Potencial de membrana 2011
31
Centro Científico y Tecnológico de Excelencia Centro de Envejecimiento y Regeneración, CARE Laboratorio de Neurobiología Molecular Facultad de Ciencias Biológicas Pontificia Universidad Católica de Chile Bases iónicas del potencial de membrana
-
Upload
jorge-parodi -
Category
Education
-
view
939 -
download
1
Transcript of Potencial de membrana 2011

Centro Científico y Tecnológico de Excelencia
Centro de Envejecimiento y Regeneración, CARE
Laboratorio de Neurobiología Molecular
Facultad de Ciencias Biológicas
Pontificia Universidad Católica de Chile
Bases iónicas del potencial de membrana

Musschenbroeck "botella de Leyden”Abad Nollet
SwammerdamFranklinGalvani
Galvani-VoltaHermann Ludwig Ferdinand von Helmholtz
Nobili, galvanometroOhm
FaradayBerstein, potencial de accion
Tschagovetz- Potassium*Overton- sodium
Neher y Sakmann, Nobel 1991

Ligandos
Efectos
Ca+2
Ca+2
quinasas
Proteínas
Proteínas

CNQX + Bic
CNQX + Bic
+ TTX

Centro Científico y Tecnológico de Excelencia
Centro de Envejecimiento y Regeneración, CARE
Laboratorio de Neurobiología Molecular
Facultad de Ciencias Biológicas
Pontificia Universidad Católica de Chile
Electrónica, electricidad y células

Trabajo, Desplazamiento resultante de la aplicación de una fuerza. Su unidad Joule, que son Newton por metro
Por ende, obtenemos Peso que seria Masa por Aceleración
Peso es Masa x 9.8 m/s2
Coulomb, carga que se mueve en un segundo, por la sección transversal de un conductorDe donde obtenemos que F=KQq/r2
en donde K es el medio donde se mueve la carga y Q son las cargas.

C es Coulomb y corresponde a 6.25x1018 electrones
e es electrones y corresponde a -1.5x10-19 C
K es una constante que vale 9x109 N x m2/C2
Campo eléctrico, se define el lugar donde un carga eléctrica sufre un fuerza eléctrica
E =F/Q, pero F=QERedefiniendo F, nos queda que E=KQ/r2

Volt, será la relación entre la energía potencial del campo y la cargaSera entonces Joules x CPor ende nos queda V=KQ/r
Una unidad gigante, para los electronicos, que la remplazan por eVolt = -1.6x10-19 Joules
Capacitancia = Q/V

Capacitor. Dos conductores que transportan cargas iguales pero opuestas en signo
Corriente, definido como el movimiento de cargas por unidad de tiempo I=Q/t1 amperio = 1Coulomb/1 seg
EjercicioPasa una corriente de 1 amperios por 5 segundo, en un conductor. Cuanta masa de electrones se movió ( 9x10-31Kg)
0,28 pG

RIV
QcapQionQlQt
dT
dVCme
V
RQ
La corriente que circula por un conductor, es directamente proporcional a la diferencia de potencial en sus puntos extremos

Capacitancis y resistencia, son propiedades independientes de un conductorDonde resistencia es = ρ(I/A) Donde I es la longitud y A el area del conductor.
Resistividad, ρ = RA/I o sea Ωm

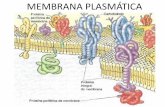
Dendritas: canales pasivos permeables a K+
K+

V
QCm
Corriente del condensador
dt
dVC
dt
dQI m
Corriente del ion (supongamos un solo ion)
Dos componentes para el potencial debido al paso de iones:
1. Potencial de Nerst (debido a la diferencia de concentraciones
del ion en el exterior y el interior de la neurona)
int][
][
ion
ionLn
zF
RTV extNerst

2. Potencial debido a corriente eléctrica (asumimos Ley de Ohm)
ionOhm RIV
ionNerstOhmNerst RIVVVV
Potencial total del ion
)( NerstNerst
ion VVgR
VVI
Corriente del ion

Masa……. No es importanteMovimiento de Cargas
Potencial de equilibrio o nerst, define “la fuerza” de un ion, o energía potencial del mismo
Goldman, incluye “varios” iones y introduce permeabilidad
Hodkin-Huxley introduce el “gating” en un modelo complejo de relaciones de cambio en el tiempo. Modela el Potencial de acción


Si ves que algo gira – ¡¡¡necesitas vacaciones!!!Mira detenidamente cada circulo separadamente y veras que ellos estan
¡quietos!

Canales, responsables de propiedades eléctricas

Membrana neuronal como circuito
Ley de Kirchoff (conservación de la carga)
0 ionC II

V
QCm
Corriente del condensador
dt
dVC
dt
dQI m
Corriente del ion (supongamos un solo ion)
Dos componentes para el potencial debido al paso de iones:
1. Potencial de Nerst (debido a la diferencia de concentraciones
del ion en el exterior y el interior de la neurona)
int][
][
ion
ionLn
zF
RTV extNerst

2. Potencial debido a corriente eléctrica (asumimos Ley de Ohm)
ionOhm RIV
ionNerstOhmNerst RIVVVV
Potencial total del ion
)( NerstNerst
ion VVgR
VVI
Corriente del ion


Nature 1963, Rojas & Luxoro
Demostración de la naturaleza, péptidica de los canales iónicos.

Tipos de canales iónicos. La clasificación de canales iónicos se ha realizado tradicionalmente siguiendo un criterio genético, No obstante, el campo de la cristalografía aplicada a los canales ha avanzado y está avanzando enormemente (y prueba de ello es el reciente Premio Nobel en el año 2003 concedido a Roderick McKinnon por su importante contribución).
Los canales iónicos se pueden clasificar en las siguientes familias o superfamilias

a) La superfamilia de canales iónicos sensibles al voltaje.
a.1) Canales de potasio• Subunidades de 6 helices transmembrana (6TM). “delayed
rectifiers” (Shaker y sus homólogos) y los “slow delayed rectifiers” (KCNQ), los canales de potasio dependientes de Calcio y voltaje (BK, SK e IK), los canales ether-a-go-go (eag o KCNH). Asimismo, pertenecen a esta subfamilia los canales de cationes no selectivos HCN y CNG. • Subunidades de dos hélices transmembrana (2TM) o KCNH. Son
canales rectificadores anómalos o hacia dentro, divididos a su vez en “clásicos” y sensibles a ATP (Kir 6.1) o acoplados a proteínas G (Kir 8.1). • Subunidades de 4 segmentos transmembrana (4TM) , TWIK o
KCNK. También conocidos como canales “de leak”, que presentan dos poros en lugar de uno.

a.2) Canales de calcio . Los componentes de esta familia se diferencian en su dependencia de voltaje, velocidad de inactivación, selectividad iónica y sensibilidad a determinados fármacos. Estos canales se clasifican en: • Canales tipo L. Deben su nombre a que presentan una gran (“Large”) conductancia de canal.
Su función es mediar la entrada de calcio en las células que se contraen o secretan en respuesta a largas o constantes despolarizaciones. • Canales tipo T. Denominados así porque producen una corriente Transitoria y una
conductancia muy pequeña (“Tiny”). Inactivan muy rápido y sólo se observan al principio de pulsos relativamente negativos. • Canales de calcio neuronales (tipo N). Presentan características intermedias entre los tipos L
y T. Dentro de este grupo, canales tipo P/Q sensibles a w-Aga IVA; y canales tipo R que son Resistentes a estas toxinas. Son los canales que se expresan de forma dominante en terminales presinápticas que secretan neurotransmisores en respuesta a potenciales de acción presinápticos.

a. 3) Canales de Na + . Este canal es uno de los principales responsables de la transmisión del potencial de acción. La familia de canales de Na + dependientes de voltaje es la que presenta menor diversidad, y todos sus miembros muestran propiedades netamente voltaje-dependientes muy similares en todos los tejidos en los que se expresa.
a.4) Canales de Cloro dependientes de voltaje. Los canales de cloruro son permeables a bastantes aniones de pequeño tamaño, como
Br-, I-, NO3-, HCO3- y SCN-, así como a varios ácidos inorgánicos. Por lo tanto podría generalizarse su denominación a “ canales aniónicos ”. Los canales de Cloruro dependientes de voltaje se dividen en cuatro clases
dependiendo de si son activados por Ca 2+ intracelular, hiperpolarización de la membrana, disminución del volumen celular o fosforilación dependiente de ATP. ClC y el CFTR, responsable de la secreción en epitelio, regulado por fosforilación. El
primer gen en mamíferos fue el ClC-1, en el músculo liso.

c) Canales operados por ligando. Este grupo engloba a una serie de canales que están especializados en la mediación de
respuestas químicas rápidas en la transmisión sináptica. El receptor de acetilcolina (AchR), el receptor de glutamato (GluR), receptor de glicina (GlyR), el receptor del acido g -aminobutírico (GABA A R), serotonina (5HT 3 R)
Los receptores de Glutamato se clasifican a su vez en tres grandes grupos: N-metil-D aspartato (NMDA), kainato y AMPA (ácido a -amino-3-hidroxi-5-metil-4-isoxalon propiónico). En general, los canales operados por ligando se expresan en la membrana postsinaptica de forma
concentrada y se abren rápidamente después de la unión de varias moléculas de agonista. La mayoría de estos canales poseen una amplia selectividad iónica, preferentemente a cationes monovalentes o aniones, y por lo tanto su función es estimular o inhibir la excitación eléctrica de la célula postsináptica. El receptor de NMDA es una excepción: es más permeable a calcio que a cationes monovalentes, y parece que su papel en el control de la concentración de calcio intracelular es mucho más importante que el electrogénico.

Ligandos
Efectos
Ca+2
Ca+2
quinasas
Proteínas
Proteínas

Patch-Clamp, célula completa
V clamp
I célula
+
–
R f
Neurona