PLAN DE VACUNACION CONTRA SALMONELLA EN … · reclutamiento y la transmigración de neutrófilos...
30
PLAN DE VACUNACION CONTRA SALMONELLA EN GALLINAS PONEDORAS Principios Inmunitarios de la Vacunación contra Salmonella en Aves Ponedoras de Huevos de Consumo Dr. Sergio Rivera P. Profesor Titular Emérito Facultad de Ciencias Veterinarias Universidad del Zulia Maracaibo, Venezuela [email protected] INTRODUCCIÓN El principal interés para prevenir la infección con Salmonella en las aves, principalmente las ponedoras es la de evitar la infección en el transcurso del levante con Samonellas específicas del ave como la Gallinarum y la Pullorum, así como la contaminación de los huevos de consumo con Salmonella entérica serovares Enteritidis o Typhimurium en Venezuela. Sabemos que la S. Gallinarum y la S.Pullorum son Salmonellas inmóviles, de crecimiento intracelular facultativo, están adaptadas especialmente a las aves como huéspedes definitivos causando patologías diferentes a edades disímiles. La S.Gallinarum atacando principalmente aves adulta, provocando una altísima mortalidad que culmina con una Fiebre Entérica de carácter hemorrágico posterior a la invasión y excreción bacteriana a nivel del intestino.
Transcript of PLAN DE VACUNACION CONTRA SALMONELLA EN … · reclutamiento y la transmigración de neutrófilos...

PLAN DE VACUNACION CONTRA SALMONELLA EN GALLINAS
PONEDORAS
Principios Inmunitarios de la Vacunación contra Salmonella en Aves Ponedoras de Huevos
de Consumo
Dr. Sergio Rivera P.
Profesor Titular Emérito
Facultad de Ciencias Veterinarias
Universidad del Zulia
Maracaibo, Venezuela
INTRODUCCIÓN
El principal interés para prevenir la infección con Salmonella en las aves, principalmente
las ponedoras es la de evitar la infección en el transcurso del levante con Samonellas
específicas del ave como la Gallinarum y la Pullorum, así como la contaminación de los
huevos de consumo con Salmonella entérica serovares Enteritidis o Typhimurium en
Venezuela.
Sabemos que la S. Gallinarum y la S.Pullorum son Salmonellas inmóviles, de crecimiento
intracelular facultativo, están adaptadas especialmente a las aves como huéspedes
definitivos causando patologías diferentes a edades disímiles. La S.Gallinarum atacando
principalmente aves adulta, provocando una altísima mortalidad que culmina con una
Fiebre Entérica de carácter hemorrágico posterior a la invasión y excreción bacteriana a
nivel del intestino.

La S.Pullorum ataca principalmente aves jóvenes produciendo una sintomatología menos
dramática que la fiebre tifoidea, caracterizada principalmente por diarreas blanquecinas,
abdomen inflamado, retardo en el crecimiento y establecimiento de un estado de
portador. Causa una infección sistémica con hepato-esplenomegalia y lesiones puntuales
blanquecinas en Hígado y bazo, en ambos sexos. Infecta los huevos por lo que los
embriones nacen infectados internamente en sus sacos vitelinos. Tempranamente los
machos son mayormente infectados posteriormente en ambos sexos se establece el
estado de portadores en el 60% de las aves infectadas camuflajeandose en el interior de
los Macrófagos principalmente a nivel del Bazo. En su forma aguda, la pullorosis es una
enfermedad prácticamente exclusiva de los polluelos y el agente se puede recuperar a
partir de casi todos los órganos, los tejidos y las heces. En las aves mayores que también
llegan a ser portadoras, S. Pullorum se aísla sobre todo a partir de los óvulos y del
oviducto y sólo ocasionalmente de otros órganos y tejidos, incluyendo el tracto digestivo.
Esto asegura a la bacteria, en el caso de las reproductoras, infectar a los pollitos recién
nacidos y en el de las ponedoras, perpetuarse en el ambiente con la ayuda de otros
vectores. Ambas Salmonellas pertenecen al serotipo D pues comparten las estructuras
antigénicas de los antígenos O y LPS de superficie, identificables por pruebas serológicas.
Esto favorece enormemente su control vacunal. Ninguna afecta al Humano.
Por el contrario, las Salmonellas Enteritidis y Typhimurium entre otras, son Salmonellas
móviles producto de su Flagelo contentivo del Antígeno H o flagelar. No son específicas
de ningún Huésped, afectan a la mayoría de las especies animales incluyendo al ser
Humano. Las aves jóvenes podrían verse afectadas con una baja mortalidad producto de
una Enteritis con diarreas profusas. En adultos infectan la cascara del huevo desde su
postura, aprovechando los poros dilatados y permitiendo el ingreso de la bacteria por esta
vía. Sin embargo, los Huevos de consumo pueden llegar a infectarse posterior a la
colonización e invasión con estas Salmonellas, indirectamente a través del tracto
reproductivo del ave, incorporándose en Macrófagos y células Epiteliales del útero al
inicio de la postura, llegando hasta la Yema del Huevo, atravesando a nado la Albúmina
gracias a los movimientos de su Flagelo. La Salmonella Enteritidis pertenece igualmente al

serogrupo D mientras que la S.Typhimurium pertenece al serogrupo B, planteando un serio
problema para su control vacunal por la ausencia de reactividad cruzada entre estas dos
especies bacterianas, las cuales, a pesar de comportarse de la misma manera durante la
patogénesis, difieren en la estructura de sus antígenos de superficie O y LPS. Varios
Autores reseñan protección cruzada entre las vacunas vivas de S.Enteritidis y S.Gallinarum,
menos o ninguna con la S.Typhimurium.
MECANISMOS DE INVASIÓN DE SALMONELLA
La migración de los fagocitos infectados a otros órganos de sistema retículoendotelial
probablemente facilita la diseminación de la bacteria en el huésped. El análisis de la
estructura genética de las bacterias patógenas revelan que los genes de virulencia se
encuentran en regiones localizadas del cromosoma denominadas “islas de patogenicidad
de la Salmonella” o “SPI”. Las SPI -1, codifican por los genes de invasión de las células
epiteliales, inducción de la secreción intestinal y la respuesta inflamatoria.
Las SPI-2, codifican los genes de la replicación intracelular y el establecimiento de la
infección sistémica proveniente del epitelio intestinal. Estas SPI en conjunto con otras,
codifican por la producción de proteínas virulentas dentro del huésped denominadas
“sistema de secreción tipo III” o “TTSSs”. Los TTSSs modifican las funciones de las células
del huésped a través de la translocación de proteínas virulentas bacterianas dentro de su
citoplasma. Estas proteínas de translocación bacteriana alteran las funciones de señales
de transducción, arquitectura del citoesqueleto, tráfico de membrana y la expresión de los
genes de Citocinas en la célula huésped.
La SPI-1 no es esencial para la invasión de la Salmonella entérica serovar Gallinarum,
escasamente puede jugar un papel en las etapas iniciales de la infección. La función de la
expresión genética de la región SPI-2 TTSS es absolutamente necesaria para causar la
enfermedad, permitiendo que las bacterias vivan y se multipliquen dentro de los
Macrófago del Sistema Retículo Endotelial. Este sistema SPI-2 se encuentra igualmente
involucrado en la translocación de la bacteria del intestino hacia el Bazo y el Hígado. La
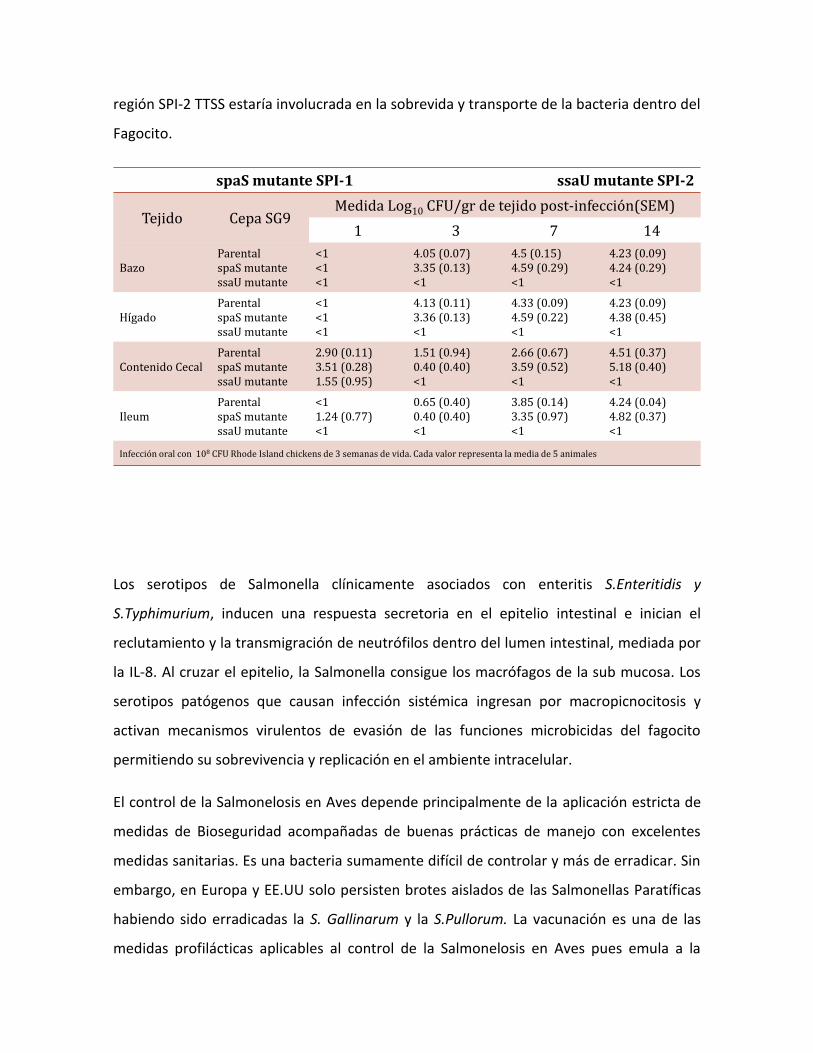
región SPI-2 TTSS estaría involucrada en la sobrevida y transporte de la bacteria dentro del
Fagocito.
spaS mutante SPI-1 ssaU mutante SPI-2
Tejido Cepa SG9 Medida Log10 CFU/gr de tejido post-infección(SEM)
1 3 7 14
BazoParentalspaS mutantessaU mutante
<1<1<1
4.05 (0.07)3.35 (0.13)<1
4.5 (0.15)4.59 (0.29)<1
4.23 (0.09)4.24 (0.29)<1
HígadoParentalspaS mutantessaU mutante
<1<1<1
4.13 (0.11)3.36 (0.13)<1
4.33 (0.09)4.59 (0.22)<1
4.23 (0.09)4.38 (0.45)<1
Contenido CecalParentalspaS mutantessaU mutante
2.90 (0.11)3.51 (0.28)1.55 (0.95)
1.51 (0.94)0.40 (0.40)<1
2.66 (0.67)3.59 (0.52)<1
4.51 (0.37)5.18 (0.40)<1
IleumParentalspaS mutantessaU mutante
<11.24 (0.77)<1
0.65 (0.40)0.40 (0.40)<1
3.85 (0.14)3.35 (0.97)<1
4.24 (0.04)4.82 (0.37)<1
Infección oral con 108 CFU Rhode Island chickens de 3 semanas de vida. Cada valor representa la media de 5 animales
Los serotipos de Salmonella clínicamente asociados con enteritis S.Enteritidis y
S.Typhimurium, inducen una respuesta secretoria en el epitelio intestinal e inician el
reclutamiento y la transmigración de neutrófilos dentro del lumen intestinal, mediada por
la IL-8. Al cruzar el epitelio, la Salmonella consigue los macrófagos de la sub mucosa. Los
serotipos patógenos que causan infección sistémica ingresan por macropicnocitosis y
activan mecanismos virulentos de evasión de las funciones microbicidas del fagocito
permitiendo su sobrevivencia y replicación en el ambiente intracelular.
El control de la Salmonelosis en Aves depende principalmente de la aplicación estricta de
medidas de Bioseguridad acompañadas de buenas prácticas de manejo con excelentes
medidas sanitarias. Es una bacteria sumamente difícil de controlar y más de erradicar. Sin
embargo, en Europa y EE.UU solo persisten brotes aislados de las Salmonellas Paratíficas
habiendo sido erradicadas la S. Gallinarum y la S.Pullorum. La vacunación es una de las
medidas profilácticas aplicables al control de la Salmonelosis en Aves pues emula a la

bacteria viva o a sus componentes antigénicos e inmunogenicos en las vacunas
inactivadas. Esta práctica ayudo a erradicar la S. Gallinarum y la S.Pullorum en Europa,
liberando a ese continente de estos dos patógenos. Sin embargo, abrió el espacio para la
implantación de las Salmonellas No Tifoideas que aún persisten en Europa.
Para diseñar un buen plan de vacunacion contra la Samonella en aves es necesario
primero conocer la Patogenesis de la enfermedad, la respuesta inmunitaria del Huesped
ante la infeccion por Salmonella, la existencia de genes de resistencia y los objetivos
trazados en el control de la enfermedad.
El ingreso de la Salmonella al huesped pasa por tres faces bien determinadas. La invasion,
la infeccion sistemica y la resolucion parcial de la misma. El ave puede controlar la
enfermedad sobre todo si se trata de lineas resistentes, puede provocar una altisima
mortalidad en aves susceptibles o utilizar al Huesped como Portador especialmente en el
caso de la S. Pullorum la cual no se asocia con altas tazas de mortalidad pero si con altas
persistencia bacteriana en el Bazo provocando infeccion del aparato reproductivo para de
esa manera contaminar por esa via el Huevo. S. Gallinarum y S.Pullorum son pobres
colonizadoras del intestino, no provocan reacciones inflamatorias en el momento de su
ingreso, no contaminan las cascaras de huevos, persisten en Hígado y Bazo, infectan el
tracto reproductivo y la yema del huevo, el ave puede curar o morir y no afectan al
Humano.
La Salmonellas No Tifoideas provocan la Salmonelosis en Humanos a traves de la
contaminacion de los huevos y de las carcasas de aves beneficiadas. En aves no provoca
gran mortalidad por el contrario afecta solo aves durante las dos primeras semanas de
vida provocando Enteritis y diarreas blanquecinas, en casos de deficiencias por otras
enfermedades virales y en el momento del inicio de la postura cuando se puede observar
un incremento de las cuentas bacterianas. La Salmonellas No Tifoideas no forman parte de
la flora intestinal normal pero pueden mantener una colonizacion por varias semanas o
por tiempo indefinido en Tonsilas cecales, parte alta del Intestino Delgado, Buche y
Proventriculo. La mayoria de las aves infectada permanecen asintomaticas como
portadores y permanecen como reservorio de la enfermedad potencialmente infectantes
para el Hombre, contaminando el ambiente y propagandose en otros individuos en la
granja. Salmonellas Enteritidis y Typhimurium no son Huésped Específicas, contaminan
principalmente el alimento, al ingreso invaden la mucosa intestinal provocando una fuerte
reacción inflamatoria que atrae polimorfonucleares y macrófagos desencadenando una
Enteritis con una altísima excreción bacteriana vía fecal, eventualmente provocan
enfermedades sistémicas, pueden provocar contaminación del tracto digestivo y ciegos sin
provocar enfermedad utilizando las aves cómo portadoras, contaminan las cascaras y la
yema de los huevos, las carcasas ambas fuentes de alimentación para el humano.
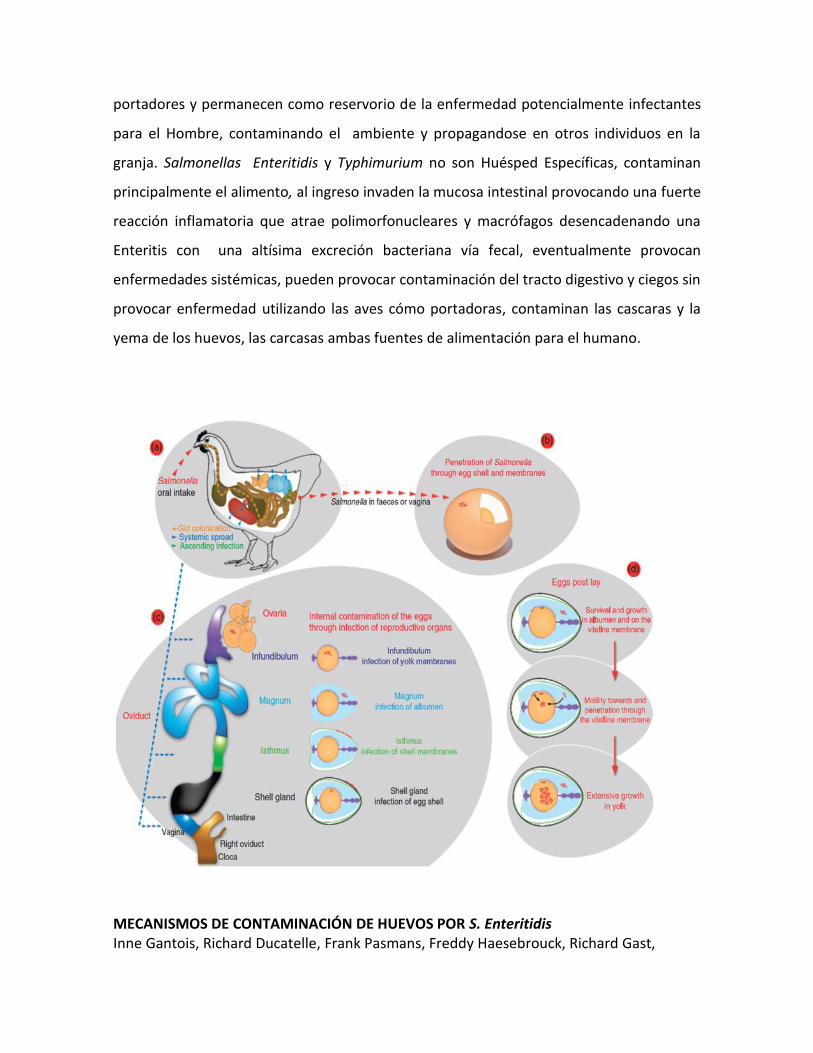
MECANISMOS DE CONTAMINACIÓN DE HUEVOS POR S. Enteritidis Inne Gantois, Richard Ducatelle, Frank Pasmans, Freddy Haesebrouck, Richard Gast,

Tom J. Humphrey & Filip Van Immerseel
La patogénesis de la contaminación del Huevo por la Salmonella se puede sintetizar de la
manera siguiente:
a) La Salmonella ingresa por la vía oral y llega al tracto intestinal. La Bacteria coloniza el
lumen intestinal e invade las células del epitelio intestinal (colonización). Como
consecuencia, las células inmunes, especialmente los Macrófagos, son atraídos al sitio de
invasión y fagocitan la Salmonella. Esto permite a la bacteria, con sus múltiples
mecanismos inhibitorios derivados de las SPI-2, sobrevivir y multiplicarse en el
microambiente intracelular del Macrófago. Los Macrófagos infectados migran desde las
organelas intestinales hasta el órgano reproductivo (diseminación sistémica). Además de
esta vía, la bacteria puede acceder al oviducto a través de la infección proveniente de la
Cloaca.
b) Una posible vía de contaminación del huevo es la penetración de la Salmonella a través
de la cascara del huevo antes de la contaminación de la misma vía la vagina o por
contaminación fecal.
c) La segunda ruta de contaminación es la contaminación directa del saco vitelino,
membranas vitelinas, albumina, membranas de la cascara y la cascara del huevo
originándose por infección del ovario, infundíbulo, magnum, istmo, y glándulas de la
cascara, respectivamente aprovechando el descenso de la respuesta celular en entre las
semanas 16 y 20 al momento del desarrollo de tracto reproductivo y el inicio de la postura
en aves ponedoras y reproductoras.
d) La Salmonella se deposita en la albumina y/o la membrana vitelina y son capaces de
sobrevivir y crecer a pesar del microambiente anti bacteriano. Ellas son capaces de migrar
y penetrar la membrana vitelina hasta alcanzar el saco vitelino donde, gracias a la
cantidad de nutrientes crecen y se desarrollan de manera extensiva. La Samonella

Enteritidis y Typhimurium pueden atravesar la albúmina hasta alcanzar la yema de huevo
gracias a su flagelo.
La disminución en la actividad proliferativa de Linfocitos T en el momento del inicio de la
postura está relacionada y coincide con una reducción del número de células inmunitarias.
Se puede observar al momento del inicio de la postura una disminución de algunas sub
poblaciones T en la cual conlleva a una inmunosupresion por la disminución de varias sub
poblaciones TCD4+, Células Tγδ y de Células Dendríticas (presentadoras de antígeno). Al
inicio de la postura, se reducen a pocas células inmunes en la mucosa del tracto
reproductivo, y una reducción fisiológica de la respuesta Th1, citosina como el IFN-γ el
cual dirige la respuesta celular capaz de eliminar la Salmonella y conferir protección
contra esta bacteria en aves se encuentra disminuido.
Es probable que estos cambios sean producidos por variaciones en las hormonas
esteroideas las cuales dirigen el tracto reproductivo en el momento del inicio de la
oviposición, semejante a la inmunosupresion observada en mamíferos durante la preñez o
el post parto. El incremento de requerimientos nutricionales acompaña este desarrollo del
tracto reproductivo en aves, el cual debería favorecer la recuperación del sistema inmune.
Hacia la semana 22 todas estas poblaciones linfocitaria se recuperan, inclusive se observa
un repunte importante de la inmunidad humoral semejante a una respuesta secundaria
de anticuerpos.
El uso de vacunas viva de Salmonella en Ponedoras durante este periodo es
contraproducente por la posibilidad de que las mismas infecten el huevo de consumo. Las
vacunas inactivadas colocadas durante este período no provocan la reacción buster
esperada para aves primo vacunadas con vacunas vivas o bacterinas. En uno de cada diez
huevos colectados de aves vacunadas durante este período se ha podido aislar la cepas
vivas vacunales de Salmonella.

ESTADO DE INMUNOSUPRESION NATURAL DE AVES HEMBRAS ENTRE LAS SEMANAS 16 Y 20. A
S.Pullorum como antígeno B. PHA (Antígeno Inespecífico)
En un estudio comparativo sobre colonización intestinal de S.Typhimurium y S.Gallinarum
se demostró que a pesar de que S.Gallinarum es una bacteria Huésped especifica del ave,
la colonización de la Bursa, los Ciegos y Macrófagos intestinales es superada
significativamente por la S.Typhimurium. Esto corrobora que aún cuando estos tejidos
intestinales son abordados por ambas salmonellas, la S.Gallinarum es más invasora de
órganos internos que colonizadora de la mucosa intestinal a través de la cual pasa sin
ocasionar ruido inflamatorio. Por otra parte, corrobora que desde el punto de vista
inmunológico ambas Salmonellas, las Paratíficas y las Huésped especifico pueden
coadyuvar un estado de inmunosupresión al atacar tejidos linfoides intestinales y
posteriormente invadir el principal órgano inmunológico secundario como lo es el Bazo, lo
cual explica la gravedad de la sintomatología en la pullorosis y la altísima mortalidad
observada en aves de todas la edades por la S.Gallinarum.
RESISTENCIA GENÉTICA A LA INFECCIÓN CON SALMONELLAS

En las razas livianas como la Leghorn son más resistentes a la pullorosis que las razas más
pesadas como Rhode Island Red, Plymouth Rock Barreada, Wyandotte Blanca o New
Hampshire. Los diferentes loci genéticos asociados a Resistencia a la Salmonelosis en aves
han sido identificados y se encuentran todos relacionados con la respuesta inmunitaria del
Ave. Se ha descrito una importante resistencia genética frente a la infección de
Salmonellas en aves y otras especies animales. Algunos genes y otros factores han sido
descritos como relacionados con esta resistencia natural entre los cuales están
slc1a1(Nramp1), el Complejo Mayor de Histocompatibilidad, el receptor Toll- like 4 y el
locus SAL1 principalmente frente al desafío con la S.Gallinarum y en menos grado frente a
la infección con S.Typhimurium. En las aves resistentes se observa disminución de la
mortalidad y de la morbilidad y disminución de las lesiones granulomatosas en Hígado
producto de la acción bacteriana. A pesar de esto no se observan diferencia en la
colonización e invasión bacteriana del tracto digestivo entre aves resistentes y
susceptibles. Sin embargo, el número de bacterias se incrementa rápidamente en Bazo e
Hígado de aves susceptibles posterior al desafío indicando que la inmunidad innata debe
impedir el acceso bacteriano de una manera más eficaz en aves resistentes. Los
Macrófagos provenientes de aves resistentes muestran una fuerte respuesta oxidativa
frente a la infección frente a una pequeña o indetectable respuesta en aves susceptibles.
El papel de los Halterófilos ha sido investigado en aves resistentes a Salmonella entérica
seovar Enteritidis, mostrando una estrecha correlación entre la función de los Heterófilos
y la resistencia a la infección.
Genes de marcadores de células T, citosinas e inmunoglobulinas han sido asociados a
resistencia a la Salmonella tales como el los genes de CD28 y TLR4, diferente expresión de
genes de IL-6, IL-18. La citosina pro-inflamatoria IL-1β incrementa su concentración 70
veces frente a la infección con S.Gallinarum en 1 hora mientras que se incrementa 60
veces frente a la infección con S.Typhimurium en 20 min, demostrando la importancia de
la reacción inflamatoria frente a las No Tifoideas. Los factores quimiotacticos de
Halterófilos CCLi2 y CXCLi1 muestran una gran expresión frente al desafío de Salmonella
en aves resistentes. Los genes de la familia MIP, quemoquina CC, CCLi2 de Macrófagos

resistentes muestran también una rápida expresión en aves resistentes. En resumen, las
citosinas pro inflamatorias IL-6, IL-8 y la citosinas Th1 IL-18 muestran una expresión
aumentada en aves resistentes a igual que la respuesta proliferativa de aves resistentes a
la Salmonella. La respuesta humoral la cual se mantiene estable en ambas. Lo cual aunado
a los estudios de Bursectomia in ovo, señala que la respuesta humoral sistémica participa
poco en el controla de la bacteria, no así la respuesta IgA dsecretoria en la mucosa la cual
es imprescindible para frenar la colonización de las Salmonellas No Tifoideas. La
respuesta vacunal de aves resistente es superior a la de las razas sensibles a la Salmonella
en un 100%.
RESPUESTA INMUNITARIA FRENTE A LA INFECCIÓN CON SALMONELLAS
La infección con Salmonellas en aves desencadena una respuesta inmunitaria compleja,
constituida por la inmunidad innata y la adaptativa en diversos grados de acuerdo a la
patogénesis de la enfermedad. A su ingreso desencadena una expresión de genes de
receptores TLR-Toll algunos comunes a varias especies como el TLR4 y otros como los
TLR5, TLR15 y TLR21, algunos específicos de las aves. Estos receptores dirigen la
trayectoria de la respuesta inmunitaria, la resistencia inmunitaria y promocionan una
respuesta adaptativa propiciando la maduración de las células Dendríticas. Igualmente se
observa la expresión de genes de citosinas pro- inflamatorias IL-6 el IFN-γ así como las
quimiotacticas IL-8 y K60, propios de la inmunidad innata. A su ingreso por la vía intestinal,
la S.Gallinarum no induce inflamación. Escasa o nula producción de la IL-6 post infección
explica la falta de reacción inflamatoria local permitiendo a la bacteria una amplia invasión
de los órganos internos como Bazo e Hígado.
BASES INMUNITARIAS DEL ESTADO DE PORTADOR Y LA INFECCIÓN DE HUEVOS
S. Pullorum persiste dentro del Macrófago en Bazo e Hígado alrededor de 12 semanas
post infección. Una fuerte producción de Anticuerpos y una ponderosa respuesta celular
pueden ser detectadas posteriores a la infección. Ulterior al estado agudo inicial de la
infección, el número de bacteria disminuye en Bazo e Hígado.

Al momento de la madurez sexual, 18 semanas aprox. se produce un incremento
significativo del número de bacterias en hembras, mas no en machos. La Salmonella se
puede recuperar del aparato reproductivo de las hembras pero no se aísla de los testículos
de los machos. Ambos sexos desarrollan el estado de portador cuando son infectados por
Salmonella. La infección del tracto reproductivo y el incremento del número de bacterias
llegada la madurez sexual, están restringidas solamente a las aves hembras. Ninguna
diferencia en la producción de Anticuerpos es observada entre ambos sexos. Las hembras
infectadas retardan la postura una semana.
La respuesta Humoral no declina en el momento del inicio de la postura. Una fuerte
respuesta humoral se observa hacia la semana 5a post infección. Hacia la semana 22 se
observa un incremento importante de la respuesta de anticuerpos semejante a una
respuesta secundaria en animales infectados con Salmonella. A pesar de la importante
respuesta Humoral IgG, esta no participa en la eliminación de la Salmonella. El repunte
hacia la sem. 22 se encuentra correlacionado con el aumento sistémico de la Salmonella
Pullorum.
Una respuesta celular T, antígeno específica se puede observar igualmente entre la 5a y 9a
semana post infección. A la semana 17 post infección se observa una drástica disminución
de la respuesta celular T. Esta baja de respuesta coincide con el inicio de la postura. La
respuesta celular inespecífica a la PHA muestra el mismo comportamiento que la
respuesta T antígeno específica para Salmonella.
El estado de portador de aves infectadas con S.Pullorum coincide con una disminución o
falta de expresión de los genes de INF-γ e IL-18, Citosinas reveladoras de un patrón de
respuesta Celular TH1. Por el Contrario, muestran una alta expresión de IL-4 Citosina del
patrón Humoral TH2 que coincide con la producción de elevadas concentraciones de IgG
específicas de S.Pullorum. La infección con S. Pullorum beneficiaría una respuesta Th2,
con una fuerte producción de anticuerpos para favorecer el estadio de portador. La
respuesta inmunitaria a la infección primaria es superior a la reinfección.

Posterior a la infección primaria con S.Enteritidis y S.Typhimurium, se observa un
incremento del IFN-γ, TGF-β e IL-1β, así como un incremento marcado de la proliferación
in vitro de esplenocitos frente a los antígenos flagelares y otras proteínas de membrana
vivas o inactivadas. Solamente una fuerte respuesta celular T se encuentra asociada a la
resistencia frente a la infección con S.Enteritidis y S.Typhimurium. El incremento de la
respuesta humoral no se encuentra asociada con la resistencia a la enfermedad a pesar
de que forma parte de las características del huésped. El incremento del IFN- y de la IL-1
son características de la Edad pero tampoco se encuentran asociadas a resistencia frente a
las Samonellas No Tifoideas.
La flora normar que coloniza al ave al nacer se caracteriza por Enterobacteriaceae (phylum
Proteobacteria) y diferentes representantes del phylum Firmicutes. La Salmonella no
forma parte de la flora intestinal. Las β defensinas forman parte de la protección de esa
mucosa y se incrementan por efecto de la microflora que incentiva la maduración. Las
gallinacins 1, 2, 4 y 6 incrementan su expresión los tres primeros días de vida,
posteriormente aparece un pico de expresión de las citosinas IL-8 y la IL-17 las cuales
propician la maduración del sistema inmune intestinal y la desaparición de las defensinas.
Todo esto producto de la acción de la flora intestinal. Las respuestas primarias tipo Th1
requieren 10 días de maduración posterior al nacimiento. La infección con S. Enteritidis,
induce una expresión de IL-8 e IL-17 significativamente mayor de la inducida por la
microflora. Igualmente, la infección con S.Enteritidis incrementa la expresión de IL-1 β, IL-
22, iNOS, IFN- γ e IL-18, tres días después de la infección al día 1, 4 o 16 posterior al
nacimiento. Ninguna de las gallinacins 1, 2, 4 o 6 (defensinas), incrementan su expresión
posterior a la infección con S.Enteritidis. Aun cuando la IL-22 y la IL-17 se expresan al
mismo tiempo producto de la excreción de las células Th17, independientemente, la IL-22
se ocupa de la protección y reparación de las células tejido epitelial de la mucosa puesto
que sus receptores se ubican solamente sobre células no inmunes. Por el contrario los
receptores de la IL-17 solo se observan en células Dendríticas, Macrófagos y Linfocitos T.
Ambas incrementan la resistencia de la mucosa frente a la infección bacteriana y
propenden su reparación, posterior al daño causado por el proceso inflamatorio. Las

células Th17 estimuladas posterior a la infección con S.Enteritidis, solo a la semana 16
posterior al nacimiento, impiden la diseminación e invasión del Bazo e Hígado por parte
de la bacteria, lo cual es reflejo de una madurez plena del aparato inmune intestinal y
pudienendo ser utilizado este resultado para mejorar los planes de vacunación.
Las aves infectadas con Salmonella Enteritidis o Typhimurium muestran altos niveles de
anticuerpos, respuesta de células T, citosinas y quemoquinas. Se han descrito poblaciones
TCD4+ cooperadoras y TCD8+ citotoxicas. La respuesta inmunitaria local en el intestino es
más efectiva y se asocia a más a la eliminación de la S.Enteritidis del tracto gastrointestinal
que la respuesta sistémica. Una fuerte respuesta celular local se observa en animales
expuestos a la Salmonella con modificaciones de las poblaciones celulares T
especialmente la CD8+TcR1+(γδ) en ciegos posterior a la inoculación de S.Typhimurium en
aves de 1 día de edad como señalamos anteriormente.
La IgA contribuye a la eliminación tardía de la Salmonella en intestino mientras que la
respuesta celular es la responsable de eliminarla en los tejidos por medio de mecanismos
aun no totalmente esclarecidos. La Bursectomia no afecta la eliminación de la Salmonella
en aves. La reinfección de la Salmonella provoca una rápida eliminación de la Salmonella,
evidenciando una importante respuesta memoria lo cual favorece el empleo de vacunas
como parte del control de la bacteria.
VACUNAS COMO MEDIDA DE CONTROL DE LA SALMONELLA
En aves, las vacunas pueden prevenir la colonización intestinal y cecal disminuyendo la
excreción fecal, impidiendo la infección sistémica y la transmisión vertical y contaminación
de los huevos. La vacunación con vacunas vivas o inactivadas reducen la susceptibilidad
del ave a la infección con Salmonella. Ambas, vivas e inactivadas protegen en mayor o
menor grado contra el desafío bacteriano. Las vacunas muertas incrementan la
inmunidad humoral y reducen la prevalencia de la Salmonella pero no son capaces de
disminuir significativamente las Salmonellas del medio ambiente de la granja.

Las vacunas inactivadas por calor o formalina más adyuvante no son capaces de generar
una buena respuesta celular que al final, resulta la más importante para eliminar la
Salmonella, además, no son capaces de inducir IgA a nivel de la superficie de la mucosa
intestinal lo cual evitaría la colonización bacteriana. Las vacunas de subunidades, así como
otras proteínas de membrana, porinas, fracción ribosomal han sido evaluadas para
prevenir la Salmonella pero ellas no son eficaces sin adyuvantes adecuados o sistemas
portadores. Las vacunas vivas y atenuadas han sido mundialmente utilizadas y su eficacia
demostrada ante el desafío bacteriano.
Las vacunas vivas de Salmonella son cepas cuya virulencia ha sido reducida sin
comprometer su inmunogenicidad; Incluye cepas semi rugosas como la 9R contra la S.
Gallinarum. Las bacterias confieren por lo tanto una protección parcial contra la colonización
intestinal, excreción fecal, diseminación sistémica y contaminación vertical en aves, por lo que ha
sido planteado que los anticuerpos específicos producidos por ejemplo contra Salmonella
Enteritidis en aves vacunadas con vacuna inactivas, no protegen a las aves contra infección y que
las vacunas vivas aumentan las respuesta inmune mediada por células, con un incremento de la
eliminación de Salmonella Enteritidis en las aves.
Algunos trabajos han indicado que los anticuerpos maternos no reducen
significativamente la excreción de la bacteria en la progenie, sin embargo reducen la
mortalidad puesto que buena parte de ellos son vertidos al intestino antes de la eclosión.
La protección ofrecida por las vacunas inactivadas no es tan fuerte como aquella
proporcionada por las vacunas vivas atenuadas, no obstante la naturaleza de la
atenuación de éstas vacunas y la respuesta inmune que ellas generan aún son
pobremente entendidas.
Por otro lado, un requisito indispensable para la reducción o ausencia de excreción de
Salmonella en las aves, es la existencia de la producción de SIgA específica en las mucosas,
nivel de los tractos digestivo y reproductivo. Esta estimulación en las mucosas no es
posible con la utilización de vacunas inactivadas de Salmonella.

A pesar de su variable eficacia y amplia utilización como herramienta en la prevención del
Salmonelosis en las aves, estos productos tienen algunas desventajas: a) los costos de
manejo para la aplicación en las aves; b) el estrés en la vacunación y post vacunación de
las aves; c) las eventuales lesiones por la reacción inflamatoria en el lugar de aplicación y
d) la interferencia con el monitoreo serológico en programas de prevención y control.
A pesar de que algunas vacunas inactivadas contra Salmonella pueden conferir protección
contra la re-infección en muchas especies animales, la potencial superioridad de las
vacunas vivas atenuadas al compararse con preparaciones inactivadas, ha servido de
punto de partida para amplias investigaciones dirigidas al desarrollo de cepas mutantes de
Salmonella que puedan ser utilizadas como vacunas en medicina humana y veterinaria.
La vacunación es una herramienta eficaz en la prevención de las infecciones por
Salmonella, las vacunas disponibles contra la salmonelosis pueden ser divididas en tres
grandes grupos: vacunas inactivadas conteniendo bacterias completas, vacunas sub-
unidades y vacunas vivas atenuadas, las cuales son utilizadas actualmente en aves. La
eficacia de las preparaciones de vacunas es evaluada generalmente por el nivel de
colonización intestinal y sistémica así como la morbilidad y mortalidad después de la
vacunación. Sin embargo, el nivel de protección de un biológico depende de otros
factores, entre los cuales se pueden mencionar: la cepa de desafío, la vía de
administración, la dosis infectante, la edad o línea de aves utilizada, entre otras. En
consecuencia, es difícil comparar la eficacia de las vacunas que actualmente están
disponibles para la prevención o el control de la salmonelosis en aves.
La vacunación en aves como método de prevención y control de Salmonella, está dirigido
básicamente a tres aspectos: a) prevenir o reducir la colonización intestinal; b) prevenir la
infección sistémica y; c) reducir la excreción fecal. La vacuna ideal debería cumplir con los
objetivos que se muestran en la Figura 1. Utilizando la vacunación se previene la infección
sistémica, se reduce la colonización en el tracto reproductivo y por lo tanto se previene la
transmisión vertical del microorganismo; y reduciendo la colonización intestinal y

excreción fecal se reduce la contaminación de la carcasa, huevo y el medio ambiente
(cama), relacionada principalmente a la transmisión horizontal de la bacteria.
La vacunación es considerada como un costo adicional en la industria avícola. La decisión
de establecer programas de vacunación debe estar de acuerdo con la gravedad del
problema en la región o área, y con los programas establecidos por las industrias o por los
servicios oficiales, después de un estudio epidemiológico serio. En líneas generales, se
puede inferir que cuando la prevalencia de los lotes es alta, la vacunación puede ser una
herramienta muy útil para reducir la prevalencia y excreción de salmonellas, aplicada
junto con las otras acciones dirigidas al control de la salmonelosis en la granja. Cuando la
prevalencia es baja la vacunación puede ser utilizada como una medida preventiva para
mantener la baja prevalencia o disminuirla aún más.
VACUNAS INACTIVADAS
Estos productos contienen la bacteria, que ha sido inactivada y suspendida usualmente en
una emulsión agua-aceite o con adyuvante hidróxido de aluminio. Estos productos
estimulan altos niveles de anticuerpos circulantes ya que son utilizados para la

administración parenteral. Estas vacunas conteniendo células bacterianas inactivadas
inducen una pobre respuesta inmune mediada por células.
Algunos estudios han demostrado la eficacia de las bacterinas en la reducción significativa
de la excreción fecal de Salmonella Enteritidis, así como la disminución de la persistencia
de Salmonella en los órganos y los huevos, demostrando que la administración de
productos inactivados reducen la infección sistémica por Salmonella Enteritidis por la
inducción de una buena respuesta humoral, y la ausencia de una buena inmunidad
mediada por células, e inmunidad a nivel de mucosas. Otros muestran que la protección
es variable, y que los niveles de contaminación en aves vacunadas y no vacunadas puede
ser el mismo.
En las aves, algunos estudios han demostrado que la vacunación con productos
inactivados tienen una respuesta excelente en la producción de anticuerpos. Esta
producción de anticuerpos está considerada como una medida de inmunidad activa, sin
embargo, a pesar de producir una buena respuesta humoral, es insuficiente en la
activación las células Th1 - linfocitos T ayudadores relacionados con la inmunidad mediada
por células.
Las bacterinas confieren por lo tanto una protección parcial contra la colonización
intestinal, excreción fecal, diseminación sistémica y contaminación vertical en aves; por lo
que ha sido planteado que los anticuerpos específicos producidos por ejemplo contra
Salmonella Enteritidis en aves vacunadas con vacuna inactivas, no protegen a las aves
contra infección, y que las vacunas vivas aumentan las respuesta inmune mediada por
células, con un incremento de la eliminación de Salmonella Enteritidis en las aves.
Algunos trabajos han indicado que los anticuerpos maternos no reducen
significativamente la excreción de la bacteria en la progenie, sin embargo reducen la
mortalidad. La protección ofrecida por las vacunas inactivadas no es tan fuerte como
aquella proporcionada por las vacunas vivas atenuadas, no obstante la naturaleza de la

atenuación de éstas vacunas y la respuesta inmune que ellas generan aún son
pobremente entendidas.
Por otro lado, un requisito indispensable para la reducción o ausencia de excreción de
Salmonella en las aves, es la existencia de la producción de SIgA específica en las mucosas,
nivel de los tractos digestivo y reproductivo. Esta estimulación en las mucosas no es
posible con la utilización de vacunas inactivadas de Salmonella.
A pesar de su variable eficacia y amplia utilización como herramienta en la prevención del
Salmonelosis en las aves, estos productos tienen algunas desventajas: a) los costos de
manejo para la aplicación en las aves; b) el estrés en la vacunación y post vacunación de
las aves; c) las eventuales lesiones por la reacción inflamatoria en el lugar de aplicación y
d) la interferencia con el monitoreo serológico en programas de prevención y control.
A pesar de que algunas vacunas inactivadas contra Salmonella pueden conferir protección
contra la re-infección en muchas especies animales, la potencial superioridad de las
vacunas vivas atenuadas al compararse con preparaciones inactivadas, ha servido de
punto de partida para amplias investigaciones dirigidas al desarrollo de cepas mutantes de
Salmonella que puedan ser utilizadas como vacunas en medicina humana y veterinaria.
VACUNAS SUB-UNIDADES
Estos productos están licenciados para su uso en humanos y también han sido utilizados
en aves. En el caso de humanos para S. Typhi esta vacunas son seguras e inmunogénicas y
son producidas del polisacárido de la cápsula. Otras vacunas producidas a partir de LPS,
extractos celulares, polisacáridos O y conjugados O, han sido probadas en modelos
experimentales y han probado ser menos eficaces. En aves vacunas producidas con
proteínas de la membrana externa con adyuvante han sido utilizadas para disminuir la
eliminación de S. Enteritidis. Otra vacuna producida a partir de un polipéptido de fliC (un
gen relacionado con la colonización de S. Enteritidis) ha confirmado que esta subunidad es
el antígeno eficaz en vacunas inactivadas de S. Enteritidis y se ha sugerido que esta única

subunidad es necesaria para suprimir efectivamente la colonización de la bacteria en el
intestino de las aves.
VACUNAS VIVAS ATENUADAS
La potencial superioridad de las vacunas vivas atenuadas en comparación con productos
inactivados ha sido motivo de numerosas investigaciones, con el desarrollo de cepas
mutantes de Salmonella tanto para uso humano como veterinario. Se ha relatado que la
vacunación con vacunas vivas disminuye la excreción de Salmonella Enteritidis
considerablemente, cuando fueron comparados con grupos vacunados con vacunas
inactivadas o sin vacunación, lo cual concuerda con los resultados obtenidos por otros
autores que confirman la eficacia de vacunas vivas en comparación con vacunas
inactivadas.
Al mismo tiempo, se reafirma la importancia de la inmunidad mediada por células (CMI)
en la eliminación de Salmonella en aves, lo que ya había sido demostrado en roedores, y
que ha tenido una confirmación en las aves por el grupo de Beal en el Reino Unido,
quienes confirman que la eliminación de Salmonella Typhimurium es independiente de la
función de las células B y que a pesar de que se inducen altos niveles de anticuerpos en las
aves, las células B no juegan un papel en la eliminación de una infección primaria y
tampoco en la mejora de la respuesta ante un segundo desafío.
El desarrollo de vacunas vivas atenuadas contra las infecciones por Salmonella ha sido
basado mayormente en conocimientos empíricos, pero la disponibilidad de informaciones
sobre el genoma, y métodos avanzados de identificación de los genes de virulencia, han
sido herramientas invaluables en la generación de cepas mutantes, candidatas potenciales
a la producción de vacunas.
No obstante, el diseño racional y la selección de las cepas deben depender del
conocimiento de la patogénesis e inmunobiología de las enfermedades. Por ejemplo, en
relación a la inmunidad, ha sido demostrado en roedores que la inmunización oral con

Salmonella Typhimurium desarrolla una respuesta en la producción de IgA
simultáneamente en la lamina propia del intestino y en el bazo; por lo que se puede
considerar la ruta de inmunización oral una vía confiable para inducir una respuesta
inmune de mucosas con la producción de IgA en el intestino.
Las vacunas vivas contra Salmonella generalmente confieren mejor protección que
vacunas inactivadas, por la estimulación de la inmunidad mediada por células y la
inmunidad humoral. El uso de vacunas vivas se ha extendido en las aves, sin embargo, a
pesar de la inocuidad demostrada en estos productos debe tenerse cuidado en respetar
los periodos de retiro del producto, para asegurar que los productos finales (carne y
huevos) no están contaminados con las cepas vacunales ni con residuos de los otros
componentes de la vacuna.
Las cepas vacunales son generalmente atenuadas o mutantes, generalmente por la
manipulación de genes que han sido blanco para la construcción de vacunas vivas
atenuadas. Por ejemplo la S. Typhimurium aro A, cepa que carece de la capacidad de
replicación en el hospedero por la ausencia o la insuficiencia de algunos metabolitos
necesario para el crecimiento bacteriano.
Las mutantes con el sistema PhoP/PhoQ no funcional, son altamente atenuadas in vivo,
este sistema está asociado con la resistencia de la bacteria a los mecanismos innatos de
defensa. Otras cepas vacunales son deficientes en el loci SPI-2 (isla de patogenicidad 2),
este sistema es requerido para la infección sistémica y para la sobrevivencia bacteriana en
los fagocitos.
Otra cepa atenuada utilizada ampliamente desde la década del 50 es la 9R con estructura
rugosa del lipolisacárido, aunque la naturaleza de su atenuación sea desconocida se utiliza
hace más de 50 años con resultados variables. A pesar de que esta cepa es altamente
atenuada comparada con la cepa original Salmonella Gallinarum 9, la cepa 9R causa una
enfermedad sistémica con lesiones en el hígado y bazo, y persistencia por algunas
semanas en estos órganos.
Por lo tanto, la cepa vacunal produce una forma leve de salmonelosis sistémica, lo que ha
permitido que se utilice como modelo biológico para caracterizar la inmunidad en la tifosis
aviar. Una vacuna eficiente en la protección de Salmonella Enteritidis y protección cruzada
contra Salmonella Gallinarum es la producida con una cepa mutante metabólica de
Salmonella Enteritidis habiéndose demostrado su eficacia también en ponedoras
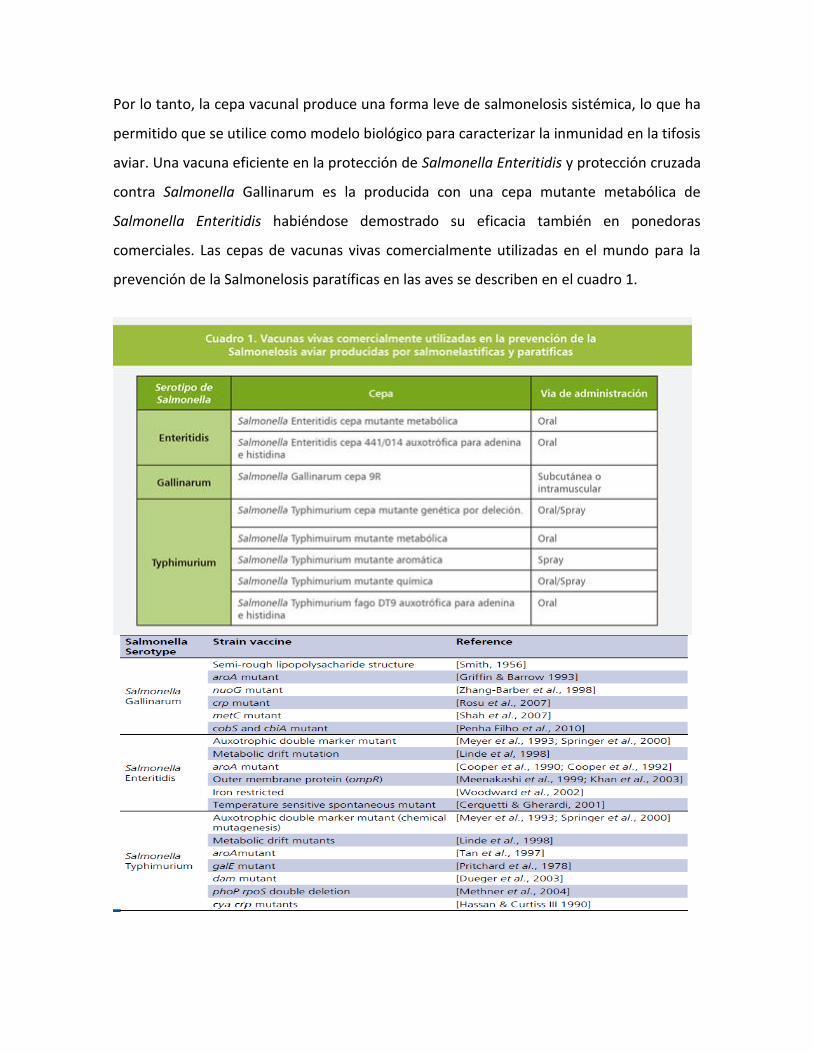
comerciales. Las cepas de vacunas vivas comercialmente utilizadas en el mundo para la
prevención de la Salmonelosis paratíficas en las aves se describen en el cuadro 1.

MECANISMOS DE PROTECCIÓN: INMUNIDAD PROPICIADA POR VACUNAS
El cuadro 2, muestra las diferencias en la activación del sistema inmune y de los
participantes de la respuesta inmune, que explica porque la eficacia de las vacunas vivas
es superior a las vacunas inactivadas en varios aspectos, especialmente en reducir la
colonización y excreción, en activar la respuesta inmune mediada por células y en generar
clones de células memoria.
Cuadro 2. Diferencias en la activación del sistema inmune y de los participantes de la
respuesta
inmune
Después de la inmunización con una vacuna viva atenuada de Salmonella, se desarrolla la
memoria inmunológica con clones de LT. Las respuestas séricas y de la mucosa obtenidas
por las vacunas vivas son directamente dirigidas a un amplio espectro de antígenos. En
animales y humanos expuestos a una vacuna viva contra Salmonella, las respuestas

inmunes son iniciadas por linfocitos cooperadores de la subpoblación Th1, directamente
relacionada con la inmunidad mediada por células, que es la responsable de la eliminación
de organismos intracelulares como la Salmonella.
Se debe tener en cuenta que dependiendo de los serotipos de Salmonella se observarán
diferencias en las respuestas inmunes y en aves se pueden establecer dos modelos básicos
relacionados a las salmonellas hospedero específicas (S. Gallinarum y S. Pullorum), y a los
grupos inespecíficos (S. Enteritidis, S. Typhimurium y otras). Es necesario mencionar
también que la respuesta inmune a antígenos complejos es raramente exclusiva mediada
por Th1 o Th2, sin embargo existe una combinación y dominancia de algunas citoquinas
que dirigen la respuesta a una respuesta predominantemente humoral o celular.
El cuadro 3, muestra las diferencias en la activación del sistema inmune y de los
participantes de la respuesta inmune, que explica porque la eficacia de las vacunas vivas
de administración oral es superior a las vacunas vivas inyectables en varios aspectos,
especialmente en reducir la colonización y excreción, en activar la respuesta inmune
mediada por células y específica en las mucosas.

Administrando la bacteria oralmente en aves recién nacidas, el resultado es una extensa
colonización del intestino y un fuerte estímulo de la respuesta inmune adquirida, en aves
que son inmunológicamente inmaduras.
Sin embargo, la colonización ejerce una variedad de eventos con una rápida respuesta
protectora dentro de un máximo de 72 horas. Esto incluye la inhibición específica de la
colonización, por el mecanismo de exclusión competitiva, en el cual la bacteria protectora
ejerce una resistencia profunda al establecimiento y colonización de otra bacteria
relacionada.
Este debe ser un atributo primario de la cepa vacunal que también puede envolver
competencia por los lugares de colonización. La presencia de una gran cantidad de
bacterias vacunales en el intestino puede inducir la infiltración de heterófilos en la pared
intestinal, lo que proporciona resistencia no sólo a la invasión sino a la diseminación
sistémica de una cepa patógena.

Esto abre la perspectiva para el uso de vacunas en pollos de carne o asaderos, ponedoras
y reproductores; pero también en otros animales que muestran susceptibilidad
aumentada a la infección por su juventud o por otras razones, como la profilaxis o
antibioticoterapia, donde la pérdida de la microflora nativa intestinal es una cuestión que
deja abierta las puertas para la instalación de patógenos, especialmente los entéricos.
Las vacunas vivas atenuadas de administración oral reproducen la vía natural de infección,
con lo que se tornan los productos más adecuados para la estimulación de la inmunidad
de las mucosas; con lo cual existe una colonización previa del intestino con la cepa vacunal
que ejerce un efecto similar al producido por los productos de exclusión competitiva o
probióticos, con la ventaja de que con la vacunación oral se estimula el sistema inmune
local de la mucosa intestinal, con la activación del sistema linfoide asociado al intestino
(GALT) y la consecuente activación de la inmunidad innata y adquirida.
La inmunidad generada en las mucosas es un factor importante en el sistema inmune,
pues puede reflejar en la inmunidad en otros tejidos o sistemas fisiológicos del organismo,
contra agentes infecciosos. La respuesta inmune intestinal contra Salmonella abarca una
serie de interacciones complejas, que incluyen, citoquinas, linfocitos, células epiteliales y
otros factores presentes en el ecosistema intestinal.
El sistema inmune común de las mucosas está respaldado por una serie de investigaciones
que demuestran que este sistema ha evolucionado paralelamente con el sistema inmune
sistémico, como consecuencia de esta dicotomía, solamente las respuestas iniciadas en los
lugares inductores de la mucosa pueden resultar en una respuesta inmune eficaz. El
sistema inmune secretor proporciona una red integrada ligada de órganos de las mucosas
que trabajan independientemente de la inmunidad sistémica.
Este concepto de “sistema inmune común de las mucosas” está respaldado por la
migración de células efectoras y la secreción de moléculas de SIgA que aseguran que la
experiencia antigénica en una mucosa puede ser reflejada en la expresión del sistema
inmune efector en una superficie distante. Por ejemplo, la estimulación del sistema

inmune en el intestino puede resultar en la producción y detección de anticuerpos
específicos en la tráquea, o la inmunización intraocular puede producir un estímulo a nivel
del tracto gastrointestinal en aves, con la producción de IgA en niveles significativos.
La ruta de administración de una vacuna es de vital importancia e influencia en la
respuesta inmune en el lugar donde el patógeno invade y donde la protección debe ser
más eficaz, especialmente para agentes intracelulares como la Salmonella. Esta bacteria
requiere de estrategias de vacunación que activen herramientas múltiples del sistema
inmune tanto la inmunidad innata como la adquirida.
Ha sido ampliamente demostrado que la respuesta inmune protectora en las mucosas es
más eficaz cuando es inducida por esa misma vía, es decir a través de vacunaciones orales,
intranasales o intracloacales, porque el sistema común de las mucosas en las aves ha sido
demostrado y las respuestas incluyen no sólo el sistema inmune innato, como la
producción de anticuerpos en la mucosa SIgA especialmente y linfocitos citotóxicos
localizados en los tejidos próximos a la inmunización necesarios para una protección
eficiente.
Como las superficies mucosas son el mayor lugar de entrada de muchos agentes
infecciosos, es importante el conocimiento del sistema inmune intestinal para entender
los mecanismos de protección de las vacunas y las nuevas estrategias de protección
utilizando productos vivos atenuados aplicados por la ruta natural de infección.
BIBLIOGRAFIA CONSULTADA
1. ANDREW POTTERA, PHILIP GRIEBEL Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides, Molecular Immunology 46 (2009) 3163–3170.
2. ATTERBURY, R.J., DAVIES, R.H., CARRIQUE-MAS, J.J., MORRIS, V., HARRISON, D., TUCKER, V. and ALLEN V.M. (2010) Effect of delivery method on the efficacy of Salmonella vaccination in chickens. The Veterinary Record 167:161-164
3. BEAL R., POWERS C., WIGLEY P., BARROW P., KAISER P. AND SMITH A. A Strong

Antigen-Specific T-Cell Response Is Associated with Age and Genetically Dependent Resistance to Avian Enteric Salmonellosis, INFECTION AND IMMUNITY, Nov. 2005, p. 7509–7516 Vol. 73, No. 110019-9567/05/$08.00_0 doi:10.1128/IAI.73.11.7509–7516.2005.
4. BERGHAUS, R.D., THAYER, S.G., MAURER, J.J. and HOFACRE C.L. (2011) Effect of vaccinating breeder chickens with a killed Salmonella vaccine on Salmonella prevalence and loads in breeder and broiler chicken flocks. Journal of Food Protection 74:727-734
5. BERNDT A., WILHELM, A., JUGERT, C., PIEPER, J., SACHSE, K. and METHNER U. (2007) Chicken cecum immune response to Salmonella enterica serovars of different levels of invasiveness. Infection and Immunity 75:5993-6007.
6. BROWNLIEA R., ZHUB J., ALLANA B., MUTWIRIA G., BABIUKC L., Salmonella pathogenicity island 2 type III secretion systems play a major role in pathogenesis of systemic disease and gastrointestinal tract colonization of Salmonella enterica serovar Typhimurium in the chicken, Avian Pathology (June 2007) 36(3), 199_203
7. CHAUSSÉ, A.M., GREPINET, O., BOTTREAU, E., LE VERN,Y., MENANTEAU, P., TROTERAU, J., ROBERT, V., WU, Z., KERBOEUF, D., BEAUMONT, C. and VELGE P. (2011) Espression of TLR4 and downstream effetors in selected cecal cell subpopulations of chicks resistant or susceptible to Salmonella carrier-state. Infection and Immunity, doi: 10.1128/IAI.00025-11.
8. CHADFIELD M., BROWN, D., AABO, S., CHRISTENSEN, J. P. and OLSEN J.E. (2003) Comparison of intestinal invasion and macrophage response of Salmonella Gallinarum and other host-adapted Salmonella enterica serovars in the avian host. Veterinary Microbiology 92:49-64.
9. CRHANOVA, M., HRADECKA, H., FALDYNOVA, M.,MATULOVA M., HAVLICKOVA, H., SISAK, F. and RYCHLIK I. (2011) Immune response of chicken gut to natural colonization by gut microflora and to Salmonella enterica serovar Enteritidis infection. Infection and Immunity, doi:10.1128/IAI.01375-10
10. DESIN T., KÖSTER W AND POTTER A., Salmonella vaccines in poultry: past, present and future
11. DIEYEY., AMEISS K., MELLATA M AND CURTISS III R., The Salmonella Pathogenicity Island (SPI) 1 contributes more than SPI-2 to the colonization of the chicken by Salmonella enterica serovar Typhimurium, BMC Microbiology 2009, 9:3 doi:10.1186/1471-2180-9-3 Expert Rev. Vaccines 12(1), 87–96 (2013)
12. JOHNSTON C., HARTLEY C., SALISBURY A.M., WIGLEY P. Immunological Changes at Point-of-Lay Increase Susceptibility to Salmonella enterica Serovar EnteritidisInfection in Vaccinated Chickens, PLOS ONE | www.plosone.org 1 October 2012 | Volume 7 | Issue 10 | e48195
13. KWON, H.J. and CHO. S.H. (2011) Pathogenicity of SG 9R, a rough vaccine strain against fowl typhoid. Vaccine 29:1311-1318.
14. ARPAIA, N., GODEC, J., LAU, L. SIVICK, K.E., MCLAUGHLIN, L.M., JONES, M.B., DRACHEVA, T.
15. PETERSON, S.N., MONACK, D.N. and BARTON. G. M. (2011) TLR signaling is required for Salmnonella Typhimurium virulence. Cell 144:675-688. BARROW P.A.

(2007) Salmonella infections: immune and non-immune protection with vaccines. Avian Pathology 36:1-13
16. KOLESAROVA M., SPISAKOVA V., M. MATULOVA M. , CRHANOVA M., SISAK F.,RYCHLIK I. Characterisation of basal expression of selected cytokines in the liver, spleen, and respiratory, reproductive and intestinal tract of hens, Veterinarni Medicina, 56, 2011 (7): 325–332
17. MCKELVEY J.,YANG M, JIANG Y, ZHANGA S., Salmonella enterica Serovar Enteritidis Antimicrobial Peptide Resistance Genes Aid in Defense against Chicken Innate Immunity, Fecal Shedding, and Egg Deposition, Infection and Immunity December 2014 Volume 82 Number 12 p. 5185–5202
18. METHNER U., BAROW, P.A., BERNDT, A. and RYCHLIK. I. (2011) Salmonella Ennteritidis with double deletion in phoPfliC-A potential live Salmonella vaccine candidate with a novel characteristics for use in chickens. Vaccine 29:3248-3253.
19. NANDRE, R.M., MATSUDA, K., CHAUDHARI, A.A., KIM, B. and HWA LEE J.A. (2011) Genetically engineered derivative Salmonella Enteritidis as a novel live vaccine candidate for salmonellosis in chcikens. Research Veterinary Science, doi:10.1016/..rvsc.2011.11.005.
20. NERREN J., HAIQI HE, GENOVESE K., KOGUT M. Expression of the avian-specific toll-like receptor 15 in chicken heterophils is mediated by Gram-negative and Gram-positive bacteria, but not TLR agonists, Veterinary Immunology and Immunopathology 136 (2010) 151–156
21. PENG, W., SI, W., YIN, L., LIU, H., YU, S., LIU, S., WANG, C., CHANG, Y., ZHANG, Z., HU, S. and HU Y. (2011) Salmonella enteritidis ghost vaccine induces effective protection against lethal challenge in specificpathogen-free chicks. Immunobiology 216:558-565
22. PENHA FILHO R.A., DE PAIVA, J.B., DA SILVA, M.D., DE ALMEIDA, A.M. and BERCHIERI. A. J. (2010) Control of Salmonella Enteritidis and Salmonella Gallinarum in birds using live vaccine containing attenuated Salmonella Gallinarum mutant strain. Vaccine 28: 2853-2859.
23. REVOLLEDO L. Avian Salmonellosis, vaccines and immune mechanisms of protection: present and future perspectives. XXIV World´s Poultry Congress 5 - 9 August - 2012 • Salvador - Bahia – Brazil
24. REVOLLEDO L. DVM, Sc., PhD. Vacunas en la Prevención de Salmonelosis Aviares Parte IVLiliana Revolledo DVM, Sc., PhD, Actualidad Agropecuaria, Lima-Perú 21/10/2013.
25. SHAUGHNESSY, R.G., MEADE, K.G., CAHALANE, S., ALLAN, B., REIMAN, C., CALLANAN, J.J., FARRELLY, C.O., Innate immune gene expression differentiates the early avian intestinal response between Salmonella and Campylobacter, Veterinary Immunology and Immunopathology (2008), doi:10.1016/j.vetimm.2009.06.007
26. SILVA E.N., SNOEYENBOS, G.H., WEINACK, O.M. andm SMYSER, C.F. (1981) Studies on the use of 9R strain of Salmonella gallinarum as a vaccine in chickens. Avian Diseases 25:38-52

27. TÉLLEZ T. Y DOMÍNGUEZ L.. Vacunas altamente inmunógenas: el método más eficaz de lucha frente a Salmonella, SELECCIONES AVÍCOLAS • MAYO 2012 • Pág. 17
28. VANDEPLAS, S., DUBOIS DUPHIN, R., BECKERS, Y.,THONARTY, P. and THÉWIS, A. (2010) Salmonella in chicken: current and developing strategies to reduce contamination at farm level. Journal of Food Protection 73:774-785.
29. VARMUZOVA K., ELSHEIMER MATULOVA M., SEBKOVA A., SEKELOVA Z., HAVLICKOVA H., SISAK F., BABAK V., RYCHLIK I., The Early Innate Response of Chickens to Salmonella enterica Is Dependent on the Presence of O-Antigen but Not on Serovar Classification, PLOS ONE | www.plosone.org , April 2014 | Volume 9 | Issue 4 | e96116
30. WIGLEY P. Immunity to bacterial infection in the chicken, Developmental and Comparative Immunology 41 (2013) 413–417
31. WIGLEY P.,HULME S., ROTHWELL L.,BUMSTEAD N., KAISE P., AND BARROW P. Macrophages Isolated from Chickens Genetically Resistant or Susceptible to Systemic Salmonellosis Show Magnitudinal and Temporal Differential Expression of Cytokines and Chemokines following Salmonella enterica Challenge, INFECTION AND IMMUNITY, Feb. 2006, p. 1425–1430 Vol. 74, No. 2, 0019-9567/06/$08.00_0 doi:10.1128/IAI.74.2.1425–1430.2006.