricha apicina 2 2
Xerosecta pomissa 1 1
Ferusssacia follicula 3 3
Candidula gigaxii 1 5 6
Helicellino/Helicella 1 2 25 6 34
Otala lactea 1 2 1 6 10
Rumina decollata 2 3 3 8
Theba pisana 7 7
∑ PULMONADOS 2 1 2 4 59 13 6 87
MOLLUSCA indet. 2 2 6 10
INDUSTRIA ÓSEA 3 5 8
TOTAL 1 2 1 1 2 2 4 61 1 1 2 1 20 7 5 7 1 1 3 23 146
NMI HUMANOS 1 13 4 2 23 3 8 7 3 3 3 3 5 1 1 6 3 1 2 8 1 101
MARFIL/HUES0 1 2 2 1 7 22 3 38
HUEVO AVESTRUZ 1 1
Tab. 7. NR de moluscos recuperados en las estructuras funerarias desglosados por taxones con datos complementarios de interés.

81
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESPECIE/NMI/AJUARES 10.012
10.015
10.016
10.022
10.023
10.031
10.032
10.034
10.036
10.037
10.038
10.040
10.042
10.042-49
10.049
10.067
10.073
10.074
10.081
10.083
10.088 ∑
Glycymeris sp. 1 1
Pecten maximus 1 1 1 1 1 1 3 9
Pecten jacobeus 1 1
Pecten sp. 1 1 1 1 1 2 7
Cymbula nigra 1 1 1 1 4
Eastonia rugosa 1 1
Ruditapes cf. decussatus 17 17
Charonia lampas 1 1
∑ MOLUSCOS MARINOS 1 1 1 1 2 1 1 2 1 2 5 1 3 17 41
Xerotricha madritensis 13 3 16
Xerotricha apicina 2 2
Xerosecta pomissa 1 1
Ferusssacia follicula 3 3
Candidula gigaxii 1 5 6
Helicellino/Helicella 1 2 25 6 34
Otala lactea 1 2 1 6 10
Rumina decollata 2 3 3 8
Theba pisana 7 7
∑ PULMONADOS 2 1 2 4 59 13 6 87
MOLLUSCA indet. 2 2 6 10
INDUSTRIA ÓSEA 3 5 8
TOTAL 1 2 1 1 2 2 4 61 1 1 2 1 20 7 5 7 1 1 3 23 146
NMI HUMANOS 1 13 4 2 23 3 8 7 3 3 3 3 5 1 1 6 3 1 2 8 1 101
MARFIL/HUES0 1 2 2 1 7 22 3 38
HUEVO AVESTRUZ 1 1
Tab. 7. NR de moluscos recuperados en las estructuras funerarias desglosados por taxones con datos complementarios de interés.
como también la estructura 10.031, excavada en la roca, han sido recuperados dos pequeños pulmona-dos del interior de los cráneos de dos individuos.
La fauna de moluscos marinos aporta luz sobre esta cuestión tafonómica como consecuencia de tres hechos fundamentales (Tab. 7):
1) Estado de conservación. Como antes se comentó, los moluscos marinos son, de entre toda la fauna estudiada del sector PP4-Montelirio, el único conjunto que presenta un aceptable grado de con-servación y escasa fragmentación. Por ello cabe suponer que los depósitos de estos moluscos son primarios y que este grupo, en el caso de las tum-bas, parece haber permanecido en los lugares en donde fueron originalmente depositados.
2) Representatividad específica. Aunque de las cuando menos siete especies representadas
en estas muestras tres no se documentan en los depósitos no funerarios, lo más interesante es que la almeja fina (Ruditapes decussatus) presenta aquí una distribución restringida a la estructura 10.088 (es decir, al 5% de los depó-sitos funerarios con fauna), mientras que, por el contrario, con una muestra muy similar, en las estructuras no funerarias esta almeja es un elemento recurrente, que documentamos en el 66% de los depósitos (Tab. 5). Cabe notar por tanto que una de las especies más apreciadas a efectos de consumo aparece circunscrita a una única estructura funeraria, mientras que aparece repartida en una mayoría de las no-funerarias.
3) Disposición de las conchas (Fig. 2). A diferencia de la restante fauna de Valencina, muchas de estas conchas marinas, por lo general grandes y llama-tivas, parecen haber sido cuidadosamente depo-sitadas junto a los cadáveres. En algunos casos

82
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
se sitúan cerca de la cadera del inhumado (p.ej. en las estructuras 10.016, Fig. 2A; 10.040, 10.081) mientras que en otros se encuentran próximas a la cabeza (estructura 10.073, Fig. 2D) o los miem-bros inferiores (Fig. 2C). Además, en todos los casos, las valvas han sido depositadas de forma que la concavidad esté vuelta hacia arriba, hecho que, unido al tamaño de los especímenes, sugiere un uso a modo de contenedores. Todo ello per-mite postular para estas conchas una condición de ajuar funerario que rompe la indefinición funcional que ha venido siendo norma en las faunas recu-peradas de este sector. También parece claro que
el número de conchas que forman parte de esta categoría no se correlaciona de forma directa con el número de inhumados por cuanto tanto tumbas individuales (10.012, 10.074) como dobles (10.022, 10.081) y colectivas (10.016, 10.023, 10.036, 10.037, 10.040, 10.073) presentan una única concha. Ello podría sugerir que tales conchas tenían para este grupo humano un simbolismo específico asociado a la muerte, algo que no se ha constatado en otros sectores del yacimiento. En este sentido, es des-tacable la pauta de deposición de moluscos mari-nos en la estructura funeraria más importante del sector PP4-Montelirio, la tumba 10.042-10.049. En
Fig. 2. Selección de estructuras funerarias con los moluscos marinos documentados in situ:
A. Estructura 10.016 con una inhumación colectiva y sus ajuares. Una valva derecha (cóncava) de viera colocada en el centro de la tumba.
B. Estructura 10.083 con una inhumación colectiva y sus ajuares. Dos valvas derechas (cóncavas) de viera fueron colocadas junto a los indi-viduos. Detalle de una tercera valva de viera junto a un recipiente cerámico.
C. Estructura 10.067 con una inhumación colectiva y un conjunto de moluscos próximo a los individuos. A la izquierda, detalle del conjunto, donde en un plano inferior lateral se ubican una valva de viera y una lapa, mientras que en un plano superior, con la cara cóncava a la vista, otra de almeja y de la valva de Eastonia rugosa.
D. Estructura 10.073 con una Inhumación colectiva. Lapa de la especie Cymbula nigra con la cara cóncava a la vista colocada cerca del occi-pital de un individuo en decúbito lateral izquierdo. Detalle de la pieza durante la excavación.

83
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
la cámara ‘principal’ de esta estructura (10.049) no se recuperaron ni conchas ni huesos de mamí-feros, a pesar de que en ella se documentan los hallazgos faunísticos ‘exóticos’ más espectacula-res de Valencina, incluyendo un colmillo de ele-fante asiático y numerosas piezas de marfil de elefante africano, así como un huevo de avestruz, que se suman a un cuantioso depósito de objetos de prestigio, que incluye además objetos exóticos como ámbar, cinabrio y una hoja de puñal en cris-tal de roca (García Sanjuán et al., 2013b). En cam-bio, en la antecámara de esta estructura (10.042), donde se han identificado restos de 5 individuos, se encontraron una concha del tritón (Charonia lampas) y una valva de viera (Pecten sp.).
Otras 12 tumbas situadas en torno a la 10.042-10.049 presentan vieras, lo que nos confirma a este género como un elemento recurrente del ajuar, dándose la circunstancia de que estas tumbas no se encuentran agrupadas ni distribuidas de ningún modo peculiar, sino que están dispersas en torno a la gran tumba 10.042-10.049, que parece tener un papel “central” en este sector (Fig. 3). A pesar de que los estudios
antropológicos todavía están en curso y no conoce-mos el género ni la edad de la mayoría de los indivi-duos inhumados, no descartamos que pudieran ser un símbolo de una identidad de género, o incluso grupal, ya que también aparecen puntualmente en otra necrópolis calcolítica como es la de Los Millares (Leisner, 1943).
En el caso de los mamíferos, en cambio, la condi-ción de depósitos intencionales dista mucho de estar resuelta. Así, con independencia de la existencia de elementos intrusivos (ratón), o potencialmente intru-sivos (¿conejo?), desconocemos su ubicación con respecto a los esqueletos humanos y carecemos de evidencias que nos señalen la presencia de elemen-tos óseos en conexión anatómica, pautas para reco-nocer ofrendas funerarias de carácter alimentario en vertebrados. Por el contrario, tanto la presencia de restos craneales de vacuno (10.023), como los de oveja y de ciervo de la estructura 10.042 (Fig. 4), apuntarían en el sentido de depósitos intencionales por tratarse de defensas cuya connotación simbólica ha sido invocada en repetidas ocasiones. En el caso del ciervo, todos los restos son esquirlas de asta, sin
Fig. 3. Plano del sector PP4- Montelirio con la localización de aquellas estructuras funerarias que tienen entre los elementos del ajuar una o va-rias especies de moluscos marinos, dejando constancia del número de piezas en cada una de ellas. Diseño gráfico: José Manuel Durán Moreno.

84
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
que hayamos sido capaces de determinar si se trata de un asta o más, o de si se trata, como parece, de desmogues o de piezas unidas al cráneo, además de otras que pudieran haber formado parte de algún enmangue (Tab. 6 y Fig. 4 D-G). No parecen argu-mentos definitivos para demostrar de forma conclu-yente un estatus simbólico, aunque el contexto indu-dablemente apunta en tal sentido.
Pero frente al discurso lineal, aparentemente con-vincente, de la recurrente presencia de restos depo-sitados intencionalmente habremos de incorporar otros datos que nos obligan a la reflexión. En efecto, tanto la estructura 10.042-10.049 como la 10.088, que alberga restos de perro, uro, vacuno y suido, presen-tan evidencias de alteraciones post-deposicionales. Así, la antecámara de la estructura 10.042-10.049 registra una alteración que afectó tanto a los restos humanos como a los de fauna (Robles Carrasco y Díaz-Zorita Bonilla, 2013: 378). La estructura 10.088, por su parte, quedó dañada por la excavación de una trinchera durante la intervención arqueológica de 1989 (Mora Molina, 2011) que explicaría el elevado
número de esquirlas documentadas en este depó-sito, que suponen el 52% de los restos de mamíferos no identificados en esta estructura (Fig. 1 y Tab. 6) así como también la presencia de animales intrusivos; pero en qué medida afecta la trinchera la calidad del registro faunístico tomado en su conjunto, es algo sobre lo que sólo cabe especular en estos momen-tos, aunque sí destacar que en esta tumba numero-sos huesos con huellas de combustión parecen indi-car un tratamiento del conjunto relacionado con los rituales de enterramiento del inhumado.
Por último, como “industria ósea” (actualmente en proceso de estudio) tenemos un reducido lote de huesos y piezas en asta muy fragmentadas hallado en las estructuras 10.042-10.049 y 10.024. Predomi-nan piezas con un escaso grado de transformación entre las que se encuentran espátulas-alisadores y varios apuntados. Mientras que puede ser discutible si estas piezas han formado parte del ajuar original de la estructura funeraria 10.042-10.049 o su pre-sencia se debe a alteraciones posteriores, sí merece la pena destacar que, con sólo tres piezas, el recinto
Fig. 4. Conjunto de restos de fauna recuperados de la estructura 10.042-10.049:
A. Dos fragmentos de clavija ósea, probablemente de un carnero.
B. y C. Lote de piezas en asta de ciervo, tanto de asta principal como fragmentos de luchaderas.
D-G. Piezas en asta de ciervo con la superficie exterior alisada que en su extremo inferior presentan un reborde perimetral. Debido al acusado grado de fragmentación y alteración del conjunto no es posible determinar si se trata de unos rebordes resultado del rebaje intencional de la superficie externa o si pueden ser el vestigios rebajados y alisados de característico perlado de la rueda del asta, ¿fragmentos de enmangues?

85
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
circular - estructura 10.024 - no permite avanzar interpretación alguna en torno a procesos de trans-formación relacionados con artefactos óseos.
5. CONCLUSIONES
Valencina de la Concepción es un yacimiento inhe-rentemente complejo. Esta complejidad tiene, lógi-camente, mucho que ver con su extensión, gigan-tesca en el marco referencial de la Prehistoria Reciente y Protohistoria ibéricas, pero sobre todo con la heterogeneidad espacial y con un registro nada sencillo, por su diacronía y por la complejidad estratigráfica de las estructuras negativas. En espa-cios en apariencia rituales y funerarios, como es el caso del sector PP4-Montelirio, habremos de añadir a todo lo anterior una disposición, tanto de tumbas como de inhumaciones y materiales asociados, que deberemos únicamente valorar sobre la base de una inhumación fragmentaria, cuando no inconexa, con frecuencia muy dañada por el paso del tiempo. La historia tafonómica de este sector, por último, no es tampoco ajena a episodios de expolio, remo-ción y otras alteraciones (incluyendo intervenciones arqueológicas) que en modo alguno permiten reali-zar una lectura simplista de las evidencias.
Especial atención de cara a futuros estudios merece el recinto circular definido por la estructura 10.024 y su posible relación con las estructuras funerarias, los rituales de inhumación y de cierre de las tumbas, donde numerosas cuestiones están pendientes de estudio (depósitos de materiales cerámicos, líticos etc., posibles prácticas de comensalidad). Es, por tanto, aquí como en algunos registros funerarios un problema recurrente en lo que se refiere a la valo-ración de las evidencias faunísticas y la contigüidad espacial de ciertos restos. Como bien sabemos, con-tigüidad no necesariamente implica asociación, pero también nos consta que la mente humana tiende a obcecarse en estos casos. En el nuestro parece ser que tenemos un ejemplo claro en la UE 554 de la estructura 10.024 donde, aparentemente encajada entre las dos ramas de una mandíbula de vacuno, aparece una escápula de este grupo. Lo improbable de esta disposición espacial invita a plantear la posi-bilidad de considerar este conjunto como evidencia de un depósito intencional, lógicamente con alguna connotación simbólica, ritual, etc. Pero ¿hasta qué punto no es más probable que se trate de una mera contingencia carente de intencionalidad alguna? En
el estado actual de la investigación sobre el sector PP4-Montelirio, y con los datos disponibles, es impo-sible decantarse por una u otra opción y por ello adoptamos la postura que entendemos más parsi-moniosa (es decir, conexión casual, por mucho que se encuentre el hallazgo en una estructura circular próxima a los enterramientos). Un segundo ejemplo de esta “casualidad espacial” de tan difícil valoración en el PP4-Montelirio lo constituye el conjunto repre-sentado por una serie de moluscos aparecidos en la estructura 10.067 donde la especie Eastonia rugosa presenta una ubicación cenital junto a una valva de Glycymeris, una lapa (Cymbula nigra) y una viera (Pecten sp.) (Fig. 2C). Ello apunta a que las cuatro pie-zas pudieron haber sido depositadas dentro de algún contenedor (cesto, cajita, tejido) confeccionado con materiales perecederos que se desintegraron den-tro de un espacio colmatado. Tal hipótesis cobraría más fuerza por situarse el acúmulo a los pies de uno de los individuos inhumados, pero ¿cómo podemos refutar esta hipótesis? en ausencia de datos verifica-bles, la hipótesis se convierte en opinión y escapa por completo del dominio que llamamos ciencia.
Por todo lo anterior, en tanto no dispongamos de información sobre la sedimentología de los depó-sitos y la tierra que los circundaba, lo máximo que podemos decir es que la zona analizada destaca, ante todo, por una escasez manifiesta de fauna y que un sector importante de la recuperada nada tiene que ver con la actividad humana pretérita en ese sector de Valencina, la cual sin duda fue importante. Del mismo modo, podemos afirmar, que, frente a los que es la situación documentada en la tumba central (10.042-10.049), toda la fauna estudiada puede con-siderarse local, lo que incluye también a las especies de moluscos marinos, por tratarse de taxones que aparecen en el litoral próximo a la desembocadura del Guadalquivir. La presencia de uro, especie pre-sumiblemente extinguida en la Península Ibérica desde época romana, y la de las náyades, apuntan al consumo de unas especies silvestres que no parece frecuente en la Prehistoria peninsular, siempre que podamos ser capaces de confirmar su carácter ali-mentario (p.ej. a través del estudio de las huellas de uso, estudios isotópicos, etc).
Con todo y con ello, entendemos que es dentro de los moluscos marinos -todos ellos necesariamente transportados al lugar- donde tenemos las pau-tas más claras de una interrelación estrecha con la fauna. El que dicha relación lo sea en unos contextos

86
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
funerarios, donde las interpretaciones se complican sobremanera, obligaría, en un futuro, a establecer una más estrecha colaboración con otros especia-listas así como una exhaustiva contextualización arqueológica, que permitiesen bien corroborar bien refutar no pocas de las cuestiones que permanecen pendientes de estudio tras este análisis.
AGRADECIMIENTOS
Deseamos agradecer al Grupo de Investigación ATLAS (HUM-694) de la Universidad de Sevilla, y en especial al Dr. Leonardo García Sanjuán la tramita-ción y obtención de los necesarios permisos para el traslado del material desde el Museo Arqueológico de Sevilla al LAZ de la UAM, además de correr con los gastos asociados. También agradecemos a D. José Manuel Durán Moreno la colaboración prestada en la realización de las Figuras 1 y 3, y a D. Arantxa Daza Perea la composición de las Figuras 2 y 4. Este trabajo ha sido financiado por el proyecto HAR 2011-28731, Plan Nacional I+D+i del Ministerio de Econo-mía y Competitividad, Gobierno de España.
BIBLIOGRAFÍA
ABRIL LÓPEZ, D., NOCETE CALVO, F., RIQUELME CANTAL, J. A., RODRÍGUEZ BAYONA, M. e INACIO, N. (2010): “Zooarqueología del III milenio ANE: El barrio metalúrgico de Valencina de la Concepción (Sevilla)”, Complutum 21(1), pp. 87-100.
BARONE, R. (1976): Anatomie Comparée des Mam-miferes Domestiques. Ostèologie. Texte et Atlas. Vigot Frères. París.
BERNÁLDEZ SÁNCHEZ, E., BERNÁLDEZ SÁNCHEZ, M. y GARCÍA-VIÑAS, E. (2013): “¿Campos de hoyos, campos de compost? Estudio tafonómico y paleo-biológico del sector de La Gallega del yacimiento de Valencina de la Concepción (Sevilla)”, El Asenta-miento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R. eds.), Sevilla, Universidad de Sevilla, pp. 421-44.
BILLAMBOZ, A. (1979): Les vestiges en bois de cer-vide dans les gisements de l’époque holocene. Essai d’identification de la ramure et de ses diffé-rentes composantes pour l’étude technologique et l’interprétation palethnographique, L’indus-trie de l’os néolithique et de l’age du Métaux, 1 (Camps-Fabrer, H. ed.), Paris, pp. 93-129.
BOESSNECK, J. (1980): “Diferencias osteológicas entre las ovejas (Ovis aries Linne) y cabras (Capra hircus Linne)”, Ciencia en Arqueología, (Brothwell, D; Higgs, E. y Clark, G. eds.), Fondo de Cultura Económica. México, pp. 338-366.
CARDOSO, J., VILSTRUP, J., EISENMANN, V. y ORLANDO, L. (2013): “First evidence of Equus asinus, L. in the Chalcolithic disputes the Phoe-nicians as the first to introduce donkeys into the Iberian Peninsula”, Journal of Archaeological Science 40, pp. 4483-4490.
CLASON, A. T. (1972): “Some remarks on the use and presentation of archaeological data”, Helli-nium 12, pp. 139-153.
DRIESCH, VON DEN, A. (1976): A guide to the mea-surement of animal bones from Archaeological sites. Peabody Museum Bulletin 1. Harvard.
GARCÍA SANJUÁN, L. (2013): “El asentamiento de la Edad del Cobre de Valencina de la Concepción: estado actual de la investigación, debates y pers-pectivas”, El Asentamiento Prehistórico de Valen-cina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R. eds.), Sevilla, Universidad de Sevilla, pp. 21-59.
GARCÍA SANJUÁN, L., VARGAS JIMÉNEZ, J. M., HURTADO PÉREZ, V., RUIZ MORENO, T. y CRUZ-AUÑÓN BRIONES, R. (Eds.) (2013a): El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, Sevilla, Universidad de Sevilla.
GARCÍA SANJUÁN, L., LUCIAÑEZ TRIVIÑO, M., SCHU-HMACHER, T., WHEATLEY, D. W. y BANERJEE, A. (2013b): “Ivory craftsmanship, trade and social significance in the southern Iberian Copper Age: the evidence from the PP4-Montelirio sector of Valencina de la Concepción (Seville, Spain)”, Euro-pean Journal of Archaeology 16 (4), pp. 610-635.
GIFFORD, D. P. (1981): “Taphonomy and Paleoe-cology”, Advances in Archaeological Method and Theory, (Schiffer, M. ed.), Academic Press, Nueva York, pp. 365-438.
GILBERT, A. S. y SINGER, B. H. (1982): “Reases-sing zooarchaeological quantification”, World Archaeology 14 (1), pp. 21-40.
GOFAS, S., MORENO, D. y SALAS, C. (coords.) (2012): Moluscos Marinos de Andalucía. Málaga: Ser-vicio de Publicaciones e Intercambio Científico, Universidad de Málaga. Málaga.
GÓMEZ PÉREZ, J. L., BLASCO BOSQUED, C., TRAN-CHO, G., RÍOS, P., GRUESO, I. y MARTÍNEZ, Mª.S. (2011): “Los protagonistas”, Yacimientos Calcolí-ticos con Campaniforme de la Región de Madrid. Nuevos Estudios (Blasco Bosqued, C., Liesau

87
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
Von Lettow, C. y Ríos Mendoza, eds.), Patrimonio Arqueológico de Madrid, 6. UAM, pp. 100-132
GRAYSON, A. K. (1984): Quantitative Zooarchaeo-logy. Academic Press. Nueva York.
HAIN, F. H. (1982): Kupferzeitliche Tierknochenfunde aus Valencina de la Concepción, Sevilla. Studien über frühe Tierknochenfunde von der Iberischen Halbinsel nº 8. München. Institut für Palaeoana-tomie, Domestikationsforschung und Geschichte der Tiermedizin der Universität München, Múnich.
HURTADO PÉREZ, V. (2013): “Los ídolos del asenta-miento de Valencina de la Concepción (Sevilla): una revisión”, El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R., eds.), Sevilla, Uni-versidad de Sevilla, pp.311-327.
LEISNER, G. y LEISNER, V. (1943): Die Megalith-gräber der Iberischen Halbinsel. Der Süden. Römisch Germanische Forschungen 17. Berlín.
LIESAU, C. (1988): “Estudio de la Industria en Asta de Ciervo de El Soto de Medinilla”, Cuadernos de Prehistoria y Arqueología de la Universidad Autónoma de Madrid 15, pp. 183-213.
LIESAU, C. (1998): El Soto de Medinilla: Faunas de Mamíferos de la Edad del Hierro en el valle Medio del Duero (Valladolid, España), Archaeofauna, 7.
LUCIAÑEZ TRIVIÑO, M., GARCÍA SANJUÁN, L. y SCHUHMACHER, T. (2014): “Restaurierung von archäologischem Elfenbein am Beispiel von vier chalkolithischen Elfenbeinobjekte aus der Sied-lung von Valencina de la Concepción (Sevilla)”, Restaurierung und Archäologie 6, pp. 71-87.
MÁRQUEZ ROMERO, J. E. y JIMÉNEZ JÁIMEZ, V. (2010): Recintos de Fosos. Genealogía y sig-nificado de una tradición en la Prehistoria del suroeste de la Península Ibérica (IV-III milenios AC). Universidad de Málaga. Málaga.
MIGUEL DE, F. y MORALES, A. (1984): “Catálogo para una unificación de las medidas del esque-leto postcraneal de los mamíferos en España”, Primeras Jornadas de Metodología de Investi-gación Prehistórica (Soria, 1981). Ministerio de Cultura, Soria, pp. 299-305.
MORA MOLINA, C., GARCÍA SANJUÁN, L., PEI-NADO CUCARELLA, J. y WHEATLEY, D. (2013): “Las estructuras de la Edad del Cobre del Sector PP4-Montelirio del sitio arqueológico de Valencina de la Concepción-Castilleja de Guzmán (Sevilla)”, El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jimé-nez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y
Cruz-Auñón Briones, R., eds.), Sevilla, Universi-dad de Sevilla, pp. 261-279.
MORALES, A. 1976: Contribución al estudio de las faunas mastozoológicas ssociadas a yacimientos prehistóricos españoles. Tesis Doctoral Inédita. Universidad Complutense de Madrid, Madrid.
NOCETE CALVO, F., VARGAS JIMÉNEZ, J. M., SCHU-HMACHER, T. X., BANERJEE, A. y DINDORF, W. (2013): “The ivory workshop of Valencina de la Concepción (Seville, Spain) and the identification of ivory from Asian elephant on the Iberian Peninsula in the first half of the 3rd millennium BC”, Journal of Archaeological Science 40, pp. 1579-1592.
PAJUELO PANDO, A. y LÓPEZ ALDANA, P. (2013): “Estudio arqueozoológico de estructuras signi-ficativas de c/ Mariana de Pineda s/n (Valencina de la Concepción, Sevilla)”, El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R., eds.), Sevilla, Universidad de Sevilla, pp. 445-458.
PAYNE, S. (1972): “On the interpretation of bone samples from archaeological sites”, Papers in Economic Prehistory, (Higgs, E. ed.), Cambridge of the University Press, Cambridge, pp. 65-81.
PRUMMEL, W. y FRISCH, H. J. (1986): “A guide for the distinction of species, sex and body side in bones of sheep and goat”, Journal of Archaeolo-gical Science 13, pp. 567-577.
RIXSON, D. (1988): “Butchery evidence on animal bones”, Circaea 1, vol. 6, pp. 49-52.
ROBLES CARRASCO, S. y DÍAZ-ZORITA BONILLA, M. (2013): “Análisis bioarqueológico de tres contex-tos-estructuras funerarias del sector PP4-Monte-lirio del yacimiento de Valencina de la Concepción –Castilleja de Guzmán (Sevilla)”, El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R., eds.), Sevi-lla, Universidad de Sevilla, pp.369-386.
SARDÁ PIÑERO, D. (2013): “Estructuras negativas multifuncionales en Avenida de Andalucía nº 9 Valencina de la Concepción (Sevilla)”, El Asenta-miento Prehistórico de Valencina de la Concep-ción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García San-juán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R., eds.), Sevilla, Universidad de Sevilla, pp. 151-156.
SCHMID, E. (1972): Atlas of Animal Bones for Pre-historians, Archaeologists and Quaternary Geo-logist. Elsevier Science Publishers. Amsterdam.

88
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
SCHUHMACHER, T., BANERJEE, A., DINDORF, W., NOCETE CALVO, F. y VARGAS JIMÉNEZ, J. M. (2013): “Los marfiles del yacimiento de Valencina de la Concepción (Sevilla) en el contexto del Cal-colítico del suroeste peninsular”, El Asentamiento Prehistórico de Valencina de la Concepción. Investigación y Tutela en el 150 Aniversario del Descubrimiento de La Pastora, (García Sanjuán, L., Vargas Jiménez, J. M., Hurtado Pérez, V., Ruiz Moreno, T. y Cruz-Auñón Briones, R., eds.), Sevi-lla, Universidad de Sevilla, pp. 495-510.
VARGAS JIMÉNEZ, J. M. (2004): “Elementos para la definición territorial del yacimiento prehistórico de Valencina de la Concepción (Sevilla).” Spal. Revista de Prehistoria y Arqueología 12, pp. 125-144.
VARGAS JIMÉNEZ, J. M., MEYER, C. y ORTEGA GORDILLO, M. (2012): “El tholos de La Pastora y su entorno: el sector oriental del yacimiento de Valencina de la Concepción (Sevilla) a través de
la geofísica.” Menga. Revista de Prehistoria de Andalucía 3, pp. 121-140.
VARGAS JIMÉNEZ, J. M., NOCETE CALVO, F. y SCHU-HMACHER, T. (2012a): “Contextos de producción de objetos de marfil en Valencina de la Concep-ción (Sevilla)”, Marfil y Elefantes en la Península Ibérica y el Mediterráneo. Actas del Coloquio Internacional en Alicante (26-27 de Noviembre de 2008), (Banerjee, A., López Padilla, J. A. y Schuh-macher, T. eds.), Mainz. Deutsches Archäologis-ches Institut y Museo Arqueológico de Alicante, pp. 69-82
WHEATLEY, D. W., STRUTT, C., GARCÍA SANJUÁN, L., PEINADO CUCARELLA, J. y MORA MOLINA, C. (2012): “New evidence on the spatial organisation of the Valencina de la Concepción Copper Age settlement: the geophysics between La Pastora and Montelirio.” Trabajos de Prehistoria 69 (1), pp. 65-79.
APÉNDICE 1. FAUNAS PROCEDENTES DE LOS HORIZONTES NO CALCOLÍTICOS DE MON-TELIRIO-PP4, DESGLOSADOS POR ESPECIES.
GRUPO PREHISTÓRICO ROMANO-MEDIEVAL TOTAL
MAMÍFEROS 31 54 85
MOLUSCOS MARINOS 5 8 13
NÁYADES 2 4 6
MOLUSCOS TERRESTRES 8 45 53
MOLUSCOS INDET 0 3 3
MAMÍFEROS INDET 29 51 80
INDUSTRIA ÓSEA 2 1 3
TOTAL 77 166 243
Número de restos (NR) de faunas procedentes de los horizontes no calcolíticos de Montelirio-PP4, desglosadas por grupos.
ESPECIEPREHISTÓRICO MEDIEVAL
NR % NMI % PESO % NR % NMI % PESO %
Bos taurus 13 38,2 6 33,3 379 45,2 11 20,4 1 8,3 230 41,1
Ovis aries 3 5,5 3 25,0
55,4Ovis/Capra 10 29,4 4 22,2 109 13,0 33 61,1 3 25,0 310
Capra hircus 4 7,4 4 33,3
Sus sp. 4 11,8 3 16,7 16 1,9 3 5,5 1 8,3 20 3,5
Equus sp. 1 2,9 1 5,6 30 3,6
Bos primigenius 2 5,9 1 5,6 95 11,3
Cervus elaphus 3 8,8 2 11,1 170 20,3
Sus scrofa 1 2,9 1 5,6 39 4,7
TOTAL 34 100 18 100 838 100 54 100 12 100 560 100
Número de restos (NR), individuos (NMI) y peso de los mamíferos de los horizontes no calcolíticos de Montelirio-PP4, desglosados por especies.

89
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
APÉNDICE 2: RELACIÓN DE LA FAUNA ESTUDIADA SEGÚN ESTRUCTURAS.
VALENCINA DE LA CONCEPCIÓN PP4 DJ 07/23
CONTEXTOS FUNERARIOS UE TIPO DE ESTRUCTURA E
INDIVIDUOS INHUMADOSGRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
CONTEXTOS FUNERARIOS CALCOLÍTICOS
10.012 246 e. neg. sin piedra NMI=1 Molusco marino
1 1 Pecten cf. jacobeus.1 valva D completa, aunque el borde ventral algo fracturado y falta la orejuela posterior algo fracturada. AM- (( 90 )). El borde la charnela presenta algo de pigmento rojo. Concrecionada.
10.015-21-61 416(9)
e. neg. con lajas y piedras tumba colectiva. NMI=13
Pulmonado 1
1
1
1
Candidula gigaxii? NMI=11 ejemplar adulto pequeño.Helicellino spp.?1 ejemplar adulto completo.
10.016 365 e. neg. sin piedra. NMI=4 Molusco marino
1 1 Pecten maximus. 1 valva D completa bien conservada. AM -(118); H- (110). concrecionada
10.022 935 e.neg. con solo 1 laja de pizarra hincada en el interior de la estructura.
NMI=2
Molusco marino
1 1 Pecten maximus.1 valva D completa bien conservada. AM -(140); H -((122)). Ejemplar de gran talla.
10.023 526 e. neg. con lajas o pie-dras pequeñas cubriendo
la estructura
Molusco marino
1 1 Pecten sp.1 valva D fragmentada en 3 piezas, concreciones. AM -((92)).
524 e. neg. funeraria con elementos pétreos. B5: nivel inferior de
enterramientoNMI=23
Bos taurus 7 1 (2,5-3 años), subad.
150 Dientes: 5 Molares superiores, 1 M1 S (++/+++); 1 M2 S (+/++); 1 M3 S (+); 1 M2 S (+/++); 1 . M3 D (+); todos con fractura reciente en la raíz.1 P4 D (+) inferior. Parece corresponder todo a un solo individuoMandíbula: 1 frag. de rama horizontal muy alterado en superficie y con fractura reciente.
524 SIN IDENTIFICAR
1 15 Macromamífero: 1 e. d. muy alterada, craqueladuras y fractura reciente.
424 Pulmonado 1 1 Otala lactea.1 ejemplar adulto, descolorido, con una rotura en la última vuelta (la rotura es grande por la parte de abajo).
10.031 453 e. neg. excavada en la roca
Humano 7 Cráneo: 7 frag. craneales del individuo 2.
10.031 453 Cráneo 1 Pulmonado 1 1 Helicellino spp.?1 fragmento del ápice.
10.031 453 Individuo 2 Pulmonado 1 1 Helicellino spp.?1 fragmento del ápice.
10.031 453 Cráneo 2 Microfauna 1 1 Micromamífero:1 posible metacarpo de roedor
10.032 447 e. neg. megalítica: NMI=8 Pulmonado 4 4 Rumina decollata.1 ejemplar adulto con rotura en la boca. 1 ejemplar juvenil entero.Otala lactea.2 ejemplares adultos, descoloridos. El ejemplar grande presenta rotura en el ápice, parte de la vuelta de espira y en la columnela. El ejemplar pequeño tiene el ápice hundido y presenta rotura siguiendo la vuelta de espira, pero conservando el ápice dentro.
10.032 511 e. meg. funeraria SIN IDENTIFICAR
1 5 Macromamífero: frag. dist. de falange? quemado (negro).
10.034 500 e. megalítica con lajas de pizarra sin función sus-
tentadora. UE asociada al ajuar de los inhumados:
NMI= 7
Molusco marino
1 1 Pecten sp.1 valva D algo fracturada en el borde de tamaño mediano AM -(aprox. 90).Posible Pecten jacobeus.
500 (12)
Molusco marino
1 1 Pecten cf. maximus.1 valva D completa bien conservada aunque con los bordes erosionados. AM- (117);H -((113))
10.034 499 Cráneo 1 Pulmonado 21 20 Theba pisana.1 ejemplar pequeño.Helicellino spp.?19 ejemplares indeterminados. 1 fragmento indeterminado.
10.034 499 Cráneo 2 Pulmonado 7 7 Ferussacia follicula.2 ejemplares, un adulto y un juvenil.Xerotricha madritensis?5 ejemplares.
10.034 499 Cráneo 3 Pulmonado 3 3 Candidula gigaxii.3 ejemplares adultos.

90
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
CONTEXTOS FUNERARIOS UE TIPO DE ESTRUCTURA E
INDIVIDUOS INHUMADOSGRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
10.034 499 Cráneo 4 Pulmonado 1 1 Rumina decollata. 1 ejemplar pequeño roto.
10.034 499 Cráneo 6 Pulmonado 20 20 Rumina decollata. 2 ejemplares juveniles.Theba pisana.4 ejemplares juveniles.2 ejemplares?Candidula gigaxii.2 ejemplares adultos.Xerotricha madritensis.8 ejemplares?Xerotricha apicina.2 ejemplares.
10.034 499 Cráneo 7 Pulmonado 4 4 Xerosecta pomissa.1 ejemplar adulto pequeño.Helicellino spp.?3 ejemplares.
499 Pulmonado 1 1 Ferussacia follicula.1 fragmento grande de la zona de la boca.
10.034 499 Pulmonado 2 2 Helicellino spp.?2 ejemplares inderteminados
10.036 512 e. neg. no megalítica con lajas de pizarra hincadas en el corredor. UE es el
ajuar asociado a los inhu-mados: NMI=3
Molusco marino
1 1 Pecten cf. maximus.1 valva D con fractura reciente de los bordes cercanos a las orejuelas y las orejuelas.
Hay foto de la excavación.
10.037 515 e. neg. con lajas o pie-dras pequeñas cubriendo la estructura. UE asocia-
da al ajuar. NMI=3
Molusco marino
1 1 Pecten sp. 1 valva posterior D.
10.038 529(1)
e. neg. sin piedra. UE asociada al ajuar. NMI= 3
Molusco marino
1 1 Cymbula nigra. Industria ósea.1 completa. Gran tamaño, pero no fósil. Concha trabajada y rebajada en ambos bordes para crear una pieza ovalada más o menos simétrica. La zona anterior ha sido agudizada para crear un pequeño apéndice de morfología semicircular. L-90.Hay foto de la excavación.
10.038 529(2)
Molusco marino
1 1 Pecten cf. maximus.1 valva completa D bien conservada AM-91,6, (H- 89). Concre-cionada.
10.040 565 e. neg. sin piedra. UE asociada al ajuar. NMI=3
Molusco marino
1 1 Pecten sp.1 valva de pequeño tamaño. AM-67,3; H-63.
10.042 211 e. megalítica con lajas de pizarra delimitando el espacio sin función
sustentadora. UE asociada al ajuar de la
primera cámara. NMI en la primera cámara=5
Molusco marino y
Pulmonado
1
2
1
2
Charonia lampas.1 frag. de ápice, se ve la columella y con fractura antigua de las primeras vueltas. Presenta al menos tres perforaciones que requieren de una exhaustiva limpieza para averiguar su posible origen.
Otala lactea.1 ejemplar adulto descolorido con algo de rotura en la boca.Rumina decollata.1 ejemplar adulto con algo de rotura en la boca.
640 Inhumación de varón joven con un colmillo de elefante en nivel inferior y un ajuar de extraordi-
naria riqueza
Cervus elaphus
17 1? (380) Asta: 17 fragmentos en pésimo estado de conservación, al menos tres piezas parecen haber sido modificados antrópicamente y presentan una superficie totalmente alisada y un reborde peri-metral. Posiblemente se trate de un rebaje de la roseta o ha sido creado artificialmente. Varias están además termoalteradas por contacto con fuego. El material está tan alterado que no resulta posible saber si se trata de un asta mudada. La extracción no ha sido completa, lo que dificulta la reconstrucción de las piezas con un mínimo de fiabilidad.
Ovis aries 2 1 79 Cuerno: 2 frag. de clavija ósea de un carnero.
640 SIN IDENTIFICAR
8 45 4 e. d. de macro y 4 de meso.
Pulmonado 4 2 Helicellino spp.?1 fragmento del ápice. 2 fragmentos indeterminados.Xerotricha madritensis.1 ejemplar?

91
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
CONTEXTOS FUNERARIOS UE TIPO DE ESTRUCTURA E
INDIVIDUOS INHUMADOSGRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
10.042 615 Oryctolagus cuniculus
2 1 1 Húmero: 1 e.d.Metatarso: 1 frag. de metatarso III prox.Por el estado tan fragmentario parece intrusivo.
615 Pulmonado 5 4 Rumina decollata. 2 ejemplares adultos.Helicella sp.1 ejemplar de pequeño tamaño, rota por un lateral.Helicellino spp.?1 ejemplar algo fragmentado. 1 fragmento indeterminado.
648 Pertenece a la 10.042, la primera de las dos
cámaras de esa estructu-ra megalítica.
Molusco marino y
Pulmonado
1
2
1
1
Pecten sp.1 frag. de valva con fracturas antiguas.
1 ápice pequeño indeterminado.1 frag. indeterminado.
10.049 667 e. neg. sin piedra. Humano 7 Parece que todos los restos son humanos, uno de ellos con huellas de descarnado.
“Interior” de las dos cámaras de que consta este gran monumento
megalítico.
Molusco 1 1 Molusca indet. 1 Pieza de reducido tamaño muy fragmentada.
10.042-10049 211 Megalito calcolítico Ajuar asociado los
inhumados de la primera cámara.
Molusca 1 Molusca indet. 1 frag. muy pequeño.
SIN IDENTIFICAR
2 5 1 e.d. de macromamífero y otra de mesomamífero.
648 Junto con la UE 211, la UE 648 corresponde al “ajuar asociado los
inhumados de la primera cámara.
Bos taurus 5 1 36 Dientes: 1 frag. de molariforme y un frag. de raíz de molariforme quemada (marrón-negro).Escápula: 4 frag. de lámina.Vértebra: 1 frag. de proceso espinoso con fractura reciente.Pelvis: 1 frag. de acetábulo.
Ovis aries/Capra hircus
2 2: 1 su-bad-ad y 1 inf.
15 Dientes: 1 M1/M2 superior (++) (2-4 años).Radio: 1 diáfisis de un animal de talla reducida (infantil).
SIN IDENTIFICAR
18 507 Macro: 18 e.d. y frag. de vértebras con fractura reciente.Mesoungulado: 9 (3 láminas costales y 6 e.d.).
Industria ósea
5 1 frag. de luchadera de ciervo con un reborde, ¿enmangue?1 frag. de escápula de bovino? Muy concrecionado y superficie alterada.1 e.d. de macro.1 hueso de macro. con un rebaje a modo de ranura semicircular.1 lámina costal con pulimento con fractura antigua.
Humano? 2 3 e.d?
10.067 907 e. neg. sin piedra. UE asociada al ajuar.
NMI=6
Molusco marino
5 5 Pecten sp.1 frag. de valva D con charnela AM-59.1 frag. ventral de valva D. NMI=2.Glycymeris sp.1 valva casi completa de talla mediana (no medible). NMI= 1Cymbula nigra. Industria ósea.1 completa. Posible rebaje antrópico en la zona anterior con tierra en su interior.L- 62,5. NMI=1. Eastonia rugosa (Mactridae).1 Valva casi completa (Hay foto de la excavación).
10.073 867 e.neg. con lajas o piedras pequeñas cubriendo la
estructura. UE asociada al ajuar. NMI= 3
Molusco marino
1 1 Cymbula nigra. Industria ósea. Pieza completa. Rebaje antrópico en toda la zona anterior.(L- 62,5). NMI=1. Pulimento en uno de los bordes como posible zona de agarre. Probable “uso en vida” de esta pieza como pocillo o contenedor.
373 Pulmonado 6 1 Otala lactea.1 ejemplar adulto, con rotura en el ápice y en las primeras vueltas.5 fragmentos pequeños.
10.074 929 e. neg. sin piedra. UE asociada al ajuar.
NMI=1
Molusco marino
1 1 Pecten cf.maximus.1 frag. D dorsal de un ejemplar de talla grande.
10.081 952 e. neg. Sin el. pétreos. UE los individuos inhumados.
NMI=2
Molusco marino
1 1 Cymbula nigra.1 pieza incompleta, en el interior presenta manchas de colorante rojo (¿cinabrio?). Borde posterior posiblemente rebajado de for-ma semicircular. Fractura reciente. Pieza en estrecho contacto con los esqueletos.

92
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
CONTEXTOS FUNERARIOS UE TIPO DE ESTRUCTURA E
INDIVIDUOS INHUMADOSGRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
10.083 871 e.neg. con lajas o piedras pequeñas cubriendo la
estructura. UE asociada al ajuar. NMI=8
Molusco marino
2 2 Pecten cf.maximus.1 frag. D de valva casi completa de un ejemplar de talla grande (no medible).1 frag. D de valva casi completa de un ejemplar de talla media (no medible).
10.083 871 (4)
Molusco marino
1 1 Pecten cf.maximus.1 frag. D de valva completa de un ejemplar de talla grande con restos de cinabrio en su borde ventral. Parece pieza rodada, y recogida en la playa. AM-122,7; H-113,5.
10.088 237 Cámara con corredor: e. neg. con piedras
pequeñas o medianas en el corredor. UE
corresponde a un cráneo seccionado.Material muy alterado y fracturado de antiguo y por el proceso
de extracción.NMI=1
Bos primigenius
3 1 112 Radio: 1 porción prox. D y dos esquirlas de diáfisis de gran talla y peso.Húmero: 1 e.d., con fracturas y abundantes fisuras postdeposi-cionales.
Bos taurus 3 194 Diente: 1 frag. de M1/M2 inferior (+/++) de juv-subad.Escápula: 1 pieza S muy incompleta con parte de la cavidad glenoidea, espina y lámina (15 frag.).Húmero: 1 porción distal de diáfisis con fractura reciente.
Sus sp. 2 6 Diente: 1 canino inferior S con fractura antigua y reciente.Húmero: 1 e.d. muy alterada en superficie.
Canisfamiliaris
3 1 12 Ulna: 1 frag. de porción articular y diáfisis muy alterado.Tibia: 2 frag. de diáfisis proximales D y S de un individuo de talla más bien reducida, con fractura reciente (probablemente de inf-juv).
SIN IDENTIFICAR
12 39 Mesofauna: e.d. y de epífisis (3 con huellas de combustión, negro).
Molusco Marino
17
6
4
2
Ruditapes cf. decussatus.1 valva casi completa D. AM -(57) H-(37,7).2 frag. De valva D.3 frag. de valva S.2 frag. S con charnela.9 frag. de valva .NMI=2BIVALVIA INDET.2 frag. de valva de un individuo de gran talla.1 frag. de valva.3 frag. de valva quemados de un individuo de gran talla.NMI= 2

93
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESTRUCTURA UE TIPO DE ESTRUCTURA E INDIVIDUOS INHUMADOS
GRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
ESTRUCTURAS CALCOLÍTICAS NO FUNERARIAS
10.024 333 “Pequeño recinto circular” de unos 18 m de diámetro. Con varias estructuras negativas
dentro de la estructura 10.024.
Bos taurus 2 1 2 Mandíbula: 1 frag. de rama ascendenteVértebra cervical: 1 frag.
Cervus elaphus
2 1 20 Asta: 1 frag. de luchadera quemado (gris).Pelvis: 1 frag. de íleon.
Ovis aries 1 1 1 Astrágalo: 1 pieza algo erosionada en superficie.
Ovis aries/Capra hircus
4 1 25 Mandíbula: 1 frag. de rama horizontal D.Diente: 1 M1/M2 (++) y 1 molariforme.Radio: 1: e.d. quemada (color gris).
SIN IDENTIFICAR
13 61 Esquirlas óseas muy alteradas, 4 quemadas (negro).
Industria ósea
1 1 espátula realizada a partir de la diáfisis de un metapodio de bovino.
Una de las zanjas del recinto, con medio millar
de piezas cerámicas, algunas a torno. Calcolí-tico con intrusiones más
recientes.
Moluscos marinos
y de agua dulce
4
5
3
5
Ostrea sp.1 Frag. de valva de pieza con los bordes redondeados (útil?), muy concrecionada. NMI=1.Glycymeris sp.1 frag. valva de NMI=1.Ruditapes sp. NMI=1.1 frag. de valva anterior con charnela.1 frag. de valva ventral.Potomida litoralis. NMI=5.5 frag. anteriores y 1 cardinal (2D y 4 S).
554 Bos taurus 27 3 (1 ad. y 1 inf)
1070 Neurocráneo: 3 frag.Viscerocráneo: 1 con M1/M2 (++).Vértebra cervical: 1 frag.Dientes: 2, 1 molar superior (+/++) y 1 molariforme.Mandíbula: 2 esquirlas de rama horizontal S. 1 D+S completa con M3 (++) (4-6 años), rama vertical fracturada y con huellas de combustión (negro), todo extraído en bloque de la excavación.Escápula: 1 muy fracturada colocada entre las dos ramas.Radio: 1 frag. de diaf. S grande, ♂ ?Pelvis: 1 frag. S.Costilla: 8 frag. lámina.Astrágalo: 1 S con dos percusiones en la cara medial.LML (71,9) LMM (63,7) AC (48,8), GLM (38,7).FI: 5 todas fusionadas menos una de talla muy pequeña (inf.).LMP- (58) - 63,8 AP- (35) - - AMD- (30) (28,8) 27,9AD (31,2) - 33,9a/p a ? a
10.024 Ovis aries/Capra hircus
2 1 20 Radio-ulna: 1 frag.Tibia: 1 e.d.
10.024 Sus sp. 1 1 15 Pelvis. 1 frag.
10.024 554 SIN IDENTIFICAR
20 105 e.d. de mesomamífero (1 quemado, negro).
10.024 148 Humano 1 malar .
10.024 148 Equus sp. 1 1 55 FI: 1 pieza con varias fracturas recientes y huellas de combus-tión en la cara dorsal (negro).
10.024 148 Bos taurus 6 1 35 Dientes:4: 3 frag, de molariformes y 1 M3 (-) (juv).Costillas: 1 frag.de lámina.Ulna: 1 frag. S de porción articular con diáfisis distal.
10.024 148 Bos primigenius
2 1 553 Escápula: 2 bastante completas, probablemente del mismo individuo, muy fragmentadas por la extracción, con fisuras y al-teraciones propias de una exposición prolongada a la intemperie. Medidas de la escápula D: LD- ((90.7)); LMC-(72,1); la escápula S: LMC-72,1; AA-62,3.
10.024 148 Ovis aries/Capra hircus
12 2 45 Dientes: 3 frag. de molariformes (1 M1/M2 (+); otro con (++) y 1 frag. de raíz).Viscerocráneo: 1 frag. de premaxilar D.Mandíbula: 1 frag. rama vertical S (zona 4).Radio: 2 e.d.Metapodio: 1 e .d.Pelvis: 3: 1 ileon S 1 frag. de pubis D y 1 frag. de isquion.Centrotarsal: 1 S.
10.024 148 Industria ósea
2 1 hueso largo de mamífero: ¿punta de un punzón?1 pieza en asta de ciervo: 1 punta de luchadera con huellas de uso.

94
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESTRUCTURA UE TIPO DE ESTRUCTURA E INDIVIDUOS INHUMADOS
GRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
10.024 148 Molusco marino
Pulmonado
7
17
5
9
Ruditapes cf. decussatus.1 valva completa D de. AM (49) H (32).2 frag. Anteriores D con charnela.1 frag. S con charnela.3 frag. de valva D. NMI=4.Ostrea sp. 1 frag. de valva. NMI= 1.Otala lactea. 7 ejemplares descoloridos (2 con rotura en ápice y 2 con rotura en la boca).Theba pisana.1 ejemplar adulto.1 columnela.8 fragmentos pequeños indeterminados.
10.024 594 Molusco agua dulce
2 2 Unio delphinus.2 valvas casi completas D, una conserva el relleno.
558 SIN IDENTIFICAR
1 1 4 Escápula: 1 frag. de lámina de macromamífero.
10.024 547 contiene dos frag., cerá-micos hechos a torno.
Bos taurus 16 2(1 juv)
159 Dientes: 4, 1 molariforme inferior, 1 pd 4 (++) (6-18 meses), 1 incisivo. 1M1 /M2 superior muy alterado en superficie.Atlas: 1 fragmento.Costillas: 2 frag. de lámina.Húmero: 4 e.d.Metapodio: 3 e.d. Radio: 2 e.d. muy alteradas por exposición a la intemperie y erosión radicular.
10.024 Ovis aries 1 1 Metacarpo: 1 porción proximal D. AP: 22.
Ovis aries/Capra hircus
13 2(1 subad y 1 ad)
86 Dientes: 5, superiores: 1 M1/M2 y 1 P3 (++). Inferiores: 1 frag-mento de M3 (+/++) (2-4 años, subad) y un M1/M2 (++/+++).Mandíbulas: 2 S, una con P4 (+).Vértebra: 1 cervical.Metacarpo: 1 fragmento de porción articular distal.Escápula: 1 frag dist. sin cavidad glenoidea.Costilla: 1 frag. de lámina.Fémur: 1 e.d.Húmero: 1 e.d.
10.024 Sus sp. 9 2(1 inf y 1 juv.)
71 Neurocráneo: 2:1 frag. de frontal y 1de occipital.Maxilar: 2: 1 frag. con un incisivo de leche y 1 frag. de maxilar S con M1 (+/++) y M 2(+) (1,5-2 años).Mandíbula: 1rama horizontal con incisivos de leche.Escápula: 1 porción distal (infantil).Radio: 1porción distal epífisis (-) (inf-juv).Metapodio: 1porción distal epífisis (-).Metacarpo: 1 Mtc V con epífisis distal (-).
10.024 Gallus gallus
1 1 Tibiotarso: 1 porción distal D.
10.024 SIN IDENTIFICAR
40 100 Esquirlas de huesos muy alteradas en superficie.
10.024 560 SIN IDENTIFICAR
1 Macro: 1 e.d. muy alterada erosión múltiple.
627 e. neg. de planta circular.
Molusco marino
1 1 Ruditapes sp.1 frag. de valva.
631 e. neg. de planta circular con piedras de molino,
lítica y cerámica.
Molusco marino
3 3 Ruditapes sp.1 frag. de valva.Cerastoderma edule.1 valva en 4 fragmentos y a falta de umbo.Antalis sp.1 pieza completa.
656 e. neg. de planta circular. Molusco marino
1 1 Ruditapes sp.1 frag. de valva.
663 e. neg. de planta de planta irregular.
Molusco marino
1 1 Ruditapes sp.1 frag. de valva.
703 e. neg. de planta circular con cerámica y 4 piedras,
una con cazoletas.
Molusco marino
1 1 Charonia lampas. 1 completa, Anchura de la boca: 55; L de la boca: 90. Rellena de tierra. Cierta erosión por abrasión en la playa.
704 e. neg. de planta circular. Molusco marino
1 1 Pecten sp.1 frag. de valva S con los bordes erosionados, tal vez por su recogida en la playa o por acción antrópica.
908 e. neg. de planta circular. Moluscos de agua dulce y marinos
2 2 Unio delphinus.1 frag. borde post. D.Spisula cf elliptica.1 valva en tres fragmentos.
944 e. neg. Molusco marino
2 2 Ruditapes sp.1 frag. de valva y 1 umbo.

95
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESTRUCTURA UE TIPO DE ESTRUCTURA E INDIVIDUOS INHUMADOS
GRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
ESTRUCTURAS PREHISTÓRICAS DE CRONOLOGÍA DUDOSA O CON INTRUSIONES POSTERIORES
10.019 363 e.neg. con intrusiones romanas.
Bos taurus 1 1 30 Escápula: 1 frag. dist. con cavidad glenoidea, muy alterada por el proceso de extracción. Fracturas recientes.
10.019 363 Ovis aries/Capra hircus
1 1 juv. 50 Mandíbula: 1 rama horizontal D con serie dentaria P3-M3 (M2 (+);M3 (+/-): 1,5-2 años, juv. Pieza muy concrecionada y alterada postdeposicionalmente. Fractura reciente.
10.019 363 Cervus elaphus
1 1 150 Cráneo: 1 frag. de frontal y occipital (fracturado en 4 piezas) de un macho, cuyos pedículos presentan superficies de fractura irregulares. Las concreciones impiden ver detalles. ¿Posible modificación intencionada?
10.019 363 SIN IDENTIFICAR
2 5
10.001 20 ¿? Molusco marino y de agua dulce
2
1
3
1
Pecten sp. 4 frag. de 1 valva S.Ruditapes sp.4 frag. de 1 valva.Bivalvia (Unionidae) indet.1 frag. de valva.
SIN IDENTIFICAR
2 4 Macro.: 1 e.d.Meso.: 1 e. d. todos muy alterados por procesos de disolución.
10.007 55 ¿? Bos taurus 9 2 280 Viscerocráneo: 1 frag. de premaxilar.Vértebra lumbar: 3 frag., uno con proceso espinoso. ant. (+/- ).Costilla: 1 frag. de lámina.Sacro: 3 sacros muy fracturados.Vértebra caudal: 1 frag de cuerpo (ant. y post. (-)).
10.007 55 Sus sp. 1 1 (inf.-juv)
5 Metacarpo: 1 metacarpo III D. Epif. dist (-); inf-juv.
167 e. neg. prehistórica. Molusco marino
1 1 Pecten sp. 2 frag. de valva D. Pieza muy alterada, posiblemente por proc. posdeposicionales.
SIN IDENTIFICAR
1 4 1 e. d. macromamífero muy alterada.
10.111 373 e.neg. prehistórica. Bos primigenius
2 1 95 Costilla: 1 frag. de porción articular de gran talla con concrecio-nes y leve combustión en la cara externa.Tibia: 1 frag. de porción dist. D muy alterada con huellas de combustión y alteración postdeposicional.
Bos taurus 2 21 subad;
1 su-bad-ad.
35 Dientes: 2 M3 inferiores S; 1 con (+/-) y otro con un desgaste de (+/++).NMI= 2: 1 de 2,5-3 años. 1 de 3-5 años.
10.111 373 Sus sp. 2 1 5 Dientes: 2 frag. de molariformes (+/++).
10.111 373 Cervus elaphus
1 1 20 Metatarso: 1 frag. de diáfisis dist. muy alterado cuya línea sagital presenta un surco muy profundo y una morfología en la cara dorsal con tendencia circular. Las alteraciones postdeposio-nales en la pieza son muy acusadas y afectan a toda la diáfisis. Erosiones múltiples y fisuras de la diáfisis con desplazamiento de su trayectoria, por lo que no resulta claro si podría tratarse de un material descartado de la industria ósea, un útil o un mero desecho alimentario. Presenta leves huellas de combustión.Asta: dos frag. de asta principal muy alterados en superficie (no se pesa).
Equus sp. 1 1 30 Diente: 1 molar inferior D, parcialmente quemado (negro), concrecionado y con fractura reciente.
SIN IDENTIFICAR
3 5 Macro: 1 e.d. de hueso largo muy alterado en superficie.Meso.: 1 e.d., tibia o/c?
377 Pulmonado 8 2 Otala lactea. 1 ejemplar adulto, bastante descolorido.1 ápice con varias vueltas de espira.6 fragmentos de esta especie de los que 2 son grandes laterales, 1 de la base de la columnela y el resto más pequeños laterales.
10.0111 620 Humano 48 A- apófisis estiloides y temporal? D con leve huella de combustión.B20 -restos craneales en mal estado y 1 lámina costal combus-tionada (negro).
10.069 562 Molusco marino
1 1 Pecten cf maximus.1 frag. de valva de un individuo de gran talla. ¿Posible rebaje intencionado en un lateral?
10.006 600 ¿? Bos taurus 1 1 34 Radio: 1 frag. prox. S muy alterado con fractura reciente.
10.006 600 Sus scrofa? 1 12-3
años subad
39 Viscerocráneo: 1 maxilar D con P4, M1 fragmentados y algo quemados y M3 incompleto (+). Am (19).Por la talla y peso de la pieza es probable que corresponda a un jabalí.
10.006 600 SIN IDENTIFICAR
6 34 Macro.: huesos craneales muy alterados. 2 piezas con leve pulimento.

96
CORINA LIESAU VON LETTOW-VORBECK ET AL.
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESTRUCTURA UE TIPO DE ESTRUCTURA E INDIVIDUOS INHUMADOS
GRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
607 B-17
¿¿ Ovis aries/Capra hircus
6 1 subad –ad.1 ad.
43 Dientes: inferiores 2: 1 M3 S y otro M3 D ( +/++) 2-4 años subad.-ad.Superiores: 2 M3 con unos desgastes irregulares (++) ad.Tibia: 1 e.d. con fisuras y erosión radicular.Costillas: 1 frag. de lámina .
607 B-17
SIN IDENTIFICAR
11 47 Esquirlas óseas muy alteradas por erosión radicular.
614 SIN IDENTIFICAR
2 4 Meso.: 3 e.d. muy alteradas.Macro: 1 frag. de lámina costal.
630 SIN IDENTIFICAR
1 8 Macro: 1 esquirla de diáfisis combustionada (color gris-blan-quecino).
10.058 724 Humano 3: 2 ad. y 1 inf.
Bolsa y conjunto único de huesos craneales y apendiculares de al menos 3 individuos. Por los radios, se trata de dos adultos, posiblemente 1 de ellos un varón muy robusto y otro 1 infantil. Parece tratarse de un enterramiento secundario.
10.111 745 (9)
Ovis aries/Capra hircus
3 1 (2-4 años)
16 Dientes: 2 superiores: M1/M2 con (+/++). 1 inferior: 1 M3 (+). Piezas muy alteradas por erosión radicular y probablemente por también por procesos postdepo-sicionales.
10.111 745 Industria ósea
2 (78) Escápula: 1 lámina escapular de Bos sp. con la espina rebajada por uso?. Pieza muy alterada.Fémur?? 1 e.d. con pulimento, acusada alteración radicular .
10.111 745 (9)
Molusco marino
1 1 Ostrea edulis.1 aglomerado con al menos 3 individuos. No se descarta que sea fósil.
10.111 745 Sus sp. 1 1 6 1 M3 inferior S (++/+++). LM-32,2; AM- 9,3.
10.111 777 SIN IDENTIFICAR
1 2 1 e.d. mesomamífero.
919 Molusco de agua dulce
1 1 Potomida litoralis. NMI=1.1 anterior S, talla grande.
10.074 923 Humano 2 2 frag. de calota craneal con restos de pigmento rojo que parece cinabrio.
ESTRUCTURAS ROMANAS/MEDIEVALES DESCONOCIDAS
384 e. neg. de planta circular romana o moderna.
Moluscos marinos
y de agua dulce
4 4 Unio delphinus.1 frag. ventral y 1 valva casi completa.Acanthocardia cf ecchinata.1 frag. de valva incompleta.Murex brandaris.1 pieza con la última vuelta y sifón fracturados. Fracturas recientes.
75 e. neg. de planta circular medieval.
Molusco marino
1 1 Glycymeris sp. Industria ósea.1 valva S con umbo fracturado y también fractura irregular en toda la superficie del borde. Zona centro de la valva con cierto pulimento, ¿alisador?
736b e.neg. de planta circular medieval.
Molusco marino y
Pulmonado
5
2
5
2
Cerastoderma edule.1 valva completa pero abrasionada (recogida en la playa).BIVALVIA.3 frag. de valvas, una de ellas desconchada, lo que no permite su identificación. La otra pieza parece un borde, pero también está abrasionada en todos sus lados.Glycymeris glycymeris.1 frag. de valva con el umbo fracturado de antiguo, pieza abrasio-nada (recogida en la playa).Otala lactea. NMI=2.2 ejemplares adultos descoloridos.
10 ¿? Pulmonado 9 7 1 ápice de bivalvo indeterminado. Xerotricha madritensis?1 ejemplar adulto grande completo.2 ejemplares pequeños juveniles.Candidula gigaxii.2 ejemplares adultos pequeños.Rumina decollata.2 ejemplares juveniles.1 fragmento de la boca.
72 ¿? Pulmonado 1 1 Rumina decollata.1 ejemplar adulto completo.
73 ¿? Pulmonado 1 1 Otala lactea.1 ejemplar adulto completo poco descolorido.
83 Pulmonado 20 9 Otala lactea.4 ejemplares adultos completos.1 ejemplar sin ápice.1 fragmento grande de la zona del ombligo.Theba pisana.4 ejemplares adultos. (2 ejemplares sin ápice + 2 ápices sueltos).

97
LA FAUNA DEL SECTOR PP4-MONTELIRIO DEL YACIMIENTO PREHISTÓRICO DE VALENCINA DE LA CONCEPCIÓN
MENGA. REVISTA DE PREHISTORIA DE ANDALUCÍA // Nº 05. 2014. PP. 69-97. ISSN 2172-6175 // DOSSIER
ESTRUCTURA UE TIPO DE ESTRUCTURA E INDIVIDUOS INHUMADOS
GRUPO/TAXON NR NMI PESO
EN GR DESCRIPCIÓN DE LOS RESTOS
112 ¿? Pulmonado 2 2 Theba pisana.1 ejemplar adulto.Xerosecta cespitum.1 ejemplar adulto descolorido, con bandas.
436 ¿? Pulmonado 1 1 Rumina decollata.1 ejemplar adulto.
470 Desconocida prehistórica Pulmonado 1 1 Theba pisana.1 ejemplar juvenil.
480 ¿? Pulmonado 2 2 Xerosecta cespitum.2 ejemplares adultos.
628 ¿? Pulmonado 2 0 Otala lactea.1 fragmento grande de la zona del ombligo.1 ápice.
628b Pulmonado 1 1 Theba pisana.1 ejemplar adulto descolorido.
521 Pulmonado 1 1 Otala lactea.1 ejemplar adulto.
10.027 364 ¿? Molusco marino
2 2 BIVALVIA.1 frag. de valva.Ruditapes sp.1 frag. de valva.
329 SIN IDENTIFICAR
1 9 1e.d. macro.
624 Molusco de agua dulce
2 1 Unionidae.2 frag. ventrales de posible Potomida.
736b Pulmonado 2 2 Otala lactea. 2 ejemplares adultos descoloridos.
10.101 544 Bos taurus 11 1 230 Costilla: 7 frag. de lámina quemados (4 negro y 1 gris-marron).Escápula: 2 S: 1 frag. de lámina y 1 frag. de arranque de espina.Pelvis: 1 acetábulo S con isquion percutido y rebajado.FII: 1 quemada (marron-negro).
10.101 544 Ovis aries 3 1♂+ 1♀ 310 Cuerno: 1 frag.de clavija ósea S de carnero.Neurocráneo: 1 frag de una hembra mocha.Fémur: 1 e.d.S.
10.101 544 Ovis aries/Capra hircus
33 3: (1 ad., 1 juv/
subad, 1 subad/
ad.)
Neurocráneo: 2: frag. de frontal con huella percusión con instru-mento metálico para eliminar el cuerno y 1 frag. de parietal.Viscerocráneo: 1 con M3 (+) subad.Atlas: 1 mordido por carnívoro.V.cervical: 1 frag. con cuerpo ant. y post. sin fusionar.Dientes: 7: superiores: 3: 1 M3 (+++) ad y 1 fag. De M1/M2 y 1 M3 (++) ad.; Inferior: 1 incisivo y 3 frag. molariformes.Mandíbula: 3 S: 1 ángulo mandibular; 1 mandíbula S algo quemada con M1 y M2 (++) subad/ad. y una casi completa con M3(++/+++) ad.Costilla: 1 frag. láminaHúmero: 1 e.d. quemado (negro).Fémur: 2 e.d. quemados (negro).Metacarpo: 1 D con huella de percusión.Metatarso: 6: e.d. y 2 porciones prox. con diferentes grados de combustión.Tibia: 6 e.d., 5 de ellos quemados (negro).
10.101 544 Capra hircus
4 4: 3♀? 1♂
Cuernos: 4 clavijas óseas S, pero incompletos con percusiones en su base, 1 es muy grande, posible ♂, y las otros parecen ser de ♀.Neurocráneo: 1 frag.
10.101 544 Sus sp. 3 1 juv. 20 Neurocráneo: 1 S quemado (marron-negro).Viscerocráneo: 1 D con P4, M1(++) y M2(+) (juv).Mandíbula: 1 rama horizontal D mordida.
10.101 544 SIN IDENTIFICAR
50 55 Esquirlas óseas muy alteradas y con fractura reciente.
907 Molusco marino
1 1 Cymbula nigra con relleno de tierra e impronta de una valva de pecten? Abrasión antrópica en el borde.
Abreviaturas:
Ad.: adultoa/p: anterior/posteriorant.: anteriorDist.: distale.d.: esquirla de diáfisise.neg.: estructura negativafrag.: fragmentoD: dexter (derecho)Inf.: infantil
Inf-juv: infantil-juvenilJuv-subad.= juvenil-subadultoMacro.: macromamíferoMeso.: mesomamíferoMNI: Número mínimo de individuosO/C: ovicaprinoPost.: posteriorProx.: proximalS: sinister (izquierdo)Subad.: subadulto