
UNIVERSIDAD NACIONAL AGRARIA LA MOLINA · La maricultura de estos pectínidos tiene carácter de...
33
UNIVERSIDAD NACIONAL AGRARIA LA MOLINA FACULTAD DE PESQUERIA PROYECTO DE TESIS: “TASA DE CONSUMO DE OXÍGENO Y ACLARAMIENTO DE LA CONCHA DE ABANICO Argopecten purpuratus EN RELACIÓN AL TAMAÑO EN DIFERENTES ZONAS DE LA BAHÍA DE SECHURA” EJECUTORA: Pamela Evelyn Cabezas Huatuco ASESOR: Dr. Jaime Mendo La Molina, Lima- Perú 2013
Transcript of UNIVERSIDAD NACIONAL AGRARIA LA MOLINA · La maricultura de estos pectínidos tiene carácter de...

UNIVERSIDAD NACIONAL AGRARIA LA MOLINA
FACULTAD DE PESQUERIA
PROYECTO DE TESIS:
“TASA DE CONSUMO DE OXÍGENO Y ACLARAMIENTO DE LA
CONCHA DE ABANICO Argopecten purpuratus EN RELACIÓN AL
TAMAÑO EN DIFERENTES ZONAS DE LA BAHÍA DE SECHURA”
EJECUTORA:
Pamela Evelyn Cabezas Huatuco
ASESOR:
Dr. Jaime Mendo
La Molina, Lima- Perú 2013

I. INTRODUCCIÓN
En el Perú, la producción de conchas de abanico se ha incrementado aceleradamente en las
últimas décadas debido a su gran demanda en los mercados internacionales. (Mendo et al.
2008). La maricultura de estos pectínidos tiene carácter de exportación, destinándose los
productos preponderantemente en estado congelado. (Mincetur, 2008).
Una de las zonas que sustenta las exportaciones de concha de abanico en el país es la Bahía
de Sechura en la Región Piura cuya producción se basa principalmente en el cultivo de
fondo (Gonzales y Yépez 2007). Esta región ha tenido un crecimiento significativo en los
últimos años por las exportaciones, representando un 82% de la producción de concha de
abanico en el Perú, según informó la directora Regional de la Producción al diario el Correo
en el 2012. Sin embargo esta actividad ha significado una siembra masiva sin sustento
técnico que podría poner en riesgo su sostenibilidad. (Mendo, 2011).
Es conocido que la sobrecarga de las zonas de cultivo puede alterar el equilibrio natural de
la Bahía en aspectos como calidad química de aguas y fondo, equilibrios sedimentarios y
condiciones físicas que deriven en una pérdida de la productividad de la Bahía (Boynton et
al. 1980, Dame et al. 1980). Por esto, la importancia de la determinación de la capacidad de
carga que necesita el conocimiento de los procesos fisiológicos como son la tasa de
respiración y filtración para entender las relaciones del organismo con el medio.
En este contexto, la determinación de los requerimientos energéticos, ecología y fisiología
de la concha de abanico Argopecten purupuratus in situ es importante para conocer los

flujos energéticos que pueden ser usados en la estimación de la capacidad de carga en
términos productivos y ecológicos (Phillipson 1975). La tasa de respiración y la tasa de
aclaramiento son dos parámetros importantes y necesarios para contribuir a la
determinación de la capacidad de carga y se sabe que estos a su vez están muy relacionados
al tamaño de los individuos y a diferentes parámetros ambientales (Flores 2012 y Duarte
2003).
Es por ello que en esta investigación se pretende estimar las tasas de respiración y
aclaramiento de la concha de abanico considerando además de la talla, diferentes habitas
que representan las áreas más comunes usadas en el repoblamiento en la Bahía de Sechura.

II. PROBLEMA DE INVESTIGACIÓN
El éxito de un cultivo de la concha de abanico Argopecten purpuratus está relacionado a la
disponibilidad de alimento y al oxígeno disponible del medio ya que ello condiciona la
densidad de siembra. Se conoce que existe un mejor crecimiento de la concha de abanico a
densidades bajas y por lo tanto mayor disponibilidad de oxígeno y alimento.
La Bahía de Sechura presenta una gran producción de conchas de abanico que ha
conducido a un incremento de las áreas de repoblamiento y al aumento de las densidades de
siembra, presentando en algunas zonas densidades de hasta 200 individuos por metro
cuadrado. (com. Pers. Luis Ysla). La disponibilidad de oxígeno y alimento en esta bahía
puede ser afectada por cambios ambientales del medio, así como por el consumo tanto de
los organismos cultivados como por el metabolismo microbiano en la degradación de las
biodeposiciones, esto indica que la cantidad de oxígeno disuelto del medio podría ser en
algún momento insuficiente para soportar la demanda energética de crecimiento de estos
organismos.
Por otro lado, no todas las áreas de repoblamiento dentro de la bahía presentan
características iguales en crecimiento, que si bien es cierto puede explicarse por factores
genéticos también puede ser resultado de factores exógenos como son la temperatura,
salinidad, oxígenos disuelto, etc propios de cada área. (Sicard et al, 1999).
Las condiciones ambientales de determinadas épocas del año producen alteraciones bio-
oceanográficas que en diversos grados afectan principalmente los cultivos de concha de
abanico. Gonzáles et al. (2012) concluyen que las zonas de impacto no son siempre

las mismas y que obedecería a condiciones locales en la que se generaría modificaciones
en el sistema de circulación a microescala, permanencia de eventos de floración algal, entre
otros factores.
Aún no se han realizado investigaciones sobre crecimiento en diferentes zonas para poder
determinar si existen variaciones dentro de la bahía, así como tampoco se han realizado
investigaciones in situ que determinen las tasas fisiológicas como las tasas de consumo de
oxígeno y filtración que presentan los organismos en diferentes áreas de repoblamiento de
la bahía y estaciones del año. La mayoría de los trabajos relacionados con la alimentación y
estudios fisiológicos de pectínidos se realizan en condiciones controladas que no
necesariamente reflejan el hábitat natural de la especie

III. OBJETIVOS DE LA INVESTIGACIÓN
3.1. Objetivo general
Determinar y comparar las tasas de consumo de oxígeno y aclaramiento en relación
al tamaño de la Argopecten purpuratus en diferentes áreas de repoblamiento y en dos
estaciones del año.
3.2.Objetivos específicos
Determinar la relación entre la tasa de consumo de oxígeno de Argopecten
purpuratus en diferentes zonas de la bahía de Sechura.
Determinar la tasa de aclaramiento en relación al tamaño de Argopecten purpuratus
en diferentes zonas de la bahía de Sechura.
Comparar las tasas de consumo de oxígeno y aclaramiento durante verano e
invierno.

7
IV. JUSTIFICACIÓN DE LA INVESTIGACIÓN
La producción de conchas de abanico se ha incrementado en los últimos años, siendo esta
actividad de suma importancia en términos socioeconómicos. Ante esto, es necesaria la
construcción de modelos que pueden ser conectados a medidas ecológicas de capacidad de
carga para lograr que el cultivo no cause un impacto significativo al ambiente y así el
cultivo sea sostenible. Según Phillipson (1975), una de las formas de asegurar la
explotación racional de un recurso consiste en conocer el flujo energético que atraviesa en
el sistema. Desde este punto de vista ecológico el registro de los principales procesos
fisiológicos en los organismos es un paso indispensable para entender las relaciones del
organismo con el medio. Uno de estos procesos fisiológicos es la respiración que está
estrechamente relacionada a la actividad metabólica del individuo, ya que de la energía
transformada por la respiración muchas funciones metabólicas son posibles.
La tasa de consumo de oxígeno es una variable importante a incorporar al momento de
realizar el modelo ecológico. Este modelo requiere de datos confiables de la variación
estacional de la tasa de respiración (George 1997) y al igual que la tasa de aclaramiento esta
se encuentra influenciada por parámetros ambientales principalmente temperatura y
salinidad. (Smaal y Widdows, 1994)
En este sentido el conocimiento de las necesidades energéticas de los individuos y de los
procesos fisiológicos que componen el presupuesto energético como son las tasas de
aclaramiento y de respiración, relacionados a los principales parámetros ambientales en tres
áreas usadas en el repoblamiento de la Bahía de Sechura es fundamental y más aún si se
realiza in situ ya que bajo condiciones de laboratorio no necesariamente refleja el hábitat
natural de la especie.

8
V. MARCO TEÓRICO
5.1. Aspectos fisiológicos
Según Flores (2012) los procesos metabólicos varían principalmente por estadios de
vida, estado reproductivo, proceso de digestión y estado de salud, así como por la
variación ambiental (temperatura, disponibilidad y composición de alimento,
composición y variación fisicoquímica del agua entre otros) y puede ser registrado
este estado metabólico a través de los índices de consumo de oxígeno (CO) y
excreción de amonio.
Balducci y Mendo (2011) afirman que la concha de abanico Argopecten purpuratus
al ser un bivalvo filtrador utiliza este proceso para filtrar al agua circundante y
adquirir tanto oxígeno para su respiración como células fitoplanctónicas del medio
para su alimentación. Según Bayne et al (1977) el proceso de la respiración ocurre
principalmente en la superficie de las branquias. Estas últimas tienen también la
función de la alimentación es decir tienen una función dual. Esto conlleva
necesariamente a relacionar la respiración con la alimentación y por lo tanto de las
entradas energéticas que toman lugar en el organismo.
Los bivalvos filtradores se alimentan de la oferta de alimento (seston) que se
encuentra en suspensión a unos pocos centímetros sobre el fondo. Retienen
partículas orgánicas en suspensión del agua circundante por medio del sistema ciliar
en los filamentos branquiales (Laudien et al 2006 y Vahl 1973).

5.1.1. Respiración
De los diferentes componentes que incluye el balance energético Bayne y Newell
(1983) indican que la respiración representa en general la mayor proporción de las
pérdidas energéticas. Esto refuerza la idea de que la pérdida energética debida a la
respiración tiene gran importancia para los estudios de manejo del recurso
específicamente en la acuacultura.
La respiración se puede definir como la energía perdida por el organismo en forma
de calor y es parte de la energía asimilada proveniente de los alimentos, que es
convertida a calor directamente o mediante trabajo mecánico. Células y organismos
son sistemas altamente ordenados y complejos por lo que el mantenimiento de sus
estructuras requiere una continua inversión de energía (Brey, 2001).
La respiración puede ser medida tanto por vía directa (producción de calor) como
por vía indirecta (consumo de oxigeno) (Brey, 2001). El oxígeno disuelto es un
factor regulador metabólico en los organismos acuáticos, que puede limitar la
capacidad metabólica y consecuentemente la producción de biomasa (Rosas et al.,
1998), y generalmente se mide, mediante la determinación del consumo de este gas
en un respirómetro (Hernández et al., 2007). El consumo de oxígeno en bivalvos
marinos ha sido estudiado por varios autores, la mayoría de estos estudios se han
realizado en ambientes controlados, como muestran los trabajos realizados sobre
tasas de respiración con especies como Placopecten magellanicus por Bricelj y
Shumway,(1991), o con Argopecten irradians por Bricelj et al., 1987).
La mayor parte de los estudios relacionan la tasa de consumo de oxígeno al tamaño
de los individuos, obteniendo relaciones alométricas, que muestran iguales
resultados que los obtenidos para la tasa de filtración. Así los estudios realizados
por James et al. (2001) para P. canaliculus, Resgalla et al. (2006) para P. perna,
Järnegren & Latín (2006) para A. excavata y Marsden (1999) para P. donacina,

10
entre otros, muestran que las tasas de respiración por individuo (mg. h‾ ¹) se
incrementan con el tamaño de los individuos, mientras que las tasas de respiración
por gramo de individuo (mg. h‾ ¹.g‾ ¹) decrecen al aumentar el tamaño de los
individuos y que estas variables se relacionan de forma exponencial con pendientes
positivas o negativas, según se trate de tasas respiratorias por individuo o por gramo
respectivamente.
Balducci y Mendo (2011) obtuvieron una mayor tasa de respiración Argopecten
purupuratus de menor tamaño (45-55mm) que en individuos de mayor tamaño (75-
85mm), registrando valores más altos durante el día en la Bahía de Sechura. En las
investigaciones de Chávez (2009) en la Bahía de Paracas y Vílchez (2011) en
Sechura se obtuvieron resultados similares en el que el consumo de oxígeno varió
claramente en relación directa con la talla.
Estudios sobre la variación mensual de la tasa de respiración, muestran que las
mayores tasas de respiración se obtienen en los meses cálidos. Resgalla et al. (2004)
muestra que las tasas de respiración promedio de P. perna, muestran
significativamente los mayores valores (0.8 ml. h‾ ¹.g‾ ¹) durante los meses más
cálidos (p < 0.01). Además Rueda & Smaal (2003) encontraron un patrón estacional
en las tasas de respiración de S. subtruncata, con mínimos valores (0.08 ml. h‾ ¹.g‾
¹) durante el invierno y máximos valores durante el verano (0.2 – 0.4 ml. h‾ ¹.g‾ ¹)
Por otro lado, Peñuela (2007) indica que el consumo de oxígeno específico (COE)
de la Argopecten purpuratus, evidencia que el estado fisiológico tiene una
influencia directa con el nivel metabólico confirmando la existencia de una relación
inversa entre el desarrollo corporal y el consumo de oxígeno por unidad de peso.
Farías et al. (2001) observó el mismo resultado en individuos reproductores de
Argopecten purpuratus que fueron expuestos a dietas con diferente contenido
proteico.

11
5.1.2. Filtración
Los bivalvos filtradores se alimentan removiendo el material particulado que se
encuentra suspendido en el agua el que además de fitoplancton incluye partículas
inertes que componen el detritus, pudiendo ser este último de origen orgánico o
inorgánico. La remoción de partículas suspendidas en el agua recibe el nombre de
tasa de aclaramiento y se entiende como el volumen de agua liberada de sus
partículas por unidad de tiempo (litros/hora). (Navarro, 2001).
La cantidad de alimento consumido por los filtradores está determinado por la tasa
de filtración y la velocidad de flujo de agua (Pares Sierra, 1996) además de la
eficiencia de retención de partículas y el tamaño de los individuos del cual se
conoce que los de mayor tamaño consumen mayor alimento que los pequeños.
Existen dos metodologías para la medición de la actividad alimentaria, el método
directo que separa y mide el flujo inhalante y exhalante en moluscos sifonados
(Owen, 1974) conocido como “tasa de bombeo” y el método indirecto que se basa
en la remoción de partículas en un volumen conocido como “tasa de filtración” y es
calculada como una función de la corriente alimenticia y de la retención de
partículas. (Coughlan, 1969).
Gosling (2003) indica que la tasa de filtración en bivalvos puede ser determinada
directa o indirectamente. El método directo es más eficiente cuando es aplicado a
aquellas especies donde las intensidades de inhalación y exhalación pueden ser
separadas. El método indirecto se utiliza para determinar la tasa de aclaramiento. El
bivalvo es colocado en un volumen conocido de agua conteniendo a una suspensión
de partículas por un periodo de tiempo. La filtración resulta en un decremento de la
concentración de partículas en un periodo de tiempo, la tasa de aclaramiento es
determinada por la disminución exponencial de la concentración algal.

12
La tasa de aclaramiento se ha definido como el volumen de agua que es limpiado
(filtrado) completamente de sus partículas, durante un tiempo determinado y
depende principalmente del tamaño corporal del organismo (Navarro 1983, Winter
1973). La tasa de filtración se mide en forma indirecta suspendiendo algún tipo de
partícula en el volumen determinado de agua y observando la disminución en la
concentración de éstas como consecuencia de la actividad metabólica de los
animales a lo largo de un tiempo determinado (Wong et al, 1997).
Se han realizado estudios de la tasa de filtración como el volumen de agua que pasa
a través del sistema de filtración por unidad de tiempo (también llamada tasa de
aclaramiento). En la mayoría de estos estudios se emplearon indistintamente el
método tradicional de aclaramiento, propuesto por Coughlan (1969), en el que se
utiliza el número de partículas en suspensión para determinar la cantidad de agua
filtrada.
Respecto al método tradicional de aclaramiento propuesto por Coughlan (1969), el
número de partículas en suspensión, puede ser estimado usando diferentes métodos
entre los que están: método tradicional de conteo de células descrito por Uthermohl
(1958), la lectura de la absorbancia a 750 nm y el método de cuantificar la biomasa
del fitoplancton a través de la clorofila-a, descrito por Parsons et al. (1984).
Resgalla et al. (2004) hacen una comparación entre estos métodos de cuantificación
de partículas para Perna perna, no encontrando diferencias significativas entre
dichos métodos, sin embargo dice que los datos obtenidos utilizando la clorofila-a
como estimador de la biomasa fitoplanctónica, son los más adecuados para los
análisis a realizar; confirmando así lo dicho por Fanslow et al. (1995), el cual
demostró que la clorofila-a es un parámetro sensible a las concentraciones del
seston en un determinado tiempo y lugar.

13
La actividad de filtración en bivalvos se ve influenciada por varios factores como la
concentración de fitoplancton, calidad, cantidad y talla de las partículas de alimento
y talla del animal. La tasa de filtración de invertebrados bentónicos ha sido
tradicionalmente medida como una tasa de remoción de material suspendido
(fitoplancton o partículas inorgánicas) por algunos bivalvos, en algunos periodos
cortos de tiempo, utilizando recipientes experimentales confinados y bajo
condiciones estáticas (Cole et al., 1992).
Flores (2012) observó mayores valores promedio de la tasa de filtración para
ejemplares adultos y los menores para las semillas de Argopecten purpuratus a
diferentes temperaturas, corroborando así el resultado del experimento de Winter et
al., 1984 y Uribe et al., 1996 en el que se observó que la tasa de filtración de otros
bivalvos se incrementaba a medida que aumentaba el tamaño corporal. Vilchez
(2011) determinó que el consumo de oxígeno varía en relación directa con la talla.
En relación a tasa de filtración obtuvo que la concha de abanico filtra con mayor
intensidad a una menor profundidad. Balducci y Mendo (2011) obtuvieron una
mayor tasa de filtración de los individuos de menor tamaño que los grandes con una
mayor actividad de filtración durante el día.
Según Winter et al. (1984) afirma que en bivalvos en general la tasa de filtración
decrece con el incremento de la concentración de alimento debido a que el exceso
de concentración algal comienza a tener un efecto negativo en la tasa de filtración
Este autor, observó también que la alta concentración de alimento no es sinónimo de
un mejor crecimiento y que incluso éste, podría ser negativo debido a la alta tasa de
biodeposición producida por los moluscos filtradores, las cuales se van acumulando
en el área de cultivo. Así concluyó que los organismos no son capaces de
seleccionar el alimento al filtrar, por lo que una alta concentración de partículas no
nutritivas disminuye significativamente el crecimiento.

14
Chávez (2009) determinó la tasa de filtración y la tasa de respiración de individuos
de Argopecten purpuratus de tres tallas obteniendo una relación inversa entre las
tasas de filtración por gramo y las tallas así como determinó la influencia de los
cambios estacionales sobre la respiración de los individuos.
En un estudio realizado por Laing (2004)) con Pecten maximus se observó una
relación inversa entre la concentración algal y la tasa de filtración, teniendo un
rango de 0,94- 9.66 mg/l-1 que correspondería a una concentración de 20- 200
células ul-1. Schulte (1975) reporta haber encontrado la misma relación inversa
entre la cantidad de alimento y la tasa de filtración, mencionando que en los casos
que se utilizaron concentraciones algales muy altas a menudo se encontraron valores
muy bajos e incorrectos, debido a que el mecanismo de filtración de los animales se
vio perturbado por la sobrecarga de partículas en suspensión (Schulte, 1975; Laing,
2004).
5.1.3. Parámetros ambientales
El metabolismo y crecimiento en moluscos bivalvos con relación a los factores
como temperatura, salinidad, oxígeno, cantidad y calidad de alimento y velocidad
de corriente, han sido investigados por muchos autores. Los resultados reportados
difieren ligeramente entre los distintos autores debido a que se trabajó con
diferentes especies, distintas latitudes y tipos de Bahías.
La respuesta de los organismos filtradores en el medio ambiente es compleja, por lo
que se deben analizar principalmente los factores que sean o puedan hacerse
limitantes. La temperatura es uno de los factores que produce cambios en la
fisiología de los organismos (His 1977), principalmente en la tasa de filtración de
los moluscos bivalvos y en el crecimiento en esta especie (Rivas y Masse 1983,
Malouf y Breese 1978).

15
Según estudios realizados (Widdows, 1978; Griffiths, 1980; Navarro y Winter,
1982; Iglesias et al., 1992, entre otros) demuestran que la temperatura está entre los
factores ambientales más influyentes en la fisiología de los moluscos bivalvos.
El efecto de la temperatura en el consumo de oxígeno específico (CEO) para cada
estadio de desarrollo es evidente. Un incremento de este factor acelera la mayoría de
procesos fisiológicos como la tasa de respiración (Griffiths & Griffiths, 1987).
Flores (2012) obtuvo incrementos desde hasta 40 veces al comparar los valores de
CEO obtenidos entre 17° y 22°C, debido a que, por estar los individuos
acondicionados a una mayor temperatura (22ºC) los requerimientos energéticos son
muchos más elevados (Araya et al., 1993). Sin embargo, Marsden (1999) para P.
donacina creo una regresión múltiple para determinar la importancia de las
variables masa corporal, temperatura, estación y consumo de oxígeno, sobre la tasa
de respiración, demostrando que la variable tasa de respiración era explicada un
63% por el peso corporal y 27% por la temperatura del medio. Vahl (1978) (citado
por Shumway et al 1988) concluyó que la variación estacional del consumo de
oxigeno de Chlamys islandica no estuvo explicada por la variación que se vio en la
temperatura, sino más bien por la cantidad de alimento disponible, también mostró
una variación significativa en el consumo de oxigeno que es explicado por el peso
corporal, además Bayne et al 1976 (Citado por Masrden 1999) encontró la misma
relación en Paphies donacina. Así como Lértora (2008) no obtuvo una relación
significativa entre las tasas de respiración y la temperatura con la almeja Gari solida
en una investigación in situ.
Por otro lado, Navarro y González (1998) encontraron que la salinidad tenía un
efecto directo sobre la actividad metabólica de Argopecten purpuratus, así, el
consumo de oxígeno aumenta significativamente al bajar la salinidad. En el mismo
estudio se encontró que el consumo de oxígeno disminuyo a menores salinidades
experimentales lo que estaría relacionado con la disminución de la alimentación
indicando un aislamiento para evadir el estrés osmótico. Patrones similares de

16
consumo de oxígeno en relación a fluctuaciones de salinidad han sido descritos para
otras especies de bivalvos como Mytilus edulis (Widdows 1973).
De acuerdo a Navarro y González (1998), cuando ejemplares de la concha de
abanico son expuestos a bajas salinidades, la tasa de filtración disminuye, de manera
que un ejemplar que filtra 2.6 L h-1 en una salinidad de 30 ‰, baja a 0.12 L h-1, si
es expuesto a una salinidad de 18 ‰.

VI. METODOLOGÍA
El estudio se llevará a cabo en cuatro áreas de repoblamiento dentro de la Bahía de
Sechura, localizada en el Distrito del mismo nombre, Región Piura. Se realizará en dos
épocas distintas caracterizadas por diferencias de temperaturas y salinidad del mar, invierno
y verano, durante los meses de Agosto y Enero. La primera estación experimental
localizada frente a Barrancos (5°47´ 20.3¨ S - 80°54´ 17.3¨ W), la segunda frente a Vichayo
(5°47´ 39¨S - 80°57´ 10.9¨ W), la tercera frente a Parachique (5° 44´ 27.9¨S - 80°55´ 32.3¨
W) y la cuarta frente a Las Delicias (5°43´ 35.7¨ S - 80°54´ 3.4¨ W).
Figura 1 : Ubicación de las zonas de estudio donde se
realizaran los experimentos considerados en este estudio.

6.1. Tipo de investigación
La investigación es de tipo experimental explicativa debido a que a lo largo del
presente estudio se expondrá el comportamiento de las variables tasa de respiración
y aclaramiento en función de otras variables como talla y peso, requiriendo de
control y cumpliendo criterios de causalidad.
6.2. Formulación de hipótesis
No existen diferencias en la relación entre la talla y tasa de consumo de oxígeno y
aclaramiento de la concha de abanico en las cuatro zonas de repoblamiento y en
ambas estaciones del año.
6.3. Identificación de variables
Las variables dependientes o respuestas son la tasa de respiración y la tasa de
aclaramiento ya que estas son influenciadas por factores parámetros bióticos y
abióticos.
Las variables independientes o explicativas son las cuatro zonas de investigación,
talla y peso de las conchas de abanico.
6.4. Definiciones operacionales
Ejemplares de Argopecten purpuratus de cuatro diferentes grupos de tamaños 40 a
50 mm, 60 a 70 mm y 80 a 90 mm y de 90 mm a 11 mm colectados aleatoriamente
en una misma zona serán empleados para la medición de tasas de respiración y
aclaramiento. Estas tasas serán medidas dentro de un sistema cerrado de circulación

19
empleando una batería de 5 cilindros o respirómetros, como se puede apreciar en la
figura 2.
Figura 2: Batería de respirómetros empleados en la investigación.
Para cada experimento se empleará cinco cilindros acrílicos provistos de un o-ring
que impedirá el intercambio de agua. Cuatro de estos cilindros conformaran el
grupo de unidades experimentales y uno el grupo control. En cada respirómetro el
agua que ingresa y sale será conectada vía mangueras de 1cm de diámetro a una
bomba sumergible de 30-40W de potencia y con un flujo de 15L\min marca Cornet
modelo GEO-inline PLUS 12V para crear un sistema de recirculación. La energía
empleada será proporcionada por una batería de 11 placas de marca ETNA, a la que
será conectada una consola de mando con la que se controlará el encendido de las
bombas.
6.4.1. Instalación
Previo a la experimentación, los ejemplares colectados serán lavados
cuidadosamente para retirar los sedimentos y epibiontes eliminando así cualquier
organismo que pudiera interferir la medición Cada individuo será colocado al azar

20
en los respirómetros previamente medido. En cada respirómetro se colocará al azar
un individuo previamente medido. Así mismo se introducirá un oxímetro de la
marca HOBO modelo U26-001 para el registro continuo de datos de oxígeno y
temperatura. El oxígeno será registrado a un intervalo de cinco minutos por el data
logger HOBO en un periodo de una hora.
El set de respirómetros será colocado en el fondo mediante un sistema de cuerdas.
y se activarán las bombas con las entradas de agua provistas de un filtro de 0.08mm
el cuál evitará la entrada de algún organismo que pueda obstruir las bombas. Así
cada individuo permanecerá 20 minutos aproximadamente dentro de los cilindros
con el sistema abierto para que pueda adaptarse. Una vez adaptado, se cerrará el
sistema y se registrará la primera medida de oxígeno y se tomará una muestra inicial
de 100 ml de agua para realizar el análisis de recuento de células fitoplanctónicas.
Luego de una hora, el sistema se abrirá el sistema y se tomará la muestra final de
agua para ser analizada en el laboratorio y así se realizará cuatro veces que será el
número de repeticiones.
6.4.2. Análisis de muestras
Las muestras de conchas de abanico serán llevadas al laboratorio, donde se realizará
el peso seco de cada una, para ello se empleará el secador de marca MEMMERT en
el que se colocará las muestras a 60°C por 48 horas.
Por otro lado para la determinación de la tasa de aclaramiento se emplearán
muestras de agua que serán almacenadas en frascos plásticos color ámbar y
conservadas en formol al 15% previamente a su traslado al laboratorio para el
análisis el cual se realizará siguiendo la técnica de Utermohl (1958).

21
6.4.2.1. Tasa de consumo de oxígeno
Para la determinación de la tasa de consumo de oxígeno se empleará la
ecuación de Widdows y Johnson(1988): RR=[(Cto-Ct1)x (V/ ΔT)]-f, donde
RR= Tasa de consumo de oxígeno (ml O2.h-1), Cto= concentración de
oxígeno en el tiempo cero (mL O2.h-1), Ct1=Concentración de oxígeno al
final del experimento (ml O2.h-1), V= Volumen de la cámara de prueba (L),
ΔT=Intervalo de tiempo de prueba (h) y f=Factor de corrección en la cámara
de control obtenido con la misma ecuación.
6.4.2.2. Tasa de aclaramiento
La tasa de aclaramiento mide el descenso de la densidad de microalgas del
medio experimental. Entonces, la tasa de aclaramiento se hallará mediante la
fórmula de Coughlan (1969) RC=V*[(LnCto-LnCt)-f)]/tN donde RC= Tasa
de aclaramiento en ml/hora, Cto= concentración inicial de fitoplancton,
Ct=Concentración de fitoplancton al final del experimento, V= Volumen del
cilindro de prueba (mL), N=número de organismos por cámara,
ΔT=Intervalo de tiempo de prueba (hora) y f=Factor de corrección de
fitoplancton para el control calculado con la misma fórmula.
Para mejorar las comparaciones entre los rangos de tallas se calcularan las tasas de
consumo de oxígeno y aclaramiento específicas es decir, por gramo de peso seco de
cada individuo.

22
6.5. Diseño de investigación
El experimento se realizará bajo cuatro tratamientos que serán los diferentes grupos
de tallas de conchas de abanico para las que se determinaran las tasas de consumo
de oxígeno y aclaramiento .Los tratamientos serán repetidos cuatro veces para cada
experimento es decir, para las cuatro zonas de repoblamiento en investigación. Los
individuos seleccionados para cada grupo de talla serán asignados al azar a cada
unidad experimental que son los respirómetros.
La siguiente tabla muestra las tasas de consumo de oxígeno o aclaramiento bajo los
cuatro rangos de tallas, con las respectivas repeticiones. Los factores en estudio son:
A: Zonas o estaciones de experimentación (a1= Barrancos, a2= Vichayo, a3=
Parachique y a4 = Las Delicias)
B: Rango de tallas (b1= 40-50 mm, b2= 60-70mm, b3=80-90mm, =90-110mm)
Tabla 1: Diseño para cada experimento
6.6. Población y muestra
La población objeto de observación y estudio serán las conchas cultivadas en las
áreas de repoblamiento. La muestra serán las 64 conchas de abanico empleadas para
cada experimento. El muestreo se realizará aleatoriamente.
a1 a2 a3 a4
b1 b2 b3 b4 b1 b2 b3 b4 b1 b2 b3 b4 b1 b2 b3 b4
R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1 R1
R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2 R2
R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3 R3
R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4 R4

23
6.7. Instrumentos de colecta de datos
El estudio se realizará experimentalmente in situ y se empleará un set de cinco
respirómetros dentro del cual se ingresará un oxímetro de la marca HOBO que
permitirá registrar los datos de oxígeno, para la determinación de la tasa de
aclaramiento se tomarán muestras de agua que serán colectadas en pequeñas bolsas
de plástico para luego ser conservadas en frascos del mismo material.
6.8. Procedimiento de análisis de datos
Para determinar la tasa de consumo de oxígeno y aclaramiento, siguiendo la
ecuación de Widdows y Johnson (1988) y la fórmula de Coughlan (1969)
respectivamente, para cada rango de tallas y para cada zona de estudio se hará uso
del programa Excel 2010 para realizar los respectivos cálculos y gráficas.
La relación existente entre la talla y las tasas de consumo de oxígeno y aclaramiento
en cada zona de estudio se determinará con un análisis de regresión simple para
medir el grado de correlación existente entre las variables. Para determinar si estas
relaciones son diferentes o no en entre zonas y estaciones del año se realizará una
comparación de las líneas de regresión. Así se comparará primero las varianzas
residuales, luego las pendientes y, finalmente, las elevaciones. En términos del
modelo sería: Y=αi + βiXij + εij , donde i=1,2,3,4 que significarán las cuatro zonas.
Primeramente se comparará las varianzas residuales σ12 y σ2
2, luego β1 y β2 , y
finalmente las elevaciones de las líneas α1 y α2 .
Por otro lado, se realizará también el análisis multivariado de regresión lineal
múltiple, en a que se relacionará las variables dependientes, tasa de consumo de
oxígeno y aclaramiento con las variables independientes cualitativa talla e
indicadora o dummy zonas. Este modelo permitirá establecer los parámetros

24
diferentes entre las tasas fisiológicas de las cuatro zonas dado el mismo grupo de
tallas. El modelo de este análisis es: Y/x1,x2,x3 = β 0+ β1X1+ β2X2+ β3X3+εi
.
Los dos análisis de regresión se realizaran con ayuda del paquete estadístico SPSS y
el programa Excel 2011.
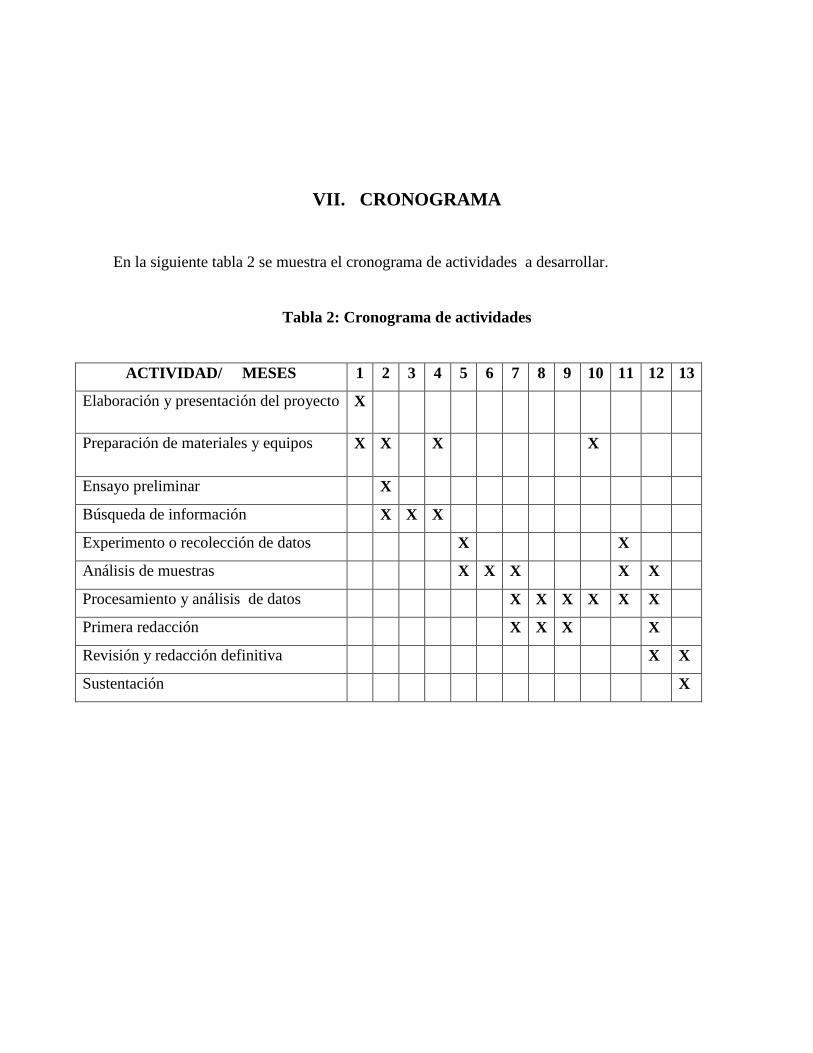
VII. CRONOGRAMA
En la siguiente tabla 2 se muestra el cronograma de actividades a desarrollar.
Tabla 2: Cronograma de actividades
ACTIVIDAD/ MESES 1 2 3 4 5 6 7 8 9 10 11 12 13
Elaboración y presentación del proyecto X
Preparación de materiales y equipos X X X X
Ensayo preliminar X
Búsqueda de información X X X
Experimento o recolección de datos X X
Análisis de muestras X X X X X
Procesamiento y análisis de datos X X X X X X
Primera redacción X X X X
Revisión y redacción definitiva X X
Sustentación X
26
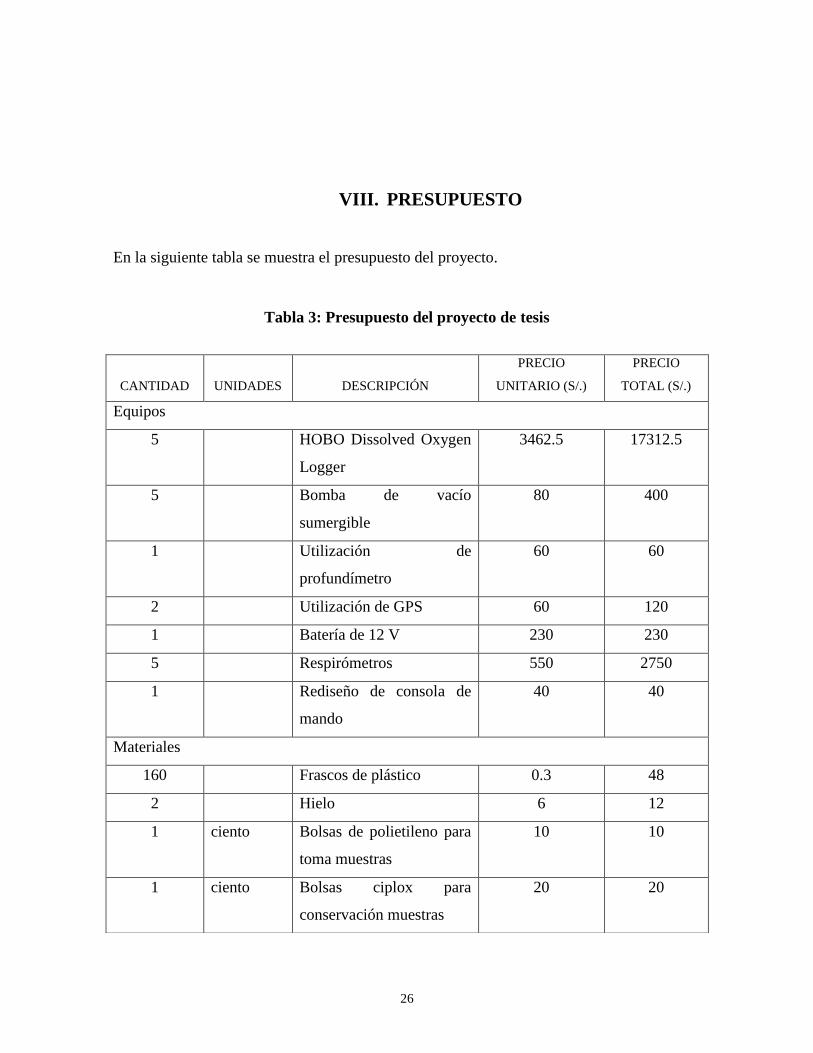
VIII. PRESUPUESTO
En la siguiente tabla se muestra el presupuesto del proyecto.
Tabla 3: Presupuesto del proyecto de tesis
CANTIDAD
UNIDADES
DESCRIPCIÓN
PRECIO
UNITARIO (S/.)
PRECIO
TOTAL (S/.)
Equipos
5 HOBO Dissolved Oxygen
Logger
3462.5 17312.5
5 Bomba de vacío
sumergible
80 400
1 Utilización de
profundímetro
60 60
2 Utilización de GPS 60 120
1 Batería de 12 V 230 230
5 Respirómetros 550 2750
1 Rediseño de consola de
mando
40 40
Materiales
160 Frascos de plástico 0.3 48
2 Hielo 6 12
1 ciento Bolsas de polietileno para
toma muestras
10 10
1 ciento Bolsas ciplox para
conservación muestras
20 20
27
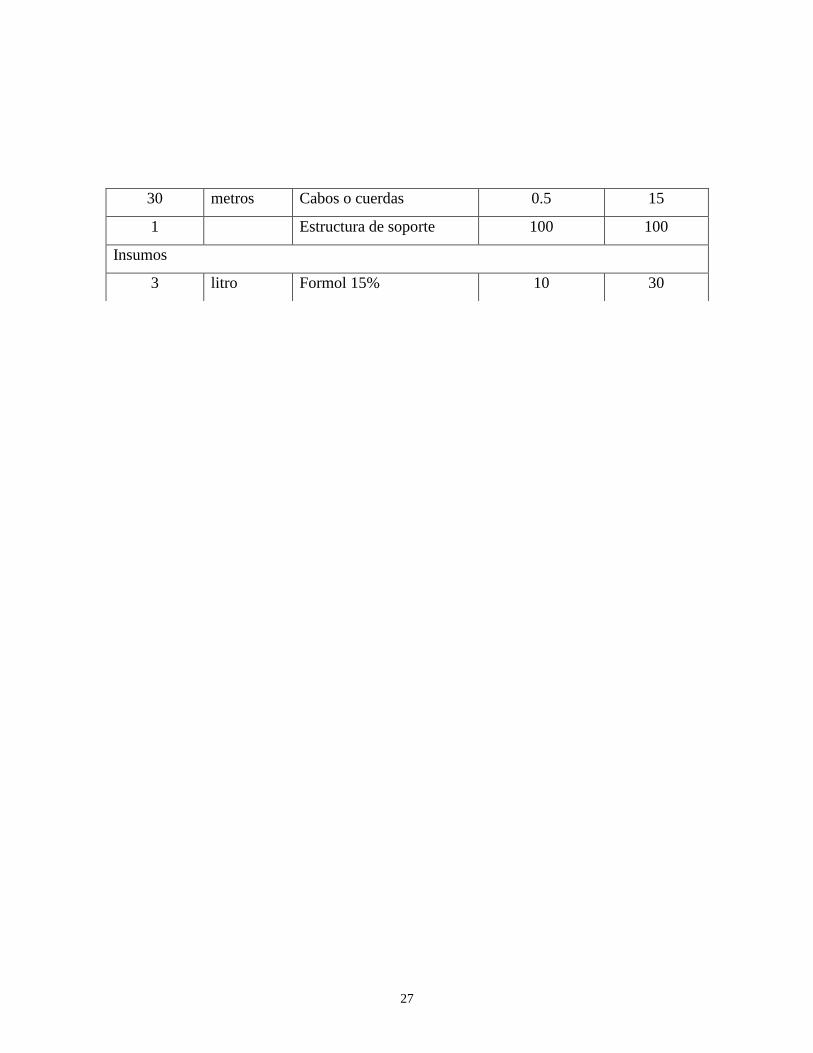
30 metros Cabos o cuerdas 0.5 15
1 Estructura de soporte 100 100
Insumos
3 litro Formol 15% 10 30

28
IX. COLABORADORES
Patrocinador: Dr. Jaime Mendo Aguilar (Profesor principal de la Facultad de Pesquería
UNALM)
Centro Leibniz de Ecología Marina Tropical- Alemania: ZMT Bremen.
Ministerio Federal de Educación e Investigación- Alemania (BMBF)
Universidad Nacional Agraria La Molina

29
X. REFERENCIAS BIBLIOGRÁFICAS
- Araya R, Jorquera M & Riquelme C. 1999. Asociación de bacterias al ciclo de vida
de Argopecten purpuratus. Revista Chilena de Historia Natural. 72: 261-271.
- Balducci N. y Mendo J. 2011. Variación circadiana de la tasa de respiración y
filtración de Argopectrn purpuratus en la Bahía de Sechura, Piura, Perú. Libro de
resúmenes del Seminario “Bases científicas para el manejo y repoblamiento de la concha de
abanico en la Bahía de Sechura” 11 de noviembre, Piura.
- Bayne, B. L. and D.R. Livingstone. 1977. Responses of Mytilus edulis to low
oxygen tension: Acclimation of the rate of oxygen consumption. Journal of comparative
physiology. Vol 114, pp 129- 142.
- Brey, T. 2001. Population dynamics in benthic invertebrates. Virtual Handbook on
PopulationDynamics.
- Bricelj, M., J. Epp and R. E. Malouf. 1987. Comparative physiology of young and
old cohorts of bay scallop Argopecten irradians irradians (Lamarck): mortality, growth,
and oxygen consumption. Journal of experimental Marine Biology and Ecology. Vol.112,
pp. 73-91.
- Boynton, W.R.; W.M. Kemp & C.G. Osborne. 1980. Nutrient fluxes across the
sediment-water interface in the turbid zone of a coastal plain estuary. pp.93-111. .En: V.S.
Kennedy (ed.). Estuarine Perspectives. Academic Press, New York. USA.
- Chávez, C. 2009. Determinación de la tasa de filtración y la tasa de respiración de la
concha de abanico Argopecten purpuratus en playa Atenas, Bahía de Paracas. Tesis para
optar el título de Ingeniero Pesquero, Facultad de Pesquería. UNALM

- Cole Be, Thompson JK, Cloern JE. 1992. Measurement of filtration rates by
infaunal bivalves in a recirculating flume. Marine Biology. 113: 219-225.
- Coughlan, J. 1969. The estimation of the filtering rate from the clearance of
suspensions. J ,Mar,Biol. Vol 2 pp. 356- 358.
- Dame, R., R. Zingmark, H. Stevenson & D. Nelson. 1980. Filter fedeer coupling
Between the estuarine water column and benthic subsystems. pp. 521-526. En: V.S.
Kennedy (ed.). Estuarine Perspectives. Academic. Press, New York, USA
- Farías A. 2001. Nutrición en Moluscos Pectínidos IN: MAEDA ALFONSO .Los
Moluscos Pectínidos de Iberoamérica: Ciencia y Acuicultura . MEXICO : A. Maeda -
Martínez (Ed.) Limusa ; p. 84-104
- Flores A. 2012. Área Funcional de Investigaciones en Acuicultura, DGIA,
IMARPE. Estudio bio oceanografico para determinación de la capacidad de carga en la
bahía de Sechura
- George A. 1997. Determinación de la tasa de respiración de la almeja Catarina
argopecten ventricosus en función de la talla bajo condiciones de laboratorio. Universidad
Autónoma de Baja California sur área interdisciplinaria de ciencias del mar departamento
de biología marina.
- Gosling E. 2003. Bivalve Molluscs. Biology, Ecology and culture. Fishing News
Books. Oxford. Pág 88-89.
- Hernández Z.M Hernández-Arevalo J., Matus P. Cruz-Casallas E. 2007. Consumo
de oxígeno en cachama blanca (Piaractus brachypomus) durante diferentes etapas de
desarrollo corporal. Universidad de Los Llanos. Colombia. Orinoquia, vol. 11, núm. 1,
2007, pp. 49-55.
- His, E. 1977. Aspects biologiques du stockage des hiutres en bassin conseils
practiques aux ostreiculteurs. Science et Peche, Bull. Inform. Inst. Peches marit. 272:1-4.

31
- Laing, I. 2004. Filtration of king scallops Pecten maximus .Aquacualture n 240, pp
369-384.
- Laudien, J., J.-M. Gili, and W. E. Arntz. 2006. Filter-feeding impact of the
Mediterranean surf clams Donax trunculus (Bivalvia, Donacidae) on the plankton
community. p 2-7.
- Lértora G. 2008. Evaluación in situ de la Tasa de Filtración y Respiración de la
“almeja” Gari solida (Gray, 1828) en la Bahía de Paracas. Tesis para optar el título de
Biólogo Facultad de Biología. UNALM
- Malouf, R.E & W. P. Breese. 1978. Intensive culture of the pacific oyster
Crassostrea gigas (Thunberg) in heated effluents. Agricultural Exptl. Station Bull. 627:41.
- Marsden, I.D. 1999. Respiration and feeding of the surf clam Paphies donacina
from New Zealand. Hydrobiologia, vol. 405, pp. 179- 188
- Mendo J, Ysla L, Orrego H, Miglio M, Gil P, Del Solar A. 2011. Bases Científicas y
Tecnológicas para incrementar la productividad del cultivo de concha de abanico en áreas
de repoblamiento en la Bahía de Sechura. Proyecto FINCYT – Contrato Nº 01 – 2009.
- Mendo J., Wolff M., Carbajal W., Gonzales I. y Badjeck M. 2008. Manejo y
explotación de los principales bancos naturales de concha de abanico (Argopecten
purpuratus) en la costa peruana. FAO. Actas de Pesca y Acuicultura Nro. 12:101-114.
- Mincetur.2008. Perfil del Mercado y Competitividad Exportadora de la Concha de
Abanico.
- Navarro, J.M and C.M. Gonzalez. 1998. Physiological responses of the chilean
scallop Argopecten purpuratus to decreasing salinities. Aquaculture, vol.167, pp. 315- 327.
- Navarro, J.M. 2001. Fisiología energética de pectínidos iberoamericanos. Los
moluscos pectínidos de iberoamérica: ciencia y acuicultura. Capítulo 3: p61 - p76.
- Osorio O. e Ibarra E. 1987. Métodos estadísticos aplicados a la investigación
agrícola. IICA-Honduras p27-p56.

32
- Pares-Sierra, G.M. 1999. Efecto del flujo y la concentración de alimento sobre el
crecimiento tasa de filtración de escalopas juveniles (Nodipecten subnodosus). Tesis de
maestría. Universidad Autónoma de Baja California. Instituto de Investigaciones
Oceanológicas. Revista de Ciencias Marinas (2000), 26(4):621-641
- Peñuela Z, Hernandez G, Corredor J, Cruz P. 2007. Consumo de oxígeno en
carachama blanca (Piaractus brachipomus) durante diferentes etapas de desarrollo corporal.
Orinoquia. Vol 11, número 001. Villavicencio, Colombia, pp. 49 – 55
- Phillipson J 1975 Rainfall, primary production and "carrying capacity" of Tsavo
National Park, Kenya. East African Wild life Journal 18 (4): 171 – 201
- Resgalla Jr., C, E. S Brasil, and L. C. Salomao. 2006. Physiological rates in
different classes of sizes of Perna perna (Linnaeus, 1758) submmited to experimental
laboratory conditions. Brazilian Journal of Biology, vol. 66, pp. 325- 336.
- Supo J. 2012. Seminarios de Investigación Científica: Metodología de la
investigación para las ciencias de la salud Editorial:Bioestadistico EIRL..
- Shumway, S.E., Newell R.. 1984. Energy resource allocation in Mulinia lateralis,
an opportunist bivalve from shallow wáter sediments. Ophelia 23:101-118.
- Schulte, E. H. 1975. Influence of algal concentration and temperature on the
filtration rate of Mytilus edulis. En: Marine biology 30, p331 – p341. La Spezia, Italia.
- Utermöhl, H., 1958. Perfeccionamiento del método cuantitativo del fitoplancton. H
L - E. Schweizerbart'sche Verlagsbuchhandlung, Science Publishers, Stuttgart. Alemania
- Valdivia V. 2008. Diseños experimentales. Selección según objetivo de la
investigación. Estadística II. Universidad Católica Agropecuaria del Trópico Seco

33
- Walne, P. 1972. The Influence of Current Speed, Body Size and Water Temperature
on the Filtration Rate of Five Species of Bivalves. Journal of the Marine Biological -
Association of the United Kingdom, 52, pp 345-374.
- Vahl, O. 1973. Efficiemcy of particle retention in Chlamys islandica.(O.F. Muller)
Astarte 6:21-25
- Widdows, J. 1973. Effect of temperature and food on the heart beat, ventilation rate
and oxygen uptake of Mytilus edulis. Journal of Marine Biology. vol 20, pp 269- 276. 4


















