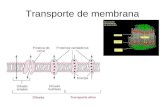
Transporte a Traves de La Membrana
17
SEMINARIO 1. Breve estudio de la estructura de la membrana celular 1.1. Bicapa fosfolipidica Los fosfogliceridos, los esfingolipidos y los esteroles son los principales lípidos de la membrana. Estos componentes ocupan el 50% de la masa de la mayoría de las membranas de las células animales y casi todo el resto es proteínas. Todas las moléculas lipídicas de las membranas son antipáticas. Los lípidos de membrana mas abundantes son los fosfolípidos. Estos tienen una cabeza polar y dos colas hidrocarbonadas hidrofobias. En las células de los animales, plantas y de las bacterias suelen ser ácidos grasos y pueden tener diferente longitud. Una de las colas presenta uno o más dobles enlaces cis (insaturada) mientras que la otra normalmente no tiene dobles enlaces (saturada). Cada enlace cis genera y una curvatura en la cadena. Las diferencias de longitud y grado de saturación entre las colas hidrocarbonadas son importantes porque afectan al modo en que se empaqueta las moléculas de fosfolípidos unas contra otras y determinan la fluidez de la membrana. Los principales fosfolípidos de la mayoría de las membranas celulares son los fosfogliceridos, que tienen como esqueleto al glicerol y tienen tres átomos de carbono. Dos AG de cadena larga están unidos mediante enlaces ester a dos átomos de adyacentes del glicerol, el tercer atomo de C está unido a un grupo fosfato , el cual esta unido a un tipo de cabeza hidrofilica. En las membranas celulares, los principales son la foafatidiletanolamina, la fosfatidiloserina y la
-
Upload
jose-rivera-ulloa -
Category
Documents
-
view
25 -
download
0
description
Pequeña imformacion de transporte a traves de la membrana
Transcript of Transporte a Traves de La Membrana
SEMINARIO1. Breve estudio de la estructura de la membrana celular1.1. Bicapa fosfolipidicaLos fosfogliceridos, los esfingolipidos y los esteroles son los principales lpidos de la membrana. Estos componentes ocupan el 50% de la masa de la mayora de las membranas de las clulas animales y casi todo el resto es protenas. Todas las molculas lipdicas de las membranas son antipticas.Los lpidos de membrana mas abundantes son los fosfolpidos. Estos tienen una cabeza polar y dos colas hidrocarbonadas hidrofobias. En las clulas de los animales, plantas y de las bacterias suelen ser cidos grasos y pueden tener diferente longitud. Una de las colas presenta uno o ms dobles enlaces cis (insaturada) mientras que la otra normalmente no tiene dobles enlaces (saturada). Cada enlace cis genera y una curvatura en la cadena. Las diferencias de longitud y grado de saturacin entre las colas hidrocarbonadas son importantes porque afectan al modo en que se empaqueta las molculas de fosfolpidos unas contra otras y determinan la fluidez de la membrana.Los principales fosfolpidos de la mayora de las membranas celulares son los fosfogliceridos, que tienen como esqueleto al glicerol y tienen tres tomos de carbono. Dos AG de cadena larga estn unidos mediante enlaces ester a dos tomos de adyacentes del glicerol, el tercer atomo de C est unido a un grupo fosfato , el cual esta unido a un tipo de cabeza hidrofilica. En las membranas celulares, los principales son la foafatidiletanolamina, la fosfatidiloserina y la fosfatidilcolina. Otro fosfolpido importante es la esfingomielina que esta formada a partir de esfingosina en lugar de glicerol. La esfingosina es una larga cadena acilo con un grupo amino y dos grupos hidroxilo (OH) en un extremo de la molcula. En la esfingomielina, una cola de AG esta unida al grupo amino y un grupo fosfocolina est unido al grupo hidroxilo terminal, quedando por tanto un grupo hidroxilo libre. Este grupo hidroxilo libre contribuye a las propiedades polares del grupo cabeza adyacente ya que puede formar enlaces de hidrogeno con el grupo cabeza de un lpido vecino, una molcula de H2O o una protena de membrana. Ademas de los fosfolpidos , las bicapas lipdicas de muchas membranas celulares contienen colesterol ( se inserta dentro de la membrana de los fosfolpidos) y glucolpidos ( se encuentran en la capa externa de la membrana con sus residuos hidrocarbonados expuestos a la superficie celular).
Estos fosfolpidos se distribuyen de manera asimtrica entre las dos mitades de la bicapa de la membrana. La capa externa de la membrana plasmtica esta compuesta principalmente poR fosfatidilcolina y esfingomielina, mientras que la fosfatidiletanolamina y la fosfatidilserina son los predominantes en la cpa interna. Las cabezas, tanto la fosfatidilserina como el fosfatidilinositol estn cargados negativamente , de forma que su predominancia en las caras internas resulta en una carga neta negativa en la cara citosolica de la membrana plasmtica. Un quinto fosfolpido, el fosfatidilinositol , se encuentra tambin localizado en la capa interna de la membrana. A temperaturas altas el colesterol interfiere con el movimiento de las cadenas de AG de los fosfolpidos, lo que disminuye la fluidez de la parte externa de la menraban y reduce su permeabilidad a molculas pequeas. A bajas temperaturas, el colesterol interfiere con las interacciones entre las cadenas de los AG protegiendo a las membranas de congelarse y mantiene la fluidez.El colesterol y los esfingolipidos (esfingomielina y glucolipidos) forman las balsas lipdicas, estos grupos de desplazan se desplazan en el interior de la membrana y se asocian con protenas de membranas especificas.
1.2. Proteinas de membranaMientras que los lpidos son los elementos estructuralews fundamentales de la membrana , las protenas son las responsables de realizar funciones especficas en la misma . Protenas integrales: son aquellas que penetran el corazn hidrofobico de la bicapa lipdica. Estas son protenas transmenbrana , tienen dominios que sobresalen por los lados extracelular y citoplasmtico de la membrana. Tienen capacidades como Funcionar como receptores que se unen con sustancias especificas en la superficie de la membrana Conductos ye permite el desplazamientos de solutos a travs de la membrana Agentes que trasfieren electrones durante la fotosntesis y la respiracinLas protenas de la membrana, al igual que los fosfolpidos son anfipaticos. La extraccin de estas protenas de membrana requiere del uso de un detergente como el detergente ionico que desnaturaliza protenas o el detergente no ionico que casi nunca altera la estructura terciaria de una protena. Los detergentes son molculas anfipaticas, sus colas hidrocarbonadas se unen a las regiones hidrofobicas de las protenas integrales de membrana formando un comlejo detergente-proteina que es soluble en solucin acuosa. Las protenas integrales solo pueden ser liberadas por tratamientos que rompan la bicapa fosfolipidica. Tambin suelen estar en hlice de 20 1 25 aa hidrofobicos que se insertan dentro de la membrana del RE durante la sntesis de la cadena polipedtidica. Estas protenas se transportan en vesculas desde el RE al aparato de Golgi y desde all a la membrana plasmtica. Protenas perifricas: son aquellas que se situan completas fuera de la bicapa lipdica, ya sea en el lado citoplasmtico o extracelular, aunque se relaciona con la superficie de la membrana mediante enlaces no covalentes, enlaces electrostticos dbiles. Las protenas perifricas casi siempre pueden solubilizarse mediante la extraccin con soluciones salinas muy concentradas que debilitan los enlaces electrostticos que mantienen unidas a estas protenas con la membrana, o tambin con agentes no polares como soluciones de PH extremo que no rompen la bicapa. Estas protenas brindan soporte estructural a la membrana y actan como ancla para las protenas inttegrales de la membrana. Otras protenas perifricas de la superficie interna de la membrana funcionan como enzimas, cubiertas especializadas o factores que trasmiten seales a travs de la membrana.
Protenas ancladas a lpidos: son aquellas que se localizan fuera de la bicapa lipdica, en la superficie extracelular o en la citoplasmtica, pero que tienen enlaces covalentes con una molcula de lpido que se encuentran dentro de la clula. Estan unidas a la membrana mediante un pequeo oligosacrido complejo vinculado con la molecula de fosfatidilinositol que esta sepultada en la hoja externa de la bicapa. Algunas protenas estn ancladas en la hoja externa de la membrana por el glucosilfosfatidil inositol (GPI)
Cuando una protena se une a la membrana a travs de GPI de anclaje se distingue fcilmente utilizando la enzima denominada fosfolipasada C especifica para fofatidilinositol. Esta enzima separa de forma especfica estas protenas de sus anclajes liberndola de la membrana.Un tipo raro de anemia , la hemoglobinuria paroxstica nocturna se debe a la sntesis deficiente de GPI lo que vuelve a los eritrocitos susceptibles a la destruccin.
2. Algunas caractersticas diferenciales de la membrana plasmtica y las citoplasmticas
3. Factores que influyen en la velocidad de transporte a travs de la membranaLa membrana plasmtica es una barrera selectivamente permeable (semipermeable). Esta permeabilidad asegura que las molculas esenciales como la glucosa, aminocidos y lpidos entren rpidamente en la clula, que los compuestos intermediarios queden retenidos en la clula y que los compuestos de deshecho dejen la clula. La permeabilidad de la membrana depende de varios factores: Tamao del soluto: En trminos generales, bicapas lipidicas son ms permeables a las molculas pequeas que a las grandes. Las molculas pequeas ms relevantes para la funcin celular son el agua, el oxgeno y el dixido de carbono. Las membranass son bastantes permeables a estas molculas, no se requiere de mecanismos de transporte especficos para introducirlos a la celula Temperatura: a mayor temperatura , mayor velocidad Concentracin de la molecula: la velocidad a la que una sustancia difunde hacia dentro es proporcional a la concentracin de las molculas en el exterior y viceversa
Presencia de canales y transportadores: al fijar las molculas al trahnsportan sufren un cambio de comformacin Carga: la carga positiva atrae iones negativos y viceversa , independientemente de la concentracin Presin: significa la suma de todas las fuerzas de las diferentes molculas que chocan contra una superficie en un momento dado. Permeabildad de la membrana: las membranas de las neuronas son 20 veces ms permeables al K que al Na Superficie de difusin: las microvellosidades incrementan el rea de difusin Liposolubilidad Polaridad de la molecula: La medicin de la polaridad (o no polaridad) de un soluto es su cociente de reparto que es la relacin entre su solubilidad en un disolvente organico (como el octanol aceite vegetal) y su solubilidad en agua. Polaridad a iones: La relativa impermeabilidad a las sustancias polares en general y a los iones en particular, es debida a su fuerte asociacin con las molculas de agua, que forman un escudo de hidratacin4. Movimiento de molculas pequeas a travs de la membrana4.1. Transporte pasivo: el pasaje de solutos ocurre sin gastos de energa (ATP) y ocurre a favor del gradiente de concentracin.4.1.1. Difusin simple: La forma ms sencilla por la que un soluto puede pasar de un lado a otro de la membrana es la difusin simple, movimiento neto no asistido de un soluto desde una regin donde su concentracin es mayor, a otra donde sta es menor. Dado que las membranas presentan una zona central hidrfoba, la difusin simple es un proceso relevante slo para molculas pequeas y relativamente poco polares. Tales molculas acceden a la bicapa lipdica por un lado, difunden pasivamente a travs de ella y emergen por el otro lado, desde donde pasan, de nuevo, al medio acuoso. El oxgeno es un modelo de molcula pequea y no polar, que atraviesa fcilmente la bicapa lipdica hidrfoba, por difusin simple. Esta circunstancia permite que los erotrocitos del sistema circulatorio tomen oxgeno en los pulmones y lo liberen luego en los tejidos. En los capilares de los tejidos corporales, donde la concentracin de oxgeno es baja, ste es liberado desde Ia hemoglobina y difunde pasivamente desde el citoplasma del eritrocito hacia el plasma sanguneo y desde ali a las clulas que limitan los capilares.E n los capiliares pulmonares se verifica el proceso contrario: el oxgeno difunde desde el aire inhalado en los pulmones, donde su concentracin es mayor, al citoplasma de los eritrocitos, donde la concentracin es menor.Las molculas hidrofilicas tambin pueden difundir con la condicin que no estn cargadas y que posean pequeo tamao. De esta manera pasan el metanol, etanol y el glicerol.
4.1.2. Difusin facilitada: La mayora de las sustanciase en las clulas son demasiado grandes o demasiado polares para atravesar las membranas por difusin simple con tasas de transporte razonables, pese a ser un proceso exergnico. Estos solutos entran y salen de las clulas y de los orgnulos de forma eficaz gracias a la asistencia de protenas transportadras que intervienen en el movimiento de solutos a travs de la membrana. Si el proceso es exergonico se denomina difusin facilitada pues el slutos difunde en el sentido impuesto por el gradiente de concentraciones (en molculas sin carga) o el gradiente electroqumico ( en iones). Como ejemplo de difusin facilitada consideraremos el paso de glucosa a travs de la membrana plasmtica del un eritrocit. La concentracin de glucosa es mayor en la sangre que en el eritrocito por lo que su entrada es exergonica, es decir no requiere aporte energtico, sin embargo la glucosa es muy grande y muy polar para poder difundir sin asistencia a travs de la membrana, se requiere de una protena de transprte para facilitar su entrada. Una diferencia fisiolgica fundamental entre la difusin simple y la difusin facilitada lo es su cinetica. En la difusin simple la velocidad de pasaje es siempre proporcional a la diferencia entre las concentraciones a ambos lados de la membrana, mientras que la difusin facilitada la velocidad de transporte aumenta rpidamente con la diferencia de concentraciones pero se llega a un tope. Cuand todas las PT estn funcionando al mximo de su velocidad se dice que el sistema esta saturado.
a) Mediada por canal: Estos forman poros o conductos hidrofilicos que recorren el espesor de todas las membranas celulares y permiten en flujo pasivo de iones a travs de estas. Son altamente especficos por lo que en general facultan el paso de un solo tipo de ion. Exiten canales que siempre permanecen abiertos y otros que se abren y cierran regulados por seales qumicas, elctricas o mecnicas que provocan cambios conformacionales en la protena. Canales regulados por voltaje: se abren y cierran como respuesta a cambios en el potencial de membrana. Existen canales regulados para sodio, potasio y calcio. Los canales de sodio de las clulas musculares permanecen cerrados cuando la membrana plasmtica mantiene su potencial de reposo. Cuando se despolariza un punto de la membrana influye sobre regiones cargadas de las protenas del canal y provoca su apertura. Canales regulados por ligando: se trata de canales-receptores que, al unirse cn su ligando especifico experimentan un cambio cnformacional que determina su apertura. Un ejemplo lo constituye el receptor de acetilcolina, canal de sodio regulado que en la sinapsis se encarga de iniciar la despolarizacin de la membrana postsinaptica. Los ligando pueden unirse al lado extracelular del canal, como los neurtransmisores o al dominio citosolico. Canales regulados mecnicamente: responden a fuerzas mecnicas que actan en la membrana. Se abren regulados por el estiramiento de la membrana, Se piensa que la tensin es trasmitida a las protenas del canal por elementos del citoesqueleto. En el caso de las clulas neuroepiteliales del odo interno, las fuerzas que actan sobre estos canales se transmiten a travs de fibrillas insertadas en la cara extracelular.
b) Difusin mediada por acarreador: PORINAS :Comparados con los canales inicos, los poros de las membranas externas de las mitocondrias, cloroplastos y muchas bacterias, son bastante ms grandes y mucho menos especficos. Estos poros se formanpor protenas denominadas porinas, que atraviesan varias veces la membranaSon muy especficos ya que discriminan incluso entre ismeros como es el caso de la glucosa y la galactosa que requieren permeasas diferentes EJEMPLO: GLUCOSA
c) Aquaporinas: Ms bien facilitan la entrada y salida rpida de molculas de agua en tejidos que as lo requieren. Por ejemplo los tbulos proximales del rin reabsorben agua durante la formacin de la orina; las clulas en este tejido tienen una elevada densidad de AQPs en sus membranas plasmticas. Lo mismo ocurre en los eritrocitos, que deben ser capaces de expandirse o retraerse rpidamente, en respuesta a los cambios repentinos de presin osmtica, que tienen lugar cuando se desplazan por las arterias renales o de otros rganos.
4.2. Transporte activo: en algunos casos las protenas transportadoras deben mediar el pasaje de iones o de molculas en contra del gradiente electroqumico. En esta circunstancia se requiere el uso de energa y se habla de transporte activo.
4.2.1. Transporte primario: Las permeasas especiales que utilizan ATP directamente como fuente de energa.a) Bombas clase p: Son protenas transmembrana multipaso funcionalmente y estructuralmente relacionadas entre si. Se denominan de tipo P porque se autofosforilan durante el ciclo de bombeo. Esta clase incluye muchas bombas ionicas responsables de generar y mantener gradientes de Na+ , K+, H+ y Ca2+ a travs de la membrana. Entre las propiedades que comparten estas AIPasas, estn las siguientes: estn formadas por un nico pptido con 8 a 10 segmentos, que zigzaguean atravesando varias veces la membrana, todos ellos son transportadores de cationes y por ultimo son sensibles a la inhibicin por el ion vanadato. La mayor parte de las bombas de tipo P se localizan en la membrana celular, donde son las responsables del mantenimiento del gradiente inico a ambos lados de Ia membrana plasmtica.
Bomba de sodio potasio ( Na+ / K+) : Un rasgo caracterstico de la mayora de las clulas animales es el alto nivel intracelular de iones potasio y el bajo nivel de iones sodio.La concentracin del ion potasio se mantiene generalmente en torno a 100-150m M en el interior de la mayora de las clulas animales mientras que la concentracin externa es mucho menor que sta pudiendo fluctuar ampliamente. Por el contrario ,la concentracin intracelular de iones de sodio est en torno a 10-15 mM mucho menor que la del medio circundante. El resultado de esto es que, tanto la entrada de iones potasio como la salida de iones sodio son procesos que requieren energa.La bomba de Na*/K* es una protena alostrica que presenta dos estados conformacionales designados E1 y E2. La conformacin E1 , est abierta hacia el interior celular y es muy affin por los iones sodio, mientras que Ia E2, se abre hacia el exterior y une potasio con gran afinidad. La presencia de sodio desencadena la fosforilacin de la enzima, estabilizndola en la conformacin E2.Funciones: Mantener diferencias de concentracin de Na y K intra y extra celulares Es electrogenica, el lado citosolico es normalmente mas negativo que el espacio extracelular Regula el volumen celular
b) Bomba clase F: Son protenas semejantes a turbinas, construidas a partir de diferentes subunidades. Se diferencian desde un punto de vista estructural de las de tipo P y se encuentran en la membrana plasmtica de las bacterias en la membrana interna de las mitocondrias y en la membrana mitocoidal de los cloroplastos. A menudo se denominan ATP sintetasa porque normalmente actan en sentido opuesto: en lugar de utilizar la hidrolisis de ATP para impulsar un gradiente de H, utilizan un gradiente de H a travs de la membrana para impulsar la sntesis de ATP a partir de ADP y fosfato.Las ATPasas de tipo F confirman un principio energtico importante: El ATP no es slo Ia fuente energtica necesaria para generar y mantener los gradientes inicos sino que estos gradientes pueden servir como fuente de energa para obtener ATP.
c) Bomba clase V: relacionadas con las de tipo F, que por lo general bombean H en lugar de sintetizar ATP. Bombean H hacia el interior de loso orgnulos como lisosomas, vesculas sinpticas y vacuolas de las plantas para acidificar el interior de estos orgnulos.
d) Superfamilia ABC: bombean sobre todo molculas pequeas a travs de la membrana celular en contraste con las ATPasas de tipo P y F que exclusivamente transportan iones.
Las ATPasas de tipo ABC tienen cuatro dominios, dos de ellos son protenas integrales de membrana, altamente hidrfobas mientras que los otros dos son protenas perifricas localizadas en la cara citoplasmtica de la membrana. Cada uno de los dos dominios integrales consta de 6 segmentos transmembrana que probablemente forman el elemento de paso de las molculas de soluto. Los dos dominios perifrico son los casetes que unen ATP y acoplan su hidrolisis al proceso de transporte . Estos cuatro dominios s on polipeptidos independientes en la mayora de las celilas, especialmente procariotas. Sin embargo, se conocen tambin ejemplos en los cuales los cuatro dominios son parte de un gran polipedtido multifuncional . Mientras que las otras tres clases de ATPasas transportan solo cationes, las de tipo ABC utilizan una notable diversidad de solutos. La mayora son especficas para un determinado soluto o un grupo de solutos relacionados entre si.
4.2.2. Transporte activo secundario: utiliza la energa potencial contenida en el gradiente favorable de la sustancia cotransportada. El gradiente electroqumico generado por la bomba de sodio potasio proporciona suficiente energa para propiciar el transporte activo de otras molculas escenciales.
a) Contratransporte: Se caracteriza por la sustancia a transportar es intracelular por ejemplo un contra-transporte de sodio por calcio, siendo el calcio la molecula a contra transportar, el sodio se une a una protena transportadora en la superficie externa de la membrana mientras que el calcio lo hace a su lugar de unin de la protena pero en el lado interno de la membrana(LIC)Una vez fijado ambas sustancias se produce un cambio de conformacin y la energa derivada del gradiente de concentracin para el sodio produce al desplazamiento hacia el interior y del calcio hacia el exterior.
b) Cotransporte: En caso del sodio con glucosa la energa de difusin que supone la separacin de dos medios con diferentes concentraciones de Na es aprovechado para mover la glucosa junto con el Na hacia el interior de la clula. Para ello es necesario la participacin de una protena transportadora que se encuentra en la membrana con un lugar de unin para el Na y otro para la glucosa. Una vez que se fija ambos se produce un cambio de conformacin de la protena y la energa del movimiento del Na produce el desplazamiento de la glucosa hacia el interior. Existen otros contransportadores importante como el: Sodio con aminocidos Sodio-Potasio-Cloroo Potasio-Cloro
5. Movimiento de agua a travs de la membrana : osmosis y presin osmticaOSMOSIS: El agua atraviesa una membrana como respuesta diferencias en la concentracin de solutos. El agua tiende a difundir desde el lugar con menor concentracin al de mayor concentracin de soluto. Esta difusin del agua, denominada smosis se observa fcilmente cuando una membrana de permeabilidad selectiva separa dos compartimientos, uno de los cuales contiene el soluto para el cual la membrana no es permeable. El movimiento osmtico de entrada o salida de agua en unaClula est relacionado con la osmolaridad o concentracin relativa de soluto en la solucin en que se encuentra la propia clula. Las disoluciones con mayor concentracin de soluto que el medio intracelular se denominan hipertnicas, mientras que las de menor concentracin son las hipotnicas. Las soluciones hipertnicas hacen que las molculas de agua difundan hacia el exterior de la celula y las hipotnicas que difundan hacia el interior. En otras palabras, el movimiento osmtico de agus se produce siempre desde la disolucin hipotnica a la hipertnica.