
¿PUEDE EL ANÁLISIS DE ENDEMICIDAD USARSE PARA...
12
Los trazos generalizados, empleados en los análisis panbiogeográficos, representan biotas ancestrales ampliamente distribuidas en el pasado y que fueron fragmentadas por eventos geofísicos (Morrone 2000). Actualmente existen varios métodos que permiten la identificación de los trazos generalizados. El primer método fue propuesto por Croizat (1958, 1964). Éste es conocido como método manual, en el cual los mapas de los trazos individuales se superponen para identificar aquellos que son congruentes en su topología, de tal manera que la superposición de dos o más trazos individuales da lugar a un trazo generalizado (Croizat 1958, 1964, Morrone 2004). Sin embargo, éste método genera imprecisiones, al depender de la apreciación del observador. Dada esta problemática, se desarrollaron otros métodos de índole cuantitativa, pero algunos de ellos no han sido usados ampliamente por varias razones (Morrone y Escalante 2009). Page (1987) propuso el método de matrices de conectividad e incidencia, que utiliza un algoritmo para calcular la congruencia entre trazos individuales (Espinosa-Organista et al. 2002). En este método, una vez delineados los trazos individuales, se construye una matriz de conectividad (n x n) por cada trazo (donde n= número de localidades o puntos) y una matriz ¿PUEDE EL ANÁLISIS DE ENDEMICIDAD USARSE PARA LA IDENTIFICACIÓN DE TRAZOS GENERALIZADOS? F OCUS A RTICLE 15 B IOGEOGRAFÍA 7 de conectividad global, en donde se suma la información de todos los trazos individuales para determinar si los trazos son homólogos o no a través de índices de conectividad. Después se construyen matrices de incidencia (i x j) por cada trazo individual (donde i= número de localidades o puntos y j= segmento del trazo involucrado). Éstas matrices identifican elementos comunes a los distintos trazos individuales. Craw propuso el método de compatibilidad de trazos en 1988. En este método se construye una matriz de presencia- ausencia de áreas por trazos individuales, que se analiza para buscar la compatibilidad entre los trazos. El conjunto de trazos compatibles entre sí representa un “clique”, el cual se emplea para construir el trazo generalizado (Craw 1988). El clique puede evaluarse estadísticamente, generando matrices al azar del mismo tamaño que la matriz real. Entre mayor sea el número de matrices obtenidas al azar que presenten los mismos cliques o cliques extras de mayor tamaño, menor validez estadística tendrá el trazo generalizado (Craw 1988, Espinosa et al. 2002). Es posible también utilizar el programa CLIQUE del paquete PHYLIP (Felsestein 1986) para llevar a cabo el método de compatibilidad de trazos (Morrone y Lopretto 1994). Leonardo Fernández-Badillo, Irene Goyenechea y Tania Escalante
Transcript of ¿PUEDE EL ANÁLISIS DE ENDEMICIDAD USARSE PARA...

Los trazos generalizados, empleados en los
análisis panbiogeográficos, representan biotas
ancestrales ampliamente distribuidas en el
pasado y que fueron fragmentadas por eventos
geofísicos (Morrone 2000). Actualmente existen
varios métodos que permiten la identificación de
los trazos generalizados.
El primer método fue propuesto por
Croizat (1958, 1964). Éste es conocido como
método manual, en el cual los mapas de los
trazos individuales se superponen para
identificar aquellos que son congruentes en su
topología, de tal manera que la superposición de
dos o más trazos individuales da lugar a un trazo
generalizado (Croizat 1958, 1964, Morrone
2004). Sin embargo, éste método genera
imprecisiones, al depender de la apreciación del
observador. Dada esta problemática, se
desarrollaron otros métodos de índole
cuantitativa, pero algunos de ellos no han sido
usados ampliamente por varias razones
(Morrone y Escalante 2009). Page (1987)
propuso el método de matrices de conectividad e
incidencia, que utiliza un algoritmo para calcular
la congruencia entre trazos individuales
(Espinosa-Organista et al. 2002). En este
método, una vez delineados los trazos
individuales, se construye una matriz de
conectividad (n x n) por cada trazo (donde n=
número de localidades o puntos) y una matriz
¿PUEDE EL ANÁLISIS DE ENDEMICIDAD USARSE PARA
LA IDENTIFICACIÓN DE TRAZOS GENERALIZADOS? FOCUS ARTICLE
15 B IOGEOGRAFÍA 7
de conectividad global, en donde se suma la
información de todos los trazos individuales para
determinar si los trazos son homólogos o no a
través de índices de conectividad. Después se
construyen matrices de incidencia (i x j) por
cada trazo individual (donde i= número de
localidades o puntos y j= segmento del trazo
involucrado). Éstas matrices identifican
elementos comunes a los distintos trazos
individuales.
Craw propuso el método de
compatibilidad de trazos en 1988. En este
método se construye una matriz de presencia-
ausencia de áreas por trazos individuales, que
se analiza para buscar la compatibilidad entre
los trazos. El conjunto de trazos compatibles
entre sí representa un “clique”, el cual se
emplea para construir el trazo generalizado
(Craw 1988). El clique puede evaluarse
estadísticamente, generando matrices al azar
del mismo tamaño que la matriz real. Entre
mayor sea el número de matrices obtenidas al
azar que presenten los mismos cliques o cliques
extras de mayor tamaño, menor validez
estadística tendrá el trazo generalizado (Craw
1988, Espinosa et al. 2002). Es posible también
utilizar el programa CLIQUE del paquete
PHYLIP (Felsestein 1986) para llevar a cabo el
método de compatibilidad de trazos (Morrone y
Lopretto 1994).
Leonardo Fernández-Badillo, Irene Goyenechea y Tania Escalante

16 B IOGEOGRAFÍA 7
Rojas-Parra (2007) propuso un método
para obtener trazos individuales de manera
computarizada, mediante el uso de una extensión
llamada Trazos2004, en el programa ArcView
GIS 3.2 (ESRI 1998). Trazos2004 se basa en el
algoritmo de Prim para obtener los trazos
individuales, a partir de los cuales dibuja un área
de influencia que permite identificar los trazos
generalizados (Rojas-Parra 2007). Por otro lado,
Liria (2008) propuso un método combinado
basado en Sistemas de Información Geográfica y
análisis espacial.
Otra programa que permite llevar a cabo
análisis de trazos es Croizat (Calvanti 2009). En
él es posible construir trazos individuales
mediante la implementación del algoritmo de
Prim, pero utiliza también el algoritmo de Bron y
Kerbosch (1973) para analizar la compatibilidad
de trazos, tal como se hace en el programa
CLIQUE (Felstestein 1986). En Croizat los trazos
son evaluados a partir de grafos aleatorios
regulares, con base en la propuesta de Craw
(1989) y Henderson (1989, 1991). Además, este
programa permite llevar a cabo análisis nodales
para cuantificar la importancia de los nodos.
MartiTracks (Echeverría-Londoño y
Miranda-Esquivel 2011) es otro programa que se
utiliza para realizar análisis de trazos mediante
un nuevo algoritmo que utiliza aproximaciones
geométricas para construir los trazos individuales
y para encontrar la congruencia entre ellos. Este
programa permite evaluar los trazos repetidos
mediante un análisis de similitud de trazos
individuales o generalizados y determinar cuáles
de ellos pertenecen al mismo trazo.
Uno de los métodos más utilizado para
la identificación de trazos generalizados es el
Análisis de Parsimonia de Endemismos o PAE
(Morrone y Escalante 2009), en el cual se
obtiene un cladograma de áreas donde los
trazos individuales que justifican el
agrupamiento de áreas son considerados como
trazos generalizados (Luna-Vega et al. 2000,
Morrone 2004). Por otra parte, una modificación
en la interpretación del PAE en
Panbiogeografía, es el PAE con eliminación
progresiva de caracteres llamado PAE-PCE
(Luna-Vega et al. 2000, García-Barros et al.
2002). El método básico se realiza de acuerdo
con Morrone (2004) y Echeverry y Morrone
(2010) construyendo los trazos individuales y el
mapeo de éstos trazos sobre un universo de
estudio elegido. Después se construye una
matriz de presencia-ausencia (r x c) donde las
filas (r) son las unidades geográficas y las
columnas (c) son los trazos individuales.
Además se añade un área externa con
sólo ausencias para enraizar el cladograma.
Las unidades geográficas que se utilizan son
las unidades predefinidas en las que se
subdividió el área de estudio. Éstas áreas
pueden ser polígonos regulares o irregulares,
naturales o arbitrarios (por ejemplo:
cuadrículas, provincias biogeográficas, o
países, entre otras). La matriz se analiza con un
algoritmo de parsimonia, y se obtiene el o los
cladogramas más parsimoniosos.

17 B IOGEOGRAFÍA 7
En el caso de obtener más de un
cladograma igualmente parsimonioso, se realiza
un consenso estricto de ellos. Posteriormente se
conectan las áreas incluidas en cada clado
sustentado por al menos dos trazos individuales
como parte de un mismo trazo generalizado.
Para realizar un PAE-PCE, se desconectan los
trazos individuales que sustentan los clados
obtenidos en un primer análisis para detectar
clados sustentados por otros, se realiza un
segundo análisis y el proceso se repite
sucesivamente hasta que ya no haya
sinapomorfías en los cladogramas. Finalmente,
se dibujan en un mapa los trazos generalizados.
Si se considera que un trazo generalizado
se forma a partir de la congruencia de dos o más
trazos individuales (Croizat 1958, 1964, Morrone
2004), y las áreas de endemismo se conforman a
partir de la congruencia en los rangos de
distribución de por lo menos dos especies
(Szumik et al. 2006), se hace evidente que para
ambos conceptos es necesario detectar la
congruencia distribucional en dos o más taxones.
Por esta razón se ha considerado que los trazos
generalizados son análogos a las áreas de
endemismo (García-Marmolejo et al. 2008) y por
tanto es posible entonces utilizar el método de
optimización o análisis de endemicidad
desarrollado para la búsqueda de áreas de
endemismo (Szumik et al. 2002, Szumik y
Goloboff 2004) como un método para la
detección de trazos generalizados.
PROPUESTA DEL MÉTODO
De acuerdo con la propuesta de Fernández-
Badillo (2013), para detectar a los trazos
generalizados, se plantea llevar a cabo un
análisis de endemicidad (en adelante AE),
mediante el uso del programa NDM/VNDM
(Goloboff 2002). Este programa incluye el
algoritmo de optimización basado en el concepto
de área de endemismo, mediante el cual es
posible evaluar cuántos y que tan endémicos son
los taxones de un área determinada, de tal
manera que aquellas áreas caracterizadas por al
menos dos taxones e índices de endemicidad
definidos por el investigador, serán seleccionadas
como áreas de endemismo (Szumik et al. 2006).
Esas áreas las consideraremos como unidades
equivalentes a los trazos generalizados.
En la selección de los parámetros de
NDM/VNDM, Fernández-Badillo (2013), sugiere
seleccionar en la opción de llenado, un valor de
cero para el caso de los registros inferidos y un
valor de 10 para los registros asumidos. Esto con
la finalidad de no ampliar de manera artificial la
distribución de los organismos.
En la ventana para llevar a cabo el AE en
NDM/VNDM, se propone que se puede modificar
ligeramente el valor mínimo de endemismo para
el área. En este caso el índice de endemismo (E)
del área puede ser de 1.5, valor por debajo de lo
establecido por defecto en el programa (E: 2.0).
El E igual a 2 indica que se requieren como
mínimo dos especies que se ajusten
perfectamente a un área (es decir, que cada una
posea un valor de índice de endemicidad de 1.0)

18 B IOGEOGRAFÍA 7
para considerar un área de endemismo. Sin
embargo, para la búsqueda de los trazos
generalizados se decidió no ser tan estrictos
(E:1.5), debido a que los trazos individuales que
forman un trazo generalizado deben ser
congruentes en la mayoría de su topología pero
no necesariamente idénticos. Por ejemplo, dos
especies con valores individuales de 0.75 serían
suficientes para conformar un posible patrón de
congruencia. De cualquier modo, al generalizar a
una cuadrícula en NDM, es necesario regresar a
los datos originales para verificar la superposición
de los trazos individuales.
En la opción de áreas superpuestas se
sugiere “retener áreas superpuestas si 40 % de
las especies son únicas”, sin embargo, este valor
puede modificarse y evaluar los distintos
resultados. Por ejemplo, es posible que un trazo
generalizado no tenga especies que se
compartan con ningún otro trazo, y entonces
puede seleccionarse este valor incluso por arriba
del 90% (ver Noguera-Urbano 2013).
A las áreas de endemismo resultantes
puede aplicárseles la opción de “área consenso”,
por ejemplo con similitud mínima del 60%,
aunque este valor también puede modificarse
(ver Aagesen et al. 2013). Las áreas consenso
obtenidas pueden ser consideradas como trazos
generalizados potenciales. Sin embargo, para
poder establecer qué áreas en realidad forman
un trazo generalizado, es necesario utilizar un
Sistema de Información Geográfica (SIG), por
ejemplo, ArcView (ESRI 1998), ArcGIS (ESRI
2008), QuantumGIS (QGIS Development Team
2014), GlobalMapper (Blue Marble Geographics
2014), DIVAGIS (Hijmans et al. 2002), entre
otros. En el SIG es necesario sobreponer los
trazos individuales de las especies que integran
cada una de las áreas de endemismo
identificadas en el análisis, de esta manera es
posible observar cuáles especies presentan un
patrón congruente de distribución que podría
considerarse como un trazo generalizado, tal
como se hace en el método manual propuesto
por Croizat para la identificación de trazos
generalizados.
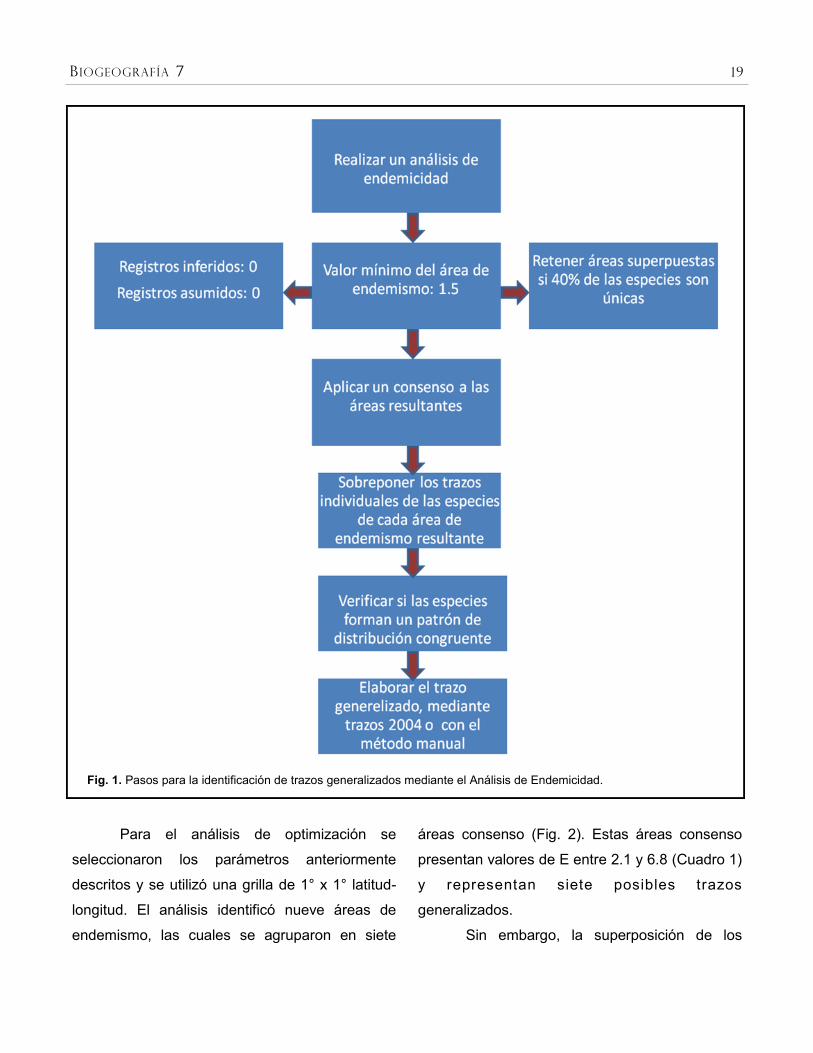
En la Figura 1 se presenta el esquema de
la secuencia de pasos para detectar trazos
generalizados por medio del análisis de
endemicidad.
ESTUDIO DE CASO: LA HERPETOFAUNA DE
LA PENÍNSULA DE BAJA CALIFORNIA,
MÉXICO
Para ejemplificar el método antes mencionado,
se utilizó parte de la base de datos utilizada en el
trabajo de Fernández-Badillo (2013), de la cual
se seleccionaron 2286 registros de 37 especies
de lagartijas y serpientes distribuidas en la
península de Baja California, en México. Dicha
información se obtuvo a partir de bases de datos
en línea así como de la base de datos
proporcionada por el Centro de Investigaciones
Biológicas del Noroeste A. C. (CIBNOR). Los
trazos individuales de las 37 especies se
obtuvieron con la herramienta automatizada
Trazos 2004 (Rojas-Parra 2007), implementada
en el programa ArcView 3.3 (ESRI 1998).
19 B IOGEOGRAFÍA 7
Fig. 1. Pasos para la identificación de trazos generalizados mediante el Análisis de Endemicidad.
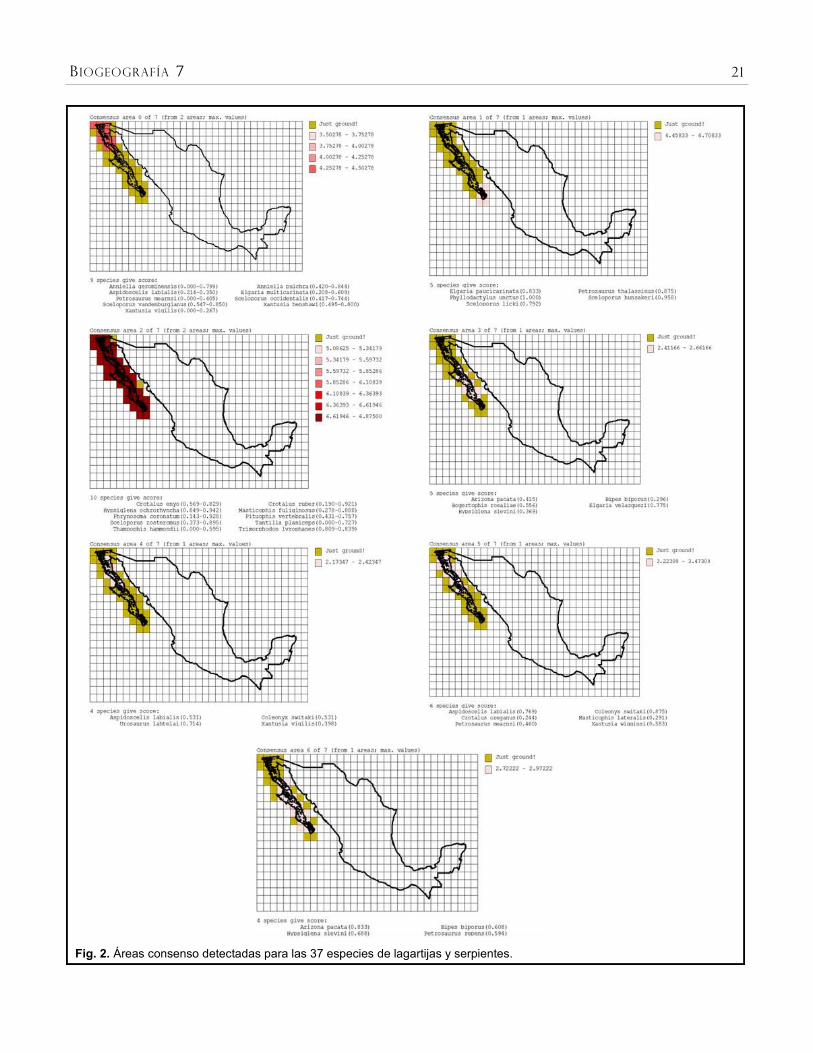
Para el análisis de optimización se
seleccionaron los parámetros anteriormente
descritos y se utilizó una grilla de 1° x 1° latitud-
longitud. El análisis identificó nueve áreas de
endemismo, las cuales se agruparon en siete
áreas consenso (Fig. 2). Estas áreas consenso
presentan valores de E entre 2.1 y 6.8 (Cuadro 1)
y representan siete posibles trazos
generalizados.
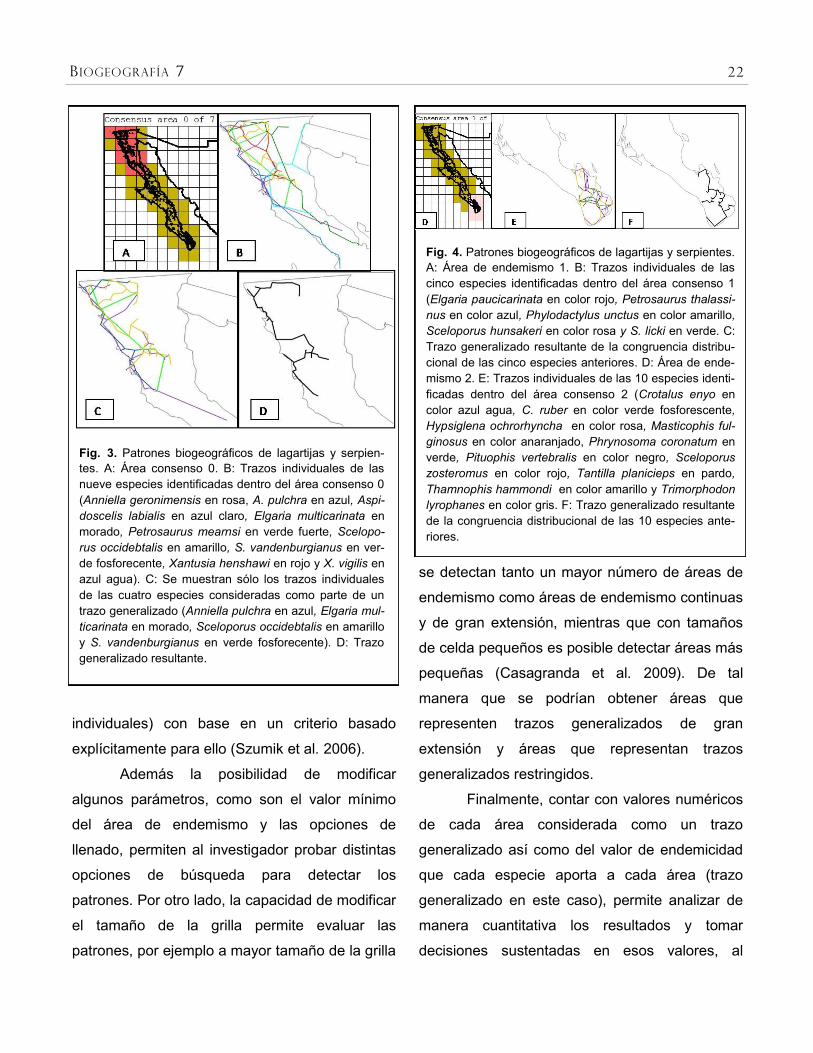
Sin embargo, la superposición de los

20 B IOGEOGRAFÍA 7
trazos individuales de 21 especies que integran
cada una de las áreas consenso obtenidas,
permitió la identificación de únicamente cuatro
trazos generalizados (Figs. 3, 4, 5 y 7). Estos
trazos generalizados corresponden a las áreas 0,
1, 2, 3 y 5.
Al sobreponer los trazos individuales de
las especies que integran las áreas 4 y 6, se
observó que no existía congruencia suficiente en
sus distribuciones para considerarlas como un
trazo generalizado (Figs. 6 y 8). Por ejemplo,
aunque las especies del área 4 se localizan en la
parte norte y centro de la Península de Baja
California, cada una muestra una distribución
distinta, lo cual puede deberse en parte a los
bajos puntajes individuales de las especies (Fig.
6). Lo mismo sucede con las especies del área 6,
localizadas en la parte centro y sur de la
Península (Fig. 8), por lo tanto no fue posible
ubicar un patrón congruente y por ello no fueron
consideradas como un trazo generalizado.
DISCUSIÓN
El uso del método de optimización para la
detección de trazos generalizados presenta
diferentes ventajas, como son la capacidad de
detectar las congruencias distribucionales entre
las especies (o en este caso los trazos
Área de
consenso
Número de
celdas
Valor de
Endemicidad Especies endémicas
0 12 3.502-4.502
Anniella geronimensis (0.0-0.799), A. pulchra (0.420-0.844), Aspidoscelis labialis (0.214-0.350), Elgaria multicarinata (0.288-0.609), Petrosaurus mearnsi (0.0-0.405), Sceloporus occidentalis (0.417-0.744), S. vandenburgianus (0.547-0.850), Xantusia henshawi (0.495-0.800), X. vigilis (0.0-0.267)
1 6 4.458-4.708 Elgaria paucicarinata (0.833), Petrosaurus thalassinus (0.875), Phyllodactylus unctus (1.000), Sceloporus hunsakeri (0.958), S. licki (0.792)
2 49 5.086-6.875
Crotalus enyo (0.569-0.8.29), C. ruber (0.190-0.921), Hypsiglena ochrorhyncha (0.849-0.942), Masticophis fuliginosus (0.278-0.888), Phrynosoma coronatum (0.143-0.928), Pituophis vertebralis (0.431-0.757), Sceloporus zosteromus (0.373-0.895), Tantilla planiceps (0.0-0.727), Thamnophis hammondi (0.0-0.595), Trimorphodon lyrophanes (0.809-0.839)
3 36 2.411-2.661 Arizona pacata (0.415), Bipes biporus (0.296), Bogertophis rosaliae (0.556), Elgaria velazquezi (0.775), Hypsiglena slevini (0.369)
4 13 2.173-2.423 Aspidoscelis labialis (0.531), Coleonyx switaki (0.531), Urosaurus lahtelai (0.714), Xantusia vigilis (0.398)
5 60 3.223-3.473 Aspidoscelis labialis (0.769), Coleonyx switaki (0.0875), Crotalus oreganus (0.244), Masticophis lateralis (0.291), Petrosaurus mearnsi (0.460), Xantusia wigginsi (0.583)
6 19 2.722-2.972 Arizona pacata (0.833), Bipes biporus (0.608), Hypsiglena slevini (0.688), Petrosaurus repens (0.594)
Cuadro 1. Áreas consenso resultantes del análisis de endemicidad para las 37 especies de lagartijas y serpientes de la península de Baja California. Se indica el número de celdas del área consenso, su valor de endemicidad y entre paréntesis el índice de endemicidad para cada especie.
21 B IOGEOGRAFÍA 7
Fig. 2. Áreas consenso detectadas para las 37 especies de lagartijas y serpientes.
22 B IOGEOGRAFÍA 7
individuales) con base en un criterio basado
explícitamente para ello (Szumik et al. 2006).
Además la posibilidad de modificar
algunos parámetros, como son el valor mínimo
del área de endemismo y las opciones de
llenado, permiten al investigador probar distintas
opciones de búsqueda para detectar los
patrones. Por otro lado, la capacidad de modificar
el tamaño de la grilla permite evaluar las
patrones, por ejemplo a mayor tamaño de la grilla
se detectan tanto un mayor número de áreas de
endemismo como áreas de endemismo continuas
y de gran extensión, mientras que con tamaños
de celda pequeños es posible detectar áreas más
pequeñas (Casagranda et al. 2009). De tal
manera que se podrían obtener áreas que
representen trazos generalizados de gran
extensión y áreas que representan trazos
generalizados restringidos.
Finalmente, contar con valores numéricos
de cada área considerada como un trazo
generalizado así como del valor de endemicidad
que cada especie aporta a cada área (trazo
generalizado en este caso), permite analizar de
manera cuantitativa los resultados y tomar
decisiones sustentadas en esos valores, al
Fig. 3. Patrones biogeográficos de lagartijas y serpien-
tes. A: Área consenso 0. B: Trazos individuales de las
nueve especies identificadas dentro del área consenso 0
(Anniella geronimensis en rosa, A. pulchra en azul, Aspi-
doscelis labialis en azul claro, Elgaria multicarinata en
morado, Petrosaurus mearnsi en verde fuerte, Scelopo-
rus occidebtalis en amarillo, S. vandenburgianus en ver-
de fosforecente, Xantusia henshawi en rojo y X. vigilis en
azul agua). C: Se muestran sólo los trazos individuales
de las cuatro especies consideradas como parte de un
trazo generalizado (Anniella pulchra en azul, Elgaria mul-
ticarinata en morado, Sceloporus occidebtalis en amarillo
y S. vandenburgianus en verde fosforecente). D: Trazo
generalizado resultante.
Fig. 4. Patrones biogeográficos de lagartijas y serpientes.
A: Área de endemismo 1. B: Trazos individuales de las
cinco especies identificadas dentro del área consenso 1
(Elgaria paucicarinata en color rojo, Petrosaurus thalassi-
nus en color azul, Phylodactylus unctus en color amarillo,
Sceloporus hunsakeri en color rosa y S. licki en verde. C:
Trazo generalizado resultante de la congruencia distribu-
cional de las cinco especies anteriores. D: Área de ende-
mismo 2. E: Trazos individuales de las 10 especies identi-
ficadas dentro del área consenso 2 (Crotalus enyo en
color azul agua, C. ruber en color verde fosforescente,
Hypsiglena ochrorhyncha en color rosa, Masticophis ful-
ginosus en color anaranjado, Phrynosoma coronatum en
verde, Pituophis vertebralis en color negro, Sceloporus
zosteromus en color rojo, Tantilla planicieps en pardo,
Thamnophis hammondi en color amarillo y Trimorphodon
lyrophanes en color gris. F: Trazo generalizado resultante
de la congruencia distribucional de las 10 especies ante-
riores.

23 B IOGEOGRAFÍA 7
considerar o no a una especie como parte de un
trazo. En general, los puntajes bajos (tanto de las
especies como de los trazos) permiten suponer
que la sobreposición no será completa y por lo
tanto el trazo generalizado pueda ser sólo un
artificio metodológico.
El uso de técnicas cada vez más precisas
para la identificación de los trazos, tanto
individuales como generalizados posibilita el
desarrollo de la panbiogeografía, cuyos principios
metodológicos han quedado establecidos desde
tiempo atrás. Por lo tanto, consideramos que en
la medida en que se avance en dicho
conocimiento, ésta será cada vez una disciplina
más robusta.
Fig. 5. Patrones biogeográficos de lagartijas y serpientes.
A: Área de endemismo 3. B: Trazos individuales de las
cinco especies identificadas dentro del área consenso 3
(Arizona pacata en color azul, Bipes biporus en color rojo,
Bogertophis rosaliae en color anaranjado, Elgaria velaz-
quezi en color verde e Hypsiglena slevini en color rosao-
lor gris. C: Se muestran sólo los trazos individuales de las
tres especies consideradas como parte de un trazo gene-
ralizado (Arizona pacata, Bogertophis rosaliae e Hypse-
glena slevini). D: Trazo generalizado resultante de la con-
gruencia distribucional de las tres especies anteriores.
Fig. 6. Patrones biogeográficos de lagartijas y serpientes.
.A: Área de endemismo 4. B: Trazos individuales de las
cuatro especies identificadas dentro del área consenso 4
(Aspidoscelis labialis en color azul, Coleonyx switaki en
color rojo, Urosaurus lahtelai en color amarillo y Xantusia
vigilis en color azul agua).
Fig. 7. Patrones biogeográficos de lagartijas y serpientes.
A: Área de endemismo 5. B: Trazos individuales de las
cinco especies identificadas dentro del área consenso 3
(Aspidoscelis labialis en color azul, Coleonyx switaki en
color rojo, Crotalus oreganus en color azul claro, Masti-
cophis lateralis en color rosa, Petrosaurus mearnsi en
color verde y Xantusia wigginsi en color amarillo. C: Tra-
zos individuales de las dos especies consideradas como
parte de un trazo generalizado (Crotalus oreganus y Mas-
ticophis lateralis). D: Trazo generalizado resultante de la
congruencia distribucional de las dos especies anteriores.

24 B IOGEOGRAFÍA 7
AGRADECIMIENTOS
A la Dra. Patricia Galina Tessaro del Centro de
Investigaciones Biológicas del Noreste S.C. por la
base de datos proporcionada. Al Dr. Juan José
Morrone por los comentarios durante la
realización de la tesis de maestría del primer
autor, de la cual surge este trabajo. Al CONACyT
por la beca de posgrado, número de becario
247535 y finalmente al proyecto FOMIX-
CONACyT Hidalgo 2012-01-191908.
REFERENCIAS
Aagesen L, Szumik C. Goloboff P. 2013.
Consensus in the search for areas of
endemism. Journal of Biogeography 40
(11): 2011-2016.
Blue Marble Geographics. 2014. Global Mapper
v15.2. Recuperado de h t tp: / /
www.bluemarblegeo.com/products/global-
mapper.php.
Bron C, Kerbosch J. 1973. Algorithm 457: Finding
all cliques of an undirected graph.
Communications of the Association for
Computing Machinery 16: 575-577.
Calvanti MJ. 2009. Croizat: A software package
f o r q u a n t i t a t i v e a n a l y s i s i n
panbiogeography. Biogeografica 4: 4-6.
Craw RC. 1988. Continuing the synthesis
between panbiogeography, phylogenetic
systematics and geology as ilustrated by
empirical studies on the biogeography of
New Zealand and the Chatham Islands.
Systematic Zoology 37: 291-310.
Croizat L. 1958. Panbiogeography. Vols. 1 y 2.
Caracas: Publicado por el autor.
Croizat L. 1964. Space, time, form: The biological
synthesis. Caracas: Publicado por el
autor.
Echeverría-Lodoño S. Miranda-Esquivel DR.
2011. MartiTracks: A geometrical aproach
for identifying geographical patterns of
d i s t r i b u t i o n . P l o S O N E 6 ( 4 ) :
e 1 8 4 6 0 . d o i : 1 0 . 1 3 7 1 /
journal.pone.0018460.
Echeverry A, Morrone JJ. 2010. Pasimony
Fig. 8. Patrones biogeográficos de lagartijas y serpien-
tes. A: Área de endemismo 7. B: Se muestran los trazos
individuales de las cuatro especies identificadas dentro
del área consenso 6 (Arizona pacata en color azul, Hyp-
siglena slevini en color rosa, Bipes biporus en color ver-
de y Petrosaurus repens en color rojo).

25 B IOGEOGRAFÍA 7
analys is of endemic i ty as a
panbiogeographical tool: an analysis of
Caribbean plant taxa. Biological Journal of
the Linnean Society 101: 961-976.
ESRI. 1998. ArcView GIS 3.2. Enviromental
Systems Research Institute. Redlands,
California.
ESRI. 2008. ArcGis 9.3. Enviromental Systems
Research Institute.California.
Espinosa-Organista D, Morrone JJ, Llorente-
Bousquets J, Flores Villela O. 2002.
Introducción al análisis de patrones en
biogeografía histórica. México, D. F. Las
Prensas de Ciencias, Facultad de
Ciencias, UNAM.
Felsestein J. 1986. Phylip user's manual. Seattle:
University of Washington.
Fernández-Badillo L. 2013. Patrones de
distribución de las lagartijas y las
serpientes en los desiertos mexicanos.
Tesis de Maestría. Pachuca: Universidad
Autónoma del Estado de Hidalgo.
García-Barros E, Guerra P, Luciáñez MJ, Cano
JM, Munguira ML , Moreno JC, Sainz H,
Sainz MJ, Simón JC. 2002. Parsimony
analysis of endemicity and its application
to animal and plant geographical
distributions in the Ibero-Balearic región
(western Mediterranean). Journal of
Biogeography 29: 109-124.
García-Marmolejo G, Escalante T, Morrone JJ.
2008. Establecimiento de propiedades
para la conservación de mamíferos
terrestres neotropicales de México.
Mastozoología Neotropical 15: 41-65.
Goloboff P. 2002. NDM and VNDM: Programs for
the identification of areas of endemism, v.
1.6. Recuperado de www.zmuc.dk/public/
phylogeny.
H e n d e r s o n I . 1 9 8 9 . Q u a n t i t a t i v e
panbiogeography: An investigation into
concepts and methods. New Zealand
Journal of Zoology 16: 495-510.
Henderson I. 1991. Biogeography without area?
Australian Systematic Botany 4: 59-71.
Hijmans RJ. Guarino L, Bussink C, Barrantes I,
Rojas E. 2002. DIVA-GIS, versión 2.
Sistema de Información Geográfica para
el Análisis de Datos de Biodiversidad.
Manual. Lima: International Potato Center.
Liria J. 2008. Sistemas de información geográfica
y análisis espaciales: un método
combinado para realizar estudios
panbiogeográficos. Revista Mexicana de
Biodiversidad 79: 281-284.
Luna-Vega I, Alcántara O, Morrone JJ, Espinosa
D. 2000. Track analysis and conservation
priorities in the cloud forest of Hidalgo,
Mexico. Diversity and Distributions 6: 137-
143.
Morrone JJ. 2000. Entre el escarnio y el encomio:
León Croizat y la Panbiogeografía.
Interciencia 25(1): 41-47.
Morrone JJ. 2004. Homología Biogeográfica: Las
coordenadas espaciales de la vida.
Cuadernos 37. Instituto de Biología.
México, D. F. Universidad Nacional
Autónoma de México.

26 B IOGEOGRAFÍA 7
Morrone JJ, Lopretto EC. 1994. Distributional
patterns of freshwater Decapoda
(Crustacea: Malacostraca) in southern
South America: a panbiogeographic
approach. Journal of Biogeography 21: 97
-109.
Morrone J, Escalante T. 2009. Panbiogeografía y
biogeografía cladística: Enfoques
complementarios para entender la
evolución biótica. En: Morrone JJ, Magaña
P. (Eds.), Evolución biológica: Una visión
actualizada desde la revista Ciencias.
México D. F. Universidad Nacional
Autónoma de México.
Noguera-Urbano E. 2013. Identificación de áreas
de endemismo de los mamíferos de la
región Neotropical utilizando el método de
optimización (NDM). Tesis de Maestría.
México, D. F. Universidad Nacional
Autónoma de México.
Page RDM. 1987. Graphs and generalized tracks:
quantifying Croizat´s panbiogeography.
Systematic Zoology 36: 1-17.
QGIS Development Team. 2014. QGIS
Geographic Information System. Open
Source Geospat ia l Foundat ion
Project. Recuperado de http://
qgis.osgeo.org
Rojas-Parra CA. 2007. Una herramienta
automatizada para realizar análisis
panbiogeográficos. Biogeografía 1: 31-33.
Szumik C, Cuezzo F, Goloboff PA., Chalup AE.
2002. An optimally criterion to determinate
areas of endemism. Systematic Biology
51: 806-818.
Szumik C, Goloboff PA. 2004. Areas of
endemism. An Improved Optimally
Criterion. Systematic Biology 53: 968-977.
Szumik C, Casagranda D, Roig-Juñet S. 2006.
Manual de NDM/VNDM: Programas para
la identificación de áreas de endemismo.
Instituto Argentino de Estudios
Filogenéticos 5: 1-26.
Leonardo Fernández-Badillo Instituto de Ciencias Básicas e Ingeniería Centro de Investigaciones Biológicas Universidad Autónoma del Estado de Hidalgo Ciudad del Conocimiento Km 4.5 Carretera Pa-chuca-Tulancingo, Mineral de la Reforma, 42181 Hidalgo, México. [email protected] Irene Goyenechea Instituto de Ciencias Básicas e Ingeniería Centro de Investigaciones Biológicas Universidad Autónoma del Estado de Hidalgo Ciudad del Conocimiento Km 4.5 Carretera Pa-chuca-Tulancingo, Mineral de la Reforma, 42181 Hidalgo, México. [email protected]. Tania Escalante Museo de Zoología “Alfonso L. Herrera” Departamento de Biología Evolutiva Facultad de Ciencias Universidad Nacional Autónoma de México Apdo. Postal 70-399, 04510 México, D. F.


















