
TENDENCIAS POBLACIONALES DE LA AVIFAUNA MADRILEÑA: DIEZ ... · primavera, entre mediados de abril...
20
I N T R O D U C C I Ó N E n el año 2005 se han cumplido los primeros diez años de muestreo de aves dentro del programa SACRE (acróni- mo de “Seguimiento de Aves Comunes Reproductoras en España”) que en 1996 puso en marcha SEO/BirdLife (figu- ra 1). Su objetivo principal es proporcionar estimas numéri- cas de la evolución temporal de las poblaciones de aves nidi- ficantes a escala nacional. Este tipo de información sólo estaba disponible hasta ahora para un reducido número de especies socialmente “emblemáti- cas” y fácilmente censables en gran parte de su área de distri- bución (véanse las síntesis existentes, por ejemplo, para el águi- la imperial ibérica Aquila adalberti —González y Oria 2001—, el quebrantahuesos Gypaetus barbatus —Antor 2001—, la cigüe- ña blanca Ciconia ciconia —Molina y Del Moral 2005—, o la avutarda común Otis tarda —Alonso et al. 2005). Desconocer esta tendencia para la inmensa mayoría de las especies dificulta mucho la cuantificación objetiva de su grado de rareza y de su vulnerabilidad, dos parámetros imprescindibles para establecer las prioridades de conservación (UICN 2001; Madroño et al. 2004). No obstante, gracias a los muestreos estandarizados realizados anualmente por los participantes del programa SACRE, actualmente podemos inferir con bastante fiabilidad la evolución de los efectivos poblacionales de casi el 50% de las especies reproductoras de España (www.seo.org/?seguimiento- deaves), para la mayoría de las cuales este dato era hasta ahora puramente especulativo o totalmente desconocido. Puesto que la Comunidad de Madrid es una provincia privi- legiada en cuanto al número de aficionados y su predisposi- David PALOMINO 1 Virginia ESCANDELL 2 Juan Carlos DEL MORAL 3 SEO/BirdLife C/ Melquiades Biencinto 34 28053 Madrid (1) Autor para correspondencia [email protected] (2) [email protected] (3) [email protected] R E S U M E N Se han analizado los datos obtenidos en el programa SACRE en la Comunidad de Madrid entre 1996 y 2005. El objetivo es identificar la ten- dencia temporal de las pobla- ciones de aves nidificantes a escala regional, un parámetro imprescindible para establecer adecuadamente prioridades de conservación. Los muestreos han sido realizados por colabo- radores voluntarios basándose en una misma metodología, basada en puntos de escucha repetidos anualmente. Los resultados indican un predo- minio de especies con tenden- cias poblacionales positivas. Algunas especies presentan diferentes evoluciones en sub- TENDENCIAS POBLACIONALES DE LA AVIFAUNA MADRILEÑA 24 TENDENCIAS POBLACIONALES DE LA AVIFAUNA MADRILEÑA: DIEZ AÑOS DE PROGRAMA SACRE
Transcript of TENDENCIAS POBLACIONALES DE LA AVIFAUNA MADRILEÑA: DIEZ ... · primavera, entre mediados de abril...

I N T R O D U C C I Ó N
E n el año 2005 se han cumplido los primeros diez años demuestreo de aves dentro del programa SACRE (acróni-mo de “Seguimiento de Aves Comunes Reproductoras en
España”) que en 1996 puso en marcha SEO/BirdLife (figu-ra 1). Su objetivo principal es proporcionar estimas numéri-cas de la evolución temporal de las poblaciones de aves nidi-ficantes a escala nacional.
Este tipo de información sólo estaba disponible hasta ahorapara un reducido número de especies socialmente “emblemáti-cas” y fácilmente censables en gran parte de su área de distri-bución (véanse las síntesis existentes, por ejemplo, para el águi-la imperial ibérica Aquila adalberti—González y Oria 2001—,el quebrantahuesos Gypaetus barbatus—Antor 2001—, la cigüe-ña blanca Ciconia ciconia —Molina y Del Moral 2005—, o laavutarda común Otis tarda —Alonso et al. 2005). Desconoceresta tendencia para la inmensa mayoría de las especies dificultamucho la cuantificación objetiva de su grado de rareza y de suvulnerabilidad, dos parámetros imprescindibles para establecerlas prioridades de conservación (UICN 2001; Madroño et al.2004). No obstante, gracias a los muestreos estandarizadosrealizados anualmente por los participantes del programaSACRE, actualmente podemos inferir con bastante fiabilidadla evolución de los efectivos poblacionales de casi el 50% de lasespecies reproductoras de España (www.seo.org/?seguimiento-deaves), para la mayoría de las cuales este dato era hasta ahorapuramente especulativo o totalmente desconocido.
Puesto que la Comunidad de Madrid es una provincia privi-legiada en cuanto al número de aficionados y su predisposi-
David PALOMINO1
Virginia ESCANDELL2
Juan Carlos DEL MORAL3
SEO/BirdLifeC/ Melquiades Biencinto 34
28053 Madrid(1) Autor para [email protected](2) [email protected](3) [email protected]
R E S U M E N
Se han analizado los datosobtenidos en el programaSACRE en la Comunidad deMadrid entre 1996 y 2005. Elobjetivo es identificar la ten-dencia temporal de las pobla-ciones de aves nidificantes aescala regional, un parámetroimprescindible para estableceradecuadamente prioridades deconservación. Los muestreoshan sido realizados por colabo-radores voluntarios basándoseen una misma metodología,basada en puntos de escucharepetidos anualmente. Losresultados indican un predo-minio de especies con tenden-cias poblacionales positivas.Algunas especies presentandiferentes evoluciones en sub-
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
24
TENDENCIAS POBLACIONALES DE LAAVIFAUNA MADRILEÑA : D IE Z AÑOS DE
PROGRAMA SACRE

ción a colaborar en las iniciativas promovidas porSEO/BirdLife, los datos acumulados tras estos diez años deseguimiento son particularmente abundantes. Hay quecomentar que en los análisis de tendencias poblacionales amenudo se suelen descartar los primeros años de muestreos,pues se supone que debido a un “efecto de aprendizaje” losdatos recogidos pueden ser demasiado imprecisos. No obs-tante, creemos que en el caso concreto de Madrid esta cir-cunstancia es poco relevante, pues desde sus inicios el equi-po de colaboradores regulares ha estado integrado en granmedida por aficionados muy cualificados (o incluso porexpertos ornitólogos profesionales).
Así, en este trabajo se resumen brevemente las tendenciastemporales de un gran número de especies nidificantes en laComunidad de Madrid entre 1996 y 2005, con el fin deaportar nueva información cuantitativa y de corte macroeco-lógico que ayude a evaluar el estatus de conservación denuestra avifauna (véanse también en esta línea: Carrascal et al.2003, 2006).
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
25
regiones geográficas distintas(es decir, área de la sierra oárea de la fosa del Tajo). Porgrupos ecológicos, las avesligadas a medios urbanos, asícomo las forestales y genera-listas han experimentado unatendencia claramente positiva.En cambio, las de mediosabiertos y arbustivos se hanmantenido estables durantetoda la década de estudio.
PALABRAS CLAVE: avesreproductoras, prioridadesde conservación, evoluciónpoblacional.

M É T O D O S
E l trabajo de campo consiste en la realización de muestre-os de 5 minutos de duración, registrando todas las avesvistas u oídas (sin límite de distancia), en 20 puntos esta-
blecidos a lo largo de un recorrido diseñado por cada cola-borador, dentro del perímetro correspondiente a una cuadrí-cula UTM de 10x10 km. Los puntos se distribuyen, en lamedida de lo posible, entre todos los hábitats presentes enese territorio de manera proporcional a la superficie de cadauno. Los puntos de muestreo son visitados dos veces cadaprimavera, entre mediados de abril y mediados de junio. Losmuestreos realizados durante la primera visita (entre el 15 deabril y el 15 de mayo) sirven para obtener la tendencia de las
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
26
Figura 1. Cuadrículas UTM 10x10 km cubiertas algún año por el programa SACRE para toda España entre 1996 y 2005.

especies sedentarias y presaharianas, mientras que con losmuestreos de la segunda visita (entre el 15 de mayo y el 15de junio) se obtienen las tendencias de las especies transaha-rianas, en general reproductores más tardíos. En cada puntose anotaron las características básicas de la vegetación, con elfin de comparar las tendencias de las aves en distintos hábi-tats.
La Comunidad de Madrid se dividió en dos áreas biogeográ-ficas (zona de la sierra y zona de la fosa del Tajo) para ana-lizar las tendencias de las aves en sub-regiones ecológicamen-te distintas (figura 2). Esta división se ha realizado siguien-do las consideraciones generales sobre orografía, clima ybotánica de la Comunidad (Izco 1984). En la figura 2 semuestra qué cuadrículas han sido muestreadas alguna vezentre 1996 y 2005 por los participantes del programaSACRE, especificando aquellas realizadas al menos durantecuatro años y que han sido consideradas en el análisis.
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
27
Figura 2. Cuadrículas con parte o todo su territorio en la Comunidad de Madrid.

Las evoluciones poblacionales de las especies han sido calcu-ladas mediante el programa estadístico TRIM (TRends forIndices and Monitoring data; Pannekoek y Van Strien 2003), queanaliza series temporales de datos mediante regresión log-lineal. Este programa ha sido específicamente diseñado parasolventar varios de los problemas más frecuentes en este tipode estudios, como son la existencia de vacíos en la matriz dedatos a analizar (por ausencia de muestreo durante algún añoen localidades concretas), o la autocorrelación temporal enel número de aves observadas en cada localidad. Para sinteti-zar de manera sencilla la tendencia de las especies durante losdiez años de estudio se optó por extraer modelos lineales, demanera que la pendiente de la recta obtenida y el error están-dar de su estima se pueden interpretar en términos biológi-cos como el porcentaje de variación anual (o tasa instantáneade crecimiento: positivo en el caso de un aumento de efecti-vos, negativo en caso de declive), y su intervalo de confianzaal 95% (±1,96 veces el error estándar). Además del análisisconsiderando conjuntamente todas las cuadrículas muestrea-das en Madrid, se realizó otro incluyendo como covariable lapertenencia de cada zona al área de influencia de la sierra ode la fosa del Tajo.
Se han considerado sólo aquellas especies observadas en más deun tercio de las visitas a las cuadrículas con presencia constata-da de la especie algún año, pues así quedan excluidas las espe-cies más raras o de presencia esporádica cuyas tendenciaspoblacionales no son adecuadamente evaluadas con métodosextensivos, como el del programa SACRE. Para analizar laspoblaciones de aves ligadas a determinados hábitats, éstas sehan clasificado en cuatro grandes categorías ambientales (tabla1), mediante la consulta de diversos trabajos con informacióncuantitativa sobre selección y preferencias de hábitat de las avesreproductoras de España (Díaz et al. 1996; Tellería et al. 1996;Martí y Del Moral 2003; Carrascal et al. 2006): a) aves demedios urbanos (7 spp.), incluyendo tanto grandes ciudadescomo pueblos y urbanizaciones; b) aves de medios forestales(11 spp.), como pinares, robledales y encinares, pero excluyen-do formaciones arbóreas poco compactas, como las dehesas defresno o encina y los bosques-galería; c) aves de matorrales (8spp.), considerando como tales masas continuas de piornos,brezos, jaras, carrascas o retamas; y d) aves de medios abiertos
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
28

en sentido amplio (13 spp.), incluyendo eriales, pastizales, cul-tivos de cereal y viñedos. Además, también se seleccionó algrupo de especies más generalistas y capaces de ocupar unagran variedad de hábitats diferentes (10 spp.). El índice mul-tiespecífico que resume la evolución global de cada grupo deaves se obtuvo promediando los índices anuales obtenidos paracada una de las especies reunidas en cada categoría. La contri-bución a este índice de cada una de las especies particulares fueponderada según su frecuencia de aparición en los muestreos,para evitar que la tendencia de las especies más raras influyesedesproporcionadamente en la evolución conjunta.
R E S U L T A D O S
Cobertura
De las 115 cuadrículas que presentan todo o parte de suterritorio en la Comunidad de Madrid, 88 (76%) se hancensado alguna vez desde 1996. De éstas, 52 (45%) se hantenido en cuenta para los análisis por haber sido realizadasal menos durante 4 años (figura 2). Las 36 cuadrículas res-tantes (31%) que ya disponen de datos pero no durante sufi-
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
29

cientes temporadas, podrán ser tenidas en cuenta en pocotiempo si continúan prospectándose. Sólo hay 27 cuadrícu-las que no han sido realizadas ningún año, muchas de ellasdebido a que presentan dificultades de acceso a la mayorparte de su territorio.
Tendencias generales
Se han obtenido los parámetros temporales para 79 especies,observándose un claro predominio de especies con evolucio-nes positivas frente a las especies de tendencias negativas. Latabla 1 resume estos resultados. Para ilustrar cómo se debeninterpretar estos datos, describiremos a modo de ejemplo losobtenidos para el carricero tordal (Acrocephalus arundinaceus).Así, esta especie muestra una tendencia poblacional a lolargo de los diez años de estudio significativamente crecien-te; o lo que es lo mismo, no cumple la hipótesis nula de unaevolución temporal estable, pues de acuerdo a los datos obte-nidos cabría menos de un 1% de posibilidades (p < 0,01)de obtener por puro azar la pendiente media observada. Enpromedio esta especie ha estado aumentando tanto como un16,1% cada año, si bien existe un margen de error en estaestima media del ±11,6 (es decir, el crecimiento real de lapoblación, con una elevadísima seguridad —al 95%—, estáincluido en el intervalo entre 4,5% y 27,7%). Nótese queeste margen de error varía mucho entre especies, estandoinversamente relacionado con el tamaño muestral disponibley con la regularidad en abundancia intrínseca de cada espe-cie, por lo que aquéllas con más datos permiten realizar esti-mas más precisas (en el caso del carricero tordal, este errores particularmente amplio, principalmente porque sólo estu-vo presente en el 49,5% de las cuadrículas muestreadas a lolargo de los diez años).
Para el conjunto de la Comunidad, 26 especies muestran unatendencia positiva estadísticamente significativa, con un por-centaje de aumento mayor al 10% en los siguientes casos:tórtola turca (Streptopelia decaocto), curruca mirlona (Sylvia hor-tensis), carricero tordal, paloma bravía (Columba livia), trepa-dor azul (Sitta europaea), arrendajo (Garrulus glandarius), jilgue-ro (Carduelis carduelis), chochín común (Troglodytes troglodytes),petirrojo europeo (Erithacus rubecula) y mosquitero papialbo
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
30

(Phylloscopus bonelli). En cambio, únicamente tres especiesmuestran una reducción significativa del número de avesobservadas a escala regional en este periodo: chova piquirro-ja (Pyrrhocorax pyrrhocorax), buitrón (Cisticola juncidis) y golon-drina dáurica (Hirundo daurica).
Las 50 especies que no presentan ni aumentos ni descensosglobalmente significativos a lo largo de los diez años de estu-dio (esto es, las que muestran un porcentaje de variaciónanual más cercano a cero), se pueden dividir en dos catego-rías. Por una parte, hay un grupo de especies con nulo cam-bio interanual y muy pequeño error estándar de este paráme-tro (reflejado en su estrecho intervalo de confianza). Esto es,se trata de especies caracterizadas por la alta estabilidad desus efectivos durante el periodo analizado (% Ev. Anual enel intervalo de ±1,0 y un I.C. < 4, subjetivamente estableci-do como umbral para distinguir a las especies con mayorestabilidad poblacional): abubilla (Upupa epops), golondrinacomún (Hirundo rustica), perdiz roja (Alectoris rufa), ruiseñorbastardo (Cettia cetti) o triguero (Emberiza calandra). Por el con-trario, hay otras especies que teniendo un cambio interanualen el número de aves también prácticamente nulo, sin embar-go presentan un error estándar elevado (% Ev. Anual en elintervalo de ±1,0 pero un I.C. > 4; tabla 1): carbonerogarrapinos (Parus ater), grajilla (Corvus monedula), lavanderablanca (Motacilla alba), paloma zurita (Columba oenas), o zorzalcharlo (Turdus viscivorus). Estudios demográficos más detalla-dos ayudarían a discernir en qué medida este patrón tan irre-gular se corresponde con su dinámica poblacional real.
Tendencias por zonas
Al incluir el efecto zona como covariable (sierra vs. fosa delTajo; tabla 1), 14 especies presentan evoluciones significati-vamente distintas en cada área. Es especialmente reseñableque 9 de ellas no muestren, en cambio, ninguna tendenciasignificativa a escala de toda la comunidad autónoma. Losdeclives más acusados en la zona de la fosa del Tajo lo pre-sentan la corneja negra (Corvus corone), la lavandera blanca, elcolirrojo tizón (Phoenicurus ochruros), y la chova piquirroja; encambio, declives en la zona de la sierra corresponden a lacogujada común (Galerida cristata) y la grajilla. Por el contra-
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
31
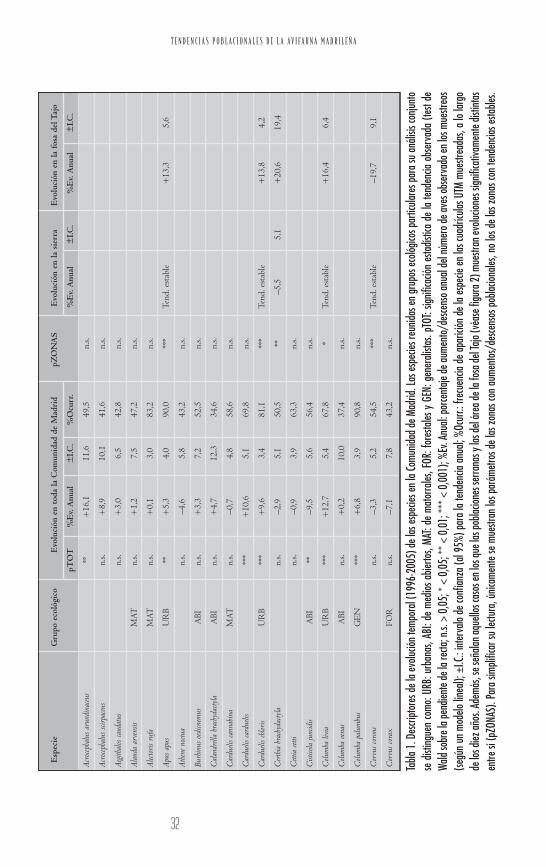
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
32
Especie
Grupo ecológico
Evolución en toda la Comunidad de Madrid
pZONAS
Evolución en la sierra Evolución en la fosa del Tajo
pTOT
%Ev. Anual
±I.C.
%Ocurr.
%Ev. Anual
±I.C.
%Ev. Anual
±I.C.
Acrocephalus arundinaceus
**+16,1
11,6
49,5
n.s.
Acrocephalus scirpaceus
n.s.
+8,9
10,1
41,6
n.s.
Aegithalos caudatus
n.s.
+3,0
6,5
42,8
n.s.
Alauda arvensis
MAT
n.s.
+1,2
7,5
47,2
n.s.
Alectoris rufa
MAT
n.s.
+0,1
3,0
83,2
n.s.
Apus apus
URB
**+5,3
4,0
90,0
***
Tend. estable
+13,3
5,6
Athene noctua
n.s.
–4,6
5,8
43,2
n.s.
Burhinus oedicnemus
ABI
n.s.
+3,3
7,2
52,5
n.s.
Calandrella brachydactyla
ABI
n.s.
+4,7
12,3
34,6
n.s.
Carduelis cannabina
MAT
n.s.
–0,7
4,8
58,6
n.s.
Carduelis carduelis
***
+10,6
5,1
69,8
n.s.
Carduelis chloris
URB
***
+9,6
3,4
81,1
***
Tend. estable
+13,8
4,2
Certhia brachydactyla
n.s.
–2,9
5,1
50,5
**–5,5
5,1
+20,6
19,4
Cettia cetti
n.s.
–0,9
3,9
63,3
n.s.
Cisticola juncidis
ABI
**–9,5
5,6
56,4
n.s.
Columba livia
URB
***
+12,7
5,4
67,8
*Tend. estable
+16,4
6,4
Columba oenas
ABI
n.s.
+0,2
10,0
37,4
n.s.
Columba palumbus
GEN
***
+6,8
3,9
90,8
n.s.
Corvus corone
n.s.
–3,3
5,2
54,5
***
Tend. estable
–19,7
9,1
Corvus corax
FOR
n.s.
–7,1
7,8
43,2
n.s.
Tabla
1. D
escripto
res d
e la e
voluc
ión te
mporal
(199
6-20
05) d
e las es
pecie
s en l
a Com
unida
d de M
adrid
. Las es
pecie
s reunid
as en
grup
os ec
ológic
os pa
rticular
es pa
ra su
análisis co
njunto
se disting
uen como
: URB
: urban
as, A
BI: d
e me
dios a
bierto
s, MA
T: de m
atorra
les, F
OR: forestal
es y GEN
: generalista
s. pTOT: sign
ificació
n estad
ística
de la
tendencia
observada (te
st de
Wald sobre la p
endie
nte d
e la r
ecta;
n.s. > 0,05
; * < 0,05; **
< 0,01; **
* < 0,001
); %E
v. An
ual: p
orcentaje
de au
mento/
desce
nso a
nual del n
úmero d
e aves o
bservado en
los m
uestr
eos
(segú
n un
modelo
lineal); ±I.C
.: int
ervalo
de co
nfian
za (a
l 95%
) para l
a tendencia
anua
l; %O
curr.: frecuencia
de ap
arición
de la
especie
en la
s cua
drícu
las UTM
muestr
eadas, a l
o largo
de los d
iez añ
os. A
demá
s, se se
ñalan
aquellos casos en
los q
ue las p
oblac
iones se
rrana
s y las d
el área de
la fo
sa de
l Tajo
(véase figu
ra 2)
muestr
an ev
olucio
nes sign
ificativam
ente
distin
tasentre
sí (p
ZONA
S). P
ara s
implificar su
lectu
ra, ú
nicam
ente
se m
uestr
an lo
s parám
etros de
las z
onas co
n au
mentos/d
escensos po
blacio
nales
, no l
os de
las z
onas co
n ten
dencias
estab
les.

A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
33
Tabla
1 (Continuació
n). D
escripto
res de
la ev
olució
n tem
poral (19
96-200
5) de
las e
species
en la Co
munid
ad de
Madrid
. Las es
pecie
s reunid
as en
grupos ec
ológic
os pa
rticular
es pa
ra su
análisis
conju
nto se
distinguen c
omo: UR
B: urbanas, AB
I: de m
edios
abier
tos, M
AT: d
e mato
rrales
, FOR
: forestales y
GEN: ge
neralist
as. p
TOT: sig
nifica
ción e
stadís
tica d
e la t
endencia observada (
test d
eWa
ld sobre la pendien
te de la
recta
; n.s. > 0,05
; * < 0,05
; ** <
0,01
; *** < 0,00
1); %
Ev. A
nual: porcenta
je de aum
ento/
desce
nso a
nual
del n
úmero
de a
ves o
bservado en
los m
uestreos
(según u
n modelo
lineal); ±I.C.: i
ntervalo
de co
nfian
za (a
l 95%
) para la t
endencia anual; %
Ocurr.:
frecuencia
de ap
arició
n de la e
specie en las cuadrícu
las UTM
muestreadas, a lo largo de
los
diez a
ños. Ad
emás, se s
eñala
n aquellos ca
sos e
n los qu
e las po
blacio
nes serr
anas y
las de
l área
de la fo
sa de
l Tajo
(véase figura 2) m
uestran ev
olucio
nes sign
ificativame
nte distinta
s entr
e sí
(pZO
NAS).
Para simp
lificar su lectur
a, única
mente
se m
uestran los p
arám
etros de
las z
onas co
n aum
entos
/descensos po
blacio
nales
, no los de
las z
onas co
n tendencias
estab
les.
Especie
Grupo ecológico
Evolución en toda la Comunidad de Madrid
pZONAS
Evolución en la sierra Evolución en la fosa del Tajo
pTOT
%Ev. Anual
±I.C.
%Ocurr.
%Ev. Anual
±I.C.
%Ev. Anual
±I.C.
Corvus monedula
n.s.
–0,1
4,7
73,6
**–8,3
7,2
Tend. estable
Coturnix coturnix
ABI
n.s.
–4,2
6,0
44,4
n.s.
Cuculus canorus
***
+9,5
3,7
70,2
n.s.
Cyanopica cooki
***
+9,8
5,4
82,7
n.s.
Delichon urbicum
URB
**+8,7
6,2
60,7
n.s.
Dendrocopos major
FOR
n.s.
+3,8
7,6
36,6
n.s.
Emberiza calandra
ABI
n.s.
–0,1
2,8
87,6
n.s.
Emberiza cia
MAT
n.s.
+1,5
6,7
51,2
n.s.
Erithacus rubecula
FOR
***
+10,3
5,2
59,4
n.s.
Falco tinnunculus
n.s.
+4,0
5,4
50,0
n.s.
Fringilla coelebs
FOR
***
+7,2
3,2
77,8
*+5,6
3,4
+16,1
8,9
Galerida cristata
ABI
n.s.
–1,2
2,8
81,2
**–9,5
6,0
Tend. estable
Galerida theklae
n.s.
+4,5
6,1
58,9
n.s.
Garrulus glandarius
FOR
**+11,0
8,4
56,4
n.s.
Hippolais polyglotta
GEN
n.s.
–5,7
6,7
49,1
n.s.
Hirundo daurica
*–13,1
10,2
39,6
n.s.
Hirundo rustica
URB
n.s.
–0,8
3,6
97,2
n.s.
Lanius meridionalis
ABI
n.s.
–3,0
6,4
44,4
n.s.
Lanius senator
n.s.
+1,1
4,6
56,5
n.s.
Lullula arborea
*+4,5
4,1
68,9
n.s.
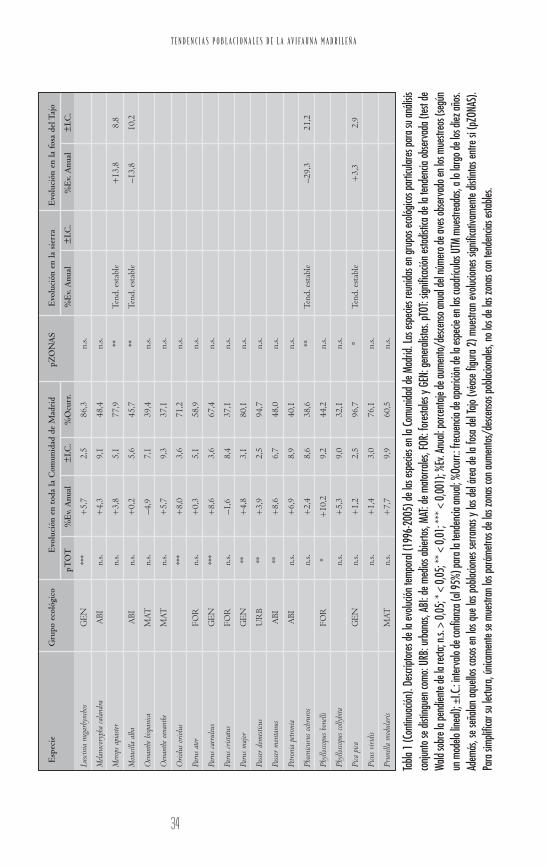
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
34
Especie
Grupo ecológico
Evolución en toda la Comunidad de Madrid
pZONAS
Evolución en la sierra Evolución en la fosa del Tajo
pTOT
%Ev. Anual
±I.C.
%Ocurr.
%Ev. Anual
±I.C.
%Ev. Anual
±I.C.
Luscinia megarhynchos
GEN
***
+5,7
2,5
86,3
n.s.
Melanocorypha calandra
ABI
n.s.
+4,3
9,1
48,4
n.s.
Merops apiaster
n.s.
+3,8
5,1
77,9
**Tend. estable
+13,8
8,8
Motacilla alba
ABI
n.s.
+0,2
5,6
45,7
**Tend. estable
–13,8
10,2
Oenanthe hispanica
MAT
n.s.
–4,9
7,1
39,4
n.s.
Oenanthe oenanthe
MAT
n.s.
+5,7
9,3
37,1
n.s.
Oriolus oriolus
***
+8,0
3,6
71,2
n.s.
Parus ater
FOR
n.s.
+0,3
5,1
58,9
n.s.
Parus caeruleus
GEN
***
+8,6
3,6
67,4
n.s.
Parus cristatus
FOR
n.s.
–1,6
8,4
37,1
n.s.
Parus major
GEN
**+4,8
3,1
80,1
n.s.
Passer domesticus
URB
**+3,9
2,5
94,7
n.s.
Passer montanus
ABI
**+8,6
6,7
48,0
n.s.
Petronia petronia
ABI
n.s.
+6,9
8,9
40,1
n.s.
Phoenicurus ochruros
n.s.
+2,4
8,6
38,6
**Tend. estable
–29,3
21,2
Phylloscopus bonelli
FOR
*+10,2
9,2
44,2
n.s.
Phylloscopus collybita
n.s.
+5,3
9,0
32,1
n.s.
Pica pica
GEN
n.s.
+1,2
2,5
96,7
*Tend. estable
+3,3
2,9
Picus viridis
n.s.
+1,4
3,0
76,1
n.s.
Prunella modularis
MAT
n.s.
+7,7
9,9
60,5
n.s.
Tabla
1 (C
ontinuació
n). D
escripto
res de
la ev
olució
n tem
poral (19
96-200
5) de
las e
species
en la Co
munid
ad de
Madrid
. Las es
pecie
s reunid
as en
grupos ec
ológic
os pa
rticular
es pa
ra su
análisis
conju
nto se
distinguen c
omo:
URB:
urbanas, AB
I: de m
edios
abier
tos, M
AT: d
e mato
rrales
, FOR
: forestales y
GEN: ge
neralist
as. p
TOT: sig
nifica
ción e
stadís
tica d
e la t
endencia observada (
test d
eWa
ld sobre la p
endie
nte de
la re
cta; n.s. > 0,
05; * < 0,
05; ** <
0,01
; *** < 0,
001); %
Ev. A
nual: po
rcenta
je de au
mento
/descenso a
nual del núm
ero de
aves ob
servado e
n los m
uestreos (según
un m
odelo
lineal); ±I.C.: inte
rvalo
de co
nfian
za (a
l 95%
) para la t
endencia anual; %
Ocurr.: fre
cuencia
de ap
arició
n de la e
specie en las cuadrícu
las UTM
muestreadas, a lo largo de
los d
iez añ
os.
Adem
ás, se s
eñala
n aquellos c
asos en
los q
ue la
s poblac
iones se
rranas y
las d
el área de
la fo
sa de
l Tajo
(véase figura 2) m
uestran ev
olucio
nes sign
ificativame
nte di
stinta
s entr
e sí (pZON
AS).
Para simp
lificar su lectur
a, única
mente
se m
uestran los p
arám
etros de
las z
onas co
n aum
entos
/descensos po
blacio
nales
, no los de
las z
onas co
n tendencias
estab
les.

Especie
Grupo ecológico
Evolución en toda la Comunidad de Madrid
pZONAS
Evolución en la sierra Evolución en la fosa del Tajo
pTOT
%Ev. Anual
±I.C.
%Ocurr.
%Ev. Anual
±I.C.
%Ev. Anual
±I.C.
Pyrrhocorax pyrrhocorax
**–12,2
8,4
38,2
**Tend. estable
–19,9
9,2
Regulus ignicapilla
FOR
n.s.
+7,0
10,1
37,5
n.s.
Remiz pendulinus
n.s.
+8,9
14,1
50,0
n.s.
Saxicola torquata
GEN
n.s.
+3,8
5,2
53,5
n.s.
Serinus serinus
GEN
n.s.
+2,0
2,9
93,8
n.s.
Sitta europaea
FOR
**+11,6
8,9
52,2
n.s.
Streptopelia decaocto
URB
***
+35,5
11,3
42,5
n.s.
Streptopelia turtur
n.s.
+1,9
4,4
56,1
n.s.
Sturnus unicolor
***
+5,0
2,6
97,6
n.s.
Sylvia atricapilla
n.s.
+4,7
5,4
48,3
*+7,9
6,0
Tend. estable
Sylvia cantillans
n.s.
+2,5
7,3
44,0
n.s.
Sylvia hortensis
**+18,6
14,2
48,7
n.s.
Sylvia melanocephala
n.s.
–2,4
4,9
56,2
n.s.
Sylvia undata
MAT
n.s.
–3,0
6,4
38,9
n.s.
Tetrax tetrax
ABI
n.s.
–4,0
5,5
65,4
n.s.
Troglodytes troglodytes
GEN
***
+10,4
6,3
55,4
n.s.
Turdus merula
GEN
***
+5,5
2,9
85,4
n.s.
Turdus viscivorus
FOR
n.s.
–0,4
6,5
52,3
n.s.
Upupa epops
n.s.
+0,0
2,7
86,0
n.s.
Tabla
1 (C
ontinuació
n). D
escripto
res de
la ev
olució
n tem
poral (19
96-200
5) de
las e
species
en la Co
munid
ad de
Madrid
. Las es
pecie
s reunid
as en
grupos ec
ológic
os pa
rticular
es pa
ra su
análisis
conju
nto se
distinguen c
omo:
URB:
urbanas, AB
I: de m
edios
abier
tos, M
AT: d
e mato
rrales
, FOR
: forestales y
GEN: ge
neralist
as. p
TOT: sig
nifica
ción e
stadís
tica d
e la t
endencia observada (
test d
eWa
ld sobre la p
endie
nte de
la re
cta; n.s. > 0,
05; * < 0,
05; ** <
0,01
; *** < 0,
001); %
Ev. A
nual: po
rcenta
je de au
mento
/descenso a
nual del núm
ero de
aves ob
servado e
n los m
uestreos (según
un m
odelo
lineal); ±I.C.: inte
rvalo
de co
nfian
za (a
l 95%
) para la t
endencia anual; %
Ocurr.: fre
cuencia
de ap
arició
n de la e
specie en las cuadrícu
las UTM
muestreadas, a lo largo de
los d
iez añ
os.
Adem
ás, se s
eñala
n aquellos c
asos en
los q
ue la
s poblac
iones se
rranas y
las d
el área de
la fo
sa de
l Tajo
(véase figura 2) m
uestran ev
olucio
nes sign
ificativame
nte di
stinta
s entr
e sí (pZON
AS).
Para simp
lificar su lectur
a, única
mente
se m
uestran los p
arám
etros de
las z
onas co
n aum
entos
/descensos po
blacio
nales
, no los de
las z
onas co
n tendencias
estab
les.
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
35
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
36
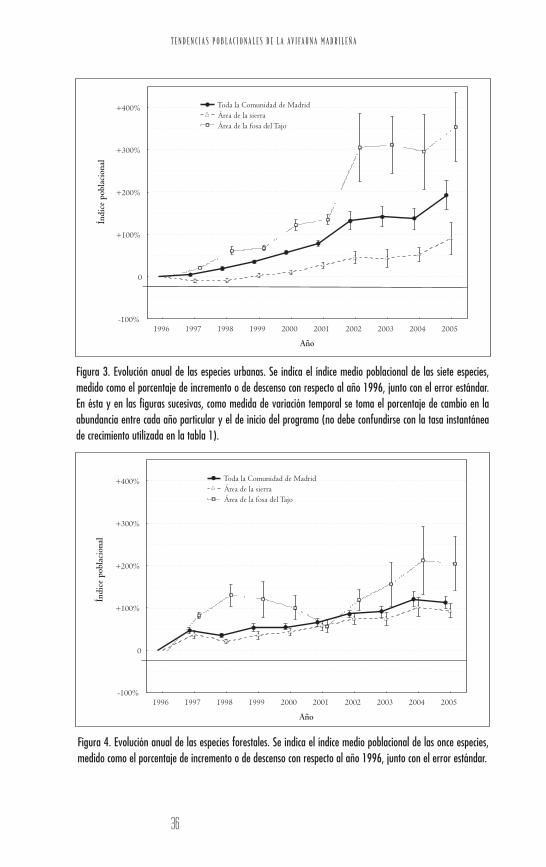
Figura 3. Evolución anual de las especies urbanas. Se indica el índice medio poblacional de las siete especies,medido como el porcentaje de incremento o de descenso con respecto al año 1996, junto con el error estándar.En ésta y en las figuras sucesivas, como medida de variación temporal se toma el porcentaje de cambio en laabundancia entre cada año particular y el de inicio del programa (no debe confundirse con la tasa instantáneade crecimiento utilizada en la tabla 1).
Figura 4. Evolución anual de las especies forestales. Se indica el índice medio poblacional de las once especies,medido como el porcentaje de incremento o de descenso con respecto al año 1996, junto con el error estándar.

Figura 5. Evolución anual de las especies más generalistas. Se indica el índice medio poblacional de las diezespecies, medido como el porcentaje de incremento o de descenso con respecto al año 1996, junto con el errorestándar.
Figura 6. Evolución anual de las especies de matorrales. Se indica el índice medio poblacional de las ocho especies,medido como el porcentaje de incremento o de descenso con respecto al año 1996, junto con el error estándar.
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
37
rio, los incrementos más significativos en la zona de la fosalo presentan el vencejo común (Apus apus), el verderón común(Carduelis chloris), la paloma bravía, el abejaruco europeo(Merops apiaster) y la urraca (Pica pica), mientras que en la zonade la sierra sólo aumenta claramente la curruca capirotada(Sylvia atricapilla). El agateador común (Certhia brachydactyla) yel pinzón vulgar (Fringilla coelebs) constituyen dos casos parti-culares: el agateador aumenta muy significativamente en lazona de la fosa pero curiosamente tiende a disminuir en lasierra, que a priori le sería más favorable; el pinzón aumentasignificativamente en ambas zonas, si bien lo hace muchomás marcadamente en la zona de la fosa del Tajo.
Tendencias por hábitats
Se han muestreado todos los grandes hábitats presentes en laComunidad de Madrid. El mayor incremento durante estosdiez años (casi el triple de efectivos detectados en 2005 queen 1996), se registró en las aves ligadas a medios urbanos(figura 3), aunque tanto las aves forestales como las avesgeneralistas (figuras 4 y 5, respectivamente) mostraron tam-
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
38
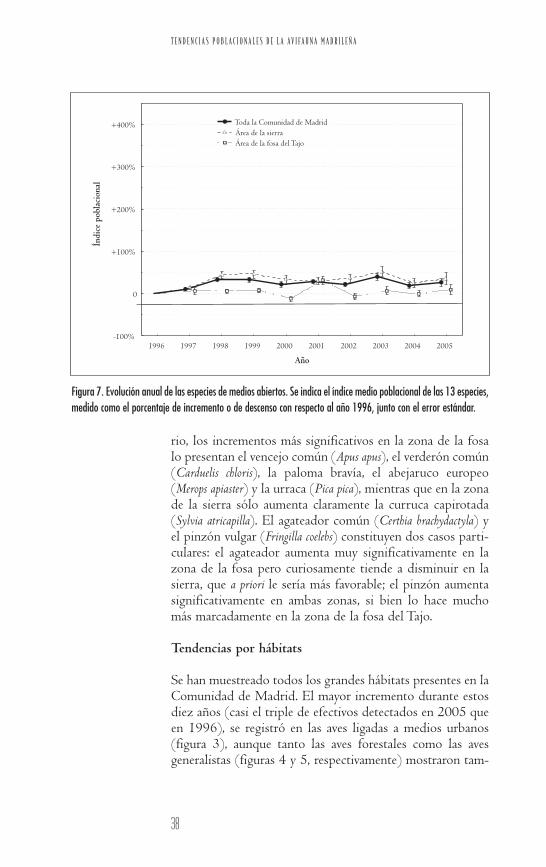
Figura 7. Evolución anual de las especies de medios abiertos. Se indica el índice medio poblacional de las 13 especies,medido como el porcentaje de incremento o de descenso con respecto al año 1996, junto con el error estándar.

bién una clara y progresiva tendencia positiva. En estos trescasos la evolución es más acusada en la fosa del Tajo que enla sierra. Las aves propias de los medios arbustivos y abier-tos (figuras 6 y 7, respectivamente) se caracterizan por unagran estabilidad durante la mayor parte del periodo de estu-dio, no siendo tan acusadas las diferencias entre las dos gran-des sub-regiones distinguidas.
D I S C U S I Ó N
Los resultados de este trabajo a escala regional reflejan enparte los obtenidos para el conjunto de España(www.seo.org/?seguimientodeaves), y apoyan la idea de
que las poblaciones de aves en la Comunidad de Madrid,globalmente, están experimentando una evolución favorable.Como conclusiones particulares se pueden destacar que:
` Una buena cobertura del programa SACRE a escalaprovincial permite conocer evoluciones poblacionales localesque resultarían poco visibles si únicamente se atendiese a losanálisis de tendencia a escala nacional. Por ejemplo, la apa-rente vulnerabilidad actual en Madrid de la golondrina dáu-rica o de la chova piquirroja no se refleja en su aumentogeneral a escala nacional (www.seo.org/?seguimientodeaves).La identificación de este tipo de patrones es fundamentalpara gestionar adecuadamente los recursos ofrecidos por lasadministraciones medioambientales regionales.
` De manera análoga, a escala regional es importante dis-tinguir las tendencias seguidas por una especie en zonas eco-lógicamente distintas dentro de Madrid. Así, la mayoría delas especies que presentan evoluciones diferentes en la sierray en el área de la fosa del Tajo, no muestran, en cambio, nin-guna tendencia significativa a escala de toda la comunidadautónoma. Obviamente, esta situación puede resultar espe-cialmente conflictiva en el caso de las seis especies en progre-siva disminución local (agateador común, cogujada común,colirrojo tizón, corneja negra, grajilla y lavandera blanca),pues para ellas no estaría justificado aplicar medidas preven-tivas en base a su estabilidad regional. Por tanto, aún cuandolos planes regionales de conservación deban aspirar a ofrecer
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
39

medidas de gestión altamente generalizables a escalas espacia-les amplias, no se debe olvidar que la mejoría efectiva de algu-nas especies debe pasar por valorar sus pautas poblacionaleslocales. Esta circunstancia plantea, además, interesantes cues-tiones en el ámbito del estudio teórico de la biogeografía decada especie, como por ejemplo examinar si estas especies quemuestran evoluciones diferentes en cada zona comparten ras-gos ecológicos que las predisponen a ello (p. ej. preferenciasde hábitat o altitudinales), o si los postulados actuales sobrecambio climático tienen alguna relación (las dos zonas consi-deradas son muy distintas en términos bioclimáticos).
` Los aumentos poblacionales más importantes se estándando en el área de la fosa del Tajo. El aumento de las pobla-ciones de aves ligadas a medios urbanos puede explicarse confacilidad por la continua expansión de pueblos y ciudades deesta zona. No obstante, las causas precisas del rápido creci-miento de las aves forestales y generalistas requiere estudiosmás detallados.
` Las especies propias de hábitats abiertos y de hábitatsarbustivos merecen un seguimiento especial de aquí en ade-lante, pues no presentan el marcado aumento poblacionalque sí se observa en el resto de grupos ecológicos (véase tam-bién De Juana 2004). Este hecho puede relacionarse con lasusceptibilidad de este tipo de ambientes a sufrir el impara-ble desarrollo urbanístico de la Comunidad de Madrid, puescultivos, eriales, carrascales o espartales están escasamentecubiertos por la actual red de espacios protegidos y pierdensuperficie año tras año. Además, los medios agrícolas tien-den a un cambio de uso cada vez más intensivo (periodosmás cortos de cosecha, mayor uso de herbicidas e insectici-das, pérdidas de lindes, mecanización, etc.), que puede estarperjudicando muy seriamente a las poblaciones de aves(Suárez 2004).
En definitiva, con este trabajo queremos hacer ver que dedi-cando únicamente dos mañanas de campo cada primavera yaplicando un sencillo protocolo de muestreo, todos los afi-cionados a la ornitología pueden contribuir a generar unavaliosísima información directamente aplicable a la conser-vación y conocimiento de la avifauna en su región. De la
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
40

misma forma que ahora se dispone de esta información enMadrid, se podría llegar a la misma situación en el resto delas comunidades autónomas. Por tanto, queremos hacer unllamamiento a grupos ornitológicos locales, especialmente alos de aquellas provincias con más baja cobertura en el pro-grama SACRE (figura 1), para que se animen a participar eneste estudio. Este trabajo redundará en un gran aumento delconocimiento del estado de conservación de las aves a escalaregional y estatal, principal objetivo de cualquier grupo localde SEO/BirdLife.
A G R A D E C I M I E N T O S
Q ueremos agradecer la participación voluntaria de todaslas personas que realizan el trabajo de campo, y muyespecialmente la de aquellas que colaboran desde el ini-
cio de este programa. Agradecemos a Pedro Silos la creaciónde la base de datos en la que se recopilan todos los muestre-os, y las modificaciones necesarias que ha tenido que realizarpara extraer la información adecuada para su análisis estadís-tico. Agradecemos al Ministerio de Medio Ambiente y a laComunidad de Madrid su apoyo económico. Javier Seoaneenriqueció notablemente con sus comentarios una versiónpreliminar del texto.
E Q U I P O D E T R A B A J O
Los colaboradores voluntarios, con cuyo trabajo de campo hasido posible la realización de este trabajo, son, por ordenalfabético: Agustín Marín Acedos, Aída Caño Mayor,
Alejandro Martínez Martino, Alfonso Barrada Ferreiros,Alfonso Pol Gray, Alfredo Esteban Corral, Alison Barrett,Álvaro Ramírez García, Ana Grandal Martín, Ángel PérezMenchero, Antonio González Castillo, Antonio Jesús PauleSánchez, Antonio Padilla Gutiérrez, Antonio Ruiz Heredia,Blas Molina Villarino, Carlos Barcala España, Carlos CelayaCarrillo, Carlos Figaredo Alvargonzález, Carlos J. CarpinteroRubio, Carlos Sánchez Delicado, Carlos Talabante Ramírez,Daniel Díaz Díaz, Daniel Sánchez Garrosa, Eduardo GonzálezHernández, Eduardo Hervás Domínguez, Elena Abati, Elena
A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
41

Baonza Díaz, Enrique Ayllón López, Eva Pérez Toledano, FélixAguado Pérez, Fernando Hortelano Coronado, FranciscoCalvet Zanon, Francisco Castellanos Toledo, Francisco JavierGómez Sanz, Francisco Javier López del Pozo, Francisco JavierPenalba Sanz, Francisco José Conde Jiménez, Francisco LópezIsac, Ignacio Fernández Aransay, Ignacio Torres Ruiz-Huerta,Iñaqui Rodríguez Prieto, Isabel González Caballero, Iván de laHera Fernández, Jaime Fernández de Gamboa, Javier CanoSánchez, Javier Fernández García-Almendral, Javier Martín delRío, Jesús Alonso Sotillo, Jesús Gamarra Ibáñez, Jesús RamírezVillar, Jesús Ruiz Rodrigo, Jorge Bartolomé Zofio, Jorge RubioCasado, José Aguilera Díez, José Antonio López Constante,José Antonio Rodríguez Crespo, José Enrique JiménezSánchez, José Gómez Moreno, José Luis Aparicio Calles, JoséLuis Armendáriz Sanz, José Luis Blas Priego, José LuisHerranz Alonso, José Luis Manzanaro Asenjo, José LuisPizarro, José Luis Talavera Rivera, José Manuel Altares Crespo,José Manuel Delgado Pérez, José Manuel Palacios Alberti, JoséMaría Martín Nández, José Mercader Herrero, José MiguelAbarca Antón, José Miguel López Fuentes, José PeñaHernández, Juan A. Malo de Molina Martínez, Juan AlbertoDíaz Reyes, Juan Bedmar Perlado, Juan Carlos Atienza, JuanCarlos del Moral González, Juan Carlos del Villar Berdión,Juan Carlos Ulecia Zaldívar, Juan José Castellanos, Juan LinazaSebastián, Justo Cilleros Prados, Laura Martínez Pascual, LeónGarcía Comendador Alonso, Leonor Zarza Caballero, LuisEnrique Díez Blanco, Luis Fernando Hernando Benavides,Maite Samblas de Miguel, Manuel Medina del Valle,Marcelino Martín Pajares, Mariano Velázquez Martín, MarinaOjeda Jiménez, Mario González Pérez, Marta ArenasRomasanta, Martín Francisco Arévalo Sánchez, MercedesBagazgoitia Barrera, Michael Schneider, Miguel Ángel CalvoGuerrero, Miguel Ángel Cosme Muñoz, Miguel ÁngelGonzález Arroyo, Miguel Ángel Letón Fernández, MiguelHigueras Ortega, Miguel Juan Martínez, Miguel SevillaCallejo, Montserrat Soria, Óscar Moreno Aparicio, PabloRodríguez Palenzuela, Pascual Alcázar Fernández, PascualCampos Rus, Ramón Gómez de la Cuesta Laporta, RamónRevuelta Gutiérrez, Ricardo Gómez Calmaestra, RicardoMéndez Maroto, Rosario Cruz Violero, Ruth Cristina CandilMartín, Tomás Díaz Pulido, Tomás Perdiguero Pérez, VirginiaEscandell González.
T E N D E N C I A S P O B L A C I O N A L E S D E L A A V I F A U N A M A D R I L E Ñ A
42

A N U A R I O O R N I T O L Ó G I C O D E M A D R I D 2 0 0 5
43
B I B L I O G R A F Í A
` Alonso, J. C.; Palacín, C. y Martín, C. A. 2005. La Avutarda Común en la
península Ibérica: población actual y método de censo. SEO/BirdLife. Madrid.
` Antor, R. 2001. Population status and conservation of the Bearded
Vulture (Gypaetus barbatus) in the Pyrenees. En: Abstracts 4th Eurasian Congress on
Raptors: 8. EBD-CSIC y Raptor Research Foundation. Sevilla.
` Carrascal, L. M.; Seoane, J.; Alonso, C. L. y Palomino, D. 2003. Estatus
regional y preferencias ambientales de la avifauna madrileña durante el invier-
no. Anuario Ornitológico de Madrid 2002: 22-43.
` Carrascal, L. M.; Palomino, D. y Seoane, J. 2006. Fundamentos ecológicos
y biogeográficos de la rareza de la avifauna madrileña. Una propuesta de modi-
ficación del catálogo regional de especies amenazadas. Graellsia, 62: 485-509.
` De Juana, E. 2004. Cambios en el estado de conservación de las aves en
España, años 1954 a 2004. Ardeola, 51: 19-50.
` Díaz, M.; Asensio, B. y Tellería, J. L. 1996. Aves ibéricas. I. No paseriformes.
J. M. Reyero Editor. Madrid.
` González, L. M. y Oria, J. 2001. La frágil recuperación del Águila
Imperial Ibérica. Quercus, 190: 20-28.
` Izco, J. 1984. Madrid Verde. Instituto de Estudios Agrarios, Pesqueros y
Alimentarios. Madrid.
` Madroño, A.; González, C. y Atienza, J. C. (eds.) 2004. Libro Rojo de las Aves
de España. Dirección General para la Biodiversidad y SEO/BirdLife. Madrid.
` Martí, R. y Del Moral, J. C. 2003. Atlas de las aves reproductoras de España.
Dirección General de Conservación de la Naturaleza y SEO/BirdLife. Madrid.
` Molina, B. y Del Moral, J. C. 2005. La Cigüeña Blanca en España. VI censo
internacional (2004). SEO/BirdLife. Madrid.
Pannekoek, J. y Van Strien, A. 2003. TRIM (TRends & Indices for Monitoring data).
Statistics Netherlands. (http://www.cbs.nl/en-GB/menu/themas/milieu-
natuur-ruimte/natuur/methoden/trim/default.htm).
` Suárez, F. 2004. Aves y agricultura en España peninsular: una revisión
sobre el estado actual de conocimiento y una previsión sobre su futuro. En:
Tellería, J. L. (ed.). La Ornitología Hoy. Homenaje al Profesor Francisco Bernis
Madrazo: 223-265. Editorial Complutense. Madrid.
` Tellería, J. L.; Asensio, B. y Díaz, M. 1999. Aves ibéricas. II. Paseriformes. J.
M. Reyero Editor. Madrid.
` UICN 2001. Categorías y criterios de la Lista Roja de la UICN: Versión
3.1. Comisión de Supervivencia de Especies de la UICN.
(http://www.iucn.org/themes/ssc/redlists/RLcats2001booklet.html).
(Recibido 26.5.2006; Aceptado 26.10.2006)


















