







Idiomas
Páginas
Jurídico


Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Fracciones de tamaño de fitoplancton deFracciones de tamaño de fitoplancton delas lagunas de la turbera de Rancholas lagunas de la turbera de Rancho
Hambre (Tierra del Fuego):Hambre (Tierra del Fuego):caracterización y relación con los factorescaracterización y relación con los factores
bióticos y abióticosbióticos y abióticos
González Garraza, Gabriela
2013-04-05
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
González Garraza, Gabriela. (2013-04-05). Fracciones de tamaño de fitoplancton de las lagunasde la turbera de Rancho Hambre (Tierra del Fuego): caracterización y relación con los factoresbióticos y abióticos. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.
Cita tipo Chicago:
González Garraza, Gabriela. "Fracciones de tamaño de fitoplancton de las lagunas de la turberade Rancho Hambre (Tierra del Fuego): caracterización y relación con los factores bióticos yabióticos". Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2013-04-05.

UNIVERSIDAD DE BUENOS AIRES
Facultad de Ciencias Exactas y Naturales
Fracciones de tamaño del fitoplancton de las lagunas de la turbera de Rancho Hambre (Tierra del Fuego):
caracterización y relación con los factores bióticos y abióticos
Tesis presentada para optar al título de Doctor de la Universidad de Buenos Aires en el área de Ciencias Biológicas
Autor: Gabriela González Garraza
Director de Tesis: Dra. Gabriela Mataloni
Consejero de estudios: Dra. Irina Izaguirre
Lugar de trabajo: Instituto de Investigación e Ingeniería Ambiental (3iA)
Universidad Nacional de Gral. San Martin
Buenos Aires, 2012

ÍNDICE
Resumen…………………………………………………………………………………i-ii
Agradecimientos………………………………………………………………………..iii-iv
Dedicatoria…………………………………………………………………………………v
Introducción…………………………………………………………………………….1- 9
Objetivos e Hipótesis………………………………………………………………….10-11
Materiales y métodos………………………………………………………………….12-26
Resultados
1) Análisis de los parámetros ambientales a- Características meteorológicas……………………………………...27-30 b- Características hidrológicas…………………………………………31-32 c- Variación de la intensidad luz y de las sustancias húmicas en la columna
de agua ……………………………………………………………..32-34 d- Características físico-químicas……………………………………...34-38 e- Relación entre los parámetros ambientales…………………………39-40
2) Caracterización de la comunidad fitoplanctónica Nano- y microfitoplancton………………………………………………….41-58
a- Riqueza específica y composición taxonómica……………………..41-53 b- Abundancia, biovolumen y diversidad……………………………..53-59
Picofitoplancton…………………………………………………………….59-62 Fracciones de tamaño del fitoplancton……………………………………...62-64 Clorofila a………………………………………………………………………65 Estrategias nutricionales…………………………………………………….66-67
3) Análisis de los posibles predadores de la comunidad fitoplanctónica a- Nanoflagelados heterotróficos………………………………………… 68 b- Protozooplancton (ciliados)…………………………………………68-69 c- Metazooplancton (rotíferos y crustáceos)…………………………..70-71
4) Relación entre los factores bióticos y abióticos y los distintos descriptores de la comunidad fitoplanctónica………………………………………………...72
5) Análisis de los grupos funcionales basados en características morfológicas (GFBM) sensu kruk et al. (2010) y sus posibles predadores……………73-80
6) Relación entre los grupos funcionales basados en características morfológicas (GFBM) y los factores bióticos y abióticos………………..80-83

Discusión……………………………………………………………………………...84-103
Análisis de los factores ambientales……………………………………………………84-89
La comunidad fitoplanctónica………………………………………………………….89-96
Relación entre las distintas comunidades planctónicas con énfasis en el estudio del fitoplancton…………………………………………………………………………….96-97
Los grupos funcionales basados en características morfológicas (GFBM) y su relación
con los factores bióticos y abióticos ……………………………………………….....97-103
Conclusiones generales……………………………………………………………..104-106
Bibliografía………………………………………………………………………….107-126
Anexo………………………………………………………………………………...127-128

RESUMEN
i
FRACCIONES DE TAMAÑO DEL FITOPLANCTON DE LAS LAGUNAS DE LA
TURBERA DE RANCHO HAMBRE (TIERRA DEL FUEGO): CARACTERIZACIÓN Y
RELACIÓN CON LOS FACTORES BIÓTICOS Y ABIÓTICOS.
Se estudió la composición, estructura y dinámica de la comunidad fitoplanctónica
en relación a parámetros bióticos y abióticos en cinco lagunas de la turbera de Rancho
Hambre, situada dentro de la Reserva Natural y Paisajística del Valle de Tierra Mayor,
(Tierra del Fuego). El análisis de las características físico-químicas y su relación con
parámetros meteorológicos, hidrológicos y morfométricos reveló que la microtopografía
determinó en gran medida la conectividad hidrológica superficial, la que influyó en las
características físico-químicas y moduló los cambios en el nivel hidrométrico. Esto, junto
con la morfometría, dió como resultado una gran diversidad ambiental. En segundo lugar,
se caracterizó la comunidad fitoplanctónica de mayor tamaño la cual fue muy rica en
especies (305 taxones). Esta riqueza estuvo representada por Zygnematophyceae
(Desmidiaceae), Chlorophyceae y Bacillariophyceae revelando floras distintivas en cada
cuerpo de agua, mientras que la fracción picofitoplanctónica estuvo compuesta
exclusivamente por picoalgas eucariotas. El pH y la temperatura explicaron las variaciones
espaciales (riqueza específica y clorofila a) y temporales (abundancia y diversidad),
respectivamente de la comunidad fitoplantónica, que en general estuvo numéricamente
dominada por picoalgas y pequeños flagelados. Finalmente, mediante un análisis
multivariado (CCA) se relacionaron los grupos morfo-funcionales del fitoplancton sensu
Kruk et al. (2010) con factores bióticos y abióticos, observándose que las abundancias de
ciliados y de los macrofiltradores de zooplancton (Daphnia commutata), así como el pH
resultaron fundamentales en la determinación de las estrategias predominantes en las
distintas comunidades fitoplanctónicas.
Palabras claves: Fitoplancton, fracciones de tamaño, grupos morfo-funcionales,
limnología, turberas, Tierra del fuego.

RESUMEN
ii
PHYTOPLANKTON SIZE FRACTIONS OF WATER HOLLOWS FROM RANCHO
HAMBRE PEAT BOG (TIERRA DEL FUEGO PROVINCE): CHARACTERISATION
AND RELATION WITH BIOTIC AND ABIOTIC FEATURES
This Ph. D. thesis studied the composition, structure and dynamics of phytoplankton
communities in relation to biotic and abiotic parameters in water bodies from Rancho
Hambre peat bog, located in, Tierra del Fuego province, Argentina. First, physical and
chemical characteristics were analyzed in relation to meteorological, hydrological and
morphometric features. We observed that small-scale local topography determined
hydrological connectivity, which in turn dictated physical and chemical features and
modulated changes in water level. This, together with morphometrical features, resulted in
a high environmental diversity. Then their phytoplankton communities were characterized,
revealing 305 taxa, most of them belonging to the Zygnematophyceae (Desmidiaceae),
Chlorophyceae and Bacillariophyceae with regards to the nano- and microphytoplankton
fraction, while eukaryotic algae entirely represented the picophytoplankton. For the whole
system, the community was numerically dominated by picoalgae and small flagellates.
Abundance and diversity varied widely according to temperature, while pH was the key
factor determining species richness and the concentrations of phytoplanktonic chlorophyll
a. Finally, a multivariate analysis (CCA) related the morpho-functional groups of the
phytoplankton sensu Kruk et al. (2010) with biotic and abiotic features. As a result of this,
we observed that the abundances of ciliates, zooplankton macrofilterers (Daphnia
commutata) and pH mainly dictated the prevailing strategies in the different phytoplankton
communities.
Keywords: Phytoplankton, size fractions, morpho-functional groups, limnology, peat bogs,
Tierra del Fuego.

AGRADECIMIENTOS
iii
AGRADECIMIENTOS
En primer lugar, MUCHAS PERO MUCHAS GRACIAS Gaby por la confianza, la
paciencia y por compartir conmigo todo tu conocimiento y ser la persona que me dio la
posibilidad de conocer ese hermoso lugar llamado Rancho Hambre.
Un gracias enorme!!!! a todo el laboratorio de limnología, Guillermo, Irina, Inés,
Haydee, Rubén, Ali, Rodri, Pati, Paulita, Luchi, Sole, Sol, Romi, Lau, Gri, Euge y Juan a
todos gracias por el aguante, los consejos y la buena onda de siempre!!!.
A mis compañeros de campañas, gracias por los recuerdos juntos, los momentos
lindos y por compartir conmigo todo su conocimiento. Gracias Cris, Silvina, Gabis, Ali,
Memo, Fer, Rubén, Vicky, Guillermo y Adrián!!!!!!. Quiero agradecer muy profundamente
a la Dirección de Recursos Hídricos, Secretaría de Desarrollo Sustentable y Ambiente de la
provincia de Tierra del Fuego, en especial a la persona que estuvo siempre ayudándome,
gracias Sergio!!!!!!!, no me quiero olvidar de Rodolfo Iturraspe y Adriana Urciuolo que
pusieron a mi disposición toda su ayuda siempre y los chicos de la secretaría Rodrigo y
José por acompañarme en mi segundo muestreo. Gracias al dream team por los recuerdos y
las campañas fabulosas que pasamos!!!!!!!!!!!. No quiero olvidarme de la gente que conocí
en el CADIC, los cuales siempre estuvieron dispuestos a ayudarme, Daniel Fernández
GRACIAS!!!!. Quiero agradecer profundamente al Dr. Andrés Boltovskoy por determinar a
los dinoflagelados de la turbera de Rancho Hambre.
Gracias papas, hermana y mi pequeña sobri!!! por comprenderme siempre y
apoyarme en todas mis locuras!!!!!!!
Mi amor que te puedo decir, gracias por aguantarme y darme siempre todo tu apoyo
en estos últimos años y a mi hijo perruno, Roco gracias por estar conmigo cuando empecé a
escribir y entender que mamá tenía que trabajar.
A mis amigotas, que supieron entender mis ausencias en estos últimos años, gracias
Ine, Pitu y Betty!!!!. A lore y Marcelo por aguantarme en su casa cuando tenía que viajar a
Ushuaia.
Al 3iA, que supo abrirme las puertas en esta una nueva etapa, gracias Alberto por
preguntarme siempre como iba la tesis!!!!. Quiero agradecer, en particular al grupo de

AGRADECIMIENTOS
iv
Biodiversidad, Limnología y Biología de la Conservación por acompañarme en estos
últimos momentos, gracias Rubén, Lizi, Andre, Ana y Vale. A mis compañeros del instituto
Diana, Cristina, Iani y Gaby!!!!.
Por último, a todas las personas que me acompañaron en esta etapa de aprendizaje
muchas pero muchas gracias!!!.
MUCHAS GRACIAS A TODOS!!!!
Las campañas de muestreo fueron financiadas por un subsidio de la Agencia
Nacional de Promoción Científica y Tecnológica (PICT 1697) y del Consejo Nacional de
Investigaciones Científicas y Técnicas (CONICET PIP 11220090100050). La presente tesis
fue financiada por la la Agencia Nacional de Promoción Científica y Tecnológica y el
Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Algunos cursos y congresos fueron financiados con la ayuda económica otorgada en
el 2011 por la Neotropical Grassland Conservancy (Ayuda económica 2000 pesos).

DEDICATORIA
v
A mis padres, a mi hermana y sobrina

INTRODUCCIÓN

INTRODUCCIÓN
1
INTRODUCCION
Los turbales son humedales de importancia global con una distribución altamente
heterogénea. La mayoría se ubica en el hemisferio norte, y sólo un 4% en América de Sur
(Parish et al. 2008) (Figura 1). Estos ecosistemas constituyen más del 50% de los
humedales del mundo (Iturraspe 2010) cubriendo una superficie equivalente al 3% del
planeta (Joosten & Clarke 2002).
En cuanto a la definición de turbales, primero es necesario definir la turba. La
turba es un conjunto de restos tanto de origen animal como vegetal que se acumulan bajo
condiciones de anegamiento permanente y aguas ácidas, las que dan como resultado
condiciones de anoxia y baja tasa de descomposición del material vegetal (Roig & Roig
2004, Rydin & Jeglum 2006). Los turbales son ecosistemas que tienen la capacidad de
acumular y almacenar turba. Dentro de estos ecosistemas, se encuentran extensiones
donde la turba está siendo producida y acumulada, denominados “turberas” (Roig & Roig
2004).
Figura 1. Distribución mundial de los turbales (modificado de Lappalainen 1996).
<5%, 5-10% y >10% de la superficie del terreno cubierta por turbales.

INTRODUCCIÓN
2
Los ecosistemas de turberas se pueden diferenciar en minerotróficos y
ombrotróficos en base a sus componentes botánicos, origen y cantidad de nutrientes,
hidrología y topografía (Roig & Roig 2004, Iturraspe 2010).
Las turberas minerotróficas generalmente se sitúan en depresiones con superficies
planas o cóncavas, con aportes de agua subterránea y superficial, ricas en nutrientes, de
moderada acidez y mayormente dominadas por ciperáceas (Iturraspe 2010) (Figura 2-A).
Las turberas ombrotróficas, en cambio, se encuentran sobreelevadas respecto del
terreno circundante con superficies convexas en forma de domo, planas o con pendiente
sin aportes de agua subterránea, pobres en nutrientes, muy ácidas y dominadas por el
musgo Sphagnum (Rydin & Jeglum 2006, Iturraspe 2010) (Figura 2-B).
Figura 2. Tipos de turbera. A- turbera minerotrófica, B- turbera ombrotrófica.

INTRODUCCIÓN
3
La evolución temporal de una turbera típica comienza desde un estadío inicial
minerotrófico hasta uno final ombrotrófico asociado con cambios en el estado trófico del
sistema (Wetzel 1981).
La estructura de una turbera define su comportamiento hidrológico y los procesos
que regulan el ciclo del carbono (Iturraspe 2010).Una turbera ombrotrófica elevada tiene
una estructura diplotélmica, donde se distinguen dos estratos denominados acrotelmo
(estrato superior de la turba) y catotelmo (estrato inferior de la turba), el primero
corresponde a una zona hidrológicamente activa con flujo horizontal reducido mientras
que el segundo se encuentra constantemente saturado con baja permeabilidad (Iturraspe
& Roig 2000, Roig & Roig 2004, Iturraspe 2010).
Las interacciones hidrológicas a nivel de la turbera y adyacencias son esenciales
para el desarrollo y funcionamiento del humedal. Las turberas tienen la capacidad de
almacenar grandes cantidades de agua dulce y constituyen así reservorios fundamentales
para regulación de la cuenca hídrica (Iturraspe 2010). En una turbera ombrotrófica
elevada, el agua libre fluctúa como respuesta a condiciones climáticas. Aumenta por
efecto de las precipitaciones o descongelamiento de nieve y disminuye principalmente
por evapotranspiración, y en menor medida por drenaje, que se encuentra comúnmente en
forma lateral, hacia un colector superficial marginal o bien hacia el escurrimiento
subterráneo presente en el suelo mineral (Iturraspe & Roig 2000) (Figura 3).
Generalmente, la precipitación excede la evapotranspiración potencial y esta es
significativamente mayor generando un excedente. Las turberas ombrotróficas de
Sphagnum presentan la napa freática paralela a la superficie convexa de la turbera.
Además, en los extremos marginales del domo, donde se incrementa la pendiente, las
fluctuaciones de los niveles de agua y la circulación de flujo son mayores (Köpke 2005,
Baumann 2006, Iturraspe 2010).

INTRODUCCIÓN
4
Figura 3. Ciclo hidrológico en una turbera ombrotrófica (modificado de Iturraspe 2010).
Las turberas de Tierra del Fuego
El 95% de las turberas de la Argentina se encuentra en la provincia de Tierra del
Fuego (Rabassa et al. 1996), abarcando una extensión de 2700 km2 (Iturraspe & Urciuolo
2004). Las primeras observaciones sobre las turberas fueguinas, realizadas por Darwin
(Mataloni 1999) se remontan al siglo XIX, mientras que el primer relevamiento y mapeo
completo de estos sistemas fue realizado por Bonarelli (1917). Este trabajo y los que le
siguieron enfatizan sobre la génesis de las turberas en relación con el clima, la macroflora
y la turba como recurso económico, tal como la exhaustiva descripción de Auer (1965).
Roig (2004) hace una valiosa enumeración de publicaciones sobre la importancia de estos
ambientes como reservorio de indicadores paleoclimáticos.
Los estudios enfocados en la hidrología de turberas en Tierra del Fuego
comenzaron con los trabajos de Iturraspe et al. (1998) e Iturraspe & Roig (2000). En el
primero de estos estudios, los autores realizaron una caracterización hidrológica de los
valles de Tierra Mayor y Carbajal, mientras que en el segundo se estudiaron los aspectos
hidrológicos de una turbera ombrotrófica de Sphagnum centrado en el funcionamiento de

INTRODUCCIÓN
5
sus diferentes estratos. Luego, Iturraspe & Urciuolo (2000) dividieron la sección
argentina de la isla de Tierra del Fuego en cuatro cuencas hidrológicas en base a criterios
de homogeneidad y sistemas de regulación dominantes. La cuenca de la zona norte
(estepa) comprende a todos los ríos localizados al norte del río Grande, donde las tasas de
flujo de la cuenca son pequeñas, y los ambientes lénticos son escasos, someros y de
origen eólico. La cuenca de la zona central (transición) comprende el área del Ecotono y
presenta un amplio rango de predominancia de llanuras onduladas con cursos de agua.
Aquí una gran proporción del flujo de los valles está ocupado por humedales y turbales.
La cuenca de la zona sur (cordillera) está delimitada por los andes fueguinos al norte y
por el canal de Beagle al sur. Un gran número de lagos de origen glaciar caracterizan esta
área. La red de drenaje es densa y caracterizada por ríos con pendiente hacia los valles de
turberas, los cuales presentan pequeños cuerpos de agua. Dentro de esta cuenca, se
encuentra el valle de Tierra Mayor, que incluye a la localidad de Rancho Hambre, área de
estudio en esta tesis doctoral. Por último, la cuenca de la zona este (turbales) ocupa la
Península Mitre, un área baja, mayormente cubierta por turberas con un muy lento
drenaje (Figura 4).
Figura 4. Cuencas hídricas de Tierra del Fuego (modificado de Iturraspe & Urciuolo 2000).
Ushuaia

INTRODUCCIÓN
6
En cuanto a la biodiversidad ficológica de los ambientes acuáticos de turberas de
Tierra del Fuego, los primeros relevamientos de la flora de microalgas fueron hechos
tempranamente por Borge (1906). Más tarde, Guarrera & Kühnemann (1949),
Thomasson (1955, 1956, 1957), Yacubson (1963), Tell & Mosto (1982), Guarrera (1986)
Lenzenweger (1993), Vigna & Kristiansen (1995 a, b, 1996), Guarrera & Echenique
(1998), Kristiansen & Vigna (1996, 2002) y Maidana (2000) estudiaron la ocurrencia de
distintos grupos taxonómicos en Tierra del Fuego, incluyendo a las turberas entre otros
sitios de estudio. Los trabajos específicos sobre la flora algal de turberas fueguinas son
más escasos (Mataloni 1991, 1995 a, b, 1997, Vigna 1993, Tell 1995).
Recientemente, como parte de un gran estudio limnológico, Tell et al. (2011)
estudiaron la distribución geográfica y biodiversidad de Chlorococcales (Chlorophyceae)
en 33 cuerpos de agua a lo largo de una transecta latitudinal desde el norte patagónico
hasta Tierra del Fuego. Entre estos, 11 se localizaron en esta última provincia incluyendo
lagos profundos, someros y cuerpos de agua de turberas y temporarios. Estos autores
registraron 24 especies de Chlorococcales, la mayor riqueza específica para este grupo
en Tierra del Fuego.
La primera investigación limnológica regional realizada en Tierra del Fuego fue
llevada a cabo por Mariazzi et al. (1987). Estos autores estudiaron 19 lagos y un gran
número de cuerpos de agua temporarios, incluyendo un charco localizado en Península
Mitre. Dentro de este estudio se midieron numerosas características limnológicas y se
estudiaron en detalle el zooplancton y el fitoplancton de red de veinte cuerpos de agua.
Luego, los sistemas acuáticos fueron clasificados de acuerdo a sus características físicas,
químicas y geoambientales.
En cuanto a la turbera de Rancho Hambre, el primer estudio limnológico fue
realizado por Mataloni & Tell (1996). Estos autores compararon la comunidad
fitoplanctónica de cinco cuerpos de agua y del río Lasifashaj en relación a datos
morfométricos, químicos y físicos. Los cuerpos de agua se separaron en dos grupos según
su composición específica. Las características morfométricas (área, profundidad media) y
la conductividad explicaron estas diferencias florísticas, mientras que los factores

INTRODUCCIÓN
7
químicos influyeron fuertemente en las frecuencias relativas de algunos grupos
taxonómicos. En un trabajo posterior, Mataloni (1999) estudió las comunidades
microalgales a lo largo de un gradiente desde aguas abiertas a condiciones de sequedad
en cuerpos de agua de seis turberas pertenecientes a distintos valles glaciarios localizados
en el extremo sudoeste de Tierra del Fuego (Figura 5). La evolución de estos sistemas
hacia condiciones “terrestres” se evidenció con un incremento de la conductividad y una
disminución del pH y la humedad. A lo largo de estos gradientes hubo una disminución
de la riqueza específica y un aumento de las frecuencias relativas de algunas pocas
especies tolerantes a condiciones de sequedad, acidez y conductividad elevada. El
ordenamiento de los puntos de muestreo en un análisis de correspondencia basado en las
frecuencias relativas de las 35 especies dominantes, demostró que estas sucesiones eran
convergentes. Por otra parte, los ojos de agua de las distintas turberas se agruparon entre
sí en función de sus características minerotróficas u ombrotróficas, más que por su
cercanía geográfica o pertenencia a una misma cuenca.
Figura 5. Mapa de la zona sur de la provincia Tierra del Fuego indicando los sitios de muestreo de
Mataloni (1999).

INTRODUCCIÓN
8
A nivel mundial, Gilbert & Mitchell (2006) recopilaron la información sobre los
distintos grupos microbianos de ecosistemas de turbales, que incluye a bacterias,
cianobacterias y algas eucariotas, flagelados heterotróficos, ciliados, tecamebas y
rotíferos. Allí se tratan aspectos de su estructura (diversidad, abundancia y biomasa) y
relaciones funcionales (descomposición, producción primaria, bucle microbiano). Según
esta recopilación, los únicos trabajos limnológicos en turberas del hemisferio sur son las
investigaciones de Mataloni & Tell (1996) y Mataloni (1999) en Tierra del Fuego, lo que
destaca la escasez de información general sobre el funcionamiento y biodiversidad de las
turberas fueguinas.
Problemática
Las turberas juegan un papel importante en cuanto al mantenimiento de la calidad
del agua y en la regulación del recurso hídrico. Así como también cumplen un rol
fundamental en la atenuación del cambio climático global, actuando como sumideros de
carbono (de la Balze et al. 2004). Estos ambientes albergan un tercio del carbono
existente en el suelo y el 10% del agua dulce disponible (Joosten & Clarke 2002). A
partir de 1800, el área global de turberas se ha reducido en el 20-30%, siendo la actividad
extractiva humana la principal causa de dicha reducción tanto a escala local como global
(Joosten & Clarke 2002).
Para la explotación de turba como mineral es necesario drenar la turbera, lo cual
estimula las pérdidas de sus propiedades de humedal y finalmente la transformación en
otro tipo de suelo, generalmente degradado. En Tierra del Fuego, hace algunos años, esta
actividad se desarrolló en mayor medida en la zona central de la isla, en el área de
influencia de la localidad de Tolhuin. En menor medida, se extrajo turba en los valles
próximos a Ushuaia, donde las turberas tienen gran importancia por su alto valor
paisajístico (Roig & Roig 2004).
En los últimos años, la provincia de Tierra del Fuego ha creado áreas de turberas
protegidas. En 2001 el gobierno de la provincia declaró al Valle de Tierra Mayor como
Reserva Natural y Paisajística y en 2009 el Glaciar Vinciguerra y Turberas asociadas,

INTRODUCCIÓN
9
que incluye al valle de Andorra, fue declarado como sitio RAMSAR. Recientemente, el
gobierno de la provincia impulsó el desarrollo de un ordenamiento territorial de las
turberas basado en aspectos ecosistémicos, sociales y económicos. Este ordenamiento
propuso áreas a conservar como el Valle de Andorra, Península Mitre, Valle de Carbajal,
Turberas de altura (Moat) y la Reserva Turístico- Paisajística de Tierra Mayor, que
incluye a la localidad de Rancho Hambre.
El informe mundial de los Recursos de Humedales (Secretaría de la Convención
de Ramsar 2010) reconoció que las turberas son algunos de los ecosistemas de humedales
más vulnerables y amenazados por la pérdida y la degradación de los hábitats, y que por
consiguiente necesitan acciones urgentes y prioritarias para garantizar su conservación y
uso racional, que incluyen un inventario de la biodiversidad y de otras funciones que
cumplen las turberas locales; así como el desarrollo de conocimientos básicos para
monitorear su comportamiento y sus alteraciones. En este sentido, el proyecto de
investigación titulado “Las lagunas de la turbera de Rancho Hambre (Tierra del Fuego)
un estudio limnológico integrado”, en el que se enmarca esta tesis doctoral, se propuso
fundar una línea de investigación interdisciplinaria dedicada al estudio de la estructura y
funcionamiento de los ecosistemas limnéticos de la turbera de Rancho Hambre (Tierra
del Fuego), tomándola como caso de estudio de este tipo de humedales. En el marco de
este proyecto se realizaron muestreos durante el período de aguas libres (octubre-abril) en
cinco cuerpos de agua de distintas características morfométricas. En ellos se relevaron los
parámetros hidrológicos y físico-químicos, así como también la diversidad de las
comunidades planctónicas. En particular, esta tesis doctoral se propone aportar
información de base necesaria para el inventario de biodiversidad propuesto por la
Convención de Ramsar, contribuyendo así a la futura planificación de su uso sustentable.

OBJETIVOS
E
HIPÓTESIS

OBJETIVOS E HIPÓTESIS
10
OBJETIVOS
Objetivos generales
-Estudiar la estructura y variación temporal de la comunidad fitoplanctónica y su
relación con parámetros morfométricos y ambientales en cinco cuerpos de agua de la
turbera de Rancho Hambre.
Objetivos específicos
-Estudiar la abundancia de las distintas fracciones de tamaño del fitoplancton en
dichos cuerpos de agua y su variación estacional e interanual.
-Estudiar, además, la composición taxonómica, riqueza específica y diversidad de
las fracciones de mayor tamaño (> 2 micrones) de la comunidad fitoplanctónica.
-Analizar cómo se relacionan estas características con las variables
morfométricas, hidrológicas, físicas y químicas que caracterizan a cada uno de estos
ambientes, y con las abundancias de otras comunidades planctónicas (protozoo- y
metazooplancton).
-Analizar si las especies del fitoplancton responden a las condiciones comunes del
ambiente a través de rasgos funcionales comunes y similares, representados por grupos
morfo-funcionales.

OBJETIVOS E HIPÓTESIS
11
HIPÓTESIS
-Las características morfométricas e hidrológicas de los ojos de agua determinan
en gran parte las propiedades físicas y químicas de sus aguas.
-Estas propiedades, especialmente las concentraciones de nutrientes, pH y
conductividad, influyen a su vez sobre la composición y estructura del fitoplancton.
-En cada cuerpo de agua, la composición y estructura de las comunidades
planctónicas responde, además, a las interacciones que estas comunidades presentan entre
sí. En particular, el fitoplancton responde a sus interacciones con el protozoo- y el
metazooplancton.
-Las características distróficas de los cuerpos de agua determinan la dominancia
de los organismos mixótrofos en el fitoplancton.

MATERIALES
Y
MÉTODOS

MATERIALES Y MÉTODOS
12
MATERIALES Y MÉTODOS
Área de estudio y Diseño de muestreo
La turbera de Rancho Hambre (54° 47´S, 68° 19´W) (Figura 6) se encuentra
dentro del Valle de Tierra Mayor, a 50 km de la ciudad de Ushuaia. Tiene una cuenca
de drenaje densa, con muchos aportes de agua desde los valles transversales y de las
pendientes de las montañas (Iturraspe et al. 1998). El área está sobreelevada respecto al
terreno circundante, en forma de domo, y fue clasificada como una turbera ombrotrófica
(Roig 2004), donde el único aporte de agua es por precipitación y descongelamiento de
nieve (Roig & Roig 2004). La vegetación está ampliamente dominada por el musgo
Sphagnum magellanicum Bridel. Una gran proporción de la superficie de Rancho
Hambre está cubierta por cuerpos de agua. Cinco de ellos representativos, localizados a
lo largo de la sección transversal de la turbera y de diferentes características
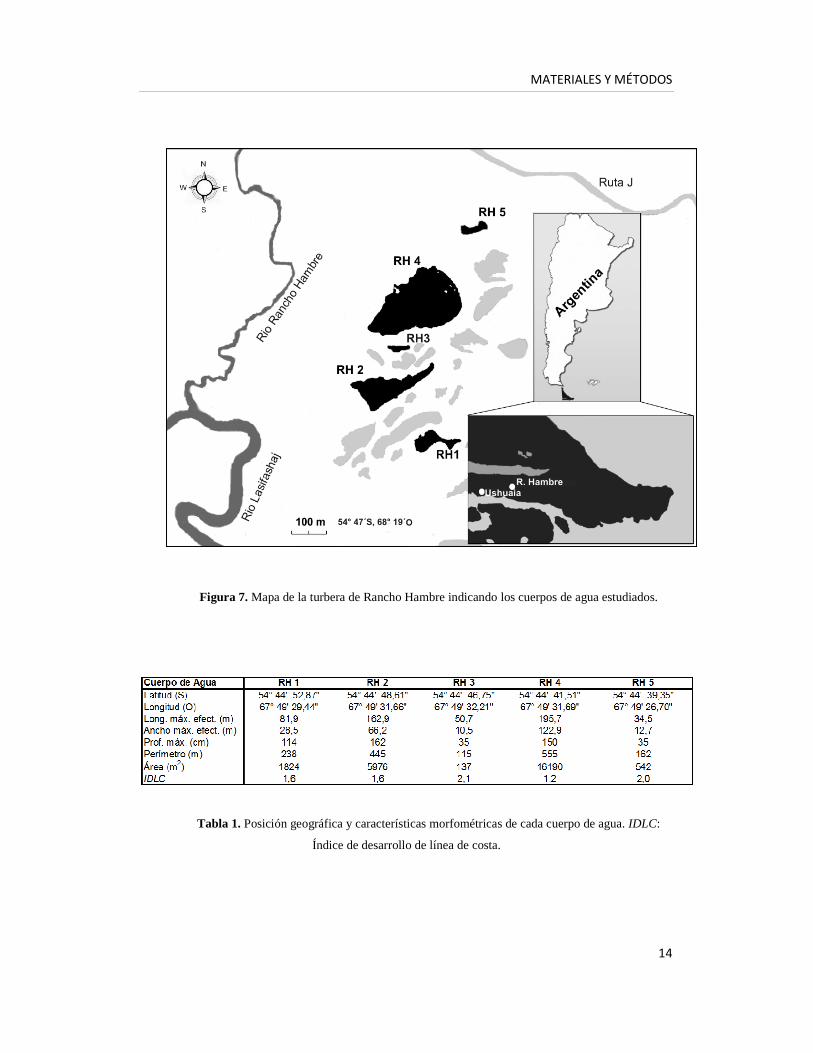
morfométricas fueron seleccionados para este estudio (Figuras 6 y 7, Tabla 1). RH1,
RH2 y RH4 son cuerpos de agua grandes y relativamente profundos, entre los cuales
RH4 es el de mayor superficie, mientras que RH3 y RH5 son sistemas someros
(profundidad <50 cm) (Figura 8). Los cuerpos de agua se muestrearon en ocho
ocasiones durante el período de aguas libres (octubre-abril) entre octubre de 2008 y
abril de 2010. En cada ojo de agua se seleccionaron desde uno a cuatro puntos de
muestreo dependiendo de su tamaño. En RH1 y RH2 se establecieron tres puntos de
muestreo (orilla, centro-superficie y centro-fondo), y cuatro puntos (orilla sur y norte,
centro-superficie y centro-fondo) en RH4. Los pequeños cuerpos de agua RH3 y RH5
se muestrearon sólo desde la orilla.

MATERIALES Y MÉTODOS
13
Figura 6. Fotografía de la turbera de Rancho Hambre.
MATERIALES Y MÉTODOS
14
Figura 7. Mapa de la turbera de Rancho Hambre indicando los cuerpos de agua estudiados.
Tabla 1. Posición geográfica y características morfométricas de cada cuerpo de agua. IDLC:
Índice de desarrollo de línea de costa.

MATERIALES Y MÉTODOS
15
Figura 8. A- Cuerpos de agua profundos de la turbera de Rancho Hambre (RH1, RH2 y RH4).
Figura 8. B- Cuerpos de agua someros de la turbera de Rancho Hambre (RH3 y RH5).

MATERIALES Y MÉTODOS
16
Posición geográfica y características morfométricas
La posición geográfica de cada laguna fue establecida con un GPS Garmin
Etrex. Las profundidades máximas fueron estimadas mediante sondeos con una
plomada desde el bote o desde orilla en el caso de las más someras. Las características
morfométricas (largo máximo efectivo, ancho máximo efectivo y perímetro) fueron
medidas a través de imágenes de alta resolución obtenidas de Google Earth y sus áreas
se determinaron con un planímetro polar digital. El Índice de Desarrollo de Línea de
Costa (IDLC), el cual describe la regularidad de costa fue calculado como IDLC= P/
2*(πA)1/2, donde P es el perímetro y A es el área del cuerpo de agua (Hutchinson 1975).
Así, para una determinada área, un cuerpo de agua totalmente circular tiene el mínimo
IDLC posible (1) y el IDLC aumenta con el incremento de la irregularidad del contorno
del cuerpo de agua.
Mediciones meteorológicas
Las variables meteorológicas se midieron en forma continua con una estación
meteorológica automática DAVIS localizada cerca del río Rancho Hambre, a un 1 km
de los cuerpos de agua, desde febrero de 2009. Las variables medidas incluyeron a la
temperatura del aire, precipitación, y velocidad y dirección del viento. Desde junio de
2008 hasta mayo de 2010, se midió la temperatura subsuperficial (0,25―0,30 m de
profundidad) del agua en los cuerpos de agua de menor y mayor superficie (RH3 y
RH4, respectivamente) así como también la temperatura de la turba (0,30 m de
profundidad) usando un registrador de temperatura Ondotori TR-52i (precisión: ± 0,3
°C). Las mediciones fueron tomadas cada hora y se calcularon valores promedio diarios
y mensuales.
Mediciones hidrológicas
Los niveles hidrométricos se midieron cada tres semanas desde diciembre de
2008 usando tubos de plástico (PVC) de 40 mm de diámetro insertados verticalmente en
la turba ubicados próximos a cada cuerpo de agua. Los niveles hidrométricos se
midieron como las profundidades hasta el tope de los tubos de PVC utilizando una cinta
métrica. Luego, se realizó un muestreo topográfico usando una estación total PENTAX

MATERIALES Y MÉTODOS
17
R-326EX para referenciar las mediciones del nivel hidrométrico de todos los cuerpos
de agua a metros sobre el nivel del mar. Por último, se calculó el coeficiente de
variación de los niveles hidrométricos de cada cuerpo de agua a lo largo del estudio
utilizando la siguiente fórmula:
CV (%) = (desvío estándar/media)*100.
Mediciones de la intensidad lumínica
La radiación fotosintéticamente activa (PAR) se midió in situ en los cuerpos de
agua profundos (RH1, RH2 y RH4) a intervalos de 20 cm usando un radiómetro
equipado con un sensor cuántico esférico sumergible (Li- Cor, USA). El coeficiente de
atenuación vertical de la luz (Kd) se calculó como la pendiente de la relación entre ln
(Irradiancia) vs. profundidad (Kirk 1994).
Mediciones de sustancias húmicas
A partir de octubre de 2009, se tomaron muestras de agua, las cuales se filtraron
por un filtro Millipore APFF (0,7µm diámetro de poro). A partir del filtrado, se
realizaron mediciones de absorbancia a distintas longitudes de onda (254, 320, 440 y
750 nm) usando como blanco agua destilada. Los coeficientes de absorción se
calcularon como a= 2,303 D/r, donde D es la absorbancia y r es el camino óptico (Kirk
1994). La absorbancia a 254 nm (A254) fue utilizada como estimador de la materia
orgánica disuelta (Keskitalo & Eloranta 1999), el coeficiente de absorción a 320 nm
(a320) como indicador del carbono orgánico disuelto coloreado (Williamson et al. 1999)
y el coeficiente de absorción a 440 nm (a440) fue utilizado como indicador del color
del agua ( Kirk 1994).
Mediciones de los factores físico-químicos
En cada fecha de muestreo se midió in situ, la temperatura, el pH, la
conductividad y el oxígeno disuelto (OD) con una sonda multiparamétrica (HORIBA,
Japón). El porcentaje de saturación de oxígeno fue estimado usando los datos de OD,
altitud y temperatura.

MATERIALES Y MÉTODOS
18
Las muestras de agua para análisis químicos fueron tomadas en botellas de
plástico lavadas con una dilución de ácido clorhídrico al 2%. Las concentraciones de
nutrientes y la dureza total se midieron con un espectrofotómetro Hach (Hach
Company, USA) con los reactivos apropiados para cada análisis y utilizando los
protocolos del espectrofotómetro Hach 2800 (www.hach.com). En el laboratorio en
Ushuaia, las muestras para nutrientes disueltos fueron filtradas a través de un filtro
Millipore APFF (diámetro de poro: 0,7 µm). Las muestras para medición de amonio
(NH4-N) se conservaron a pH 2 y a 4 °C y se analizaron posteriormente según el
método del salicilato (N° 8155). Las muestras para nitratos (NO3-N) y fosfatos (PO4-P)
se conservaron a -20 °C y se analizaron según los métodos de la reducción del cadmio
(N° 8192) y del ácido ascórbico (N° 8048), respectivamente. El nitrógeno inorgánico
disuelto (NID) se calculó como la suma entre el amonio y el nitrato. Las muestras sin
filtrar para el análisis de nitrógeno total (NT), el fósforo total (PT) y la dureza se
preservaron a -20 °C hasta su análisis. NT y PT fueron determinados por digestión con
persulfato de potasio y ácido bórico (APHA 2005) seguida de las determinaciones de
NO3-N y PO4-P. La dureza de los iones de calcio y magnesio fue determinada por el
método colorimétrico de la calmagita (n° 8030), luego se estimó la dureza total según
APHA (2005). Para determinar la concentración de sólidos suspendidos (SS) se filtró
una muestra de agua a través de un filtro Millipore APFF calcinado y pre-pesado. El
material retenido en el filtro se secó hasta peso constante y los sólidos se estimaron por
diferencia de peso:
SS (mg L-1)= Peso filtro con material–Peso filtro sin material (APHA 2005)
Las determinaciones de carbono orgánico disuelto (COD) se realizaron desde
octubre de 2009 filtrando las muestras de agua a través de un filtro Millipore de 0,7 µm
de tamaño de poro previamente calcinado a 500 °C. Las muestras se colocaron en
frascos oscuros luego de ser llevadas a pH 2 y se conservaron a 4 °C hasta su análisis en
el Laboratorio Experimental de Tecnologías Sustentables- Instituto Nacional del Agua
(INA). Aquí, se determinaron utilizando un método de oxidación catalítica Pt de alta
temperatura (Analizador Shimadzu TOC-5000A, técnica SM5310B, límite de
detección: 0,1 mg/L) siguiendo las recomendaciones de Sharp et al. (1993).

MATERIALES Y MÉTODOS
19
Mediciones de la clorofila a fitoplanctónica
Para estimar la clorofila a fitoplanctónica se filtró un volumen conocido de
agua a través de un filtro Millipore APFF (diámetro de poro: 0,7 µm), los filtros se
preservaron a -20 °C. Los pigmentos fotosintéticos se extrajeron con etanol caliente
(60–70 °C) y se mantuvieron por 24 horas a 4 °C en oscuridad (Nusch 1980). La
concentración de clorofila a libre de feopigmentos se determinó por espectrofotometría
midiendo a 665 y 750 nm antes y después de acidificar con ácido clorhídrico 0,1N. Se
utilizaron las ecuaciones de Marker et al. (1980).
Análisis de la comunidad fitoplanctónica
� Picofitoplancton (fracción < 2µm)
-Análisis cuantitativo
Para el análisis cuantitativo del picoplancton autotrófico se tomaron muestras en
frascos de plástico de 120 ml y se fijaron con glutaraldehído filtrado y frío
(concentración final 2%) y se conservaron a 4 °C. Antes de las 24 horas de tomada la
muestra se filtró un volumen conocido (2, 4 ó 5 ml de acuerdo a la densidad presente) a
través de filtros de policarbonato negro de 0,2 µm de poro. Estos se montaron entre
porta y cubreobjetos con aceite de inmersión para epifluorescencia. Los preparados se
conservaron a -20 °C. Los recuentos del picoplancton autotrófico se realizaron mediante
microscopio óptico para epifluorescencia con un aumento de 1000× (Olympus BX40F4,
Japón). El error de los recuentos se calculo según Venrick (1978) (Error=
[media/(n1/2)*t*100]*DS -1) aceptando un error máximo del 20% para la especie
dominante. Las picoalgas (eucariotas y procariotas) se identificaron por su
autofluorescencia dada por los pigmentos fotosintéticos ante la excitación con luz azul
(450- 490 nm) y verde (546 nm) (Callieri y Pinolini 1995). Las picoalgas eucariotas
excitadas bajo luz azul se ven rojas y la picocianobacterias se ven amarillas (células
ricas en ficoeritrina) o rojas oscuras (células ricas en ficocianina), dependiendo de la
presencia o ausencia de la ficobiliprotenía, ficoeritrina. Bajo luz verde, las
picocianobacterias se ven amarillas/naranjas (células ricas en ficoeritrina) y rojas

MATERIALES Y MÉTODOS
20
(células ricas en ficocianina) (Callieri 2007). El biovolumen del picofitoplancton se
calculó multiplicando la abundancia por el volumen celular que se estimó a través de
análisis de imágenes (Image Pro-Plus), utilizando las fórmulas y los criterios propuestos
por Hillebrand et al. (1999).
� Nano- y microfitoplancton (fracción > 2µm)
-Análisis cuali- y cuantitativo
Para el análisis cualitativo del fitoplancton de mayor tamaño se tomaron
muestras con red de plancton de 18µm de diámetro de poro, se fijó una fracción con
formol al 4% y se mantuvo otra fracción sin fijar para el análisis de los organismos
vivos bajo microscopio óptico con un aumento de 1000× (Olympus BX40F4, Japón).
Para la determinación específica de las Bacillariophyceae (diatomeas), las muestras
fueron oxidadas en caliente con peróxido de hidrógeno al 30% y luego se lavaron con
agua destilada (Battarbee 1986) y se realizaron preparados permanentes utilizando la
resina Styrax como medio de montaje. Los distintos ejemplares se identificaron
taxonómicamente utilizando una vasta cantidad de bibliografía; en particular los
trabajos florísticos de Desmidiaceae de Coesel (1996), Coesel & Krienitz (2008),
Croasdale (1962), Cvijan & Lausevic (1991), Grönblad (1942), Kirk et al. (1976),
Kouwets (1987), Krieger (1937), Presccott et al. (1972, 1977), Stastný J. (2010), Teiling
(1966), West & West (1904, 1905, 1908, 1912, 1922) y Williamson (1992); los de
Xanthophyceae de Ettl (1978), los de Chrysophyceae de Hüber-Pestalozzi (1941),
Starmach (1985), los de Cryptophyceae de Javornický (2003), Lepistö & Holopainen
(2003), los de Cyanobacteria de Komárek & Anagnostidis (1999, 2005), el de
Chlorophyceae de Komárek & Fott (1983) y los de Bacillariophyceae de Krammer
(1992, 1997 a, b), Krammer & Lange-Bertalot (1986, 1988, 1991 a, b) y Van de Vijver
et al. (2002), los de turberas del mundo de Ireneé- Marie (1938), Kitner et al. (2004),
Kulikovskiy et al. (2010), Stepánkova et al. (2008) y por último los específicos de los
diferentes ambientes acuáticos de la provincia de Tierra del Fuego: Lenzenweger
(1993), Maidana (2000), Mataloni (1991, 1995, 1997), Tell (1995), Tell & Conforti

MATERIALES Y MÉTODOS
21
(1986), Yacubson (1963), entre otros. Para el análisis cuantitativo se tomaron muestras
por duplicado en frascos de plástico de 120 ml y se fijaron con solución ácida de Lugol
al 1%. Las muestras se dejaron sedimentar 24 horas en cámaras de 10 ml y los recuentos
se realizaron utilizando un microscopio invertido Olympus bajo un aumento de 400×
(Utermöhl 1958). El error de los recuentos se calculó de acuerdo a Venrick (1978)
aceptando un error máximo del 20% para la especie dominante. El biovolumen del
nano- y microfitoplancton se estimó multiplicando la abundancia por el volumen algal
por especie, utilizando las fórmulas y los criterios propuestos por Hillebrand et al.
(1999) y Sun & Liu (2003).
Otras comunidades planctónicas estudiadas en el marco del mismo proyecto de
investigación a cargo de diferentes especialistas
� Nanoflagelados heterotróficos (Lic. Quiroga M.V & Dr. Unrein)
Para el análisis cuantitativo de los nanoflagelados heterotróficos (NFH) se
tomaron muestras de agua en frascos de plástico y se fijaron con glutaraldehído filtrado
y frío a una concentración final de 2% y se conservaron a 4°C. Antes de las 24 horas de
tomada la muestra, se filtró un volumen conocido (10, 15, 20 ml dependiendo de la
abundancia presente) a través de membranas de policarbonato negras de 0,8 µm de poro
y se tiñeron con DAPI (50µl) utilizando la técnica estándar (Porter & Feig 1980). Los
filtros se montaron entre porta y cubreobjetos con aceite de inmersión de baja
fluorescencia. Los preparados se conservaron a -20°C. Los recuentos se realizaron
mediante microscopio óptico para epifluorescencia bajo un aumento de 1000×
(Olympus BX40F4, Japón) y el error de los recuentos fue calculado según Venrick
(1978) aceptando un error máximo del 20%. Los nanoflagelados heterotróficos se
cuantificaron por medio de la excitación con luz azul (450- 490 nm) y UV para
discriminar los organismos heterótrofos de los autótrofos.
� Protozooplancton (Ciliados) (Dra. Küppers M.G)
Para el estudio de los ciliados planctónicos en cada punto de muestreo se
tomaron muestras por duplicado en bidones de 1 litro y se fijaron con lugol acético al

MATERIALES Y MÉTODOS
22
2% (Finlay & Guhl 1992). Posteriormente se dejaron decantar por 24 horas en
oscuridad y se redujo el volumen a 120 ml. Las muestras se dejaron sedimentar por 24
horas en cámaras de 10 ml y los recuentos se realizaron bajo el microscopio invertido
(Utermöhl 1958) con un aumento de 150× y 600×. Las especies fueron identificadas a
partir de la observación de muestras vivas y de organismos impregnados con protargol
(Wilbert 1975). Las identificaciones de las especies se basaron en los trabajos de Kahl
(1930, 1931, 1932, 1935), Foissner et al. (1991, 1992, 1994, 1995, 1999), Foissner &
Berger (1996), y en artículos taxonómicos específicos.
� Metazooplancton (rotíferos y crustáceos) (Lic. Marinone M.C & Dra. Menu Marque S.)
Las muestras cualitativas del metazooplancton se tomaron por arrastre con redes
de plancton de 120 µm de poro en el caso de los microcrustáceos (cladóceros y
copépodos), y de 55 µm para los rotíferos. En ambos casos se fijaron con alcohol 96%.
Para el análisis cuantitativo se extrajeron muestras por duplicado, obteniéndose
volúmenes de agua de 5 litros en el centro de las lagunas profundas y 20 litros en orillas
y ambientes someros. Las muestras se filtraron in situ por un tamiz de 55 µm de malla y
se preservaron en formol al 4%. El microzooplancton (nauplii y rotíferos) se
submuestreó con una pipeta de Hensen-Stempel y se contó en una cámara de Sedgwick-
Rafter de 1 ml bajo microscopio óptico. Las alícuotas del mesozooplancton
(copepoditos, copépodos adultos y cladóceros) se extrajeron con un submuestreador de
Russell y se enumeraron en una cámara de Bogorov de 5 ml bajo estereomicroscopio.
Se contaron las alícuotas necesarias para mantener el error de recuento por debajo del
10%. Las identificaciones taxonómicas se realizaron según Bayly (1992) y Reid (1985)
para Copepoda, Benzie (2005), Orlova-Bienkowskaja (1998), Paggi (1979, 1995) y
Smirnov (1992) para Cladocera, y Boltovskoy & Urrejola (1977), Ruttner-Kolisko
(1974), y Voigt & Koste (1978) para Rotifera. Se clasificó al metazooplancton en los
siguientes gremios alimenticios siguiendo los criterios propuesto por diferentes autores,
tales como Heywood (1970 a, b), Weller (1977) Tranvik & Hansson (1997), Green &
Shiel (1999), Almada (2002) e Izaguirre et al. (2003):
-Microfiltradores: larvas nauplii de Copepoda, Cladocera y Rotifera
-Macrofiltradores: Daphnia commutata y Simocephalus cf. Mixtus

MATERIALES Y MÉTODOS
23
-Cladocera perifitófagos: Familia Chydoridae: Alona spp., Chydorus spp. y Pleuroxus
spp.
-Copepoda Calanoida filtradores: Boeckella poppei y sus copepoditos
-Copepoda macrófagos herbívoros: copepoditos de Cyclopoida, adultos y copepoditos
de Harpacticoida
-Predadores o macrófagos carnívoros: adultos de Cyclopoida: Diacyclops sp., Eucyclops
sp., Paracyclops sp. y Tropocyclops prasinus meridionalis; Rotifera: Asplanchna girodi
(<600 µm) y Ploesoma truncatum.
Análisis de datos
- Para determinar la existencia de diferencias significativas para las variables bióticas y
abióticas entre los distintos puntos de muestreo en cada uno de los cuerpos de agua
profundos (RH1, RH2 y RH4), en el caso de los parámetros físico-químicos,
picofitoplancton y nanoflagelados heterotróficos, se realizó un análisis de varianza
multivariado en bloques aleatorizados, considerando los períodos de muestreo como
bloques y los puntos de las lagunas como factores fijos. Además, para cada variable se
realizó un análisis de varianza de dos factores utilizando el modelo III de bloques
aleatorizados sin réplicas (Zar 2010). Para el resto de las comunidades planctónicas, en
cada cuerpo de agua y fecha de muestreo se realizó un análisis de varianza de un factor
(Zar 2010). En ambos casos se utilizó el programa SPSS Statistics 17.0.
- Para relacionar las variables meteorológicas (temperatura del aire; precipitación
acumulada a los 1, 3, 7 y 14 días previos a la fecha de muestreo; velocidad del viento) y
limnológicas (temperatura del agua, pH, conductividad, oxígeno disuelto, sólidos
suspendidos, dureza total, NID, NT, PO4-P y PT) se realizó una matriz de correlación
para cada cuerpo de agua utilizando los datos registrados entre febrero de 2009 y abril
de 2010. Como muchas de estas variables no siguen una distribución normal se utilizó
el coeficiente de correlación no paramétrico de Spearman (Zar 2010) (programa SPSS
Statistics 17.0).

MATERIALES Y MÉTODOS
24
- Para determinar qué factores afectan a la intensidad de luz en la columna de agua se
realizó una matriz de correlación entre el coeficiente de extinción vertical de la luz (Kd)
y todas las variables medidas. Se utilizó el programa SPSS Statistics 17.0 y, dado que la
mayoría de las variables no siguen una distribución normal se empleó el coeficiente de
correlación no paramétrico de Spearman (Zar 2010).
- Para relacionar los diferentes estimadores de sustancias húmicas (A254, a320 y a440)
con los parámetros medidos se realizó una matriz de correlación. Dado que la mayoría
de las variables no sigue una distribución normal se empleó el coeficiente de correlación
no paramétrico de Spearman (Zar 2010) (programa SPSS Statistics 17.0).
- Para analizar la dinámica de las características físico-químicas de los cinco cuerpos de
agua en el mismo período en que se midieron los parámetros meteorológicos e
hidrológicos, se realizó un Análisis de Componentes Principales (PCA) utilizando los
datos registrados entre febrero de 2009 y abril de 2010 usando el programa NTSYSpc
2.2.
- Para detectar asociaciones específicas en la composición del fitoplancton de los
distintos cuerpos de agua se realizó un análisis de agrupamiento y además se calcularon
índices de similitud por cuerpo de agua (NTSYSpc 2.2.) basados en la matriz de
presencia-ausencia de especies utilizando el índice de similitud de Jaccard (J), el cual
compara los sitios de a pares (Magurran 2004):
J = a/ (n-d)
donde a representa el número de especies presentes en ambos sitios; n el número de
especies presentes en toda la muestra y d representa el número de especies ausentes en
ambos sitios. Este índice adquiere el valor 1 cuando todas las especies son compartidas
entre las comunidades que se están comparando y vale 0 cuando la composición de las
dos muestras es totalmente diferente.

MATERIALES Y MÉTODOS
25
- Para cada una de las muestras obtenidas en cada cuerpo de agua a lo largo de los 8
muestreos se calculó la diversidad fitoplanctónica utilizando el índice de Shannon-
Weaver (Magurran 2004):
H= ∑ pi*ln pi
donde pi es la abundancia de la especie i
y la equitatividad
E= H/ln (S)
donde S es la riqueza total de cada cuerpo de agua en cada fecha de muestreo.
- Para determinar la existencia de diferencias significativas en el volumen celular del
picofitoplancton entre los cuerpos de agua profundos se realizó un análisis de varianza
de un factor utilizando el programa SPSS Statistics 17.0.
- Para determinar qué factores bióticos y abióticos influyen sobre los distintos
descriptores de la comunidad fitoplanctónica (abundancia, diversidad, riqueza específica
y clorofila a) se realizaron regresiones múltiples aplicando cuando fuera necesario la
transformación y= ln (x) a las variables dependientes para obtener una distribución
normal, y un análisis de correlación utilizando el coeficiente de correlación no
paramétrico de Spearman (SPSS Statistics 17.0), ya que la mayoría de las variables no
siguen una distribución normal (Zar 2010).
- Las especies fitoplanctónicas fueron clasificadas en Grupos Funcionales Basados en
las características Morfológicas (GFBM) sensu Kruk et al. (2010).
-Para relacionar la abundancia y el biovolumen de los GFBM del fitoplancton y las
abundancias de sus posibles predadores (proto- y metazooplancton) se realizaron
correlaciones múltiples usando el coeficiente de correlación no paramétrico de
Spearman (SPSS Statistics 17.0) dado que muchas variables no siguen una distribución
normal (Zar 2010).
- Para relacionar el biovolumen de los grupos funcionales basados en las características
morfológicas (Kruk et al. 2010) con los valores de los parámetros bióticos y abióticos se

MATERIALES Y MÉTODOS
26
utilizó un Análisis de Correspondencia Canónica (CCA) ya que el biovolumen de los
GFBM presentó una respuesta unimodal testeada mediante un DCA (Detrended
Canonical Analysis). En ambos casos se utilizó el programa CANOCO 4.5. La
significación de los ejes canónicos fue testeada mediante el Test de Monte Carlo.

RESULTADOS

RESULTADOS
27
RESULTADOS
1) Análisis de los parámetros ambientales
a- Características meteorológicas
La Figura 9 muestra la temperatura del aire, del agua (RH3 y RH4) y de la turba
desde junio de 2008 hasta junio de 2010. Durante el invierno, la temperatura de la turba se
mantuvo por encima de la temperatura del agua. El descongelamiento de los cuerpos de
agua ocurrió en ambos años a principios de octubre, junto con un marcado incremento de la
temperatura del aire. La temperatura en el cuerpo de agua de menor superficie (RH3)
mostró una marcada variación diaria en el período de aguas libres de hielo, mientras que en
el cuerpo de agua de mayor superficie (RH4) y en la turba, las temperaturas fueron
claramente más estables.
Figura 9. Variación temporal de la temperatura del aire, del agua (RH3 y RH4) y de la turba. El mal
funcionamiento del datalogger impidió el registro de las temperaturas de la turba y de RH4 entre enero y
junio de 2009.

RESULTADOS
28
El rango de variación mensual de la temperatura del aire registrado durante nuestro
estudio (-0,7―8,7°C) estuvo dentro de los registros medidos para Rancho Hambre desde
1999 (-4,1―11,4°C), mientras que los mínimos y máximos absolutos de temperatura
durante el período fueron -12,9 y 21,1°C, respectivamente (Figura 10). El cálculo de la
temperatura media anual por año hidrológico (período de aguas libres, desde octubre hasta
el siguiente marzo) mostró que el período octubre 2009―marzo 2010 fue el más frío de los
últimos siete años.
Figura 10. A-Temperatura del aire mensual, B- precipitación mensual registrada en la turbera de Rancho
Hambre desde marzo de 2009. ND: no hay dato. Las barras indican ± DS.

RESULTADOS
29
En particular, la Figura 11 muestra el diferente aspecto de RH2, el último cuerpo de
agua en descongelarse, durante los muestreos de primavera de los dos años de estudio.
Figura 11. La laguna RH2 en octubre A- de 2008 y B- de 2009.
El promedio de la precipitación mensual registrada dentro del período de estudio fue
de 60 mm con valores máximos en agosto de 2009, enero de 2010 y febrero de 2010 (>100
mm) y mínimos en septiembre de 2009 (<27 mm). Tanto la temperatura del aire como la
precipitación mensual variaron en forma similar desde septiembre de 2009 a abril de 2010,
excepto en diciembre de 2009, donde la temperatura aumentó y las precipitaciones
disminuyeron. Este último parámetro mostró un fuerte incremento al mes siguiente (enero

RESULTADOS
30
de 2010) (Figura 10). Los vientos predominantes en el área de estudio fueron de dirección
oeste, coincidiendo con la orientación oeste-este del valle de Tierra Mayor en general y de
los cuerpos de agua en particular. Las velocidades del viento promedio para los meses de
muestreo oscilaron entre 5,3 y 8,7 km h-1 (Figura 12) siendo el máximo valor registrado
104,6 km h-1.
Figura 12. Frecuencia de la dirección del viento (%). Se calcularon las velocidades del viento promedio para
cada mes de muestreo (se indican en color verde).

RESULTADOS
31
b- Características hidrológicas
Observaciones de campo mostraron que cada uno de los cuerpos de agua presenta su
propia cuenca de aportes, cuyos límites están determinados topográficamente por sectores
elevados de turba que soportan vegetación arbustiva (renovales de Nothofagus sp.). Por otra
parte, varios de los cuerpos de agua en la turbera de Rancho Hambre están interconectados
mediante canales naturales superficiales. Dentro de los cuerpos de agua más grandes, RH1
y RH4 tienen entradas y/o salidas, mientras que RH2 está aislado hidrológicamente.
Los niveles hidrométricos de los cinco cuerpos de agua se registraron durante el
período de aguas libres desde diciembre de 2008 (Figura 13). Los cuerpos de agua
pequeños y someros (RH3 y RH5) mostraron similar tendencia a lo largo del período de
estudio, con coeficientes de variación (CV) de 18 y 11% respectivamente. RH2 mostró el
nivel hidrométrico más variable (CV= 29%) debido a su aislamiento hidrológico, RH1 un
valor intermedio (CV= 16%) y el de mayor superficie (RH4) fue el más estable, mostrando
el más bajo CV (7%). Los niveles hidrométricos de todos los cuerpos de agua se muestran
en la Figura 13-f. A partir de este gráfico, se observa que no hay evidencia de
interconexiones entre los cuerpos de agua estudiados.

RESULTADOS
32
Figura 13. a-e: Variación temporal del nivel hidrométrico (metros sobre el nivel del mar) de cada cuerpo de
agua. f: Variación temporal de los niveles hidrométricos de todos los cuerpos de agua estudiados.
c- Variación de la intensidad de luz y de las sustancias húmicas en la columna de agua
El coeficiente de atenuación vertical de la luz (Kd) promedio en los sistemas
profundos a lo largo del período de estudio (RH1, RH2 y RH4) fue de 2,5 m-1 con un rango
de variación entre 1,68 y 3,45 m-1. Este parámetro varió tanto estacionalmente como entre
los dos años de estudio, y mostró diferentes patrones en cada cuerpo de agua (Figura 14).
No se encontraron correlaciones significativas entre el Kd y las variables medidas en este
estudio.

RESULTADOS
33
Figura 14. Variaciones temporales del coeficiente de extinción vertical de la luz (Kd) en los cuerpos
de agua profundos, ND: no hay dato.
Las estimaciones de las sustancias húmicas se realizaron desde octubre de 2009
hasta abril de 2010. En general, los valores de absorbancia a 254 nm (A254) (estimador de
materia orgánica disuelta) (Keskitalo & Eloranta 1999) y los coeficientes de absorción a
320 nm (a320) (estimador del carbono orgánico disuelto coloreado) (Williamson 1999) y a
440 nm (a440) (estimador del color del agua) (Williamson et al. 1999, Huovinen et al.
2000) estuvieron altamente correlacionados con las concentraciones carbono orgánico
disuelto (COD) (rA254=0,89; ra320=0,88; ra440=0,69; en todos los casos p=0,01) y fueron

RESULTADOS
34
mayores en los cuerpos de agua someros (RH3 y RH5) que en el resto de los sitios (Figura
15). A su vez, estos estimadores mostraron tendencias similares, presentando un patrón
estacional más marcado en los sistemas someros (RH3 y RH5), con valores bajos en la
primavera (octubre) (Figura 15); a causa de esto las sustancias húmicas se correlacionaron
positivamente con la temperatura (rA254nm= 0,54; ra320=0,52; ra440= 0,50; en todos los casos
p<0,05).
Figura 15. Sustancias húmicas (A254, a320 y a440) en los distintos cuerpos agua y fechas de muestreo.
d- Características físico-químicas
A lo largo del período de estudio, los parámetros físicos y químicos no mostraron
diferencias significativas entre los distintos puntos de muestreo en los cuerpos de agua
profundos (RH1, RH2 y RH4) (ANOVA p>0,05). Dados estos resultados, para el análisis
en esta sección se promediaron los valores obtenidos en cada fecha para cada cuerpo de
agua.

RESULTADOS
35
En la Tabla 2 se resumen las características físico-químicas de los cinco cuerpos de
agua de la turbera de Rancho Hambre para el período de estudio.
Tabla 2. Características físico-químicas registradas en cada punto de muestreo a lo largo del período de
estudio. Se calcularon valores promedio, con los mínimos y máximos entre paréntesis (nRH1 = 23, nRH2 =22,
nRH3 =8, nRH4 =32, nRH5 =8). Cond: conductividad, OD: oxígeno disuelto, SO: % de saturación de oxígeno,
Sólidos susp: sólidos suspendidos, NID: nitrógeno inorgánico disuelto, COD: carbono orgánico disuelto.
RH1 RH2 RH3 RH4 RH5Temperatura 8,8 8,6 11,8 8,5 10,6
(°C ) (2,2 - 17,3) (1,1 - 17,0) (3,2 - 25,0) (2,4 - 15, 0) (1,7 - 20,0)
pH 5,7 4,5 4,5 6,3 4,6(5,0 - 7,1) (3,8 - 5,5) (3,6 - 5,4) (5,8 - 7,0) (4,1 - 5,4)
Cond 23,6 22,8 33,1 29,8 25,6
(µS cm-1) (13,6 - 50,0) (8,7 - 40,0) (10,0 - 82,0) (16,0 - 60,0) (5,5 - 50,0)OD 10,5 11,3 10,4 10,6 9,9
(mg L-1) (8,2 - 14,0) (7,6 - 14,4) (8,7 - 12,9) (7,6 - 12,2) (8,6 - 11,5)
SO 93 101 100 94 93(%) (65 - 114) (63 - 135) (73 - 134) (63 - 118 ) (71 - 112)
Sólidos susp 2,1 2,5 5,5 6,1 4,1
(mg L-1) (0,7 - 3,7) (0,9 - 8,3) (0,4 - 20) (1,6 - 23,9) (0,3 - 10,9)NID 46 53 55 44 36
(µg L-1) (7 - 102) (7 - 239) (10 - 103) (19 - 107) (0 - 73)N total 5317 6293 7305 6859 9479
(µg L-1) (1430 - 10100) (1980 - 12870) (1980 - 11330) (1073 - 26000) (3410 - 30000)
PO4-P 62 58 61 34 31
(µg L-1) (27 - 93) (23 - 157) (30 - 130) (10 - 60) (20 - 50)
P total 206 172 169 164 195
(µg L-1) (113 - 477) (92 - 330) (90 - 308) (88 - 290) (77 - 420)
Dureza total 25,6 24,2 22 30,5 22,3
(mg equiv CaCO3 L-1) (7,0 - 41,4) (6,8 - 46,2) (7,5 - 43,3) (11,0 - 42,5) (10,9 - 36,4)
COD 7,3 7,5 10,4 5,4 8,4
(mg L-1) (5,4 - 9,2) (5,1 - 9) (2,8 - 13,4) (4,4 - 7,0) (3,9 - 11,6)
TN:TP 36,0 40,4 52,1 50,2 62,9(por peso) (10,0 - 69,4) (10,1 - 64,4) (15,0 - 104,4) (7,5 - 128,4) (11,7 - 132,9)
DIN:PO4-P 0,9 0,8 1,2 1,5 1,6(por peso) (0,08 - 1,5) (0,1 - 1,5) (0,08 - 2,4) (0,7 - 3,1) (0 - 3,7)
La temperatura varió estacionalmente con valores máximos y mínimos
correspondientes a las estaciones de verano y de otoño/primavera, respectivamente (Figura
16). Los cuerpos de agua más grandes (RH1, RH2 y RH4) presentaron menores valores
promedios y rangos de variación en comparación con los sistemas someros (RH3 y RH5)
(Tabla 2 y Figura 16).

RESULTADOS
36
Figura 16. Variación temporal de la temperatura de cada cuerpo de agua a lo largo del período de
estudio.
Además, los ambientes someros (RH3 y RH5) también mostraron altos valores
promedio y amplios rangos para la conductividad y los sólidos suspendidos. La dureza
total, en cambio, mostró valores bajos y poco variables para todos los cuerpos de agua. El
pH mostró un comportamiento estable a lo largo del tiempo, pero separó los cuerpos de
agua en dos grupos diferentes: RH1 y RH4 (pH promedio= 6,0) por un lado, y RH2, RH3 y
RH5 (pH promedio= 4,5) por el otro. Para todos los cuerpos de agua, el valor medio de
oxígeno disuelto fue >9,9 mg L-1 con un rango de variación del porcentaje de saturación de
oxígeno que osciló entre 63 y 135 %. El rango de variación de la concentración de carbono
orgánico disuelto varió entre 2,8 y 13,4 mg L-1 registrándose estos extremos en el cuerpo de
agua más pequeño (RH3) (Tabla 2).
Con respecto a los nutrientes, la mayor proporción de nitrógeno inorgánico disuelto
(NID) en estos sistemas acuáticos estuvo presente como amonio, mientras que el nitrato fue
no detectable en la mayoría de las muestras. El NID fue bajo en todos los sitios y representó
una muy pequeña fracción del nitrógeno total. Por otro lado, alrededor del 25% del fósforo
total estuvo presente como fosfato (Tabla 2). En las Figuras 17 y 18 se muestra el distinto
comportamiento de las variaciones temporales de los nutrientes disueltos (NID y PO4-P) en
los cuerpos de agua. Mientras que el NID en general separó los sistemas profundos (RH1,
RH2, RH4) de los someros (RH3 y RH5), el comportamiento del PO4-P fue muy variable

RESULTADOS
37
(Figuras 17 y 18). Para este último parámetro, los sistemas ligeramente ácidos (RH1 y
RH4) reflejaron las mismas condiciones en octubre de ambos años, mostrando luego
comportamientos diferentes (Figura 18).
Figura 17. Variación temporal de la concentración de nitrógeno inorgánico disuelto (NID) en los distintos
cuerpos de agua. ND: no hay dato.

RESULTADOS
38
Figura 18. Variación temporal de la concentración de fosfato (PO4-P) en los distintos cuerpos de agua.
La proporción NT: PT fue siempre mayor a la proporción de Redfield (7,2:1 por
peso), la cual representa la proporción de N y P necesarios para el crecimiento del
fitoplancton (Guilford & Hecky 2000). Los valores mínimos de esta proporción para los
diferentes cuerpos de agua variaron desde 7,5 a 15,0 y fueron encontrados después del
descongelamiento de primavera, mientras que los máximos oscilaron entre 66,3 y 132,9 y
no se relacionaron con una fecha de muestreo específica. En cambio, la proporción NID:
PO4-P fue mucho menor que la proporción de Redfield, variando entre ≈ 0 y 3,66. La
concentración tanto de los nutrientes disueltos como totales en todos los cuerpos de agua se
encontraron dentro del mismo rango de variación, así las fluctuaciones temporales dentro
de cada uno de ellos superaron en general la variabilidad espacial del sistema en un
momento dado (Tabla 2).

RESULTADOS
39
e- Relaciones entre los parámetros ambientales
La conductividad mostró bajos valores asociados con las precipitaciones previas al
día de muestreo (tres y siete días) a través de una correlación negativa entre ambos
parámetros (rRH1= -0,61, -0,56; rRH4= -0,65, -0,77; en todos los casos p<0,05), evidenciando
un efecto de dilución en RH1 y RH4. Dentro de las correlaciones realizadas entre variables
meteorológicas y limnológicas, las mencionadas anteriormente fueron las únicas que
resultaron significativas.
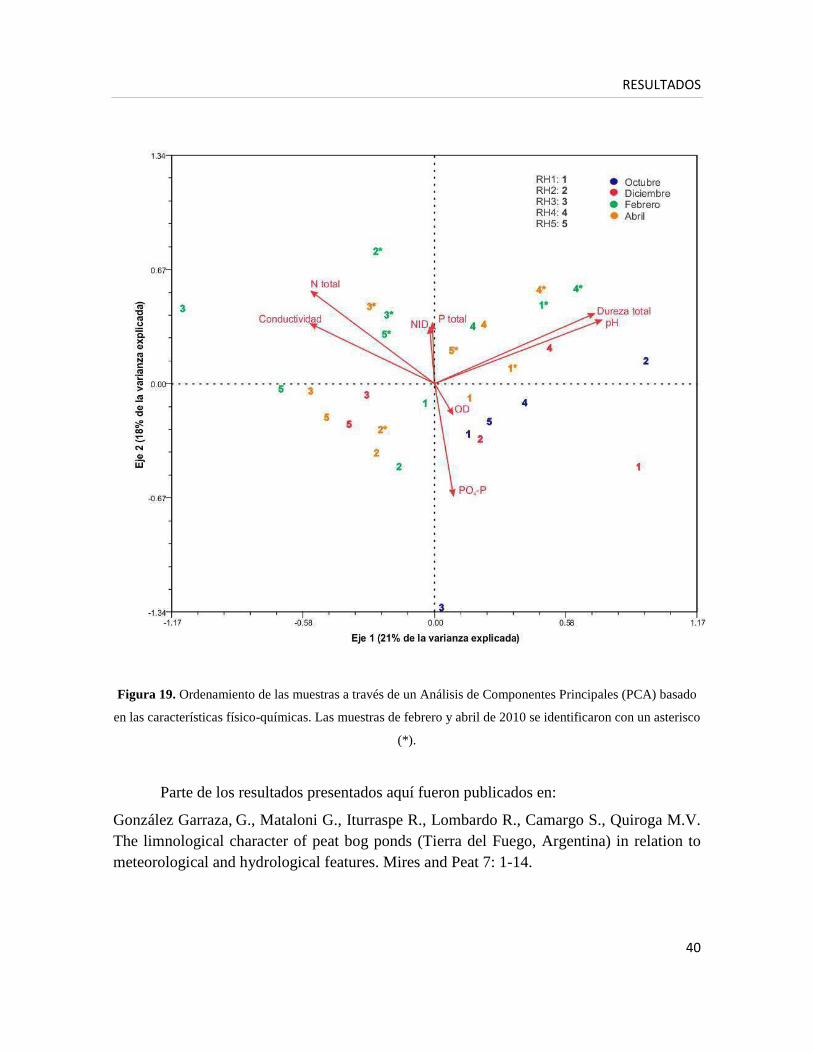
La Figura 19 muestra los resultados del Análisis de Componentes Principales (PCA)
realizado en base a las características físico-químicas de cada cuerpo de agua entre febrero
de 2009 y abril de 2010, en los cuales los dos primeros ejes explican el 39% de la varianza.
El ordenamiento de las muestras a lo largo del primer eje (21% de la varianza) reflejó un
gradiente del estado minero-ombrotrófico. RH2, RH3 y RH5 se localizaron en el extremo
izquierdo del diagrama, mientras que RH1 y RH4 se ubicaron a la derecha, asociados a
altos valores de pH (autovector= 0,75) y dureza total (autovector= 0,72), que reflejan
condiciones minerotróficas. En el segundo eje (18% de la varianza) las muestras se
ordenaron a lo largo de un gradiente temporal, con las muestras de fines de verano (febrero)
localizadas generalmente en la parte superior del diagrama, mientras que las condiciones de
bajas temperaturas (octubre) son representadas en la parte inferior. Éstas se asociaron con
altos valores de fosfatos (autovector= -0,67) y bajas concentraciones de N total
(autovector= 0,55). La ubicación de la mayoría de las muestras de febrero-abril 2009 en la
parte inferior del diagrama, y las de febrero-abril 2010 en la parte superior evidenció la
amplia variación interanual de los parámetros físico-químicos que dependen de la
temperatura y varían estacionalmente.
RESULTADOS
40
Figura 19. Ordenamiento de las muestras a través de un Análisis de Componentes Principales (PCA) basado
en las características físico-químicas. Las muestras de febrero y abril de 2010 se identificaron con un asterisco
(*).
Parte de los resultados presentados aquí fueron publicados en:
González Garraza, G., Mataloni G., Iturraspe R., Lombardo R., Camargo S., Quiroga M.V. The limnological character of peat bog ponds (Tierra del Fuego, Argentina) in relation to meteorological and hydrological features. Mires and Peat 7: 1-14.

RESULTADOS
41
2) Caracterización de la comunidad fitoplanctónica
• Nano- y microfitoplancton
a- Riqueza específica y composición taxonómica
A partir del estudio taxonómico detallado de la flora algal de los cinco cuerpos de
agua de la turbera de Rancho Hambre a lo largo de ocho campañas de muestreo se
registraron 305 taxones (Tabla 3) entre los cuales predominaron las Zygnematophyceae
(Desmidiaceae) (33%), las Chlorophyceae (22%), las Bacillariophyceae (20%) y las
Cyanobacteria (13%).
Tabla 3. Lista florística de todas las especies en cada cuerpo de agua. Los taxones presentes en todos
los cuerpos de agua se indican con un asterisco (* ).
Taxones RH1 RH2 RH3 RH4 RH5
Cyanobacteria
Alternantia geitleri Schil. * ● ● ● ● ●
Anabaena sp. 1 ● ●
Anabaena sp. 2 ● ● ● ●
Aphanocapsa elachista W. et G.S.West * ● ● ● ● ●
Aphanocapsa incerta (Lemm.) Cronberg et Kom. * ● ● ● ● ●
Aphanocapsa planctonica (G.M.Smith) Kom. et Anag. ● ● ● ●
Aphanothece elabens (Bréb.) Elenkin ● ● ● ●
Aphanothece minutissima (W.West) Komárkova-Legnerová et Cronberg ● ● ● ●
Aphanothece nidulans Richter * ● ● ● ● ●
Aphanothece stagnina (Sprengel) A.Braun * ● ● ● ● ●
Calothrix sp. ●
Chroococcus limneticus Lemm. * ● ● ● ● ●
Chroococcus minimus (Keissler) Lemm. * ● ● ● ● ●
Chroococcus minor (Kütz.) Näg. ● ● ● ●
Chroococcus minutus (Kütz.) Näg. * ● ● ● ● ●
Chroococcus turgidus (Kütz.) Näg. * ● ● ● ● ●
Chroococcus sp. ● ● ●
Cyanodictyon reticulatum (Lemm.) Geitler * ● ● ● ● ●
Cyanosarcina burmensis (Skuja) Kovacik ● ●
Cyanothece aeruginosa (Näg.) Kom. ●

RESULTADOS
42
Taxones RH1 RH2 RH3 RH4 RH5
Cylindrospermum sp. ● ●
Eucapsis minor (Skuja) Elenkin * ● ● ● ● ●
Geitlerinema splendidum (Greville) Anag. ●
Gloeocapsopsis pleurocapsoides (Novacek) Kom. et Anag. * ● ● ● ● ●
Hapalosiphon cf. hibernicus W. et G.S.West ● ● ●
Leptolyngbya crassior (Skuja) Anag. ● ● ●
Merismopedia elegans A.Braun * ● ● ● ● ●
Merismopedia punctata Meyen * ● ● ● ● ●
Microcystis smithii Kom. et Anag. * ● ● ● ● ●
Nostoc sp. ●
Phormidium chlorinum (Kütz.) Anag. ● ●
Phormidium aff. chlorinum (Kütz.) Anag. ● ●
Phormidium simplicissimum (Gom.) Anag. et Kom. ● ●
Pseudanabaena catenata Lauterborn ● ● ● ●
Pseudanabaena sp. ●
Rhabdoderma lineare Schmidle et Lauterborn * ● ● ● ● ●
Rhabdogloea linearis (Geitler) Kom. * ● ● ● ● ●
Rhabdogloea smithii (R. et F.Chodat) Kom. ● ●
Rhabdogloea sp. * ● ● ● ● ●
Scytonema sp. ●
Subtotal 32 25 26 32 28
Bacillariophyceae
Achnanthes sp.1 ● ● ●
Achnanthes sp.2 ● ●
Achnanthidium minutissimum (Kütz.) Czarnecki ● ● ● ●
Brachysira brebissonii Ross ● ●
Brachysira cf. intermedia (Oestrup) L.-B. ● ● ●
Cocconeis placentula var. lineata (Ehr.) V.H. ● ● ● ●
Cymbella cf. heteropleura (Ehr.) Kütz. ● ●
Cymbopleura naviculiformis Auerswald ● ●
Encyonema elginense (Krammer) D.G.Mann ●
Encyonema neogracile Krammer ● ● ●
Encyonema perpusillum var. chilense Krammer, Rumrich et L.-Bert. ● ● ● ●
Eunotia bilunaris (Ehr.) Mills * ● ● ● ● ●
Eunotia exigua (Bréb.) Rabenhorst * ● ● ● ● ●
Eunotia flexuosa (Bréb.) Kütz. ● ● ● ●
Eunotia aff. gracillima (Krasske) Nörpel ● ● ●
Continuación de la Tabla 3.

RESULTADOS
43
Taxones RH1 RH2 RH3 RH4 RH5
Eunotia intermedia (Krasske) Nörpel et L.-B. * ● ● ● ● ●
Eunotia minor (Kütz.) Grun. * ● ● ● ● ●
Eunotia monodon Ehr. ● ● ●
Eunotia muscicola Krasske ●
Eunotia naegelii Migula * ● ● ● ● ●
Eunotia veneris (Kütz.) De Toni * ● ● ● ● ●
Fragilaria capucina Desmazieres morpho 1 ● ●
Fragilaria capucina Desmazieres morpho 2 ● ●
Fragilaria exigua Grun. ● ●
Fragilaria germainii Reichardt et L.-B. ●
Frustulia rhomboides var. rhomboides (Ehr.) De Toni ● ● ● ●
Frustulia rhomboides var. saxonica (Rabenh.) De Toni * ● ● ● ● ●
Frustulia rhomboides var. saxonica fo. capitata (Mayer) Patrick * ● ● ● ● ●
Frustulia rhomboides var. saxonica fo. undulata Hust. ● ● ●
Gomphonema acuminatum var. coronatum (Ehr.) Smith ●
Gomphonema exilissimum (Grun.) L.-B. et Reichardt ● ● ●
Gomphonema gracile Ehr. ●
Gomphonema aff. subclavatum (Grun.) Grun. ●
Gomphonema truncatum var. capitatum (Ehr.) Grun. ● ●
Gomphonema sp. ●
Kobayasiella sp. ● ● ● ●
Microstatus sp. ● ●
Navicula cf. radiosa Kütz. ●
Neidium cf. affine (Ehr.) Pfitzer ● ● ●
Neidium ampliatum (Ehr.) Krammer ● ●
Neidium sp. ● ●
Nitzschia aff. fonticola Grun. ●
Nitzschia gracilis Hantzsch ● ● ●
Nitzschia palea (Kütz.) W. Smith ● ● ● ●
Pinnularia borealis Ehr. ● ●
Pinnularia divergens var. decrescens (Grun.) Krammer ● ● ● ●
Pinnularia mesolepta (Ehr.) W. Smith morpho 1 ● ● ● ●
Pinnularia microstauron (Ehr.) Cl. * ● ● ● ● ●
Pinnularia neomajor Krammer ● ● ●
Pinnularia obscura Krasske ●
Pinnularia aff. subcapitata Gregory ● ●
Pinnularia subgibba Krammer ● ● ● ●
Pinnularia viridiformis Krammer ● ● ●
Continuación de la Tabla 3.

RESULTADOS
44
Taxones RH1 RH2 RH3 RH4 RH5
Sellaphora laevissima Kütz. ●
Stauroneis phoenicenteron (Nitzsch) Ehr. ● ● ● ●
Staurosira venter (Ehr.) Cl. et Möller ●
Staurosirella aff. pinnata Ehr. ● ●
Stenopterobia intermedia (Lewis) V.H. ● ●
Surirella linearis var. linearis W. Sm. ● ● ●
Surirella linearis var. constricta (Ehr.) Grun. ● ●
Surirella cf. pseudolinearis Krasske ●
Surirella sp. ●
Synedra acus Kütz. ● ●
Tabellaria flocculosa (Roth) Kütz. ● ● ● ●
Subtotal 46 32 15 59 25
Chrysophyceae
Dinobryon sertularia Ehr. * ● ● ● ● ●
Mallomonas sp.1 ● ● ● ●
Mallomonas sp.2 ● ● ●
Salpingoeca sp. ●
Synura sp. ● ● ● ●
Chrysophyceae 1 ● ●
Chrysophyceae 2 ● ●
Chrysophyceae 3 ●
Chrysophyceae 4 ● ●
Chrysophyceae 5 ● ● ●
Chrysophyceae 6 ●
Chrysophyceae 7 ●
Chrysophyceae cysts ● ● ● ● ●
Subtotal 11 7 5 5 6
Dinophyceae
Hemidinium nasutum Stein ●
Peridinium centenniale * ● ● ● ● ●
Peridinium inconspicuum Lemm. * ● ● ● ● ●
Peridinium wierzejskii Woloszynka * ● ● ● ● ●
Peridinium willei Hutfeld-Kaas * ● ● ● ● ●
cf. Amphidium sp. ●
Subtotal 5 5 4 4 4
Continuación de la Tabla 3.

RESULTADOS
45
Taxones RH1 RH2 RH3 RH4 RH5
Tribophyceae
Pseudostaurastrum cf. lobulatum (Pasch.) Fott ● ● ●
Subtotal 1 1 0 1 0
Chlorophyceae
Actinochloris sphaerica Kors. ● ● ●
Ankistrodesmus falcatus (Corda) Ralfs ●
Ankistrodesmus fasciculatus (Lundb.) Kom.-Legn. ● ●
Ankistrodesmus fusiformis Corda ● ● ● ●
Ankyra judayi (G.M.Smith) Fott ●
Binuclearia cf. tectorum (Kütz.) Berger * ● ● ● ● ●
Botryococcus braunii Kütz. * ● ● ● ● ●
Chlamydomonas sp. 1 ●
Chlamydomonas sp. 2 ●
Chlamydomonas sp. 3 ●
Chlamydomonas sp. 4 ●
Chlamydomonas sp. 5 ●
Chlamydomonas sp. 6 ●
Chlamydomonas spp. ● ● ● ●
Chloromonas angustissima (Ettl) Gerloff ● ●
Chloromonas sp. ●
Coelastrum indicum Turn. ● ● ●
Coenochloris planconvexa Hind. ●
Coenochloris sphagnicola Hind. ●
Coenocystis subcylindrica Kors. ●
Dictyochlorella globosa (Kors.) Silva ●
Dictyosphaerium ehrenbergianum Näg. ●
Dictyosphaerium pulchellum Wood ● ●
Dictyosphaerium sphagnale Hind. ●
Enallax cf. alpinus Pasch. ● ● ●
Enallax coelastroides (Bohl.) Skuja ●
Eutetramorus fottii (Hind.) Kom. ● ● ●
Fusola cf. viridis Snow ●
Geminella sp. ●
Kirchneriella irregularis var. irregularis (G.M.Smith) Kors. ● ●
Kirchneriella irregularis var. spiralis Kors. ●
Kirchneriella microscopica Nyg. * ● ● ● ● ●
Korshikoviella michailovskoensis (Elenk.) Silva ● ●
Continuación de la Tabla 3.

RESULTADOS
46
Taxones RH1 RH2 RH3 RH4 RH5
Lobocystis planctonica (Tiff. et Ahlstr.) Fott ●
Microspora palustris Wichm. * ● ● ● ● ●
Monoraphidium griffithii (Berk.) Kom.-Legn. ● ●
Monoraphidium minutum (Näg.) Kom.-Legn. ●
Oocystis lacustris Chod. ● ● ●
Oocystis submarina Lagerh. ● ● ●
Pandorina morum (O.F.Müller) Bory ●
Pediastrum angulosum (Ehr.) ex Menegh. ●
Polyedriopsis cf. spinulosa (Schmidle) Schmidle ●
Quadrigula closterioides (Bohl.) Printz ● ● ● ●
Quadrigula lacustris (Chod.) G.M.Smith ● ●
Quadrigula sp. ● ●
Saturnella saturna (Steinecke) Fott ● ●
Scenedesmus brevispina (G.M.Smith) Chod. ● ● ● ●
Scenedesmus cf. dispar (Bréb.) Rabenh. ●
Scenedesmus ecornis (Ehr.) Chod. ● ● ● ●
Scenedesmus heimii Bourr. ● ● ●
Scenedesmus obliquus (Turp.) Kütz. ●
Scenedesmus quadrispina Chod. ● ● ●
Scenedesmus serratus (Corda) Bohl. * ● ● ● ● ●
Scenedesmus cf. spinosus Chod. ● ●
Scenedesmus sp. ● ●
Schroederia sp. ●
Sphaerellocystis sp. ●
Sphaerocystis cf. bilobata Broady ● ● ●
Stigeoclonium sp. ● ●
Treubaria setigera (Arch.) G.M.Smith ●
Chlorophyceae 1 ●
Chlorophyceae 2 ●
Chlorophyceae 3 ● ● ●
Chlorophyceae 4 ●
Chlorophyceae 5 ●
Chlorophyceae 6 ●
Chlorophyceae 7 ●
Oedogoniales ● ● ● ● ●
Subtotal 35 22 20 43 19
Continuación de la Tabla 3.
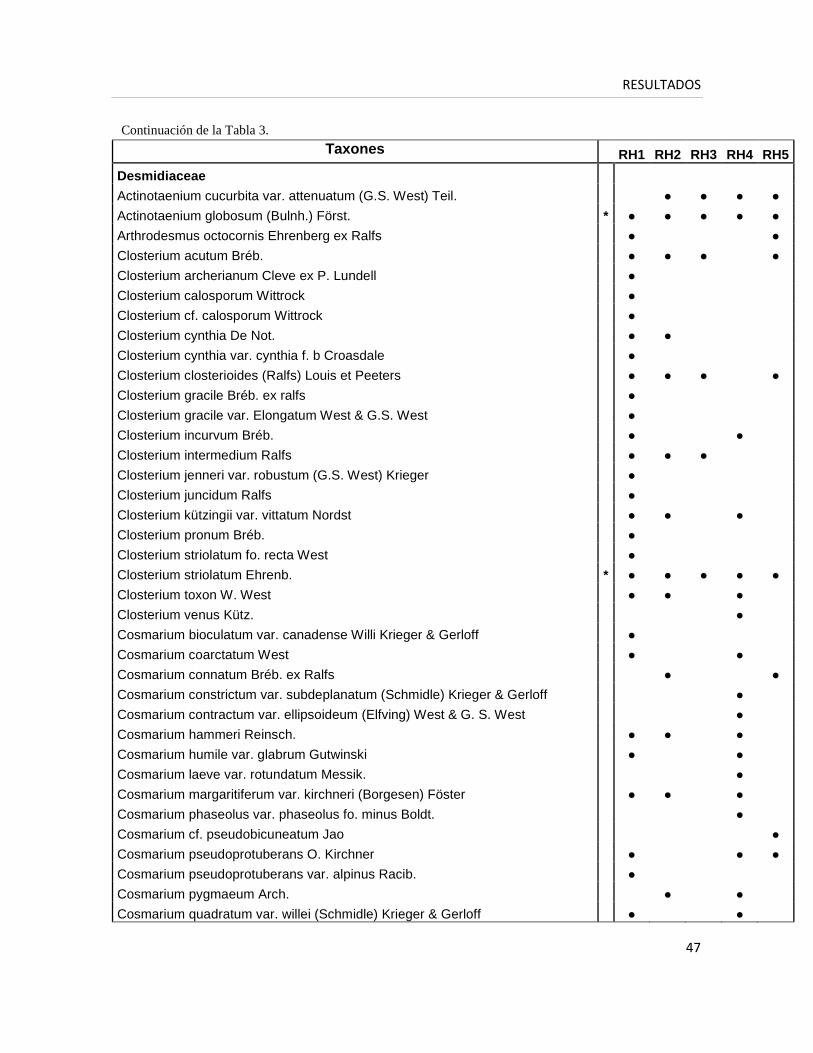
RESULTADOS
47
Taxones RH1 RH2 RH3 RH4 RH5
Desmidiaceae
Actinotaenium cucurbita var. attenuatum (G.S. West) Teil. ● ● ● ●
Actinotaenium globosum (Bulnh.) Först. * ● ● ● ● ●
Arthrodesmus octocornis Ehrenberg ex Ralfs ● ●
Closterium acutum Bréb. ● ● ● ●
Closterium archerianum Cleve ex P. Lundell ●
Closterium calosporum Wittrock ●
Closterium cf. calosporum Wittrock ●
Closterium cynthia De Not. ● ●
Closterium cynthia var. cynthia f. b Croasdale ●
Closterium closterioides (Ralfs) Louis et Peeters ● ● ● ●
Closterium gracile Bréb. ex ralfs ●
Closterium gracile var. Elongatum West & G.S. West ●
Closterium incurvum Bréb. ● ●
Closterium intermedium Ralfs ● ● ●
Closterium jenneri var. robustum (G.S. West) Krieger ●
Closterium juncidum Ralfs ●
Closterium kützingii var. vittatum Nordst ● ● ●
Closterium pronum Bréb. ●
Closterium striolatum fo. recta West ●
Closterium striolatum Ehrenb. * ● ● ● ● ●
Closterium toxon W. West ● ● ●
Closterium venus Kütz. ●
Cosmarium bioculatum var. canadense Willi Krieger & Gerloff ●
Cosmarium coarctatum West ● ●
Cosmarium connatum Bréb. ex Ralfs ● ●
Cosmarium constrictum var. subdeplanatum (Schmidle) Krieger & Gerloff ●
Cosmarium contractum var. ellipsoideum (Elfving) West & G. S. West ●
Cosmarium hammeri Reinsch. ● ● ●
Cosmarium humile var. glabrum Gutwinski ● ●
Cosmarium laeve var. rotundatum Messik. ●
Cosmarium margaritiferum var. kirchneri (Borgesen) Föster ● ● ●
Cosmarium phaseolus var. phaseolus fo. minus Boldt. ●
Cosmarium cf. pseudobicuneatum Jao ●
Cosmarium pseudoprotuberans O. Kirchner ● ● ●
Cosmarium pseudoprotuberans var. alpinus Racib. ●
Cosmarium pygmaeum Arch. ● ●
Cosmarium quadratum var. willei (Schmidle) Krieger & Gerloff ● ●
Continuación de la Tabla 3.

RESULTADOS
48
Taxones RH1 RH2 RH3 RH4 RH5
Cosmarium quadrifarium Lundeil ● ● ● ●
Cosmarium regnesii var. regnesii Reinsch. ●
Cosmarium regulare Schmidle ● ● ●
Cosmarium sinostegos var. obtusius Gutwinski ● ● ●
Cosmarium subtumidum Nordst. ● ●
Cosmarium trilobulatum fo. retusum Gutwinski ●
Cosmarium truncatellum Perty ● Cosmarium venustum var. excavatum (B. Eichler & Gutwinski) West & G. S. West ● ●
Cosmarium cf. venustum fo. minor (Wille) West & G. S. West ● ●
Cosmarium sp. 1 ● ●
Cosmarium sp. 2 ●
Cylindrocystis brebissonii var. brebissonii Menegh. * ● ● ● ● ●
Cylindrocystis brebissonii var. jenneri (Ralfs) Hansgirg ● ●
Euastrum attenuatum var. lithuanicum f. pulchellum Prescott & Scott ● ●
Euastrum binale (Turp.) Ehrenberg ● ●
Euastrum insulare var. insulare (Wittr.)Roy * ● ● ● ● ●
Euastrum obesum var. obesum Josh ● ● ● ●
Euastrum obesum var. trapezicum (Börg.)Krieg. * ● ● ● ● ●
Euastrum sphyroides Nordst. ●
Euastrum sp. ● ●
Gonatozygon pilosum Wolle ● ●
Hyalotheca dissiliens var. dissiliens (Smith) Bréb. ● ● ●
Micrasterias radiosa Ralfs ●
Mougeotia sp. ● ● ●
Netrium digitus var. digitus (Ehr.) Hzigs et Rothe ● ●
Netrium sp. ● ●
cf. Octacanthium bifidum (Bréb.) P. Compère ●
Penium cylindrus Bréb. ex Ralfs ● ●
Penium phymatosporum Nordst. ● ●
Sphaerozosma vertebratum Bréb. ex Ralfs ●
Staurastrum borgeanum fo. minus Schm. ● ●
Staurastrum crenulatum (Näg.) Delp. ● ● ● ●
Staurastrum floriferum W. & G. S. West ●
Staurastrum cf. gracile var. coronulatum Boldt. ●
Staurastrum inconspicuum Nordst. ●
Staurastrum iotanum Wolle ● ● ● ●
Staurastrum laeve W. West ● ● ●
Staurastrum lapponicum Grönbl. ● ●
Continuación de la Tabla 3.

RESULTADOS
49
Taxones RH1 RH2 RH3 RH4 RH5
Staurastrum margaritaceum (Ehr.) Menegh. ● ● ●
Staurastrum muricatum Bréb. ex Ralfs ● ●
Staurastrum paradoxum Meyen ● ● ● ●
Staurastrum polytrichum (Perty) Rabenh. * ● ● ● ● ●
Staurastrum subavicula West et West ● ●
Staurastrum subnudibrachiatum West et West ● ● ● ●
Staurastrum tetracerum Ralfs ● ●
Staurastrum sp. sensu Lenzenweger 1993 ● ● ● ●
Staurodesmus connatus (P. Lundell) Thomasson ● ●
Staurodesmus convergens var. convergens Teil. ● ● ●
Staurodesmus crassus (West & G.S. West) M. B. Florin * ● ● ● ● ●
Staurodesmus cuspidatus var. cuspidatus (Bréb) Teil. ● ● ● ●
Staurodesmus dejectus (Bréb.) Teil. ● ●
Staurodesmus dickiei (Ralfs) S. Lillieroth ●
Staurodesmus extensus var. joshuae (Gutwinski) Teil. ● ● ●
Staurodesmus extensus var. vulgaris (Eichle et Racib.) Croas. * ● ● ● ● ●
Staurodesmus mamillatus (Nordst.) Teil. ● ●
Staurodesmus cf. mamillatus ● ●
Staurodesmus patens (Nordst.) Croas. * ● ● ● ● ●
Staurodesmus phimus var. robustus Teil. ●
Staurodesmus quiriferus Teil. ● ● ●
Staurodesmus triangularis (Largerheim) Teil. ● ● ● ●
Staurodesmus sp. ●
Teilingia excavata (Ralfs ex Ralfs) Bourr. ● ● ●
Teilingia granulata (Roy et Biss.) Bourr. ● ● ● ●
Tetmemorus brebissonii (Menegh.) Ralfs ● ● ● ●
Tetmemorus granulatus (Bréb.) Ralfs ● ● ●
Xanthidium smithii Arch. * ● ● ● ● ●
Subtotal 79 47 26 65 32
Euglenophyta
Euglena sp. ●
Phacus onyx Pochm. ● ●
Phacus polytrophos Pochm. ● ●
Phacus pyrum (Ehr.) Stein ●
Phacus swirenkoi Skv. ●
Trachelomonas lacustris var. ovalis Drez. emend. Defl. ● ●
Trachelomonas volvocina var. punctata Playf. ● ●
Continuación de la Tabla 3.

RESULTADOS
50
Taxones RH1 RH2 RH3 RH4 RH5
Subtotal 4 0 1 6 0
Cryptophyceae
Cryptomonas sp. ●
Cryptophyceae 1 ●
Subtotal 1 0 0 1 0
Rhodophyta
Batrachospermum sp. ● ● ● ●
Subtotal 0 1 1 1 1
Total 215 141 98 217 116
Las especies más “ubicuas”, presentes en todos los cuerpos de agua, pertenecieron a
los grupos Cyanobacteria (18 taxones), Zygnematophyceae (10 taxones) y
Bacillariophyceae (9 taxones). En especial, las Zygnematophyceae y las Bacillariophyceae
realizaron un gran aporte a la riqueza total de los ambientes estudiados contribuyendo con
más del 42% de los taxones. Estos dos grupos, junto con las Chlorophyceae, revelaron una
mayor cantidad de taxones en los ambientes grandes y profundos, y conectados
hidrológicamente (RH1 y RH4) (Tabla 3). Contrariamente, los sistemas más ácidos (RH2,
RH3 y RH5) fueron menos ricos en especies, particularmente los más pequeños (Figura
20).
Continuación de la Tabla 3.

RESULTADOS
51
Figura 20. Porcentajes relativos de la riqueza en cada cuerpo de agua respecto al total de especies relevadas
en la turbera de Rancho Hambre.
A través de la matriz de presencia/ausencia de especies en los distintos cuerpos de
agua se realizó un análisis de agrupamiento utilizando el índice de similitud de Jaccard (J).
Tanto RH1 como RH4 mostraron composiciones florísticas características y constantes. Por
otro lado, los cuerpos de agua someros (RH3 y RH5) fueron similares entre sí en
composición y poco variables a lo largo del período de estudio, mientras que la
composición florística de RH2 sufrió una fuerte variación temporal (Figura 21).

RESULTADOS
52
Figura 21. Análisis de agrupamiento basado en la matriz de presencia-ausencia de especies. Referencias: O:
octubre, D: diciembre, F: febrero, A: abril, mes-año-sitio, por ejemplo O-08-RH1.
En cuanto al “pool de especies”, término que se refiere al total de especies
registradas durante todo el estudio en cada cuerpo de agua, en la Tabla 4 se muestra el
resultado del cálculo del índice de Jaccard para todas las combinaciones posibles de
cuerpos de agua. La similitud en “pool de especies” en los cinco cuerpos de agua varió
entre 0,31 y 0,54, observándose los mayores índices de similitud entre RH1 y RH4 por un
lado y en los ambientes someros (RH3 y RH5) por el otro. A pesar de esto, en general los

RESULTADOS
53
índices registrados fueron más bajos que los esperados para sistemas tan cercanos,
indicando que la flora fitoplanctónica fue bastante característica de cada sistema.
RH1 RH2 RH3 RH4 RH5RH1 1RH2 0,50 1RH3 0,36 0,48 1RH4 0,54 0,40 0,31 1RH5 0,38 0,49 0,54 0,35 1
Tabla 4. Valores del índice de Jaccard entre pares de cuerpos de agua considerando la sumatoria de
todos los muestreos en cada sistema.
b- Abundancia, biovolumen y diversidad
En los sistemas profundos (RH1, RH2 y RH4), la densidad y el biovolumen del
nano- y microfitoplancton no mostraron diferencias significativas (ANOVA p>0,05; en
todos los casos) entre los distintos puntos de muestreo dentro de cada cuerpo de agua. Por
esta razón se calcularon valores promedio para cada fecha de muestreo en estos sistemas.
La variación temporal de la densidad del nano- y microfitoplancton (fracción > 2
µm) se describió a partir de las especies dominantes, definiendo como tales a aquellas que
representaron en algún cuerpo de agua más del 40% de la densidad total. La abundancia de
esta fracción varió entre 2,1 103 (octubre 2008) y 2,5 104 (diciembre 2009) ind mL-1. Los
valores de abundancia promedio en los cuerpos de agua profundos (RH1, RH2 y RH4)
variaron dentro del mismo orden de magnitud (4,93 103―7,64 103 ind mL-1) mientras que
los sistemas someros superaron a éstos en un orden de magnitud (1,13 104―1,38 104 ind
mL-1).
A lo largo del período de estudio, la distribución de la abundancia fue generalmente
unimodal con máximos en verano (diciembre o febrero) (Figura 22). Las variaciones de la
abundancia de esta fracción del fitoplancton en RH1 y RH2, estuvieron principalmente
causadas por una pequeña Chrysophyceae flagelada (cf. Ochromonas sp. 2). Por su parte,
los sistemas someros mostraron picos de abundancia debidos a cf. Plagioselmis sp. 2 y cf.
Ochromonas sp. 2. La abundancia de RH4 fue muy estable durante el primer año, mientras

RESULTADOS
54
que en el segundo mostró un pico de abundancia de la especie Kirchneriella microscopica
Nyg. (Figura 22).
Figura 22. Variación temporal de las abundancias de las especies dominantes del fitoplancton en cada cuerpo
de agua.
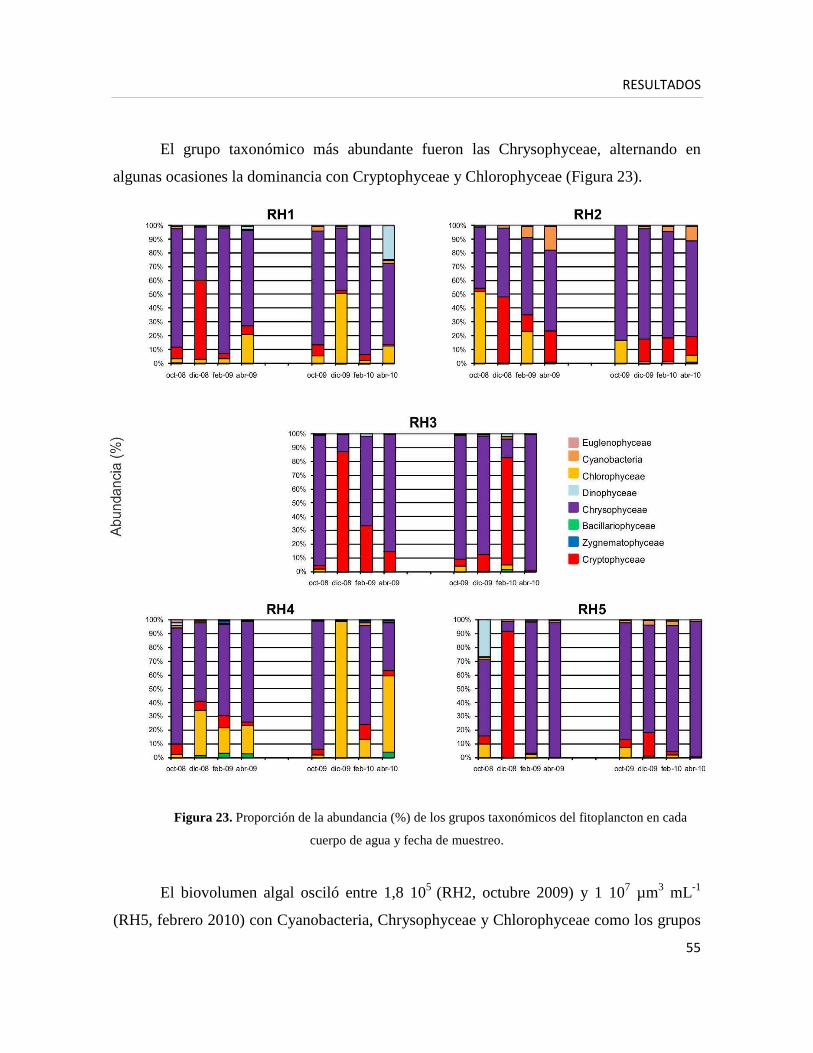
RESULTADOS
55
El grupo taxonómico más abundante fueron las Chrysophyceae, alternando en
algunas ocasiones la dominancia con Cryptophyceae y Chlorophyceae (Figura 23).
Figura 23. Proporción de la abundancia (%) de los grupos taxonómicos del fitoplancton en cada
cuerpo de agua y fecha de muestreo.
El biovolumen algal osciló entre 1,8 105 (RH2, octubre 2009) y 1 107 µm3 mL-1
(RH5, febrero 2010) con Cyanobacteria, Chrysophyceae y Chlorophyceae como los grupos

RESULTADOS
56
mayormente representados. En general, para todos los cuerpos de agua el biovolumen
promedio varió dentro del mismo orden de magnitud (2,0 106―2,8 106 µm3 mL-1) salvo en
la laguna RH2 donde se encontró un orden por debajo (9,23 105 µm3 mL-1). A pesar de que
la densidad de Cyanobacteria fue baja durante el estudio, el gran tamaño de las colonias
mucilaginosas tuvo como resultado una importante contribución al biovolumen (10―95 %)
(Figuras 24 y 25).
Figura 24. Variación temporal del biovolumen de los grupos taxonómicos del nano- y microfitoplancton en
cada cuerpo de agua y fecha de muestreo.

RESULTADOS
57
Figura 25. Proporción del biovolumen (%) de los grupos taxonómicos del fitoplancton en cada
cuerpo de agua y fecha de muestreo.
Las variaciones del biovolumen del nano- y microfitoplancton en RH1 estuvieron
causadas por el aporte de los diferentes grupos taxonómicos que dominaron en diferentes
ocasiones (Cryptophyceae, Cyanobacteria y Zygnematophyceae). En RH4, la
Chlorophyceae Kirchneriella microscopica realizó un aporte de hasta el 80% al
biovolumen. Ambos sistemas mostraron una composición similar luego del
descongelamiento de los cuerpos de agua (octubre) siendo posteriormente dominados por
diferentes grupos taxonómicos. Las variaciones en RH2 estuvieron principalmente dadas

RESULTADOS
58
por Aphanothece minutissima (W.West) Komárkova-Legnerová et Cronberg, con aportes
mayores al 30%. En los sistemas someros, RH3 y RH5, se observaron grandes
contribuciones al biovolumen de Chrysophyceae (cf. Ochromonas spp.) dominando
exclusivamente los muestreos de otoño, y de Cyanobacteria (Aphanothece minutissima). En
particular, RH5 mostró el mayor aporte de Dinophyceae (Peridinium inconspicuum Lemm.
y Peridinium willei Huitfeld-Kaas) en relación al resto de los sistemas estudiados (Figura
25).
Tanto la diversidad (índice de Shannon-Weaver) como la equitatividad para todos
los cuerpos de agua durante el período de estudio variaron en forma similar (Figuras 26 y
27). La diversidad promedio para todos los sistemas estudiados fue de 1,38 con los valores
más bajos registrados en diciembre (media=1,0) y los máximos en octubre y abril
(media=1,62). Los sistemas someros (RH3 y RH5) mostraron grandes variaciones
temporales con máximos en primavera (Octubre) y mínimos en verano asociados con una
menor equitatividad dada por la dominancia de Cryptophyceae (Figura 26 B). En cambio,
los sistemas profundos (RH1, RH2 y RH4) fueron más estables en el primer año, RH1
mostró una mayor diversidad en abril mientras que en RH2 y RH4 se observó en febrero
(Figura 26 A).
Figura 26. Diversidad (índice de Shannon- Weaver) en los distintos cuerpos de agua: A- Variación temporal
de la diversidad en los cuerpos de agua profundos (RH1, RH2 y RH4) y B- en los cuerpos de agua someros
(RH3 y RH5), registrada durante el período de estudio.

RESULTADOS
59
El cuerpo de agua más profundo (RH2) demostró ser el sistema más equitativo
(valor promedio ERH2= 0,67) en relación al resto de los cuerpos de agua estudiados (valor
promedio: ERH1= 0,53, ERH3= 0,54, ERH4= 0,55 y ERH5= 0,53) (Figura 27).
Figura 27. Equitatividad en los diferentes cuerpos de agua en cada fecha de muestreo. A- Variación temporal
de la equitatividad en los cuerpos de agua profundos (RH1, RH2 y RH4) y B- en los cuerpos de agua someros
(RH3 y RH5), registrada durante el período de estudio.
• Picofitoplancton
Tanto la abundancia como el biovolumen de la comunidad picofitoplanctónica
(fracción <2 µm) no mostraron diferencias significativas (ANOVA p>0,05) entre los
diferentes puntos de muestreo en los cuerpos de agua profundos (RH1, RH2 y RH4). Por
esta razón, para el análisis de esta fracción se consideraron valores promedio por cuerpo de
agua.
El picofitoplancton estuvo exclusivamente compuesto por organismos eucariotas,
distinguiéndose dos morfos, uno de ellos característico de los sistemas someros y otro de
los profundos (Figura 28).
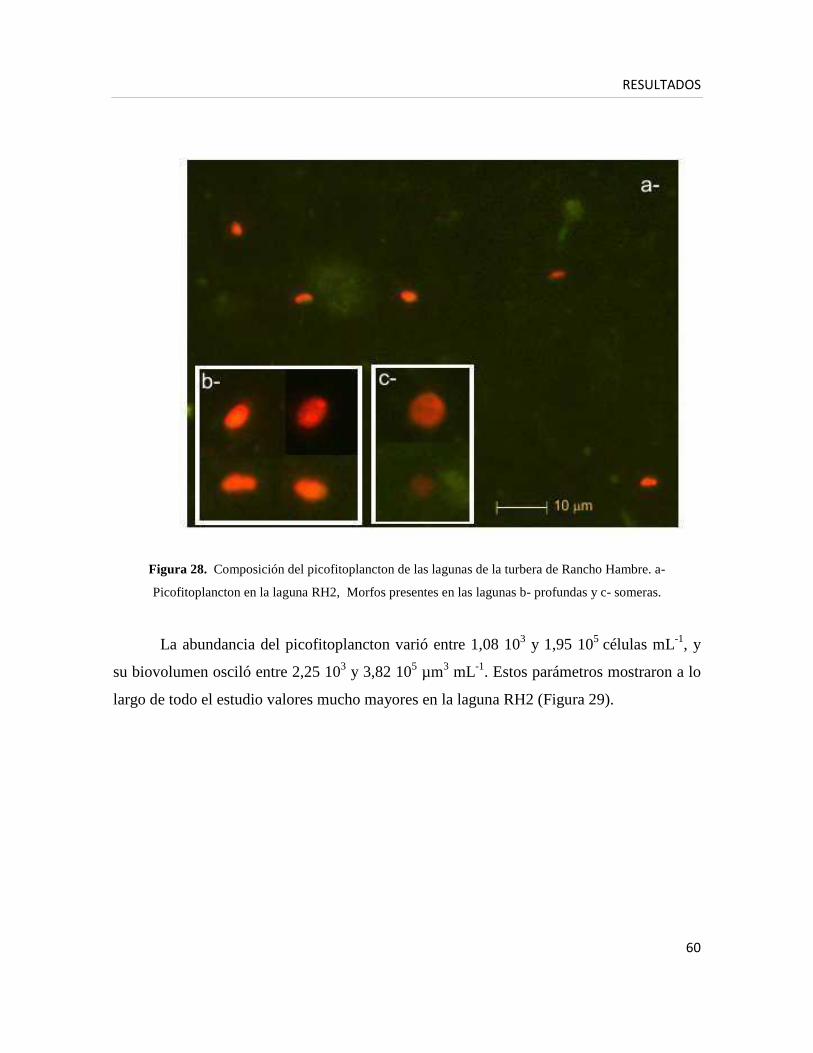
RESULTADOS
60
Figura 28. Composición del picofitoplancton de las lagunas de la turbera de Rancho Hambre. a-
Picofitoplancton en la laguna RH2, Morfos presentes en las lagunas b- profundas y c- someras.
La abundancia del picofitoplancton varió entre 1,08 103 y 1,95 105 células mL-1, y
su biovolumen osciló entre 2,25 103 y 3,82 105 µm3 mL-1. Estos parámetros mostraron a lo
largo de todo el estudio valores mucho mayores en la laguna RH2 (Figura 29).

RESULTADOS
61
Figura 29. Variación temporal de la abundancia (a) y del biovolumen (b) del picofitoplancton durante el
período de estudio. c- Variación del volumen celular promedio del picoplancton autotrófico entre octubre de
2008 y abril de 2009.

RESULTADOS
62
El volumen celular promedio estimado durante el primer año de muestreo
(2008―2009) mostró en RH1 y RH4 los coeficientes de variación más bajos (19 y 18%
respectivamente) en comparación con el resto de los sistemas analizados (CVRH2=28,
CVRH3=43 y CVRH5= 38%). En general, los volúmenes celulares promedio variaron
estacionalmente y mostraron diferentes tendencias en cada cuerpo de agua. El mayor valor
promedio se registró en RH3 (4,5 µm3) mientras que dentro de los sistemas profundos, RH2
mostró diferencias significativas en volumen celular con RH1 y RH4 a pesar de mostrar
igual morfología (p 2-1=0,036; p 2-4=0,01) (Figura 29).
• Fracciones de tamaño del fitoplancton
Las proporciones de las distintas fracciones de tamaño del fitoplancton mostraron
variaciones estacionales, las que fueron consistentes en ambos períodos de estudio (Figuras
30 y 31). En términos de abundancia, se observaron dos patrones de variación: mientras que
en RH2 y RH4 el picofitoplancton mostró un incremento en su proporción a lo largo del
tiempo, en RH1, RH3 y RH5 esta fracción tendió a disminuir su aporte durante el verano.
El picofitoplancton (porcentajes entre 45 y 95% en RH2 y RH4) y el nanofitoplancton
(porcentajes entre 40 y 90% en RH1, RH3 y RH5) fueron las fracciones que mostraron la
mayor contribución a la densidad total. En particular, el picofitoplancton realizó un aporte
significativo al biovolumen total en los sistemas más profundos (RH2 y RH4) (hasta un
40%) mientras que en general, el microfitoplancton (fracción >20 µm) representó más del
50% del biovolumen total en casi todos los sitios y fechas de muestreo (Figuras 30 y 31).

RESULTADOS
63
Figura 30. Proporciones de las distintas fracciones de tamaño del fitoplancton en relación a la abundancia y
el biovolumen totales en RH1, RH2 y RH3.

RESULTADOS
64
Figura 31. Proporciones de las distintas fracciones de tamaño del fitoplancton en relación a la abundancia y
el biovolumen totales en RH4 y RH5.
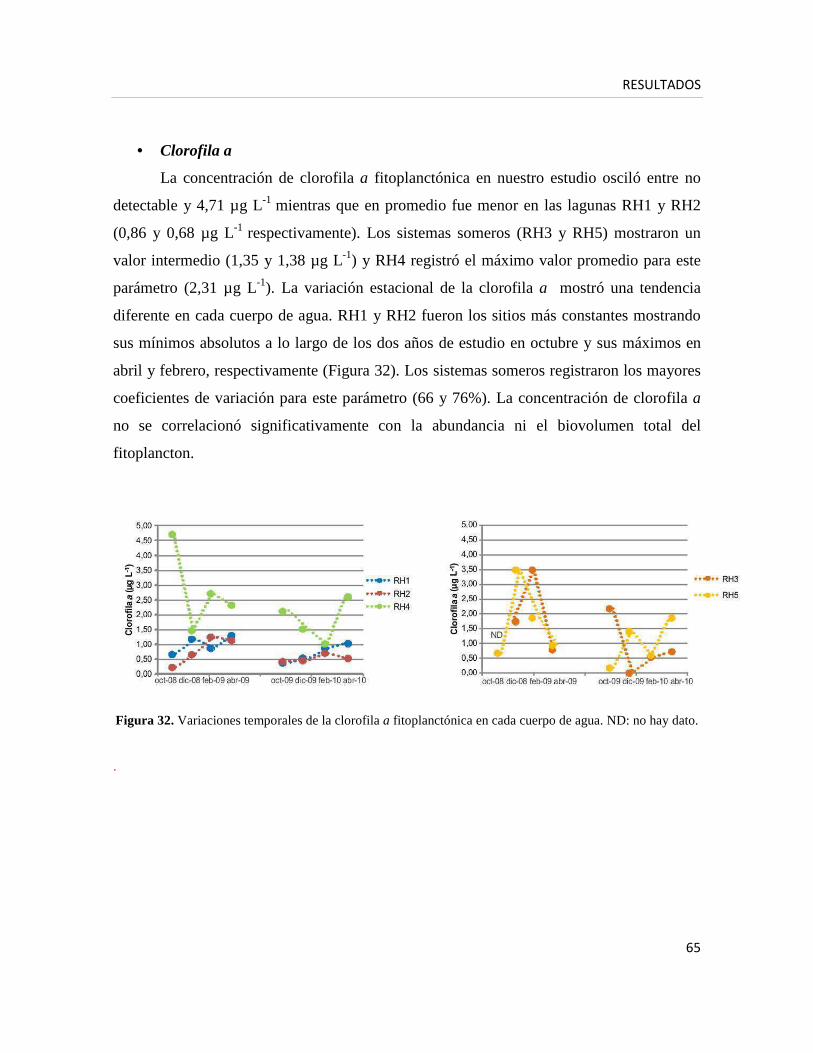
RESULTADOS
65
• Clorofila a
La concentración de clorofila a fitoplanctónica en nuestro estudio osciló entre no
detectable y 4,71 µg L-1 mientras que en promedio fue menor en las lagunas RH1 y RH2
(0,86 y 0,68 µg L-1 respectivamente). Los sistemas someros (RH3 y RH5) mostraron un
valor intermedio (1,35 y 1,38 µg L-1) y RH4 registró el máximo valor promedio para este
parámetro (2,31 µg L-1). La variación estacional de la clorofila a mostró una tendencia
diferente en cada cuerpo de agua. RH1 y RH2 fueron los sitios más constantes mostrando
sus mínimos absolutos a lo largo de los dos años de estudio en octubre y sus máximos en
abril y febrero, respectivamente (Figura 32). Los sistemas someros registraron los mayores
coeficientes de variación para este parámetro (66 y 76%). La concentración de clorofila a
no se correlacionó significativamente con la abundancia ni el biovolumen total del
fitoplancton.
Figura 32. Variaciones temporales de la clorofila a fitoplanctónica en cada cuerpo de agua. ND: no hay dato.
.

RESULTADOS
66
• Estrategias nutricionales
Los organismos mixótrofos, representados principalmente por las Chrysophyceae
nanoflageladas cf. Ochromonas spp., dominaron numéricamente todos los cuerpos de agua
luego de su descongelamiento (> 50% de la abundancia total). Esta dominancia continuó en
los cuerpos de agua someros (RH3 y RH5) y en RH1 a lo largo del verano. En cambio,
RH2 y RH4 estuvieron dominados por el picofitoplancton autotrófico (Figura 33 a).
Figura 33-a. Proporción de mixótrofos y autótrofos (%) en función de la abundancia total del fitoplancton
incluyendo todas las fracciones de tamaño.
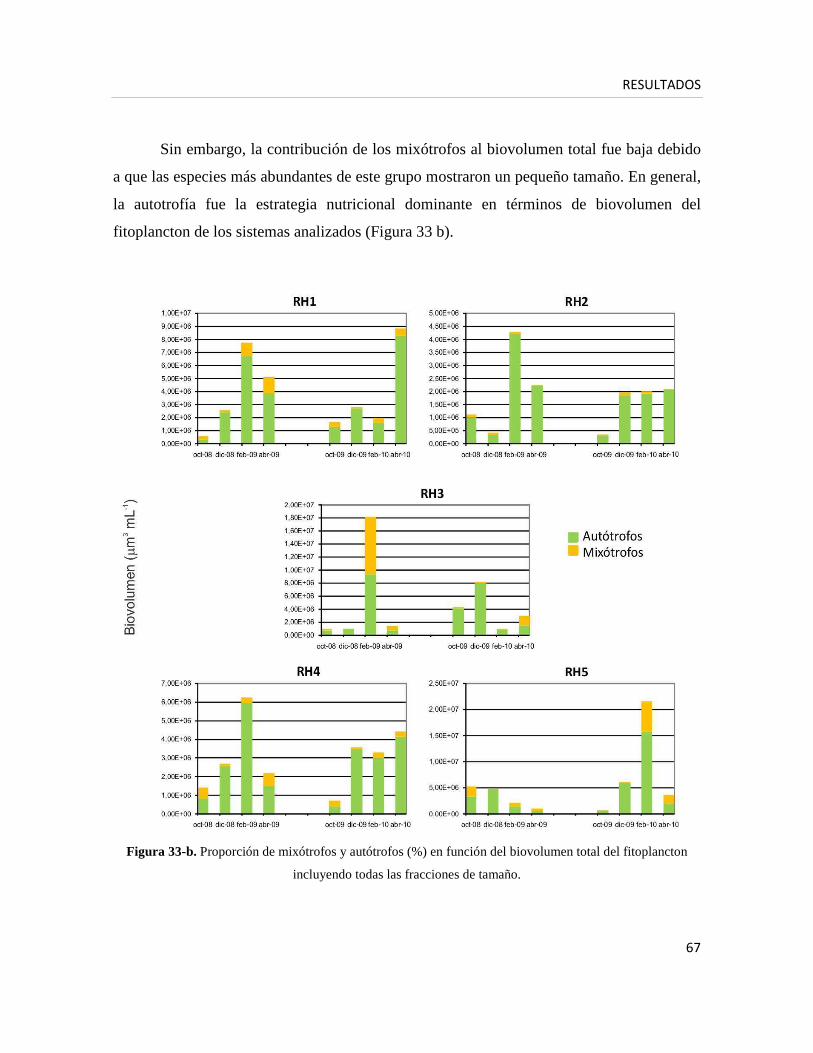
RESULTADOS
67
Sin embargo, la contribución de los mixótrofos al biovolumen total fue baja debido
a que las especies más abundantes de este grupo mostraron un pequeño tamaño. En general,
la autotrofía fue la estrategia nutricional dominante en términos de biovolumen del
fitoplancton de los sistemas analizados (Figura 33 b).
Figura 33-b. Proporción de mixótrofos y autótrofos (%) en función del biovolumen total del fitoplancton
incluyendo todas las fracciones de tamaño.

RESULTADOS
68
3) Análisis de los posibles predadores de la comunidad fitoplanctónica.
Como se mencionó anteriormente, esta tesis doctoral forma parte de un proyecto de
investigación que tuvo como objetivo el estudio limnológico integral de las lagunas de la
turbera de Rancho Hambre, en el cual diferentes investigadores se especializaron en el
estudio de las distintas comunidades planctónicas. Con el objetivo de poder relacionar las
distintas fracciones de tamaño del fitoplancton con sus posibles predadores en los cinco
cuerpos de agua estudiados, se calcularon las abundancias totales (para los nanoflagelados
heterotróficos) y de las distintas especies y grupos funcionales identificados (para ciliados y
metazooplancton). Las abundancias de los nanoflagelados heterotróficos y del proto- y
metazooplancton revelaron una distribución espacial homogénea, en la que no mostraron
diferencias significativas (ANOVA p>0,05) entre los distintos puntos de muestreo en los
cuerpos de agua profundos (RH1, RH2 y RH4). Por esta razón, se calcularon valores
promedio por cuerpo de agua.
a- Nanoflagelados heterotróficos (NFH)
El tamaño de los NFH relevados en los cinco cuerpos de agua fue menor a 5 µm. La
abundancia promedio de estos organismos para todos los sistemas fue de 5,4 103 ind mL-1
oscilando entre 1,3 103 y 3,4 104 ind mL-1 (Quiroga et al. en prep.). La abundancia de los
NFH no se correlacionó significativamente con ninguna de las fracciones de tamaño del
fitoplancton. Según estudios previos de predación, los NFH menores a 5�m consumen
principalmente picocianobacterias y picoplancton heterotrófico (Sherr & Sherr 1991, Sherr
et al. 1991, Stockner 1991, Weisse 1993). Dado estos antecedentes y nuestros resultados
decidimos excluir a estos organismos del posterior análisis multivariado.
b- Protozooplancton (ciliados)
En este estudio se relevaron 125 especies de ciliados, con Balanion planctonicum
(Foissner, Oleksiv & Müller) Foissner, Berger & Kohmann, Cyclidium sp.,
Pelagostrombidium fallax (Zacharias) Krainer, Rimostrombidium sp., Halteria grandinella

RESULTADOS
69
(Müller) Dujardin, Urotricha sp.1 y Urotricha sp.2 como las más abundantes en estos
sistemas (Küppers et al. en prep.). De acuerdo a la bibliografía existente, todas las especies
mencionadas pueden consumir tanto bacterias como algas.
La densidad de los ciliados mostró un rango de variación para el sistema de 3 102―
2 105 ind L-1. Las lagunas profundas e hidrológicamente conectadas (RH1 y RH4) fueron
similares en composición y comportamiento y mostraron sus máximos de abundancia en
abril, mientras que el resto de los sistemas estudiados (RH2, RH3 y RH5) mostraron picos
de dominancia de distintas especies y en diferentes momentos (Figura 34).
Figura 34. Variación temporal de las abundancias de las especies dominantes de ciliados en cada cuerpo de
agua.

RESULTADOS
70
c- Metazooplancton (rotíferos y crustáceos)
En el transcurso de este estudio se registró un total de 72 taxones. Los organismos
dominantes fueron los rotíferos Conochilus unicornis Rousselet, Keratella valdiviensis
Thomasson y Polyathra dolichoptera Idelson, los cladóceros Alona spp., Bosmina chilensis
Daday, Ceriodaphnia cf. dubia Richard, Chydorus sp., Daphnia commutata Ekman y
Pleuroxus sp., y los copépodos Boeckella poppei Mrázek y Tropocyclops prasinus
meridionalis Kiefer (Marinone & Menu Marque en prep.).
La densidad de rotíferos y crustáceos osciló entre 0 y 6,8 103 ind L-1. En general, los
Rotifera dominaron los cuerpos de agua estudiados salvo en la laguna RH4, dominada por
los crustáceos Cladocera y Copepoda. La abundancia total del metazooplancton varió
estacionalmente, mostrando dos tipos de comportamiento interanual diferentes, uno
asociado a los sistemas profundos (RH1, RH2 y RH4) y otro a los someros (RH3 y RH5)
(Figura 35).

RESULTADOS
71
Figura 35. Variación temporal de la abundancia de los grupos taxonómicos dominantes del metazooplancton
(rotíferos y crustáceos).

RESULTADOS
72
4) Relación entre los factores bióticos y abióticos y los distintos descriptores de la
comunidad fitoplanctónica
Los descriptores de la comunidad fitoplanctónica en los cuerpos de agua estudiados
(abundancia, biomasa y diversidad) se relacionaron con los diferentes parámetros bióticos y
abióticos medidos. La abundancia del nano- y microfitoplancton varió estacionalmente y
como consecuencia estuvo correlacionada con la temperatura (r=0,52 p=0,01). A su vez,
este último parámetro se correlacionó negativamente con la diversidad (r= -0,502 p=0,01) y
la equitatividad (r= -0,504 p=0,01), asociado con la dominancia de algunas pocas especies.
Por su parte, el pH explicó las variaciones de la biomasa fitoplanctónica (estimada a través
de la clorofila a) y de la riqueza específica a través de una regresión lineal significativa (β=
0,20 p= 0,021; β=16,39 p< 0,0001 respectivamente). Las abundancias de las distintas
fracciones de tamaño del fitoplancton se asociaron a las densidades del proto- y
metazooplancton. Por un lado, las abundancias del picofitoplancton eucariota y del ciliado
Halteria grandinella (Müller) Dujardin se correlacionaron negativamente (r=-0,366
p=0,05), debido principalmente al comportamiento de ambos organismos en la laguna RH2
(rRH2= -0,846 p=0,01). Por el otro, para el resto de los cuerpos de agua, la abundancia del
picofitoplancton eucariota se correlacionó positivamente con las densidades de los
copépodos filtradores (cof) (Boeckella poppei Mrázek) (r=0,50 p=0,01).

RESULTADOS
73
5) Análisis de los Grupos Funcionales Basados en características Morfológicas (GFBM)
sensu Kruk et al. (2010) y sus posibles predadores.
A partir del análisis cuantitativo del fitoplancton se registraron 137 especies que
fueron clasificadas en Grupos Funcionales Basados en características Morfológicas
(GFBM) sensu Kruk et al. (2010) (Tabla 5). Este análisis se realizó con el objetivo de
identificar los grupos de especies con rasgos morfo-funcionales comunes y similares que
responden a las distintas condiciones ambientales establecidas en los cinco cuerpos de agua
estudiados. En general, se observó una gran riqueza morfo-funcional en todos los cuerpos
de agua debido a la ocurrencia simultánea de todos los GFBM. El análisis de la Tabla 5
reveló que algunos de los organismos pequeños (grupo I, representado principalmente por
el picofitoplancton) y los flagelados pequeños (grupo II) mostraron una mayor constancia
que el resto de los GFBM, apareciendo en la mayoría de los muestreos en todos los cuerpos
de agua. Por su parte, los flagelados grandes (grupo V) revelaron una alta fidelidad a los
sistemas profundos con entradas y/o salidas superficiales de agua (RH1 y RH4). El grupo
VII (colonias mucilaginosas) se asoció principalmente a RH4. Mientras que los organismos
con exoesqueleto silíceo (grupo VI) mostraron una mayor constancia pero con una baja
frecuencia en los ocho muestreos en RH1, los individuos de tamaño medio sin estructuras
especializadas (que no contienen flagelados, aerotopos, mucílago ni exoesqueleto silíceo)
(grupo IV) lo hicieron en RH4.

RESULTADOS
74
Tabla 5. Lista florística de cada laguna. GFBM (Grupo Funcional Basado en las características
Morfológicas sensu Kruk et al. 2010), especies registradas entre 0- 25% ( ), 25-50% ( ), 50-75% ( ) y 75-
100%( ) de los 8 muestreos realizados en cada cuerpo de agua. I: organismos pequeños, II: flagelados
pequeños, II: grandes filamentos con aerotopos, IV: organismos de tamaño medio sin estructuras
especializadas, V: flagelados de tamaño medio a grande, VI: organismos con exoesqueleto silíceo, VII:
grandes colonias mucilaginosas.
Taxones GFBM RH1 RH2 RH3 RH4 RH5
cf. Cyanobium sp. I
Kirchneriella irregularis var. irregularis (G.M.Smith) Kors. I
Kirchneriella irregularis var. spiralis Kors. I
Kirchneriella microscopica Nyg. I
Schroederia sp. I
Treubaria setigera (Arch.) G.M.Smith I
Dictyochlophyceae I
Filamentos de cyanobacteria I
Picofitoplancton eucariota I
Dinobryon sertularia Ehr. II
Mallomonas akrokomos Ruttner II
Mallomonas sp. 1 II
Mallomonas sp. 2 II
cf. Ochromonas sp. 1 II
cf. Ochromonas sp. 2 II
cf. Ochromonas sp. 3 II
Chrysophyceae 1 II
Chrysophyceae 3 II
Chrysophyceae 4 II
Chrysophyceae 5 II
Chrysophyceae 6 II
Chrysophyceae 7 II
Chrysophyceae 9 II
Chrysophyceae 10 II
Quistes de Chrysophyceae II
Anabaena sp. 2 III
Actinochloris sphaerica Kors. IV
Actinotaenium globosum (Bulnh.) Förster IV
Ankistrodesmus fasciculatus (Lundb.) Kom.-Legn. IV
Ankistrodesmus fusiformis Corda IV
Ankyra judayi (G.M.Smith) Fott IV

RESULTADOS
75
Taxones GFBM RH1 RH2 RH3 RH4 RH5
Arthrodesmus octocornis Ehr. IV
Closterium incurvum Bréb. IV
Closterium kützinguii var. vittatum Nordst. IV
Closterium striolatum Ehr. IV
Closterium venus Kütz. IV
Cosmarium bioculatum var. depressum (Schaarschmidt) Schmidle IV
Cosmarium hammeri Reinsch IV
Cosmarium pseudoprotuberans Kirchn. IV
Cosmarium pygmaeum Archer IV
Cosmarium quadrifarium Lundell IV
Cosmarium regulare Schmidle IV
Cosmarium trilobulatum fo. retusum Gutw. IV
Cosmarium venestum var. excavatum (Eichl. et Gutw.) W. et G.S.West IV
Cylindrocystis brebissonii var. brebissonii Menegh. IV
Cylindrocystis brebissonii var. jenneri (Ralfs) Hansgirg IV
Cylindrospenium sp. IV
Euastrum insulare (Wittrock) Roy IV
Euastrum obesum Josh IV
Gonatozygon pilosum Wolle IV
Korshikoviella michailovskoensis (Elenk.) Silva IV
Monoraphidium arcuatum (Kors.) Hind. IV
Monoraphidium contortum (Thur.) Kom.-Legn. IV
Monoraphidium griffithii (Berk.) Kom.-Legn. IV
Monoraphidium sp. IV
Leptolyngbya crassior (Skuja) Anag. IV
Phormidium chlorinum (Kütz.) Anag. IV
Pseudanabaena catenata Lauterborn IV
Quadrigula closterioides (Bohl.) Printz IV
Quadrigula lacustris (Chod.) G.M.Smith IV
Scenedesmus brevispina (G.M.Smith) Chod. IV
Scenedesmus heimii Bourr. IV
Scenedesmus serratus (Corda) Bohl. IV
Scenedesmus cf. spinosus Chod. IV
Staurastrum crenulatum (Näg.) Delponte IV
Staurastrum paradoxum Meyen IV
Staurodesmus extensus var. vulgaris (Eichl. et Raciborski) Croasdale IV
Staurodesmus cuspidatus (Bréb.) Teil. IV
Staurodesmus crassus (W. et G.S.West) Florin IV
Continuación de la Tabla 5.

RESULTADOS
76
Taxones GFBM RH1 RH2 RH3 RH4 RH5
Staurodesmus phimus var. robustus Teil. IV
Staurodesmus triangularis (Lagerh.) Teil. IV
Teilingia excavata (Ralfs) Bourr. IV
Teilingia granulata (Roy et Bisset) Bourr. IV
Tetmemorus granulatus (Bréb.) Ralfs IV
Chlamydomonas sp. 1 V
Chlamydomonas sp. 2 V
Chlamydomonas sp. 5 V
Chloromonas angustissima (Ettl) Gerloff V
Chloromonas sp. V
Cryptomonas cf. tatrica Czosnowski V
cf. Amphidium sp. V
Gloeodinium montanum Klebs V
Hemidinium nasutum Stein V
Peridinium centenniale (Playfair) Lefèvre V
Peridinium inconspicuum Lemm. V
Peridinium umbonatum Stein V
Peridinium wierzeyskii Woloszynka V
Peridinium willei Hutfeld-Kaas V
Phacus polytrophos Pochm. V
cf. Plagioselmis sp. 1 V
cf. Plagioselmis sp. 2 V
cf. Plagioselmis sp. 3 V
cf. Plagioselmis sp. V
Vitreochlamys sp. V
Trachelomonas volvocina var. punctata Playf. V
Cryptophyceae 1 V
Brachysira cf. intermedia (Oestrup) L.-Bert. VI
Encyonema neogracile Krammer VI
Eunotia exigua (Bréb.) Rabenhorst VI
Eunotia flexuosa (Bréb.) Kütz. VI
Eunotia aff. gracillima (Krasske) Nörpel VI
Eunotia intermedia (Krasske) Nörpel et L.-Bert. VI
Eunotia naegelii Migula VI
Eunotia veneris (Kütz.) De Toni VI
Fragilaria capucina Desmazieres VI
Microstatus sp. VI
Neidiun cf. affine (Ehr.) Pfitzer VI
Continuación de la Tabla 5.

RESULTADOS
77
Taxones GFBM RH1 RH2 RH3 RH4 RH5
Nitzschia aff. fonticola Grun. VI
Nitzschia gracilis Hantzsch VI
Nitzschia palea (Kütz.) W.Smith VI
Pinnularia microstauron (Ehr.) Cl. VI
Pinnularia subgibba Krammer VI
Stenopterobia intermedia (Lewis) V.H. VI
Tabellaria flocculosa (Roth) Kütz. VI
Aphanocapsa elachista W. et G.S.West VII
Aphanocapsa incerta (Lemm.) Cronberg et Kom. VII
Aphanocapsa planctonica (G.M.Smith) Kom. et Anag. VII
Aphanothece minutissima (W.West) Kom.-Legn. et Cronberg VII
Aphanothece nidulans Richter VII
Botryococcus braunii Kütz. VII
cf. Coelosphaeirum sp. VII
Chroococcus minimus (Keissler) Lemm. VII
Chroococcus minor (Kütz.) Näg. VII
Chroococcus turgidus (Kütz.) Näg. VII
Coenocystis subcylindrica Kors. VII
Cyanodictyon reticulatum (Lemm.) Geitler VII
Dictyochlorella globosa (Kors.) Silva VII
Dictyosphaerium chlorelloides (Naum.) Kom. et Perm. VII
Dictyosphaerium pulchellum Wood VII
Dictyosphaerium sp. 2 VII
cf. Dictyosphaerium sp. VII
Gloecapsopsis pleurocapsoides (Novácek) Kom. et Anag. VII
Merismopedia elegans Braun VII
Merismopedia punctata Meyen VII
Merismopedia tenuissima Lemm. VII
Lobocystis planctonica (Tiff. et Ahlstr.) Fott VII
Rhabdogloea linearis (Geitler) Kom. VII
Sphaerocystis cf. bilobata Broady VII
La abundancia de los GFBM en RH1, RH3 y RH5 estuvo principalmente
representada por los grupos I, II y V, mientras que en RH2 y RH4 los pequeños organismos
(grupo I, representado principalmente por picofitoplancton eucariota y Kirchneriella
microscopica) fueron el grupo morfo-funcional dominante. A su vez, los sistemas
Continuación de la Tabla 5.

RESULTADOS
78
estudiados mostraron en general diferentes comportamientos temporales de los GFBM
excepto en los sistemas someros donde las variaciones fueron similares (Figura 36).
Figura 36. Variación temporal de la abundancia de los GFBM. I: pequeños organismos; II: pequeños
flagelados; III: grandes filamentos con aerotopos; IV: organismos de tamaño medio sin aerotopos, flagelos,
heterocistos, mucílago ni exoesqueleto silíceo; V: flagelados de tamaño medio a grande; VI: organismos con
exoesqueleto silíceo; VII: grandes colonias mucilaginosas.

RESULTADOS
79
Por su parte, la variación temporal del biovolumen de los GFBM mostró un similar
comportamiento en RH1 y RH4 con la presencia del grupo IV. RH2 estuvo claramente
dominado por las grandes colonias mucilaginosas (grupo VII). Los sistemas someros
evidenciaron diferentes comportamientos y estuvieron dominados por distintos grupos
(grupo II en RH3 y grupos II, V y VII en RH5) (Figura 37).
Figura 37. Variación temporal del biovolumen de los GFBM. I: pequeños organismos; II: pequeños
flagelados; III: grandes filamentos con aerotopos; IV: organismos de tamaño medio sin aerotopos, flagelos,
heterocistos, mucílago ni exoesqueleto silíceo; V: flagelados de tamaño medio a grande; VI: organismos con
exoesqueleto silíceo; VII: grandes colonias mucilaginosas.

RESULTADOS
80
A partir de los resultados del análisis de correlación entre los descriptores
(abundancia y biovolumen) de los distintos grupos morfo-funcionales del fitoplancton y las
abundancias del proto- y metazooplancton (agrupados en gremios alimenticios, Anexo 1),
observamos que en cada cuerpo de agua las relaciones entre los GFBM y sus potenciales
predadores fueron diferentes. La falta de correlaciones entre parámetros permite afirmar
que las relaciones entre las distintas comunidades son complejas y que dependen de varios
factores, lo que justifica el posterior análisis multivariado.
6) Relación entre los Grupos Funcionales Basados en características Morfológicas
(GFBM) y los factores bióticos y abióticos.
Las Figuras 38 y 39 muestran el ordenamiento resultante del análisis de
correspondencia canónica (CCA) tanto de los grupos morfo-funcionales del fitoplancton
como de las muestras, en base a la composición del biovolumen de los GFBM y a los
parámetros bióticos y abióticos. El porcentaje de varianza de la relación especie-ambiente
explicada por los dos primeros ejes fue del 82%. Tanto el test de significancia de Monte
Carlo del primer eje canónico como el del conjunto de los ejes arrojaron valores de p<
0,002. Las variables con mayor influencia sobre el eje 1 (horizontal) fueron la abundancia
de ciliados (coeficiente de correlación: -0,59) y el pH (coeficiente de correlación: 0,46);
mientras que la variable asociada en mayor medida al eje 2 (vertical) fue la abundancia de
macrofiltradores (principalmente representado por el cladócero Daphnia commutata)
(coeficiente de correlación: 0,55). El grupo I (pequeños organismos) se asoció a bajas
abundancias de macrofiltradores y menor pH. Los pequeños flagelados (grupo II) se
asociaron fuertemente con altas abundancias de ciliados. El grupo III (grandes filamentos
con aerotopos, representado por una única especie en nuestro estudio), el IV (organismos
de tamaño medio sin estructuras especializadas) y el VI (diatomeas) se asociaron a las
muestras de verano y otoño en ambientes con mayor pH y presencia de macrofiltradores
(RH1 y RH4). Por último, los muestreos de verano en los ambientes más someros, ácidos y

RESULTADOS
81
con altas abundancias de fósforo total, microfiltradores y copépodos filtradores (Boeckella
poppei) se caracterizaron por los grupos V y VII.
Figura 38. Resultado del Análisis de Correspondencia Canónica basado en el biovolumen de los grupos
morfo-funcionales de fitoplancton y los parámetros bióticos (flechas en verde) y abióticos (flechas en rojo).
NT: nitrógeno total; PT: fósforo total.

RESULTADOS
82
Figura 39. Ordenamiento de los sitios en función de los parámetros bióticos (fechas en verde) y abióticos
(flechas en rojo) a partir de un análisis multivariado (CCA). NT: nitrógeno total; PT: fósforo total. Las
muestras del período 2009-2010(segundo año) se indican con un asterisco (*).
En general, el ordenamiento de las muestras de los diferentes cuerpos de agua en el
CCA estuvo determinado por la combinación entre los factores bióticos y abióticos,
diferenciándose claramente dos grupos de cuerpos de agua. En la parte superior derecha del
diagrama se ubican las muestras de RH1 y RH4 con presencia de macrofiltradores
(principalmente Daphnia commutata) y mayores valores de pH, y donde se encuentran
organismos del fitoplancton de tamaño medio sin estructuras especializadas o con
exoesqueleto silíceo (grupos IV y VI). En cambio, en la parte inferior del diagrama se
localizan la mayoría de las muestras (RH2, RH3 y RH5) con bajo pH y altas abundancias

RESULTADOS
83
de fósforo total y del resto de los gremios alimenticios del metazooplancton
(microfiltradores, predadores tope, copépodos filtradores y copépodos macrófagos
herbívoros), donde las características morfo-funcionales dominantes del fitoplancton son
grandes colonias mucilaginosas (grupo VII) y grandes organismos flagelados (grupos V).

DISCUSIÓN

DISCUSIÓN
84
DISCUSIÓN
Análisis de los factores ambientales
La naturaleza de la turbera depende de la procedencia del agua que la recarga, la que
determina las características químicas de los cuerpos de agua (Holden 2006, Iturraspe
2010). Cuando las turberas son parcialmente alimentadas por la napa freática (condiciones
minerotróficas), el pH y la dureza tienden a valores relativamente elevados, mientras que en
las turberas en forma de domo que se alimentan solo por precipitación, las aguas
especialmente las que se encuentran entre los capítulos del musgo Sphagnum muestran
valores más bajos de pH y dureza (condiciones ombrotróficas) (Holden 2006, Rydin &
Jeglum 2006, Iturraspe 2010). Esto es, particularmente, debido a la gran capacidad de
intercambiador de cationes de este musgo, el cual captura cationes y libera protones (Clymo
1964, Roig & Roig 2004, Rydin & Jeglum 2006).
En una turbera ombrotrófica elevada de Sphagnum, Iturraspe & Roig (2000)
encontraron una disminución en el flujo de agua horizontal a pocos decímetros de
profundidad en el acrotelmo (capa superior de la turba) y que no hay flujo vertical a través
del catotelmo (capa inferior). En el caso de Rancho Hambre, esta fuerte restricción del flujo
sub-superficial hace que la microtopografía local sea un factor clave para la diversidad de
ambientes limnéticos dentro de una misma turbera a través de la regulación de la
conectividad hidrológica (presencia de entradas y/o salidas superficiales). Esto explicaría
las diferentes condiciones limnológicas de los cuerpos de agua que, si bien se hallan
cercanos, no evidencian interconexiones entre ellos y muestran diferente variabilidad en el
nivel hidrométrico, sugiriendo que pertenecen a diferentes sub-cuencas separadas por
domos de Sphagnum.
Mientras que el equilibrio entre la temperatura y la precipitación determinó en
general las fluctuaciones en los niveles hidrométricos observados en este estudio -
notablemente el descenso general durante la estación seca y cálida, diciembre 2009- las
diferencias en la conectividad hidrológica (presencia de entradas y/o salidas superficiales) y
el tamaño explicaron los distintos patrones de fluctuación de los niveles hidrométricos entre

DISCUSIÓN
85
los cuerpos de agua: el de mayor superficie (RH4) tuvo el nivel hidrométrico más estable,
mientras que el más profundo pero aislado hidrológicamente (RH2) fue el más variable.
La influencia del tamaño sobre los parámetros físicos y químicos se evidenció
también a partir de la temperatura del agua, ya que el cuerpo de agua pequeño mostró no
sólo mayores valores de temperatura sino también una variación térmica más amplia. Por lo
tanto, la morfometría es un factor fundamental que influye en la hidrología de los cuerpos
de agua y en los parámetros físico-químicos que dependen de la temperatura.
Los valores bajos de pH y dureza total revelaron el carácter ombrotrófico de los
cuerpos de agua someros (RH3 y RH5), mientras que los grandes, hidrológicamente
conectados (RH1 y RH4) fueron más minerotróficos (altos valores de dureza y pH). Estas
diferencias morfométricas e hidrológicas explican la heterogeneidad limnológica
encontrada bajo condiciones meteorológicas idénticas. A partir de lo mencionado
anteriormente, se verificó la hipótesis planteada de que las características morfométricas e
hidrológicas de los cuerpos de agua determinan gran parte de las propiedades físicas y
químicas de sus aguas.
Un recurso esencial para el fitoplancton es la luz (Litchman & Klausmeier 2008).
En los ambientes acuáticos, la luz es atenuada por las sustancias disueltas y las partículas
suspendidas, incluyendo al fitoplancton, resultando en un gradiente vertical en intensidad y
distribución espectral (Kirk 1994). Los coeficientes de atenuación vertical de la luz (Kd)
medidos en los cuerpos de agua profundos (RH1, RH2 y RH4) de este estudio se
encontraron dentro del rango de variación de aquellos estimados en una laguna de turbera
de Tierra del Fuego (laguna Victoria) (Tell 2011, Saad 2011) y en lagos húmicos (Lindell et
al. 1996). Este parámetro varió estacionalmente y mostró diferentes tendencias en cada
sistema. Sin embargo, según la profundidad del disco de Secchi, la luz llegó siempre hasta
el fondo de cada cuerpo de agua. Esta no limitación explicaría la falta de correlación entre
el Kd y la concentración de clorofila a fitoplanctónica que podría haberse esperado en los
cuerpos de agua profundos. Sin embargo, la producción primaria en estos cuerpos de agua
de turberas posiblemente se encuentre limitada por luz en el invierno, momento en que los
sistemas están cubiertos por una capa de hielo y nieve.

DISCUSIÓN
86
El carbono orgánico disuelto coloreado (CODC) es de gran importancia en los
sistemas acuáticos (Klug 2002). El CODC se encuentra compuesto principalmente por
sustancias húmicas que determinan, por ejemplo en lagos húmicos, su clásico color
caramelo. El CODC es un importante regulador del crecimiento del fitoplancton autotrófico
ya que influye sobre la disponibilidad de luz y de nutrientes -a través de la formación de
complejos principalmente con el fósforo biodisponible para el fitoplancton- (Jones 1992,
Hessen & Tranvik 1998).
A partir de nuestros resultados, observamos que los distintos estimadores de
sustancias húmicas (A254, a320 y a440) estuvieron altamente correlacionados con las
concentraciones de COD (carbono orgánico disuelto) y sus variaciones temporales se
correlacionaron significativamente con la temperatura. A su vez, A254 y a320 fueron los
mejores estimadores de la concentración de COD en los sistemas estudiados, ya que
mostraron los mayores coeficientes de correlación.
En cuanto a las concentraciones de COD, el rango de variación medido en nuestro
estudio (2,8―13,4 mg L-1) incluyó los valores registrados para la laguna Victoria, sistema
acuático asociado a turberas de la provincia de Tierra del Fuego (Tell et al. 2011, Saad
2011) (4,6―10,8 mg L-1). A su vez, las concentraciones de COD y los contenidos de
sustancias húmicas (A254= 0,093―0,497) se encontraron dentro de los valores mínimos
reportados por Jasser et al. (2009) en dos lagos de Polonia con contenidos húmicos
contrastantes (oligo y polihúmico) (COD=7,1―21,0 mg L-1, A254= 0,4331―2,331)
sugiriendo que los sistemas acuáticos de turberas estudiados podrían clasificarse como
ambientes oligohúmicos. Por su parte, el rango de variación de uno los estimadores de
sustancias húmicas del agua empleados en este estudio (a440= 1,38―8,75 m-1) fue mayor
al reportado por Davies- Colley & Vant (1987) para tres lagos de turberas de Nueva
Zelanda (a440= 1,26―4,87 m-1) y los obtenidos por Huovinen et al. (2003) para lagos
húmicos finlandeses (a440= 0,63―6,75 m-1). Las variaciones temporales del nivel
hidrométrico, reflejadas por la temperatura, y las diferencias morfométricas entre los
cuerpos de agua, la que determina la influencia de la cuenca sobre los sistemas, explicaron
el amplio rango de valores encontrado aquí, con las mayores concentraciones de COD y

DISCUSIÓN
87
sustancias húmicas en los sistemas someros y pequeños. A su vez, en estos cuerpos de agua
se registraron las mayores abundancias del nano- y microfitoplancton, lo que posiblemente
se relaciona con las propiedades de absorción del COD, las que pueden favorecer al
fitoplancton actuando como una protección a la radiación UV como ocurre en lagos
húmicos (Kirk 1994, Lindell et al. 1996, Huovinen et al. 2003). Por otra parte, Górniak et
al. (2003) registraron en un embalse con aportes de COD provenientes de una turbera
minerotrófica, un aumento en las concentraciones de COD junto con un incremento en la
concentración de clorofila a mostrando que las sustancias húmicas estimulan el desarrollo
del fitoplancton (Grabowska et al. 2003). A pesar de que en las capas más profundas de la
mayoría de los lagos la alta concentración de COD es perjudicial debido a que disminuye la
luz disponible para la fotosíntesis (Jones 1992, Carpenter et al. 1998), ese fenómeno no
ocurre en estos cuerpos de agua someros y bien mezclados, en los que la luz llega hasta el
fondo.
La concentración de nutrientes es otro factor que puede ser fundamental en la
regulación de la producción del fitoplancton (así como también la biomasa de todas las
comunidades) (Tilman et al. 1982) y de sus cambios anuales e internuales (Padisák &
Dokulil 1994, Sommer et al. 1986, Reynolds 1997). Los bajos contenidos de nutrientes
disueltos encontrados en general coinciden con los reportados para otras turberas del
mundo, desde los lagos de turberas de Hungría (Borics et al. 2003) hasta los de pequeñas
turberas tropicales de Bhutan (Sharma & Bhattari 2005). La variación temporal de los
nutrientes en un mismo cuerpo de agua durante todo el estudio superó largamente a las
diferencias entre los cuerpos de agua para un mismo muestreo. Estos resultados nos
permiten rechazar parte de la hipótesis planteada, en la cual se propone que las diferencias
en concentraciones de nutrientes son determinantes de las diferencias en la composición y
estructura del fitoplancton entre los cuerpos de agua estudiados.
Los rangos de concentraciones de NT excedieron a los de PT por 1 ó 2 órdenes de
magnitud, en concordancia con los resultados obtenidos por Mataloni (1999) para seis
turberas de Tierra del Fuego. Según Downing & McCauley (1992), la relación NT: PT
refleja la fuente de nutrientes del sistema, con altas proporciones caracterizando ambientes

DISCUSIÓN
88
naturales en cuencas no perturbadas, en las cuales se exporta más nitrógeno que fósforo.
Esto explicaría que los valores más bajos se hayan registrado luego del descongelamiento
primaveral (por lo tanto reflejando las “condiciones internas iniciales” de cada sistema), así
como el posterior incremento en los cuerpos de agua por la alimentación de la cuenca y las
precipitaciones, siendo ambas más ricas en nitrógeno que en fósforo. A su vez, estas altas
relaciones fueron mayores a 22,6 (por peso) ó 50 (molar) lo que reflejarían una limitación
por fósforo para la biomasa del fitoplancton (Guildford & Hecky 2000). En cuanto a la
proporción de NID: PO4-P, su valor máximo fue aproximadamente la mitad de la
proporción de Redfield, lo cual junto con los bajos valores de nutrientes disueltos indica
que la disponibilidad de NID podría afectar al crecimiento algal. Por lo tanto, la cuestión de
si alguno de estos elementos es efectivamente limitante para la producción autotrófica se
debe abordar de forma experimental. Mientras tanto, la diferencia entre las relaciones N: P
totales y disueltos hace que sea altamente conveniente continuar caracterizando las
condiciones de nutrientes de estos sistemas a través de ambos cocientes.
Según Mataloni & Tell (1996) la conductividad fue uno de los parámetros que
explicó las diferencias en composición específica del fitoplancton entre cinco cuerpos de
agua de la turbera de Rancho Hambre. En nuestro estudio, el rango de variación de la
conductividad medido en esta misma turbera (8,7―82,0 µS cm-1) fue mayor al registrado
por estos autores (12,6―22,3 µS cm-1) en un único muestreo. El amplio rango de variación
obtenido aquí se debe a variaciones temporales más que a las espaciales. En este sentido, el
rango de variación entre los cuerpos de agua en una misma fecha de muestreo no fue
determinante para reflejar cambios en la composición y estructura de las comunidades
fitoplanctónicas lo que nos permite rechazar la hipótesis planteada en la cual se postuló que
la conductividad determina las diferencias en composición y estructura del fitoplancton
entre los cinco cuerpos de agua estudiados.
Las relaciones establecidas entre las variables meteorológicas, hidrológicas,
morfométricas y limnológicas nos permitieron proponer un modelo interpretativo para la
caracterización limnológica de estos ambientes, que se proyecta validar en otros sistemas
de turberas fueguinas. Según este modelo, la topografía determinó en gran medida la

DISCUSIÓN
89
conectividad superficial hidrológica, la que influyó a su vez en el estado
minero/ombrotrófico de los cuerpos de agua a través de la existencia de fuentes de agua
adicionales al aporte directo de la lluvia. La morfometría reveló su influencia en los
sistemas someros, donde las pérdidas por evaporación originaron amplias variaciones en la
temperatura del agua, y la mayor influencia de la cuenca se manifestó en concentraciones
más altas y variables de carbono orgánico disuelto. El efecto del tiempo actuó a dos escalas
diferentes: mientras que un número de características abióticas tales como las
concentraciones de fosfato y nitrógeno total mostraron cambios estacionales, nuestros
resultados muestran que la variabilidad interanual de las condiciones meteorológicas tiene
la capacidad de regular la intensidad de dichos cambios.
Como resultado de esto, las propiedades limnológicas de estos cinco cuerpos de
agua de la turbera de Rancho Hambre revelaron una gran diversidad ambiental temporal y
espacial. Según Mataloni & Tell (1996), esta diversidad en las condiciones abióticas es
fundamental para la biodiversidad de las comunidades planctónicas de los cuerpos de agua
de la turbera de Rancho Hambre.
La comunidad fitoplanctónica
Diversos autores (Mataloni 1999, Borics et al. 2003, Krivograd Klemenčič et al.
2010) destacan que las condiciones particulares que se encuentran en los ecosistemas de
agua dulce de turberas (ambientes distróficos de aguas ácidas coloreadas) dan lugar a una
flora fitoplanctónica característica y muy diversa, que se ha estudiado alrededor del mundo
desde principios del siglo XX (Mataloni 1999). Las lagunas de turberas estudiadas aquí
revelaron una alta riqueza específica (305 taxones) coincidiendo con lo reportado por esta
autora. Los grupos mayormente representados fueron las Zygnematophyceae
(Desmidiaceae), Bacillariophyceae y Chlorophyceae, en concordancia con los estudios
realizados en cuerpos de agua de otras turberas del mundo (Nováková 2002, Borics et al.
2003, Rakowska & Sitkowska 2007, Krivogrand Klemenčič et al. 2010). Borics et al.
(2003) y Krivogrand Klemenčič et al. (2010) reportaron a las Zygnematophyceae

DISCUSIÓN
90
(Desmidiaceae) como el grupo con mayor cantidad de taxones coincidiendo con lo
registrado en este estudio. En cambio, Mataloni & Tell (1996), Mataloni (1999), Nováková
(2002) y Rakowska & Sitkowska (2007) muestran a las Bacillariophyceae como el grupo
de mayor riqueza. A pesar del gran aporte de las Desmideaceae a la riqueza específica total,
estas algas no dominaron cuantitativamente la comunidad fitoplanctónica de los sistemas
estudiados, al igual que ocurre en ambientes limnéticos de regiones templadas (Brook
1981). Este grupo taxonómico es mucho más importante cuantitativamente en lagos
tropicales que en sistemas acuáticos templados (Huszar & Reynolds 1997). Gesst & Coesel
(2012) atribuyeron este resultado al fenómeno de atelomixis, el cual involucra la mezcla de
la columna de agua durante el día debido a las diferencias de temperatura del aire entre el
día y la noche disminuyendo así las tasas de hundimiento de las células de desmidiáceas.
Lo postulado por estos autores no explica los resultados obtenidos en nuestro estudio, ya
que los cuerpos de agua estudiados son poco profundos (30 a 150 cm de profundidad) y se
encuentran sometidos a la acción de viento lo que promueve una mezcla continua de la
columna de agua. En el caso de las Bacillariophyceae, su baja abundancia posiblemente se
asocie, como ocurre en lagos húmicos, con una deficiencia de sílice disuelto (Arvola et al.
1999).
En nuestro estudio la mayoría de las Desmidiaceae y Bacillariophyceae prefirieron
ambientes grandes, profundos y de condiciones más minerotróficas confirmando resultados
previos obtenidos en esta misma turbera por Mataloni & Tell (1996) y con lo postulado
luego por Mataloni (1999), quien estableció que estas características ambientales antes que
la posición geográfica determinaron las diferencias en riqueza específica entre cuerpos de
agua de diferentes turberas fueguinas. Dentro de estos dos grupos taxonómicos, sólo veinte
especies (indicadas con un asterisco en la Tabla 3) se relevaron en todos los cuerpos de
agua. Las especies mencionadas son cosmopolitas, encontrándose presentes en turberas de
todo el mundo (Kingston 1982, Mataloni &Tell 1996, Mataloni 1999, Nováková 2002,
Negro et al. 2003, Rakowska & Sitkowska 2007, Štěpánková et al. 2008, Krivogrand
Klemenčič 2010, Rakowska & Sitkowska 2010).

DISCUSIÓN
91
La composición de especies en los sistemas grandes, profundos, conectados
hidrológicamente y de condiciones más minerotróficas (RH1 y RH4) fue distintiva y
constante a lo largo del tiempo, como demuestra el análisis de agrupamiento. Además, estos
cuerpos de agua fueron más ricos y mostraron una mayor similitud (Jaccard) entre sus pool
de especies. A pesar de que los sistemas estudiados se encuentran cercanos entre sí, la
similitud entre pool de especies en general fue baja destacando la gran diversidad ficológica
que albergan los cuerpos de agua y su importancia a la hora de relevar la biodiversidad de
ambientes de turberas (Mataloni 1997).
Uno de los factores más importantes que explica las variaciones en la riqueza
específica entre los cuerpos de agua de la turbera de Rancho Hambre es el pH, dato
revelado a través de una regresión lineal positiva entre ambos parámetros. Además, la
riqueza de especies estuvo positivamente correlacionada con el área de cada cuerpo de agua
coincidiendo con lo reportado por Mataloni & Tell (1996) para esta misma turbera. Por lo
tanto, corroboramos la hipótesis generada a partir de antecedentes previos en turberas de
Tierra del Fuego (Mataloni & Tell 1996, Mataloni 1999) en la cual se postula que el pH
influye en la composición de la comunidad fitoplanctónica.
Los descriptores de la comunidad fitoplanctónica (abundancia y biovolumen) no
mostraron diferencias significativas entre los distintos puntos de muestreo en los sistemas
profundos estudiados (p>0,05) debido principalmente a su escasa profundidad (profundidad
máxima registrada: 1,5 m) y al efecto del viento de dirección oeste que coincide con la
orientación del valle de Tierra Mayor en general y con los cuerpos de agua estudiados en
particular, lo que promueve la mezcla y consiguiente homogeneización de la columna de
agua.
La abundancia del nano- y microfitoplancton mostró un claro patrón estacional a
través de una alta correlación con la temperatura. Este último parámetro, se destacó como
un factor clave en la regulación de la dinámica de las comunidades fitoplanctónicas en
lagos húmicos de latitudes altas (Arvola et al.1999). Por su parte, las densidades de esta
fracción estuvieron dentro del mismo orden de magnitud que las reportadas por Mataloni
& Tell (1996) en cuerpos de agua de esta misma turbera comparando la misma estación de

DISCUSIÓN
92
muestreo. Las variaciones de la abundancia del fitoplancton >2�m fueron principalmente
causadas por Chrysophyceae, confirmando los resultados previos en dos cuerpos de agua
de la turbera de Rancho Hambre (Mataloni y Tell op.cit.) y con lo obtenido por Kuczyńska-
Kippen (2006) y Druvietis et al. (2010) en cuerpos de agua de turberas del hemisferio norte.
Dentro de este grupo taxonómico, la especie más abundante fue cf. Ochromonas sp 2., un
género con características pioneras que suele dominar lagos extremadamente ácidos
(Kristiansen 2005). Por el contrario, en términos de biovolumen los sistemas profundos y
conectados hidrológicamente (RH1 y RH4) mostraron una composición similar luego del
descongelamiento de los sistemas, momento en el cual las condiciones de fosfatos fueron
iguales. A pesar de esto, ambos cuerpos de agua luego estuvieron dominados por diferentes
grupos taxonómicos (RH1: Cyanobacteria, RH4: Chlorophyceae). Por su parte, en el
sistema más profundo (RH2) dominaron las Cyanobacteria durante casi todo el estudio,
mientras que en los cuerpos de agua someros (RH3 y RH5) se registraron picos de
Chrysophyceae durante el otoño. Estas diferencias en el comportamiento del biovolumen
entre los cuerpos de agua se asocian con las distintas composiciones en grupos algales y
especies del fitoplancton, los cuales responden diversamente a los distintos factores
ambientales registrados en cada sistema.
El índice de diversidad de Shannon- Weaver ha sido el más utilizado para el estudio
de las comunidades fitoplanctónicas tanto de ambientes marinos como de agua dulce
(Revelante & Gilmartin 1982, Romo & Miracle 1995, Cottingham et al. 1998). En nuestro
estudio, el índice de diversidad promedio estimado a lo largo de ocho campañas de
muestreo para los cinco cuerpos de agua de la turbera de Rancho Hambre (media= 1,38;
rango= 0,30―2,11) estuvo dentro del rango de variación (H= 0,56―2,18) medido por
Jasser et al. (2009) en lagos con diferentes concentraciones de sustancias húmicas de
Polonia, mientras que tanto el valor medio como el rango de variación fueron menores a los
registrados por Mataloni & Tell (1996) (media= 3,24; rango de variación= 2,57―4,29)
debido a una mayor equitatividad reportada por estos autores en cinco cuerpos de agua de
esta misma turbera. La temperatura y la diversidad se correlacionaron negativamente
debido a la equitatividad, ya que las temperaturas más altas favorecieron la dominancia de

DISCUSIÓN
93
las Cryptophyceae, especialmente en los sistemas someros. Por su parte, el sistema más
profundo (RH2) fue el más equitativo debido a que las mismas especies mostraron una
similar proporción durante todo el período de estudio. Por lo tanto, la temperatura es un
factor muy importante del cual depende la abundancia y diversidad de la comunidad
fitoplanctónica en los sistemas acuáticos estudiados.
Actualmente existen varios estudios de la comunidad picofitoplanctónica (fracción
de 0,2―2 µm de tamaño) en lagos húmicos (Søndergaard 1990, Jasser & Arvola 2003,
Drakare et al. 2003 y Jasser et al. 2009). Algunos resultados de estos autores indican que el
contenido de ácidos húmicos y el pH influyen en la composición del picofitoplancton: en
sistemas oligohúmicos (Jasser et al. 2009) y ácidos (Søndergaard 1991) domina el
picofitoplancton eucariota, mientras que a medida que aumentan el contenido de ácidos
húmicos y el pH, el aporte del picofitoplancton procariota (picocianobacterias) es mayor.
Coincidentemente, en los cuerpos de agua aquí estudiados (oligohúmicos y de bajo pH)
dominaron exclusivamente las picoalgas eucariotas, dentro de las cuales se distinguieron
dos tipos de morfos; uno característico de los sistemas profundos y otro de los someros.
Además, el volumen celular promedio mostró diferencias significativas dentro de los
cuerpos de agua profundos, lo que nos permite hipotetizar que existen distintas poblaciones
asociadas a un mismo morfo.
Según una recopilación realizada por Callieri (2007) para interpretar la dinámica
del picofitoplancton en sistemas acuáticos de agua dulce, es muy importante considerar las
características morfométricas y abióticas de los cuerpos de agua. A pesar de que en nuestro
estudio consideramos dichas variables, no encontramos ninguna relación estadísticamente
significativa entre los descriptores de la fracción picofitoplanctónica (abundancia y
biovolumen) y los parámetros morfométricos y abióticos estudiados. Drakare et al. (2003)
observaron para lagos húmicos una relación inversa entre la proporción del
picofitoplancton y la concentración de COD (carbono orgánico disuelto) coincidiendo con
lo observado en los sistemas someros estudiados aquí, los cuales mostraron mayores
concentraciones de COD y menores abundancias del picofitoplancton. Jasser et al. (2009)
reportaron en un lago oligohúmico de Polonia un importante aporte del picofitoplancton

DISCUSIÓN
94
eucariota (>60%) a la biomasa fitoplanctónica en concordancia con lo registrado en los
sistemas más profundos de la turbera de Rancho Hambre, en los cuales esta fracción realizó
un aporte de hasta el 40% al biovolumen total.
El fitoplancton de las lagunas de la turbera de Rancho Hambre estuvo dominado
numéricamente por pequeñas algas (cf. Ochromonas sp. 2 y picoeucariotas) capaces de
desarrollar episodios breves de crecimiento rápido, seguido de fuertes disminuciones.
Según las categorías de Reynolds debido a su pequeño tamaño, alta relación
superficie/volumen y altas tasas de crecimiento, estas algas son típicas C-estrategas
(Reynolds 2006), por lo que la comunidad fitoplanctónica de estos ambientes está
numéricamente dominada por especies colonizadoras. Este resultado es particularmente
novedoso para sistemas de turberas, ya que esta es la primera investigación en donde se
caracterizan todas las fracciones de tamaño del fitoplancton en estos sistemas acuáticos, por
lo que no pueden establecerse comparaciones con resultados de estudios en otras turberas
del mundo.
Con respecto a la clorofila a estimador de la biomasa fitoplanctónica, el rango de
variación medido en nuestro estudio (0―4,71 µg L-1) incluyó los valores bajos
registrados por Tell et al. (2011) en las lagunas Negra (0,10 µg L-1) y Victoria (0,68 µg L-1)
ambas sistemas acuáticos asociados a turberas de la provincia de Tierra del Fuego; y a los
obtenidos luego en un muestreo posterior en esta última laguna por Saad (2011) (1,63 µg L-
1). La clorofila a fue muy variable y no mostró un claro patrón temporal para los cuerpos
de agua estudiados. Sin embargo, las variaciones de este parámetro dependieron del pH a
través de una regresión lineal positiva entre ambas variables. Por lo tanto, en los sistemas
acuáticos estudiados, el pH fue el factor abiótico que más explicó las variaciones espaciales
de la riqueza específica y la clorofila a, mientras que la temperatura influyó sobre los
cambios temporales de la abundancia y diversidad de la comunidad fitoplanctónica.
Las estrategias nutricionales mixotróficas de muchas especies son bien conocidas
desde los primeros estudios taxonómicos de los organismos planctónicos (Sanders & Porter
1988). Sin embargo, el creciente conocimiento del bucle microbiano o “microbial loop”
como una vía importante de flujo de carbono y nutrientes en ecosistemas acuáticos (Azam

DISCUSIÓN
95
et al. 1983) ha producido un creciente interés sobre el rol de los organismos mixótrofos en
las comunidades pelágicas (Jones 2000). Las especies mixótrofas combinan los modos de
nutrición fagotrófico y fototrófico, lo que les brinda un amplio espectro de estrategias
nutricionales que les permiten adaptarse a una gran variedad de condiciones ambientales
(Sigee 2005). Sin embargo, en los sistemas distróficos estudiados, los organismos
mixótrofos dominaron todos los cuerpos de agua sólo al principio del período de aguas
libres coincidiendo con lo observado en lagos antárticos (Izaguirre et al. 1993, 2001, 2003,
Marshall & Laybourn-Parry 2002) en los cuales esta dominancia ocurre en el invierno
cuando los sistemas están cubiertos por hielo y la luz es un recurso limitante. En estas
condiciones, los organismos mixótrofos se ven favorecidos, ya que tienen la capacidad de
obtener energía de la ingestión de bacterias (Nygaard & Tobiesen 1993, Urabe et al. 2000,
Callieri et al. 2002). Una situación similar fue descripta para lagos húmicos en Suecia
(Jansson et al. 1999), donde los flagelados mixótrofos asumen la dominancia a través de la
bacterivoría. Nuestros resultados demuestran que en general, la estrategia nutricional
dominante en los ambientes estudiados durante el período de aguas libres es la autotrofía,
ya que tanto las condiciones lumínicas como los nutrientes no serían limitantes para el
crecimiento autotrófico. A partir de lo mencionado anteriormente, no podemos rechazar ni
corroborar la hipótesis planteada en la cual se postuló que las características distróficas en
los ambientes estudiados determinaría la dominancia de los organismos mixótrofos, ya que
posiblemente la mixotrofía sea la estrategia nutricional dominante en el invierno, sugerida a
través de su dominancia luego del período de descongelamiento, reemplazándose
posteriormente por especies autótrofas. A pesar de que no realizamos mediciones de
producción primaria (PP) y bacteriana (PB) en este estudio, ambientes de características
similares mostraron una relación PP/PB mayor a uno (Saad 2011). Por lo tanto,
probablemente la captación de energía y fijación de carbono en las lagunas de turberas esté
dada principalmente por el fitoplancton, contrariamente a lo observado en lagos húmicos
donde dominan las vías heterotróficas (Hessen et al. 1990, Nürnberg & Shaw 1999,
Drakare et al. 2002). Para corroborar esta hipótesis, sería de gran interés realizar estudios

DISCUSIÓN
96
de producción primaria y bacteriana en futuras investigaciones en las lagunas de la turbera
de Rancho Hambre.
Relación entre las distintas comunidades planctónicas con énfasis en el estudio del
fitoplancton
El bucle microbiano, en los ecosistemas acuáticos, está constituido por materia
orgánica, bacterias, flagelados heterotróficos y ciliados. Particularmente, los nanoflagelados
heterotróficos (NFH) son considerados los principales consumidores de bacterias y
picoalgas (Porter et al. 1985, Fenchel 1986, Sherr et al. 1987, Bloem et al. 1989). Sin
embargo, en este estudio no se encontró ninguna relación entre las abundancias de los NFH
y la abundancia y el biovolumen del picofitoplancton. Una posible explicación podría ser
que estos organismos menores a 5�m se encuentren regulados por sus predadores “top-
down”. Otra posibilidad, se relaciona con el tamaño, ya que según estudios de predación
(“grazing”) los NFH menores a 5�m consumen principalmente picoplancton procariota
(Sherr & Sherr 1991, Sherr et al. 1991, Stockner 1991, Weisse 1993), por lo que muy
probablemente no ejerzan un efecto de predación importante sobre el picofitoplancton
eucariota.
Los ciliados juegan un rol central en la estructura del bucle microbiano (microbial
loop) ya que pueden consumir bacterias, flagelados heterotróficos y pico- y
nanofitoplancton, constituyendo así el vínculo con los niveles tróficos superiores (Sanders
et al. 1989, Šimek et al. 1990, Sherr et al. 1987). En este caso, la mayoría de las especies
más abundantes de ciliados registrados puede consumir tanto bacterias como algas (Šimek
et al. 1996; Küppers et al. en prep.). Para la laguna RH2, se observó una correlación
negativa entre las abundancias del picofitoplancton eucariota y del ciliado Halteria
grandinella, que posiblemente indique un efecto de regulación predador-presa. Este ciliado,
presente únicamente en esta laguna, muestra un comportamiento de escape (movimientos
en forma de saltos y natación rápida) muy efectivo ante la predación de rotíferos como los

DISCUSIÓN
97
que dominan este cuerpo de agua (Gilbert 1994), resultando posiblemente en una mayor
presión de predación sobre la comunidad picofitoplanctónica.
Por su parte, para el resto de los cuerpos de agua las abundancias de los copépodos
filtradores (Boeckella poppei) se correlacionaron positivamente con las abundancias del
picofitoplancton, lo que posiblemente sugiera un efecto de cascada trófica positivo sobre
esta fracción. Almada et al. (2004) demostró en experimentos de microcosmos en un lago
antártico dominado por Boeckella poppei que el nanofitoplancton es controlado
eficientemente por este copépodo y que no hay un efecto significativo sobre el
picofitoplancton. Entonces, en los sistemas estudiados posiblemente Boeckella poppei
tenga un efecto de cascada trófica positivo sobre el picofitoplancton disminuyendo la
presión de predación sobre esta fracción o a través de una mayor disponibilidad de
nutrientes debido a un fuerte control sobre sus competidores. Estos resultados nos permiten
plantear las hipótesis de que en RH2 el ciliado Halteria grandinella tiene un importante
efecto de predación sobre la comunidad picofitoplanctónica, mientras que en el resto de los
sistemas los copépodos tienen un efecto de cascada trófica positivo sobre el
picofitoplancton. Para testear estas hipótesis sería necesario realizar en próximas
investigaciones en las lagunas de la turbera de Rancho Hambre experimentos en los cuales
se excluyan los diferentes componentes del plancton, observándose luego las tasas de
crecimiento de las distintas fracciones de tamaño del fitoplancton en cada tratamiento, tales
como los realizados por Allende & Pizarro (2006) y Allende (2009) en lagos antárticos.
Los Grupos Funcionales Basados en características Morfológicas (GFBM) y su relación
con los factores bióticos y abióticos.
Las primeras clasificaciones del fitoplancton basadas en las propiedades funcionales
fueron realizadas por Reynolds (1980, 1984), en las cuales propuso 14 asociaciones en
función de características fisiológicas, morfológicas y ecológicas basadas en observaciones
del fitoplancton de lagos de Inglaterra. Esta clasificación fue sucesivamente refinada y
expandida (Reynolds 1997, Padisák & Reynolds 1998). Actualmente, se proponen 40

DISCUSIÓN
98
asociaciones de acuerdo al aporte de diferentes especialistas (Reynolds 2002, Padisák et al.
2003, 2006, 2009). En este marco conceptual, las distintas estrategias de los organismos del
fitoplancton están fuertemente relacionadas con las diferencias en las dimensiones
geométricas y en las formas. Las características morfológicas son condiciones esenciales
que influyen en las propiedades de hundimiento, tasas de crecimiento, eficiencia en la
utilización de luz y nutrientes y en la susceptibilidad a la predación (Salmaso & Padisák
2007). Sobre esta base, Kruk et al. (2010) realizaron una clasificación funcional del
fitoplancton basada en simples características morfológicas, en la cual propusieron siete
grupos morfo-funcionales. La ventaja de esta clasificación es que es objetiva, independiente
de afiliaciones taxonómicas y su aplicación a la mayoría de las especies es relativamente
fácil. La utilización de grupos funcionales nos permite identificar las respuestas comunes
de la comunidad fitoplanctónica a determinadas condiciones ambientales. La escasa
información sobre la autoecología de muchas especies del fitoplancton características de las
lagunas de turberas (Mataloni 1997) hace imposible la utilización de la clasificación de
Reynolds (2002). Por esta razón utilizamos la clasificación propuesta por Kruk et al. (2010)
para el estudio del fitoplancton de las lagunas de la turbera de Rancho Hambre.
De acuerdo a los presentes resultados, algunos organismos de grupo I (pequeños
organismos) y II (flagelados pequeños) fueron muy abundantes y mostraron una mayor
constancia que el resto de los GFBM, apareciendo en un alto porcentaje de los muestreos en
la mayoría de los cuerpos de agua. El pequeño tamaño permite la rápida captación de
nutrientes a través de una alta relación superficie/volumen (Kruk et al. 2010, Kruk &
Segura 2012), estrategia muy importante en ambientes oligo a mesotróficos (Naselli-Flores
et al. 2007). A pesar de que estos organismos son muy palatables (Kruk et al. 2010),
presentan altas tasas de crecimiento poblacional, lo que les permite tolerar las altas pérdidas
por predación (Sommer et al. 2003). Por su parte, los flagelados de tamaño medio a grande
fueron muy abundantes pero revelaron una mayor fidelidad a los sistemas profundos,
conectados y ligeramente ácidos (RH1 y RH4). Su mayor dimensión lineal máxima y la
presencia de flagelos les proporcionan una sustancial tolerancia a la predación
principalmente del zooplancton especialista (Reynolds 1997). Los organismos sin

DISCUSIÓN
99
estructuras especializadas (grupo IV) representados mayormente en este estudio por
Desmidiaceae y los individuos con exoesqueleto silíceo (Bacillariophyceae, grupo VI) se
revelaron con una baja frecuencia durante el período de estudio en RH1 y RH4,
respectivamente. Las grandes colonias mucilaginosas asociadas con un mayor aporte de
Chlorococcales (por ejemplo, Dictyochlorella globosa (Kors.) Silva, Dictyosphaerium
chlorelloides (Naum.) Kom. et Perm., Dictyosphaerium pulchellum Wood) mostraron una
mayor fidelidad al sistema de mayor superficie, conectado hidrológicamente y más
minerotrófico (RH4). Sin embargo, este grupo mostró su mayor aporte al biovolumen,
representado por colonias de Cynobacteria (Aphanothece minutissima) en el cuerpo de
agua más profundo, aislado hidrológicamente y ácido (RH2). Mataloni & Tell (1996)
registraron una preferencia de este grupo taxonómico por ambientes de bajo pH a través de
su mayor abundancia. Por el contrario, en este estudio sólo fueron importantes en términos
de biovolumen.
Según el resultado del ordenamiento del biovolumen de los grupos morfo-
funcionales del fitoplancton y las variables bióticas y abióticas a partir del análisis de
correspondencia canónica (CCA), las abundancias de ciliados, macrofiltradores del
metazooplancton y el pH fueron los factores que resultaron fundamentales en la
determinación de las estrategias predominantes en las distintas comunidades
fitoplanctónicas de las lagunas de la turbera de Rancho Hambre. Los grupos IV
(mayormente Desmidiaceae) y VI (Bacillariophyceae) son indicadores de altos pH y mayor
abundancia de macrofiltradores (Daphnia commutata). Este resultado confirma lo obtenido
por Mataloni & Tell (1996) en esta misma turbera, donde establecieron anteriormente que
las Zygnematophyceae (Desmidiaceae) y Bacillariophyceae prefieren ambientes grandes y
de condiciones más minerotróficas (altos valores de pH y dureza). Por su parte, las especies
del género Daphnia son claves en la dinámica de las tramas tróficas de los lagos e influyen
en la sucesión estacional del fitoplancton (Sommer et al. 1986). Este gran cladócero es un
importante predador y tiene altos requerimientos de fósforo para su crecimiento (Balseiro et
al. 2007). A pesar que puede consumir y digerir tanto Desmidiaceae (Coesel 1997) como
Bacillariophyceae (Carotenuto & Lampert 2004) presenta un mayor impacto sobre el

DISCUSIÓN
100
picoplancton (Callieri et al. 2002, Modenutti et al. 2003, Balseiro et al. 2004),
nanoflagelados y ciliados (Jürgens 1994, Modenutti et al. 2003).
En síntesis, la caracterización de los cinco cuerpos de agua de la turbera de Rancho
Hambre que se muestra en la Tabla 6 reveló que:
- Los ambientes profundos, conectados hidrológicamente, ligeramente ácidos y con
altas concentraciones de fosfatos (RH1 y RH4) fueron los sistemas más minerotróficos y
ricos en especies, particularmente representados por Zygnematophyceae, Chlorophyceae y
Bacillariophyceae. Además, las especies dominantes numéricamente del fitoplancton >2�m
fueron cf. Ochromonas sp. 2 y Kirchneriella microscopica, esta última especie además
realizó un gran aporte al biovolumen. La composición del protozooplancton fue similar y
ambos coincidieron en la presencia del cladócero Daphnia commutata, especie clave en la
estructuración de las relaciones tróficas entre las diferentes comunidades planctónicas. La
estrategia morfo-funcional del fitoplancton que dominó numéricamente fueron los
organismos pequeños (GFBM I) representados principalmente por picofitoplancton y
Kirchneriella microscopica, mientras que el biovolumen se caracterizó por la presencia de
organismos sin estructuras especializadas (GFBM IV) y colonias mucilaginosas (GFBM
VII).
- El ambiente profundo pero aislado hidrológicamente (RH2) con grandes
fluctuaciones en el nivel hidrométrico mostró un bajo pH, mayores concentraciones de
fosfatos y valores intermedios de riqueza específica. La abundancia del fitoplancton >2�m
estuvo principalmente representada por una Chysophyceae nanoflagelada (cf. Ochromonas
sp. 2) mientras que en biovolumen dominaron las grandes colonias mucilaginosas de
Aphanothece minutissima (GFBM VII). La composición y dinámica del protozooplancton
fue característica, revelando además en este sitio el mayor aporte en abundancia y
biovolumen de la fracción picofitoplanctónica (GFBM I).
- Los ambientes someros y ácidos (RH3 y RH5), con grandes variaciones térmicas y
bajas concentraciones de fosfatos fueron los sistemas más pobres en especies. La
abundancia del fitoplancton >2�m estuvo caracterizada por dos especies nanoflageladas (cf.

DISCUSIÓN
101
Plagioselmis sp. 2 y cf. Ochromonas sp. 2.) mientras que en biovolumen realizaron un
gran aporte las Chrysophyceae (cf. Ochromonas spp.) y las Cyanobacteria (Aphanothece
minutissima). Estos sistemas mostraron una gran variación en la composición y dinámica
del protozooplancton, aunque no así en el metazooplancton. Se caracterizaron por la
presencia de estrategias morfo-funcionales consistentes en organismos pequeños (GFBM I)
y flagelados (GFBM II y V) en términos de abundancia, mientras que en términos de
biovolumen estuvieron dominados por diferentes GFBM (RH3: II, RH5: VII, V y II).
Tabla 6. Caracterización de los cinco cuerpos de agua de la turbera de Rancho Hambre.

DISCUSIÓN
102
A lo largo de este trabajo de Tesis Doctoral se reveló una gran diversidad
limnológica en la turbera de Rancho Hambre, la cual determinó una metacomunidad
fitoplanctónica muy rica en especies, la que reveló a su vez una flora distintiva en cada
cuerpo de agua. Esto corrobora la hipótesis previamente postulada por Mataloni & Tell
(1996) en esta misma turbera, que establece que la diversidad limnológica es fundamental
para la biodiversidad de las comunidades planctónicas en estos sistemas acuáticos. Estas
diferencias reflejaron distintos patrones temporales y espaciales en la estructura, dinámica y
estrategias morfo-funcionales del fitoplancton, revelando a las algas pequeñas como las
más abundantes (Chrysophyceae nanoflageladas y picoeucariotas). Los parámetros
abióticos que explicaron las variaciones espaciales y temporales de la comunidad
fitoplanctónica fueron el pH y la temperatura, respectivamente (pH: riqueza específica y
clorofila a, temperatura: abundancia y diversidad), mientras que los factores bióticos más
importantes fueron las abundancias del ciliado Halteria grandinella y las del copépodo
Boeckella poppei. A continuación se muestra un esquema del funcionamiento de los
cuerpos de agua estudiados en la turbera de Rancho Hambre (Figura 40).

DISCUSIÓN
103
Figura 40. Esquema del funcionamiento de los cuerpos de agua de la turbera de Rancho Hambre.
Las fechas punteadas indican posibles relaciones no demostradas en esta Tesis Doctoral.

CONCLUSIONES
GENERALES

RESUMEN Y CONCLUSIONES GENERALES
104
Principales conclusiones
• La topografía de la turbera determinó en gran medida la conectividad hidrológica
superficial (entradas y/o salidas superficiales de agua) que junto con la morfometría
influyó en el estado minero/ombrotrófico de los cuerpos de agua a través de la
existencia de fuentes de agua adicionales al aporte directo de la lluvia y además
moduló los cambios en el nivel hidrométrico. Esto permitió caracterizar los sistemas
en: profundos conectados hidrológicamente, profundo y aislado hidrológicamente, y
ambientes someros.
• Los cuerpos de agua de la turbera de Rancho Hambre fueron sistemas acuáticos
homogéneos en sus propiedades físico-químicas y en la distribución espacial de sus
comunidades planctónicas durante el período de aguas libres debido a que la
dirección del viento coincide con la orientación del valle y de los cuerpos de agua,
lo que promueve la mezcla permanente de la columna de agua.
• La cantidad de carbono orgánico disuelto y sustancias húmicas permitió clasificar a
estos cuerpos de agua como oligohúmico. Dado que estos cuerpos de agua fueron
poco profundos y bien mezclados, el color dado por las sustancias húmicas no
afectó la disponibilidad de luz para la comunidad fitoplanctónica.
• El pH y la temperatura fueron los factores abióticos más importantes que modularon
las variaciones espaciales y temporales, respectivamente, de los descriptores de
comunidad fitoplanctónica, (pH: riqueza especifica y clorofila a, temperatura:
abundancia y diversidad), mientras que las abundancias del ciliado Halteria
grandinella y las del copépodo Boeckella poppei fueron los factores bióticos más
relevantes para el fitoplancton.
• En los cuerpos de agua estudiados, la similitud entre pool de especies fue baja a lo
esperado para cuerpos de agua tan cercanos con una flora característica en cada
sistema. Esto confirma la hipótesis previamente planteada por Mataloni & Tell
(1996) que establece que esta diversidad limnológica es fundamental para la
biodiversidad de las comunidades planctónicas de los cuerpos de agua de turberas.

RESUMEN Y CONCLUSIONES GENERALES
105
• Pese a las diferencias en la composición de especies y grupos taxonómicos entre los
cuerpos de agua estudiados, en general las algas pequeñas fueron las más
abundantes (Chrysophyceae nanoflageladas y picoeucariotas) y la autotrofía fue la
estrategia nutricional dominante del fitoplancton durante el período de aguas libres,
ya que la luz y los nutrientes no resultaron limitantes para el crecimiento
autotrófico.
• La clasificación en Grupos Funcionales Basados en las características Morfológicas
(GFBM) sensu Kruk et al. (2010) fue muy útil para clasificar el fitoplancton de
lagunas de turberas dando como resultado a los grupos morfo-funcionales IV
(mayormente Desmidiaceae) y VI (Bacillariophyceae) como indicadores de
condiciones más minerotróficas (alto pH) y presencia del cladócero Daphnia
commutata. Esta clasificación será una herramienta muy útil para el monitoreo de
estos humedales ya que se podrá detectar alteraciones que afecten los sistemas a
través de cambios en la composición de GFBM.
Perspectivas
Las lagunas de turberas son sistemas con una alta biodiversidad (Mataloni 1997) y
su conocimiento desde lo estructural y funcional es de gran importancia en función del
ordenamiento territorial de estos humedales planificado por la provincia de Tierra del
Fuego en base a aspectos ecosistémicos, sociales y económicos, y que involucra la
protección de las turberas fueguinas. En ese sentido, el proyecto que enmarca a esta tesis
doctoral estudió integralmente por primera vez los ecosistemas acuáticos en estos frágiles
humedales fueguinos, aportando información sobre el funcionamiento y biodiversidad de
las comunidades planctónicas en los sistemas acuáticos de turberas.
Esta tesis doctoral es la primera caracterización realizada a lo largo de un período de
dos años de cuerpos de agua de turberas, y de la estructura y dinámica de sus comunidades
fitoplanctónicas y su relación con las características bióticas y abióticas. Dentro del marco
antes planteado, el presente estudio será una herramienta muy útil ya que aporta el
conocimiento de base necesario para poder comparar las comunidades fitoplanctónicas de
diferentes cuerpos de agua de turberas en el marco de una caracterización regional de estos

RESUMEN Y CONCLUSIONES GENERALES
106
sistemas, dado que el enfoque basado en los grupos morfo-funcionales sensu Kruk et al.
(2010) puede ser utilizado en monitoreos a largo plazo y para comparar una gran cantidad
de cuerpos de agua (Izaguirre et al. 2012).
También, esta tesis nos permitió obtener una primera aproximación a la
identificación de cuáles son los factores tanto bióticos como abióticos que modelan la
comunidad fitoplanctónica en las lagunas de turberas, determinando además de sus distintas
estrategias morfo-funcionales. En este caso, se propone corroborar las siguientes hipótesis:
- La captación de energía y fijación de carbono en las lagunas de turberas está dada
principalmente por el fitoplancton. Esta hipótesis se pondrá a prueba realizando estudios de
producción primaria y bacteriana.
- En RH2 el ciliado Halteria grandinella tiene un importante efecto regulador sobre la
fracción de tamaño picofitoplanctónica, mientras que en el resto de los sistemas estudiados
los copépodos tienen un efecto de cascada trófica positivo sobre el picofitoplancton. Estas
hipótesis se podrán a prueba realizando experimentos en los cuales se excluyan los
diferentes componentes del plancton, observándose luego las tasas de crecimiento de las
distintas fracciones de tamaño del fitoplancton.
A partir de esta tesis doctoral, surgieron interrogantes sobre la identidad taxonómica
de las fracciones del fitoplancton más abundantes en los ambientes estudiados (pico y
nanofitoplancton) ya que su estudio es difícil mediante las técnicas tradicionales de
microscopía. Por esto, para lograr una correcta clasificación taxonómica es necesario
adoptar un enfoque polifásico (Komárek 2006), el cual integra la información surgida a
partir de diferentes técnicas de análisis: morfología, ecología y biología molecular. Este
punto está pensado para una próxima etapa, en la que se plantea una caracterización
molecular de la fracción picoplanctónica mediante DGGE y bibliotecas genéticas,
citometría de flujo y microscopía de epifluorescencia en el marco de una beca posdoctoral.
Ambas líneas de investigación nos permitirán aportar información original y útil
tanto para el inventario de biodiversidad como para el conocimiento de base propuestos por
la Convención Internacional de Ramsar como una herramienta fundamental para garantizar
la conservación y el uso racional de las turberas.

BIBLIOGRAFÍA

BIBLIOGRAFÍA
107
BIBLIOGRAFÍA
Almada P. (2002) Estudios experimentales sobre el efecto del pastoreo del copépodo
Boeckella poppei sobre el fitoplancton de un lago antártico (Bahía Esperanza,
Psula. Antártica). Tesis de Licenciatura, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, Buenos Aires, 1–40 p.
Almada P., Allende L., Tell G., Izaguirre I. (2004) Experimental evidence of the grazing
impact of Boeckella poppei on phytoplankton in a maritime Antarctic lake. Polar
Biology 28 (1): 39-46.
Allende L. (2009) Combined effects of nutrients and grazers on bacterioplankton and
phytoplankton abundance in an Antarctic lake with even food-chain links. Polar
Biology 32: 493-501.
Allende L. & Pizarro H. (2006) Top-down control on plankton components in an
Artarctic pond: experimental approach to the study of low-complexity food webs.
Polar Biology 29: 893-901.
American Public Health Association (APHA) (2005) Standard methods for the
examination of water and wastewaters. 21st edition. APHA, AWWA, WEF,
Washington, DC, 1368 pp.
Arvola L., Eloranta P., Järvinen M., Keskitalo J. & Holopainen A. (1999)
Phytoplankton. En: Keskitalo J. & Eloranta P. (eds.) Limnology of humic waters,
Backhuys Publishers, Leiden, Netherlands, 135- 171 p.
Auer V. (1965) The Pleistocene of Fuego-Patagonia. Part IV: Bog profiles. Ann. Acad.
based on light microscopy. Hydrobiologia 502: 271-283.
Azam F., Fenchel T., Field J., Gray J., Meyer-Reil L. & Thingstad F. (1983) The
ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 10:257-
263.
Balseiro E., Modenutti B., Queimaliños C. & Reissig M. (2007) Daphnia distribution in
Andean Patagonian lakes: effect of low food quality and fish predation. Aquatic
Ecology 41: 599-609.

BIBLIOGRAFÍA
108
Balseiro E., Queimaliños C. & Modenutti B. (2004) Grazing impact on autotrophic
picoplankton in two south Andean lakes (Patagonia, Argentina) with different
light: nutrient ratios. Revista Chilena de Historia Natural 77: 73-85.
Battarbee E.W. (1986) Diatom analysis. En: Berglund B.E. (ed) Handbook of holocene
palaeoecology and palaeohydrology, Wiley, New York, 527–570 p.
Baumann M. (2006) Water flow, spatial patterns, and hydrological self regulation of a
raised bog in Tierra del Fuego (Argentina). Diploma- thesis (unpubl.), Univ. of
Greifswald, Institute of Botany and Landscape Ecology.
Bayly I.A.E. (1992) Fusion of the genera Boeckella and Pseudoboeckella (Copepoda)
and revision of their species from South America and sub-Antarctic islands. Rev.
Chil. Hist. Nat. 65: 17-63.
Benzie J.A.H. (2005) Cladocera: The Genus Daphnia (including Daphniopsis)
(Anomopoda: Daphniidae). Backhuys Publishers, Leiden, 368 pp.
Bloem J., Ellenbroek F.M., Bär-Gilissen M.J.B. & Cappenberg T.E. (1989) Protozoan
grazing and baterial production in stratified Lake Vechten estimated with
fluorescently labeled bacteria and by thymidine incorporation. Appl. Environ.
Microbiol. 55: 1787-1795.
Bonarelli G. (1917) Tierra del Fuego y sus turberas. Anales Min. Agric. Nac. - Secc.
Geol., Mineral. y Minería 12 (3): 119 pp.
Boltovskoy A. & Urrejola R. (1977) Dos nuevas especies del género Keratella
(Rotatoria) de Tierra del Fuego, Argentina. Limnobios 1: 181-187.
Borge O. (1906) Süsswasser-Chlorophyceen von Feuerland und Isla Desolación. Bot.
Stud. tillagn. F.R. Kjellmann, Uppsala.
Borics G., Tothmérész B., Grigorszky I., Padisák J., Várbíro G. & Szabó S. (2003)
Algal assemblage types of bog-lakes in Hungary and their relation to water
chemistry, hydrological conditions and habitat diversity. Hydrobiologia 502: 145-
155.
Brook A.J. (1981) The biology of desmids. En: Burnett J.H. (ed) Botanical Monographs
16, University of California Press, Berkeley, Los Angeles, 276 pp.
Callieri C. (2007) Picophytoplankton in freshwater ecosystems: the importance of
small-size photothophs. Freshwater Reviews 1:1-28.

BIBLIOGRAFÍA
109
Callieri C., Bertoni R. & Corno G. (2002) Dynamics of bacteria and mixotrophic
flagellates in an Alpine lake in relation to Daphnia population development. J.
Limnol. 61 (2): 177-182.
Callieri C. & Pinolini M.L. (1995) Picoplankton in Lake Maggiore, Italy. International
Revue der gesamten Hydrobiologie 80: 491-501.
Carotenuto Y. & Lampert W. (2004) Ingestion and incorporation of freshwater diatoms
by Daphnia pulicaria: do morphology and oxylipin production matter?. Journal
Plankton Research 26 (5): 563-569.
Carpenter D.L., Cole J.J., Kitchell J.F. & Pace M.L. (1998) Impact of dissolved organic
carbon, phosphorus, and grazing on phytoplankton biomass and production in
experimental lakes. Limnology and Oceanography 43: 73-80.
Clymo R.S. (1964) The origin of acidity in Sphagnum bogs. The Bryologist 67: 427-
431.
Coesel P.F.M. (1996) Biogeography of desmids. Hydrobiologia 336: 41-53.
Coesel P.F.M (1997) The edibility of Staurastrum chaetoceras and Cosmarium
abbreviatum (desmidiaceae) for Daphnia galeata/hylina and the role of desmids
in aquatic food web. Aquatic Ecology 31: 73-78.
Coesel P.F.M. & Krienitz L. (2008) Diversity and geographic distribution of desmids
and other coccoid green algae. Biodivers. Conserv. 17: 381-392.
Cottingham K.L., Carpenter S.R. & St. Amand A.L (1998) Responses of epilimnetic
phytoplankton to experimental nutrients enrichment in three small seepage lake. J.
Plankton Res. 20: 1889-1914.
Croasdale H. (1962). Freshwater algae of alaska III. Desmids from the cape Thompson
area. Trans. Amer. Microsc. Soc. 81: 12-42.
Cvijan M. & Lausevic R. (1991) Desmids of Vlasinsko Lake- from peat bog to lake.
Arch. Protistenkd. 139: 21-37.
Davies-Colley R.J. & Vant W.N. (1987) Absorption of light by yellow substance in
freshwater lakes. Limnology and Oceanography 32 (2):416-425.
de la Balze V.M., Blanco D.E. & Loekemeyer N. (2004) Aspectos sobre el uso y
conservación de los turbales patagónicos. En: Blanco D.E. & de la Balze V.M.
(eds) Los Turbales de la Patagonia, bases para su inventario y la conservación de

BIBLIOGRAFÍA
110
su biodiversidad, Fundación Humedales / Wetlands International, Buenos Aires,
Argentina, 129–145 p.
Downing J.A. & McCauley E. (1992) The nitrogen: phosphorus relationship in lakes.
Limnology and Oceanography 37(5): 936–945.
Drakare S., Blomqvist P., Bergström A.K. & Jansson M. (2002) Primary production and
phytoplankton composition in relation to DOC input and bacterioplankton
production in humic lake Örträsket. Freshwater Biology 47: 41-52.
Drakare S., Blomqvist P., Bergström A.K. & Jansson M. (2003) Relationships between
picophytoplankton and environmental variables in lakes along a gradiend of water
color and nutrient content. Freshwater Biology 48: 729-740.
Druvietis I., Springe G., Briede A., Kokorite I. & Parele E. (2010) A comparative
assessment of the bog aquatic environment of the ramsar site of Teiči nature
reserve, Latvia. En: Klavinš M. (ed) Mires and peat, University of Latvia Press,
Riga 19-40 p.
Ettl H. (1978) Xanthophyceae. En: Ettl H., Gerloff J. & Heynig H. (eds),
Süsswasserflora von Mitteleuropa, Gustav Fischer Verlag, Suttgart, 530 pp.
Fenchel T. (1986) The ecology of heterotrophic microflagellates. Adv. Microb. Ecol. 9:
57-97.
Finlay B. J. & Guhl B.E. (1992) Plankton sampling - freshwater. En: Lee J.J. & Soldo
A.T. (eds) Protocols in protozoology, Society of Protozoology, Allen Press,
Lawrence KS, USA, B1.1- B2.5.
Foissner W. & Berger H. (1996) A user-friendly guide to the ciliates (Protozoa,
Ciliophora) commonly used by hydro- biologists as bioindicators in rivers, lakes,
and waste waters, with notes on their ecology. Freshw. Biol. 35: 375-482.
Foissner W., Berger H. & Kohmann F. (1992) Taxonomische und ökologische Revision
der Ciliaten des Saprobiensystems - Band II: Peritrichia, Heterotrichida,
Odontostomatida. Informationsberichte des Bayer, Landesamtes für
Wasserwirtschaft, München, 5/92, 502 pp.
Foissner W., Berger H. & Kohmann F. (1994) Taxonomische und ökologische Revision
der Ciliaten des Saprobiensystems - Band III: Hymenostomata, Prostomatida,

BIBLIOGRAFÍA
111
Nassulida. Informationsberichte des Bayer, Landesamtes für Wasserwirtschaft,
München, 1/94, 548 pp.
Foissner W., Berger H., Blatterer H. & Kohmann F. (1995) Taxonomische und
ökologische Revision der Ciliaten des Saprobiensystems - Band IV:
Gymnostomates, Loxodes, Suctoria. Informationsberichte des Bayer,
Landesamtes für Wasserwirtschaft München, 1/95, 540 pp.
Foissner W., Berger H. & Schaumburg J. (1999) Identification and ecology of limnetic
plankton ciliates. Informationsberichte des Bayer, Landesamtes für
Wasserwirtschaft, München, 3/99, 793 pp.
Foissner W., Blatterer H., Berger H. & Kohmann F. (1991) Taxonomische und
ökologische Revision der Ciliaten des Saprobiensystems - Band I: Cyrtophorida,
Oligotrichida, Hypotrichia, Colpodea. Informationsberichte des Bayer,
Landesamtes für Wasserwirtschaft, München, 1/91, 478 pp.
Gesst A.V. & Coesel P. (2012) Desmids from lake Nabugabo (Uganda) and adjacent
peat bogs. Fottea (1): 95- 110.
Gilbert D. & Mitchell E.A.D. (2006) Micobial diversity in Spahgnum peatlands. En:
Martini I.P., Martínez Cortizas A. & Chesworth W. (eds) Peatlands: Evolution
and records of environmental and climatic changes, Elsevier Press, 289-320 p.
Gilbert J.J. (1994) Jumping behaviorhe in oligotrich ciliates Strobilidium velox and
Halteria grandinella grandinella, and its significance as a defense against rotifer
predator. Microbial Ecology 27:189-200.
Górniak A., Grabowska M., Jekatierynczuk-Rudczyk E., Zieliński P. & Suchowolec T.
(2003) Long- term variations of phytoplankton primary production in a shallow,
polyhumic reservoir. Hydrobiologia 506-509: 305-310.
Grabowska M., Górniak A., Jekatierynczuk-Rudczyk E. & Zieliński P. (2003) The
influence of hydrology and water quality on phytoplankton community
composition and biomass in a humoeutrophic reservoir, Siemianówka reservoir
(Poland). Ecohydrol. Hydrobiol. 3: 185-196.
Green J.D. & Shiel R.J. (1999) Mouthpart morphology of three calanoid copepods from
Australian temporary pools: evidence for carnivory. N. Z. J. Mar. Freshwater Res.
33: 385–398.

BIBLIOGRAFÍA
112
Grönblad R. (1942) Algen, Hauptsächlich Desmidiaceen, aus dem finnischen,
norwegischen and schwedischen lappland. Mit berücksichtigung der organismen
des phytoplanktons und des zooplanktons. Acta societatis scientiarum fennicae.
Nova Serie B., Tom. II Nº 5.
Guarrera S.A. (1986) Orden Chroococcales. En: Guarrera S., Gamundi I. & Halperin D.
(eds) Flora criptogámica de Tierra del Fuego, Buenos Aires, 63 pp.
Guarrera S.A. & Echenique R.O. (1998) Hormogonophycidae. En: Guarrera S.A.,
Gamundi de Amos I. & Matteri C.M (eds), Flora criptogámica de Tierra del
Fuego, CONICET, Buenos Aires, 93 pp.
Guarrera S.A. & Kühnemann O. (1949) Catálogo de las “Chlorophyta” y “Cyanophyta”
de agua dulce de la República Argentina, Lilloa 19: 219-533.
Guilford S.T. & Hecky R.E. (2000) Total nitrogen, total phosphorus, and nutrient
limitation in lakes and oceans: is there are common relationship? Limnology and
Oceanography 45(6): 1213–1223.
Hessen D.O., Andersen T. & Lyche A. (1990) Carbon metabolism in a humic lakes;
pool sizes and cycling through zooplankton. Limnology and Oceanography 35:
84-99.
Hessen D.O. & Tranvik L.J. (eds) (1998) Aquatic humic substances: ecology and
biogeochemistry. Ecology Studies vol. 133, Springer- Verlag, Berlin, 346 pp.
Heywood R.B. (1970a) Ecology of the fresh-water-lakes of Signy Island, South Orkney
Islands: III. Biology of the copepod Pseudoboeckella silvestri Daday (Calanoida,
Centropagidae). Br. Antarct. Surv. Bull. 23: 1–17.
Heywood R.B. (1970b) The mouthparts and feeding habits of Parabroteas sarsi (Daday)
and Pseudoboeckella silvestri (Daday) (Copepoda, Calanoida). En: Holdgate M.
W. (ed), Antarctic Ecology, Academic Press, London, 639–650 p.
Hillebrand H., Dürselen C.D., Kirschtel D., Pollingher U. & Zohary T. (1999)
Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 35: 403-424.
Holden J. (2006) Peatland hydrology. En: Martini I.P., Martínez Cortizas A. &
Chesworth W. (eds) Peatlands: Evolution and records of environmental and
climate changes, Elsevier, Amsterdam, 319–346 p.

BIBLIOGRAFÍA
113
Hüber-Pestalozzi G. (1941) Chrysophyceen. En: Thienemann A. (ed) Die
Binnengewässer. Das Phytoplankton des Süsswassers, 16/2. Nägele and
Obermiller, Stuttgart, 1-247.
Huovinen P.S., Penttilä H. & Soimasuo M.R. (2003) Spectral attenuation of solar
ultraviolet radiation in humic lakes in Central Finland. Chemosphere 51: 205-214.
Huszar V.L.M & Reynolds C.S. (1997) Phytoplankton periodicity and sequences of
dominance in a Amazonian flood-plain lake (lago Batata, Pará, Brazil): responses
to gradual envioronmental changes. Hydrobiologia 346: 169-181.
Hutchinson G.E. (1975) A treatise on limnology 1. Geography, physical and chemistry.
Wiley, New York, 1015 pp.
Ireneé- Marie F.I.C. (1938) Flore Desmidiale de la Region de Montreal. Laprairie,
Canada, 547 pp.
Iturraspe R. (2010) Las turberas de Tierra del Fuego y el Cambio Climático global.
Fundación Humedales / Wetlands International, Buenos Aires, Argentina, 26 pp.
Iturraspe R., Gaviño M. & Urciuolo A. (1998) Caracterización hidrológica de los valles
de Tierra Mayor y Carbajal, Tierra del Fuego. Acta del XVII Congreso Nacional
del Agua y II Simposio de Recursos Hídricos del Cono Sur. Santa Fe, Argentina,
147–156 p.
Iturraspe R. & Roig C. (2000) Aspectos hidrológicos de turberas de Sphagnum de Tierra
del Fuego, Argentina. En: Coronato A. & Roig C. (eds) Conservación de
ecosistemas a nivel mundial con énfasis en las turberas de Tierra del Fuego,
Disertaciones y conclusiones. Ushuaia, Argentina, 85–93 p.
Iturraspe R. & Urciuolo A. (2000) Clasificación y caracterización de las cuencas
hídricas de Tierra del Fuego. Acta del XVIII Congreso Nacional del Agua.
Santiago del Estero, Argentina, CDRom, 17 pp.
Iturraspe R. & Urciuolo A. (2004) Les tourbieres de la Terre de Feu en Argentine: un
patrimonie natural trés menacé. Geocarrefour 79 (4): 143- 152.
Izaguirre I., Allende L., Escaray R., Bustingorry J., Pérez G. & Tell G. (2012)
Comparison of morpho-functional phytoplankton classifications in human-
impacted shallow lakes with diferent stable states. Hydrobiologia 1-14.

BIBLIOGRAFÍA
114
Izaguirre I., Allende L. & Marinone M.C. (2003) Comparative study of the planktonic
communities of three lakes of contrasting trophic status at Hope Bay (Antarctic
Peninsula). Journal Plankton Research 25 (9): 1079- 1097.
Izaguirre I., Mataloni G., Allende L. & Vinocur A. (2001) Summer fluctuations of
microbial planktonic communities in a eutrophic lake, Cierva Point, Antarctica.
Journal Plankton Research 23 (10): 1095- 1109.
Izaguirre I., Mataloni G., Vinocur A., Tell G. (1993) Temporal and spatial variations of
phytoplankton from Boeckella lakes (Hope Bay, Antarctic Peninsula). Antarctic
Science 5(2): 137-141.
Jansson M., Bergström A.K., Blomqvist P., Isaksson A. & Jonsson A. (1999) Impact of
allochthonous organic carbon on microbial food web dynamics and structure in
lake Örträsket. Archiv. Fur Hydrobiologie 144: 409-428.
Jasser I. & Arvola L. (2003) Potential effects of abiotic factor on the abundance of
autotrophic picoplankton in four boreal lakes. Journal of plankton research 25(8):
873-883.
Jasser I., Kostrzewska-szlakowska I., Ejsmont-karabin J., Kalinowska K. & Węgleńska
T. (2009) Autotrophic versus heterotrophic production and components of trophic
chain in humic lakes: the role of microbial communities. Polish journal of ecology
53 (3): 423-439.
Javornický P. (2003) Taxonomic notes on some freshwater planktonic Cryptophyceae
based on light microscopy. Hydrobiologia 502: 271-283.
Jones R. (1992) The influence of humic substances on lacustrine planktonic food chains.
Hydrobiologia 229: 73- 91.
Jones R. (2000) Mixotrophy in planktonic protists: an overview. Freshwater Biology 45:
219-226.
Joosten H. & Clarke D. (2002) Wise use of mires and peatlands. Background and
principles incluing a framework for decision-marking. International Mire
Conservation Group and International Peat Society, Finland, 304 pp.
José de Paggi S. & Paggi J.C. (1995) Determinación de la Abundancia y Biomasa
Zooplanctónica. En: Lopretto E. & Tell G. (eds) Ecosistemas de Aguas
Continentales, Metodologías para su estudio, Ediciones Sur, La Plata, 1: 315-321.

BIBLIOGRAFÍA
115
Jürgens K. (1994) Impact of Daphnia on planktonic microbial food webs, a review.
Marine Microbial Food Webs 8: 295- 324.
Kahl A. (1930) Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 1.
Allgemeiner Teil und Prostomata. En: Dahl F. (ed) Die Tierwelt Deutschlands 18,
Gustav Fischer, Jena 1-180 p.
Kahl A. (1931) Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 2.
Holotricha außer den im 1. Teil behandelten Prostomata. En: Dahl F. (ed) Die
Tierwelt Deutschlands 21, Gustav Fischer, Jena 181-398 p.
Kahl A. (1932) Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 3.
Spirotricha. En: Dahl F. (ed) Die Tierwelt Deutschlands 25, Gustav Fischer, Jena
399-650 p.
Kahl A. (1935) Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 4.
Peritricha und Chonotricha; Nachtrag I. En: Dahl F. (ed) Die Tierwelt
Deutschlands 30, Gustav Fischer, Jena 651-886 p.
Keskitalo J. & Eloranta P. (1999) Limnology of humic water. Backhuys publishers,
Leiden, The Netherlands, 284 pp.
Kingston J.C. (1982) Association and distribution of common diatoms in surface
samples from northern Minnesota peatlands. Ex Beiheft Zur Nova hedwigia 73:
333- 346.
Kirk J. (1994) Light and phosynthesis in aquatic ecosystems. Cambridge University
Press, Cambridge, 509 pp.
Kirk W.L., Postek M.T & Cox E.R. (1976) The Desmid genera Sphaerozosma,
onychonema and teilingia: an historical appraisal. J. Phycol. 12: 5-9.
Kitner M., Poulícková A., Novotny R. & Hájek M. (2004) Desmids
(Zygnematophyceae) of the spring fens of part of West Carpathians. Czech
Phycology, Olomouc, 4: 43-61.
Klug J.L. (2002) Positive and negative effects of allochthonous dissolved organic matter
and inorganic nutrients on phytoplankton growth. Can. J. Fish. Sci. 59: 85-95.
Komárek J. (2006) Cyanobacterial taxonomy: current problems and prospects for the
integration of traditional and molecular approaches. Algae 21 (4): 249-375.

BIBLIOGRAFÍA
116
Komárek J. & Anagnostidis K. (1999) Cyanoprokaryota. 1. Chroococcales. En: Fischer
G. (ed), Süsswasserflora von Mitteleuropa 19/1, Jena, Stuttgart, Lübeck, Ulm, 548
pp.
Komárek J. & Anagnostidis K. (2005) Cyanoprokariota. 2. Teil: Oscillatoriales. En:
Büdel B., Gärtner G., Krienitz L. & Schagerl M. (eds) Süsswasserflora von
Mitteleuropa, Elsevier, Heidelberg, 759 pp.
Komárek J. & Fott B. (1983) Chlorophyceae (Grünalgen) Ordnung: Chlorococcales. En:
Huber Pestalozzi G., Heynig H. & Mollenhauer D. (eds) Das Phytoplankton des
Süsswassers7/1, die Binnengewässer, Stuttgart, 1044 pp.
Köpke K. (2005) Musterbildung in einem feuerländischen Regenmoor. Thesis Inst. Bot.
and Landscape Ecol., University of Greifswald, Germany.
Kouwets F.A.C. (1987) Desmids from the Auvergne (France). Hydrobiologia. 146: 193-
263.
Krammer K. (1992) Pinnularia, eine Monographie der europaischen Taxa. Bibliotheca
Diatomologica 26: 1-353.
Krammer K. (1997a) Die cymbelloiden Diatomeen. Teil 1. Allgemeines und
Encyonema part. Bibliotheca Diatomologica 36: 1-382.
Krammer K. (1997b) Die cymbelloiden Diatomeen. Teil 2. Encyonema part.,
Encyonopsis und Cymbellopsis. Bibliotheca Diatomologica 37: 1-469.
Krammer K. & Lange-Bertalot H. (1986) Bacillariophyceae. 1. Naviculaceae. En: Ettl
H., Gerloff J., Heying H. & Mollenhauer D. (eds), Süsswasserflora von
Mitteleuropa 2/1, Gustav Ficher Verlag, Jena, 876 pp.
Krammer K. & Lange-Bertalot H. (1988) Bacillariophyceae. 2. Bacillariaceae,
Epithemiaceae, Surirellaceae. En: Ettl H., Gerloff J., Heying H. & Mollenhauer D.
(eds), Süsswasserflora von Mitteleuropa 2/2, Gustav Ficher Verlag, Jena, 596 pp.
Krammer K. & Lange-Bertalot H. (1991a) Bacillariophyceae. 3. Centrales,
Fragilariaceae, Eunotiaceae. En: Ettl H., Gerloff J., Heying H. & Mollenhauer D.
(eds) Süsswasserflora von Mitteleuropa 2/3, Gustav Ficher Verlag, Jena, 576 pp.
Krammer K. & Lange-Bertalot H. (1991b) Bacillariophyceae 4. Achnanthaceae,
kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema
Gesamtliteraturverzeichnis Teil 1-4. En: Ettl H., Gerloff J., Heying H. &

BIBLIOGRAFÍA
117
Mollenhauer D. (eds) Süsswasserflora von Mitteleuropa, Fischer, Stuttgart, Jena,
437 pp.
Krieger W. (1937) Die Desmidiaceen. Europas mit Berücksichtigung der
auβereuropäischen Arten. Rabenhorst´s Kryptogammen- Flora 13 (2): 173-712.
Kristiansen J. (2005) Golden algae, A Biology of Chrysophytes. A.R.G Gantner Verlag
K.G., India, 167 pp.
Kristiansen J. & Vigna M.S. (1996) Bipolarity in the distribution of silica-scaled
chrysophytes. En: Kristiansen J. (ed.) Biogeography of freshwater algae, Dev.
Hydrobiol. 118/Hydrobiol. 336: 121-126.
Kristiansen J. & Vigna M.S. (2002) Chrysophyceae y Synurophyceae de Tierra del
Fuego (Argentina). Monogr. Mus. Argentino Cienc. Nat. 3: 1-45.
Krivograd Klemenčič A., Smolar-Žvanut N.,Istenič D. & Griessler- Bulc T. (2010)
Algal community patterns in Slovenian bogs along envioronmental gradients.
Biology 65 (3): 422- 437.
Kruk C., Huszar V.L.M., Peeters E.T.H.M., Bonilla S., Costa L., Lürling M., Reynolds
C.S. & Scheffer M. (2010) A morphological classification capturing functional
variation in phytoplankton. Freshwater Biology 55: 614-627.
Kruk C. & Segura A.M. (2012) The habitat template of phytoplankton morphology-
based functional groups. Hydrobiologia 698 (1): 191-202.
Kuczyńska-Kippen N., Klimaszyk P. & Szelag-Wasielewska E. (2006) Diel changes in
the plankton communities of the ecotone zone a peat mat of the dystrophic lake
glodne jeziorko (drawienski national park). Acta Agrophysica 7(2): 423-431.
Kulikovskiy M. S., Lange-Bertalot H., Witkowski A., Dorofeyuk N. I. & Genkal S. I.
(2010) Diatom Assemblages from Sphagnum bogs of the world. I: Nur Bog in
northern Mongolia. Bibliotheca Diatomologica 55: 1-326.
Lappalainen E. (ed) (1996) Global Peat Resources. International Peat Society and
Geological Survey of Finland, Juskä, 359 pp.
Lenzenweger R. (1993) Beitrag zur Kenntnis der Desmidiaceenflora von Feuerland
(Argentinia). Ach. Protistenkd. 143: 143-152.
Lepistö L. & Holopainen A. (2003) Ocurrence of Cryptophyceae and katablepharids in
boreal lakes. Hydrobiologia 502: 307-314.

BIBLIOGRAFÍA
118
Lindell M.J., Wilhelm Granéli H. & Tranvik L.J. (1996) Effects of sunlight on bacterial
growth in lakes of different humic content. Aquatic Microbial Ecology 11: 135-
141.
Litchman E. & Klausmeier C.A. (2008) Trait- based community ecology of
phytoplankton. Annu. Rev. Ecol. Evol. Syst. 39: 615-639.
Magurran A.E. (2004) Measuring Biological Diversity. Blackwell Science Ltd., Estados
Unidos, 256 pp.
Maidana N.I. (2000) Achnanthaceae (Bacillariophyceae) continentales de Tierra del
Fuego, Argentina. I. Boletín de la Sociedad Argentina de Botánica 35: 49-61.
Mariazi A., Conzonno V., Ulibarrena J., Paggi J. & Donadelli J. (1987) Limnological
investigation in Tierra del Fuego, Argentina. Biología Acuática 10: 1-74.
Marker A.F.H., Crowther C.A. & Gunn R.J.M. (1980) Methanol and acetone as solvents
for estimating chlorophyll-a and phaeopigments by spectrophotometry.
Ergebnisse der Limnologie 14: 52-69.
Marshall W. & Laybourn-Parry J. (2002) The balance between photosynthesis and
grazing in Antarctic mixotrophic cryptophytes during summer. Freshwater
Biology 47: 2060-2070.
Mataloni G. (1991) Remarks on the distribution and ecology of some desmids from
Tierra del Fuego (Argentina). Nova Hedwigia 53: 433-443.
Mataloni G. (1995a) Cyanodictyum reticulatum (Lemm.) Geitl. and Alternantia geitleri
Schill. (Cyanophyceae): Two rare species from Tierra del Fuego (Argentina) peat
bogs. Cryptogamic Botany 5:1-4.
Mataloni G. (1995b) Ecological notes on some interesting desmids from Tierra del
Fuego (Argentina) peat bogs. Nova Hedwigia 60 (1-2): 135-144.
Mataloni G. (1997) Flora algal de las turberas de Tierra del Fuego. Contribución del
Instituto Antártico Argentino 467: 1-81.
Mataloni G. (1999) Ecological studies on algal communities from Tierra del Fuego peat
bogs. Hydrobiologia 391: 157–171.
Mataloni G. & Tell G. (1996) Comparative analysis of the phytoplankton communities
of Rancho Hambre peat bog (Tierra del Fuego, Argentina). Hydrobiologia 325:
101-112.

BIBLIOGRAFÍA
119
Modenutti B., Queimaliños C., Balseiro E. & Reissig M. (2003) Impact of different
zooplankton strutures on the microbial food web of a south andean oligotrophic
lake. Acta Oecologica 24: S289- S298.
Naselli-Flores L., Padisák J. & Albay M. (2007) Shape and size in phytoplankton
ecology: do they matter?. Hydrobiologia 578: 157- 161.
Negro A.I., de Hoyos C. & Aldasoro J.J (2003) Diatom and desmid relationships with
the environment in mountain lakes and mires of NW Spain. Hydrobiologia 505: 1-
13.
Nováková S. (2002) Algal flora of subalpine peat bog pools in the Krkonoše Mts.
Preslia, Praha 74: 45-56.
Nürnberg G.K. & Shaw M. (1999) Productivity of clear and humic lakes: nutrients,
phytoplankton, bacteria. Hydrobiologia 382: 97-112.
Nusch E.A. (1980) Comparison of different methods for chlorophyll and phaeopigment
determination. Arch Hydrobiology Beih Ergebn Limnol 14: 14-36.
Nygaard K. & Tobiesen A. (1993) Bacterivory in algae: a survival strategy during
nutrient limitation. Limnology and Oceanography 38: 273-279.
Orlova-Bienkowskaja M.Y. (1998) A revision of the cladoceran genus Simocephalus
(Crustacea: Daphniidae). Bull. nat. Hist. Mus. Lond. (Zool.) 64: 1-62.
Padisák J., Borics G., Fehér I., Grigorszky I., Oldal I., Schmidt A. & Zámbóné-Doma Z.
(2003) Dominant species and frequency of equilibrium phases in late summer
phytoplankton assemblages in Hungarian small shallow lakes. Hydrobiologia 502:
157-168.
Padisák J. Crossetti L.O & Naselli-Flores L. (2009) Use and misuse in the application of
phytoplankton functional classification: a critical review with updates.
Hydrobiologia 621: 1-19.
Padisák J. & Dokulil M. (1994) Meroplankton dynamics in a saline, turbulent, turbid
shallow lake (Neusiedlersee, Austria and Hungary). Hydrobiologia 289: 23- 42.
Padisák J., Grigorszky I., Borics G. & Soróczki-Pintér É. (2006) Use of phytoplankton
assemblages for monitoring ecological status of lakes within the Water
Framework Directive: the assemblage index. Hydrobiologia 553: 1-14.

BIBLIOGRAFÍA
120
Padisák J. & Reynolds C.S. (1998) Selection of phytoplankton associations in lake
Balaton, Hungary, in response to eutrophication and restoration mesuares, with
special reference to the cyanoprokaryotes. Hydrobiologia 384: 41-53.
Paggi J.C. (1979) Revisión de las especies argentinas del género Bosmina Baird
agrupadas en el subgénero Neobosmina Lieder (Crustacea: Cladocera). Acta
Zoológica Lilloana 35: 137-162.
Paggi J.C. (1995) Crustacea Cladocera. En: Lopretto E.C. y Tell G. (eds) Ecosistemas
de aguas continentales, Metodologías para su estudio. Ediciones Sur, La Plata, 3:
909-951.
Parish F., Sirin A., Charman D., Joosten H., Minaeva T. & Silvius M. (eds) (2008)
Assessment on peatlands, biodiversity and climate change. Global Environment
Centre, Kuala Lumpur and Wetlands International Wageningen, 179 pp.
Porter K.G. & Feig Y.S. (1980) The use of DAPI for identifying and counting aquatic
microflora. Limnology and Oceanography 25: 943-948.
Porter K.G., Sherr E.B., Sherr B.F., Pace M.L. & Sanders R.W. (1985) Protozoa in
planktonic food webs. J. Protozool. 32: 409-415.
Presccott G.W., Croasdale H.T & Vinyard W.C. (1972) A Synopsis of North American
Desmids. Part II. Desmidiaceae: Plascodermae. Section I.University of Nebraska
Press, Lincoln, London, 275 pp.
Presccott G.W., Croasdale H.T & Vinyard W.C. (1977) A Synopsis of North American
Desmids. Part II. Desmidiaceae: Plascodermae. Section II.University of Nebraska
Press, Lincoln, London, 413 pp.
Rabassa J., Coronato, A. & Roig, C. (1996) The peat-bogs of Tierra del Fuego,
Argentina. En: Lappalainen E. (ed) Global Peat Resources, International Peat
Society Publisher, Jyskä, Finland, 261-266 p.
Rakowska B. & Sitkowska M. (2007) Bacillariophyceae and Chlorophyceae of the
Zdzisławów peat-bog at village of Kwilno near the town of Piątek (Łódź
province). Oceanological and Hydrobiological studies 36 (1): 107-112.
Rakowska B. & Sitkowska M. (2010) Algae assemblages in the Rąbién peat-bog
reserve. Oceanological and Hydrobiological Studies, University of Gdańsk,Poland
34 (2): 63-74.

BIBLIOGRAFÍA
121
Reid J.W. (1985) Chave de identificação e lista de refrências bibliográficas para as
espécies continentais sulamericanas de vida livre da ordem Cyclopoida (Crustacea
Copepoda). Boletim de Zoologia, Universidade de São Paulo 9: 17-143.
Revelante N. & Gilmartin M. (1982) Dynamics of phytoplankton in the Great Barrier
Reef Lagoon. J. Plankton Res. 4: 47-76.
Reynolds C.S. (1980) Phytoplankton associations and their periodicity in stratifying
lake systems. Holartic Ecology 3: 141-159.
Reynolds C.S. (1984) The ecology of freshwater phytoplankton, Cambridge University
Press, Cambridge, 384 pp.
Reynolds C.S (1997) Vegetation process in the pelagic: a model for ecosystem theory.
Excellence in ecology. Ecology Institute Oldendorf, 371 pp.
Reynolds C.S (2006) Ecology of phytoplankton. Ecology, Biodiversity and
Conservation, Cambridge, 535 pp.
Reynolds C.S. Huszar V., Kruk C., Naselli-Flores L. & Melo S. (2002) Towards a
functional classification of the freshwater phytoplankton. Journal of plankton
Research 24: 417-428.
Roig C. (2004) Antecedentes sobre turberas en Tierra del Fuego. En: Blanco D.E. & de
la Balze V.M. (eds) Los Turbales de la Patagonia, bases para su inventario y la
conservación de su biodiversidad (Patagonia Peatlands, foundations for
inventories and biodiversity conservation), Fundación Humedales / Wetlands
International, Buenos Aires, Argentina, 33-44 p.
Roig C. & Roig F.A. (2004) Consideraciones generales. En: Blanco D.E. & de la Balze
V.M. (eds) Los Turbales de la Patagonia, bases para su inventario y la
conservación de su biodiversidad, Fundación Humedales / Wetlands International,
Buenos Aires, Argentina, 5-21 p.
Romo S. & Miracle M.R. (1995) Diversity of the phytoplankton assemblages of a
polimictic hypertrophic lake. Arch. Hydrobiol. 132: 363-384.
Ruttner-Kolisko A. (1974) Plankton rotifers. Biology and taxonomy. Die
Biennegewässer 26/1 suppl., E. Schweizerbart'sche Verlagsbuchhandlung (Nägele
u. Obermiller), Stuttgart, 146 pp.

BIBLIOGRAFÍA
122
Rydin H. & Jeglum J. (2006) The biology of peatlands. Oxford University Press,
Oxford, UK, 360 pp.
Saad J. (2011) Impacto de depredación de protistas fagotróficos y su relación con la
producción primaria y bacteriana en un lago oligotrófico y uno distrófico de
Tierra del Fuego (Argentina). Tesis de Licenciatura, Facultad de Ciencias Exactas
y Naturales, Universidad de Buenos Aires, Buenos Aires, 1–45 p.
Salmaso N. & Padisák J. (2007) Morpho-functional groups and phytoplankton
development in two deep lakes (lake Garda, Italy and lake Stechlin, Germany).
Hydrobiologia 578: 97-112.
Sanders R.W. & Porter K.G. (1988) Phagotrophic phytoflagellates. En: Marshall K.C.
(ed), Advances in microbial Ecology 10: 167- 192.
Sanders R.W., Porter K.G., Bennett S.J. & deBiase A.E. (1989) Seasonal patterns of
bacterivory by flagellates, ciliates, rotifers and cladocerans in a freshwater
planktonic community. Limnology and Oceanography 34: 673-687.
Secretaría de la Convención de Ramsar (2010) Uso racional de los humedales:
Conceptos y enfoques para el uso racional de los humedales. Manuales Ramsar
para el uso racional de los humedales, 4ta. Edición, vol.1. Secretaría de la
Convención de Ramsar, Gland (Suiza).
Sharma B.K. & Bhattarai S. (2005) Hydrobiological analysis of a peat bog with
emphasis on its planktonic diversity and population dynamics in Bumdeling
Wildlife Sanctuary, eastern Bhutan. Limnology 6: 183-187.
Sharp J.H., Peltzer E.T., Alperin M.J., Cauwet G., Farrington J.W., Fry B., Karl D.M.,
Martin J.H., Spitzy A., Tugrul S. & Carlson C.A. (1993) Procedures subgroup
report. Mar Chem 41:37-49.
Sherr B.F. & Sherr E.B. (1991) Proportional distribution of total numbers, biovolume,
and bacterivory among size classes of 2-20 �m nonpigmented marine flagellates.
Marine Microbial Food Webs 5 (2): 227-237.
Sherr B.F, Sherr E.B. & Fallon R.D. (1987) Use of monodispersed fluorescently labeled
bacteria to estímate in situ protozoan bacterivory. Appl. Environ. Microbial 53:
958-965.

BIBLIOGRAFÍA
123
Sherr E.B., Sherr B.F. & McDaniel J. (1991) Clearance rates of < 6 �m fluorescently
labeled algae (FLA) by estuarine protozoa: potential grazing impact of flagellates
and ciliates. Marine Ecology Progress Series 69: 81-92.
Sigee D. (2005) Freshwater microbiology. Biodiversity and dynamic interactions of
microorganisms in the aquatic environment. John Willey & Sons Ltd P7, New
Jersey, 524 pp.
Šimek K., Macek M., Pernthaler J., Straškrabová V. & Psenner R. (1996) Can
freshwater planktonic ciliates survive on a diet of picoplankton?. Journal of
Plankton Research 18 (4): 597-613.
Šimek K., Macek M., Seda J. & Vyhnálek V. (1990) Possible food chain relationships
between bacterioplankton, protozoans, and cladocerans in a reservoir. Int. Rev.
ges. Hydrobiol. 74: 583-596.
Smirnov N.N. (1992) The Macrothricidae of the world. Guides to the identification of
the Microinvertebrates of the Continental Waters of the World. 1. SPB Academic
Publishing, The Hague, Netherlands, 143 pp.
Sommer U., Maciej Gliwicz Z., Lampert W. & Duncan A. The PEG-model of seasonal
succession of planktonic events in fresh water (1986). Arch. Hydrobiol. 106 (4):
433-471.
Sommer U., Sommer F., Santer B., Zollner E., Jurgen K., Jamieson C., Boersma M. &
Gocke K. (2003) Daphnia versus copepod impact on summer phytoplankton:
functional compensation at both trophic levels. Oecologia 135: 639-647.
Søndergaard M. (1991) Phototrophic picoplancton in temperate lakes: seasonal
abundance and importance along a trophic gradient. Int Rev Gesamten Hydrobiol
76:505-522.
Starmach K. (1985) Chrysophyceae und Haptophyceae. En: Fischer G. (ed), Pascher
Süsswasserflora von Mitteleuropa, Suttgart, New York, 594 pp.
Stastný J. (2010) Desmids (Conjugatophyceae, Viridiplantae) from the Czech Republic;
new and rare taxa, distribution, ecology. Fottea 10 (1): 1-74.
Štěpánková J., Vavruskova J., Hasler P., Mazalová P. & Poulícková A. (2008) Diversity
and Ecology of desmids of peat bogs in the Jizerské hory Mst. Biologia 63(6):
891-896.

BIBLIOGRAFÍA
124
Stockner J.T. (1991) Autotrophic picoplankton in freshwater ecosystems: the view from
the summit. Internationale Revue der gesamten Hydrobiologie 76: 483- 492.
Sun J. & Liu D. (2003) Geometric models for calculating cell biovolume and surface
area for phytoplankton. Journal of Plankton Research 25 (11): 1331-1346.
Teiling E. (1966) The desmid genus Staurodesmus. A taxonomic study. Du Rietz G.E. y
Lundqvist. Arkiv för Botanik Band 6 nr 11, 467-628 p.
Tell G. (1995) Taxonomía y distribución geográfica de algunas algas de turberas de
Tierra del Fuego (Argentina). Bol. Soc. Arg. Bot. 31 (1-2): 103-112.
Tell G. & Conforti V. (1986) Euglenophyta pigmentadas de la Argentina. Bibliotheca
Phycologica 75: 1-301.
Tell G., Izaguirre I. & Allende L. (2011) Diversity and geographic distribution of
Chlorococcales (Chlorophyceae) in contrasting lakes along a latitudinal transect
in Argentinean Patagonia. Biodiversity and Conservation, 20 (4): 703-727.
Tell G. & Mosto P. (1982) Orden Chlorococcales. En: Guarrera S., Gamundi I. &
Halperin D. (eds), Flora criptogámica de Tierra del Fuego. Buenos Aires, 147 pp.
Thomasson K. (1955) Studies on South-America freshwater plankton.3. Plankton from
Tierra del Fuego and Valdivia. Acta Horti Gotoburgensis 19 (6): 193-225.
Thomasson K. (1956) Reflections on Artic an Alpine lakes. Oikos 7: 117-143.
Thomasson K. (1957) Studies on South-America freshwater plankton. 1. Notes on the
plankton from Tierra del Fuego and Valdivia. Annales Societatis Tartuensis and
Res Naturae Investigandas Constitutae 1: 54-64.
Tilman D., Kilham S.S. & Kilham P. (1982) Phytoplankton community ecology: The
role of limiting nutrients. Annual Review of Ecology and Systematics 13: 349-
372.
Tranvik L.J. & Hansson L.A. (1997) Predator regulation of aquatic microbial abundance
in simple food webs of sub-Antarctic lakes. Oikos, 79: 347–356.
Urabe J., Gurung T.B., Yoshida T., Sekino T., Nakanishi M., Maruo M. & Nakayama E.
(2000) Diel changes in phagotrophy by Crytomonas in Lake Biwa. Limnology
and Oceanography 45: 1558-1563.
Utermöhl H. (1958) Zur Vervollkommung der quatitativen Phytopankton-Methodik.
Mitt. Int. Ver. Theor. Angew. Limnol. 9: 1-38.

BIBLIOGRAFÍA
125
Van de Vijver B., Frenot Y. & Beyens L. (2002) Freshwater diatoms from Ile de la
Possession (Crozet Archipielago, Subantarctica). Bibliotheca Diatomologica 46:
1-412.
Venrick E.L. (1978) How many cells to count? En: Sournia A. (ed) Phytoplankton
Manual. UNESCO, 167-180 p.
Vigna M.S. (1993) Chrysophyta escamosas (Synurophyceae) de una turbera de Tierra
del Fuego (Argentina). Darwiniana 32 (1-4): 299-302.
Vigna M.S & Kristiansen J. (1995 a) Mallomonas fueguina (Synurophyceae), a new
species of silica-scaled chrysophytes from Tierra del Fuego, Argentina. Nord. J.
Bot. 15: 439-442.
Vigna M.S & Kristiansen J. (1995 b) Mallomonas heterospina and M. americana
(Synurophyceae) in Tierra del Fuego, Argentina, with remarks on their
taxonomical interrelation. Darwinia 33: 61-65.
Vigna M.S & Kristiansen J. (1996) Biogeographic implications of new records of scale-
bearing chrysophytes from Tierra del Fuego (Argentina). Arch. F. Protistenk 147:
137-144.
Voigt M. & Koste W. (1978) Rotatoria. Die Rädertiere Mitteleuropas. Überordnung
Monogononta. Bd. I, Textband II. Aufl. Gebrüder Borntraeger, Berlin, Stuttgart,
673 pp.
Voigt M. & Koste W. (1978) Rotatoria. Die Rädertiere Mitteleuropas. Überordnung
Monogononta. Bd II. Textband II. Aufl. Gebrüder Borntraeger, Berlin, Stuttgart,
234 T.
Weisse T. (1993) Dynamics of autotrophic picoplankton in marine and freshwater
ecosystems. En: Jones J. G. (ed) Advances in Microbial Ecology, Kluwer
Academic Publisher, New Yord 13: 327- 370 p.
Weller D.L.M. (1977) Observations on the diet and development of Pseudoboeckella
poppei (Calanoida, Centropagidae) from an Antarctic lake. Br. Antarct. Surv.
Bull. 45: 77–92.
West W. & West G.S. (1904) A monograph of the Bristish Desmidiaceae. Ray Society,
London, 1: 224 pp.

BIBLIOGRAFÍA
126
West W. & West G.S. (1905) A monograph of the Bristish Desmidiaceae. Ray Society,
London, 2: 206 pp.
West W. & West G.S. (1908) A monograph of the Bristish Desmidiaceae. Ray Society,
London, 3: 274 pp.
West W. & West G.S. (1912) A monograph of the Bristish Desmidiaceae. Ray Society,
London, 4: 197 pp.
West W. & West G.S. (1922) A monograph of the Bristish Desmidiaceae. Ray Society,
London, 5: 300 pp.
Wetzel R. (1981) Limnología. Omega, Barcelona, 679 pp.
Wilbert N. (1975) Eine verbesserte Technik der Protargolimprägnation für Ciliaten.
Mikrokosmos 64: 171-179.
Williamson C.E., Morris D.P., Pace M.L. & Olson O.G. (1999) Dissolved organic
carbon and nutrients as regulators of lake ecosystems: resurrection of a more
integrated paradigm. Limnology and Oceanography 44: 795-803.
Williamson D.B. (1992) A contribution to our Knowledge of the Desmid Flora of the
Shetland Islands. Bot. J. Scotl. 46 (2): 233-285.
Yacubson S. (1963) Desmidiaceas de Lapataia (Tierra del Fuego). Revista del museo
Argentino de Ciencias Naturales Bernadino Rivadavia e Instituto Nacional de
Investigación de las Ciencias Naturales. Hidrobiologia 1(5): 157-178.
Zar, J.H. (2010) Biostatistical Analysis. 5th edition, Pearson Prentice-Hall, Upper Saddle
River, New Jersey, 944 pp.

ANEXO

ANEXO 1
127
Variación temporal de la abundancia de los gremios alimenticios del metazooplancton en
los cinco cuerpos de agua de la turbera de Rancho Hambre.
Referencias: Mic: microfiltradores; Mac: Cladóceros macrofiltradores; Per: Cladóceros perifitófagos; Cof:
Copépodos Calanoida filtradores; Com: Copépodos macrófagos herbívoros; Pre: Predadores o macrófagos
carnívoros.

ANEXO 1
128
Variación temporal porcentual de la abundancia de los gremios alimenticios del metazooplancton en los cinco cuerpos de agua de la turbera de Rancho Hambre.
Referencias: Mic: microfiltradores; Mac: Cladóceros macrofiltradores; Per: Cladóceros perifitófagos; Cof:
Copépodos Calanoida filtradores; Com: Copépodos macrófagos herbívoros; Pre: Predadores o macrófagos
carnívoros.
Top Related