
UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAM4274.pdf · ... por ponerme en el...
34
UNIVERSIDAD AUTONOMA METROPOLITANA UNIDAD IZTAPALAPA SERVICIO SOCIAL DINAMICA DE CRECIMIENTO Y COLONIZACION DE DOS ESPECIES DE PASTOS MARINOS DEL CARIBE MEXICANO Halodule writhii Ascher y Svrinaoditcm filiforme Kiitzing mayo 1995.
-
Upload
truongnguyet -
Category
Documents
-
view
214 -
download
0
Transcript of UNIVERSIDAD AUTONOMA METROPOLITANA …148.206.53.84/tesiuami/UAM4274.pdf · ... por ponerme en el...

UNIVERSIDAD AUTONOMA METROPOLITANA UNIDAD IZTAPALAPA
SERVICIO SOCIAL
DINAMICA DE CRECIMIENTO Y COLONIZACION DE DOS ESPECIES DE PASTOS MARINOS DEL
CARIBE MEXICANO
Halodule writhii Ascher y Svrinaoditcm filiforme Kiitzing
mayo 1995.

NOMBRE: María Aimé Rodriguez Román.
MATRICULA: 88341408
LICENCIATURA: Hidrobiología Ciencias Biológicas y de la Salud Unidad Iztapalapa
TITULO DEL TRABAJO: "Dinámica de crecimiento y colonización de dos especies de pastos marinos del Caribe Mexicano: Halodule wrigthii (Ascher) y Syringodium filvorme (Kiitzing)" .
ASESOR EXTERNO: Dr. Martin Merino Ibarra. Investigador Titular 'IC" de Tiempo Completo Instituto de Ciencias del Mar y Limnología U N A M .
Profesor Titular " A l de Tiempo Completo U A " I
ASESOR INTERNO: M. en C. Abel Sentíes Granados.
LUGAR DE REALIZACIóN: Instituto de Ciencias del Mar y Limnología U N A M .
FECHA DE INICIO: 29 de Octubre de 1993
FECHA DE TERMINACI~N: 29 de Abril de 1994 (25 Mayo de 1995)
CLAVE: H. 036.93
NOMBRE DEL PROYECTO: "Recolonización por pastos marinos en zonas perturbadas del Caribe Mexicano y Mediterraneo Español." Y el proyecto "Importancia de la disponibilidad de nutrientes en angiospermas marinas del Caribe Mexicano" Proyecto CONACyT 361 8-T-UNAM.

INSTITUTO DE CIENCIAS DEL M A R Y LIMNOLOGIA
APARTADO POSTAL 7 0 - 3 0 5 MEXICO 04510. LL F. M E X I C O
UNIVERSIDAD NACIONAL AUTONOMA DE
M E X I C O México, D.F., 25 de Mayo de 1995.
M. en C. ROSAURA GRET.HER GONZÁLEZ, DIRECTORA DE LA DIVISIÓN DE CIENCIAS BIOLóGICAS Y DE LA SALUD. UAM -1ZTAPALAPA.
Por este conducto me permito informarle que la alumna María Aimé Rodriguez Román, con número de matricula 88341408 de la Licenciatura en Hidrobiología, realizó un trabajo de investigación exhaustivo, consistente en: análisis de datos, búsqueda Bibliográfica, viajes al Caribe Mexicano para la investigación en campo y pruebas en el Laboratorio del Instituto de Ciencias del Mar y Limnología, razón por la cual, la presentación de su Informe de
~, Servicio Social, denominado: "Dinámica de crecimiento y colonización de
_-..
.T ) dos especies de pastos marinos del Caribe Mexicano Sirinqodium I - filiforme Kützing y Halodule wriathii Ascher" se hace en esta fecha,
Agradezco la atención que se sirva brindar a la presente.
A t e n t a m e n t e .
DR. MARTiN MERINO IBARRA. Investigador Titular.

..
INSTITUTO DE CIENCIAS DEL M A R Y LiMNOLOGlA
APARTAOO POSTAL 70-30s MEXICO 04510. a F. M E X I C O
UNIVERSIDAD NACIONAL AUTONOMA D E
M E X l C O
México, D.F., 25 de Mayo de 1995.
M. en C. ROSAURA GRETHER GONZÁLEZ, DIRECTORA DE LA DIVISIóN DE CIENCIAS BlOLÓGlCAS Y DE LA SALUD. UAM -1ZTAPALAPA.
Por este conducto me dirijo a usted, para comunicarle que la alumna María Aimé Rodriguez Román, con número de matrícula 88341408 de la Licenciatura en Hidrobiología, terminó satisfactoriamente con su Servicio Social denominado "Dinámica de crecimiento y colonizaci6n de dos especies de pastos marinos del Caribe Mexicano Sirinnodium filiforme Kützing y Halodule wriathii Ascher", el cual tuvo una duración del 29 de octubre de 1993 al 25 de mayo de 1995.
Sin otro particular, me despido quedando de usted,
A t e n t a m e n t e .
DR. MARTiN MrRINO IBARRA. Investigador Titular.

Casa abierta al tiempo
UNIVERSIDAD AUTONOMA METROPOLITANA DNISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD SERVICIO SOCIAL
Junio 1, 1995 SS.CBS.369.95 H.036.93
LIC. JULIO DE LARA ISASSl Coordinador de Sistemas Escolares P r e s e n t e
Por medio de la presente se hace constar que el alumno cuyos datos se describen a continuación, concluyó su Servicio Social:
NOMBRE: RODRIGUEZ ROMAN MARIA AlME
MATRICULA: 8834 1408
LICENCIATURA: HlDROBlOLOGlA
PROYECTO: Dinámica de crecimiento y colonización de dos especies de pastos marinos del Caribe Mexicano: Halodule wrightii (Ascher) y Syringodium filiforme (Kützing).
Se extiende la presente para los fines que I interesado convengan, al primer día del mes de junio de mil novecientos noventa y cinco.
A T E N T A M E N T E "CASA ABIERTA AL TIEMPO"
/
DIRECTORA
I *LPC
UNIDAD IZTAPALAPA
Av. Michoacán y La Purísima Irtapalapa 09340 México, D.F. A.P. 55-535 Fax: (5) 612-80-83 Tels 77.: '5 8 1 'y 85

AGRADECIMIENTOS.
Al Dr. Martin Merino Ibarra y al M. en C. Abel Sentíes Granados, por su excelente asesoría, por invertir parte de su tiempo en la revisión del presente trabajo, por sus consejos y por su incondicional amistad.
Al Biól. Marco Aurelio Pe rk H. por su ayuda en el manejo estadístico de 10s datos, por su paciencia y por su valiosa amistad.
A la Dra. Margarita E. Gallegos Martinez, por ponerme en el camino de la botánica acuática, dentro de un grupo de investigación realmente valioso, y por muchas cosas más.
Al Biól. Francisco Contreras E, del laboratorio de Ecosistemas Costeros de la UAM-1, por permitir la elaboración de este informe en dicho laboratorio, por su ayuda y consejos.
Al Biól. Felipe Flores A. jefe de la estación de Puerto Morelos del ICMyL, por facilitar las instalaciones para la realización del trabajo de campo.

DINÁMICA DE CRECIMIENTO Y COLONIZACIÓN DE DOS ESPECIES DE PASTOS MARINOS
DEL CARIBE MEXICANO: Halodule wrightii (ASCHERS) Y
Syringodiumfiliforme (KÜTZING).
Los pastos marinos son plantas vasculares que se encuentran en las aguas costeras del mundo entre los círculos Artico y Antártico, formando extensas praderas. Son monocotiledóneas y estan ubicados en dos familias: Hydrocharitaceae (3 géneros, 14 especies) y Potamogetonaceae (9 géneros, 34 especies) (Phillips & Meñez, 1988); perteneciendo a ésta última las dos especies del presente estudio. Forman ecosistemas extremadamente complejos que desempeñan dos papeles en las zonas costeras: uno trófico y otro estructural (Duarte, 1989), siendo el primero el que contribuye directamente con producción primaria al ecosistema, así como el aporte a la cadena del detritus; y el segundo con múltiples funciones: estabilizan y atrapan sedimentos aún bajo el enorme estres de huracanes y tormentas (Zieman, 1974); reducen la velocidad de la corriente promoviendo la sedimentación y reduciendo la resuspensión (Fonseca et al., 1982); sirven de guarida y refugio para residentes y transeúntes de adultos y juveniles de especies de importancia económica como: Peneidos, Pomadasydos, Lutjanidos, Centropominos, entre otros (De la Lanza y Tovilla, 1986); son importantes sustratos para la fijación de epibiontes y muestran notable eficiencia en el reciclamiento de nutrientes de las aguas marinas y sedimentos (Dawes, 1986). Las comunidades de pastos sufren fluctuaciones en el área que cubren, por alteraciones debidas a fenómenos naturales como tormentas (Thomas et al.), al impacto de las actividades humanas y a la mortalidad ocasional (Short et al., 1988; Williams, 1988; Orth & Moore, 1983). Siendo de gran importancia la recuperación de las praderas, que depende de la reproducción sexual y de las tasas de crecimiento horizontal (Duarte & Sand-Jensen, 1990). Las comunidades de pastos marinos presentan un claro patrón sucesional donde, las especies colonizadoras tienen gran importancia. El patrón de actividad del meristem0 terminal determina las diferencias entre los patrones de crecimiento vegetativo para las

especies de pastos marinos. De acuerdo con las características fisiológicas la "dependencia del meristemo" determina que especie será colonizadora y cual clímax, ya que este determina cuando la ramificación y producción de haces nuevos es proliferativa o meramente regenerativa (Tomilson, 1974). El Único género que presenta un patrón diferente es Syringodium en donde los meristemos laterales se desarrollan como haces, es un tipo de proliferación ramificada, pero donde esta no es ordenada por el haz mismo, sino por la perturbación del ambiente (Phillip & Meñez op cit).
En las camas de pasto del Caribe Mexicano, existe una dominancia de Thalassia
testudinum, seguida en orden de importancia, por macroalgas, Syringodium fillforme y Halodule wrigtii. Sin embargo la tasa de crecimiento horizontal de Thalassia es de 2 a 4 veces menor que la de las especies colonizadoras (Gallegos et al., 1993), por lo que la recolonización le tomaría muchos años, resaltando así, la hnción de Halodule wrightii y Syringodium fillforme ya que estas atrapan sedimento propiciando un ambiente óptimo para el crecimiento de los parches de Thalassia testudinum.
México cuenta con extensas zonas de pastos marinos, las cuales se han estudiado escasamente, es probable que el grado de deterioro ecológico de origen antropogénico sea mayor en su avance, que el conocimiento de estos (De la Lanza y Tovillas op cit.). Debido a esto es necesario el conocimiento de este recurso que permita un uso y manejo racional. Con este estudio se pretende estimar el crecimiento y productividad en escalas de tiempo (anuales, mesuales y en términos del intervalo de plastocrono), mediante observaciones más sencillas y de menor frecuencia que las utilizadas hasta ahora. Así como su capacidad de recuperación ante las fluctuaciones, la cual depende de la formación de nuevos parches y del crecimiento de los mismos, así como de las tasas de expansión de los ya existentes. Con esto se pretende contribuir al conocimiento de estos sistemas, y en particular de los procesos de recolonización de estas dos especies del Caribe Mexicano.
CARACTER~STICAS Y DISTRIBUCI~N DE LAS ESPECIES.
Halodule wrightii.
Presenta internodos de rizoma de 0.75 a 3.5 cm de largo, cada nodo contiene entre 2 y 5
raíces y un haz. La vaina de la hoja tiene un largo de 1.5 a 4 cm y la hoja de 3.5 a 32 cm, con un ancho de 0.3 a 2.2 m m . La flor macho presenta un pedúnculo de 12.5 a 23 mm de largo y antenas de 3 .5 a 5 mm. Los ovarios de la flor hembra son elipsoides con 1.5 a 2 mm de largo, pistilo de 10 a 28 m m ; el fruto es ovoide globoso de 1.5- 2 mm (Fig 1). Esta especie se distribuye ampliamente más abajo de la zona intermareal y por arriba de la zona submareal en sustratos arenosos y fangosos, en localidades cubiertas, así como
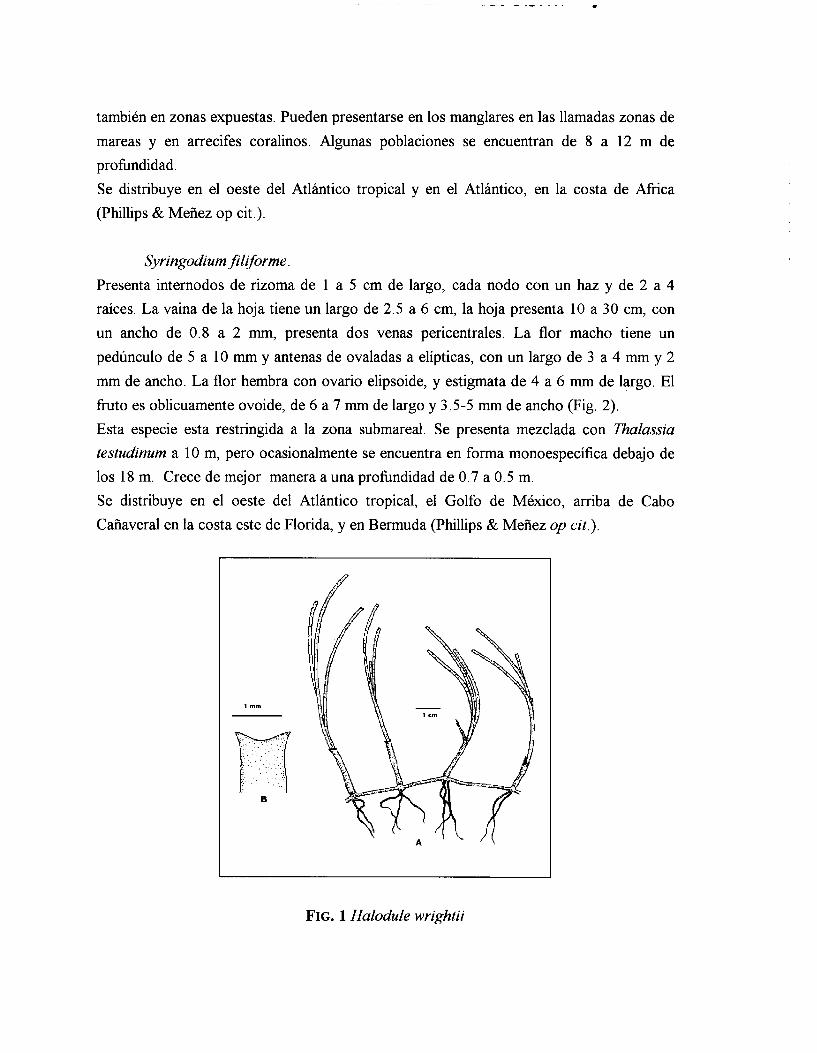
también en zonas expuestas. Pueden presentarse en los manglares en las llamadas zonas de mareas y en arrecifes coralinos. Algunas poblaciones se encuentran de 8 a 12 m de profundidad. Se distribuye en el oeste del Atlántico tropical y en el Atlántico, en la costa de Africa (Phillips & Meiiez op cit.).
Syringodium filforme .
Presenta internodos de rizoma de 1 a 5 cm de largo, cada nodo con un haz y de 2 a 4
raíces. La vaina de la hoja tiene un largo de 2.5 a 6 cm, la hoja presenta 10 a 30 cm, con un ancho de 0.8 a 2 mm, presenta dos venas pericentrales. La flor macho tiene un pedúnculo de 5 a 10 mm y antenas de ovaladas a elípticas, con un largo de 3 a 4 mm y 2 mm de ancho. La flor hembra con ovario elipsoide, y estigmata de 4 a 6 mm de largo. El fruto es oblicuamente ovoide, de 6 a 7 mm de largo y 3.5-5 mm de ancho (Fig. 2). Esta especie esta restringida a la zona submareal. Se presenta mezclada con Thalassia
testudinum a 10 m, pero ocasionalmente se encuentra en forma monoespecífica debajo de los 18 m. Crece de mejor manera a una profimdidad de 0.7 a 0.5 m. Se distribuye en el oeste del Atlántico tropical, el Golfo de México, arriba de Cabo Cañaveral en la costa este de Florida, y en Bermuda (Phillips & Meiiez op cit.).
1 mm
.,. . . . . . . . . . . . , . . . .. . _. . . .
B
~~~
FIG. 1 Halodule wrightii

FIG 2. Syringodium fillforme
ANTECEDENTES.
Existen pocos trabajos en el Caribe Mexicano y específicamente en la laguna arrecifal, apesar de la importancia de sus comunidades bióticas. Se pueden citar aquellos referentes a moluscos de Aguirre (1988), Ekdale (1974) y Espejel (1983); de macrofauna bentónica Arriaga (1 985); Gómez y Green (1 984) trabajo sobre esponjas; Garcia-Saenz et al (1 987) de peces arrecifales; siendo la mayor contribución los relacionados con los arrecifes coralinos entre estos encontramos: Jordan (1 979a), Jordan (1 979b), Jordan (1 980), Jordan y Nuget (1979, Jordan et al ( 1981), Jordan et al (1985), Iglesias (1986), Merino et al (1980) y Moreno et al (1982) entre otros. Las contribuciones en lo que respecta a vegetación son aun menores, encontrando para macroalgas dos trabajos León y Jordan (1 978) y Taylor (1 93 5). En pastos marinos el de Nuget et al (1978) y los recientemente realizados Gallegos et al. (1992), Gallegos et al. (1 993), Marbá et al. (1 993), Gallagos et al., (1 994). Y por último un trabajo reciente para la especie Syrigodiumfrllforme, realizado en la laguna arrecifal de Pto. Morelos de Van Tussenbroek (1 994).
.

OBJETIVO GENERAL.
Determinar la dinámica de crecimiento y la producción en términos de biomasa de Halodule wrightii y Syringodium filgorme, con miras a caracterizar el origen de su capacidad de colonización de sedimentos libres.
OBJETIVOS PARTICULARES.
- Estimar las tasas de crecimiento de rizoma, haces y hojas de ambas especies.
- Estimar la producción en términos de biomasa de hojas, rizoma y raíces.
- Comparar las tasas de crecimiento y producción de ambas especies y determinar la importancia de cada una en el proceso de colonización.
- Realizar una comparación e integración de resultados entre los dos métodos utilizados, conteo de cicatrices y marcaje.
- Estimar la producción de rizomas y raíces, y su distribución por profbndidad, de las dos especies y compararlas con la producción de la especie climax Thalassia testudinum.
AREA DE ESTUDIO.
El presente trabajo se realizó en la laguna arrecifal cercana al pueblo de Puerto Morelos, a 20 km al sur de Cancún (entre 20"51' y los 20" 52' de latitud norte y 86" 54' longitud oeste) (Fig. 3).
La laguna arrecifal esta comprendida entre la línea de costa y la barrera arrecifal, esta depresión alcanza una anchura que varía entre 350 y 1600 m. Hacia el sur termina donde la barrera arrecifal también acaba, primero formando bancos aislados para después desaparecer por completo. Hacia el norte termina a la altura de Punta Tanchacté, donde el arrecife y la costa se acercan hasta una distancia de aproximadamente 150 m y con profbndidad escaza de menos de un metro, estrangulándose la laguna en este lugar. La mayor parte del fondo de la laguna arrecifal se encuentra cubierto por densas praderas de pastos marinos, dominadas por Thalassia testudinum. En alguna zonas se localizan fondos arenosos con escasa presencia de pastos marinos y algas coralinas.

La zona presenta influencia de diferentes masas de aire, que determinan en gran medida el clima de la región. La mayor parte del tiempo (entre febrero y julio) se encuenta bajo la influencia de las masas de aire marítimo tropical transportadas por los vientos alisios del Caribe y el Atlántico. Posteriormente sigue una época de transición (de julio a septiembre). En invierno (de octubre a enero) la influencia es de los llamados "nortes", que aunque llegan a Yucatan y Quinta Roo bastante modificados, influyen negativamente sobre la meteorología, asociados con frentes fríos se presentan nublados y chubascos. La región presenta un clima de tipo Aw 1 (x')(i')g. Clima cálido con temperatura media mayor a los 22" C, y la del mes más fiío mayor a los 18" C; clima subhúmedo, con lluvias en verano, precipitación del mes más seco menor a 60 mm y un porcentaje de lluvia invernal entre 5 y 10.2 de la anual. Corresponde a un tipo intermedio de los subhúmedos con un cociente precipitaciódtemperatura de entre 43.2 y 55.3. En este caso la precipitación invernal es mayor al 10.2 por ciento de la anual. Con poca oscilación térmica, de entre 5 y 7" C. Temperatura tipo Ganges, presentándose el mes más caliente del año antes de junio (Merino y Otero, 1991).
Factores Físicos. Corrientes. Existen pocos estudios relacionados con corrientes en esta zona, pero se sabe que las velocidades oscilan alrededor de los 10 c d s , y en algunos casos cercanas a 50
c d s , presentándose estas en la boca o aberturas que se forman en la barrera arrecifal o entre ésta y la playa. La dirección dominante de las mismas es de norte-noroeste y sur- suroeste, es decir, en dirección paralela a la costa, esto es probablemente debido a la topografia del sitio como a la dirección de los vientos dominates. Oleaje. La barrera arrecifal hnciona como un eficiente disipador de la energía del oleaje, por lo que se ve hertemente transformado por su paso a través de la misma, observaciones de 1983 indican una altura media de oleaje de 14 cm en la zona de la playa frente a la estación de la UNAM, y un período de éste de aproximadamente 2 segundos. Marea. No existen datos para determinar cual es el comportamiento de la marea en la zona, y se mencionan los registros tomados en la isla de Cozumel, y datos frente a Pto. Morelos. En ambos casos el régimen de marea es mixto y semidiurno, presentándose dos pleamares y dos bajamares durante el día. Los valores medios de pleamares y bajamares presentan un valor absoluto de 10 cm, por lo que es de esperarse en el área un rango de oscilación diurna promedio de alrededor de 20 cm. Temperatura. Este parámetro es uno de los más medidos y por lo que sus valores resultan más confiables. La temperatura media es de 27.74 "C, oscilando entre los 24.96 y

los 30.52 "C. Las temperaturas más bajas se registran durante la madrugada y la temperatura máxima en zonas someras durante el verano. Salinidad. El valor medio es de 35.72 O/oo, y su oscilación es de 34.92-36.51 O/oo. La zona presenta un caracter océanico, dominado hidrológicamente por las aguas oceánicas adyacentes. Oxígeno disuelto. El valor promedio es de 4.99 ml/l, superior al porciento de saturación para la temperatura y salinidad del área, el mínimo es de 3.53 y el máximo de 6.45 ml/l. Los valores mínimos registrados se han presentado durante la madrugada (Merino y Otero, 199 1).
4
I
FIG. 3 Area de Estudio.

METODOLOGÍA.
Se estudiaron praderas monoespecíficas de Halodule wrightii y Syringodim jilrforme en la laguna arrecifal, durante agosto de 1991, el tiempo de máxima biomasa en la zona (Van Tussenbrok, 1994) y durante la primavera de 1992 y 1993. El muestre0 se realizó con equipo autónomo de buceo (SCUBA). Las muestra heron tomadas con un nucleador de acero inoxidable de 20.5 cm de diámetro. Se colectaron 6 y
3 núcleos de Syringodium jilrforme y Halodule wrightii respectivamente. El nucleador h e enterrado hasta 40 cm dentro del sedimento para colectar los haces desde su inserción en el rizoma, lo cual es necesario para la estimación de edades (Gallegos et al., 1993). Las muestras heron lavadas, separándolas totalmente de los sedimentos carbonatados. Se realizó un conteo de haces vivos determinando su edad (número de cicatrices más hojas vivas presentes) según la técnica propuesta por Patriquin (1973), determinación de haces muertos datándolos (contando el número de cicatrices, de solo aquellos que terminaban en punta redondeada, indicando que el haz no se había roto) según lo propuesto por Gallegos et al (1993). Las unidades de edad se obtuvieron en intervalos de plastocrono (PIS) las cuales representan el promedio del intervalo de tiempo entre la iniciación de dos hojas sucesivas en un haz (Erickson & Michelini, 1957; Brouns 1985). Los internodos verticales de los haces heron medidos con un microscopio de disección, con ocular calibrado con reglilla de 20 p, para examinar el curso del tiempo en los haces más viejos de la muestra, identificando ciclos anuales en la longitud de los internodos verticales (Duarte et al., 1994; Marbá et al., 1994) el inverso del número de internodos en un ciclo (número de hojas al año) corresponde al promedio anual de PI (Duarte et al. 1994; Marbá et al., 1994) el cual fue utilizado para convertir los PIS a unidades de tiempo absolutas. En muestreos siguientes se obtuvieron datos de frecuencia de floración, edades máximas de haces y secuencias adicionales de longitudes internodales de haces viejos. El tamaño de muestra de los núcleos h e de 2000 y 4000 haces para Syringodium jilrforme y Halodule
wrightii respectivamente. El PI de rizoma h e calculado por regresión del número de internodos de rizoma entre haces a lo largo del rizoma contra sus diferencias de edad (Patriquin, 1973; Gallegos et al.,
1993). La tasa de crecimiento vertical h e calculado por una regresión de la longitud del haz contra edad (Gallegos et al., 1993) y por medio del marcaje contando el número de rizomas y la longitud en un tiempo conocido. La producción de raíces insertadas al rizoma presentes en cada nodo, se obtuvo midiendo la longitud total y contando el número de raíces presentes, de vivas y muertas. Este cálculo se puede relacionar con la densidad de haces y rizoma, ya que la relación 1 : 1 de :

un internodo de rizoma por cada haz producido, se puede complementar con las raíces, teniendo una relación 1 : 1 :2, para las dos especies. Se obtuvo el promedio de raíces vivas y raíces muertas. Para la producción de raíces en haces se obtuvieron 2 núcleos de ambas especies, midiendo edad de haces, por conteo de cicatrices más hojas vivas presentes y el número de raíces vivas y muertas (aquellas con punta necrozada), longitud de raices, peso húmedo y peso seco. Se aplicó una regresión lineal simple, entre el número de raíces totales (suma de raíces vivas, muertas y rotas) y la edad. La estructura de edades h e descrita utilizando datos en haces vivos y muertos (Gallegos et al., 1993; Duarte et al., 1994). El promedio de edad de haces vivos representa el total de la edad de la población, pero esta sobrestima la esperanza de vidad, la cual esta mejor representada por las edades de los haces muertos (Gallegos et al., 1993; Duarte et al.,
1994). La mortalidad de haces (k en unidades de Ldaño) (Duarte & Sand-Jensen, 1990b) f ie calculado desde la distribución de edades de haces muertos (Gallegos et al., 1993; Duarte et al., 1994) con la siguiente ecuación:
Nt = No e -kt donde No= número de haces con edad igual a la moda; y Nt= número de haces mayores que la clase modal por el tiempo (t). El muetreo de marcaje se realizó durante las siguientes visitas a la zona de estudio, para el cual se localizaron puntas de crecimiento en las orillas de los parches, de Halodule
wrightii y Syringodium jilgorme se marcaron 33 rizomas en tres parche, desenterrando el rizoma y marcandolo con una etiqueta númerada en el tercer haz, se registró la longitud de la marca hasta la punta de crecimiento. La recolecta se realizó después de 6-8 días, la recuperación de rizomas marcados h e de 10 YO para Halodule wrightii y 39.9% para Syringodium Jilgorme. Se midiendo longitud final, número de internodos, número de haces y hojas nuevas, estos datos heron obtenidos en PIS y en escalas de tiempo absolutas. Tambien se realizó la colecta de rizomas, para complementar los datos de número de hojas nuevas, midiendo la longitud y número de internodos entre haces de diferente edad. Los datos de PI de hoja y tasa de elongación de rizoma, heron comparados con los obtenidos mediante el método de reconstrucción (Duarte et al., 1994). Por último se colectaron cinco núcleos en praderas donde las tres especies estuvieran presentes, para determinar la producción subterránea de estas, se realizaron cortes de los
núcleos cada 3 cm, separando raíces y rizomas de H. wrightii, S. jilgorme y T. testudinum.
Se realizó un análisis de varianza entre núcleos, para raíces y rizomas, no encontrando diferencias significativas entre cada núcleo, pudiendo obtener un promedio para la

comparación entre las tres especies y se realizó el análisis de varianza entre las tres especie. Se obtuvo una submuestra de cada componente registrando peso húmedo, longitud y peso seco, determinando la biomasa de rizoma y raíz por prohndidad.
RESULTADOS
Halodule wrightii y Syringodium filrforme, desarrollan abundantes praderas monoespecíficas en la laguna arrecifal, registrando una alta densidad de haces y una alta biomasa de haces y rizomas (Tabla 1). Los internodos de rizoma de S. filrforme heron mas grandes que los de H. wrightii, teniendo una longitud promedio de 30.6 * 0.25mm/internodo y forma un densa red de 228,5 m de rizomd m2, y los de H. wrightii de 20.2 * 1 .O mndinternodo, y formando una red 2 16.8 m de rizomdm2 respectivamente. La densidad de haces h e relacionada con la densidad de internodos de rizoma (Tabla l), porque ambas especies producen un haz por cada internodo de rizoma. El número de internodos entre los haces conectados y su diferencia de edades h e estrechamente corelacionado (p<O.OOOl). La pendiente de la ecuación de regresión, indica que H. wrightii y S. jilrforme producen 1.81 * O. 17 y 2.81 * 0.27 internodosP1, respectivamente.
Variable
Biomasa de haces (gPS/m2) Densidad de haces (hacedm2) Biomasa de rizoma (gPS/m2) Densidad de rizoma (rizamdm2) Intervalo de plastocrono de hoja (d) Hojas al año Elongación de rizoma (cm/año/ápice) Crecimiento vertical (cm/año/haz) Densidad de raíces de nodos (raícedm2) Biomasa de raíces de nodos (gPS/m2) Raíces de haces (raíces/año/haz) Reclutamiento de haces (In unidadedaño) Tasa de mortalidad de haces (In unidadedaño) Producción de hoja (grPS/m2/año) Producción de rizoma (grPS/m2/año)
S. jilrforme
446.9 2 48.4 7139 21366
23 3 7468 60.8
6 51.6 3.36
14278 89.38 4.44 0.77
2.04 2 0.39 2234 466
H. wrightii
519 5 86 14872i2444
97 10732 16.5 22.1 80.9 2.65
22962 81.86 17.017 4.18
3.54 0.37 4619 3 20

El crecimiento vertical h e calculado por análisis de regresión entre la longitud de haces y la edad del haz, fue de 0.56 A 0.13 y 0.12 0.01 cm/PI para S. filIfome y H. wrightii,
respectivamente (Fig. 4). 32
4
4
b
b
I
I -
-
l - .d
L 2
I I I
b
+ +/
I
I -
-
l - .d
L 2 I I
4 8 16 Edad de Haces [PI]
I
4 8 16 Edad de Haces [PI]
16.D , I I
- +
0.1 2 I I
2 4 8 16 32 E d a d de Haz [PI]
I I
FIG. 4. Relación entre la longitud del rizoma vertical y la edad del haz. Para Syringodium
fillforme y Halodule wrightii.
Los distintos patrones en la secuencia de las longitudes de internodos verticales (Fig. 5) se obtuvo el cálculo del número promedio de internodos verticales (= núm de hojas)
2 2 2 7 7 6

producidas por un haz por año, y el promedio de PI para las 2 especies (Tabla 1) los cuales heron utilizados para calcular las tasas de elongación de rizoma de las tasas por PI (Tabla 1). Las ramificaciones del rizoma en 3.2 % y 9.3% de los internodos examinados para S. fillforme y H. wrightii respectivamente.
15 I I I I I
CI
E E "
I
h n - -
c
-
o
i . . ' l b c
On I I I I I
3 6 9 12 15 18 Hasta el presente [PI]
I I I I I I I
01 I I I I I I I O 10 20 30 40 50 60 70
Hasta el presente [PI]
FIG. 5 Syringodiumfillforme y Halodule wrightii. Secuencias de internodos registradas de los haces más viejos. Se indican los ciclos anuales.
Las raíces insertadas en cada nodo heron de 1 a 2 en número, para H. wrightii, teniendo un porcentaje de 45.83% de nodos con una sola raíz y 54.16% con dos raíces. El promedio de raíces vivas y muertas fue de 1.36 .76 y 0.27 f O. 53. Teniendo la relación de un internodo de rizoma por cada haz producido y las proporciones de número de raíces se considera un produción de 22962 raíces/m2. La longitud promedio h e de 12.2 6.96
I

cm. Teniendo una biomasa de raíces vivas de 8 1.86 gr DWlm2. Y para S. fillforme, las raíces totales heron de 2 5 0.27, teniendo un promedio de raíces vivas y muertas de 0.69 5
0.76 y 0.45 5 0.63, la mayor proporción encontrada f ie de raíces rotas, por lo que los datos pueden estar subestimados. La producción de raíces es de 14278 raíceslm2. La longitud promedio f ie de 25.1 1 cm. Teniendo una biomasa de raíces vivas de 89.38 gr DWlm2 (Tabla 1). Para la producción de raices en haces se calculó una regresión lineal simple entre el número de raices totales contra la edad del haz, teniendo una producción de 0.77 raices/PI con un coeficiente de correlación de ,978, para H. wrightii, teniendo así una producción de 17.017 raíceslaño. Y de 0.74 raíces/PI con un coeficiente de correlación de 0,7157 (Fig. 6) para S. filiforme, con 4.44 raíceslaño. Los haces "mas viejos" analizados en la producción de raices correspondian a 2.48 años para H. wrightii y de 2.8 para S. fillforme.
I
2 2 2 7 7 6

m
m m
o 3 6 9 12 15 18
Edad de haces
FIG. 6 Relación de raíces totales contra edad. H. wrightii y S. fillforme
El promedio de edad de haces de S. fillforme f ie de 1 año (6.7 5 O. 1 PIS) y sobretodo de haces que han sido producidos en el año en curso (1993) (i.e. edad<7 PIS; Fig. 8)
indicativo de una alta tasa de reclutamiento (Tabla 1). La distribución de edad de haces (Fig. 8) presenta un claro declive exponencial en la sobrevivencia con incrementos de edad que dejaron calcular las tasas de mortalidad (Tabla 1).
1 3 5 7 9 11 13 15 17 19 Edad de haces [PI]

60
51 m
=I u u u t 34
17
1 3 5 7 10 12 16 18 Edad de haces (PI]
FIG. 8. Distribución de edades de haces vivos y muertos de Syringodiumfilvorme.
El promedio de la esperanza de vida de haces de S. fillforme h e cerca de medio año (3 2
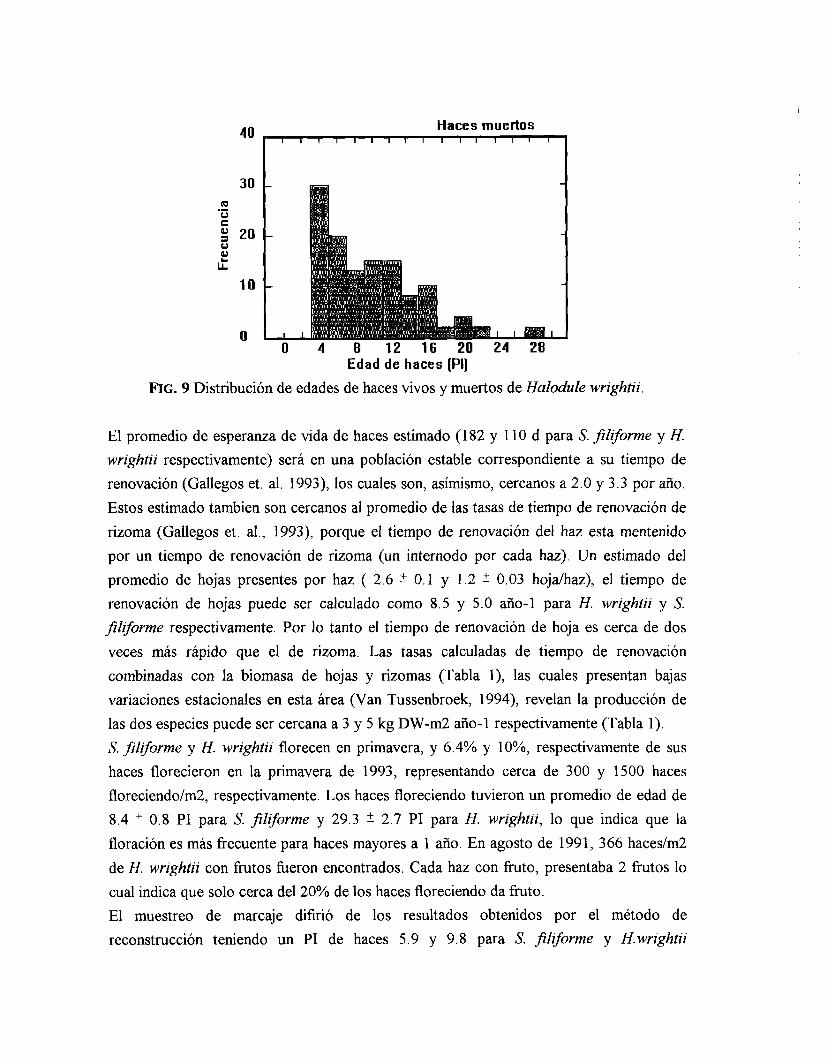
O. 16 PIS) y los haces más viejos encontrados heron de cerca de 7 años (46 PIS, n>2000). Casi todos los haces vivos de H. wrightii heron producidos durante el mismo año (Fig. 9) indicando tasas de reclutamiento muy altas (Tabla 1). Las altas tasas son necesarias compensarlas con las altas tasas de mortalidad (Tabla l), reflejado por el rápido declive exponencial en la abundancia de haces con “edades mayores” (Fig. 9). La esperanza de vida de haces de H. wrightii es de solo tres meses 6.9 0.4 PIS, y los haces más viejos vivos heron de solo 3 años (70 PIS n>4000).
Haces vivos
2 6 10 14 18 22 26 Edad de Haces [PI]
40 l"--l Haces muertos
30 IR u C
.- 2 20
10
u
LL E
0 O 4 8 1 2 16 20 24 28
Edad de haces [PI)
FIG. 9 Distribución de edades de haces vivos y muertos de Halodule wrightii.
El promedio de esperanza de vida de haces estimado (182 y 1 10 d para S. fillforme y H. wrightii respectivamente) será en una población estable correspondiente a su tiempo de renovación (Gallegos et. al. 1993), los cuales son, asimismo, cercanos a 2.0 y 3.3 por año. Estos estimado tambien son cercanos al promedio de las tasas de tiempo de renovación de rizoma (Gallegos et. al., 1993), porque el tiempo de renovación del haz esta mentenido por un tiempo de renovación de rizoma (un internodo por cada haz). Un estimado del promedio de hojas presentes por haz ( 2.6 0.1 y 1.2 0.03 hojdhaz), el tiempo de renovación de hojas puede ser calculado como 8.5 y 5 .O año- 1 para H. wrightii y S. fillfurme respectivamente. Por lo tanto el tiempo de renovación de hoja es cerca de dos veces más rápido que el de rizoma. Las tasas calculadas de tiempo de renovación combinadas con la biomasa de hojas y rizomas (Tabla l), las cuales presentan bajas variaciones estacionales en esta área (Van Tussenbroek, 1994), revelan la producción de las dos especies puede ser cercana a 3 y 5 kg DW-m2 año-1 respectivamente (Tabla 1). S. filiforme y H. wrightii florecen en primavera, y 6.4% y lo%, respectivamente de sus
haces florecieron en la primavera de 1993, representando cerca de 300 y 1500 haces floreciendo/m2, respectivamente. Los haces floreciendo tuvieron un promedio de edad de 8.4 5 0.8 PI para S. fillforme y 29.3 5 2.7 PI para H. wrightii, lo que indica que la floración es más frecuente para haces mayores a 1 año. En agosto de 1991, 366 haces/m2 de H. wrightii con frutos heron encontrados. Cada haz con fruto, presentaba 2 frutos lo cual indica que solo cerca del 20% de los haces floreciendo da fruto. El muestre0 de marcaje difirió de los resultados obtenidos por el método de reconstrucción teniendo un PI de haces 5.9 y 9.8 para S. fillforme y H.wrightii

respectivamente. Y el intervalo de plastocrono de hoja (d) de 6 1.8 y 37.01
respectivamente (Tabla 2). Las tasas de elongación de rizoma heron de 196 d a ñ o para S. fillforme y de 292 cmlaño para H. wrightii ,
TABLA 2. Comparación entre los dos métodos utilizados.
Variable Reconstrucción1 Marcaje Reconstrucción1 Marcaje S. fillforme H. wrightii
Intervalo de plastocrono de hoja (d)
401.5 80.9 197.1 51.6 (cmlañolápice) Tasa de elongación de rizoma
37.01 22.1 5.9 6 Hojas / año 9.8 16.5 61.8 60.8
En la producción de material subterráneo el análisis de varianza entre las tres especies para rizoma y raiz h e significativo (nivel de significancia 0.0 12 y de .O00 1 para rizoma y raíz, respectivamente). El análisis de rangos múltiples (Tabla 3 y 4) indicó que existian diferencias significativas para H. wrightii y T. testudinum, y para S. fillforme y T. testudinum; pero no significativas para H. wrightii y S. fillforme.
TABLA 3. Análisis de rangos múltiples para variaciones de raíz entre las tres especies. 1 Nivel IN 1 Promedio 1 Gruuos homogéneos 1
2
X 1.6623077 13 3
X ,2261385 13 1
X ,0649769 13 2
X 1.6623077 13 3
X ,2261385 13 1
X ,0649769 13
contraste
0.72514 * -1.5973 2-3
0.72514 * -1.43617 1-3
0.72514 0.16116 1-2
limites diferencia + I -

. . S.
* diferencias significativas 1 Halodule wrightii 2 Syringodium fillforme 3 Thalassia testudinum .
La mayor biomasa para H. wrightii en rizoma f ie en los dos primeros cortes de tres cm de 13.3 1 gr/m2 y de 6,62 gr/m2, llegando a encontrarse a una profundidad máxima de 15- 18 cm con una biomasa de 0.21 gr/m2. S. filiforme esta especie se encuentra verticalmente distribuida en cuanto a su rizoma solo en los primeros 9 cm teniendo su máximo en la capa de 0-3 cm, con una biomasa de 8.77 gr/m2. T. testudinum la especie climax tiene una distribución marcadamente diferente teniendo mayores biomasas de rizoma 3-18 cm encontrando su máximo de 6 a 9 cm de profbndidad, con una biomasa de 42.51 gr/m2, llegando a encontrarse hasta de 27 a 30 cm con una biomasa de 0.44 gr/m2 (Fig. 10).
I 1
I O 10 20 30 40
O 5 21-24 24-27
30-33 3x36 3639
g 27-30:
I I FIG. 10 Biomasa de rizoma por prohndidad de las tres especies.
2 2 2 7 7 6

Mientras que la raíz la mayor biomasa para H. wrightii se encuentra de O a 6 cm con 2.18 y 2.64 gr/m2, encontrándose hasta los 33 cm de profundidad. Para S. filiforme la raíz presenta una distribución semejante a la de H. wrightii, de O a 9 cm, con biomasas de 2.53, 2.57 y 2.05 gr/m2, presentando el máximo en la capa de 3 a 6 cm y llegando hasta 30-33 cm de profundidad con una biomasa de 0.03 gr/m2. Y por último para T. testudinum presenta sus mayores distribuciones de 6 a 21 cm encontrando el máximo en el intervalo de 9 a 12 cm, con una biomasa de 24.38 gr/m2 y se llega a encontrar hasta los 39 cm de profundidad, posiblemente la raíz se presente en capas más profundas, pero el muestreados cuenta con una profundidad de 40 cm (Fig. 11).
grlm2 O 5 10 15 20 25 I
Halodule wrightii 24-27
0 Syringodium filfiforme I -- 27-30 +
~ m 30-33
33-36 36-39 f FIG. 11 Biomasa de raíz por profundidad de las tres especies.
DISCUSI~N En base a los resultados obtenidos, ambas especies desarrollan praderas altamente productivas, en el Caribe Mexicano. Las densidades de haces fueron mayores a las reportadas anteriormente por Eleuterius (1 987) para el Golfo de México, las cuales van de 300 a 3200 haces/m2 para H. wrightii y de 150 a 1600 haces/m2 para S. fillforme;
difiriendo al igual con los resultados obtenidos por Short et al. (1993) para S. filiforme
(1563 2 1263 haces/m2) y los de Pangallo y Bell (1988) para H. wrightii (1275 a 4000
haces/m2). El número de haces por unidad de área es dificil de obtener, debido a que existen variaciones considerables entre praderas y en las praderas mismas (Eleuterius op. cit.) sin embargo, en trabajos que reportan ambas especies, la densidad de H. wrightii
siempre es superior a la de S. fillforme, que puede deberse al rápido crecimiento vegetativo de H. wrightii. Y se puede considerar que la región del Caribe Mexicano es según datos reportados, la más densa, para ambas especies.

Las tasas de crecimiento de rizoma para Syringodium fillforme son semejantes a las encontradas en el Oeste de Oceáno Atlántico de 0.5 d a ñ o (Short et al., 1993). Mientras que la obtenida por Eluterius (1987) de 134.6 cdaño es similar a la obtenida en este trabajo por el método de marcaje. Por otro lado, para H. wrightii, la tasa de elongación de rizoma de 80.9 (cdañolápice), es menor a la obtenida por el método de marcaje (Tabla 2) y que la reportada por Eleuterius (1987). El crecimiento en pastos presenta variaciones estacionales por lo que las tasas de elongación obtenidas por el método de marcaje nos reflejan la velocidad de crecimiento para una época precisa, no siendo correcto la extrapolación a un año completo. Realizando una comparación con Thalassia testudinum,
la especie clímax, se observa que S. filrforme y H. wrightii crecen de 2 a 4 veces más rápido, que esta (22.3 cdaño) (Gallegos et al., 1993). En términos de biomasa los resultados obtenidos, al igual que la densidad, son mayores a los reportados en los trabajos anteriores (Eleuterius, 1987; Short et al., 1993), los resultados obtenido, pueden ser diferentes por el método de muestre y el tamaño de muestra, no obstante existe un valor mayor en biomasa de S. fillforme para la misma zona de estudio en el Caribe Mexicano, de 1 KgDW/m2 (Van Tussenbroek, 1994). En suma a sus altas tasas, S. filrforme y H. wrightii se ramifican cada 92 y 21 .8 cm, mientras que X testudinum presenta una ramificación cada 6300 cm (Gallegos et al. unpublished). La recuperación de praderas depende de el crecimiento horizontal y de la formación de nuevos parches, por tanto la capacidad de floración de las especies colonizadoras es de suma importancia, encontrando así que la densidad de floración por haz, para ambas especies, es de cerca de 10 a 50 veces mayor que la de T. testudinum,
contribuyendo así a la colonización de sedimentos libres más rápido. Las altas tasas de crecimiento asociadas con las altas tasas de mortalidad de S. JUrforme y H. wrightii (Tabla I), implican que son especies pioneras, o bien, facilitadoras de acuerdo con el modelo propuesto por Connel & Slatyer (1977), así como lo sugerido por Williams (1990), con requerimientos mayores de nutrientes (Fourqurean et al., 1992; Short et al.,
1993; Duarte, 1994). La rápida perdida de material de S. fillforme y H. wrightii
representadas por las grandes cantidades de material depositado en las costas, asociado con la corta vida de las hojas y haces, reduce su capacidad para el almacenamiento de nutrientes y el reciclamiento interno. En contraste T. testudinum crece más despacio y el tiempo de vida de las hojas es mayor (Gallegos et. al., 1993) y de acuerdo a esto tiene requerimientos menores de nutrientes (Fourqurean et. al., 1992; Duarte, 1994) y pueden ser más capaces de almacenar y reciclar los nutrientes (Patriquin, 1972). El papel que juega S. filrforme es intermedio entre H. wrightii y 7: testudinum. debido a sus tasas de crecimiento. Teniendo así las etapas de sucesión: macroalgas-H. wrightii-S.

fillforme-T. testudinum. Las tres especies se pueden encontrar mezcladas, sin embargo la dominancia de T. testudznum, es evidente, tanto en biomasa de haces y hojas, como en biomasa subterránea. Williams (1987) propone que la competencia por recursos, entre T. testudinum y S. filflorme, es por luz, y la producción de S. fillforme, disminuye, con la presencia de la sombra producida por 7: testudinum. Tomasko (1992) da evidencias de que H. wrightii varia la forma de crecimiento, en competencia con T. testudinum.
Teniendo así, en praderas mezcladas, H. wrightzi, en los primeros centímetros de profundidad, con sus dos componentes rizomas y raíces, S. jillforme tambien en los primeros centímetros con rizoma, pero sus raíces se llegan a encontrar más profundas que en el caso anterior. Y T. tetudinum, con rizomas hasta los treinta centímetros de profundidad y sus raíces a los 40 cm. Por otro lado cabe destacar que la utilización de dos métodos diferentes, h e complementaria para el caso de S. fillforme, mientras que para H. wrightii no hubo concordancia alguna. El método de reconstrucción del intervalo de plastocrono (PI)
utilizando determinación de edades está sujeto a diferentes fuentes de incertidumbre por lo que se recomienda, el uso simultáneo del método de marcaje. Sin embargo, Duarte et. al. (1994), demuestran que la estimación de la media anual de producción de hojas por haz obtenidas por reconstrucción, están estrechamente correlacionadas a las observadas, por técnica de marcaje. Se puede pensar que para el caso de H. wrightzi, el problema de las discrepancias entre ambos métodos, pudo deberse a las deficiencias en la técnica de marcaje, esta especie presenta hojas muy pequeñas y de poco grosor, donde las marcas resultan fácilmente confusas, así como de la perdida total de rizomas marcados, teniendo una recuperación menor al 10%. Aunado a los inconvenientes de la técnica de solo reflejar tasas de crecimiento de una época del año (Mayo), cuando se realizó el muestreo. El método de reconstrucción, propuesto aquí, tiene la ventaja de que además de obtener tasas de crecimiento y biomasa, que se pueden obtener por marcaje, nos permite realizar medidas poblacionales, tales como estructura de edades, tasas de mortalidad, tasas de reclutamiento y esperanza de vida.
CONCLUSIONES. Ambas especies, son altamente productivas en el Caribe Mexicano, y juegan un papel primordial en la colonización de sedimentos libres, facilitando de esta manera, la entrada de la especie clímax, 7: testudinum. Syringodium filqorme, por sus tasas de crecimiento de rizoma, haces y hojas, es intermediario en el proceso sucecional, entre H. wrightii y T. testudinum.

El método de reconstrucción del intervalo de plastocrono, es más eficiente. Ya que requiere menor esfberzo de muestre0 y por tanto, menor presupuesto de investigación, pudiéndo obtenerse mayor información.
RECOMENDACIONES.
Se recomienda, investigar métodos alternativos de marcaje, para ambas especies, en el caso de H. wrzghtiz, el pasto es muy frágil, y el peso de la marca, debe ser menor, para que no se produzca daño y pérdidas del material. Para S. filforme, las hojas no debe ser pinchadas, ya que se origina la perdida de la hoja o bien su muerte. México, cuenta con extensas praderas de pastos marinos, las cuales no han sido estudiadas, la técnica de reconstrucción del intervalo de plastocrono, es una alternativa para el conocimiento de éstas. Teniendo posibilidades de en poco tiempo, conocer el estado de diferentes praderas y realizar comparaciones, en todo el litoral del Golfo de México.
ACTIVIDADES REALIZADAS.
Durante el Servicio Social, se desarrollaron las siguientes actividades: Búsqueda de información bibliográfica. Análisis de base de datos, de los muestreos anteriores realizados por el grupo de trabajo, este incluye datos obtenidos en agosto de 199 1, mayo de 1992, y mayo de 1993, Publicación de artículo en la revista Murine Ecology Progress Series.
Muestre0 en campo: en marzo y agosto de 1994. Trabajo de laboratorio, realizado en la estación de Puerto Morelos, Quinta Roo, del Instituto de Ciencias del Mar y Limnología U N A M . Y trabajo de laboratorio realizado en ICLyM UNAM, México, D.F. Procesamiento estadístico de datos. Análisis y Discusión de resultados. Elaboración del informe final. Así mismo, existe la colaboración en investigaciones realizadas en la especie Thalassia
testudinum tanto en dinámica poblacional, limitación por nutrientes (análisis de nutrientes en agua intersticial y contenido de hierro en hojas) y medidas de fotosíntesis (curva P-I fotosíntesis irradianza y medidas de fluorescencia).

LITERATURA CITADA.
Aguirre, B.L.A., 1988. Estructura espacio - temporal de la malaco-fauna bentónica del arrecife de Puerto Morelos, Quintana Roo, México. Tesis de Maestría. CINVESTAV- IPN, Unidad Mérida
73 PP.
Arriaga, R., 1985. Estudio preliminar de la macrofauna de invertebrados de las playas arenosas de Quintana Roo y Yucatán (México) y su relación con el sedimento. Tesis de Licenciatura, Facultad de Ciencias, U N A M .
Brouns, J.J.W.M., 1985a. The plastochrone interval method for the study of productivity of seagrasses; possibilities and limitations. Aquat. Bot. 21: 71-88.
Brouns, J.J.W.M., 1985b. A comparison of the annual production and biomass in the monospecific seagrass beds of Thalassia hemprii (Ehrenb.) Aschers. Aquat. Bot. 23: 149- 175.
Connell, J.H. & Slatyer, R.O., 1977. Mechanims of succession in natural communities and their role un community stability and organisation. American Naturalist 1 1 1 : 1 1 19- 1 144.
Dawes, C.J., 1986. Botánica Marina. Limusa. México.
De la Lanza, E. G. y C. Tovilla H., 1986. Una revisión sobre taxonomía y distribución de pastos marinos. Universidad y Ciencia 3(6): 17-38.
Duarte, C.M., 1989. Temporal biomass variability and productionhiomass relationships of seagrass comunities. Mar. Ecol. Prog. Ser. 51: 269-276.

Duarte, C.M., Marbá, N,, Agawin, N,, Cebrián, J., Enríquez, S., Fortes, M.D., Gallegos, M.E., Merino, M., Olesen, B., Sand-Jensen, K., Uri, J. & Vermaat, J., 1994. Reconstruction of seagrass dynamics: age determinations and associated tools for the seagrass ecologist. Mar. Ecol. Prog. Ser. (in press).
Duarte, C.M. & K. Sand-Jensen, 1990. Seagrass colonization: patch formation and patch growth in Cymodocea nodosa. Mar. Ecol. Prog. Ser. 65: 193-200.
Ekdale, A.A., 1974. Marine molluscs from shallow-water environments (O a 60 meters) off the Northeast Yucatan Coast., Mexico. Bull. Mar. Sci. 24: 638-668.
Eleuterius, L. N,, 1987. Seagrass ecology along the coast of Alabama, Louisiana, and Mississippi. In: Durako, M. J., Phillips, R.C., Lewis, R. R. 111 (eds) Proc. Symp. Subtropical-Tropical Seagrasses Southeaster United States. Flarnar. Res. Publ. 42: 1 1-20.
Erickson, R.O., Michelini, F.J., 1957. The plastocrhon index. Am. J. Bot. 44: 297-305.
Espejel, J.M., 1983. Análisis de la distribución y abundancia del género Cyphoma ( Gasteropoda: Orulidae) en el arrecife de Puerto Morelos Quintana Roo. Tesis de Licenciatura. Facultad de Ciencias, U N A M .
Fonseca, M.S., Fisher, J.J., Zieman, J.C., Thayer, G.W., 1982. Influence of the seagrass, Zostera marina L., on current flow. Estuar. Coast ShelfSci. 15:
351-364.
Fourqurean, J.W., Zieman, J.C., Powell, G.V.N., 1992. Relationship between porewater nutrient and seagrasses in a subtropical carbonate environment, Mar. Biol. 1 14: 57-65.

Gallegos, M. E., M. Merino, N. Marbá & C.M. Duarte, 1992. Flowering of Thalasia testudinum Banks ex Konig in the Mexican Caribbean: age- dependence and interannual varability. Aquat. Bot. 43: 249-255.
Gallegos, M.E., M. Merino, N. Marbá y C.M. Duarte, 1993. Biomass and dynamics of Thalassia testudinum in the Mexican Caribbean: elucidating rhizome growth. Mar Ecol. Prog. Ser. 95: 185-192.
Garcia - Saenz, C. y Espejel, J. J., 1987. Feeding preferences of Cyphoma gzbbosum (Linnaeus, 1758) on different zones of a reef. In: IX Int, Malacol. Congress. Edimburgo, Escocia.
Gómez, L.P. y G. Green, 1984. Sistemática de las esponjas marinas de Puerto Morelos, Quintana Roo, México. An. Inst. Cienc. Mar. y Limnol. Univ. Nul. Autón. México. l l(1): 65-90
Iglesias, R.P., 1986. Mecanismos de la fotoadaptación en gorgonáceos. Tesis de Maestría. UACPyP del CCH, UNAM, México.
Jordan, E.D., 1979a. Estructura y composición de arrecifes coralinos en la región noreste de la Península de Yucatán, México. An. Centr. Cienc. Mar. y Limnol. Univ. Nul. Autón. México. 6 (1): 69- 86.
Jordan, E.D., 1979b. An analysis of a Gorgonian community in a reef calcareus plataform on the Caribbean coast of Mexico. An. Centr. Cienc. Mar. y Limnol. Univ. Nul. Autón. México. 9(1): 87-
96.
Jordan, E.D., 1980. Arrecifes coralinos del noreste de la Península de Yucatán: estructura comunitaria, un estimador del desarrollo arrecifal. Tesis Doctoral, UACPyP del CCH, U N A M , México.

Jordan, E.D. y R. S. Nuget, 1978. Evaluación poblacional de Plexaura homomalla (Esper) en la costa noreste de la Península de Yucatán. Air. Centr. Cienc. Mar. y Limnol. Univ. Nal. Autón. México. 5(1): 189-200.
Jordan, E.D., M. Merino, O. Moreno y E. Martin, 198 l . Community structure of coral reefs in the Mexican Caribbean. In: Proceedings of the IV International Coral Reef Symposium, Manila. Vol. 2.
Jordan, E.D., O. Moreno, E. Martin, P. Thome y M. Merino, 1985. Informe final de la tercera etapa del Proyecto "Ecologia y Evaluación de Poblaciones de Plexaura homomalla y su explotación racional con el fin de preservar el recurso". Marzo, 1985, presentado a CONACyT.
León, H. y E. Jordan D., 1978. Asociaciones de macroalgas del arrecife posterior de Puerto Morelos, Quintana Roo. VI Congreso Nacional de Oceanografla. Ensenah, B. C., México.
Marbá, N,, Gallegos, M.E., Merino, M. y Duarte, C.M., 1994. Vertical growth of Thalassia testudinum: seasonal and interannual variability. Aquat. Bot. 47: 1-11.
Merino. M., E. Martin, O. Moreno y E. Jordan, 1980. El estudio del ambiente marino de Quintana Roo. In: Simposio Quintana Roo, Problemática y Perspectivas. Ed. por el Centro de Investigaciones de Quintana Roo y el Instituto de Geografla de la UN&.
Merino, M., y L. Otero D., 1991. Atlas Ambiental Costero, Puerto Morelos - Quintana Roo. Centro de Investigaciones de Quintana Roo (CIQRO). Fondo de Publicaciones y Ediciones. 80 p.
Moreno, O., Merino, M. y E. Jordan, 1982. Estimación de la edad en un organismo colonial: Plexaura hornomalla (Esper) (Octocorallia: Plexauridae). An. Inst. Cienc. del Mar. y Limnol., Univ. Nal. Aut6.n. México., 9(1): 345-358.

Nuget, R.B., E. Jordan y R. de la Torre, 1978. Investigaciones preliminares de la biomasa de Thalassia testudinum (Konig) en la costa del Caribe Mexicano. An. Centr. Cienc. Mar. y Limnol. Univ. Nul. Autón. México 5(1): 247-254.
Orth, R. J., Moore, K.A., 1983. Chesapeake Bay: an unprecedent decline in submerged aquatic vegetation. Sczence 222:
51-53.
Pangallo, R. A. & Bell, S., 1988. Dynamics of the aboveground and belowground structure of the seagrass Halodule wrightii. Mar. Ecol. Prog. Ser. 43 : 297-301.
Patriquin, D., 1972. The origen of nitrogen and phosphorus for growth of the marine angiosperm Thalassia testudinum Konig. Mar. Biol. 15:35-46.
Patriquin, D., 1973. Estimation fo growth rate, production and age of the marine angiosperm Thalassza testudinum Koning. Caribb. J. Sci. 13: 11 1-123.
Phillips, R.C. and E.G., Meñez, 1988. Seagrasses. Smithsonian Institution Press, Washington, D.C. 103 pp.
Short, F.T., Ibelings, B.W., Den Hartog, C., 1988. Comparison of a current eelgrass disease to the wasting disease inthe 1930s. Aquat. Bot. 30: 295-304.
Short, F. T., Montgomery, J., Zimmermann, C.F., Shor, C.A., 1993. Production and nutrient dynamics of a Syringodiumfilrforme Kutz. seagrass bed in Indian River Lagoon, Florida. Estuaries 16:323-334.
Taylor, W.R., 1935. Marine algae from the Yucatan Peninsula. Botany of the Maya Area; Vol. Publications of the Carnegie Institution of Washington 41 6: 1 1 5 - 124.

Thayer, G.W., Wolfe, D.A., Williams, R.B., 1975. The impact of man on seagrass systems. Am. Scient. 63: 288-296.
Thomas, L.P., Moore, D.R., Work, R.L., 1961. Effects of hurricane Donna on the turtle-grass beds of Biscayne Bay Florida. Bull. Mar. Sci. 11: 191-197.
Tomasko, D. A,, 1992. Variation in growth form of shoal grass (Halodule wrightii) due to changes in the spectral composition of ligth below a canopy of turtle grass (Thalassia testudinum). Estuaries I5
(2): 214-217.
Tomilson, P. B., 1974. Vegetative morphology and meristem dependence - the foundation of productivity in seagrasses. Aquaculture 4: 107- 130.
Van Tussenbroek, B., 1994. Spatial and seasonal variability in biomass and leaf morphology of the manatee grass, Syringodiumfllqorme in a tropical coral reef lagoon, Mexico. An. Inst. Cienc. del Mar y Limnol. WM. Mexico. (in press).
Williams, S., 1988. Disturbance and recovery of a deep-water Caribbean seagrass bed. Mar. Ecol. Prog. Ser. 42: 63-71.
Zieman, J.C.,1974. Methods for the study of the growth and production of the turtle grass, Thalassia testudinum Koning. Aquaculture 4: 13 9- 143.


















