
Presentación de PowerPoint - artropodosysalud.com€¦ · efecto en la salud, humana, animal y...
46
Enero-Junio 2016. Vol-. 1 No. 1 Editorial Entomología Forense Por: Dr. Humberto Quiroz Contenido : 1. - Editorial: Zika Virus 2. - Monografía: Anita Hoffmann 3. - WHO reveals its shopping list for weapons against Zika 4. - Mosquitos Silvestres ( Diptera : Culicidae ) de la Ría de Celestún Yucatán 5. - Brief Description Of: Zika Virus 6. - Mecanismos No Químicos De Comunicación En Insectos 7. - Virus De La Tristeza De Los Cítricos 8. - Tifus Murino : Generalidades
Transcript of Presentación de PowerPoint - artropodosysalud.com€¦ · efecto en la salud, humana, animal y...

Enero-Junio 2016. Vol-. 1 No. 1
Editorial
Entomología Forense
Por: Dr. Humberto
Quiroz
Contenido:
1.- Editorial: Zika Virus
2.- Monografía: Anita Hoffmann
3.- WHO reveals its shopping list for weapons
against Zika
4.- Mosquitos Silvestres (Diptera: Culicidae)
de la Ría de Celestún Yucatán
5.- Brief Description Of: Zika Virus
6.- Mecanismos No Químicos De Comunicación
En Insectos
7.- Virus De La Tristeza De Los Cítricos
8.- Tifus Murino: Generalidades

CONTENIDOEnero-Junio 2016. Vol-. 1 No. 1
1.- Editorial: Zika Virus
Por. Publicación Lancet
2.- Monografía: Anita Hoffmann.
Ángel Muñoz y Cristian Escobedo
3.- WHO reveals its shopping list for
weapons against Zika.John Maurice
4.- Mosquitos silvestres (Diptera: Culicidae)
de la Ría de Celestún Yucatán..Pablo Manrrique Saide
5.- Brief Description of: Zika Virus.
Omar Eduardo Tova-Herrera
6.-Mecanismos no Químicos de
Comunicación en Insectos
Biol. Mara Ivonne Garza-Rodríguez
7.- Virus De La Tristeza De Los Cítricos
Pedraza Muñoz Raymundo
8.- Tifus Murino: GeneralidadesCaballero Treviño Yesica
Fotografías
Portada: Fumigación aérea, por Gerardo Trujillo
Puerto Vallarta, Jal.
Contenido: Gustavo Ponce García
DIRECTORIO
Ing. Rogelio G. Garza Rivera
Rector
Dra. Carmen del Rosario de la Fuente García
Secretario General
Dr. Juan Manuel Alcocer González
Secretario Académico
Dr. Celso José Garza Acuña
Secretario de Extensión y Cultura
Dr. Celso José Garza Acuña
Director de Publicaciones
Dr. Antonio Guzmán Velasco
Director de la Facultad de
Ciencias Biológicas
Dr. José Ignacio González
Sub-Director de la Facultad de
Ciencias Biológica
Dr. Gustavo Ponce García
Editor Responsable
Dr. Pedro Cesar Cantú Martínez
Redacción
Ing. Oscar Manuel Loaiza Jiménez
Dr. Saúl Lozano Fuentes
Diseño
Artrópodos y Salud, Año 3, Nº 1. Es una publicación
semestral, editada por la Universidad Autónoma de Nuevo
León, a través de la Facultad de Ciencias Biológicas.
Domicilio de la publicación: Lab. de Entomología Medica,
Ave. Universidad s/n, Ciudad Universitaria, 2º piso, Unidad
B, San Nicolás de los Garza, Nuevo León, México, C.P.
66450. Teléfono: + 52 81 83294111. Fax: + 52 81
83294111. www.artropodosysalud.com. Editor
Responsable: Dr. Gustavo Ponce García. Reserva de
derechos al uso exclusivo No. 04-2013-120916500700-
102. ISSN en trámite, ambos otorgados por el Instituto
Nacional del Derecho de Autor, Registro de marca ante el
Instituto Mexicano de la Propiedad Industrial: En trámite.
Responsable de la última actualización de este Número,
Unidad Informática, Ing. Oscar Manuel Loaiza Jiménez,
Albino Espinoza 1308, Col. Obrera, C.P. 64010,
Monterrey, Nuevo León México. Fecha de última
modificación: 1 de enero de 2016.
Las opiniones expresadas por los autores no
necesariamente reflejan la postura del editor de la
publicación.
Prohibida su reproducción total o parcial de los contenidos
e imágenes de la publicación sin previa autorización del
Editor.
Todos los derechos reservados
© Copyright 2014

A Los Lectores:
Estimados lectores bienvenidos a la edición número uno de la
revista de divulgación Artrópodos y Salud, agradeciendo el interés
por la lectura de este número. Esta publicación será publicada
tetramestralmente, en la cual les presentamos una serie de
información sobre tópicos relacionados con los artrópodos y su
efecto en la salud, humana, animal y vegetal.
En nuestra sección Editorial citamos a la publicación The Lancet,
donde se da una semblanza sobre Zika Virus, tema abordado de
manera general.
En la sección de monografías, se habla de la Dra. Anita
Hoffmann, sus obras y legado.
En el artículo, Brief Description Of: Zika Virus, nos da un
panorama general de los principales descubrimientos en este
tema, así también se abordan otros temas como es la Mosquitos
Silvestres (Diptera: Culicidae) de la Ría de Celestún Yucatán,
Mecanismos No Químicos De Comunicación En Insectos,
Virus De La Tristeza De Los Cítricos, Tifus Murino:
Generalidades y WHO reveals its shopping list for weapons
against Zika.
Los invitamos de la manera más atenta a que disfrute del
contenido de esta publicación, cuyo objetivo es divulgar
conocimiento dentro del apasionante tema de los Artrópodos y su
efecto en la Salud en general.
CONSEJO EDITORIAL
1

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Editorial: Zika Virus
2
Editorial
Zika Virus
Is an arbovirus of the Flaviviridae family,
which includes dengue, West Nile, yellow fever,
and Japanese encephalitis viruses. Like other
members of the genus, Zika virus contains a
positive, single-stranded genomic RNA encoding
a polyprotein.
Zika virus was first isolated in 1947 from a
rhesus monkey in the Zika forest of Uganda.
Transmitted by the Aedes genus of mosquito, for
decades, the disease mainly affected monkeys in
a narrow equatorial belt stretching across Africa
and into equatorial Asia. It occasionally affected
human beings causing a mild disease.
In 2007, the first documented outbreak of
Zika occurred in the Federated States of
Micronesia where 185 suspected cases were
reported, of which 49 were confirmed and 59
were considered probable. Between 2013 and
2014, four additional Pacific Island nations
documented large Zika outbreaks. In French
Polynesia, the Zika outbreak was associated with
neurological complications, but at a time when
the virus was co-circulating with dengue.
In May, 2015, WHO confirmed the presence
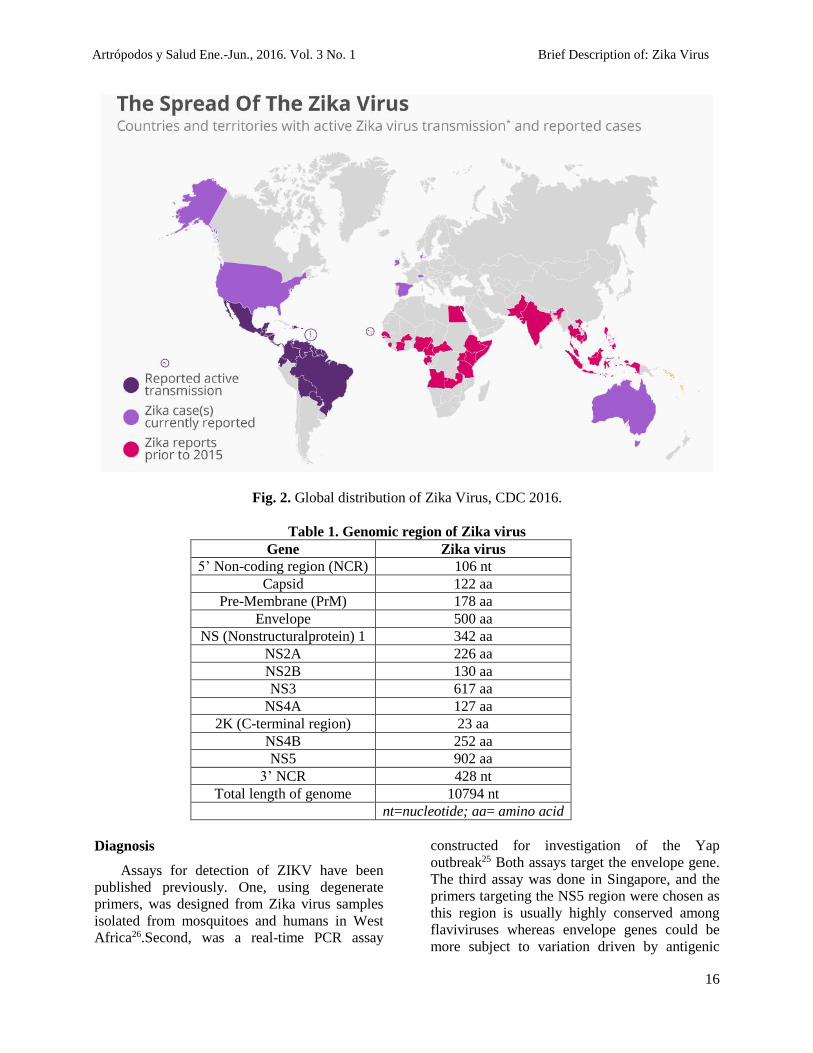
of Zika virus in the Americas. As of Feb 1, 2016,
active transmission has been reported in 28
countries and territories, mainly in the
Americas, but also in Africa (Cape Verde)
and Oceania (American Samoa, Samoa).
At-risk population: As of Feb 1, 2016,
active transmission of Zika virus was being
reported in 28 countries and territories (figure),
mostly in the Americas, including Brazil,
Colombia, Venezuela, Mexico, Haiti, and
Barbados. Sylvain Aldighieri, head of
International Health Regulations Epidemic Alert
and Response at the Pan American Health
Organization (PAHO), explained to reporters in
a WHO briefing on Jan 28 that the fast
dissemination of the virus in the Americas was
down to two reasons: the population being
immunologically naive to Zika virus and the
ubiquitous presence of the vector—Aedes
mosquitoes. The vector can be found in the
region from southern USA to northern Argentina
(excluding continental Chile), an area
encompassing around 500 million people. WHO
estimates that there will be 3–4 million cases of
Zika infection (including asymptomatic cases) in
the Americas in the next 12 months.
The Lancet. Febrero 2016.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Monografia: Anita Hoffmann
3
Dra. Ana Esther Hoffmann Mendizábal
(Anita Hoffmann)
(Marzo 1919 - Octubre 2007)
Anita Hoffmann nació en el estado de
Puebla, México, en 1919. Su pasión por los
insectos se dio gracias a su padre, el Dr. Carlos
Cristian Hoffmann (1876-1942), reconocido
entomólogo mexicano de origen alemán, quien
llevaba a Anita para que participara en sus
excursiones por todo México. En 1939, cuando
se funda la Facultad de Ciencias de la UNAM,
ella entra a dicha institución para formar parte de
la primera generación de estudiantes,
comenzando la Maestría en Biología, finalizando
sus estudios cuatro años más tarde, en 1941.
En 1991 la doctora Anita Hoffmann donó al
Instituto de Biología de la Universidad Nacional
Autónoma de México un trabajo que ocupó más
de 50 años de su vida: la más importante
colección acarológica de México, conformada
por más de 100 mil ejemplares pertenecientes
aproximadamente a mil especies de 149 familias
de ácaros, única en el país e incluida desde 1979
en el Índice de colecciones de acarología del
mundo. Junto con ella, cedió también otras tres
colecciones de relevancia científica: una
aracnológica, con más de 400 especies; otra, que
incluye especies de siete órdenes y trece familias
de quilópodos y diplópodos (ciempiés y milpiés,
respectivamente) y una más de insectos
ectoparásitos, con ejemplares de importancia
médica y veterinaria. Investigadora, educadora,
académica y reconocida bióloga mexicana
especializada en Acarología y Parasitología,
pionera en el estudio de los arácnidos y ácaros en
México. Fundó el Laboratorio de Acarología (el
primero en América latina) en la Escuela
Nacional de Ciencias Biológicas del Instituto
Politécnico Nacional en 1965, así como del
Laboratorio de Acarología de la Facultad de
Ciencias de la Universidad Nacional Autónoma
de México en 1977. Entre sus publicaciones más
Importantes se encuentran:
2000 - Biodiversidad de los ácaros en
México. CONABIO 2002 - Biodiversidad,
Taxonomía y Biogeografía de Artrópodos de
México. Hacia una Síntesis de su Conocimiento.
CONABIO, Acari, Volumen III, 223-276.2004 -
Cap. 13: Los artrópodos de las cavernas de
México. 229-326. Biodiversidad, Taxonomía y
Biogeografía de Artrópodos de México: Hacia
una Síntesis de su Conocimiento. CONABIO,
Facultad de Ciencias, IBUNAM, Vol. IV.
Por: Ángel Alberto Muñoz Ochoa y Cristian
Daniel Escobedo Garza (2015).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 World Report: WHO reveals its shopping list for
weapons against Zika
4
World Report
WHO reveals its shopping list for weapons
against Zika
Expunging its weak performance against the recent Ebola epidemic, WHO takes
swift steps to make sure that the world will have new ways to battle Zika virus.
If viruses had ears, Zika virus would be
in bad shape after listening to a WHO official
describing the armamentarium the world is
getting ready to launch against it. “Based on
our experience with research and
development during the west Africa Ebola
epidemic, WHO’s [R&D] response is
proceeding very quickly for Zika”, Marie-
Paule Kieny, WHO Assistant Director-
General, Health Systems and Innovation, told
a press conference on Feb 12 at the agency’s
headquarters in Geneva, Switzerland. WHO,
she said, has identified industry and research
institutions interested in developing the tools
needed to combat the Zika epidemic now
taking hold in 26 countries (panel). There is
an urgent need for vaccines, diagnostic tests,
and prophylactic medicines against Zika as
well as innovative technologies to pit against
the mosquitoes that carry the virus.
In the vaccine arena, the chase is on
between 15 research groups, most of which
are in Europe. Nearly all are still in the early
stages of vaccine development. Leading the
pack by a small margin is a team at the US
National Institutes of Health working on
several vaccines and another team working in
an Indian firm, Bharat Biotech. Candidate
vaccines in the pipeline are either based on
attenuated viral vectors, as is the case for the
recently licensed dengue vaccine, or on
inactivated products (DNA, recombinant
proteins, or killed Zika virus), as is the case
for some of the licensed Japanese encephalitis
vaccines.
An encouraging factor in developing a
vaccine against Zika virus, Kieny noted, is
that “Zika is a fl avivirus and vaccines have
been successfully produced against other
flaviviruses, such as yellow fever, Japanese
encephalitis, and dengue”. A somewhat
discouraging factor she noted is that “all the
Zika vaccine candidates are at a very basic
level and it could be something like 18
months before large-scale trials could begin”.
The challenge to produce reliable diagnostic
tests needed to facilitate research, clinical
management, and surveillance activities has
attracted
“We need to be sure that
poorquality or fake Zika tests will not
be around, as was the case with the
Ebola epidemic.”
ten biotech companies. Three types of
tests are being developed. One is needed to
detect the presence of the virus in an infected
person and to determine in what body fl uid it
is found. Another is an antibody-based test
for detecting a past infection. And a third is a
rapid diagnostic test that is field-friendly and
can give a result on a dipstick within minutes.
“So far none of these tests have been
independently validated and none have
regulatory approval”, said Kieny. “We need
to be sure that poorquality or fake Zika tests
will not be around, as was the case with the
Ebola epidemic.”
“Studies”, Kieny pointed out, “are also
underway on antiviral medications,
particularly prophylactic products that could

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 World Report: WHO reveals its shopping list for
weapons against Zika
5
prevent infection in vulnerable people,
especially pregnant women”.
Research and development efforts are
also targeting the Aedes aegypti mosquito
vector of Zika virus. “Some innovative
methods of preventing transmission of Zika
to people seem promising”, Kieny said. One
strategy reduces the local mosquito
population by releasing into the environment
bacteria that can prevent the reproduction of
the insects. Another achieves the same
purpose by using male mosquitoes engineered
to carry a lethal gene that alters the viability
of the eggs that they have fertilised.
A lesson WHO learned from the Ebola
epidemic, Kieny explained, is the need,
during a life-threatening epidemic, for a fast-
track system that allows the deployment of
new promising tools that have not run the full
gamut of safety and effi cacy testing but
could save lives. “WHO”, Kieny said, “has
set up a WHO Emergency Assessment and
Listing procedure, which guarantees a certain
level of quality and performance, balancing a
small risk against a high risk of mortality and
morbidity from an epidemic”.
WHO is also preparing a so-called R&D
Blueprint roadmap “that will facilitate a swift
research and development response to an
epidemic”. Last December, a group of experts
met in WHO’s Geneva headquarters to draw
up a list of diseases qualifying for activation
of the R&D Blueprint. Zika was high on the
list.
John Maurice
Published Online February 16, 2016
http://dx.doi.org/10.1016/ S0140-
6736(16)00390-1.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
6
Mosquitos silvestres (Diptera: Culicidae)
de la Ría de Celestún Yucatán. P. Manrique-Saide1, 2, Gustavo Ponce-García3, E. Ceh-Pavía1, J. Góngora-Salinas1, A. Medina-Barreiro2, E. Koyoc-
Cardeña2, S. Dzib-Florez 2 1Departamento de Zoología. Campus de Ciencias Biológicas y Agropecuarias, UADY. Km. 15.5 Carr. Mérida-
X’matkuil s/n Mérida, Yucatán 97315, México. [email protected] 2Unidad Colaborativa para Bioensayos Entomológicos (UCBE), Universidad Autónoma de Yucatán, Km. 15.5 Carr.
Mérida-X’matkuil s/n Mérida, Yucatán 97315, México. 3Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Av. Universidad s/n Cd. Universitaria San
Nicolás de los Garza, N.L. 66455.
Resumen
Se llevaron a cabo colectas de mosquitos por 14 días consecutivos mediante siete colectas en el
amanecer y siete crepusculares; en cinco puntos de muestreo a lo largo de 2.5 km en la Ría de la localidad
costera de Celestún. Se colectaron un total de 2, 202 mosquitos de cinco especies: Anopheles crucians
(Wiedemann), Coquilletidia venezuelensis (Dyar y Knab 1906), Culex quinquefasciatus (Say 1823),
Ochlerotatus sollicitans (Walker 1856), Ochlerotatus taeniorhynchus (Wiedemann 1821) y Psorophora
confinnis. Hubo mas registro de mosquitos en el amanecer (1122) en comparación con el crepúsculo
(1080). El cebo humano presentó mas altos registros con 1081 ejemplares (51.2% del total colectado). Oc.
taeniorhynchus, fue la especie mas abundante en todo el estudio con 2,099 especímenes (95.32% del total
colectado). An. crucians, Cx. quinquefasciatus, Oc. sollicitans y Oc. taeniorhynchus, han sido reportados
con interés zoonótico para localidad de Celestún y los habitantes pudieran estar en riesgo potencial de
infección por microfilarias.
Palabras Clave: Mosquitos de la Ría de Celestún, Cebo humano, trampeo
Introducción
Hasta la fecha han sido reportadas 3300
especies de mosquitos pertenecientes a 41
géneros, agrupados en la familia Culicidae (1).
En México se reconocen 228 especies
pertenecientes a 21 géneros de las cuales solo 40
se han registrado en el estado de Yucatán),
representando el 18% de la diversidad reportada
para el país (1-3). Los estudios de campo que
incluyen el monitoreo de los ambientes de
crianza y de las poblaciones de mosquitos, así
como la actividad de las especies que actúen
como vectores de enfermedades, nos permite un
control más eficiente de mosquitos y poder tener
información de cuando y donde se deben llevar a
cabo medidas de protección (2).
En la localidad costera de Celestún, se ha
reportado la más alta prevalencia (59.8%) de
dirofilariosis canina en todo México (4) y en
estudios de incriminación de mosquitos
involucrados con este padecimiento en secas y
lluvias, se demostró la presencia de nueve
especies que estuvieron infectados con
microfilarias y se reportó por primera vez en
México al mosquito Ochlerotatus
taeniorhynchus como vector principal en el ciclo
doméstico de transmisión de Dirofilaria immits
en perros microfilarémicos del centro de la
localidad (5,6). Sin embargo hasta el momento
no existe un reporte de los mosquitos presentes
en áreas aledañas cercanas al centro de la
localidad. Por tal motivo del presente estudio fue
el de colectar e identificar las especies de
mosquitos silvestres en un manchón de
vegetación de manglar y humedal en la Ría de
Celestún, Yucatán, México, en los alrededores
de la localidad y determinar si son las mismas
con interés zoonótico ya colectadas con
anterioridad y que pudieran representar un riesgo
potencial para los habitantes.
Materiales y métodos
Área de estudio
El estudio se llevó a cabo en la en la
localidad costera de Celestún (Cabecera del

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
7
municipio del mismo nombre) (Fig. 1),
localizada en la costa Oeste del estado de
Yucatán en el Golfo de México (20º51’39” N, 90
º23’33”). El clima es cálido y semi-árido, con
temperaturas máximas mensuales de 35-40ºC,
humedad relativa de 65-95% y un promedio de
precipitación anual de 767 mm. La temporada de
lluvias va de junio a septiembre que es cuando se
presenta la precipitación anual total, la
temporada de secas va de octubre a mayo; y está
inmersa en uno de los sistemas de humedales del
sureste de México, en una línea de litoral
bordeado por un lado por el Golfo y por otro por
una laguna costera (SEMARNAT, 2000).
Figura 1. Localización de los sitios de colecta en el Municipio de Celestún.
Colecta e identificación de mosquitos
silvestres.
Para la colecta de mosquitos se realizaron
colectas diarias durante 14 días durante el mes
de Agosto 2009, que consistieron en siete
colectas en el amanecer (6.00-8.00 hrs) y siete
colectas crepusculares (17.00-19.00 hrs.) en
cinco puntos de muestreo a lo largo de 2.5 km
(500 m entre cada punto), a lo largo de la Ría de
Celestún. Se utilizaron dos aspiradoras portátiles
de mochila (battery powered Modified CDC
Backpack Aspirator Model 1412 John W. Hock
Co.®).
Se establecieron dos metodologías (Fig.2): i)
Colectas en vegetación para capturar los
mosquitos que estuvieran reposando sobre las
hojas de los árboles, arbustos y plantas; y ii)
Cebo humano que consistió en colectar todos los
mosquitos que se posaban sobre los voluntarios
en un intento por alimentarse, aspirando brazos y
piernas; esto de acuerdo a la metodología
utilizada por Jones et al. (2004). Los mosquitos
capturados se transportaron al laboratorio de
DUMAC (Ducks Unlimited de México A.C.)
donde se sacrificaron por congelación y se
separaron por sexo y especie; la se realizó
cotejando sus características morfológicas con
las descripciones en Carpenter y LaCasse (1995)
y Darsie y Ward (1981).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
8
Figura 2. Métodos de colecta de mosquitos:
vegetación (A y B); y cebo humano (C y D).
Análisis de los datos
Este fue un estudio descriptivo transversal
en el cual se calculó la abundancia relativa de las
especies capturadas: número de mosquitos
capturados de cada especie entre el número total
colectado; número de especies capturadas en el
amanecer y el crepúsculo entre el total de
especies colectado; la abundancia de las
especies colectadas en cada sitio entre el total
colectado y las abundancias de acuerdo a las
colectadas en cebo humano y en vegetación.
Resultados
Especies de mosquitos capturadas
Se capturaron en total en todo el estudio,
2,202 mosquitos adultos representados en 5
géneros y seis especies anteriormente reportados
para Celestún: Anopheles crucians
(Wiedemann), Coquilletidia venezuelensis
(Dyar y Knab 1906), Culex quinquefasciatus
(Say 1823), Ochlerotatus sollicitans (Walker
1856), Ochlerotatus taeniorhynchus
(Wiedemann 1821) y Psorophora confinnis
(Cuadro 5). La especie Oc. taeniorhynchus
(comúnmente llamado “mosquito del mangle”)
fue definitivamente la especie predominante en
todos los métodos de colecta combinados
(95.32%). El número de mosquitos capturados y
su abundancia relativa se presentan en el cuadro
1.
Cuadro 1. Especies de mosquitos (♀) capturadas (en orden descendente en cuanto su abundancia).
Especies Número total Abundancia
relativa (%)
Oc. Taeniorhynchus (Wiedemann 1821) 2,099 95.32
Oc. Sollicitans (Walker 1856) 46 2.08
An. crucians (Wiedemann), 44 1.99
Cq. Venezuelensis (Dyar y Knab 1906) 6 0.27
Cx. quinquefasciatus (Say 1823) 4 0.18
Ps. Confinnis (Lynch Arribálzaga) 3 0.13
Total 2,202 100
El mayor número de registro de mosquitos en todos los sitios, fue en las colectas realizadas durante el
amanecer, con respecto al crepúsculo (51% y 49%) respectivamente; y en ambos periodos hubo registro
de todas las especies capturadas en todo el estudio. Tanto en amanecer como en crepúsculo Oc.
taeniorhynchus continuó siendo la especie más abundante (Cuadro 2).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
9
Cuadro 2. Especies de mosquitos (♀) capturados en colectas durante el amanecer y el crepúsculo (en
orden descendente en cuanto su abundancia).
Especies Amanecer Crepúsculo
Oc. taeniorhynchus 1,084 1,015
Oc. sollicitans 20 26
An. crucians 10 34
Cq. venezuelensis 2 4
Cx. quinquefasciatus 4 0
Ps. confinnis 2 1
Total 1,122 1,080
Cuadro 3. Especies de mosquitos (♀) capturados sobre vegetación y con cebo humano en los diferentes
puntos de colecta (en orden descendente en cuanto su abundancia).
Especies Sitios Total
Colectas en vegetación 1 2 3 4 5
Oc. taeniorhynchus 431 136 206 188 57 1,018
Oc. sollicitans 0 1 0 3 0 4
An. crucians 23 0 0 0 0 23
Cq. venezuelensis 1 2 1 0 0 4
Cx. quinquefasciatus 2 0 0 0 0 2
Ps. confinnis 2 0 0 0 0 2
Total 459 139 207 191 57 1,053
Colecta en cebo humano 1 2 3 4 5
Oc. taeniorhynchus 323 304 204 165 85 1,081
Oc. sollicitans 13 8 12 6 3 42
An. crucians 20 1 0 0 0 21
Cq. venezuelensis 0 1 1 0 0 2
Cx. quinquefasciatus 2 0 0 0 0 2
Ps. confinnis 1 0 0 0 0 1
Total 359 314 217 171 88 1,149
Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
10
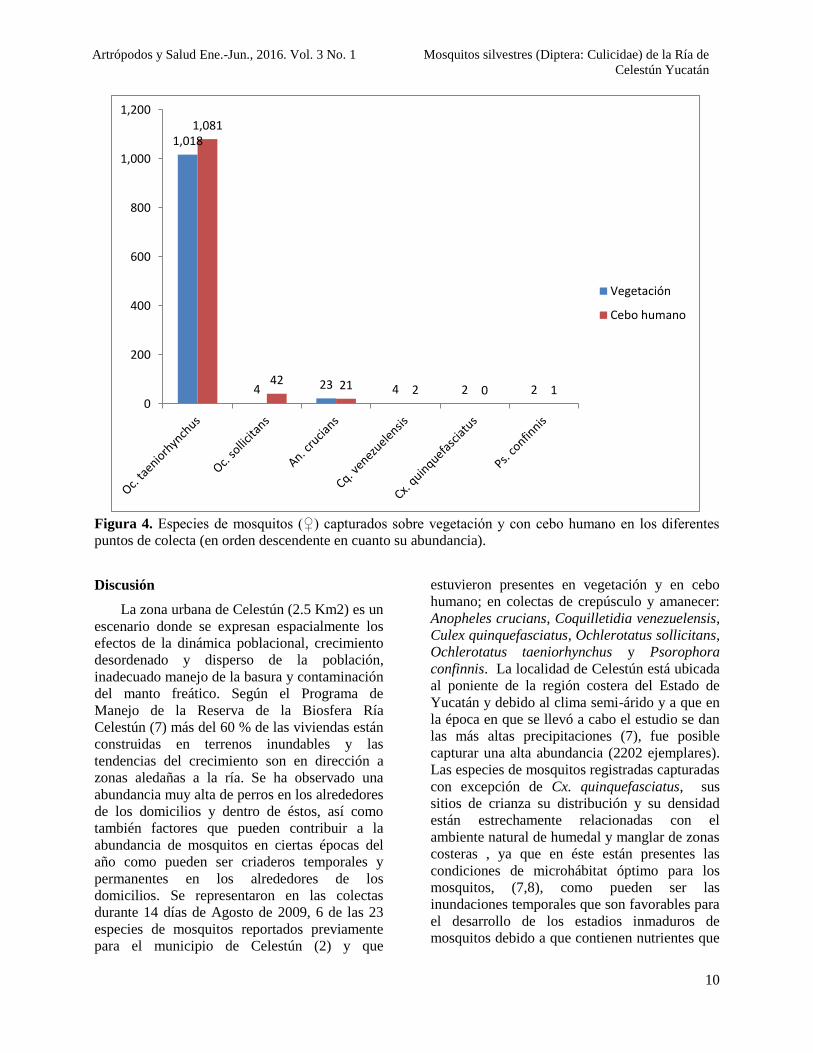
Figura 4. Especies de mosquitos (♀) capturados sobre vegetación y con cebo humano en los diferentes
puntos de colecta (en orden descendente en cuanto su abundancia).
Discusión
La zona urbana de Celestún (2.5 Km2) es un
escenario donde se expresan espacialmente los
efectos de la dinámica poblacional, crecimiento
desordenado y disperso de la población,
inadecuado manejo de la basura y contaminación
del manto freático. Según el Programa de
Manejo de la Reserva de la Biosfera Ría
Celestún (7) más del 60 % de las viviendas están
construidas en terrenos inundables y las
tendencias del crecimiento son en dirección a
zonas aledañas a la ría. Se ha observado una
abundancia muy alta de perros en los alrededores
de los domicilios y dentro de éstos, así como
también factores que pueden contribuir a la
abundancia de mosquitos en ciertas épocas del
año como pueden ser criaderos temporales y
permanentes en los alrededores de los
domicilios. Se representaron en las colectas
durante 14 días de Agosto de 2009, 6 de las 23
especies de mosquitos reportados previamente
para el municipio de Celestún (2) y que
estuvieron presentes en vegetación y en cebo
humano; en colectas de crepúsculo y amanecer:
Anopheles crucians, Coquilletidia venezuelensis,
Culex quinquefasciatus, Ochlerotatus sollicitans,
Ochlerotatus taeniorhynchus y Psorophora
confinnis. La localidad de Celestún está ubicada
al poniente de la región costera del Estado de
Yucatán y debido al clima semi-árido y a que en
la época en que se llevó a cabo el estudio se dan
las más altas precipitaciones (7), fue posible
capturar una alta abundancia (2202 ejemplares).
Las especies de mosquitos registradas capturadas
con excepción de Cx. quinquefasciatus, sus
sitios de crianza su distribución y su densidad
están estrechamente relacionadas con el
ambiente natural de humedal y manglar de zonas
costeras , ya que en éste están presentes las
condiciones de microhábitat óptimo para los
mosquitos, (7,8), como pueden ser las
inundaciones temporales que son favorables para
el desarrollo de los estadios inmaduros de
mosquitos debido a que contienen nutrientes que
1,018
4 23 4 2 2
1,081
42 21 2 0 10
200
400
600
800
1,000
1,200
Vegetación
Cebo humano

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
11
pueden ser renovados constantemente. Todas son
picadores persistentes que presentan actividad de
picadura tanto en el día como en la noche; estas
especies atacan a sus hospederos cuando estos
irrumpen en sus sitios de reposo lo que los
convierte en seria amenaza ya que en la zona de
estudio específicamente en el primer sitio, se
encuentra una salinera artesanal que es una de
las actividades a las que se dedican los
habitantes de la localidad además de la pesca y el
ecoturismo, y están expuestos a las picaduras de
estos mosquitos. Otra particularidad de estas
especies es que pueden viajar grandes distancias
desde sus sitios de crianza (1,3,9), razón por la
cual también han sido colectados en la zona
habitada de la localidad de Celestún. En los dos
estudios previos (5,6), llevados a cabo en la zona
centro de la localidad, se había registrado la
presencia de Cx. quinquefasciatus infectados
naturalmente con microfilarias; no así en el
presente estudio, esta especie es uno de los
vectores de Virus del Oeste del Nilo (VON) y su
ausencia podría estar influenciada por sus
hábitos biológicos, ya que presenta una fuerte
asociación con el ambiente doméstico y se le
encuentra en sitios de crianza de inmaduros
alrededor de las viviendas y reposando dentro de
las habitaciones de las mismas (9), lo que le da
importancia zoonótica.
Tanto en amanecer como en crepúsculo los
mosquitos fueron similarmente muy activos y
Oc. taeniorhynchus continuó siendo la especie
más abundante. A pesar de que en el amanecer
hubo una aparente mayor abundancia numérica,
ésta realmente se debió a que hubo un registro
mayor de Oc. taeniorhynchus (Cuadro 2), sin
embargo fue en el crepúsculo en el que hubo
mayores registros tanto de otras especies
colectadas como de mosquito del mangle; lo
anterior podría deberse a que en las horas de
colecta establecidas algunas especies no hayan
iniciado aún su actividad de picadura a pesar de
estar presentes en la zona.
Como era de esperarse, todas las especies de
mosquitos estuvieron más atraídos por los
colectores que sirvieron de cebo humano, en
comparación con la colecta de vegetación, ya
que los mosquitos antropofílicos (atraídos por
humanos) responden preferentemente al CO2
liberados en la respiración al ácido láctico y a
una variedad de compuestos del sudor (12, 13,
14). La poca presencia de mosquitos en
vegetación podría deberse a que en el momento
de la colecta, había pocos ejemplares en las
plantas de cada sitio en el momento en que se
pasó el aspirador. El conocimiento de la
distribución de las especies de es esencial para
determinar áreas de potencial riesgo de
transmisión de patógenos (3), dado por el
contacto que estos podrían tener con las personas
ya que según el Programa de Manejo de la
Reserva de la Biosfera Ría Celestún (8) más del
70% de las viviendas están construidas en
terrenos inundables y las tendencias del
crecimiento son en dirección a zonas aledañas a
la ría en donde se crían las distintas especies
mosquitos que se distribuyen por toda la zona,
enfatizando la importancia de este conocimiento
por seguridad y salud de habitantes y visitantes
de Celestún como centro turístico.
Debido a que se colectaron cuatro especies
ya reportadas en la localidad de interés zoonótico
por Dirofilaria immitis: An. crucians, Cx.
quinquefasciatus, Oc. sollicitans y Oc.
taeniorhynchus (6, 7), existe un riesgo potencial
de que la población de Celestún este expuesta a
los mosquitos infectados con microfilarias, ya
que habitan en una zona de alta prevalencia de
Filariosis (5), de igual manera están en contacto
o comparten los mismos espacios, con perros
(hospederos) de esas áreas, con el vector
infectado y con el parásito, por lo tanto, no sería
sorprendente que estudios futuros reportaran
evidencia de la enfermedad en humanos para
esta localidad.
Conclusiones
En el amanecer se registraron los mayores
números de mosquitos. Oc. taeniorhynchus, fue
la especie más abundante en todo el estudio.
Todas las especies de mosquitos capturados
estuvieron más atraídas hacia el cebo humano.
An. crucians, Cx. quinquefasciatus, Oc.
sollicitans y Oc. taeniorhynchus pueden estar en
contacto con las personas que entren a su hábitat
y presentar un riesgo zoonótico potencial.
Referencias
1. Ibáñez-Bernal, S., Strickman, D. y Martínez-
Campos, C. (1996) Culicidae (Diptera).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mosquitos silvestres (Diptera: Culicidae) de la Ría de
Celestún Yucatán
12
Capítulo 38. pp. 591-602. En: Llorente-
Bousquets, J., García-Aldrete, A., González-
Soriano, E. (Eds.). Biodiversidad,
taxonomía y biogeografía de artrópodos de
México: hacia una síntesis de su
conocimiento. UNAM-CONABIO, México.
2. Ibáñez-Bernal S. 1999. Los Diptera
hematófagos y taxa relacionados de dos
Áreas Protegidas del Estado de Yucatán,
México (Insecta). México: InDRE
[Secretaría de Salud, Instituto Nacional de
Diagnóstico y Referencia Epidemiológicos,
Laboratorio de Entomología e Insectario].
3. Kline DL. 2006. Mosquito population
surveillance techniques. Techical Bulletin of
Florida. Mosquito Control Association. 7: 2-
8.
4. Gaffigan, T., Wilkerson, R., Pecor, J., Stoffer,
J. y Anderson, T. (2011) Systematic catalog
of Culicidae. The Walter Reed Byosistematic
Unit [En línea], disponible en:
http://www.mosquitocatalog.org/ [Accesado
el día 19 de junio de 2013].
5. Caro-González JA, Bolio-González ME,
Escobedo-Ortegón FJ, Manrique-Saide P,
Rodríguez-Vivas RI, Rodríguez-Buenfil JC,
Sauri-Arceo CH. 2011. Prevalence of
Dirofilaria immitis Infection in Dogs from
Celestun, Mexico, Using Polymerase Chain
Reaction Test. Vector Borne & Zoonotic
Diseases, 11(2):193-6.
6. Manrique-Saide P, Bolio-González M, Sauri-
Arceo C, Dzib-Florez S, Zapata-Peniche A.
2008. Ochlerotatus taeniorhynchus: a
probable vector of Dirofilaria immitis in
coastal areas of Yucatan, Mexico. Journal of
Medical Entomology 45: 169–171.
7. Manrique-Saide P, Escobedo-Ortegón J,
Bolio-González M, Sauri-Arceo C, Dzib-
Florez S, Guillermo-May G, Ceh-Pavía E,
Lenhart A. 2010. Incrimination of the
mosquito, Aedes taeniorhynchus, as the
primary vector of heartworm, Dirofilaria
immitis, in coastal Yucatan, México.
Veterinary Entomology 24:456–460.
8. Secretaría de Medio Ambiente y Recursos
Naturales (SEMARNAT). 2000. Programa
de Manejo de la Reserva de la Biosfera Ría
Celestún. Comisión Nacional de Áreas
Naturales Protegidas, Mexico, DF.
9. Mercer, DR, Sheeley SL, Brown EJ. 2005.
Mosquito (Diptera: Culicidae) Development
within Microhabitats of an Iowa Wetland.
Journal of Medical Entomology 42(4):685-
693.
10. Subra R. 1981. Biology and control of Culex
pipiens quinquefasciatus Say, 1823
(Diptera, Culicidae) with special reference
to Africa. Insect Science Application. 1(4):
319-338.
11. Carpenter S., LaCasse W. (1955).
Mosquitoes of North America (North of
Mexico). University of California, Press,
Berkeley, CA.
12. Torres-Estrada JL, Rodríguez MH. 2003.
Physic-chemical signals involved in host
localization and induction of disease vector
mosquito bites. Salud Pública de México.
45:497-505.
13. Rebollar-Téllez EA. 2005. Human body
odor, mosquito bites and the risk of Disease
transmission. Folia Entomológica Mexicana.
44 (2): 247-265.
14. Day JF. 2005 Host-Seeking Strategies of
Mosquito Disease Vectors. Journal of the
American Mosquito Control Association
21(4) Supplement: 17–22.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
13
Brief Description of: Zika Virus Omar Eduardo Tova-Herrera1, Olga Karina Villanueva-Segura2, Jorge Martínez Dávila3, Mayra A. Gomez-Govea4,
Aron Medina-Sanchez5, Michelle de Jesús Zamudio-Osuna6, Laura E. Martínez-de-Villarreal6, Beatriz Lopez-
Manroy3, Irám Pablo Rodríguez-Sanchez6
1,2,4 Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas. 3Facultad de Medicina, 5Secretaria de
Salud de Jalisco. Guadalajara, Jalisco, México, 6Facultad de Medicina, Departamento de Genética. San Nicolás de
los Garza, Nuevo León 66451, México.E-mail:[email protected]
Abstract
Zika is a mosquito-borne viral disease caused by Zika virus (ZIKV), a flavivirus from the Flaviviridae
family, initially identified in 1947 in the Zika forest in Uganda in the Rhesus macaque population.
Comprehensive genomic comparison showed different sub-clades reflecting the existence of two main
lineages, one African and one Asia lineage. The main clinical symptoms in patients are low-grade fever (<
38.5 °C), transient arthritis/arthralgia with posible joint swelling (mainly in the smaller joints of the hands
and feet) and maculo-papular rash (that often starts on the face and then spreads throughout the body),
conjunctival hyperaemia or bilateral non-purulent conjunctivitis with general non-specific symptoms such
as myalgia, asthenia and headaches. Clinical symptoms of Zika disease appear after an incubation period
ranging between 3 and 12 days. The disease symptoms are usually mild and short lasting (2–7 days), and
infection may go unrecognised or be misdiagnosed as dengue. Association with neurological
complications such as Guillain-Barré syndrome remains under investigations. A high rate of asymptomatic
infection with ZIKV is expected, similar to other flaviviral infections, such as dengue and West Nile fever.
Approximately one in four people infected with ZIKV are believed to develop symptoms. Most people
fully recover without severe complications, and hospitalisation rates are low. To date, there have been no
reported deaths associated with ZIKV infection. The purpose of this article is to inform people about the
virus and create awareness of prevention against the mosquito.
Keywords: Zika Virus, Flavivirus, Aedes aegypti.
Introduction
The genus Flavivirus of the family
Flaviviridae comprises around 70 viruses such
as: dengue (DEN), Japanese encephalitis (JE),
St. Louis encephalitis (SLE), and yellow fever
(YF), which are important human pathogens.
The difficulty encountered with Flavivirus
classification partly derives from the extensive
geographic distribution and the diversity of the
arthropod vectors or vertebrate hosts associated
with biological transmission of these viruses1. In
this genus is found the Zika Virus (ZIKV), an
emerging arthropod-borne virus (arbovirus)
isolated in 1947 from a rhesus monkey in the
Zika Forest near Entebbe, Uganda during a
project researching Yellow Fever virus2,3. This
was the first human ZIKV infection reported in
Uganda in 1964, and the whole genome was
sequenced in 20064
Since 1947, sporadic human infections were
reported in Africa and Asia, been during 2007 in
Yasp State, the Federated States of Micronesia
the first well documented ZIKV outbreak,
approximately three-quarters of Yap residents
were infected with Zika virus5. By October 2013,
an outbreak occurred in French Polynesia with
approximately 11% of population infected6.
Also, there have been reports of returning
travellers from non-endemic countries including
Japan, Germany, Italy, Canada, Australia and the
United States7,8.
ZIKV has been isolated from Ae. africanus,
Ae. apicoargenteus, Ae. luteocephalus, Ae.
aegypti, Ae. vitattus, and Ae. furcifer
mosquitoes9,10 and the most common clinical
manifestations in patients with Zika infections
included high fever, malaise, stomach ache,
diarrhea, conjunctivitis, dizziness, and anorexia;

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
14
other less frequent manifestations included
myalgia, headache, retro-orbital pain, edema,
and vomit3,11
A brief description of the Zika virus
The Aedes mosquito that deposits the virus
in the epidermis and dermis of the bitten host
during a blood meal transmits ZIKV. Indeed,
both skin fibroblasts and epidermal keratinocytes
were found to be highly permissive to infection
with ZIKV Infection of skin fibroblasts rapidly
resulted in the presence of high levels of RNA
copy numbers and a gradual increase in the
production or ZIKV particles over time,
indicating active viral replication in the infected
cells12.
Although a ZIKV entrance mechanism into
the cell is poor understood, it is known that
flaviviruses mechanism is mediated by the E
glycoprotein and cell surface receptors13. The
initial contact between flaviviruses and the host
cell is mediated by glycosaminoglycans (GAGs),
such as heparan sulfate proteoglycans or
syndecans14, 15. GAGs are long, unbranched,
sulfated polysaccharides normally found linked
to core proteins attached to cellular surfaces
(proteoglycans) 16.
Virus-GAG interactions is due to an
electrostatic interaction between the positive
amino acids of E glycoprotein and the negative
charge of sulfate groups on the polysaccharide,
making this interaction for concentrating the
viral particles around the cell before involving
the receptors for entry14.
Despite several researchers trying to
elucidate the receptors that mediate the flavivirus
entry, so for this process is not well understood.
A large number of molecules have been
described as flavivirus candidate receptors in
different cell types, but their precise role in virus
endocytosis remains obscure13. Currently, the
best-characterized protein families that bind to
and enhance flavivirus infection are C-type
(calcium-dependent) lectin receptors and the
recently identified phosphatidylserine receptors,
T-cell immunoglobulin and mucin domain (TIM)
and TYRO3, AXL and MER (TAM), andαvß3
integrin13.
In the case of ZIKV, it was reported that
entry is mediated by DC-SIGN, AXL, Tyro3
and, although to a lesser extent, by TIM-1. Once
the virus is transported into the cell, a fight
against the host immune response starts. The
infection of fibroblasts in vitro with ZIKV
strongly induced the expression of several
antiviral gene clusters, such as RIG-I, MDA-5
and TLR3 that are able to detect the presence of
PAMPs15, this for the rapid elimination of the
infection.
Epidemiology and world distribution
After the first appearance in 1947, ZIKV has
been detected in humans from Nigeria during
from 1968–1975 reporting that 40% of Nigerians
had Zika virus neutralizing antibodies. Human
isolates were obtained from febrile children 10
months, 2 years (2 cases), and 3 years of age, all
without other clinical details described, and from
a 10-year-old boy with fever, headache, and
body pains17,18.
Also, from 1951 through 1981, ZIKV
infections have been reported in countries such
as Uganda, Tanzania, Egypt, Central African
Republic, Sierra Leone, and Gabon23-25, and
some countries from Asia such as India,
Malaysia, the Philippines, Thailand, Vietnam,
and Indonesia9,11.
An outbreak of the ZIKV was reported in
2007 on the island of Yap, an island of the
Federated States of Micronesia, physicians of
this island noted an outbreak of illness with
symptoms of rash, conjunctivitis, subjective
fever, arthralgia, and arthritis, similar but with
some impression of clinically differences with
dengue. They took samples and sent to Centers
for Disease Control and Prevention (CDC);
ZIKV was confirmed in 49 cases and 59 were
probable cases. Patients with confirmed or
probable Zika virus disease had IgM antibody
against ZIKV and neutralizing antibodies. No
hospitalization or death was reported. Analysis
of water-holding containers shows that 43%
were infected with mosquito larvae or pupae,
being the Ae. hensilli the predominant species
identified as this was present in 36% of the
containers5. Primates are thought to be wild
reservoirs, although another reservoir host has
not been excluded19. Interestingly, sexual

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
15
transmission of Zika virus between heterosexual
couples was proposed, suggesting another
transmission route for these adaptable viruses11.
Recently, Zika virus was detected in saliva
samples20.
By October 2013 in the French Polynesia, a
53-year-old woman and two other relatives
experienced a mild dengue-like illness consisting
of low fever (<38°C), asthenia, wrist and fingers
arthralgia, headache, and rash, ZIKV was
confirmed by RT-PCR. Ten weeks later the first
report 5,895 patients with suspected ZIKV
infections were recordeded. ZIKV tested serum
from 584 patients; 294 samples were positive.
Regarding the vector, the French Polynesia hosts
several mosquito species, notably Ae. aegypti,
but also other potential vectors, such as Ae.
Polynesiensis21.
On 7 May 2015, the Pan American Health
Organization (PAHO)/World Health
Organization (WHO) issued a recommendation
to states in the Region of Americas to establish
managements for ZIKV infection, as well as to
reduce the presence of ZIKV vectors22. On 15
May 2015, the Ministry of Health of Brazil
confirmed the circulation of ZIKV in the
country. This is the first report of autochthonous
ZIKV infection in Brazilk23. On 20 May 2015,
the state of Sao Paulo notified the detection of a
confirmed case without travel24.
Genetic aspects of Zika virus
Zika virus like other members of the
Flavivirus genus, contains a positive single-
stranded genomic RNA, containing 10,794
nucleotides encoding 3,419 amino acids, giving
a polyprotein that is processed into three
structural proteins. The capsid (C), the precursor
of membrane (prM) and the envelope (E), and
nonstructural proteins NS1 to NS5 (Table 1)4
Virus replication occurs in the cellular
cytoplasm1.
ZIKV is most closely related to Spondweni
virus, there appear to be three lineages of Zika
virus based on NS5 homology, the East-African
lineage, a West-African lineage, and the more
distally related Yap lineage25. The genome
sequences of Zika virus are published in
GenBank. The prototype MR766, isolated from
rhesus monkey 766 in Zika Forest 1947
(GenBank accession number AY632535.1 and
LC002520.1) and Zika EC 2007, obtained from
the Yap outbreak (Gen Bank Accession number
EU545988.1).
Fig 1. Monopartite, linear, ssRNA(+) genome of
of 10,794 bp.The virion RNA is infectious and
serves as both the genome and the viral
messenger RNA. The whole genome is
translated in a polyprotein 3,419 aa long, which
is processed co- and post-translationally by host
and viral proteases.
Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
16
Fig. 2. Global distribution of Zika Virus, CDC 2016.
Table 1. Genomic region of Zika virus
Gene Zika virus
5’ Non-coding region (NCR) 106 nt
Capsid 122 aa
Pre-Membrane (PrM) 178 aa
Envelope 500 aa
NS (Nonstructuralprotein) 1 342 aa
NS2A 226 aa
NS2B 130 aa
NS3 617 aa
NS4A 127 aa
2K (C-terminal region) 23 aa
NS4B 252 aa
NS5 902 aa
3’ NCR 428 nt
Total length of genome 10794 nt
nt=nucleotide; aa= amino acid
Diagnosis
Assays for detection of ZIKV have been
published previously. One, using degenerate
primers, was designed from Zika virus samples
isolated from mosquitoes and humans in West
Africa26.Second, was a real-time PCR assay
constructed for investigation of the Yap
outbreak25 Both assays target the envelope gene.
The third assay was done in Singapore, and the
primers targeting the NS5 region were chosen as
this region is usually highly conserved among
flaviviruses whereas envelope genes could be
more subject to variation driven by antigenic

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
17
pressure. No Zika virus was detected in patient
samples, suggesting that Zika virus not be a
widespread infection in Singapore27. In patients
in French Polynesia, Zika virus RNA has been
detected in the urine more than ten days after
onset of disease28.
ZIKV specific IgM/IgG antibodies can be
detected by ELISA and immunofluorescence
assays in serum specimens, usually from day
five or six of symptomatic illness. Interpretation
of serological results should be considered very
carefully as false positive dengue IgM cross
reactivity both by indirect immunofluorescence
assay and rapid test has been reported in both
primary Zika virus infected patients and also
those with a probable history of other prior
flaviviral infection. There are no commercially
available serological assays for the detection of
Zika virus-specific antibodies25,29,30.
Prevention
We need to remain alert on new emerging or
re-emerging viral agents that could cause a
significant damage in the human population.
ZIKV-related virus such as Chikungunya
(CHIKV), West Nile (WNV) and dengue
(DENV) are the most known of human
pathogenic arthropod-borne viruses (arboviruses)
with altogether rates of morbidity and mortality
surpassing the Ebola´s rates.
Arboviruses present a high capability of
adaptation reflected by the fact of being
transmitted by arthropods (mosquitoes, ticks,
sandflies, midges, bug) and vertebrates during
the life cycle of the virus31. (approximately 300
types of mosquito can transmit arboviruses).
This diversity of species and the wide
distribution of these transmission vectors explain
why arboviruses are so successful in dispersing
globally via the mechanisms highlighted earlier.
A high proportion of arboviruses associated with
human and animal disease circulate in tropical,
and subtropical regions, where mosquitoes and
other flying insects tend to be abundant.
However, many arboviruses also circulate
amongst wildlife species in temperate parts of
the world.
The definition of arthropod-borne disease
transmission was developed in 1881 by Carlos
Finlay32 from the studies of Josiah Clark33, by
that time Finlay suggested that mosquitoes were
the agents for transmission of yellow fever, this
theory was later the base for methods, such as
mosquito eradication, which was used very
successfully in Cuba33, along with the
development of a yellow fever vaccine in the
protection of millions of humans from infection
by this virus34,35.
Currently, advances in both antiviral
molecules and vaccine development have help to
control diseases in patients with viral infections
such as AIDS, Swine influenza, etc. But
considering the eradication of RNA viruses is
not that simple, because of their high mutation
rate and particularly the case of arboviruses, it
has been reported that mosquitoes carry large
numbers of known and unknown viruses that
infect humans, primates, mammals, birds,
insects, and plants35,36.
Preventing or reducing arboviruses
transmission depends entirely on control of the
mosquito vectors or interruption of human–
vector contact.
Environmental management
Control of mosquitoes would be a disaster
that will impact on the survival of many wildlife
species, nonetheless, implementation of
temporary localized arthropod control measures
during epidemics, for example in high density
urbanized areas, can still play an important but
transient role in reducing the impact on humans
and animals of emerging arboviruses.
Control measures consist of integrated
strategies to eliminate or reduce the presence of
mosquitoes including the basics methods as
information to authorities and residents,
composed in urged to residents to remove
breeding site with a specific door-to-door
distributed leaflet.
Three types of environmental management
are defined:
The physical transformations to reduce
vector larval habitats, such as installation of a
reliable piped water supply in communities The
temporary changes involving frequent emptying
of water containers, cleaning of gutters,
sheltering stored tyres from rainfall, recycling or
proper disposal of discarded containers and

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
18
tyres, vigilance from the vicinity of homes of
plants such as ornamental that collect water in
the leaf axils. The human habitation or behavior
of contact, such as installing mosquito nets on
entry points (windows, doors).
Chemical control: larvicides Another strategy widely used but that should
be considered as complementary to
environmental management (restricted to
containers that cannot otherwise) is based on
chemical agents such as methoprene,
pyriproxyfen, and temephos. Two or three
application rounds carried out annually promptly
with proper monitoring of efficacy may suffice,
especially in areas where the main transmission
season is short. Precaution with these molecules
is because of safety of the active ingredients in
the final formulation varies from product to
product, making to required further studies, thus
placing chemicals in domestic water, particularly
drinking water, is often viewed with suspicion in
some communities. Also, current environmental
laws have discontinued the use of these
molecules because the impact on non-target
species, contamination of soil and water and
subsequent mosquito resistance to insecticides37.
Biological control
This control based on the introduction of
organisms that compete with or otherwise reduce
populations of the target species. The most
studied of these is Cyclopoidea (small freshwater
crustaceans) have proved to be effective in
operational contexts in specific container
habitats. Limitations of this methods are: the
expense and task of rearing the organisms on a
large scale, difficulty in applying them and their
limited utility in aquatic sites where temperature,
pH, and organic pollution may exceed the
narrow requirements of the organism. Biological
control methods are effective only against the
immature stages of vector mosquitoes in the
larval habitat where they are introduced38.
In addition, if we want to win the war
against the arboviruses, we need to focus on:
drugs and vaccine development (with
cooperation between academic and private
institutions); vector-controlled strategies (not
affecting the human health and others species);
encourage the development of research
programmes to understand the underlying
mechanisms of arboviral pathogenicity,
evolution, emergence and dispersal; improve
public health measures to inform and educate
citizens in local arboviral disease control
measures, including monitoring and reporting.
Finally, the lessons of the past let us know
that human race can face adversity, just like
arbovirus, humankind can also adapt to
environmental hard times just like did against
Black death, 1918 flu pandemic, Plague of
Justinian, third plague pandemic and more
recently avian and swine influenza46-47.
Acknowledgements
The authors gratefully acknowledge the
critical reading of the manuscript by Sergio
Lozano-Rodríguez, MD.
References
1. Kuno G., Chang G.J., Tsuchiya K.R.,
Karabatsos N. and Cropp C.B. 1998.
Phylogeny of the genus Flavivirus. J Virol.
Jan.72(1):73-83.
2. Dick G.W., Kitchen S.F. and Haddow A.J.
1952. Zika virus. I. Isolations and
serological specificity. Trans R Soc Trop
Med Hyg. Sep.46(5):509-20.
3. Simpson D.I. 1964 Zika Virus Infection in
Man. Trans R Soc Trop Med Hyg. Jul.
58:335-8.
4. Kuno G. and Chang G.J. 2007. Full-length
sequencing and genomic characterization of
Bagaza, Kedougou, and Zika viruses. Arch
Virol.152(4):687-96.
5. Duffy M.R., Chen T.H., Hancock W.T.,
Powers A.M., Kool J.L. and Lanciotti R.S.
2009.Zika virus outbreak on Yap Island,
Federated States of Micronesia. N Engl J
Med. Jun 11.360(24):2536-43.
6. Hancock W.T., Marfel M.and Bel M. 2013
Zika virus, French Polynesia, South Pacific.
Emerg Infect Dis. Nov;20(11):1960.
7. Foy B.D., Kobylinski K.C., Chilson Foy
J.L., Blitvich B.J., Travassos da Rosa A,
Haddow AD. 2011. Probable non-vector-
borne transmission of Zika virus, Colorado,
USA. Emerg Infect Dis. May;17(5):880-2.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
19
8. Fonseca K., Meatherall B., Zarra D., Drebot
M., MacDonald J. and Pabbaraju K. 2014.
First case of Zika virus infection in a
returning Canadian traveler. Am J Trop Med
Hyg. Nov;91(5):1035-8.
9. Marchette N.J., Garcia R. and Rudnick A.
1969. Isolation of Zika virus from Aedes
aegypti mosquitoes in Malaysia. Am J Trop
Med Hyg. May;18(3):411-5.
10. McCrae A.W. and Kirya B.G. 1982. Yellow
fever and Zika virus epizootics and
enzootics in Uganda. Trans R Soc Trop Med
Hyg. 76(4):552-62.
11. Olson J.G., Ksiazek T.G., Suhandiman and
Triwibowo.1981. Zika virus, a cause of
fever in Central Java, Indonesia. Trans R
Soc Trop Med Hyg. 75(3):389-93.
12. Hamel R., Dejarnac O., Wichit S.,
Ekchariyawat P., Neyret A., Natthanej L.,
Perera-Lecoin M., Surasombatpattana P.,
Talignani L., Thomas F., Cao-Lormeau
V.M., Choumet V., Briant L., Després P.,
Amara A., Yssel H. and Missé D. 2015.
Biology of Zika virus infection in human
skin cells. J. Virol JVI. 00354-15.
doi:10.1128/JVI.00354-15
13. Perera-Lecoin M., Meertens L., Carnec X.
and Amara A. 2014. Flavivirus Entry
Receptors: An Update. Viruses 6: 69-88.
DOI: 10.3390/v6010069.
14. Chen Y., Maguire T., Hileman R.E., Fromm
J.R., Esko J.D., Linhardt R.J., 1997. Dengue
virus infectivity depends on envelope
protein binding to target cell heparan
sulfate. Nat Med. Aug;3(8):866-71.
15. Hilgard P. and Stockert R. 2000. Heparan
sulfate proteoglycans initiate dengue virus
infection of hepatocytes. Hepatology.
Nov;32(5):1069-77.
16. Zhang L. 2010. Glycosaminoglycan (GAG)
biosynthesis and GAG-binding proteins.
Prog Mol Biol Transl Sci.93:1-17.
17. Moore D.L., Causey O.R., Carey D.E.,
Reddy S., Cooke A.R. and Akinkugbe F.M.,
1975. Arthropod-borne viral infections of
man in Nigeria, 1964-1970. Ann Trop Med
Parasitol. Mar;69(1):49-64.
18. Fagbami A.H. 1979. Zika virus infections in
Nigeria: virological and seroepidemiological
investigations in Oyo State. J Hyg (Lond).
Oct;83(2):213-9.
19. Hayes E.B. 2009. Zika virus outside Africa.
Emerg Infect Dis. Sep;15(9):1347-50.
20. Musso D., Roche C., Nhan T.X., Robin E.,
Teissier A., Cao-Lormeau V.M. 2015.
Detection of Zika virus in saliva. J Clin
Virol. Jul;68:53-5.
21. Cao-Lormeau VM, Roche C, Teissier A,
Robin E, Berry AL, Mallet HP, et al. Zika
virus, French polynesia, South pacific, 2013.
Emerg Infect Dis. 2014 Jun;20(6):1085-6.
22. Pan American Health Organization (PAHO)
/ World Health Organization (WHO). Zika
virus infection [Internet]. Washington:
Regional Office for the Americas of the
World Health Organization; 2015 [updated
7 May 2015; cited 2015 7 May 2015].
Available from:
http://www.paho.org/hq/index.php?option=c
om_docman&task=doc_view&Itemid=270
&gid=30075&lang=en.
23. Ministério da Saúde (Brazil). Situação
Epidemiológica / Dados, [Internet]. 2015
[updated 24 May 2015; cited 2015 25 May
2015]. Available from:
http://portalsaude.saude.gov.br/index.php/sit
uacaoepidemiologica-dados dengue-2.
24. Ministério da Saúde (Brazil). Confirmação
do Zika Vírus no Brasil, [Internet]. Brasília:
Ministério da Saúde (Brazil); 2015 [updated
14 May 2015; cited 2015 14 May 2015].
Available from:
http://portalsaude.saude.gov.br/index.php/ci
dadao/principal/agencia saude/17701-
confirmacao-do-zikavirus-no-brasil.
25. Lanciotti R.S., Kosoy O.L., Laven J.J.,
Velez J.O., Lambert A.J. and Johnson AJ,
2008. Genetic and serologic properties of
Zika virus associated with an epidemic, Yap
State, Micronesia, 2007. Emerg Infect Dis.
Aug;14(8):1232-9.
26. Faye O., Dupressoir A., Weidmann M.,
Ndiaye M. and Alpha Sall A. 2008. One-

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Brief Description of: Zika Virus
20
step RT-PCR for detection of Zika virus. J
Clin Virol. Sep;43(1):96-101.
27. Balm M.N., Lee C.K., Lee H.K., Chiu L.,
Koay E.S. and Tang J.W. 2012. A
diagnostic polymerase chain reaction assay
for Zika virus. J Med Virol. Sep;84(9):1501-
5.
28. Gourinat A.C., O'Connor O., Calvez E.,
Goarant C. and Dupont-Rouzeyrol M. 2015.
Detection of Zika virus in urine. Emerg
Infect Dis. Jan;21(1):84-6.
29. Tappe D. Rissland J., Gabriel M., Emmerich
P., Gunther S. and Held G. 2014. First case
of laboratory-confirmed Zika virus infection
imported into Europe, November 2013.
Euro Surveill. 19(4).
30. European Centre for Disease Prevention and
Control. 2015. Rapid risk assessment: Zika
virus infection outbreak, Brazil and the
Pacific region. Stockholm: ECDC.
31. Calisher C.H. 1994. Medically important
arboviruses of the United States and
Canada. Clin Microbiol Rev. Jan;7(1):89-
116.
32. White C.B. 1881 The Yellow Fever
Epidemic at New Orleans in 1878. Public
Health Pap Rep.7:201-4.
33. bloda Gessa J.A. and Figueredo Gonzalez
R. 1986. Application of environmental
management principles in the program for
eradication of Aedes (Stegomyia) aegypti
(Linneus, 1762) in the Republic of Cuba,
1984. Bull Pan Am Health Organ.
20(2):186-93.
34. Bloch H. 1973. Yellow Fever in city of New
York. Notes on epidemic of 1798. N Y State
J Med. Oct 15;73(20):2503-5.
35. Ng T.F., Willner D.L., Lim Y.W.,
Schmieder R., Chau B. and Nilsson C.,
2011. Broad surveys of DNA viral diversity
obtained through viral metagenomics of
mosquitoes. PLoS One. 6(6):e20579.
36. Cook S., Chung B.Y., Bass D., Moureau G.,
Tang S. and McAlister E. 2013. Novel virus
discovery and genome reconstruction from
field RNA samples reveals highly divergent
viruses in dipteran hosts. PLoS One.
8(11):e80720.
37. Conis E. 2010. Debating the health effects
of DDT: Thomas Jukes, Charles Wurster,
and the fate of an environmental pollutant.
Public Health Rep. Mar-Apr;125(2):337-42.
38. Special Programme for Research and
Training in Tropical Diseases., World
Health Organization. Dengue : guidelines
for diagnosis, treatment, prevention, and
control. New ed. Geneva: TDR : World
Health Organization; 2009.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
21
Mecanismos no químicos de
comunicación en insectos
Biol. Mara Ivonne Garza-Rodríguez
Universidad Autonoma de Nuevo León [email protected]
Resumen
El éxito de los insectos proviene de su capacidad para percibir e interpretar su entorno. Los insectos
pueden identificar y responder de forma selectiva a las señales de un entorno heterogéneo. Como los
insectos son generalmente de corto ciclo de vida, el comportamiento y condición reproductiva deben estar
sincronizados así que este es uno de los procesos en los que tiene participación la comunicación. La
mayoría de los insectos tienen algo de visión, y muchos poseen sistemas visuales altamente desarrollados,
siendo el ojo compuesto el más especializado, también presentándose en algunos estructuras como los
ocelos y stematas. Los estímulos agrupados en la mecanorrecepción son aquellos asociados con la
distorsión causada por movimiento mecánico como resultado del propio medio ambiente, la relación del
insecto con el medio ambiente o las fuerzas internas derivadas de los músculos. En conclusión los insectos
tienen en la comunicación la forma fundamental de percibir estímulos que los ayudan a responder de una
forma apropiada de acuerdo a sus necesidades biológicas.
Abstract
The success of insects comes from their ability to perceive and interpret their environment. Insects
can identify and selectively respond to signals form a heterogeneous environment. Because they are
generally short life cycle, behavior and reproductive condition must be synchronized so this is one of the
processes in which communication is involvement. Most insects have some vision, and many have highly
developed visual system, the eye being the most specialized compound, also appearing in some structures
such as ocelli and stemmatas. Stimuli grouped mechanoreception are those associated with the distortion
caused by mechanical movement as a result of the environment itself, the relationship of the insect to the
environment or the internal forces arising from muscles. In conclusion insects have in communicating the
fundamental way of perceiving stimuli that help them respond in an appropriate way according to their
biological needs.
Palabras clave: comunicación, reproducción, recepción, acústica, visual

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
22
1. Introducción
Los insectos son importantes miembros de
las comunidades bióticas, por dos razones:
Existen una gran cantidad de ellos y hay muchos
tipos diferentes. La abundancia promedio de
cualquier especie es simplemente una función de
lo pequeño que es, qué tan grande es su fuente de
recursos y cuántos enemigos se aprovechan de
ella. Pero los insectos son inusuales entre los
organismos en la abundancia de especies. De
acuerdo con la teoría ecológica convencional,
cada especie ocupa un nicho único, una manera
única de vivir que funciona mejor que cualquier
otra especie y que se define por el lugar donde se
vive y lo que consume. En algunas comunidades
de especies de insectos están tan apretadas que
ampliamente se solapan nichos, lo que resulta en
una competencia, pero la separación y la
inestabilidad ambiental previenen la exclusión de
un competidor por otro (1).
El éxito de los insectos proviene, al menos
en parte, de su capacidad para percibir e
interpretar su entorno. Los insectos pueden
identificar y responder de forma selectiva a las
señales de un entorno heterogéneo. Pueden
diferenciar entre los hospederos, tanto plantas
como animales, y distinguir entre los muchos
factores microclimáticos, tales como las
variaciones de humedad, temperatura y flujo de
aire. La complejidad sensorial permite tanto
comportamientos simples como complejos en los
insectos (2).
La comunicación animal no sólo es
fascinante, su estudio también ha permitido
importantes avances en nuestra comprensión
general de los sistemas motores y sensoriales, la
evolución y la especiación. Un atractivo
importante del estudio de la comunicación es que
un investigador puede cuantificar que tan
biológicamente importante puede ser la
información codificada, en particular las
propiedades físicas de una señal y luego
experimentalmente determinar si los propios
animales utilizan esta información.
Para algunos sistemas, las consecuencias
selectivas de las decisiones de comportamiento
basadas en dicha información también se pueden
cuantificar (3).
Debido a que gran actividad de los insectos
es nocturna, la orientación y la navegación no se
basan únicamente en las señales visuales
convencionales, en muchas especies activas
durante la noche, olores y sonidos juegan un
papel importante en la comunicación. La serie de
la información sensorial utilizada por insectos
difiere de la de los humanos. Dependemos
mucho de la información visual y aunque
muchos insectos tienen una visión bien
desarrollada, la mayoría hacen un mayor uso del
olfato y el oído de lo que lo hacen los humanos.
El insecto está aislado de su entorno externo
por una barrera cuticular relativamente
inflexible, insensible e impermeable. La
respuesta al enigma de cómo el insecto blindado
puede percibir su entorno más cercano se
encuentra en las modificaciones cuticulares
frecuentes y abundantes que detectan los
estímulos externos. Órganos de los sentidos
sobresalen de la cutícula o a veces se encuentran
dentro o debajo de ella. Las células
especializadas detectan los estímulos que pueden
ser categorizados como mecánicos, térmicos,
químicos, y visuales (2).
2. Comunicación en la reproducción.
La mayoría de los insectos son sexuales y
por lo tanto los machos y hembras adultos deben
estar presentes en el mismo tiempo y lugar para
que la reproducción tenga lugar. Como los
insectos son generalmente de corto ciclo de vida,
el comportamiento y condición reproductiva
deben estar sincronizados. Esto requiere
respuestas fisiológicas finamente sintonizadas y
complejas al ambiente externo. El cortejo puede
incluir representaciones visuales, sobre todo por
parte de los machos, incluyendo movimientos de
las piezas del cuerpo, tales como antenas,
pedúnculos, las alas y movimientos ritualizados.
Sistemas de cortejo auditivo y sistemas de
reconocimiento del apareamiento son comunes
en muchos insectos por ejemplo en Hemiptera,
Orthoptera y Plecoptera (2).
2.1 Canto de llamada del macho en
saltamontes
Las señales acústicas de los machos y la
información que transmiten a menudo son
determinantes críticos en la elección de la
hembra. Los saltamontes “bladder” (Bullacris

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
23
membracioides) es uno de los numerosos
taxones de ortópteros que utilizan el sonido
como base del cortejo y apareamiento en última
instancia. Se han examinado las respuestas
acústicas de las hembras a las llamadas de
reproducción de los machos (4).
A continuación se presentan oscilogramas
representando la amplitud de onda de los cantos
de advertencia emitidos por estos insectos,
mientras que el último muestra el sonido que la
hembra produce como respuesta al canto del
insecto del macho de su misma especie. Figura1
(4).
3. Comunicación visual
La mayoría de los insectos tienen algo de
visión, y muchos poseen sistemas visuales
altamente desarrollados. Los componentes
básicos que se necesitan para la visión son una
lente para enfocar la luz sobre los fotorreceptores
(células que contienen moléculas sensibles a la
luz) y un sistema nervioso lo suficientemente
complejo como para procesar la información
visual. La estructura fotorreceptora es el
rabdoma, que comprende varias células
adyacente retinula (o nervios) y que consta de
microvellosidades de empaquetamiento
compacto que contienen pigmento visual. La luz
que cae sobre el rabdoma cambia la
configuración del pigmento visual, lo que
provoca un cambio de potencial eléctrico a
través de la membrana celular.
3.1 Ojos compuestos
El órgano visual más sofisticado en insectos
es el ojo compuesto. Prácticamente todos los
adultos y ninfas tienen un par de ojos
compuestos prominentes, que a menudo cubren
cerca de 360 ° del espacio visual. El ojo
compuesto se basa en la repetición de muchas
unidades individuales llamadas omatidias (2).
Figura 2. Secciones longitudinales de los ojos simples: (a) sistema sencillo de una larva de lepidóptero (5);
(b) un ocelo adaptado a la luz ocelo medio de una langosta (6).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
24
Figura: Detalles del ojo compuesto de: (a) una
vista que muestra la disposición de la omatidia;
(b) una sola omatidia con una ampliación
transversal (7 y 8).
Los ojos compuestos son los órganos
visuales más evidentes y conocidos de insectos,
pero hay otros tres medios por los cuales un
insecto puede percibir la luz: detección dérmica,
stematas y ocelos.
3.2 Detección dérmica
En los insectos capaces de detectar la luz a
través de su superficie corporal, hay receptores
sensoriales por debajo de la cutícula del cuerpo,
pero sin sistema óptico con estructuras de
enfoque. La evidencia de esta capacidad de
respuesta general a la luz
proviene de la persistencia de las respuestas
fóticas después de cubrir todos los órganos
visuales, por ejemplo en las cucarachas y las
larvas de lepidópteros. Algunos insectos ciegas
que viven en cuevas, sin órganos visuales
reconocibles, responden a la luz, al igual que las
cucarachas decapitadas. En la mayoría de los
casos, las células sensibles y su relación con el
sistema nervioso central aún no se han
descubierto. Sin embargo, dentro del cerebro
mismo, los áfidos tienen células sensibles a la
luz que detectan cambios en la duración del día,
una señal ambiental que controla el modo de
reproducción (es decir, ya sea sexual o
partenogenética). El ajuste del reloj biológico se
basa en la capacidad de detectar el fotoperíodo
(2).
3.3 Stematas
Los únicos órganos visuales de los insectos
holometábolos larvarios son los stematas, a
veces llamados ocelos larvales. Estos órganos se
encuentran en la cabeza y varían de un único
punto pigmentado en cada lado hasta seis o siete
stematas más grandes, cada uno con numerosos
fotorreceptores y células nerviosas asociadas. En
el stemma más simple, una lente cuticular se
superpone a un cuerpo cristalino secretado por
varias células. La luz es enfocada por la lente en
un solo rabdoma. Cada punto de la stemma va en
una dirección diferente de modo que el insecto
ve sólo unos pocos puntos en el espacio de
acuerdo con el número de stematas (2).
3.4 Ocelos
Muchos insectos adultos, así como algunas
ninfas, tienen ocelos dorsales además de ojos
compuestos. Estos ocelos no están relacionados
embriológicamente al stemata. Normalmente, los
tres pequeños ocelos se encuentran en un
triángulo en la parte superior de la cabeza. La
cutícula que cubre un ocelo es transparente y
puede ser curva como una lente. Se recubre las
células epidérmicas transparentes, de manera que
la luz pasa a través de una retina extendida
compuesta de muchos rabdomas. Así pues, los
ocelos integran la luz sobre un campo visual
grande, de forma óptica y neurológicamente. Son
muy sensibles a la intensidad de la luz baja y los
cambios sutiles en la luz, pero no están
diseñados para alta resolución de visión. Ellos
parecen funcionar como "detectores de
horizonte" para el control de los movimientos de
balanceo y cabeceo en vuelo y para registrar los
cambios cíclicos en la intensidad de la luz que se
correlacionan con los ritmos de comportamiento
diurnos (2).
3.5 Producción de luz
Las representaciones visuales más
espectaculares de insectos involucran la
producción de luz o bioluminiscencia. El papel
principal de la emisión de luz se dice que es en la
señalización de cortejo. Esto implica la variación
específica en la especie de la duración, número y
tasa de destellos en un patrón, y la frecuencia de
repetición del patrón (2). El estudio del
mecanismo de emisión de luz en la luciérnaga
Photinus pyralis puede ser típicos de un

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
25
coleoptera luminiscente. La enzima luciferasa
oxida un sustrato, luciferina, en presencia de una
fuente de energía de trifosfato de adenosina
(ATP) y oxígeno, para producir oxiluciferina,
dióxido de carbono, y luz. La variación en la
liberación de ATP controla la velocidad del
destello, y las diferencias en el pH puede
permitir variación en la frecuencia (color) de la
luz emitida (2).
Los patrones de destello de machos de un
número de especies de luciérnaga Photinus
(Coleoptera: Lampyridae), cada uno de los
cuales genera un patrón distintivo de señales con
el fin de obtener una respuesta de hembras de la
misma especie (22).
4. Mecanorrecepción
Los estímulos agrupados aquí son aquellos
asociados con la distorsión causada por
movimiento mecánico como resultado del propio
medio ambiente, la relación del insecto con el
medio ambiente o las fuerzas internas derivadas
de los músculos. Los estímulos mecánicos
detectados incluyen el tacto, estiramiento
corporal y el estrés, la posición, la presión, la
gravedad y las vibraciones, incluidos los
cambios de presión del aire y del sustrato
involucrado en la transmisión del sonido y la
audición. El cuerpo de los insectos está revestido
con proyecciones cuticulares. Estas se llaman
microtriquios si muchos surgen de una célula o
pelos, cerdas, setas o macrotriquia sí son de
origen multicelular. Las sensilias táctiles son
estimuladas solamente durante el desplazamiento
real del pelo. La sensibilidad es variable, siendo
algunos tan sensibles que responden a las
vibraciones de las partículas de aire causadas por
el ruido (2).
Sección longitudinal de una sensila tricoidea
que muestra la disposición de las tres células
asociadas (21).
4.1 Sonido
El sonido es una fluctuación de la presión
transmitida en una forma de onda a través del
movimiento del aire o el sustrato, incluyendo el
agua. La audición permite también la detección
de los depredadores, como los murciélagos
insectívoros, que utilizan ultrasonido en la caza.
Probablemente cada especie de insecto detecta el
sonido dentro de uno o dos rangos de frecuencias
relativamente estrechas que se relacionan con
estas funciones. El método más común de
producción de sonido de los insectos es por
estridulación, en el que una parte del cuerpo
especializado, el rascador se frota contra otro.
(2).
La descendencia de muchas especies de
animales solicita alimentos de los padres usando
señales que lo piden, mientras que los padres
manipulan el comportamiento de la descendencia
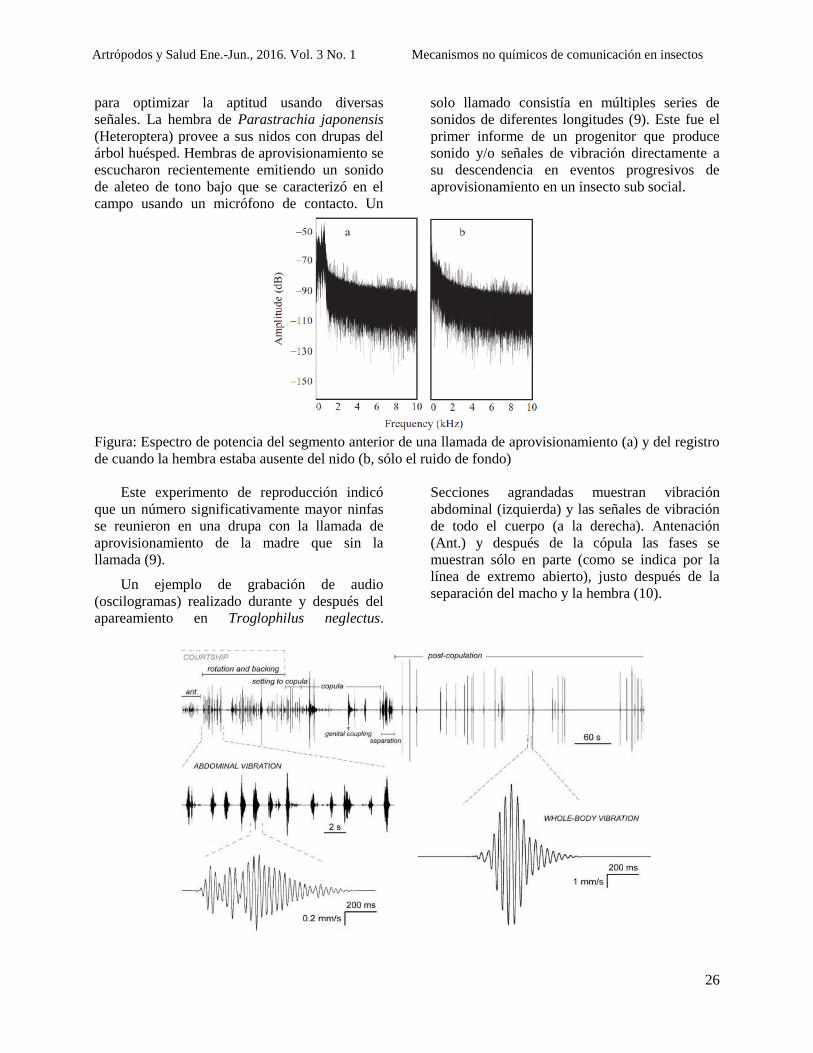
Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
26
para optimizar la aptitud usando diversas
señales. La hembra de Parastrachia japonensis
(Heteroptera) provee a sus nidos con drupas del
árbol huésped. Hembras de aprovisionamiento se
escucharon recientemente emitiendo un sonido
de aleteo de tono bajo que se caracterizó en el
campo usando un micrófono de contacto. Un
solo llamado consistía en múltiples series de
sonidos de diferentes longitudes (9). Este fue el
primer informe de un progenitor que produce
sonido y/o señales de vibración directamente a
su descendencia en eventos progresivos de
aprovisionamiento en un insecto sub social.
Figura: Espectro de potencia del segmento anterior de una llamada de aprovisionamiento (a) y del registro
de cuando la hembra estaba ausente del nido (b, sólo el ruido de fondo)
Este experimento de reproducción indicó
que un número significativamente mayor ninfas
se reunieron en una drupa con la llamada de
aprovisionamiento de la madre que sin la
llamada (9).
Un ejemplo de grabación de audio
(oscilogramas) realizado durante y después del
apareamiento en Troglophilus neglectus.
Secciones agrandadas muestran vibración
abdominal (izquierda) y las señales de vibración
de todo el cuerpo (a la derecha). Antenación
(Ant.) y después de la cópula las fases se
muestran sólo en parte (como se indica por la
línea de extremo abierto), justo después de la
separación del macho y la hembra (10).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
27
4.2 Ultrasonido
Aunque la depredación que lleva a cabo el
murciélago de los insectos se produce en la
oscuridad de la noche, es evidente que una serie
de taxones de insectos pueden detectar los
ultrasonidos de los murciélagos y tomar una
acción evasiva adecuada. La respuesta
conductual al ultrasonido, es llamada la
respuesta de sobresalto acústico e involucra
contracciones musculares muy rápidas y
coordinadas. Esto conduce a reacciones como
"congelamiento", desviación impredecible en
vuelo o rápido cese de vuelo y caída hacia el
suelo. Hasta la fecha, los insectos pertenecientes
a cinco órdenes han demostrado ser capaces de
detectar y responder a los ultrasonidos: crisopas
(Neuroptera), escarabajos (Coleoptera), mantis
religiosa (Mantodea), polillas (Lepidoptera),
langostas, saltamontes y grillos (Orthoptera) (2).
4.3 Vibratoria
Muchos insectos pueden detectar
vibraciones transmitidas a través de un sustrato
en barreras sólido-aire o sólido-agua o a lo largo
de una superficie de agua-aire. La percepción de
las vibraciones del sustrato es particularmente
importante para los insectos que habitan en el
suelo, las especies especialmente nocturnas e
insectos sociales que viven en nidos oscuros (2).
Figura: En una investigación se observó que el comportamiento reproductivo de pentatómidos está
mediada por las señales de comunicación de diferentes modalidades, a continuación se observa un
oscilograma y espectro de frecuencia de un pulso de cantos vibratorios en Edessa meditabunda: (a)
canción macho 1 (MS1); (b) canción rival macho (MR); (c) canción macho 2 (MS2); (d) canción hembra 1
(FS1).
La comunicación vibratoria comienza con la
emisión de la primera canción del macho (MS1).
Esta canción fue producida cuando un macho
estaba solo en la arena, en presencia de una
hembra o como respuesta a una canción
femenina. Durante el cortejo, la MS1 puede
cambiar a MS2 o interrumpirla durante algunos
minutos. En respuesta a las hembras MS2 emite
su primera canción (FS1), y, después de la
emisión de algunos impulsos, la pareja comienza
a intercambiar FS1 y MS2 primero intercaladas y
más tarde con impulsos superpuestos (11).
5. Termorrecepción
Los insectos detectan variación en la
temperatura, como se ve por su comportamiento,
sin embargo, la función y la ubicación de los
receptores es poco conocida. Los más estudiados
detectan la temperatura con sus antenas: la
amputación conduce a una respuesta térmica
diferente de la de los insectos con antenas
intactas. En las hormigas cortadoras de hojas
(especies Atta), hay sensillas termosensibles que
se agrupan en el flagelómero antenal apical y
responden tanto a los cambios de temperatura del
aire como a radiaciones de calor (2).
La temperatura tiene efectos drásticos en las
propiedades básicas de las células nerviosas y
musculares, tales como la tasa de pico, la
velocidad de conducción, la integración
sináptica, etc. (12, 13, 14). Por lo tanto, es un
problema grave para los animales poiquilotermos
mantener sus funciones neuronales y motoras

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
28
que opere a pesar de los cambios rápidos en la
temperatura del ambiente (15). En la parte
efectora, los cambios de temperatura afectan
enormemente las velocidades de contracción de
los músculos y por lo tanto el rendimiento motor
(16, 17). En las vías aferentes, el procesamiento
de las señales sensoriales se ve influenciado por
los cambios de temperatura de una manera
similar (18, 19, 20).
Las diferencias en la temperatura del cuerpo
entre el emisor y el receptor pueden inducir un
desajuste entre los parámetros de las señales y
las propiedades del receptor, evitando de este
modo de reconocimiento de señal. Los
saltamontes, por ejemplo, producen patrones de
sonido específicos de cada especie, moviendo
rítmicamente sus patas traseras a través de sus
alas delanteras. Estos movimientos se hacen más
rápido a temperaturas elevadas, y por lo tanto las
subunidades de la señal se acortan y se repiten a
un ritmo mayor - cambios por un factor de dos y
más de longitud subunidad no son poco comunes
(23).
6. Conclusiones
La comunicación en los insectos se da
debido a la necesidad del organismo por
satisfacer una necesidad biológica ya sea
alimentarse, reproducirse, huir de sus
depredadores, entre otras. Al detectar estímulos
externos, el insecto tiene la capacidad de
reaccionar y preparar una respuesta, la
comunicación puede ser visual, auditiva,
vibratoria entre otras pero aunque cada uno de
los procesos se ha estudiado individualmente,
estos guardan una estrecha relación, debido a
que de acuerdo a la exigencia de cada especie,
serán los tipos de comunicación que realizará
dependiendo de su ambiente, la competencia que
se le presente, duración del ciclo de vida,
alimentación o depredadores. El éxito del insecto
dependerá de su habilidad para detectar estos
estímulos, responder y con esto realizar las
funciones vitales que necesite.
7. Literatura Citada
1.- Triplehorn, C.A. y N.F. Johnson. 2005.
Borror and DeLong’s Introduction to the
study of Insects. 7ma. Edición. Thomson
Brooks/Cole. 93-94.
2.- Gullan, P.J. y P.S. Cranston. 2010. The
insects: and outlife of entomology. 4ta
Edición. Ed. Wiley-Blackwell. Pp. 92-122.
3.- Gerhardt, H.C. y F. Huber. 2002. Acoustic
communication in insects and anourans:
common problems and diverse solutions.
The Universisty of Chicago Press. ISBN 0-
226-28832-3.
4.- Couldridge, V. C. K. y M. J. van Staaden.
2006. Female Preferences For Male Calling
Songs In The Bladder Grasshopper
Bullacris Membracioides. Behaviour.
Academic Search Complete. 1439-1456.
5.- Snodgrass, R.E. 1935. Principles of Insect
Morphology. McGraw-Hill, New York.
6.- Wilson, M. 1978. The functional organisation
of locust ocelli. Journal of Comparative
Physiology 124, 297–316.
7.- CSIRO.1970. The Insects of Australia.
Melbourne University Press, Carlton.
8.- Rossel, S. 1989. Polarization sensitivity in
compound eyes. In: Facets of Vision.
Springer-Verlag, Berlin. 298–316.
9.- Nomakuchi, S., T. Yanagi, N. Baba, A.
Takahira, M. Hironaka, L. Filippi y R.
Knell. 2012. Provisioning call by mothers of
a subsocial shield bug. Journal Of Zoology,
50-56.
10.- Stritih, N. y A. Čokl, 2012. Mating
Behaviour and Vibratory Signalling in Non-
Hearing Cave Crickets Reflect Primitive
Communication of Ensifera', Plos ONE.
Academic Search Complete. 1-10.
11.- Silva, C., R. Laumann, J. Cavalcante
Ferreira, M. Blassioli Moraes, M. Borges y
A. Čokl. 2012. Reproductive Biology,
Mating Behavior, and Vibratory
Communication of the Brown-Winged Stink
Bug, Edessa meditabunda (Heteroptera:
Pentatomidae). Psyche: A Journal Of
Entomology, Academic Search Complete. 1-
9.
12.- Burrows, M. 1989. Effect of temperature on
a central synapse between identified motor
neurons in the locust. J Comp Physiol A
165:687–695.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Mecanismos no químicos de comunicación en insectos
29
13.- Hille, B. 1992. Ionic channels of excitable
membranes. Sinauer, Sunderland.
14.- Janssen, R. 1992. Thermal influences on
nervous system function. Neurosci
Biobehav Rev 16:399–413.
15.- Montgomery, J.C. y J.A. Macdonald. 1990.
Effects of temperature on nervous system:
implications for behavioural performance.
Am J Physiol 28:R191-R196 .
16.- Josephson R.K. 1981. Temperature and
mechanical performance of insect muscle.
In: Heinrich B (ed) Insect thermoregulation.
Wiley, New York. 20–44
17.- Full R.J. y A. Tullis. 1990. Capacity for
sustained terrestrial locomotion in an insect:
energetics, thermal dependence, and
kinematics. J Comp Physiol B 160:573–581
18.- Kodavová, B. 1996. Resolution of
pheromone pulses in receptor cells of
Antheraea polyphemus at different
temperatures. J Comp Physiol A 179:301–
310.
19.- Warzecha A.K., W. Horstmann y M.
Egelhaaf. 1999. Temperature dependence of
neuronal performance in the motion
pathway of the blowfly Calliphora
erythrocephala. J Exp Biol 202: 3161–3170.
20.- Tatler B., D.C. O’Carroll y S.B. Laughlin.
2000. Temperature and the temporal
resolving power of fly photoreceptors. J
Comp Physiol A 186:399–407.
21.- Chapman, R.F. 1991. General anatomy and
function. In: The Insects of Australia, 2da
Edición. (CSIRO). Melbourne University
Press, Carlton. 33–67.
22.- Lloyd, J.E. 1966. Studies on the flash
communication system in Photinus fireflies.
University of Michigan Museum of
Zoology, Miscellaneous Publications 130,
1– 95.
23.- Helversen D. von y O. von Helversen. 1997.
Recognition of sex in the acoustic
communication of the grasshopper
Chorthippus biguttulus (Orthoptera,
Acrididae). J Comp Physiol A 180: 373–386

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
30
UNA AMENAZA AGRÍCOLA MUNDIAL: EL VIRUS DE LA
TRISTEZA DE LOS CÍTRICOS Y SU VECTOR EL PULGÓN
CAFÉ Toxoptera citricida.
Pedraza Muñoz Raymundo; Ponce Samaniego J. León; Quintanar Alvarado Eduardo A.; Valdés Mejorado
David; Villareal Ramírez Gerardo.
Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Ap. Postal 109-F, San Nicolás de los
Garza, NL 66450, México e-mail de contacto: [email protected]
RESUMEN:
El virus de la tristeza de los cítricos (CTV) es causante de la enfermedad denominada “tristeza de
los cítricos” llamada así debido a los efectos de decaimiento, clorosis y defoliación causados en este tipo
de plantas, ocasionando una de las enfermedades más destructivas en el cultivo de los cítricos. Ha sido
considerada como la virosis más importante y devastadora de la citricultura a escala mundial, llegando a
causar pérdidas muy importantes e incluso millonarias a los agricultores. Dicha enfermedad es provocada
por un virus de la familia Closteroviridae, género Closterovirus y es transmitido por injertos y por áfidos
(pulgones) en forma semipersistente. El vector más eficiente es el pulgón café, Toxoptera citricida, este es
un insecto diminuto que pertenece al orden Homoptera, este grupo se caracteriza por que sus integrantes
son fitófagos y al alimentarse de las plantas transmiten patógenos (principalmente virus) de importancia
fitosanitaria. El combate contra la enfermeda involucra principalmente el uso de material de siembra libre
del virus, y evitar la entrada de T. citricida en áreas donde no está presente ya que una vez que se
establece su combate es muy difícil. Actualmente existen en México y en el mundo una alta normatividad
para establecer los cultivos libres del patógeno y del vector, además que se trabaja arduamente con
métodos de control biológico como lo son el uso de depredadores naturales y hongos entomopatógenos.
INTRODUCCIÓN:
El virus de la tristeza (CTV) causante de la
“tristeza de los cítricos” tiene distribución
mundial y ocasiona una de las enfermedades más
destructivas en el cultivo de los cítricos (1). Ha
sido considerada como la virosis más importante
y devastadora de la citricultura a escala mundial
(2), llegando a causar pérdidas muy importantes
e incluso millonarias a los agricultores.
Dicha enfermedad afecta prácticamente a
todos los tipos de cítricos, pero principalmente al
naranjo, toronjo y lima, pertenecientes al género
Citrus (3); es provocada por un virus de la
familia Closteroviridae, género Closterovirus y
es transmitido por injerto (Figura 1 y 2) y por
áfidos en forma semipersistente. El vector más
eficiente es el pulgón café, Toxoptera citricida
(Figura 3).
Figura 1.- Injerto http://www.jardin-
undani.com/empelts/Xip-mallorquin.htm
Figura 2.- Pulgones transmisores del CTV
http://www.cambiodigital.com.mx/mosno.php?nota=8
0962&seccion=Principal

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
31
Figura 3.- El pulgón café Toxoptera citricida
http://biblioteca.inifap.gob.mx:8080/jspui/bitstream/h
andle/123456789/698/6.pdf?sequence=1
La enfermedad de la tristeza produce el
colapso y decaimiento de los cítricos mediante
desecación y marchitamiento más o menos
repentinos de sus hojas, seguidos por la muerte
del árbol o muerte descendente de sus ramas y
recuperación parcial de estas últimas, además de
mostrar picaduras en el tallo (3).
Aunque la enfermedad afecta a algunos
árboles aun cuando se desarrollen sobre sus
propias raíces (plántulas), la tristeza produce
daños más severos en árboles que se injertan o
geman en los patrones de otras especies, como
los árboles del naranjo dulce que se desarrollan
en los patrones del naranjo agrio. (3)
Los síntomas de la tristeza varían en los
diferentes hospedantes e incluso entre los
mismos hospedantes en caso de que se
desarrollen en diferentes patrones. Los síntomas
típicos de esta enfermedad se pueden dividir en
tres:
1. Tristeza: siendo éste el más importante,
donde ocurre un decaimiento rápido o lento;
el decaimiento se caracteriza por una
clorosis y defoliación de los árboles,
asociado a una severa pudrición de raicillas y
a una pérdida de material de reserva a nivel
del tejido cortical bajo la zona de unión del
injerto (Figura 4).
2. Acanaladuras en la madera: se caracteriza
por la presencia de estrías o acanaladuras
longitudinales a nivel del xilema con
proyecciones puntiagudas de la corteza, las
que calzan perfectamente con las cavidades
presentes en la madera. Ello conduce a una
pérdida de productividad de las plantas
afectadas.
3. Amarillez de plantas francas de pie o
“seedling yellows”: donde ocurre una
clorosis intensa y normalmente se da bajo
condiciones de invernadero. (1)
Estos síntomas siempre se asocian a un
menor vigor de los árboles, a un crecimiento
deficiente y al desarrollo de frutos pequeños y
deformes.
Figura 4.- Síntomas de la planta afectada http://articulos.infojardin.com/PLAGAS_Y_ENF/Enfermedades/virus-tristeza.htm
El combate involucra principalmente el uso
de material de siembra libre del virus, y evitar la
entrada de T. citricuda en áreas donde no está
presente ya que una vez que se establece su
combate es muy difícil. (3)
Características de los Closterovirus
Los closterovirus son conocidos como un
grupo viral que infectan plantas de importancia
económica y son productores de enfermedades
devastadoras como el amarillamiento de la
remolacha y la tristeza de los cítricos. Son
partículas filamentosas no envueltas (Figura 5),
es decir, sin membrana de lípidos y
excepcionalmente largo aproximadamente 1250-
2200 nm de longitud y de 10 a 13 nm de
diámetro (3).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
32
Figura 5.- Partículas filamentosas del CTV http://cicshare.dep.usal.es/avelino/Bacteriologia/Tema%2017%20C
losterovirus%20A.pdf
El cuerpo del virión es montado por la
principal proteína de la cápside (CP) y la cola de
la proteína de la cápside menor (CPM).
Pertenecen al grupo 4 de la clasificación de
Baltimore, es decir, poseen un RNA de cadena
sencilla con polaridad positiva (4).
Cuentan con genomas mono catenarios de
15.5 -19.3kb, en los que se distinguen dos grupos
de genes altamente conservados, con diferentes
estrategias de expresión.
El primer bloque incluye los genes
implicados en la replicación que codifican para
la replicasa viral (RdRp) y las posibles helicasas
(HEL) y metiltransferasas (MT) y para una o dos
proteasas similares a la papaína. Este bloque de
genes presenta dos estrategias de expresión, una
mediado por la síntesis de una poli proteína
(ORF 1a) y cambio en la pauta +1 de lectura
traduccional (ORF 1b). El procesamiento
proteolítico, catalizado por las proteasas virales,
genera las posibles HEL y MT, mientras que la
RdRp es generada mediante el cambio en la
lectura traduccional.
La expresión del segundo bloque de genes,
localizado en el extremo 3`del RNA genómico,
se realiza en forma de RNA´s subgenómicos, y
codifica para los genes exclusivos de esta familia
HSP70, CPd y CP y para las proteínas p&4, p6,
que probablemente están implicadas en el
movimiento célula a célula (4).
Los miembros de este género están
distribuidos mundialmente y su rango de
hospedantes es amplio, afectando a especies
cultivadas, tanto herbáceas como leñosas de
interés agrícola y ornamental (3).
Características del vector
Los áfidos son insectos diminutos de
aproximadamente 1/8 de pulgada de largo
(Figura 6).
Figura 6.- Áfidos debajo de una hoja http://academic.uprm.edu/ofarrill/HTMLobj-227/afidos.pdf
Los áfidos pertenecen al grupo de los
homópteros al igual que las escamas,
chicharritas y la mosquita blanca. Se parecen
mucho a los hemípteros, pero sus partes bucales,
que también forman un punzón, surgen desde la
parte posterior de la cabeza. Tienen gran
importancia en la agricultura, pues todas sus
especies son herbívoras y suelen transmitir
enfermedades virales (5).
Tienen el cuerpo blando y su parte posterior es
redondeada en forma de pera. Se caracterizan
porque en la parte posterior poseen dos
estructuras tubulares de color oscuro. A estas
estructuras se les llama cornículos (6). El color
varía desde tonos amarillosos hasta colores
oscuros dependiendo de las diferentes especies
de áfidos (1) (Figura 7). Normalmente, los áfidos
no tienen alas, pero las pueden desarrollar para
migrar a nuevas áreas a causa del hacinamiento
o la escasez de alimento. Estos insectos se
reproducen en grandes números en un tiempo
relativamente corto. Pueden completar su ciclo
de vida en aproximadamente 10 a 14 días, que
consta de tres etapas: huevo, ninfa y adulto (6).
Los áfidos se alimentan de las hojas, los
renuevos, las flores, los frutos, las ramas, los
tallos y las raíces de una gran diversidad de
plantas, árboles y arbustos.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
33
Figura 7.- Áfidos a través de un microscopio http://academic.uprm.edu/ofarrill/HTMLobj-227/afidos.pdf
Además del daño que causan por sí solos,
los áfidos transmiten virus que causan
enfermedades serias en las plantas (3). También,
estas plagas secretan una sustancia azucarada
tipo melaza que promueve el crecimiento de un
hongo negro conocido como fumagina o moho
de hollín. Este hongo no parasita las plantas, los
árboles ni los arbustos, pero les desluce su
apariencia y disminuye la acción fotosintética
del follaje. Las hormigas también se alimentan
de esta secreción azucarada y tienen una relación
simbiótica con los áfidos. Esta relación
simbiótica consiste en que las hormigas protegen
a los áfidos y los transportan de una planta a otra
a cambio de la secreción azucarada (6).
Situación Geográfica
La enfermedad es endémica en las
principales áreas citrícolas de Asia (China y
Japón), África del Sur y Australia y en muchas
de las zonas citrícolas de América (Argentina,
Brasil, Uruguay Venezuela). Se encuentra
ampliamente extendida en casi todo el mundo
(Figura 8).
El pulgón café de los cítricos se describió
originalmente de especímenes colectados en
Hawaii en 1907 por Kirkaldy y algunos autores
indican que es originario del este asiático
(Blackman y Eastop, 2000). Desde inicios de la
mitad del siglo XX su distribución abarcaba gran
parte de Asia, Nueva Zelanda, Australia, algunas
islas del Pacífico, y el sur del Sahara y
Madagascar en África. Actualmente el insecto se
encuentra distribuido en los cinco continentes (7)
Situación en México
De acuerdo con el reporte del SENASICA
de junio de 2012 (7), Toxoptera citricida se
encuentra distribuido en los estados de la
Península de Yucatán, Tabasco, Veracruz,
Chiapas, Puebla e Hidalgo y en algunos
municipios de Guerrero, Oaxaca, Morelos,
Querétaro, San Luis Potosí, Jalisco y Nayarit.
Figura 8.- Distribución mundial de Toxoptera
citricida http://www.cesaveson.com/fckeditor/editor/filemanager/connectors
/aspx/UserFiles/file/Pulgoncafe.pdf
El SENASICA a través de la Dirección
General de Sanidad Vegetal, coordina la
campaña contra el Virus Tristeza de los Cítricos
y su vector (Toxoptera citricida) en la cual la
principal acción a realizar es el control biológico
del pulgón café de los cítricos. (7).
Impacto económico
En todas estas zonas citrícolas (figura 8) el
patrón naranjo amargo desapareció hace años, ya
que las variedades injertadas sobre él decayeron
rápidamente, causando la muerte de unos 40
millones de árboles. En los países citados, el
CTV se dispersa muy fácilmente, ya que el
pulgón pardo de los cítricos (Toxoptera
citricida), está presente. Este pulgón es el vector
más eficaz de la enfermedad y además, causa por
si solo una importante plaga en los cítricos (8).
En otras zonas citrícolas importantes, los
aislados mayoritarios de CTV son comunes o
poco agresivos, pero el virus infectará en un
futuro próximo al 100% de los árboles en
España, Israel y Estados Unidos de América
(Florida y California).

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
34
El insecto se caracteriza por ser un eficiente
transmisor del Virus de la tristeza de los cítricos
(VTC) el cual en el mundo ha provocado la
muerte de más de 116 millones de árboles,
establecidos sobre naranjo agrio (Citrus
aurantium L.) y que se intensifica al interactuar
con el pulgón café de los cítricos (T. citricida) ya
que el insecto tiene la facultad de transmitir
específicamente las variantes más severas del
virus. Las regiones citrícolas de Brasil,
Argentina, España, Venezuela, EUA (California,
Florida) han sido las más afectadas (7).
Medidas Fitosanitarias
La SENASICA-DGSV ha enlistado una
serie de acciones emprendidas correspondientes
a la normatividad aplicable en material de
cítricos en general o específicamente aplicable al
CTV. Entre ellas se encuentran la NORMA
Oficial Mexicana NOM-079-FITO-2002,
Requisitos fitosanitarios para la producción y
movilización de material propagativo libre de
virus tristeza y otros patógenos asociados a
cítricos (7)
Regulatorias
Al tratarse de una plaga con alto potencial
de transmisión del VTC, se recomienda la pronta
acción en el momento de su detección en zonas
donde el virus ya está presente. En México, una
vez que se detecta y confirma la presencia del
pulgón café de los cítricos se lleva a cabo un
programa de Manejo Integrado de Plagas, éste
está descrito en la NOM-031-FITO-2000 e
incluye entre otras:
a) Confinamiento de la plaga en área afectada y
regulación de la movilización, mediante la
operación de puntos de verificación interna de
los cordones fitozoosanitarios o los interestatales
que sean necesarios.
b) Aplicación de control químico con productos
autorizados por la Comisión Intersecretarial para
el Control del Proceso y Uso de Plaguicidas,
Fertilizantes y Sustancias Tóxicas
(CICOPLAFEST).
c) Liberación de Harmonia axyridis u otros
organismos benéficos con efectividad biológica
comprobada en contra del pulgón café de los
cítricos.
d) Intensificación del muestreo y diagnóstico del
virus tristeza de los cítricos y del pulgón café de
los cítricos.
e) Eliminación de plantas positivas al virus
tristeza de los cítricos.
Protección
Control químico:
No existe una lista de insecticidas autorizados
por CICOPLAFEST para el combate específico
de Toxoptera citricida en México; no obstante,
existe una lista autorizada para Toxoptera
aurantii. Se recomienda leer las etiquetas de
cada producto, seguir las recomendaciones de
seguridad, no exceder las dosis indicadas,
aplicarlos oportunamente y respetar el intervalo
de seguridad. Adicionalmente, se recomienda
consultar a los técnicos de la Campaña contra
VTC, o técnicos de Sanidad Vegetal (7).
Control biológico:
Éste es uno de los pilares en la estrategia de
manejo integrado que recomienda el gobierno
federal, y está contenido en la NOM-031-FITO-
2000. A nivel mundial se han identificado al
menos 113 especies de enemigos naturales entre
parasitoides, depredadores y entomopatógenos
que atacan al pulgón café de los cítricos (Figura
9 y 10). En México se encuentra presentes, al
menos, 18 de ellos (Michaud, 1998; López-
Arroyo et al., 2008). En México, el control
biológico se ha enfocado al uso de depredadores,
principalmente Harmonia axyridis, y hongos
entomopatógenos.
Figura 9.- Colonia de T. citricida con masas de
huevos de la catarinita Cycloneda sanguinea.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
35
http://www.cesaveson.com/fckeditor/editor/filemanager/connectors
/aspx/UserFiles/file/Pulgoncafe.pdf
Figura 10.- Adulto Cycloneda sanguinea
depredando en colonias de T. citricida en brotes
de naranjo. http://www.cesaveson.com/fckeditor/editor/filemanager/connectors
/aspx/UserFiles/file/Pulgoncafe.pdf
Con respecto a los entomopatógenos se
conoce la presencia de, al menos, cinco especies
de hongos con potencial en el control de esta
plaga. De éstas Paecilomyces fumosoroseus es la
especie que se ha considerado con mayor
potencial pues se han alcanzado niveles de
mortalidad superiores al 90 % en campo
(Hernández-Torres et al., 2006) (Figura 11).
Figura 11.- Evaluación de Hongos
Entomopatógenos para el Control del Pulgón
Café de los Cítricos. http://biblioteca.inifap.gob.mx:8080/jspui/bitstream/handle/123456
789/698/6.pdf?sequence=1
Control legal:
México, al igual que la mayoría de los
países, tiene una legislación fitosanitaria
conformada por una serie de normas emergentes
u oficiales para impedir que lleguen al territorio
nacional plagas y/o enfermedades que pondrían
en riesgo su agricultura. En el caso del pulgón
café de los cítricos las medidas cuarentenarias
están definidas en la NOM-011-FITO-1995, por
la que se establece la cuarentena exterior contra
las plagas de los cítricos, y la NOM-007-FITO-
1995, por la que se establecen los requisitos
fitosanitarios para la importación de material
vegetal propagativo.
Bibliografía:
1.- Rocha-Peña, M.A., R. F. Lee, R. Lastra, C. L.
Niblett, F. M. OchoaCorona, S. M. Garnsey,
and R. K. Yokomi. 1995. Citrus tristeza
virus and its aphid vector Toxoptera
citricida. Threats to citrus production in
Caribbean and Central and North América.
Plant Disease 79: 437-445.
2.- Cambra, M., Moreno, P. 2000. Tristeza. Pags.
77-81. In: Enfermedades de los cítricos. P.
Moreno, N. Durán-Vila (eds.).
3.- Agrios GN, Fitopatologia 2da edición, 1999,
Editorial LIMUSA, págs. 718-721.
4.- Bar-Joseph M, Che X, Mawassi M, Gowda S,
Satyanarayana T, Ayllón M, Albiach-Martí
M, Garnsey SM, Dawson WO. The
Continuous Challenge of Citrus tristeza
virus Molecular Research. Fifteenth IOCV
Conference, 2002- Citrus Tristeza Virus.
5.- Barrientos Llosa Zaidett, Zoología general
Primera edición 2003, Editorial Universidad
Estatal a Distancia San José págs. 285-290.
6.- O'Farrill Nieves Hipólito, Buitrago González
Misael, Las plagas del hogar y jardín 2005
Servicio de extensión agrícola Universidad
de Puerto Rico.
7.- http://www.senasica.gob.mx/2015
8.- Rodríguez Leyva Esteban; Lomeli Flores
Refugio J. Ficha Técnica Pulgón café de los
cítricos Toxoptera citricida (Kirkaldy),
Colegio de Postgraduados 2012 SAGARPA
Servicio nacional de sanidad, inocuidad y
calidad agroalimentaria
Silva-Vara Sergio, 2001. Distribution of
citrus tristeza virus in three commercial
groves in Nuevo Leon, México, p. 441-450.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Una amenaza agrícola mundial: El virus de la tristeza de
los cítricos y su vector el pulgón café Toxoptera citricida.
36
Arauz L.F., Fitopatología : Un Enfoque
Agroecológico, Editorial Universidad de
Costa Rica, 1998, pág. 81
Berlanga-Padilla, A. M. & J. I. López
Arroyo. 2006. Efecto de hongos
entomopatógenos en depredadores del
pulgón café de los cítricos (Homoptera:
Aphididae), p. 384-388. In: Memorias del
XXIX Congreso Nacional de Control
Biológico. Manzanillo, Colima, México.
Monografía de la Sociedad Española de
Fitopatología. N°2. Madrid. Ediciones
Mundi-Prensa. 213 pp
Mora Aguilera Gustavo Ficha técnica CTV
Virus de la Tristeza de los Cítricos Colegio
de Postgraduados SAGARPA Servicio
nacional de sanidad, inocuidad y calidad
agroalimentaria.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
37
TIFUS MURINO:
GENERALIDADES Caballero Treviño Yesica, Gómez Santillán Alejandro, Meléndez Villanueva Mayra Alejandra, Vargas López
Armando. [email protected].
Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas, San Nicolás de los Garza
INTRODUCCION
La participación en la historia de la
humanidad de las antropozoonosis, que abarcan
los grupos de fiebres manchadas y tifo, han
mermado la vida de más seres humanos que las
guerras.
La enfermedad del tifus murino ha originado
gran número de epidemias y ha estado presente
desde antes de la conquista, se dice que es
asociada a pobreza, hacinamiento, malas
condiciones higiénicas y guerras.
El tifus murino o endémico es causado por
Rickettsia typhi (Rickettsia mooseri), la cual se
transmite habitualmente por la pulga de la rata
(Xenopsylla cheopis), ocasionalmente puede ser
transmitida por la pulga del gato
(Ctenocephalides felis), al igual que por la pulga
del ratón (Leptopsyllia segnis). (1)
Las rickettsias son bacterias pequeñas,
cocobacilos Gram negativos, pleomorfos,
intracelulares obligados. Por esas dos
características, existieron dudas mucho tiempo
sobre si pertenecían a los virus o a las bacterias.
Son muy sensibles y raramente sobreviven fuera
del huésped (reservorio o vector), a excepción
de Coxiella burnetii (productora de la fiebre Q)
que es resistente a la desecación, al calor y la luz
solar. (2)
La incidencia de esta enfermedad es mayor
en los meses de verano y otoño cuando las
pulgas de las ratas son más activas. En la
mayoría de los casos se asocia a grandes
poblaciones de ratas. (1)
Etiología
El Tifus murino es causado por una bacteria
intracelular obligada del orden de los
Rickettsiales, Rickettsia typhi (11,16). Como otra
Rickettsias, Rickettsia typhi es pequeña (0.4 X
1.3 µm), gram negativa, intracelular obligado
que depende de los artrópodos hematófagos
(ejemplo pulgas y garrapatas) y mamíferos para
mantener su ciclo de vida. (4)
Rickettsia typhi existe en la naturaleza en
ciclos enzoóticos que involucran roedores y
ectoparásitos, es transmitida a los humanos
principalmente por Xenopsylla cheopis la pulga
de la rata oriental aunque otras especies de
pulgas, piojos y ácaros ocasionalmente se
pueden ver implicados. (7)
Las complicaciones son raras y la tasa de
letalidad para todas las edades es menor al uno
por ciento, sin embargo esta aumenta con la
edad. (5)
Reservorios y vectores
Es una zoonosis mantenida en ratas (rattus
rattus y rattus norvegicus) y transmitida a los
humanos a través de la infección con heces de la
pulga oriental de la rata (Xenopsylla cheopis
(Figura 1)) de la piel lastimada (12). Igualmente,
se ha sugerido que la pulga del gato
(Ctenocephalides felis) y las zarigüeyas podrían
actuar como vectores. La infección persiste en la
naturaleza por el ciclo rata – pulga – rata. (5)
Figura 1 Xenopsylla cheopis

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
38
Modo de transmisión
La infección ocurre en el momento de la
picadura de la pulga en el periodo de
rickettsemia. El agente se multiplica en el
intestino sin causar daño aparente a la pulga. El
vector elimina R. typhi por sus heces durante
toda su vida pero no por la saliva. No hay
transmisión de la infección de X. cheopis a su
progenie y la infección de nuevas generaciones
de pulgas se produce siempre por medio de sus
huéspedes. La infección en otras especies de
pulgas sigue iguales condiciones (5).
El periodo de incubación tiene un promedio
de doce días con un rango de variación entre 6 y
14 días. En el hombre, la difusión de la
enfermedad está determinada por el nivel de
enzootia entre las ratas y el grado de contacto
con estos animales y sus pulgas. La
susceptibilidad es general y la enfermedad
confiere inmunidad. Las pulgas permanecen
infectadas de por vida (hasta 1 año). (5)
La infección al hombre ocurre cuando la
pulga de la rata (u otra pulga como C. felis) lo
pica y defeca sobre su piel. Al rascarse el
hombre introduce la materia fecal contaminada a
través de la picadura u otra abrasión de la piel.
Es probable que el hombre también pueda
adquirir la infección por otras vías, tales como la
conjuntival o por inhalación. Estos modos de
trasmisión son sin embargo poco importantes. (5,
4, 9)
En la siguiente esquema se muestra el ciclo
de infección de Rickettsia typhi tanto en el
hombre como susreservorios (Figura 2).
Epidemiología
La distribución de tifus endémico es
mundial. Se localiza en zonas donde las
habitaciones humanas están infestadas de ratas y
ratones. La incidencia es mayor en los meses de
verano y otoño, cuando las pulgas de las ratas
son más activas. Si bien la enfermedad ocurría
sobre todo en áreas urbanas, especialmente en
edificios infestados de roedores, actualmente se
observa su extensión a áreas rurales.
En el siguiente mapa se muestran los países
afectados por el tifus murino en el siglo XX.

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
39
http://www.sci.unal.edu.co/scielo.php?script=sci_artt
ext&pid=S1657-
95342012000200011&lng=es&nrm=iso
En 1947, fue reportada R. typhi en la
República Mexicana en los estados de Guerrero,
Hidalgo, Jalisco, México, Michoacán, Nayarit,
Nuevo León, Oaxaca, Querétaro, Tamaulipas y
Zacatecas.
El tifo endémico ha estado presente en
México a lo largo de la historia, sobre todo en
los estados de Sinaloa, Coahuila, Guanajuato,
Oaxaca y Chiapas, sin embargo desde 1997 no se
reportan casos de tifus epidémico en México.
Respecto al tifus murino hubo 1,016 casos de
1988 a 2004; hasta el mes de septiembre del
2005 se habían presentado nueve casos. La
Secretaría de Salud reportó en Dr. Arroyo,
Nuevo León dos casos de rickettsiosis.
Fisiopatogenia
El periodo de incubación de las rickettsiosis
suele ser de 14 días tras la inoculación ya sea por
picadura del vector, el contacto con aerosoles de
material infectante o la contaminación
conjuntiva con heces de pulgas (Xenopsylla
cheopis), parásitos de ratas, perros y gatos
domésticos, liebres, ardillas voladoras y
zarigüeyas, entre otros mamíferos. (1,3)
En la patogenia de la enfermedad por
rickettsias interviene también el sistema inmune,
aunque de una forma todavía no bien conocida.
Al parecer, las rickettsias originan una
inmunosupresión leve del tipo de la
hipersensibilidad retardada, si bien no se sabe si
es un efecto directo de la infección o una
respuesta inmunomoduladora del huésped. (12)
Manifestaciones clínicas del tifus murino
La enfermedad tiene una sintomatología
similar a la del tifus epidémico transmitido por
piojos pero es de curso más corto y benigno. Se
presenta con fiebre, cefalalgia intensa y dolores
generalizados, a los 5 o 6 días del comienzo de la
fiebre aparece la erupción macular (Figura 3),
que se observa primero en el tronco y luego en
las extremidades, pero que no afecta las palmas
de las manos ni la planta de los pies o la cara, la
sintomatología incluye también tos, nerviosismo,
náuseas y vómitos. En el campo de refugiados en
Tailandia, los síntomas principales fueron fiebre
persistente, cefalalgia retroorbital y mialgias.
También en los EUA se registraron 200 casos
que se presentaron en el sur de Texas en donde
la erupción cutánea se manifestó solo en 58.1%
de los pacientes y la náusea en 44.9%.
Las complicaciones son raras. En los
pacientes no tratados la convalecencia puede
extenderse por varios meses. La letalidad
aumenta con la edad; en los EUA actualmente es
inferior al 1% en todas las edades. (15)
Figura 3. Erupción macular en el tronco
Diagnostico
El diagnóstico del tifo epidémico, tifo murino y
fiebre maculosa, se basa en el cuadro clínico y
antecedentes de residencia o procedencia de
áreas endémicas con transmisión de la
enfermedad. La confirmación por medio de
inmunofluorescencia indirecta con un valor
mínimo de 1:64 es considerada el valor mínimo
para diagnóstico presuntivo de tifo epidémico.
Para el diagnóstico del tifo transmitido por
piojos se requiere un alto índice de sospecha en
las zonas endémicas teniendo en cuenta factores
como: datos clínicos, condiciones de clima frío,
el hacinamiento, la poca higiene personal y la

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
40
presencia de piojos. Por laboratorio se lo puede
diferenciar de la enfermedad de Brill-Zinsser por
la presencia de anticuerpos IgM.
Pruebas de Laboratorio
Anticuerpos inmunofluorescentes. No
diferencia entre el tifus epidémico y el murino,
salvo que se practique absorción diferencial con
el respectivo antígeno de Rickettsia antes de
hacer la prueba o se tipifique con un equipo
específico. La confirmación de casos se deberá
hacer por: Identificación de las Rickettsias en
frotis de sangre teñidos con alguna de las
siguientes técnicas: Giemsa, Macchiavello, Ruiz
Castañeda o Giménez.
Serología. Para IgG, tomando dos muestras
pareadas, una en la etapa aguda, y otra con dos
semanas de diferencia respecto a la primera. Un
incremento de cuatro veces o más en los títulos
obtenidos, confirma el diagnóstico. Las pruebas
de anticuerpos IgM, se tornan positivas al sexto
día después de iniciado el cuadro.
En la enfermedad de Brill-Zinsser, el anticuerpo
inicial es la IgG, a títulos altos >1:512.
Aislamiento de rickettsias por cultivo. Se
realiza de muestras provenientes de reservorios
humanos y vectores.
Esta técnica sólo se puede efectuar en
laboratorios especializados y de alta seguridad.
El diagnóstico de las fiebres manchadas al
inicio de la erupción es clínico y epidemiológico.
En el laboratorio puede lograrse el aislamiento
de la R. rickettsii en sangre y en biopsias
cutáneas por inmunofluorescencia directa,
técnicas histoquímica o la reacción en cadena de
la polimerasa, durante la fase aguda.
Habitualmente se detectan anticuerpos séricos
específicos de grupo en la fase de convalecencia
por hemoaglutinación indirecta (con títulos de
1:128), inmunofluorescencia indirecta (IFI),
aglutinación del latex con títulos de 1:64, y
fijación de complemento (FC) con valores de
1:16.
Algunas herramientas para el diagnostico
son la reacción de Weil- Felix, que determina de
forma cruzada los anticuerpos de rickettsias
midiendo anticuerpos para antígenos de Proteus
OX19, la serología, la inoculación en ratones
lactantes para después identificar después las
bacterias en frotis teñidos por giemsa, biopsia
con tinción de inmunoflorescencia, la reacción
en cadena de la polimerasa (PCR).
Tratamientos
Las rickettsias son resistentes a
cefalosporinas, aminoglucósidos y penicilinas;
los antibióticos con sulfas no son eficaces y
pueden exacerbar las fiebres manchadas. El
tratamiento con tetraciclina o cloramfenicol se
originó por la necesidad de las rickettsias de
sintetizar proteínas OmpB, responsables de la
virulencia de la bacteria y de la formación de su
membrana celular a partir del sitio de unión
Ku70, ligasa receptora para células huésped.
Prevención
El tifus murino puede ser prevenido
efectivamente, a través del control de las pulgas
en las mascotas especialmente en los gatos
domésticos.
Se debe realizar podado, con el fin de no
proveer guarida a roedores, zarigüeyas o gatos
salvajes extraviados. Se deben colocar mallas en
las ventanas para impedir la entrada de animales
a la casa. No se deben permitir fuentes de
alimento para los animales salvajes tales como
latas, desperdicios de alimento o alimento para
mascotas (4).
En la mayoría de los casos, la prevención
depende de una cultura de protección ante los
riesgos; la vestimenta de colores claros y
brillantes facilita la visibilidad de posibles
vectores, y los pantalones y botas, además de
repelentes contra insectos y el retiro e inspección
corporal tras una exposición, disminuyen el
riesgo de infección. (17)
Es importante desparasitar los contactos
donde ocurre la infección y considerar la dosis
única de doxiciclina (200 mg) para tratar tanto
estos como los residentes de áreas infectadas, así
como personal médico que atienda los casos. (6)
Conclusiones
La enfermedad del Tifus epidémico está
asociada fundamentalmente con condiciones de

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
41
precariedad y carencias de saneamiento
ambiental por eso la intervención adecuada es
prevenir las malas condiciones de higiene en los
hogares y en las personas.
En la enfermedad del tifus epidémico existe
poca información acerca de medidas
epidemiológicas especialmente de incidencia.
Esto es una dificultad dado que no se pueden
trazar líneas de base para medir las
intervenciones realizadas.
Los roedores junto con las pulgas tienen el
papel más importante en la transmisión de R.
typhi, por ello las acciones de prevención y
control deben estar encaminadas a prevenir la
infestación de los hogares mediante la
educación.
El cambio climático y el aumento de
temperatura y pluviosidad en nuestro país puede
incidir en un incremento de la transmisión de
estos padecimientos a humanos desde
reservorios animales, como perros y gatos
domésticos, portadores de garrapatas y pulgas de
roedores, que tras un tiempo de sequía estarían
libres de predadores, o luego de lluvias
abundantes, aumentarían el alimento disponible.
(10)
Bibliografía
1.- Bernabeu-Wittel W, Segura-Porta F.
“Enfermedades producidas por rickettsias”.
Enf Infecc Microbiol Clin 2005; 23(3): 163-
172 .
2.- Blair P J, Schoeler G B, Moron C, Anaya E,
Caceda R, Céspedes M, et al. Evidence of
rickettsial and leptospira infections in
Andean Northern Peru. Am J Trop Med Hyg
2004; 70: 357-63.
3.- Boostrom A, Beier M, Macaluso J.
“Geographic association of Rickettsia felis
infected opossums with human murine
typhus, Texas”.Emerg Infec Dis2002;8:
549-554.
4.- Civen R, Ngo V. Murine typhus: an
unrecognized suburban vector borne
disease. Clin Infect Dis. 2008;46(6):913-8
5.- Colombia. Instituto Nacional de Salud.
Subdirección de Vigilancia y Control de
Salud Pública. Sistema Nacional de
Vigilancia de Eventos de Interés en Salud
Pública, Sivigila 2006:V.1. Protocolo de
vigilancia de tifus [CD– ROM]. Bogotá:
Instituto Nacional de Salud; 2006.
6.- Cowan G. “Rickettsial infections”. En Cook
G, Zumla A, Manson’s Tropical Diseases,
21ava. ed., cap. 50, Londres, Saunders,
2003, 891-906.
7.- Hidalgo M, Salguero E, de la Ossa A,
Sánchez R, Vega JF, Orejuela L, Valbuena
G. Murine typhus in Caldas, Colombia. Am
JTrop Med Hyg. 2008;78(2):321-2.
8.- La Colaboración Cochrane, Oxman Andrew
y Clarke Mike.Manual de Revisores 4.;
2000.
9.- Letaïef AO, Kaabia N, Chakroun M, Khalifa
M, Bouzouaia N, Jemni L. Clinical and
laboratory features of murine typhus in
central Tunisia: a report of seven cases. Int J
Infect Dis.2005; 9(6):331-4.
10.- López-Vélez R, Molina-Moreno R.
“Cambio climático en España y riesgo de
enfermedades infecciosas y parasitarias
transmisibles por artrópodos y garrapatas”.
RevEsp Salud Pública2005; 79: 177-190.
11.- Marshall GS, Stout GG, Jacobs RF, Schutze
GE, Paxton H, Buckingham SC,DeVincenzo
JP, Jackson MA, San Joaquin VH,Standaert
SM, Woods CR; Tick-Borne Infections in
Children Study Group. Antibodies reactive
to Rickettsia rickettsii among children living
in the southeast and south central regions of
the United States. Arch Pediatr Adolesc
Med. 2003; 157(5):443-8.
12. Morón et al. Tifus exantemático. Ministro de
Salud, Lima, 2001
13.- Mouffok N, Parola P, Raoult D. Murine
typhus, Algeria.Emerg Infect Dis.
2008;14(4):676-8.
14.- Nogueras MM, Cardeñosa N, Sanfeliu I,
Muñoz T, FontB,Segura F. Evidence of
infection in humans with Rickettsia

Artrópodos y Salud Ene.-Jun., 2016. Vol. 3 No. 1 Tifus Murino: Generalidades
42
typhiand Rickettsia felis in Catalonia in the
North east of Spain. Ann N YA cad Sci.
2006;1078:159-61.
15.-Pedro N. Acha y Boris Szyfres Zoonosis y
enfermedades transmisibles comunes al
hombre y a los animales, 3a edición. Vol. II
Clamidiosis, rickettsiosis y virosis pp 50-54
16.- Razzaq S, Schutze GE. Rocky Mountain
spotted fever: aphysician's challenge.
Pediatr Rev. 2005;26(4):125-30.
17.- Walker D. “Targetting rickettsia (clinical
implication ofbasic research)”.New Engl J
Med2006; 354: 1418-1420.

Instituciones Participantes



















