







Idiomas
Páginas
Jurídico


ALGUNOS ASPECTOS SOBRE LA BIOLOGÍA Y ECOLOGÍA DE LA
GUAPUCHA, Grundulus bogotensis (Humboldt, 1821)
(TELEOSTEI, CHARACIDAE), EN EL ALTIPLANO
CUNDIBOYACENSE, COLOMBIA.
CAMILO ANDRÉS ROA FUENTES
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BIOLOGÍA
Bogotá, D.C.
Agosto de 2007

ALGUNOS ASPECTOS SOBRE LA BIOLOGÍA Y ECOLOGÍA DE LA
GUAPUCHA, Grundulus bogotensis (Humboldt, 1821)
(TELEOSTEI, CHARACIDAE), EN EL ALTIPLANO
CUNDIBOYACENSE, COLOMBIA.
CAMILO ANDRÉS ROA FUENTES
TRABAJO DE GRADO
Presentado como requisito parcial
para optar al título de
BIÓLOGO
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
CARRERA DE BIOLOGÍA
Bogotá, D.C.
Agosto de 2007

Artículo 23 de la Resolución N° 13 de Julio de 1946
“La Universidad no se hace responsable por los conceptos emitidos por sus
alumnos en sus trabajos de tesis. Solo velará por que no se publique nada
contrario al dogma y a la moral católica y por que las tesis no contengan
ataques personales contra persona alguna, antes bien se vea en ellas el
anhelo de buscar la verdad y la justicia”.

ALGUNOS ASPECTOS SOBRE LA BIOLOGÍA Y ECOLOGÍA DE LA
GUAPUCHA, Grundulus bogotensis (Humboldt, 1821)
(TELEOSTEI, CHARACIDAE), EN EL ALTIPLANO
CUNDIBOYACENSE, COLOMBIA.
CAMILO ANDRÉS ROA FUENTES
APROBADO
____________________________ ____________________________
Saúl Prada Pedreros, Biólogo Ph.D. Carlos Rivera Rondón, Biólogo MSc.
Director Asesor
____________________________ ____________________________
Jorge E. Forero Useche, Biólogo Andrés Acosta Galvis, Biólogo
Jurado Jurado

ALGUNOS ASPECTOS SOBRE LA BIOLOGÍA Y ECOLOGÍA DE LA
GUAPUCHA, Grundulus bogotensis (Humboldt, 1821)
(TELEOSTEI, CHARACIDAE), EN EL ALTIPLANO
CUNDIBOYACENSE, COLOMBIA.
CAMILO ANDRÉS ROA FUENTES
APROBADO
____________________________ ____________________________
Ángela Umaña, M. Phil Andrea Forero, Bióloga
Decano Académico Directora Carrera de Biología

A la casi infinita paciencia de mis padres,
y a ellos, por supuesto…

vi
Agradecimientos
De manera especial a Saúl Prada Pedreros, Ricardo Álvarez Zamora y
Carlos Rivera Rondón, sin su apoyo y contribución la consecución del mismo
hubiese sido imposible.
A las siguientes personas, en orden alfabético, por su valiosa ayuda en las
diferentes etapas del proyecto: Ángela Zapata, Angélica Pérez, Armando
Ortega Lara, Diana Galindo, Diana Morales, Iván Arce, Javier Maldonado
Ocampo, Javier Táutiva, Juana Andrade, familia Maldonado Ocampo,
Vladimir Páez, y a los integrantes del grupo de estudios en ictiología y
ecosistemas acuáticos de la Universidad Javeriana.
De igual forma expreso mi agradecimiento a Javier Molina y Francisco Ciri de
la Corporación Autónoma de Boyacá por su desinteresada colaboración.
Aunque no fueron parte activa dentro de este trabajo, manifiesto mi sincera
gratitud con Carlos Lasso, Juan Carlos Rodríguez, Alejandro Giraldo y Julián
Mora del Museo de Historial Natural La Salle.
Finalmente, al Instituto de Investigaciones Alexander von Humboldt y a la
Pontificia Universidad Javeriana por la co-financiación del proyecto Nº 287.
Camilo Roa
Bogotá
Agosto de 2007

vii
TABLA DE CONTENIDO
1. Introducción ............................................................................................ 3 2. Marco teórico y revisión de literatura ...................................................... 5 2.1 Antecedentes ....................................................................................... 5 2.2 Sistemática y taxonomía ...................................................................... 6 2.3 Distribución .......................................................................................... 7 2.4 Aspectos de la biología y ecología....................................................... 8 2.4.1 Hábitat ............................................................................................... 8 2.4.2 Estimativos poblacionales ................................................................. 9 2.4.3 Proporción de sexos.......................................................................... 9 2.4.4 Relación longitud – peso ................................................................. 10 2.4.5 Dieta y amplitud de nicho trófico...................................................... 12 2.5 Grado de amenaza y medidas de conservación ................................ 14 3. Formulación del problema y justificación. ............................................. 15 3.1 Formulación del problema.................................................................. 15 3.2 Preguntas de investigación ................................................................ 16 3.3 Justificación........................................................................................ 16 4. Objetivos............................................................................................... 18 4.1 Objetivo general ................................................................................. 18 4.2 Objetivos específicos ......................................................................... 18 5. Área de Estudio .................................................................................... 19 5.1 Altiplano Cundiboyacense.................................................................. 19 5.1.1 Temperatura media anual ............................................................... 19 5.1.2 Clima ............................................................................................... 20 5.1.3 Aspectos geológicos........................................................................ 20 5.2 Cuenca hidrográfica del río Bogotá.................................................... 21 5.2.1 Distribución temporal de la precipitación ......................................... 21 5.2.2 Distribución espacial de la precipitación.......................................... 22 5.2.3 Localidades de muestreo cuenca del río Bogotá............................. 23 5.3 Cuenca hidrográfica del río Suárez.................................................... 29 5.3.1 Distribución temporal de la precipitación ......................................... 29 5.3.2 Distribución espacial de la precipitación.......................................... 29 5.3.3 Localidades de muestreo cuenca del río Suárez............................. 30 5.4 Cuenca hidrográfica de lago de Tota ................................................. 34 5.4.1 Localidades de muestreo cuenca del lago de Tota ......................... 35 6. Materiales y Métodos............................................................................ 36 6.1 Selección de localidades.................................................................... 36 6.2 Captura y preservación de los peces ................................................. 36

viii
6.3 Datos de captura y caracterización del hábitat................................... 37 6.4 Proporción de sexos .......................................................................... 38 6.5 Relación longitud – peso.................................................................... 38 6.6 Contenido estomacal ......................................................................... 39 7. Análisis de la información ..................................................................... 40 7.1. Abundancia y caracterización del hábitat ........................................... 40 7.2. Proporción de sexos .......................................................................... 40 7.3. Relación longitud – peso.................................................................... 40 7.4. Contenido estomacal ......................................................................... 41 8. Resultados............................................................................................ 48 8.1 Abundancia de G. bogotensis y caracterización del hábitat ............... 48 8.1.1 Parámetros fisicoquímicos en cuerpos de agua lóticos................... 51 8.1.2 Parámetros fisicoquímicos en cuerpos de agua lénticos................. 51 8.2 Proporción de sexos .......................................................................... 55 8.3 Relación longitud – peso.................................................................... 57 8.4 Aspectos tróficos................................................................................ 60 9. Discusión .............................................................................................. 68 9.1 Abundancia de G. bogotensis y caracterización del hábitat ............... 68 9.2 Proporción de sexos .......................................................................... 72 9.3 Relación longitud – peso.................................................................... 74 9.4 Aspectos tróficos................................................................................ 76 10. Conclusiones ........................................................................................ 81 10.1 Abundancia y caracterización del hábitat ........................................... 81 10.2 Proporción de sexos .......................................................................... 81 10.3 Relación longitud - peso..................................................................... 82 10.4 Dieta................................................................................................... 82 11. Recomendaciones ................................................................................ 83 12. Referencias........................................................................................... 85 13. Anexos.................................................................................................. 99

ix
ÍNDICE DE TABLAS
Tabla 1. Localidades de muestreo en la parte media y alta del río Bogotá... 23
Tabla 2. Localidades de muestreo en la cuenca del río Suárez y cuenca del
lago de Tota. ................................................................................................. 30
Tabla 3. Códigos asignados para los ítems alimentarios.............................. 47
Tabla 4. Tipo de cuerpo de agua, macróftica dominante, abundancia de G.
bogotensis y tiempo de pesca....................................................................... 48
Tabla 5. Abundancia de G. bogotensis por cuencas..................................... 50
Tabla 6. Parámetros fisicoquímicos para las 16 localidades de estudio. ...... 53
Tabla 7. Proporción de sexos en G. bogotensis por localidad. ..................... 55
Tabla 8. Proporción de sexos por intervalos de longitud total de todos los
individuos capturados de G bogotensis en las 16 localidades de estudio. ... 56
Tabla 9. Relación longitud – peso de G. bogotensis en 11 localidades de
estudio .......................................................................................................... 58
Tabla 10. Riqueza de ítems alimentarios, amplitud de nicho determinado por
los índices de Levins (B), Shannon (H’), y su valor de equidad (J’), para las
11 localidades de estudio. ............................................................................ 60
Tabla 11. Valores del índice de preponderancia (IP) de los ítems
alimentarios................................................................................................... 67

x
ÍNDICE DE FIGURAS
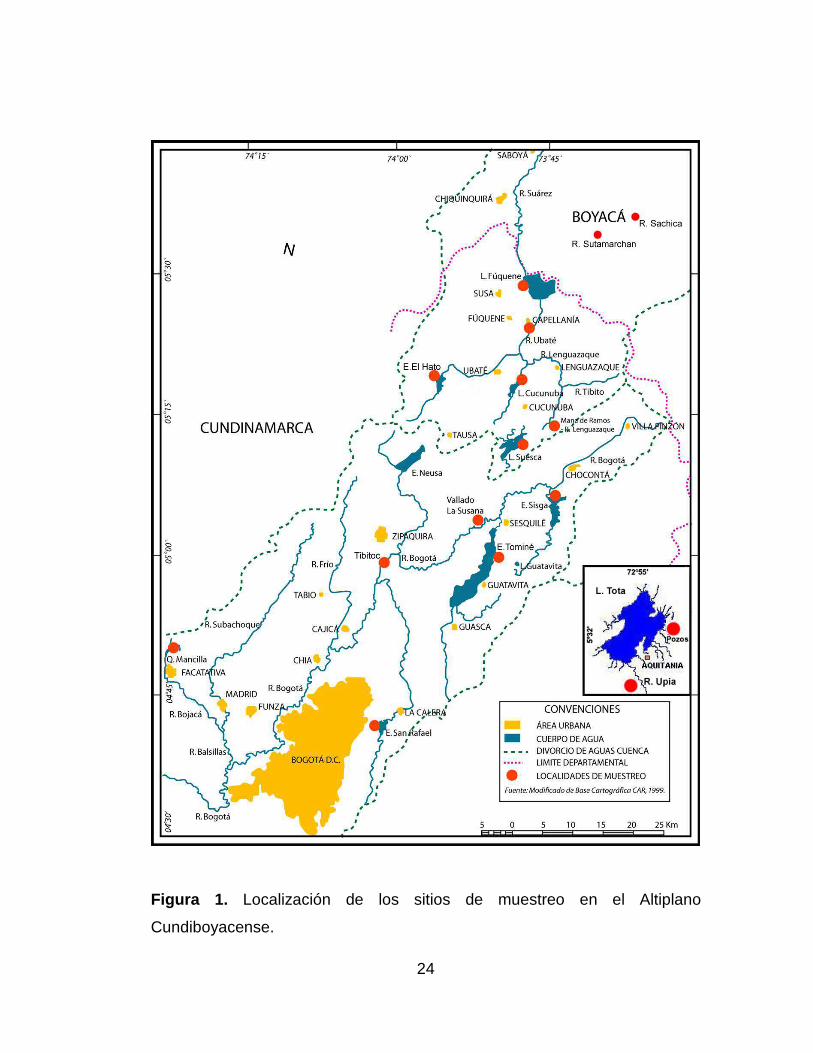
Figura 1. Localización de los sitios de muestreo en el Altiplano
Cundiboyacense. .......................................................................................... 24
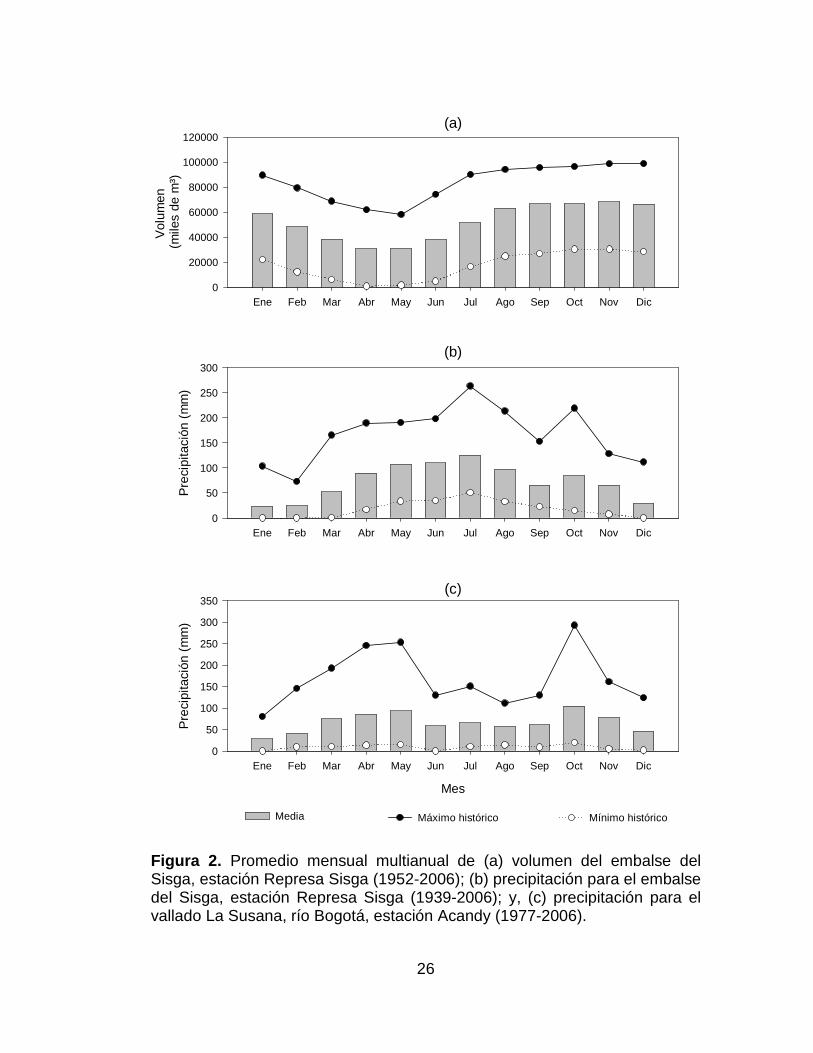
Figura 2. Promedio mensual multianual de (a) volumen del embalse del
Sisga, estación Represa Sisga (1952-2006); (b) precipitación para el embalse
del Sisga, estación Represa Sisga (1939-2006); y, (c) precipitación para el
vallado La Susana, río Bogotá, estación Acandy (1977-2006). .................... 26
Figura 3. Promedio mensual multianual de precipitación para (a) la quebrada
Mancilla, río Bogotá, estación Tesoro (1932-2006); (b) el embalse de Tominé,
estación Guatavita (1941-2006); y, (c) embalse de San Rafael, estación La
Cabaña (1956-dato no disponible)................................................................ 28
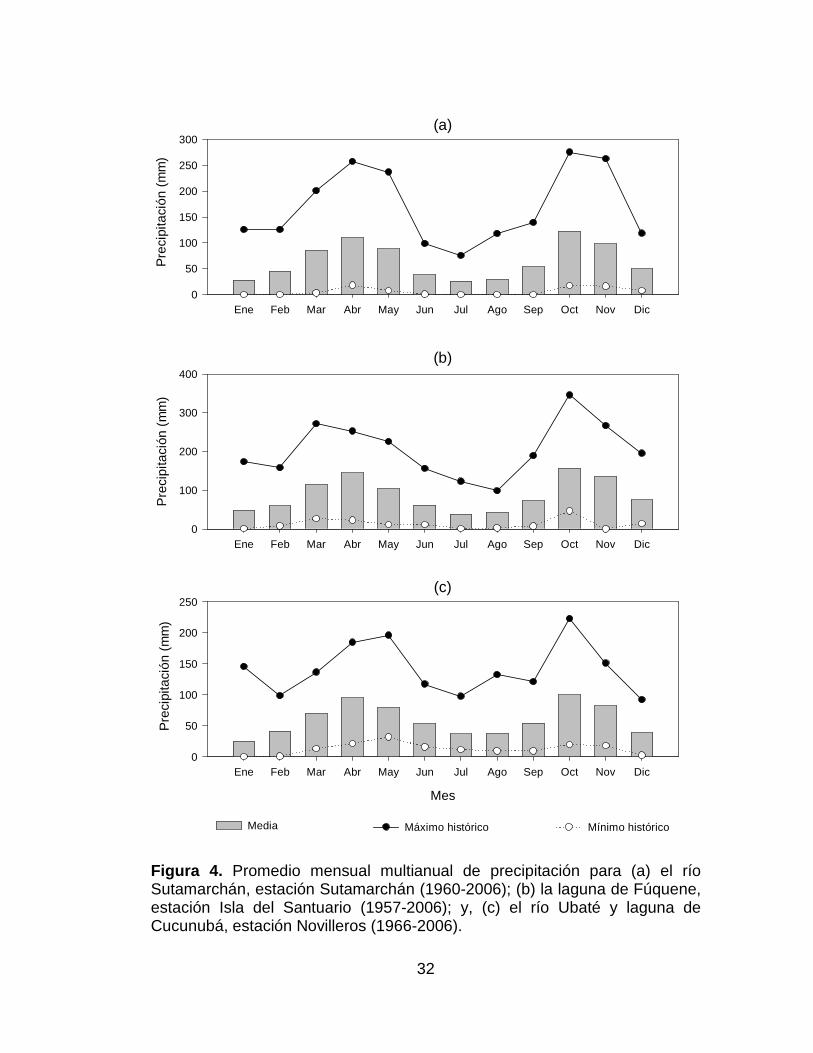
Figura 4. Promedio mensual multianual de precipitación para (a) el río
Sutamarchán, estación Sutamarchán (1960-2006); (b) la laguna de Fúquene,
estación Isla del Santuario (1957-2006); y, (c) el río Ubaté y laguna de
Cucunubá, estación Novilleros (1966-2006). ................................................ 32
Figura 5. Promedio mensual multianual de precipitación para (a) el embalse
El Hato, estación El Hato (1961-2006); y, (b) la laguna de Suesca y quebrada
Mana de Ramos, río Lenguazaque, estación Carrizal (1960-2006).............. 34
Figura 6. Guía para la interpretación del método gráfico de Costello
modificado por Amundsen et al. (1996). ....................................................... 43
Figura 7. Proporción de abundancia de G. bogotensis en cada localidad. ... 49
Figura 8. Porcentaje de abundancia de G. bogotensis por tipo de cuerpo de
agua. LO-BA=lótico corrientes bajas; LE=léntico; LO-MA=lótico corrientes
medialtas. ..................................................................................................... 50
Figura 9. Relación del porcentaje de abundancia de G. bogotensis con la
conductividad. ............................................................................................... 54
Figura 10. Proporción de sexos por intervalo de longitud para la totalidad de
ejemplares de G. bogotensis capturados en las 16 localidades de estudio.. 56

xi
Figura 11. Porcentaje de volumen de los ítems de alimento en las localidades
de (a) laguna de Fúquene, (b) río Ubaté, (c) laguna de Cucunubá, y (d)
embalse El Hato. Para codificación remitirse a la tabla 3. ............................ 62
Figura 12. Porcentaje de volumen de los ítems de alimento en las localidades
de (a) Tibitoc, (b) lago de Tota, caño Los Pozos, (c) lago de Tota, nacimiento
del río Upía, y (d) quebrada Mana de Ramos, río Lenguazaque. Para
codificación remitirse a la tabla 3. ................................................................. 63
Figura 13. Porcentaje de volumen de los ítems de alimento en las localidades
de (a) vallado La Susana, (b) quebrada Mancilla, y (c) embalse de Tominé.
Para codificación remitirse a la tabla 3. ........................................................ 64
Figura 14. Representación gráfica de la dieta de G. bogotensis mediante el
método de Costello modificado por Amundsen et al. (1996). Para codificación
de ítems remitirse a la tabla 3. ...................................................................... 66

xii
ÍNDICE DE ANEXOS
Anexo 1. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la laguna de Fúquene.............................. 99
Anexo 2. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la laguna de Fúquene (continuación). ... 100
Anexo 3. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el río Ubaté............................................ 101
Anexo 4. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el río Ubaté (continuación). ................... 102
Anexo 5. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la laguna de Cucunubá. ........................ 103
Anexo 6. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el embalse El Hato. ............................... 104
Anexo 7. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el embalse El Hato (continuación)......... 105
Anexo 8. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en Tibitoc.................................................... 106
Anexo 9. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el caño Los Pozos, lago de Tota. .......... 107
Anexo 10. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el caño Los Pozos, lago de Tota
(continuación). ............................................................................................ 108
Anexo 11. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el nacimiento del río Upía, lago de Tota.109
Anexo 12. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mana de Ramos, río
Lenguazaque. ............................................................................................. 110

xiii
Anexo 13. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mana de Ramos, río
Lenguazaque (continuación)....................................................................... 111
Anexo 14. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el vallado La Susana, río Bogotá........... 112
Anexo 15. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mancilla, río Bogotá........... 113
Anexo 16. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el embalse de Tominé. .......................... 114
Anexo 17. Gráficas de la relación longitud – peso para la totalidad de
individuos capturados. ................................................................................ 115
Anexo 18. Gráficas de la relación longitud – peso en la laguna de Fúquene.
.................................................................................................................... 116
Anexo 19. Gráficas de la relación longitud – peso en el río Ubaté.............. 117
Anexo 20. Gráficas de la relación longitud – peso en la laguna de Cucunubá.
.................................................................................................................... 118
Anexo 21. Gráficas de la relación longitud – peso en el embalse El Hato. . 119
Anexo 22. Gráficas de la relación longitud – peso en Tibitoc. .................... 120
Anexo 23. Gráficas de la relación longitud – peso en lago de Tota, quebrada
Los Pozos. .................................................................................................. 121
Anexo 24. Gráficas de la relación longitud – peso en lago de Tota, nacimiento
del río Upía. ................................................................................................ 122
Anexo 25. Gráficas de la relación longitud – peso en la quebrada Mana de
Ramos, río Lenguazaque............................................................................ 123
Anexo 26. Gráficas de la relación longitud – peso en el vallado La Susana,
río Bogotá. .................................................................................................. 124
Anexo 27. Gráficas de la relación longitud – peso en la quebrada Mancilla, río
Bogotá. ....................................................................................................... 125
Anexo 28. Gráficas de la relación longitud – peso en el embalse de Tominé.
.................................................................................................................... 126

xiv
Anexo 29. Volumen (mm³) de cada ítem alimentario por localidad............. 127
Anexo 30. Frecuencia de ocurrencia de cada ítem alimentario para las 11
localidades de estudio. ............................................................................... 128

1
Resumen
La abundancia en términos de captura por unidad de esfuerzo, el hábitat, la
proporción de sexos, la relación peso – longitud, y aspectos de la dieta del
carácido Grundulus bogotensis fueron estudiados en 16 localidades
pertenecientes a tres cuencas: Bogotá, Suárez y Tota. La distribución
geográfica de G. bogotensis está restringida al Altiplano Cundiboyacense
colombiano, la especie está inscrita dentro de la categoría “casi amenazada”
y las investigaciones sobre aspectos de su biología y ecología son escasas
además de que se restringen a unas pocas localidades. Se realizaron
muestreos empleando el método de electropesca entre marzo y junio de
2006, donde se colectaron 338 machos, 284 hembras y 130 alevinos. La
especie se capturó en cuerpos de agua con presencia de macrófitas como
Eichhornia crassipes, Egeria densa y Schoenoplectus sp.. El sitio con mayor
número de ejemplares capturados fue la quebrada Mana de Ramos, cuenca
del río Suárez, con 127 individuos representando el 16,71% del total. La
proporción de sexos en algunas localidades fue diferente de 1:1, con una
predominancia de machos en las clases de mayor talla. El parámetro b de la
relación peso – longitud varió entre 2.38 para los alevinos de la Laguna de
Fúquene y 4.64 para las hembras de la Laguna de Cucunubá. El análisis del
contenido estomacal para 281 individuos mostró que la dieta fue diversa y
diferente entre localidades. Los insectos (cf. Chrysomelidae, Chironomidae) y
los microcrustáceos (Hyalella sp., Daphniidae, Cyclopoida) fueron los ítems
más importantes. El papel de los cuerpos de agua lénticos como hábitat y
sitios de alimentación, y la importancia de las condiciones ambientales
locales para los aspectos estudiados en G. bogotensis fueron discutidos.

2
Abstract.
The abundance in terms of catch per unit of effort (CPUE), habitat, sex ratio,
relation between length and weight, and diet aspects were studied for
characid fish Grundulus bogotensis in sixteen different localities distributed in
three basins: Bogotá, Suárez and Tota. Geographic distribution of G.
bogotensis is restricted to the Colombian cundiboyacense plateau and
recently was listed under the category “near threatened”. Researches about
their biological and ecological aspects are scarce; also they are restricted to a
few localities. Samplings were conducted with electrofishing from march to
june in 2006, were captured 338 males, 284 females and 130 alevines. G.
bogotensis was only captured in 11 of the sixteen localities always associated
with macrophytes (Eichhornia crassipes, Egeria densa and Schoenoplectus
sp.). In Mana de Ramos stream, Suárez basin, were captured 127 individuals
(16,71%). The sex ratio in some localities was different that 1:1, with a male
predominance in greater size classes. In the length – weight relationship the
value of the parameter b ranged from 2.38 for alevines in Fúquene lagoon to
4.64 for females in Cucunubá lagoon. The analyses of stomach contents of
281 individuals showed that the diet was different between localities and
diverse, where the insects (cf. Chrysomelidae, Chironomidae) and
microcrustaceans (Hyalella sp., Daphniidae, Cyclopoida) were the most
important items. The role of the lentic habitats as habitat and site of feeding
and local environmental conditions importance for the aspects of biology and
ecology of G. bogotensis was discussed.

3
1. Introducción Dentro de los Andes colombianos se presenta una variabilidad geográfica,
que combinada con factores climáticos y una heterogeneidad geológica,
generan un gran número de hábitats y ecotonos que permiten la presencia
de grupos taxonómicos exclusivos (Mittermeier et al. 1999 en Rodríguez et
al. 2004, Murgueitio y Calle 1999, Chará y Murgueitio 2005).
La pérdida de biodiversidad y transformación del paisaje de la zona andina
ha sido tan drástica en las últimas décadas que ecosistemas completos y un
alto número de especies, entre las que se encuentran aquellas endémicas de
la región, están hoy amenazados de desaparición (Rodríguez et al. 2004).
La guapucha, Grundulus bogotensis (Humboldt, 1821) es una especie
endémica del Altiplano Cundiboyacense. El área de su distribución natural se
encuentra en una de las regiones más intervenidas del territorio colombiano,
producto del desarrollo agrícola e industrial sin las debidas medidas de
control y al mal manejo de los residuos generados por, quizá, la mayor
densidad de población humana del país. Bajo estas circunstancias, la
especie se incluyó en la categoría nacional “casi amenazada” en el 2002, con
lo cual se recomendó realizar estudios que aportaran un mayor conocimiento
sobre su biología y ecología (Álvarez-León et al. 2002).
En la actualidad se reconoce que el estudio de los diferentes aspectos de la
biología y ecología de la ictiofauna continental son claves para establecer
una gestión apropiada, determinar el papel en los ecosistemas acuáticos de
cualquier población de peces sujeta a explotación o bajo algún grado de
amenaza, y por lo tanto, para mantener el equilibrio dentro de un ecosistema
(Lobón-Cerviá 1991, Lizama y Ambrósio 2002).

4
Con este trabajo se pretende aportar al conocimiento básico de la ecología y
biología de G. bogotensis en diferentes localidades ubicadas en el Altiplano
Cundiboyacense. Aportes sobre aspectos como: abundancia, hábitat,
proporción de sexos, relación longitud – peso, y dieta.
El estudio se desarrolló en el marco del proyecto Nº 287: “Variación
morfológica y abundancia relativa de poblaciones de Grundulus bogotensis
(Humboldt, 1821) (Ostariophysi: Characidae), en el altiplano
Cundiboyacense, Colombia”, cofinanciado por la Pontificia Universidad
Javeriana y el Instituto de Investigaciones Alexander von Humboldt.

5
2. Marco teórico y revisión de literatura
2.1 Antecedentes El primer trabajo que abordó aspectos de la biología y ecología de la
guapucha lo llevó a cabo Forero y Garzón (1974). En él se realizó un
seguimiento anual de la población en la represa del Neusa, departamento de
Cundinamarca, generando conocimientos sobre su hábitat, relación longitud
– peso, proporción de sexos, ecología trófica y aspectos reproductivos.
Gompertz (1986) trabajó en detalle la distribución de la guapucha en la
Sabana de Bogotá, incluyendo además algunos comentarios sobre la dieta y
preferencia de hábitat.
Mora et al. (1992) realizan un estudio sobre algunos aspectos biológicos y
ecológicos de los peces del lago de Tota, departamento de Boyacá, entre los
que se incluye a la guapucha. Se abordan características reproductivas como
fecundidad y estados de madurez junto con una valoración de la dieta en 13
ejemplares.
Por otro lado, González et al. (1992) mediante la comparación de cariotipos
de machos y hembras de G. bogotensis demostró que no existe dimorfismo
sexual cromosómico en la especie. Álvarez-León et al. (2002) incluyeron a la
guapucha en el libro rojo de peces dulceacuícolas en la categoría nacional
“casi amenazada”.
Recientemente, León et al. (2005) estudiaron aspectos biométricos de la
especie en el embalse de San Rafael entre los que se incluyen la relación
longitud – peso, mientras que Rivera-Rondón et al. (2007) valoran los efectos
de diferentes especies de macrófitas sobre la distribución espacial de G.
bogotensis.

6
2.2 Sistemática y taxonomía La guapucha, Grundulus bogotensis (Humboldt, 1821), pertenece a la familia
Characidae, diferentes autores la han incluido en la subfamilia
Cheirodontinae por poseer una sola hilera de dientes en la premaxila
(Eigenmann 1915, 1917, 1921, Dahl 1971, Miles 1971, Géry 1977), sin
embargo, dentro de la familia su relación filogenética es desconocida y el
género de posición incierta (Malabarba 1998).
Se consideran sinonimias de la especie a la combinación original, Poecilia
bogotensis Humboldt, 1821, y al sinónimo junior, Ctenocharax bogotensis
Regan, 1907 (Miles 1971, Álvarez-León et al. 2002, Maldonado-Ocampo et
al. 2005, Froese y Pauly 2006).
Este pez no supera los 10 cm de longitud total, su cuerpo es robusto, corto y
está recubierto por escamas de tipo cicloideo excepto en el área predorsal
que se encuentra desnudo. Posee una aleta dorsal con 9 radios; 2 aletas
pectorales con 15 radios cada una; 2 aletas pélvicas con 7 radios cada una;
una aleta anal corta con 21 radios; y una aleta caudal tipo homocerca, con 32
radios. Presenta línea lateral incompleta, con 1 a 6 escamas perforadas
únicamente, ligeramente curva y se extiende hasta un poco después de la
aleta pectoral (Forero y Garzón 1974, Román-Valencia et al. 2003). La vejiga
natatoria está compuesta por dos cámaras, siendo la anterior más pequeña y
de forma circular, la posterior es de tamaño doble y alargada (Posada-
Arango 1909, Forero y Garzón 1974). Los dientes son estrictamente cónicos,
forman una sola hilera (Forero y Garzón 1974, Géry 1977); en el dentario
presenta de 8 a 9 y en el maxilar de 4 a 7 (Román-Valencia et al. 2003). Hay
pérdida de la aleta adiposa (Géry 1977, Román-Valencia et al. 2003).

7
2.3 Distribución Eigenmann (1921) anota que la guapucha se encuentra en la cuenca del río
Magdalena pero su distribución se limita a un río dentro de esta. Eigenmann
(1922, en Forero y Garzón 1974) registra la presencia de la guapucha en los
cuerpos de agua de las altiplanicies de la cordillera Oriental, desde la
vecindad de Bogotá hasta la quebrada de Suaita en el departamento de
Santander, pasando por sitios intermedios como el río Chiquinseco en el
departamento de Boyacá. Fowler (1942) también es algo impreciso y anota
que la especie se distribuye en la cuenca del río Magdalena, río Bogotá, y en
la quebrada de Suaita, Santander. Según Dahl (1971) y Miles (1971) esta
habita en la Sabana de Bogotá y hacia el norte hasta el departamento de
Santander, este último señala que es una de las tres especies nativas de la
Sabana de Bogotá.
Forero y Garzón (1974) apuntan que es una especie endémica de la sabana
de Bogotá y del valle de Ubaté. Mojica (1999) y Álvarez-León et al. (2002)
indican que la especie habita en el Altiplano Cundiboyacense, mientras que
Román-Valencia et al. (2003) señalan que se distribuye en la Sabana de
Bogotá, cuenca del río Magdalena.
Gompertz (1986) realiza un estudio detallado sobre la distribución de la
especie en la sabana de Bogotá y sus alrededores, la registra para los ríos
Bogotá a la altura de los municipios de Chocontá, Suesca, Sesquilé,
Gachancipá y Tocancipá, Neusa a la altura de Zipaquirá, Frío a la altura de
Chía, Siecha a la altura de Guasca, Teusacá a la altura de La Calera,
Subachoque a la altura de Subachoque y Madrid, Serrezuela a la altura de
Madrid, Los Andes a la altura de Facatativa, Checua a la altura de Facatativá
y Nemocón, Susagua a la altura de Cogua, La Vieja a la altura de Nemocón,
Chiquito a la altura de Sopó, y, Balsillas a la altura de Madrid. También la
registra en los siguientes cuerpos de agua lénticos: laguna Pantano Redondo

8
en el municipio de Zipaquirá, laguna Palacios en Ubaté, laguna de
Cucunubá, laguna de Fúquene, laguna de Suesca, embalse de Tominé,
embalse del Sisga, embalse de Facatativá en Facatativa, y, embalse de
Neusa en Zipaquirá. Concluye que a G. bogotensis se le encuentra repartida
en la sabana de Bogotá en cuerpos tanto lénticos como lóticos.
Aunque la distribución natural de la especie estaba restringida únicamente a
un área de drenaje menor a los 30.000 km² (Álvarez-León et al. 2002), ha
sido ampliada producto del transplante a diferentes cuerpos de agua, entre
ellos la laguna de la Cocha y el lago de Tota en los departamentos de Nariño
y Boyacá respectivamente (Alvarado y Gutiérrez 2002).
2.4 Aspectos de la biología y ecología
2.4.1 Hábitat El término genérico “hábitat” se utiliza para describir los componentes físicos
y químicos, en este caso de un cuerpo de agua, que proveen un ambiente
ideal para la colonización de la biota. Al estudiar el hábitat se pueden
proporcionar conceptos unificados que conducen al mismo tiempo hacia
aspectos ecológicos de tolerancia fisiológica, forrajeo y alimentación,
reproducción e historias de vida. En el nivel más fundamental, al seleccionar
un hábitat se está haciendo lo mismo con un área donde los individuos
probablemente encuentran condiciones favorables dentro de sus tolerancias
fisiológicas, como en el caso de la temperatura, salinidad y concentración de
oxígeno. De igual manera, al seleccionar un hábitat se está haciendo lo
mismo con: el grado de exposición a los depredadores a través de la
naturaleza y cantidad de protección dada en el hábitat, y el régimen de
disponibilidad de alimento por la probabilidad de ocurrencia de las presas y
competidores en el hábitat elegido o cerca de él (Rice 2005).

9
La literatura disponible sustenta que la guapucha habita principalmente hacia
las bocas de los afluentes (Forero y Garzón 1974, Mora et al. 1992), canales
de drenaje de los cultivos también denominados “vallados” (Mora et al 1992),
y a la orilla de ambientes lénticos con presencia de macrófitas (Román-
Valencia et al. 2003).
2.4.2 Estimativos poblacionales El uso de la captura por unidad de esfuerzo (CPUE) como medida de la
abundancia es un índice comúnmente empleado en la evaluación de
poblaciones de peces (Haggarty y King 2006). En algunos casos se
considera que la CPUE es parcializada y no es estrictamente proporcional a
la abundancia (Harley et al. 2001), debido a que bajo ciertas situaciones las
tasas de captura pueden permanecer altas si la abundancia disminuye,
fenómeno conocido como hiperestabilidad, y se da por diferentes causas, ya
sea el incremento en la eficiencia de la pesca (capturabilidad), cambios en
los hábitos de los pescadores, o por comportamientos de los peces como la
hiperagregación y la selección del hábitat. Sin embargo, estos factores
pueden, en cierto grado, ser controlados mediante el empleo de métodos
estandarizados que incluyan duración de pesca, localidades y equipos pre-
determinados (Haggarty y King 2006).
Para G. bogotensis no se conocen estimativos de la abundancia de sus
poblaciones (Álvarez-León et al. 2002) y de hecho para ninguna especie
íctica de Colombia se conoce el tamaño poblacional (Mojica et al. 2002).
Existen anotaciones como las de Mora et al. (1992) donde se afirma que la
especie es abundante en el lago de Tota.
2.4.3 Proporción de sexos
La proporción sexual en peces varía a lo largo del ciclo de vida en función de
eventos sucesivos, que actúan de modo distinto sobre los individuos de cada

10
sexo. La mortalidad es uno de los factores que pueden afectar de modo
diferencial esta proporción (Vazzoler 1996).
Forero y Garzón (1974) para una muestra de 770 ejemplares capturados a lo
largo de un año en el caño Guanquica, represa del Neusa – Cundinamarca,
encontraron que el 44,29% eran machos adultos y el 55.71% hembras
adultas, valores que estadísticamente fueron significativos indicando que se
desviaban de la proporción 1:1. En este trabajo encontraron diferencias en la
distribución porcentual de los sexos para diferentes intervalos de longitud, el
porcentaje de hembras disminuye gradualmente hasta llegar al límite
marcado entre los 84 y 88 mm de longitud total, a partir del cual la población
se encuentra representada casi únicamente por machos. Afirman, que los
posibles factores de la resultante proporción de sexos en G. bogotensis
pueden ser la segregación por sexos a través de diferentes períodos a lo
largo del año, diferencias en relación a la mortalidad natural y depredación
de acuerdo al sexo.
Mora et al. (1992) afirman que en el lago de Tota, departamento de Boyacá,
para una muestra de 57 ejemplares colectados durante el mes de noviembre
de 1989 la proporción de hembras y machos es “normal”, aunque no se
habla de la composición para los datos de captura.
2.4.4 Relación longitud – peso
La relación entre el peso y la longitud se usa frecuentemente para comparar
el efecto de factores bióticos y abióticos sobre la salud y bienestar de una
población de peces (Cone 1989), y para estimar el peso correspondiente a
determinada longitud (Froese 2006). El conocimiento de esta relación es una
herramienta importante para el estudio de la biología de cualquier especie
(Lizama y Ambrósio 2002). La relación entre la longitud (L) y el peso (W)
generalmente se expresa mediante la ecuación 1:

11
baLW =
Ecuación 1
donde el parámetro a es el coeficiente de la relación aritmética entre longitud
– peso (Ecuación 1) o el intercepto en la forma logarítmica (Ecuación 3) y el
parámetro b es el exponente de la forma aritmética de la relación longitud –
peso (Ecuación 1) o la pendiente de la regresión lineal en su forma
logarítmica (Ecuación 3) (Froese 2006). El valor de b debería encontrarse
normalmente entre 2.5 y 3.5 (Carlander 1969, en Froese 2006). Cuando b=3,
el aumento del peso se denomina isométrico, significando que peces
pequeños en la muestra bajo consideración tienen la misma forma y
condición que los peces grandes. Cuando b≠3 el aumento del peso se
denomina alométrico, y puede ser positivo (b>3) significando que los peces
grandes han incrementado en peso más que en longitud, o negativo (b<3)
indicando que los peces grandes han cambiado la forma del cuerpo para ser
más alongados o que los pequeños estaban en una mejor condición
nutricional en el momento del muestreo (Froese 2006). En definitiva, b es la
razón de la proporción de crecimiento para la longitud y el peso (Le Cren
1951).
Forero y Garzón (1974) determinaron que en la represa del Neusa los
valores promedio de peso y longitud total para machos eran de 7,9 gr y 76,19
mm; para hembras 5,07 gr y 67,56 mm; y para alevinos 0,28 gr y 23,82 mm.
De acuerdo al coeficiente de regresión obtenido para el total de la población
(b = 3,16) se concluyó que el peso aumentaba un poco más que el cubo de
la longitud, mientras que por tipo de población se obtuvo para machos (b =
2,08), hembras (b = 3,05) y en alevinos (b = 3,4); con lo que concluyen que a
partir de la pérdida de la bolsa vitelina los individuos presentan una tasa de
crecimiento muy elevada.

12
León et al. (2005) determinaron la relación longitud-peso para 144
ejemplares capturados en el embalse de San Rafael encontrando que el
coeficiente de regresión (b = 3,60) indica un crecimiento mayor en peso que
en longitud para los individuos capturados en esta localidad que el reportado
por Forero y Garzón (1974) en la represa del Neusa.
2.4.5 Dieta y amplitud de nicho trófico
La descripción de la estructura trófica en peces y el entendimiento de las
relaciones de alimentación entre organismos son fundamentales para una
efectiva conservación y manejo de la ictiofauna (Luz-Agostinho et al. 2006).
Además, el entender y conocer cómo los organismos utilizan los recursos
permite identificar factores que pueden afectar su distribución y abundancia
(Deus y Petrere-Junior 2003).
Hutchinson (1957, en Whittaker et al. 1973) propuso que las variables físicas
y biológicas que afectan a la especie pueden ser concebidas como un
conjunto de n coordenadas y que para cada una de estas coordenadas
existen valores limitantes dentro de los cuales las especies pueden sobrevivir
y reproducirse. Los rangos de coordenadas dentro de los valores límites
definen un “hipervolumen” n-dimensional, a este hipervolumen se le
denomina nicho fundamental; la respuesta de la población de la especie
dentro de este hipervolumen describe su nicho (Whittaker et al. 1973).
La estimación de la amplitud de nicho es usada principalmente como una
medida inversa de la especialización ecológica. Para su cálculo los datos son
colectados de una serie de estados del recurso que representan una
categoría ecológica importante para la especie, por ejemplo el tipo de
alimento lo que se denominaría amplitud de nicho trófico (Colwell y Futuyma
1971, Smith 1982).

13
El primer estudio dietario para G. bogotensis lo llevó a cabo por Forero y
Garzón (1974) basado en 126 ejemplares colectados en la represa del
Neusa. A partir de este estudio se encontró que no existen cambios en los
hábitos de alimentación relacionados con el aumento de la talla de individuos
entre 20 hasta los 90 mm de longitud total. Se aprecia que la dieta de los
alevinos es menos variada, debido al alto porcentaje formado por crustáceos
como el copépodo Cyclops spp. y Cladocera, así como por estadios larvarios
de dípteros pertenecientes a las familias Chironomidae y Ceratopogonidae.
En individuos adultos los ítems encontrados en mayor proporción fueron las
familias Chironomidae y Ceratopogonidae, con más del 50%, Cyclops spp. e
insectos no identificados. También se registraron huevos de invertebrados,
restos vegetales y detrito, pero todos en menor proporción. Finalmente
clasifican a la guapucha como una especie con alimentación carnívora.
Gompertz (1986) señala que para las 7 especies estudiadas en la sabana de
Bogotá, entre ellas G. bogotensis, la mayoría presentan una dieta carnívora.
Los ítems alimentarios los trabajó a nivel de clase y los registrados en mayor
porcentaje fueron Insecta con 21% y Crustacea con 16%, seguidos por Algae
y dos ítems no identificados (uno de origen animal y otro de origen vegetal)
cada uno con el 10%.
Mora et al. (1992), analizaron el contenido estomacal de 13 individuos
colectados en noviembre de 1989 en el lago de Tota, y determinaron que la
dieta está dominada por el copépodo Boeckella gracilis con el 95%, pero
aclaran que la población adulta consume con preferencia el anfípodo Hyalella
sp. y los cladóceros Eurycercus sp., Neobosmina sp., Graptoleberis sp. y
Chydorus sp., así como larvas de Chironomidae, pupas de Diptera y el
bivalvo del género Pisidium.

14
Pantoja-Agreda y Pantoja-Agreda (2003) determinaron a partir del contenido
de 64 estómagos que G. cochae, una especie emparentada con G.
bogotensis, es carnívora consumidora directa de zooplancton que presenta
una dieta poco diversa con predominio de microcrustáceos, principalmente
del copépodo Diaptomus sp. (56,10%), los cladóceros Bosmina sp. (24.15%)
y Chydorus sp. (19,09%).
2.5 Grado de amenaza y medidas de conservación
Es una estimación conservadora que el 20% de las especies de peces
dulceacuícolas del mundo están prontas a desaparecer o a declinar (Moyle y
Leidy 1992, en Magurran y Phillip 2001). Tradicionalmente se ha asumido
que la pérdida de especies está acompañada por un cambio en la
distribución de la abundancia de log normal a serie geométrica, lo que en
otras palabras significa que asambleas no perturbadas, donde la abundancia
de especies está relativamente distribuida de manera uniforme, son
reemplazadas por unas con baja riqueza de especies con alta dominancia.
Además, otro de los inconvenientes acerca de la pérdida de biodiversidad es
que el ecosistema se ve afectado funcionalmente (Magurran y Phillip 2001).
Álvarez-León et al. (2002) ubican a la guapucha en la categoría nacional casi
amenazada (NT) por considerar que en un futuro cercano podría pertenecer
a la categoría vulnerable (VU) que indica que la especie afronta o estaría
muy cerca de afrontar un deterioro poblacional a mediano plazo.
Gompertz (1986) recomienda realizar estudios ictiológicos con miras a
repoblar los cuerpos de agua de la sabana de Bogotá con especies nativas
con problemas de conservación como G. bogotensis, o por los menos
estudios para mantener sus poblaciones. También apunta que se deben
realizar investigaciones sobre la dinámica de la población de cada especie
íctica de la sabana.

15
Para G. bogotensis no existen medidas de conservación pero se
recomiendan la protección y restauración de su hábitat natural, y el apoyo a
programas de investigación científica que aporten conocimientos de la
biología básica y profundicen los ya existentes (Álvarez-León et al. 2002).
3. Formulación del problema y justificación.
3.1 Formulación del problema
La guapucha, G. bogotensis, es un pez endémico del Altiplano
Cundiboyacense que se encuentra en la categoría nacional “casi
amenazada” (NT). Actualmente su hábitat natural presenta procesos
avanzados de deterioro ambiental generados principalmente por el
vertimiento de aguas no tratadas propias de labores agropecuarias e
industriales, sumado a las desechos producidos en las cabeceras
municipales (Álvarez-León et al. 2002, CAR 2001).
Es poco lo que se conoce, y además se restringe a unas pocas localidades,
sobre la abundancia, hábitat, proporción de sexos, relación longitud – peso, y
aspectos de la dieta en G. bogotensis. Los diferentes estudios abordan
elementos como la biología general en la represa del Neusa (Forero y
Garzón 1974), distribución en la Sabana de Bogotá (Gompertz 1986),
aspectos reproductivos y dieta en el lago de Tota (Mora et al. 1992), relación
longitud – peso en el embalse de San Rafael (León et al. 2005) y distribución
espacial en la laguna de Fúquene (Rivera-Rondón et al. 2007). Por lo
anterior se ha recomendado el desarrollo de un mayor número de
investigaciones no sólo para ampliar el conocimiento existente sino también
para el diseño de programas e implementación de acciones encaminadas al
manejo y conservación de la especie (Álvarez-León et al. 2002).

16
3.2 Preguntas de investigación
Para este trabajo se proponen las siguientes preguntas de investigación:
¿Se presentan variaciones en los valores de abundancia de G. bogotensis en
las localidades de estudio, y se podrían relacionar estas con el tipo de cuerpo
de agua o alguno de los parámetros fisicoquímicos medidos?
¿Hay diferencias en la proporción de sexos y en el valor del parámetro b de
la relación longitud-peso entre machos, hembras y alevinos de G. bogotensis
en las localidades de estudio?
¿Cambia la composición y abundancia de la dieta de G. bogotensis en las
localidades estudiadas?
3.3 Justificación
Dentro de los Andes colombianos se presenta una variabilidad geográfica,
que combinada con factores climáticos y una heterogeneidad geológica,
generan un gran número de hábitats y ecotonos que permiten la presencia
de grupos taxonómicos exclusivos y gran número de especies con rangos de
distribución que están restringidos a elevaciones específicas u otras
unidades biogeográficas (Mittermeier et al. 1999 en Rodríguez et al. 2004,
Murgueitio y Calle 1999, Chará y Murgueitio 2005).
La pérdida de biodiversidad y transformación del paisaje de la zona andina
ha sido tan drástica en las últimas décadas que ecosistemas completos y un
alto número de especies, entre las que se encuentran aquellas endémicas de
la región, están hoy amenazados de desaparición y por la tanto trayendo
consecuencias para los múltiples bienes y servicios que la región brinda para
el desarrollo de las actividades sociales y económicas de las comunidades
locales (Rodríguez et al. 2004), llegando de esta manera a ubicarse como la

17
primera región en el listado global de prioridades para conservación
(Mittermeier et al. 1999, en Rodríguez et al. 2004).
En la actualidad la región andina colombiana es la más poblada del país, y
soporta una gran actividad agrícola y ganadera. Se estima que el 70% de la
cobertura boscosa ha sido transformada y que el 80% de esta área
deforestada está ocupada por cultivos para consumo humano y pasturas
destinadas a actividades ganaderas (Murgueitio e Ibrahim 2001, Chará y
Murgueitio 2005).
Los impactos más comunes sobre ecosistemas acuáticos que se encuentran
inmersos en una matriz bajo uso agrícola y pecuario son la modificación de
las riberas y los canales, regulación del flujo, fragmentación, contaminación
química (metales pesados, pesticidas, fertilizantes), y polución orgánica.
Todas estas alteraciones están ligadas a una degradación ecológica
extensiva haciéndolos no sostenibles en la provisión de bienes y servicios
(Ej.: descenso en la disponibilidad y calidad del agua); y, generando cambios
en la distribución, composición y estructura de la biota acuática (Watzin y
McIntosh 1999, Anbumozhi et al. 2005, Tejerina-Garro et al. 2005).
En el contexto regional, los cuerpos de agua del Altiplano Cundiboyacense
presentan fuertes procesos de deterioro ambiental que influyen sobre las
diferentes comunidades bióticas que los habitan, y posiblemente han
afectado a las poblaciones de G. bogotensis restringiéndolas a unos pocos
sectores aún no modificados de forma importante, motivando así a que la
especie recientemente se haya incluido en la categoría nacional casi
amenazada (NT) (Álvarez-León et al. 2002).
En la actualidad se reconoce que el estudio de los diferentes aspectos de la
biología y ecología de la ictiofauna continental son claves para establecer

18
una gestión apropiada, determinar el papel en los ecosistemas acuáticos de
cualquier población de peces sujeta a explotación o bajo algún grado de
amenaza, y por lo tanto, para mantener el equilibrio dentro de un ecosistema
(Lobón-Cerviá 1991, Lizama y Ambrósio 2002).
4. Objetivos
4.1 Objetivo general
Estudiar diversos aspectos de la biología y ecología de G. bogotensis en
diferentes localidades del Altiplano Cundiboyacense, Colombia.
4.2 Objetivos específicos
• Estimar la abundancia de G. bogotensis para cada una de las localidades.
• Caracterizar el hábitat de la guapucha en las diferentes localidades de
estudio.
• Determinar la proporción de sexos en cada una de las localidades de
estudio.
• Estimar la relación longitud-peso, considerando machos, hembras y
alevinos, de G. bogotensis en cada una de las localidades de estudio.
• Identificar los hábitos alimentarios de la especie en las localidades
estudiadas.

19
5. Área de Estudio
Los muestreos se realizaron en 16 localidades diferentes, ubicadas en el
Altiplano Cundiboyacense, entre los meses de marzo a junio de 2006. De
ellas, 6 hacían parte de la cuenca del río Bogotá, 8 a la cuenca del río
Suárez y 2 a la cuenca del lago de Tota.
5.1 Altiplano Cundiboyacense
El Altiplano Cundiboyacense está localizado en los departamentos de
Cundinamarca y Boyacá, Colombia, tiene una extensión de 19.529 km² de
los cuales 9.510 pertenecen al departamento de Cundinamarca, 8.830,8 al
departamento de Boyacá y 1.188,2 a la ciudad de Bogotá. Geográficamente
se encuentra entre los 3º45’13’’ y los 6º03’13’’ de latitud norte y entre los
72º39’40’’ y los 74º52’30’’ de longitud oeste (IGAC 1984).
La región presenta dos grandes paisajes: uno de altiplanicies y valles,
conformado por la Sabana de Bogotá, la altiplanicie de Tunja y los valles de
Ubaté y Chiquinquirá, y otro paisaje montañoso de relieve quebrado a
escarpado, constituido por las cuencas altas del altiplano y por las vertientes
oriental y occidental de la cordillera oriental (IGAC 1984). Sus ríos tributarios
son: Sumapaz, Bogotá, los tres ríos Seco, Negro, Minero, Ubaté y Suárez.
Las vertientes hacia los llanos orientales alimentan las cuencas del río
Guaviare y del Meta (CAR 2002).
5.1.1 Temperatura media anual
En el altiplano drenado por los ríos Bogotá, Ubaté y Suárez, se registran
temperaturas entre los 12º C y 14º C, pero que descienden en el sector
suroriental de Bogotá, sobre el páramo de Sumapáz, alcanzando valores
inferiores a 10º C. En la mayor parte del territorio, en enero y en julio se
presentan las temperaturas promedio más altas, mientras que en abril y
noviembre se registran las menores (CAR 2002).

20
5.1.2 Clima
La Sabana de Bogotá, y la planicie de los ríos Ubaté y Suárez presentan,
siguiendo la clasificación de Caldas-Lang, un clima semihúmedo, intercalado
por una franja árida que se extiende a lo largo del río Bogotá y por las
lagunas de Suesca y de Cucunubá, además cruza sectores de los municipios
de Soacha, Cucunubá, Ubaté y Fúquene. En los cerros que rodean la
Sabana de Bogotá y las cuencas de los ríos Ubaté y Suárez, por los flancos
occidental y oriental, el clima es de páramo bajo semihúmedo (CAR 2002).
5.1.3 Aspectos geológicos
Las rocas que afloran en las cuencas de la laguna de Fúquene y del río
Bogotá son resultado de la acumulación y la litificación de sedimentos que
tienen edades entre los 200 millones de años y el presente. Estas rocas han
sido deformadas por los movimientos regionales de las placas, que han
causado su emersión y su erosión, originando los detritos que se han
depositado en las cuencas formadas en la parte central y en las vertientes de
la cordillera oriental. A causa de este proceso los cursos de los ríos que
drenan esta región han sido, de alguna forma, controlados estructuralmente
(CAR 2002).
La estructura geológica de la región, está formada por rocas sedimentarias
de diferentes edades, que van desde el jurásico hasta el cuaternario reciente,
sin embargo, su mayor extensión corresponde a formaciones del terciario.
Las diferentes formaciones geológicas presentan múltiples materiales
rocosos, mezclados o en capas alternas, constituidos por areniscas
consolidadas o friables de grano grueso a fino, lutitas, limolitas, shales,
cherts (plaeners), calizas y arcillas negras, pardas y rojizas (IGAC 1984).
En una considerable extensión, las formaciones geológicas sedimentarias
fueron cubiertas por mantos de cenizas volcánicas provenientes de la

21
cordillera central, siendo más importante su acumulación hacia el flanco
occidental de la cordillera oriental principalmente entre Silvania, Facatativá,
Cota, Subachoque y Chiquinquirá (IGAC 1984).
5.2 Cuenca hidrográfica del río Bogotá
La planicie fluviolacustre del río Bogotá se ubica en el centro del
departamento de Cundinamarca, en una franja alargada con dirección
noreste – suroeste. La variación altitudinal oscila entre lo 2600 msnm en la
zona plana y 3000 msnm en los cerros que la circundan. Tiene una superficie
de 569.460 hectáreas. El río tiene una longitud de 335 km; en Chocontá su
caudal medio es de 2,6 m³/s; en Alicachín, al límite de la Sabana de Bogotá
por el sur, llega a 29 m³/s, y en su desembocadura en el río Magdalena
asciende a 52 m³/s. El caudal medio anual (oferta natural global) de la
cuenca es de 52,56 m³/s (CAR 2002).
Por sus características topográficas y poblacionales, asociadas al
aprovechamiento y a la degradación del recurso hídrico, se definen tres
sectores en la cuenca: alta, media y baja. Las partes alta y media, sectores
donde se llevó a cabo parte de los muestreos, se extienden desde el
nacimiento del río hasta el sector de Alicachín (CAR 2002). La parte media
de la cuenca comprende la denominada Sabana de Bogotá, que es la
cuenca de un antiguo lago de alta montaña localizado en el centro del
departamento de Cundinamarca, con una superficie de 1450 km² (IGAC
1974, en: Gompertz 1986).
5.2.1 Distribución temporal de la precipitación
El río Bogotá en su cuenca alta muestra un comportamiento monomodal
debido a la influencia del clima de la Orinoquia, en el resto de la cuenca se
presenta un régimen bimodal determinado por el paso de la Zona de
Confluencia Intertropical – ZCTI, que es un franja de la atmósfera con bajas

22
presiones, que se sitúa aproximadamente paralela al Ecuador, dando origen
a la formación de grandes masas nubosas y de abundantes precipitaciones
(CAR 2002).
La zona con régimen unimodal tiene un máximo de 110 mm en julio, un
mínimo de 20 mm en enero y un total anual de 818 mm. La otra parte de la
cuenca, con régimen bimodal, presenta precipitaciones máximas en el
segundo periodo lluvioso del año; en las partes media y alta se registran
durante el mes de octubre, valores máximos que oscilan entre los 100 y 130
mm, mientras que en la parte baja de la cuenca los máximos se dan durante
noviembre, con valores que alcanzan los 200 mm (CAR 2002).
En las partes alta y media de la cuenca se observan valores mínimos durante
el primer periodo seco, con oscilaciones entre los 20 y los 40 mm en enero
(CAR 2002).
5.2.2 Distribución espacial de la precipitación
En las partes alta y media se presentan dos zonas características: 1) la zona
húmeda y montañosa, con valores anuales entre 1000 y 1400 mm que
pertenece a los cerros que bordean la Sabana de Bogotá, por el oriente y por
el occidente, sobre los cuales se acumulan las masas de humedad durante
todo el año; y 2) la zona correspondiente a la Sabana de Bogotá, en donde
se registran precipitaciones anuales inferiores a los 1000 mm. Hay sectores
muy secos en los alrededores del municipio de Suesca y en una franja
aledaña al río Bogotá, en las poblaciones de Soacha, Mosquera, y Madrid,
con valores cercanos a los 600 mm. En el sector central de la Sabana, desde
Tocancipá hasta Funza y en el área del municipio de Sesquilé, se localizan
núcleos de precipitación de 600 mm a 800 mm; en el resto de la sabana los
valores son similares: entre 800 y 1000 mm anuales (CAR 2002).

23
5.2.3 Localidades de muestreo cuenca del río Bogotá
Fueron seis las localidades estudiadas en las partes media y alta de la
cuenca del río Bogotá. Se inició en Tibitoc el 8 de abril de 2006 y finalizó el
13 de junio de 2006 en el embalse de San Rafael (Figura 1).
5.2.3.1 Tibitoc
El muestreo se realizó el 8 de abril de 2006, la localización exacta fue
04º58'38.2''N y 73º58'01''O, municipio de Tocancipá, Cundinamarca, a una
altura de 2563 m.s.n.m. (Tabla 1). El río Neusa transporta el caudal
descargado del embalse de Neusa más los aportes de la cuenca que se
suman al caudal no regulado del río Checua. Pocos metros aguas abajo de
la desembocadura del río está localizada la compuerta de El Espino con la
que se regula la captación de agua con el propósito de ser conducida a la
planta de tratamiento de Tibitoc y someterla al proceso de potabilización
(IGAC 1984, CAR 2002).
Tabla 1. Localidades de muestreo en la parte media y alta del río Bogotá.
Localidad Cuenca Municipio Departamento Latitud N Longit ud WAltura
(msnm)
Tibitoc Bogotá Tocancipá CUN 04º58'38.2'' 73º58'01'' 2563
Embalse del Sisga Bogotá Chocontá CUN 05º05'28,5'' 73º43'26,3'' 2688
R. Bogotá-La Susana Bogotá Sesquilé CUN 05º03'50,3'' 73º49'51,8'' 2573
Quebrada Mancilla Bogotá Facatativa CUN 04º51'09,7'' 74º21'01,8'' 2582
Embalse de Tominé Bogotá Guatavita CUN 04º59'10,7'' 73°4 9'09'' 2622
Embalse San Rafael Bogotá La Calera CUN 04º42'32,9'' 73°59' 55'' 2799
24
Figura 1. Localización de los sitios de muestreo en el Altiplano
Cundiboyacense.

25
5.2.3.2 Embalse del Sisga
El muestreo se realizó el 17 de mayo de 2006, la localización exacta fue
05º05'28,5''N y 73º43'26,3''O, municipio de Chocontá, Cundinamarca, a una
altura de 2688 m.s.n.m. El embalse se construyó en el año de 1951, tiene
como única función la de regular los afluentes que controla, siendo el más
importante el río San Francisco. Tiene una capacidad de almacenamiento de
102 millones de metros cúbicos (Mm³) y regula 2,6 m³/s (Tabla 1, Figura 2a y
1b) (IGAC 1984, CAR 2002).
5.2.3.3 Vallado La Susana – Río Bogotá
El muestreo se realizó el 18 de mayo de 2006, la localización exacta fue
05º03'50,3''N y 73º49'51,8''O, municipio de Sesquilé, Cundinamarca, a una
altura de 2573 m.s.n.m. (Tabla 1,Figura 2c).
5.2.3.4 Quebrada Mancilla
El muestreo se realizó el 20 de mayo de 2006, la localización exacta fue
04º51'09,7''N y 74º21'01,8''O, municipio de Facatativa, Cundinamarca, a una
altura de 2582 m.s.n.m. La quebrada Mancilla es un afluente del río Bojacá,
pertenece a la cuenca del río Balsillas, que a su vez está conformada por las
subcuencas de los ríos Balsillas, Subachoque y Bojacá. La cuenca se
localiza al oeste de la Sabana de Bogotá con áreas de los municipios de
Funza, Mosquera, Madrid, Bojacá, El Rosal, Subachoque y Facatativa; tiene
663,34 km² (Tabla 1,Figura 3a) (CAR 2002).
26
(b)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Pre
cipi
taci
ón (
mm
)
0
50
100
150
200
250
300
Máximo histórico
(a)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Vol
umen
(m
iles
de m
³)
0
20000
40000
60000
80000
100000
120000
(c)
Mes
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Pre
cipi
taci
ón (
mm
)
0
50
100
150
200
250
300
350
Mínimo históricoMedia
Figura 2. Promedio mensual multianual de (a) volumen del embalse del Sisga, estación Represa Sisga (1952-2006); (b) precipitación para el embalse del Sisga, estación Represa Sisga (1939-2006); y, (c) precipitación para el vallado La Susana, río Bogotá, estación Acandy (1977-2006).

27
5.2.3.5 Embalse de Tominé
El muestreo se realizó el 23 de mayo de 2006, se localizó a los 04º59'10,7''N
y 73°49'09''O, municipio de Guatavita, Cundinamarca , a una altura de 2622
m.s.n.m. El punto de muestreo hace parte de los predios del club náutico
“Refugio de Tominé”. El embalse se construyó en el año de 1962, posee una
capacidad de almacenamiento de 690 Mm³, y regula 3,9 m³/s. Además de
regular los afluentes de los ríos Aves, Chipatá y Siecha, regula el flujo del río
Bogotá en forma directa, bombeando una parte de su caudal para
almacenarlo (Tabla 1,Figura 3b) (IGAC 1984, CAR 2002).
5.2.3.6 Embalse de San Rafael
El muestreo se realizó el 13 de junio de 2006, la localización exacta fue
04º42'32,9''N y 73°59' 55''O, municipio de La Caler a, Cundinamarca, a una
altura de 2799 m.s.n.m. El embalse de San Rafael, con capacidad total de 75
Mm³, entró en operación en febrero de 1997; su función principal es la de
abastecer el sistema cuando se requiera suspender las descargas del
embalse de Chuza, a la conducción Chuza-Wiesner, y cuando se necesite
controlar las crecientes del río Teusacá (Tabla 1, Figura 3c) (CAR 2002).

28
(a)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
300
(b)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
Máximo histórico
(c)
Mes
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
300
Mínimo históricoMedia
Pre
cipi
taci
ón (
mm
)P
reci
pita
ción
(m
m)
Pre
cipi
taci
ón (
mm
)
Figura 3. Promedio mensual multianual de precipitación para (a) la quebrada Mancilla, río Bogotá, estación Tesoro (1932-2006); (b) el embalse de Tominé, estación Guatavita (1941-2006); y, (c) embalse de San Rafael, estación La Cabaña (1956-dato no disponible).

29
5.3 Cuenca hidrográfica del río Suárez
La cuenca de primer orden de los ríos Ubaté y Suárez se ubica al norte del
departamento de Cundinamarca y al suroccidente de Boyacá; tiene una
superficie de 196.910 hectáreas y en Cundinamarca comprende los
municipios de Tausa, Sutatausa, Ubaté, Carmen de Carupa, Fúquene, Susa,
Guachetá, Simijaca, Lenguazaque y Cucunubá; y en Boyacá los municipios
de Chiquinquirá, Caldas, San Miguel de Sema, Ráquira, y Saboya (CAR
2002).
La planicie fluviolacustre de los ríos Ubaté y Suárez tiene su centro en la
laguna de Fúquene, con dirección sur – norte, relieve plano, cóncavo y de
valles de inundación, que son drenados por los ríos Ubaté, Suárez y sus
afluentes; altitudinalmente varía entre los 2600 y los 2800 msnm. Los
principales accidentes hidrográficos se localizan en la parte plana y
corresponden a las lagunas de Fúquene, Palacios, Cucunubá y Suesca
(CAR 2002).
5.3.1 Distribución temporal de la precipitación
Esta cuenca presenta bajas precipitaciones, con promedios anuales
inferiores a los 1000 mm en la zona plana y régimen bimodal. Las máximas
se dan durante el segundo periodo húmedo, en octubre hay lluvias con
valores cercanos a los 100 mm en la parte alta, los que se incrementan hasta
alcanzar los 160 mm en la zona de Saboya, parte baja de la cuenca. Los
valores mínimos de precipitación se presentan en enero con registros
cercanos a los 30 mm (CAR 2002).
5.3.2 Distribución espacial de la precipitación
Por su localización geográfica presenta un comportamiento similar al de la
Sabana de Bogotá, aunque un poco más húmeda. En la zona montañosa del
occidente de la planicie fluviolacustre la precipitación anual supera los 1000

30
mm, en el resto del terreno los valores fluctúan entre 800 mm en el área de
Cucunubá y el valle del río Ubaté y 1200 mm al norte de la laguna de
Fúquene. Finalmente, se presenta una zona de altas precipitaciones en la
cuenca del río Suárez, en el sector de Saboya, allí la precipitación alcanza
los 2400 mm al año (CAR 2002).
5.3.3 Localidades de muestreo cuenca del río Suárez
Fueron ocho las localidades estudiadas en la cuenca hidrográfica del río
Suárez. Se inició en el río Sáchica el 31 de marzo de 2006 y finalizó el 17 de
mayo de 2006 en la quebrada Mana de Ramos (Figura 1).
5.3.3.1 Río Sáchica
El muestreo se realizó el 31 de marzo de 2006, la localización exacta fue
05º36'48.5''N y 73º33'41.4''O, municipio de Sáchica, Boyacá, a una altura de
2115 m.s.n.m. (Tabla 2).
Tabla 2. Localidades de muestreo en la cuenca del río Suárez y cuenca del lago de Tota.
Localidad Cuenca Municipio Departamento Latitud N Longit ud WAltura
(msnm)
Río Sáchica Suárez Sáchica BOY 05º36'48.5'' 73º33'41.4'' 2115
Río Sutamarchán Suárez Sutamarchán BOY 05º37'09.2'' 73º37'13.1'' 2109
Laguna Fúquene Suárez Ubaté CUN 05º28'05.4'' 73º46'20.7'' 2551
Río Ubaté Suárez Ubaté CUN 05º23'00.8'' 73º45'53.9'' 2559
Laguna Cucunubá Suárez Cucunubá CUN 05º17'01.4'' 73º47'11.9'' 2563
Embalse El Hato Suárez Carmen de Carupa CUN 05º17'33.8'' 73º54'31.2'' 2867
Tota-Q. Los Pozos Tota Aquitania BOY 05º33'35,5'' 72º53'08,8'' 3037
Tota - Nac.Upía Tota Aquitania BOY 05º29'29'' 72º56'0,97'' 3042
Laguna de Suesca Suárez Suesca CUN 05º11'44,5'' 73º46'29,7'' 2880
Q. Mana de Ramos Suárez Suesca CUN 05º12'08,9'' 73º44'38,7'' 2842

31
5.3.3.2 Río Sutamarchán
El muestreo se realizó el 31 de marzo de 2006, la localización exacta fue
05º37'09.2''N y 73º37'13.1''O, municipio de Sutamarchán, Boyacá, a una
altura de 2109 m.s.n.m. (Tabla 2,Figura 4a).
5.3.3.3 Laguna de Fúquene
El muestreo se realizó el 1 de abril de 2006, la localización exacta fue
05º28'05.4''N y 73º46'20.7''O, municipio de Ubaté, Cundinamarca, a una
altura de 2551 m.s.n.m. (Tabla 2, Figura 4b).
5.3.3.4 Río Ubaté
El muestreo se realizó el 2 de abril de 2006, la localización exacta fue
05º23'00.8''N y 73º45'53.9''O, municipio de Ubaté, Cundinamarca, a una
altura de 2559 m.s.n.m. El río Ubaté nace en el municipio de Carmen de
Carupa, por la confluencia de los ríos Hato y La Playa, y sus principales
afluentes son los ríos Suta y Lenguazaque; recorre 42,7 km. El área de
drenaje alcanza 778 km²; es el principal afluente de la laguna de Fúquene y
el eje de recolección de aguas en el valle (Tabla 2, Figura 4c) (CAR 2002).
5.3.3.5 Laguna de Cucunubá
El muestreo se realizó el 2 de abril de 2006, la localización exacta fue
05º17'01.4''N y 73º47'11.9''O, municipio de Cucunubá, Cundinamarca, a una
altura de 2563 m.s.n.m. La laguna de Cucunubá se encuentra en una zona
de bajas precipitaciones, está conectada por canales con el río Lenguazaque
y por consiguiente con la laguna de Fúquene, además cuenta con un canal
perimetral. Sus aguas se utilizan principalmente para riego (Tabla 2, Figura
4c) (Van der Hammen 2001).
32
(a)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
300
Media
(b)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Pre
cipi
taci
ón (
mm
)
0
100
200
300
400
Máximo histórico
(c)
Mes
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
Mínimo histórico
Pre
cipi
taci
ón (
mm
)P
reci
pita
ción
(m
m)
Figura 4. Promedio mensual multianual de precipitación para (a) el río Sutamarchán, estación Sutamarchán (1960-2006); (b) la laguna de Fúquene, estación Isla del Santuario (1957-2006); y, (c) el río Ubaté y laguna de Cucunubá, estación Novilleros (1966-2006).

33
5.3.3.6 Embalse El Hato
El muestreo se realizó el 3 de abril de 2006, la localización exacta fue
05º17'33.8''N y 73º54'31.2''O, municipio de Carmen de Carupa,
Cundinamarca, a una altura de 2867 m.s.n.m. Fue construido entre los años
de 1990-1993 para el abastecimiento del acueducto del municipio de Cármen
de Carupa y con el propósito de controlar las periódicas crecientes del río
Ubaté para disminuir su incidencia en las fluctuaciones de los niveles de
agua, especialmente en la laguna de Fúquene. Tiene una capacidad de
almacenamiento de 14,4 Mm³ de agua y ocupa un área de 100 ha (Tabla 2,
Figura 5a) (CAR 2002).
5.3.3.7 Laguna de Suesca
El muestreo se realizó el 17 de mayo de 2006, la localización exacta fue
05º11'44,5''N y 73º46'29,7''O, municipio de Suesca, Cundinamarca, a una
altura de 2880 m.s.n.m. Esta laguna es una cuenca cerrada y forma parte de
la cuenca mayor de los ríos Bogotá y Suárez y de la subcuenca laguna de
Suesca, según el acuerdo 09 de 1979 de la CAR (Tabla 2, Figura 5b)
(Escobar 1991).
5.3.3.8 Quebrada Mana de Ramos
El muestreo se realizó el 17 de mayo de 2006, la localización exacta fue
05º12'08,9''N y 73º44'38,7''O, municipio de Suesca, Cundinamarca, a una
altura de 2842 m.s.n.m. Pertenece a la subcuenca del río Lenguazaque
(Tabla 2, Figura 5b).

34
(a)
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic0
50
100
150
200
250
300
(b)
Mes
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Pre
cipi
taci
ón (
mm
)
0
50
100
150
200
250
MediaMáximo históricoMínimo histórico
Pre
cipi
taci
ón (
mm
)
Figura 5. Promedio mensual multianual de precipitación para (a) el embalse El Hato, estación El Hato (1961-2006); y, (b) la laguna de Suesca y quebrada Mana de Ramos, río Lenguazaque, estación Carrizal (1960-2006).
5.4 Cuenca hidrográfica de lago de Tota
Se encuentra ubicada en la cordillera oriental, en el departamento de Boyacá
en los municipios de Aquitania, Tota, Cuítiva y Sogamoso. El área de la
cuenca es de 20.100 ha de las cuales 6.000 corresponden al espejo de agua
del lago (Roa 1998).

35
Hacia el sur y el oriente se encuentra rodeada por montañas de elevación
media de 3800 m. al occidente se encuentra la zona interandina media de la
cordillera. Los mayores aportes de volumen de agua lo realizan el río Tobal y
Olarte, y las quebradas Mugre, Pozos y Hato Laguna (Roa 1998).
La temperatura media mensual oscila entre 9.9 y 11.7ºC; los vientos que
recorren a la cuenca en los diferentes meses del año corresponden al
sistema de los Alisios. La precipitación promedio anual en el municipio de
Aquitania es de 945 mm (Roa 1998).
Las aguas son de buena calidad y se captan para uso doméstico, riego e
industria. El territorio está ocupado por extensos cultivos de cebolla en el
área adyacente al lago a una altura de 3050 m.s.n.m., por otros cultivos
hasta los 3300 m., y pastoreo en terrenos de protección llegando en
ocasiones hasta los 3600 m. (Roa 1998).
5.4.1 Localidades de muestreo cuenca del lago de To ta
Fueron dos las localidades estudiadas en la cuenca del lago de Tota. Ambos
muestreos, en el caño Los Pozos y el nacimiento del río Upía, se realizaron
el 16 de mayo de 2006 (Figura 1).
5.4.1.1 Lago de Tota – Caño Los Pozos
El muestreo se realizó el 16 de mayo de 2006, la localización exacta fue
05º33'35,5''N y 72º53'08,8''O, municipio de Aquitania, Boyacá, a una altura
de 3037 m.s.n.m. (Tabla 2).
5.4.1.2 Lago de Tota – Nacimiento del río Upía
El muestreo se realizó el 16 de mayo de 2006, la localización exacta fue
05º29'29''N y 72º56'0,97''O, municipio de Aquitania, Cundinamarca, a una

36
altura de 3042 m.s.n.m. En la región se le conoce con el nombre de “El
Desaguadero” (Tabla 2).
6. Materiales y Métodos
6.1 Selección de localidades
La selección de las localidades de captura se realizó con ayuda de un mapa
del Altiplano Cundiboyacense escala 1:400.000 (fuente: CAR 2002). En
campo, la localización exacta del punto de muestreo (latitud, longitud y
altura) se georreferenció con ayuda de una unidad de GPS (Global
Positioning System) marca Garmin, modelo GPS 72.
La información de aspectos generales y clima del área de estudio se obtuvo
mediante revisión y análisis de documentos publicados para el área.
Además, se analizaron datos climáticos provenientes de diferentes clases de
estaciones (pluviográfica, pluviométrica, meteorológica especial,
climatológica principal, limnigráfica, y limnimétrica) administradas por la
Corporación Autónoma Regional de Cundinamarca (CAR), Corporación
Autónoma Regional de Boyacá (CORPOBOYACA), y el Instituto de
Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM).
6.2 Captura y preservación de los peces
La captura de los peces se realizó mediante un equipo de pesca eléctrica
portátil con peso aproximado de 25 kg. Consta de un generador marca
Honda a gasolina, de potencia máxima de 2 kw, potencia de salida de 1,6
kw, voltaje de 120V y frecuencia de 60 s.f.; un cátodo terminado en una rejilla
metálica de 32x33 cm; un ánodo o nasa compuesta de un aro en cobre de
diámetro interno de 46 cm y externo de 49 cm, al cual se le adapta una malla
de 90 cm, con entre-nodo de 1 mm, y un mango de 115 cm con un cable de

37
50 metros de largo. Para otras especificaciones del método de electropesca
ver Barbour et al. (1999) y Maldonado-Ocampo et al. (2005).
En cada localidad se seleccionaron 100 metros del cuerpo de agua como
longitud de colecta, distancia recomendada para obtener una muestra
representativa (Barbour et al. 1999). Se empleó una hora de electropesca
como unidad de esfuerzo de captura estándar, sin embargo, si durante los
primeros 30 minutos no se realizaban capturas se finalizaba el muestreo.
Los peces se fijaron en campo en una solución de formaldehído al 10%
(Uieda y Castro 1999), el material se separó de acuerdo a localidad, se llevó
un registro fotográfico y finalmente en el laboratorio se conservaron en una
solución acuosa de etanol al 70% para su ingreso en la Colección de Peces
del Museo de Historia Natural de la Pontificia Universidad Javeriana (MPUJ)
Bogotá, Colombia.
6.3 Datos de captura y caracterización del hábitat
Se entiende por datos de captura: información de posicionamiento global
(latitud, longitud y altura), localidad específica, parámetros fisicoquímicos,
caracterización del hábitat, y otros datos detallados. Para la anotación de
este tipo de información se utilizó la plantilla desarrollada para la toma de
datos de campo de la Colección de Peces del Museo de Historia Natural de
la Pontificia Universidad Javeriana (MPUJ) Bogotá, Colombia.
La caracterización fisicoquímica del agua se llevó a cabo en el mismo lugar
de colecta antes de emplear el método de electropesca. Se midieron la
temperatura del agua; oxígeno disuelto; porcentaje de saturación de oxígeno
disuelto; pH; y conductividad, todos a nivel de superficie y fondo, este último
variando de acuerdo al punto de colecta y no superando los 150 cm. Se

38
empleó un potenciómetro (pH/Cond10) y un oxímetro, ambos de marca
Oakton.
Se realizaron anotaciones de las macrófitas existentes en el lugar de colecta,
hora y profundidad promedio de captura; además se llevó a cabo una
clasificación cualitativa de cada cuerpo de agua pudiendo ser: léntico, lótico
de corrientes bajas o lótico de corrientes medialtas.
6.4 Proporción de sexos
La diferenciación de sexos se realizó mediante el reconocimiento
macroscópico externo para individuos desde los 40 mm de longitud total. En
el macho a esta longitud hay aparición de una pigmentación externa a lo
largo de la región ventral, principalmente en el istmo, y sobre el dorso. En las
hembras se presenta una coloración plateada metálica en la región del istmo
(Forero y Garzón 1974). Ejemplares comprendidos entre 28 (o incluso un
poco menor) y 39 mm se les llevó a cabo un corte ventral desde el ano hasta
la cintura pectoral para la extracción de las gónadas y su posterior
diferenciación tomando en cuenta que los ovarios inmaduros son de
coloración blancuzca o en algunas zonas translúcidos, con tamaño reducido
pero su diferenciación es posible a simple vista, y en el caso de los testículos
estos aparecen como un par de filamentos translúcidos que recorren antero-
posteriormente la cavidad abdominal que en estado de inmadurez presentan
ondulaciones poco acentuadas (Forero y Garzón 1974). Para lo anterior se
utilizó un estereomicroscopio marca Olympus.
6.5 Relación longitud – peso
La totalidad de los ejemplares se pesaron y midieron, se utilizó una balanza
electrónica marca Sartorius con precisión de 0,1 g y un calibrador digital con
una precisión de 0.01mm. Para la consideración respectiva de machos y
hembras se siguió el mismo procedimiento utilizado para la proporción de

39
sexos. Se consideraron alevinos a los ejemplares de menos de 28 mm de
longitud total.
6.6 Contenido estomacal
Para la extracción de las vísceras se realizó un corte ventral desde el ano
hasta la cintura pectoral, se separó el tubo digestivo conformado por el
esófago, estómago, ciegos pilóricos e intestino. Se seleccionaron 30
estómagos con contenido estomacal por localidad, procurando mantener
tallas similares y representación de ambos sexos. El número de estómagos
examinados mostró variaciones de acuerdo a la abundancia presentada por
localidad.
El contenido estomacal se agrupó por localidad. Para su cuantificación se
utilizó el método volumétrico de análisis directo, que consiste en la expresión
en unidad de volumen de la categoría de alimento o ítem determinado. Cada
ítem fue separado en cajas de petri y se depositó en una celda cuyo volumen
era ajustable mediante una pieza móvil (ver: Prejs y Colomine 1981, y
Marrero 1994); la distancia recorrida por esta, multiplicada por la profundidad
y altura de la celda proporcionaba el volumen en mm³. El volumen de cada
categoría de alimento se expresó como un porcentaje del volumen total de
los contenidos examinados por localidad o composición porcentual
volumétrica (PA) (Ecuación 2) (Hyslop 1980, Prejs y Colomine 1981, Marrero
1994).
100___
_ ×=localidadítemstotalVolumen
ítemAVolumenPA
Ecuación 2
Como ayuda para la identificación de los contenidos, se realizó una colección
de referencia conformada por organismos o materiales que potencialmente

40
pudieran ser consumidos por los peces, se procedió a su identificación en el
laboratorio. Los ítems alimentarios fueron identificados a la mejor resolución
taxonómica posible, en su mayoría hasta familia. Para su determinación se
utilizó bibliografía especializada: McCafferty y Provonsha (1981), Merritt y
Cummins (1984), Roldán (1996), y Gaviria y Aranguren (2003). Para su
observación se empleó un estereoscopio marca Olympus.
7. Análisis de la información
7.1. Abundancia y caracterización del hábitat
La abundancia por localidad se expresó como captura por unidad de
esfuerzo (CPUE) o el número de peces capturado por hora de electropesca.
Para el análisis de la caracterización del hábitat se utilizó estadística
descriptiva con base en los datos de las hojas de campo. Para relacionar las
capturas con los valores de conductividad se realizó una regresión de
acuerdo a Alabaster y Hartley (1962).
7.2. Proporción de sexos
El material se analizó como proporción de machos y hembras capturados en
total y en cada una de las localidades; también se calculó esta proporción por
intervalos de longitud para la totalidad de los ejemplares capturados de
acuerdo a los rangos propuestos para la especie por Forero y Garzón (1974).
La significancia de la diferencia de la proporción 1:1 fue calculada utilizando
una prueba de Chi-cuadrado X2 (α = 0.05) (Vazzoler 1996, Sirangelo y
Ferreira 2004).
7.3. Relación longitud – peso
La relación longitud – peso se estimó por localidad y para el total de
localidades a nivel de alevinos, hembras, machos, y número total de
ejemplares capturados. Los parámetros a y b se estimaron mediante la forma

41
linealizada de la Ecuación 1, que toma logaritmos (base 10) en ambos lados
de la ecuación (Pauly 1984).
( ) ( ) ( )LtbaWt logloglog +=
Ecuación 3
donde a es el intercepto y b es la pendiente de la regresión lineal en su forma
logarítmica (Froese 2006).
Se halló el valor de t-estadístico para establecer si el resultado del
exponente b era significativamente diferente de 3
21
3
..
..2
)(
)( −∗−−
∗= nr
b
ds
dst
y
x
Ecuación 4
donde s.d.(х) es la desviación estándar de los valores de log Lt (longitud
total), y s.d.(y) la desviación estándar de los valores de log Wt (peso total), n
es el número de peces utilizados para el cálculo. Se considera que el valor
de b es diferente de 3 si el t estimado es mayor que el valor de t tabulado con
n-2 grados de libertad. El coeficiente de determinación (r²) expresa la
proporción de la varianza que es explicada por la regresión (Pauly 1984). Su
validez se comprobó al 0.05 y 0.01 de nivel de confianza.
7.4. Contenido estomacal
Para determinar los ítems dominantes de la dieta y la estrategia alimentaria
de la especie se adaptó, para utilizarlo a nivel de localidades, el método
gráfico de Costello (1990, en Amundsen et al. 1996 y Marshall y Elliott 1997)
modificado por Amundsen et al. (1996). En este método se realizan gráficos
de dispersión donde el eje y representa la abundancia específica de la presa

42
(AEP) que para este caso indica el porcentaje de una determinada presa o
ítem, incluyendo todas las demás, sólo en las localidades en la que esta se
presentó (Ecuación 5), es decir:
100×
=∑∑
i
i
St
SAEP
Ecuación 5
donde Si es el contenido estomacal en volumen comprendido por el ítem i en
todas las localidades y Sti es el volumen total ocupado por el contenido
estomacal de sólo las localidades donde se presentó el ítem i (Amundsen et
al. 1996). Por otro lado, el eje x representa la frecuencia de ocurrencia (FO),
que para este caso expresa la frecuencia de localidades, del total
examinado, en la que se presenta un determinado ítem (Ecuación 6) (Hyslop
1980, Amundsen et al. 1996).
slocalidadedeTotal
ítemAconsLocalidadeFO
−−−−=
Ecuación 6
La gráfica permite un análisis de la dieta en tres aspectos: importancia de la
presa, estrategia alimentaria, y amplitud de nicho; su interpretación se realiza
respecto a la posición de los puntos a lo largo de las diagonales y los ejes
(Figura 6).

43
Figura 6. Guía para la interpretación del método gráfico de Costello modificado por Amundsen et al. (1996).
De acuerdo con Amundsen et al. (1996), la diagonal que atraviesa la gráfica
desde el ángulo inferior izquierdo hasta el ángulo superior derecho
proporciona la medida de importancia de la presa, indicando presas raras o
no importantes en la parte baja y dominantes en la parte alta. El eje vertical
representa la estrategia alimentaria de la especie en términos de
especialización o generalización. Determinada especie se ha especializado
en una presa cuando el punto correspondiente a esta se localiza en la parte
alta de la gráfica, mientras que se reconoce una generalización en la dieta
cuando los puntos se encuentran en la parte baja de la misma. La
localización de las presas en la parte superior izquierda de la gráfica podría
ser un indicativo de especialización en localidades individuales, y aquellos en
la parte superior derecha pueden representar la especialización en todas.

44
Finalmente, en términos de amplitud de nicho, los puntos que se ubican ya
sea en la esquina superior izquierda o en la inferior derecha representan
tipos de presas que contribuyen de igual manera a la dieta de la especie,
pero indican estrategias alimentarias diferentes en localidades individuales.
Que para este caso se refiere a que las presas con alta abundancia
específica y una baja frecuencia de ocurrencia (esquina superior izquierda)
han sido consumidas en pocas localidades de manera abundante indicando
quizá algún grado de especialización, mientras que presas con una baja
abundancia específica y una alta ocurrencia (esquina inferior derecha) han
sido consumidas en la mayoría de las localidades (Amundsen et al. 1996).
Las diferencias en la estrategia alimentaria están relacionadas con la inter- y
la intra- contribución fenotípica a la amplitud de nicho. En una localidad
donde los ítems representen un alto componente inter-fenotípico (BPC, por
sus siglas en inglés), diferentes individuos se especializan en diferentes tipos
de recurso mientras que en localidades donde representen un alto
componente intra-fenotípico (WPC, por sus siglas en inglés) la mayoría de
los individuos utilizan varios recursos al mismo tiempo. El patrón de
distribución de los puntos a lo largo de dicha diagonal indica entonces la
contribución de los componentes inter- e intra- fenotípicos a la amplitud de
nicho, si no hay puntos en la parte superior derecha del diagrama sino que
se localizan a lo largo de esta la población del depredador tendrá un alto
valor para la amplitud de nicho. Cabe hacer énfasis en que estos dos últimos
componentes, la variación interfenotípica e intrafenotípica, no fueron
evaluados en este estudio ya que se requiere de un análisis estomacal
individual dentro de la localidad (Figura 6).
Para determinar de mejor manera los ítems dominantes de la dieta, adicional
al método gráfico de Costello, se siguieron las recomendaciones de Marshall

45
y Elliott (1997) utilizando el índice de preponderancia (IP) (Ecuación 7), el
cual genera un único valor para cada atributo basado en el porcentaje de
ocurrencia (Ecuación 6), y la composición porcentual volumétrica (Ecuación
2).
∑=
⋅
⋅=n
i
OiVi
OiViIP
1
)(
Ecuación 7
donde Vi y Oi son el porcentaje de volumen y ocurrencia respectivamente. Al
comparar los valores obtenidos con este índice se genera una clasificación
de la presa en orden de importancia dentro de la dieta (Marshall y Elliot
1997).
Como estimadores de la amplitud de nicho trófico se utilizó el índice de
Levins (B), que es el recíproco del índice de diversidad de Simpson (Krebs
1999), y el índice de Shannon-Wiener (H’). El primero se calculó de la
siguiente manera:
∑=
2
1
jpB
Ecuación 8
donde pj es la proporción de la dieta que es representada por la presa j
(Marshall y Elliott 1997). El índice tiene un valor mínimo de 1.0 cuando sólo
un tipo de presa es encontrado en la dieta y un máximo de n, donde n es el
número total de categorías de presa (Svanbäck y Persson 2004).

46
El índice de Shannon-Wiener (H’) y su respectiva medida de equidad (J’) se
calcularon de la siguiente manera:
∑−= PiPiH ln'
Ecuación 9
n
HJ
ln
''=
Ecuación 10
donde Pi es la proporción de observaciones encontradas en la categoría i y n
el número total de estados del recurso (Krebs 1999). Un valor único es
arrojado por cada localidad y puede ser comparado (Marshall y Elliott 1997).
Como se recomienda en Marshall y Elliott (1997), pa ra los ítems restos de insectos y restos de crustáceos se cuantificó su volumen pero no fueron tomados en cuenta para los análisis de dieta ya que generan una considerab le pérdida de resolución en los resultados de tipo taxonómico. Para facilitar la in terpretación de las tablas y gráficas, los ítems siguieron la codificación indicada en la
Tabla 3.

47
Tabla 3. Códigos asignados para los ítems alimentarios.
CódigoInsecta
ColeopteraChrysomelidae cf………………………………………….. CHRElmidae…………………………………………………….. ELM
DipteraCeratopogonidae CERChironomidae Larvas……………………… CHI-LChironomidae Pupas……………………… CHI-PCulicidae…………………………………………………… CULSimuliidae………………………………………………….. SIMDiptera sin det…………………………………………….. RDI
HemipteraCorixidae…………………………………………………… CORNotonectidae………………………………………………. NOTRestos Hemiptera…………………………………………. HNN
HymenopteraVespidae…………………………………………………… VES
OdonataAeshnidae………………………………………………….. AESCoenagrionidae……………………………………………. COEOdonata sin det…………………………………………… RZY
TrichopteraLeptoceridae……………………………………………….. LEPTrichoptera sin det………………………………………… TRI
Restos Insecta…………………………………………………………….. RINAcarina…………………………………………………………………………………………. ACACrustacea
AmphipodaHyalellidae Hyalella sp………………… HAY
CladoceraBosminidae Bosmina sp………………. BOSChydoridae Chydorus sp………………. CHY
Eurycercus sp……………. EURDaphniidae…………………………………………………. DAP
CopepodaCyclopoida…………………………………………………. CYC
Ostracoda………………………………………………………………….. OSTRestos Crustacea…………………………………………………………. RCR
MolluscaGastropoda
Physidae…………………………………………………… PHYBivalvia……………………………………………………………………… BIV
Huevos Invertebrados…………………………………………………………………………. HUENematoda……………………………………………………………………………………… NEMPlantae
Semillas…………………………………………………………………….. SEMRestos plantas…………………………………………………………….. RPL
Escamas………………………………………………………………………………………. ESCDetrito………………………………………………………………………………………….. DAT
Ítems Alimentarios

48
8. Resultados
8.1 Abundancia de G. bogotensis y caracterización del hábitat
Se capturaron 759 individuos de G. bogotensis para un esfuerzo total de
pesca de 843 minutos. El mayor valor de abundancia se presentó en la
quebrada Mana de Ramos, 127 individuos representando el 16,71% del total,
seguido por el embalse El Hato con 105, 13,82%, y la laguna de Fúquene
con 102, 13,42%. En 5 localidades, ríos Sáchica y Sutamarchán, embalses
del Sisga y San Rafael, y la laguna de Suesca, no se colectaron ejemplares
de la especie (Tabla 4, Figura 7).
Tabla 4. Tipo de cuerpo de agua, macróftica dominante, abundancia de G. bogotensis y tiempo de pesca.
1 Tibitó LE EC 12 602 Embalse del Sisga LE PA 0 303 Vallado La Susana LO-BA PA/OT 78 604 Quebrada Mancilla LO-MA PA 15 605 Embalse de Tominé LE EC 30 606 Embalse San Rafael LE PA/OT 0 607 Río Sáchica LO-MA OT 0 608 Río Sutamarchán LO-MA OT 0 309 Laguna Fúquene LE EC/ED/SC 102 50
10 Río Ubaté LO-BA EC 92 6011 Laguna Cucunubá LE ED 70 6012 Embalse El Hato LE PA/OT 105 4313 Laguna de Suesca LE SC 0 3014 Q. Mana de Ramos LO-BA EC/PA/OT 127 6015 Tota-Pozos LO-BA ED/SC 89 6016 Tota-Upía LE MA 39 60
LE=léntico; LO-BA=lótico corrientes bajas; LO-MA=lótico corrientes medialtasEC= Eichhornia crassipes ; ED= Egeria densa ; SC= Schoenoplectus sp ; MA= Myriophyllum acuaticum; PA= Pastos; OT= Otras
Macrófita
dominante
Tipo de cuerpo de
aguaLocalidad
Abundancia
(nº de indiv)
Minutos de
pesca

49
Sáchica
Suta/chán
FúqueneUbaté
CucunubáEl H
atoTibitó
Tota-Pozos
Tota-UpíaSisga
Suesca
M.Ramos
La Susana
MancillaTominé
San Rafael
Abu
ndan
cia
(%)
0
2
4
6
8
10
12
14
16
18
Figura 7. Proporción de abundancia de G. bogotensis en cada localidad.
Respecto al tipo de cuerpo de agua, 9 localidades se clasificaron como tipo
léntico dónde se presentó el 47.11% del total de individuos, 4 como lóticos de
corrientes bajas, con el 50.79%, y 3 como lóticos de corrientes medialtas, con
el 1,97% (Tabla 4, Figura 8). Las macrófitas encontradas a lo largo del
estudio fueron el buchón de agua (Eichhornia crassipes), la elodea (Egeria
densa), el junco (Schoenoplectus sp.), el pinito de agua (Myriophyllum
acuaticum), su dominancia varió de acuerdo a la localidad (Tabla 4).

50
Tipo de cuerpo de agua
LO-BA LE LO-MA
% d
e ab
unda
ncia
0
10
20
30
40
50
Figura 8. Porcentaje de abundancia de G. bogotensis por tipo de cuerpo de agua. LO-BA=lótico corrientes bajas; LE=léntico; LO-MA=lótico corrientes medialtas.
El valor de abundancia por cuencas muestra que en las localidades que
pertenecen a la del río Suárez, con un esfuerzo de pesca de 393 minutos, se
capturaron 496 individuos que representan el 65,26% de las capturas totales,
en la cuenca media del río Bogotá, con 330 minutos pesca, se capturaron
135 individuos (17,89%), y en la cuenca del lago de Tota, con 120 minutos
de pesca, se atraparon 128 ejemplares (16,84%) (Tabla 5)
Tabla 5. Abundancia de G. bogotensis por cuencas.
Cuenca Individuos Esfuerzo de pesca (min)
Suárez 496 393Bogotá 135 330Tota 128 120
Total 759 843

51
8.1.1 Parámetros fisicoquímicos en cuerpos de agua lóticos
En los ríos Sáchica y Sutamarchán se presentaron temperaturas del agua
superiores a 20ºC, las más altas para cuerpos de agua de tipo lótico durante
el estudio, mientras que la quebrada Mancilla y el río Ubaté presentaron los
valores de temperatura más baja (14 y 16.5ºC respectivamente). A nivel de
superficie, el mayor valor para oxígeno disuelto (OD) se encontró en la
quebrada Mancilla (7.57 ppm) seguido de cerca por la quebrada Mana de
Ramos y el río Sutamarchán, mientras que el más bajo se localizó en el
vallado La Susana (2.8 ppm) y en el río Ubaté (2.88 ppm). La mayor
diferencia de este parámetro entre fondo y superficie fue en la quebrada
Mana de Ramos, que además obtuvo el menor valor a nivel de fondo (0.38
ppm) seguida por el río Ubaté (0.39) que de igual manera presenta una
variación importante con respecto al valor de superficie, en las restantes
localidades la diferencia entre superficie y fondo no fue mayor a 1 ppm. El
porcentaje de saturación de oxígeno disuelto en superficie osciló entre 31.1
en el río Ubaté y 101.7 en la quebrada Mancilla, en fondo entre 4.1 en el río
Ubaté y 99 en la quebrada Mancilla (Tabla 6).
El pH en todas las localidades osciló entre 6 y 7 tanto en fondo como
superficie. La diferencia entre los valores de conductividad en fondo y
superficie no presentaron mayores variaciones, el mayor se registró en el río
Sáchica con 982 µS/cm y el menor en la quebrada Mana de Ramos 36
µS/cm seguida muy de cerca por la quebrada Mancilla con 36.2 µS/cm
(Tabla 6).
8.1.2 Parámetros fisicoquímicos en cuerpos de agua lénticos
La temperatura del agua no presentó mayores diferencias entre fondo y
superficie, la mayor temperatura se registró en la laguna de Fúquene con
24.7ºC y la menor en la laguna de Tota, caño Pozos, con 13.5ºC. El oxígeno
disuelto presentó mayor variación entre fondo y superficie en la laguna de

52
Suesca 0.41 - 5.34 y en la de Fúquene 1.64 - 4.97 ppm. A nivel de superficie
el menor valor registrado para OD fue 0.91 ppm en Tibitoc, mientras que el
mayor fue 7.1 en la laguna de Tota, nacimiento del río Upía; en fondo el
mayor se registró en la laguna de Tota, nacimiento del río Upía, 6.54 ppm
mientras que 0.41 ppm en la laguna de Suesca fue el menor. En superficie el
menor porcentaje de saturación de oxígeno disuelto se encontró en Tibitoc
12.7 seguido por la laguna de Cucunubá, el mayor porcentaje 98.8 se
presentó en el lago de Tota, nacimiento del río Upía. En fondo la laguna de
Suesca con 6.4 obtuvo el menor valor para este parámetro y presentando
además una diferencia importante respecto al valor de superficie, al igual que
en la laguna de Fúquene; en el lago de Tota, nacimiento del río Upía, se
mantuvo el mayor valor para porcentaje de saturación 95.3 (Tabla 6).
Similar a los cuerpos de agua lóticos el valor de pH osciló entre 6 y 7, a
excepción de la laguna de Suesca con 8.3. Los valores de conductividad en
fondo y superficie no presentaron mayores diferencias, al mayor valor para
este parámetro fue de 1099 µS/cm en la laguna de Cucunubá, seguido por la
laguna de Suesca con 898 µS/cm. Los embalses del Sisga y de Tominé
presentaron los valores más bajos de conductividad con 17.06 y 36 µS/cm
respectivamente (Tabla 6).

53
Tabla 6. Parámetros fisicoquímicos para las 16 localidades de estudio.
Superf. Fondo Superf. Fondo Superf. Fondo Superf. Fondo Superf. Fondo
Tibitó 20 17.9 15.4 0.91 0.57 12.7 7.7 6.5 6.3 120.5 102.4
Sisga 40 18.5 18.3 6.42 5.65 94.3 82.9 7.4 7.5 17.06 17.06
La Susana 20 18.0 18.0 2.8 2.64 40.2 37.8 7.0 6.8 152.3 152.7
Mancilla 40 14.0 13.9 7.57 7.41 101.7 99 6.6 6.6 36.2 36.3
Tominé 30 17.9 17.3 6.39 4.54 93.2 63.5 6.5 6.7 36 36.1
San Rafael 40 17.5 17.7 6.1 5.56 87 81.5 6.1 6.5 47.9 48.5
Sáchica 20 24.3 23.8 5.03 5.47 68.3 46.8 7.6 7.7 982 990
Sutamarchán - 21.8 - 6.45 - 77.1 - 6.8 - 188 -
Fúquene 150 24.7 19.8 4.97 1.64 63.7 17.4 6.2 6.0 143.1 130.1
Ubaté 170 16.5 16.2 2.88 0.39 31.1 4.1 6.2 6.4 158.2 220
Cucunubá 150 17.3 17.3 3.65 3.24 41.1 35.1 6.9 6.9 1099 1086
El Hato - 16.2 - 6.38 - 70.4 - 7.4 - 87.9 -
Suesca 30 20.6 20.0 5.34 0.41 84.2 6.4 8.3 8.3 880 898
Mana de Ramos 25 19.7 19.9 6.39 0.38 94.9 5.6 6.9 6.8 36 35.8
Tota-Pozos 30 14.0 13.5 6.81 6.38 94.8 87.7 6.5 6.8 68.1 71.4
Tota-Upía 45 15.9 15.7 7.1 6.54 98.8 95.3 7.3 7.3 73.6 73.5
Cue
nca
río
Bog
otá
Cue
nca
río
Suá
rez
Cue
nca
Tot
a
LocalidadOxí. Disuelto (ppm) % saturación de O.D. pH Conductivi dad (µS/cm)Fondo
(cm)
Temperatura (ºC)
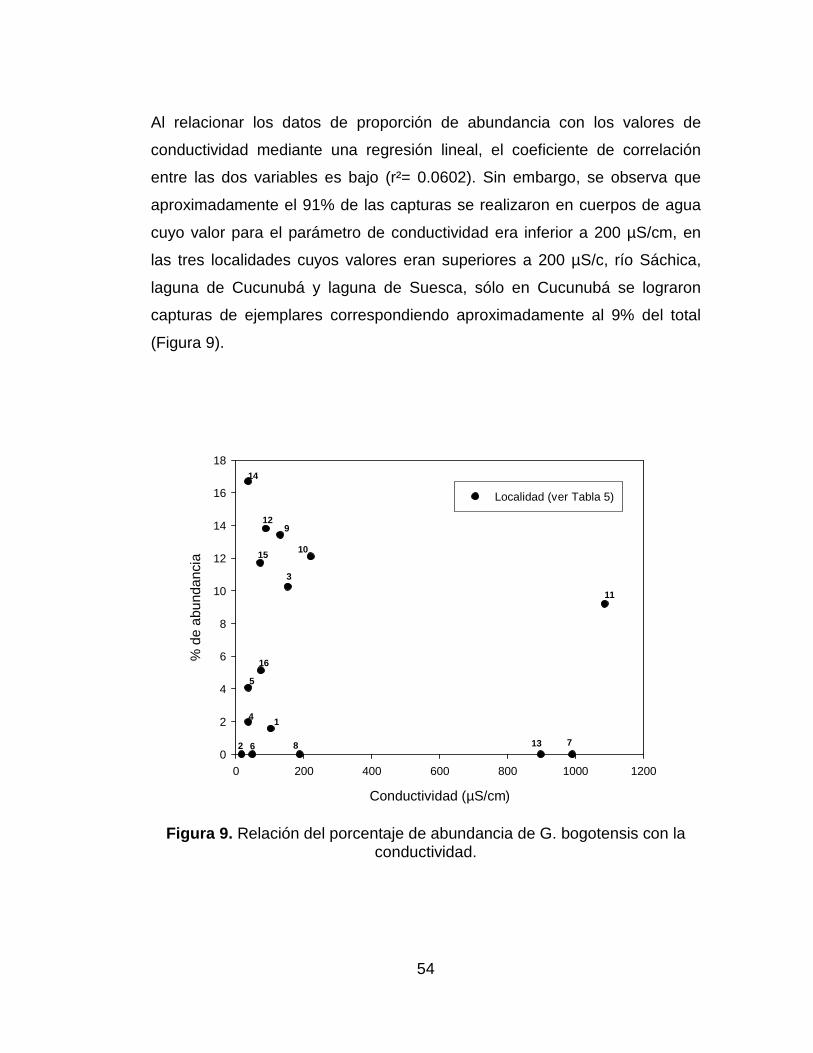
54
Al relacionar los datos de proporción de abundancia con los valores de
conductividad mediante una regresión lineal, el coeficiente de correlación
entre las dos variables es bajo (r²= 0.0602). Sin embargo, se observa que
aproximadamente el 91% de las capturas se realizaron en cuerpos de agua
cuyo valor para el parámetro de conductividad era inferior a 200 µS/cm, en
las tres localidades cuyos valores eran superiores a 200 µS/c, río Sáchica,
laguna de Cucunubá y laguna de Suesca, sólo en Cucunubá se lograron
capturas de ejemplares correspondiendo aproximadamente al 9% del total
(Figura 9).
Conductividad (µS/cm)
0 200 400 600 800 1000 1200
% d
e ab
unda
ncia
0
2
4
6
8
10
12
14
16
18
Localidad (ver Tabla 5)
7132
16
1
11
9
8
14
10
12
15
3
4
5
6
Figura 9. Relación del porcentaje de abundancia de G. bogotensis con la conductividad.

55
8.2 Proporción de sexos
La diferencia de la proporción 1:1 de machos y hembras capturados en total
no fue significativa (X2= 0.74; α=0.05). En la laguna de Fúquene (X2= 33.35;
α=0.05), laguna de Cucunubá (X2= 31.64; α=0.05), lago de Tota, nacimiento
del río Upía, (X2= 4.50, α=0.05), y embalse de Tominé (X2= 14.79; α=0.05) la
proporción de machos fue significativamente mayor; por el contrario, en el río
Ubaté (X2= 8.80; α=0.05), y en Tibitoc (X2= 25.00; α=0.05) la proporción de
hembras fue significativamente mayor. En las restantes localidades la
diferencia de la proporción 1:1 no fue significativa (Tabla 7).
Tabla 7. Proporción de sexos en G. bogotensis por localidad.
Machos Hembras Machos HembrasFúquene 56 15 78.9 21.1 3.7 33.35*Ubaté 32 59 35.2 64.8 0.5 8.80*Cucunubá 50 14 78.1 21.9 3.6 31.64*El Hato 25 28 47.2 52.8 0.9 0.32Tibitó 3 9 25.0 75.0 0.3 25.00*Tota 39 44 47.0 53.0 0.9 0.36Upía 20 13 60.6 39.4 1.5 4.50*Mana de Ramos 50 49 50.5 49.5 1.0 0.01La Susana 38 37 50.7 49.3 1.0 0.02Mancilla 7 8 46.7 53.3 0.9 0.44Tominé 18 8 69.2 30.8 2.3 14.79*TOTAL 338 284 54.3 45.7 1.2 0.74
*=significativo con α = 0.05
Localidad Número % X²M:H
En el intervalo de longitud total de 48.10 hasta 52.09 mm hay predominancia
de hembras (X2= 7.11; α=0.05) mientras que en los intervalos comprendidos
entre 40.10 y 48.09 mm y 68.10 y 76.09 mm la diferencia de la proporción
1:1 es significativa, siendo mayor el número de machos. Se alcanzaron a
diferenciar hembras con longitud total inferior a 28.09 mm (hasta 26.00), en
ejemplares con longitud superior a 80.10 mm no se presentaron hembras. En
los restantes intervalos la diferencia de la proporción 1:1 no fue significativa
(Tabla 8, Figura 10).
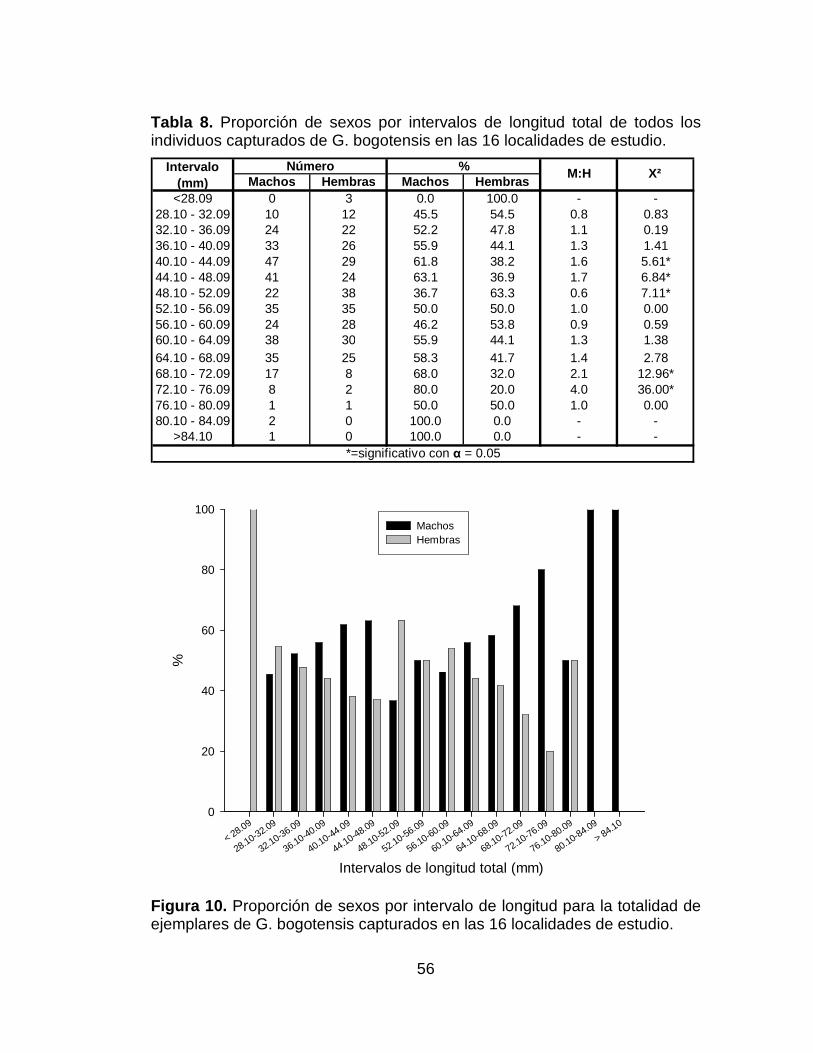
56
Tabla 8. Proporción de sexos por intervalos de longitud total de todos los individuos capturados de G. bogotensis en las 16 localidades de estudio.
Machos Hembras Machos Hembras<28.09 0 3 0.0 100.0 - -
28.10 - 32.09 10 12 45.5 54.5 0.8 0.8332.10 - 36.09 24 22 52.2 47.8 1.1 0.1936.10 - 40.09 33 26 55.9 44.1 1.3 1.4140.10 - 44.09 47 29 61.8 38.2 1.6 5.61*44.10 - 48.09 41 24 63.1 36.9 1.7 6.84*48.10 - 52.09 22 38 36.7 63.3 0.6 7.11*52.10 - 56.09 35 35 50.0 50.0 1.0 0.0056.10 - 60.09 24 28 46.2 53.8 0.9 0.5960.10 - 64.09 38 30 55.9 44.1 1.3 1.3864.10 - 68.09 35 25 58.3 41.7 1.4 2.7868.10 - 72.09 17 8 68.0 32.0 2.1 12.96*72.10 - 76.09 8 2 80.0 20.0 4.0 36.00*76.10 - 80.09 1 1 50.0 50.0 1.0 0.0080.10 - 84.09 2 0 100.0 0.0 - -
>84.10 1 0 100.0 0.0 - -*=significativo con α = 0.05
Número % M:H X²Intervalo (mm)
Intervalos de longitud total (mm)
< 28.09
28.10-32.09
32.10-36.09
36.10-40.09
40.10-44.09
44.10-48.09
48.10-52.09
52.10-56.09
56.10-60.09
60.10-64.09
64.10-68.09
68.10-72.09
72.10-76.09
76.10-80.09
80.10-84.09
> 84.10
%
0
20
40
60
80
100MachosHembras
Figura 10. Proporción de sexos por intervalo de longitud para la totalidad de ejemplares de G. bogotensis capturados en las 16 localidades de estudio.

57
8.3 Relación longitud – peso
Se sometieron 751 ejemplares de G. bogotensis al análisis de relación
longitud – peso, presentaron una longitud total (Lt) promedio de 46.00 mm y
un peso total (Wt) promedio de 1.60 g. Los machos presentaron promedios
de longitud y peso menores que los de las hembras, mientras que en los
alevinos fue de 24.23 mm y 0.20 g respectivamente. En la quebrada Mancilla
tanto machos, 71.69 mm y 8.00 g, como hembras, 68.39 mm y 6.35 g,
presentaron los valores promedio de longitud y peso más altos mientras que
en el embalse El Hato los machos, 34.50 mm y 0.70 g, y hembras, 33.98 mm
y 0.60 g, se colectaron los ejemplares de menor promedio para las dos
variables (Tabla 9, Anexo 1 al Anexo 16).
Para los 751 ejemplares capturados el valor de b= 3.285 fue
significativamente mayor a 3 (r²= 0.9676; t= 13.402; α= 0.01). Por sexos,
machos (b= 3.531; r²= 0.9781; t= 18.430; α= 0.01) y hembras (b= 3.556; r²=
0.9592; t= 12.706; α= 0.01) presentaron valores significativamente mayores a
3 (Tabla 9, Anexo 17 al Anexo 28).
El valor del exponente b varió significativamente, tanto para la totalidad de
ejemplares como para machos y hembras, en algunos de los sitios
estudiados. Cuando se estimó el valor del exponente b para el total de
individuos capturados por localidad, en el lago de Tota - río Upía, quebrada
Mana de Ramos, y embalse de Tominé no fue significativamente diferente de
3 y el coeficiente de determinación fue mayor a 0.97 en todos los casos. En
las restantes localidades se obtuvieron valores significativamente superiores
a 3, con coeficientes de determinación no menores a 0.875, fluctuando desde
b= 3.110 en la laguna de Fúquene hasta b= 3.623 en la quebrada Mancilla
(Tabla 9, Anexo 17 al Anexo 28).

58
Tabla 9. Relación longitud – peso de G. bogotensis en 11 localidades de estudio. Sexo: macho (M), hembra (H), y alevinos (A), número de individuos (n); características de longitud total (Lt): mínima, máxima, y promedio; características de peso total (Wt): mínimo, máximo, y promedio; exponente b; coeficiente de determinación (r²), t estimado (t); significancia (SN).
Fúquene Total 101 14.85 55.90 40.45 0.05 3.6 1.00 3.110 0.9702 2.008 ♪
M 56 36.38 55.90 43.85 0.50 3.6 1.30 3.114 0.746 0.461 ns H 15 30.84 55.14 40.44 0.50 2.9 1.00 3.142 0.9382 0.635 ns A 30 14.85 33.32 18.74 0.05 0.5 0.10 2.383 0.716 -2.176 ns
Ubaté Total 92 28.99 67.84 49.39 0.40 5.9 2.10 3.524 0.9261 4.998 ♫
M 32 36.86 64.56 53.41 0.70 5.3 2.75 3.561 0.9606 4.257 ♫
H 59 36.09 67.84 48.13 0.60 5.9 1.90 3.558 0.899 3.536 ♫
A 1 28.99 28.99 - 0.40 0.4 - - - - -Cucunubá Total 70 22.74 58.99 42.69 0.10 3.6 1.20 3.603 0.886 3.844 ♫
M 50 32.87 52.91 42.95 0.40 2.3 1.20 3.287 0.9085 1.906 ♪
H 14 32.75 58.99 43.41 0.10 3.6 1.10 4.639 0.7769 2.283 ♪
A 6 22.74 26.07 24.30 0.10 0.4 0.15 3.341 0.13 0.079 ns Hato Total 105 16.27 68.90 29.94 0.10 5.0 0.40 3.295 0.9106 2.901 ♫
M 25 29.94 48.34 34.50 0.40 2.1 0.70 3.740 0.9686 5.275 ♫
H 28 29.53 68.90 33.98 0.30 5.0 0.60 3.238 0.9487 1.608 ns A 52 16.27 31.97 27.55 0.10 0.5 0.30 2.631 0.688 -1.453 ns
Tibitó Total 12 33.01 57.87 47.68 0.70 4.8 2.35 3.437 0.9893 3.826 ♫
M 3 34.73 47.97 47.24 0.80 2.3 2.30 3.343 0.9982 2.418 ns H 9 33.01 57.87 52.43 0.70 4.8 2.80 3.456 0.9875 3.103 ♫
A 0 - - - - - - - - - -Tota Total 85 21.55 77.46 63.42 0.10 11.0 5.50 3.352 0.8755 2.554 ♫
M 39 54.67 77.46 65.95 3.20 11.0 6.10 3.574 0.9036 2.988 ♫
H 43 50.21 70.82 61.62 2.40 7.9 5.20 3.453 0.9227 2.900 ♫
A 3 21.55 23.87 23.67 0.10 0.2 0.10 3.063 0.1887 0.010 ns Nac.Upía Total 38 17.90 67.98 58.80 0.10 7.3 4.40 3.065 0.9914 1.372 ns
M 20 41.06 67.98 61.90 1.70 6.9 4.95 2.808 0.9099 -0.922 ns H 13 53.25 67.41 56.24 2.80 7.3 4.00 3.460 0.9501 1.927 ♪
A 5 17.90 30.53 19.27 0.10 0.5 0.20 2.847 0.8899 -0.264 ns Mana Ramos Total 127 16.42 74.85 52.51 0.10 9.6 2.70 2.936 0.9758 -1.555 ns
M 50 30.09 74.85 62.73 0.50 9.6 5.00 3.426 0.9797 5.977 ♫
H 49 30.68 66.50 50.98 0.70 6.8 2.50 3.188 0.9654 2.140 ♪
A 28 16.42 29.93 23.57 0.10 0.6 0.30 3.318 0.7501 0.848 ns V.Susana Total 76 28.16 75.17 56.39 0.30 9.3 3.50 3.452 0.9942 14.764 ♫
M 38 32.10 74.27 56.39 0.50 8.7 2.95 3.387 0.9942 8.973 ♫
H 37 30.48 75.17 56.98 0.40 9.3 3.80 3.528 0.9933 10.768 ♫
A 1 28.16 28.16 - 0.30 0.3 - - - - -Q.Mancilla Total 15 42.01 86.76 68.88 1.20 14.7 7.10 3.623 0.9886 5.769 ♫
M 7 43.92 86.76 71.69 1.40 14.7 8.00 3.735 0.9924 5.028 ♫
H 8 42.01 76.11 68.39 1.20 10.5 6.35 3.455 0.9857 2.677 ♪
A 0 - - - - - - - - - -Tominé Total 30 20.80 62.10 46.66 0.20 3.9 1.85 2.890 0.9926 -2.343 ns
M 18 29.51 62.10 53.09 0.50 3.9 2.55 2.932 0.9853 -0.765 ns H 8 26.00 50.52 41.69 0.30 2.2 1.25 2.965 0.9925 -0.338 ns A 4 20.80 25.24 22.94 0.20 0.3 0.25 2.464 0.7335 -0.510 ns
TOTAL Total 751 14.85 86.76 46.00 0.05 14.7 1.60 3.295 0.9676 13.402 ♫
M 338 29.51 86.76 49.79 0.40 14.7 2.10 3.531 0.9781 18.430 ♫
H 283 26.00 76.11 50.34 0.10 10.5 2.30 3.556 0.9592 12.706 ♫
A 130 14.85 33.32 24.23 0.05 0.6 0.20 2.542 0.6283 -2.653 ns ♫= significativo al 1%; ♪= significativo al 5%; ns= no significativo
SNWprom
(g)b r²
Ltmax (mm)
Ltprom (mm)
Wmin (g)
Wmax (g)
Localidad Sexo nLtmin (mm)

59
Para los machos analizados en la laguna de Fúquene, Tibitoc, lago de Tota –
río Upía, y embalse de Tominé el valor de b no fue significativamente
diferente de 3 y el coeficiente de determinación fue mayor a 0.90, a
excepción de Fúquene (0.746). En las demás localidades estudiadas el valor
de b fue significativamente mayor a 3 encontrándose entre 3.287 en la
laguna de Cucunubá y 3.740 en el embalse El Hato, todas con coeficiente de
determinación no menor a 0.9036 (Tabla 9, Anexo 17 al Anexo 28).
En el caso de las hembras, en la laguna de Fúquene, embalse El Hato, y
embalse de Tominé el valor de b no fue significativamente diferente de 3 y el
coeficiente de determinación fue mayor a 0.93. En las demás localidades
estudiadas el valor de b fue significativamente diferente de 3, con
coeficientes de determinación no menores a 0.7769, y los valores fluctuaron
entre 3.188 en la quebrada Mana de Ramos hasta 4.639 en la laguna de
Cucunubá (Tabla 9, Anexo 17 al Anexo 28).
En la laguna de Cucunubá, Tibitoc, lago de Tota – río Upía, y vallado La
Susana las hembras presentaron un mayor valor del parámetro b que los
machos, mientras que en el río Ubaté, embalse El Hato, lago de Tota – caño
Los Pozos, quebrada Mana de Ramos, y quebrada Mancilla, fueron los
machos los que obtuvieron un mayor valor para b (Tabla 9, Anexo 17 al
Anexo 28).
El análisis de la relación longitud – peso para los alevinos sólo se realizó en
7 de las 11 localidades, ya que en 4 de ellas no se capturaron individuos
para esta clase. Para estos el valor del exponente b no es significativamente
diferente de 3 en ninguna de las localidades, en algunas el coeficiente de
determinación presentó valores bajos, la laguna de Cucunubá (r²= 0.1300) y
el lago de Tota, caño Los Pozos (r²= 0.1887), y el valor más alto del

60
coeficiente en alevinos se obtuvo en el lago de Tota, nacimiento del río Upía
(r²= 0.8899) (Tabla 9, Anexo 17 al Anexo 28).
8.4 Aspectos tróficos
Se examinaron 281 estómagos con contenido estomacal, la menor cantidad
de estómagos observados fue 12 en Tibitoc seguido por la quebrada Mancilla
con 14. Se determinaron 35 ítems incluyendo restos de insectos y restos de
crustáceos, la riqueza varió entre 10 en el río Ubaté, Tibitoc y vallado La
Susana, hasta 17 en el lago de Tota, nacimiento del río Upía (Tabla 10).
Tabla 10. Riqueza de ítems alimentarios, amplitud de nicho determinado por los índices de Levins (B), Shannon (H’), y su valor de equidad (J’), para las 11 localidades de estudio.
Riqueza Levins (B) Shannon (H') Equidad (J')
Fúquene 12 6.93 2.17 0.87
Ubaté 10 1.43 0.74 0.32
Cucunubá 11 5.39 1.91 0.80
El Hato 11 3.23 1.50 0.63
Tibitó 10 3.52 1.63 0.71
Tota-Pozos 11 5.67 2.00 0.84
Tota-Upía 17 9.14 2.48 0.88
Lenguazaque 13 1.43 0.70 0.27
La Susana 10 2.63 1.39 0.60
Q. Mancilla 12 3.45 1.59 0.64
Tominé 11 7.43 2.14 0.89
En general el material presente en la dieta fue de origen autóctono, los
grupos más abundantes fueron insectos de los órdenes Coleoptera
(Chrysomelidae cf.), Diptera (larvas y pupas de Chironomidae) y en menor
cantidad Trichoptera (no determinado) y Odonata (Aeshnidae); detrito; restos
de plantas; y microcrustáceos principalmente el anfípodo Hyalella sp.;

61
cladóceros de la familia Daphniidae y en menor cantidad Bosminidae; y
copépodos del orden Cyclopoida (Anexo 29).
La composición de la dieta y la abundancia porcentual volumétrica de los
ítems presentaron variaciones por localidad (Figura 11 a Figura 13). En tres
localidades, laguna de Fúquene (Figura 11a), laguna de Cucunubá (Figura
11c), y lago de Tota, río Upía (Figura 12c), el ítem más abundante fue restos
de insectos con hasta un 35.4% del volumen total en el caso de Cucunubá.
Las pupas de Chironomidae fueron la presa más abundante en el río Ubaté
(Figura 11b), mientras que en estado larvario lo fue en el embalse El Hato
(Figura 11d). Otras localidades donde los insectos se presentaron como el
grupo más abundante dentro de la dieta fueron en la quebrada Mana de
Ramos con el coleóptero de la familia Chrysomelidae cf. (Figura 12d), y en el
vallado La Susana con Trichoptera no identificado (Figura 13a). En la
quebrada Mancilla el detrito se presentó como el ítem más abundante (43%)
seguido por formas larvarias de dípteros de la familia Chironomidae (Figura
13b).
Los microcrustáceos también se presentaron en varias localidades como el
grupo más abundante dentro de la dieta. El ítem restos de crustáceos fue el
más abundante en el embalse de Tominé (Figura 13c), los cladóceros de la
familia Daphniidae lo fueron en Tibitoc (Figura 12a), y en el lago de Tota-Los
Pozos, el anfípodo Hyalella sp. (Figura 12b).

62
(a)
RINRPL
DAPHAY
RCRNOT
ESCCHI-L TRI
OSTCYC
CHYBOS
ANN
Vol
umen
(%
)
0
5
10
15
20
25
n= 30
(b)
CHI-P RINCHI-L
RCRRDI
TRICYC
PHYCUL
HUEHAY
CHY0
10
40
50
n= 30
(c)
Ítems
RINRCR
AESCOE
BOSLE
PVES
CHI-LOST
RPLDAP
CYCACA
Vol
umen
(%
)
0
10
20
30
40
(d)
ÍtemsCHI-L
BOSRZY
RINPHY
CYCRCR
RPLRDI
OSTTRI
CULDAP
0
10
20
30
40
50n= 30n= 30
Figura 11. Porcentaje de volumen de los ítems de alimento en las localidades de (a) laguna de Fúquene, (b) río Ubaté, (c) laguna de Cucunubá, y (d) embalse El Hato. Para codificación remitirse a la tabla 3.
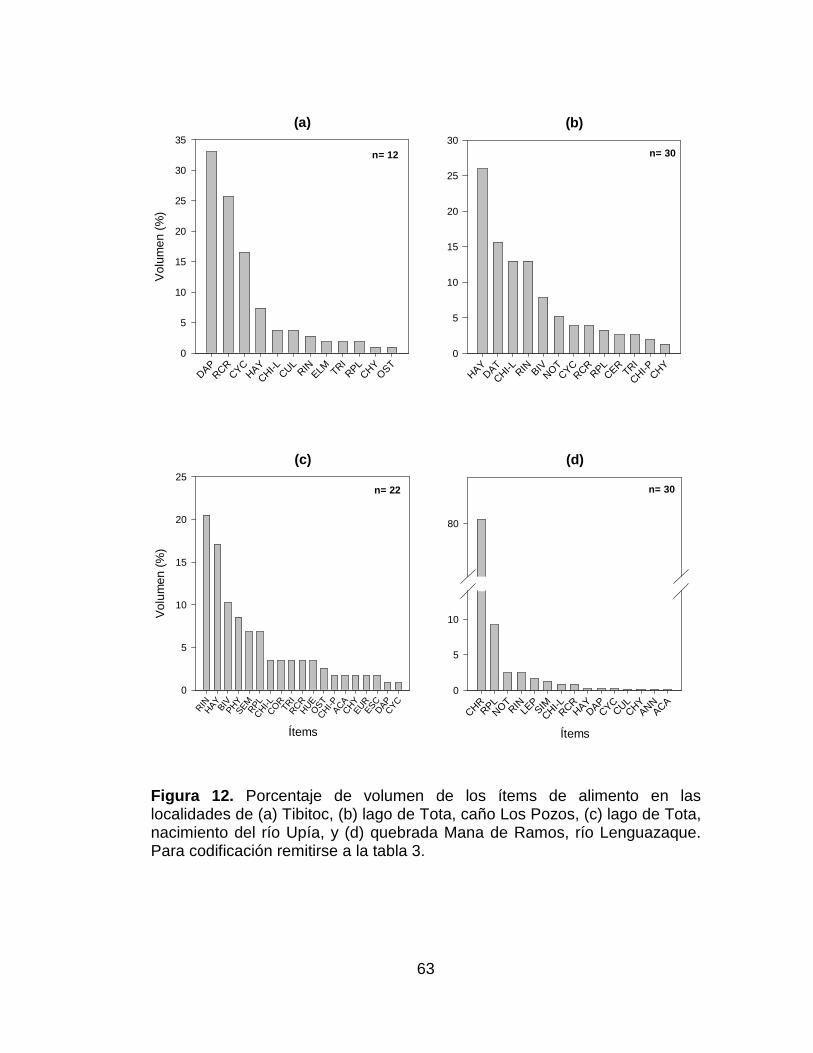
63
(c)
Ítems
RINHAYBIV
PHYSEMRPL
CHI-LCO
RTR
IRCRHUEO
STCHI-PACACHYEURESCDAPCYC
Vol
umen
(%
)
0
5
10
15
20
25n= 22
(a)
DAPRCR
CYCHAY
CHI-L CULRIN
ELM TRIRPL
CHYOST
Vol
umen
(%
)
0
5
10
15
20
25
30
35
n= 12
(d)
Ítems
CHRRPL
NOTRIN
LEPSIM
CHI-LRCR
HAYDAP
CYCCUL
CHYANN
ACA0
5
10
80
n= 30
(b)
HAYDAT
CHI-L RIN BIVNOT
CYCRCR
RPLCER
TRI
CHI-PCHY0
5
10
15
20
25
30n= 30
Figura 12. Porcentaje de volumen de los ítems de alimento en las localidades de (a) Tibitoc, (b) lago de Tota, caño Los Pozos, (c) lago de Tota, nacimiento del río Upía, y (d) quebrada Mana de Ramos, río Lenguazaque. Para codificación remitirse a la tabla 3.

64
(a)
TRIRIN
RCRCYC
CHI-L RPLHAY
DATCHY
OSTACA
DAP
Vol
umen
(%
)
0
10
20
30
40
n= 30
(b)
DATCHI-L
AESRIN
HAYRPL
CERSEM
ACACYC
ANNHNN
CHY0
10
20
40
45
50
n= 14
(c)
Ítems
RCREUR
CHI-LDAP
HAYRPL
RINCHY
TRICYC
OSTPHY
ACA
Vol
umen
(%
)
0
10
20
30
40
50
n= 23
Figura 13. Porcentaje de volumen de los ítems de alimento en las localidades de (a) vallado La Susana, (b) quebrada Mancilla, y (c) embalse de Tominé. Para codificación remitirse a la tabla 3.

65
De acuerdo al método gráfico de Costello modificado por Amundsen et al.
(1996), en la dieta de G. bogotensis no se presentan presas dominantes.
Gran parte de los puntos se localizaron en la esquina inferior izquierda (baja
frecuencia de ocurrencia y baja abundancia específica de la presa) indicando
así que son ítems raros y su aporte individual a la dieta es inferior al 5%,
entre ellos se encuentran insectos de los órdenes Coleoptera (Elmidae),
Diptera (Ceratopogonidae, Simuliidae, otros no identificados), Hemiptera
(Corixidae, otros no identificados), Hymenoptera (Vespidae), Odonata
(Aeshnidae, Notonectidae, otros no identificados), Trichoptera
(Leptoceridae), el cladócero del género Eurycercus sp., moluscos, semillas, y
escamas (Figura 14, Anexo 30).
Respecto a la estrategia alimentaria, en general, por la disposición de los
puntos dentro de la gráfica se infiere que la especie presenta una
generalización de la dieta; sin embargo, en localidades puntuales se
presentó preferencia hacia alguna presa como en la quebrada Mana de
Ramos con el coleóptero de la familia c.f. Chrysomelidae ubicado en el
ángulo superior izquierdo de la gráfica, no obstante el alcance de este
resultado en particular debe ser tomado con cautela ya que esta presa (un
solo ejemplar de gran tamaño) fue consumida por un solo individuo de los 30
examinados. La misma tendencia pero menos marcada ocurre en el río
Ubaté para el ítem pupas de Chironomidae.
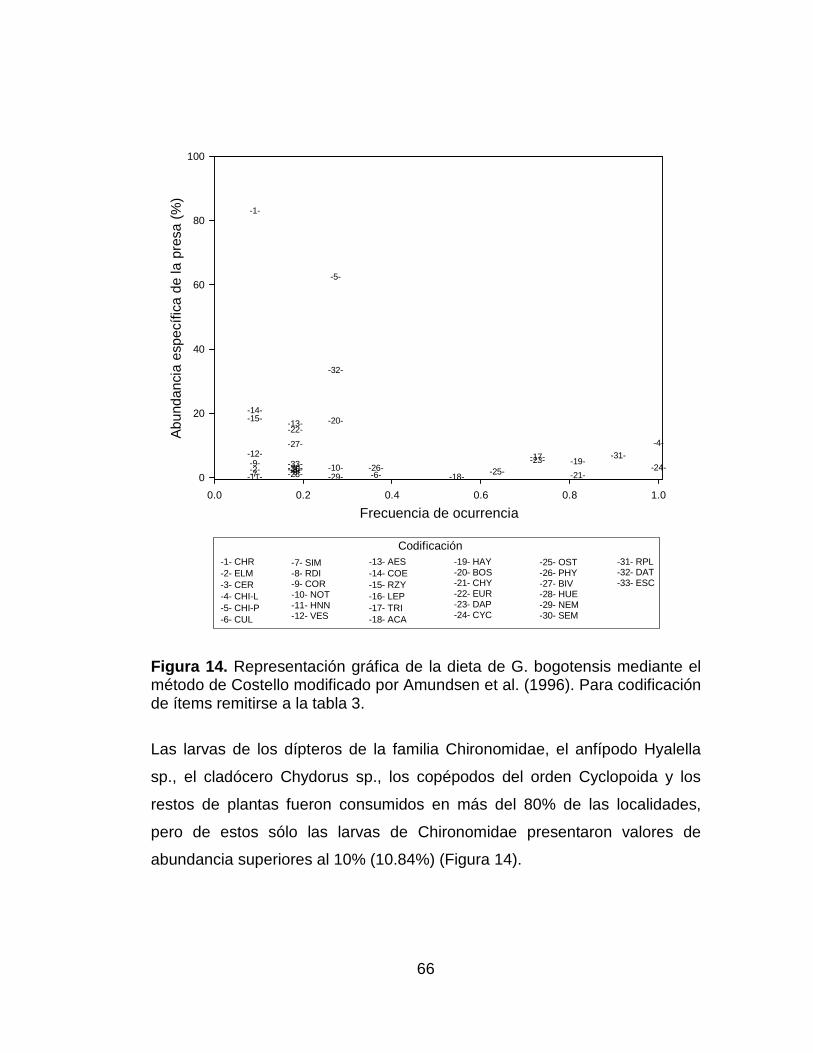
66
Frecuencia de ocurrencia0.0 0.2 0.4 0.6 0.8 1.0
Abu
ndan
cia
espe
cífic
a de
la p
resa
(%
)
0
20
40
60
80
100
-1-
-2- -3-
-4-
-5-
-6--7- -8--9- -10--11-
-12-
-13-
-14--15-
-16-
-17-
-18-
-19-
-20-
-21-
-22-
-23--24--25--26-
-27-
-28- -29--30-
-31-
-32-
-33-
Codificación-1- CHR-2- ELM-3- CER-4- CHI-L-5- CHI-P-6- CUL
-7- SIM-8- RDI-9- COR-10- NOT-11- HNN-12- VES
-13- AES-14- COE-15- RZY-16- LEP-17- TRI-18- ACA
-19- HAY-20- BOS-21- CHY-22- EUR-23- DAP-24- CYC
-25- OST-26- PHY-27- BIV-28- HUE-29- NEM-30- SEM
-31- RPL-32- DAT-33- ESC
Figura 14. Representación gráfica de la dieta de G. bogotensis mediante el método de Costello modificado por Amundsen et al. (1996). Para codificación de ítems remitirse a la tabla 3.
Las larvas de los dípteros de la familia Chironomidae, el anfípodo Hyalella
sp., el cladócero Chydorus sp., los copépodos del orden Cyclopoida y los
restos de plantas fueron consumidos en más del 80% de las localidades,
pero de estos sólo las larvas de Chironomidae presentaron valores de
abundancia superiores al 10% (10.84%) (Figura 14).

67
Casi la totalidad de las presas se localizaron debajo o a lo largo de la
diagonal que recorre la gráfica desde la esquina superior izquierda a la
inferior derecha lo que implica en general un alto valor para amplitud de
nicho de la especie (Figura 14).
De acuerdo al índice de preponderancia (IP) el ítem más importante dentro
de la dieta de G. bogotensis fueron las larvas de Chironomidae, seguido por
restos de plantas, pupas de Chironomidae y el anfípodo del género Hyalella
sp. (Tabla 11).
Tabla 11. Valores del índice de preponderancia (IP) de los ítems alimentarios.
Ítem IP Ítem IP
CHI-L 0.2666 RZY 0.0039RPL 0.1269 EUR 0.0031
CHI-P 0.1032 BIV 0.0029HAY 0.0877 ACA 0.0025CYC 0.0779 RDI 0.0023CHR 0.0642 COE 0.0023DAP 0.0629 CER 0.0020TRI 0.0557 SEM 0.0020DAT 0.0481 SIM 0.0010BOS 0.0183 HUE 0.0010CHY 0.0136 NEM 0.0009AES 0.0134 VES 0.0008OST 0.0089 ESC 0.0005PHY 0.0088 ELM 0.0003NOT 0.0084 COR 0.0002CUL 0.0052 HNN 0.0001LEP 0.0046
Al comparar los valores de los índices utilizados para calcular la amplitud de
nicho, Shannon y Levins, no se aprecian mayores diferencias. Para ambos
índices las localidades con menor amplitud de nicho fueron la quebrada
Mana de Ramos y el río Ubaté, y las de mayor amplitud fueron el lago de
Tota-río Upía y el embalse de Tominé, según el índice de Shannon, y el lago

68
de Tota-Upía y la laguna de Fúquene pero seguido muy de cerca por el
embalse de Tominé, según el de Levins (Tabla 10).
9. Discusión
9.1 Abundancia de G. bogotensis y caracterización del hábitat
Los valores de algunos parámetros fisicoquímicos, bajo oxígeno disuelto
(Miller et al. 2002) y alta conductividad (Lobón-Cerviá 1991, Pinto-Coelho et
al. 2005), sumado a la presencia extensiva de macrófitas, como Eichhcornia
crassipes y Egeria densa, que es el caso específico de la laguna de
Fúquene, río Ubaté, laguna de Cucunubá, Tibitoc, laguna de Suesca, lago de
Tota, son indicadores de su probable estado eutrófico. En un diagnóstico
reciente del estado de la cuenca de los ríos Ubaté – Suárez (CAR 2006) se
identifica a la laguna de Fúquene -y sistemas lagunares asociados-, laguna
de Cucunubá, y laguna de Suesca como cuerpos de agua eutróficos, donde
el vertimiento directo de aguas residuales domésticas e industriales y en
menor intensidad la disposición inadecuada de residuos sólidos domésticos
son los elementos asociados a su contaminación. Por otra parte, la cuenca
del río Bogotá recibe descargas contaminantes propias de actividades en
cabeceras municipales, labores agropecuarias, especialmente cultivos de
flores, papa y café, y adicionalmente desechos de la industria de
curtiembres, lácteos y minera (CAR 2002). En el lago de Tota la utilización de
herbicidas, fertilizantes y fungicidas en el cultivo de cebolla junca es la
principal causa del deterioro de este cuerpo de agua (CORPOBOYACÁ,
2002).
El pH en la laguna de Suesca fue el más alto de las localidades estudiadas
mientras que el valor para conductividad estuvo cercano a los 900 µS/cm.
Escobar (1991) reporta valores de conductividad entre 500 y 545 µS/cm,
indicando un grado de mineralización debido principalmente al calcio,

69
cloruros y en menor grado a los sulfuros. La conductividad probablemente
está estrechamente relacionada con el valor de pH reportado en este
estudio. En las restantes localidades los valores fluctuaron dentro del rango
aceptado para la mayoría de los ríos y lagos del mundo (6.0-8.0) (Bolner y
Baldisserotto 2007).
G. bogotensis se ha reportado en sistemas acuíferos cuyos valores mínimos
para oxígeno disuelto llegan hasta 4,25 ppm en el embalse del Neusa
(Forero y Garzón 1974), 4,7 ppm en el río Balsillas (Gompertz 1986), y
recientemente, Rivera-Rondón et al. (2007) la reportan en la laguna de
Fúquene con valores que oscilan entre 4 y 12 ppm en el primer metro de
profundidad, y sugieren que su permanencia en sectores con lecturas <1
ppm puede estar restringida a cortos intervalos de tiempo, de manera que los
individuos realizarían movimientos verticales en la columna de agua.
En el presente estudio los valores registrados para OD se encuentran dentro
de los rangos de reportes anteriores a excepción de la localidad de Tibitoc
donde el valor de OD tanto en superficie como fondo fue <1 ppm, llegando
incluso a estar por debajo de 0.6 ppm valor registrado como letal bajo
condiciones de laboratorio para algunas especies de peces neotropicales
que presentan respiración en la capa superficial del agua como adaptación
para afrontar bajas concentraciones de oxígeno (Soares et al. 2006).
También se ha encontrado que la reducción del oxígeno disuelto puede
afectar la calidad del hábitat, la supervivencia, el crecimiento y la
reproducción (McKinsey y Chapman 1998, USEPA 2000). Sin embargo, para
G. bogotensis no se han llevado a cabo estudios que indiquen las
concentraciones mínimas de OD a las cuales puede sobrevivir o
adaptaciones bajo condiciones cercanas a la anoxia.

70
La captura de G. bogotensis en cuerpos de agua de carácter léntico con
presencia de macrófitas como E. crassipes, E. densa y Schoenoplectus sp.
concuerda con lo reportado por Forero y Garzón (1974), Gompertz (1986),
Mora et al (1992), Román-Valencia et al. (2003) y Rivera-Rondón et al.
(2007).
Para el embalse de San Rafael, León et al. (2005) había reportado la
presencia de la guapucha, sin embargo, en el presente trabajo no se
capturaron ejemplares en esta localidad. Quizás el factor que podría explicar
este resultado es la ausencia de macrófitas particularmente de las especies
nombradas arriba. Algunos estudios han determinado que la abundancia de
los peces se ve afectada por el tipo de macrófita (Grenouillet et al. 2002).
Para el caso de los ríos Sáchica y Sutamarchán probablemente por ser
cuerpos de agua lóticos con corriente media-alta no se genera un hábitat
propicio para el desarrollo de la especie. En la quebrada Mancilla
probablemente fue la oferta de microhábitats, como pocetas o sectores con
corrientes de baja velocidad, uno de los factores que permitió la presencia de
la especie. Algunos estudios han relacionado la abundancia de los peces en
función de la abundancia del zooplancton (Rozas y Odum 1988, Grenouillet
et al. 2002). En ambientes lóticos, el incremento en la velocidad de flujo del
agua, especialmente en temporada de lluvias, ha sido considerado como uno
de los principales factores que determinan la reducción de las abundancias
locales de organismos acuáticos como insectos bentónicos y
microcrustáceos (Bispo et al. 2006).
La relación entre el esfuerzo de captura y el bajo número de individuos
capturados en la cuenca del río Bogotá, respecto a la cuenca del río Suárez
y lago de Tota, quizás sea explicada por la calidad de sus aguas. En esta
cuenca se encuentran 483 vertimientos de origen municipal, 672 de índole

71
agropecuario y 2822 de tipo industrial, siendo esta la cuenca del país más
contaminada (CAR 2002) y donde las concentraciones de metales pesados
están por encima de los niveles máximos permitidos (Mancera-Rodríguez y
Álvarez-León 2006).
Gompertz (1986) reporta a G. bogotensis en la laguna de Suesca pero no
aporta valores de abundancia u otros datos de captura. En el presente
estudio no se capturaron ejemplares en esta localidad. Probablemente se
deba a un fenómeno de extinción a nivel local, ya que de acuerdo con
información de habitantes de la zona aledaña a la laguna recientemente se
ha venido presentando un proceso de desecación importante, ayudado por la
particularidad de que es una cuenca cerrada (Escobar 1991) y a que la
laguna se ha ido llenando lentamente con materiales de diferentes orígenes,
llevando sus áreas periféricas a procesos de consolidación convirtiéndolas
en un sistema terrestre (CAR 2006).
En el embalse del Sisga, un cuerpo léntico y donde Gompertz (1986) registró
a la guapucha, la baja conductividad probablemente fue el parámetro que
más afectó la eficiencia de captura. Se considera que la conductividad tiene
efectos sobre el voltaje y la densidad de línea corriente, lo que la convierte
en el parámetro fisicoquímico más importante a la hora de implementar el
método de electropesca (Bohlin et al. 1989). La eficiencia de captura se ve
reducida significativamente a conductividades menores de 20 µS/cm (Sternin
et al. 1972, en: Bohlin et al. 1989) llegando a niveles mínimos
aproximadamente a los 5 µS/cm (Bohlin et al. 1989).
Se consideran aguas de baja conductividad, y donde el desempeño de la
pesca eléctrica es bajo, a las que presentan valores menores de 75-100
µS/cm, (Lobón-Cerviá 1991). Según Alabaster y Hartley (1962) la eficiencia
de captura no supera el 15% bajo estos valores de conductividad. En este

72
estudio la mayoría de las capturas se produjeron a valores de conductividad
inferiores a 100 µS/cm. En estudios llevados a cabo por Lasso y Castroviejo
(1992, en Penczak et al. 1997) y Penczak et al. (1997) también se ha
relatado la captura de peces en cuerpos de agua donde la conductividad era
inferior a 75 µS/cm, llegando incluso por debajo de los 20 µS/cm, mediante el
método de electropesca. Entonces quizá el valor del coeficiente de
correlación (r²= 0.0602) entre la proporción de abundancia por localidad y la
conductividad se debe al número de localidades que presentaron valores de
conductividad mayor a 200 µS/cm.
9.2 Proporción de sexos
En muchas especies de peces se observa una proporción 1:1 entre hembras
y machos (Vazzoler 1996). En el presente estudio, para la totalidad de
individuos capturados, la proporción sexual fue 1:1. En la evaluación
individual de las localidades la misma situación se presenta en el embalse El
Hato, lago de Tota – Los Pozos, quebrada Mana de Ramos, vallado La
Susana y quebrada Mancilla.
Algunas localidades presentaron diferencias significativas en la composición
de sexos. En el río Ubaté y Tibitoc se presentó predominancia significativa de
las hembras fenómeno que se había reportado para el caño Guanquica,
embalse de Neusa, por Forero y Garzón (1974).
Cuando el análisis de la proporción se realiza a nivel de clases de talla o
intervalos de longitud se puede determinar la predominancia de individuos de
alguno de los dos sexos (Vazzoler 1996). Al calcular la proporción por
intervalos de longitud para la totalidad de individuos capturados se observó el
mismo fenómeno de disminución paulatina de hembras y aumento de
machos reportado por Forero y Garzón (1974), pero desde una longitud total
de 40.10 mm osea inferior a los 44.00 mm de ese estudio, hasta llegar a una

73
ausencia de hembras en longitudes superiores a los 80.10 mm. Se debe
aclara que hubo una variación en uno de los intervalos (48.10 – 52.09 mm)
donde predominaron de nuevo las hembras.
Lo anterior podría apoyar la hipótesis de Forero y Garzón (1974) quienes
sostienen que probablemente en G. bogotensis la diferencia en la proporción
de sexos está relacionada con una ventaja particular para alguno de los dos
sexos o con una mortalidad diferencial, eventos que ya han sido relatados
como causantes de la diferencia en la proporción sexual en otras especies
de peces (Reay 1989, en Cognato y Fialho 2006, Vazzoler 1996). También
se podría deber a que los machos presentarían una tasa de crecimiento
mayor a la de hembras llegando de esta manera a predominar en los
intervalos de mayor longitud como se ha reportado también para algunas
especies (Vicentini y Araújo 2003).
Sumado a lo anterior, se ha encontrado que la variación en la proporción 1:1
en peces también puede estar influenciada por la disponibilidad de alimento
(Vicentini y Araújo 2003), segregación sexual debido a ocupación de
diferentes hábitats o grupos sociales (Robbins 2007), factores abióticos como
el pH (Rubin 1985) e incluso por interacciones entre la temperatura y el
genotipo (Conover y Kynard 1981).
Forero y Garzón (1974) establecen dos períodos bien marcados de
reproducción en la especie que coinciden con la temporada de mayor
precipitación, la primera postura se da en los meses de marzo-abril y la
segunda entre septiembre-octubre. En estos períodos se produce una
migración por parte de las hembras desde los caños hacia la laguna, lugar
donde procederán a la elaboración de los nidos y posterior cuidado parental
de manera turnada. En el presente trabajo, no son claros los efectos de la
migración sobre la proporción de sexos quizás por la escala temporal

74
empleada en este estudio y/o variedad de ambientes donde se llevaron a
cabo las capturas. No obstante, cabe anotar que Mora et al. (1992)
encontraron que la mayoría de las hembras colectadas en el lago de Tota
para el mes de noviembre se encontraban en estadio gonadal de madurez
avanzado muy próximo al desove, indicando una posible época reproductiva
para comienzos de año.
9.3 Relación longitud – peso
Forero y Garzón (1974) reportan que en el embalse de Neusa los promedios
de longitud y peso fueron de 65.23mm y 5.54g para la población, 76.19mm y
7.90g para machos, 67.56mm y 5.07g para hembras, y 23.82mm y 0.28g
para alevinos. León et al. (2005) indican que para el embalse de San Rafael,
donde no se separó por machos, hembras o alevinos, los promedios de
longitud y peso fueron de 48.45mm y 2.15g respectivamente.
En el presente estudio los promedios de talla y peso en el río Ubaté, Tibitoc y
embalse de Tominé se asemejaron a los reportados para el embalse de San
Rafael; mientras que en el lago de Tota-Los Pozos los valores fueron
similares a los reportados en el Neusa. Rivera-Rondón et al. (2007) señalan
que en zonas dominadas por la macrófita E. densa los individuos de G.
bogotensis son de menor tamaño, sin embargo, esa condición no fue clara
en este estudio probablemente porque los muestreos no fueron selectivos,
encontrándose sectores donde no había dominancia de una sola especie de
macrófita (ej: laguna de Fúquene).
A nivel de sexos, los machos del río Ubaté (3.561), laguna de Cucunubá
(3.287), embalse El Hato (3.740), lago de Tota-Los pozos (3.574), quebrada
Mana de Ramos (3.426), vallado La Susana (3.387), y quebrada Mancilla
(3.735) presentaron crecimiento alométrico positivo (b>3). En las restantes el
crecimiento en machos fue isométrico (b=3) y en ninguna localidad se

75
presentó crecimiento alométrico negativo (b<3) como lo reportó Forero y
Garzón (1974) para los machos de la represa del Neusa (b=2.08). Esta
variación intraespecífica en la relación longitud – peso puede ser substancial,
dependiendo de la temporada, la población, o las diferencias anuales en las
condiciones ambientales (Froese 2006).
En las hembras, las del río Ubaté (3.558), laguna de Cucunubá (4.639),
Tibitoc (3.456), lago de Tota-Los Pozos (3.453), lago de Tota-Upía (3.460),
quebrada Mana de Ramos (3.188), vallado La Susana (3.528) y quebrada
Mancilla (3.455) presentaron crecimiento alométrico positivo (b>3). En las
restantes se presentó isometría (b=3).
Se reconoce que las diferencias en el exponente b se deben a factores entre
los que se incluyen la fase de crecimiento, temporada, grado de llenura de
los estómagos, madurez gonadal, sexo, técnicas de preservación, y salud y
condición general del pez (Tesch 1971, en Harrison 2001). Aunque la
mayoría de estos factores no se tomaron en cuenta durante el desarrollo del
presente trabajo, el alto valor del parámetro b que se obtuvo para las
hembras de G. bogotensis en algunas localidades posiblemente está
influenciado por el número considerable de hembras que poseían huevos
maduros y cuyo alto peso originó un efecto alométrico positivo, cabe
destacar que los meses de muestreo corresponden a la primera temporada
de reproducción reportada para la especie (Forero y Garzón 1974). El
anterior evento ha sido reportado en otras especies de peces continentales
(Ruiz-Campos et al. 2006).
El rango dentro del cual se debería encontrar el parámetro b es entre 2.5 y
3.5 (Le Cren 1951, Froese 2006). Cuando se estimó la relación longitud –
peso para la totalidad de capturas por localidad, en el río Ubaté, laguna de
Cucunubá, y quebrada Mancilla b superó el establecido 3.5 (considerando

76
tres decimales), sucedió lo mismo con los machos del río Ubaté, embalse El
Hato, lago de Tota-Los Pozos, quebrada Mancilla, y con las hembras del río
Ubaté, laguna de Cucunubá, y vallado La Susana. Carlander (1977, en
Froese 2006) demostró que valores del parámetro b <2.5 ó >3.5 se derivan
generalmente de muestras con un n bajo para los diferentes rangos de talla,
estos resultados fueron además confirmados por Froese (2006) para una
muestra de 1773 especies donde el 90% de los valores de b se encontraban
entre 2.7 y 3.4.
En general, el coeficiente de determinación (r²) de la mayoría de localidades
fue superior a 0.90 indicando que un alto porcentaje de la variabilidad de los
datos se explica por la asociación entre las variables longitud – peso (Pauly
1984).
9.4 Aspectos tróficos
Los grupos más abundantes dentro de la dieta de G. bogotensis fueron
insectos pertenecientes a los órdenes Coleoptera (Chrysomelidae cf.) y
Diptera (larvas y pupas de Chironomidae), detrito, restos de plantas, además
de microcrustáceos, principalmente el anfípodo Hyalella sp., cladóceros de la
familia Daphniidae y Bosminidae, y copépodos del orden Cyclopoida. En
otras especies de pequeño porte pertenecientes a la familia Characidae y
que habitan en zonas con presencia de macrófitas también se ha encontrado
que su dieta se compone principalmente de ítems autóctonos, representados
por microcrustáceos e insectos inmaduros del orden Diptera (Casatti et al.
2003).
De acuerdo a la distribución de los puntos en la diagonal que valora la
importancia de la presa dentro del gráfico de Costello modificado por
Amundsen et al. (1996), la dieta de G. bogotensis no exhibió presas
dominantes. La mayoría de los puntos se ubicaron en el ángulo inferior

77
izquierdo del gráfico (baja frecuencia de ocurrencia y baja abundancia
específica). Los ítems raros y que además presentaron un bajo valor para el
índice de preponderancia (IP), el caso de huevos de invertebrados, escamas,
semillas, larvas de Ceratopogonidae, y Hemiptera (Corixidae, Notonectidae),
también fueron considerados por Forero y Garzón (1974), para la represa del
Neusa, como poco importantes en la dieta.
En el eje dentro del gráfico de Costello modificado por Amundsen et al.
(1996) que se refiere a la estrategia alimentaria, la mayoría de los puntos se
localizaron en la parte baja de la gráfica sugiriendo una generalización de la
dieta. Es decir, que no hay una preferencia acentuada por una fuente de
alimento lo que se traduce en un amplio espectro de alimentos consumidos
(33 ítems diferentes). Esta condición generalista, que se presenta en un gran
número de especies del trópico, significa que la especie es potencialmente
capaz de utilizar todos los recursos que sean adecuados a su aparato
alimentario, capacidad digestiva y táctica alimentaria (Gerking 1994, en
Abelha et al. 2001), entendiendo por táctica las características variables en el
patrón, en respuesta a las fluctuaciones del ambiente (Vazzoler 1996).
Tanto los resultados de la importancia de la presa como los de la estrategia
alimentaria coinciden con estudios realizados para G. bogotensis, donde se
reporta que su dieta se compone principalmente de insectos y
microcrustáceos. Forero y Garzón (1974) indican que los ítems con mayor
abundancia fueron las familias de dípteros Chironomidae y Ceratopogonidae,
con más del 50%, y el copépodo Cyclops spp. (35%) además de otros
insectos no identificados. Mora et al. (1992) en el lago de Tota, mediante el
análisis de 13 estómagos, determinaron que la dieta de la guapucha está
dominada por el copépodo Boeckella gracilis con el 95%, pero indican que la
población adulta consume con preferencia el anfípodo Hyalella sp. y los
cladóceros Eurycercus sp., Neobosmina sp., Graptoleberis sp. y Chydorus

78
sp., así como larvas de Chironomidae, pupas de Diptera y el bivalvo Pisidium
sp., mientras que Gompertz (1986) señala que la dieta se compone
principalmente de los órdenes Insecta con 21% y Crustacea con 16%. De
acuerdo a la composición de la dieta diferentes autores han convenido en
clasificar a la especie como carnívora (Forero y Garzón 1974, Gompertz
1986) no obstante basándose en literatura reciente convendría clasificarle
como invertívora concepto que define a una especie que se alimenta de
invertebrados (Casatti et al. 2003, Gomiero y Braga 2005).
Los puntos en la gráfica de Costello localizados en la esquina superior
izquierda e inferior derecha representan tipos de presas que contribuyen de
manera similar a la dieta de la especie, pero indican localidades donde las
estrategias alimentarias fueron diferentes (Amundsen et al. 1996). La
posición de presas como larvas de Chironomidae, restos de plantas, Hyalella
sp., Cyclopoida, Daphniidae, y Trichoptera no determinado (esquina inferior
derecha), muestra que estos recursos se utilizaron en varias localidades al
mismo tiempo y presentan abundancia similar; mientras que la posición del
coleóptero Chrysomelidae cf., y en cierto grado pupas de Chironomidae,
señala que este ítem ha sido consumido en pocas localidades (tan sólo en
una para Chrysomelidae cf.).
Lo anterior puede estar relacionado con que la dieta y en particular la
plasticidad trófica de los peces de agua dulce, que es la habilidad para
aprovechar una fuente de alimento más ventajosa en determinado tiempo
(Gerking 1994, en Abelha et al. 2001), está influenciada por las condiciones
ambientales y espaciales particulares (Sendacz et al. 2006), por la biología
de cada especie, abundancia específica de la presa, variaciones individuales,
y estrategias alimentarias -patrón general de alimentación mostrado por una
especie (Vazzoler 1996)- (Abelha et al. 2001). Aunque en este estudio no se
propuso distinguir un patrón general entre estas variables y la dieta, en la

79
literatura es ampliamente citada su relación y a partir de la cual se podrían
sugerir algunas de las posibles causas de los resultados en ciertas
localidades.
En este estudio la composición de la dieta y la abundancia porcentual
volumétrica de los ítems presentaron variaciones por localidad, apoyando la
tendencia que se observa en los resultados presentados en Forero y Garzón
(1974), Gompertz (1986) y Mora et al. (1992). Bispo et al. (2006) sugieren
que una parte importante de la variación de las abundancias de insectos
acuáticos está determinada por la estacionalidad climática regional y su
interacción dentro de las características de los cuerpos de agua (descarga,
cobertura de la vegetación, y estabilidad del sustrato).
Rozas y Odum (1988) observaron que los anfípodos, dípteros
(Chironomidae), Odonata y otras larvas de insectos son abundantes sobre
los tallos y raíces de macrófitas sumergidas, además de que poseen un
mayor tamaño que en las zonas sin vegetación acuática. Stansfield et al.
(1997) determinaron que la abundancia de algunos géneros de cladóceros
de la familia Daphniidae ha mostrado una positiva asociación, relacionada
con la utilización como refugio, con las macrófitas y el incremento de su
cobertura, pero aclaran que este aprovechamiento está restringido a cortos
intervalos de tiempo debido al comportamiento ecológico del grupo que es
típicamente pelágico (aguas abiertas).
Respecto a la distribución espacial y explotación de los recursos para
alimento, la composición de la dieta indica que G. bogotensis consume
presas que se localizan tanto en el fondo como a lo largo de la columna de
agua. La presencia de granos de arena y detrito junto con larvas bentónicas
de insectos (ej: Chironomidae, Ceratopogonidae) se ha considerado como un
indicador de que el alimento está tomándose del fondo (Casatti et al. 2003).

80
Mientras que el consumo de presas que son típicamente pelágicas, el caso
de cladóceros de la familia Daphniidae (Stansfield et al. 1997), sustenta esta
apreciación. La utilización de toda la columna de agua por parte de la
guapucha ha sido recientemente relatada por Rivera-Rondón et al. (2007)
para la laguna de Fúquene.
Los valores más bajos para amplitud de nicho corresponden a localidades
donde se evidencia dominancia de uno o dos ítems. En el caso del río Ubaté
las pupas de Chironomidae y los restos de insectos representaron más del
85% del alimento consumido, mientras que en la quebrada Mana de Ramos
el coleóptero de la familia Chrysomelidae cf. representó más del 80%. En el
caso del río Ubaté la posible causa sea una alta disponibilidad de un ítem en
particular (pupas de Chironomidae) ya que el fenómeno de emergencia en
poblaciones de esta familia de dípteros muestra una directa correlación con
la temperatura del agua mediada por la temporada de lluvias (Lobinske et al.
1996).
Para el caso de la quebrada Mana de Ramos se debe destacar que aunque
el análisis de la dieta se realizó mediante la agrupación de los contenidos
estomacales de cada localidad sólo un individuo adulto consumió la larva de
Chrysomelidae cf., al respecto se ha encontrado que en algunos peces el
grado de especialización individual aumenta con el incremento de la talla en
la población adulta (Svanbäck y Persson 2004).

81
10. Conclusiones
10.1 Abundancia y caracterización del hábitat
En las localidades que pertenecían a la cuenca del río Bogotá, y donde se
empleó un esfuerzo de pesca similar a las del Suárez, los valores de
abundancia fueron menores, lo cual podría indicar que el hábitat de G.
bogotensis presenta mejores condiciones en esta última cuenca.
G. bogotensis habita en cuerpos de agua lénticos y lóticos con corrientes de
baja velocidad, y se encuentra asociada principalmente a las macrófitas E.
crassipes, E. densa y Schoenoplectus sp.
La velocidad de la corriente (ríos Sáchica y Sutamarchán) y la ausencia de
macrófitas (embalses del Sisga y San Rafael) pudieron influir para que en
determinadas localidades no se colectaran ejemplares.
La captura de G. bogotensis en cuerpos de agua con características
eutróficas (bajas concentraciones de oxígeno disuelto, alta conductividad, y
gran cantidad de macrófitas), demuestra que este pez puede tolerar
condiciones con algún grado de disturbio.
No se pudo establecer relación entre los valores de conductividad para cada
localidad con el número de ejemplares colectados en ella.
10.2 Proporción de sexos
La proporción de sexos cambió de acuerdo a la localidad. No fue clara la
razón de las diferencias de las proporciones, principalmente por la escala
temporal empleada en este estudio; sin embargo, una causa probable sea
los diferentes tipos de cuerpos de agua en los que se desarrolló el estudio, o
los eventos migratorios que obedecen a un comportamiento reproductivo y
que ya han sido reportados para la especie.

82
La disminución de hembras a partir de los 40.10mm de longitud total y su
desaparición a partir de los 80.10mm apoyan la hipótesis formulada por otros
autores donde se sostiene que en G. bogotensis la diferencia en la
proporción de sexos está relacionada con una ventaja particular para uno de
los sexos o con una mortalidad diferencial.
La diferenciación entre machos y hembras se logró realizar desde los
26.00mm de longitud total en hembras y 29.51mm en machos.
10.3 Relación longitud - peso
El valor del parámetro b de la relación longitud – peso en G. bogotensis
presentó variaciones de acuerdo a la localidad.
En el río Ubaté, laguna de Cucunubá, lago de Tota-Los Pozos, quebrada
Mana de Ramos, vallado La Susana, y quebrada Mancilla, el crecimiento
tanto de hembras como de machos fue alométrico positivo (b>3). Los
machos del embalse El Hato, y las hembras de Tibitoc y lago de Tota-Upía,
también presentaron crecimiento alométrico positivo (b>3). En las demás
localidades los ejemplares presentaron crecimiento isométrico (b=3).
En el caso de las hembras que presentaron un alto valor del parámetro b
este está influenciado por el número considerable de hembras que poseían
huevos maduros y cuyo alto peso originó un efecto alométrico positivo.
10.4 Dieta
La abundancia porcentual volumétrica y la composición de la dieta de G.
bogotensis indica que, aún cuando se presentaron variaciones entre
localidades, la especie es carnívora (invertívora de acuerdo a literatura
reciente).

83
Los insectos y microcrustáceos de origen autóctono son la principal fuente de
alimento para G. bogotensis. De los primeros el orden Diptera (larvas y
pupas de Chironomidae) fueron los más importantes, y de los segundos el
anfípodo Hyalella sp., cladóceros de las familias Daphniidae y Bosminidae, y
los copépodos del orden Cyclopoida.
En la dieta de G. bogotensis no se presentaron presas dominantes. La
estrategia alimentaria es generalista aunque existieron localidades donde,
por probables comportamientos individuales o abundancia de la presa, se
consumió con preferencia uno o dos ítems. En los casos de preferencia por
una presa en particular los valores para amplitud de nicho presentaron los
menores valores.
El consumo de larvas bentónicas y microcrustáceos de hábitos pelágicos
indica que la especie está utilizando la columna de agua tanto a nivel de
fondo como a lo alto de la misma.
11. Recomendaciones
Para la laguna de Suesca se recomienda el desarrollo de estudios que
tengan como objetivo comprobar si se están generando fenómenos de
extinción a nivel local.
Se deben llevar a cabo investigaciones donde se consideren un mayor
número de variables que puedan estar afectando el método de electropesca,
esto con el fin de estandarizar el método para cuerpos de agua altoandinos.
Se deben desarrollar investigaciones donde se determinen los rangos de
oxígeno disuelto y pH a los cuales la guapucha puede sobrevivir.

84
Se deben realizar estudios que profundicen en aspectos que puedan estar
influenciando la abundancia y la proporción sexual intraespecífica. En el caso
de la migración a cuerpos de agua aledaños para desove, se recomienda
debe realizar un seguimiento que incorpore más períodos de muestreo y
observaciones detalladas sobre el comportamiento de la especie.
Para determinar que variables están afectando el valor del parámetro b en la
relación longitud – peso, en futuros estudios de deben incluir aspectos como
fases de crecimiento y condiciones ambientales locales por un periodo
extendido de tiempo.
En los casos donde el valor del parámetro b fue inferior a 2.5 o mayor a 3.5,
al estimar la relación longitud – peso se debe buscar que los rangos de talla
estén representados por un mayor número de individuos.
Aunque la agrupación de los contenidos estomacales por localidad
representó una ganancia significativa de tiempo, la pérdida de datos e
información individual resultante debe considerarse al momento de
implementar el método gráfico utilizado en este estudio.

85
12. Referencias
Abelha, M.C.F., A.A. Agostinho y E. Goulart. 2001. Plasticidade trófica em
peixes de água doce. Acta Scientiarum 23(2): 425-434.
Alabaster, J.S. y W.G. Hartley. 1962. The efficiency of a direct current electric
fishing method in trout streams. J. Anim. Ecol. 31: 385-388.
Alvarado, H. y F.P. Gutiérrez. 2002. Especies hidrobiológicas continentales
introducidas y transplantadas y su distribución en Colombia. Ministerio
de Medio Ambiente – Instituto de Investigaciones Alexander von
Humboldt. Bogotá D.C. 167p.
Álvarez-León, R., J.A. González y J.E. Forero. 2002. Grundulus bogotensis.
Pp.200-202. En: Mojica, J.I., Castellanos, C., Usma, S. y Álvarez, R.
(Eds.). 2002. Libro rojo de peces dulceacuícolas de Colombia. La serie
Libros Rojos de Especies Amenazadas de Colombia. Instituto de
Ciencias Naturales, Universidad Nacional de Colombia, Ministerio del
Medio Ambiente, Bogotá, Colombia.
Amundsen, P.A., H.M. Gabler, y F.J. Staldvik. 1996. A new approach to
graphical analysis of feeding strategy from stomach contents data –
modification of the Costello (1990) method. Journal of Fish Biology 48:
607-614.
Anbumozhi, V., J. Radhakrishnan, y E. Yamaji. 2005. Impact of riparian buffer
zones on water quality and associated management considerations.
Ecological Engineering 24: 517–523.

86
Barbour, M.T., J. Gerritsen, B.D. Snyder, y J.B. Stribling. 1999. Rapid
bioassessment protocols for use in streams and wadeable rivers:
periphyton, benthic macroinvertebrates and fish. Segunda edición.
EPA 841-B-99-002. U.S. Environmental Protection Agency; Office of
Water; Washington, D.C. 339 p.
Bispo, P.C., L.G. Oliveira, L.M. Bini, y K.G Sousa. 2006. Ephemeroptera,
Plecoptera and Trichoptera assemblages from riffles in mountain
streams of central Brazil: environmental factors influencing the
distribution and abundance of immatures. Brazilian Journal of Biology
66(2B): 611-622.
Bohlin, T., S. Hamrin, T.G. Heggberget, G. Rasmussen y S.J. Saltveit’. 1989.
Electrofishing - Theory and practice with special emphasis on
salmonids. Hydrobiologia 173: 9-43.
Bolner, K.C.S. y B. Baldisserotto. 2007. Water pH and urinary excretion in
silver catfish Rhamdia quelen. Journal of Fish Biology 70: 50–64.
CAR. 2002. Atlas Ambiental CAR 2001. Primera edición. Corporación
Autónoma Regional de Cundinamarca, CAR. Bogotá. 175 p.
CAR. 2006. Diagnóstico prospectiva y formulación de la cuenca hidrográfica
de los ríos Ubaté y Suárez - Plan de Ordenamiento de la Cuenca de
los Ríos Ubaté y Suárez. Informe General. 289 p.
Casatti, L., H.F. Mendes y K.M. Ferreira. 2003. Aquatic macrophytes as
feeding site for small fishes in the Rosana reservoir, Paranapanema
river, southeastern Brazil. Brazilian Journal of Biology 63(2): 213-222.

87
Chará J. y E. Murgueitio. 2005. The role of silvopastoral systems in the
rehabilitation of Andean stream habitats. Livestock Research for Rural
Development 17 (2);
http://www.cipav.org.co/lrrd/lrrd17/2/char17020.htm [Consulta: 17 mar.
2006]
Cognato, D. de P. y C.B. Fialho. 2006. Reproductive biology of a population
of Gymnotus aff. carapo (Teleostei: Gymnotidae) from southern Brazil.
Neotropical Ichthyology 4(3): 339-348.
Colwell, R.K. y D.J. Futuyma. 1971. On the Measurement of Niche Breadth
and Overlap. Ecology 52(4): 567-576.
Cone, R.S. 1989. The need to reconsider the use of condition indices in
fishery science. Transactions of the American Fisheries Society 118:
510-514.
Conover, D.O. y B.E. Kynard. 1981. Environmental sex determination:
Interaction of temperature and genotype in a fish. Science 213(4507):
577-579.
CORPOBOYACÁ. 2002. Plan de gestión ambiental regional 2002-2011 -
Resumen ejecutivo. 215 p.
Dahl, G. 1971. Los peces del norte de Colombia. Ministerio de Agricultura,
Instituto de Desarrollo de los Recursos Naturales Renovables –
INDERENA. Bogotá. 390 p.

88
Deus, C.P. y Petrere-Junior, M. 2003. Seasonal diet shifts of seven fish
species in an atlantic rainforest stream in Southeastern Brazil.
Brazilian Journal of Biology 63(4): 579-588.
Eigenmann, C.H. 1915. The Cheirodontinae, a subfamily of minute characid
fishes of South America. Memoirs Carnegie Museum 7: 1-99.
Eigenmann, C.H. 1917. The American Characidae. Mem. Mus. Comp. Zoöl.
XLIII Part 1: 1-102.
Eigenmann, C.H. 1921. The origin and distribution of the genera of the fishes
of South America West of the Maracaibo, Orinoco, Amazon, and
Titicaca basins. Proceedings of the American Philosophical Society
60(1): 1-6.
Escobar, B.N. 1991. Fitoplancton y aspectos fisicoquímicos de la laguna de
Suesca, departamento de Cundinamarca. Trabajo de grado
presentado como requisito parcial para optar al título de biólogo.
Pontificia Universidad Javeriana, Facultad de Ciencias Básicas.
Bogotá, Colombia.
Forero, J.E. y M.R. Garzón. 1974. Ciclo biológico de la guapucha, Grundulus
bogotensis (Humboldt, 1821) (Pisces: Characidae), de la Sabana de
Bogotá. Tesis de Pregrado, Universidad Nacional de Colombia,
Facultad de Ciencias, Bogotá, Colombia.
Fowler, H.W. 1942. Lista de peces de Colombia. Rev. Acad. Colomb. Cien.
5(17): 128-138.

89
Froese, R. 2006. Cube law, condition factor and weight–length relationships:
history, meta-analysis and recommendations. J. Appl. Ichthyol. 22:
241–253.
Froese, R. y D. Pauly (Eds). 2006. FishBase. World Wide Web electronic
publication. www.fishbase.org, version (06/2006). [Consulta: 29 sept.
2006].
Gaviria, S. y N. Aranguren. 2003. Guía de laboratorio para identificación de
Cladóceros (Anomopoda y Ctenopoda) y Copépodos (Calanoida y
Cyclopoida). Universidad Pedagógica y Tecnológica de Colombia –
UPTC. Tunja, Boyacá, Colombia.
Géry, J. 1977. Characoids of the world. T.F.H. Publications, New Jersey. 672
p.
Gomiero, L.M. y F.M. Braga. 2005. Uso do grau de preferência alimentar para
a caracterização da alimentação de peixes na APA de São Pedro e
Analândia. Acta Sci. Biol. Sci. 27(3): 265-270.
Gompertz, M.P. 1986. Distribución de la fauna ictiológica en la sabana de
Bogotá y alrededores. Trabajo de grado presentado como requisito
parcial para optar al título de biólogo. Pontificia Universidad Javeriana.
Facultad de Ciencias. Bogotá, Colombia.
González, A.J., M.L. Bueno y J.E. Forero. 1992. Caracterización
cromosómica de dos especies ícticas nativas; Guapucha, (Grundulus
bogotensis) y Capitán, (Eremophilus mutisii), de la sabana de Bogotá.
Acta Biológica Colombiana 7-8: 45-54.

90
Grenouillet, G., Pont, D., y Seip, K.L. 2002. Abundance and species richness
as a function of food resources and vegetation structure: juvenile fish
assemblages in rivers. Ecography 25: 641–650.
Haggarty, D.R. y J.R. King. 2006. CPUE as an index of relative abundance
for nearshore reef fishes. Fisheries Research 81(1): 89-93.
Harley, S.J., R.A. Myers, y A. Dunn. 2001. Is catch-per-unit-effort proportional
to abundance? Canadian Journal of Fisheries and Aquatic Sciences
58(9): 1760-1772.
Harrison, T.D. 2001. Length-weight relationships of fishes from South African
estuaries. J. Appl. Ichthyol. 17: 46-48.
Hyslop, E.J. 1980. Stomach contents analysis – A review of methods and
their application. Journal of Fish Biology 17: 411-429.
IGAC. 1984. Estudio regional integrado del Altiplano Cundiboyacense.
Proyecto IGAC-ORSTOM. Bogotá. 498 p.
Krebs, C.J. 1999. Ecological methodology. 2nd. ed., Addison Wesley
Longman, NY, USA. 620 p.
Le Cren, E.D. 1951. The length-weight relationship and seasonal cycle in
gonad weight and condition in the Perch (Perca fluviatilis). Journal of
Animal Ecology 20(2): 201-219.
León, L.D., J.A. González y J.E. Forero. 2005. Aspectos biométricos de la
guapucha, Grundulus bogotensis (Pisces: Characidae). Dahlia - Rev.
Asoc. Colomb. Ictiol. 8:71-77.

91
Lizama, M. de los A. P. y A.M. Ambrósio. 2002. Condition factor in nine
species of fish of the Characidae family in the upper Paraná river
floodplain, Brazil. Brazilian Journal of Biology 62(1): 113-124.
Lobinske R.J., A. Arshad y I.J. Stout. 1996. Qualitative and quantitative
studies on Chironomidae (Diptera) and selected physico-chemical
parameters in two tributaries of the Wekiva river, central Florida.
Florida Entomologist 79(4): 531-542.
Lobón-Cerviá, J. 1991. Dinámica de poblaciones de peces en ríos. Pesca
eléctrica y métodos de capturas sucesivas en la estima de
abundancias. Monografías del Museo Nacional de Ciencias Naturales.
Centro de Investigaciones de Agua (C.S.I.C.). Madrid. 156 p.
Luz-Agostinho, K.D.G. L.M. Bini, R. Fugi, A.A. Agostinho, y H.F. Júlio Jr.
2006. Food spectrum and trophic structure of the ichthyofauna of
Corumbá reservoir, Paraná river Basin, Brazil. Neotropical Ichthyology
4(1):61-68.
Magurran, A.E. y D.A.T. Phillip. 2001. Implications of species loss in
freshwater fish assemblages. Ecography 24: 645–650.
Malabarba, L.R. 1998. Monophyly of the Cheirodontinae, characters and
major clades (Ostariophysi: Characidae). Pp. 193-233 en: Malabarba,
L.R., R.E. Reis, R.P. Vari, Z.M.S. Lucena y C.A.S. Lucena (eds.).
Phylogeny and classification of Neotropical fishes. EDIPUCRS. Porto
Alegre, Brasil.

92
Maldonado-Ocampo, J.A., A. Ortega-Lara, J.S. Usma, G. Galvis, F.A. Villa-
Navarro, L. Vásquez, S. Prada-Pedreros y C. Ardila. 2005. Peces de
los Andes de Colombia. Instituto de Investigaciones de Recursos
Biológicos «Alexander von Humboldt». Bogotá – Colombia. 346 p.
Mancera-Rodríguez, N.J. y R. Álvarez-León. 2006. Estado del conocimiento
de las concentraciones de mercurio y otros metales pesados en peces
dulceacuícolas de Colombia. Acta Biológica Colombiana 11(1): 3-23.
Marshall, S. y M. Elliott. 1997. A comparison of univariate and multivariate
numerical and graphical techniques for determining inter- and
intraspecific feeding relationships in estuarine fish. Journal of Fish
Biology 51: 526-545.
Marrero, C. 1994. Métodos para cuantificar contenidos estomacales en
peces. Museo de Zoología - Universidad de los Llanos Ezequiel
Zamora (UNELLEZ). 37 p.
McCafferty, W.P. y A.V. Provonsha. 1981. Aquatic entomology. Science
Books Internacional, Boston. 448 p.
McKinsey, D.M. y L.J. Chapman. 1998. Dissolved oxygen and fish distribution
in a Florida spring. Environmental Biology of Fishes 53: 211-223.
Merritt, R.W. y K.W. Cummins. 1984. An introduction to the aquatic insects of
North America. Second Edition. Kendall/Hunt Publishing Co. Dubuque,
Iowa. 722 p.
Miles, C. 1971. Los peces del río Magdalena. 2ª. Edición, Universidad del
Tolima, Ibagué, Colombia. 214 p.

93
Miller, D.C., S.L. Poucher, y L. Coiro. 2002. Determination of lethal dissolved
oxygen levels for selected marine and estuarine fishes, crustaceans,
and a bivalve. Marine Biology 140: 287-296.
Mojica, J.I. (1999). Lista preliminar de las especies de peces dulceacuícolas
de Colombia. Rev. Acad. Colomb. Cienc. 23 (Supl. Especial): 547-566.
Mojica, J.I., C. Castellanos, S. Usma y R. Álvarez-León (Eds.). 2002. Libro
Rojo de peces dulceacuícolas de Colombia. La serie Libros Rojos de
Especies Amenazadas de Colombia. Instituto de Ciencias Naturales,
Universidad Nacional de Colombia, Ministerio del Medio Ambiente,
Bogotá, Colombia. 288 p.
Mora, G., L.S. Téllez, P. Cala y G. Guillot. 1992. Estudio bioecológico de la
ictiofauna del lago de Tota (Boyacá-Colombia), con énfasis en la
trucha arco iris, Oncorhynchus mykiss. Rev. Acad. Colomb. Cienc.
18(70): 409-422.
Murgueitio, E.M. y Z. Calle. 1999. Diversidad biológica en sistemas de
ganadería bovina en Colombia. En: Sánchez, M. y M. Rosales (Eds.)
Agroforestería para la producción animal en Latinoamérica. Estudio
FAO sobre la producción y sanidad animal 143. Roma pp 53-88.
Murgueitio, E. y M. Ibrahim. 2001. Agroforestería pecuaria para la
reconversión de la ganadería en Latinoamérica. Livestock Research
for Rural Development (13) 3;
http://www.cipav.org.co/lrrd/lrrd13/3/murg133.htm

94
Pantoja-Agreda, F. y C. Pantoja-Agreda. 2003. Hábitos alimentarios de
Grundulus sp. (Pises: Characidae) en el lago Guamuez, departamento
de Nariño, Colombia. Dahlia – Rev. Asoc. Colomb. Ictiol. 6: 113-115.
Pauly, D. 1984. Fish population dynamics in tropical waters: A manual for use
with programmable calculators. International Center for Living Aquatic
Resources Management – ICLARM. Manila, Philippines. 325 p.
Penczak, T., A.A. Agostinho, Ł. Głowacki y L.C. Gomes. 1997. The effect of
artificial increases in water conductivity on the efficiency of electric
fishing in tropical streams (Paraná, Brazil). Hydrobiologia 350: 189–
201.
Pinto-Coelho, R.M., J.F. Bezerra-Neto, y C.A. Morais-Jr. 2005. Effects of
eutrophication on size and biomass of crustacean zooplankton in a
tropical reservoir. Brazilian Journal of Biology 65(2): 325-338.
Posada-Arango, A. 1909. Los peces: contribución al estudio de fauna
colombiana. Pp. 285-322. En: Estudios científicos del doctor Andrés
Posada con algunos otros escritos suyos sobre diversos temas y con
ilustraciones o grabados. Imprenta Oficial, Medellín, Colombia. 432 p.
Prejs, A. y G. Colomine. 1981. Métodos para el estudio de los alimentos y las
relaciones tróficas de los peces. Instituto de Zoología Tropical (IZT) –
Universidad Central de Venezuela, Imprenta Universitaria. Caracas.
129 p.
Rice, J.C. 2005. Understanding fish habitat ecology to achieve conservation.
Journal of Fish Biology 67 (Suppl. B): 1-22.

95
Rivera-Rondón, C.A., S. Prada-Pedreros, D. Galindo y J.A. Maldonado-
Ocampo. 2007. Efecto de la vegetación acuática sobre la distribución
espacial de Grundulus bogotensis (Humboldt, 1821) (Pisces:
Characidae). Documento de trabajo, 33 p.
Roa, N. 1998. Delimitación, diagnóstico y priorización de los ecosistemas
estratégicos en la jurisdicción de CORPOBOYACÁ. Documento
técnico: 31-44.
Robbins, R.L. 2007. Environmental variables affecting the sexual segregation
of great white sharks Carcharodon carcharias at the Neptune Islands
South Australia. Journal of Fish Biology 70: 1350–1364.
Rodríguez, N., D. Armenteras, M. Morales y M. Romero. 2004. Ecosistemas
de los Andes colombianos. Instituto Alexander von Humboldt, Bogotá.
155 p.
Roldán, G. 1996. Guía para el estudio de los macroinvertebrados acuáticos
del departamento de Antioquia. Universidad de Antioquia, Facultad de
Ciencias Exactas y Naturales. Centro de Investigaciones - CIEN,
Medellín. Fondo FEN Colombia, COLCIENCIAS, Bogotá. 217 p.
Román-Valencia, C., Hans-Joachim Paepke y F. Pantoja. 2003 (“2001”). Una
especie nueva de Grundulus (Teleostei: Ostariophysi: Characidae)
para Colombia y redescripción de Grundulus bogotensis (Humboldt y
Valenciennes, 1833). Mem. Fund. La Salle Cienc. Nat. 155: 51-72.
Román-Valencia, C., Ruiz R.I. y Barriga, R. 2005. Una nueva especie
ecuatoriana del género de peces andinos Grundulus (Characiformes:
Characidae). Rev. Biol. Trop. (Int. J. Trop. Biol.) 53 (3-4): 537-544.

96
Rozas, L.P. y W.E. Odum. 1988. Occupation of submerged aquatic
vegetation by fishes: testing the roles of food and refuge. Oecologia
77:101-106.
Rubin, D.A. 1985. Effect of pH on sex ratio in Cichlids and a Poecilliid
(Teleostei). Copeia 1985(1): 233-235.
Ruiz-Campos, G., A. F. González-Acosta, y J. De La Cruz-Agüero. 2006.
Length–weight and length–length relationships for some continental
fishes of northwestern Baja California, México. J. Appl. Ichthyol. 22:
314–315.
Sendacz, S., S. Caleffi, y J. Santos-Soares. 2006. Zooplankton biomass of
reservoirs in different trophic conditions in the state of São Paulo,
Brazil. Brazilian Journal of Biology 66(1B): 337-350.
Sirangelo, A. y N. Ferreira. 2004. Reproductive biology of Menticirrhus
littoralis in southern Brazil (Actinopterygii: Perciformes: Sciaenidae).
Neotropical Ichthyology 2(1): 31-36.
Smith, E.P. 1982. Niche breadth, resource availability, and inference. Ecology
63(6): 1675-1681.
Soares, M.G., N.A. Menezes, y W.J. Junk. 2006. Adaptations of fish species
to oxygen depletion in a central Amazonian floodplain lake.
Hydrobiologia 568:353–367.
Stansfield, J.H., M.R. Perrow, L.D. Tench, A.J.D. Jowitt y A.A.L. Taylor. 1997.
Submerged macrophytes as refuges for grazing Cladocera against fish

97
predation: observations on seasonal changes in relation to macrophyte
cover and predation pressure. Hydrobiologia 342/343: 229–240.
Svanbäck, R. y L. Persson. 2004. Individual diet specialization, niche width
and population dynamics: implications for trophic polymorphisms.
Journal of Animal Ecology 73: 973-982.
Tejerina-Garro, F.L., M. Maldonado, C. Ibáñez, D. Pont, N. Roset y T.
Oberdorff. 2005. Effects of natural and anthropogenic environmental
changes on riverine fish assemblages: A framework for ecological
assessment of rivers. Brazilian Archives of Biology and Technology
48(1): 91-108.
USEPA. 2000. Ambient aquatic life water quality criteria for dissolved oxygen
(saltwater): Cape Cod to Cape Hatteras. EPA-822-R-00-012.
Narrangansett, RI: United States Environmental Protection Agency. 55
p.
Uieda, V.S. y R.M.C. Castro. 1999. Coleta e fixação de peixes de riachos. Pp.
1-22. En: Caramaschi, E.P., R. Mazzoni, y P.R. Peres-Neto (Eds.).
Ecologia de peixes de riachos. Série Oecologia Brasiliensis vol. VI.
PPGE-UFRJ. Rio de Janeiro, Brasil. 260 p.
Van der Hammen, T. 2001. Bases para un plan de mejoramiento de la laguna
de Fúquene y su cuenca hidrográfica. 33-55 pp. En: Memorias del
Comité de Expertos para la Recuperación de la laguna de Fúquene.
Corporación Autónoma Regional de Cundinamarca, CAR. Bogotá. 89
p.

98
Vazzoler, A.M. 1996. Biologia da reprodução de peixes teleósteos: Teoria e
prática. EDUEM : Editora da Universidade Estadual de Maringá.
Maringá, Brasil. 169 p.
Vicentini, R.N. y F.G. Araújo. 2003. Sex ratio and size structure of
Micropogonias furnieri (Desmarest, 1823) (Perciformes, Sciaenidae) in
Sepetiba Bay, Rio de Janeiro, Brazil. Brazilian Journal of Biology
63(4): 559-566.
Watzin, M.C. y A.W. McIntosh. 1999. Aquatic ecosystems in agricultural
landscapes: A review of ecological indicators and achievable
ecological outcomes. Journal of Soil and Water Conservation 54: 636-
644.
Whittaker, R.H., S.A. Levin y R.B. Root. 1973. Niche, habitat, and ecotope.
The American Naturalist 107(955): 321-338.

99
13. Anexos
Anexo 1. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en la laguna de Fúquene.
Ejemplar
No.Lt (mm) Ls (mm) Wt (g) Sexo
1 55.90 46.12 3.6 macho2 49.72 41.24 2.1 macho3 53.48 44.72 2.7 macho4 55.14 46.54 2.9 hembra5 48.10 41.16 2.1 macho6 52.24 42.74 2.2 macho7 54.32 45.82 2.7 hembra8 48.08 39.90 1.9 macho9 49.42 41.24 2.1 macho10 47.90 40.00 2.0 hembra11 48.12 40.38 1.7 macho12 47.56 40.10 1.6 macho13 47.90 40.30 1.8 macho14 42.58 34.70 1.3 macho15 45.96 37.54 1.5 macho16 45.20 38.20 1.6 macho17 48.30 40.70 2.0 hembra18 43.80 36.90 1.4 macho19 42.28 35.10 1.2 macho20 46.10 39.31 1.6 macho21 44.28 37.00 1.3 macho22 43.90 35.94 1.3 macho23 44.62 37.18 1.3 macho24 44.96 37.76 1.6 macho25 43.10 36.00 1.3 macho26 36.38 38.74 1.8 macho27 45.90 38.40 1.6 macho28 45.30 38.20 1.5 macho29 45.40 33.84 1.4 macho30 43.00 36.26 1.3 macho31 40.44 33.53 0.8 hembra32 44.13 35.96 1.4 macho33 40.17 32.63 1.2 hembra34 37.94 32.04 0.8 macho35 38.65 31.85 0.9 hembra36 40.79 33.82 1.0 hembra37 41.91 34.03 1.0 macho38 40.45 33.61 0.8 macho39 42.48 34.96 1.0 macho40 39.47 32.53 0.9 macho
Ejemplar
No.Lt (mm) Ls (mm) Wt (g) Sexo
41 38.01 31.33 1.0 macho42 40.33 33.38 1.1 macho43 41.18 34.29 1.2 macho44 40.27 32.14 1.0 macho45 38.39 31.77 0.8 hembra46 38.79 31.66 1.1 macho47 27.45 21.61 0.2 alevino48 20.96 16.97 0.1 alevino49 22.29 17.80 0.2 alevino50 19.37 16.03 0.1 alevino51 17.60 14.59 0.1 alevino52 33.32 27.17 0.5 alevino53 14.85 11.71 0.1 alevino54 16.44 12.99 0.1 alevino55 18.73 14.92 0.1 alevino56 25.27 20.45 0.3 alevino57 29.07 22.92 0.2 alevino58 18.87 15.35 0.1 alevino59 17.88 13.15 0.1 alevino60 19.23 16.23 0.1 alevino61 41.09 33.98 1.2 macho62 44.77 37.42 1.5 macho63 40.19 32.30 1.0 macho64 45.05 37.09 1.5 macho65 41.85 34.85 1.3 macho66 41.58 34.57 1.1 hembra67 42.76 34.54 1.2 macho68 45.01 36.77 1.6 macho69 44.58 36.58 1.5 macho70 43.99 35.90 1.4 macho71 45.84 38.16 1.9 macho72 44.40 36.55 1.5 macho73 41.64 34.87 1.2 macho74 42.35 34.06 1.2 macho75 40.83 33.60 1.2 hembra76 39.12 32.04 0.9 macho77 37.74 30.39 0.5 macho78 30.84 25.70 0.6 hembra79 38.24 31.54 1.0 macho80 40.51 33.28 1.0 macho

100
Anexo 2. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en la laguna de Fúquene (continuación).
Ejemplar
No.Lt (mm) Ls (mm) Wt (g) Sexo
81 37.80 34.96 0.9 macho82 43.33 30.96 1.3 macho83 35.32 26.46 0.7 hembra84 32.68 24.29 0.5 hembra85 36.97 33.26 0.7 hembra86 25.16 19.08 0.2 alevino87 19.25 15.78 0.1 alevino88 19.28 16.26 0.1 alevino89 19.03 15.71 0.1 alevino90 18.23 15.38 0.1 alevino91 17.58 14.36 0.1 alevino92 18.69 15.75 0.1 alevino93 18.52 15.11 0.1 alevino94 18.63 14.90 0.1 alevino95 16.40 12.78 0.05 alevino96 18.87 15.64 0.1 alevino97 17.55 13.89 0.05 alevino98 18.75 15.94 0.1 alevino99 17.86 13.62 0.05 alevino
100 17.65 13.01 0.1 alevino101 17.76 13.54 0.05 alevino102 Det. Det. Det. alevino
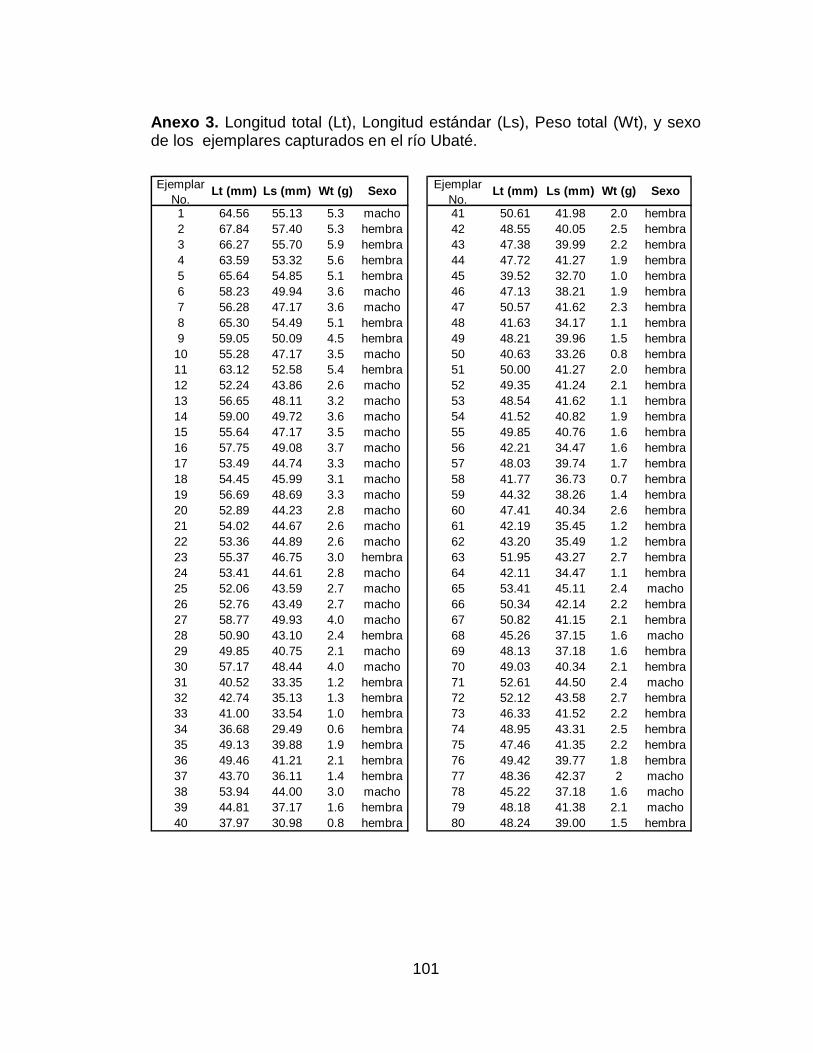
101
Anexo 3. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en el río Ubaté.
Ejemplar
No.Lt (mm) Ls (mm) Wt (g) Sexo
1 64.56 55.13 5.3 macho2 67.84 57.40 5.3 hembra3 66.27 55.70 5.9 hembra4 63.59 53.32 5.6 hembra5 65.64 54.85 5.1 hembra6 58.23 49.94 3.6 macho7 56.28 47.17 3.6 macho8 65.30 54.49 5.1 hembra9 59.05 50.09 4.5 hembra
10 55.28 47.17 3.5 macho11 63.12 52.58 5.4 hembra12 52.24 43.86 2.6 macho13 56.65 48.11 3.2 macho14 59.00 49.72 3.6 macho15 55.64 47.17 3.5 macho16 57.75 49.08 3.7 macho17 53.49 44.74 3.3 macho18 54.45 45.99 3.1 macho19 56.69 48.69 3.3 macho20 52.89 44.23 2.8 macho21 54.02 44.67 2.6 macho22 53.36 44.89 2.6 macho23 55.37 46.75 3.0 hembra24 53.41 44.61 2.8 macho25 52.06 43.59 2.7 macho26 52.76 43.49 2.7 macho27 58.77 49.93 4.0 macho28 50.90 43.10 2.4 hembra29 49.85 40.75 2.1 macho30 57.17 48.44 4.0 macho31 40.52 33.35 1.2 hembra32 42.74 35.13 1.3 hembra33 41.00 33.54 1.0 hembra34 36.68 29.49 0.6 hembra35 49.13 39.88 1.9 hembra36 49.46 41.21 2.1 hembra37 43.70 36.11 1.4 hembra38 53.94 44.00 3.0 macho39 44.81 37.17 1.6 hembra40 37.97 30.98 0.8 hembra
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 50.61 41.98 2.0 hembra42 48.55 40.05 2.5 hembra43 47.38 39.99 2.2 hembra44 47.72 41.27 1.9 hembra45 39.52 32.70 1.0 hembra46 47.13 38.21 1.9 hembra47 50.57 41.62 2.3 hembra48 41.63 34.17 1.1 hembra49 48.21 39.96 1.5 hembra50 40.63 33.26 0.8 hembra51 50.00 41.27 2.0 hembra52 49.35 41.24 2.1 hembra53 48.54 41.62 1.1 hembra54 41.52 40.82 1.9 hembra55 49.85 40.76 1.6 hembra56 42.21 34.47 1.6 hembra57 48.03 39.74 1.7 hembra58 41.77 36.73 0.7 hembra59 44.32 38.26 1.4 hembra60 47.41 40.34 2.6 hembra61 42.19 35.45 1.2 hembra62 43.20 35.49 1.2 hembra63 51.95 43.27 2.7 hembra64 42.11 34.47 1.1 hembra65 53.41 45.11 2.4 macho66 50.34 42.14 2.2 hembra67 50.82 41.15 2.1 hembra68 45.26 37.15 1.6 macho69 48.13 37.18 1.6 hembra70 49.03 40.34 2.1 hembra71 52.61 44.50 2.4 macho72 52.12 43.58 2.7 hembra73 46.33 41.52 2.2 hembra74 48.95 43.31 2.5 hembra75 47.46 41.35 2.2 hembra76 49.42 39.77 1.8 hembra77 48.36 42.37 2 macho78 45.22 37.18 1.6 macho79 48.18 41.38 2.1 macho80 48.24 39.00 1.5 hembra

102
Anexo 4. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el río Ubaté (continuación).
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
81 49.52 41.50 2.1 macho82 50.55 41.55 2.2 hembra83 51.80 42.78 2.2 macho84 45.66 38.56 1.6 macho85 48.53 43.32 2.3 hembra86 40.65 33.34 1.1 hembra87 36.86 30.13 0.7 macho88 37.33 30.95 0.7 hembra89 36.11 29.72 0.6 hembra90 36.16 30.18 0.7 hembra91 36.09 29.21 0.6 hembra92 28.99 25.27 0.4 alevino

103
Anexo 5. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en la laguna de Cucunubá.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 46.07 37.84 1.5 hembra2 45.04 37.65 1.5 hembra3 43.36 36.78 1.3 macho4 44.81 37.52 1.3 macho5 45.37 38.79 1.6 macho6 44.13 36.02 1.2 macho7 42.72 35.48 1.4 macho8 44.35 36.59 1.3 macho9 43.86 36.07 1.2 macho10 46.32 38.87 1.3 macho11 44.00 35.60 1.2 hembra12 44.59 36.97 1.4 macho13 44.44 36.23 1.3 macho14 45.62 37.31 1.4 macho15 42.65 34.45 1.2 macho16 52.91 44.50 2.3 macho17 49.50 41.54 1.7 macho18 50.63 42.41 2.3 macho19 51.06 42.82 2.3 macho20 58.99 50.04 3.6 hembra21 46.73 39.27 1.7 macho22 47.57 39.39 1.8 hembra23 48.41 39.84 1.7 macho24 50.57 41.37 2.0 macho25 46.73 39.33 1.5 macho26 51.09 41.91 1.9 macho27 49.21 41.43 1.8 macho28 45.26 38.45 1.4 macho29 49.91 38.38 1.6 macho30 43.23 36.31 1.4 macho31 44.73 35.81 1.3 hembra32 39.55 32.37 1.0 macho33 35.22 27.89 0.6 hembra34 36.30 29.92 0.8 hembra35 37.40 31.15 0.7 macho36 42.82 34.21 1.0 hembra37 44.81 38.48 1.2 hembra38 42.34 34.24 1.0 macho39 41.98 35.58 1.1 macho40 41.28 34.44 1.1 macho
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 41.15 33.72 1.0 macho42 37.85 31.07 0.8 macho43 39.72 31.52 0.9 macho44 40.48 33.02 1.0 macho45 39.77 33.11 0.9 macho46 44.97 37.28 1.7 macho47 42.83 35.98 1.1 macho48 41.27 33.33 1.0 macho49 37.73 31.42 0.8 hembra50 41.63 33.90 1.0 macho51 42.27 34.64 1.2 macho52 41.04 33.89 1.1 macho53 38.93 31.88 0.9 macho54 43.07 35.49 1.2 macho55 40.99 33.74 1.5 macho56 38.31 31.69 0.8 macho57 39.95 32.69 0.8 macho58 38.98 31.35 0.8 hembra59 36.53 29.53 0.7 macho60 32.87 25.42 0.4 macho61 38.76 32.50 0.9 macho62 37.57 30.13 0.7 hembra63 38.12 31.47 0.8 macho64 32.75 25.90 0.1 hembra65 25.24 19.84 0.4 alevino66 25.76 21.09 0.2 alevino67 26.07 21.07 0.1 alevino68 23.03 17.80 0.1 alevino69 22.74 17.63 0.1 alevino70 23.35 17.94 0.2 alevino

104
Anexo 6. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en el embalse El Hato.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 37.57 30.82 1.0 macho2 45.10 36.71 1.6 hembra3 35.98 29.29 0.9 hembra4 36.99 30.61 0.8 hembra5 48.34 39.86 2.1 macho6 68.90 57.41 5.0 hembra7 40.10 32.12 1.3 macho8 43.02 34.62 1.4 macho9 34.90 29.01 0.7 hembra
10 42.63 35.23 1.3 macho11 33.60 26.58 0.6 hembra12 37.16 30.88 0.8 hembra13 35.64 28.61 0.8 hembra14 34.98 27.92 0.7 macho15 36.51 29.41 0.8 hembra16 40.49 33.33 1.1 macho17 35.91 29.10 0.8 macho18 39.15 32.00 1.0 hembra19 36.16 29.03 0.7 macho20 34.72 28.57 0.7 hembra21 34.15 28.35 0.7 macho22 35.20 28.64 0.7 macho23 34.05 27.72 0.6 macho24 34.50 27.96 0.7 macho25 33.82 27.14 0.6 macho26 34.72 27.88 0.7 hembra27 34.21 27.78 0.6 hembra28 36.78 29.90 0.8 hembra29 35.40 29.31 0.8 macho30 34.61 28.40 0.7 macho31 30.97 25.03 0.4 macho32 29.94 23.81 0.4 macho33 30.75 24.41 0.4 hembra34 29.53 23.48 0.3 hembra35 32.76 26.25 0.5 hembra36 30.22 23.47 0.4 hembra37 28.31 22.45 0.3 alevino38 26.31 20.77 0.2 alevino39 29.57 23.62 0.4 alevino40 25.15 19.98 0.2 alevino
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 28.77 22.47 0.3 alevino42 31.68 25.65 0.4 hembra43 32.72 26.17 0.5 hembra44 28.43 22.45 0.3 alevino45 27.83 21.69 0.3 alevino46 31.46 24.90 0.4 macho47 27.83 21.98 0.3 alevino48 28.03 22.22 0.3 alevino49 30.47 24.30 0.4 hembra50 27.93 21.55 0.3 alevino51 27.49 21.50 0.2 alevino52 27.61 22.13 0.3 alevino53 31.25 25.16 0.4 macho54 26.98 21.53 0.3 alevino55 31.97 25.95 0.5 alevino56 26.22 21.14 0.2 alevino57 39.93 29.82 0.7 hembra58 29.68 23.59 0.4 alevino59 22.61 17.98 0.1 alevino60 31.09 24.79 0.4 macho61 31.05 25.10 0.4 macho62 26.71 21.43 0.3 alevino63 25.55 19.94 0.2 alevino64 30.76 24.52 0.4 hembra65 21.59 17.29 0.1 alevino66 29.70 23.70 0.1 alevino67 19.88 15.69 0.1 alevino68 26.94 21.85 0.2 alevino69 29.29 22.93 0.4 alevino70 29.52 23.32 0.4 alevino71 26.94 20.77 0.3 alevino72 27.71 21.34 0.3 alevino73 29.49 23.46 0.4 alevino74 28.80 22.43 0.3 alevino75 30.33 24.22 0.4 hembra76 31.83 23.36 0.5 hembra77 26.78 21.83 0.2 alevino78 28.52 22.64 0.3 alevino79 27.62 21.18 0.2 alevino80 25.16 19.97 0.2 alevino

105
Anexo 7. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en el embalse El Hato (continuación).
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
81 31.79 26.31 0.4 hembra82 30.17 24.47 0.4 macho83 27.66 20.84 0.3 alevino84 34.39 27.73 0.6 macho85 27.48 21.81 0.3 alevino86 30.13 24.49 0.4 macho87 29.11 23.46 0.4 alevino88 33.74 26.44 0.5 hembra89 28.00 21.74 0.3 alevino90 28.37 22.35 0.3 alevino91 28.69 22.82 0.3 alevino92 33.03 26.11 0.5 hembra93 26.34 20.58 0.2 alevino94 25.11 20.23 0.2 alevino95 27.87 22.32 0.3 alevino96 24.54 19.19 0.2 alevino97 22.08 17.48 0.2 alevino98 25.03 19.97 0.3 alevino99 25.16 20.21 0.2 alevino
100 25.40 20.88 0.3 alevino101 20.99 16.54 0.1 alevino102 17.67 14.87 0.1 alevino103 16.27 13.35 0.1 alevino104 19.26 15.40 0.1 alevino105 29.12 22.80 0.3 alevino

106
Anexo 8. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en Tibitoc.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 52.77 43.90 3.4 hembra2 52.43 43.33 3.4 hembra3 57.87 50.21 4.8 hembra4 47.38 40.18 2.4 hembra5 47.97 39.82 2.3 macho6 37.44 30.39 1.0 hembra7 33.01 26.62 0.7 hembra8 52.48 44.26 3.3 hembra9 35.79 28.99 0.8 hembra
10 52.95 44.03 2.8 hembra11 47.24 40.17 2.3 macho12 34.73 27.65 0.8 macho

107
Anexo 9. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo de los ejemplares capturados en el caño Los Pozos, lago de Tota.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 75.74 64.50 11.0 macho2 72.71 61.90 9.7 macho3 77.46 64.77 9.6 macho4 71.09 60.65 8.4 macho5 69.94 58.11 7.6 hembra6 69.19 58.77 7.4 macho7 71.47 60.08 7.1 macho8 66.13 56.39 6.3 macho9 71.09 60.10 7.2 macho
10 69.72 58.42 7.8 hembra11 65.50 54.62 6.0 macho12 66.03 55.11 6.1 hembra13 66.03 55.97 7.0 macho14 64.89 55.17 7.2 hembra15 66.26 56.26 6.9 hembra16 64.23 54.20 5.9 macho17 65.03 53.90 6.5 hembra18 67.99 57.79 7.6 macho19 64.17 53.94 5.3 macho20 66.91 57.12 6.7 macho21 70.66 60.44 8.5 macho22 69.37 59.40 8.1 macho23 35.56 55.85 7.3 hembra24 65.94 55.66 7.4 hembra25 72.33 62.55 8.9 macho26 71.02 60.37 9.2 macho27 63.30 53.98 6.4 hembra28 67.39 57.18 7.2 macho29 67.34 57.89 6.9 macho30 70.82 60.73 7.9 hembra31 60.06 50.48 5.2 hembra32 62.27 52.95 5.8 macho33 64.58 54.57 5.5 macho34 59.19 49.36 4.5 hembra35 56.92 48.40 3.9 hembra36 63.46 53.44 6.1 hembra37 61.51 51.65 5.3 hembra38 67.83 56.68 6.4 hembra39 62.32 52.62 6.2 hembra40 65.43 55.94 6.4 hembra
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 64.68 54.39 6.1 hembra42 62.08 52.04 5.1 hembra43 55.72 46.17 3.8 hembra44 56.37 47.78 3.7 macho45 64.73 54.15 5.4 macho46 63.68 53.63 5.9 macho47 65.12 54.68 5.2 hembra48 56.41 47.46 4.1 hembra49 60.17 50.51 4.4 macho50 60.84 52.26 4.9 macho51 67.37 55.97 8.5 macho52 63.64 53.64 5.3 macho53 59.40 50.24 4.8 hembra54 58.54 49.33 4.4 hembra55 56.26 47.65 3.6 hembra56 65.13 55.40 6.1 macho57 62.90 53.85 5.3 hembra58 60.80 51.66 4.4 hembra59 57.14 48.43 3.5 hembra60 64.06 53.75 6.0 hembra61 65.97 56.08 6.2 macho62 63.05 53.60 5.2 macho63 60.70 51.19 4.9 hembra64 50.21 42.04 2.4 hembra65 63.09 54.05 5.9 macho66 54.67 45.53 3.2 macho67 65.95 55.51 5.6 macho68 62.68 52.91 4.6 macho69 63.19 53.30 5.4 hembra70 60.57 50.59 4.8 hembra71 61.05 51.62 4.8 macho72 60.65 51.85 5.2 hembra73 57.11 47.94 3.7 hembra74 61.99 51.17 5.1 hembra75 54.84 45.81 3.5 hembra76 63.38 52.95 5.4 hembra77 61.13 51.80 4.9 hembra78 63.50 54.16 5.5 macho79 61.62 51.95 5.2 hembra80 55.23 45.89 3.4 hembra

108
Anexo 10. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el caño Los Pozos, lago de Tota
(continuación).
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
81 61.18 51.48 4.8 hembra82 62.13 52.01 4.5 macho83 56.25 48.13 3.9 hembra84 23.87 19.61 0.1 alevino85 21.55 17.30 0.1 alevino86 23.67 19.82 0.2 alevino87 56.45 46.81 3.7 -88 60.62 50.96 4.5 -89 59.29 50.93 4.3 -

109
Anexo 11. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el nacimiento del río Upía, lago de Tota.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 64.65 54.41 5.8 macho2 61.84 52.72 5.7 macho3 63.80 54.24 4.9 macho4 58.58 49.30 4.6 macho5 62.86 53.24 5.5 macho6 67.41 57.05 7.3 hembra7 65.63 56.00 6.5 hembra8 56.54 47.57 3.9 hembra9 65.27 56.33 6.1 macho10 63.95 54.25 5.8 macho11 62.26 52.42 4.4 macho12 62.30 52.83 5.4 macho13 67.98 57.94 6.9 macho14 61.40 51.43 5.8 macho15 54.03 45.49 3.4 hembra16 58.72 49.56 4.3 hembra17 61.95 52.23 4.4 macho18 63.91 53.80 5.9 macho19 60.94 51.62 5.0 macho20 58.88 50.17 4.4 macho21 60.44 51.21 4.6 hembra22 56.24 47.70 3.7 hembra23 57.63 48.91 4.3 macho24 59.42 49.51 4.5 macho25 56.18 46.50 4.0 hembra26 53.79 45.14 3.3 hembra27 54.70 46.44 3.7 hembra28 56.52 48.07 4.3 hembra29 56.20 47.15 4.0 hembra30 59.94 51.37 4.7 macho31 53.25 43.24 2.8 hembra32 20.75 17.60 0.2 alevino33 41.06 32.38 1.7 macho34 54.99 43.87 3.0 macho35 30.53 23.77 0.5 alevino36 19.27 15.45 0.2 alevino37 18.24 15.12 0.1 alevino38 17.90 14.77 0.1 alevino39 33.40 26.60 0.7 -

110
Anexo 12. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mana de Ramos, río
Lenguazaque.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 72.00 61.79 8.8 macho2 69.69 60.00 7.7 macho3 67.85 57.07 7.6 macho4 74.85 63.51 9.1 macho5 71.09 60.25 9.1 macho6 65.41 55.11 6.5 macho7 66.21 55.88 6.1 macho8 63.60 53.56 6.6 macho9 74.63 63.86 9.5 macho10 71.43 58.91 7.5 macho11 70.32 58.89 9.6 macho12 72.67 61.16 9.1 macho13 69.81 59.36 7.5 macho14 68.86 59.80 7.7 macho15 62.64 53.48 6.0 macho16 64.28 53.13 4.9 macho17 66.27 54.32 5.4 macho18 61.87 50.90 4.7 macho19 63.54 52.85 5.5 macho20 66.12 54.85 5.7 macho21 66.38 56.13 7.1 macho22 68.82 57.31 6.6 macho23 64.24 56.03 6.8 macho24 65.20 55.81 6.5 hembra25 64.55 54.18 5.6 hembra26 64.17 53.15 6.0 macho27 66.43 55.91 6.5 macho28 63.45 52.92 5.6 hembra29 66.50 56.46 6.8 hembra30 65.81 55.15 6.5 macho31 56.40 47.41 4.0 hembra32 54.93 46.38 3.7 hembra33 53.44 44.75 3.3 hembra34 53.89 44.80 2.8 hembra35 53.71 44.88 3.0 hembra36 55.44 46.16 3.4 hembra37 59.46 48.99 4.7 hembra38 53.12 43.12 2.9 hembra39 63.44 53.20 4.6 hembra40 56.33 46.74 3.4 hembra
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 47.28 39.76 2.1 hembra42 46.99 41.35 2.3 hembra43 50.53 41.37 2.4 hembra44 48.84 41.13 2.3 hembra45 50.98 41.62 2.2 hembra46 53.57 44.61 2.9 macho47 61.16 49.97 4.4 hembra48 53.97 44.41 3.1 hembra49 53.94 44.35 2.7 hembra50 59.90 48.98 3.9 hembra51 51.43 41.76 2.8 hembra52 47.42 37.87 1.9 hembra53 49.57 41.59 2.5 hembra54 49.83 39.89 2.1 hembra55 49.13 40.22 2.3 hembra56 50.45 42.70 2.3 hembra57 62.73 52.29 5.1 macho58 62.73 52.60 4.9 macho59 61.10 51.21 4.4 macho60 60.76 49.91 4.7 macho61 55.22 46.59 3.6 hembra62 53.39 44.46 2.5 hembra63 54.00 44.07 2.7 macho64 53.92 44.50 2.9 macho65 54.97 46.05 3.3 macho66 55.69 46.90 3.0 macho67 55.05 45.50 3.0 macho68 56.54 46.93 3.5 macho69 53.33 44.35 2.7 macho70 52.51 43.91 2.7 macho71 52.65 43.95 2.8 hembra72 42.18 35.18 1.3 hembra73 48.37 39.45 2.0 hembra74 59.16 49.19 4.3 macho75 56.94 53.23 3.5 macho76 52.85 43.52 2.7 hembra77 52.29 45.35 3.2 hembra78 49.04 40.63 2.3 hembra79 55.12 45.70 3.1 macho80 41.72 34.05 1.3 hembra

111
Anexo 13. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mana de Ramos, río
Lenguazaque (continuación).
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
81 43.06 35.40 1.5 hembra82 45.71 37.35 1.8 hembra83 43.24 36.07 1.6 hembra84 38.21 30.90 0.9 hembra85 37.13 29.87 0.9 hembra86 45.41 37.19 1.5 macho87 36.72 29.77 0.9 hembra88 45.26 35.70 1.8 macho89 41.30 32.79 1.8 hembra90 39.88 32.47 1.1 macho91 30.68 27.55 0.7 hembra92 35.79 29.39 0.8 hembra93 33.11 27.24 0.5 macho94 34.94 27.79 0.7 hembra95 30.09 26.28 0.6 macho96 36.70 29.95 0.8 hembra97 35.79 29.52 0.8 macho98 40.02 32.20 1.1 macho99 39.29 33.09 1.1 macho100 25.55 20.14 0.3 alevino101 27.64 22.11 0.6 alevino102 24.97 20.17 0.4 alevino103 23.90 19.35 0.3 alevino104 26.05 21.25 0.5 alevino105 23.97 19.15 0.4 alevino106 24.10 18.97 0.3 alevino107 25.32 20.85 0.3 alevino108 24.01 19.40 0.3 alevino109 22.72 18.45 0.3 alevino110 29.93 23.67 0.5 alevino111 26.31 21.81 0.5 alevino112 26.92 21.85 0.5 alevino113 24.00 18.31 0.4 alevino114 20.99 17.35 0.3 alevino115 23.24 18.49 0.5 alevino116 19.82 16.18 0.3 alevino117 18.71 14.55 0.2 alevino118 19.52 16.06 0.3 alevino119 22.34 17.85 0.3 alevino120 24.36 19.03 0.3 alevino
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
121 19.84 15.41 0.1 alevino122 18.19 14.56 0.1 alevino123 16.83 12.86 0.1 alevino124 17.90 14.21 0.1 alevino125 19.21 15.28 0.1 alevino126 16.42 13.37 0.1 alevino127 19.49 16.07 0.1 alevino

112
Anexo 14. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el vallado La Susana, río Bogotá.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 74.07 62.39 8.2 macho2 75.17 63.28 9.3 hembra3 74.27 61.93 8.7 macho4 68.22 57.47 6.4 hembra5 67.57 56.32 5.9 macho6 69.95 59.89 6.6 macho7 65.26 54.55 5.8 hembra8 63.47 52.82 4.5 macho9 65.73 55.30 6.1 macho
10 65.85 54.64 6.1 macho11 66.44 56.58 6.7 hembra12 68.15 57.19 6.8 hembra13 65.65 55.63 5.8 macho14 61.03 51.70 4.9 hembra15 63.02 52.34 5.0 macho16 63.42 54.08 5.1 macho17 66.86 57.38 6.1 macho18 62.59 51.53 5.2 macho19 62.87 52.01 5.3 macho20 62.71 52.22 4.9 macho21 66.53 55.70 6.1 hembra22 63.30 53.62 5.6 hembra23 63.17 53.09 5.1 hembra24 62.55 52.89 4.8 hembra25 65.55 54.93 6.0 hembra26 64.74 53.88 5.5 hembra27 63.13 53.07 4.8 macho28 62.93 53.69 5.1 macho29 63.46 52.92 5.3 hembra30 58.30 47.84 4.1 hembra31 55.99 47.09 3.8 hembra32 44.64 8.42 1.7 hembra33 63.73 52.50 4.9 hembra34 52.61 43.74 2.7 hembra35 48.76 40.00 1.9 hembra36 56.15 46.27 3.4 hembra37 56.98 46.82 3.6 hembra38 52.83 43.30 2.7 hembra39 61.40 52.87 4.7 hembra40 59.56 50.21 3.9 hembra
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
41 52.25 43.59 2.6 hembra42 52.24 44.15 2.8 hembra43 52.84 44.13 2.9 hembra44 45.83 37.95 1.8 hembra45 57.29 49.70 4.2 macho46 58.84 49.32 4.6 hembra47 50.81 42.48 2.2 hembra48 59.47 49.34 4.2 macho49 56.41 46.27 2.8 macho50 56.37 47.46 3.1 macho51 36.88 29.63 0.7 hembra52 39.92 33.77 1.0 macho53 36.48 29.25 0.8 macho54 41.68 33.86 1.2 macho55 32.10 24.01 0.5 macho56 39.50 32.24 1.1 macho57 39.30 31.72 1.1 hembra58 34.00 27.55 0.6 macho59 34.32 27.30 0.7 macho60 54.16 46.16 2.7 hembra61 42.90 35.33 1.3 hembra62 39.38 32.75 1.1 macho63 40.36 32.95 1.2 macho64 41.44 34.32 1.1 macho65 35.00 28.55 0.7 macho66 39.22 34.70 1 macho67 33.12 25.78 0.5 hembra68 34.72 27.11 0.6 macho69 32.96 26.25 0.6 macho70 34.20 27.36 0.6 macho71 32.48 26.89 0.5 hembra72 35.92 29.63 0.7 macho73 28.16 21.56 0.3 alevino74 37.70 28.65 0.8 macho75 30.48 25.84 0.4 hembra76 38.24 30.52 0.8 macho77 33.70 26.66 0.6 -78 36.40 29.59 0.7 -

113
Anexo 15. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en la quebrada Mancilla, río Bogotá.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 81.74 71.14 13.8 macho2 86.76 74.00 14.7 macho3 82.65 71.11 13.3 macho4 70.95 59.88 7.3 hembra5 68.88 58.39 7.1 hembra6 50.01 42.24 2.3 hembra7 76.11 68.83 10.5 hembra8 54.61 45.15 2.8 macho9 43.92 35.98 1.4 macho10 66.48 56.18 5.1 hembra11 42.01 34.40 1.2 hembra12 71.69 61.14 8.0 macho13 48.89 40.11 1.6 macho14 67.90 56.70 5.6 hembra15 74.08 63.21 8.5 hembra

114
Anexo 16. Longitud total (Lt), Longitud estándar (Ls), Peso total (Wt), y sexo
de los ejemplares capturados en el embalse de Tominé.
Ejemplar No.
Lt (mm) Ls (mm) Wt (g) Sexo
1 58.96 48.71 3.9 macho2 53.70 43.99 3.0 macho3 47.01 37.93 1.8 macho4 48.72 41.48 2.1 macho5 29.51 23.69 0.5 macho6 48.51 40.54 2.2 macho7 48.48 40.68 2.2 hembra8 54.63 46.16 2.9 macho9 55.46 45.98 2.7 macho
10 55.10 45.85 3.2 macho11 44.99 36.47 1.5 hembra12 23.49 18.76 0.3 alevino13 22.39 18.25 0.2 alevino14 33.00 16.50 0.6 macho15 26.16 20.90 0.3 hembra16 26.00 20.75 0.3 hembra17 26.98 21.88 0.4 hembra18 52.48 43.83 2.4 macho19 46.31 38.40 1.8 macho20 59.82 50.69 3.8 macho21 54.76 46.25 3.2 macho22 46.00 38.32 1.9 macho23 59.34 48.90 3.4 macho24 41.25 34.23 1.2 hembra25 35.82 29.50 0.8 macho26 25.24 20.75 0.3 alevino27 42.13 34.52 1.3 hembra28 50.52 41.45 2.2 hembra29 62.10 51.61 3.7 macho30 20.80 16.78 0.2 alevino

115
Anexo 17. Gráficas de la relación longitud – peso para la totalidad de
individuos capturados.
Todas Localidades
Wt = 6E-06*Lt3.2952
r2 = 0.9676n= 751
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
0 20 40 60 80 100LT (mm)
Todos Machos
Wt = 2E-06*Lt3.5313
r2 = 0.9781n= 338
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
0 20 40 60 80 100LT (mm)
Todas Hembras
Wt = 2E-06*Lt3.5558
r2 = 0.9592n= 284
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 20 40 60 80LT (mm)
Todos Alevinos
Wt = 7E-05*Lt2.5418
r2 = 0.6283n= 130
0.0
0 .1
0.2
0.3
0.4
0.5
0.6
0.7
0 10 20 30 40LT (mm)

116
Anexo 18. Gráficas de la relación longitud – peso en la laguna de Fúquene.
Fúquene - Todo
Wt = 0.00001*Lt3.110
r2 = 0.9702n= 101
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
0 20 40 60
LT (mm)
Fúquene - Machos
Wt = 0.00001*Lt3.114
r2 = 0.746n= 56
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
0 20 40 60
LT (mm)
Fúquene - Hembras
Wt = 0.000009*Lt3.142
r2 = 0.9382n= 15
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0 20 40 60
LT (mm)
Fúquene - Alevinos
Wt = 0.00009 * Lt2.383
r2 = 0.716n= 30
0 .0
0.1
0 .2
0 .3
0 .4
0.5
0 .6
0 10 20 30 40
LT (mm)

117
Anexo 19. Gráficas de la relación longitud – peso en el río Ubaté.
Ubaté - Todo
Wt = 2E-06*Lt3,5242
r2 = 0,9261n= 92
0 .0
1.0
2 .0
3 .0
4 .0
5.0
6 .0
7.0
0 20 40 60 80
LT (mm)
Ubaté - Machos
Wt = 2E-06*Lt3,5607
r2 = 0,9606n= 32
0 .0
1.0
2 .0
3 .0
4 .0
5.0
6 .0
0 20 40 60 80LT (mm)
Ubaté - Hembras
Wt = 2E-06*Lt3,5583
r2 = 0,899n= 59
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
0 20 40 60 80LT (mm)
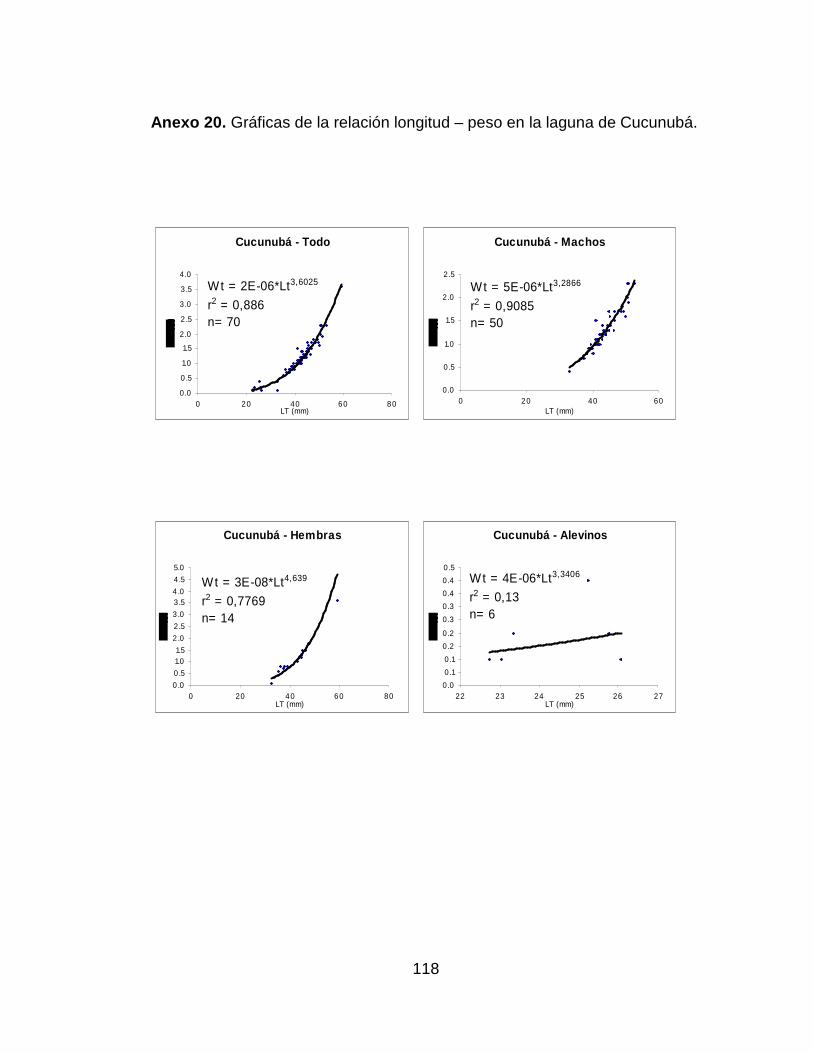
118
Anexo 20. Gráficas de la relación longitud – peso en la laguna de Cucunubá.
Cucunubá - Todo
Wt = 2E-06*Lt3,6025
r2 = 0,886n= 70
0.0
0 .5
1.0
1.5
2.0
2 .5
3.0
3 .5
4.0
0 20 40 60 80LT (mm)
Cucunubá - Machos
Wt = 5E-06*Lt3,2866
r2 = 0,9085n= 50
0.0
0.5
1.0
1.5
2.0
2.5
0 20 40 60LT (mm)
Cucunubá - Hembras
Wt = 3E-08*Lt4,639
r2 = 0,7769n= 14
0 .0
0.5
1.01.5
2 .0
2.5
3 .0
3.54 .0
4.5
5.0
0 20 40 60 80LT (mm)
Cucunubá - Alevinos
Wt = 4E-06*Lt3,3406
r2 = 0,13n= 6
0.0
0 .1
0 .1
0.2
0.2
0.3
0.3
0.4
0.4
0.5
22 23 24 25 26 27LT (mm)

119
Anexo 21. Gráficas de la relación longitud – peso en el embalse El Hato.
El Hato - Todos
Wt = 5E-06*Lt3,2951
r2 = 0,9106n= 105
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
0 20 40 60 80LT (mm)
El Hato - Machos
Wt = 1E-06*Lt3,74
r2 = 0,9689n= 25
0 .0
0.5
1.0
1.5
2 .0
2.5
0 20 40 60LT (mm)
El Hato - Hembras
Wt = 6E-06*Lt3,2376
r2 = 0,9487n= 28
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
0 20 40 60 80
LT (mm)
El Hato - Alevinos
Wt = 4E-05*Lt2,6308
r2 = 0,668n= 52
0
0.1
0 .2
0 .3
0 .4
0.5
0 .6
0 10 20 30 40
LT (mm)

120
Anexo 22. Gráficas de la relación longitud – peso en Tibitoc.
Tibitó - Todo
Wt = 4E-06*Lt3,4369
r2 = 0,9893n= 12
0.0
1.0
2.0
3.0
4.0
5.0
6.0
0 20 40 60 80LT (mm)
Tibitó - Machos
Wt = 6E-06*Lt3,3434
r2 = 0,9982n= 3
0 .0
0.5
1.0
1.5
2 .0
2.5
0 20 40 60LT (mm)
Tibitó - Hembras
Wt = 4E-06*Lt3,4562
r2 = 0,9875n= 9
0.0
1.0
2.0
3.0
4.0
5.0
6.0
0 20 40 60 80LT (mm)

121
Anexo 23. Gráficas de la relación longitud – peso en lago de Tota, quebrada
Los Pozos.
Tota (Pozos) - Todos
Wt = 5E-06*Lt3,3521
r2 = 0,8755n= 85
0 .0
2 .0
4 .0
6 .0
8 .0
10 .0
12 .0
0 20 40 60 80 100LT (mm)
Tota (Pozos) - Machos
Wt = 2E-06*Lt3,5736
r2 = 0,9036n= 39
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 20 40 60 80 100LT (mm)
Tota (Pozos) - Hembras
Wt = 3E-06*Lt3,4529
r2 = 0,9227n= 43
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
0 20 40 60 80LT (mm)
Tota (Pozos) - Alevinos
Wt = 8E-06*Lt3,063
r2 = 0,1887n= 3
0
0.05
0.1
0.15
0.2
0.25
21 22 23 24LT (mm)

122
Anexo 24. Gráficas de la relación longitud – peso en lago de Tota,
nacimiento del río Upía.
Tota (Upía) - Todos
Wt = 2E-05*Lt3,0653
r2 = 0,9914n= 38
0 .0
1.0
2 .0
3 .0
4 .0
5.0
6 .0
7.0
8 .0
0 20 40 60 80LT (mm)
Tota (Upía) - Machos
Wt = 5E-05*Lt2,8081
r2 = 0,9099n= 20
0 .0
1.0
2 .0
3 .0
4 .0
5.0
6 .0
7.0
8 .0
0 20 40 60 80LT (mm)
Tota (Upía) - Hembras
Wt = 3E-06*Lt3,4602
r2 = 0,9501n= 13
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
0 20 40 60 80LT (mm)
Tota (Upía) - Alevinos
Wt = 3E-05*Lt2,8473
r2 = 0,8899n= 5
0.0
0 .1
0.2
0.3
0.4
0.5
0.6
0 10 20 30 40LT (mm)

123
Anexo 25. Gráficas de la relación longitud – peso en la quebrada Mana de
Ramos, río Lenguazaque.
Lenguazaque - Todos
Wt = 3E-05*Lt2,9357
r2 = 0,9758n= 127
0 .0
2 .0
4 .0
6 .0
8 .0
10 .0
12 .0
0 20 40 60 80LT (mm)
Lenguazaque - Machos
Wt = 4E-06*Lt3.4255
r2 = 0.9797n= 50
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 20 40 60 80
LT (mm)
Lenguazaque - Hembras
Wt = 9E-06*Lt3,1884
r2 = 0,9654n= 49
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
0 20 40 60 80LT (mm)
Lenguazaque - Alevinos
Wt = 9E-06*Lt3,3182
r2 = 0,7501n= 28
0 .0
0.1
0 .2
0 .3
0 .4
0.5
0 .6
0.7
0 .8
0 10 20 30 40
LT (mm)
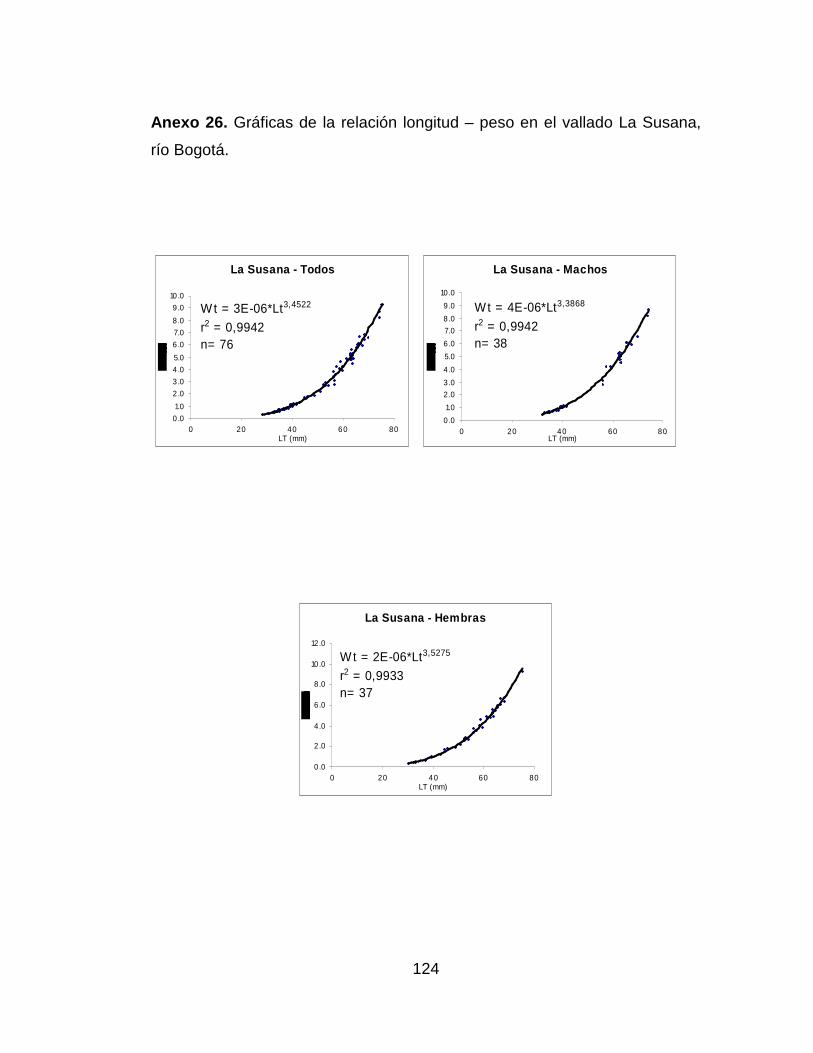
124
Anexo 26. Gráficas de la relación longitud – peso en el vallado La Susana,
río Bogotá.
La Susana - Todos
Wt = 3E-06*Lt3,4522
r2 = 0,9942n= 76
0 .0
1.0
2 .0
3 .0
4 .0
5.0
6 .0
7.0
8 .0
9 .0
10 .0
0 20 40 60 80LT (mm)
La Susana - Machos
Wt = 4E-06*Lt3,3868
r2 = 0,9942n= 38
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 20 40 60 80LT (mm)
La Susana - Hembras
Wt = 2E-06*Lt3,5275
r2 = 0,9933n= 37
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 20 40 60 80LT (mm)

125
Anexo 27. Gráficas de la relación longitud – peso en la quebrada Mancilla,
río Bogotá.
Mancilla - Todos
Wt = 1E-06*Lt3,6227
r2 = 0,9886n= 15
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
0 20 40 60 80 100LT (mm)
Mancilla - Machos
Wt = 9E-07*Lt3,7351
r2 = 0,9924n= 7
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
18.0
0 20 40 60 80 100LT (mm)
Mancilla - Hembras
Wt = 3E-06*Lt3,4545
r2 = 0,9857n= 8
0.0
2.0
4.0
6.0
8.0
10.0
12.0
0 20 40 60 80LT (mm)

126
Anexo 28. Gráficas de la relación longitud – peso en el embalse de Tominé.
Tominé - Todo
Wt = 3E-05*Lt2,8895
r2 = 0,9926n= 30
0 .0
0.5
1.0
1.5
2 .0
2.5
3 .0
3.5
4 .0
4.5
0 20 40 60 80LT (mm)
Tominé - Machos
Wt = 2E-05*Lt2,9315
r2 = 0,9853n= 18
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
0 20 40 60 80LT (mm)
Tominé - Hembras
Wt = 2E-05*Lt2,9645
r2 = 0,9925n= 8
0.0
0.5
1.0
1.5
2.0
2.5
0 20 40 60LT (mm)
Tominé - Alevinos
Wt = 0,0001*Lt2,4637
r2 = 0,7335n= 4
0.0
0 .1
0 .1
0.2
0.2
0.3
0.3
0.4
0 10 20 30LT (mm)

127
Anexo 29. Volumen (mm³) de cada ítem alimentario por localidad.
Ítem Fúquene Ubaté Cucunubá El Hato Tibitoc Tota-Pozos Tota-Up ía Mana de Ramos La Susana Mancilla TominéCHR 0.00 0.00 0.00 0.00 0.00 0.00 0.00 392.00 0.00 0.00 0.00ELM 0.00 0.00 0.00 0.00 2.00 0.00 0.00 0.00 0.00 0.00 0.00CER 0.00 0.00 0.00 0.00 0.00 2.00 0.00 0.00 0.00 4.00 0.00
CHI-L 1.00 18.00 4.00 63.00 4.00 10.00 1.00 4.00 3.00 34.00 6.00CHI-P 0.00 208.00 0.00 0.00 0.00 1.50 0.50 0.00 0.00 0.00 0.00CUL 0.00 3.00 0.00 0.50 4.00 0.00 0.00 0.50 0.00 0.00 0.00SIM 0.00 0.00 0.00 0.00 0.00 0.00 0.00 6.00 0.00 0.00 0.00RDI 0.00 5.00 0.00 2.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00COR 0.00 0.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.00 0.00NOT 1.00 0.00 0.00 0.00 0.00 4.00 0.00 12.00 0.00 0.00 0.00HNN 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.50 0.00VES 0.00 0.00 5.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00AES 0.00 0.00 20.00 0.00 0.00 0.00 0.00 0.00 0.00 21.00 0.00COE 0.00 0.00 14.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00RZY 0.00 0.00 0.00 24.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00LEP 0.00 0.00 6.00 0.00 0.00 0.00 0.00 8.00 0.00 0.00 0.00TRI 0.50 4.00 0.00 1.00 2.00 2.00 1.00 0.00 30.00 0.00 2.00RIN 4.00 195.00 48.00 8.00 3.00 10.00 6.00 12.00 15.00 20.00 4.00ACA 0.00 0.00 0.25 0.00 0.00 0.00 0.50 0.25 0.25 1.00 0.25HAY 1.50 1.00 0.00 0.00 8.00 20.00 5.00 1.00 2.00 16.00 5.00BOS 0.25 0.00 12.00 25.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00CHY 0.50 0.50 0.00 0.00 1.00 1.00 0.50 0.50 1.00 0.25 4.00EUR 0.00 0.00 0.00 0.00 0.00 0.00 0.50 0.00 0.00 0.00 9.00DAP 3.00 0.00 1.00 0.50 36.00 0.00 0.25 1.00 0.25 0.00 6.00CYC 0.50 4.00 0.50 3.00 18.00 3.00 0.25 1.00 10.00 1.00 2.00OST 0.50 0.00 2.00 2.00 1.00 0.00 0.75 0.00 0.50 0.00 1.00RCR 1.00 6.00 21.00 3.00 28.00 3.00 1.00 4.00 15.00 0.00 30.00PHY 0.00 4.00 0.00 6.00 0.00 0.00 2.50 0.00 0.00 0.00 1.00BIV 0.00 0.00 0.00 0.00 0.00 6.00 3.00 0.00 0.00 0.00 0.00HUE 0.00 2.00 0.00 0.00 0.00 0.00 1.00 0.00 0.00 0.00 0.00NEM 0.25 0.00 0.00 0.00 0.00 0.00 0.00 0.50 0.00 1.00 0.00SEM 0.00 0.00 0.00 0.00 0.00 0.00 2.00 0.00 0.00 4.00 0.00RPL 3.00 0.00 2.00 3.00 2.00 2.50 2.00 45.00 3.00 10.00 5.00DAT 0.00 0.00 0.00 0.00 0.00 12.00 0.00 0.00 2.00 84.00 0.00ESC 1.00 0.00 0.00 0.00 0.00 0.00 0.50 0.00 0.00 0.00 0.00

128
Anexo 30. Frecuencia de ocurrencia de cada ítem alimentario para las 11
localidades de estudio.
ÍtemFrecuencia Ocurrencia
ÍtemFrecuencia Ocurrencia
CHI-L 1.000 RDI 0.182CYC 1.000 AES 0.182RPL 0.909 LEP 0.182HAY 0.818 EUR 0.182CHY 0.818 BIV 0.182TRI 0.727 HUE 0.182DAP 0.727 SEM 0.182OST 0.636 ESC 0.182ACA 0.545 CHR 0.091CUL 0.364 ELM 0.091PHY 0.364 SIM 0.091
CHI-P 0.273 COR 0.091NOT 0.273 HNN 0.091BOS 0.273 VES 0.091NEM 0.273 COE 0.091DET 0.273 RZY 0.091CER 0.182
Top Related