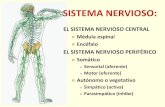
Sistema nervioso - sld.cu
32
Entre las propiedades características y exclusivas de los seres vivos está la capacidad, más o menos evi- dente, de percibir los estímulos, los cambios de las con- diciones ambientales, y de reaccionar ante estos, es decir, de presentar una sensibilidad o irritabilidad. Estas pro- piedades son algunos de los elementos que permiten distinguir a los seres vivos de los cuerpos inanimados. Filogenia del sistema nervioso Línneo expresaba: Los vegetales crecen y viven, los animales crecen, viven y sienten. Aunque hay ani- males, como las esponjas, que no presentan reacciones evidentes y vegetales, como la mimosa púdica (vergon- zosa) que demuestra mayor sensibilidad que algunos animales. Se señalan brevemente algunos elementos filogenéticos (Fig. 15.1): 1. En los protozoos, el protoplasma ejecuta todas las funciones vitales. 2. En los espongiarios, que son animales pluricelulares, cuyo tipo de organización es el más simple de los metazoos, el sistema nervioso está dado por células epiteliales del epitelio de revestimiento; las que se contraen como respuesta a cambios de presión o composición del agua que las rodea, y reciben el nombre de porositos. Este tipo de sistema nervioso tan primitivo es denominado neuroide y está repre- sentado en el hombre por el epitelio ciliado del apa- rato respiratorio. 3. Los celenterados, están representados por dos ti- pos diferentes de organización: el pólipo y la medu- sa, la primera más primitiva y sencilla, la hidra de Capítulo 15 Sistema nervioso agua dulce, y la segunda con mayor organización. Se observa un nuevo eslabón en el desarrollo del sistema nervioso; algunas células simples, se espe- cializan convirtiéndose en muy irritables a ciertos estímulos y siendo capaces de transmitir su irritabi- lidad a otras células especializadas. En las formas más avanzadas de los celenterados aparece un ter- cer eslabón en la evolución del sistema nervioso; la célula receptora, la cual envía prolongaciones subepiteliales que se ramifican formando un plexo de fibras entre las que se encuentran otras células, las intercaladas, que envían a su vez una prolonga- ción hacia la célula ejecutora. 4. En los platelmintos ya se observan puntos fotosensitivos en la cabeza, órganos sensoriales dis- puestos en hileras, y se alcanza, por tanto, la cefalización. El paso siguiente es la segmentación, que se aprecia en los anélidos y que alcanza grados de especialización en los artrópodos. 5. En los cordados primitivos ya aparece la formación del tubo neural. 6. En los vertebrados menos evolucionados hay un marcado desarrollo de la médula espinal y evolutivamente aparecen los plexos nerviosos. 7. En especies más evolucionadas, comienza el desarrollo del extremo cefálico con la diferen- ciación de sus distintas porciones. 8. En los mamíferos, las funciones nerviosas son re- guladas por la corteza cerebral. 9. El sistema nervioso humano presenta una mayor complejidad que el de todas las especies preceden- tes y repite en su desarrollo embrionario algunas etapas ancestrales.
Transcript of Sistema nervioso - sld.cu

Entre las propiedades características y exclusivasde los seres vivos está la capacidad, más o menos evi-dente, de percibir los estímulos, los cambios de las con-diciones ambientales, y de reaccionar ante estos, es decir,de presentar una sensibilidad o irritabilidad. Estas pro-piedades son algunos de los elementos que permitendistinguir a los seres vivos de los cuerpos inanimados.
Filogenia del sistema nerviosoLínneo expresaba: Los vegetales crecen y viven,
los animales crecen, viven y sienten. Aunque hay ani-males, como las esponjas, que no presentan reaccionesevidentes y vegetales, como la mimosa púdica (vergon-zosa) que demuestra mayor sensibilidad que algunosanimales.
Se señalan brevemente algunos elementosfilogenéticos (Fig. 15.1):
1. En los protozoos, el protoplasma ejecuta todas lasfunciones vitales.
2. En los espongiarios, que son animales pluricelulares,cuyo tipo de organización es el más simple de losmetazoos, el sistema nervioso está dado por célulasepiteliales del epitelio de revestimiento; las que secontraen como respuesta a cambios de presión ocomposición del agua que las rodea, y reciben elnombre de porositos. Este tipo de sistema nerviosotan primitivo es denominado neuroide y está repre-sentado en el hombre por el epitelio ciliado del apa-rato respiratorio.
3. Los celenterados, están representados por dos ti-pos diferentes de organización: el pólipo y la medu-sa, la primera más primitiva y sencilla, la hidra de
Capítulo 15
Sistema nervioso
agua dulce, y la segunda con mayor organización.Se observa un nuevo eslabón en el desarrollo delsistema nervioso; algunas células simples, se espe-cializan convirtiéndose en muy irritables a ciertosestímulos y siendo capaces de transmitir su irritabi-lidad a otras células especializadas. En las formasmás avanzadas de los celenterados aparece un ter-cer eslabón en la evolución del sistema nervioso; lacélula receptora, la cual envía prolongacionessubepiteliales que se ramifican formando un plexode fibras entre las que se encuentran otras células,las intercaladas, que envían a su vez una prolonga-ción hacia la célula ejecutora.
4. En los platelmintos ya se observan puntosfotosensitivos en la cabeza, órganos sensoriales dis-puestos en hileras, y se alcanza, por tanto, lacefalización. El paso siguiente es la segmentación,que se aprecia en los anélidos y que alcanza gradosde especialización en los artrópodos.
5. En los cordados primitivos ya aparece la formacióndel tubo neural.
6. En los vertebrados menos evolucionados hay unmarcado desarrollo de la médula espinal yevolutivamente aparecen los plexos nerviosos.
7. En especies más evolucionadas, comienza eldesarrollo del extremo cefálico con la diferen-ciación de sus distintas porciones.
8. En los mamíferos, las funciones nerviosas son re-guladas por la corteza cerebral.
9. El sistema nervioso humano presenta una mayorcomplejidad que el de todas las especies preceden-tes y repite en su desarrollo embrionario algunasetapas ancestrales.

214
Embriología especial
Inducción en la formacióndel sistema nervioso
La interrelación inductiva, entre la notocorda y electodermo, en el desarrollo del sistema nervioso fuedescrita en los primeros años del siglo XX, medianteexperimentos realizados en anfibios. Sin embargo, enlos efectuados en mamíferos se ha demostrado que losmecanismos fundamentales de la inducción neural sonsimilares en los vertebrados.
Varias investigaciones experimentales han com-probado, que la inducción no es un simple mecanismogeneral, sino que existe una gran especificidad regio-nal. Esto implica que existen inductores que estimulanlas porciones anteriores o cefálicas, y otros que estimu-lan las porciones posteriores o caudales en el desarrollode las diferentes estructuras nerviosas.
En la actualidad se han identificado moléculas es-pecíficas relacionadas con la inducción neural en aves
y mamíferos. Dos moléculas de señales, noggin ychordin secretadas por la notocorda son agentesinductores. Inicialmente se pensó que estas moléculasactuaban de manera directa sobre el ectodermo dorsal,pero investigaciones en anfibios han mostrado que es-tos agentes inductores actúan bloqueando la acción deun inhibidor del ectodermo dorsal, el BMP-4 (proteínamorfogenética ósea), cuya expresión evita la formaciónde la placa neural. Esta interacción entre ambos tiposde moléculas lleva al ectodermo suprayacente a lanotocorda, a convertirse en tejido neural, al bloquearsela expresión de BMP-4 por las moléculas noggin ychordin. Este proceso es el primer paso en la forma-ción del sistema nervioso. El segundo es laregionalización del sistema nervioso central (SNC), quese refiere a la subdivisión de este en las diferentes re-giones en sentido craneocaudal.
Las moléculas noggin y chordin, además de in-tervenir en la formación del tejido neural, son responsa-bles de la formación de las estructuras neurales máscraneales. Si además se añade la presencia de FGF-8,entonces se forman las estructuras caudales, como elcerebro posterior y médula espinal. Se afirma, que elmesodermo tiene un importante efecto en el proceso decaudalización del ectodermo neural.
La placa precordal tiene gran importancia y espe-cificidad en la formación del cerebro anterior. Esta pla-ca por sí sola no tiene efecto sobre el ectodermo paraformar el tejido neural; sin embargo, una vez producidala inducción por las moléculas noggin y chordin, laplaca precordal desempeña una función importante enla regionalización del cerebro anterior del SNC.
Mediante la acción de moléculas de señal comoLim-1 y una proteína denominada cerberus, elmesodermo de la placa precordal ejerce su influenciapara la formación inicial del cerebro anterior en la placaneural, al mismo tiempo que la parte rostral de la cabe-za comienza a formarse.
Desarrollo y morfogénesis del tuboneural
El primer esbozo del SNC es la placa neural (Fig. 15.2),la cual aparece al inicio de la tercera semana en formade un engrosamiento del ectodermo, a nivel de la líneamedia y cefálicamente con respecto a la fosita primiti-va. En este proceso, la notocorda actúa como inductorprimario. Ya formada la placa neural, las células adop-tan formas primero cuboideas y, más tarde, cilíndricas.Sus bordes laterales se elevan y forman los plieguesneurales.
Fig. 15.1. Algunos estadios en la evolución de la funciónnerviosa.

215
Capítulo 15. Sistema nervioso
Los pliegues neurales se elevan más, el centro sedeprime y forma el surco neural, que ya es notorio en elembrión de 17 días, los pliegues se acercan en la líneamedia y por último se fusionan formando el tubo neural.
Esto ocurre en el periodo de siete somitas, y lafusión se inicia a nivel de la cuarta somita. La uniónprogresa en dirección cefálica y caudal. El tubo neuralqueda abierto por sus dos extremos; dichos orificios reci-ben el nombre de neuroporo anterior y posterior (Fig. 15.3).El cierre del neuroporo anterior se efectúa en el estadio de20 somitas (25 días) y el del posterior, en el de 25 somitas(27 días).
El sistema de trasplante pollo-codorniz se ha usa-do para descubrir, qué partes de los pliegues neuralesdan origen a las diferentes porciones del cerebro. Enestos experimentos, una pequeña porción del pliegueneural del pollo es reemplazado con la porción homólogade una codorniz, y las estructuras que se desarrollan delas células de la codorniz se identifican por sus nucléolosprominentes.
Después de cerrarse el tubo neural, su porción me-dia y el extremo caudal son estrechos y regulares, dandoorigen a la médula espinal. La parte anterior, más ancha,forma después el encéfalo. Esta zona adquiere un gran
desarrollo y se levanta sobre el nivel del resto del discoembrionario, proyectándose hacia delante del plieguecefálico del embrión, recibiendo el nombre de procesocefálico, que representa la etapa de vesícula única.
A finales de la tercera semana, antes que laneuralización comience, aparecen tres dilataciones en estazona anterior, denominadas vesículas encefálicas prima-rias, debido a la formación de dos surcos que señalan lasconstricciones del tubo neural. Estas vesículas son:
1. Prosencéfalo o cerebro anterior.2. Mesencéfalo o cerebro medio.3. Rombencéfalo o cerebro posterior.
Al tiempo que aparecen dos acodaduras o curva-turas: curvatura cervical, en la unión del cerebro poste-rior y la médula espinal, y curvatura cefálica situada enla región del mesencéfalo, de tal manera, la tercera se-mana se corresponde con la etapa de tres vesículas.
Ya en la quinta semana, las tres vesículas cere-brales primarias que continúan flexionándose, se trans-forman con la aparición, en el rombencéfalo, de unatercera curvatura de concavidad dorsal, denominadacurvatura pontina y que divide a este último en dos ve-sículas cerebrales secundarias: mielencéfalo ymetencéfalo. La primera dando origen al bulbo raquídeoo médula oblongada, y la segunda, a la protuberancia y elcerebelo. El límite entre estas dos porciones está marca-do por un pliegue, denominado pliegue protuberancial, eneste caso la quinta semana se corresponde con la etapade cinco vesículas (Fig. 15.4).
Fig. 15.2. Formación del tubo neural en cortes transversales.
Fig. 15.3. Cierre del tubo neural.

216
Embriología especial
Neuralización
Neuralización primariaEs el plegamiento de la placa neural para formar
el tubo neural. Está asociada con cambios característi-cos en la forma de las células neuroepiteliales, por locual estas cambian, de cuboidal, a una forma de cuña.Estos cambios se creen sean producidos por micro-túbulos intracelulares y microfilamentos, conducidos porfuerzas que son ambas intrínsecas y extrínsecas alneuroepitelio.
En los mamíferos, la situación es más complejaque en aves y anfibios, se ha mostrado, en el pollo, quelos cambios en la forma de las células neuroepitelialesresultan de interacciones inductivas con la futuranotocorda. La expansión de la matriz extracelular demesénquima influye en la elevación de los plieguesneurales. Los factores que son importantes en lamorfogénesis del sistema nervioso incluyen cambios enla forma celular, la migración de las células, la prolifera-ción diferencial y la muerte celular diferencial.
Neuralización secundariaLa porción caudal del tubo neural no surge por
fusión de los pliegues neurales, sino que se desarrollade una masa de células conocidas como la eminenciacaudal (o brote terminal) (Fig. 15.5. A). Después que elneuroporo caudal se ha cerrado, el tejido neural continúahacia abajo como cordón neural, dentro del cual se extien-de la cavidad ya formada del tubo neural (Fig. 15.5. B).Este proceso se denomina neuralización secundaria. El cualcomienza en el sitio del cierre del neuroporo caudal,que se encuentra a nivel de la somita 31, que corres-ponde al plano vertebral futuro de S2.
Fig. 15.4. A. Etapa de tres vesículas cerebrales o primarias. B.Etapa de cinco vesículas cerebrales o secundarias.
Fig. 15.5. Desarrollo de la porción caudal del tubo neural.
A inicios del segundo mes (quinta semana), en elprosencéfalo aparece la curvatura telencefálica, muyacentuada en el desarrollo humano, y que divide alprosencefálico en:
1. Telencéfalo o cerebro terminal, que a su vez pre-senta una parte media y dos evaginaciones latera-les, los hemisferios cerebrales.
2. Diencéfalo, de gran importancia por intervenir en laformación de los ojos, mediante la evaginación delas vesículas ópticas.
El mesencéfalo no se divide y está separadodel rombencéfalo por un surco profundo, el istmodel rombencéfalo o de His. Este último se diferen-cia a su vez en metencéfalo y mielencéfalo.
En resumen, de las tres vesículas cerebrales pri-marias se derivan cinco secundarias; que en dirección

217
Capítulo 15. Sistema nervioso
cefalocaudal son: telencéfalo, diencéfalo, mesencéfalo,metencéfalo y mielencéfalo.
La luz cada vez más pequeña de la médulaespinal, conducto ependimario, mantiene continui-dad con las cavidades de las vesículas encefálicas.Así, la cavidad del rombencéfalo se correspondecon el cuarto ventrículo; la del diencéfalo con el ter-cer ventrículo; y las de los hemisferios cerebralescon los ventrículos laterales. El tercero y cuartoventrículo comunican entre sí por el acueducto deSilvio, que corresponde a la luz del mesencéfalo muyestrecha. Los ventrículos laterales comunican conel tercer ventrículo a través de los agujeros deMonro. Todo este sistema intercomunicado de lascavidades ventriculares, permite la circulación nor-mal del líquido cefalorraquídeo formado en los plexoscoroideos.
La mayor parte del tubo neural se sitúadorsalmente con respecto a la notocorda, pero su ex-tremidad craneal (proceso cefálico), se proyecta másallá de esta, y por encima del estomodeo. Durante untiempo, su porción caudal está en comunicación conel intestino posterior por medio de una pequeña aber-tura, el canal neuroentérico, que representa alblastoporo y al canal notocordal (Fig. 15.6). La dife-renciación del tejido neural de la porción caudal de lamédula espinal no se completa, hasta que ha termina-do el periodo de formación de las somitas.
gruesa denominada capa neuroepitelial o neuroepitelio(Fig. 15.7). Estas células se extienden por todo el gro-sor de la pared y forman un grueso epitelioseudoestratificado, y están conectadas entre sí porcomplejos de unión.
Histogénesis de sistema nerviosoA partir de estudios realizados en embriones de
anfibios se demostró que, el número de células queforman la placa neural es comparativamente pequeño(alrededor de 125 000), y que esta cifra se modificapoco durante la formación de tubo neural. Sin embar-go, después que el tubo neural se cierra, la prolifera-ción celular se acentúa de manera marcada y, en cor-to plazo, la capa de células epiteliales cilíndricas queformaban la placa neural se transforma en una epitelial
Fig. 15.6. Formación del canal neuroentérico.
Fig. 15.7. Evolución del neuroepitelio del tubo neural. A. Antesdel cierre. B. Después del cierre.
Las células neuroepiteliales, cuando inician la di-visión mitótica se acortan hasta quedar sus núcleospróximos a la luz del tubo neural. Durante el cierre deeste último, el huso acromático de estas divisiones sedispone paralelo a la superficie interna del neuroepitelio, locual hace que las dos nuevas células neuroepiteliales, asíformadas, se dispongan una junto a la otra (Fig. 15.8. A).Después, al producirse nuevas divisiones, el huso dedivisión se dispone perpendicular, una de las célulashijas permanece y continúa como célulaneuroepitelial, y la otra se desprende y se dirige haciala superficie externa del neuroepitelio, donde se di-ferencian en otro tipo celular. Estas son las célulasnerviosas primitivas o neuroblastos que rodean ex-ternamente al neuroepitelio formando la capa delmanto, las cuales pierden la capacidad de dividirse ypor último dan origen a la sustancia gris de la médulaespinal (Fig. 15.8. B).

218
Embriología especial
Células del tejido nerviosoLos neuroblastos, inicialmente poseen una prolon-
gación que se extiende hacia la luz y recibe el nombre deneuroblasto monopolar (Fig. 15.9). Luego, esta prolon-gación desaparece y los neuroblastos adquieren formaredondeada u ovoide y se denominan neuroblastosapolares. Al continuar la diferenciación aparecen dos nue-vas prolongaciones citoplasmáticas en los lados opuestosdel cuerpo celular, y se forma el neuroblasto bipolar. Laprolongación de un extremo de la célula se alarga y daorigen al axón primitivo, el otro extremo presenta peque-ñas arborizaciones citoplasmáticas que son las dendritasprimitivas. Ahora la célula se denomina neuroblastomultipolar, que más tarde origina la neurona definitiva.
El mecanismo de orientación del crecimiento axonaly de la migración neuronal durante el desarrollo del sis-tema nervioso se debe al quimiotropismo, planteado porCajal en 1890, y confirmado a partir de 1985 hasta elmomento actual, por trabajos experimentales que demues-tran la presencia de moléculas quimiotrópicas pertene-cientes a las familias de las netrinas y de las semaforinas.
Las neuroglias inician su diferenciación al concluirla producción de neuroblastos, las células neuroepitelialescomienzan a formar las células de sostén primitivas oglioblastos los cuales emigran, desde la capa neuroepitelial,a la capa del manto (Fig. 15.10), donde se diferencian enastrocitos protoplasmáticos y astrocitos fibrosos.
Fig. 15.8. División celular en elneuroepitelio. A. Antes del cierredel tubo neural. B. Después delcierre.
Fig. 15.9. Formación de neuronas multipolares.

219
Capítulo 15. Sistema nervioso
La zona periférica del tubo neural, que está des-provisto de núcleos de células nerviosas en desarrollopero que contiene las células de neuroglia denominadasoligodendroglias, encargadas de formar las vainas demielina de los axones ascendentes y descendentes, ylas prolongaciones de las células nerviosas en el siste-ma nervioso central, se denomina capa marginal.
Otro tipo más de glia denominada microglia y úni-ca de origen mesenquimatoso aparece en la segundamitad del desarrollo fetal, la cual tiene propiedadesfagocíticas.
Por último, las células neuroepiteliales al dejar deproducir neuroblastos y glioblastos, se diferencian enlas células ependimarias que recubren las cavidadesventriculares del sistema nervioso central y la cubiertade los plexos coroideos.
La capa del manto se convierte en la sustanciagris, y la capa marginal en la sustancia blanca del SNC.Este esquema de desarrollo descrito en las células ner-viosas y las neuroglias, es el patrón característico de lamédula espinal, aunque se puede aplicar, con algunasvariaciones, al encéfalo.
Además de las células originadas en elneuroepitelio, hay otros tipos celulares que se originande las denominadas crestas neurales. Estas estructurasaparecen justamente al cerrarse el tubo neural, un gru-po de sus células se separan de la parte dorsal de estasy comienzan a distribuirse por todo el cuerpo del em-
brión, por lo que algunas veces se le llama a las crestasneurales la cuarta hoja embrionaria del cuerpo.
Las crestas neurales se originan como una faja deectodermo especializado que bordea por cada lado a laplaca neural durante su plegamiento, y está interpuesta,entre esta última, y el ectodermo somático. Sus célulasse denominan células de la cresta neural.
Al producirse el cierre de los pliegues neuralespara formar el tubo neural, y al fusionarse el ectodermosomático situado dorsalmente, la cresta neural de cadalado aparece como una columna de células aisladas si-tuadas a lo largo de la cara dorsal del tubo neural. Des-pués, sus células migran en dirección ventrolateral parasituarse a lo largo de la cara dorsolateral del tubo ex-tendiéndose, desde la futura región mesencefálica, has-ta el nivel de las somitas caudales. Algunas células ori-ginan los ganglios sensitivos o ganglios de la raíz dorsalde los nervios raquídeos.
Los neuroblastos de los ganglios sensitivos pre-sentan dos prolongaciones, una que penetra en la por-ción dorsal del tubo neural (médula espinal), termina enel asta dorsal o ascienden por la capa marginal hastalos centros cerebrales superiores. Estas prolongacio-nes reciben el nombre de raíz sensitiva dorsal del nervioraquídeo.
Otras prolongaciones se unen a las fibras de laraíz motora ventral y, de tal modo, contribuyen a la for-mación del tronco del nervio raquídeo o espinal. Por lo
Fig. 15.10. Diferenciación de las célulasneuroepiteliales.

220
Embriología especial
tanto, los neuroblastos de los ganglios sensitivos danorigen a las neuronas de la raíz dorsal.
Las rutas de migración de las células de la crestaneural, de las diferentes partes de la placa neural, sehan mapeado por estudios de células marcadas en em-briones de aves y rata. Estas investigaciones utilizanmarcadores naturales o artificiales para seguir las víasde migración y de linaje celular. Se argumenta que uncomponente en la migración celular es principalmentepasivo, sin embargo, parece ser que las células de lacresta también migran de manera activa y que estasestán guiadas por moléculas en la matriz extracelular,tales como la fibronectina y los glicosaminoglicanos.
Las células de la cresta neural migran a muchaslocalizaciones específicas en el cuerpo, donde se dife-rencian en una variedad marcada de estructuras. Estetipo de tejido inusual surgió, primeramente, durante laevolución temprana de los vertebrados. La migraciónse produce en dirección cefalocaudal, desde elmesencéfalo, a la parte final del tubo neural espinal.
Los principales derivados, descritos brevemente son:1. Cresta neural cefálica: ganglio parasimpático del
III nervio craneal; tejido conectivo alrededor de losojos y nervio óptico; músculos de la pupila y delcuerpo ciliar; mesénquima craneal cefálico al nivelde mesencéfalo, piamadre y aracnoides, arcosfaríngeos de cabeza y cuello (varios cartílagos yhuesos de la nariz, cara, oído medio y cuello). Alnivel de mesencéfalo y rombencéfalo también for-ma la dermis; músculo liso y grasa de la cara yporción ventral del cuello; los odontoblastos de losdientes en desarrollo; las células de la cresta en laporción más caudal del rombencéfalo origina las cé-lulas C del tiroides; las neuronas y células gliales delos ganglios parasimpáticos de los nervios cranealesVII, IX y X; neuronas y células gliales de los gangliossensitivos craneales V, VII, VIII, IX y X. En el sis-tema olfatorio, en cambio, sus neuronas más distalesse originan en el ectodermo superficial.
2. Cresta neural occipital y espinal: el sistema nerviosoperiférico del cuello, tronco y extremidades incluyelos tres tipos de neuronas periféricas siguientes:a) Neuronas sensitivas periféricas, cuyos cuerpos
residen en los ganglios de las raíces dorsales ex-plicadas. La supervivencia y la diferenciación deestos ganglios puede depender de una pequeñaproteína denominada factor neurotrópico deriva-do del cerebro (BDNF) segregada por el tuboneural adyacente, así como, otros factores decrecimiento que incluyen miembros de la familiaproteína morfogenética ósea (BMP).
Existen siete pares cervicales de glanglios de lasraíces dorsales, 12 torácicos, 5 lumbares, 5 sa-cros y 1 coccígeo. El más craneal cervical apa-rece en el día 28 y los demás en los siguientesdías en sucesión craneocaudal.
b) Las neuronas parasimpáticas posganglionares delas vísceras se derivan de células de la cresta neuralque migran a las paredes del corazón, estómago yla vejiga. Los cuerpos celulares de estas neuronasresiden en los ganglios parasimpáticos periféricosque proveen inervación motora parasimpática. Losganglios parasimpáticos periféricos de la pared deltubo digestivo, denominados ganglios entéricos, seconectan al SNC por axones que transcurren, yasea en el nervio vago o en los nervios esplácnicospélvicos de los niveles sacros 2; 3 y 4.
c) La cadena ganglionar simpática: algunas célulasde la cresta neural de la médula espinal migran auna zona situada ventralmente al futuro gangliode la raíz dorsal, donde forman la cadenaganglionar del sistema simpático autónomo.
Al contrario de la raíz dorsal, la cadena ganglionarno depende del BDNF para la supervivencia, sinoque obedece a otros factores de crecimiento pare-cidos a la insulina (IGE) y el factor de crecimientonervioso (NGF). Estas neuronas motoras simpáti-cas centrales están localizadas 12 en los nivelestorácicos y en los 3 primeros lumbares.
Además, células de la cresta neural cervical se con-gregan junto a las ramas mayores de la aorta dorsalpara formar los ganglios prevertebrales o preaórticossituados en la base de la arteria celiaca, otros gangliosmás difusos se desarrollan en asociación con la arte-ria mesentérica superior, las arterias renales y la ar-teria mesentérica inferior. Estas últimas formadas porcélulas de la cresta neural torácica y lumbar.
3. Cresta neural de la médula espinal: estructuras noneuronales. Células de la cresta neural espinal for-man la piamadre y la aracnoides (cubiertasmeníngeas) de la médula espinal; así como célulasgliales de los ganglios espinales. Algunas de las cé-lulas de la cresta neural se diferencian en célulasde Schwan, las cuales forman las vainas de mielina(neurilema) de los nervios periféricos. También lascélulas de la cresta neural espinal se diferencian enlas células cromafines neurosecretoras de la mé-dula suprarrenal, y en células neurosecretoras delcorazón y los pulmones. Las células de la crestaneural también dan lugar a los melanocitos (célulaspigmentarias) y contribuyen igualmente a la forma-ción de las almohadillas endocardiacas y del tabi-que troncoconal en el corazón.

221
Capítulo 15. Sistema nervioso
MielinizaciónEs un proceso por el cual las fibras nerviosas
periféricas y la mayoría del SNC adquieren vainas es-pecializadas que contienen lecitina, y reciben el nombrede vainas de mielina. Las vainas de los nerviosperiféricos, tanto las fibras sensitivas, como las moto-ras, se forman a partir de células de neurilema (célu-las de Schwan) que se enrollan alrededor del axón deuna neurona, y forman cadenas que siguen el trayectode las fibras nerviosas. En los nervios periféricos, lavaina de mielina está interrumpida por constriccionesregulares sin mielina denominadas nudos de Ranvier.Esta disposición facilita el incremento de la velocidadde transmisión del impulso nervioso.
En el SNC las células de Schawn son reemplaza-das por los oligodendrocitos (productores de mielina enel SNC), pero el patrón de mielinización en este casopresenta algunas diferencias con la periférica, ya quecada oligodendrocito forma mielina en varios axones ala vez y las de Schwan solo en uno.
En el humano, la mielinización comienza en elcuarto mes de la vida fetal (100 mm) pero no se com-pleta hasta el segundo o tercer año después del naci-miento; empieza cerca de la célula nerviosa y de allíse extiende a lo largo de la fibra nerviosa, hasta cercade su terminación.
La mielinización aparece primero en los nerviosque funcionan más precoces o que son, filogenética-mente, más antiguos. En el momento que alcanzan sumadurez funcional, los troncos nerviosos estánmielinizados por completo.
Mielinización de la médula espinalLa porción cervical de la médula espinal es la
primera en mielinizarse y a partir de esta, el procesose extiende en dirección caudal. Las fibrasintersegmentarias que están en contacto con la co-lumna gris anterior son las primeras en mielinizarsey, más tarde, le siguen las fibras comisurales anterio-res. Las fibras de la raíz ventral lo hacen antes quelas de la dorsal. Las columnas posteriores de sustan-cia blanca comienzan su mielinización en el sexto mesde vida fetal, los haces espinocerebrales yespinotalámicos en el séptimo mes, y las fibras mo-toras descendentes (piramidales y rubroespinales) co-mienzan a mielinizarse en el momento del nacimiento
pero no se completa hasta el segundo año de vidaposnatal.
Mielinización del encéfaloEn los nervios craneales del rombencéfalo y del
mesencéfalo, la mielinización comienza en el sextomes de vida fetal. Las fibras motoras se mielinizanantes que las sensitivas. El nervio vestibular es el pri-mer nervio sensitivo que se mieliniza. La mielinizacióndel nervio óptico comienza en el feto a término. El hazlongitudinal medio es el primero de los tractos cere-brales que se mieliniza, y las conexiones cerebrales enel octavo mes. La corteza cerebral se puede clasificaren numerosas zonas mielogenéticas diferentes, segúnel momento en que sus fibras se mielinizan.
El momento y la secuencia en que aparece lamielinización del SNC tienen un gran valor para el estu-dio del origen, de la evolución y de la terminación de losgrupos de fibras nerviosas dentro del SNC y es de apli-cación fundamental en la clínica neurológica.
Médula espinalEn embriones del periodo somítico (25 pares)
de 28 a 30 días, el canal se ha cerrado, y el tuboneural formado es una estructura dorsoventral consus caras laterales engrosadas y las paredes de susbordes dorsal y ventral, adelgazadas; estas últimasreciben el nombre de placa de techo y de piso, res-pectivamente (Fig. 15.11).
Con el aumento de los neuroblastos en la capa delmanto, a cada lado del tubo neural se producen dosengrosamientos, uno ventral y otro dorsal. Los ventraleso placas basales, que incluyen a las células motoras delas astas ventrales (columnas grises anteriores o astasanteriores), forman las áreas motoras de la médulaespinal; mientras que los dorsales o placas alares, for-man la áreas sensitivas, compuestas por neuronas deasociación, que dan lugar a las columnas grises poste-riores o astas posteriores. Los engrosamientos ventralesson más notorios, lo que provoca el estrechamiento delconducto central en esta zona, y la aparición de un sur-co en la cara lateral que separa la zona ventral de ladorsal, el surco limitante.
Entre las dos áreas señaladas se sitúan un grupode neuronas que forma después el asta lateral o inter-media, más pequeña, la cual contiene neuronas del sis-tema nervioso autónomo.

222
Embriología especial
Las placas del techo y del piso de la médulaespinal permanecen delgadas; están integradas porcélulas ependimarias y neuroglia, no se formanneuroblastos y sirven de paso a prolongaciones ner-viosas dispuestas en fibras comisurales.
Como consecuencia del desarrollo de las pla-cas basales a ambos lados y en dirección ventral,en la superficie externa de la médula se estableceel surco ventral, en el que se aloja la arteria espinalanterior. Las placas alares se dirigen hacia la líneamedia, donde se fusionan y forman el tabique medioposterior.
Durante el desarrollo y diferenciación de la mé-dula espinal se producen cambios de posición de lamedula, con respecto al crecimiento de la columnavertebral. Así, al comienzo del tercer mes (30 mm)se extiende hasta la región coccígea; el mesodermoque la rodea da origen a los huesos y cartílagos dela columna vertebral, que posteriormente crece enlongitud más rápido que esta, lo que conduce a queen el momento del nacimiento el extremo caudal deesta última quede situado al nivel de la tercera vér-tebra lumbar y permanezca una zona adelgazada enforma de filamento denominada filum terminalis,quedando unido al periostio de la primera vértebracoccígea.
En el adulto, la médula espinal termina a la altu-ra de L2-L3. El filum terminalis se halla rodeado de
Fig. 15.11. Evolución del desarrollo de la médula espinal.
Fig. 15.12. Evolución de la porción caudal de la médulaespinal.
fibras nerviosas que se extienden por el interior delcanal medular en la región lumbosacra, donde con-forman la cola de caballo (cauda equina).
La formación de la cola de caballo depende,fundamentalmente, de que los nervios raquídeos, alprincipio, salen por los agujeros intervertebrales máscercanos a su origen en la médula espinal, con unatrayectoria casi transversal, pero debido al creci-miento desigual entre la médula espinal y el raquis,los nervios raquídeos toman una dirección caudal,desde su segmento de origen, ahora más cefálico,hasta el agujero intervertebral por donde salen de lacolumna vertebral situado mucho más caudal. Laduramadre permanece unida a la columna vertebralen la región coccígea. Las características señaladasresultan de interés al realizar una punción lumbar, parano dañar la médula espinal (Fig. 15.12).

223
Capítulo 15. Sistema nervioso
En cuanto a la incidencia geneticomolecular deldesarrollo de la médula espinal, inicialmente en el es-tadio de placa neural se expresan los factores de trans-cripción PAX3, PAX7, MSX1 y MSX2. Este cuadrode expresión es alterado por Sonic hedgehog, Shhexpresado en la notocorda, así como las “proteínasmorfogenéticas óseas” (BMP4 y BMP7), expresadosen el ectodermo no neural en el borde de la placaneural. La señal de Shh reprime la expresión de PAX3y PAX7 y de MSX1 y MSX2; de este modo, en laregión ventral se forma una placa del piso, la cual tam-bién expresa Shh. La expresión de BMP4 y BMP7mantiene y regula a PAX3 y PAX7 en la mitad dorsaldel tubo neural, donde se forma después la placa alar.Estos factores de transcripción son requeridos para laformación de las células de la cresta neural en la cimade los pliegues neurales. El Shh producido por la pla-ca del piso inhibe la expresión de los genes PAX3 yPAX7 en la porción ventral de tubo neural.
Anomalías en el desarrollode la médula espinal
La mayoría de los defectos de la médula espinalocurren durante la segunda mitad del primer mes deldesarrollo, como resultado de la formación y cierre anor-mal de los pliegues neurales.
Defectos de tubo neural al nivelde la médula espinal
Las anormalidades resultantes, denominadas de-fectos del tubo neural, pueden implicar a las meninges,vértebras, músculos y a la piel.
Espina bífidaSe produce por la falta de unión de los arcos
neurales de las vértebras y no formación de la apófisisespinosa. Es una hendidura de los arcos vertebrales,que puede o no afectar al tejido neural vecino. La inci-dencia de defectos severos del tubo neural varía entrediferentes poblaciones.
Espina bífida ocultaEs un defecto de los arcos vertebrales que está
cubierto por la piel y frecuentemente no afecta al tejidoneural subyacente, por lo general se presenta en la re-gión lumbosacra.
Fig. 15.13. Malformaciones congénitas al nivel de la médulaespinal provocada por la espina bífida.
Espina bífida abiertaEn este caso el tejido neural, las meninges o ambos
protruyen a través de un defecto en los arcos vertebralesy piel para formar un saco herniario. La mayoría de laspersonas con esta anomalía presentan déficit neurológico.Cuando solo líquido y meninge protruyen a través deldefecto se le denomina, espina bífida con meningocele;en otros casos el tejido neural está incluido en el saco,espina bífida con mielomeningocele. En algunos, el tuboneural permanece aplanado y origina la espina bífida conmielosquisis o raquisquisis (Figura 15.13).
Los arcos vertebrales se pueden visualizar a fina-les del tercer mes de embarazo. La espina bífida sepuede diagnosticar prenatalmente por ultrasonido y pordeterminación de los niveles de alfafetoproteína en elsuero materno o en el líquido amniótico.
En la actualidad se puede corregir el defectomediante cirugía fetal in útero, aproximadamente alos siete meses de embarazo. El bebé se expone porcesárea, el defecto se repara y el infante se coloca denuevo en el útero.
En el origen de la mayoría de los defectos del tuboneural intervienen varios factores, por lo que esmultifactorial; la posibilidad de tener un niño con tal de-fecto aumenta, si existe descendencia previa con taldefecto.

224
Embriología especial
EncéfaloEl encéfalo es la porción del SNC que ocupa la
cavidad craneal y está conformada por el cerebro, elcerebelo, el istmo, la protuberancia y el bulbo raquídeo omédula oblongada. La porción más voluminosa corres-ponde al cerebro, que cubre a las demás porciones,excepto al nivel de la base. El desarrollo del cerebro tie-ne algunas características que lo hacen diferir sustan-cialmente de la médula espinal.
La importancia de las placas alares aumenta demanera progresiva, de tal forma, que en las vesículasmás cefálicas presentan un desarrollo mayor; al tiem-po, en que las células de estas placas emigran en senti-do ventral, dicha migración es más marcada en las ve-sículas más cefálicas. Paralelamente a esto, disminuyela importancia de las placas basales. Como consecuen-cia de todo ello el diencéfalo y el telencéfalo presentansolo células derivadas de las placas alares, que consti-tuyen toda su pared.
Además, el telencéfalo presenta un desarrollo muymarcado, crece lateralmente y origina los hemisferioscerebrales, como consecuencia de este crecimiento, cu-bre al diencéfalo. La migración de células de la capa delmanto hacia la superficie da lugar a la formación de lascortezas cerebral y cerebelosa. El mesencéfalo, que deinicio crece, queda al final del proceso oculto por los he-misferios cerebrales. La diferenciación de cada una deestas vesículas se describe a continuación (Fig. 15.14).
MielencéfaloCorresponde a la vesícula más caudal derivada
del rombencéfalo, se extiende, desde la médula espinal(al nivel del primer nervio cervical), hasta la curvaturapontina. Del mielencéfalo se origina el bulbo raquídeo omédula oblongada.
La pared dorsal del mielencéfalo o techo del cuar-to ventrículo se modifica notablemente por el crecimientosimultáneo de la separación, en sentido lateral, de lasplacas alares y la acentuación de la curvatura pontica(Fig. 15.15). Esto trae como consecuencia que, el te-cho del cuarto ventrículo se adelgace de manera consi-derable hasta quedar integrado solo por la capaependimaria, a partir de esos cambios se le denominavelo medular.
La cavidad del tubo neural al nivel del mesencéfaloforma la fosa romboidea o futuro cuarto ventrículo.
Vista de la cara dorsalEn la formación inicial del mielencéfalo se distin-
guen fácilmente el surco limitante, las placas alares ybasales, el suelo y el techo, elementos señalados en lamédula espinal.
Placa basalLas células de la placa basal del mielencéfalo se
diferencian en tres grupos de núcleos motores:1. Grupo medial o eferente somático.2. Grupo intermedio o eferente visceral especial.3. Grupo lateral o eferente visceral general.
Cada uno de estos grupos celulares origina las fi-bras motoras que integran diferentes nervios, estos son:
1. Grupo medial o eferente somático: contiene lasneuronas motoras que forman la continuacióncefálica de las células del asta anterior. Dado queeste grupo eferente somático continúa en direcciónrostral hacia el mesencéfalo, se le denomina colum-na motora eferente somática. En el mielencéfalo re-presenta a las neuronas del nervio hipogloso (XII par)que se distribuyen en los músculos de la lengua,derivados de las somitas occipitales.
2. Grupo intermedio o eferente visceral especial: estegrupo se extiende hasta el metencéfalo y forma lacolumna motora eferente visceral especial. Susneuronas motoras se distribuyen en los músculosestriados de los cuatro arcos branquiales (faríngeos).Al nivel de mielencéfalo forma la parte de los nerviosFig. 15.14. Desarrollo del encéfalo a partir de las cinco vesículas.

225
Capítulo 15. Sistema nervioso
Fig. 15.15. Diferenciación del mielencéfalo. A, B y C. Cortes transversales.
vagos (X par), del glosofaríngeo (IX par) y la porciónbulbar del nervio espinal (XI par). Se considera que suporción caudal se diferencie en los núcleos del nervioespinal y después emigre ventralmente hacia el pisodel ventrículo donde forma el núcleo ambiguo.
3. Grupo lateral o eferente visceral general: este dalugar a la formación de la columna eferente visceralgeneral, que se encuentra en posición medial conrespecto al surco limitante, y da origen a fibras delos pares X (vago) y IX (glosofaríngeo) que inervanla musculatura lisa de los sistemas respiratorio ydigestivo, así como al corazón. Este grupo celularqueda definitivamente representado por el núcleodorsal del vago y el salival inferior que, a través delnervio glosofaríngeo, transmite la informacióneferente a la glándula parótida. Todas las fibrasmotoras originadas en los núcleos eferentesviscerales especiales y generales abandonan el bul-bo en una posición dorsolateral, con respecto al nú-cleo olivar. Dicho punto de salida está en relacióncon la llegada de las fibras sensitivas.
Placa alarInicialmente las células situadas en la porción más
lateral de la placa alar proliferan y emigran hacia aba-
jo y delante, situándose en la zona marginal, y en posi-ción ventrolateral con respecto a los derivados de laplaca basal.
Este desplazamiento recibe el nombre de prolon-gación bulbopontina (Fig. 15.16) y da origen, en su por-ción caudal, al complejo nuclear de la oliva bulbar, lacefálica se desplaza hasta la protuberancia donde for-ma los núcleos grises pontinos.
Al separarse la prolongación bulbopontina, el res-to de las células de la placa se diferencian en tresgrupos eferentes: lateral, intermedio y medial. Cadauno de estos establece sinapsis con fibras sensitivasque integran diferentes nervios formando núcleos sen-sitivos de relevo:
1. Grupo lateral: forma el grupo aferente somático(sensitivo) que recibe impulsos del oído y la super-ficie de la cabeza, por medio de los nervios vestí-bulo coclear (estatoacústico) VIII par y fibrasbulboespinales descendentes del nervio trigémino(V par). Hacia la parte final del cuarto ventrículo,las células más caudales de este grupo lateral seacercan a las del lado opuesto y se diferencian enel núcleo del cordón delgado (de Goll) y el núcleocuneiforme (de Burdach).

226
Embriología especial
2. Grupo intermedio: forma el grupo aferente visceralespecial o aferente branquial, y recibe fibras de laregión branquial (botones gustativos de la lenguay del paladar, bucofarínge y epiglotis) por mediode los nervios glosofaríngeo (IX par) y vago (Xpar). Las células de este grupo emigran en direc-ción ventral y forman los núcleos del fascículo so-litario.
3. Grupo medial: forma el grupo aferente visceral ge-neral, se sitúa en posición lateral con respecto alsurco limitante y contribuye a la creación del nú-cleo sensitivo dorsal del nervio vago. Recibe infor-mación interoceptiva del aparato gastrointestinal ydel corazón.
La capa marginal, en la porción correspondienteal mielencéfalo, presenta marcado crecimiento prove-niente de fibras nerviosas que pasan hacia la médulaespinal o en sentido contrario, hasta las porciones másrostrales.
MetencéfaloProviene de la porción cefálica del rombencéfalo
situada, desde la flexura pontina, hasta el istmo delrombencéfalo. En esta, las paredes se acercan a lalínea media y, al igual que en el mielencéfalo, las pla-cas basales y alares y el surco limitante se identificancon facilidad. Sin embargo, se forman dos nuevos com-ponentes:
1. El cerebelo, que actúa como centro de coordina-ción para el equilibrio (postura y movimiento).
2. El puente o protuberancia, que sirve para el paso delas fibras nerviosas, desde la médula espinal hacialas cortezas cerebral y cerebelosa.
Además de fibras nerviosas, la protuberancia po-see los denominados núcleos pontinos o protuberanciales,originados al nivel de las placas alares del metencéfaloy el mielencéfalo (Fig. 15.17).
Placa basalContiene a cada lado tres grupos de neuronas
motoras:1. El grupo eferente somático medial: que da origen al
núcleo del nervio motor ocular externo (VI par).2. El grupo eferente visceral especial: sus células ori-
ginan, en la porción caudal, al núcleo del nervio fa-cial (VII par) y en la cefálica, al núcleo del nerviotrigémino (V par), que inervan los músculos del pri-mero y el segundo arcos faríngeos.
3. El grupo eferente visceral general: sus células for-man el núcleo salival superior del VII par que inervalas glándulas submaxilares, sublinguales, lagrimalesy nasales.
Fig. 15.17. Diferenciación del metencéfalo.
Fig. 15.16. Prolongación bulbopontina y olivar.

227
Capítulo 15. Sistema nervioso
Placa alarEn su desarrollo se divide en dos porciones:
ventromedial y dorsolateral. En la primera se diferen-cian tres grupos celulares sensitivos:
1. Grupo lateral, aferente somático general: las célu-las de la porción cefálica de este integran la parteprotuberancial del nervio trigémino (V par), las dela porción caudal participan en una pequeña partedel complejo vestibulococlear (VIII par).
2. Grupo intermedio, aferente visceral especial: formala porción cefálica del núcleo del fascículo solitario.
3. Grupo medial, aferente visceral general: intervieneen la formación del núcleo sensitivo dorsal del ner-vio vago (X par) en su porción más cefálica.
La porción dorsolateral forma los labios rómbicosque se dirigen hacia la línea media y, en parte, se pro-yectan hacia el interior del cuarto ventrículo y cubrenparcialmente la placa del techo. La evolución ulteriorde los labios rómbicos origina el cerebelo.
Puente o protuberanciaComo se ha señalado, está formado por el au-
mento marcado de la capa marginal en la porción
correspondiente a las placas basales, debido a queactúa como zona de tránsito de las fibras nerviosasque comunican en ambos sentidos a la médula con elcerebro y el cerebelo.
Los pedúnculos cerebelosos medios, que se en-cuentran al nivel del puente, están conformados por lasfibras que se dirigen hacia el cerebelo, desde los nú-cleos pontinos, originados a su vez de la prolongaciónbulbopontina del mielencéfalo y de la placa alar delmetencéfalo, dichos núcleos están situados en el espe-sor de la protuberancia o puente.
CerebeloYa se explicó cómo las porciones dorsolaterales
de las placas alares crecen, se hacen más gruesas yforman los labios rómbicos (Figs. 15.17 y 15.18), loscuales se proyectan hacia el interior del cuarto ventrículo,originando una porción intraventricular, la cual crecenotablemente. En la superficie del metencéfalo perma-nece una porción extraventricular. En la porción caudaldel metencéfalo los labios rómbicos están muy separa-dos, pero de manera inmediata, por debajo se acercanen la línea media y forman la placa cerebelosa por de-bajo del techo del metencéfalo, que contribuye a la for-mación de las capas de la corteza cerebelosa.
Fig. 15.18. Estadios en la formación del cerebelo.

228
Embriología especial
Inicialmente, el esbozo del cerebelo es intraven-tricular pero después comienza a desplazarse hacia elexterior, hasta hacerse visible, proceso que recibe elnombre de eversión del cerebelo (Fig. 15.18).
Antes del final del tercer mes del desarrollo, casitodo el cerebelo es extraventricular y comienza, en-tonces, la formación de los pliegues en la cortezacerebelosa. Las porciones laterales del cerebelo cre-cen marcadamente y originan los lóbulos laterales ohemisferios cerebelosos que quedan unidos en la líneamedia por el vermis cerebeloso.
Hacia el final del cuarto mes aparecen las fisuras.La primera en aparecer es la denominada fisuraposterolateral que separa el flóculo, en posición lateral,de los hemisferios cerebelosos, y al nódulo, en posiciónmedial, del vermis cerebeloso (Fig. 15.19). El lóbulofloculonodular es la parte del cerebelo más antiguofilogenéticamente y establece conexiones con los nú-cleos vestibulares. Con posterioridad continúan apare-ciendo fisuras secundarias, primero al nivel del vermisy, luego, de los hemisferios cerebelosos, que le dan unaspecto característico, debido a la formación de laslaminillas del cerebelo.
Histogénesis del cerebeloLa sustancia gris del cerebelo forma los núcleos
cerebelosos (nuclei cerebelli) profundos y la cortezacerebelosa externa. Cuatro núcleos profundos se for-man a cada lado: el dentado, el globuloso, el emboliformey el núcleo fastigii o del techo, que representan los sis-temas de las vías de conducción, es decir, toda la entra-da y salida de la corteza cerebelosa se realiza a travésde estos núcleos.
Fig. 15.19. Formación de los hemisferios cerebelosos en una vista dorsal.
La corteza cerebelosa (sustancia gris) tiene unacitoarquitectura extremadamente regular, que es si-milar en toda la extensión del cerebelo. Los tipos ce-lulares de la corteza están dispuestos en capas. Losnúcleos profundos y la corteza del cerebelo son origina-dos por un proceso complejo de diferenciación.
Como en cualquier otra parte del tubo neural, elneuroepitelio de los labios rómbicos metencefálicos pre-senta una proliferación inicial para producir las capasventriculares, del manto y la marginal (Fig. 15.20. A).Sin embargo, en el tercer mes, una segunda capa decélulas proliferantes aparece en la capa más superficialde la zona marginal. La capa proliferante ventricular esdenominada ahora capa germinativa interna y la capanueva es nombrada germinativa externa (también capagranulosa externa) (Fig. 15.20. B).
Al comienzo del cuarto mes, las capas germinalesexterna e interna presentan divisiones celulares, altamentereguladas, que producen las poblaciones diferentes deneuroblastos cerebelosos. La capa germinativa internada lugar a los neuroblastos nucleares primitivos, los cua-les migran para formar los núcleos cerebelosos. Ade-más, esta capa produce dos tipos de neuroblastos quemigran a la corteza: los neuroblastos de Purkinje primiti-vos, los cuales se diferencian para formar las células dePurkinje, y los neuroblastos de Golgi, los cuales se dife-rencian y originan las células de Golgi (Fig. 15.20. C).
Según cada neuroblasto de Purkinje primitivo migrahacia la corteza, va dejando un axón que mantiene elcontacto sináptico con los neuroblastos situados en losnúcleos cerebelosos en desarrollo. Estos axones repre-sentan los únicos eferentes de la corteza cerebelosaprofunda. Las células de Purkinje forman una capa decélulas de Purkinje distintiva, justo debajo de la capagerminativa externa.

229
Capítulo 15. Sistema nervioso
La capa germinativa externa presenta tres olea-das de proliferación celular para dar origen, en suce-sión, a las tres poblaciones de neuroblastos remanentesde la corteza cerebelosa: los neuroblastos en cesta, losde los granos y los estelares (Fig. 15.20. D).
Los neuroblastos de los granos y algunos de losde cesta y estelares son desplazados a una localiza-ción profunda a las células de Purkinje, donde confor-man la capa de la granulosa de la corteza definitiva. Elmecanismo de este desplazamiento no está aún acla-rado. Las células en cesta y estelares remanentespermanecen superficiales en asociación con las célu-las de Purkinje y forman la capa molecular de la cor-teza definitiva.
Mientras que las oleadas de neurogénesis se su-ceden, las capas germinales producen los glioblastos
del cerebelo, los cuales se diferencian en astrocitos(que incluyen las células de Bergman cerebelosas es-pecializadas) y oligodendrocitos.
La corteza del cerebelo alcanza su desarrollo de-finitivo después del nacimiento, aunque los núcleoscerebelosos profundos alcanzan su posición final antesde este.
Los pedúnculos cerebelosos superiores están com-puestos, en su mayor parte, por los axones de las célu-las de Purkinje que llegan al núcleo dentado, y las fi-bras de este último se dirigen hacia el mesencéfalodando lugar a la parte de las fibras eferentes. Las co-nexiones aferentes del cerebelo provienen de distintasfuentes de origen y llegan principalmente a través delpedúnculo medio, un poco por el inferior y menos porel superior.
Fig. 15.20. Histogénesis del cerebelo.

230
Embriología especial
El estudio de la evolución filogenética del cerebelo ylos detalles de su embriología demuestran que se originancomo una especialidad del sistema de núcleos vestibulares.Cada vez se dispone de más información sugestiva de queel cerebelo no solo interviene en la regulación del compor-tamiento motor, sino que afecta también al sistema nervio-so vegetativo, a conductas emocionales, a procesos de tipoperceptivo y cognitivos, así como su lesión produce déficitdiversos en la esfera neuropsicológica.
Techo del cuarto ventrículoLa placa del techo se adelgaza al producirse la
curvatura pontina y adquiere una forma romboidea. Aesta placa del techo adelgazada, formada por célulasependimarias, se le suma más tarde, por su parte exter-na, la piamadre, que contiene vasos sanguíneos y reci-be el nombre de tela coroidea.
Los plexos coroideos se forman, a su vez, porplegamientos de la tela coroidea en el interior del cuartoventrículo y son los encargados de producir el líquidocefalorraquídeo (LCR). La fisura coroidea en la super-ficie, indica la localización del plexo coroideo (Fig. 15.21).
La porción de la tela coroidea situada entre la fi-sura y el cerebelo se transforma en el velo medularposterior (válvula de Tarin). Durante el tercer mes, pordebajo de la fisura coroidea y en la línea media, el re-vestimiento ependimario desaparece parcialmente yforma el agujero de Magendie, que comunica al cuartoventrículo con el espacio subaracnoideo.
Al constituirse la fosa romboidea, sus porciones la-terales reciben el nombre de recesos laterales, con pos-terioridad a la formación del agujero de Magendie, enestos recesos se forman los agujeros de Luschka, que
comunican el cuarto ventrículo con el espacio subarac-noideo permitiendo el paso del líquido cefalorraquídeo adicho espacio.
El espacio subaracnoideo se extiende alrededorde todo el SNC, haciéndose más amplio en ciertos nive-les para formar las cisternas. Por último, la porción detela coroidea que se encuentra entre el cerebelo y elmesencéfalo se transforma en el velo medular anterior(válvula de Vieussens).
MesencéfaloEs la más primitiva de las vesículas encefálicas, li-
mita en la porción caudal con el istmo del rombencéfaloy en la cefálica, con el diencéfalo. Forma ventralmentelos pedúnculos cerebrales y dorsalmente los tubérculoscuadrigéminos. Debido a la presencia de la curvaturacefálica, esta vesícula es la más alta en su inicio, aunqueluego queda oculta como consecuencia del desarrollo deltelencéfalo. El crecimiento de las paredes es marcado,no así el de la cavidad de la vesícula que resulta másestrecha y al final, representa el acueducto de Silvio quecomunica el tercer ventrículo con el cuarto (Fig. 15.22).
Fig. 15.22. Diferenciación del mesencéfalo.Fig. 15.21. Cuarto ventrículo en un corte transversal.

231
Capítulo 15. Sistema nervioso
Placa basalLas células de esta placa forman dos grupos celu-
lares motores:1. Grupo medial: eferente somático, representado por
los nervios oculomotor (motor ocular común, IIIpar) y troclear (patético, IV par), que inervan alos músculos del ojo.
2. Grupo lateral: pequeño, eferente visceral general,representado por el núcleo de Edinger-Westphal,que inerva al esfínter de la pupila por medio de fi-bras viscerales incorporadas al III par.
Placa alarLas células del manto de la placa alar se despren-
den y agrupan dorsalmente y forman la láminacuadrigémina, dividida en dos prominencias por un sur-co en sentido sagital. Después, un segundo surco lasdivide en sentido transversal. Así se originan cuatromasas: los tubérculos cuadrigéminos superiores(colículos anteriores) y los tubérculos cuadrigéminosinferiores (colículos posteriores). Los colículos se for-man por oleadas de neuroblastos que emigran hacia lazona marginal suprayacente, donde se disponen en ca-pas estratificadas.
Los colículos posteriores funcionan como centrossinápticos de relevo para los reflejos auditivos; los ante-riores sirven como centros de correlación y de reflejospara los estímulos visuales. Otros grupos de célulasemigran en dirección ventral y dan lugar a la formacióndel locus niger, en forma de semiluna y el núcleo rojo,de aspecto redondeado.
Por último, en la capa marginal de cada placa basal,ventralmente con respecto al locus niger se agrupanfibras motoras y, dorsalmente, fibras sensitivas que for-man unas prominencias denominadas pedúnculos cere-brales. Los pedúnculos sirven de vía para las fibras ner-viosas que descienden, desde la corteza cerebral, hacialos centros inferiores de la protuberancia y la médulaespinal.
DiencéfaloSe origina de la porción medial del prosencéfalo
al principio del segundo mes. Su límite caudal estáseñalado por un plano que pasa por detrás de la glán-dula pineal (epifisiaria) y de los tubérculos mamilares,que lo separa del mesencéfalo. El límite cefálico con-siste en un plano que pasa por delante del quiasmaóptico y rodea al agujero interventricular o de Monro(Fig. 15.23). Además, la placa del techo, la del piso y el
epéndimo del diencéfalo dan lugar a varias estructurasa través de mecanismos que son relativamente únicos.Estas estructuras incluyen el plexo coroides y órganoscircunventriculares, el lóbulo posterior de la glándulahipofisiaria (neurohipófisis) y las vesículas ópticas.
Las estructuras diencefálicas se derivan en su to-talidad de las dilatadas placas alares y del techo, ya quelas placas basales y del piso solamente llegan hasta elmesencéfalo (es interesante señalar que, un marcadorde línea media ventral, sonic hedgehog (Shh), es ex-presado en el piso del diencéfalo, lo que sugiere queexista una placa del piso).
Placa del techoLa placa del techo del diencéfalo permanece
epitelial en su carácter, está compuesta por una capade células ependimarias cubiertas por mesénquimavascular y piamadre. Esta capa se proyecta en el inte-rior y forma los plexos coroideos del tercer ventrículo.En otras partes del tercer ventrículo, el epéndimo for-ma un número de estructuras secretorias únicas queincorpora metabolitos específicos y neuropéptidos al lí-quido cefalorraquídeo. Estas estructuras, colectivamenteconocidas como órganos circunventriculares, incluyenel órgano subfornical, el organum vasculosum de lalámina terminalis y el órgano subcomisural.
En la porción caudal de la placa del techo se pro-duce una evaginación durante la séptima semana y daorigen al cuerpo pineal o epífisis (Figs. 15.23 y 15.24),que se sitúa finalmente en el mesencéfalo y sirve comoregulador de la acción de la luz y la oscuridad sobre losritmos endocrinos y del comportamiento.
La placa del techo contribuye a formar el epitálamo,el cual está formado por un grupo de núcleos situadosjunto y a cada lado de la glándula pineal, denominadosganglios de la habénula. Estos ganglios participan en laconducción olfatoria y están unidos por fibras que atra-viesan la línea media por delante y por detrás del cuer-po pineal, lo que da lugar a las comisuras interhabenulary posterior.
Placa alarEn las caras ventriculares de las placas alares apa-
recen al final de la quinta semana unos surcoslongitudinales superficiales, estos son los hipotalámicosque dividen a estas caras en expansión en las regionestalámicas e hipotalámicas. El tálamo crece despro-porcionadamente después de la séptima semana y seconvierte en la estructura más grande del diencéfalo.

232
Embriología especial
Debido a la rápida expansión de las carasventriculares, las dos regiones talámicas se fusionan enla línea media y forman la masa intermedia o comisuragris intertalámica. En el tálamo se forman los núcleostalámicos (anterior, ventral, medial y lateral) del cuerpogeniculado medio y del lateral. Al final de la sexta se-mana, un surco poco profundo denominado sulcusdorsalis separa al tálamo del epitálamo originado, esteúltimo, en el borde dorsal de la pared diencefálica y laplaca del techo adjunta.
El tálamo actúa primero como centro de relevopara la corteza cerebral, este recibe toda la informa-ción (sensoria y otras) que se proyecta a la corteza desdeestructuras subcorticales, las procesa, si es necesario,y las envía a las áreas corticales apropiadas. Dentro deltálamo, el sentido de la vista es manipulado por el cuer-po geniculado lateral y el sentido de la audición por elcuerpo geniculado medio.Fig. 15.24. Corte sagital del diencéfalo.
Fig. 15.23. Evolución del diencéfalo.

233
Capítulo 15. Sistema nervioso
El hipotálamo está formado por gran número depequeños núcleos que regulan la actividad endocrina dela hipófisis, así como respuestas autonómicas. Participaen el sistema límbico, el cual controla la emoción y co-ordina el estado emocional con las respuestas visceralesapropiadas. El hipotálamo también regula el sueño, lavigilia, la digestión y la temperatura corporal. Posterior-mente, una parte de estos núcleos originan los cuerposo tubérculos mamilares en dirección cefalocaudal.
Las fibras de la retina, desde la cúpula óptica seproyectan a los cuerpos geniculados por medio del ner-vio óptico. Antes que penetren al cerebro, las fibras deambos ojos se encuentran para formar el quiasma ópti-co, una estructura de unión en la cual algunas de lasfibras de cada lado se entrecruzan (decusan). Los ha-ces resultantes de fibras ipsolaterales y contralateralesse dirigen entonces atrás en los cuerpos geniculadoslaterales, donde hacen sinapsis para formar un mapadel campo visual.
No todas las fibras de la retina se proyectan en loscuerpos genitales laterales; sino que algunas de estasterminan en el colículo superior, donde estos median elcontrol del reflejo ocular.
Glándula pituitaria o hipófisisLa hipófisis, también conocida como glándula
pituitaria, tiene función endocrina de origen dual, com-puesta por la adenohipófisis y la neurohipófisis. Laprimera aparece durante la tercera semana, delectodermo del techo del estomodeo en forma de bol-sa, denominada bolsa de Rathke, y la segunda, deuna prolongación descendente del piso del diencéfalo,el infundíbulo (Fig. 15.25. A y B).
Hacia el final del segundo mes, la bolsa de Rathkepierde su conexión con la cavidad bucal (Fig. 15.25. C),y las células de su pared ventral proliferan y forman lapars distalis; una pequeña parte de esta porción rodeael infundíbulo creando la pars tuberalis. En cambio, lapared posterior se desarrolla muy poco y da origen a lapars intermedia (Fig. 15.25. D y E). Del infundíbulo sederivan el tallo infundibular, que permanece conectadoal diencéfalo y, en su extremo distal, la neurohipófisis opars nervosa. La luz del infundíbulo se oblitera por esteproceso pero una fosita proximal pequeña, el recesoinfundibular, persiste en el piso del tercer ventrículo. Laneurohipófisis está integrada por una masa de célulasde neuroglia modificada, que reciben el nombre depituicito, y por fibras nerviosas provenientes delhipotálamo.
TelencéfaloEste segmento prosencefálico es el extremo
cefálico (rostral) del encéfalo y está formado por dosevaginaciones laterales que aparecen durante la quintasemana, las vesículas telencéfalicas, y que contienenlos ventrículos laterales (Fig. 15.26). Están unidas en elcentro por la lámina terminal y una porción media, quees continuación, y contiene la parte anterior del tercerventrículo, la que comunica con los ventrículos lateralesa través de los agujeros interventriculares o agujeros deMonro. La porción total y visible exteriormente de loshemisferios cerebrales, representa el palio. Deltelencéfalo se originan los hemisferios cerebrales, lascomisuras y otras estructuras que se les unen. Tambiénforma los bulbos olfatorios y tractos olfatorios, los cua-les, junto con los centros olfatorios y tractos de los he-misferios cerebrales, constituyen el rinencéfalo (nariz-cerebro).
Al igual que el diencéfalo, el telencéfalo se derivade las láminas alares extendidas. Las vesículastelencéfalicas desarrollan tres estructuras principales:el lóbulo olfatorio (rinencéfalo), el cuerpo estriado, con-tinuación directa del tálamo y la corteza cerebral(neopalio) (Fig. 15.26).
Fig. 15.25. Desarrollo de la hipófisis.

234
Embriología especial
En el sitio donde la pared del hemisferio se une conel techo del diencéfalo no existen neuroblastos y la zonaes muy delgada, ya que solo tiene una sola capa de célu-las ependimarias cubiertas por mesénquima vascularizadoque forman el plexo coroideo. Este último debía haberdado lugar al techo del hemisferio pero, como consecuen-cia del crecimiento desproporcionado de varias partesdel hemisferio (Fig. 15.27), sobresale en el ventrículo la-teral siguiendo una línea denominada fisura coroidea.
RinencéfaloSe debe señalar inicialmente que, las placodas
nasales aparecen a finales de la cuarta semana. Muytemprano, algunas células en la placoda nasal se dife-rencian para formar las células neurosensoriales pri-marias del futuro epitelio olfatorio. Al final de la quintasemana, estas células emiten axones que cruzan la dis-
Fig. 15.26. Formación de las cavidades del telencéfalo.
Fig. 15.27. Diferenciación del telencéfalo.
tancia corta para penetrar en la porción final más cra-neal del telencéfalo. La osificación subsecuente delhueso etmoides, alrededor de estos axones, crea la pla-ca cribiforme perforada (Fig. 15.28).
En cuanto al rinencéfalo es la parte olfatoria delcerebro. El aparato olfatorio también forma parte delpalio. En la sexta semana, en el área de la superficieventral de cada hemisferio cerebral, donde los axonesde las células neurosensoriales primarias comienzan ahacer sinapsis, forman unos abultamientos denomina-dos lóbulos olfatorios, que muy pronto se dividen en dosporciones: una ventral integrada por el bulbo olfatorio ylas cintillas olfatorias; y otra dorsal que origina la sus-tancia perforada anterior y el área paraolfatoria.
La porción proximal de los lóbulos olfatorios for-ma el paleopalio; al margen de cada hemisferio contri-buirá a formar el hipocampo o arquipalio. Esta estruc-tura engrosada, situada por arriba de la fisura coroidea,tiene función olfatoria y sobresale gradualmente en elventrículo lateral. En la superficie del hemisferio, elhipocampo está indicado por un surco longitudinal quecorresponde a la fisura o surco del hipocampo, que esparalelo y está situado por encima de la fisura coroidea.
Cuerpo estriadoDurante la sexta semana se produce una marca-
da proliferación de la capa del manto, en la porciónbasal de cada hemisferio, que produce una prominen-cia dentro del ventrículo lateral y también en el pisodel agujero de Monro, con presencia de crestaslongitudinales en formas de estrías, por lo que recibeel nombre de cuerpo estriado.

235
Capítulo 15. Sistema nervioso
Corteza cerebralLa corteza no olfatoria (neopalio) en el hombre
sobrepasa en su crecimiento al resto del telencéfalo,por lo cual las demás partes quedan ocultas. Abarcatodo el hemisferio, excepto la formación hipocámpica.
Después de la aparición del paleopalio, las célulasde la capa del manto emigran, a partir de la séptimasemana, hacia la capa marginal (visibles en formaestratificada en el recién nacido) y producen una capacortical superficial que en su porción motora está com-puesta por células piramidales y, en la sensitiva, porcélulas granulosas.
Los hemisferios crecen y se expanden rápido, pri-mero ventralmente formando los lóbulos frontales, des-pués de manera cefálica y lateral, desarrollando los ló-bulos parietales y, de inmediato, las expansiones dorsa-les y caudales constituyen los lóbulos occipitales y tem-porales. La corteza que cubre el núcleo lenticular per-manece como área deprimida o fija, que recibe el nom-bre de ínsula de Reil, y queda oculta en el piso de lacisura de Silvio, debido al crecimiento de los lóbulosadyacentes (Fig. 15.29).
Fig. 15.28. Evolución del rinencéfalo.
Al principio, el cuerpo estriado está separado deltálamo por una hendidura, la que desaparece gradual-mente con el desarrollo, y ambas estructuras formanuna masa continua. El cuerpo estriado se divide en dospartes: una ventrolateral que origina el núcleo lenticular,y otra dorsomedial en la cual se forma el núcleo caudadoque se mantiene en contacto con el tálamo.
Esta división se produce como consecuencia deque el cuerpo estriado es atravesado por un númerocada vez mayor de cilindroejes formando así, la cápsulainterna. Posteriormente, el núcleo lenticular, por la mis-ma causa, se divide en una porción lateral, el putamen,y otra medial, el globus pallidus.
Las fibras de proyección cortical no pasan todas através de la cápsula interna; algunas lo hacen por fueradel núcleo lenticular formando la cápsula externa, la cualsepara al núcleo lenticular de un grupo de células apla-nadas, conocidas con el nombre de claustrum que es-tán profundamente situadas en la corteza insular. Lasfibras continúan y pasan entonces a través de la regiónsubtalámica y penetran en los pedúnculos delmesencéfalo.

236
Embriología especial
Fig. 15.30. Desarrollo de la corteza cerebral.
Al principio, las paredes de los hemisferios cere-brales son lisas pero, a partir del cuarto mes, la superficiede los hemisferios cerebrales crece con más rapidez,debido al aumento de la corteza o sustancia gris, que lacapa medular blanca situada por debajo, por lo cual sepliega y forma las circunvoluciones separadas por sur-cos que, cuando son muy pronunciados, reciben el nom-bre de cisuras (Fig. 15.30). El cuadro detallado de lascircunvoluciones que se forman finalmente en los hemis-ferios cerebrales, varía un tanto de un individuo a otro.
ComisurasDespués del cierre del neuroporo anterior, la pa-
red anterior del prosencéfalo se denomina lámina
terminalis; se continúa hacia arriba con la delgada pla-ca del techo del prosencéfalo y, hacia abajo, con su pla-ca gruesa del piso.
Después de la aparición del hemisferio cerebral,la lámina terminalis se engruesa y forma la placacomisural, por la cual pueden pasar las vías de un he-misferio cerebral a otro. Cuando estas fibras conectanregiones correspondientes de las dos mitades del cere-bro, se denominan fibras comisurales. Otras fibras quecruzan de un lado a otro del cerebro se decusan. En elcerebro anterior, tanto las fibras comisurales, como lasque se decusan, se agrupan por lo general comocomisuras cerebrales (Fig. 15.31).
La primera en aparecer, octava semana, en laporción inferior de la placa comisural, es la comisuraanterior, que está formada por fibras que conectan lacorteza piriforme con el bulbo olfatorio de un hemis-ferio con los del lado opuesto. Poco después aparecela segunda comisura, la del hipocampo del trígono odel fórnix, sus fibras nacen en el hipocampo y con-vergen en la lámina terminal, cerca de la placa deltecho del diencéfalo; sus fibras relacionan las corte-zas de las formaciones hipocámpicas de ambos he-misferios.
Fig. 15.29. Crecimiento de los hemisferios cerebrales.

237
Capítulo 15. Sistema nervioso
La mayor y más importante de las comisuras es elcuerpo calloso, que aparece hacia la décima semana, yque conecta las áreas corticales no olfatorias de los doshemisferios cerebrales. Su crecimiento se debe a laexpansión rápida y continua del neopalio. La parte másanterior se forma primero y su extensión posterior (elsplenium) se forma tarde en la vida fetal, se extiendeprimero hacia delante, uniendo los lóbulos frontales,después, en la región dorsal con el desarrollo de los ló-bulos parietales, se arquea sobre el delgado techo deldiencéfalo. El área entre el cuerpo calloso y la comisu-ra del hipocampo se hace muy delgada y forma el septolúcido. Las comisuras descritas y el septo lúcido en sudesarrollo forman parte de la lámina embrionaria y re-presentan prolongaciones de esta.
Además de las tres comisuras señaladas, apare-cen otras tres. Dos de las cuales, las comisuras poste-rior y superior (habenular) se encuentran inmediatamen-te por delante y por debajo del tallo de la epífisis, laprimera está formada por fibras que unen ambos haceslongitudinales medios y la segunda por fibras queinterconectan ambas regiones epitalámicas. La terce-ra, el quiasma óptico, se forma de manera precoz. Ensu porción anterior de la lámina terminal está compues-ta por fibras de la mitad interna de la retina que cruzanhacia el lado opuesto para alcanzar el cuerpo geniculadoy el tubérculo cuadrigémino superior.
En cuanto a la incidencia geneticomolecular deldesarrollo del cerebro hay que tener en cuenta que, comose describió al inicio del capitulo, la inducción primariadel sistema nervioso da como resultado la formación dela placa neural por encima de la notocorda. Estudios
moleculares realizados fundamentalmente en anfibioshan permitido conocer mejor algunos mecanismos de lainducción neural.
En los embriones, durante la etapa de gastrulación, electodermo dorsal produce la molécula de señal BMP-4que inhibe la formación de tejido neural. Sin embargo,moléculas producidas por la notocorda como noggin ychordin, así como folistatina en anfibios, bloquean laacción inhibidora del BMP-4 sobre el ectodermo dor-sal, por lo que se forma la placa neural.
Bajo la acción de noggin y chordin, la placa neuraldesarrolla el cerebro anterior, al tiempo en que, por laacción del factor de crecimiento fibroblástico (FGF-8) sedesarrolla la porción caudal de la placa neural, es decir,la médula espinal. Luego el ectodermo neural inducidoy bajo la acción de noggin, chordin y FGF-8, juntasdesarrollan el cerebro medio y posterior. Casi de inme-diato, después de la inducción neural, la placa neural co-mienza el proceso de formación del tubo de igual nombre.
Como se observa, en estas etapas tempranas, yase define el eje o patrón anteroposterior e inicio de laformación de cada una de las regiones del encéfalo,con patrones específicos que caracterizan las diferen-tes regiones del cerebro, además de establecer los lími-tes entre regiones. Luego se continúa el desarrollo denumerosas regiones o segmentos denominadosrombómeras en la región del cerebro posterior, y divi-siones menos precisas conocidas como prosómeras enel cerebro anterior.
En el caso del cerebro posterior se consideranimportantes las acciones del factor de transcripciónKsox-20 y genes tipo homeobox, en la determinaciónde los segmentos del cerebro posterior. Si se elimina
Fig. 15.31. Formación de las comisuras cerebrales.

238
Embriología especial
Ksox-20 las rombómeras tres y cinco no se forman.Estos genes por tanto, confieren un valor posicional a lolargo del eje anteroposterior y determinan la identidad delas rombómeras, así como especifican sus derivados.
Aún no está claro cómo ocurre esta regulación,aunque parece que el ácido retinoico puede desempe-ñar alguna función en el control sobre la expresión delos genes tipo Hox (homeobox). Una deficiencia en elácido retinoico trae como consecuencia la formaciónde un cerebro posterior pequeño con ausencia de lasrombómeras posteriores.
Los mecanismos que intervienen para la defini-ción del patrón en la región del cerebro medio se en-cuentran situados en un centro de señales molecularesen el límite, entre el mesencéfalo y el metencéfalo. Laprincipal molécula es la FGF-8, la que se expresa en unestrecho anillo en el límite anterior del primer rombómero,una subdivisión del metencéfalo. El FGF-8 actuando conla molécula de señal Wnt-1 induce la expresión de losgenes engrailed En-1 y En-2. El En-1 regula el desarrollodel cerebro medio dorsal (tectum) y la parte anteriordel cerebro posterior (cerebelo), el En-2 participa soloen la diferenciación del cerebelo.
Estudios moleculares recientes en algunas especiesanimales indican que el límite, entre el futuro cerebromedio y posterior, no se corresponde con la constricciónanatómica entre el mecencéfalo y el romboencéfalo (ist-mo), sino que este límite se localiza craneal a esta cons-tricción en un plano marcado por el límite posterior de laexpresión de los genes homeobox Otx-2.
Anomalías congénitas del encéfaloLas malformaciones del SNC representan alre-
dedor de 10 % de todas las malformaciones y, al me-nos, la mitad de estas son evidentes al nacimiento.Los defectos del tubo neural (DTN) son mucho másfrecuentes en embriones que en recién nacidos, y secree que más de 90 % de embriones afectados sepierden durante el periodo embrionario.
En los humanos, los virus (rubéola y citomega-lovirus), los rayos X, la hipoxia, y los trastornos here-ditarios y cromosómicos (ejemplo trisomías) son los másimportantes en la producción de estas afecciones.
Alrededor de 95 % de estos son espina bífida yanencefalia. La prevalencia de los defectos del tuboneural varía de 1 a 5/1 000 nacidos vivos y el índice deocurrencia es aproximadamente de 3 %.
Para el diagnóstico prenatal, los métodos principa-les utilizados para la detección de los defectos del tuboneural, especialmente para anencefalia y espina bífidaabierta son la ultrasonografía, la evaluación dealfafetoproteína y acetilcolinesterasa y la citologíaexfoliativa del líquido amniótico.
La alfafetoproteína es producida inicialmentepor el saco vitelino pero muy pronto es sustituida porel hígado (Fig. 15.32). Entra al líquido amniótico pormedio de la orina fetal y es extraída por ingestiónfetal y digestión intestinal. Una comunicación direc-ta o indirecta, entre el líquido cefalorraquídeo y ellíquido amniótico, ocurre en la espina bífida y en laanencefalia.
Fig. 15.32. Relación de los defectos del tubo neural y laalfafetoproteína.
Si ocurre alguna malformación, el resultado de laprueba diagnóstica de alfafetoproteínas, que se realizaen el líquido amniótico y en el suero materno, es deniveles más altos de lo normal. Por lo que esto se puededetectar prenatalmente.

239
Capítulo 15. Sistema nervioso
Niveles elevados se encuentran en la gastrosquisis,en la atresia esofágica o intestinal, mientras que el niveldisminuido se presenta en la trisomía 21 y 18.
PrevenciónUna dieta materna mejorada y preparaciones
multivitamínicas se recomiendan para la prevención dedefectos del tubo neural. Además, a causa de que los cam-bios metabólicos del folato son considerados como factorimportante de defectos del tubo neural, se recomienda ac-tualmente la administración de ácido fólico (4 mg/d) du-rante las etapas de embarazo y antes del embarazo.
Defectos del tubo neural en el encéfaloLos diferentes tipos principales de espina bífida ya
descritos tienen sus contrapartes en la región cerebral:1. La placa neural permanece abierta.2. El tubo neural se exterioriza.3. Solo las meninges se exteriorizan.4. Puede suceder que un simple defecto esquelético
sea evidente.
Se pueden presentar las anomalías siguientes:1. Anencefalia: el tipo más severo de disrafia cerebral
se presenta como la ausencia parcial del cerebro yde la bóveda craneal suprayacente. Las partes quefaltan generalmente son el prosencéfalo, elmesencéfalo y la parte rostral del rombencéfalo.Muchos nacen muertos pero algunos sobrevivenpocas horas o días. Esta condición se puede combi-nar con un defecto espinal adyacente (craneorra-quisquisis). La incidencia varía de manera conside-rable con la localización geográfica. Es cuatro ve-ces más frecuente en las mujeres que en los hom-bres. Debido a que falta en el feto el mecanismo dedeglución, en los dos últimos meses del embarazose presenta hidramnios. La anormalidad se puedereconocer en los rayos X, ya que la bóveda cranealestá ausente. La anencefalia se ha producido deforma experimental, por ejemplo, en ratas conhipervitaminosis A (Fig. 15.33).
2. Meningocele, meningoencefalocele y meningohidroen-cefalocele: son todas causadas por un defecto de osi-ficación en los huesos del cráneo. Con frecuencia sonreferidas todas como encefalocele. El hueso que másse observa afectado es la porción escamosa del huesooccipital, la cual puede faltar de manera parcial o to-tal. Si la apertura del hueso occipital es pequeña, solo
las meninges protruyen a través de esta, pero si eldefecto es grande, parte del cerebro y aun parte delventrículo pueden penetrar a través de la apertura den-tro del saco meníngeo. Las dos últimas malformacio-nes son conocidas como meningoencefalocele ymeningohidroencefalocele, respectivamente.
3. Síndrome de Meckel-Gruber: en esta afección existeencefalocele occipital, acompañado de riñonespoliquísticos y polidactilia.
Fig. 15.33. Algunos ejemplos de anomalías cerebrales.
4. Hidrocefalia: se caracteriza por aumento anormaldel líquido cefalorraquídeo dentro del sistemaventricular. La hidrocefalia puede tener causa pre-natal, siendo un ejemplo la infección viral otaxoplasmosis o, posnatal, como por ejemplo la neo-plasia. Algunos casos son familiares. Los ventrículosestán siempre aumentados en tamaño, con marca-do agrandamiento de la cabeza poco antes del naci-miento. La anormalidad se puede aliviar con un “cor-tocircuito” del líquido cefalorraquídeo del sistemaventricular a la cavidad peritoneal o dentro de laaurícula derecha. La hidrocefalia se subdivide endos tipos principales: no obstructiva y obstructiva:a) Hidrocefalia no obstructiva: se refiere a un
agrandamiento relativo de los ventrículos y de

240
Embriología especial
los espacios subaracnoideos, causados por pér-dida de tejido cerebral.
b) Hidrocefalia obstructiva: es la que se asocia condeterioro de la circulación o de la absorción delíquido cefalorraquídeo. Se subdivide en comuni-cante y no comunicante:
– Hidrocefalia comunicante: se refiere a un ac-ceso libre al espacio subaracnoideo, aunque unaobstrucción ocurre fuera del sistema ventricular.Las causas incluyen infección (por ejemplo,citomegalovirus o Toxoplasma) y hemorragiaintracraneal.
– Hidrocefalia no comunicante: se refiere a faltade comunicación entre el sistema ventricular yel espacio subaracnoideo, ambas craneal yespinal. La obstrucción es más frecuente en elacueducto de Silvio, pero puede estar en el cuar-to ventrículo o sus aperturas. Aunque la causade la estenosis del acueducto es por lo generaldesconocida, puede resultar de gliosis, inflama-ción o neoplasia.
5. La malformación de Arnold-Chiari: descrita porCleland en 1883, es un desplazamiento caudal yherniación de las estructuras cerebelosas y médula,incluyendo la porción caudal del cuarto ventrículo através del foramen magnum. La malformación deArnold-Chiari se presenta de manera virtual en cadacaso de espina bífida quística y está con frecuenciaacompañada de hidrocefalia (Fig. 15.34).
6. Holoprosencefalia: es la pérdida de las estructurasde la línea media en el cerebro y la cara. En loscasos severos, los ventrículos laterales emergencomo una vesícula telencefálica única, los ojos es-tán fusionados y hay una cámara nasal única al ladode otros defectos faciales de la línea media. Estaanomalía es heterogénea e incluye formasautosómicas dominantes y recesivas. Puede serparte también de síndromes monogénicos. Latrisomía 13 y 18 puede estar asociada. La diabetesmaterna y los medicamentos antiepilépticos puedenactuar como teratógenos. Las mutaciones del Shh,gen que regula el establecimiento de la línea mediaventral en el SNC, resultan en algunas formas deholoprosencefalia. Esta última es mucho más fre-cuente en embriones que en recién nacidos, y sepiensa que más de 90 % de los embriones afecta-dos se pierden durante el periodo embrionario.
7. Síndrome de Smith-Lemoli-Ortiz: es un defecto de labiosíntesis del colesterol, estos niños presentan de-fectos craneofaciales y de las extremidades inferio-res y 5 % presentan holoprosencefalia. Este síndro-me se debe a anormalidades en la 7-dihidrocolesterolreductasa la cual metaboliza al 7-dihidrocolesterola colesterol. Mucho de los defectos, incluyendo losdel cerebro y las extremidades inferiores, se pue-den deber a señalamiento anormal de Shh, ya queel colesterol es necesario para que este gen ejerzasus defectos. Todavía otra causa de holoprosen-cefalia es el abuso del alcohol el que, en estadiostempranos del desarrollo, destruye de manera se-lectiva a las células de la línea media.
8. Microcefalia: una cabeza pequeña es el resultado,con frecuencia, de una reducción general de la masadel cerebro, debida a una hipolasia primaria o a unadestrucción secundaria del cerebro que se puedeestablecer prenatal o posnatal. Factores medioam-bientales como, radiación en la madre, anoxia perinatal,trisomía 21 (síndrome de Down), así como factoresgenéticos (autosómica recesiva) son causas de esta
Fig. 15.34. Característica de la malformación Arnold-Chiari.

241
Capítulo 15. Sistema nervioso
afección. La toxoplasmosis fetal puede causar cal-cificación precoz cerebral y de los huesos del crá-neo. La microcefalia es la base de muchos casosde retardo mental.
9. Retardo mental: puede, aun en casos severos, no es-tar asociado con anormalidades cerebralesmorfológicamente. El retardo mental puede resultarde anormalidades genéticas (síndrome de Down yde Klinefelter) o de exposición a teratógenos(malnutrición y saturnismo) o a agentes infecciosos(rubéola, citomegalovirus y Toxoplasma) u otras cau-sas como fenilcetonuria y cretinismo. La deficienciade iodo es una causa importante prevenible a causade que el iodo es necesario para el desarrollo delcerebro, así como para la función de la glándulatiroides. Pero la causa líder del retardo mental es, sindudas, el abuso materno del alcohol.
Estas anormalidades señaladas son las más seriasy pueden ser incompatibles con la vida. Otros numero-sos defectos del SNC se pueden presentar sin manifes-taciones externas. Por ejemplo, el cuerpo calloso, pue-de estar ausente de manera parcial o total sin muchotrastorno funcional. También puede haber ausencia par-cial o completa del cerebelo con trastornos ligeros de lacoordinación.
Sistema nervioso periféricoEstá formado por dos sistemas principales: el
cerebroespinal y el vegetativo, ambos están integra-dos por nervios dispuestos en formas pares, que emi-ten sus prolongaciones hacia regiones simétricas delcuerpo. Estos nervios están conformados por fibraseferentes, que transmiten impulsos motores prove-nientes, tanto de los centros nerviosos de neuroblastossituados en la placa basal del sistema nervioso cen-tral, como por fibras eferentes que llevan impulsossensitivos hacia el sistema nervioso central, y se ori-ginan de células nerviosas situadas en la cresta neurala lo largo del tubo neural.
Sistema nervioso cerebroespinal
Nervios raquídeosSe disponen segmentariamente de acuerdo con los
miotomas que inervan. Presentan dos raíces: una dor-
sal (posterior) asociada a un ganglio y otra ventral (an-terior). Hacia el final de la cuarta semana se puedenobservar las fibras de la raíz ventral (que crecen de losneuroblastos de la capa del manto de la médula) y losganglios raquídeos como expansiones de la cresta neural,todavía continua. En la quinta semana, las neuronas delos ganglios emiten dos tipos de prolongaciones: centrí-petas, que penetran en la capa marginal de la médulaformando las raíces dorsales y periféricas, que se unena las raíces ventrales y son los troncos nerviosos.
En la sexta semana, desaparecen los puentes dela cresta neural que unían a los ganglios raquídeos y sehacen evidentes los nervios raquídeos. Esta diferencia-ción se efectúa en dirección cefalocaudal.
Del tronco de cada nervio raquídeo, poco despuésde la unión, se emite una rama ventral (motora) y otradorsal (sensitiva).
La rama ventral, de mayor grosor, se prolonga yemite la rama comunicante que se dirige al ganglio sim-pático, y las ramas terminales lateral y ventral queinervan los músculos y la piel de las paredes laterales yventrales del cuerpo respectivamente (Fig. 15.35).
Plexos nerviososEn las regiones correspondientes a los miembros
donde los músculos no presentan la regularidad de lamusculatura del tronco, las ramas lateral y ventral emi-ten asas comunicantes de un nervio raquídeo a otro, yforman así los plexos nerviosos que son ya evidentes enembriones de seis semanas. Los plexos de los miem-bros superiores son los branquiales y los de los miem-bros inferiores los lumbosacros.
Los nervios dorsales de los plexos inervan losmúsculos extensores y, los ventrales los músculosflexores.
Nervios cranealesLos nervios craneales están formados por 12 pa-
res y hacen su aparición durante la quinta y sexta se-manas. Se caracterizan por no adoptar una disposiciónsegmentaria ordenada, además de los componentesgenerales, sensitivos y motores, contienen fibras espe-ciales distribuidas en los principales órganos de los sen-tidos y en los músculos derivados de los arcosbranquiales. Sus núcleos sensitivos y motores están dis-puestos en masas y columnas definidas de las respecti-vas placas alar y basal.

242
Embriología especial
c) VI motor ocular externo.d) XII hipogloso mayor.
3. Sensitivos y motores viscerales:a) V trigémino.b) VII facial.c) IX glosofaríngeo.d) X neumogástrico o vago (incluye fibras del XI
par espinal o accesorio).
Sistema nervioso autónomoo vegetativo
Se divide desde el punto de vista funcional en: sis-tema nervioso simpático, que abarca la regióntoracolumbar y sistema nervioso parasimpático, corres-pondiente a las regiones craneal y sacra.
Fig. 15.35. Formación de los nervios raquídeos y simpáticos.
A diferencia de los nervios raquídeos o espinales,la composición funcional de los nervios craneales varíade manera amplia, los sensitivos especiales y los moto-res somáticos tienen una sola clase de fibras, mientrasque los sensitivos tienen varias, especialmente los ner-vios glosofaríngeo y neumogástrico, cada uno de loscuales contiene cinco tipos de fibras diferentes. La agru-pación funcional de estos nervios se resume a conti-nuación:
1. Sensitivos especiales:a) I olfatorio.b) II óptico.c) VIII estatoacústico.
2. Motores somáticos:a) III motor ocular común.b) IV patético.

243
Capítulo 15. Sistema nervioso
Sistema nervioso simpáticoEstá compuesto de neuronas (motoras viscerales)
ubicadas en el sistema nervioso central, cuyas fibras seextienden hasta los ganglios simpáticos, y por otro gru-po de neuronas, que tienen sus cuerpos celulares en unganglio y cuyos axones salen fuera de este y activan lascélulas glandulares o los músculos involuntarios de lasvísceras y los vasos sanguíneos.
En la quinta semana, ciertas células de la crestaneural emigran a través de las raíces dorsales de losnervios espinales y se disponen en masas pares situa-das dorsolateralmente con respecto a la aorta. Prontose fusionan debido a su crecimiento y forman un par debandas longitudinales en las cuales aparecen agrupa-ciones segmentarias de neuroblastos que son los gangliossimpáticos (Fig. 15.35. B). Estos neuroblastos se dife-rencian en células ganglionares multipolares,encapsuladas por células satélites que se originan tam-bién en la cresta neural.
Los ganglios simpáticos se comunican entre sípor fibras nerviosas y forman de este modo las ca-denas simpáticas, lo cual ocurre durante la séptimasemana.
Desde su posición inicial en el tórax, grupos deneuroblastos emigran hacia la región cervical ylumbosacra, y se fusionan al nivel de la región cervi-cal. Algunos neuroblastos emigran a la superficie ven-tral de la aorta y forman los ganglios preaórticos (celía-co, mesentérico superior y mesentérico inferior) yotros emigran al corazón, el pulmón y el sistema di-gestivo y originan los plexos viscerales simpáticos(Fig. 15.35. A).
Al nivel de los segmentos toracolumbares de lamédula espinal se originan fibras nerviosas que pene-tran en los ganglios simpáticos, y hacen sinapsis conlos neuroblastos en desarrollo a este nivel. Algunas deestas fibras se extienden en lugares más altos o másbajos en las cadenas simpáticas o, a los gangliospreaórticos. Estas se denominan fibras preganglionaresy poseen vainas de mielina. Su función es estimular lascélulas de los ganglios simpáticos. Al salir de los ner-vios raquídeos hacia los ganglios simpáticos recibenel nombre de ramas comunicantes blancos. Esta co-lumna visceroeferente se extiende solo del primer seg-mento toráxico al segundo lumbar de la médula espinal(Fig. 15.35. B).
Los cilindroejes de las células ganglionares sim-páticas se denominan fibras posganglionares y care-cen de vaina de mielina. Se dirigen a otros niveles dela cadena simpática al corazón, los pulmones y el apa-rato digestivo. Existen otras fibras conocidas comoramos comunicantes grises que salen de la cadena sim-pática y se unen con los nervios raquídeos, de dondese dirigen a los vasos sanguíneos periféricos, el pelo ylas glándulas sudoríparas. Estos ramos comunicantesgrises se encuentran en todos los niveles de la médulaespinal.
Sistema nervioso parasimpáticoAl mismo tiempo que se forma el sistema sim-
pático, otro grupo de neuroblastos emigra hacia losórganos que han de inervar y forman los ganglios ter-minales, los cuales se localizan cerca o en el interiormismo del órgano inervado; su origen se discute. Seencuentran en la cabeza, el corazón, las vísceraspelvianas y los plexos submucosos y mientéricos deltubo digestivo. La acción de las fibras parasimpáticases antagónica con respecto a los efectos funcionalesdel sistema simpático.
Sistema cromafínAlgunas células de los ganglios se transforman
en células satélites y de neurilema, asociadas con lasneuronas, otras se transforman en elementos endocrinosque tienen afinidad por las sales de cromo, por lo cualse denominan células cromafines, tal afinidad es atri-buible a la presencia de la hormona epinefrina. El inte-grante principal de este sistema es la médulasuprarrenal.
Glándulas suprarrenalesLa glándula suprarrenal tiene doble origen; uno de
sus componentes es la porción mesodérmica, que for-ma la corteza y el otro, la porción ectodérmica, que ori-gina la médula.
En los peces, la corteza y la médula son órganosindependientes. En los animales más evolucionados sevan asociando progresivamente, hasta que en los ma-míferos la corteza en forma de cápsula envuelve a lamédula.
Corteza suprarrenal. En el embrión de cincosemanas, las células del epitelio celómico, situadas

244
Embriología especial
Fig. 15.36. Diferenciación de la glándula suprarrenal.
entre la raíz del mesenterio y las gónadas (Figs. 15.35. Ay 15.36), proliferan, se introducen en el mesénquimasubyacente y se convierten en órganos acidófilosvoluminosos denominados corteza fetal o primiti-va (Fig. 15.36. B). Muy pronto, esta corteza que-da envuelta por una segunda migración de célulasde igual origen que forma posteriormente la corte-za definitiva o permanente, característica de losprimates (Fig. 15.36. C y D).
Después del nacimiento, la corteza fetal comien-za a involucionar, mientras que la permanente crecehacia el centro.
Médula suprarrenal. Durante la séptima semana,células provenientes de los ganglios simpáticos del plexocelíaco comienzan a invadir el lado interno del esbozocortical. La migración continuada forma cúmulos celu-lares que ocupan una posición central en la glándula(Fig. 15.36). Poco antes del nacimiento cesa este procesoy el tejido cromafín queda agrupado en cordones y masas.