
Precipitación, capacidad de carga y potencial de uso de ungulados en un bosque tropical seco del...
25
Consuelo Lorenzo Eduardo Espinoza Jorge Ortega EDITORES
-
Upload
salvador-mandujano-rodriguez -
Category
Documents
-
view
265 -
download
9
Transcript of Precipitación, capacidad de carga y potencial de uso de ungulados en un bosque tropical seco del...

Consuelo LorenzoEduardo Espinoza
Jorge OrtegaEDITORES

PRECIPITACiÓN, CAPACIDAD DE CARGAY PRODUCCIÓN DE UNGULADOS
EN UN BOSQUE TROPICAL SECO DEL PAciFICO MEXICANO
Salvador Mandujano
De¡XlfwmellfO de Biodil"el"sidad y Ecología Animal, fl/sli/1I10 de Ecolugía A. C. Kili. 1.5 Cflt"l"(!lemAllligll(1 a COa/eJ}ec #351, Congregación (Iel Haya. Apm"wdo 1>0.\'la/63. el' 91070 XlIlapa,
Veracrll::, México. CE: safl'(ulvr.mlllld/ljaflo@illecol.('(lu./IIx
Resumen: Se analizaron las interrelaciones entre la precipitación. la disponibilidad de alimento,la capacidad de carga y la producción pOlencial para aprovechamiento humano del venado colablanca (Ot/ocoilells virgillimllls) y pecad de collar (Peca"; tajacu) Cllllll bosque tropical seco deChamela, México. Durante los allOS del El NiJio la lluvia en la época húmeda (junio a octubre)disminuyó y aumentó durante la época seca (noviembre a mayo); mientras que durante La Nilia seohservó UIl efecto contrario. 1'.1'1.<;0 podría tener un efecto ;ldieional sohre la disronihilid;ld dealimento para los ungulados, principalmente en la época seca. Se definió capacidad de cargacomo la densidad en el punto de equilibrio del crecimiento poblacional más el número de animalesremovidos por los depredadores. La capacidad de carga se estimó en 16.5 a 17.2 vcnados/km2 y9.3 II 9.5 pccaries/k.m2• y una biomasa combinada de 841 a 874 kg/kmz. Se estimó un;t producciónpotencial para aprovechamiento humano de 2.1 venadoslkm l y 4.4 pccaríes7km l
. Los datos indiClln que en el bosquc tropical seco de Chamela la densidad y biomasa de los ungulados silvestrespuede ser similar o mayor en comparación con otros bosques neotropicales.
Palabras clave: Bosque tropical seco, capacidad de carga. densidad, liNSO. Ot/ocoifeusJlirgilliclJIlIS, rmixima cosecha sustentablc, Pecari /(Ijac/{. precipitación.
Abst raet: Data are provided on the illterrelatiOllS amOllg minfal!. food. carrying capacity andpotential productioll for sustainablc human use of white-tai led deer (Ot/ocoilells ¡-¡rgillialllls) andcollared peccary (PeCClri lajaclI) in a proteclcd tropical dry forest at Chamela 011 the Pacific coastof Mexico. During El Nilio years raillfall decrcascd during the wet scason (Junc to OClober) andincreased during the dry season (Novernber to May), with the opposilc elTecl during La Niñayears. The results provide evidence tha! ENSO events alTeel deer food availability. partieularly inthe dry season. The carrying capacity was defined as the equilibrium dcnsity plus thc numbcr ofanimals removed by predators. Carrying capacity estimates wcre 16.5 to 17.2 decrlkmzand 9.39.5 peccarieslkm1, representing a eombined biomass of841-874 kglkm2. A potcntial productionfor human use of2.1 dcerlkm1 and 4.4 peecaricslkm" WllS estimatcd. The dala suggest that, in theprotected tropical dry forest of Chamela, the dcnsity and biomass of wild ungulates can maintaina similar or greatcr dCllSity and biornass than otller Neotropical forests.
Kcy words: Carrying capacity, EN50; dCllSity; Mexico; Ot/ocoileus I'irgillialllls: Pecari lajacl/;potential production; minfall; tropical dry rores!.
LoreIlZO. c.. E. Espino7.a y J. Onega (ed$.). 2008. Amllcl'.~ en l'i Esllldio de /o.v Ml,mifero.< dt, México. Publicaciones·1 l' ' lCiadÓI1 I\",,~;···"'a ¡le MaS!l)7.oologia. A. c., México. D. F.

Ungulados en selva baja
INTRODUCCIÓN
Importancia de la lluvia para los unguladosL:l variación clinuitic<l asociada con los fenómenos meteorológicos conocidos como la Oscilacióndel Atlántico Norte (N,\O. "North Atlantic Oscillation") y la El Niii.o/Oscilación del Sur (ENSO. "ElNiii.o/Southern Oscillatiol1"). ticnen gran influencia sobre la producción primaria de la biosferJ ysobre la dinámica poblacional de muchos organismos;1 nivel mundial (Behrel1feld el al., 2001;Stenseth el al., 2003). Estas fluctuacioncs clirmiticas a gran cscala afectan el clima local a travésde patrones de tc!eeonexión (Stenseth er al.. 2003). Por ejemplo, los episodios ENSO están asociados
con los patrones de lluvia en la región del Pacifico lo cual afecta tamo a los ecosistemas marinoscorno terrestres (Jaksic. 2001).
Tanto el c!illln corno los eventos del ENSO pueden tener consecuencias importantes en lademografia de herbívoros terrestres (e.g.. Owen-Smith. 1990: Langvatn el al.. 1996; S:cther, 1997;McKinlley el al.. 2001: Marshal er al., 2002: Georgiadis ef al.. 2003: Ogutu y Owen-Srnith. 2003).Se debe a que estos eventos influyen sobre la fenología de las plalltns lo cual afecta la calidad y 1:1producción de biomasa, aspectos que detenninan la capacidad de carga del habit;ll (Coe et al..1976; Sinclairel al.. 1985; Langvatn el al.. 1996: S:cther. 1997; Post y Stenseth, 1999). Evidenciasde estos efectos sobre las poblaciones de ungulados han sido generadas principalmentc cn regionestempladas. en varias partes del mundo. y en regiones tropicales de África. En col1lraste. existenmuy pocos estudios en 1" región neotropical (c.g., Wright el al.. 1999: Ticktin. 2003).
Cal}acidad de cargaEl COl1cepto de capacidad de carga (A., es usualmente empleado en el manejo de fauna silvestre.
Sin embargo. dependiendo de los objetivos y del enfoque, puede variar en su definición, 10 quetiene implicaciones importantes en el método para estimar K (Millcr y Wentwonh, 2000). Ladefinición corminlllente empleada de K es: el número m:íximo de animales que una poblacióndada puede ser sostenida en función de los recursos disponibles (McCullough. 1992). Desde unaperspectiva demogr.ifica el término se refiere a la densidad en equilibrio a la que el crecimiento de
la población se estabiliza cuando las tasas de natalidad y mortalidad son iguales (Ak~akaya el al..
1999). En contraste, desde una perspectiva de uso y disponibilidad de habitat, K se refiere almaximo número de individuos de una población que puede ser sostenido sin que exista un deteriorodel habitat (Gallina. 1993: Groot-l3minderink y H:lzeborek. 1995). En el caso de ungulados. se haasumido que K depende del valor nutricional de las plantas (MeCa]] el al.. 1997).
Sin embargo. existe diferencia entre lo que se cntiendc por capacidad de carga desde una
perspectiva ecológica que desde una visión económica (Scoolles, 1993). Desde esta ultima. elconcepto de K toma en consideración el nllJllero max imo de animales que pueden existir sin quehaya una repercusión económica negativa para intereses hum:ll1OS específicos (\Vest y Parkhurst,2002). 01ra definición es el número m[¡ximo de animales que pueden cosecharse de manerasustentable (DeC:llesta y Stout. 1997). En el caso de comunidades humanas habitando bosques

M:lI1dujano 639
tropicales, K es definida como el nt:i.ximo número de personas que pueden ser mantenidas sin quese afecte la biodiversidad de un Silio (Robinson y Bennet. 1999). Otras definiciones más delalladaspueden ser consultadas en los trabajos de McCullough (1992) y Miller y Wentworth (2000).
Ilroducción máxima sustentableEl modelo más común para evaluar la sustenl,lbilidad de la cacería de fauna silvestre en
regiones tropicales es el conocido como de crecimiento poblaciona1. propuesto por Robinsoll yRcdford (1991) O modelo de producción (Bodmer y Robinson. 200"'). Éste ha sido aplicado ennumerosos cSludios en África y Latinoamérica (e.g., Filzgibbon el 01.. 1995: AIvard ('Ia/., 1996;
Wilkie el al.. 1998) y rccientemcnle en Mcxico (Namnjo el al.. 2004a: Mandujano. 2007). Estemodelo requiere información especifica de la capacidad de c:lfga (Al. la lasa finita m:ixirna decrecimiento poblacional (A",) y la densidad (D). con el fin de estimar la máxima cosecha sustel1lable(.usr. "Maxil1lul1l $uslainable Yield"; Robinson y Redford. 1991). Además. en este modelo seasume que las poblaciones tienen un crecimiento denso-dependie11le y alc,Ltlza la produccióllllláximacuando la población está al 60"10 de su capacidad de carga. es decir O.6K. Por lo t:mto, la producción
máxima (P""") se caleula multiplicando la densidad (estimada como 0.610 por la lasa finita decrccimiellto poblacional (A"..). y luego se le SUSlrae la densidad del año anlerior. es decir: p.." =
(O.6K . Á~) - O.6KHabitualmente K se estima a partir de muestreos de poblaciones en Silios pocos perturbados.
y se considera que la densidad poblacional se encuentra a su capacidad de carga. es decir K'" D.Sin cmbargo. dos factores podrían afectar este supuesto. Primcro. las poblaciones en equilibro no
necesariamcnte se encuenlran a su capacidad de carga, especialmente en aquellas áreas donde el
crecimienlo poblacional eslá fuertemenle regulado por los depredadores (Bodmer y Robinson.2004). Segundo. la cflpacidad de carga no es un valor estálico sino que cambia depcndicndo de ladinámica. variación estacional. deprcdación y necesidades humanas (McCullough. 1987; 1·lutllery Rungc. 2004). En consecucncia, en sistemas nalurales habilualmenle existen llllcluaciones delas condiciones ambientales y las poblacioncs en realidad lluctllan alrededor de K. Por lo tamo.eslirnar sólo una vez la densidad en áre¡¡s muy poco pcrturbadas podria llevar 11 una estimaciónsesgada dc K. Desafortunadamcnte. existen muy pocos estudios en el Ncolrópico donde se hayaIlloniloreado los cambios ambientales con la finalidad de considerar este aspecto tanto en la
eslimación de K como de MSl' (Bodmer y Robinson. 2004).
Ungulados de importancia económica en MéxicoEl venado cola blanca (Odocoilells \'irgilliallll.~)y el pecari de collar (PeClll"i ((IjoclI) son las
dos especies de ungulados que tienen la mayor {lrca de distribución geogn'lfica en el continenteAmericano (Hall. 198 1: Eisenberg, 1989). Son especies ampliamentc aprovechadas por grupos
indígenas y poblaciones nlraJes para complemento de su diela y para comercio de carne y piel y,en algunos sitios. se les aprovecha como trofeos cinegéticos (Mandujano y Rico-Gray. 1991;Escamilla el al.. 2000, Naranjo el al.. 200411). En consecuencia. evaluar el potencial de producciónde estos ungulados cs un aspecto crucial cn aquellos silios donde se prclcnde aprovecharlos de

640 Ungulados en selva baja
manera sustentable, por ejemplo en las Unidades de Conservación. Manejo y AprovechamientoSustentable de la Vida Silvestre (UMAS) extensivas. Sin embargo, la estimación de K y su variacióna través dclliempo es un aspecto muy poco conocido; ademas, hay muy pocos sitios en los cuales
estos ungulados no están perturbados por presiones alllrópicas. En este sentido, en algunas áreasnaturales protegidas se puede generar infomlllción básica sobre la dinámica de estos ungulados yasi proveer con los datos necesarios para paramclrizar el modelo Robinson y Rcdford (1991).
Ungulados en el bos<)ue tropical seco de ChamelaEl presente estudio se realizó en el bosque tropical seco o selva baja de Chamcla localizado
en la costa dcl Pacifico mcxicano. Hasta la fecha. cslc siguc siendo el Siliocon selva baja dondc sehan realizado los estudios más extcnsos con venado en el pais, y lal vez Latinoamérica. Unasíntcsis dc los trab¡tios sc prescnt¡l cn Mandujano el al. (2002).
La marcada estacíonalidad dc las lluvias es el principal factor que influyen la estructura ydinámica de este ecosistema (Murphy y Lugo, 1986). Las comunidades de pl:mtas en este sitio sonafectadas por la variación interanual de los vicntos Alisios, huracanes del Pacífico y eventos ENSO
(Bullock, 1986; Garcia-Oliva el al.. 1991). Además, la topografía y otros factores influyen en ladisponibilidad de agua en el suelo, produciendo áreas extensivas de bosque tropical seco en las
ladcras y lomas de las montañas y parches de bosque tropical sllbcaducifolio en los arroyos y ríos(Lon el a/.. 1987: Bullock y Solís-Magallanes, 1990). Por lo tamo, este lugar se caracteriza por la
variación teml>oral Y espacial en la disponibilidad y calidad del alimento, fllentes de agua, ycobertura de protección para el pecad y venado cola blanca (Martinez-Rornero y Mandujano,1995; Mandujano y Martinez-Rornero, 1997,2002; Silva-Villalobos el al.. 1999; Mandujano el
al.. 2004: Arceo el al.. 2005). Las dcnsidades promedio del venado cola blanca y el pecari decollar sc han cstimado dc 12 y 9 ind./knr. respectivamente (Mandlljano y Gallina. 1993; 1995b:Mandujano. 1999: Mandujnllo y Martíncz-Romero, 2002). En comparllción con otras rcgionesdel país, cn Cha1l1cla existc relativamcnte una baja presión humana. En contraste, la dcpredaciónpor puma (Pullla COI/color). jaguar (POli/llera 011(:0). ocelotc (Leopordus pardo/is) y coyote (Co/lis lalrol1S: Nur'lcz el al., 2000: Hidalgo~Miharl el a/., 2001; Villa-Meza el al.. 2002) ticne un
papel importantc sobre las poblacioncs dc cstos ungulados.
Objctivos dcl capítuloEn la primera pm1e del prescntc capitulo se analiza el efecto de ENSO sobre el patrón de
precipitación cn el sitio de cstudio empleando datos colcetados en 26 <I1'1os. En la segunda partedcl trabajo se provee de evidencia preliminnr de la importancia dc ENSO el1 modificar ladisponibilidad temporal del alimento empleando datoscolcetados en 4 aijos. En la tercera parte se
reportan cstimaciones de la capacidad de carga y la producción potencial de venado cola blanca ypecari de collar. II p"rtir estinlllr la densidad en equilibrio y el número de animales removidos porlos depredadores. En conSi,,'(:uencin. en cste estudio K no se considera simplemente como lól densidaddel aJ'Io-i. cs decir: K'" D¡. En cambio. K es estimada considerando la rclación denso-dependienteentre la tllsn finita de crecimiento poblllcional y la densidad estimada eOIl datos colectados en 5

Mandujan.,o'-- _ 641
años. En la cuarta parte de este capítulo se analiza la posible variación de K basada en datos dcprecipitación colectados durante 26 alIaS y sc discute cómo podría emplearse para definir cstrategiasde manejo en este tipo de hábilat. Mientras que cn la última parte sc prescnta gráficamentc lafluctuación poblacional en relación a la precipitación y biomasa vegetal. El enfoque conceptual yde análisis del presente capítulo (Fig. 1) no se había abordado previamente en México y puedeconstituir una aportación interesante en el Neotrópieo (Mandlljallo y G¡lllina. 2005; Mandlljano.
2006. 2007).
•Capacidad
de carga
Topogr.ofí", ouelo.faclOlCS h,SlÓf1COS,
,mclOd,ma
V""IOS Alisio»H"r:>cancli Padfi<o
l:NSO
•
•
•Lluvia
•Plantas
biomasa/calidad
•o Densidad en Tas;o dee
nalalidad.o •~
equilibrio largo sob",,-,"cncoa.~ mot!alidado plazo •~ •~eo •~
" Densidad t>cp",dadorcs y Ie.s
aclual enTelmeJadeso.¡;~
Producciónmáxima
sustentable. MSY
•Nllmero deanimales a
extraer•
~Icrtadu.
leg,slaclÓn.cul'"",,
' .
Figurn l. Represe11laeión diagnnmitiea del enfoque conceptual del prcsente esludio.

642
ÁREA DE ESTUDIO
Ungulados en selva baja
El trabajo se realizó en la Estación de Biología Chamela de la Universidad Nacional Autónoma de
Mex;co. situada en la costa de Jalisco. México (19" 30' N Y 105" 00' W). La Estación tiene unaextensión de 3200 ha y fOfma parte de la Reserva de la Biosfera Chmncla-Cuixmala(www.ibiologia.1l113m.rnx/cbcharnclalHIST2.html). El sitio se camcleriza por una topografía irre
gular. con pendientes de 21" a 34"; el intervalo altitudinal es de 30 a 580 m. aunque la mayor parte
del terreno no sobrepasa los 150 m. El clima es tropical con una marcada estacionalidad. Latemperatura media afiual es 25"C, con los meses más calientes cntre mayo y septiembre (Bullock,
1986). La precipitación promedio anual de 1977-1997 ha sido de 740 mm (SO = 196). La temporada
de lluvias inicia la última semana de junio. y de julio a octubre cae 80% de la lluvia anual y tiene
una duración lllcdia de 126 días.
La vegetación dominante es el bosque tropical caducifolio el que se localiza en lomcríos con
suelos someros, el estrato arbóreo tiene una altura de 4 a 15 m, presenta un sotobosque biendesarrollado y numerosas especies arbóreas y arbustivas pierden sus hojas durante la epoca seca.
Algunas de las especies más comunes son el botoncilio (Cordia alliodora [Ruiz y Pav.] Oken).
vara blanca (LonchocarplIs lanceo/mus Benth.) e iguanero (Caesalpil/ia erioslachys Benth.).
También se encuentra el bosque tropical subperennifolio que se desarrolla a lo largo de los arro
yos principales, en los lugares protegidos sobre suelos profundos y el estrato arbóreo tiene ulla
altura de lO a 25 m. Las especies más comunes son el eulebro (AslroniulII gral·eo/el1s Jacq.),
mojote (Brosimu/11 alicaslmm Sw.) y tempisque (Sideroxy/on capiri A. OC.; Lott ef al.. 1987).
MÉTODOS
Análisis de la precipitaciónEn este estudio se consideró como inicio del año en el mes cuando cae la primera lluvia
imponante lo cual, en esta localidad, típicamente sucede alrededor de la tercera semana de junio
(Bullock. 1986). La probabilidad de que llueva 100 mm define una estación húmeda de cinco
meses (junio a octubre) y una seca de siete meses (noviembre a mayo: Garcia-Oliva ef al., 1991).
Datos de precipitacióll del periodo 1977-2003 se obtuvieron de la estación climatológica del sitio.
Infonnación de ENSO se obtuvo de la base de datos del Oepanment of Commerce de Estados
Unidos (hup://www.pmel.noaa.gov/tao/elnino/el-boy-story.htm1).asicomo del Climate Diagnos
lics Center (http://www.cdc. noaa.gov.lENSO/enso.difTcrenLhtm1) y de University ofll1inois
(hup:l/www.ww20l0.atmos.uiuc.edtt/(G I)/wwhlpr/guides/mtr/e1n1def.rxml). De estas bases se
obtuvieron los palrones tcmporalesdcl ¡ndice de la Oscilación del Sur (SOl. "Sotlthem OsciJlalion
Index") del periodo 1977-2003. El SOl fue calculado agrupando datos de junio a mayo. Valores
negativos del 501 están asociados con los eventos del El Nilio (fase caliente), y valores positivos
con La Niña (fase fria: Stenseth el al.. 2003). La relación entre el 501 y la lluvia anllal y estacional,
fue analizada empleando correlaciones lineales simples.

Mandujano 643
Estimación disponibilidad de alimentoPara estimar la disponibilidad del alimcnto se establecieron aleatoriamcnte 25 parcelas
permancntes (10 x 10 m cada una) en el bosque tropical scco y 25 cn el bosquc tropicalsubcadllcifolio. Pam mayordctalle consultc Mandujano el al. (2004). La disponibilidad del alimcntosc estimó como biomasa en pie y como riqucza de cspccies. Como ambas variables cstánsignificativamentc conc1acionadas (r= 0.79. P = 0.02). se cmplcó el indicc de Shannon-Wicner(H') como una medida dc la disponibilidad dcl alimcnto.
Estimaciones de la densidad y biomasa poblacionalPara estimar la densidad poblacional. sc empleó cl método de contco directo dc anirnlllcs cn
transectos diumos durante todo cl mio. La densidad reponada en este capitulo se cstimó con clprograma DISTANCE4.1 vcrsión 2 program (Thomas el al., 2003). Infonmción dctalla de los métodosdc estimación dc ambas espccies de ungulados sc pucdcn consultar en trabajos prcviamcntcpublicados (Mandujano y Gallina. 1993, 1995a; Mandujano, 1999,2007). Para estimar la biomasapoblacional de cada ungulado. se multiplicó la densidad anual por el peso promedio de animalesadultos: 15.7 kg para el pccarí dc collar y 42.1 kg del vcnado cola bllmca (Naranjo el al.. 2004b).
Estimación de la tasa finita de crecimiento poblacionalLa tasa finita de crecimiento anual (A, lambda) se estimó como: 1..= N,./N,. dondc N,,/ es la
abundancia dc individuos cn el año 1+/ y N, cs la abundancia dcl año prcvio. La abund:ll1ci:1 seobtuvo multiplicando la dcnsidad anual por la superficic de estudio (aproximadamente 500 ha).La tasa finita máxima de crecimiento poblacional se estimó como 1..-.-. = e--. La tasa intrínseca dccrecimiento natural (r"",) solo puede ser calculada para poblaciones no limitadas por cl alimento,espacio, competencia por recursos y depredadores (Robinson y Redford, 1986). Se estimó "MMusando la ecuación de Cole (1954. ver Robinson y Redford, 1991). la cual tiene que resolvcrseitcrativamcntc como: 1= e-' ..... + be _""'U (,.!_ be -'-(.... /). donde l/ es la edad a la que ocurre el
primer evento reproductivo. IV es la edad del último evento reproductivo, y b es la tasa anual denacimiento de crías hembras. Los parámetros l/. b, Y 1\1 fueron 1.5,0.69 Y7. Y 1,2. Y7 para el
vcnado y pccad cn este sitio (Manduj:mo y Maninez-Rolllcro, 2002; Mandujano y Gallina, 2005).
Estimaciones de la capacidad de cargaLa capacidad de carga (K) se estimó calculando primero la densidad en cquilibrio (/J,,) de
acuerdo con la relación de denso-depcndencia entre el crecimiento dc la población y 1<1 densidada partir de un modelo logístico (Akpkaya el al., 1999); luego se le sumó el número dc individuosrcmovidos por los deprcdadorcs. P. En consecuenciu en este estudio K - D<q + P. Para calcular ladensidad cn equilibrio se empIcó a la densidad COIllO variable independiente y la tasa finita decrecimiento corno variable dependiente. El análisis se hizo para cada ungulado por separado.Valores de la tasa finita mayores a llllO indican que la población está aumentando, micntras quemenores a uno indican decrecimiento. El punto en el cual la línea de regresión intcrccpta a la lineade remplazamicnto (cuando A= 1) se considcra como la dcnsidad en equilibrio (Ak~akaya et al.,

644 Ungulados en selva baja
1999). Los estudios con depredadores en este silio sugieren que anualmente remueven de 36.742.7%. Yde 9-11 % de las poblaciones de venado cola blanca y pecarí de collar, respectivamente(Nuiicz el al.. 2000: Hidalgo-Mihar! el al.. 2001: Villa-Meza et 01.. 2002). El número de animales
removidos se calculó considerando estos porcentajes multiplicado por la densidad promedio. Elresultado se sumó a la densidad en equilibrio, asumiendo que el porcentaje de remoción es constantea través de 10$ :lilas. El resultado es una estimación de la caJXlcidad de carga.
EslinlllciOIlCS de la Ilroducción potencialLa producción máxima potencial (P ) se estimó como: P = (O.6K • A..J- O.6K (Robinson- -y Rcdford. 1991). La producción para uso humano se consideró como PRR =Q.4QP"",,' El factor
0.40 se empleó considerando que estas especies tienen su último evento rcproductivo antes de los10 de edad (Slade ef at., 1998). En olras palabras, se asume que hasta 40% dc la producciónmaxima puede scr usada sin afectar adversamente a la población.
Análisis de la variación dc K cn función de la lluviaPara cOllocer la posible variación de K durantcel periodo 1977 a 2002. se analizó el patrón de
lluvia asumiendo que la disponibilidad del alimenlO depende de la precipitación (Mandujano,
2006). McCullough (1987) propuso algunos modelos esquematicos donde se puede mostrar ladistribución de frecuencia de la ocurrencia de años "buenos" y anos "malos" en diferentes ambientes.En este trabajo la clasificación de un a110 como "bueno" o corno "malo" se hizo dependiendo de ladcsviación de la lluvia. de un año detenninado, contra el promedio de la lluvia del pcriodo de 26años aquí considerado. La desviación se estandarizó y expresó como porcentaje respeclO a la
mcdia. Entonccs, un valor dc cero representa la media. un valor < Orepresenta un "3110 malo".mientras quc un valor> Orepresenta un "ano bucno". Los valores dc la media. mediana, asimetriay kurtosis fueron empleados como un índice para eVllluar cuando la distribución de frecuencia dela precipitación corresponde a un ambicnte "estable o inestable" y años "buenos o malos", Unadistribución 1cptokúrtica (y~ > O) o platikúrtica (Y
l<O) representan un ambiente altamente estable
o uno alt:mlellte inestable. respectivamente: mientras que una asimetría positiva (Y1
> O) o unaasimetría negativa (y I < O) representa un ambiente malo y un ambiente bueno, respectivamente. El
anúlisis se hizo en el programa STATISTICA para Windows, versión 5.5 (Statsofi lnc. 2000).
RESULTADOS
Patrón dc lluviaEl analisis de datos mostró que de 1977-1984 la lluvia almal estuvo muy cercana al promedio
de los 26 años analizados que corresponde a 755 mm. Se presentaron dos periodos con años secos1985-1988 y 2001-2003. El periodo 1989-2000 fue una década con mueha variación anual en laIluvi:1 (Fig. 2). El promedio de lluvia durante la época hllmeda Gunio a octubre) fue de 665 ± 222mm (rango 368-1288 lllm), mientras que en la época seca (noviembre a mayo) [\Ie de 90 ± 148

Mandujano
1500
-EE 1000~
e"O·0
.ª 500c.·0o
"..O
-•••• 080 :;:~¡ :;: :::
Año
precipitación ..... venado cola blanca -&-pecari de collar
15
"E1O~
."o
5 "C.,5Q
o
645
Figura 2. Serie de tiempo de la lluvia anual de 1977 a 2002 y densidad poblacional del venado cola blanca(e) y del pecari dc collar (O) durante el periodo de estudio 1989 a 1993_
mm (rango 0-721 111m). La precipitación de la lluvia en la época seca fue independiente de la quecayó en la época húmeda previa (,.1 = 0.07, F= 1.73, P = 0.20).
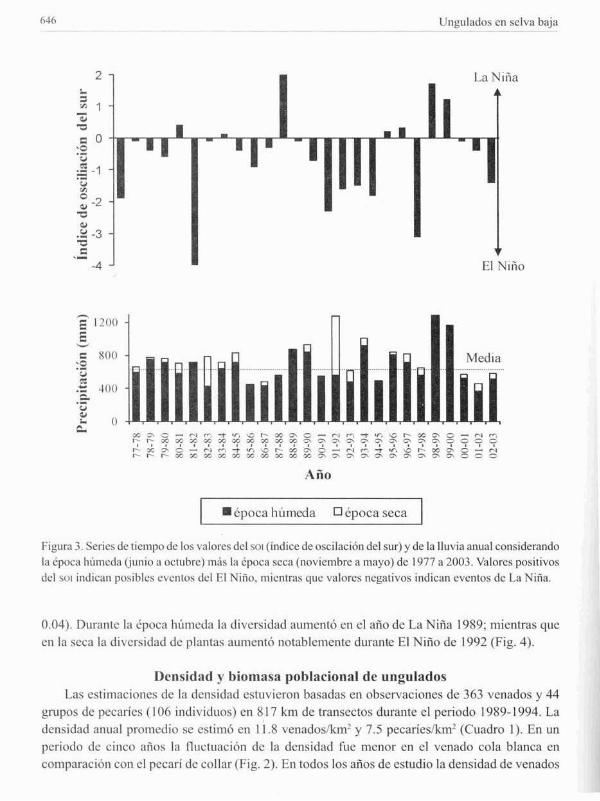
Dumnte los 26 ;"11105 analizados se detectaron tres fuertcs evenlos del El Niiio (SOl> -2): 1982
83, 1991-92, y 1997-1998; Ycinco menos evidentes (SOl -1.5 to -1.9) durante 1977-78, 1992-93,1993-94, 1994-95, y 1998-99 (Fig. 3). La Nina (SOt positivo) fue evidente durante 1988·89. 199899 y 1999-2000. No hubo relación significativa entre el SOl Y la lluvia anual durante el periodo1977-2003 (1.1 =0.07, F= 1.83, P =0.(9). Por el contrario, el SOl estuvo positivamente asociadocon la lluvia de la epoca húmeda (1.1 = 0.40, F= 16.2, P = 0.0005). Durante los años El Nir'lo laprecipitación fue baja en la época húmeda; mientras que durante los anos de La Nir'la la precipitaciónfue alta en esa misma época. Por otro tado, durante la época seca el SOl estuvo negativamenteasociado con la lluvia (,..2 = 0.28, F = 9.4, P = 0.005). En particular. durante El Niño (1982-83 y1991-92) la lluvia fue alta cnla época seca, mientras que en La Niña no hubo precipitación en estamisma época (Fig. 3).
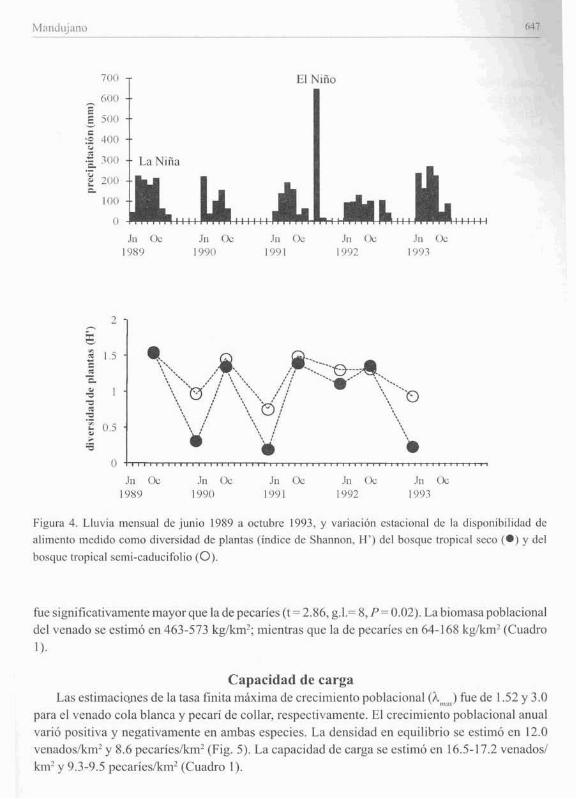
Producción de plantasLa distribución mensual de la lluvia y las estimaciones de diversidad de especies de plantas
durante [as épocas húmedas y sceas del periodo 1989-1993, se presentan en la Fig. 4. La diversidadfue mas :lita en la época h(llneda que en la seca excepto en 1992 (F = 30.9. P = 0.0001). Ladiversidad fue similar en ambos tipos de bosques durante la época húmeda: mientras que en laseca fue más alta en el tropical subeaducifolio en comparación con el tropical seco (F= 5.22, p=
646 Ungulados en selva baja
2 La Niria
",~ 1
"""-5 O
'C;¡
~ -1·0~
0_2~
""~.:: -3
""=-4 El NUlo
E 1200E~
= ",O Media-o·0o.~ 400"-o~
" O..~~~~~~~~~~~~~~~~~~~~~~~99~~~R~"~~~~~~~~g~~~;~~~~~8~8
Año
-epoca húmeda Oepoca seca
Figura 3. Series de tiempo de los valores del SOl (indice de oscilación del sur) y de la lluvia anual considerandola época húmeda (junio ,1 octubre) mús la epoca seC;1 (noviembre a m,IYO) de 1977,12003. V,llores positivos
del SOl indican posibles eventos del El NiTio. mientms que valores neg:llivos indican eventos de La Niña.
0.04). Durantc la época hÚlllcda la divcrsidad aumcntó cn el aTio de La Nilla 1989: micntras quccn la scca la divcrsidad dc plantas aumcntó notablcmcntc durantc El Nillo de 1992 (Fig. 4).
Dcnsi<hld y biomasa poblacional de unguladosLas estimaciones de la densidad estuvicron basadas cn obscrvacioncs dc 363 venados y 44
grupos dc pcearies (106 individuos) en 8 [7 km de transeetos durante el periodo 1989-1994. Ladcnsidad anual promcdio sc eslimó cn 11.8 vcnados/km! y 7.5 pecaríes!km! (Cuadro 1). En unperiodo dc cinco afios la fluctuación dc la dcnsidad fuc mcnor cn cl vcnado cola blanca encomparación con el peeari dc collar (Fig. 2). En todos los años de estudio la densidad de vcnados
r-I;llluujano
700
600
15()()
•:: 400o
".~ )00
.~ 200
100
o
B 15•"...
Jn (),;
19W)
Jn Oc19S9
Jn Oc1990
Jn ().:1990
.In ().,
1991
Jn o..:199)
Jn ().,1992
Jn ~A:
1992
Jn (X:
1993
JII Oc19')1
Figura 4. Lluvia mcnsual dc junio 1989 a octubre 1993. y varillci6n eSlacional de la disponibilidad dealimento medido como diversidad dc plantas (indice dc Shannon. H') dcl bosquc tropical scco (e) y delbosquc tropical scmi-caducifolio (O).
fue significativamente mayor que la de pecaries (1:0 2.86, g.I.:o 8, p:o 0.02). La biomasa poblaciollaldel venado se estimó en 463-573 kglkm~; mientras que l:l de pecarics en 64-168 kglkm2 (Cuadro
1).
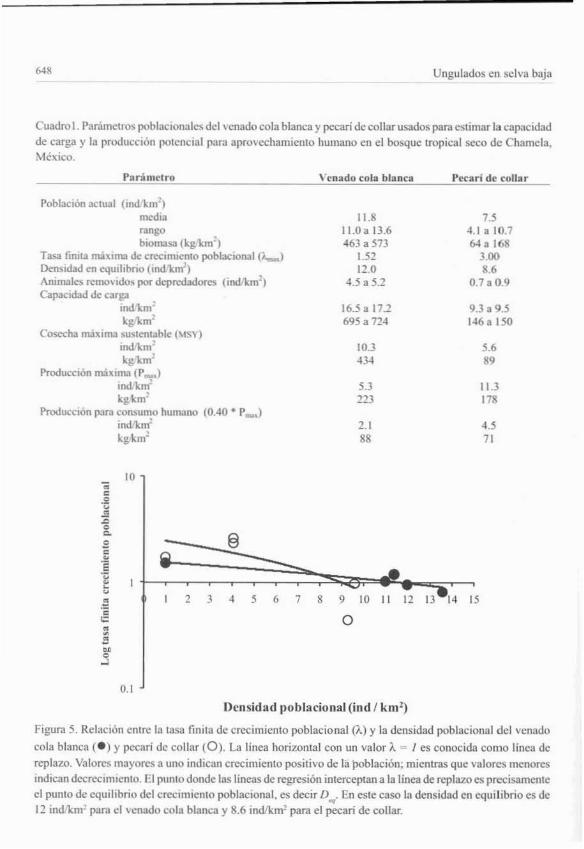
Capacidad de cargaLas estimacio.nes de la tasa finita máxima de crecimiento poblacional (A-..) fue de 1.52 y 3.0
para el venado cola blanca y pecarí de collar. respectivamente. El crecimiento poblaciollal anualvarió positiva y negativamente en ambas especies. La densidad en equilibrio se eslimó en 12.0venadosikm! y 8.6 peeariesikm! (Fig. 5). La capacidad de carga se estimó en 16.5-17.2 venados!km~ y 9.3-9.5 pecaríes/km! (Cuadro 1),
.., Ungulados en selva baja
Cuadro l. Parámclros poblacionalcs del \'cnadocola blanca y pecari de collar usados para estimar la cap.'lcidad
de carga y la producción potencial para aprovechamiento humano en el bosque tropical seco de Charnela.México.
l'lIrimelru
Población aclual (uld.l::ml)mediarangob)¡)masa (kg,'km!)
Tasa finita mhima de cn:cimlcnlo pobbclOnal (~)Densidad en equilibrio (indll"rr)Animales remoVidos por depredadores (indlkml)Capacidad de carga
lOd'km~
kg.'lm!Cosecha mál[ill'l3 SUSllllllabk ('-lS")
lndlml
kg.'km!Producción milxuna {P_.>
lnd.'lm!kg.l:m!
Producción pal11 consumo humano (0.40 • P_)",d'km'kg.kml
Venado cola blanca
11.811.0 a 13.6463:1 S73
1.5212.0
4.5 a 5.2
ló.5 a 17.2695 a 724
10.343-1
5.3223
2.188
Pec:arl de collar
7.54.1310.764 a 168
3.008.'
0.7&0.9
9.] a 9.5146.150
5.'89
11.3178
4.571
10••~
"S~•~••.,.5
•"•.,~
•••~•~
0.1
€l
2 3 4 5 6 7 8 9 10 11 12 13 14 IS
O
Densidad poblacional (ind I km!)
Figura 5. Relación entre la tasa finita de crecimiento poblacional (A) y la densidad poblacional del vcnado
cola bl:mca (e) y pccari de collar (O). La linea horizontal con un valor A = I es conocida como linea dc
replazo. Valores mayores a uno indican crccimicnlo positivo de la población: mientras que valores menores
indican decrecimu:nto. El punto donde las lineas de regresión interceptan ala línea de replazo es precisamenteel punto de equilibrio del erecnnlento poblaciona1. es decir O... En este caso la densidad en equilibrio es de12 indlkml para el \cnado cola blanca y 8.6 indlkml para el pecan de collar.
M¡mdujallo 649
Producción potencial máximaLa producción máxima sustcntable (Msr) se estimó en 10.3 venados/km! y 5.6 pecarieslkm!
(Cuadro 1). Mientras que la producción máxima se estimó corno 5.2-5.4 venadoslkm2 y 11.1-11.4pccaríeslkm! (Cuadro 1). Considerando una cosecha de 40% de la productividad máxima, el
potencial para uso humano se calculó en 2 venadoslkm2 y 4 pecades/km!. Dado el mayor pesocorporal del venado. el rendimiento en biomasa puede ser mayor para este ungulado (Cuadro \).
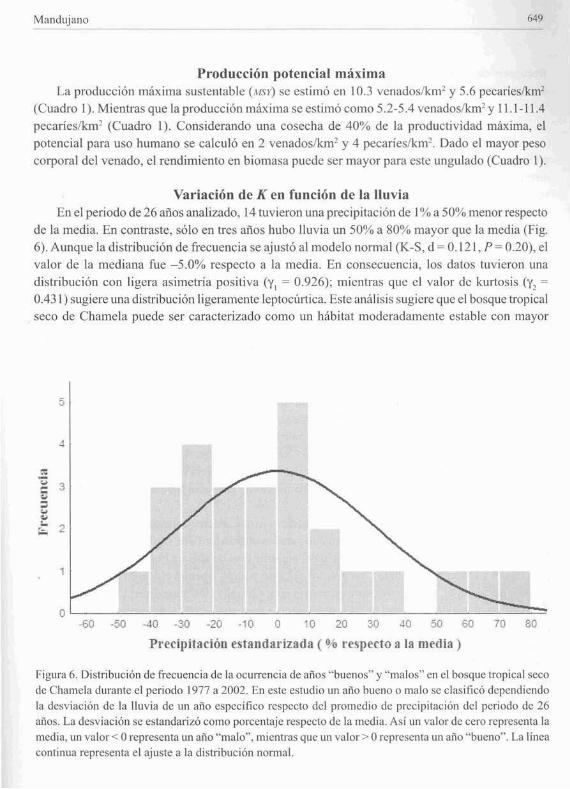
Variación de K en función de la lluviaEn el periodo de 26 años analizado. 14 tuvieron lIna precipitación de 1% a 50% menor respecto
de la media. En contraste, sólo en tres anos hubo lluvia un 50% a 80% mayor que la media (Fig.6). Aunque la distribución de frecuencia se ajustó al modelo nomlal (K+S, d:= 0.121, P = 0.20), elvalor de la mcdiana fue ~5.0% respecto a la media. En consccucncia, los datos tuvicron unadistribución con ligera asimetria positiva (y) := 0.926); mientras quc el valor de kurtosis (y! =
0.431) sugiere una distribución ligeramente ]eptocúrtica. Este análisis sugiere que el bosque tropicalseco de Charnela puede ser caracterizado como un hábitat moderadamcntc cstable con mayor
,J
..." 3ti="•-2'""
1
O·60 -50 -40 ~ ~ -10 O 10 ~ ~ ~ ~ M 70 M
Precipitación eslalldarizada (00 ¡'especto a 1ft media)
Figufil 6. Distribución de frecuencia de la ocurrencia de anos "buenos" y "malos" en el bosquc tropical secodc Chamcla durante el periodo 1977 a 2002. En cste estudio un año bueno o malo se clasificó dependiendola desviación de la lluvia de un año especifico respecto del promedio de precipitación del periodo de 26años. La des\'iación se cstandarizó como porcentaje respecto de la media, Asi un v,llor de cero representa lamedia, un valor < Orepresenta un año "malo", mientfils que un valor> Orepresenta un año "bueno". La lineacontinua representa el ajuste a la distribución nomlal.

650 Ungulados en selva baja
frecuencia de "mIos malos" pero ocasionalmente se presentan "anos muy buenos" en algunos
casos relacionados con F.NSO.
Tendencia CSI¡ICional de la lluvia, plantas y unguladosLa Oucluación estacional de la lluvia, biomasa de plantas y densidad de ungulados durante el
periodo de estudio se presenta en la Fig. 7. Los datos de lluvia y biomasa estacional fuerondirectamente obtenidos en campo. Por otro lado. para elaborar la curva de tendencia de la densidadse procedió a restar de la estimación de la densidad de la cpoca hllmeda el número de animalesremovidos por los depredadores y el resullmlo se asumió como la densidad final en la época seca.Este procedimiento pemlitió crear una curva de tendencia estacional. Los resultados sugieren
mayor ajuste de la tendcncia dc la densidad dcl vcnado cola blanca con la lluvia y con la biomasa;micntras que la tcndencia fuc menos evidcnte para el pecarí de collar (Fig. 7).
DISCUSiÓN
Lluvia y SllS cfcctos sobrc cl alimento y KEl análisis de la precipitación del periodo de 26 años sugiere una asociación significativa entre elENSO y la lluvia estllcional en el bosque tropical de Charnela. Se observó que durante la época secacae una inllsualmayor cantidad de Iluvill cuando se presenta el cvento del El Niño; mientras quedurante La Nil'la elle mayor cantidad de lluvia de 13 nonnal en la época húmeda y nada durante laépoca seca. Lima er al. (1999) encontraron un patrón similar al analizar un periodo de 99 años enuna región semilÍrida de Chile; y Marshal el al. (2002) lo hizo para el desierto Sonorensc duranteun periodo de 47 alios. Por lo tanto, los eventos del ENSO aumentan la variación de la lluvia, lo cual
tiene importantes consecuencias en el funcionamiento de este ecosistema (García-Oliva el al..2002).
La lluvia que cae durante la época húmeda controla el crecimiento de las plantas y enconsecuencia la producción anual del alimento para los ungulados (Cocer al.. 1976; Owen-Smith.1990; McKinney el al.. 2001). La evidencia del presente estudio sugiere que. similar a otros sitios(e.g. Ogutu y Owen-Smith, 2003). los eventos del ENSO podrán tener un efecto adicional sobre lasplantas; por ejemplo, esto flle evidente durante la época seca de 1992 cuando en 15 días, en el mes
de enero. cayó una lluvia record de 649 111111 (Garcia-Oliva el al.. 2002). Sin embargo. este cfectopodria dcpender del mes particular cuando la lluvia cae. Por cjemplo, durantc los años del f.NSO
1983 y 1997 cayeron 177 y < 100 mm en mayo, respectivamente. Lo cual pudo no haber tcnidoconsecucllcia cvidcnte Cll la producción de plantas debido a lo avanzado de la época scca. Por lotanto. considerando la notable diferencia en la composición norística y fomlas de vida entrc losbosqucs tropicales seco y subcaduci folía en ellÍrea de estudio (Lotl el al.. 1987), se podría esperar
una relación compleja entre ENSO. la precipitación y la respucsta de las plantas como se ha mostradoen otros estudios (e.g.. \Vright el al., 1999: Connell y Green, 2000; Wil1iamsoll y Ickes, 2002;Rolim er al.. 2005).
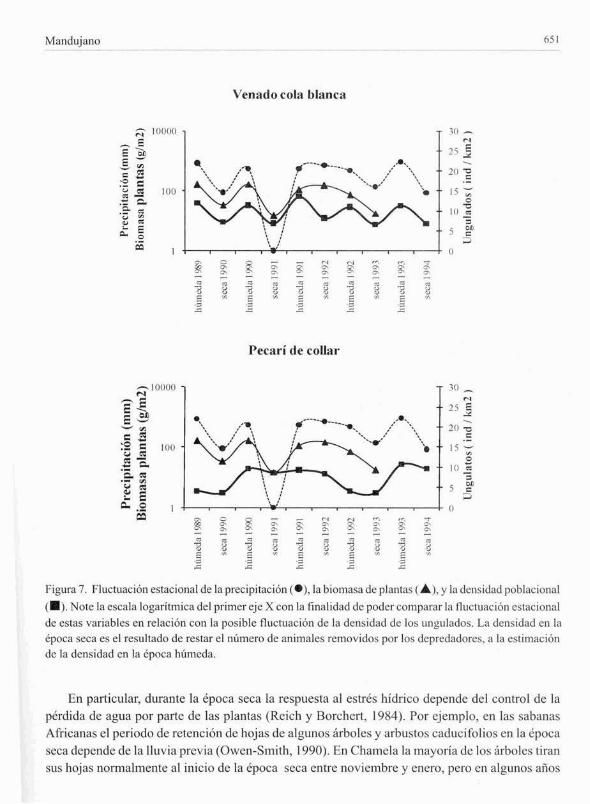
Malldujano 651
VenlHlo ('ola blanCll
"'25 ]
lO "ª
,
~o _
15 -:o
10]5 ~
5
"'25 ::~
20:;;~
•!.!
•,,,
,.....-.----... / ...../ ........,,/ \"..
•!
\ .•./\,1---~-~-'-:'h-_- __-_-_--+"
FigurJ. 7. Fluctuación estacional de la precipitación (e). 1<1 biomasa de planl¡IS (.....). y hl densid¡¡d pobhleiollal
(.). Note la escala logaritmica del primer eje X con la linalidad de poder comp¡¡rar la t1uctuaeión estacionalde eSlas variables en relación con la posible fluctuación de la densid:.d de los ungulados. La densidad en laépoca seca es el resultado de restar el número de animales removidos por los depredadores. a la estimaciónde la densidad en la epoca humcda.
En particular, durantc la época seca la respuesta al estrés hidrico dependc del control de la
pérdida de agua por parle de las plantas (Reich y Borchert, 1984). Por ejcmplo. cn las sabanas
AfriClmas el periodo de rctención de hojas de algunos árboles y arbustos caducifolios en la época
seca depende de la lluvia previa (Owen-Smith. 1990). En Chamela la mayorí:. de los {lrboles tiran
sus hojas nomlalmCllte al inicio de la época seca entre noviembre y enero, pero en algunos 1l11os

652 Unguhldos en selva baj:l
las hojas pueden pcnnaneccr más tiempo en respuesta a la presencia de lluvias tardías o invemarlcs(Bullock y Solis-Magallallcs. 1990). Enlonccs. en algunos ai'tos del ENSO hay un aumento de laslluvias durante la época seca lo que ¡Wlllcnta la probabilidad de que las plantas mtlntcngan sus
hojas verdes y esto mejora la disponibilidad de alimento para los ungulados. Adicionalmente. 1:IS
condiciones creadas por el ENSQ mejoran la producción de frulos en algunas especies tropicales(c.g.. \Vrighl el (//.. 1999; Williamson e Ickcs. 2002). Considerando que el venado cola blanca
aumenta su consumo de fmlos (Arcca ef ,,1.. 2005), entonces es posible que el ENSQ tenga unefecto adicionalll1cjorando la capacidad de carga del hábitat durante el periodo de sequía.
Producción de unguladosLos datos presemados aquí sugieren que en el bosque tropical scco dc Chamela la densidad
(15.1 a 24.3 individuoslkm') y biomasa (527 11 741 kglkm!) dc los ungulados silvestres esrelativamente 'lIla si se compara con datos de otros bosques neotropicales. Por ejemplo. en unbosque tropical scco con pareilcs dc hábit:n semi-abierto en Venezuela y Brasil. la biomasa deungullldos silvestres cs de 224 a 275 kg/km' (Robinson y Bellnell, 1999). En bosques tropicales
lluviosos donde pueden habitar hasta cinco especies dc ungulados (Tapirus lerresrris, Tayas.mpecar;, Pee:tlri /o)ac/I, A"o=(III/(/ americalla y M. goua=ollbira) llegan a tener una densidad y biomasade 28.3 individllos!km2 y 761 kglkm2, respectivamelllc(Robinson y Redrord, (991); aunque datosde campo de Pan:11l1á, Perú, Brasil y Venezuela sugieren que la biomasa varía de 270 a 542 kglkm2
(Robinson y Bennetl. 2004). Porotm parte, en la Selva Laeandona, Mcxico, la densidad y biomasa
de cinco especies dc ungulados en un area con baja presión de caceria se han estimado en 9.9indi\'iduos/km2 y 337 kgfkm! (Naranjocr a/., 2004b). Estas comparaciones sugieren que el bosquetropical scco de Chamela, con menor riqueza de especies de ungulados, puede m:mtcner densidadessimilares o mayores cn comparación con otros I1cotropicalcs (e.g., Robinsoll y Redrord, 1991;Bodmcr y Robinson, 2004; Naranjo el al.. 2004b). Esto podria deberse, por un lado, al tipo dehistoria de vida de las especies prescntcs en Charnela y, por otro, a la mayor capacidad de carga
dcl bosque tropical seco. Por ejemplo, se ha documentado que especies con poblaciones conabundancia alta y rápida reproducción, como el peeari de collar y venado cola, podrían ser menos
vulnerables a la sobrecaeeria (McCullough, 1987; Bodmer y Robinson, 2004).Elmodc1o de Robinsol1 y Redrord (1991) asume que K =: O en poblaciones con poco o
ninguna presión de caza, lo eual110 es selll;illo de encontrar. Los datos que se provecn en estetrabajo, obtenidos en un área relativamcnte protcgida. mostraron que la densidad de ambosungulados varia dc un afi.o a otro tanto por posibles sesgos de muestreo como por cambios biológicosreales de las poblacioncs. En consecuencia, una única estimación de O podria resultar sesgada de
K. Para evitar esto, Bodmer y Robinson (2004) sugiere monilOrcar la población, el hábitat y lacacería con el fin de incorporar esta variación cnlos modelos para evaluar la sustcntabilidad de lacacería. En este sentido, la estimación de la densidad en equilibrio (D"l) obtenida en este estudioconsidera la variación interanual del cambio poblaciona1. Obviamente, estc procedimiento requieremonitorear la población por vllTios a¡ios, lo cual no es sencillo. Sin embargo, esto es importanteporque la dinilluica de las plantas y los ungulados es muy compleja (McCullough, 1987; Marshal

Mandujano 653
el al., 2002; Ogutu y Owen-Smith, 2003). Es importante clarificar que la dcnsidad en equilibriono es simplemente la densid¡ld promedio anual durante el periodo de estudio. Por el contTllrio. laD se calcula según la relación denso-dependiente entre el crecimiento anual y la actual densidad,,según un modelo logistico. En consecuencia, la densidad en equilibrio representa la tendencia dela población a lo largo del tiempo.
Importancia de la depredación en la estimación de K.y MSI'
Según la aproximación adoptada en este trabajo, en ausencia de perturbaciones importantesla densidad actual fluetuflfá alrededor del vlllor 1) . En lllgunos casos se asume erróneamente que,1)<q = K. lo cual es cierto solo en el caso raro que la depredación no tenga un papel importante encontrolar el crecimiento poblaeional. Sin embargo, la D"I no necesariamente ocurre al mismonivel de K en aquellas áreas donde el crecimiento poblacional est:i fuel1emente regulada por ladepredación (l3odrner y Robinson, 2004). Para el bosque tropical seco de Chamela, Nuiiezef al.(2000) proponen que, en ausencia de depredadores, las poblaciones de ungul<ldos podrian alcanzary sobrepasar la capacidad de carga del hábitat. Esos datos parecen corroborar 10 encontrado enotros estudios que indican que los depredadores mantienen 11 los ungulados por debajo de K(Skogland, 1991). Sin embargo, para distinguir si este equilibrio es resultado de los depredadores
o por limitación de recursos o por llna combinación de estos dos factores, se necesita conocer silos depredadores, o los recursos o ambos, están afectando las tasas de nacimientos y de mortandad(Caughley y Sinclair, 1994). Desafortunadamente, este nivcl de detalle no esl~ disponible panl elsitio de estudio. Por lo tanto, aun y cuando los depredadores mantienen a las poblaciones deungulados por de bajo de K, es compleja la relación entre la disponibilidad del alimcnto. la densidad
poblacional y los factores de mortalidad. por 10 que se requiere de muchos anos para monitoreary comprcnder mejor los mecanismos subyacellles en esta relación (McCullough, 1992; S¡ether,1997).
Un resultado importante de presente estudio es que el nlJmero de animales removidos por losdepredadores se deben adicionar a la densidad en equil ibrio con el fin de cstimar adecuadamentela K y el .l/sr. Este aspecto es mas importante para aquellas especies con alto porcentaje de
depredación, como el caso del venado cola blanca en la zona de estudio. Una subestimación delMsr podría poner en riesgo a 1:1 población presa (McCullough. 1987). Para clarificar este puntohay que considerar los siguientes argumentos (ver Bodmer y Robinson, 2004). La cosechasustentable puede obtenerse <l cualquicr tamaiio poblacional pero sólo hay un punto al cual lacosecha es máxima, el conocido .l/Sl'. Cosechar a una población a su valor teórico deluwo cuandoc1t<lmaíio poblacional es menor que estc valor (a la izquierda del Msr) puede ser una estrategia demanejo riesgosa y debe evitarse. Esto se debe a que si se comenten pcquerlos errores en la estimación
del MSl'podría llevar a Ullll sobrecacería 10 cual podría rcsultaren una disminución de la poblaciónen los siguientes años 10 cual podría rápidamente llcvarla una sobre-explotación y eventualextirpación. En contraste, cosechar poblacioncs con alta abundancia (a la derecha del valor teóricoMsr) es una estrategia de manejo segura que pucdc ser usada para el aprovechamicnto a largoplazo (McCullough, 1987).

'" Ungulodos en selva baja
En consecuencia, cosechar una especie con aha abundanda de sus poblaciones es una estrategiade manejo segura que puede lener menos probabilidad de que se extirpe localmente. Esto esespecialmente importante para especies limitadas por la depredación debido a que sus densidadesestán por debajo de K. En conclusión, una subcstim<Lción de K puede llevar a una subestimacióndel MSl'Y ti llna interpretación errónea de la relación enlre la abundancia y el valor del AlSl". Para el
área de estudio los rcsull<ldos sugieren que una estrategia de manejo segura es cazar a los venados
y recaríes sólo cuando la densidad poblacional es mayor a 10.3 y 5.6 individuos/km l .
respectivamente. Esta estrategia tiene la vClItajll adicional de protcger a los depredadores como eljaguar. puma y ocelotc. allllanejar sllslentablementc a los ungulados prcsas.
Consideraciones para el manejoEl analisis a largo plazo del plltrón dc prccipitación cn un arca espccífica, pucde ser una
aproximación muy útil y sencilla pam desarrollar guias de manejo para los ungulados en unaregión detenninada. El ana lisis de la precipitación en la región de Chamela sugiere que el bosquetropical seco puede considemrse como un habitat moderadamente cstable camcterizado por aiiosmalos con otros aijos con lluvias cspor:idicamcntc intensas. Tanto la varianza ambiental como ladistribución de frecucncia dc 3110s "buenos" y "malos" puedcn influir la precisión de las estrategiasde manejo que deben aplicarse (McCullough, 1987). Por cjcmplo, en un ambiente con una variaciónaleatoria o periódica de la capacidad de carga. [as poblaciones con alta tasa de crecimiento natural
(como pequeños mamíferos) ticnden a seguir la variación del ambiente; mientras que poblacionesCOIl una tasa de crecimicnto baja ticnden a promediar el ambientc (May, 1976). Debido al t:ullaiiogrande y las caracteristicas particulares de la historia de vida, las poblaciones de los ungulados
típicamente estan muy cercanas a la capacidad de carga del hábitat (McCullough. 1992). Porejemplo. en un periodo de cinco allos de estudio la fluctuación de la densidad poblacional fuemenor pura el venado que tiene m:lyor tasa de crecimiento que para el pecad con mayor tasa Am>Jl'
MeCullough (1987) sugierc que en ambicntes con alta variabilidad el manejo debe ser {jd IIOC
para responder a variables con poco control humano. En contrastc, cn habitats con muchos aitosbuenos el manejo puede ser muy preciso y sólo debe atenderse los años malos. En sitios comoChamcla el manejo puede scr rclativamente preciso debido a que es mareadamcnte cstacional,
moderadamente estable y con una frecuencia de atlos con precipitación menor al promedio yalgunos a110s con abundancia de lluvias influenciados por eventos del ENSO y otros. En este tipo de
hábitat se debe tener precaución cuando se preselllan varios 3110s consecutivos secos pam evitar lasobreexplotación de los ungulados.
CONCLUSiÓN
El análisis dc 1:1 relación entre 1:1 lluvia. la capacidad de carga y la producción potencial de unguladospara aprovechamiento humano llluestra. por un lado, 10 complejo de esta rclación y la necesidadde monilOrcar estas vllriables durante el mayor tiempo posible; por otro, la utilidad del análisis

M:llldujao'o'-- _
para proveer bases sólidas para el manejo al incorporar la variación anual de la lluvia. En este
sentido, en aquellos sitios donde las poblaciones de venado cola bl:ll1ca y pecad dc collar cst:'ln
sometidas a manejo con tines de aprovechamiento. se recomienda ampliamente analizar el patrón
de precipitación por el mayor numero de años posibles pues esto pernlile Icner una idea muy
precisa delmallejo genewl de las poblaciones y de la atención en al105 o épocas particulares. En
este sentido. las :'Ireas naturales protegidas pueden servir como labomtorios naturales para
monitorear y estudiar esta relación compleja entre las variablcs elim{l\icas, el hábitat y las
poblacioncs de fauna silvestre. El valor de esas áreas no sólo sigue siendo importante desde una
perspectiva de conservación sino, además, como sitios para la generación de información búsica
para aplicar a poblaciones en condiciones ecológicas similares sujetas a aprovechamiento. Bajo
esta perspectiva, si bien la información de ungulados presentllda en este eapílulo fue obtenida
hacc más de 15 años, su vigencia es actual para proveer de un marco conceplual para el
aprovechamiento de estas especies de ungulados. Por lo tanto, el bosque tropical seco de la Reserva
de la Biosfera Chamela-Cuixmala es un sitio con un altísimo valor tanto para la conservación
como para el aprovechamiento sustentable de la vida silvestre.
LITERATURA CITADA
Akyakaya, H.R., M.A. Burgman y L.R. Ginzhurg. 1999. Applied Popllla/ion Ecology: Principies
al/d COII/pl//er Exercises IIsil/g RAMAS® EcoLab 2.0. Second edition. Sinauer Assoeiatcs, lne.
Sunderland, Massachusetts.
Alvard, M.S., J.G Robinson, K. H. Redford y H. Kaplan, H. 1996. Thc sustainability 01" subsis
tenee hunting in the Neotropies. COllseJ1latiOIl Biology, II :977-982.
Arceo. G, S. Mandujano, S. Gallina y L. A. Perez-Jirncncz. 2005. Diet divcrsity of white-Iailed
deer (Odocoile/ls lIirgilliolllls) in a tropical dry forest in Mexico. MallJlllolia, 69: 159-168.Behrenfeld, M. J., J. T. RUlldersoll, C. R. McClain, G C. Feldrnan, S. Q. Los, C. J. Tuckcr, P. G.
Falkowski, C. B. Field, R. Frouin, W. E. Esaias, D. D. Kolbcr y N. 1-1. Pollack. 200\. Bios
pheric primary produclion during an ENSO transition. Science, 291 :2594-2597.
Bodmer, R.E. y J.G Robinson. 2004. Evaluating the sllstainability of hunting in Ihe NCOlropics.
Pp. 299-323, il/ People il/ Na/l/re: Wildlife COI/serva/ion in SOllt¡' amI Cel/I/'al America (Silvus,
K.M., R.E. Bodmer y J.M.Y. Fragoso, eds.). Columbia University Press, New York.
BlIllock, S. H. 1986. Climate ofCharnela, Jalisco, and trcnds in sOllth eoastal region of Mexico.
Al'ch. Meteoro. Geoph Bioclimatol., 36:297-316.
Bullock, S. H. YJ. A. Solis-Magallanes. 1990. Phenology ofcanopy trees ofa tropical dcciduous
forest in Mcxico. Bio/ropica, 22:22-35.
Caughley. G y A.R.E. Sincbir. 1994. Wildlife Ecology and Management. Blackwell Science,
Cambridge, Massachusetts.
Coe, M. J.. D. H. Cumming y J. Phillipson. 1976. Biomass and produetion oflarge Al"rican herbi
vores in relatíon to rainfall and primary production. Oecologia, 22:341-354.

'56 Ungulados en selva baja
Conncll. J. H. YP. T. Grccn. 2000. $ccdling dynamics aver thirty-IWO ycars in a tropical rdil1 Cores!Irce. Ecolog)', 81 :568-584.
DcCa1csta. D. S. y S. L. SlOut. 1997. Rclative deer density alld sllstainability: a conceptual framcwork ror inlcgraling dccr l111magcmcnt with ccosyslcm managcmcnt. Wildlife Society Bulle/ilJ,
25:252-258.Eisenbcrg. J. F. 1989. Ma1111lluls orIlle Ncotropics. The Northcm Ncotropics. Vol. l. The Univer
sity ofChicago Prcss. Chicago.Escamilla. A.. M. $al1viccntc. M. Sosa y C. Galindo-LcaJ. 2000. Habita! mosaie, wildlifc availa
bility, and hunling in lhe tropical rores! ofCalakmul. Mcxico. COlIselwllioll Biology. 14: 15921601.
Fitzgibbol1, C. D., H. Mogaka y J. H. Fanshawc. 1995. Subsistence hUI11ing in Arabuko~Sokoke
foresl, Kenya, and ils effects on mammal poplllalions. COI/serva/ion Biology, 9: 1116-1126.Gallina, S. 1993. Biolllasa disponible y capacidad de carga en la reserva la Michilia, Durango, Pp.
437~453, in Awmces en el Es/udio de los Mamíferos de México (Medel1ín, R. A. Y G A.Ceballos, eds.). Publicaciones Especiales, VoL l. Asociación Mexicana de Mastozoología A.
C. México.Garcia~Oliva, F.. E. Ezeurra y L Galieia, 1991. Pattem ofrainfall distribution in the central Pa~
cific coast of Mcxieo. Geografiska AmI., 73: 179~ 186.Garda-Oliva, F., A. Camou y J. M. Maass. 2002. El clima de la región central de la costa del
Pacífico mexicano. Pp. 3-10, il/ HiSToria Nalural de C/wmefa (Noguera, F., J. H. Vega, A. N.García-Aldrete y M. Quesada-Avcndaño, eds.). Instituto dc Biologia,uNAM. México.
Georgiadis, N., M. Haek y K. Turpin. 2003. The influenee ofrainfall on zebra population dyna
mics: implieations for rnanagemellt. Joumal ofApplied Eeology, 40: [25-136.Groot-Bruinderink. QW.T.A. y E. Hazebrock. [995. Modelling eanying capacity for wild boar
Sus serojá serofa in a forcstlheatland eeosystem. Wildlife Biology, 1:81-88.Hall, E.R. 1981. rhe Mammals ofNor/h America. 2nd ed. John Wilcy and Sons, Ncw York.Hidalgo-Mihart, M.. L. Cantú-Salazar, C. A. Lópcz-González, E. Martinez.Mcyery A. GOl1z{llcz~
Romero. 2001. Coyote (eauis la/rol/s) food habits in a tTopical dcciduous forcst ofweslcmMcxieo. Americal/ Midlolld NOfllralis/, 146:210-216.
Hunter, C. M. y M. C. Runge. 2004. The importanee ofenvironmental variability and management control crror to oplimal harvesl poliees. JOllmal of lVildlife Mallagemem, 68:585-594.
Jaksic, F. M. 2001. Ecological cffcctsofEI Nino in tcrrestrial ecosystemsofwestem $outhAmcricaEcogrophy, 24:241-250.
Langvatn, R.• S. D. Albon, T. Bllrkcy y T. H. Cltltton~Brocks. 1996. Climate, plant phcnology andvarialion in age of firsl reproduclion in a temperate herbivorc. JO/ll"1lal 01Applied Ecology,65:653-670.
Lima, M., P. A. Marquet y F. M. Jaksic. 1999. El Niño evenls, precipilation pattcms, and rodentolllbreaks are slatisticlllly associatcd in scmiarid Chile. Ecography, 22:213~218.
Lot!. E. 1. S. H. BlIllock y J. A. Solis~Magllllanes. 1987. Floristic diversity lInd struclure ofuplandand arroyo forcsls in coastal Jalisco. BiollVpica, 19:228~235.

Mandujano 657
Mandujano, S. 1999. Varialion in herd size ofcollarcd pcccaries in a lllexicantropical lores!. TheSOllfllll"eslem Nalllra/isl. 44: 199-204.
Mandujano, S. 2005. Calibration of tracks count lo eslirnate population density of white-lailed
deer (Odocoilells v;rginialllls) in a Mcxiean tropienl rores\. rhe SOlllhll'esfern N(Illlmli.\·I.
50:223-229.
MandlUano. S. 2006. Prelilllinary evidenee ofthe importanee of rainfall all(II,NSO in lTlodify food
availibility for white-tailed deer in a Mexicalllropical dry fores\. Biol/"Opica. 38:695-699.
Mandluano, S. 2007. Carrying capacity and pOlentíal production ofungulalcs for human use in a
Mexican tropical dry fores!. Biollvpica, 39:519-524.
Mandujano. S. y S. Gallina. 1993. Densidad del venado cola blancn basada en conteos en lranseclos
en un bosque tropical de Jalisco. Acla Zoológica !J.'lexic(Il1ll (n. s.). 56: 1-37.
Mandujano. S. y S. Gallina. 1995a. Comparison of decr censusing rnethods in 11 tropical dry
foresl. Wi/dlife Sociel)' BIII/etill, 23: 180-186.Mandujano, S. y S. Gallina. 1995b. Disponibilidad de agua para el venado cola blanca cn un
bosque tropical caducifolio de México. Vida Si/I'eslre Neoll'Opico/. 4: 107-1 J8.Mnndujano, S. y S. Gallina. 2005. Dinámica poblacional del venado cola blanca (Odocoi/e/ls
virgilliolllls) en un bosque tropical eaducifolío de Jalisco. Pp. 335-348. in COlIll"ibllcione~'
""as/o=oológicllS en Homenaje al Doctor Bemardo Vil/a (Sánchcz-Cordero. V. y R. Medellin,
eds.). Institutos dc Biología y Ecología-uNAM /CONAlJIO. México. DI'.
Mandujano, S. y L. E. Martínez-Romero. 1997. Fruit falt caused by chachalacas (Ona/is
/Joliocepha/a) on red mombim trecs (Spondias pllrplll"ea): impact on terreslríal fruit consu
mers, especially the whilc-tailed dcer (Odocoilells l'irgilliclIIlfs). Sllldie~' Neollvpical Faulla
alld ElIl'iroJIIllent, 32: 1-3.
Mandujano, S. y L. E. Martíncz-Romero. 2002. Pccarí de collar (Pecarl /ajaclI SOJ/orellsi.\'. Meams
1897). Pp. 411-41 S. in Historia NCIIIfI'(lI de CJIC/llle!c. (Noguera, F. A.. J. 1-1. Vega. A. N. Gareía
Aldrete y M. Quesada, eds.). Institulo de Biología UNAM, México. Df.
Malldujano, S. y V. Rico-Gray. 1991. Hunting, use and knowledge ofthe biology ofthe whilC
lailed deer. Odocoilells l'irgilliGlllfs (Hays), by Ihe maya of cenlral Yucatan. Mexico. ./o/il'l/al
ofETlmobi%gy. 11: 175-183.
Mandujano, S.. S. Gallína, G Arceo. G Sánehez~Rojas y M. G Silva-Villa lobos. 2002. Ddocoi/clIs
virgil/lclll/ls silla/oae (J. A. A1Ien 1903): Venado cola blanca. Pp. 415-422. ill His/oria Narlll'(ll
de Challlela (Noguera, F.. J. H. Vega. A. N. García-Aldrete y M. Quesada.Avendailo. cds.).
InstitUlo de Biologia-uNAM, México. 01'.
Mandujano, S., S. Gallina, G. Arceo y L. A. Pérez-Jiménez. 2004. Varíación estacional del uso y
prefcrencia de los tipos vegctacionales por el venado cola blanca en un bosque tropícal de
Jalisco. Ac/(/ Zoológica Mexicana (n. s.), 20:45-67.
Marshal, J. P., P. R. K.rausman, V. C. Bleích. \V. B. Ballard y J. S. McKecver. 2002. RainfalL El
Niño. and dynamics ofmule deer in the SOlloran desen. California . ./Olll"llal ofWildlife Ma
lIagemelll, 66: 1283-1289.Manlnez-Romero. L E. YS. Mandujano. 1995. Hábitos alimentarios del pecari de collar en un

658 Ungulados en selva baja
bosque tropical caducifolio de Jalisco. Acla Zoológica Mexicana (n. s.), 64: 1-20.
May. R. M. 1976. Modcls for single populations. Pp. 4-25 in Theorerical Eco/agy.o PrincipIes alldApplicaliofls (May, R.M., eJ.). w,s. Saundcrs, Philadelphia.
McCall, T.e.. R. D. Brown y L.e. Bender. 1997. Comparison oftcchniques for dClcnllining (he
nutrilional carrying capacity for white-taitcd deer. Jouma/ ofRonge Managemellf. 50:33-38.
McCullough, D. R. 1987. The theory and managemcnt ofOdocoi/eus populations. Pp. 535-549,
ill Bio/ag)' ond mallagemelll aflhe Cerl'idae: proceedings ola symposiwlI (Wemmer, C. M.,
eJ.). Smithsoniall Insl. Prcss, Washington, De.
McCullough, D. R. 1992. Conccptsoflargc hcrbivorepopulation dynamics. Pp. 967-984,il/ Wildlife
2001: populalions (McCullough, D. R. Y R. H. Barret, eds.). EIsevier Applicd Scicncc, Lon
don, UK.
McKinncy. T.. T. W. Smith y 1. D. Hanna. 200 l. Prccipitation and descrt bighom sheep in the
Mazatzal mountains, Arizona. Tite SOl/lhll'estem Nall/ralist, 46;345-353.
Millcr. K.Y. y J. M. Wcntworth. 2000. Carrying capacity. Pp. 140-155, in Ecology and Manage
11/efl( of Large Mallllllals in Norlh America (Dcmarais, S. y P. R. Krausman, cds.). Prcnticc
Hall, Uppcr Saddlc River, New Jerscy.
Murphy, P. G yA. E. Lugo. 1986. The ccology of tropical dry forest. Amlllal RevielV of Ecol.S)':;/ematin. 17:67-88.
Naranjo, E. 1., M. M. Guerra, R. E. Bodmcr y 1. E. Bolaños. 2004a. Subsistcl1cc hllnting by Ihree
elhnic groups ofthc Lacandon forcst, México. JOl/mal ofEr/mobiology, 24:233-253.
Naranjo, E. J., J. E. Bolaños, M. M. Guerra y R. E. Bodmer. 2004b. Hunting sustainability ofungulatcs populations in the Lacandon forcst, Mexico. Pp. 324-343, iI¡ People ill Nature:
Wildlife COlIserra/ioll in SOllt/¡ alUl Celllral America (Silvus, K.M., R.E. Bodmcr y J.MV
Fragoso, cds.). Columbia Univcrsily Prcss, Ncw York.
NlIi'í.cz. R.. B. Millcr y F. Lindzcy. 2000. Food habits of jaguars and pumas in Jalisco, Mexico.
Joumal ofZoology (Lolldon). 254:373-379.
Ogutu.1. O. y N. Owcn-Smith. 2003. ENSO, rainfall and temperature inOuences on extremc popu
lation dcclincs among African savanna ungulatcs. Ecological Leller~', 6:412-419.
Owcn-Smith, N. 1990. Demography ofa large herbivore, Ihe grcalcr kudu Tragelaphus slrepsiceros,
in rclation \O rainfall. Joumal ofAnimal Ecology, 59:893-913.
POSI. E. y N. C. Stenscth. 1999. Climatic variabiliry, plant phcnology, and northem ungulates.
Ecology, 80: 1322-1339.
Rcieh. P. B. Y R. Borehcrt. 1984. Water slrcss and trce phenology in a tropical dry forest in the
lowlands ofCosta Rica. JOl/mal ofEcology, 72:61-74.
Robinson, J. G Y E.L. Bennctt. 1999. Carrying capacity limits to sustainable hunting in tropical
foresto Pp. 13-30, in HUlllingfor sustainabilit)' in tropicalforest (Robinson, J.G y E. L. Bcnllclt,
cds.). Columbia UniversilY Prcss, Ncw York.
Robinson, 1. G YE.L. Bcnllctt. 2004. Having you wildlife and cating it loo: an analysis ofhunting
suslainability across tropical ccosystems. Animal Consen1a/io/l, 7:397-408.
Robinson, J. G y K. H. Redford. 1986. Intrinsic rate of natural increase in Neotropical forest

Mandujano 659
mammals: rclationship to phytogcny alld dicL Oecologia, 68:516-520.Robinson. J. G. Y K.H. Rcdford. 1991. Sustainablc harvest ofncotropical forcst mammals. Pp.
415-429, in Neorropicalll'i1dlife use alld cOI/serva/ion (Robinson, J.G. YK.H. Rcdford, eds.).
University ofChicago Press, JIIinois.Rolirn. S. G, R. M. JCStlS, H. E. M. Nascimento, H. T. Z. do CoulO y J. Q. Chambcrs. 2005.
Biomass changc in an Allanlic tropical moist forcst: thc ENSO cITcct in pcrrnancnt samplcplots over a 22-ycar periodo Oecologia, 142:238-246.
Slcther, B-E. 1997. Environmcntal stochasticity and popu1ation dynamics oflargc herbivorcs: asearch formcchanisms. TREE, 12:143-149.
Scoones, l. 1993. Economic and ccological carrying capacity: apptications to pastoral systcrns inZimbabwc. Pp. 96-117, in Ecollomics alld ecology: lIelV jiV/lfiers (111(1 Sllsfaillahle developmellf (Barbicr, E. B., ed.). Chapman & Hall LId, Londoll.
S¡lva-Villalobos, G, S. Mandujano, G. Arceo, S. Gallina y L. A. Pérez-Jiménez. 1999. Nutritiollalvaluc ofplants consumcd by Ihe white-tailed dccr in a tropical forest ofMexico. Vida Si/ves/reNeotropical.8:1-6.
Sinclair, A. R. E., H. Dublin y M. Bomer. 1985. Population rcgulation ofScrcllgcti Wildebccsl: ates! ofthc food hypothesis. Oecologia, 65:266-268.
Skogland, T. 1991. What arc lhe efTccts of predators 011 large ungulate populalions? Oikos, 61 :40 1
411.Slade, N.A., R. Gomulkicwicz y H.M. Alexandcr. 1998. Altemativcs to RobinSOIl and Redford's
melhod of asscssing over harvest from incomplete demographic data. ConserWl/iOJl Biolog)'.
12:148-155.SlatSOft, Inc. 2000. STAT1ST1C for Windows: Compuler Manual. Slatsofl, lne., Tlllsa, OK.
Stcllselh, N.C., G Oltersen, J. W. Hurrell, A. Myslcrud, M. Lima, K. Chan, N. G Yoecoz y B.Adlandsvik. 2003. Studying climate cffccts on ccology through the use ofclimate indices: lheNorlh Atlantic Oscillalion, El Nino Soulhern Oscillalion aud bcyond. P. Roy. Soco Lond. B
Bio., 270:2087-2096.TIlomas, L., J. L. Laake, S. Strindbcrg, F.F.C. Marques, S.T. Buckland. D.L. Borchcrs, D.R.Ander
son, K.P. Bumham. S.L. Hcdlcy, J.H. Pollard y J.R.B. Bishop. 2003. Distance 4.1 Release 2.Research Unit for Wildlife Population Asscssmcnl, University of SI. Andrews, UK. http://www.rmvpa.st-and.ac.uk/distance/.
Ticktin, T. 2003. Rclationships bctwecll El Niño Southcm Oscillation and demographic patlemsin a sllbslitule foo<! for collared pcccarics in Panama. Biotl"opica, 35: 189-197.
Villa-Meza, A., E. Martincz-Meyer y C. A. LÓpez-González. 2002. Occlot (Leopardlls pardalis)food habits in a tropical dcciduous forest of Jalisco, Mcxieo. American Mildland NatllraliSf,
148:146-154.\Vesl, Re. y J.A. Parkhurst. 2002. Interaclions bctwcen decr damagc, dcer density, and stakc
holdcr attin.dcs in Virginia. Wildlife Socie')' Bullerill, 30: 139-147.\Vilkie. D.S., B. Curran, R. Tshombe y GA. Morelli. 1998. Modclling the suslainability ofsubsis
tence fanning and hunting in Ihe lture foresl ofZairc. Comervafioll Biology. 12: 137-147.

660 Ungulados en selva baja
Williamson, GB. y K. Jckcs. 2002. Mas! fmiting and ENSQ cycles - does (he cue betray a cause?Oikos. 97:459-461.
Wright. S. 1., C. Carrasco, O. Calderon y S. ratono 1999. The El Niño Southem Oscillalion,variable fnJÍI production, amI famine in a tropical rores\. Ec%gy, 80: 1632-1647.


















