
Tema 14 de Biología NS Diploma BI Curso 2011-2013 · PDF fileTema 14 de Biología...
71
Tema 14 de Biología NS Diploma BI Curso 2011-2013 Metabolismo 1/70 Respiración celular y Fotosíntesis 3ª Parte: Fotosíntesis
Transcript of Tema 14 de Biología NS Diploma BI Curso 2011-2013 · PDF fileTema 14 de Biología...

Tema 14 de Biología NSDiploma BI
Curso 2011-2013
Metabolismo 1/70
Respiración celular y Fotosíntesis3ª Parte: Fotosíntesis

Metabolismo 2/70
Anabolismo� A partir de los precursores metabólicos obtenidos en las reacciones
catabólicas, la célula puede realizar la biosíntesis (anabolismo) de estructurasy moléculas propias más complejas, con consumo de energía (ATP) tambiénaportado por el catabolismo.
� Como la biosíntesis o anabolismo son reacciones de reducción, se necesitapoder reductor, también obtenido del catabolsimo en forma de NADH o deNADPH.
� Las reacciones de biosíntesistambién están catalizadasenzimáticamente y requieren unaporte energético proporcionadopor la hidrólisis del ATP.

Metabolismo
Metabolismo 3/70

Tipos de anabolismo
Metabolismo 4/70
� Se pueden distinguir dos tipos de anabolismo:
- Anabolismo autótrofo: Consiste en la síntesis de moléculas orgánicassencillas a partir de precursores inorgánicos tales como CO2, H2O y NH3. Sólola realizan las células autótrofas y existen dos modalidades:
* Fotosíntesis: que utiliza la energía de la luz (células fotolitotrofas).
* Quimiosíntesis: que utiliza la energía liberada en reaciones redox(células quimiolitotrofas).
- Anabolismo heterótrofo: Consiste en la síntesis de moléculas orgánicasprogresivamente más complejas a partir de otras más sencillas. Todas lascélulas pueden llevarlo a cabo (incluídas las autótrofas). Utilizan la energíadel ATP y coenzimas reducidos obtenidos del catabolismo.

Fotosíntesis
Metabolismo 5/70
� Concepto: Proceso anabólico autótrofo por el que las células vegetales, lasalgas y algunas bacterias utilizan sustancias inorgánicas y energía luminosa,que absorben por los pigmentos, para transformarlas en sustancias orgánicas.
� Se pueden presentar por la siguiente reacción de oxidorreducción:
H2X + Y H2Y + X
sustancia reducida + molécula inorgánica molécula orgánica + sustancia oxidada
- En las células vegetales, las algas y algunas bacterias, la sustancia X esel oxígeno, H2X es agua y el producto final X es el O2. Y puede ser CO2 o salesminerales, que se reducen con el H del agua para dar moléculas orgánicas H2Y.Este proceso se llama fotosíntesis oxigénica.
- En otro grupo de bacterias, la sustancia X puede ser azufre, por lo que elreactivo inicial sería H2S y un producto final X sería azufre. Y puede ser CO2 osales minerales, que se reducen con el H hasta moléculas orgánicas. Como nose libera oxígeno, este proceso se llama fotosíntesis anoxigénica.

Etapas de la fotosíntesis
Metabolismo 6/70
� Para explicar la fotosíntesis oxigénica, vamos a suponer que la planta utilizacomo nutrientes CO2 y H2O, por lo que la materia orgánica resultante seránmoléculas hidrocarbonadas (glúcidos y lípidos sencillos).
� Para formar otros lípidos complejos, Aa y nucleótidos, que poseen nitrógeno,fósforo, etc., tendrán que incorporarlos como nitratos, fosfatos, etc.
� Su ecuación general es:
El oxígeno liberado enla fotosíntesis procededel agua, no del CO2.

Etapas de la fotosíntesis
Metabolismo 7/70
� La fotosíntesis transcurre en dos fases o etapas: La fase luminosa y la faseoscura.

Células 8/70
Micrografía electrónicas de cloroplastosMembrana cloroplastídica(interna y externa)
Estroma(ribosomas 70S y ADN circular) Reacciones independientes de la luz
Membrana tilacoidal
Agrupación de tilacoides (grana) Reacciones dependientes de luz
Gránulo almidón Gota lipídica
Reserva de energía química(productos de la fotosíntesis)

Células 9/70
Micrografía electrónicas de cloroplastos� Rotula las regiones señaladas y calcula la magnificación de la imagen.

Fase Luminosa (reacciones dependientes de luz)
Metabolismo 10/70
� Conjunto de reacciones dependientes de la luz.
� Tiene lugar en la membrana de los tilacoides de los cloroplastos, donde selocalizan los fotosistemas y otros complejos multiproteicos que participan enla captación de la energía lumínica y su conversión en energía química.
� Consta de dos acontecimientos inportantes, de los que derivan 3consecuencias fundamentales:
- Fotolisis del agua: La luz rompe la molécula de agua, liberándose O2(consecuencia 1) a la atmósfera y reteniendo el H por el coenzima NADP, quepasa a su forma reducida NADPH (obtención poder reductor: consecuencia2).
- Fotofosforilación: La energía luminosa se emplea en la formación deATP (consecuencia 3).
� La reacción que representa esta fase sería:
2H2O + 2 NADP+ + ADP + Pi + LUZ O2 +2 NADPH + ATP

H2O
Reacciónesdependientes
de la luz
Cloroplasto
Luz
ATP
NADPH
O2
Fase Luminosa
Metabolismo 11/70
Fotolisis
Fotofosforilación
Video4

Fase Oscura (reacciones independientes de la luz)
Metabolismo 12/70
� Conjunto de reacciones independientes de la luz, pero que se realizan conluz y sin ella.
� Tiene lugar en el estroma de los cloroplastos y en ella ocurre la reducciónde la materia inorgánica, que se convierte en orgánica.
� El H necesario lo proporciona el NADPH y la energía el ATP, que se produjeronen la fase luminosa.
� La reacción que representa esta fase sería:
CO2 + 2 NADPH + ATP (CH2O) + 2 NADP+ + ADP + Pi
� El CH2O representa una molécula orgánica hidrocarbonada que podría ser unmonosacárido, la glicerina, un ácido graso, etc.

H2O
Reacciones dpendientes
de la luz
Cloroplasto
Luz
ATP
NADPH
O2
NADP+
CO2
ADPP+ i CICLO de
CALVIN
[CH2O](azúcar)
Fase Oscura
Metabolismo 13/70
Web westlake city

Espectro electromagnético
Metabolismo 14/70
� La luz es una forma de energía electromagnética, también denominadaradiación electromagnética.
� Al igual que otros tipos de energía electromagnética, la luz viaja medianteondas rítmicas.
� La longitud de onda es ladistancia entre las crestas de dosondas consecutivas. La longitudde onda determina el tipo deenergía electromagnética.
� La luz también se comporta comosi consistiera de partículasdiscretas, denominadas fotones.

Espectro de luz visible
Metabolismo 15/70
� La luz del sol está compuesta por unconjunto de radiaciones con distintaslongitudes de onda.
� Sólo una pequeña parte del espectroelectromagnético es perceptible por elojo humano, conocida como espectrovisible (400-700 nm), donde seencuentran los colores.
Longitud de onda
Video2

Pigmentos fotosintéticos
Metabolismo 16/70
� Los pigmentos son sustancias que absorben la luz visible. Existen diferentespigmentos que absorben diferentes longitudes de onda.
� Las longitudes de onda que no son absorbidas son reflejadas o transmitidas. Lashojas son verdes porque la clorofila refleja y transmite la luz verde.
� La clorofila es el más importante de los pigmentos fotosintéticos. Es unamolécula formada por un anillo porfirínico, semejante al grupo hemo de lahemoglobina, pero con un ión Mg2+ en lugar de Fe3+, unido a un terpeno (fitol).

Espectro de luz visible
Metabolismo 17/70
Luz blanca = todos loscolores (longitudes de onda)
La luz verde esreflejada
Luz de longitudde onda roja yazul es absorbida
¿Por qué aparecen las hojas de color verde?
La clorofila es el principalpigmento fotosintético.

Espectro de luz visible
Metabolismo 18/70
Cloroplasto
LuzLuz
reflejada
Luzabsorbida
Luz transmitida
Grana

Espectro de acción
Metabolismo 19/70
� ¿Pueden las plantas usar toda laradiación procedente del Solpara hacer la fotosíntesis?
� El espectro de acción muestrael rango de longitudes de ondadel espectro visible que puedenusarse en las reaccionesdependientes de luz.
� Las longitudes de ondacorrespondientes con el azul yel rojo-anaranjado producenla mayor tasa fotosintética.
Violeta-azul verde-amarillo naranja-rojo

Espectro de absorción
Metabolismo 20/70
� El espectro de absorción muestra el rango de longitudes de onda del espectrovisible que son absorbidos por la clorofila, el principal pigmento fotosintético.
� La clorofila a y bson químicamentesemejantes.
� El espectro de acción dela fotosíntesis y el deabsorción de la clorofilasolapan, mostrandoque la clorofila es elpigmento fotosintéticomás importante.
We Universidad AberdeenVideo3

Espectro de absorción
Metabolismo 21/70
� Otros pigmentos funcionancomo pigmentos antena(carotenos y ficobilinas).
Video4

Reacciones dependientes de luz: Fotosistemas
Metabolismo 22/70
� Los fotosistemas son las unidades de las membrananas tilacoidales en lasque se produce la captación de la energía solar y su utilización para liberarelectrones.
� Cada fotosistema está formado por un complejo antena y por un centro dereacción.
� El complejo antena es un centro colector de luz especialmente diseñado paracaptar energía luminosa y que está formado por cientos de moléculas declorofila y otros pigmentos accesorios.

Tilacoide
FotónComplejosantena
FotosistemaCentro dereacción
ESTROMA
Aceptor primariode electrones
e–
Transferenciade energía
Moléculasespecialesde clorofila a
Moléculas depigmentos
ESPACIOTILACOIDAL
Mem
bran
atil
acoi
dal
Reacciones dependientes de luz: Fotosistemas
Metabolismo 23/70
� Cuando una de las moléculasdel complejo antena recibe unfotón, se excita, transfiriendoesta energía de excitación aotra molécula cercana porresonancia, y en una reacciónen cadena, esa energía llegahasta el centro de reacción.
� El centro de reacción estásituado en una proteínatransmembrana y tiene dosmoléculas especiales declorofila a que dirigen los e-hacia la cadena de transporteelectrónico en la membranatilacoidal.

Reacciones dependientes de luz: Fotosistemas
Metabolismo 24/70
� En la membrana de los tilacoides de los cloroplastos de las células vegetalessuperiores hay dos tipos de fotosistemas:
� PSI se localiza en la membrana de los tilacoides en contacto con el estroma.Su centro de reacción contiene dos moléculas de clorofila a denominadas P700,puesto que tienen su máxima absorción de luz a una longitud de onda de 700nm.
� PSII se localiza en la membrana de los tilacoides que se apilan para formarlos grana. Su centro de reacción contiene dos moléculas de clorofila adenominadas P680, puesto que tienen su máxima absorción de luz a unalongitud de onda de 680 nm. Aquí se lleva a cabo la reación de oxidación porel que el agua da lugar a O2 por acción de una proteína llamada complejoproductor de O2.

Metabolismo 25/70
� Los dos fotosistemas pueden actuar en serie, con lo que se produce un flujolineal de electrones desde la molécula de agua (donador inicial de e-) hasta elNADP+ (aceptor final de e-), que es reducido a NADPH.
� En el PSII se encuentra el complejo productor de O2, que puede separarlos e- y los protones de los átomos de H de una molécula de agua situada enla cara interna de la membrana tilacoidal, liberando O2.
Reacciones dependientes de luz: Fotofosforilación no cíclica
� Los e- del agua son cedidos ala molécula de clorifila a delPSII, ya que pierden sus e-por acción de la luz, a medidaque los fotones inciden en loscomplejos antena y la energíallega hasta ella.
� El proceso de rotura del agua,para que la clorofila arecupere sus e- perdidos porla luz, en oxígeno, protones yelectrones se denominafotolisis del agua.
Video5

Metabolismo 26/70
� Los e- liberados por la clorofila a del PSII entran en la cadena de transportede electrones tilacoidal, siendo captados en primer lugar por la feofitina(FeO).
� Los e- son transportados siguiendo el gradiente de potencial redox, hasta quela Plastocianina (PC) los cede a la clorofila a del PSI, que a su vez habíaperdido sus e- al recibir la energía procedente de la incidencia de un fotón ensu complejo antena.
� El PSI cede los e- a laFerredoxina (Fd), quejunto con loshidrógenos necesariosllegan hasta el NADP+
que se reduce a NADPH(poder reductor).
Reacciones dependientes de luz: Fotofosforilación no cíclica

Metabolismo 27/70
� El transporte de e- en la membrana del tilacoide produce la energía necesariapara que se bombeen H+ desde el estroma hasta el interior del tilacoide.
� Es el complejo citocromo b/f el que funciona como una bomba que introduceprotones en el espacio tilacoidal, creando un gradiente electroquímico.
� La hipótesis quimiosmóticade Mitchell explica la síntesisde ATP de forma muysemejante a como ocurre en lamitocondria.
� Cuando los H+ salen a favor degradiente desde el interior deltilacoide, a través del canal deprotones de la ATP sintasa,se libera nergía que se utilizapara fabricar ATP.
ESTROMA
ESPACIO TILACOIDAL
Reacciones dependientes de luz: Fotofosforilación no cíclica

Metabolismo 28/70
Reacciones dependientes de luz: Fotofosforilación no cíclica
Animación1

Metabolismo 29/70
Light P680
e–
Photosystem II(PS II)
Primaryacceptor
[CH2O] (sugar)
NADPH
ATP
ADP
CALVINCYCLE
LIGHTREACTIONS
NADP+
Light
H2O CO2
Ene
rgy
ofel
ectr
ons
O2
Reacciones dependientes de luz: Fotofosforilación no cíclica

Light P680
e–
Photosystem II(PS II)
Primaryacceptor
[CH2O] (sugar)
NADPH
ATP
ADP
CALVINCYCLE
LIGHTREACTIONS
NADP+
Light
H2O CO2
Ene
rgy
ofel
ectr
ons
O2
e–
e–
+2 H+
H2O
O21/2
Metabolismo 30/70
Reacciones dependientes de luz: Fotofosforilación no cíclica

Metabolismo 31/70
Light P680
e–
Photosystem II(PS II)
Primaryacceptor
[CH2O] (sugar)
NADPH
ATP
ADP
CALVINCYCLE
LIGHTREACTIONS
NADP+
Light
H2O CO2
Ene
rgy
ofel
ectr
ons
O2
e–
e–
+2 H+
H2O
O21/2
Pq
Cytochromecomplex
Pc
ATP
Reacciones dependientes de luz: Fotofosforilación no cíclica

Light P680
e–
Photosystem II(PS II)
Primaryacceptor
[CH2O] (sugar)
NADPH
ATP
ADP
CALVINCYCLE
LIGHTREACTIONS
NADP+
Light
H2O CO2
Ene
rgy
ofel
ectr
ons
O2
e–
e–
+2 H+
H2O
O21/2
Pq
Cytochromecomplex
Pc
ATP
P700
e–
Primaryacceptor
Photosystem I(PS I)
Light
Metabolismo 32/70
Reacciones dependientes de luz: Fotofosforilación no cíclica

LightP680
e–
Photosystem II(PS II)
Primaryacceptor
[CH2O] (sugar)
NADPH
ATP
ADPCALVINCYCLE
LIGHTREACTIONS
NADP+
Light
H2O CO2
Ene
rgy
ofel
ectr
ons
O2
e–
e–
+2 H+
H2O
O21/2
Pq
Cytochromecomplex
Pc
ATP
P700
e–
Primaryacceptor
Photosystem I(PS I)
e–e–
NADP+
reductase
Fd
NADP+
NADPH+ H+
+ 2 H+
Light
Metabolismo 33/70
Reacciones dependientes de luz: Fotofosforilación no cíclica

ATP
Photosystem II
e–
e–
e–e–
MillmakesATP
e–
e–
e–
Photosystem I
NADPH
Metabolismo 34/70
Reacciones dependientes de luz: Fotofosforilación no cíclica

Metabolismo 35/70
� El PSI puede funcionar independientemente del PSII, en cuyo caso se realizaun transporte cíclico de los electrones.
� En este caso, los e- del PSI son cedidos desde la Fd al cit b/f, volviendo alPSI que los perdió.
� De esta manera se produceATP sin que haya producciónde NADPH ni de O2, siendonecesario para cubrir lasnecesidades energéticas dela fase oscura, donde seconsume más cantidad deATP que de NADH.
Animación2
Reacciones dependientes de luz: Fotofosforilación cíclica

Metabolismo 36/70
Fotofosforilación vs Fosforilación oxidativa� Similitudes:
- Bombean iones H+ al espacio intermembrana/tilacoidal para generar ungradiente de concentración de protones.
- Implica una cadena de transporte de electrones embebida en la membrana delas crestas/tilacoides.
- La energía se libera cuando los e- se intercambian de un transportador a otro.
- Usan la ATPsintasa para generar ATP a partir de la quimiosmosis de los H+.
� Diferencias (Respiración Celular):
- Los transportadores de e- (NADH yFADH2) proveen los e- y H+ .
- El oxígeno es el aceptor final deelectrones (y H+), produciéndoseagua como producto (fosforilaciónoxidativa).
- No utiliza la luz.
- Usa citocromos.
� Diferencias (Fotosíntesis):
- La fotolisis del agua provee lose- y H+.
- El NADP+ es el aceptor final deelectrones (y H+), produciéndoseNADPH para ser usado en elciclo de Calvin.
- Utiliza la luz.
- Usa fotosistemas.

Metabolismo 37/70
� Ocurre en el estroma de los cloroplastos.
� Fase biosintética en el que el NADPH y el ATP producido en la fase luminosa seemplea para reducir moléculas inorgánicas sencillas que tienen un alto estadode oxidación, como el CO2, nitrato (NO3
-) o sulfato (SO42-).
Fase oscura: Reacciones independientes de la luz
� Estas reducciones serealizan conindependencia de la luzy permite a las célulasobtener compuestosorgánicos sencillos.
� Uno de los procesos defijación y reducción másimportantes es el delCO2, que transcurre enuna serie de reaccionesconocidas como ciclode Calvin.

Metabolismo 38/70
� También conocido como ciclo/metabolismo C3, ya que tanto el pimerproducto de la fijación del C como la mayoría de los metabolitos intermediariostienen 3C, es un proceso cíclico que se produce en el estroma del cloroplasto,donde se fija el carbono atmosférico (CO2) a la materia orgánica. Sediferencian tres etapas.
Reacciones independientes de luz: Ciclo de Calvin
Carboxilación
Reducción
Regeneración

Metabolismo 39/70
1) Carboxilación. La enzima ribulosa 1,5 bifosfato carboxilasaoxigenasa (RuBisCO) cataliza la combinación de la ribulosa 1,5 bifosfato(5C) con el CO2, formándose un compuesto intermedio e inestable (6C), quese descompone en dos moléculas de 3-fosfoglicérico (3C).
Carboxilación
Reacciones independientes de luz: Ciclo de Calvin

Metabolismo 40/70
2) Reducción. Mediante la energía que suministra el ATP y los hidrógenosque suministra el NADPH, el fosfoglicérico se reduce a gliceraldehído 3-fosfato (triosa fosfato).
Reducción
X 2
X 2
Reacciones independientes de luz: Ciclo de Calvin
Metabolismo 41/70
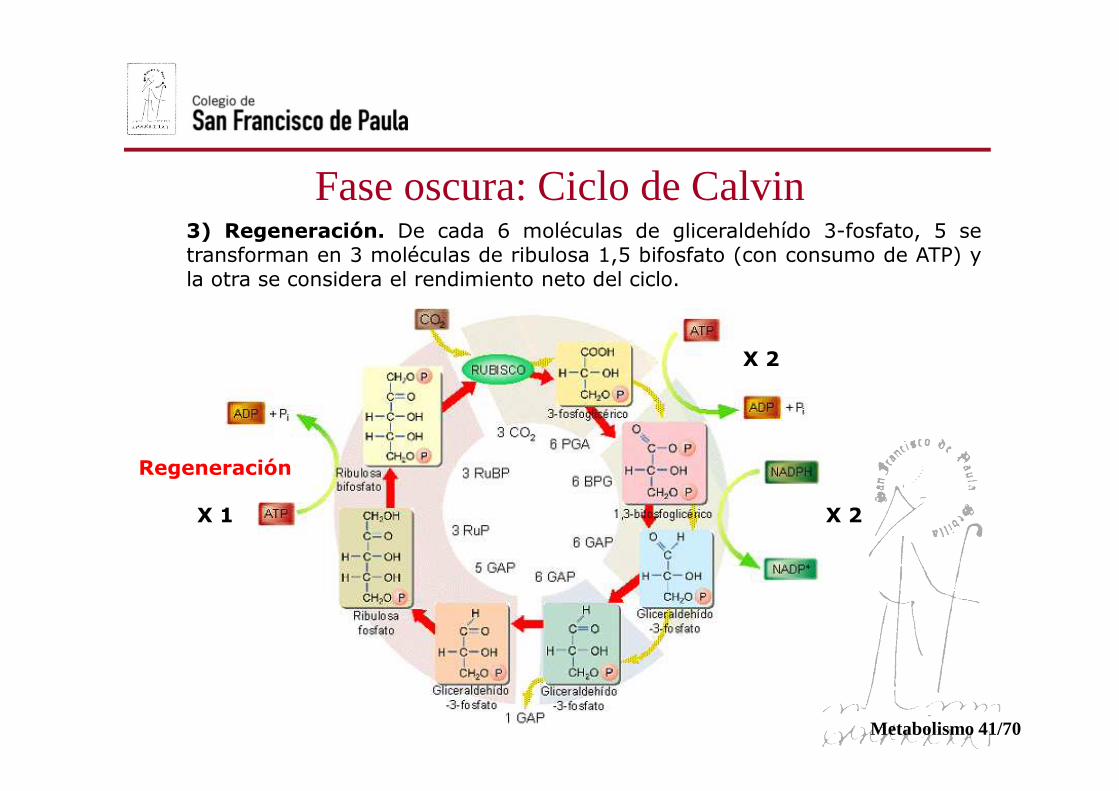
3) Regeneración. De cada 6 moléculas de gliceraldehído 3-fosfato, 5 setransforman en 3 moléculas de ribulosa 1,5 bifosfato (con consumo de ATP) yla otra se considera el rendimiento neto del ciclo.
Fase oscura: Ciclo de Calvin
Regeneración
X 2
X 2X 1
Metabolismo 42/70
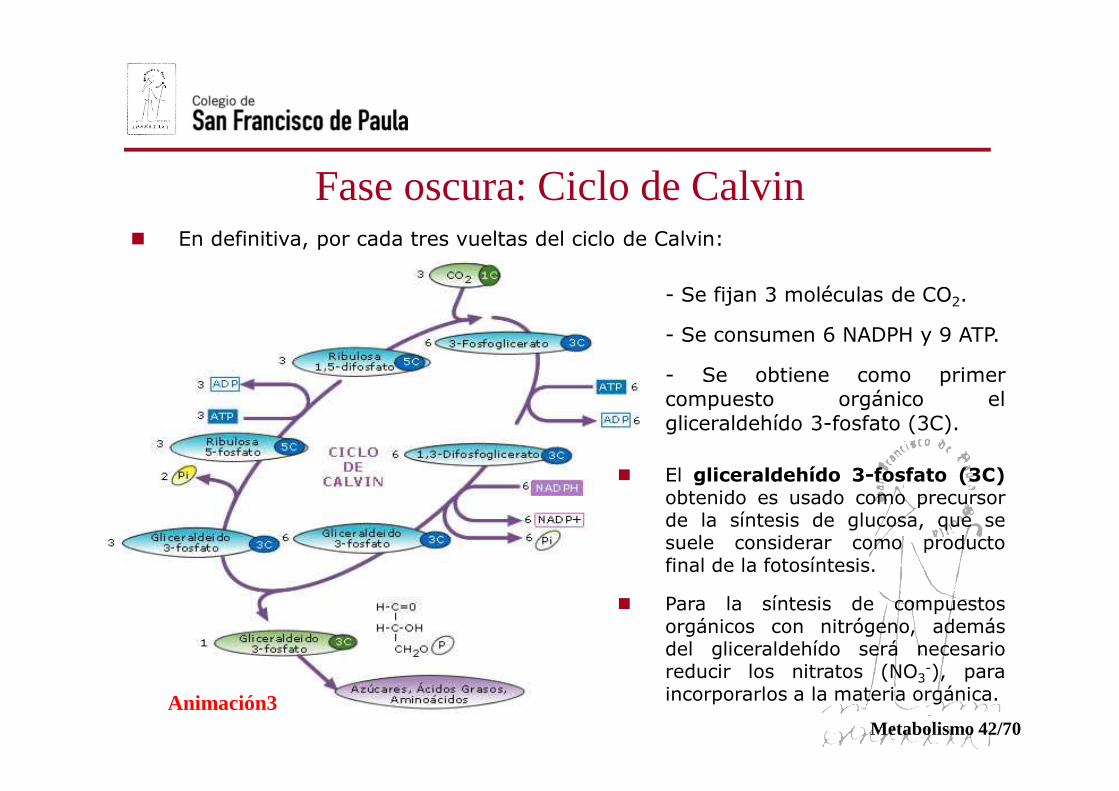
� En definitiva, por cada tres vueltas del ciclo de Calvin:
Fase oscura: Ciclo de Calvin
- Se fijan 3 moléculas de CO2.
- Se consumen 6 NADPH y 9 ATP.
- Se obtiene como primercompuesto orgánico elgliceraldehído 3-fosfato (3C).
� El gliceraldehído 3-fosfato (3C)obtenido es usado como precursorde la síntesis de glucosa, que sesuele considerar como productofinal de la fotosíntesis.
� Para la síntesis de compuestosorgánicos con nitrógeno, ademásdel gliceraldehído será necesarioreducir los nitratos (NO3
-), paraincorporarlos a la materia orgánica.Animación3

Metabolismo 43/70
� Podemos deducir por tanto, que en el ciclo de Calvin, por cada molécula deCO2 fijado, se consumen 2 NADPH y 3 ATP.
Fase oscura: Ciclo de Calvin
CO2
RuBP (C5) 2 Fofoglicerato (C3)
Gliceraldehído 3-P+
Gliceraldehído 3-P
(C3)
2 ATP + 2 NADPH
2 ADP + 2 Pi + 2 NAD+
Síntesis de glúcidos
1 ATP
ADP + Pi
CarboxilaciónRuBisCO
ReducciónRegeneración(3 vueltas)

Metabolismo 44/70
� Toda la energía producida en la Fase luminosa es consumida durante la Faseoscura para la fijación del carbono atmosférico desde moléculas inorgánicasoxidadas (CO2) a moléculas orgánicas simples y reducidas (gliceraldehído-3P).
Balance de la Fotosíntesis

Metabolismo 45/70
� Membrana de los tilacoides y granas:- Presenta un gran área superficial para la absorciónde luz y las reacciones dependientes de ellas.- Fotosistemas y ATPasa unidas a membrana.- Alta densidad de clorofilas para la absorción de luz
Relación estructura-función del cloroplasto
� Pequeño volumen espacios tilacoidales:- Rápida generación del gradiente de H+
para quimiosmosis.� Estroma rico en enzimas:
- Contiene la enzima RuBisCO para lacarboxilación de la RuBP.- Fluido para una fácil difusión.
� Alta densidad de cloroplastosen el parénquima en
empalizada:- Eficiente absorción de la luz.
Metabolismo 46/70
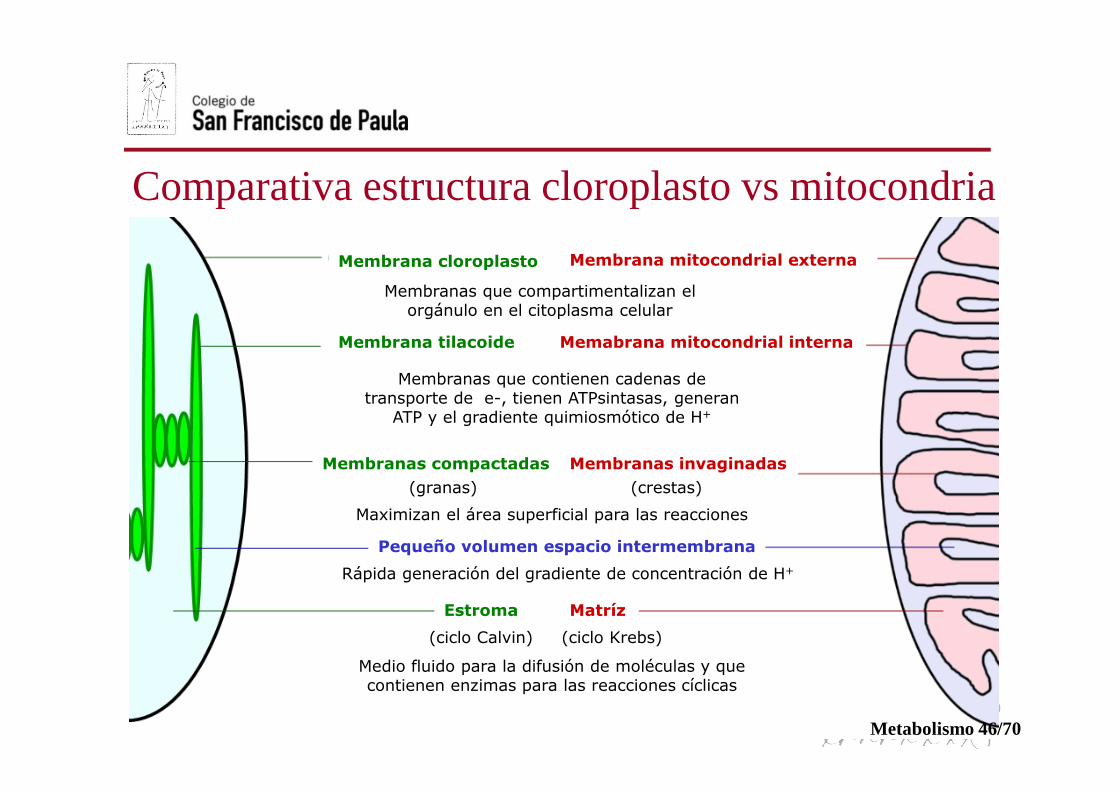
Comparativa estructura cloroplasto vs mitocondria
Membrana cloroplasto
Membrana tilacoide
Membranas compactadas
Estroma Matríz
Membranas invaginadas
Memabrana mitocondrial interna
Membrana mitocondrial externa
Pequeño volumen espacio intermembrana
Membranas que compartimentalizan el orgánulo en el citoplasma celular
Membranas que contienen cadenas de transporte de e-, tienen ATPsintasas, generan
ATP y el gradiente quimiosmótico de H+
Rápida generación del gradiente de concentración de H+
Maximizan el área superficial para las reacciones
(granas) (crestas)
(ciclo Calvin) (ciclo Krebs)
Medio fluido para la difusión de moléculas y que contienen enzimas para las reacciones cíclicas

Metabolismo 47/70
Comparativa estructura cloroplasto vs mitocondriaDiferenciasestructurales
* La membrana interna mitocondrial forma crestas internas y la plastidial no; * Los cloroplastos presentan tilacoides y las mitocondrias no; * Los cloroplastos presentan fotosistemas I y II y las mitocondrias no.
Semejanzasestructurales
* Doble membrana, * Espacio intermembrana, * Matriz o estroma, * ADN circular, * Ribosomas 70S, * ATP sintasas.
Diferenciasfuncionales
* Ciclo de Calvin / ciclo de Krebs;* Fuente de energía lumínica / energía de reacciones químicas; * Obtención de electrones del H2O / obtención de electrones de compuestos orgánicos; * Productos finales de la respiración [CO2, NADH + H+, FADH2, GTP (ATP)] / productos finales de la fotosíntesis (O2, triosa).
Semejanzas funcionales
* División por bipartición, * Cadena de transporte de electrones, * Síntesis de ATP, * Síntesis propia de proteínas.

Metabolismo 48/70
� La enzima RuBisCO también puede catalizar la unión de O2 a la RuBP, en unproceso que desprende CO2 y que se conoce como fotorrespiración.
� Este proceso ocurre porque el O2 compite con el CO2 como sustrato de laenzima, que también puede comportarse como oxigenasa además decarboxilasa.
Fotorrespiración

Metabolismo 49/70
Fotorrespiración
Estoma cerradoEstoma abiertoO2 CO2
CO2 O2
ribulosa biP + O2
ác. Fosfoglicérico (3C) + ác. Fosfoglicólico (2C)
ciclo de Calvin oxidación
oxidación
CO2 + otros productos orgánicos
cloroplasto peroxisoma
rubisco
� En un ambiente cálido y seco, se cierran los estomas para evitar la pérdida deagua, con acumulación de O2 y escasez de CO2. La RuBisCO oxida la RuBP.
� La RuBisCO oxida la RuBPgenerando una moléculade 2C (fosfoglicólico) yotra de 3C, en lugar dedos de 3C.
� El fosfoglicólico se oxida hasta CO2 en los peroxisomas, es decir, se consumeoxígeno y se produce CO2 (fotorrespiración), y el fosfoclicérico va al Calvin,pero sólo 1 molécula, por lo que funciona a la mitad de su eficacia (limitación).

Metabolismo 50/70
Fotorrespiración� Para solucionar este problema, las plantas de climas secos y cálidos realizan la
ruta de Hatch-Slack de las plantas C4.
� Las plantas C3 tienen un su paréquima un sólo tipo de células fotosintéticas,sin embargo, en las plastas C4 hay dos tipos de células en las hojas, las célulasenvolventes del haz vascular (de la vaina) y las del mesófilo (parénquima).

Metabolismo 51/70
Fotorrespiración� Las células envolventes el haz vascular
son más internas y contienen la RuBisCO parafijar el CO2 mediante el ciclo de Calvin.
� Las células del mesófilo son más externasy están en contacto directo con las anteriores,que no poseen RuBisCO y poseen la rutaHatch-Slack, por la que fijan el CO2 en uncompuesto de 3C, el fosfoenolpiruvato, dandouno de 4C.
� Este ácido de 4C se transforma en ácidomálico que pasa a las células envolventes(vaina) donde se descarboxila y cede el CO2al Calvin.
� Este sistema supone una ventaja adaptativapara estas plantas que viven en ambientessecos (plantas tropicales y muchos cereales),donde la fotorrespiración sería un graveproblema. Lo evitan concentrando todo el CO2posible en las células de la vaina.

Metabolismo 52/70
¿Cómo medir la tasa de fotosíntesis?� La tasa de fotosíntesis puede ser determinada directamente a partir de:
1) La producción de oxígeno.
- Contando burbujas de oxígeno
- Colectándolo en un jeringa
- Usando un sensor de O2 (data-logger)

Metabolismo 53/70
¿Cómo medir la tasa de fotosíntesis?2) La fijación de dióxido de carbono usando un sensor CO2 (data-logger).

Metabolismo 54/70
¿Cómo medir la tasa de fotosíntesis?� La tasa de fotosíntesis puede ser determinada indirectamente por el
incremento de biomasa.

Metabolismo 55/70
Factores que limitan la tasa de fotosíntesis� Varios son los factores que pueden afectar a la tasa de fotosíntesis, y cuyos
niveles deben encontrase dentro de un determinado valor para que lafotosínstesis sea óptima.
� El término factor limitante se usa para describir el factor que controla unareacción particular a su máximo rendimiento.

Metabolismo 56/70
Factores que limitan la tasa de fotosíntesis� Concentración de CO2: La fotosíntesis es una ruta metabólica donde una
enzima cataliza la carboxilación de la RuBP. Si hay bajos niveles de CO2, seproduce menos gliceraldehído-3P.
� La concentración de CO2 afecta a la fase oscura (independiente de luz). El CO2es uno sustrato en la reacción, por lo que actúa limitando la reacción al igualque cualquier otro sustrato en una reacción enzimática: A mayor CO2 mayoractividad fotosintética, hasta que se satura porque todos los sitios activosestán ocupados.
Un incremento de laconcentración implicaun incremento de latasa de reacción
El CO2 es un sustrato en la reacción, porlo que la curva se comporta como unacurva de concentración de sustrato
A altas concentraciones, un incrementoposterior no tiene efecto sobre la tasade reacción, al ser el CO2 fijado con lamáxima eficiencia
Tasa defotosíntesis

Metabolismo 57/70
� Intensidad de luz: La luz es necesaria para la fotoactivación de los e- y lafotolisis del agua. Por tanto, la intensidad de luz afecta a la fase luminosa(dependiente de luz).
� Bajas intensidades de luz resultan en una baja producción de NADPH + H+ y deATP, lo que provoca que se pare el ciclo de Calvin.
Un incremento enla intensidad deluz implica unincremento de latasa de reacción
A altas intensidades de luz, unincremento posterior no tiene efectosobre la tasa de reacción, al estartodos los cloroplastos funcionandocon la máxima eficienciaTasa de
fotosíntesis
Intensidad de luz
Factores que limitan la tasa de fotosíntesis

Metabolismo 58/70
� Temperatura: La fotosíntesis es una ruta metabólica catalizada por unaenzima. La temperatura afecta a la afecta las reacciones independientes deluz, ya que la temperatura afecta a la actividad enzimática.
� Bajas temperaturas ralentizan las reacciones debido a una menor energía,mientras que a medida que aumenta la temperatura, incrementa la tasa defotosíntesis, hasta que se alcanza la temperatura óptima, punto a partir delcual las enzimas pueden desnaturalizarle.
Un incremento implicaun incremento de la tasade reacción, ya que lasmoléculas presentanmás energía
Las enzimas sedesnaturalizan: elsito activo cambiay no funciona
Tasa defotosíntesis
Temperatura óptima
Temperatura
Factores que limitan la tasa de fotosíntesis
Video6

Metabolismo 59/70
Óptimo de tasa fotosintéticaen el rango del rojo y azul
Factores que limitan la tasa de fotosíntesis

Metabolismo 60/70
Factores limitantes y TdC� Para maximizar la producción agrícola, los agricultores manipulan los factores
limitantes y controlan el ambiente en el que se enuentran las plantas,permitiendo que crezcan en países de donde no son nativas.
� Existe una necesidad de realizarexperimentos controlados con sumocuidado. Si queremos investigar elefecto de un factor, hay que controlar elresto de factores que pudieran influir.
� Por ejemplo, los vegetarianos tienen una mayoresperanza de vida que las personas que comencarne; sería incorrecto concluir que comer carnereduce la esperanza de vida, a menos quepudiéramos demostrar que la única diferencia ennuestro ensayo entre el grupo de vegetarianos yel de las personas que comen carne fuera el deincluir carne en la dieta.
� En el ámbito de la fotosíntesisla situación es relativamentesimple y podemos garantizarque los factores diferentes delfactor objeto de nuestrainvestigación se mantienen aun nivel constante y óptimo.
En otros ámbitos los problemasque se plantean son muchomayores. En la mayoría de lasinvestigaciones en el campo dela salud humana casi siempreintervienen complejos factoresque complican la investigación.

Importancia biológica de la fotosíntesis
Metabolismo 61/70
� La fotosíntesis sirve como soporte de la vida en la Tierra, desde suorigen evolutivo en el pasado hasta la actualidad.
� Desde el punto de vista evolutivo, la aparición de la fotosíntesis sobre laTierra marcó un hito que condujo hacia la actual situación en nuestro planeta.
- El tipo de metabolismo más primitivo serían reacciones semejantes a lasactuales fermentaciones, ya que empleaba la abundante materia orgánicapara obtener energía en condiciones anaerobias.

Importancia biológica de la fotosíntesis
Metabolismo 62/70
- La adquisición por algunos seres del complejo molecular del fotosistema I,en un momento en el que la materia orgánica escaseaba, supuso podertransformar la energía lumínica en química por fotofosforilación cíclicaaprovechando el H2S como agente reductor para formar materia inorgánica apartir del CO2, NO2, etc.

Importancia biológica de la fotosíntesis
Metabolismo 63/70
- El éxito de la fotosíntesis anoxigénica se completaría con la aparición delfotosistema II, con el que poder emplear la fotolisis del agua para poder haceruna fotofosforilación acíclica más rentable, dando como producto de desechoel O2. Surgió así la fotosíntesis oxigénica, que pasó a ser dominante y queenriqueció la atmósfera e hidrosfera, permitiendo la aparición de organismosaerobios.

Importancia biológica de la fotosíntesis
Metabolismo 64/70
- Al mismo tiempo, el O2 sufrió el efecto oxidante de los rayos ultravioleta delSol, que hasta entonces impedía la vida fuera del agua, y se fuetransformando en ozono.
- La acumulación de ozono en la atmósfera produjo el efecto de filtro paraesas radiaciones, lo que facilitó la salida y posterior evolución de los seresvivos del medio acuático al áreo.

Importancia biológica de la fotosíntesis
Metabolismo 65/70
� En la actualidad la fotosíntesis sigue siendo el proceso responsable delmantenimiento del ciclo de materia y el flujo de energía en losecosistemas.
- Ciclo de materia: Intervieneen todos los ciclosbiogeoquímicos, transformandola materia inorgánica enorgánica y ligando así el medioabiótico y biótico, al tiempoque mantiene en equilibrio losprocesos aerobios de larespiración al producir oxígeno.
- Flujo de energía: la materiafabricada en la fotosíntesis esutilizada por los propios seresfotosintéticos y por losheterótrofos como fuente deenergía en al cadena trófica.

Importancia biológica de la fotosíntesis
Metabolismo 66/70
� La fotosíntesis es uno de los procesos bioquímicos más importante de laBiosfera por varios motivos:
1. La síntesis de materia orgánica a partir de la inorgánica se realizamediante la fotosíntesis y luego pasará de unos seres vivos a otrosmediante las cadenas tróficas, para ser transformada en materiapropia por los diferentes seres vivos.
2. Produce la transformación de la energía luminosa en energíaquímica, necesaria y utilizada por los seres vivos.
3. En la fotosíntesis se libera oxígeno, que será utilizado en larespiración aerobia.
4. La fotosíntesis fue causante del cambio producido en la atmósferaprimitiva, que era anaerobia y reductora.
5. De la fotosíntesis depende también la energía almacenada encombustibles fósiles como carbón, petróleo y gas natural.
6. El equilibrio necesario entre seres autótrofos y heterótrofos no seríaposible sin la fotosíntesis.

Tipos de anabolismo
Metabolismo 67/70
� Se pueden distinguir dos tipos de anabolismo:
- Anabolismo autótrofo: Consiste en la síntesis de moléculas orgánicassencillas a partir de precursores inorgánicos tales como CO2, H2O y NH3. Sólola realizan las células autótrofas y existen dos modalidades:
* Fotosíntesis: que utiliza la energía de la luz (células fotolitotrofas).
* Quimiosíntesis: que utiliza la energía liberada en reaciones redox(células quimiolitotrofas).

Metabolismo 67/70
� Proceso anabólico autótrofo por el que algunas bacterias (quimiolitotrofas=quimiosintéticas) transforman sustancias inorgánicas en orgánicas,utilizando como fuente de energía química de enlace procedente de reaccionesde oxidación a partir de moléculas inorgánicas que se encuentran en el mediocelular.
Quimiosíntesis
� Por analogía con la fotosíntesis, puede considerarse dividada en dos etapas:

Metabolismo 69/70
Quimiosíntesis
2.- Etapa biosintética donde se fija CO2 para formar distintos compuestoshidrocarbonados. Etapa semejante a la fase oscura de la fotosíntesis, donde sereduce el CO2 por reacciones semejantes al ciclo de Calvin.
1.- Obtención de poder reductor en forma de NADPH + H+ y energía enforma de ATP. Para ello, los electrones de los H procedentes de esas reaccionesde oxidación son llevados a una cadena de transporte de e- en la membranabacteriana, donde se libera energía, que se emplea para crear un gradiente deprotones y finalmente ATP por fosforilación.

Importancia biológica de la Quimiosíntesis
Metabolismo 70/70
� La importancia de la quimiosíntesis redica en dos aspectos:
1. Cierran los ciclosbiogeoquímicos en elecosistema al terminar demineralizar la materia orgánicay ponen a disposición de losorganismos fotosintéticos lossulfatos, nitratos, etc., paraque puedan formar másmateria orgánica.
2. Constituyen el nivel de losproductores en pequeños ecositemassituados en hábitats extremos (chimeneashirotermales).

Metabolismo 70/70
Integración del Catabolismo y del Anabolismo


















