UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAMI10463.pdf · células dos tipos de...
41
Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 1 UNIVERSIDAD AUTONOMA METROPOLITANA IZTAPALAPA. CIENCIAS BIOLOGICAS Y DE LA SALUD (CBS) TRABAJO PARA OBTENER EL GRADO DE: LICENCIATURA EN BIOLOGIA EXPERIMENTAL TITULO: Técnicas Aplicadas al Estudio de Polimorfismos Genéticos Asociados con el Sistema Principal de Histocompatibilidad. FRAGOSO LONA JOSE MANUEL. TUTORES: Dr. GILBERTO VARGAS ALARCÓN Dr. JOSE LUIS GÓMEZ O. COTUTORA: QBP. GUADALUPE HERNANDEZ PACHECO 27 DE ENERO DEL 2003
Transcript of UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAMI10463.pdf · células dos tipos de...

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 1
UNIVERSIDAD AUTONOMA METROPOLITANA
IZTAPALAPA.
CIENCIAS BIOLOGICAS Y DE LA SALUD(CBS)
TRABAJO PARA OBTENER EL GRADO DE:
LICENCIATURA EN BIOLOGIA EXPERIMENTAL
TITULO:Técnicas Aplicadas al Estudio de PolimorfismosGenéticos Asociados con el Sistema Principal de
Histocompatibilidad.
FRAGOSO LONA JOSE MANUEL.
TUTORES:
Dr. GILBERTO VARGAS ALARCÓN
Dr. JOSE LUIS GÓMEZ O.
COTUTORA:
QBP. GUADALUPE HERNANDEZ PACHECO
27 DE ENERO DEL 2003

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 2
TITULO
Técnicas aplicadas al estudio de Polimorfismos genéticosasociados con el Sistema Principal de Histocompatibilidad
a) INTRODUCCIÓN
Hoy en día las técnicas de biología molecular se han convertido en una
herramienta de gran importancia en el ámbito de la investigación básica, así
como en el ámbito clínico para la identificación de distintas enfermedades y elestudio de su etiología.
Las técnicas de biología molecular se pueden aplicar a la búsqueda de
los Polimorfismos genéticos que se asocian con el desarrollo de enfermedadescardiovasculares en distintas poblaciones mexicanas. Para esto se busca la
tasa de mutaciones, que es una característica de cada proteína y que
presumiblemente está en función en parte de su flexibilidad a la hora de que
ocurren los cambios. Dentro de cada especie, las proteínas evolucionan
mutaciones que se eliminan o se fijan dentro del grupo reproductivo (por
selección natural). La presencia de dos o más variantes alélicas (querepresentan posibles formas alternativas de un gen) en la población a tratar sedenomina polimorfismo (1).
La biología molecular es el estudio bioquímico de la estructura y
expresión de los genes. Este campo sé inicio con el estudio genético de
organismos simples como los virus y las bacterias, y posteriormente seextendió a organismos más complejos. En los mamíferos, la biología molecular
sé empezó a utilizar en el estudio de la expresión génica de proteínas como la
hemoglobina y las inmunoglobulinas muy abundantes sintetizadas por células
de fácil acceso (células sanguíneas circulantes o de medula ósea). Con la
creación y el refinamiento de la ingeniería genética y de la metodología del
ácido desoxirribonucleico (ADN) recombinante, se abrió camino hacia elestudio de genes que producen proteínas menos abundantes, así comomutaciones genéticas y polimorfismos (2).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 3
Los polimorfismos parecen ser muy comunes en los genes de ácido
ribonucleico ribosomal (ARNr) y ácido ribonucleico de transferencia (ARNt) de
los que a menudo existen formas alternativas, con la ausencia ó presencia de
intrones. La presencia de dos o más variantes alélicas en la población se
denomina polimorfismo. Un determinado polimorfismo puede ser estable, en
cuyo caso ninguna de las formas tiene ventajas sobre la otra, o transitorio,como debe ocurrir durante el reemplazamiento de una variante por otra (1).
Complejo Principal de Histocompatibilidad
El sistema inmune se apoya en numerosos mecanismos que se
encargan de mantener su capacidad de responder hacia los diferentes agentes
infecciosos y algunos tejidos neoplásicos a los que es susceptible, pero no los
enfrenta con un sólo esquema, sino que se vale de todo un microcosmos de
eventos celulares y moleculares bastante complejos que son controlados engran medida por un grupo de genes denominado Sistema Principal deHistocompatibilidad (MHC) (del inglés, major histocompatibility complex).
Este no tiene ubicación especifica en los mamíferos es un grupo de
genes está localizado en el brazo corto del cromosoma 6 en el humano, en el
cromosoma 17 en el ratón y en el cromosoma 20 en la rata. Tal grupo de genes
fue identificado en sus inicios por el efecto que estos tenían sobre el trasplantede piel, y posteriormente en el control de la respuesta inmunológica (5).
Este sistema abarca entre tres mil y cuatro mil pares de bases de
nucleótidos y se ha podido localizar gracias a estudios de electroforesispulsada en el gel con el fin de estudiar megabases de DNA (6).
Las moléculas de histocompatibilidad son de dos clases, de la clase I yclase II y pueden presentarse varias versiones. Todas pueden coexistir de
manera simultánea en la membrana de la misma célula, aunque en diferentes
proporciones.
Las moléculas de histocompatibilidad son proteínas de membrana
formadas por dos cadenas y existe una región polimórfica (α1α2 y α1β1,
respectivamente) que une peptidos que interaccionan con TCRαβ (receptores
para antígeno a la célula T) y una región monomórfica (βα3 y α2β2) que
interacciona con el co-receptor (CD8 y CD4 respectivamente). Las células de

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 4
todos los tejidos disponen de moléculas de clase I excepto los eritrocitos. Sin
embargo unas pocas células con funciones inmunológicas expresan, además
moléculas de clase II: son presentadas en las células presentadoras deantígeno (abreviatura del inglés APC o antigen presenting cells).
La cadena alfa (α) consta de tres dominios α1, α2, α3 en donde los
dominios α1 y α2 son los más externos y donde radica la mayor parte del
polimorfismo de la molécula, el dominio α3 es no polimórfico y es la región
transmembranal de la molécula.
Los datos acerca de la estructura tridimensional de las moléculas clase I
surgen a partir de 1987, donde se realizan los primeros estudios para dilucidar
la estructura de estás moléculas y definir las características funcionales que
dirigen la presentación de péptidos por parte de estas moléculas. La
información acerca de la estructura tridimensional de las moléculas de clase II
es menos extensa, ya que los primeros datos sobre su estructura datan de1993.
Moléculas Clase I del Sistema Principal de Histocompatibilidad
Las moléculas de clase I se expresan sobre la superficie de la mayoría
de las células nucleadas, mientras que las moléculas clase II sólo se expresansobre la superficie de las células presentadoras de antígeno como el
macrófago, linfocito B, T y la célula dendrítica interdigitante. Hasta la fecha, se
conocen una gran cantidad de alelos en los genes de las moléculas (de laabreviatura en inglés HLA; human leukocyte antigens) de clase I y de clase II(7).
El análisis de estas estructuras se realizó por medio de cristalografía de
rayos X y se determinaron varios alelos de esta región clase I: HLA-A2, HLA
Aw68 y HLA-B27 y a su vez estas moléculas estaban unidas a una mezcla de
péptidos heterólogos. Estos ensayos establecieron los principios generales que
determinan la unión entre el péptido al sitio de unión de este en la molécula
clase I. Así mismo, permitieron establecer la hipótesis que postula que en las
porciones terminales del péptido se presentan determinantes esenciales paraunirse a la molécula de clase I con una alta afinidad y especificidad.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 5
Cada versión está codificada por un gen diferente. En humanos las
versiones de las moléculas de la clase I se denominan HLA-A, HLA-B y HLA-C
y las de la clase II HLA-DR, HLA-DQ y HLA-DP, dentro de esta región de clase
II se incluyen los determinantes HLA-Dw los cuales se determinan por cultivomixto de linfocitos (6). Esta nomenclatura se considera no adecuada, ya que
las moléculas HLA se expresan también en células que no son leucocitos.Todas las células pueden por lo tanto presentar péptidos a los linfocitos Tutilizando cualquiera de sus moléculas de la clase I.
Moléculas Clase II del Sistema Principal de Histocompatibilidad
Los genes ubicados en la región de clase II codifican los antígenos HLA-
DP, DQ y DR que son un heterodímero constituido por una cadena α de
aproximadamente 30 kda y una cadena β de aproximadamente 28 kda.
Los estudios estructurales realizados sobre ésta molécula son
relativamente recientes, la comparación con los de las moléculas clase I hanpermitido entender los mecanismos por los cuales une los péptidos para ser
presentados a las células T, que se realizan a través de uniones entre los
hidrógenos del péptido antigénico y del sitio de unión en la molécula HLA claseII (9).
Las células presentadoras de antígeno pueden hacerlo, con cualquierade sus moléculas de la clase II. Las moléculas de histocompatibilidad de cada
versión tampoco son todas iguales, cada individuo puede expresar en sus
células dos tipos de moléculas (alotipos) que a menudo son distintos (HLA-A2 y
HLA-A11) una codifica para un gen de origen materno y otra para origenpaterno. Lo mismo ocurre para las demás moléculas de clase I y de clase II.
En conjunto el polimorfismo de las moléculas de histocompatibilidad esextraordinario, pero también lo es su trabajo, la presentación de péptidos a los
linfocitos Tαβ. Conviene aclarar que la variabilidad del TCRαβ es diferente del
polimorfismo de las moléculas de histocompatibilidad. Los genes que codifican
las cadenas del TCR son muy parecidos o iguales en todos los individuos; su
combinación al azar en cada individuo es lo que genera el polimorfismo,
aunque cada linfocito T expresa un único tipo de TCRαβ (4).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 6
La molécula clase II tiene cuatro dominios externos: α1, α2, β1 y β2,
análogos a los dominios α1, α2,α3 y a la β2-microglobulina de la molécula de
clase, respectivamente. Los dominios α2 y β2 son del tipo de los dominios de
las inmunoglobulinas, muy conservados y los dominios α1 y β1 son
polimorficos. El polimorfismo de las moléculas de clase II se encuentra en lascadenas beta de los antígenos HLA-DP, -DQ y -DR.
En todas las especies de mamíferos estudiadas se ha constatado que
los genes responsables de las porciones polimórficas de las moléculas de clase
I (cadena α) y de clase II (cadenas α y β) se encuentran juntos en una pequeña
zona del genoma, a dicha zona se le ha denominado complejo principal dehistocompatibilidad o MHC (4).
En 1989 se encontraron dos loci de la proteína de choque térmico
(HSP70-1 y HSP70-2), situados entre los genes de clase III y el del factor de
necrosis tumoral alfa. Los genes clase I, II y III se heredan de maneramendeliana codominante. Las moléculas clase I se expresan en la membrana
de todas las células nucleadas del cuerpo, excepto en neuronas y trofoblastos
maduros; las moléculas de clase II se expresan solo en ciertas células como
macrófagos, linfocitos B, células endoteliales, células dendriticas y células deLangerhans (6).
En el humano, los genes del MHC se ubican en el brazo corto del
cromosoma 6, y esta es una región altamente polimorfica, dentro del genoma
humano, y que se divide a su vez en tres regiones denominadas sistema HLA(Human Leucocyte Antigens): HLA clase I, HLA clase II y una región no HLA
denominada clase III, que se encuentra entre la I y la II, cuyos genes codifican
para los componentes del complemento C2, factor B (fB), y C4A y C4B, asícomo los genes estructurales de la 21 - hidroxilasa A y B (21-OHA y 21-OHB).
En el sistema principal de histocompatibilidad se incluye el gen de la
glioxalasa I, los genes del factor de necrosis tumoral alfa y beta (TNFα y
TNFβ), que están entre el HL-B y la región clase III y el gen de una proteína
con estructura periódica no común llamada RD, que se encuentra entre el gen
del factor B y el de C4A. Recientemente se han encontrado otros cinco genes
asociados al locus HLA-B llamados transcritos asociados a B, que son BAT -

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 7
1,2,3,4 y 5, así como también un gen llamado B-114 que es análogo a un gencon igual nombre que el del ratón (6).
En la región de clase I del MHC se encuentran los genes HLA-A, B y C
que codifican una glucoproteína de 45 kilodaltones unida a una cadena no
polimórfica conocida como β2-microglobulina, codificada por un gen ubicado en
el cromosoma 15.
Como parte de su función, las moléculas HLA clase I requieren unir
péptidos en un intervalo de 8 a10 residuos de aminoácidos. Este espectro se
considera no infinitamente variado en cuanto a secuencia de aminoácidos,
tiene un límite pero con una cierta preferencia ubicada en la segunda posiciónaminoterminal del péptido (posición denominada P2) y para el residuo carboxi-
terminal en el caso de péptidos de 9 residuos. Sin embargo, el sitio de unión
del péptido de la molécula clase I presenta una considerable flexibilidad para
extenderse y ampliar su rango para unir péptidos de secuencias más grandes.
Estudio recientes han permitido entender con mayor exactitud los
mecanismos que influyen esta propiedad funcional de las moléculas clase I endonde la molécula de agua es capaz de actuar como extra opcional en ciertas
posiciones del sitio donde se une el pepito en la molécula HLA clase I,
realizando cambios muy finos en la conformación y en las fuerzaselectrostáticas tanto de la molécula HLA como del péptido antigénico (7)(9).
Los genes del MHC también desempeñan un papel muy importante en laresistencia a infecciones y en la susceptibilidad a un gran número de
enfermedades autoinmunes. Las primeras observaciones que indicaron que el
control de la respuesta inmune estaba estrechamente relacionado a los genes
del MHC han servido de base para desarrollar numerosas y detalladas
investigaciones en las últimas dos décadas, mismas que han llevado al
conocimiento en el ámbito molecular de los detalles más pequeños delreconocimiento celular en el sistema inmunológico (5).
Los genes del MHC desempeñan un papel central en la respuesta
inmunitaria frente a antígenos proteicos, debido a que los linfocitos T
específicos para un antígeno no lo reconocen en su forma libre ni en su forma
soluble, sino que reconocen porciones de los antígenos proteicos (es decir,
péptidos) unidos de forma no covalente a productos génicos del MHC, debido a

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 8
que las moléculas del MHC II están asociadas a membranas celulares y no se
secretan, los linfocitos T reconocen antígenos extraños sólo cuando estánunidos a estas moléculas en la superficie de otras células.
Su función principal es la de presentar péptidos propios o extraños
procesados de manera previa en el citoplasma y unidos a la parte más
extracelular de estas moléculas mediante un mecanismo complejo que
involucra la participación de otras moléculas chaperonas para finalmente ser
exportados al exterior de la membrana de la célula y ser presentados a los
linfocitos T CD8+, mientras que las moléculas de clase II presentan péptidos
procesados de origen endógeno por una ruta diferente a los presentados por
las clase I y ayudado de otro tipo de moléculas chaperonas a los linfocitos T
CD4+ iniciando así los procesos de respuesta inmune adecuada en contra delantígeno que la originó (10).
En resumen, las moléculas del MHC proporcionan un mecanismo para
presentar los péptidos antigénicos a las células T, lo que permite muestrear al
organismo en búsqueda de péptidos derivados de proteínas extrañas. Los
genes del MHC son muy polimórficos, es decir, existen numerosos alelos en
una población, que difieren en sus distintas capacidades para unirse ypresentar a los diferentes determinantes antigénicos de las proteínas (8).
En la región de clase I del MHC de los humanos (sistema HLA, verFigura 1) se han identificado los genes HLA-A, -B y C que determinan las
cadenas α de las moléculas.
En la región de clase II se encuentran los genes HLA-DRA, -DQA1 y -
DPA1, que determinan las cadenas β de las moléculas HLA-DR (1 a 18), HLA-
DQ Y HLA-DP y los genes HLA-DRB1, -DPB1, que determinan las cadenas β
de dichas moléculas. Algunos cromosomas contienen, además, un segundo
gen (que puede ser HLA-DRB3, o DRB4, o DRB5) que codifican
simultáneamente otra cadena β para las moléculas HLA-DR (4) (ver Figura 1).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 9
Figura 1. Distribución cromosómica de los genes que constituyenel Sistema Principal de histocompatibilidad en el linfocitoshumano. Existen tres conjuntos de genes implicados en la capacidadde un organismo para activar la respuesta inmunológica haciaantígenos. En la figura solo se hace referencia a dos clases demoléculas de histocompatibilidad.
El polimorfismo alélico de los antígenos de clase I y clase II se reconoce
fenotípicamente por serología. Las variantes de HLA - Dw se detectan porcultivo mixto de linfocitos. El polimorfismo se reconoce mediante secuenciación
de nucleótidos, análisis de fragmentos de restricción y la secuencia primaria deaminoácidos (6) (ver Figura 2).
Los genes de la clase III del CHM, después del HLA forman el conjunto
de marcadores genéticos más polimórfico del humano, los cuatro loci de esta
región (C2, Factor B, C4A y C4B) se heredan en bloque, pues al analizarvarios centenares de meiosis informativas, se ha observado que no hayintercambios genéticos entre ellos (ver Figura 1).
El polimorfismo estructural de C4 es mayor que el de C2 o fB y es el que
proporciona el polimorfismo de los complotipos (una combinación dada de
alelos de los loci fB, C2, C4A y C4B forman un llamado complotipo o haplotipode genes del complemento) (6).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 10
Importancia del Sistema Principal de Histocompatibilidad
La función del MHC en la respuesta inmune está influenciada por elpolimorfismo genético que existe en este complejo. El polimorfismo es la
presencia de un mayor número de variantes genéticas que las esperadas en
una población, como ya se mencionó, los genes del sistema HLA muestran un
alto grado de polimorfismo, esto proporciona la ventaja de contar con un gran
numero de moléculas HLA que aumentan el repertorio para la presentación de
antígeno, lo que permite presentar una gran cantidad de péptidos antigénicosderivados de la extensa cantidad de patógenos ambientales existentes y aún
los que no existen.
Figura 2. Variabilidad regional de aminoácidos en las proteínas delsistema principal de histocompatibilidad. En la figura se muestran lasregiones de alto y bajo recambio en aminoácidos que establecen parte delpolimorfismo en las proteínas que conforman a las moléculas dehistocompatibilidad clase I y II. Los sitios en color obscuro representansitios de alta tasa de recambio de aminoácidos.
Si el polimorfismo fuera limitado esto provocaría que poblaciones
enteras fueran susceptibles a elementos infecciosos que sufrieran de cambios
ligeros en su composición antigénica para los cuales los individuos fueranincapaces de responder. Sin embargo, en algunas ocasiones estos procesos
sufren algunas alteraciones, lo que origina problemas patológicos quedesencadenan la autoinmunidad (5).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 11
Las moléculas de MHC de clase I y clase II tienen diferente distribución
celular, lo cual refleja las diferentes funciones efectoras de las células T que las
reconoce. Las moléculas de clase I se encuentran prácticamente en todas las
células nucleadas, y presentan péptidos de agentes patógenos citosólicos,
generalmente virus. Este reconocimiento trae como consecuencia la
destrucción de la célula por parte de los linfocitos T CD8+. Por otro lado, lafunción principal de las células T CD4+ que reconocen moléculas de MHC declase II es activar a otras células efectoras del sistema inmune.
Por lo tanto, las moléculas de MHC de clase II se expresan en células
presentadoras de antígeno, como el linfocito T, los macrófagos y en las células
dendríticas.
Cuando las células T CD4+ reconocen péptidos unidos a moléculas de
HLA de clase II en las células B, estimulan la producción de anticuerpos por
parte de las células B. Del mismo modo, las células T CD4+ que reconocen
péptidos unidos a moléculas de HLA de clase II en los macrófagos, activan a
dichas células para que destruyan a los agentes patógenos que se hallan en
sus vesículas. La expresión de moléculas de HLA de clase I y II está reguladapor citocinas, en particular por los interferones (IFN).
El papel biológico de las moléculas de histocompatibilidad es la
presentación de péptidos a los linfocitos Tαβ. En condiciones normales no hay
moléculas MHC vacías en la superficie de las células de hecho todas lasmoléculas de histocompatibilidad llevan siempre un péptido pegado que es
imprescindible para que la molécula sea expresada en la membrana. La
presentación de péptidos obedece a la regla clase I/CD8, clase II/CD4; Pero
esto no es fruto de la casualidad y de hecho él mérito no es de las moléculas
de histocompatibilidad, son de los linfocitos T que son culpables de este
fenómeno por que como se sabe los linfocitos Tαβ pertenecen a dos categorías
los que tienen un TCRαβ que reconocen péptidos en moléculas de
histocompatibilidad de clase I y los que tienen un TCRαβ que prefiere hacerlo
con las de la clase II.
Los de la clase I son siempre de los CD8 y los de la clase II son los CD4.
La razón de esta selectividad está en la interacción del co-receptor (CD4 o
CD8) con la molécula de clase II o I, dentro de cada una estas dos categorías

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 12
de linfocitos Tαβ, se pueden distinguir tantas subcategorías como moléculas de
histocompatibilidad distintas tenga el individuo.
Cada variante de molécula de histocompatibilidad presenta distintos
péptidos; esta es quizá la característica mas destacada de las moléculas de
histocompatibilidad, cada molécula tiene una estructura peculiar y única
especialmente en la región donde se une el péptido. Esto hace que una a
ciertos peptidos que otros, por ejemplo la molécula HLA-B27 presentará toda
una familia de peptidos de unos nueve aminoácidos que tengan una argininaen la segunda posición (4).
Por lo que las secuencias de ácidos nucleicos pueden variar confrecuencia relativamente alta entre los individuos normales de una población,
es decir que al menos el 1 por 100 de los individuos puede expresar un gen
que difiera del gen homólogo del resto de los miembros de la población por lo
que tales genes se dice, que son polimórficos con la característica de que cada
variante común de un gen polimórfico presente en la población se le llama
alelo, como en los seres humanos ciertos alelos de HLA de loci diferentes seheredan juntos, un fenómeno llamado desequilibrio de la unión. Tales
haplotipos (grupo total de alelos del MCH presentes en cada cromosoma), en
los que múltiples genes HLA permanecen unidos, se asocian a ciertasenfermedades autoinmunitarias (ver Tabla I).
Para el año de 1973 se informó la asociación entre espondilitisanquilosante y el antígeno HLA B27, desde entonces más de 50
enfermedades han sido descritas como asociadas con uno o más antígenos del
CMH. Estas enfermedades se pueden incluir básicamente en tres grupos:
1. Enfermedades asociadas a los antígenos clase I (principalmente las
espondiloatropatias asociadas al B27).
2. Enfermedades asociadas a los antígenos clase II principalmente con ellocus HLA DR.
3. Enfermedades asociadas a los antígenos clase III.
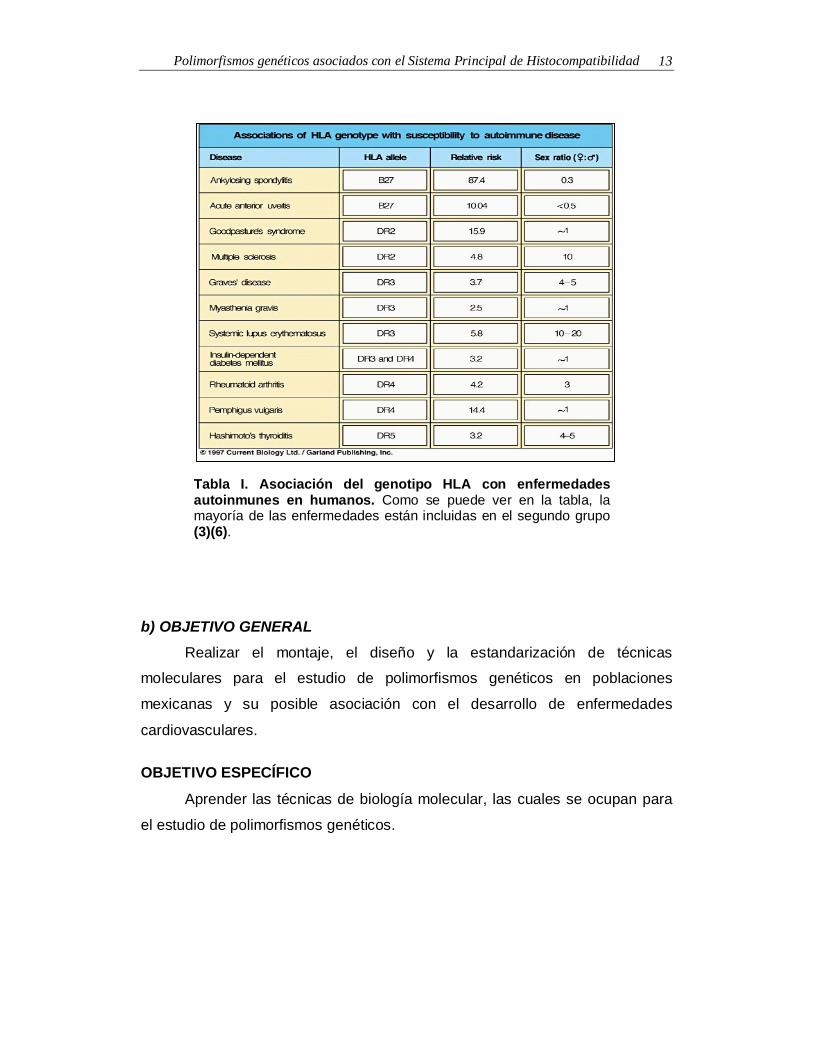
Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 13
Tabla I. Asociación del genotipo HLA con enfermedadesautoinmunes en humanos. Como se puede ver en la tabla, lamayoría de las enfermedades están incluidas en el segundo grupo(3)(6).
b) OBJETIVO GENERALRealizar el montaje, el diseño y la estandarización de técnicas
moleculares para el estudio de polimorfismos genéticos en poblaciones
mexicanas y su posible asociación con el desarrollo de enfermedades
cardiovasculares.
OBJETIVO ESPECÍFICOAprender las técnicas de biología molecular, las cuales se ocupan para
el estudio de polimorfismos genéticos.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 14
c) METODOLOGÍA UTILIZADA
Obtención del ADN
A partir de 10 ml de sangre venosa empleando EDTA 0.5M como
anticoagulante, se realizó la técnica de expulsión salina. Dicho procesoconsistió en someter a la sangre a un baja osmolaridad para lisar los eritrocitos.
1. En un tubo Falcon de 50 ml se colocaron de 5 a 10 ml de sangre total y se
añadieron 40 ml de buffer lisis para células rojas 1X que se Mezclo durante
10 minutos a temperatura ambiente.
2. El hemolizado se centrifugo a 2000 rpm durante 10 minutos. (Centrifuga IEC
Center-8R U.S.A.)
3. El sobrenadante se descartó.
4. El botón celular se volvió a resuspender en 40 ml de buffer de lisis de rojos,
y mezcló durante un minuto.
5. Se volvió a centrifugar a 2000 rpm. durante 10 minutos. (repitiendo 3 o 4
veces en las mismas condiciones)
6. Finalmente, se descarto el sobrenadante. Y el tubo de centrífuga se invirtióde posición sobre papel absorbente para retirar el exceso de liquido.
7. El botón celular se resuspendió en 160 µl de buffer lisis de leucocitos
conteniendo proteínasa K, 40 µl de proteínasa K (10 mg/ml), 40 µl de SDS
al 20% y 300µl de agua estéril (volumen final 800 µl). La muestra se
transfirió a un tubo eppendorf de 1.5 ml.8. La muestra se incubó a 37° C durante toda la noche o bien a 55° C por al
menos dos horas. En este ultimo caso añadió 20 µl de proteínasa K
después de una hora de incubación.
9. Transcurrido este tiempo, se adicionaron 200 µl de NaCl 6 M. y mezcló
vigorosamente en el vórtex para precipitar las proteínas.
10. La mezcla se centrifugó a 13,000 rpm durante 10 minutos.
11. El sobrenadante se colocó en un tubo eppendorf limpio de 1.5 ml
12. Se dividió el sobrenadante en dos tubos ependorf (aproximadamente 450 µl
por tubo) y se añadieron 900 µl de alcohol absoluto frío, mezclando por
inversión hasta observar el DNA precipitado.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 15
13. El ADN precipitado se centrifugó a 13,000 rpm durante 5 minutos, se
decantó el alcohol y se lavó con alcohol al 70%. (Pipeteando suavemente).
14. Se volvió a centrifugar a 13000 rpm durante 5 minutos, y retiró el etanol y el
tubo se colocó en un desecador hasta eliminar el exceso de liquido.
15. El DNA se hidrató con 100 µl de agua estéril y guardar el DNA a 4°C.
En el material hidrato se determinó el contenido de ADN. Para esto, se hizo
una dilución de 1 / 100 y/o 1 / 200, que dependió del tipo estudios a realizar.
1. Se tomaron 5 µl de la muestra de DNA en un tubo ependorff limpio de 1.5 ml
y se colocan 495 µl de agua estéril, mezclando con la pipeta suavemente.
2. La muestra obtenida se colocó en la celda del cuantificador de ADN(BIORAD, E.U.A.) el cual proporciona al mismo tiempo, las absorbencias a 260
nm y 280 nm que proporcionan las concentraciones de cada una de las
absorbencias. (la pureza que este presenta)
3. Después de la cuantificación, se hicieron los cálculos correspondientes para
la realizar el amplificado del ADN por reacción en cadena de la polimerasa
(PCR). Tomando en cuenta que la cantidad de DNA en ng/µl dependió del tipo
de HLA a buscar (para el diferente HLA que se deseaba tipificar).
Formula para cuantificación de ADN:
Abs(260) (50) (100) = X ng/µl
Donde:
260nm = absorbencia que da el DNA.
50 es un factor de dilución.
100 es la dilución que se hace para medir en espectrofotómetro.
Nota: Esto es por citar alguna de las cosas que se hacen en el laboratorio,
pero se hacen más cosas referente a la técnica de PCR. Como una sola
tipificación de un HLA en especifico como él -A, -B, -C, DQ y DR. Hubofamiliarización con técnicas de hibridación en Dot Blot reverso, la preparación
de geles de poliacrilamida y otras.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 16
Tipificación de los genes HLA
Para llevar acabo este procedimiento se requirió una concentración de
100 ng en 1 µl; por lo que dependiendo del HLA en estudio (se necesitaban
entre 40 y 80µl de ADN).
HLA clase I y clase II
La tipificación de los genes HLA clase I y clase II se realizó por la
reacción en cadena de la polimerasa revelado con iniciadores específicos de
secuencia (PCR-SSP) utilizando una serie de mezclas de iniciadoresespecíficos para cada alelo o grupo de alelos. (12). Dicho sistema empleó un
total de 96 mezclas diferentes que permitieron la detección de cada uno de los
alelos HLA (Pel-Freez Clinical Systems, Brown Deer, Wisconsin, USA)
Para cada placa HLA-A/B/DR se emplearon 580 µl de una solución
amortiguadora para PCR que incluye los desoxinucleótidos: dATP, dTTP,
dGTP, dCTP, 228 µl de agua estéril , 9.3 µl de Taq DNA polimerasa 5U/µl de
Thermufilus aquaticus BM (Boehringer Mannheim GmbH, Alemania), de esta
mezcla se tomaron 8µl y se colocaron en el pozo número 96 de la placa, el cual
funciona como control negativo, posteriormente a la mezcla anterior se le
agregaron 80 µl del DNA del paciente con la concentración antes mencionada,
y de esta mezcla final se colocaron 8 µl en cada una de los 95 pozos restantes
de la placa. Finalmente, la placa se sometió al siguiente perfil de amplificación:
Paso 1 1 ciclo 1 minuto a 96°CPaso 2 5 ciclos 96°C 25 segundos
70°C 50 segundos72°C 45 segundos
Paso 3 21 ciclos 96°C 25 segundos65°C 50 segundos72°C 45 segundos
Paso 4 4 ciclos 96°C 25 segundos55°C 60 segundos72°C 120 segundos
Mantenimiento 4°C

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 17
Los productos de PCR se verificaron mediante electroforesis en gel de
agarosa al 2 % (Boehringer Manheim GmbH), teñido con bromuro de etidio en
una concentración de 1 mg/dl, e interpretado con un análisis de computación
diseñado para tal propósito (Unimatch-Pel-Freez).
Tipificación de los genes LMP2 y LMP7 por PCRPara esto se emplearon los siguientes cebadores; LMP2X: 5´CTT GAA
CCA GGG AGG CGA AGT TTG 3´; y LMP2Y: 5´CAG CGT AAC CGA AGA
GTG CAT AGT 3´ para LMP-2 y LMP7X: 5´CGG ACA GAT CTC TGG GTG CT
3´; y LMP7Y 5´CTT CCC TAC TGC CCC AAG CT 3´ para LMP7.
La amplificación se llevó a cabo con 0.25 ng de DNA genómico,
utilizando 1 unidad de Taq DNA polimerasa (Promega, Madison, WI, USA), 200uM de cada dNTP, 2 mM de MgCl2, amortiguador de PCR 1X y 25 pmoles de
cada iniciador en un volumen final de 25 µl. La reaccione se realizó en un
termociclador Perkin Elmer modelo 9700 (Foster City, CA, USA) bajo las
siguientes condiciones: 30 ciclos de 94°C 60 segundos, 62°C 45 segundos, y
72°C 60 segundos para LMP7 y 30 ciclos de 94°C 60 segundos, 57°C 50
segundos, y 72°C 60 segundos para LMP2.
Preparación del gel de agarosa al 2%
Se pesaron 2 gramos de agarosa LE (Promega, Wi. USA), sé disolvió en
5ml de TBE 10X y 95 ml de agua destilada. Para un volumen final de 100 ml.
Luego, se calentó hasta que hirvió y después se dejo enfriar hasta 40° C
aproximadamente y se agrego 1.5µl de bromuro de etidio. Mezclando
perfectamente. La mezcla se vació sobre el molde del gel, dejándolo secar
cuando menos unos 25 minutos antes de usarlo.
Para correr el DNA mezclarlo con una solución gel loading de rojo de
cresol a razón de 3:10µl. Las muestras se colocaron en el gel previamente
sumergido en TBE .5X. El gel se corrió a 80 mV durante 30 minutos
aproximadamente. La separación se visualizo en una lámpara de luzultravioleta (11).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 18
Preparación de geles de poliacrilamida
Previamente, los vidrios se lavaron con detergente y se enjuagaron con
agua corriente, posteriormente se enjuagaron con agua destilada y finalmente
se limpiaron con alcohol al 96°. Se colocaron los separadores laterales entre
los vidrios y el inferior y se sujetaron con clips rodeando todo el vidrio. Se
preparo la solución de acrilamida:
Acrilamida al 30% --------------------------16.5 ml
TBE 10X-----------------------------------------5 ml
Persulfato de amonio al 10%----------------400µl
Temed--------------------------------------------80µl
Agua destilada---------------------------------28ml.
Todo esto se mezcló y se vació en el molde e inmediatamente se colocó
el peine. Se dejó polimerizar (aproximadamente 20-30 minutos) y se dispuso enla cámara de electroforesis. La cámara se cubrió con solución amortiguadora
TBE 1X. Las muestras se sometieron a la corriente eléctrica durante 5 horas a
80 volts.
Revelado de geles de poliacrilamida con nitrato de plata
El gel de acrilamida se reveló con una solución de nitrato de plata.
Previamente, se lavaron los recipientes para el revelado del gel con agua
corriente y con agua destilada, y después se enjuagó con la solución fijadora.Se desmontó el gel de acrilamida y se colocó en el recipiente con la solución
fijadora (aproximadamente 100ml), se dejó embebido con agitación suave
durante 10 a 15 minutos.
Se desecho toda la solución y se agregó la solución de tinción (100ml)
cuidando de que al vaciar la solución no caiga directamente sobre el gel. Se
dejó en agitación leve durante 5 minutos. Se eliminó toda la solución y seagregaron unos 10 ml de solución de revelado, cuidando que no caiga
directamente sobre el gel, se enjuago el recipiente con la solución y se
desecho.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 19
Finalmente, se enjuagó con agua destilada 10 ml. Y se agregaron 90 ml
de solución de revelado, durante unos 10 minutos o hasta que se visualicen las
bandas en el gel, este se tornara de color amarillo. La reacción se detuvo
cuando se añadió una solución de ácido acético al 1.5 % (00 ml).
El gel de acrilamida al 10% se colocó bajo una luz blanca
(megatoscopio) y se interpretaron las bandas de acuerdo a su polimorfismo yse tomó una foto que se imprimió en papel (polaroid) (11).
La hibridación por Dot Blot Reverso
Se hizó una desnaturalizacion de ADN, que consta en agregar al
amplificado aproximadamente 20 ó 25 µl de solución desnaturalizante e
incubarlo a temperatura ambiente aproximadamente 30 minutos. Luego el
material amplificado se pre-calentó el horno de hibridacion a 52° C + ó - ungrado; Junto con las soluciones de lavado y de hibridacion. Las tiras de
hibridación se rotularon (dependerá de lo que se quiera hibridar ya sea -A, -B, -
C, DR ó DQ) y colocaron en la charola previamente lavada con alcohol
absoluto. Se agregaron 2 ml de Buffer de hibridación a 52°C a cada tira y
posteriormente agregó el desnaturalizado y se incubó durante 30 minutos a
52°C, con constante agitación. Se eliminó el Buffer de hibridación pasados
estos minutos y lavó con 2 ml de Buffer de lavado a temperatura ambiente
(este paso es lo mas rápido posible). Se agregó 2 ml de Buffer de lavado que
esta a 52°C y se incubó a 52°c durante 15 minutos en constante agitación.
Para el revelado y el conjugado, las tiras se pasaron al recipiente de
revelado agregando 1.1 ml de la mezcla de conjugado a cada tira que esta
conformada por 1.1 ml de Buffer de lavado mas 3.3 µl de estreptividina - HRP a
temperatura ambiente (para una sola tira) durante 15 minutos, con agitación
constante. Se hicieron dos lavados con Buffer de lavado a temperatura
ambiente cada uno de 5 minutos, con agitación constante. Después se hace unlavado con 1.1 ml de Buffer de citratos por 5 minutos a temperatura ambiente.
Se reveló con 1.1 ml de la mezcla del sustrato A y B que esta conformado por
0.93 ml de solución A + 0.233 ml de la solución B, con agitación constante
hasta observar las bandas en la tira.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 20
La interpretación de las tiras resultantes y las bandas que se aprecien se
interpretaran en un programa de computadora (Dynal Patter Matching Program
v5) para identificar los posibles haplotipos existentes en los individuos en
estudio.
d) ACTIVIDADES REALIZADASDurante la estancia en el laboratorio de Biología Molecular del INCICH, el
alumno aprendió las técnicas de biología molecular aplicadas en tales estudios:
1) La extracción de ADN por expulsión salina (sating-out) de muestras deindividuos indígenas y mestizos.
2) La determinación del contenido de ADN con espectrofotómetro a unaabsorbencia de 260 nm y 280 nm, con el propósito de que los resultados
obtenidos de las absorbencias a 260 nm servirán para realizar él calculo yajustar el contenido de ADN.
3) La realización de la reacción en cadena de la polimerasa (PCR) y los
componentes que esta necesita con respecto a la tipificacion del SPH.
4) La preparación de geles de agarosa al 1.5% y 2% para verificar el nivel deamplificación de las muestras en el termociclador.
5) La preparación de geles de poliacrilamina para verificar los fragmentos derestricción después de las digestiones con enzimas de restricción.
6) La participación en otra técnica como hibridación con dot blot reverso, al
mismo tiempo que se participa en la preparación de distintas soluciones que seocupan para el desarrollo de estas técnicas, además del manejo del material de
laboratorio como micropipetas, cámaras de electroforesis, termocicladores,
centrifugas, secadores ADN, espectrofotómetro y otros.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 21
e) OBJETIVOS Y METAS ALCANZADOS
Para cumplir con el objetivo principal planteado al inicio de la prestación
de Servicio Social que era el montaje, diseño y estandarización de técnicasmoleculares para el estudio de polimorfismos genéticos en poblaciones
mexicanas fue la indispensable conocer el fundamento teórico y práctico de las
técnicas de biología molecular. Por lo que, inicialmente se aprendió desde el
manejo de instrumentos de laboratorio como: micropipetas, cámaras de
electroforesis, termocicladores, centrifugas, secadores de ADN,
espectrofotómetro entre otros. Así como, la preparación de soluciones como el
TBE 10X, las soluciones para el preparado de geles de poliacrilamina, la
proteínasa K, buffer de proteínasa K, solución de lisis de eritrocitos entre otrassoluciones.
Una vez que las metodologías estuvieron estandarizadas se emplearon
para establecer los polimorfismos genéticos en varias moléculas Clase I y
Clase del complejo principal de histocompatibilidad, así como de enzimas que
se han observado que pueden estar implicadas en algunos enfermedades
inmunológicas (paraoxonasa, polipéptidos de bajo peso componentes deproteosomas -LMP-).
Como ya se mencionó en la introducción los genes del sistema HLA
muestran un alto grado de polimorfismo, esto proporciona la ventaja de contar
con un gran número de moléculas HLA que aumentan el repertorio para la
presentación de antígeno, lo que permite presentar una gran cantidad de
péptidos antigénicos derivados de la extensa cantidad de patógenosambientales existentes.
Algo importante es que aun que no se aprendieron todas las técnicas por
lo menos se aprendieron las más aplicables para el desarrollo de estos
estudios, por lo que el desarrollo del Servicio Social tendrá un seguimiento para
el desarrollo de mi proyecto de investigación, el cual se desarrollara en elmismo instituto y en el mismo departamento.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 22
f) RESULTADOS Y CONCLUSIONES
En el Laboratorio de Biología Molecular del Departamento de Fisiología
en el Instituto Nacional de Cardiología "Ignacio Chávez" se emplean técnicas
moleculares en el análisis de los polimorfismos de algunos genes y su relación
con algunas enfermedades, principalmente de tipo cardiovascular. Por lo que,
no es de extrañar que haya incidencia de enfermedades autoinmunes enindividuos que tienen alguna molécula de histocompatibilidad en particular.
Los avances logrados en el conocimiento de la Genética y la Biología de
las moléculas clase de histocompatibilidad I, II y III, han estimulado la
investigación en el área de la asociación de enfermedades de los alelos deestos genes, especialmente por el papel que tiene en la regulación de larespuesta inmunológica.
En otras palabras se puede inferir que cualquier trastorno manifestado,
ya sea como aumento o defecto de la respuesta inmunológica, es natural que
involucre a cualquiera de los antígenos del Complejo Principal de
Histocompatibilidad (HMC de sus siglas en inglés), trayendo consecuenciasrelevantes en el tipo de respuesta inmunitaria que desarrolle un individuo enparticular (6).
Las enfermedades asociadas al CMH involucran tanto el terreno
genético del individuo como también ciertos factores ambientales que pueden
estar actuando conjuntamente para influir en la susceptibilidad o resistencia al
desarrollo de enfermedades especificas. La mayoría de los estudios en los quese ha analizado el papel de HLA (antígeno de histocompatibilidad en linfocito
humano) en las enfermedades, se han realizado en población abierta y ha
involucrado padecimientos que generalmente no ocurren en más de un
miembro de una familia. Tales estudios han generado información importante
concerniente a la prevalencia de ciertos antígenos HLA en forma aislada, ytambién han informado acerca de la frecuencia relativa de varios haplotipospara HLA para una enfermedad en poblaciones diferentes (6).
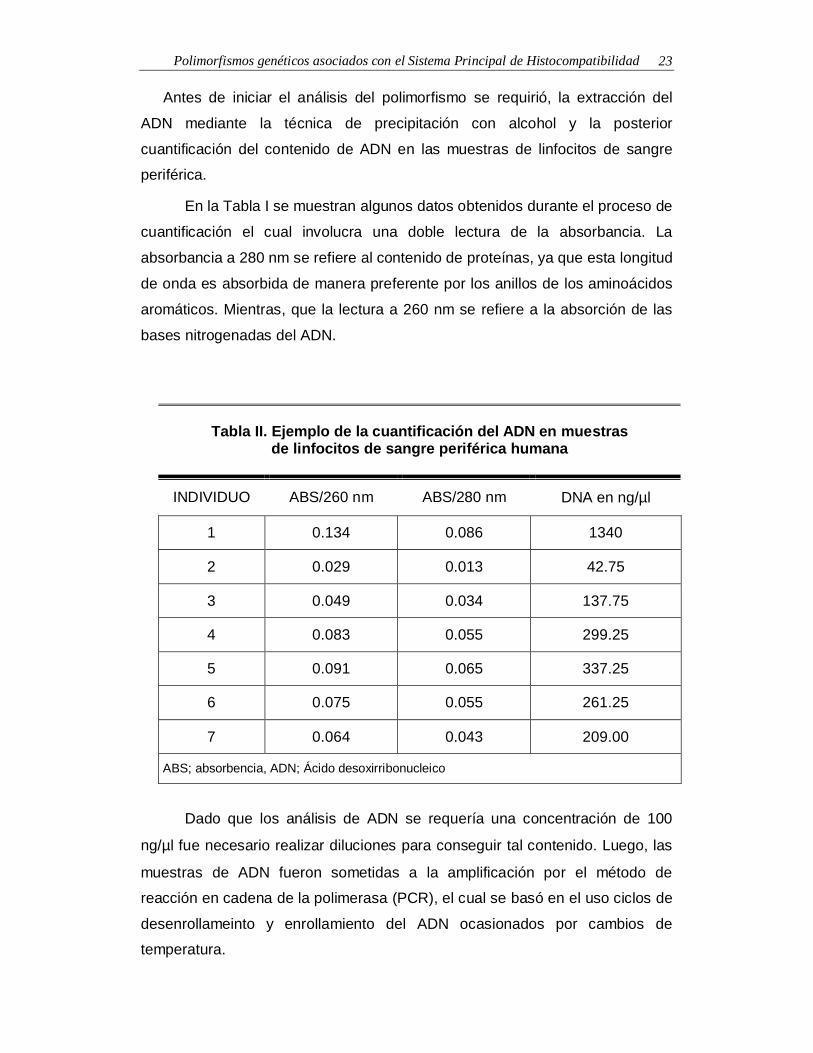
Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 23
Antes de iniciar el análisis del polimorfismo se requirió, la extracción del
ADN mediante la técnica de precipitación con alcohol y la posterior
cuantificación del contenido de ADN en las muestras de linfocitos de sangreperiférica.
En la Tabla I se muestran algunos datos obtenidos durante el proceso de
cuantificación el cual involucra una doble lectura de la absorbancia. La
absorbancia a 280 nm se refiere al contenido de proteínas, ya que esta longitud
de onda es absorbida de manera preferente por los anillos de los aminoácidos
aromáticos. Mientras, que la lectura a 260 nm se refiere a la absorción de las
bases nitrogenadas del ADN.
Tabla II. Ejemplo de la cuantificación del ADN en muestrasde linfocitos de sangre periférica humana
INDIVIDUO ABS/260 nm ABS/280 nm DNA en ng/µl
1 0.134 0.086 1340
2 0.029 0.013 42.75
3 0.049 0.034 137.75
4 0.083 0.055 299.25
5 0.091 0.065 337.25
6 0.075 0.055 261.25
7 0.064 0.043 209.00
ABS; absorbencia, ADN; Ácido desoxirribonucleico
Dado que los análisis de ADN se requería una concentración de 100
ng/µl fue necesario realizar diluciones para conseguir tal contenido. Luego, las
muestras de ADN fueron sometidas a la amplificación por el método dereacción en cadena de la polimerasa (PCR), el cual se basó en el uso ciclos de
desenrollameinto y enrollamiento del ADN ocasionados por cambios detemperatura.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 24
La temperatura influyó de manera positiva para amplificar el ADN, ya
que a las a temperaturas muy altas las hebras de ADN sufrieron un proceso de
desnaturalización (rompimiento de la doble hélice), luego al descender la
temperatura se favoreció la fijación de los primers (cebadores) por la
complementariedad de bases y finalmente, la actuación de la Taq polimerasa
(ADN polimerasa de Termophilus aquaticus) para copiar la hebra del ADN quese deseaba amplificar.
Un siguiente paso fue someter el ADN amplificado por PCR fue
someterlos a la digestión con enzimas de restricción. Este proceso se basó en
el hecho de que estas enzimas son nucleasas que tienen sitios específicos de
corte del ADN, dicho de otra manera, reconocen secuencias consenso y cortanen esa posición.
Para establecer la existencia de variantes alélicas los fragmentos de
restricción fueron sometidos a electroforesis en geles de agarosa y
poliacrilamida. El uso de las técnicas electroforéticas en la separación de los
ácidos nucleicos se vió favorecida a la carga negativa neta del ADN que se
debe a la presencia de los grupos fosfato cargados negativamente y que están
orientados hacia el exterior de la molécula. Mientras, que en la separación delas proteínas se requiere en uso de detergentes cargados negativamente,como es el caso del dodecil sulfato de sodio (SDS).
En la Figura 3 se muestra una electroforesis de una placa de ABDRDQ,
que sirvió para poder conocer los alelos de clase I (-A y -B) y clase II (DR y DQ)del Complejo Principal de Histocompatibilidad.
La interpretación para estos alelos fue la siguiente. El los pocillos del 1 al
20 se encontraban los alelos correspondientes a DR, y del pozo 24 al 30 se
localizaban los alelos DQ, del pozo 31 al 52 los alelos de A, los alelos B del
pozo 53 al 95. Una vez que ha ocurrido la separación se tiene en cuenta el
tamaño de la banda y se compara con una hoja de calculo (se anexa copia de
la hoja de cálculo). Además, el análisis estuvo asociado a un programacomputacional (ambos proporcionados por Unimatch-Pel-Freez) a partir delcual se pudo conocer el subtipo de los alelos correspondientes.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 25
(Por citar un ejemplo, el gel de agarosa que se presenta en la Figura 1
fue para las moléculas del CMH clase I y II. En las hojas de cálculo, se
remarcan los subtipos que este presentaron basándose en el amplificado de lasbandas.
Figura 3. Determinación del polimorfismo de algunas variantes de lasmoléculas I y II en el Complejo Principal de Histocompatibilidad enlinfocitos de sangre periférica humana. En esta figura se muestra un gel deagarosa al 2%, en el cual observa a los alelos para A, B, DR y DQ. En la parteizquierda de la figura se muestra el número de los pocillos (1 a 16, 17 a 32, 33 a48, 49 a 64, 65 a 80, 81 a 96). En los pocillos 5, 7, 11, 23, 26, 28, 37, 53, 54, 70,84, 89, 92 se observó una amplificación. Los pocillos 22 y 41 fueronconsiderados como falsos positivos. A continuación se muestra la hoja decalculo, así como la interpretación computacional.
Polimorfismo génico de la Paraoxonasa
El gen de la paraoxonasa es un miembro de una familia multigénica que
involucra a tres genes: PON1, PON2, y PON3 quienes codifican a proteína que
muestran una elevada una homología estructural. Los cuales se encuentran enel cromosoma 7q-21-q22 (13). Se ha determinado que el gen de PON1
presenta dos polimorfismos, uno de ellos se encuentra en el codón 192 y se
caracteriza por la substitución de una glutamina (alelo A) por una arginina (aleloB) (14). Un segundo polimorfismo se ha detectado en el codón 54 y secaracteriza por la substitución de una leucina por una metionina (15).
81
65
49
33
17
1

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 26
Mientras, que PON2 presenta una sustitución de una cisteina por una
serina en el codón 311. La actividad y el polimorfismo de los genes de PON1 y
PON2 se han asociado con el riesgo de enfermedad cardiovascular como la
hipercolesterolemia, la diabetes mellitus no insulino dependiente y la
enfermedad arterial coronaria e infarto al miocardio, así como la enfermedad de
Alzheimer´s y demencia vascular, aunque en ambas familias de genes losresultados son controversiales.
Se ha considerado que la paraoxonasa presenta tres variantes
polimórficas, uno en la posición 192 e involucra la sustitución de una arginina
(R) por glutamina (Q) y otro en la posición 54 e involucra el cambio de una
metionina (M) por leucina (L) y el tercero en la posición 311 (cisteina - serina).En la figura 4 se muestra una electroforesis en agarosa en la que observa la
distribución de fragmentos de restricción respectivos para las variantes génicasdel gen de la paraoxonasa.
Figura 4. Polimorfismo génico de la paroxonasa en linfocitos desangre periférica de sujetos mexicanos. En la figura se muestra ladistribución de fragmentos de restricción en un gel de agarosa al 2%. Elcarril 1 es el marcador de peso molecular, los carriles 2, 7 y 9corresponden a la forma heterocigota (L/M) del gen. En tanto, que loscarriles 3, 5 y 6 corresponden a la forma homocigota (L/L) y el carril 4representan a la variante homocigota (M/M).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 27
En un cultivo de células endoteliales aórticas se ha observado que la
expresión del gen PON3 previene la formación de LDLox e inactiva a las LDLox
preformadas. En contraste a PON1, la PON3 no es activa contra sustratos
sintéticos y fenilacetatos. Además, se ha determinado que la expresión de
PON3 en cultivos celulares de HepG2 no es regulada por fosfolípidos oxidados
ó en hígado de ratón alimentado con una dieta alta en grasas. Estascaracterísticas hacen importante el estudio de estos genes y sus respectivos
polimorfismos. Así, la relación que pueda existir entre ellos, los niveles delípidos y la susceptibilidad a desarrollar enfermedad.
Así mismo, se ha considerado que es necesario establecer una
correlación entre estos genes y otros genes que participan en los procesos
tanto inflamatorios (IL-1b, IL-6, la proteína C reactiva, interferón gamma,moléculas de adhesión) así como los genes que participan en los niveles delípidos.
Lo anterior se basa en los resultados de estudios recientes que han
mostrado que la paraoxonasa (PON), la cual se asocia a las lipoproteínas dealta densidad (HDL) tiene propiedades anti-oxidativas y anti-inflamatorias (16).Estudios in vitro indican que la PON puede disminuir significativamente los
niveles de lípidos peroxidados durante la oxidación de las LDL y de esta
manera asociarse a HDL y dar protección contra la aterosclerosis. Por otro
lado, la actividad serica de la PON parece ser baja en pacientes con infarto almiocardio (17) y con diabetes e hipercolesterolemia familiar (18). La PON se
une fuertemente a subfracciones de HDL que también contienenapolipoproteína (apo) A-I y apo-J (19). La PON ha sido ampliamente estudiada
por su papel en la hidrólisis de compuestos organofosforados incorporados enla formulación de algunos plaguidas (20).
Polimorfismo génico en polipéptidos de bajo peso molecular (LMP) queconstituyen a proteosomas 20S.
Dentro de los genes clase II encontramos los genes DP, DO/DZ, DQ y
DR, cada uno con un par de genes α y β, también los genes TAP1 y TAP2 y los
genes LMP2 y LMP7 (21). Su importancia radica en la participación que tienen
durante el procesamiento y presentación de los antígenos mediados por lasmoléculas clase I del sistema principal de histocompatibilidad.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 28
Los genes del LMP son polimorficos y sus productos constituyen dos
subunidades del complejo proteosomal las cuales se les a atribuido que
participan en la degradación de proteínas citosólicas y en la generación de
péptidos antigénicos. Mientras que las TAP (TAP1 y TAP2) forman un
heterodímero y participan en el transporte de péptidos generados por lasproteasas, desde el citsol hacia el retículo endoplasmico (21).
Los polipéptidos de bajo peso molecular (LMP abreviatura del inglés low
molecular weigth proteasome) se localizan dentro del sistema principal de
histocompatibilidad en la región clase II. Los genes del LMP son polimorficos y
sus productos constituyen dos subunidades del complejo proteosomal las
cuales se les a atribuido que participan en la degradación de proteínascitosolicas y en la generación de péptidos antigénicos (22). Dada su
importancia en éste último evento inmunológico, donde pueden tener un papel
restrictivo muy importante, son considerados candidatos como un factoradicional a la susceptibilidad de daño autoinmune.
El proteosoma o LMP, es una proteasa formada por multisubunidades y
es la maquinaria proteolitica central de la célula, y una de sus muchas
funciones es generar péptidos antigénicos (23). El procesamiento antígenico esmás eficiente, la célula reemplaza algunas de sus subunidades proteosomales
con subunidades mas apropiadas, formando el llamado inmunoproteosoma(23).
Componentes del Proteosoma
Los sitios activos proteoliticos del proteosoma son encerrados en una
cámara enzimática cilíndrica- la subunidad 20S- la cual tiene un complejo
regulador 19S. El proteosoma reconoce la mayoría de los sustratos por la
presencia de una multi-ubiquitina. Una de las funciones reguladoras de 19S es
reconocer las señales de la multi-ubiquitina, para que estas no destruyan lasproteínas intracelulares indiscriminadamente (24-25).
Este complejo de una subunidad 20S y dos subunidades 19S forman un
proteosoma 26S. La subunidad reguladora 19S tiene dos multisubunidades la
base y la tapa . La base, la cual esta compuesta por 6 ATPasas de una triple
familia más dos subunidades no ATPasas (26) que se unen al núcleo catalítico20S.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 29
Las ATPasa tiene una actividad semejante a las chaperonas y se cree
que ayudan al desdoblamiento de los sustratos para unirse con las proteasas(27-28).
Ocho unidades no ATPasas diferentes a la subunidad reguladora 19S
forman la tapa, la función exacta no se conoce. La partícula 20S incluye cuatro
pilas de anillos, cada una conteniendo siete subunidades relacionadasevolutivamente, pero no idénticas (ver Figura 5). Están dentro de dos
categorías, (α y β). Los anillos externos contienen la subunidad α (α1-α7), la
cual forma la compuerta por la cual el sustrato entra y el producto es liberado.
Los anillos internos incluyen la subunidad β (β1-β7), tres de los cuales -β1, β2 y
β5- protegen los seis sitios activos (dos copias de cada uno). El residuo
catalíticamente activo es una treonina localizada en el extremo amino terminal
de las tres subunidades β (29-30) y caracterizan al proteosoma como un
miembro de la familia hidrolasa nucleofilica amino-terminal (NTN). El sitio activo
de las treoninas de la subunidad β es precedido por dos pro-secuencias con
diferente secuencia y longitud.
Durante el ensamblaje del proteosoma 20S, estas pro-secuencias son
removidas por un mecanismo de dos pasos: en el primer paso, los sitios activos
vecinos cortan dentro de las pro-secuencias, mientras que el segundo paso es
autocatalítico, generando el sitio activo de la treonina y un complejo funcional20S (31-32).
El proteosoma cumple estos requerimientos y por lo tanto es
considerada como la principal maquinaria para la generación de péptidos
antígenicos, pero en casos específicos, otras proteasas (como tripeptidil
peptidasa II, furin o thimet oligopeptidasa) puede también contribuir al grupopéptidico clase I (33).

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 30
Figura 5. Modelo de la subunidad 20S del proteosoma. La subunidadestá formado por 4 anillos de 7 unidades cada uno. Los anillos externosformados por subfamilias de tipo alfa y los anillos externos de tipo beta.
Sin embargo, debido a su especificidad de corte selectivo, estasproteasas pueden generar solo un grupo limitado de péptidos que son
apropiados para la unión al SPH, y por lo tanto pueden no ser importantes para
la producción de péptidos antigénicos o sustitutos para la función delproteosoma, aun cuando se expresen juntos.
Tipificación del polimorfismo de los genes LMP2 y LMP7
Con el fin de obtener las secuencias que incluyen los sitios polimórficos
en los genes LMP2 y LMP7, se realizó una PCR utilizando los primers
específicos para LMP2X, LMP2Y,LMP7X y LMP7Y, que fueron sometidos a 30
ciclos de desnaturalización y renaturalización. Las temperaturas paraamplificación de los genes LMP fue diferente.
El polimorfismo de LMP2 se detectó cortando el fragmento amplificado
con la enzima de restricción CfoI, mientras que el polimorfismo de LMP7 sedeterminó cortando el producto amplificado con la enzima BsmI.
En la Figura 6 se muestra el patrón de restricción obtenido para los
genes LMP2 en geles de poliacrilamida al 10%, que se visualizaron en un
transiluminador de luz UV y se compararon el tamaño de los fragmentos con unmarcador de peso molecular estándar.
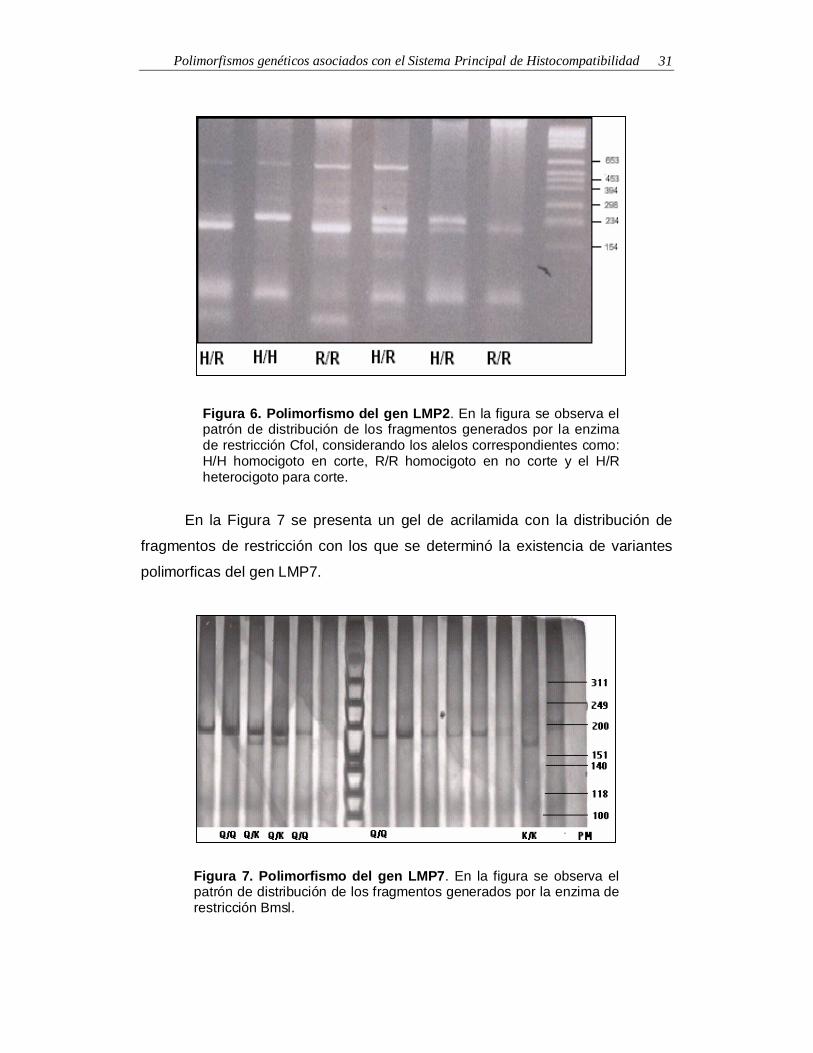
Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 31
Figura 6. Polimorfismo del gen LMP2. En la figura se observa elpatrón de distribución de los fragmentos generados por la enzimade restricción Cfol, considerando los alelos correspondientes como:H/H homocigoto en corte, R/R homocigoto en no corte y el H/Rheterocigoto para corte.
En la Figura 7 se presenta un gel de acrilamida con la distribución de
fragmentos de restricción con los que se determinó la existencia de variantes
polimorficas del gen LMP7.
Figura 7. Polimorfismo del gen LMP7. En la figura se observa elpatrón de distribución de los fragmentos generados por la enzima derestricción Bmsl.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 32
La genética de poblaciones puede ayudar a entender el posible papel en
la diversidad genética. La importancia del estudio de poblaciones para la
interpretación de las causas de los polimorfismos se considera importante. Por
lo que el estudio de poblaciones Amerindias y en particular de las poblaciones
indígenas Mexicanas, las cuales son muy diversas pueden ayudar a entenderla microevolución de los genes clásicos del HLA (34-35).
En la Tabla III se presentan las frecuencias alélicas y genotípicas para
LMP2 y el LMP7 en dos diferentes poblaciones Mexicanas. En todas las
muestras las frecuencias observadas y las frecuencias esperadas están en
equilibrio de Hardy-Weinberg.
El genotipo más frecuente para LMP2 fue el homocigoto en R/R (51.5%
para el grupo de mestizos Mexicanos, 58% en Nahuas, 62.5% en Mazatecos,56.2% en Tennek y 48.7% en Mayos) y el alelo más frecuente fue el R (73.1%
in Mestizos Mexicanos, 65.6% en Nahuas, 62.7% en Mazatecos, 76.7% en
Teenek y 73.9 en Mayos).
Por otro lado, para LMP7 el genotipo Q/Q fue el más común (79.2% en
Mestizos Mexicanos, 82.2% en Nahuas, 94.3% en Mazatecos, 82.0 en Teenek
y 74.6% en Mayos) así como el alelo Q (83.9% en Mestizos Mexicanos
Mestizos, 91.1% en Nahuas, 94.2% en Mazatecans, 91% en Teenek y 87.3%
en Mayos) fueron los más frecuentes encontrados en todas las poblaciones
El genotipo K/K fue ausente ene los Nahuas, Tennek y Mayos, y solo un
individuo Mazateco presento este genotipo. Tanto la población indígena y
mestiza Mexicana presentaron una distribución similar en el polimorfismo deLMP2 y LMP7, excepto en Nahuas y Mayos que presentaron la frecuencia mas
alta de genotipo H/H y la más baja del genotipo H/R (p<0.05), cuando se
comparó con los mestizos mexicanos.
Enfermedades relacionadas con el polimorfismo de los genes LMPDiversos estudios han tratado de establecer la participación de los genes
Clase II del SPH en la susceptibilidad de daño de diversas enfermedades. Los
trabajos realizados hasta la fecha concernientes a la participación de los genes
del LMP en algunas enfermedades son escasos y muy controversiales.
Algunos investigadores han tratado de establecer una asociación entre el
polimorfismo genético del LMP y la susceptibilidad a enfermedades tales como

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 33
la diabetes mellitus insulino-dependiente (DMID) y su relación con otros genesdel HLA (36-37). Deng y colaboradores (1995) (38) informaron una asociación
entre el polimorfismo de LMP7Q/K y DMID en una población Caucásica
(Noruega), la cual explican que no se debió a la unión de desequilibrio con los
alelos DRB1, DQA1 y DQB1.
Mientras, que para el polimorfismo del gen LMP2 H/R encontraron queconfiere una susceptibilidad adicional a DMID solo entre los individuos DR4
positivos, mientras que el polimorfismo del LMP7 confiere una susceptibilidad
independientemente del genotipo de los genes HLA clase II.
Sin embargo, Undilien y colaboradores (1997), no encontraron evidencia
en la susceptibilidad adicional conferida al polimorfismo del gen LMP entre
pacientes con DMID y DRB1, DQA1 y DQB1, incluyendo los subtipos de DR4.También, se han realizado diversos estudios en relación entre el
polimorfismo del LMP y la susceptibilidad a enfermedades autoinmunes como
son las espondiloartropatias y entre ellas la espondilitis anquilosante y la artritisreumatoide. Maksymowich y colaboradores (36-37) estudiaron la relación entre
el LMP2 y la espondilitis anquilosante.
Así como, la Uveitis aguda anterior en pacientes Mestizos Mexicanos,
mostrando que la presencia del alelo LMP2 esta asociado con un decremento
en el riesgo de daño extra-espinal en individuos con un HLA-B27 negativo, en
donde puede también tener una participación de los genes HLA DR2, DR3, y
DR8. Sin embargo, nuevamente estos hallazgos son controversiales con losreportados por Burney y colaboradores (1994) (39) el cual sugiere que esto
puede ser un reflejo de la unión de desequilibrio entre los alelos del LMP2 yHLA-DR4 (40).
Los datos reportados hasta la fecha con respecto al polimorfismo
genético del LMP y la susceptibilidad a ciertas enfermedades, principalmente
de tipo autoinmune son muy heterogénea, lo que demuestra que es necesario
el conocimiento del polimorfismo genético de los genes del HLA entre ellos losdel LMP para poder establecer de manera más contundente la participación de
estos genes en el desarrollo de ciertas enfermedades.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 34
Tabla III . Frecuencias alélicas y genotípicas del LMP2 y LMP7 en lapoblación mexicana (mestizos e indígenas)
Mestizos Nahuas Mazatecos Teenek Mayos
LMP-2 (n=95) (n=48) (n=56) (n=50) (n=63)
N (af) N (af) n (af) n (af) n (af)
H 51 (26.9) 33 (34.4) 44 (37.3) 28 (28) 44 (34.9)
R 139 (73.1) 63 (65.6) 64 (62.7) 72 (72) 82 (65.1)
N (gf) N (gf) n (gf) n (gf) n (gf)
H/H 5 (5) 13 (27)* 5 (8.9) 7 (14) 18 (28.5)*
H/R 41 (43.1)* 7 (14.5) 16 (28.5) 14 (28) 8 (12.6)
R/R 49 (51.5) 28 (58) 35 (62.5) 29 (58) 37 (58.7)
LMP7 (n=95) (n=48) (n=26) (n=50) (n=63)
N (af) N (af) n (af) n (af) n (af)
Q 170 (89.4) 88 (91.6) 49 (94.2) 91 (91) 110 (87.3)
K 20 (10.5) 8 (8.4) 3 (5.8) 9 (9) 16 (12.7)
N (gf) N (gf) n (gf) n (gf) n (gf)
Q/Q 81 (85.2) 40 (83.3) 24 (94.3) 41 (82) 47 (74.6)
Q/K 8 (8.4) 8 (16.6) 1 (3.8) 9 (18) 16 (25.3)**
K/K 6 (6.3) 0 1 (3.8) 0 0
*p<0.05 mayor en Nahuas y Mayos cuando se compararon contra los Mestizos yMazatecos para el genotipo LMP2*H, e incremento en Mestizos cuando secompararoncon Nahuas y Mayos para el genotipo LMP2*H/R.
**P<0.05 aumentado en Mayos cuando se compararon con Mestizos y Mazatecosen el genotipo LMP7*Q/K.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 35
CONCLUSIÓNLa familiarización con las técnicas de biología molecular y la
participación en la estandarización de dichas técnicas con el fin de aplicarlas a
estudios de diversos polimorfismos genéticos. Tales estudios están enfocados
al análisis de frecuencias de dichos alelos en la población tanto mestiza como
indígena de México y la posible relevancia que tiene como marcadores desusceptibilidad para el desarrollo de enfermedades principalmente autoinmunes
y cardiovasculares.
El alumno aprendió metodologías básicas e indispensables de la
biología molecular, como la reacción en cadena de la polimerasa que permite la
amplificación de regiones especificas del material genético en donde se
localizan las variaciones de los genes. La región una vez amplificada, se aplicóla metodología de fragmentos de restricción para la identificación de los
polimorfismos que se consideran como la presencia de un mayor número de
variantes genéticas de las esperadas en una población, por lo que la utilización
de enzimas de restricción que sirven para cortar en sitios específicos de la
secuencia amplificada de ADN. Posteriormente, los fragmentos resultantes de
la digestión se separan en geles de agarosa y acrilamida según el tamaño de
los productos para finalmente asignar cada uno de los alelos.
Otra metodología que el alumno aplicada fue la determinación de los
alelos del sistema principal de histocompatibilidad (SPH) por medio de la
técnica de PCR-SSP (reacción en cadena de la polimerasa-secuencia de
cebadores específicos). En general el alumno adquirió destreza para el manejodel material y el equipo del laboratorio que incluye cámaras de electroforesis,
micropipetas, termocicladores, transiluminador, centrifugas, secadores ADN,
espectrofotómetro y otros, así como la preparación de soluciones que sé
necesitan para este tipo de estudios.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 36
g) CRITERIOS DE EVALUACION
Los criterios de evaluación fueron según el desempeño del alumno dentro
del laboratorio, teniendo en cuenta las siguientes características.
- Limpieza del alumno en el trabajo
- Puntualidad en el trabajo- Las aptitudes al desarrollar el trabajo
- La habilidad para desempeñar las tareas
- En general el desenvolvimiento del alumno dentro del laboratorio y en el
ámbito clínico.
h) BIBLIOGRAFÍA
1. Lewin, B. (1993) GENES VII. 7ª edición. Ed. Reverté. España - pp. 972
2. Gamba, G. (1993) Clonación de Proteínas de membrana y los ovocitos deXenopus Laevis. Ciencia y desarrollo. 19: 40-45
3. Abul K, Andrew H. Jordan S. (1995). Inmunología celular y molecular. 2ª
edición. Mc Graw - Hill - España. Pp.44,79-80, 107-1264. Regueiro, JR. y López, C. (1997) INMUNOLOGIA-Biología y patología del
sistema inmune. 2ª edición. Ed. Panamericana. España. Pp. 9-19,19-27, 41-
55
5. Holloway W, Beghé B, Holgate T. (1999). The genetic basis of atopicasthma. Clin. Exp. Allergy. 29: 1023-1032.
6. Vargas-Alarcón, G. y Granados-Arriola, J. (1993). El Sistema Principal deHistocompatibilidad. Rev. Mex. Reumat. 8: 251-256.
7. Shi, F-D., Ljunggren H-G. y Sarvetnick, N. (2001). Innate immunity andautoimmunity from self-protection to self-destruction. Trends in Immunol. 22:
87-91
9. Rhodes D.A. Trowsdate J. (1999) Genetics and molecular genetics of theMHC. Rev. Immunogenetics.: 1: 21-31
10. Monaco J.J. (1992) A molecular model of MHC class I restricted antigenprocessing. Immunology Today: 5: 173-178
11. Sambrook - Fristsch - Maniatis. Molecular Cloning A Laboratory Manual.
2ª edicion. 1989 New York. Tomo I II y III. pp

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 37
12. Kit de PCR-SSP (reacción en cadena de la polimerasa de secuencia de
primers especificos). Pel-Freez Clinical Systems, Brown Deer, Wisconsin,
USA.
13. Parmo SLP, Sorenson RC, Teiber J, y La Du BN. (1996). The human
serum paraoxonase/arylesterase gene (PON1) is one member of amultigene family. Genomics. 33:498-507.
14. Humbert R, Adler DA, Disteche CM, Hassett C, Furlong CE. (1993). The
molecular basis of the human serum paraoxonase activity polymorphism.Nat. Genet. 3: 73-76.
15. Shangera DK, Aston CE, Saha N, Kamboh M. (1998). DNA
polymorphisms in two paraoxonase genes (PON1 and PON2) areassociated with the risk of coronary heart disease. Am J Hum Genet.,
62:36-44.
16. Watson AD, Berliner JA, Hama SY, LaDu BN, Faull KF, Fogelman AM,
Navab M. (1995). Protective effect of high density lipoprotein associated
paraoxonase. Inhibition of the biological activity of minimally oxidized lowdensity lipoprotein. J Clin Invest., 96:2882-91.
17. McElveen J, Mackness MI, Colley CM, Peard T, Warner S, Walker CH.
(1986). Distribution of paraoxon hydrolytic activity in the serum of patientsafter myocardial infarction. Clin Chem., 32: 671-3.
18. Mackness MI, Abbott C, Durriggton PN. (1991). Paraoxonase preventsaccumulation of lipoperoxides in low-density lipoprotein. FEBS Lett. 29:
152-4.19. Blatter MC, James RW, Messmer S, Barja F, Pometta D. (1993).
Identification of a distinct human high-density lipoprotein subspecies
defined by a lipoprotein-associated protein, K-45. Identity of K-45 withparaoxonase Eur J Biochem. 211:871-9.
20. Davies HG, Rebecca JR, Keifer M, Broomfield CA, Sowalla J, Furlong CE.
(1996) The effect of the human serum paraoxonase polymorphism isreversed with diazoxon, soman and sarin. Nat Genet. 14: 334-6.
21. Savi M, Ferraccioli GF, Neri TM. (1988). HLA-DR antigens and
anticardiolopin antibodies in norther italian systemic lupus erithematosuspatients. Arthritis Rheum. 31: 1568-70

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 38
22. York IA, Goldberg AL, Mo XY, Rock KL. (1990). Proteolysis and class Imajor histocompatibility complex antigen presentation. Inmmunol Rev.172: 49-66.
23. Coux O, Tanaka K, Goldberg AL. (1996). Structure and functions of the20S and 26S proteosomes. Ann. Rev. Biochem. 65: 801-47
24. Rock KL (1994). Inhibitors of the proteosome block the degradation ofmost cell proteins and generation of peptides presented on MHC class Imolecules. Cell, 27:761-771.
25. Glickman MH. (1998). A subcomplex of the proteosome regulatory particle
requiered for ubiquitin-conjugate degradation and related for the COP9-signalosome and elF3. Cell, 94:615-623.
26. Braun BC. (1999). The base of the proteosome regulatory particle exhibitsATP dependent chaperone-like activity. Nature Cell Biol. 1:221-226.
Glickman MH. (1999). Functional analysis of the proteosome regulatoryparticle. Biol. Mol. Reprod. 26:21-28.
27. Groll M. (1995). Structure of the 20S proteosome from yeast at 2.4 A
resolution. Nature 1997; 286:463-471.
28. Fenteany G, et al. Inhibition of the proteosome activity and subunit specificamino terminal modification of the lactacystin. Science 268:726-731.
29. Chen P. y Hochstrasser M. (1996). Autocatalytic subunit proccesing
couples active sities formation in the 20S proteosome to completion ofassembly. Cell 86:961-972.
30. Schmidtke G. (1996). Analysis of proteosome biogenesis: The maduration
of β subunits is an ordered two step mechanism involving autocatalysis.
EMBO J; 15:6887-6898.
31. Schwarz K. (2000). The selective proteosome inhibitors lactacystin and
expoxmycin can be used to either up or down regulate antigenpresentation at non toxic doses. J. Immunol.; 164: 6147-6157.
32. Parham P, Arnett KL, Adams EJ Little AM. (1997). Episodic evolution and
turnover of HLA-B in the indigenous human populations of the Americas.Tissue Antigens 50: 219-23.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 39
33. Belich MP, Madrigal JA, Hildebrand WH, Zemmour J, Williams RC, Luz R,
Petzl Erler ML, Parham P. (1992). Unusual HLA-B alleles in two tribes ofBrazilian Indians. Nature. 327: 326-335.
34. Van Endert PM, Liblau RS, Patel SD, Fugger L, Lopez T, Pociot F, Nerup
J, McDevitt. (1994). Major histocompatibility complex-encoded antigenprocessing gene polymorphism in IDDM. Diabetes; 43:110-117.
35. Undilien DE, Akselsen HE, Joner G, Dahl-Jorgensen K, Sovik O,
RonningenK, Thorsby E. (1997). No independent associations of LMP2and LMP7 polymorphism with susceptibility to develop IDDM. Diabetes 46:
307-312.
36. Maksymowich WP, Jhangri G, Gorodezky C, Luong M, Wong C, Burgos-
Vargas R. (1997). The LMP2 polymorphism is associated withsusceptibility to acute anterior uveitis in HLA-B27 positive juvenile andadult Mexican subjects with ankylosing spondylitis. Ann Rheum Dis., 56:
488-492.
37. Maksymowich WP, Tao S, Vaile J, Suarez-Almazor M, Ramos-Remus C,
Russell A. (2000). LMP2 polymorphism is associated with extraspinal
disease in HLA-B27 negative Caucasian and Mexican mestizo patientswith ankylosing spondylitis. J Rheumatol., 27:183-189.
38. Deng GY, Muir A, Maclaren NK, She JX. (1995). Association of LMP2 and
LMP7 genes within the mayor histocompatibility complex with insulin-
dependent diabetes mellitus: Population and family studies. Am J Hum
Genet.; 56:528-534.
39. Burney RO, Pile KD, Gibson K. (1994). Analysis of the MHC class II
encoded components of HLA class I antigen processing pathwayy inankylosing spondilitis. Ann Rheum Dis.; 53:58-60.
40. Holer T, Schaper T, Sshneider PM. (1997). No primary association
between LMP2 polymorphism and extraspinal manifestations inspondyloarthropathies. Ann Rheum Dis., 56:741-743.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 40
i) APENDICE DE TÉCNICAS
SOLUCIONES PARA LA HIBRIDACIONBuffer de lavado (1L)
50 ml de 20X SSPE concentrado
945 ml de H2O destilada5 ml de docecil sulfato de sodio (SDS) (20%)
Buffer de hibridacion (1L)
100 ml de 20X SSPE concentrado.387.5 de H2O destilada.
12.5 de SDS al 20%Buffer de citratos (600 ml)
30 ml de citratos 10X concentrado.
570 ml de agua destilada.
SOLUCIONES PARA LA EXTRACCION DE DNA
Solución de lisis para eritrocitos (SLR) (1L) (20X)
50ml de Tris 2M =12.12grs.
50ml de MgCl2 1M =10.16grs.
100ml de NaCl 1M = 5.78grs.
Ajustar el pH a 7.6 y aforar a 1L.Para preparar 1L (1X)
50ml de SLR (20X) aforar a 1000ml con H2O.
Buffer de proteinaza K (5X) (100ml)
0.375M de NaCl = 2.19grs
0.12M de EDTA =4.46grsAjustar el pH a 8 y aforar a 100ml con H2O y filtrar
Proteinasa K
Ajustarla a 10 mg/ml con H2O inyectable.

Polimorfismos genéticos asociados con el Sistema Principal de Histocompatibilidad 41
NaCl 5M 14.6 g en 50 ml de agua destilada
Preparación del TBE 10X (1L)
Tris base 54 g
Ácido bórico 27.5 g
0.5M de EDTA (pH=8.0) 20mlAforar con agua desionizada a pH 8.3
GELES DE POLIACRILAMINAAcrilamida al 30% (100 ml)
Acrilamida (grado biología molecular) 29 g
N´N´Methilenebisacrilamida 1 gDisolver en 100 ml de agua destilada
Filtrar en filtros de celulosa de 0.45µm de diámetro
Guardar en botellas ámbar a temperatura ambiente
Persulfato de amonio al 10%
10 grs de Persulfato de amonio en un volumen de 100 ml
Solución fijadora de geles (10-15 min)
Etanol al 10%-----------------------agregar 30 ml de etanol al 96%.Acido Acético al 0.5%----------- agregar 1.5 ml de ácido acético puro.
Aforar a 300 ml con agua destilada.
Solución de tinción (5 min)
AgNO3 0.2%--------- pesar 200mg
Aforar con la solución fijadora a 100ml y disolver.
Solución de revelado (10min)
NaOH (3%) agregar 3 g
Formaldehído concentrado agregar 300µl
Aforar a 100 ml. con agua destilada y disolver.
Acido acético (1.5%) - Para detener la reacción-