







Idiomas
Páginas
Jurídico


COMPARACIÓN ESTACIONAL DE LA COMUNIDAD DE MACROINVERTEBRADOS
EPIBENTÓNICOS ASOCIADOS A PRADERAS DE Thalassia testudinum EN LA
GUAJIRA, CARIBE COLOMBIANO
ADRIANA AGUIRRE AGUIRRE
TRABAJO DE GRADO Presentado como requisito parcial
Para optar por el titulo de
Bióloga
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS CARRERA DE BIOLOGIA
BOGOTA, D.C AGOSTO, 2006
PDF created with pdfFactory trial version www.pdffactory.com

NOTA DE ADVERTENCIA
Articulo 23 de la Resolución Nº 13 DE Julio de 1946
“La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en
sus trabajos de tesis. Solo velará por que no se publique nada contrario al dogma y a la
moral católica y por que las tesis no contengan ataques personales contra persona
alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia”
PDF created with pdfFactory trial version www.pdffactory.com

Bogotá, Septiembre 25 de 2006 Señores Pontificia Universidad Javeriana Ciudad Estimados Señores: Yo, Adriana Aguirre Aguirre identificada con C.C. No. 52.865.380, autora del trabajo de grado titulado “Comparación estacional de la comunidad de macroinvertebrados epibentónicos asociados a praderas de Thalassia testudinum en La Guajira, Caribe colombiano”, presentado como requisito para optar por el titulo de Bióloga en el año 2006; autorizo a la Universidad a: a) Reproducir el trabajo en medio digital o electrónico con el fin de ofrecerlo para la consulta en la Biblioteca General b) Poner a disposición para la consulta con fines académicos, en la pagina web de la facultad, de la Biblioteca General y en redes de información con las cuales tenga convenio la Universidad Javeriana c) Enviar el trabajo en formato impreso o digital, en caso de que sea seleccionado para participar en concursos de trabajos de grado d) Distribuir ejemplares de la obra, para la consulta entre entidades educativas con las que la facultad tenga convenio de intercambio de información, para que este sea consultado en las bibliotecas y centros de documentación de las respectivas entidades e) Todos los usos que tengan finalidad académica Los derechos morales sobre el trabajo son de los autores de conformidad con lo establecido en el artículo 30 de la Ley 23 de 1982 y el artículo 11 de la Decisión Andina 351 de 1993, los cuales son irrenunciables, imprescriptibles, inembargables e inalienables. Atendiendo lo anterior, siempre que se consulte esta obra, mediante cita bibliográfica se debe dar crédito del trabajo y a su autor. Este documento se firma, sin perjuicio de los acuerdos que el autor pacte con la Unidad Académica referentes al uso de la obra o a los derechos de propiedad industrial que pueda surgir de la actividad académica. Adriana Aguirre Aguirre C.C. 52.865.380
PDF created with pdfFactory trial version www.pdffactory.com

COMPARACIÓN ESTACIONAL DE LA COMUNIDAD DE MACROINVERTEBRADOS
EPIBENTÓNICOS ASOCIADOS A PRADERAS DE Thalassia testudinum EN LA
GUAJIRA, CARIBE COLOMBIANO
ADRIANA AGUIRRE AGUIRRE
APROBADO
___________________ __________________ Guillermo Duque Nivia Alberto Acosta Moreno
Biólogo, M. Sc. Ph. D. Biólogo, M. Sc. Ph. D.
Director Codirector
__________________ ___________________ Giovanni Fagua Gonzáles Néstor Ardila Espitia
Biólogo, M. Sc. Biólogo Marino, M. Sc.
Jurado Jurado
PDF created with pdfFactory trial version www.pdffactory.com

COMPARACIÓN ESTACIONAL DE LA COMUNIDAD DE MACROINVERTEBRADOS
EPIBENTÓNICOS ASOCIADOS A PRADERAS DE Thalassia testudinum EN LA
GUAJIRA, CARIBE COLOMBIANO
ADRIANA AGUIRRE AGUIRRE
APROBADO
___________________ __________________ Decano Académico Andrea Forero
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
Autor Apellidos Nombre
Aguirre Aguirre Adriana Director
Apellidos Nombre Duque Nivia Guillermo
Codirector
Apellidos Nombre Acosta Moreno Alberto
TRABAJO PARA OPTAR POR EL TITULO DE: Bióloga TITULO COMPLETO DEL TRABAJO: “Comparación estacional de la comunidad de macroinvertebrados epibentónicos asociados a praderas de Thalassia testudinum en La Guajira, Caribe colombiano” SUBTITULO DEL TRABAJO: NO FACULTAD: Ciencias PROGRAMA: Carrera NOMBRE DEL PROGRAMA: Biología CIUDAD: BOGOTA AÑO DE PRESENTACION DEL TRABAJO: 2006 NUMERO DE PAGINAS: 53 TIPO DE ILUSTRACIONES: Mapas, Fotografías, Tablas, Figuras. MATERIAL ANEXO: NO DESCRIPTORES O PALABRAS CLAVE: Macroinvertebrados epibentónicos, Composición, Densidad, Variación estacional, Guajira, Colombia. RESUMEN DEL CONTENIDO: Diferentes estudios, principalmente en las zonas templadas y subtropicales han
comprobado la variación estacional en la estructura de las praderas de pastos
marinos y la estructura de la fauna asociada, sin embargo la información acerca
de los cambios estacionales de la fauna en el trópico y especialmente en el
Caribe colombiano es escasa. Con el fin de identificar posibles variaciones, se
estudió la fauna de macroinvertebrados epibentónicos asociados a praderas de
Thalassia testudinum en La Guajira durante dos épocas climáticas: seca menor y
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
lluvias (2005). Se escogieron quince estaciones, en praderas monoespecíficas,
con un área no inferior a 600 m² y con una cobertura superior al 40%, en donde
mediante censos visuales en transectos lineales (60 m²) se cuantificó la
abundancia y se determinó la composición de las comunidades de
macroinvertebrados epibentónicos. Análisis de diversidad, de clasificación
(Jaccard y Bray-Curtis) y de varianza fueron efectuados para comparar
composición y densidad en el tiempo. Se identificaron 110 especies (con un
nuevo registro para el Caribe colombiano), 42 fueron comunes para las dos
épocas, mientras que 32 fueron exclusivas de la época seca menor y 36 de la
época de lluvias. Los moluscos fueron el grupo dominante con 30.9% del total de
especies. La densidad promedio del total de individuos de todas las especies y la
Equitatividad de Pielou (J´) fueron significativamente diferentes entre épocas
climáticas mientras que la diversidad de Shannon-Weiner (H i ) y la Riqueza de
Margalef no mostraron diferencias significativas entre épocas. La densidad
promedio de las especies dominantes, Lytechinus variegatus y Arca zebra fue
significativamente diferente entre épocas climáticas, siendo mayor en la época
de lluvias, mientras que la densidad promedio de Clypeaster rosaceus y Vasum
muricatum no mostró diferencias significativas entre épocas. Los análisis de
similitud de Bray-Curtis y Jaccard indicaron que las estaciones son muy disímiles
entre si en cuanto a abundancia y composición, por lo que la variación en la
estructura de los macroinvertebrados epibentónicos asociados a praderas de
Thalassia testudinum en esta zona del Caribe colombiano, durante la época
seca menor y lluvias puede ser en parte explicada por las diferencias a nivel
espacial y temporal.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
A mi familia y en especial a mis padres que me apoyaron y guiaron desde el momento que decidí estudiar esta carrera. Gracias por darme todo y más, a
ellos les dedico el inicio de esta nueva etapa.
A Felipe B. que ha sido parte esencial de este proceso y que me ha
acompañado incondicionalmente, gracias por dejarme hacer parte
de ese sueño.
“Hay grandeza en esta concepción de la vida….que mientras este planeta ha ido
girando según la constante ley de la gravitación, se han desarrollado y se están desarrollando, a partir de un comienzo tan sencillo, infinidad de formas cada vez
mas bellas y maravillosas”
Charles Darwin
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
VI
AGRADECIMIENTOS
Agradezco muy especialmente al Instituto de Investigaciones Marinas y Costeras “José
Benito Vives de Andréis” INVEMAR y a la Corporación Autónoma Regional de La Guajira
CORPOGUAJIRA por el apoyo logístico y económico. Al grupo de pastos marinos (Diana
Isabel Gómez, Nelson Manrique y Paola Garzón) por su colaboración en campo. A las
personas que nos colaboraron durante nuestra estadía en La Guajira y a todos los que
nos recibieron en los lugares más alejados del Departamento, en especial a nuestro guía
y conductor “Tobi” por estar pendiente de nosotros y por acompañarnos en nuestros
recorridos. A Sven Zea, Néstor Campos, Norella Cruz, Adriana Gracia, Javier Reyes y
Juan Armando Sanchéz por su ayuda en la identificación y confirmación del material
colectado. A Fabián Cortés y nuevamente a Sven Zea por su colaboración y por guiarme
en el análisis de los datos. A Juan Carlos Diez que siempre estuvo dispuesto a
colaborarme en la búsqueda de bibliografía, al personal de sistemas especialmente Axel
y Nieelsen gracias por ayudarme siempre con los problemas del computador. Agradezco
infinitamente a Nelson Manrique porque sin su ayuda jamás hubiera terminado este
trabajo, gracias por los buenos momentos, por enseñarme tantas cosas y por ser un
excelente amigo y compañero de buceo. Al SIMAC (Beto, Cata, Raúl, Sonia, Nel y
Martica) por abrirme las puertas en el Instituto y enseñarme tantas cosas, especialmente
a Raúl Navas por sus consejos y su cariño durante el tiempo que estuve en Santa Marta.
A mi director Guillermo Duque por su tiempo, apoyo, enseñanzas y por estar siempre
pendiente de mi desempeño. A Diana Isabel Gómez por estar al tanto de mi trabajo, por
su ayuda en campo, comentarios y correcciones y a mi codirector Alberto Acosta que
aún estando lejos, estuvo pendiente de mi trabajo; gracias por las correcciones y por el
tiempo dedicado.
A mis padres que son el mejor ejemplo de dedicación les agradezco por darme las bases
necesarias para empezar esta etapa de mi vida como persona y como profesional y a
mis hermanos que siempre han estado a mi lado y que también han sido parte de mi
formación. A Felipe Ballesteros que estuvo siempre apoyándome y dándome ánimos
para hacer las cosas lo mejor posible, gracias por darle sentido a todo, por la compañía y
los buenos momentos que viví en Santa Marta.
Finalmente quiero agradecer a todos los que fueron como mi familia durante este año,
solo puedo decirles que me llevo recuerdos muy buenos del tiempo que compartimos en
Santa Marta, gracias a todos por su amistad y por hacer mas agradable mi estadía.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
VII
TABLA DE CONTENIDO
1. INTRODUCCIÓN........................................................................................................ 1
2. MARCO TEÓRICO..................................................................................................... 3
2.1 ÁREA DE ESTUDIO ................................................................................................... 3 2.2 GENERALIDADES ..................................................................................................... 4 2.3 DISTRIBUCIÓN ......................................................................................................... 5 2.4 FAUNA ASOCIADA.................................................................................................... 7
3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN........................................... 12
3.1 FORMULACIÓN DEL PROBLEMA ............................................................................... 12 3.2 PREGUNTA DE INVESTIGACIÓN ................................................................................ 12 3.3 JUSTIFICACIÓN DE LA INVESTIGACIÓN ...................................................................... 12
4. HIPÓTESIS .............................................................................................................. 14
5. OBJETIVOS............................................................................................................. 15
5.1 OBJETIVO GENERAL .............................................................................................. 15 5.2 OBJETIVOS ESPECÍFICOS ....................................................................................... 15
6. MATERIALES Y MÉTODOS .................................................................................... 16
6.1 FASE DE CAMPO ................................................................................................... 17 6.2 FASE DE LABORATORIO ......................................................................................... 18 6.3 ANÁLISIS DE INFORMACIÓN..................................................................................... 18
7. RESULTADOS......................................................................................................... 21
7.1 ESTRUCTURA DE LA MACROFAUNA ASOCIADA .......................................................... 21 7.2 NUEVO REGISTRO PARA EL CARIBE COLOMBIANO. .................................................... 25 7.3 ANÁLISIS DE COMUNIDADES.................................................................................... 26 7.4 ESPECIES DOMINANTES.......................................................................................... 29
8. DISCUSIÓN ............................................................................................................. 31
8.1 ANÁLISIS DE COMUNIDADES.................................................................................... 32 8.2 ESPECIES DOMINANTES.......................................................................................... 34
9. CONCLUSIONES..................................................................................................... 38
10. RECOMENDACIONES........................................................................................... 39
11. REFERENCIAS...................................................................................................... 40
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
VIII
LISTA DE TABLAS
TABLA 1. FAMILIAS Y GÉNEROS DE PASTOS MARINOS. MODIFICADO DE SHORT Y COLES (2001).
............................................................................................................................... 5 TABLA 2. COMPARACIÓN GENERAL DE LAS PRADERAS DE PASTOS MARINOS EN SISTEMAS FRÍOS
/ TEMPLADOS, TEMPLADOS Y SUBTROPICALES Y TROPICALES (HECK Y ORTH, 1980;
VIRNSTEIN ET AL., 1984)............................................................................................ 9 TABLA 3. FRECUENCIA DE OCURRENCIA (F%), ABUNDANCIA (N), Y PROMEDIO DE LA DENSIDAD
(NUMERO DE INDIVIDUOS/M²) DE LAS ESPECIES ENCONTRADAS DURANTE: A) ÉPOCA SECA
MENOR Y B) ÉPOCA DE LLUVIAS................................................................................. 22 TABLA 4. VALORES DE DENSIDAD PROMEDIO (NÚMERO DE INDIVIDUOS/M²) Y DE LOS ÍNDICES DE
DIVERSIDAD DE SHANNON-WEINER (H i EQUITATIVIDAD DE PIELOU (J´) Y RIQUEZA DE
MARGALEF DURANTE LA ÉPOCA SECA MENOR (S) Y ÉPOCA DE LLUVIAS (LL) Y VALORES
PROMEDIO CON SU RESPECTIVO ERROR ESTÁNDAR (EE) EN CADA ESTACIÓN MUESTREADA.
............................................................................................................................. 27 TABLA 5. COMPARACIÓN DEL NÚMERO DE ESPECIES (S), NÚMERO DE INDIVIDUOS (N) Y
DENSIDAD DE INDIVIDUOS/M² (D) OBTENIDOS EN DIFERENTES ESTUDIOS DE FAUNA
ASOCIADA A PRADERAS REALIZADOS EN EL CARIBE COLOMBIANO Y EN OTRAS ZONAS DEL
CARIBE. SE MUESTRA TAMBIÉN LA FAUNA ASOCIADA ESTUDIADA (E: EPIFAUNA. I:
INFAUNA) Y EL MÉTODO DE MUESTREO. PARA EL PRESENTE ESTUDIO SE MUESTRAN LOS
DIFERENTES VALORES OBTENIDOS DURANTE LA ÉPOCA SECA MENOR (T) Y ÉPOCA DE
LLUVIAS (LL). ......................................................................................................... 31
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
IX
LISTA DE FIGURAS
FIGURA 1. DISTRIBUCIÓN DE LAS PRADERAS DE PASTOS MARINOS EN EL CARIBE COLOMBIANO.
TOMADO DE DÍAZ ET AL., 2003. .................................................................................. 4 FIGURA 2. MAPA DEL ÁREA DE ESTUDIO MOSTRANDO LA UBICACIÓN DE LAS ESTACIONES EN LA
ZONA COSTERA DEL DEPARTAMENTO DE LA GUAJIRA. ALTA GUAJIRA: PC (PUNTA
COCOS), PP (PUERTO PORTETE), W (WARREO) Y MEDIA GUAJIRA: CV1 (CABO DE LA
VELA I), CV2 (CABO DE LA VELA II), EC (EL CARDÓN), FT (TORIMA), SR2 (SANTA ROSA
II), SR1 (SANTA ROSA I), M1 (MANAURE I), M2 (MANAURE II), MU (MUSICHE), T
(TAWAYA), B (BALLENAS) Y MY (MAYAPO). FUENTE: LABORATORIO DE SISTEMA DE
INFORMACIÓN GEOGRÁFICA (SIG) INVEMAR. .......................................................... 16 FIGURA 3. REPRESENTACIÓN ESQUEMÁTICA DEL PORCENTAJE DE COBERTURA SEGÚN LA
ESCALA DE DAHL (1981). MODIFICADO DE ENGLISH ET AL. (1997)............................... 17 FIGURA 4. COMPARACIÓN DE LA RIQUEZA TOTAL DE ESPECIES PARA CADA GRUPO TAXONÓMICO
ENCONTRADO DURANTE LA ÉPOCA SECA MENOR Y ÉPOCA DE LLUVIAS .......................... 21 FIGURA 5. IMAGEN DE LA ESPECIE PILUMNUS PANNOSUS, NUEVO REGISTRO PARA EL CARIBE
COLOMBIANO ENCONTRADA EN PRADERAS DE T. TESTUDINUM DE LA GUAJIRA. ............. 26 FIGURA 6. CURVAS DE K- DOMINANCIA PARA LA ÉPOCA DE SECA MENOR (S) Y ÉPOCA DE
LLUVIAS (LL). ......................................................................................................... 27 FIGURA 7. REPRESENTACIÓN ESQUEMÁTICA DEL ÍNDICE DE SIMILITUD DE BRAY-CURTIS ENTRE
LAS ESTACIONES DE MUESTREO PC (PUNTA COCOS), PP (PUERTO PORTETE), W
(WARREO), CV1 (CABO DE LA VELA I), CV2 (CABO DE LA VELA II), EC (EL CARDÓN), FT
(TORIMA), SR2 (SANTA ROSA II), SR1 (SANTA ROSA I), M1 (MANAURE I), M2 (MANAURE
II), MU (MUSICHE), T (TAWAYA), B (BALLENAS) Y MY (MAYAPO), UTILIZANDO LOS
VALORES DE LAS ABUNDANCIAS TRANSFORMADAS CON LN (X + 1) PARA LA ÉPOCA SECA
MENOR (S) Y ÉPOCA DE LLUVIAS (LL). ....................................................................... 28 FIGURA 8. REPRESENTACIÓN ESQUEMÁTICA DEL COEFICIENTE DE SIMILITUD DE JACCARD (J´)
ENTRE LAS ESTACIONES DE MUESTREO: PC (PUNTA COCOS), PP (PUERTO PORTETE), W
(WARREO), CV1 (CABO DE LA VELA I), CV2 (CABO DE LA VELA II), EC (EL CARDÓN), FT
(TORIMA), SR2 (SANTA ROSA II), SR1 (SANTA ROSA I), M1 (MANAURE I), M2 (MANAURE
II), MU (MUSICHE), T (TAWAYA), B (BALLENAS) Y MY (MAYAPO) EN LA ÉPOCA SECA
MENOR (S) Y ÉPOCA DE LLUVIAS (LL). LOS VALORES DE SIMILITUD VAN DE 0 A 100, EN
DONDE 0 % REPRESENTA LA MENOR SIMILITUD Y 100% LA MAYOR SIMILITUD. ............... 29 FIGURA 9. DENSIDAD MEDIA (±EE) DE LAS ESPECIES DOMINANTES REGISTRADAS DURANTE LA
ÉPOCA SECA MENOR Y LA ÉPOCA DE LLUVIAS EN EL ÁREA DE ESTUDIO. ......................... 30
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
X
LISTA DE ANEXOS
ANEXO A. UBICACIÓN GEOGRÁFICA Y CARACTERÍSTICAS DE LAS QUINCE ESTACIONES
MUESTREADAS DURANTE LA ÉPOCA SECA MENOR (T) Y/O ÉPOCA DE LLUVIAS (H) EN LA
GUAJIRA. PC (PUNTA COCOS), PP (PUERTO PORTETE), W (WARREO) CV1 (CABO DE LA
VELA I), CV2 (CABO DE LA VELA II), EC (EL CARDÓN), FT (TORIMA), SR2 (SANTA ROSA
II), SR1 (SANTA ROSA I), M1 (MANAURE I), M2 (MANAURE II), MU (MUSICHE), T
(TAWAYA), B (BALLENAS) Y MY (MAYAPO). LA PROFUNDIDAD ESTÁ DADA EN METROS, LA
SALINIDAD EN UPS (UNIDADES PRACTICAS DE SALINIDAD) Y LA TEMPERATURA EN C°
(GRADOS CENTÍGRADOS). ........................................................................................ 49 ANEXO B. CUADRO SISTEMÁTICO DE LAS ESPECIES ENCONTRADAS DURANTE LAS ÉPOCAS (E)
SECA MENOR (T) Y/O DE LLUVIAS (LL) EN LAS PRADERAS DE T. TESTUDINUM DEL
DEPARTAMENTO DE LA GUAJIRA. INFORMACIÓN OBTENIDA DE ITIS (2006). ................. 50
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
XI
RESUMEN
Diferentes estudios, principalmente en las zonas templadas y subtropicales han
comprobado la variación estacional en la estructura de las praderas de pastos marinos y
la estructura de la fauna asociada, sin embargo la información acerca de los cambios
estacionales de la fauna en el trópico y especialmente en el Caribe colombiano es
escasa. Con el fin de identificar posibles variaciones, se estudió la fauna de
macroinvertebrados epibentónicos asociados a praderas de Thalassia testudinum en La
Guajira durante dos épocas climáticas: seca menor y lluvias (2005). Se escogieron
quince estaciones, en praderas monoespecíficas, con un área no inferior a 600 m² y con
una cobertura superior al 40%, en donde mediante censos visuales en transectos
lineales (60 m²) se cuantificó la abundancia y se determinó la composición de las
comunidades de macroinvertebrados epibentónicos. Análisis de diversidad, de
clasificación (Jaccard y Bray-Curtis) y de varianza fueron efectuados para comparar
composición y densidad en el tiempo. Se identificaron 110 especies (con un nuevo
registro para el Caribe colombiano), 42 fueron comunes para las dos épocas, mientras
que 32 fueron exclusivas de la época seca menor y 36 de la época de lluvias. Los
moluscos fueron el grupo dominante con 30.9% del total de especies. La densidad
promedio del total de individuos de todas las especies y la Equitatividad de Pielou (J´)
fueron significativamente diferentes entre épocas climáticas mientras que la diversidad
de Shannon-Weiner (H i ) y la Riqueza de Margalef no mostraron diferencias
significativas entre épocas. La densidad promedio de las especies dominantes,
Lytechinus variegatus y Arca zebra fue significativamente diferente entre épocas
climáticas, siendo mayor en la época de lluvias, mientras que la densidad promedio de
Clypeaster rosaceus y Vasum muricatum no mostró diferencias significativas entre
épocas. Los análisis de similitud de Bray-Curtis y Jaccard indicaron que las estaciones
son muy disímiles entre si en cuanto a abundancia y composición, por lo que la variación
en la estructura de los macroinvertebrados epibentónicos asociados a praderas de
Thalassia testudinum en esta zona del Caribe colombiano, durante la época seca menor
y lluvias puede ser en parte explicada por las diferencias a nivel espacial y temporal.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
XII
ABSTRACT
Different studies, especially in the temperate and subtropical zone have determined the
seasonal variation in the structure of the seagrass meadows and structure of associated
fauna. However the information of temporal changes of the fauna in the tropic and
especially in the Colombian Caribbean is scarce. To determine possible variations, the
epibenthic macroinvertebrates associated to seagrass meadows of Thalassia testudinum
were studied in La Guajira in two climatic seasons: minor dry and rainy (2005). Fifteen
stations were chosen in monospecific meadows, with an area greater than 600 m2 and
with a coverage higher than 40%, in which by visual surveys, diving on linear transects
(60 m2) abundance and composition of the epibenthic macroinvertebrate community were
quantified and determined. Diversity analysis, classification (Jaccard and Bray Curtis) and
variance were done to compare composition and density in time. 110 species were
identified (plus a new record for Colombian Caribbean), 42 were common for both
climatic seasons, 32 exclusive of minor dry season and 36 for rainy season. Mollusks
were the dominant group with 30.9% of the total species. Mean density of all individuals
and Evenness index (J’) differed significantly between seasons as well, while Shannon-
Weiner diversity (H i ) and Margalef richness showed no significant differences between
seasons. For the dominant species, mean density of Lytechinus variegatus and Arca
zebra differed significantly beetwen seasons, being greater in the rainy season, while
mean density of Clypeaster rosaceus and Vasum muricatum showed no differences
between seasons. Classification analysis of Bray-Curtis and Jaccard showed that
differences in composition and density may be related with spatial variation. As a result,
variation on the structure of macroinvertebrates associated with seagrass meadows of
Thalassia testudinum for this location of the Colombian Caribbean, in minor dry and rainy
seasons may be explained by structural changes in a spatial and temporal level.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
1
1. INTRODUCCIÓN
Las praderas de pastos marinos cumplen diversas funciones ecológicas que sustentan la
existencia de una gran cantidad de invertebrados que utilizan estos ecosistemas como
hábitats de alimentación, crianza y refugio. Las variaciones climáticas generan cambios
en la estructura de las praderas y por ende en la estructura de la macrofauna de
invertebrados epibentónicos en términos de su composición y abundancia. Este tema ha
sido bien documentado en las praderas de la Florida y el Golfo de México en donde
dichas variaciones tienen un comportamiento estacional marcado. Los ambientes
tropicales por su parte, suelen presentar bajos niveles de estrés físico y pequeñas
variaciones climáticas, pese a que algunos estudios afirman que el crecimiento y
productividad de los pastos guardan una estrecha relación con los cambios climáticos.
En el Caribe colombiano este tipo de información es escasa, por lo tanto se desconocen
las transformaciones que potencialmente pueden presentarse a causa de las variaciones
entre épocas climáticas y cuyos efectos podrían explicar en gran parte la presencia de
ciertos organismos así como su abundancia en un tiempo determinado.
La presencia de macroinvertebrados epibentónicos influye en procesos ecológicos como
herbivoría, flujos tróficos y redes alimentarias, razón por la cual tienen un gran valor
trófico que permite mantener la estabilidad del ecosistema. Igualmente estos organismos
son considerados como buenos indicadores de alteraciones debido a que responden de
manera más directa a los cambios generados por factores naturales y antrópicos. Esto
último resulta de gran importancia pese a que las praderas del sector de La Guajira no
presentan un grado de transformación alto ligado a factores de origen natural (p. ej.
tormentas y huracanes) y/o antrópico (p. ej. sobreexplotación, contaminación, desarrollo
de zonas industriales y turísticas) como otras zonas del Caribe colombiano. Esta
situación se debe a que el desarrollo de ciudades e industrias ha sido menos intenso,
existiendo algunas áreas que pueden ser consideradas como no perturbadas; no
obstante el uso directo de algunos recursos por parte de los pobladores ha venido
aumentando en los últimos años generando preocupación por la conservación de los
ecosistemas marino-costeros, entre ellos las praderas de pastos marinos.
En Colombia los estudios sobre fauna asociada a praderas de pastos marinos son
escasos y actualmente La Guajira, el área con la mayor extensión de praderas en el
Caribe colombiano, no cuenta con la información necesaria para identificar patrones de
variación estacional de las comunidades de macroinvertebrados epibentónicos, siendo
esto un paso elemental para la evaluación del estado de las praderas, de las
comunidades asociadas y para el entendimiento y manejo de estos ecosistemas y sus
recursos.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
2
Este trabajo está basado en la evaluación de los macroinvertebrados epibentónicos
asociados a praderas de Thalassia testudinum (Banks ex könig, 1805) en el
departamento de La Guajira durante dos épocas climáticas (época seca menor y época
de lluvias) y hace parte del proyecto “Caracterización biofísica de la zona costera del
departamento de La Guajira: Una aproximación para su manejo integrado” desarrollado
entre el Instituto de Investigaciones Marinas y Costeras INVEMAR y la Corporación
Autónoma Regional de La Guajira CORPOGUAJIRA. El propósito del presente trabajo
fue determinar la composición y densidad de las especies de macroinvertebrados
epibentónicos presentes en las praderas de Thalassia testudinum y observar posibles
cambios a causa de las variaciones climáticas durante las épocas seca menor y lluvias.
La información generada en este trabajo representa un aporte al conocimiento de las
praderas de pastos marinos del departamento de La Guajira y de la fauna de
macroinvertebrados epibentónicos asociados y es considerada como una meta a corto
plazo dentro de las acciones de cobertura regional del Programa Nacional de
investigación en Biodiversidad Marina y Costera–PNIBM (INVEMAR, 2000) e igualmente
valiosa para la implementación de la Política Nacional Ambiental para el Desarrollo
Sostenible de los espacios oceánicos y las zonas costeras e insulares de Colombia-
PNAOCI (MMA, 2001) Esta Política propone como primer paso caracterizar y
diagnosticar las Unidades Ambientales Costeras y/o Oceánicas (UACO) tanto del Caribe
como del Pacifico para llevar a cabo futuros planes de manejo integrado de zonas
costeras.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
3
2. MARCO TEÓRICO
2.1 Área de estudio
El departamento de La Guajira se encuentra localizado entre los 10º 23´ y 12º 28´ N y los
71º 06´ y 73º 39´ W, en la parte más septentrional de Suramérica y en el extremo
noroeste de Colombia. A pesar de estar integrado a la zona del Caribe colombiano, el
departamento presenta características diferentes en cuanto a relieve, aguas, clima,
suelos y vegetación (IGAC, 1986).
A nivel general, el clima del departamento depende de la posición geográfica de la
península de La Guajira y la influencia de la Sierra Nevada de Santa Marta (IGAC, 1986).
En el extremo norte, la fuerza y dirección de los vientos Alisios que determinan las
condiciones de aridez de la península y condicionan el clima de la región, transportan
gran cantidad de partículas del suelo que se depositan sobre el mar y contribuyen a la
turbidez de las aguas en algunos sectores (Montoya, 2002). El comportamiento de las
lluvias conserva los rasgos típicos del Caribe colombiano, se presentan en los meses de
abril-junio y septiembre-diciembre, siendo el periodo septiembre-noviembre el más
lluvioso. Estos periodos están alternados con otros dos de menores lluvias (época seca)
que se presentan en diciembre-marzo y julio-agosto, este último se denomina “veranillo
de San Juan” (Corredor, 1992). La temperatura es más o menos uniforme y oscila entre
los 24 ºC y 31 ºC, aunque puede variar un poco a razón del cambio en la precipitación, la
humedad y la nubosidad a lo largo del día (Borrero et al., 1996; IGAC, 1986).
Los vientos varían en intensidad y dirección dependiendo de la temporada, durante la
época seca los Alisios del norte son fuertes y uniformes, mientras que en la época de
precipitaciones, llegan más del sur o del sureste y se caracterizan por ser vientos más
lentos y cálidos (IGAC, 1986). El departamento se caracteriza por ser una zona con
surgencia permanente, este fenómeno aumenta la riqueza biológica resultando en una
elevada abundancia de recursos pesqueros que hacen de esta zona la de mayor
actividad extractiva por parte de la flota pesquera del Caribe colombiano (Criales, 2004;
Borrero et al., 1996). El fenómeno de surgencia se extiende desde Punta Chichibacoa en
el Oriente hasta el Cabo de la Aguja en el Occidente y las zonas de mayor intensidad
son el Norte del Cabo de La Vela, en las inmediaciones del Cañon del Ranchería y al
Norte del Cabo de la Aguja (Corredor, 1992).
El área de estudio incluye la zona comprendida entre Riohacha y la Bahía de Portete, sin
embargo la búsqueda de praderas de pastos marinos en este proyecto se realizó en toda
la zona costera del departamento de La Guajira desde Castilletes hasta Palomino. En la
zona comprendida entre Riohacha (11º 32´ N, 72º 56´ W) y el Cabo de La Vela (12º 08´
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
4
N, 72º 15´ W) se encuentran las praderas más amplias del Caribe colombiano, ocupando
una extensión de aproximadamente 33174 hectáreas (Figura 1). En algunos sectores las
praderas se pueden extender hasta más de 10 km afuera de la costa (Díaz et al., 2003)
debido a la amplitud de la plataforma continental en esta zona (Díaz, 1991; INVEMAR,
2003). Por su parte Bahía Portete (12º 09´- 16´ N, 71º 51´ - 72´ W), concentra 1320
hectáreas de praderas que se desarrollan entre los 0.5 m y 2 m de profundidad, sin
embargo en la parte suroccidental pueden alcanzar profundidades hasta los 5 m. La
Bahía está rodeada en su mayoría por mangle y las praderas en su mayoría son mixtas
(Thalassia. testudinum - Syringodium filiforme o T. testudinum - S. filiforme - Halodule
wrightii), sin embargo en la parte suroccidental la especie dominante es T. testudinum
(Díaz et al., 2003).
Figura 1. Distribución de las praderas de pastos marinos en el Caribe colombiano. Tomado de Díaz et al., 2003.
2.2 Generalidades
Los pastos marinos son angiospermas acuáticas que completan todo su ciclo de vida
totalmente sumergidas en un medio salino y están adaptadas para formar lechos
extensos (Durako, 1988). Existen 57 especies agrupadas en doce géneros y cuatro
familias; adicionalmente se encuentra otra especie, género y familia (Ruppia maritima,
Ruppiaceae; Tabla 1) de hábitos eurihalinos. Debido a que son plantas dependientes de
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
5
la luz solar para realizar el proceso fotosintético, están limitadas principalmente a zonas
someras en donde encuentran lugares de fijación, que deben contener una cierta
cantidad de materia orgánica y nutrientes necesarios para su supervivencia (Díaz et al.,
2003) su distribución también está controlada por factores de salinidad, temperatura y
corrientes (Durako, 1988).
Tabla 1. Familias y géneros de pastos marinos. Modificado de Short y Coles (2001).
FAMILIA GÉNERO
Zosteraceae Heterozostera
Zostera Phyllospadix Posidoniaceae Posidonia Hydrocharitaceae Enhalus Halophila Thalassia Cymodoceaceae Cymodocea Amphibolis Halodule Syringodium Thalassodendron Ruppiaceae Ruppia
Estos ecosistemas comunes en las zonas costeras de todo el mundo tanto en ambientes
tropicales como en zonas templadas (Ogden, 1980), son altamente productivos (English
et al., 1997) y tienen varias funciones ecológicas importantes como: I) producir fuentes
directas e indirectas de alimento, II) estabilizar los sedimentos para proteger la costa de
la erosión marina, III) proporcionar un hábitat de refugio, crianza y alimentación a
diferentes especies animales, IV) recircular nutrientes y V) proveer sustrato para la
fijación de epífitos, entre otras (Durako, 1988; Díaz et al., 2003; Álvarez-Guillen et al.,
1986).
Las praderas de pastos marinos son particularmente susceptibles a los efectos causados
por las actividades de origen antrópico debido a que se encuentran limitadas a zonas
someras cercanas a la costa (Díaz et al., 2003). Entre otros problemas a los que se
enfrenta el ecosistema se encuentra la eutroficación, la sobrepesca y la destrucción del
hábitat (Díaz et al., 2003; Duffy, 2006). Se considera que en el Caribe colombiano los
efectos antrópicos son de mediana magnitud, sin embargo se presentan áreas en donde
el impacto es mayor (Barrios y Gómez-López, 2001).
2.3 Distribución
Las especies dominantes en las zonas templadas son Zostera marina y Posidonia
oceanica (Kikuchi, 1980) mientras que la especie dominante y mejor estudiada en el
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
6
Atlántico occidental tropical y el Caribe es T. testudinum (Doering y Bone, 1983;
INVEMAR, 2003) que puede encontrarse mezclada con otras especies como S. filiforme
y H. wrightii (Ogden, 1980). Por lo general estas tres especies hacen parte del esquema
de sucesión ecológica que se inicia con el arribo de la especie pionera H. wrightii y
termina por lo general con una pradera dominada casi en su totalidad por T. testudinum
(Díaz et al., 2003), formando praderas monoespecíficas, características del Caribe
colombiano (Díaz y Garzón-Ferreira, 2000). De las 57 especies de pastos conocidas en
el mundo, 40 se encuentran distribuidas en la región occidental del Indo-pacífico;
mientras que solo nueve están registradas en el Atlántico occidental, incluyendo el mar
Caribe (Díaz et al., 2003).
En las zonas costeras colombianas se encuentran cinco géneros y seis especies, H.
wrightii S. filiforme T. testudinum, Halophila baillonis, Halophila decipiens (Barrios y
Gómez-López, 2001) y R. maritima (Pantaleón y Rodríguez, 2004). Ninguna de las
especies anteriores se encuentran reportadas para las costas del Pacifico debido a las
condiciones en el aporte de agua dulce de grandes ríos, turbidez del agua y falta de
sustratos adecuados (INVEMAR, 2003; Díaz y Garzón-Ferreira, 2000). En el Caribe por
su parte, la distribución discontinua de los pastos marinos se debe a la incidencia de
factores que limitan su establecimiento y/o crecimiento como la incidencia de la luz,
profundidad, temperatura, afluencia de aguas dulces, turbulencia, inestabilidad del
sustrato, entre otras (Díaz et al., 2003). En el departamento de La Guajira los principales
factores que impiden el crecimiento son las aguas turbulentas y la inestabilidad del
sustrato probablemente como consecuencia de la alta energía del oleaje provocado por
los vientos Alisios y el oleaje proveniente del noreste (Barrios y Gómez-López, 2001).
Algunas praderas presentes en el Cabo de La Vela y Riohacha crecen de forma
exuberante formando extensas áreas en donde el sustrato es principalmente de origen
litobioclástico (terrígeno, fragmentos de coral, erizos y algas calcáreas). En otros
sectores aledaños a Riohacha y en el extremo septentrional de la península de La
Guajira la ausencia de praderas es atribuible a la turbulencia generada por el oleaje y la
inestabilidad del sustrato (Díaz et al., 2003). Según Montoya (2002), las actividades de
origen antrópico parecen no tener una influencia notable sobre la estructura de las
praderas de la zona costera de La Guajira como en otras zonas del Caribe colombiano,
sin embrago, trabajos anteriores realizados en la Bahía de Portete revelan los efectos
causados por la construcción y funcionamiento de las instalaciones de Puerto Bolívar
(INVEMAR, 1988) que resultaron en la desaparición de un área de aproximadamente
11500 m² de praderas a causa de la construcción y adecuación de terrenos y fondos
para la instalación de muelles y otras obras de infraestructura.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
7
2.4 Fauna asociada
En general la fauna que habita en las praderas puede dividirse en necton (los que
nadan) y bentos (los que permanecen en el fondo), la fauna bentónica por su parte se
divide en infauna y epifauna. Los organismos representantes de la infauna son aquellos
que se encuentran enterrados en el fondo (poliquetos y bivalvos principalmente) mientras
que la epifauna se refiere a aquellos que se ubican sobre el sustrato. Kikuchi y Peres
(1977); en Heck y Orth (1980) establecieron las siguientes subunidades: I) especies que
viven en las hojas incluyendo micro y meiofauna, fauna sésil, epifauna móvil que camina
o se desliza y fauna nadadora, II) especies ubicadas en los tallos y rizomas y III)
especies bajo las hojas de los pastos. Este grupo a la vez puede dividirse en residentes
permanentes y visitantes regulares o esporádicos (Díaz et al., 2003), siendo los últimos
organismos transitorios y migratorios que visitan las praderas de manera regular u
ocasional en busca de alimento y refugio (Peña y Jiménez, 2001). La cercanía entre las
praderas y otros ecosistemas como arrecifes y manglares permite que las especies
puedan trasladarse de un hábitat a otro a diferentes horas del día y con diferentes
propósitos (Ogden, 1980) haciendo que la recirculación de nutrientes no solo se de
dentro de la cadena trófica sino por medio de la producción y exportación de nutrientes a
otros ecosistemas vecinos lo cual crea un complejo sistema de transferencia de materia
y energía (Ogden y Zieman, 1977; Díaz et al., 2003)
Estudios en todo el mundo se han dedicado a comprender los aspectos estructurales de
las comunidades de pastos marinos, concentrándose específicamente en las especies
que habitan sobre las hojas y los vástagos de los pastos. Estos grupos han sido bien
estudiados debido a que entre ellos se encuentran algunas especies con cierto valor
económico pero también porque los métodos para registrar estas especies son
relativamente sencillos (Heck y Orth, 1980). Estos organismos también se consideran
importantes porque responden de manera más inmediata a los cambios generados en el
ecosistema (Virnstein, 1987). Entre las especies más representativas se incluyen
algunos corales como Manicina areolata, Porites porites, P. astreoides, Siderastrea
radians, Cladocora arbuscula, y algunas especies del genero Millepora y Diploria así
como esponjas, tunicados, anémonas, equinodermos (p. ej. Lytechinus variegatus,
Tripneustes ventricosus, Istichopus badionotus, Oreaster reticulatus), moluscos (p. ej.
Strombus gigas, Cassis tuberosa, Modulus modulus, Cerithium spp.) y crustáceos (p. ej.
Tozeuma carolinense, Panulirus argus, Portunus spp; Ogden, 1980; Montoya, 2002).
Otros organismos como tortugas y peces juegan un importante papel en el ecosistema,
entre ellos se destacan la tortuga verde Chelonia mydas y algunos peces de las familias
Scaridae, Acanthuridae, Labridae, Scorpaenidae entre otros (INVEMAR, 2003).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
8
Cada una de las especies asociadas a las praderas cumple una función importante
dentro del ecosistema y depende directa o indirectamente de los recursos
proporcionados por los pastos ya sea refugio, alimentación o áreas de crianza
(Rodríguez y Villamizar, 2000; INVEMAR, 2003) principalmente porque las hojas y
rizoides brindan un sustrato adecuado a una gran variedad de fauna y porque los epífitos
que crecen sobre la superficie de las hojas son el principal sustento de alimento para
otros organismos (Jiménez, 1994). Como recurso directo de alimentación, los pastos son
elementales para ciertos consumidores herbívoros que se alimentan principalmente de
las hojas y/o de los epifitos que crecen sobre ellas (Ogden, 1980), los consumidores más
notables incluyen gasterópodos y equinodermos siendo los erizos los más importantes
en términos de consumo directo; en el trópico las especies sobresalientes son Diadema
antillarum, T. ventricosus y L. variegatus. Otros organismos como gasterópodos,
anfípodos, isópodos, cangrejos y otros crustáceos pueden consumir pastos
accidentalmente mientras se alimentan de las algas epifitas y el detrito (Thayer et al.,
1984; Ogden, 1980). La herbivoría es un importante medio de obtención de energía
(Valentine et al., 2000) y la vía del detrito y el material exportado a diferentes sistemas,
que es consumido a grandes distancias por otros organismos, también son vías tróficas
importantes (Thayer et al., 1984), la vía del detrito en particular es esencial en la
transferencia de energía a niveles tróficos superiores por lo que el grupo de los
detritívoros suele ser abundante en las praderas (Ansari et al., 1991; Montoya, 2002). En
otras estrategias de alimentación en las praderas, son comunes los carnívoros (corales,
anémonas, algunos crustáceos, gasterópodos y equinodermos), la comunidad de
invertebrados filtradores y detritívoros (bivalvos, gusanos poliquetos, esponjas, ascidias,
equinodermos y algunos crustáceos) y los omnívoros (algunos gasterópodos y
crustáceos), así mismo una gran cantidad de peces de diversos hábitos alimentarios
juegan un papel esencial controlando de cierto modo la estructura y composición en las
praderas. De esta manera las praderas brindan recursos a una gran cantidad de
organismos de diversos tamaños, formas de vida y estrategias de alimentación (Díaz et
al., 2003) que mantienen en equilibrio el funcionamiento del ecosistema en términos del
flujo de energía y materia (Krebs, 1985; Cruz-Ábrego et al., 1994).
En cuanto a espacios de fijación y establecimiento, los pastos brindan sustrato a una
inmensa cantidad de especies, las hojas, estolones y la base de los vástagos actúan
como microhábitats de refugio y alimentación, por lo que se ha sugerido que el número
de especies de invertebrados asociados a los pastos está directamente relacionado con
la complejidad del hábitat, principalmente con la biomasa (Virnstein et al., 1984). En este
sentido, en las praderas de pastos marinos se alberga una mayor densidad de peces e
invertebrados que en otros hábitats de fondos sin vegetación (Heck y Orth, 1980; Orth et
al., 1984). Según Duffy (2006) la complejidad de las praderas de pastos marinos no solo
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
9
se deriva de los aspectos físicos y estructurales de las plantas sino de la riqueza e
identidad de otros organismos sésiles como las esponjas que proveen un importante
hábitat para muchas especies que se refugian debajo y dentro de ellas.
En el Caribe colombiano la estructura de la comunidad está controlada, entre otros
factores, por atributos estructurales como la longitud de las hojas y el grado de epifitismo
(Montoya, 2002); no obstante, aunque la mayoría de estudios sugieren una relación
positiva entre los atributos de la pradera y la diversidad de especies, se considera que
algunas interacciones biológicas (p. ej. competencia y predación) juegan un papel
determinante (Orth et al., 1984). Como sitios de crianza o áreas nodriza, es fundamental
destacar el papel que las praderas cumplen para el desarrollo y supervivencia de
diferentes especies de importancia ecológica y comercial como el caracol pala Strombus
gigas (Stoner, 2003) y la langosta espinosa P. argus (Duffy, 2006) entre otros.
La diversidad varía considerablemente a escala local y geográfica siendo la estructura
física y la complejidad del hábitat los factores que más influyen en la composición y
abundancia de especies (Heck, 1979; Heck y Orth, 1980; Ogden, 1980). Se ha sugerido
que existe un patrón latitudinal que explica en gran parte la complejidad estructural de
las praderas de pastos marinos (Tabla 2), entre otros factores se incluye la productividad
como una variable que tiende a aumentar a medida que decrece la latitud, lo cual puede
explicar en gran parte la diversidad de los sistemas tropicales (Virnstein et al., 1984).
Tabla 2. Comparación general de las praderas de pastos marinos en sistemas fríos / templados, templados y subtropicales y tropicales (Heck y Orth, 1980; Virnstein et al., 1984)
Sistemas fríos / templados
Sistemas templados y
subtropicales
Sistemas tropicales
Patrón estacional Marcado. Marcado. Pocas variaciones climáticas anuales.
Complejidad de las praderas
Sistemas poco complejos, praderas simples con solo pastos
Sistemas medianamente complejos, praderas con pastos, algas a la deriva y esponjas
Sistemas muy complejos, praderas con pastos, algas rojas, algas calcáreas verdes y esponjas
Distribución de las praderas
Amplias praderas sin interrupción.
Amplias praderas sin interrupción.
Praderas separadas por arrecifes, parches coralinos y canales de arena.
Biomasa
Variación estacional
Variación estacional
Poca variación
Productividad
Valores bajos
Valores medios
Valores altos
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
10
Las praderas estructuralmente complejas permiten que diferentes especies encuentren
una fuente de alimento y refugios para esconderse de sus depredadores por lo que se
puede hallar un gran número de especies (muchas veces raras) y una distribución más
uniforme de las abundancias relativas entre especies (Hooks et al., 1976; Orth et al.,
1984; Virnstein, 1987). Las variaciones en el crecimiento y productividad de las praderas
de pastos marinos inciden de manera indirecta sobre otros procesos ecológicos como
herbivoría, flujos tróficos y redes alimentarias. Dichos cambios estructurales varían en el
tiempo tanto en las zonas frías y templadas (Cancemi., et al 2002; Fernández-
Torquemada y Sánchez-Lizaso, 2003) como en ambientes subtropicales y tropicales
(Solana-Arellano, 2001; Garzón-Ferreira y Rodríguez-Ramírez, 2003; Franke, 2001) y
tienen una incidencia importante sobre la fauna acompañante aún cuando se cree que
los ambientes tropicales presentan bajos niveles de estrés físico y pequeñas variaciones
climáticas intranuales (Heck, 1977).
En los sistemas fríos y templados en donde se registra una estacionalidad marcada, se
han comprobado las variaciones en la estructura de la fauna asociada a pastos marinos,
en cuanto a composición y abundancia. Trabajos como el de Nakaoka et al., 2001 han
demostrado que existe una mayor abundancia y riqueza de especies en verano y otoño y
una menor abundancia y riqueza en invierno y primavera, lo cual se encuentra
principalmente relacionado con los cambios estructurales en las praderas. En los
sistemas tropicales y subtropicales aun cuando no existe un patrón estacional marcado,
también se presentan variaciones intranuales en la composición y abundancia de
especies, relacionados principalmente con los cambios en la estructura física de la
praderas y con los cambios de algunas variables fisicoquímicas (Klumpp, 2005; Heck,
1977; Franke, 2001).
La temperatura, salinidad y transparencia del agua guardan estrechas relaciones entre si
con la presencia de los vientos Alisios y la surgencia costera en el Caribe colombiano
(Garzón-Ferreira y Rodríguez-Ramírez, 2003) y como se ha mencionado, La Guajira se
caracteriza por la presencia de estos vientos que varían en intensidad y dirección
produciendo un fenómeno de surgencia que lleva consigo nutrientes y oxígeno disuelto
proporcionando las condiciones adecuadas para el desarrollo de organismos y como
consecuencia, un aumento en la productividad biológica (Corredor, 1992). Este evento
presenta una marcada estacionalidad registrándose durante la época seca un transporte
vertical máximo mientras que en la temporada de lluvias disminuye llegando en
ocasiones a ser nulo (Álvarez-León et al., 1995). El efecto de las lluvias por su parte
parece tener un efecto sobre el crecimiento y productividad de las plantas de Thalassia
(Garzón-Ferreira y Rodríguez-Ramírez, 2003).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
11
Los cambios en la composición y abundancia de la fauna en las praderas también se
relacionan con las épocas de reproducción de las especies que pueden ocurrir en
diferentes temporadas. En las zonas tropicales la ocurrencia de especies en un
ecosistema está relacionada con el concepto de programación estacional que significa,
que a lo largo del año la producción en un sistema se mantiene por medio de diferentes
grupos de productores primarios. Esto implica que exista una diferencia en el uso
temporal y espacial del ecosistema, que coincide simultáneamente, con el desarrollo
ontogénico de una especie y los cambios de estacionalidad; principalmente patrones de
corrientes y producción primaria (García-Hernández, 2003; Yañez-Arancibia y Nugent,
1988).
Adicional a las variaciones estacionales, la estructura física de las praderas y la
estructura de la fauna asociada se ve influenciada por los cambios espaciales ya que las
praderas de pastos marinos son muy disímiles entre sí incluso en áreas cercanas (Brook,
1978; Virnstein, 1987; Angel y Polania, 2000). Los cambios espaciales pueden llegar a
ser mayores que los cambios estacionales ya que las variaciones climáticas no eliminan
las diferencias que separan a las praderas geográficamente cercanas (Franke, 2001); en
estos términos hablar de manera general sobre la dinámica de este ecosistema es
prematuro ya que cada pradera presenta diferencias importantes en su estructura y
composición (Díaz et al., 2003). En la dinámica del ecosistema juegan diferentes
variables estructurales (cobertura, densidad biomasa) y ambientales (profundidad, tipo
de sedimento, distancia de ecosistemas cercanos) por lo cual una sola variable
estructural o ambiental no puede explicar por si sola la estructura de la macrofauna
asociada a las praderas de pastos marinos (Montoya, 2002).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
12
3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN
3.1 Formulación del problema
A pesar de ser los ambientes someros más productivos y de tener una reconocida
importancia por sus funciones ecológicas, las praderas de pastos marinos no han
recibido mayor atención en Colombia en donde los estudios han sido de carácter local o
se han concentrado en grupos muy específicos de organismos con un valor ecológico o
comercial (principalmente moluscos y peces). En el trópico existe información acerca de
patrones de variación temporal en la estructura de las praderas de pastos marinos, sin
embargo se desconocen los efectos de dichas variaciones sobre la composición y
abundancia de la fauna de macroinvertebrados epibentónicos. Si bien los trabajos
desarrollados son un gran aporte al conocimiento, al evaluar la información existente
sobre las comunidades de macroinvertebrados epibentónicos asociados a praderas de T.
testudinum, es posible detectar la falta de información sobre la composición y
abundancia de especies, interacción con el ecosistema y cambios estacionales que
eventualmente puedan presentarse en las comunidades asociadas. La información en el
departamento de La Guajira, que representa algo más del 80.2% (aproximadamente
34674 hectáreas) del total de praderas existentes en el Colombia (Díaz et al., 2003) es
muy escasa si se compara con otros estudios realizados en el mundo principalmente La
Florida y el Golfo de México, esta situación impide tener un conocimiento actualizado de
las especies y de su papel dentro del ecosistema así como de los cambios intranuales
que puedan modificar la estructura de las comunidades.
3.2 Pregunta de investigación
¿Cómo varía la estructura de la comunidad de macroinvertebrados epibentónicos
asociados a praderas de T. testudinum, en dos épocas climáticas (seca menor y lluviosa)
en el departamento de La Guajira?
3.3 Justificación de la investigación
A pesar que las praderas en el sector de La Guajira no muestran un grado de amenaza
evidente en comparación con otras áreas del Caribe colombiano, su peculiaridad en
cuanto a características e importancia ecológica merece el estudio y reconocimiento de
los organismos que se encuentran en este ecosistema, así mismo es importante estudiar
y entender como los cambios climáticos generan variaciones estructurales en las
praderas y por ende, en las comunidades de macroinvertebrados epibentónicos
asociados. El análisis de la estructura y composición de los macroinvertebrados en
diferentes épocas climáticas es elemental, dada la importancia de estos dentro del
sistema trófico como componentes fundamentales para el mantenimiento del equilibrio
del ecosistema y en consecuencia, para la sobrevivencia de otras especies de
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
13
importancia ecológica y comercial. Teniendo en cuenta que los cambios climáticos
pueden afectar ciertas propiedades de las praderas influyendo sobre la fauna
acompañante, modificando procesos ecológicos tales como herbivoría, flujos tróficos y
redes alimentarias así como procesos de reproducción, reclutamiento y migración entre
otros, es importante identificar el papel ecológico de los macroinvertebrados
epibentónicos en estos ecosistemas para en un futuro definir propuestas de manejo y
conservación de especies a partir de la planeación y desarrollo de proyectos.
Los resultados de este trabajo, junto con los obtenidos durante la continuidad del
macroproyecto permitirán aumentar el conocimiento actual del estado y la dinámica
estacional de los macroinvertebrados epibentónicos en las praderas de pastos marinos
del departamento de La Guajira, con el fin de proteger la biodiversidad y evaluar la
posibilidad de iniciar proyectos de aprovechamiento de especies promisorias que
actualmente no se estén explotando debido a falta de información que impiden avanzar
en la toma de medidas para el manejo y conservación de este ecosistema y sus
recursos.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
14
4. HIPÓTESIS
H1: La densidad de la fauna de macroinvertebrados epibentónicos asociados a las
praderas de T. testudinum en La Guajira varía en las épocas seca menor y lluvias.
Predicción:
Existe una gran variación de la composición y abundancia de la macrofauna asociada a
T. testudinum en diferentes épocas climáticas debido a que las variables fisicoquímicas y
estructurales (p. ej. temperatura, salinidad, profundidad, productividad, biomasa,
densidad y tamaño de las hojas) en las praderas guardan una estrecha relación con las
variaciones climáticas, lo cual tiene una incidencia sobre otros procesos ecológicos como
herbivoría, flujos tróficos y redes alimentarías, entre otros (Garzón-Ferreira y Rodríguez-
Ramírez, 2003). Los cambios en las abundancias así como la dominancia de algunas
especies es atribuible a la variación temporal de las condiciones fisicoquímicas que
afectan el reclutamiento, la supervivencia y la reproducción, entre otros (Posey et al.,
1998; Franke, 2001).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
15
5. OBJETIVOS
5.1 Objetivo general
Evaluar la estructura de la macrofauna de invertebrados epibentónicos asociados a
praderas de T. testudinum en dos épocas climáticas (seca menor y lluvias) en la
plataforma continental en el departamento de La Guajira.
5.2 Objetivos específicos
1. Determinar la estructura de la comunidad de macroinvertebrados epibentónicos en
dos épocas climáticas en cuanto a la composición, densidad, diversidad, riqueza y
equitatividad para departamento de La Guajira.
2. Comparar la densidad, diversidad, riqueza y equitatividad de la comunidad de
macroinvertebrados epibentónicos y la densidad de las especies dominantes en dos
épocas climáticas (seca menor y lluvias).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
16
6. MATERIALES Y MÉTODOS
Esta investigación evaluó los cambios en la composición, abundancia, diversidad,
riqueza y equitatividad de la comunidad de macroinvertebrados epibentónicos asociada a
praderas de T. testudinum en dos épocas climáticas (seca menor y lluvias), así mismo se
evaluaron las densidades de las especies dominantes. En total se establecieron quince
estaciones de muestreo durante este estudio (Figura 2), catorce iniciales en la época
seca menor, de las cuales doce se repitieron en la época de lluvias (Anexo A). Las dos
restantes (Cabo de La Vela II y Mayapo) no pudieron ser muestreadas debido a las
malas condiciones de visibilidad causadas por mar de leva, pese a esto se estableció
una nueva estación en Bahía de Portete con el nombre de Puerto Portete para un total
de trece estaciones en esta época climática. La unidad de muestreo consistió en un
transecto lineal de 30 m x 2 m (60 m²) en donde se realizaron censos visuales de la
fauna de macroinvertebrados epibentónicos por medio de buceo autónomo (scuba). El
área de muestreo corresponde a la sugerida por Díaz et al., 2003.
Figura 2. Mapa del área de estudio mostrando la ubicación de las estaciones en la zona costera del departamento de La Guajira. Alta Guajira: PC (Punta Cocos), PP (Puerto Portete), W (Warreo) y Media Guajira: CV1 (Cabo de La Vela I), CV2 (Cabo de La Vela II), EC (El Cardón), FT (Torima), SR2 (Santa Rosa II), SR1 (Santa Rosa I), M1 (Manaure I), M2 (Manaure II), MU (Musiche), T (Tawaya), B (Ballenas) y MY (Mayapo). Fuente: Laboratorio de Sistema de Información Geográfica (SIG) INVEMAR.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
17
En este estudio se tuvieron en cuenta todas las especies de crustáceos, equinodermos,
moluscos, cnidarios y esponjas y los únicos poliquetos censados fueron el gusano de
fuego Hermodice carunculada y el gusano sésil Eupolymnia sp. Las estaciones fueron
establecidas en praderas monoespecíficas de T. testudinum con un área no inferior a
600 m², con una cobertura superior al 40%, según la escala de Dahl (Figura 3). Cada
estación fue georeferenciada con un geoposicionador satelital (GPS) marca GARMIN
GPS MAP 76S. Debido a las limitaciones de tiempo para encontrar praderas con las
características antes mencionadas, no fue posible establecer las estaciones a
profundidades similares (Anexo A).
Figura 3. Representación esquemática del porcentaje de cobertura según la escala de Dahl (1981). Modificado de English et al. (1997).
El trabajo en campo se realizó en el área costera y marina del departamento de La
Guajira; sin embargo las estaciones fueron ubicadas en donde se hallaron praderas con
las características anteriormente mencionadas. Quince estaciones de muestreo fueron
establecidas en total, de Norte a Sur en la Bahía de Portete y entre el Cabo de La Vela y
Riohacha de la siguiente forma: 1) Punta Cocos, 2) Puerto Portete, 3) Warreo, 4) Cabo
de La Vela I, 5) Cabo de La Vela II, 6) El Cardón, 7) Torima, 8) Santa Rosa II, 9) Santa
Rosa I, 10) Manaure I, 11) Manaure II, 12) Musiche, 13) Tawaya, 14) Ballenas y 15)
Mayapo.
La variable independiente correspondió a la época climática (época seca menor de julio-
agosto y época de lluvias en el mes de noviembre) y la diversidad, riqueza, equitatividad
y densidad fueron las variables dependientes. La composición y abundancia se
definieron a partir de la observación de cuales especies y cuantos individuos de cada
una de ellas se encontraron en el transecto y la densidad se calculó a partir de los
valores de abundancia como el número de individuos encontrados en 1 m².
6.1 Fase de Campo
A fin de entrenar a los investigadores en el reconocimiento y conteo de especies en
campo se realizó un pre-muestreo en la Bahía de Chengue (Parque Nacional Natural
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
18
Tayrona). Después de esto y ya establecidos en el área de estudio se realizaron
recorridos de reconocimiento general con observaciones rápidas que permitieron
establecer características acerca de la densidad y composición de la pradera,
adicionalmente se registraron algunas variables fisicoquímicas (profundidad, salinidad y
temperatura). Para establecer el transecto se utilizó un decámetro de 30 m amarrado en
sus extremos a dos boyas que permitieron ubicar el área a muestrear desde la
embarcación, este fue fijado al fondo con la ayuda de dos estacas metálicas a fin de
mantener estática la línea base. Luego de colocar el decámetro, dos buzos procedieron
a identificar y contar los organismos presentes, las especies que no lograron ser
identificadas en campo se colectaron en bolsas plásticas de cierre hermético y luego en
tierra se ubicaron en frascos plásticos con alcohol al 70%, debidamente rotulados y
marcados. El material no identificado en campo fue transportado al laboratorio en donde
las especies fueron identificadas hasta el nivel taxonómico más bajo posible.
La metodología sugerida por Díaz et al., 2003 incluye arrastres de fondo como método
complementario y ha sido previamente utilizada en trabajos recientes como el de
Montoya (2002) en varias zonas del Caribe colombiano. En este trabajo se realizaron
conjuntamente censos visuales y arrastres de fondo, sin embargo la información acá
presentada solo incluye los resultados de los censos visuales por lo que las especies
registradas, responden a organismos grandes y conspicuos que son observados
fácilmente. Únicamente se realizó el conteo de especies no coloniales (crustáceos,
equinodermos, moluscos y algunos anélidos) mientras que de las especies coloniales
(corales, esponjas, tunicados y algunos hidroides) se tomaron datos de presencia, esto
debido a que el conteo de estos organismos en campo se dificulta debido a su cobertura.
6.2 Fase de laboratorio
El material no identificado en campo, fue llevado al INVEMAR en donde se identificó por
medio de claves taxonómicas y con la ayuda de esteroscopio. Para equinodermos se
utilizó el trabajo de Hendler et al., (1995), para los moluscos se empleó Díaz y Puyana
(1994) y Ríos (1994), Rathbun (1925), Rathbun (1930), Chace (1972) y Holthuis (1952)
para crustáceos y se contó con la asesoría de expertos para identificación y confirmación
de diferentes especies. Esponjas (Sven Zea), crustáceos (Norella Cruz y Néstor
Campos), moluscos (Adriana Gracia), corales y octocorales (Javier Reyes y Juan
Armando Sánchez).
6.3 Análisis de información
Una vez obtenidos los datos de los censos visuales en su totalidad, la información fue
consignada en formatos de papel, así mismo se elaboró una base de datos con la
información para cada época climática. En esta base de datos se incluyó información
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
19
acerca de los grupos colectados, especies, número de individuos registrados, fecha de
colecta, estación de muestreo y coordenadas geográficas. Por otra parte, se elaboraron
tablas de frecuencia de ocurrencia, abundancia, densidad y desviación estándar de los
valores promedio de las densidades y se escogieron las especies dominantes teniendo
en cuenta principalmente la frecuencia de ocurrencia y luego la abundancia en cada
época climática. Normalmente se sugiere graficar la frecuencia de ocurrencia de las
especies y sus respectivas abundancias (Digby y Kempton, 1987) lo que permite obtener
varias categorías como las empleadas por Montoya (2002): a) especies ampliamente
distribuidas y muy abundantes, b) ampliamente distribuidas pero poco abundantes, c)
poco distribuidas pero muy abundantes, d) poco distribuidas y poco abundantes, e)
restringidas y muy abundantes, f) restringidas y poco abundantes y g) restringidas y
escasas.
Para el análisis de comunidades se realizaron matrices con los datos de las abundancias
de las especies por época climática, eliminando las estaciones no pareadas es decir
aquellas que no fueron muestreadas en las dos épocas climáticas (n= 12 estaciones).
Con los datos obtenidos se calcularon los índices de Diversidad de Shannon-Weiner
(H i ), índice de Equitatividad de Pielou (J´), índice de Riqueza de Margalef y se
realizaron curvas de k – dominancia utilizando el programa PRIMER 5. La densidad
media de individuos (número de individuos / m²) fue calculada a partir de las
abundancias con el fin de determinar si existieron o no diferencias estadísticas de estos
valores durante las dos épocas climáticas. Luego de cumplir con los requisitos de
normalidad mediante la prueba de Shapiro-Wilk y homogeneidad de varianzas a partir de
la gráfica de residuales, se realizaron análisis de varianza a una vía (ANOVA) de bloques
aleatorios utilizando las estaciones como bloques muestreados en cada época climática
(factor principal); este análisis permitió separar las variaciones en magnitud entre
estaciones mostrando las diferencias entre épocas. Empleado el programa SAS (SAS
Institute, 1996) se analizó el comportamiento de los índices calculados, los valores de
densidad total de individuos y los valores de densidad de las especies dominantes
durante las dos épocas climáticas, para lo cual las densidades fueron transformadas
usando ln (x+1).
Como método complementario se realizó un análisis de clasificación normal a partir de
los datos de las abundancias en cada estación y durante cada época climática, para esto
se elaboró una matriz de orden de magnitud basada en log (10) con el fin de escoger las
especies más representativas de la comunidad. Los datos fueron transformados con ln
(x+1) y comparados mediante el índice de similaridad de Bray-Curtis (Field et al., 1982;
Digby y Kempton, 1987). Como estrategia de agrupamiento se utilizó el promedio no
ponderado UPGMA debido a que es uno de los métodos más simples y el que se ha
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
20
encontrado más exitoso en numerosas aplicaciones (InfoStat, 2004). La transformación
de datos es importante para estandarizar los valores empleados en una matriz con el fin
de reducir cualquier efecto de dominancia que logre enmascarar aquellos datos que
tienen una significancia menor (Digby y Kempton, 1987). Por último, se analizaron las
diferencias en la composición de especies en cada época climática empleando el
coeficiente de Jaccard (J) que tiene en cuenta datos cualitativos de presencia/ausencia
(Heck, 1977; InfoStat, 2004). Dicho coeficiente varia entre 0 y 1, donde 0 indicaría la
ausencia de especies en común y 1 demostraría que las épocas son idénticas en
términos de composición.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
21
7. RESULTADOS
7.1 Estructura de la macrofauna asociada
Generalidades
Durante las dos épocas climáticas se identificaron 110 especies pertenecientes a 6 phyla
de invertebrados (Anexo B). En la época seca menor (julio-agosto) se registraron 74
especies y los moluscos fueron el grupo mejor representado (33.8%) seguido por los
cnidarios (23%), crustáceos (14.9%), esponjas, (13.5%), equinodermos (10.8%),
anélidos (2.7%) y tunicados (1.4%). En la época de lluvias (noviembre) se encontraron
79 especies, siendo los moluscos el grupo con el mayor numero de especies (26.6%), le
siguen los cnidarios (21.5%), las esponjas (20.3%), crustáceos (15.2%), equinodermos
(12.7%), anélidos (2.5%) y tunicados (1.3%; Figura 4). 42 especies fueron comunes para
las dos épocas climáticas mientras que 32 fueron exclusivas de la época seca menor y
36 de la época de lluvias.
Figura 4. Comparación de la riqueza total de especies para cada grupo taxonómico encontrado durante la época seca menor y época de lluvias
Moluscos Este grupo se destacó por su frecuencia y abundancia dentro de las comunidades
epifaunales en donde fueron el grupo dominante (Anexo B). Durante la época seca
menor las especies más frecuentes fueron el bivalvo Arca zebra (71.4%) y los
gasterópodos Vasum muricatum (28.6%) y Crucibulum planum (21.4%). A. zebra fue la
especie más abundante aportando el 35.5% del total de individuos del phylum, seguida
por el nudibranquio Bursatella leachii (30.9%) y el gasterópodo Cerithium sp. (21.8%). El
72% de las especies de moluscos estuvo representado por menos de dos individuos
(Tabla 3a) lo cual hace de éstas, no solo poco abundantes sino restringidas en el área de
estudio. En la época de lluvias A. zebra fue la especie más frecuente (76.9%), seguida
por V. muricatum (46.2%), M. modulus y Cerithium eburneum (38.5% cada una). En
cuanto a la abundancia A. zebra, contribuyó con el 86.9% del total de individuos del
phylum, seguida por M. modulus (5.4%) y C. eburneum (2.2%), las demás especies del
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
22
grupo fueron poco abundantes aportando menos del 2% del total de individuos (Tabla
3b).
Tabla 3. Frecuencia de ocurrencia (F%), abundancia (N), y promedio de la densidad (numero de individuos/m²) de las especies encontradas durante: a) época seca menor y b) época de lluvias. La frecuencia fue calculada respecto al total de estaciones de cada época. a)
Especies F (%) N Densidad media (± EE) Lytechinus variegatus 100 558 0.664 ± 0.182 Arca zebra 71.4 192 0.228 ± 0.183 Clypeaster rosaceus 42.9 23 0.027 ± 0.011 Vasum muricatum 28.6 7 0.008 ± 0.004 Crucibulum planum 21.4 7 0.008 ± 0.006 Mithraculus forceps 21.4 5 0.006 ± 0.003 Dardanus fucosus 21.4 3 0.003 ± 0.002 Oreaster reticulatus 21.4 3 0.003 ± 0.002 Cerithium sp. 14.3 118 0.140 ± 0.132 Eupolymnia sp. 14.3 32 0.038 ± 0.037 Echinaster sp.1 14.3 16 0.019 ± 0.016 Cerithium atratum 14.3 6 0.007 ± 0.006 Astraea tecta tecta 14.3 2 0.002 ± 0.002 Holothuria mexicana 14.3 2 0.002 ± 0.002 Pitho lherminieri 14.3 2 0.002 ± 0.002 Bursatella leachii 7.1 167 0.198 ± 0.199 Chama macerophylla 7.1 21 0.025 ± 0.010 Istichopus badionotus 7.1 12 0.014 ± 0.014 Petrochirus diogenes 7.1 3 0.003 ± 0.004 Cassiopea frondosa 7.1 2 0.002 ± 0.002 Cassis madagascariensis 7.1 2 0.002 ± 0.002 Modulus modulus 7.1 2 0.002 ± 0.002 Phyllonotus margaritensis 7.1 2 0.002 ± 0.002 Strombus raninus 7.1 2 0.002 ± 0.002 Alpheus norman 7.1 1 0.001 ± 0.001 Alpheus sp. 7.1 1 0.001 ± 0.001 Anadara baughmani 7.1 1 0.001 ± 0.001 Anadara notabilis 7.1 1 0.001 ± 0.001 Astrophyton muricatum 7.1 1 0.001 ± 0.001 Cerithium eburneum 7.1 1 0.001 ± 0.001 Chama sp. 7.1 1 0.001 ± 0.001 Conus amphiurgus 7.1 1 0.001 ± 0.001 Conus spurius spurius 7.1 1 0.001 ± 0.001 Crepidula plana 7.1 1 0.001 ± 0.001 Echinaster sp.2 7.1 1 0.001 ± 0.001 Hermodice carunculata 7.1 1 0.001 ± 0.001 Hindsiclava chazaliei 7.1 1 0.001 ± 0.001 Hypselodoris bayeri 7.1 1 0.001 ± 0.001 Melongena melongena 7.1 1 0.001 ± 0.001 Microphrys bicornutus 7.1 1 0.001 ± 0.001 Paguristes werdingi 7.1 1 0.001 ± 0.001 Panulirus Aarhus 7.1 1 0.001 ± 0.001 Petrolisthes galathinus 7.1 1 0.001 ± 0.001 Pilumnus pannosus 7.1 1 0.001 ± 0.001 Pinna carnea 7.1 1 0.001 ± 0.001 Pteria colymbus 7.1 1 0.001 ± 0.001 Turritella variegata 7.1 1 0.001 ± 0.001
PDF created with pdfFactory trial version www.pdffactory.com
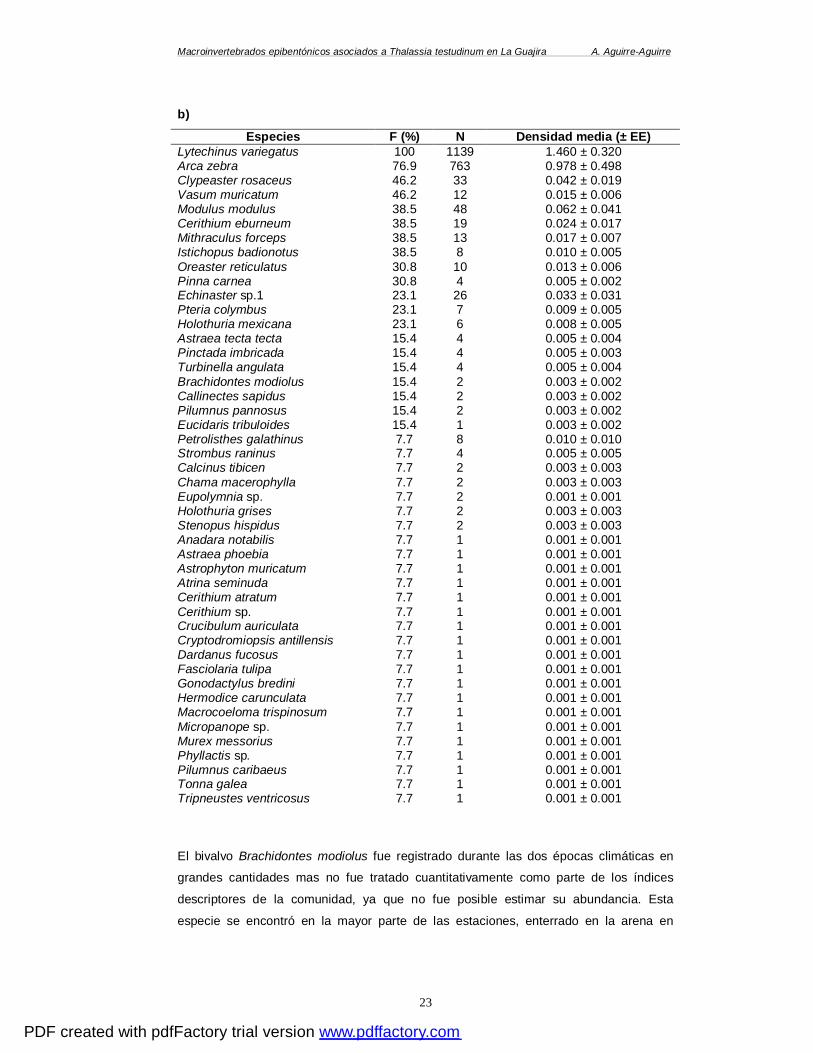
Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
23
b)
Especies F (%) N Densidad media (± EE) Lytechinus variegatus 100 1139 1.460 ± 0.320 Arca zebra 76.9 763 0.978 ± 0.498 Clypeaster rosaceus 46.2 33 0.042 ± 0.019 Vasum muricatum 46.2 12 0.015 ± 0.006 Modulus modulus 38.5 48 0.062 ± 0.041 Cerithium eburneum 38.5 19 0.024 ± 0.017 Mithraculus forceps 38.5 13 0.017 ± 0.007 Istichopus badionotus 38.5 8 0.010 ± 0.005 Oreaster reticulatus 30.8 10 0.013 ± 0.006 Pinna carnea 30.8 4 0.005 ± 0.002 Echinaster sp.1 23.1 26 0.033 ± 0.031 Pteria colymbus 23.1 7 0.009 ± 0.005 Holothuria mexicana 23.1 6 0.008 ± 0.005 Astraea tecta tecta 15.4 4 0.005 ± 0.004 Pinctada imbricada 15.4 4 0.005 ± 0.003 Turbinella angulata 15.4 4 0.005 ± 0.004 Brachidontes modiolus 15.4 2 0.003 ± 0.002 Callinectes sapidus 15.4 2 0.003 ± 0.002 Pilumnus pannosus 15.4 2 0.003 ± 0.002 Eucidaris tribuloides 15.4 1 0.003 ± 0.002 Petrolisthes galathinus 7.7 8 0.010 ± 0.010 Strombus raninus 7.7 4 0.005 ± 0.005 Calcinus tibicen 7.7 2 0.003 ± 0.003 Chama macerophylla 7.7 2 0.003 ± 0.003 Eupolymnia sp. 7.7 2 0.001 ± 0.001 Holothuria grises 7.7 2 0.003 ± 0.003 Stenopus hispidus 7.7 2 0.003 ± 0.003 Anadara notabilis 7.7 1 0.001 ± 0.001 Astraea phoebia 7.7 1 0.001 ± 0.001 Astrophyton muricatum 7.7 1 0.001 ± 0.001 Atrina seminuda 7.7 1 0.001 ± 0.001 Cerithium atratum 7.7 1 0.001 ± 0.001 Cerithium sp. 7.7 1 0.001 ± 0.001 Crucibulum auriculata 7.7 1 0.001 ± 0.001 Cryptodromiopsis antillensis 7.7 1 0.001 ± 0.001 Dardanus fucosus 7.7 1 0.001 ± 0.001 Fasciolaria tulipa 7.7 1 0.001 ± 0.001 Gonodactylus bredini 7.7 1 0.001 ± 0.001 Hermodice carunculata 7.7 1 0.001 ± 0.001 Macrocoeloma trispinosum 7.7 1 0.001 ± 0.001 Micropanope sp. 7.7 1 0.001 ± 0.001 Murex messorius 7.7 1 0.001 ± 0.001 Phyllactis sp. 7.7 1 0.001 ± 0.001 Pilumnus caribaeus 7.7 1 0.001 ± 0.001 Tonna galea 7.7 1 0.001 ± 0.001 Tripneustes ventricosus 7.7 1 0.001 ± 0.001
El bivalvo Brachidontes modiolus fue registrado durante las dos épocas climáticas en
grandes cantidades mas no fue tratado cuantitativamente como parte de los índices
descriptores de la comunidad, ya que no fue posible estimar su abundancia. Esta
especie se encontró en la mayor parte de las estaciones, enterrado en la arena en
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
24
grandes cantidades que no permitieron cuantificar con precisión un número aproximado
de individuos, no obstante se sabe que es localmente abundante en algunas praderas de
La Guajira (Montoya, 2002).
Cnidarios En la época seca menor las especies más sobresalientes fueron los octocorales
Plexaura sp. (57.1%), Pterogorgia sp. (50%) y los corales escleractinios Siderastrea
siderea y Solenastrea bournoni (42.9% cada uno). De las 17 especies registradas
durante esta época, el 76.5% fueron halladas en menos de 2 estaciones. Por su parte,
en la época de lluvias las especies más frecuentes fueron los octocorales Plexaurella
dichotoma (69.2%) y Pterogorgia sp. (38.5%), otros como Eunicea sp., Millepora
complanata y S. bournoni tuvieron una frecuencia del 30.8% durante esta época
mientras que el 70.6% de las especies se encontraron en dos o menos estaciones.
Crustáceos El 68.4% de las especies encontradas correspondieron al Orden Decápoda, mientras el
31.6% fueron del Orden Stomatopoda. Durante la época seca menor las especies más
frecuentes del grupo fueron Mithraculus forceps y el cangrejo ermitaño Dardanus fucosus
(21.4% cada uno). M. forceps también fue la especie más abundante (25% del total de
individuos) mientras que las demás representaron menos del 15%, es decir tres o menos
individuos (Tabla 3a). En la época de lluvias M. forceps fue de nuevo la especie más
frecuente (38.5%) y abundante (37.1%) mientras que el resto de especies fueron
encontradas en dos o menos estaciones y con abundancias muy bajas (Tabla 3b).
Esponjas Se registraron 21 especies de esponjas de las cuales las más frecuentes durante la
época seca menor fueron Aplysina fulva (21.4%), Chondrilla nucula y Dysidea sp. (14.3%
cada una). Las demás especies tuvieron una frecuencia muy baja (7.1%). En cuanto a la
época de lluvias, se encontró que A. fulva fue de nuevo la especie más frecuente
(28.6%) seguida por Cliona varians y Tedania ignis (21.4%). El 81.3% de las especies
restantes estuvo presente en dos o menos estaciones de muestreo.
Equinodermos
A pesar de ser un grupo con una baja riqueza de especies en relación con los demás
taxa encontrados, los equinodermos en general fueron frecuentes y abundantes durante
las dos épocas climáticas. En la época seca menor el erizo blanco L. variegatus fue la
especie más frecuente (100%) y abundante (90.6% del total de individuos del grupo).
seguida por el bizcocho de mar Clypeaster rosaceus que tuvo una frecuencia de 42.9% y
una abundancia de 3.7%. Otras especies como los pepinos de mar I. badionotus y
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
25
Holothuria mexicana fueron poco frecuentes (7.1% y 14.3% respectivamente). En la
época de lluvias. L. variegatus fue de nuevo la especies más frecuentes (100%). al igual
que C. rosaceus (46.2%) e I. badionotus (38.5%). Las tres especies anteriores mostraron
los más altos valores de abundancia del grupo respecto al total de individuos del phylum
(92.8%. 2.7% y 2.1% respectivamente).
Anélidos Durante la época seca menor se encontró que Eupolymnia sp. tuvo una frecuencia de
14.3% mientras que H. carunculata solo fue encontrado en una de las estaciones de
muestreo (7.1%). En la época de lluvias solo se encontró un individuo de cada especie.
Tunicados La especie Botrylloides sp. fue identificada para las dos épocas climáticas. Durante la
época seca menor tuvo una frecuencia de 7.1% mientras que en la época de lluvias la
frecuencia fue de 28.6%. Esta ha sido registrada recubriendo conspicuamente las hojas
de T. testudinum en otras zonas del Caribe colombiano como al interior de la Bahía de
Cartagena (Díaz y Gómez-López, 2003).
7.2 Nuevo registro para el Caribe colombiano.
Una de las especies registradas durante la elaboración de este trabajo constituye un
nuevo registro para el Caribe colombiano. La especie Pilumnus pannosus (Rathbun,
1896) (Decapoda: Brachyura), una de las mejores estudiadas en el Caribe colombiano
(Campos et al., 2004) hace parte de la familia Xanthidae. Rathbun (1930) presenta 25
especies del género Pilumnus (Pilumnidae) para las dos costas americanas. De estas 25
especies nueve han sido registradas para el Caribe colombiano (Lemaitre, 1981;
Campos et al., 2004) y la costa norte de Sudamérica (Rodríguez, 1980). Williams (1984)
describió ocho especies de este género para el Atlántico occidental, algunas de las
cuales han sido registradas para el Caribe colombiano, excepto P. pannosus. Con este
hallazgo, se incrementa a 56 el inventario que se tiene de esta familia (Campos et al.,
2004) en el Caribe colombiano.
Se encontraron tres ejemplares de la especie, uno durante la época seca menor y dos
durante la época de lluvias. En laboratorio fueron separados y preservados en etanol al
70%, se fotografiaron (Figura 5) y finalmente se identificaron utilizando las claves y
descripciones de Rathbun (1930) y Williams (1984). Los ejemplares fueron depositados
en el Museo de Historia Natural Marina de Colombia (MHNMC) en el INVEMAR, con el
número de catalogo INVCRU 4901 y INVCRU 4902.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
26
Figura 5. Imagen de la especie Pilumnus pannosus, nuevo registro para el Caribe colombiano encontrada en praderas de T. testudinum de La Guajira.
7.3 Análisis de comunidades Durante la época seca menor se colectaron 1212 individuos cuya densidad promedio
total fue de 1.442 (± 0.332) individuos/m² mientras que en la época de lluvias se hallaron
2149 individuos con una densidad promedio de 2.754 (± 0.443) individuos/m². Durante la
época seca menor el 91.5% de las especies tuvieron una frecuencia baja encontrándose
en menos de tres estaciones, de estas la mayoría fueron moluscos que también
presentaron abundancias bajas, (excepto B. leachii y Chama macerophylla) y algunos
crustáceos que en su mayoría estuvieron representados por un individuo (Tabla 3a). En
la época de lluvias por su parte, el 78.7% de las especies se encontraron en tres o
menos estaciones, de estas la mayoría fueron moluscos y crustáceos con abundancias
muy bajas.
Mediante los análisis de varianza ANOVA se identificaron diferencias significativas de los
valores de densidad promedio del total de individuos entre épocas climáticas (p =
0.0222), así mismo se registraron diferencias significativas del índice de Equitatividad de
Pielou entre épocas (p = 0.0177), siendo la época seca menor la que presentó la mayor
uniformidad (Tabla 4). Este comportamiento se pudo corroborar mediante las curvas de k
- dominancia (Figura 6) que también muestran a esta época como la más diversa. Los
índices de Diversidad de Shannon-Weiner y Riqueza de Margalef por su parte no
mostraron diferencias significativas (p = 0.1063 y p = 0.8854 respectivamente).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
27
Figura 6. Curvas de k- dominancia para la época de seca menor (S) y época de lluvias (LL).
Tabla 4. Valores de densidad promedio (número de individuos/m²) y de los Índices de Diversidad de Shannon-Weiner (H i Equitatividad de Pielou (J´) y Riqueza de Margalef durante la época seca menor (S) y época de lluvias (LL) y valores promedio con su respectivo error estándar (EE) en cada estación muestreada. Estaciones Densidad
(±EE) Diversidad Equitatividad Riqueza
PC (S) 2.650 ± 0.021 1.78 0.48 2.37 W (S) 0.233 ± 0.004 2.50 0.89 2.27
CV1 (S) 3.067 ± 0.028 1.06 0.31 1.92 CV2 (S) 0.483 ± 0.005 2.85 0.82 2.97 EC (S) 0.200 ± 0.003 2.86 0.95 2.82 FT (S) 1.867 ± 0.025 0.14 0.14 0.25
SR2 (S) 0.383 ± 0.004 3.08 0.86 3.51 SR1 (S) 0.850 ± 0.017 0.15 0.09 0.42 M1 (S) 0.400 ± 0.006 0.50 0.18 1.31 M2 (S) 1.750 ± 0.022 1.98 0.85 1.26 MU (S) 4.433 ± 0.028 0.64 0.32 0.64 T (S) 1.350 ± 0.015 0.22 0.22 0.25 B (S) 1.600 ± 0.022 1.33 0.40 1.61
MY (S) 0.933 ± 0.018 1.11 0.70 0.46 Media (± EE) 1.443 ± 0.332 1.442 ± 0.286 0.516 ± 0.085 1.576 ± 0.293
PC (LL) 3.500 ± 0.029 1.17 0.50 0.96 PP (LL) 1.067 ± 0.015 0.99 0.35 1.12 W (LL) 4.017 ± 0.028 1.45 0.42 1.82
CV1 (LL) 6.700 ± 0.043 0.66 0.21 1.33 EC (LL) 3.000 ± 0.029 0.82 0.24 1.93 FT (LL) 1.533 ± 0.022 0.32 0.11 1.12
SR2 (LL) 0.967 ± 0.008 2.87 0.72 3.69 SR1 (LL) 3.533 ± 0.034 0.34 0.15 0.88 M1 (LL) 2.217 ± 0.017 0.78 0.30 1.15 M2 (LL) 3.717 ± 0.031 2.06 0.54 2.66 MU (LL) 1.650 ± 0.023 0.83 0.30 1.11 T (LL) 2.650 ± 0.024 0.39 0.17 0.87 B (LL) 1.267 ± 0.019 1.26 0.40 1.58
Media (± EE) 2.755 ± 0.443 1.072 ± 0.202 0.338 ± 0.049 1.557 ± 0.227
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
28
Los análisis de similitud de Bray-Curtis (Figura 7) y Jaccard (Figura 8) indicaron que las
estaciones son muy disímiles entre si en cuanto a abundancia y composición y que no
existe una tendencia clara que permita definir grupos a partir de las diferencias entre las
dos épocas climáticas. Esto se pone de manifiesto al observar los resultados de los
dendogramas, en donde se aprecia la formación de diferentes grupos compuestos por
estaciones de distintas localidades en diferentes épocas climáticas, es decir que aun
cuando los análisis de varianza ANOVA indican que la densidad total de individuos fue
significativamente mayor en la época de lluvias, no existe una evidencia clara de que las
diferencias en la estructura de la fauna asociada se deban únicamente a las variaciones
entre épocas climáticas, sino que la variación espacial puede ser incluso mayor.
Figura 7. Representación esquemática del Índice de similitud de Bray-Curtis entre las estaciones de muestreo PC (Punta Cocos), PP (Puerto Portete), W (Warreo), CV1 (Cabo de La Vela I), CV2 (Cabo de La Vela II), EC (El Cardón), FT (Torima), SR2 (Santa Rosa II), SR1 (Santa Rosa I), M1 (Manaure I), M2 (Manaure II), MU (Musiche), T (Tawaya), B (Ballenas) y MY (Mayapo), utilizando los valores de las abundancias transformadas con ln (x + 1) para la época seca menor (S) y época de lluvias (LL).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
29
Figura 8. Representación esquemática del coeficiente de similitud de Jaccard (J´) entre las estaciones de muestreo: PC (Punta Cocos), PP (Puerto Portete), W (Warreo), CV1 (Cabo de La Vela I), CV2 (Cabo de La Vela II), EC (El Cardón), FT (Torima), SR2 (Santa Rosa II), SR1 (Santa Rosa I), M1 (Manaure I), M2 (Manaure II), MU (Musiche), T (Tawaya), B (Ballenas) y MY (Mayapo) en la época seca menor (S) y época de lluvias (LL). Los valores de similitud van de 0 a 100, en donde 0 % representa la menor similitud y 100% la mayor similitud. 7.4 Especies dominantes Las especies dominantes durante las dos épocas fueron L. variegatus. A. zebra. C.
rosaceus y V. muricatum (Figura 9), sin embargo el erizo blanco L. variegatus fue la
única especie que se presentó en todas las estaciones de muestreo durante las dos
épocas climáticas. Los resultados de los análisis de varianza (ANOVA) mostraron
diferencias significativas de los valores de densidad promedio de L. variegatus entre
épocas climáticas (p = 0.0160), resultando en un aumento considerable de individuos
durante la época de lluvias. De acuerdo con estos resultados L. variegatus fue
considerada como una especie ampliamente distribuida y muy abundante en La Guajira.
Los valores de densidad promedio del bivalvo A. zebra también fueron significativamente
diferentes entre épocas climáticas (p = 0.0347) registrándose un valor de densidad
media mayor durante la época de lluvias. En cuanto a C. rosaceus y V. muricatum no se
encontraron diferencia significativas en los valores de densidad durante las dos épocas
climáticas (p = 0.3768; p = 0.2097 respectivamente).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
30
Figura 9. Densidad media (±EE) de las especies dominantes registradas durante la época seca menor y la época de lluvias en el área de estudio.
PDF created with pdfFactory trial version www.pdffactory.com
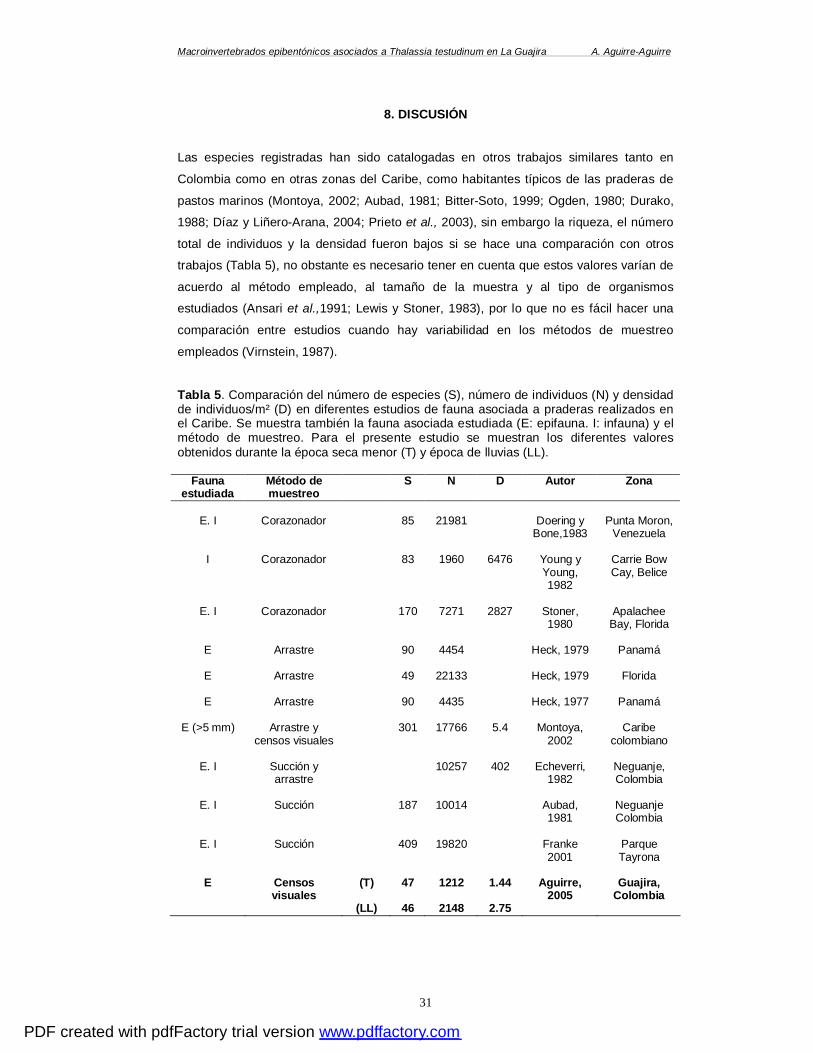
Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
31
8. DISCUSIÓN
Las especies registradas han sido catalogadas en otros trabajos similares tanto en
Colombia como en otras zonas del Caribe, como habitantes típicos de las praderas de
pastos marinos (Montoya, 2002; Aubad, 1981; Bitter-Soto, 1999; Ogden, 1980; Durako,
1988; Díaz y Liñero-Arana, 2004; Prieto et al., 2003), sin embargo la riqueza, el número
total de individuos y la densidad fueron bajos si se hace una comparación con otros
trabajos (Tabla 5), no obstante es necesario tener en cuenta que estos valores varían de
acuerdo al método empleado, al tamaño de la muestra y al tipo de organismos
estudiados (Ansari et al.,1991; Lewis y Stoner, 1983), por lo que no es fácil hacer una
comparación entre estudios cuando hay variabilidad en los métodos de muestreo
empleados (Virnstein, 1987).
Tabla 5. Comparación del número de especies (S), número de individuos (N) y densidad de individuos/m² (D) en diferentes estudios de fauna asociada a praderas realizados en el Caribe. Se muestra también la fauna asociada estudiada (E: epifauna. I: infauna) y el método de muestreo. Para el presente estudio se muestran los diferentes valores obtenidos durante la época seca menor (T) y época de lluvias (LL).
Fauna estudiada
Método de muestreo
S N D Autor Zona
E. I
Corazonador
85
21981
Doering y
Bone,1983
Punta Moron,
Venezuela I
Corazonador
83
1960
6476
Young y Young, 1982
Carrie Bow Cay, Belice
E. I
Corazonador
170
7271
2827
Stoner, 1980
Apalachee Bay, Florida
E
Arrastre
90
4454
Heck, 1979
Panamá
E
Arrastre
49
22133
Heck, 1979
Florida
E
Arrastre
90
4435
Heck, 1977
Panamá
E (>5 mm)
Arrastre y
censos visuales
301
17766
5.4
Montoya,
2002
Caribe
colombiano
E. I
Succión y arrastre
10257
402
Echeverri,
1982
Neguanje, Colombia
E. I
Succión
187
10014
Aubad, 1981
Neguanje Colombia
E. I
Succión
409
19820
Franke 2001
Parque Tayrona
E
Censos visuales
(T)
47
1212
1.44
Aguirre,
2005
Guajira,
Colombia (LL) 46 2148 2.75
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
32
Las especies registradas en este trabajo respondieron a organismos grandes y
conspicuos que fueron observados fácilmente mediante los censos visuales, resultando
en una mayor contribución de estos a la riqueza y abundancia.
La dominancia de los moluscos en las praderas de pastos marinos se atribuye
principalmente a la amplia variedad de formas de vida y estrategias de alimentación que
presenta este grupo (Díaz, 2003). En otros estudios similares también fueron un grupo
dominantes (Echeverri, 1982; Barrios y Gómez-López, 2001; Montoya, 2002; Cruz-
Ábrego et al., 1994; Brook, 1978; Young y Young, 1982; Greenway, 1995) lo que
coincide con los resultados de este trabajo en donde aportaron el 30.91% del total de
especies.
8.1 Análisis de comunidades
La variación temporal de los valores de densidad total de individuos y la dominancia de
algunos taxa se ha relacionado en otros estudios con la variación temporal de las
condiciones físicas del medio que pueden afectar el reclutamiento, la supervivencia y la
reproducción de algunas especies (Posey et al., 1998). Los resultados obtenidos en este
trabajo demuestran que existen diferencias significativas en la densidad de individuos en
dos épocas climáticas (seca menor y lluvias) y coinciden con los resultados de otros
trabajos similares realizados en el Caribe, en donde se han registrado cambios no solo
en la abundancia sino en la composición de especies; sin embargo al observar los
resultados del análisis de similitud de Bray-Curtis y Jaccard queda claro que las
diferencias espaciales pueden ser mayores que las variaciones entre épocas climáticas
por lo cual no es posible concluir que los cambios en la estructura de fauna asociada a
praderas de T. testudinum se deban únicamente a las variaciones estacionales sino que
las diferencias espaciales entre dos áreas pueden ser mayores, ya que los cambios
climáticos no eliminan las diferencias que las separan, tal como lo concluyó Franke
(2001) en su trabajo en el Parque Nacional Natural Tayrona..
En cuanto a los cambios que se presentan entre épocas climáticas, se encuentran las
variaciones en la temperatura, salinidad y transparencia del agua, factores determinantes
para el proceso de fotosíntesis y para el metabolismo, reproducción, crecimiento e
incluso supervivencia de las fanerógamas marinas (Fernández-Torquemada y Sánchez-
Lizaso, 2003). Pese a que dichos factores fueron medidos en este trabajo, no fueron
suficientes para encontrar relaciones claras como evidencia de las diferencias entre las
épocas seca menor y lluvias, sin embargo se sabe que estos factores guardan una
estrecha relación con la presencia de los vientos Alisios y la surgencia costera durante
los meses de diciembre-marzo y julio y la influencia de las lluvias durante mayo-junio y
septiembre-noviembre (Garzón-Ferreira y Rodríguez-Ramírez, 2003) y pueden afectar en
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
33
gran medida la estructura física de las praderas, influyendo de manera indirecta sobre la
estructura de la fauna asociada (Heck, 1979; Ansari et al., 1991; Knowles y Bell, 1998;
Virnstein, 1987). Según los resultados obtenidos por Garzón-Urbina (en preparación)
durante la ejecución de este proyecto los valores de biomasa, cobertura y densidad de
vástagos tomados en cada estación fueron mayores en la época de lluvias, hecho que
permitiría suponer que la estructura de las praderas se ve afectada por los cambios de
climáticos que a su vez afectan la estructura de la fauna asociada. En este sentido
Garzón-Ferreira y Rodríguez-Ramírez (2003) afirman que el efecto de las lluvias es
determinante sobre el crecimiento y productividad de las plantas de Thalassia que en
consecuencia incidiría sobre otros procesos ecológicos.
Las praderas de pastos marinos están sujetas a perturbaciones que varían tanto en
espacio como en tiempo y la diversidad de la fauna asociada se modifica a razón de
estos cambios (Heck y Orth, 1980) incluso en las zonas tropicales en donde los
fenómenos estacionales son decisivos para el funcionamiento de las comunidades
naturales (Krebs, 1985); sin embargo otras variables son fundamentales en la
organización de las comunidades; la distribución de la fauna, así como la diversidad,
riqueza y densidad de individuos son el resultado de las interacciones de diferentes
factores (Bitter-Soto, 1999) entre los que cabe mencionar la presencia de ecosistemas
cercanos como manglares, arrecifes de coral y fondos blandos (Cruz-Ábrego et al., 1994;
Montoya, 2002) y otros factores como competencia, depredación migración,
reproducción y reclutamiento (Hooks et al., 1976; Heck, 1977; 1979; Coen et al., 1981;
Young y Young, 1982; Orth et al., 1984; Livingston, 1984).
Según Montoya (2002) los cambios en la composición y abundancia de especies en las
praderas del Caribe colombiano no responden a una única variable, el origen del
sedimento y la ubicación geográfica así como la longitud de las hojas, el grado de
epífitismo y la profundidad son importantes atributos que controlan la estructura de la
macrofauna asociada a T. testudinum. En el caso de las praderas de la Guajira
muestreadas en este trabajo, las diferencias posiblemente están relacionadas con la
ubicación geográfica ya que las estaciones establecidas en Bahía Portete se encuentran
en una Bahía protegida con condiciones oceanográficas particulares muy diferentes a la
que se presentan frente a los casi 150 km de línea de costa comprendidos entre el Cabo
de La Vela y Riohacha (Díaz et al., 2003), en donde se ubicaron la mayor parte de las
estaciones (Figura 2). Adicionalmente, otras variables como la profundidad pueden haber
influido en la estructura de la macrofauna asociada teniendo en cuenta que las
estaciones no fueron ubicadas a profundidades similares y que esta variable es
determinante en la configuración y estructura de los ecosistemas acuáticos (Krebs,
1985).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
34
Se encontraron diferentes especies de macroinvertebrados epibentónicos en las
praderas de T. testudinum de La Guajira que aprovechan este ecosistema con diferentes
propósitos, solo pocas se relacionaron directamente con los pastos especialmente
aquellas que viven adheridas a la superficie de las hojas y las que se alimentan
directamente de los pastos. En este estudio sólo pocas fueron halladas dentro de esta
categoría, sin embargo otras se encontraron en las praderas posiblemente
aprovechando otros recursos (alimentación, refugio o áreas de crianza). Esta situación
ha sido observada en otros estudios, por lo que se ha propuesto que el aporte de los
pastos marinos a la diversidad de invertebrados no es tan relevante como el aporte a la
productividad y al flujo de energía hacia niveles tróficos superiores (Penagos et al., 2003)
lo que significa que algunas especies presentan por lo general rangos de hábitat muy
amplios y no son exclusivas de este ecosistema (especies raras), no obstante la
presencia de éstas es determinante para los procesos ecológicos que mantienen la
estabilidad del ecosistema y sus recursos.
8.2 Especies dominantes
Los valores de densidad promedio de individuos durante la época de lluvias fueron altos
debido en gran parte al aporte de las especies dominantes consideradas en este trabajo,
en especial de L. variegatus y A. zebra. Como lo muestran los resultados, el erizo blanco
fue la especie más frecuente y con los mayores valores de densidad en el área de
estudio, su presencia tiene un significado ecológico importante ya que su
comportamiento alimenticio es determinante dentro del ciclo de energía y nutrientes en el
ecosistema (Thayer et al., 1984). En el caso de las aguas costeras de la península de La
Guajira que son ricas en nutrientes, la abundancia de algas epifitas es alta por lo que el
papel de los herbívoros que se alimentan de las algas es muy importante para mantener
la productividad de los pastos (Orth, 1992; Rose et al., 1999; Valentine et al., 2000;
Moran y Bjorndal, 2005). Algunos estudios han considerado que L. variegatus a igual que
otros macroherbívoros tiene un papel poco trascendental (Ogden y Zieman, 1977), sin
embargo la mayoría de estos estudios han sido llevados a cabo en las zonas templadas
en donde los herbívoros son poco abundantes mientras que en el Caribe hacen parte de
un gran porcentaje de la fauna que depende directamente de los pastos (Thayer et al.,
1984; Valentine y Heck, 1999). Esta especie representa un canal energético entre los
pastos y los organismos detritívoros que se alimentan de sus desechos metabólicos y
que juegan un papel trascendental en la transferencia de energía a niveles tróficos más
altos (García y Criales, 2005; Moran y Bjorndal, 2005), esto sin contar que puede
controlar la estructura de las algas presentes en las praderas y las poblaciones de otras
especies herbívoras (Ogden, 1980).
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
35
El erizo L. variegatus fue considerada una especie altamente distribuida y muy
abundante, en este aspecto Thayer et al. (1984) aclaran que son pocas las especies
herbívoras que se alimentan directamente de los pastos, sin embargo, su abundancia
suele ser bastante alta. Los valores de densidad están relacionados según Montoya
(2002) con los altos valores de biomasa foliar y longitud de las hojas que se presentan
en esta zona del Caribe colombiano y que representan una fuente de alimento para la
especie, otra explicación puede estar relacionada con la baja abundancia de otras
especies herbívoras, teniendo en cuenta que las praderas de La Guajira en general
están poco influenciadas por ecosistemas adyacentes como manglares y formaciones
coralinas (Montoya, 2002) que representan un aporte de especies (en especial peces
herbívoros) que se mueven hacia las praderas en búsqueda de alimento. Ogden y
Zieman (1977) demostraron que en los puntos de las praderas más cercanos a las
formaciones coralinas se encuentran las hojas con los signos de pastoreo más grandes,
resultantes de poblaciones de peces loro (Scaridae) que forrajean en las praderas y que
a medida que se incrementa la distancia los arrecifes decrece no solo el tamaño sino el
numero de estos signos de pastoreo.
La diferencia en la densidad promedio de L. variegatus entre épocas climáticas se debió
probablemente a un evento de reclutamiento reciente ya que el tamaño de los individuos
observados en la época de lluvias fue mucho menor al tamaño de los individuos
encontrados durante la época seca menor, sin embargo es difícil comprobar esta
situación ya que la época reproductiva de L. variegatus no es la misma en los diferentes
lugares en donde se ha investigado su comportamiento reproductivo. Lo que si se sabe
es que a diferencia de otras especies como Echinometra lucunter y E. viridis, el erizo
blanco tiene una época reproductiva más amplia, que le permite sobrevivir ante
condiciones cambiantes de temperatura y salinidad (Cameron, 1985).
El bivalvo A. zebra fue considerado también como una especie ampliamente distribuida y
abundante en el área de estudio, este es un habitante común de las praderas de
Thalassia (Prieto et al., 2003; Díaz y Liñero-Arana, 2004); sin embargo hasta el momento
no existen estudios relacionados con la productividad biológica de la especie y sobre el
papel que desempeña en las diferentes comunidades donde habita (Prieto et al., 2003).
En el estudio de Prieto et al. (2001) en Venezuela, la abundancia de la especie se
relacionó con el enriquecimiento y fertilidad de la zona que está sometida a la acción
constante de los vientos Alisios, que favorecen el crecimiento y reproducción de la
especie por lo que su éxito en el ecosistema y particularmente en las praderas de La
Guajira está relacionado con el fenómeno de surgencia que caracteriza a esta zona
(Criales, 2004) y que incrementa la productividad del sistema resultando en un hábitat
con grandes recursos en especial para organismos filtradores como los bivalvos
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
36
(Montoya, 2002; Díaz, 2003). En cuanto a las diferencias en la densidad promedio de
individuos entre épocas climáticas, no existe información de la época reproductiva de la
especie, sin embargo Prieto et al. (2001) encontraron una mayor abundancia de la
especie en los meses de octubre y noviembre, lo que coincide con los resultados
presentados en este trabajo.
Adicionalmente, se sabe que esta especie tiene un rápido crecimiento y productividad lo
cual ha permitido soportar una de las mayores pesquerías en el Estado de Sucre,
Venezuela (Prieto et al., 2001; Vera, 1992) por lo que podría clasificarse como una
especie potencialmente explotable que podría llegar a tener un valor comercial
importante en La Guajira.
El erizo C. rosaceus por su parte fue considerada como una especie poco distribuida y
poco abundante si se compara con la frecuencia y abundancia de las dos especies
anteriores, a pesar de esto se considera una especie típica de las praderas que
aparentemente se alimenta de las hojas de los pastos en descomposición y en algunos
casos de segmentos de Halimeda (Hendler et al., 1995; Rützler et al., 2004), también
encuentra refugio en las praderas en donde utiliza las hojas para esconderse de sus
depredadores (Hendler et al., 1995). Esta especie juega un papel importante en la
producción de sedimentos finos ya que puede triturar grandes partículas de sedimento y
convertirlas en partículas finas de arena (Hendler et al., 1995). El gasterópodo V.
muricatum fue reconocido como restringido y poco abundante en el área de estudio a
pesar de ser un habitante común de las praderas (Díaz y Gómez-López, 2003). Esta
especie es carnívora por lo que su presencia en las praderas puede estar relacionada
con la abundancia de pequeños invertebrados como crustáceos que constituyen una
fuente de alimentación (Montoya, 2002).
Las bajas frecuencias y densidades de la mayoría de especies encontradas reflejan el
comportamiento característico de las comunidades en cuanto a la jerarquía de las
abundancias, con pocas especies dominantes y muchas especies raras (Posey et al.,
1998). Las especies poco abundantes fueron en su mayoría crustáceos y pequeños
moluscos que según Montoya (2002) predominan en aquellas praderas con altos valores
de biomasa y longitud de las hojas en donde encuentran refugio y espacios adecuados
para alimentarse (Hooks et al., 1976; Lewis y Stoner, 1983).
Si bien las especies dominantes consideradas en este trabajo revisten una importancia
por su predominio dentro de las comunidades epifaunales, la presencia de otras
especies corrobora el papel de las praderas como hábitats esenciales durante diferentes
etapas de vida de algunos organismos. Durante la época seca menor se registró una alta
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
37
densidad del opistobranquio B. leachii, debido posiblemente a la oferta de alimento que
brindan las praderas o a un evento reproductivo. En cualquiera de los dos casos es
pertinente aclarar que esta especie tiene un alto potencial de crecimiento cuando los
recursos son abundantes además, por ser hermafroditas, pueden aparearse con
cualquier individuo maduro disponible resultando en grandes agregaciones en un
espacio y tiempo determinado (Rudman, 2004). Adicional a esto, se encontraron algunas
masas de huevos de gasterópodos y un gran número de hembras de crustáceos estaban
ovadas al momento de la colecta lo que confirma que las praderas son utilizadas como
refugio durante periodos reproductivos (Puentes y Campos, 1992).
Se concluye que la estructura de la macrofauna epibentónica en las praderas de T.
testudinum en La Guajira está controlada por diferentes factores que varían en espacio y
tiempo por lo cual no es posible definir una sola variable que controle la composición y
abundancia de especies en este ecosistema. Este trabajo prueba algunas variaciones en
la estructura de la fauna de macroinvertebrados en dos épocas climáticas, sin embargo
las diferencias espaciales que caracterizan a las praderas de pastos marinos en áreas
adyacentes (Brook, 1978) pueden ser la explicación más acertada a los cambios
encontrados en este trabajo. Luego de analizar el método de estimación de Jacknife 2
para predecir el porcentaje de especies se hubieran hallado con un muestreo más
intenso, se encontró que durante la época seca menor se muestreo el 62% de las
especies mientras que en la época de lluvias se muestreo el 65,4% de las especies por
lo que las diferencias encontradas entre las dos épocas climáticas también pueden
atribuirse al tipo de muestreo empleado.
Finalmente, aunque el impacto de origen antrópico no es tan evidente en las praderas de
pastos en la región de La Guajira en comparación con otras zonas del Caribe
colombiano (Díaz et al., 2003), el uso extractivo en las praderas ha dado paso no solo a
la reducción de algunos macroinvertebrados, sino también a especies de peces (Gómez-
Canchong et al., 2004; Mejía y Acero, 2002) y tortugas (Ceballos-Fonseca, 2004;
Castaño y Mora, 2002). Es posible que la reducción poblacional de estas especies tenga
consecuencias importantes en las praderas alterando la estructura trófica del sistema y
generando cambios en la distribución y abundancia de otros organismos (Díaz y Gómez-
López 2003; Moran y Bjorndal, 2005; Duffy, 2006). Por esta razón, se considera
necesario implementar medidas que permitan a los pobladores de la zona tener nuevas
alternativas de ingresos de manera que el ecosistema pueda convertirse en un sistema
productivo y sostenible. Por otra parte, se podría proponer como medida de
conservación, la creación de una o varias áreas protegidas que abarquen parte de las
praderas de pastos marinos de La Guajira, como lo propusieron años atrás Díaz et al.,
(2003), lo cual podría garantizar la conservación de este ecosistema y sus recursos.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
38
9. CONCLUSIONES
Las especies de macroinvertebrados epibentónicos encontradas en este trabajo son
similares a las reportadas en otros estudios en diferentes zonas del Caribe colombiano y
del Gran Caribe. Solo una especie (P. pannosus) de las registradas no había sido
registrada en Colombia.
Se presentaron diferencias en densidad de especies asociadas a praderas de T.
testudinum en La Guajira, siendo la época de lluvias (noviembre) la que presentó los
mayores valores de densidad total de individuos.
El índice de Equitatividad fue significativamente mayor en la época seca menor mientras
que los índices de Diversidad y Riqueza no presentaron diferencias significativas entre
épocas climáticas.
La densidad promedio de las especies dominantes, L. variegatus y A. zebra fue
significativamente diferente entre épocas climáticas, registrándose una mayor densidad
durante la época de lluvias. En contraste, la densidad media de C. rosaceus y V.
muricatum no mostró diferencias significativas entre épocas climáticas.
Los cambios en la composición y densidad de la fauna de macroinvertebrados
epibentónicos parece responder en parte a un efecto de la variación estructural de las
praderas debido a los cambios generados durante la época seca menor y la época de
lluvias, sin embargo otros factores relacionados con las variaciones a nivel espacial y no
medidos en este trabajo tiene una influencia importante sobre la estructura de las
comunidades epifaunales.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
39
10. RECOMENDACIONES
Se recomiendan futuros estudios que incluyan información sobre algunos procesos
ecológicos como competencia, depredación, migración, reproducción y reclutamiento,
además de ampliar la información de factores fisicoquímicos como la temperatura,
salinidad, transparencia del agua y profundidad, que aunque fueron medidos en el
presente trabajo, no fueron representativos para detectar posibles relaciones con los
datos biológicos.
Es importante replantear la metodología cuando se hacen comparaciones estacionales,
con el fin de muestrear el mismo transecto y reducir los efectos de la variación espacial,
así mismo es esencial aumentar el número de transectos por zona a fin de tener un
mayor tamaño muestreal y una mayor representatividad.
Asumiendo que las praderas de pastos marinos son un importante ecosistema costero,
como hábitats de alimentación, crianza y refugio de muchas especies de importancia
ecológica y comercial es necesario realizar una caracterización de la pesca artesanal
para evaluar los posibles efectos de esta actividad sobre el ecosistema y la macrofauna
asociada.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
40
11. REFERENCIAS
Álvarez-Guillen, H., M.C. García-Abad., G.J. Villalobos-Zapata y A. Yañez-Arancibia. 1986.
Prospección ictioecológica en la zona de pastos marinos de la Laguna arrecifal en Puerto
Morelos. Quintana Roo. Verano 1984. Anales del Centro de Ciencias del Mar y
Limnología. UNAM.13: 317-336.
Álvarez-León, R., J. Aguilera., C.A. Andrade y P. Nowak. 1995. Caracterización general de la
zona de surgencia en La Guajira colombiana. Revista de la Academia colombiana de
Ciencias Exactas. Físicas y Naturales. 75(19): 679-694.
Angel, I.F. y J. Polania. 2000. Estructura y distribución de los pastos marinos en San Andrés Isla,
Caribe colombiano. Boletín Ecotrópica: Ecosistemas Tropicales No. 35.
Ansari, Z.A., C.U. Rivonker., P. Ramani and A.H. Parulekar. 1991. Seagrass habitat complexity
and macroinvertebrate abundance in Lakshadweep coral ref. lagoons. Arabian Sea.
Coral Reefs. 10: 127-131.
Aubad, M. 1981. Estructura de una pradera de Thalassia testudinum y la fauna de invertebrados
asociada en la Bahía de Neguanje. Parque Nacional Tayrona. Bióloga. Universidad de
Antioquia. Medellín. Colombia. 127 p.
Barrios, L.M y D.I. Gómez-López. 2001. Estado de las praderas de pastos marinos. Informe del
Estado de los Ambientes Marinos y Costeros en Colombia. Instituto de Investigaciones
Marinas y Costeras INVEMAR. Santa Marta. Colombia. 41-51 pp.
Bitter-Soto, R. 1999. Benthic communities associated to Thalassia testudinum
(Hydrocharitaceae) at the localities of Morrocoy National Park. Venezuela. Revista de
Biología Tropical. 47 (3): 443-452.
Borrero, F.J., J.M. Díaz y A. Seczon. 1996. Las ostras perlíferas (Bivalvia: Pteridae) en el Caribe
colombiano. Historia de su explotación, ecología y perspectivas para su
aprovechamiento. Primera Edición. Serie de Publicaciones Especiales. No 1. Instituto de
Investigaciones Marinas y Costeras “José Benito Vives de Andréis” INVEMAR. Santa
Marta. Colombia. 53 p.
Brook, I.M. 1978. Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum)
communities in South Florida characterized by high blade density. Bulletin of Marine
Science. 28 (1): 213-217.
Campos, N.H., M. Falk y B. Werding. 2004. Distribución de los cangrejos de la familia Xanthidae
en el mar Caribe colombiano. 197-210 pp. En: Campos. N.H. y A. Acero (eds).
Contribuciones en Ciencias del Mar en Colombia. Investigación y desarrollo de territorios
promisorios. Bogota. Colombia.
Cancemi, G., M.C. Buia and L. Mazzella. Structure and growth dynamics of Cymodocea nodosa
meadows. Scientia Marina. 66 (4): 365-373.
Castaño-Mora, O.V. 2002. Libro rojo de reptiles de Colombia. Libros rojos de especies
amenazadas de Colombia. Instituto de Ciencias Naturales-Universidad Nacional de
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
41
Colombia, Ministerio del Medio Ambiente, Conservación Internacional-Colombia. Bogota,
Colombia. 160 p.
Castillo-Torres, P. 2002. Caracterización estructural y evaluación del estado de las praderas de
pastos marinos del Caribe colombiano. Bióloga Marina. Universidad Jorge Tadeo
Lozano. Facultad de Biología Marina. Área de Ingeniería y Recursos Naturales. Santa
Marta. Colombia. 81 p.
Ceballos-Fonseca, C. 2004. Distribución de las playas de anidación y áreas de alimentación de
tortugas marinas y sus amenazas en el Caribe colombiano. Boletín de Investigaciones
Marinas y Costeras. 33. 79-99
Chace, F.A. 1972. The shrimps of the Smithsonian-Bredin Caribbean expeditions with a summary
of the West Indian Shallow-water species (Crustacea:Decapoda:Natantia). Smithsonian
Contributions to Zoology. Smithsonian Institution Press. Washington. 98-180 p.
Coen, L.D., K.L. Heck and L.G. Abele. 1981. Experiments on competition and predation among
shrimps of seagrass meadows. Ecology. 62 (6): 1484-1493.
Corredor, J .E. 1992. Dinámica espacio-temporal de la surgencia de La Guajira. Memorias del
VIII Seminario Nacional de Ciencias y Tecnologías del Mar. Congreso Centroamericano
y del Caribe de Ciencias del Mar. Santa Marta. Colombia. 206-221 pp.
Criales, M.I. 2004. Flujos de energía en el sistema de surgencia tropical en la Península de La
Guajira. Caribe colombiano. Tesis (Magíster Scientiae en Biología Marina). Universidad
Nacional de Colombia. Instituto de Investigaciones Marinas y Costeras “José Benito
Vives de Andréis” INVEMAR. Santa Marta. Colombia. 117 p.
Cruz-Ábrego, F.M., P. Hernández-Alcántara y V. Solis-Weiss. 1994. Estudios de la fauna de
poliquetos (Annelida) y moluscos (Gastropoda y Bivalvia) asociada con ambientes de
pastos marinos (Thalassia testudinum) y manglares (Rhizophora mangle) en la Laguna
de Términos. Campeche. México. Anales del Instituto de Ciencias del Mar y Limnología
Universidad Nacional Autónoma de México. 21: 1-13.
Daniel, O. 1998. Subsidios al uso del índice de Diversidad de Shannon. Congreso
Latinoamericano IUFRO (International Union of Forest Research Organizations).
Valdivia. Chile. Tema 3. CD-ROM.
Díaz, J.M., L.M. Barrios y D.I. Gómez-López. 2003. Las praderas de pastos marinos en
Colombia. Estructura y distribución de un ecosistema estratégico. Instituto de
Investigaciones Marinas y Costeras “José Benito Vives de Andréis” INVEMAR. Serie de
publicaciones especiales – No. 10. Santa Marta. Colombia. 159 p.
__________. 2003. Diversidad de moluscos en una pradera de Thalassia testudinum en el
Parque Nacional Natural Tayrona. 11-114 pp. En: Díaz, J.M., L.M. Barrios y Gómez-
López D.I (eds). 2003 Las praderas de pastos marinos en Colombia. Estructura y
distribución de un ecosistema estratégico. Instituto de Investigaciones Marinas y
Costeras “José Benito Vives de Andréis” INVEMAR. Serie de publicaciones especiales –
No. 10. Santa Marta. Colombia. 159 p.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
42
__________ y D.I. Gómez-López. 2003. Cambios históricos en la distribución y abundancia de
praderas de pastos marinos en la Bahía de Cartagena y áreas aledañas (Colombia).
Boletín de Investigaciones Marinas y Costeras. 32: 57-74.
__________ y J. Garzón-Ferreira. 2000. Estado de las praderas de pastos marinos en Colombia.
Año 2000. Informe del Estado de los Ambientes Marinos y Costeros en Colombia.
Instituto de Investigaciones Marinas y Costeras INVEMAR. Santa Marta. Colombia. 39-47
pp
__________ y M. Puyana. 1994. Moluscos del Caribe colombiano. Un catalogo ilustrado. Primera
edición. Editorial presencia. Bogota. Colombia. 329 p.
__________. 1991. Perfil Ambiental del Caribe colombiano. Ecosistemas Litorales del Caribe
colombiano. 1-34 pp. En: Consejo Regional de Planificación de la Costa Atlántica
(CORPES). Perfil Ambiental del Caribe colombiano. Documentos de Trabajo. Tomo No 1.
Santa Marta. Colombia.
Díaz, O. e I. Liñero-Arana. 2004. Comunidad de moluscos asociados a praderas de Thalassia
testudinum (Bank Et König 1805). en la Bahía de Mochima. Venezuela. Acta Científica
Venezolana. 55: 44-55.
Doering, V. and D. Bone. 1983. Composición de la fauna asociada a la pradera de Thalassia
testudinum de Punta Moron. 115-131 pp. En: Penchaszaden, P.E (ed). 1983 Ecología del
ambiente marino costero de Punta Moron (Termoeléctrica planta centro. Estado
Carabobo. Venezuela). Informe final segunda fase del proyecto CADAFE-USB.
Universidad Simón Bolívar. Instituto de Tecnología y Ciencias Marinas INTECMAR. 463
p.
Duffy, J.E. 2006. Biodiversity and the functioning of seagrass ecosystem. Marine Ecology
Progress Series. 311: 233-250.
Durako, M.J. 1988. Turtle grass (Thalassia testudinum Banks ex König) – A seagrass. En:
BAJAJ. Y.P.S. (ed). 1988. Biotechnology in Agriculture and Forestry. Vol 6 Crops II.
Springer-Verlag. New York. U.S.A. 504-520 pp.
Digby, P.G.N. and R.A. Kempton. 1987. Multivariate analysis of ecological communities.
Population and Community Biology Series. Primera Edición. Chapman & Hall. Londres.
206 p.
Echeverry, B.E. 1982. Estudio de la fauna de invertebrados de una pradera de Thalassia
testudinum Konig. - Biomasa e incidencia de algunos factores ambientales sobre el
ecosistema en la Bahía de Neguanje Parque Nacional Tayrona. Bióloga. Universidad de
Antioquia. Medellín. Colombia. 103 p.
English, S., C. Wilkinson and V. Beker. 1997. Survey manual for tropical marine resourses.
Segunda edición. Australian Institute of Marine Science. Townsville. 368 p.
Fernández-Torquemada, Y. y J.L. Sánchez-Lizaso. 2003. Efecto de un posible interacción entre
el pH y la salinidad sobre el crecimiento de Posidonia oceanica (L.) Delile. 1813. Boletín
del Instituto Español de Oceanografía. 19 (1-4): 247-252.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
43
Field, J., K. Clarke and R. Warwick. 1982. A practical strategy for analyzing multispecies
distribution pattern. Marine Ecology Progress Series. 8:37-52
Franke, R. 2001. Evaluación de las comunidades epifaunales de las praderas de Thalassia
testudinum en el Parque Nacional Natural Tayrona. Tesis de Maestría. Bióloga Marina.
Universidad Nacional de Colombia. Facultad de Ciencias. Santa Marta. Colombia. 118 p.
García, C. y M.I. Criales. 2005. Primera evaluación de la respiración de Lytechinus variegatus
(Echinodermata: Echinoidea) en la Bahía de Chengue. Parque Nacional Natural. Caribe
Colombiano. Revista de la Academia Colombiana de Ciencias Exactas. Físicas y
Naturales. 19 (110): 85-88.
García-Hernández, V.D. 2003. Influencia de la complejidad estructural de Thalassia testudinum
sobre la comunidad íctica juvenil en Laguna de Yalahau. Quintana Roo. Maestro en
Ciencias en la especialidad de Biología Marina. Centro de Investigación y de Estudios
avanzados del Instituto Politécnico Nacional. Mérida. Yucatán. México. 146 p.
Garzón-Urbina, P.A. Caracterización estructural y funcional de las praderas monoespecíficas de
Thalassia testudinum (Banks ex könig. 1805) en el departamento de la Guajira. Caribe
colombiano. (Trabajo de Grado en preparación). Universidad Jorge Tadeo Lozano.
Facultad de Biología Marina. Santa Marta. Colombia.
Garzón-Ferreira, J. y A. Rodríguez-Ramírez. 2003. Diez años de monitoreo en una pradera de
Thalassia.71-74 pp. En: Díaz, J.M., L.M. Barrios y D.I. Gómez-López (eds). 2003. Las
praderas de pastos marinos en Colombia. Estructura y distribución de un ecosistema
estratégico. Instituto de Investigaciones Marinas y Costeras “José Benito Vives de
Andréis” INVEMAR. Serie de publicaciones especiales – No. 10. Santa Marta. Colombia.
159 p.
Gómez-López, D.I., A. Rodríguez y A. Jauregui. 2005. Estado de los pastos marinos en
Colombia. 115-127 pp. En: Informe del Estado de los ambientes marinos y costeros en
Colombia: año 2004. INVEMAR Serie de Publicaciones periódicas No. 8. 214 p.
Gómez-Canchong, P., L. Manjarrés., L.O. Duarte y J. Altamar. 2004. Atlas pesquero del área
norte del Mar Caribe de Colombia. Editorial Gente Nueva Ltda. Universidad del
Magdalena. Santa Marta. Colombia. 230 p.
Greenway, M. 1995. Trophic relationships of macrofauna within a Jamaican seagrass meadow
and the role of the echinoid Lytechinus variegatus (Lamarck). Bulletin of Marie Science.
56 (3): 719-736.
Heck, K.L. and R.J. Orth. 1980. Seagrass habitats: The role of habitat complexity. competition
and predation in structuring associated fish and motile macroinvertebrate assemblages
449-464 pp. En: KENNEDY, V.S. 1980. Estuarine Perspectives. Academic Press. Nueva
York. Estados Unidos. 533 p.
Heck, K.L. 1979. Some determinants of the composition and abundance of motile
macroinvertebrate species in tropical and temperate turtlegrass (Thalassia testudinum)
meadows. Journal of Biogeography. (6): 183-200.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
44
__________. 1977. Comparative species richness. composition. and abundance of invertebrates
in Caribbean seagrass (Thalassia testudinum) meadows (Panamá). Marine Biology. 41:
335-348.
Hendler, G., J. Miller., D. Pawson and M. Porter. 1995. Echinoderms of Florida and the
Caribbean Sea Stars. Sea Urchins and Allies. Smithsonian Institution Press. Smithsonian
Institution. Washington D.C. 390 p.
Holthuis, L.B. 1952. A general revision of the Palaemonidae (Crustacea. Decapada. Natantia) of
the Americas II. The subfamily Palaemoninae. Allan Hancock Foundation Occasional
Papers. 11(1951) 1-332
Hooks, T.A., K.L. Heck and R.J. Livingston. 1976. An inshore marine invertebrate community:
Structure and habitat associations in the Northeastern Gulf of Mexico. Bulletin of Marine
Science. 26 (1): 99-109.
InfoStat. 2004. InfoStat, versión 2004. Manual del Usuario. Grupo InfoStat, FCA, Universidad
Nacional de Córdoba. Primera Edición, Editorial Brujas. Argentina. 314 p.
IGAC- Instituto Geográfico Agustín Codazzi Subdirección de Investigación y divulgación
geográfica. 1986. La Guajira: Aspectos geográficos. Sección imprenta y ediciones IGAC.
Bogotá. Colombia. 151 p.
INVEMAR. Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”.
2003. Programa holandés de asistencia para estudios en cambio climático: Colombia.
Definición de la vulnerabilidad de los sistemas bio-geofísicos y socioeconómicos debido
a un cambio en el nivel del mar en la zona costera colombiana (Caribe continental.
Caribe insular. y Pacífico) y medidas para su adaptación Informe Técnico No. 2. Tomo
VII. Santa Marta. Colombia. 530 p.
__________. 2000. Programa Nacional de Investigación en Biodiversidad Marina y Costera. Plan
de Acción 2001-2010. DÍAZ, J.M. y D.I. GÓMEZ-LÓPEZ (eds). INVEMAR, FONADE,
MMA. ISBN: 958-96972-0-8. 80 p.
__________. 1988. Diagnóstico actual de las comunidades marinas de la bahía Portete. análisis
de efectos reales por la construcción y operación de las instalaciones portuarias. Editado
por J.M. Díaz. O.D. Solano. R. Dueñas y J. Garzón – Ferreira. CARBOCOL. INTERCOL.
El Cerrejón – Zona Norte. Informe final. Santa Marta. Colombia. 156 p.
ITIS. Integrated Taxonomic Information System. 2005. [en línea].
http://www.itis.usda.gov/itis/index.html [Consulta: febrero, 2006]
Jiménez, M. 1994. Comunidad de moluscos asociada a Thalassia testudinum en la ensenada de
Reyes. Bahía de Mochima. Edo. Sucre. Venezuela. Boletín del Instituto Oceanográfico
de Venezuela. Universidad de Oriente. 33 (1 y 2): 67-76.
Kikuchi, T. 1980. Faunal relationships in Temperate seagrass beds.153-172 pp. En: Phillips, R.C.
y C.P. McRoy. Handbook of seagrass biology: An ecosystem perspective. Garland
Publishing. Inc. New York. U.S.A. 353p.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
45
Klumpp, D. 2005. (Abstract) Composition and abundance of benthic macrofauna of tropical sea-
grass bed in North Queensland, Australia. Pacific Science. 59-4:541-560.
Knowles, L.L. and S.S. Bell. 1998. The influence of habitat structure in faunal-habitat associations
in a Tampa Bay seagrass system. Florida. Bulletin of Marine Science. 62 (3): 781-794.
Krebs, C.J. 1985. Ecología. Estudio de la distribución y abundancia. Segunda Edición. Editorial
Karla. Mexico. D.F. 753 p.
Livingston, R.J. 1984. Trophic response of fishes to habitat variability in coastal seagrass
systems. Ecology. 65 (4): 1258-1275.
Lemaitre, R. 1981. Shallow-water crabs (Decapoda. Brachyura) collected en the southern
Caribbean near Cartagena. Colombia. Bulletin of Marine Science. 31 (2): 234-266.
Lewis, F.G. and A.W. Stoner. 1983. Distribution of macrofauna within seagrass beds: An
explanation for patterns of abundance. Bulletin of Marine Science. 33 (2): 296-304.
Mejía, L.S. y A. Acero. 2002. Libro rojo de peces marinos de Colombia. INVEMAR, Instituto de
Ciencias Naturales- Universidad Nacional de Colombia, Ministerio del Medio Ambiente.
La serie de Libros Rojos de Especies Amenazadas de Colombia. Bogota, Colombia. 173
p.
(MMA) Ministerio del Medio Ambiente. 2001. Política Nacional Ambiental para el Desarrollo
Sostenible de los Espacios Oceánicos y las Zonas Costeras e Insulares de Colombia
PNAOCI. INVEMAR - Ministerio del Medio Ambiente. ISBN:958969721-6. 96 p.
Montoya, P. 2002. Evaluación de la macrofauna epibentónica asociada a praderas de Thalassia
testudinum (Banks ex König) en el Caribe colombiano. Biólogo Marino. Universidad
Jorge Tadeo Lozano. Facultad de Biología Marina. Área de Ingeniería y Recursos
Naturales. Santa Marta. Colombia. 123 p.
Moran, K.L. and K.A. Bjorndal. 2005. Simulated green turtle grazing affects structure and
productivity of seagrass pastures. Marine Ecology Progress Series. 305: 235-247.
Nakaoka, M., Toyohara, T. and Matsumasa, M. 2001. Seasonal and between-substrate variation
in mobile epifaunal community in a multispecific seagrass bed of Otsuchi Bay, Japan.
Marine Ecology. 22: 379-395.
Ogden, R.J. 1980. Faunal relationships in Caribbean seagrass beds. 173-198 pp. En: PHILLIPS,
R.C. and McROY. C.P (eds). 1980 Handbook of seagrass biology: An ecosystem
perspective. Garland Publishing. Inc. New Cork. U.S.A. 353p
__________ and J.C. Zieman. 1977. Ecological aspects of coral reef-seagrass bed contacts in
the Caribbean. Proceeding. Third International Coral Reef Symposium. 377-382 p.
Orth, R.J. 1992. A perspective on plant-animal interactions in seagrasses: physical and biological
determinants influencing plant and animal abundance 147-164 pp. En: Jhon, D.M., S.J.
Hawkins and J.M. Price (eds). 1992. Plant-animal interactions in the marine benthos.
Systematics association special volumes series 46. Oxford University Press. 592 p.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
46
__________., K. Heck and J. Montfrans. 1984. Faunal communities in seagrass beds: A review
of the influence of plant structure and prey characteristics on predator-prey relationships.
Estuaries. 7 (4A): 339-350.
Pantaleón-Lizarazu, A. y D. Rodríguez-Gacha. 2004. Primer registro de Ruppia maritima en el
Caribe colombiano. Pág. 124. En: Informe del Estado de los Ambientes Marinos y
Costeros en Colombia. 2004. INVEMAR. Serie de Publicaciones Periódicas No 8. 210 p.
Penagos, G., J. Palacio y N. Aguirre. 2003. Macroepifauna móvil de una pradera de pasto marino
en Punta San Bernardo – Norte del Golfo de Morrosquillo. 102-105 pp. En: Díaz, J.M.,
Barrios. L.M y Gómez-López. D.I. 2003 Las praderas de pastos marinos en Colombia.
Estructura y distribución de un ecosistema estratégico. Instituto de Investigaciones
Marinas y Costeras “José Benito Vives de Andréis” INVEMAR. Serie de publicaciones
especiales – No. 10. Santa Marta. Colombia. 159 p.
Peña, T.A. y M. Jiménez-Prieto. 2001. Comunidad de peces en tres praderas de Thalassia
testudinum del Golfo de Cariaco. estado de Sucre. Venezuela. Boletín del Instituto
Oceanográfico de Venezuela. Universidad de Oriente. 40 (1y 2): 39-48.
Posey, M.H., T.D. Alphin., S. Banner., F. Vose and W. Linberg. 1998. Temporal variability.
diversity and guild structure of a benthic community in the Northeastern Gulf of Mexico.
Bulletin of Marine Science. 63 (1): 143-155.
Prieto, A.S., S. Sant., E. Méndez y C. Lodeiros. 2003. Diversidad y abundancia de moluscos en
las praderas de Thalassia testudinum de la Bahía de Mochima. Parque Nacional
Mochima. Venezuela. Revista de Biología Tropical. 51 (2): 413-426 p.
__________., L.J. Ruiz. N. García y M. Álvarez. 2001. Diversidad malacológica en una
comunidad de Arca zebra (Mollusca: Bivalvia) en Chacopata. Estado de Sucre.
Venezuela. Revista de Biología Tropical. 49 (2): 591-598.
Puentes, L.G. y N.H. Campos. 1992. Los camarones (Crustácea: Decapoda: Natantia) asociados
a praderas de thalassia testudinum Banks ex Koning. en la región de Santa Marta.
Caribe colombiano. Caldasia. 17 (1): 121-132.
Rathbun, M.J. 1930. The Cancroid Crabs of America of the families Euryalidae. Portunidae.
Atelecyclidae. Cancridae and Xanthidae. Smithsonian Institution Press. United States
National Museum Bulletin. Washington. 152-609 pp.
__________.1925. The spider crabs of America. Smithsonian Institution Press. United States
National Museum Bulletin. Washington. 379 p.
Rios, E. 1994. Seashells of Brazil. Segunda edición. Editora da Furg. Rió Grande. Brazil. 425 p.
Rodríguez, G. 1980. Crustáceos decápodos de Venezuela. Instituto Venezolano de
Investigaciones Científicas. Caracas. Venezuela. 494 p.
Rodríguez, C. y E. Villamizar. 2000. Fauna bentónica asociada a una pradera de Thalassia
testudinum (Hydrocharitaceae) en el Parque Nacional Morrocoy. Venezuela. Revista de
Biología Tropical. 1: 243-249.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
47
Rose, C.D., W.C. Sharp., W.J. Kenworthy., J.H. Hunt., W.G. Lyons., E.J. Prager., J.F. Valentine.,
M.O. Hall., P.E. Whitfield., and J.W. Fourqurean. 1999. Overgrazing of a large seagrass
bed by the sea urchin Lytechinus variegatus in the Outer Florida Bay. Marine Ecology
Progress Series. 190: 211-222.
Rudman, W.B. 2004. Population fluctuations [en línea] Sea Slug Forum. Australian Museum.
Sydney. http://www.seaslugforum.net/factsheet.cfm?base=popfluc [Consulta: abril. 2006]
Rützler, K., I. Goodbody., M.C. Díaz., I. Feller and I.G. Macintyre. 2004. The aquatic environment
of Twin Cays. Belize. Atoll Research Bulletin. 512: 1-48.
Short, F.T. and Coles. R.G. 2001. Global Seagrass Research Methods. Primera edición. Editorial
Elsevier. Amsterdam. Holanda. 473 p.
SAS INSTITUTE. 1996. SAS/STAT guide for personal computers. 6th edition. SAS Institute. Cary.
NC. 429 p.
Solana-Arellano, E. Utilización de métodos cuantitativos para el estudio de la dinámica de los
pastos marinos: Una revisión crítica. Revista de Biología Marina y Oceanógrafa. 36 (2):
165-180.
Stoner, A.W. 2003. What constitutes essential nursery habitat for a marine species? A case of
habitat form and function for queen conch. Marine Ecology Progress Series 257: 275-
289.
__________. 1980. The role of seagrass biomass in the organization of benthic macrofaunal
assemblages. Bulletin of Marine Science. 30 (3): 537-551.
Thayer, G.W., K.A. Bjorndal., J.C. Ogden., S.L. Williams and J.C. Zieman. 1984. Role of larger
herbivores in seagrass communities. Estuaries. 7 (4A): 351-376.
Valentine, J.H., K.L. Heck., K.D. Kirsch., and D. Webb. 2000. Role of sea urchin Lytechinus
variegatus grazing in regulating subtropical turtlegrass Thalassia testudinum meadows in
the Florida Keys (USA). Marine Ecology Progress Series. 200: 213-228.
__________ and K.L. Heck. 1999. Seagrass herbivory: evidence for the continued grazing of
marine grasses. Marine Ecology Progress Series. 176: 291-302.
Vera, B. 1992. Seagrasses of the Venezuelan Coast: Distribution and community components.
135-140 pp. En: Seeliger, U. 1992. Coastal plant communities of Latin America.
Academic Press. San Diego. California. U.S.A. 392 p.
Virnstein, R.W. 1987. Seagrass-associate invertebrate communities of the Southestern U.S.A: A
review. En: Durako, M.J., Phillips. R.C., and Lewis. R.R. Florida Marine Research
Publications. Proceedings of the symposium on Subtropical-Tropical seagrasses of the
Southeastern United States. Florida Department of Natural Resources. Bureau of Marine
Science. St. Petersburg. Florida. U.S.A. 209 p.
__________., W.G. Nelson. F.G. Lewis and R.K. Howard. 1984. Latitudinal patterns in seagrass
epifauna: Do patterns exist. and can they be explained? Estuaries. 7 (4A): 310-330.
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
48
__________., P. Mikkelsen., K. Cairns and M. Capone. 1983. Seagrass beds versus sand
bottoms: the trophic importance of their associated benthic invertebrates. Florida
Scientist. 46: 363-381.
Williams, A.B. 1984. Shrimps. lobsters and crabs of the Atlantic coast of the eastern United
States. Maine to Florida. Smithsonian Institution Press. Washington. U.S.A. 550 p.
Yáñez-Arancibia, A. y R.S. Nugent. 1988. El papel ecológico de los peces en los estuarios y
lagunas costeras. Anales de Ciencias del Mar y Limnología. Universidad Nacional
Autónoma de México. 4 (1): 107-114.
Young, D.K. and M.W. Young. 1982. Macrobenthic invertebrates in Bare Sand and seagrass
(Thalassia testudinum) at Carrie Bow Cay. Belize. En: Rützler. K. y G. Macintyre. The
Atlantic Barrier Reef Ecosystem al Carrie Bow Cay. Belize. 1: Structure and
Communities. Smithsonian Contribution to Marine Sciences (12): 115-126.
Zea, S. 1998. Estado actual del conocimiento en sistemática de esponjas marinas (Porifera) del
Caribe colombiano. Boletín Ecotrópica: Ecosistemas Tropicales. 33: 45-59.
PDF created with pdfFactory trial version www.pdffactory.com
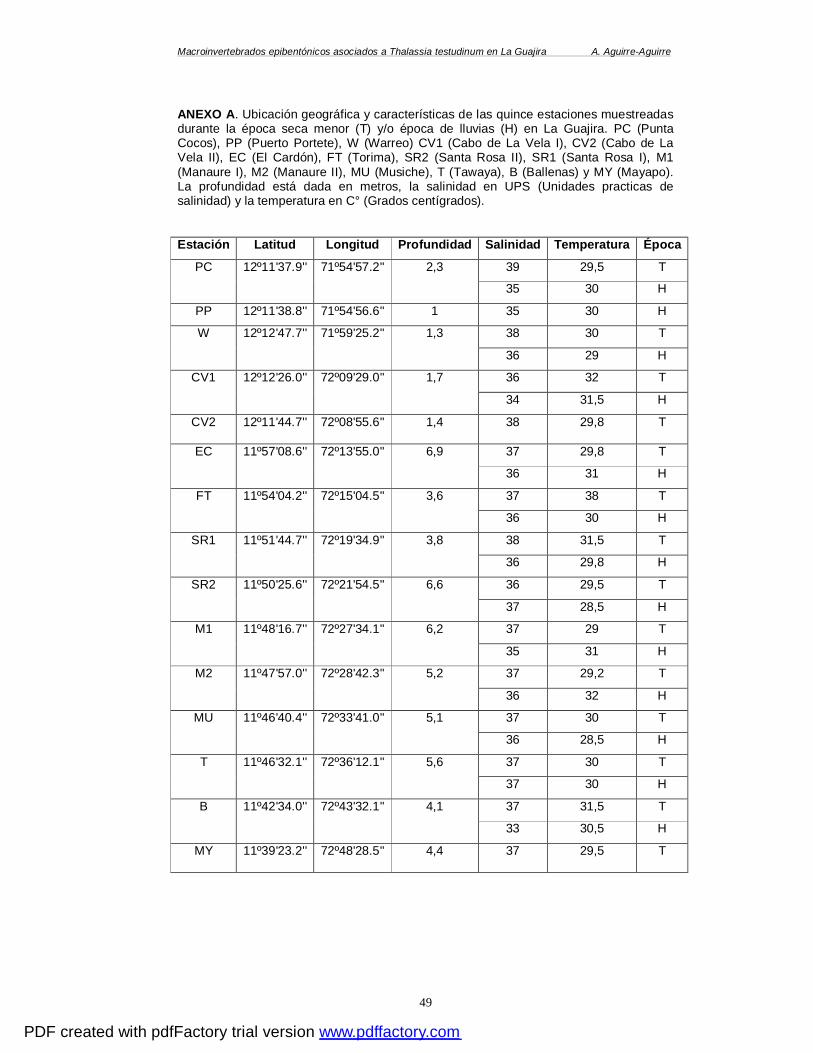
Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
49
ANEXO A. Ubicación geográfica y características de las quince estaciones muestreadas durante la época seca menor (T) y/o época de lluvias (H) en La Guajira. PC (Punta Cocos), PP (Puerto Portete), W (Warreo) CV1 (Cabo de La Vela I), CV2 (Cabo de La Vela II), EC (El Cardón), FT (Torima), SR2 (Santa Rosa II), SR1 (Santa Rosa I), M1 (Manaure I), M2 (Manaure II), MU (Musiche), T (Tawaya), B (Ballenas) y MY (Mayapo). La profundidad está dada en metros, la salinidad en UPS (Unidades practicas de salinidad) y la temperatura en C° (Grados centígrados). Estación Latitud Longitud Profundidad Salinidad Temperatura Época
39 29,5 T PC
12º11'37.9'' 71º54'57.2'' 2,3
35 30 H
PP 12º11'38.8'' 71º54'56.6'' 1 35 30 H
38 30 T W 12º12'47.7'' 71º59'25.2'' 1,3
36 29 H
36 32 T CV1 12º12'26.0'' 72º09'29.0'' 1,7
34 31,5 H
CV2 12º11'44.7''
72º08'55.6''
1,4 38 29,8 T
37 29,8 T EC 11º57'08.6''
72º13'55.0''
6,9
36 31 H
37 38 T FT 11º54'04.2''
72º15'04.5''
3,6
36 30 H
38 31,5 T SR1 11º51'44.7''
72º19'34.9''
3,8
36 29,8 H
36 29,5 T SR2 11º50'25.6''
72º21'54.5''
6,6
37 28,5 H
37 29 T M1 11º48'16.7''
72º27'34.1''
6,2
35 31 H
37 29,2 T M2 11º47'57.0''
72º28'42.3''
5,2
36 32 H
37 30 T MU 11º46'40.4''
72º33'41.0''
5,1
36 28,5 H
37 30 T T 11º46'32.1''
72º36'12.1''
5,6
37 30 H
37 31,5 T B 11º42'34.0''
72º43'32.1''
4,1
33 30,5 H
MY 11º39'23.2''
72º48'28.5''
4,4 37 29,5 T
PDF created with pdfFactory trial version www.pdffactory.com
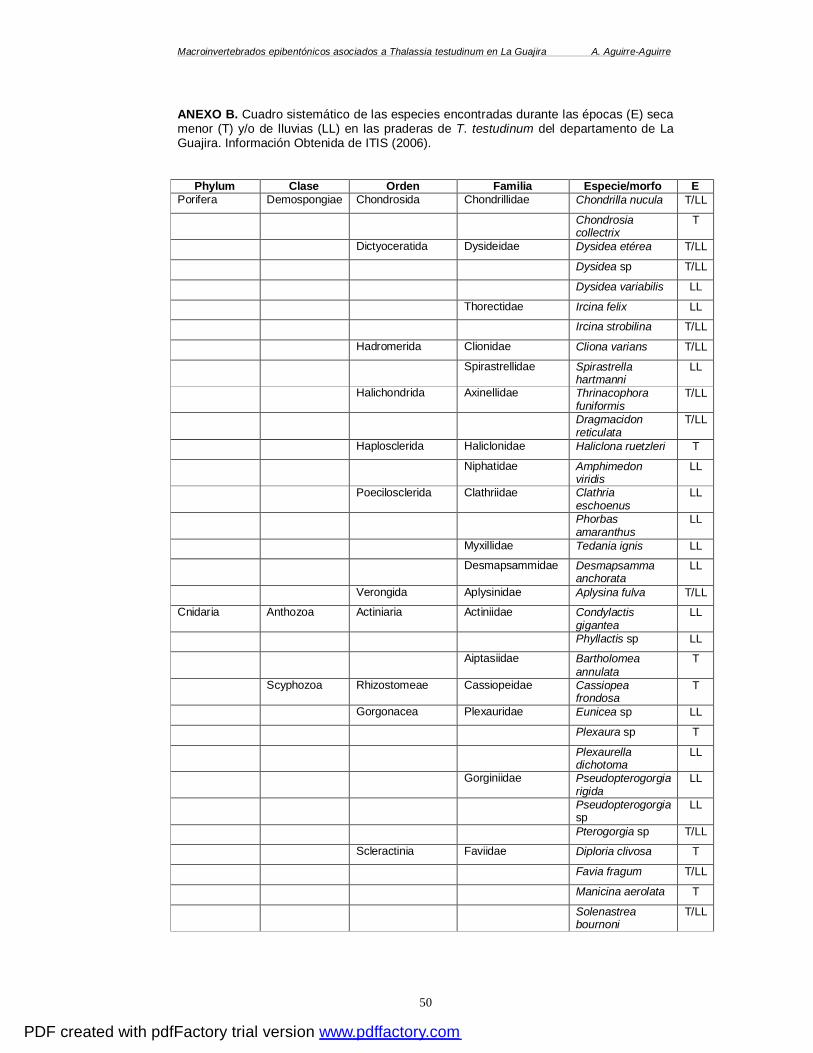
Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
50
ANEXO B. Cuadro sistemático de las especies encontradas durante las épocas (E) seca menor (T) y/o de lluvias (LL) en las praderas de T. testudinum del departamento de La Guajira. Información Obtenida de ITIS (2006).
Phylum Clase Orden Familia Especie/morfo E Porifera Demospongiae Chondrosida Chondrillidae Chondrilla nucula T/LL
Chondrosia collectrix
T
Dictyoceratida Dysideidae Dysidea etérea T/LL
Dysidea sp T/LL
Dysidea variabilis LL
Thorectidae Ircina felix LL
Ircina strobilina T/LL
Hadromerida Clionidae Cliona varians T/LL
Spirastrellidae Spirastrella hartmanni
LL
Halichondrida Axinellidae Thrinacophora funiformis
T/LL
Dragmacidon reticulata
T/LL
Haplosclerida Haliclonidae Haliclona ruetzleri T
Niphatidae Amphimedon viridis
LL
Poecilosclerida Clathriidae Clathria eschoenus
LL
Phorbas amaranthus
LL
Myxillidae Tedania ignis LL
Desmapsammidae Desmapsamma anchorata
LL
Verongida Aplysinidae Aplysina fulva T/LL
Cnidaria Anthozoa Actiniaria Actiniidae Condylactis gigantea
LL
Phyllactis sp LL
Aiptasiidae Bartholomea annulata
T
Scyphozoa Rhizostomeae Cassiopeidae Cassiopea frondosa
T
Gorgonacea Plexauridae Eunicea sp LL
Plexaura sp T
Plexaurella dichotoma
LL
Gorginiidae Pseudopterogorgia rigida
LL
Pseudopterogorgia sp
LL
Pterogorgia sp T/LL
Scleractinia Faviidae Diploria clivosa T
Favia fragum T/LL
Manicina aerolata T
Solenastrea bournoni
T/LL
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
51
Solenastrea hyades
T
Meandrinidae Dichocoenia stokesi
T/LL
Oculinidae Oculina difusa T
Poritidae Porites astreoides T/LL
Porite porites T/LL
Siderastreidae Siderastrea radians
T
Siderastrea siderea
T/LL
Zoanthidea Epizoanthidae Isaurus duchassaingi
LL
Hydrozoa Hydroida Plumuraliidae Hidroide sp 1 T/LL
Milleporina Milleporidae Millepora alcicornis
T/LL
Annelida Polichaeta Canalipalpata Artacaminae Eupolymnia sp T/LL
Amphinomidae Hermodice carunculata
T/LL
Mollusca Bivalvia Arcoida Arcidae Anadara baughmani
T
Anadara notabilis T/LL
Arca zebra T/LL
Mytiloida Mytilidae Atrina seminuda LL
Brachidontes modiolus
T/LL
Pinnidae Pinna carnea T/LL
Pterioida Pteriidae Pinctada imbricata LL
Pteria colymbus T/LL
Veneroida Chamidae Chama macerophylla
T/LL
Chama sp T
Gastropoda Achaeogastropoda Turbiniidae Astraea phoebia LL
Astraea tecta tecta T/LL
Mesogastropoda Turritellidae Turritella variegata T
Cassidae Cassis madagascariensis
T
Cerithidae Cerithium atratum T/LL
Cerithium eburneum
T/LL
Cerithium sp T/LL
Crepidulidae Crepidula plana T
Modulidae Modulus modulus T/LL
Strombidae Strombus raninus T/LL
Tonnidae Tonna galea LL
Calyptraeidae Crucibulum auriculata
LL
Crucibulum planum
T
Fasciolariidae Fasciolaria tulipa LL
Turridae Hindsiclava chazaliei
T
PDF created with pdfFactory trial version www.pdffactory.com

Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
52
Neogastropoda Muricidae Murex messorius LL
Phyllonotus margaritensis
T
Nassariidae Melongena melongena
T
Turbinellidae Turbinella angulata
LL
Vasum muricatum T/LL
Conidae Conus amphiurgus T
Conus spurius spurius
T
Anaspidea Notarchidae Bursatella leachii T
Nudibranchia Chromodorididae Hypselodoris bayeri
T
Crustacea Malacostraca Decapoda Alpheidae Alpheus normanii T
Alpheus sp T
Diogenidae Calcinus tibicen LL
Dardanus fucosus T/LL
Petrochirus diogenes
T
Portunidae Callinectes sapidus
LL
Mithracidae Macrocoeloma trispinosum
LL
Mycrophrys bicornutus
T
Mithraculus forceps
T/LL
Inachidae Micropanope sp LL
Paguridae Paguristes werdingi
T
Palaemonidae Panulirus argus T
Dromiidae Cryptodromiopsis antillensis
LL
Stomatopoda Gonodactylidae Gonodactylus bredini
LL
Porcellanidae Petrolisthes galathinus
T/LL
Xanthidae Pilumnus caribaeus
LL
Pilumnus pannosus
T/LL
Ophthalmiidae Pitho lherminieri T
Stenopodidae Stenopus hispidus H
Echinodermata Ophiuroidea Phrynophiurida Gorgonocephalidae Astrophyton muricatum
T/LL
Echinoidea Clypeasteroida Clypeasteridae Clypeaster rosaceus
T/LL
Toxopneustidae Lytechinus variegatus
T/LL
Tripneustes ventricosus
LL
Asteroida Valvatida Cidaridae Eucidaris tribuloides
LL
Oreasteridae Oreaster reticulatus
T/LL
PDF created with pdfFactory trial version www.pdffactory.com
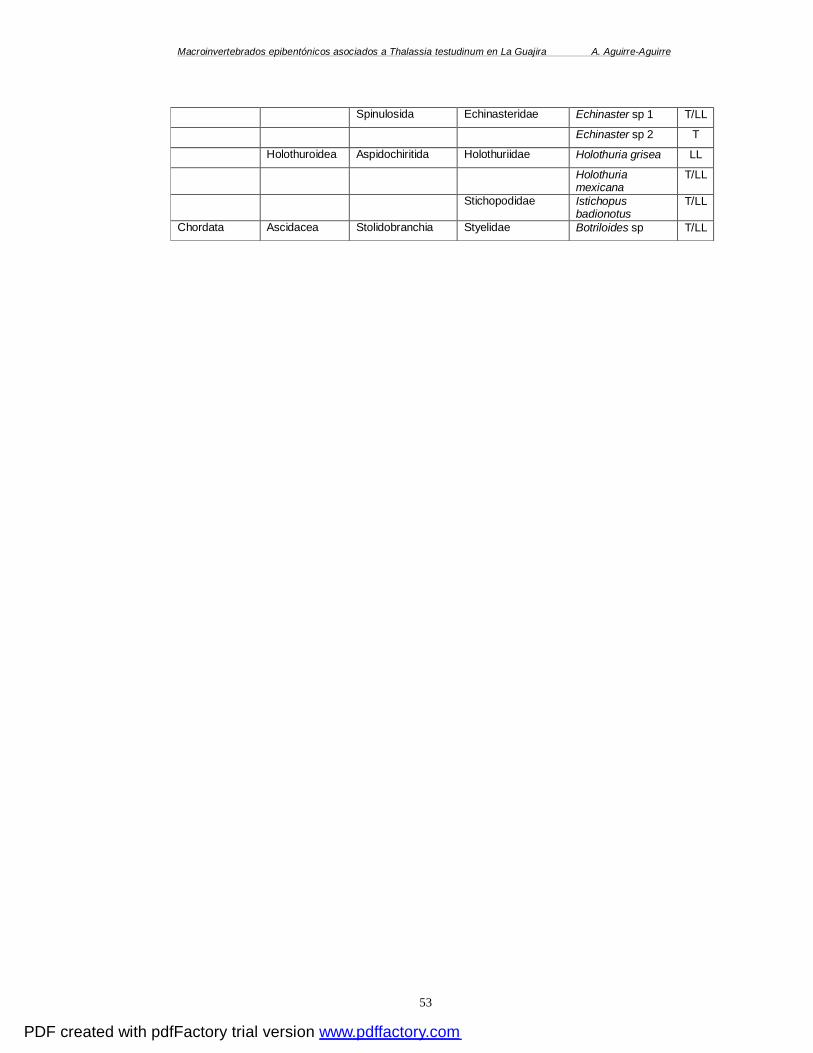
Macroinvertebrados epibentónicos asociados a Thalassia testudinum en La Guajira A. Aguirre-Aguirre
53
Spinulosida Echinasteridae Echinaster sp 1 T/LL
Echinaster sp 2 T
Holothuroidea Aspidochiritida Holothuriidae Holothuria grisea LL
Holothuria mexicana
T/LL
Stichopodidae Istichopus badionotus
T/LL
Chordata Ascidacea Stolidobranchia Styelidae Botriloides sp T/LL
PDF created with pdfFactory trial version www.pdffactory.com
Top Related