
SECCIÓN 1 CAPÍTULO I, ANTECEDENTES …tesis.uson.mx/digital/tesis/docs/19885/Capitulo1.pdf ·...
48
2 SECCIÓN 1 CAPÍTULO I, ANTECEDENTES Membranas Biológicas En este trabajo se estudian algunas propiedades físicas de membranas biológicas. Por esta razón, se consideró importante empezar este manuscrito con una discusión sobre la estructura, composición química y funciones de las membranas biológicas. Las membranas biológicas, representadas en la figura 1, son estructuras que se encuentran en todos los niveles de organización celular. La membrana plasmática separa a la célula del medio exterior; los organelos a su vez están también rodeados de membranas o están constituidos por ellas. Muchas de las funciones básicas de la célula ocurren en la membrana o están ligadas de una u otra forma a estas estructuras. Entre estas funciones se puede mencionar: endocitosis, exocitosis, fusión celular, flujo de iones desde y hacia la célula, adhesión celular, reconocimiento específico de moléculas, conversión de energía, entre otros (Lodish et al., 2000). La composición de las membranas biológicas es altamente heterogénea, y se sabe que el soporte básico de las mismas es una doble capa de fosfolípido formada por agregación espontánea. A este soporte se agrega una gran cantidad de proteínas de diferentes tipos, polisacáridos, colesterol, esfingomielina. La bicapa misma está formada por fosfolípidos de diversa naturaleza química (zwiteriónicos y cargados) y de estructura distinta es decir diferentes tipo de cabeza polar, variación en la longitud y grado de insaturación de las cadenas hidrofóbicas (Berg et al., 2002). La composición característica de una membrana particular depende de la funciones que realiza. Así, las membranas de las células del cerebro poseen una composición diferente que las del hígado y esta a su vez que las de los pulmones (Cotterril et al., 2002).
Transcript of SECCIÓN 1 CAPÍTULO I, ANTECEDENTES …tesis.uson.mx/digital/tesis/docs/19885/Capitulo1.pdf ·...

2
SECCIÓN 1
CAPÍTULO I, ANTECEDENTES
Membranas Biológicas
En este trabajo se estudian algunas propiedades físicas de membranas
biológicas. Por esta razón, se consideró importante empezar este manuscrito con
una discusión sobre la estructura, composición química y funciones de las
membranas biológicas.
Las membranas biológicas, representadas en la figura 1, son estructuras
que se encuentran en todos los niveles de organización celular. La membrana
plasmática separa a la célula del medio exterior; los organelos a su vez están
también rodeados de membranas o están constituidos por ellas. Muchas de las
funciones básicas de la célula ocurren en la membrana o están ligadas de una u
otra forma a estas estructuras. Entre estas funciones se puede mencionar:
endocitosis, exocitosis, fusión celular, flujo de iones desde y hacia la célula,
adhesión celular, reconocimiento específico de moléculas, conversión de energía,
entre otros (Lodish et al., 2000).
La composición de las membranas biológicas es altamente heterogénea, y
se sabe que el soporte básico de las mismas es una doble capa de fosfolípido
formada por agregación espontánea. A este soporte se agrega una gran cantidad
de proteínas de diferentes tipos, polisacáridos, colesterol, esfingomielina. La
bicapa misma está formada por fosfolípidos de diversa naturaleza química
(zwiteriónicos y cargados) y de estructura distinta es decir diferentes tipo de
cabeza polar, variación en la longitud y grado de insaturación de las cadenas
hidrofóbicas (Berg et al., 2002).
La composición característica de una membrana particular depende de la
funciones que realiza. Así, las membranas de las células del cerebro poseen una
composición diferente que las del hígado y esta a su vez que las de los pulmones
(Cotterril et al., 2002).

3
Figura 1. Estructura general de la membrana biológica Fuente: http://www.ugr.es/~ajerez/proyecto/imagenes/t3-2.jpg
Función de la Membrana Celular Las membranas son la estructura celular más común tanto en animales
como en plantas y están involucradas en casi todos los aspectos de la actividad
celular, desde simples funciones mecánicas tales como motilidad, captación de
nutrientes y procesos bioquímicos altamente específicos tales como energía de
transducción, reconocimiento inmunológico, conducción nerviosa y biosíntesis
(Israelachvilli, 1992).
La composición de las membranas celulares es fundamental para los
procesos biológicos por lo que alteraciones importantes de la estructura de la
membrana pueden afectar el balance hídrico y el flujo iónico y por tanto todos los
procesos dentro de la célula. Por otra parte, las deficiencias específicas o
alteraciones de los lípidos que conforman la estructura básica de la membrana
celular (debido a mutaciones, por ejemplo), pueden conducir a situaciones
patológicas tales como la enfermedad de Tangier o la enfermedad de Niemann–
Pick tipo C.
Estructura de la Membrana Celular A pesar de la considerable variación en composición, todos los lípidos de
las membranas tienen una característica importante: son anfifílicos, es decir, una

4
parte de la molécula es insoluble en agua o hidrofóbica (la cadena
hidrocarbonada o ácido graso) y la otra parte es soluble en agua o hidrofílica (la
cabeza polar). La polaridad de la cabeza se debe a la presencia de cargas
eléctricas asociadas con el grupo fosfato y la base nitrogenada si está presente
y/o al gran número de grupos oxidrilos. Cuando moléculas anfifílicas como los
fosfolípidos son puestas en solución acuosa se autoarreglan de tal forma que la
parte hidrofílica o “cabeza polar” se pone en contacto con el agua, mientras las
“colas” hidrofóbicas se orientan en dirección al aire o se esconden del contacto
con el agua en diferentes agregados (Cotterill et al., 2002). Este proceso, que
explica la formación y la estructura básica de las membranas, se conoció
gradualmente a partir de las primeras décadas del siglo XX.
El primer modelo propuesto para explicar con una base estructural las
propiedades de la membrana plasmática fue dado por Davson y Danielli en 1935
y consistía en una bicapa lipídica en la cual se encontraban proteínas globulares
adsorbidas que podían atravesar la membrana o estar embebidas en ella, es
decir, con la parte hidrofóbica de la proteína dentro de la membrana y la parte
hidrofílica en el exterior de la bicapa lipídica. Versiones revisadas de este modelo
incluyen el enrollamiento de proteínas globulares en la superficie y la presencia de
poros de proteínas alineadas, las cuales proveen una ruta a través de la cual
pueden pasar el agua, iones o solutos polares. Sin embargo, esta visión fue
solamente una etapa en la formulación del modelo de “mosaico fluido” formulado
por Singer y Nicolson más de tres décadas después (Singer y Nicholson, 1972).
En este modelo se propuso que los lípidos de la bicapa se encuentran en fase de
líquido cristalino, lo cual constituye la matriz de las membranas biológicas. Por
otra parte se consideró la interacción de proteínas periféricas e integrales con la
membrana; en este modelo, la matriz de fosfolípido junto con las proteínas
embebidas forman una especie de solución viscosa bidimensional.
Así, los grupos polares de los lípidos y las proteínas están en contacto
directo con el medio acuoso mientras la porción no polar de ambas moléculas
está asociada a la parte central de la membrana. El modelo de mosaico fluido es
el más comúnmente aceptado y sirve como base para la discusión de las

5
propiedades físicas de las membranas, incluyendo las características de
estructura y composición de los lípidos que forman parte de ella, características
que generalmente tienen significado funcional.
La existencia de proteínas globulares embebidas en una bicapa lipídica
tiene un fuerte sustento experimental, especialmente basado en la técnica de
microscopía electrónica de criofractura. Sin embargo, a pesar de la aceptación del
modelo del mosaico fluido, no se descarta la existencia de otros arreglos,
permanentes o temporales: en organelos tales como mitocondrias y cloroplastos,
los lípidos pueden estar organizados dentro de micelas en vez de bicapas (Karp
et al., 1979).
Por otra parte, experimentos que determinan el coeficiente de difusión de
fosfolípido y/o de proteínas han mostrado que los constituyentes de una
membrana tienen gran movilidad. Los movimientos laterales de proteínas dentro
de la bicapa pudieran estar relacionados con alguna forma de ordenamiento de
las proteínas en cortos períodos de tiempo con propósitos específicos
funcionales. Finalmente, cabe mencionar que los movimientos posibles que una
molécula de fosfolípido puede efectuar en una membrana son: traslación, rotación
y el llamado “flip-flop” que es el movimiento por el cual una molécula puede pasar
de una capa de la membrana a la otra, es importante mencionar que esté tipo de
movilidad de las membranas es prácticamente inexistente o altamente improbable
(Cotterill et al., 2002).
Composición de las Membranas
Composición Química de las Membranas Las membranas están compuestas principalmente por lípidos, proteínas y
carbohidratos. Estos constituyentes, así como sus proporciones varían de
acuerdo al tipo de organismo, el tipo de membrana y el tipo de tejido en el cual se
encuentre la membrana; la composición de lípidos de algunos tipos de
membranas se muestra en la tabla 1. Los análisis químicos y las características
de permeabilidad indican que las membranas biológicas son ricas en lípidos tal
como se muestra en la tabla 1. En general, la proporción total de lípido es del

6
orden de 25 a 70 % en peso (Robinson et al., 1975). Los fosfoglicéridos son
generalmente la clase de lípidos dominante, especialmente en membranas de
células de origen animal, seguidos por las esfingomielinas y el colesterol. Los
fosfoglicéridos están representados principalmente por fosfatidilcolina (FC),
fofatidiletanolamina (FE), fosfatidilserina (FS), fosfatidilinositol (FI) y
lisofosfatidilcolina (LFC). Los más abundantes son los tres primeros mencionados,
siendo la FC y la FE fosfolípidos sin carga eléctrica neta, y la FS el fosfolípido
cargado eléctricamente más abundante (carga negativa) (Finean et al., 1978).
Tabla 1. Composición de la membrana
Membrana mg lípido/mg de
proteína Lípidos Principales
Fosfolípido/col. mol/mol
Plasmática 0.5 - 1.0 Col, FC, FS, FE, Efm, 0.4 – 1.0
Mielina 3.5 -4.0 FC, FE, Efm, Col 0.7 -1.2
Golgi 1.2 FC, FE, Efm, Col 0.45 – 0.5
Retículo endoplásmico 0.2 - 0.5 FC, FE, FI 0.06 – 0.1
Lisosomal 0.3 FC, FE, Efm, Col 0.5
Mitocondría 0.4 FC, FE, FI 0.1
Nuclear 0.2 – 0.6 FC, FE, FI
Cloroplastos 0.6 Col, FC
Bacterias Gram + 0.3 – 0.5 FE, DFG,
Micoplasma 0.3 Col, FG
Fuente: Neil F. Hadley, 1985 Lípidos de mayor importancia: FC Fosfatidilcolina, FS Fosfatidilserina, FE
Fosfatidiletanolamina, FI Fosfatidilinositol, Efm Esfingomielina, Col Colesterol, DFG Difosfatidilglicerol.
Otros componentes como las esfingomielinas y el colesterol se encuentran
abundantemente en membranas como la plasmática, la del complejo de Golgi y
en las de eritrocitos. En este último caso, el contenido de colesterol es
aproximadamente equimolar al de los fosfolípidos (fosfoglicéridos y
esfingomielinas).
La composición de lípidos en las membranas de las plantas es
básicamente similar a la descrita para células de animales, excepto por la
presencia de cantidades significativas de glicosilgliceroles en las membranas de
cloroplasto.

7
Por otra parte, la distribución de lípidos en las membranas biológicas es
asimétrica (Rothman et al., 1977). Los lípidos tales como fosfatidilcolina y las
esfingomielinas se encuentran principalmente en la cara exterior de la membrana
plasmática (o en su equivalente topológico de los organelos). En contraste, los
fosfolípidos que contienen aminas como la fosfatidiletanolamina y la
fosfatidilserina se encuentran preferentemente en el lado citoplásmico de la
membrana. Por poner un ejemplo, las membranas de los eritrocitos contienen
más del 75 % de su fosfatidilcolina en la superficie externa; en dichas
membranas, el 100 % de la fosfatidilserina se encuentra en la monocapa interna,
es decir, del lado del citoplasma (Daleke et al., 2003).
Lípidos que Componen la Membrana Biológica Los principales componentes de las membranas biológicas son lípidos y
proteínas. Los carbohidratos cuentan con alrededor del 10% del peso de la
membrana plasmática e invariablemente están unidos por enlaces covalentes a
lípidos o proteínas. Las cantidades relativas de lípidos y proteínas son muy
amplias como se muestra en la tabla 2, (Pind y Kuksis, 1986). Además de la
variación natural hay variación de la proporción proteína/lípido dependiendo del
método de aislamiento. Hay un número de proteínas las cuales están débilmente
unidas a la superficie de la membrana principalmente por fuerzas electrostáticas y
las cuales es posible extraer de la superficie de la membrana. Los cambios en la
fuerza iónica, pH, o la composición de la solución amortiguadora, (ejemplo, la
adición o eliminación de un quelante tal como el EDTA) son suficientes para
remover las proteínas de la membrana y modificar la relación de peso entre
proteína/lípido. La enorme diversidad de lípidos es una característica remarcada
de las membranas biológicas la razón de esta diversidad es aún materia de
discusión, a pesar de los avance en el conocimiento de la función de los lípidos en
la membrana.
En la tabla 2 se resume de forma respectiva la composición de lípidos del
plasma de mamíferos y membranas subcelulares. Los principales fosfolípidos que
componen las membranas son fosfatidilcolina, fosfatidiletanolamina, y
fosfatidilserina, este último cargado negativamente. Por otra parte se sabe que la

8
cadena hidrocarbonada tiene un número par de carbonos, en un intervalo de 14 a
22 átomos, con una predominancia de 16 y 18 átomos de carbono. La gran
mayoría tiene configuración cis. El grado de insaturación puede varias
considerablemente, los más comunes son de entre uno y cuatro insaturaciones
con la cadena insaturada ocupando usualmente la posición sn-2, en la molécula
de glicerol.
Tabla 2. Composición de lípidos de las membranas plasmática y subcelular Porcentaje de Fosfolípidos* Fosfolípidos
(μg/mg
proteína)
Colesterol
(μg/mg
proteína)
PC
PE
PS
PI
PA
CL
LGP**
SM
Membrana de glándula rectal 50.4 35.5 8.4 <1 - - - 5.7 389 n.d.
Membrana de bordes de vellosidades 33.3 35.6 7.4 8.2 1.2 n.d 4.1 10.3 190 50
Receptores colinérgicos de membrana 37 40.5 17 <1 - <1 <1 330(480)*** 135(190)***
Membrana plasmática 39 23 9 8 1 1 2 16 672 128
Mitocondria 40 35 1 5 - 18 1 1 175 3
Microsomas 58 22 2 10 1 1 11 1 374 14
Lisosomas 40 14 2 5 1 1 7 20 156 38
Membrana nuclear 55 13 3 10 2 4 3 3 500 38
Membrana de Golgi 50 20 6 12 <1 1 3 8 825 78
Retículo sarcoplásmico 72.7 13.5 1.8 8.7 <1 <1 - 1 603 12
Nota: Se pueden encontrar más datos de la composición de lípidos de la membrana de eritrocito
en la revisión de Pind y Kuksis 1986.
* Abreviación de fosfolípidos: PC, Fosfatidilcolina; PE, FosfatidilEtanolamina; PS, Fosfatidilserina;
PI, Fosfatidilinositol; PA, Ácido fosfatídico; CL, Cardiolipina; LGP, Lisoglicerofosfolípido; SM,
Esfingomielina
** Valores para LGP en exceso se deben tomar con reserva. Altos contenidos de LGP son
probablemente resultado de la degradación de fosfolípidos durante la preparación
*** Valores para dos diferentes preparaciones
Los ácidos grasos constituyentes de las moléculas de fosfolípidos también
varían dependiendo del tipo de membrana. Por ejemplo, los ácidos grasos de
membranas de células de hígado tienen cadenas largas y son más saturados en
comparación con los encontrados en el resto de las células (Nystrom et al., 1973).
La heterogeneidad de los lípidos es un requerimiento importante de modo
que la membrana pueda realizar adecuadamente su función. Los lípidos

9
participan activamente en un número de funciones especializadas; por poner unos
ejemplos:
1. Los lípidos de la membrana forman una bicapa cristalina líquida en la
cual las proteínas están embebidas y funcionando adecuadamente.
Algunos lípidos toman importancia por razones morfológicas. Además
pueden estabilizar regiones específicas de la membrana, por ejemplo
bicapas con alta curvatura o involucradas en la formación de enlaces
con otras membranas.
2. Lípidos específicos se acumulan en regiones determinadas de la bicapa
y puede ser requeridos para optimizar la actividad enzimática
3. Los lípidos participan en el metabolismo con funciones importantes,
otros lípidos pueden estar involucrados en funciones de regulación. Un
notable ejemplo es el fosfatidilinositol y su análogo fosforilado.
4. Existe una creciente evidencia de que los lípidos están involucrados en
algunas funciones especializadas tales como la formación de pequeñas
regiones en la membrana celular denominadas dominios. Tales
regiones están involucradas en procesos como el reconocimiento
celular o la activación de linfocitos T en nuestro sistema inmunológico.
Por otra parte los glicolípidos están implicados en funciones tales como
reconocimiento celular, diferenciación celular y crecimiento celular.
Principales Lípidos de las Membranas Las membranas celulares tienen una composición heterogénea y están
constituidas de manera compleja. Las moléculas que las conforman varían de
membrana a membrana dependiendo de la función que desempeñe la célula o el
organelo en que se encuentren. Los principales constituyentes de las membranas
biológicas son los lípidos, las proteínas y los carbohidratos. A su vez los lípidos
presentan una gran variedad, siendo los fosfolípidos los más importantes desde el
punto de vista estructural. En esta sección describimos las características de los
principales lípidos que constituyen las membranas biológicas con la finalidad de
comprender su importancia y conocer su estructura química.

10
Ácidos grasos de cadena larga Los ácidos grasos de cadena larga (AGCL) son constituyentes de todos los
lípidos esterificados y complejos, por ejemplo, de los glicerofosfolípidos y las
esfingomielinas. Los ácidos grasos de cadena larga están unidos por enlaces
ester o amida y son los lípidos más importantes que forman las membranas. Los
ácidos grasos libres representan un bajo porcentaje de las componentes de las
membranas. Por otra parte, los AGCL son ampliamente usados en la industria
como precursores de emulsificantea, aditivos de alimentos y detergentes. En
células eucariotas los AGCL tienen un número típico de átomos de carbono de
entre 14 y 24 y con cero hasta seis dobles enlaces como se muestra en la figura
2. Los dobles enlaces de las cadenas de ácidos grasos mono y poliinsaturados,
usualmente tienen configuración cis.
Ácido octadecanóico trans-9-ácido octadecanóico cis-9-ácido octadecanóico cis-9-cis-12-ácido octadecadienóico cis-9,12,15-ácido octadecatrienóico cis-6,9,12-ácido octadecatrienóico cis-5,8,11,14-ácido Eicosatetraenóico
Figura 2. Ácidos grasos de cadena larga, lípidos de gran importancia en la
formación de lípidos complejos constituyentes de las membranas biológicas.

11
Esteroides El sistema fundamental de anillos para todos los esteroides es el
ciclopentanofenantreno, cuya estructura se muestra en la figura 3.
Ciclopentanofenantreno Colesterol
Colate Cenodexicolate
β -Sitosterol Estigmasterol
Lanosterol Ergosterol
Hopano 29 -(1 ´,2 ´,3 ´,4 ´-tetrahidropentil )-hopano )
Bacteriohopanotetrol
22 -hidroxihopano o diplopterol
Tetrahimanol
Ciclopentanofenantreno Colesterol
Colato Cenodexicolato
β -Sitosterol Estigmasterol
Lanosterol Ergosterol
Hopano 29 -(1 ´,2 ´,3 ´,4 ´-tetrahidropentil )-hopano )
Bacteriohopanotetrol29 -(1 ´,2 ´,3 ´,4 ´-tetrahidropentil )-hopano )
Bacteriohopanotetrol
22 -hidroxihopano o diplopterol
Tetrahimanol
Esteroides
Hopanoides
Figura 3. Estructura química de Esteroides y Hopanoides

12
Más que esteroides son alcoholes a los que se acordó llamar esteroles. Como es
evidente en su estructura todos tienen el sistema de anillos pero difieren
considerablemente en su cadena lateral, sus características estructurales
periféricas, su estereoquímica y en su número de dobles enlaces en los anillos.
El colesterol se encuentra en la membrana plasmática de las células de
mamíferos aproximadamente en un 30% del total de los lípidos, y también en
lisosomas, endosomas, y el aparato de Golgi. El ergosterol es frecuentemente
encontrado en microorganismos eucarióticos tales como las levaduras.
En las décadas de los 80s y 90s fueron descubiertos una nueva clase de
compuestos penta cíclicos similares al esterol y se nombraron hopanoides. Estos
compuestos son ampliamente distribuidos en bacterias pero también son
encontradas en algunas plantas. Éste compuesto se encuentra en la membrana
bacterial y se ha propuesto que su función es la estabilización estructural de la
membrana, similar a los esteroles en eucariotas. Esteroles y hopaniodes son
moléculas rígidas y preferentemente anfipáticas. Los grupos anfifílicos están en el
lado opuesto de la molécula: esto es, en el grupo 3-hidroxilo en el esterol y en el
poliol en la cadena lateral de los hopaniodes.
Acilgliceroles Es importante mencionar que los acilgliceroles constituyen la estructura
básica de los fosfolípidos que forman parte de nuestro sistema experimental
(SOPC y SOPS). Como se mencionó anteriormente, los ácidos grasos libres se
unen por medio de un enlace éster a un glicerol, el cual a su vez se une al grupo
fosfato en las denominadas cabezas polares, resultando la estructura general de
un fosfolípido. A continuación, mencionaremos las características más
importantes y las estructuras químicas de estos compuestos.
Un gran número de lípidos complejos tienen como parte central de su
estructura el grupo glicerol. El glicerol tiene tres grupos reactivos OH y forman
esteres con ácidos grasos. Dependiendo del número de ácidos grasos que
reaccionen con el glicerol se obtienen monoacil, diacil o triacil gliceroles como se
muestra en la figura 4.

13
Los acilgliceroles particularmente los monoacil y los diacilgliceroles son
componentes minoritarios de las membranas biológicas. Los diacilgliceroles
tienen una función importante como parte fundamental de la estructura básica de
las membranas. También tienen funciones más específicas en los procesos
metabólicos; por ejemplo actúan como segundo mensajero en la señal de
transducción. Además, muchas sustancias biológicamente activas tales como
algunas hormonas de neurotransmisores reaccionan con receptores en la
membrana celular y provocan una respuesta hacia el interior de la célula, lo cual
es debido a la conformación de los acilgliceroles.
Esteres de colesterol El colesterol puede ser esterificado con ácidos grasos de cadena larga para
formar éster de colesterol figura 4.
Figura 4. Acil y alquilgliceroles y esteres de colesterol

14
Los cuales son mucho más hidrofóbicos que los compuestos de origen. Los
ésteres de colesterol son constituyentes de lipoproteínas del suero. Por ejemplo
las Lipoproteínas de baja densidad (LDL) son ricas en colesteroil éster. Además
los ésteres de colesterol se acumulan en ciertos desórdenes (Small et al., 1977),
tales como la enfermedad por almacenamiento de colesteril ester (Fredrickson
and Ferran, 1978), arterioesclerosis (Small and Shipley, 1974),
hipercolesterolemia (Fredrickson et al., 1978), y la enfermedad de Tangier. Los
esteres de colesterol son componentes menores de las membranas celulares.
Lípidos Complejos Como se ha mencionado, los lípidos complejos son las principales
moléculas constituyentes de las membranas celulares y tiene propiedades
estructurales diversas. Su variedad en cuanto a peso molecular, caga neta, grado
de insaturación en la cola hidrofóbica, etc., confiere propiedades específicas a las
membranas, dependiendo de su composición. A continuación se describirán las
características estructurales de los lípidos más importantes que componen las
membranas celulares; más específicamente, describiremos los lípidos que forman
parte de nuestro sistema experimental para la fabricación de membranas.
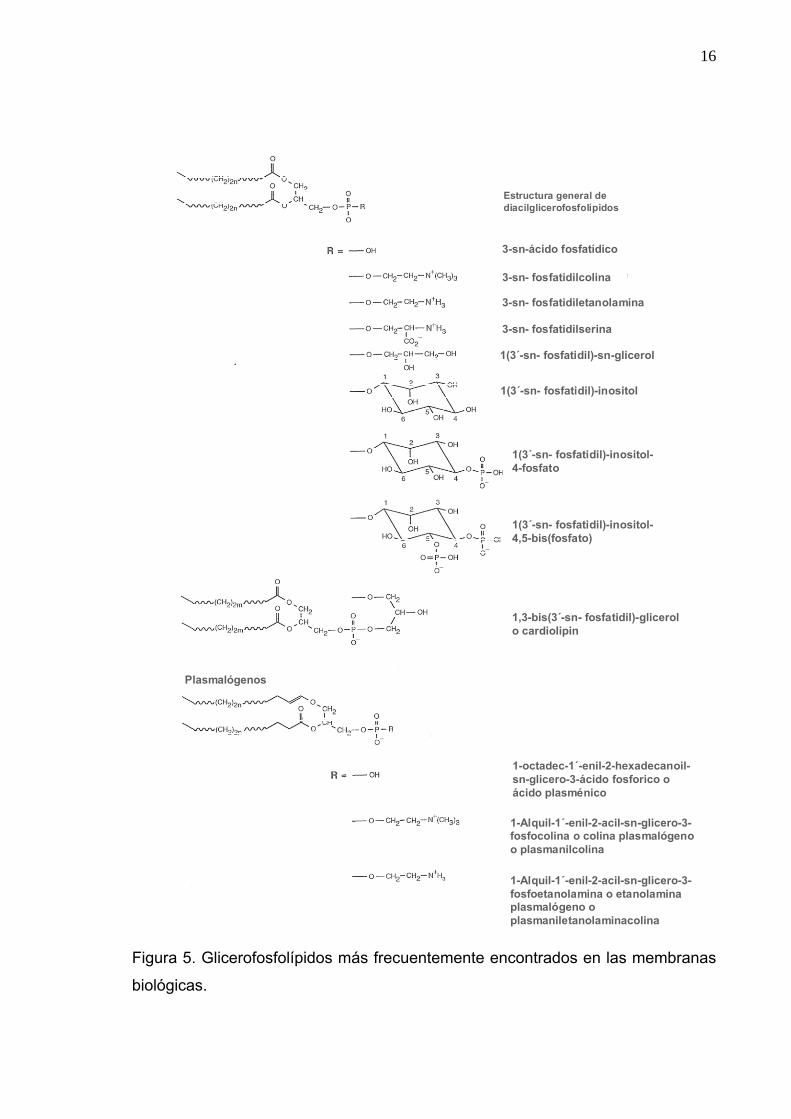
Glicerofosfolípidos Los principales lípidos constituyentes de las membranas son de la clase de
lípidos complejos y glicolípidos. Los más comúnmente encontrados son los
glicerofosfolípidos, los cuales son resumidos en la figura 5. Los glicerofosfolípidos
son derivados de sn-glicero-3-ác fosfórico, usualmente con dos ácidos grasos
esterificados en las posiciones 1 y 2 del glicerol.
Los diacilglicerofosfolípidos son los lípidos predominantes en las
membranas celulares de las células eucarióticas y procarióticas. En células el 1,2-
diacil-sn-glicero-3-fosfocolina es el principal constituyente de la membrana celular
animal y el 1,2-diacil-sn-glicero-3-fosfoetanolamina con frecuencia es el principal
constituyente de la membrana bacteriana.
La cardiolipina es el principal fosfolípido componente de la monocapa
interior de la membrana mitocondrial, cloroplasto y de algunas membranas

15
bacterianas. El plasmalógeno es un término genérico para glicerofosfolípidos en
los cuales el glicerol se une a un grupo 1-alquenil éter. La etanolamina
plasmalógeno o plasmeniletanolaminas son componentes importantes de mielina
y el retículo sarcoplásmico cardíaco (Gross et al., 1985). El término genérico para
fosfolípidos con solo una cadena hidrocarbonada es lisofosfolípido.
Como se ha comentado, los fosfoglicéridos son generalmente la clase de
lípidos dominante especialmente en membranas de células de origen animal,
seguidos por las esfingomielina y el colesterol.
Los fosfoglicéridos están representados principalmente por fosfatidilcolina
(FC), fofatidiletanolamina (FE), fosfatidilserina (FS), fosfatidilinositol (FI) y
lisofosfatidilcolina (LFC). Los más abundantes son los tres primeros (figura 5),
siendo la FC y la FE fosfolípidos con carga neta igual a cero, y la FS el fosfolípido
cargado eléctricamente más abundante (carga negativa) (Finean et al., 1978). Por
tal razón, se eligieron como sistema de estudio dos lípidos, uno neutro la
Estearoil-2-Oleil-sn-Glicero-3-Fosfatidilcolina (SOPC) y un lípido cargado
negativamente Estearoil-2-Oleil-sn-Glicero-3-Fosfatidil-L-Serina (SOPS), para la
fabricación de las vesículas que se estudiaron en la primera parte de este trabajo.
A pesar de las considerables variaciones en la composición de
glicerofosfolípidos en las membranas celulares, todos ellos tienen una
característica importante: son anfifílicos, es decir, una parte de la molécula es
insoluble en agua o hidrofóbica (la cadena hidrocarbonada) y la otra parte es
soluble en agua o hidrofílica (la cabeza polar). La polaridad de la cabeza se debe
a la presencia de cargas eléctricas asociadas con el fosfato y la base nitrogenada
si está presente y/o al gran número de grupos oxidrilos. La naturaleza anfifílica de
los lípidos de la membrana se puede claramente ilustrar analizando la estructura
de los fosfolípidos utilizados en este trabajo: la fosfatidilcolina (FC) y la
fosfatidilcolina (FS), esquematizados en la figura 5. En la figura se puede observar
la ubicación de las cargas de la FC: positiva (en el grupo colina) y negativa (en el
grupo fosfato), lo cual resulta en una carga neta cero, característica en los
zwitteriones.
16
Estructura general de diacilglicerofosfolípidos
3-sn-ácido fosfatídico
3-sn- fosfatidilcolina
3-sn- fosfatidiletanolamina
3-sn- fosfatidilserina
1(3´-sn- fosfatidil)-sn-glicerol
1(3´-sn- fosfatidil)-inositol
1(3´-sn- fosfatidil)-inositol-4-fosfato
1(3´-sn- fosfatidil)-inositol-4,5-bis(fosfato)
1,3-bis(3´-sn- fosfatidil)-glicerol o cardiolipin
Plasmalógenos
1-octadec-1´-enil-2-hexadecanoil-sn-glicero-3-ácido fosforico o ácido plasménico
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfocolina o colina plasmalógenoo plasmanilcolina
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfoetanolamina o etanolaminaplasmalógeno o plasmaniletanolaminacolina
Estructura general de diacilglicerofosfolípidos
3-sn-ácido fosfatídico
3-sn- fosfatidilcolina
3-sn- fosfatidiletanolamina
3-sn- fosfatidilserina
1(3´-sn- fosfatidil)-sn-glicerol
1(3´-sn- fosfatidil)-inositol
1(3´-sn- fosfatidil)-inositol-4-fosfato
1(3´-sn- fosfatidil)-inositol-4,5-bis(fosfato)
1,3-bis(3´-sn- fosfatidil)-glicerol o cardiolipin
Plasmalógenos
1-octadec-1´-enil-2-hexadecanoil-sn-glicero-3-ácido fosforico o ácido plasménico
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfocolina o colina plasmalógenoo plasmanilcolina
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfoetanolamina o etanolaminaplasmalógeno o plasmaniletanolaminacolina
Estructura general de diacilglicerofosfolípidos
3-sn-ácido fosfatídico
3-sn- fosfatidilcolina
3-sn- fosfatidiletanolamina
3-sn- fosfatidilserina
1(3´-sn- fosfatidil)-sn-glicerol
1(3´-sn- fosfatidil)-inositol
1(3´-sn- fosfatidil)-inositol-4-fosfato
1(3´-sn- fosfatidil)-inositol-4,5-bis(fosfato)
1,3-bis(3´-sn- fosfatidil)-glicerol o cardiolipin
Plasmalógenos
1-octadec-1´-enil-2-hexadecanoil-sn-glicero-3-ácido fosforico o ácido plasménico
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfocolina o colina plasmalógenoo plasmanilcolina
1-Alquil-1´-enil-2-acil-sn-glicero-3-fosfoetanolamina o etanolaminaplasmalógeno o plasmaniletanolaminacolina
Figura 5. Glicerofosfolípidos más frecuentemente encontrados en las membranas
biológicas.

17
Con respecto a la molécula de FS, podemos decir que es un anión a pH
neutro, debido a que dos cargas se encuentran en la serina (una positiva y una
negativa en el grupo amino y carboxilo respectivamente), además de la carga
negativa ubicada en el grupo fosfato. Así, la carga neta de la molécula es
negativa.
Esfingomielina Uno de los componentes importantes de las membranas biológicas son los
esfingolípidos, los cuales juegan un papel importante tanto en la estructura como
en la función celular. Una de sus funciones es la formación de dominios en las
membranas al interaccionar con moléculas de fosfolípidos y colesterol. Es
importante mencionar que en el presente trabajo utilizaremos mezclas de éstos
tres lípidos en diferentes proporciones para fabricar los modelos de membranas a
los cuales se le determinarán sus propiedades elásticas.
La estructura fundamental común para esfingolípidos es esfinganina, su
análogo insaturado trans-4-esfinganina o esfingosina y se muestra en la figura 6.
El más simple derivado de esfingosina es sicosina en el cual un monosacárido es
enlazado glicosídicamente al hidroxilo de la posición 1 de la esfingosina. Un
ejemplo es 1-O-(D-galactopiranosil)-esfingosina. La acilación del grupo NH2 en la
posición 2 de la esfingosina da como resultado otra estructura fundamental, N-
acil-esfingosina para la cual el nombre genérico es ceramidas.
Cerebrósidos y gangliósidos. Los lípidos esfingosina, sicosina y ceramida
son esfingolípidos; no obstante este nombre es usado tanto para la clase de
cerebrósidos y gangliósidos como para glicolípidos. Como ya se ha mencionado
el esfingolípido importante para este trabajo es la esfingomielina, la cual esta
formado por una N-Acil-esfingocina o ceramida que en su grupo hidroxilo terminal
se une usualmente a la fosfocolina o fosfoetanolamina mediante un enlace éster.
Similar a fosfolípidos, la esfingomielina contiene un grupo fosfodiester; además de
compartir características fisicoquímicas similares los glicerofosfolípidos y las
esfingomielinas encuentran frecuentemente agrupados.

18
Glicoglicerolípidos Dentro de la clasificación de los glicolípidos, se encuentran los compuestos
llamados glicoglicerolípidos; son aquellos que en la posición sn-3 del glicerol
están unido glicosídicamente un carbohidrato, por ejemplo galactosa. Estos
lípidos son los principales constituyentes de la membrana de cloroplastos. Y están
por encima del 5% de los lípidos polares de las plantas, representando uno de los
lípidos más abundantes de la naturaleza
Los glicoglicerolípidos son raros en el reino animal; contienen un grupo
sulfa unido al carbohidrato y son llamados sulfolípidos. También son encontrados
en cantidades significativas en algas azul verdes y bacterias.
Esfingosina
1-O-β-D-galactosile-esfingosina o sicosina
N-Acil-esfingosina o ceramida
Esfingomielina
Esfingosina
1-O-β-D-galactosile-esfingosina o sicosina
N-Acil-esfingosina o ceramida
Esfingomielina
Figura 6. Esfingolípidos estructuras fundamentales y sus derivados
Cerebrósidos Los cerebrósidos se encuentran dentro de los lípidos llamados
glicoesfingolípidos y su estructura fundamental es ceramida en la cual un mono o
polisacárido es enlazado glicosídicamente a su grupo OH terminal, figura 7. Los
cerebrósidos más complejos son llamados oligoglicosilceramidas usando el
nombre común del residuo mono u oligosacárido. Entendiendo que el azúcar está

19
unido glicosídicamente al C-1 del grupo del hidroxilo de la ceramida. Un ejemplo
es el lactocilceramida.
Gangliósidos Sialoglicoesfingolípidos o gangliósidos son una clase de glicoesfingolípidos
complejos que contienen polisacáridos constituidos por una cadena de cuatro
azucares unidos por enlaces glucosídicos al C-1 del grupo del hidroxilo de la
ceramida. Estos son lípidos aniónicos porque la cadena de polisacáridos contiene
una o más moléculas de ácido siálico (ácido-N-acetil neuramínico) negativamente
cargado (figura 7). El ácido siálico está enlazado glicosídicamente a uno de los
residuos del azúcar o de la cadena del oligosacárido.
Los glicoesfingolípidos están presentes en la monocapa exterior de la
membrana plasmática de mamíferos, usualmente como componente menor. Los
glicoesfingolípidos tienen propiedades antigénicas y actúan como receptores para
anticuerpos, lectinas y ciertas toxinas. Glicoesfingolípidos son constituyentes la
membrana de eritrocitos. Se cree que los glicoesfingolípidos se originan en el
sistema membranoso del retículo endoplásmico desde el cual son exportados al
aparato de Golgi.
Lipopolisacáridos Los lípidos X, Y y A que se muestran en la figura 8 son constituyentes de
las moléculas de los lipopolisacáridos, los cuales se encuentran en la parte
exterior de la membrana de bacterias Gram negativas.
Los lipopolisacáridos (figura 9), son glicolípidos responsables de la antigenicidad y
patogenicidad de Escherichia coli y otras bacterias Gram negativas. Las
lipopolisacáridos, son responsables de una gran variedad de actividades
biológicas: entre otras, median la interacción de los microorganismos con la célula
huésped, activan el sistema inmune de mamíferos y tienen actividad endotóxica.
Estas actividades son responsables del gran interés que han despertado los
lipopolisacáridos en las últimas dos décadas. El lípido A provee la porción
hidrofóbica con el cual los lipopolisacáridos se anclan a la membrana de bacterias
Gram negativas. Las estructuras químicas del lípido X y el lípido Y figura 8 se

20
elucidaron recientemente; ambos pueden ser considerados como constituyentes
de la molécula de lípido A. La estructura del lípido A de Escherichia coli se ha
estudiado extensamente y su estructura propuesta tiene una parte a la que se
unen dos residuos de 2-deoxy-2-amino-D-glucopiranosil, los cuales están unidos
por enlaces β1-6
β-D-galactosilceramida
β-D-Gal-(1-4)-β-D-Glc-ceramidaO lactosilceramida
Gangliósido GM1
N-Acetil-α-D-neuraminato
β-D-galactosilceramida-3´-sulfato
β-D-galactosilceramida
β-D-Gal-(1-4)-β-D-Glc-ceramidaO lactosilceramida
Gangliósido GM1
N-Acetil-α-D-neuraminato
β-D-galactosilceramida-3´-sulfato
Figura 7. Estructuras fundamentales y derivados de glicoesfingolípidos
(cerebrósidos y gangliósidos.)

21
Lípido X Lípido Y
Lípido A
Lípido X Lípido Y
Lípido A
Figura 8. Lipopolisacáridos. La estructura química propuesta del lípido A de E. coli
se presenta junto con la estructura química del lípido X y el lípido Y los cuales son
constituyentes del lípido A (glucosalina)
Figura 9. Estructura del 2—3-deoxy-D-manno-ceto-deoxi-D-ácido-manno-octánico

22
En las secciones anteriores se describieron las propiedades estructurales y
funciones más importantes de los lípidos que conforman las principales
membranas biológicas. A continuación, se tratarán las propiedades físicas y
químicas de los fosfolípidos. Dichas propiedades están directamente involucradas
en la agregación y auto-organización de los fosfolípidos, procesos mediante los
cuales forman bicapas, bases estructurales de las membranas biológicas.
Agregación de Fosfolípidos
Los fosfolípidos son el componente principal en las células y membranas
de organelos, lipoproteínas y tensoactivos de pulmón. La interacción de estos
compuestos con el agua es de vital importancia para la formación de estructuras
que desempeñan importantes funciones biológicas. En esta sección se describen
los agregados lipídicos de más importancia, como las bicapas y las micelas. Es
importante mencionar que la formación de agregados no solamente depende de
las propiedades de los lípidos, sino también de su interacción con el solvente, así
como variables como el pH, la fuerza iónica y la temperatura.
La auto-organización de las moléculas anfifílicas (aquellas compuestas por
una cabeza polar y una cola hidrofóbica) permite la formación de una gran
variedad de estructuras. Las fases resultantes pueden dividirse
esquemáticamente como soluciones isotrópicas, fases sólida y fases líquida
cristalina. Las soluciones isotrópicas se caracterizan por tener desorden a corta y
larga distancia, mientras que las fases líquido cristalino o mesofases presentan
orden a largo alcance. En ambos, soluciones isotrópicas y cristales líquidos, el
estado de las cadenas hidrocarbonadas de las anfifílicas se puede describir en
general como líquido; las moléculas tienen gran movilidad dentro del plano de la
membrana. En cristales, formados por debajo de la temperatura de fusión de las
cadenas hidrocarbonadas, el estado es más o menos parecido al sólido.
Las moléculas anfifílicas pueden auto-organizarse en estructuras de formas
diversas como micelas esféricas o cilíndricas, las cuales tienen hacia su interior
las cadenas hidrocarbonadas rehuyendo del agua, mientras que las cabezas
polares están en la superficie en contacto con el agua. Las micelas esféricas se

23
caracterizan por un valor bajo del llamado parámetro de empaquetamiento v/la <
1/3 y una gran curvatura espontánea. El centro hidrocarbonado de las micelas
esféricas tiene un radio similar a la longitud de las cadenas hidrofóbicas de las
moléculas. Las micelas cilíndricas también tienen un interior compuesto por
cadenas hidrocarbonadas, mientras que las cabezas polares están en la interface
con el agua. En este caso, la longitud de la micela es altamente variable, así que
las micelas son polidispersas en tamaño.
Un aspecto que es importante mencionar es que las micelas se forman a
una concentración dada de moléculas anfifílicas, la cual recibe el nombre de
concentración micelar crítica (cmc), concepto que comentamos a continuación.
Concentración Micelar Crítica Cuando moléculas anfifílicas se disuelven en agua, su característica dual
en cuanto a su polaridad (o inversamente, respecto a su hidrofobicidad) hace
que las moléculas espontáneamente formen estructuras. A concentraciones
bajas, lo primero que ocurre es la formación de una monocapa superficial en la
frontera agua-aire cuyo efecto más notorio consiste en disminuir la tensión
superficial del agua. Esta monocapa se forma porque las cabezas polares
penetran al agua pero las colas hidrofóbicas salen hacia el aire (Fennell et al.,
1999). Sin embargo, si se incrementa la concentración de moléculas anfifílicas
en el agua, estas forman agregados esféricos exponiendo sólo la parte polar al
agua y escondiendo la parte hidrofóbica en el interior de los agregados. Tales
agregados se llaman micelas (Tanford et al., 1980a). Los agregados que se
forman primero generalmente son de forma esférica y la concentración a la cual
inician su formación es conocida como concentración micelar crítica o cmc. La
cmc es una de las características más importantes de las moléculas anfifílicas y
depende de la energía libre ganada cuando un molécula anfifílica aislada en
solución forma parte de un agregado (Tanford et al., 1980a).
Dado que las propiedades de las soluciones de moléculas anfifílicas
tales como turbidez, tensión superficial, conductividad, entre otras, cambian
abruptamente cuando se alcanza la cmc, esta concentración se determina

24
siguiendo la evolución de alguna de estas propiedades. (Israelachvili, 1992). De
manera general, y en una primera aproximación al problema, se puede afirmar
que la predisposición para formar micelas de una molécula anfifílica particular
depende fuertemente de su estructura geométrica. Cuando el volumen de la
cabeza polar de la molécula anfifilica es mayor que el volumen de la cola
hidrofóbica, la molécula tiene una forma cónica. La base del cono es la cabeza
polar y el vértice queda hacia la cola hidrofóbica. En esas condiciones, cuando
las moléculas se aglomeran para formar micelas, tienen una tendencia a
cerrarse dejando la parte hidrofóbica en el interior, lo cual favorece la formación
de micelas en agua. Si por el contrario el volumen de la parte hidrofóbica es
mayor que el de la parte polar, el cono está invertido y la tendencia es a
cerrarse dejando en el interior la parte polar, esto favorece la formación de
micelas en un solvente orgánico (ejemplos: decano, iso-octano, entre otros), a
estas micelas se les conoce como micelas inversas (Israelachvili 1992). Existen
otras situaciones en donde la molécula anfifílica no puede ser concebida como
un cono. Por ejemplo, si el área asociada a la cabeza polar y la sección
transversal de la cola hidrofóbica son comparables, la molécula asemeja más
bien a un cilindro. Esto da lugar a un agregado lamelar de baja curvatura, es
decir, a la formación de bicapas o membranas.
Efecto Hidrofóbico. Las moléculas anfifílicas poseen en su estructura química una fracción
hidrofóbica que rehúye del agua, y una fracción hidrofílica que tiene afinidad por el
agua. Los fosfolípidos son de esta naturaleza, en su parte hidrofóbica
generalmente tiene una cadena hidrocarbonada y en la fracción hidrofílica tienen
un grupo derivado del ácido fosfórico. Debido a la baja solubilidad en agua de la
cadena hidrocarbonada y los fuertes enlaces de hidrógeno entre las moléculas de
agua se genera la fuerza atractiva que mantienen unidos a los fosfolípidos en
complejos moleculares.
El efecto hidrofóbico se puede describir más por una atracción entre las
moléculas de agua que por las atracciones entre moléculas de hidrocarbonos
(Hartley et al., 1936). Según esta visión, la energía libre interfacial de atracción

25
entre moléculas de agua es lo que genera la gran dificultad del contacto del agua
con las moléculas los hidrocarbonos. En términos moleculares el agua es un
líquido altamente estructurada con una configuración tetraédrica y con 4 posibles
enlaces de hidrógeno con otras moléculas de agua vecinas lo que forma un
arreglo muy estable. No obstante se pueden formar nuevos puentes de hidrógeno
con un soluto polar pero no con solutos no polares. En contacto con solutos
hidrofóbicos se rompe el ordenamiento de la red molecular en el agua, además
cuando la parte hidrofóbica de una molécula anfifílicas tiene contacto con el agua,
el área de contacto es reducida, reduciendo así la energía libre (Tanford et al.,
1980).
La fuerte inclinación de las moléculas de agua para formar puentes de
hidrógeno entre sí afecta la interacción con moléculas no polares que son
incapaces de formar puentes de hidrógeno. Ejemplos de estas son los
hidrocarbonos y los fluorocarbonos. Cuando las moléculas de agua se ponen en
contacto con dichas moléculas no polares, se enfrentan a un aparente dilema:
cualquiera que sea la forma en que se presente la molécula de agua, parecería
que una o más de sus cuatro cargas tendrá que apuntar hacia la molécula de
soluto inerte y por lo tanto se perderá la formación del enlace de hidrógeno.
Claramente la mejor configuración tendría el menor número de cargas apuntando
hacia la especie no polar de tal forma que las otras cargas puedan apuntar hacia
la fase acuosa y así estar disponible para participar en la formación de puentes de
hidrógeno. Si la molécula del soluto no polar no es muy grande, es posible que las
moléculas de agua formen una red a su alrededor sin ceder ninguno de sus sitios
disponibles para enlaces de hidrógeno. El principal efecto en la interacción del
agua con moléculas no polares es la reorientación de las moléculas de agua de
tal forma que puedan participar en la formación de enlaces de hidrógeno con el
agua del volumen.
Así gracias a la extraordinaria habilidad de coordinación de las moléculas
tetraédricas para enlazarse entre ellas alrededor de casi cualquier molécula inerte
sea la que fuera su forma o tamaño, el aparente dilema mencionado
anteriormente es a menudo fácil de resolver. De hecho ya que las moléculas de

26
agua en estado líquido participan en promedio en aproximadamente 3.0 a 3.5
enlaces de hidrógeno, al parecer alrededor de una molécula de soluto inerte las
moléculas de agua tiene en realidad una mayor coordinación (de cuatro) y por lo
tanto su arreglo puede ser más ordenado que en el volumen del líquido. Esto
frecuentemente es referido como solvatación o hidratación. En el presente no hay
una teoría simple y completa de dichas interacciones soluto-solvente que permita
explicar el efecto hidrofóbico. No obstante, estudios teóricos y experimentales
indican que la reorientación o reestructuración del agua alrededor de solutos o
superficies no polares es entrópicamente desfavorable, ya que se altera la
estructura existente del agua y se impone una estructura más ordenada. También
se observa que para diferentes hidrocarbonos, la energía libre es
aproximadamente proporcional al área superficial de las moléculas. Es claro que,
cuando las moléculas de agua en el bulto tienen un rearreglo de coordinación
para dar cabida a una molécula extraña de soluto el precio es alto; pero cuando
una molécula de agua es completamente sacada de su red de interacción el
precio es aún más alto. La inmiscibilidad de sustancias inertes en agua, y la
naturaleza principalmente entrópica de esta incompatibilidad es conocida como
efecto hidrofóbico (Tanford, 1980). Este efecto es el responsable de que el agua y
el aceite nos se mezclen. También, este efecto explica la formación de micelas y
de membranas biológicas.
Formas y Agregados de Moléculas Anfifílicas. Las moléculas anfifílicas tales como surfactantes, lípidos, copolímeros y
proteínas por su naturaleza dual pueden organizarse espontáneamente en
presencia de agua formando arreglos moleculares de forma esférica o cilíndrica
llamados micelas. Otra estructura posible son las bicapas; estas a su vez, pueden
tener una topología abierta (planos fluctuantes) o pueden formar agregados
cerrados como las vesículas. Las condiciones fisicoquímicas del sistema
(temperatura, concentración, pH, fuerza iónica) determinan cual fase es estable
en un momento dado y cuales son las transformaciones posibles entre ellas.
La explicación a éste comportamiento se hizo desde el punto de vista de
que la autoagregación se llevaba acabo termodinámicamente debido al efecto

27
hidrofóbico. Así que se asumió que en las micelas la cadena hidrocarbonada
estaba completamente confinada en su interior, donde se asumió que no había
agua. El grupo que conforma la cabeza polar se ordena en la superficie del
agregado manteniendo un fuerte enlace con el agua. (Tartar H. V. et al., 1955)
En las micelas el área por molécula de lípido es menor que en las bicapas
las cuales frecuentemente forman estructuras cerradas llamadas vesículas o
liposomas y el área por molécula calculada en las monocapas interna y externa
difieren ligeramente de una estructura lamelar plana.
Por otra parte, un aspecto importante es referente a los anfifilos con dos
cadenas hidrocarbonadas tales como los fosfolípidos, el área de la cabeza sería
la misma si la molécula tuviera una o dos cadenas hidrocarbonadas, pero el área
por cadena sería la mitad si fuera de una sola cadena en lugar de dos (Tanford et
al., 1980) por lo que es claro que la estructura química de los componentes de
una membrana puede modificar de manera importante la forma de los agregados.
Como se había comentado las diversas formas en las que se organizan los
lípidos en agua esta gobernada tanto por las propiedades estructurales de las
moléculas anfifílicas como las propiedades del solvente. Una teoría considera las
propiedades geométricas de los lípidos y su capacidad para agruparse en función
de dichas características relacionando el área óptima de superficie ao, el volumen
de las cadenas hidrocarbonadas v, la longitud crítica de la cadena lc, la longitud
crítica de la cadena es la longitud máxima al cual se puede extender la cadena.
Las cantidades mencionadas anteriormente definen el parámetro crítico de
empaquetamiento v/ao lc que determina la forma que adoptarán las moléculas
anfifílicas ensambladas (Israelachvili, 1980).
Las estructuras predichas son aquellas en las cuales cada molécula de
lípido tendrá un mínimo de energía. Por lo que, sí el valor resultante de la
ecuación del parámetro de empaquetamiento v/ao lc es menor a 1/3 corresponderá
a una molécula en forma de cono, favoreciendo la formación de micelas; si el
valor está entre 1/3 y 1/2 corresponde a una forma de cono truncado,
favoreciendo la formación de micelas cilíndricas y elipsoidales; si los valores están
entre 1/2 y 1 la forma de las moléculas corresponden a cilíndricas, y se formarán

28
bicapas o vesículas, y si el valor es mayor a 1 corresponde a moléculas con
cabezas relativamente pequeñas, formando micelas inversas o una fase
hexagonal invertida. No obstante se sabe que los diacilfosfolípidos tienen una
forma aproximada a los cilindros, así que forman principalmente bicapas es decir
estructuras lamelares. Por otra parte algunos fosfolípidos tales como
fosfatidiletanolaminas insaturadas tienden a formar fases hexagonales, ya que
ellos tienen una cabeza relativamente pequeña en comparación con el volumen
de la región de la cadena hidrocarbonada (Israelachvili, 1980).
Por otra parte para moléculas que se ensamblan para la formación de
micelas esféricas, su área de superficie óptima a0 debe ser suficientemente
grande y su volumen v suficientemente pequeño de tal forma que el radio de la
micela R no exceda la longitud crítica de la cadena lc
Es importante decir que los lípidos que forman bicapas son aquellos que no
se pueden empacar dentro de una estructura micelar debido a su pequeña área
de la cabeza polar a0 o porque su cadena hidrocarbonada es demasiado
voluminosa para caber en agregados pequeños. Para lípidos que forman bicapas,
el valor de v/ao lc se debe encontrar cercano a 1, y esto requiere que para la
misma área de la cabeza a0 y longitud de la cadena lc, su volumen v deba ser
aproximadamente dos veces mayor al de los lípidos que forman las micelas. Por
ello, los lípidos con dos cadenas hidrofóbicas tienen gran propensión a formar
bicapas. Por ejemplo, una cadena de lisolecitina forma pequeñas micelas, aunque
no de forma esférica, mientras que la lecitina,con dos cadenas tales como di-C14 y
di-C16, forma bicapas.
La existencia de la doble cadena hidrocarbonada en la estructura química
de los lípidos también afecta a otras propiedades de los agregados, tanto
estáticas como dinámicas. Esto es debido a que se incrementa la hidrofobicidad
de los lípidos, la cual a su vez reduce drásticamente su cmc. Las estructuras
autoensambladas son con frecuencia altamente dinámicas, con las moléculas en
constante movimiento térmico; además, existe un equilibrio en cuanto al
intercambio de moléculas con los monómeros que permanecen en solución.

29
Por otra parte, las bicapas también pueden formar las estructuras cerradas
conocidas como vesículas. En este caso, se vuelve más favorable el cierre de las
bicapas para generar una forma esférica que una estructura plana infinita. Esto es
debido a que en una bicapa cerrada (vesícula) se eliminan los bordes
energéticamente desfavorables. Así, cuando los lípidos en una bicapa curvada
pueden mantener su área en el valor óptimo, las vesículas son las estructuras
preferidas. Como ya se ha comentado antes, las bicapas planas existen cuando el
valor del parámetro de empaquetamiento es v/ao lc ≈ 1. Para una bicapa curvada
los lípidos de la monocapa externa deben estar disponibles para empacarse en
promedio en conos truncados, lo cual requiere que v/ao lc <1.
Fases Liotrópicas de Fosfolípidos
Nomenclatura Las moléculas anfifílicas como lo son los fosfolípidos tienen gran habilidad
de formar estructuras diversas con gran periodicidad y poco grado de desorden.
Luzzati et al, realizaron un estudio detallado de muchas fases formadas por
moléculas anfifílicas anhídridas e hidratadas.
La nomenclatura más ampliamente usada y aceptada para nombrar las
mesofases liotrópicas es la propuesta por (Luzzati et al., 1999).
En esta nomenclatura el tipo de red se denota con una letra mayúscula, L
para lamelar, H para hexagonal, y Q para cúbica. Se utilizan los subíndices I y II
para denotar si el tipo de fase es normal aceite en agua o inversa agua en aceite.
Además se usa una letra griega como subíndice para denotar la conformación de
la cadena hidrocarbonada: β para fase gel ordenada, α para fase líquida, αβ para
las regiones de coexistencia de fase gel y fase líquida y δ para la conformación de
cadenas helicoidales, además la letra c se usa frecuentemente para denotar un
arreglo cristalino. En la tabla 3 se describen las mesofases liotrópicas.
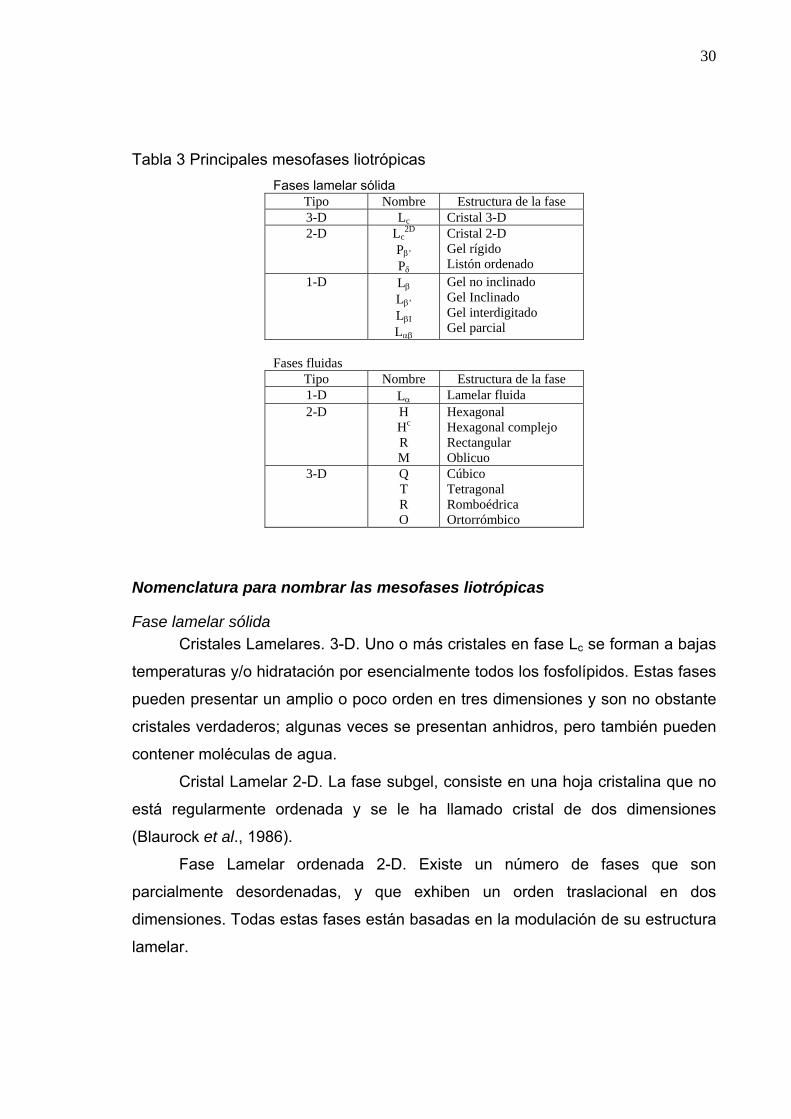
30
Tabla 3 Principales mesofases liotrópicas Fases lamelar sólida
Tipo Nombre Estructura de la fase 3-D Lc Cristal 3-D 2-D Lc
2D Pβ’ Pδ
Cristal 2-D Gel rígido Listón ordenado
1-D Lβ Lβ’ LβI Lαβ
Gel no inclinado Gel Inclinado Gel interdigitado Gel parcial
Fases fluidas
Tipo Nombre Estructura de la fase 1-D Lα Lamelar fluida 2-D H
Hc R M
Hexagonal Hexagonal complejo Rectangular Oblicuo
3-D Q T R O
Cúbico Tetragonal Romboédrica Ortorrómbico
Nomenclatura para nombrar las mesofases liotrópicas
Fase lamelar sólida Cristales Lamelares. 3-D. Uno o más cristales en fase Lc se forman a bajas
temperaturas y/o hidratación por esencialmente todos los fosfolípidos. Estas fases
pueden presentar un amplio o poco orden en tres dimensiones y son no obstante
cristales verdaderos; algunas veces se presentan anhidros, pero también pueden
contener moléculas de agua.
Cristal Lamelar 2-D. La fase subgel, consiste en una hoja cristalina que no
está regularmente ordenada y se le ha llamado cristal de dos dimensiones
(Blaurock et al., 1986).
Fase Lamelar ordenada 2-D. Existe un número de fases que son
parcialmente desordenadas, y que exhiben un orden traslacional en dos
dimensiones. Todas estas fases están basadas en la modulación de su estructura
lamelar.

31
Fase Pβ´ Gel Rígida. Ocurre a temperaturas por debajo de la fase lamelar
fluida Lα como se muestra en la figura 10 (inciso a) y esto ha sido observado en
fosfatidilcolinas (Tardieu A. et al,. 1973), fosfatidilglicerol a pH neutro, (Watts A. et
al., 1978), fosfatidiletanolamina, (Stümpel J. et al., 1980), y en ácido fosfatídico
(Harlos, K. et al., 1979) a pH altos.
Fase Lamelar ordenada 1-D. En esta fase las moléculas son ordenadas
en las bicapas la cual a su vez se ordena en una estructura de multicapas y están
separadas entre ellas por agua como se muestra en la figura 11. Las bicapas son
paralelas y equidistantes entre sí, el espesor de la capa de agua depende de
factores tales como el contenido de agua, temperatura, tamaño de la cabeza,
polaridad y carga. Respecto a la cadena hidrocarbonada esta puede ser un gel
(β), gel parcial (αβ) o helicoidal (δ).
En la fase gel las cadenas son rígidas completamente extendidas y
empacadas en dos dimensiones en una red hexagonal. Las cadenas
hidrocarbonadas pueden ser paralelas Lβ, como se muestra en el inciso b de la
figura 10, rígidas (Lβ) (inciso c) o interdigitada (LβΙ) (inciso d) (Rank, J. L. et al.,
1980).
La fase Lδ se ha observado en fosfatidilcolina seca (Doucet J. et al.,
1983), en este caso las moléculas están arregladas en forma lamelar pero las
cadenas hidrocarbonadas están en forma helicoidal con un arreglo de red
cuadrada en dos dimensiones. Las cabezas polares están orientadas
perpendicularmente a la bicapa y las cadenas de carbonos están interdigitadas
con las moléculas de la otra monocapa.
Fase fluida 2-D. Las fases fluidas de dos dimensiones mejor establecidas
son las fases de topología hexagonal HI y HII normal y inversa como se muestra
en la figura 12. Las cadenas son fluidas y están contenidas dentro de cilindros tipo
I o normal hexagonal (HI), o el agua está dentro de los cilindros y las cadenas
hidrocarbonadas fluidas llenan el espacio entre los cilindros (HII). La fase HI se
presenta comúnmente en sistemas de tensoactivos simples y no con
diacilfosfolípidos. La fase (HII) se presenta frecuentemente en sistemas de
diacilfosfolípidos como fosfatidiletanolamina, de cabeza pequeñas y débilmente

32
hidratadas así como fuertes interacciones cabeza-cabeza (Seddon et al., 1990).
Se han observado fases similares a HI pero con simetría rectangular u oblicua con
ciertos tensoactivos como el dodecil sulfato de sodio SDS y agua (Kékicheff et al.,
1987).
a) b)
c) d)
a) b)
c) d)
Figura 10. Estructuras de fase gel: a) Gel rígido Pβ’, b) Gel no inclinado Lβ, c) Gel inclinado Lβ’, d) Gel interdigitado LβΙ.
Fase fluida 3-D. En esta fase se puede observar un complejo patrón de
picos de Bragg en la región de bajo ángulo. Lα puede estar en una topología
normal (aceite en agua) o inversa (agua en aceite). La gran mayoría de las fases
líquidas de tres dimensiones hasta ahora descubiertas son de simetría cúbica,
aunque también se han detectado fases de topología inversa tales como:
romboedros, tetragonales, y ortorrómbicos en pocos sistemas de lípidos de baja
hidratación.

33
Aceite
Agua
Aceite
Agua
Aceite
Agua
Aceite
Agua
Figura 11. La fase lamelar fluida Lα en su conformación en agua y aceite
Fase de solución. La estructura de las fases puede ser formada por
fosfolípidos los cuales se pueden autoensamblar por sus propiedades en micelas,
microemulsiones, o la llamada L3 fase esponja. a)
b)
a)
b)
Figura 12. Fases hexagonales normal e inversa a) HI, b) HII

34
Las micelas pueden ser formadas por fosfolípidos de cadena corta
generalmente C6 o C8 o por lisofosfolípidos en agua; las micelas pueden tener una
variedad de formas tales como esferas, tubulares, discos, además se pueden
obtener soluciones de micelas inversas al hidratar fosfolípidos en presencia de
ciertos solventes orgánicos como lo son benceno o clorobenceno (Luisa et al.,
1985). Por otra parte se sabe que las fosfatidilcolinas en presencia de pequeñas
cantidades de agua de 2 a 8 moléculas por lípido y ciertos solventes orgánicos
como los alcanos forman geles rígidos no birrefringentes (Scartazzini R., 1988).
Las microemulsiones por otra parte son soluciones transparentes
microestructuradas formadas por una mezcla de tensoactivo, aceite y agua (de
Gennes P. G., 1982). En algunos sistemas de tensoactivos, se han encontrado
geles rígidos de microemulsiones los cuales presentan una estructura de fase
cúbica, aunque aparentemente esta basada en una monocapa interfacial mas que
en una bicapa como la fase cúbica bicontinua de fosfolípidos (Anderson D. M.,
1990; Scartazzini R. 1988; Radiman S., 1990).
Vesículas de Fosfolípido: Las vesículas son estructuras formadas por lípidos con propiedades
anfifílicas que en presencia de agua o soluciones acuosas interaccionan entre sí y
con el solvente bajo condiciones adecuadas formando bicapas lipídicas que se
cierran atrapando volúmenes de solvente; las formas que toman pueden ser
diversas: esferas, tubulares, mixtas, trenzados, en forma de collar de perlas, entre
otras (Paredes, G. et al. 2006).
En éste sentido en la década de los sesentas se mostró que los fosfolípidos
en presencia de agua formaban espontáneamente estructuras cerradas de
membranas y se propuso la aplicación de las vesículas de fosfolípido como
modelos de membranas biológicas (Bangham A. et al., 1968).
Como es sabido el estudio de las propiedades fisicoquímicas de
membranas tiene un gran interés en la actualidad para entender los fenómenos
básicos de funcionamiento y estructurales de la vida a escala celular. Una de las
interrogantes fundamentales estriba en la elucidación de la relación entre la
función de una membrana y su composición química. Otra interrogante es el

35
efecto de la composición de las membranas sobre la forma característica de los
organelos, células y microorganismos (Hotani et al., 1999). En éste sentido, en
nuestro trabajo nos proponemos estudiar de manera sistemática la relación entre
la composición química de las membranas modelo formadas por mezclas de
fosfolípidos y las formas que adquieren las membranas, lo cual está relacionado
con su función biológica. Por otra parte y paralelamente a este interés
fundamental, existe un gran interés comercial por la aplicación de los agregados
de fosfolípidos en industrias como la alimenticia y la biotecnológica (Gutiérrez et
al., 1999).
Las vesículas multilamelares son agregados esféricos formados por
bicapas lipídicas arregladas de manera concéntrica y equidistantes, separadas
por capas de agua. Las vesículas multilamelares tienen un patrón característico
en la difracción de rayos x, mostrando una serie de picos a bajo ángulo y una
reflección ancha y difusa en la región de ángulo amplio. A bajo ángulo el patrón
consiste en una serie de picos, lo cual es debido al arreglo periódico de las
bicapas.
Las vesículas unilamelares son definidas como agregados esféricos que
constan de una sola bicapa o membrana cerrada que rodea un volumen acuoso.
El patrón de difracción de rayos x de dispersiones diluidas de vesículas
unilamelares es claramente diferente a las presentadas por vesículas
multilamelares ya que se observa, como se comentó una amplia dispersión en vez
de una serie de picos a bajo ángulo.
Las vesículas unilamelares son divididas de manera arbitraria en dos
clases de acuerdo a su tamaño; las vesículas con diámetros mayores a 100 nm
son referidas como vesículas unilamelares grandes y vesículas menores a 100
nm como vesículas unilamelares pequeñas (Hope M. J. et al, 1986). Por otra
parte, a las vesículas con un tamaño aproximado a 20 μm se les denomina
vesículas gigantes; pueden ser tanto unilamelares como multilamelares.

36
Dinámica e Interacción de los Lípidos de Membrana
La matriz de lípidos de casi todos las biomembranas es un líquido cristalino
en estado lamelar caracterizado por la formación de una bicapa estable
principalmente formada por fosfolípidos que mantienes sus propiedades fluidas y
su movilidad molecular. Los grados de libertad en la movilidad de los lípidos están
inherentemente ligados a la elasticidad de la bicapa pero también a las funciones
de la membrana, tales como permeabilidad para solventes, solutos y la acción de
proteínas periféricas y proteínas integrales de la membrana. Mantener la
naturaleza de la fase líquido cristalino de los lípidos requiere una composición
específica de la membrana, rango de temperatura y estado de hidratación (Reiss-
Huson, F., 1967).
El orden y dinámica de los lípidos son estudiados por resonancia
magnética nuclear (NMR), infrarrojo y espectroscopía Raman, espectroscopía de
fluorescencia con marcadores localizados en diferentes regiones de la bicapa,
calorimetría diferencial de barrido (DSC), rayos x y experimentos de difracción de
neutrones, además de simulaciones moleculares. Las simulaciones moleculares
proveen la localización dependiente del tiempo de todos los átomos de la bicapa,
permitiendo la más detallada descripción del movimiento molecular (Pastor, R. W.
et al., 1991)
Proteínas de la Membrana Las proteínas son responsables de una gran cantidad de procesos
dinámicos llevados a cabo en la membrana, dichas membranas lipídicas forman
una barrera impermeable y por lo tanto establecen los compartimientos en la
célula. En particular, ciertas proteínas transportan químicos e información a través
de la membrana.
Las membranas difieren en su contenido de proteínas. La mielina, una
membrana que sirve como un aislante alrededor de ciertas fibras nerviosas, tiene
una baja concentración de proteínas (18%), también cuenta con una
concentración relativamente alta de lípidos que funcionan como aislamiento. En
contraste, las membranas de muchas otras células son muy activas; estas

37
contienen bombas, canales, receptores y enzimas. El contenido de proteínas de
las membranas plasmáticas, de las mitocondrias y cloroplastos, tienen el más alto
contenido de proteínas generalmente el 75%.
Las proteínas de la membrana poseen características especiales que las
diferencian de otras proteínas globulares debido a que frecuentemente contienen
una proporción elevada de aminoácidos hidrofóbicos, en especial en las zonas de
las proteínas que están embebidas en la membrana. (Berg. J. M. et al., 2002).
Interacción de Proteínas con Lípidos La facilidad o dificultad para extraer una proteína de la membrana nos
indica la fuerza de asociación entre ambas. Algunas proteínas de membrana
pueden ser solubilizadas por medios relativamente suaves, tales como extracción
por una solución de alta fuerza iónica (ej. NaCl 1M). Otras proteínas están
fuertemente enlazadas; estas pueden ser solubilizadas usando detergentes o
solventes orgánicos. Las proteínas de membrana se clasifican en periféricas o
integrales con base en su capacidad de disociarse de la membrana. Las proteínas
integrales de membrana interactúan intensamente con la cadena hidrocarbonada
de la membrana lipídica, y agentes que compitan por esta interacción no polar
pueden liberarlas. En contraste, las proteínas periféricas están unidas a la
membrana por interacciones electrostáticas y puentes de hidrógeno con la cabeza
polar de los lípidos. Estas interacciones se pueden romper por cambios en el pH o
fuerza iónica. Muchas proteínas periféricas están unidas a la superficie de las
proteínas integrales ya sea en el interior o exterior de la membrana.
Fluidez de la Membrana Los constituyentes de las membranas biológicas tienen las propiedades de
un líquido bidimensional. Así, las componentes están en movimiento térmico
continuo y pueden difundirse cuando hay gradientes de concentración. La fluidez
es una propiedad que se puede considerar inversa a la viscosidad. La fluidez de
la membrana puede ser interpretada como una medida del movimiento viscoso de
sus constituyentes, tales como las proteínas involucradas en procesos dinámicos

38
como rotación y difusión lateral. Se puede decir que también la viscosidad es una
propiedad que de alguna manera participa en la función de las membranas.
Transiciones de Fase en las Membranas Los lípidos en membranas biológicas pueden existir en un estado de alto
orden cristalino (estado gel) o en un estado menos ordenado de viscosidad
variable (líquido cristalino). El paso de un estado al otro se denomina transición de
fase y ocurre a una temperatura fija para cada fosfolípido particular. Por ejemplo,
una bicapa artificial compuesta por fosfatidilcolina con dos ácidos mirísticos (14:0)
esterificados al glicerol está en estado gel a temperaturas por debajo de 25 ºC.
Cuando la temperatura es elevada por encima de los 25 ºC la bicapa de lípidos
pasa de un gel cristalino a un cristal líquido mucho más fluido (Karp et al., 1979).
En el último estado mencionado las cadenas hidrocarbonadas adquieren
considerablemente más movilidad. Aunque el glicerol y la cabeza polar mantienen
un arreglo regular, existe evidencia de incremento en la movilidad del grupo polar
y de una reorganización del agua alrededor de este grupo durante el cambio de
fase (Chapman et al., 1975a, b). La temperatura a la cual ocurre el cambio de gel
a cristal líquido se llama temperatura de transición. Ésta se puede medir usando
varias técnicas fisicoquímicas incluyendo calorimetría, tal como se describe en la
sección de materiales y métodos.
Experimentos en sistemas artificiales y membranas naturales han mostrado
que la temperatura de transición es dependiente de la longitud de la cadena del
ácido graso, el grado de insaturación de esta cadena, la naturaleza de la cabeza
polar, el estado de hidratación de la membrana, la presencia o proporción de
colesterol y la pureza de los lípidos constituyentes (Chapman, 1975; Chapman y
Wallach, 1968).
Dominios en la Membrana Celular
En los últimos años ha tomado importancia el tema de la organización de
los lípidos y las proteínas de membrana, específicamente aquellos que se han
concentrado en regiones localizadas conocidas colectivamente como
microdominios de membrana. Por esto ha emergido el interés por un específico

39
tipo de regiones conocidas coloquialmente como (dominios) de membrana.
Precisamente, estas regiones son generalmente definidas como dominios ricos en
esfingolípido y colesterol en fase liquido ordenado. Se ha propuesto que estos
dominios están involucrados en una amplia variedad de procesos celulares tales
como, clasificación de proteínas (Simons, K. 1997), señal de transducción
(Zajchowski, L. D., 2002), transcitosis (Simionescu, N., 1983), (Anderson, R. G.
W., 1992), rutas alternativas de endocitosis (Smart, E. J., 1999), internalización de
toxinas, bacterias y virus (Parton, R. G., et al. 1994, Fivaz, M. et al 1999, Shin, J.
S. et al. 2000), proceso de ensamble y liberación en VIH-1 (Ono, A., and Freed, E.
O., 2001), y transporte de colesterol (Oram, J. F. et al., 1996, Smart, E. J. et al,
1996).
La hipótesis de los dominios es que se les ha descrito como estados de
separación discreta en fase líquida ordenada y desordenada que se presenta
cuando las membranas contienen cantidades suficientes de esfingolípido y esterol
(Brown, D. A., 2001). Un dominio se define como una región que está diferenciada
por sus características físicas que lo distinguen de sus alrededores. En las
membranas los dominios se caracterizan por un espacio ordenado de lípidos y
proteínas. Los dominios de las membranas son regiones ricas en colesterol y
esfingolípidos, y son regiones especializadas que tienen un tamaño y propiedades
bioquímicas únicas.
En las características estructurales del colesterol y de los efingolípidos se
encuentra parte de la explicación de su comportamiento en las membranas. La
esfingomielina en un típico dominio de membrana contiene una ceramida como
estructura principal a la cual se une una fosfatidilcolina para formar lo que se
llama cabeza polar. Debido a los grupos amida y oxidrilo de la esfingomielina,
estos lípidos tienen un grupo donador y receptor para la formación de puentes de
hidrógeno. Además, la cadena hidrocarbonada de la esfingomielina, a la cual se
une la amida, es generalmente saturada. Colectivamente estas propiedades
permiten a estos lípidos la facilidad de formar una red de enlaces de hidrógeno.
En contraste, los predominantes glicerofosfolípidos contienen en su estructura
básica una molécula de glicerol al cual se une un ácido graso en la posición sn-1

40
y sn-2 por medio de enlaces ester. Las cadenas hidrocarbonadas de los
fosfolípidos tienes usualmente 16 o 18 carbonos de longitud, frecuentemente
insaturada en la posición sn-2 y tienen solamente la capacidad de aceptar un
enlace de hidrógeno. No obstante se organizan dentro de la membrana de
manera distinta a los esfingolípidos. Los compuestos más abundantes que forman
la cabeza y que están unidos al glicerol por medio del un enlace fosfo-ester son
serina, colina y etanolamina.
La diferencia en organización de los fosfolípidos y esfingolípidos es clara
cuando se compara las temperaturas de transición de fase de estas especies de
lípidos (Brown, R. D. et al, 1998). El comportamiento de fase de la bicapa lipídica
en general depende del empacamiento, el grado de orden y de la movilidad de
sus lípidos constituyentes; las dos fases mas contrastantes son las fases gel y
líquido cristalino. En una mezcla homogénea de lípidos la temperatura de fusión
de la cadena larga de esfingomielina es de 40 ºC en comparación a -3 ºC para 1-
palmitoil, 2-oleil, fosfatidilcolina (POPC). Las insaturaciones que presenta la
estructura de las cadenas hidrocarbonadas impiden la linearidad de la molécula y
la cohesividad del empaquetamiento de estos lípidos. No obstante la temperatura
de transición de fase de fosfatidilcolina (PC) se incrementa dramáticamente de -3
a 49 ºC cuando la cadena monoinsaturada del ácido graso 18:1, es remplazada
por 18:0.
Los fosfolípidos y glicoesfingolípidos en membranas celulares existen en un
ambiente conteniendo varios tipos de moléculas como el colesterol y las proteínas
como se muestra en la figura 13. El colesterol esta intercalado entre cadenas
lipídicas, los cuatro anillos del colesterol permiten una pequeña flexibilidad
conformacional y no se acomoda fácilmente en la matriz hidrocarbonada. El grupo
3-hidróxi del colesterol permite a la molécula orientarse paralelamente a las
cadenas hidrocarbonadas de los lípidos, con el OH en la interfase lípido/agua.
Estas propiedades del colesterol provocan un decremento en la transición de fase
de gel a líquido cristalino tanto de esfingolípidos como de la bicapa lipídica.
(Ipsen, J. H., et al., 1987).

41
El empaquetamiento de los esfingolípidos específicamente esfingomielina y
fosfolípidos lleva a la membrana a una separación de fases. Las regiones de la
membrana ricas en esfingolípidos coexisten con dominios ricos en fosfolípidos. En
una región de la membrana, que esta enriquecida con esfingomielina y colesterol,
se puede esperar una tercera fase, una fase líquida ordenada (Io). Las cadenas
hidrocarbonadas de los lípidos están extendidos y firmemente empacados, como
en la fase gel pero tienen un alto grado de movilidad lateral.
La estructura de las membranas al parecer se mantiene unida o
estabilizada principalmente por interacciones hidrofóbicas entre ácidos grasos
saturados, esfingomielina y glicoesfingolípidos, estabilizados por moléculas de
colesterol intercaladas.
Como se sabe la monocapa externa de los dominios está selectivamente
enriquecida con glicoesfingolípidos; además, otro lípido de gran importancia en su
constitución es la ceramida (Václav, H.et al., 2005).
FL Aniónico
FL Neutro
Ác. Graso Insaturado
Ác. Graso Saturado
ColesterolFosfolípidoAnular
FosfolípidoAnular
Colesterol Anular
RaftTransmembranal
Raft
FL Aniónico
FL Neutro
Ác. Graso Insaturado
Ác. Graso Saturado
ColesterolFosfolípidoAnular
FosfolípidoAnular
Colesterol Anular
RaftTransmembranal
Raft
Figura 13. Representación esquemática de los dominios lipídicos de la membrana
plasmática. El nombre de fosfolípido anular se refiere a la primera molécula de
fosfolípido alrededor del colesterol o proteína. Y colesterol anular se le llama a la
primera molécula de colesterol alrededor de una proteína.

42
Los dominios probablemente existen en la fase líquida ordenada (Io) o en
un estado de propiedades muy similares dado las cadenas hidrocarbonadas
largas y saturadas de la esfingomielina. Los fosfolípidos naturalmente tienen una
cadena insaturada en la posición sn-2, y tienden a tener una menor afinidad por la
fase (Io) y están así excluidos de estos dominios.
Debido a que el colesterol puede promover separación de fases en
mezclas de fosfolípidos y esfingomielina (Sankaram, M. B., et al., 1991), se tiene
la hipótesis que los dominios pueden formarse en membranas celulares con
relativamente bajos niveles de colesterol. Más aún esto explica por que colapsan
los dominios y su función por la disminución de colesterol en las membranas. Es
importante considerar que los lípidos pueden estar asociados a cualquiera de los
tres estados: sólido gel, líquido ordenado, o líquido desordenado. Así, en la
membrana plasmática de las células los tres estados antes citados pueden
coexistir y se puede encontrar un tipo específico de lípido en más de un dominio.
No obstante, no todo el contenido de colesterol y glicoesfingolípido en la
membrana está asociado con los dominios.
Evidencia Experimental de la Existencia de Dominios Los dominios de lípido se detectaron por primera vez en fibroblastos como
grandes matrices proteicas insolubles a detergentes, estos complejos estaban
enriquecidos con glicoesfingolípidos, también se describe la formación de
microdominios de colesterol y glicoesfingolípidos que sirve como plataforma para
el transporte de lípidos y proteínas desde el aparato de Golgi a la membrana
plasmática (Lisanti, M. P. et al., 1988).
Por otra parte, se ha demostrado que el colesterol y la esfingomielina
interaccionan fuertemente en los microdominios de membranas. En estos
estudios se observó que una disminución en la concentración de esfingomielina
de la superficie de la membrana afecta a la biosíntesis y subsecuente localización
del colesterol. Estas observaciones y estudios anteriores ayudaron a argumentar
que es necesaria una interacción entre colesterol y esfingomielina para el proceso
normal de transporte intracelular (Scheek et al., 1997). Por otra parte se demostró
que un 33 mol % de colesterol induce la formación de la fase líquida ordenada en

43
vesículas mixtas de esfingomielina/PC, las vesículas con concentración entre 10 y
13 mol % de esfingomielina tienen una estructura similar a la de las membranas
plasmáticas (Ahmed, S. N., et al., 1997).
Una problemática de los dominios es que son pequeñas islas que no se
pueden observar fácilmente por técnicas comunes de microscopía en la superficie
de una célula intacta. No obstante, al parecer el marcaje de los dominios con
moléculas fluorescentes de anticuerpos o la toxina del cólera han dado resultados
positivos. Esto es debido a que la toxina del cólera tiene afinidad por glicolípidos
encontrados específicamente en los dominios. Esto a su vez nos indica que
probablemente el tamaño del dominios está por debajo de los límites de la
microscopía óptica (Hiscox S. et al., 2002).
Organización de los Dominios El término dominio lipídico incluye a los caveolae, estructuras generalmente
definidas tanto por su morfología y composición rica en caveolina como por sus
microdominios de líquido ordenado (Io). Se han descrito algunas características
comunes de la organización básica de los microdominios en las membranas.
Estas características se ilustran en la figura 14.
Dominio de proteínasDominio de proteínas
Figura 14. Los lípidos de los dominios en la fase líquido ordenada (Io) se
muestran rodeados por lípidos en la fase líquido desordenada (Id). Se muestra
también que el agrupamiento de los dominios de proteínas puede formar dominios
de mayor tamaño al coaleser (A) o pueden unirse al raft de mayor tamaño por
afinidad de las proteínas (B).

44
Comúnmente los dominios son ricos en colesterol y tienen un firme
empaquetamiento de esfingolípidos en la fase líquido ordenado (Io), y una
creciente afinidad por gangliósidos (Hagmann, J. et al., 1982). Además, existe una
variedad de proteínas que están asociadas a las membranas resistentes a
detergentes (MRD). Estas proteínas, las cuales incluyen caveolina-1, -2 y -3,
hemoaglutinina, entre otras, se usan frecuentemente como marcadores de
dominios (Anderson, R. G. W. et al., 2002). Resultados recientes han demostrado
que proteínas transmembranales son frecuentemente encontradas en los
dominios (Claas, C., et al., 2001). Estas proteínas intervienen en diversos
procesos incluyendo fusión de la membrana, agregación de plaquetas,
proliferación y activación de células T y B (Hemler, M. E., et al., 1996). La
superficie citoplásmica de las caveolae están cubiertas con caveolina, una familia
de proteínas entre 21 y 25 KDa que forman una capa de la membrana (Rothber,
K. G., et al., 1992). La caveolae está involucrada en la endocitosis, señal de
transducción, y contiene receptores de tirosina quinasa. Por otra parte, se ha
identificado una pequeña fracción que se denomina lipd shell (concha de lípidos)
(Anderson, R. G. W. et al., 2002). A estas estructuras lipídicas se les llama así por
que contienen una costra o concha de colesterol y esfingolípidos. La predicción de
tamaño de una concha de lípidos que contiene las proteínas membranales y 80
moléculas de colesterol-esfingolípido es aproximadamente de 7 nm. y existen
como unidades móviles en el plano de la membrana.
Función de los Dominios La asociación en la membrana de los lípidos ricos en colesterol y el firme
empaquetamiento de los dominios en la membrana permite la formación de
complejos de señalización. Los dominios contienen una alta concentración de
moléculas de señalización y se han clasificado como centros de señalización. Los
mecanismos por los cuales funcionan estos centros no están completamente
entendidos. Algunas moléculas de señalización como proteínas y lípidos se han
identificados en los dominios, estas incluyen las proteínas transmembranales,
EGF receptor (Furuchi, T. et al., 1998), receptor bradkinin B2, TCR, y BCR
(Haasemann, M. et al., 1998). Los lípidos, esfingomielina, ceramida, fosfoinositida

45
y diacilglicerol, así como la familia de las proteínas Gα, fosfolipasa Cγ, y
adenilatociclasa, entre otros, están asociados a los dominios (Lui, J. et al., 1997).
La asociación de los componentes de los procesos de la señal de
transducción con los dominios tiene importancia funcional y consecuencias en la
regulación. En el caso más simple el receptor puede ser un constituyente del
dominio listo para iniciar la señal en este sitio. Se ha propuesto que los complejos
de receptor-ligando se desplazan fuera del dominio dando como resultado una
baja regulación de la señal. Alternativamente el complejo receptor-ligando puede
desplazarse fuera del dominio asociado con otros complejos para iniciar la
respuesta.
El receptor se puede localizar fuera del dominio, pero el ligando inicia una
señal dentro del dominio seccionando y dirigiendo la señal. En todos estos
mecanismos, los dominios juegan un papel importante controlando y regulando la
señal. Queda por determinar si la composición única de lípido de los diferentes
tipos de dominios es capaz de dar la respuesta de los complejos. En este sentido,
existe evidencia que sugiere que en el caso del canal de potasio, la composición
de lípidos del dominio altera el comportamiento de las proteínas involucradas en
la señal. Ácidos grasos poliinsaturados inhiben la activación de linfocitos T por
desplazamiento de proteínas de los dominios en la membrana (Martens, J. R., et
al., 2000). Los dominios tomaron importancia tan pronto como se encontró que
están involucrados de manera importante en las etapas tempranas de la señales
de inmuno receptores (Václav, H. et al., 2005).
Dominios en Membranas de Plaquetas Las plaquetas presentan de cierta manera un escenario único al ser
sometidas a una translocación, estímulo dependiente de fosfolípidos. En una
simulación se eligieron: trombina, colágeno, ionomicina, fosfatidilserina y
fosfatidiletanolamina para realizar el proceso de translocación desde el interior al
exterior de la membrana. Esta translocación es necesaria en el proceso
hemostático para mediar la formación de un complejo de protrombinasa. Se ha
observado una correlación entre la respuesta de las plaquetas a estimulantes

46
tales como: ADP, epinefrina, trombina, tromboxano A2 y colágenos, y el contenido
de colesterol en la membrana.
No obstante, se ha observado un estímulo dependiente de translocación
del colesterol de la monocapa externa a la interna en una membrana (Boesze-
Battaglia et al., 1996). La base molecular del mecanismo responsable para estas
observaciones es desconocida, pero evidencia reciente sugiere que regiones de
la membrana ricos en colesterol dominios juegan un papel esencial.
En estudios recientes, se utilizó Triton X100 para aislar membranas de
plaquetas resistentes a detergente (DRM). Estas DRM son ricas en colesterol, con
41% del total del colesterol presente. La población de DRM fue heterogénea
consistiendo en vesículas en un intervalo de tamaños de 20 a 1000 nm de
diámetro basado en microscopía electrónica de barrido.
Estos trabajos sugieren que las DRM de plaquetas, en una manera análoga
a otros DRMs están involucradas en la señal de transducción. En trabajos
recientes se ha incrementado la información para respaldar esta hipótesis y más
aún, se tienen criterios esenciales en la caracterización de DRM en plaquetas
específicamente, una descripción detallada de la composición de lípidos DRM de
las plaquetas.
En una serie de estudios (Gousset et al, 2002) se visualizan regiones
condensadas en membranas de plaquetas las cuales se proponen como
dominios. Dichos resultados fueron obtenidos usando la técnica de microscopía
de fluorescencia.
Propiedades Mecánicas de Membranas
Hasta la fecha se ha estudiado ampliamente la composición de la
membrana de células sanguíneas y otras membranas naturales, pero aún quedan
aspectos por estudiar en términos de la participación de sus componentes en las
sus propiedades de cohesión. El análisis de ácidos grasos en la membrana de
eritrocitos muestran que la cadena de los lípidos puede contener
aproximadamente seis dobles enlaces y que aproximadamente el 17% de los
ácidos grasos contienes entre 4 y 6 dobles enlaces (Cooper et al., 1970). En otras

47
membranas de células no plasmáticas tales como los conos de la retina los lípidos
poliinsaturados con al menos seis dobles ligaduras por molécula de lípidos son
relativamente más abundantes (Drenthe et al., 1980). Otro aspecto de la cohesión
en bicapas de gran interés es la dependencia respecto a las insaturaciones
presentes en las cadenas hidrocarbonadas de los fosfolípidos y la influencia del
colesterol en estos lípidos.
Una característica importante del colesterol es que afecta fuertemente la
parte superior de la cadena hidrocarbonada de los lípidos dejándolos menos
disponibles para el cambio de conformación, mientras el centro de la bicapa
generalmente está mas desordenado (McIntosh, 1978). La interacción del
colesterol con los fosfolípidos incrementa la cohesión en la membrana y en
consecuencia su rigidez, por otra parte disminuye su permeabilidad al agua
(Evans, et al 1987).
El colesterol en las membranas artificiales fabricadas con un lípido y
aproximadamente 50% molar de colesterol afecta el comportamiento de las fases
dependientes de la temperatura (Needham, et al 1988).
La cantidad máxima de colesterol que puede ser incorporado en una
membrana lipídica es de aproximadamente 50% en mol. No obstante el límite de
solubilidad del colesterol en una membrana lipídica en fase líquida puede mostrar
una sutil dependencia respecto a la estructura molecular de los lípidos (Needham,
D. et al., 1990).
Estudios en monocapas han mostrado que el área por molécula se
incrementa con el número de dobles enlaces presentes (Demel, et al. 1972).
Moléculas de lípidos en las cuales los dobles enlaces se encuentran solamente en
una de sus cadenas muestran una reducción en el área molecular media cuando
están combinados con colesterol. Sin embargo el efecto de condensación del
colesterol es virtualmente eliminado cuando los lípidos contienen más de dos
insaturaciones en configuración cis por cadena. Estas mediciones se realizaron
en monocapas a una presión de 12 dyn/cm, (Evans y Waugh, 1977). En este
sentido toma interés examinar estos sistemas de membranas altamente
insaturados. Así pues, aunque se sabe mucho sobre sistemas de membranas

48
modelo compuestos de fosfolípidos y colesterol, quedan algunos aspectos
importantes para investigación como lo son las propiedades mecánicas.
Desde el punto de vista de la ciencia de materiales, las membranas de
lípidos se pueden describir en términos de sus propiedades físicas como se
muestra en la figura 15. En esta gráfica típica de esfuerzo vs deformación, se
traza la tensión τ producida por la aplicación de la succión con micropipetas
contra el incremento relativo en el área α de la membrana de la vesícula. Un
incremento en la tensión de la membrana causa un incremento lineal en el área
de la vesícula y se muestra una recuperación perfectamente reversible (elástica)
del área de la membrana al disminuir la tensión. La pendiente de la línea Δτ/Δα�
es el módulo elástico de expansión de área K. Posteriormente al ir incrementando
la tensión en la membrana se produce la falla y la lisis de la vesícula a un
deformación crítica del área y tensión crítica, dando� αc y τlis (τlis =� Kαc).
Figura 15. Gráfica de esfuerzo vs deformación por aspiración con micropipetas de
vesículas de lípidos (SOPC/CHOL 62/38).
Se han usado varias técnicas para cuantificar la elasticidad mecánica en
bicapas lipídicas, siendo las de mejores resultados: la técnica de succión con
micropipetas (Know W. and Evans E., 1981; Evans E. and Needham D., 1987),
microscopía de correlación de fotones y la dispersión dinámica de luz para
K
αc
τ lis Lisis de vesícula
Rigidez Tf
τ din/cm
5
10
15
20
α0.00 0.01 0.02 0.03 0.04 0.05 0

49
vesículas pequeñas bajo presión osmótica (Rutkowski et al., 1991; Hallett et al.,
1993), microscopía electrónica de criofractura de vesículas bajo presión osmótica
(Miu et al., 1993), difracción de rayos x de multicapas deshidratadas (Koening et
al., 1997). De estas técnicas solo el método de micropipetas puede ser usado
para medir la expansión de una bicapa con una resolución mejor a 0.1% en
cambios relativos de área y verificar la reversibilidad de la elasticidad (Rawicz W.
et al., 2000) es decir, que con está técnica se puede medir la reversibilidad de la
elasticidad de las membranas en el proceso de succión de vesículas con
micropipetas ya que al ir disminuyendo la tensión τ� en la membrana su área se
va recuperando de manera lineal manteniendo la misma relación Δτ/Δα, siempre y
cuando no se haya llegado al límite de su elasticidad o punto de falla (lisis). En
contraste a las propiedades elásticas del área, la pequeña rigidez de curvatura de
bicapas se ha derivado tradicionalmente de un detallado análisis de fluctuaciones
térmicas en la forma de las vesículas (Schneider et al., 1984). No obstante el
método de presurización con micropipetas también se puede usar para cuantificar
el módulo de curvatura en vesículas unilamelares; lo cual consiste en medir la
tensión de la membrana ocasionada por las fluctuaciones térmicas de la bicapa
(Evans E. and Rawicz W., 1990).


















