
Bioenergética II - Parte 1 - Ciclo de Krebs y Fosforilación Oxidativa - parte 1
Upload
lyz-honorato-soledadCategory
view
54download
2

Mitocondriasitio donde ocurre la fosforilación oxidativa en
células eucariotas (1948)
Albert L. Lehninger1917-1986
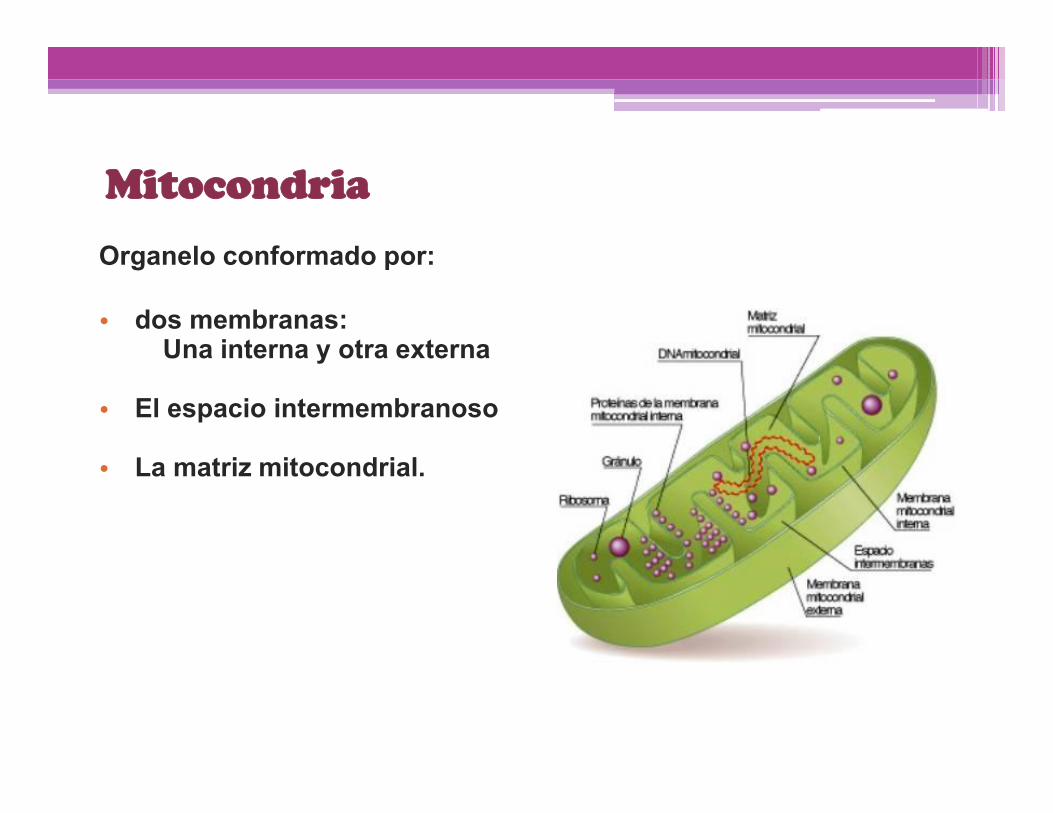
Mitocondria
Organelo conformado por:
• dos membranas:Una interna y otra externa
• El espacio intermembranoso
• La matriz mitocondrial.

Membrana Interna· Alto nivel de especialización, por lo cual desarrolla las crestas
mitocondriales
· Moléculas que contiene:-Proteínas transportadoras-Complejos enzimáticos: NADH deshidrogenasa, Citocromo-Oxidasa,- Ubiquinona y Citocromo C.-ATP sintetasa.
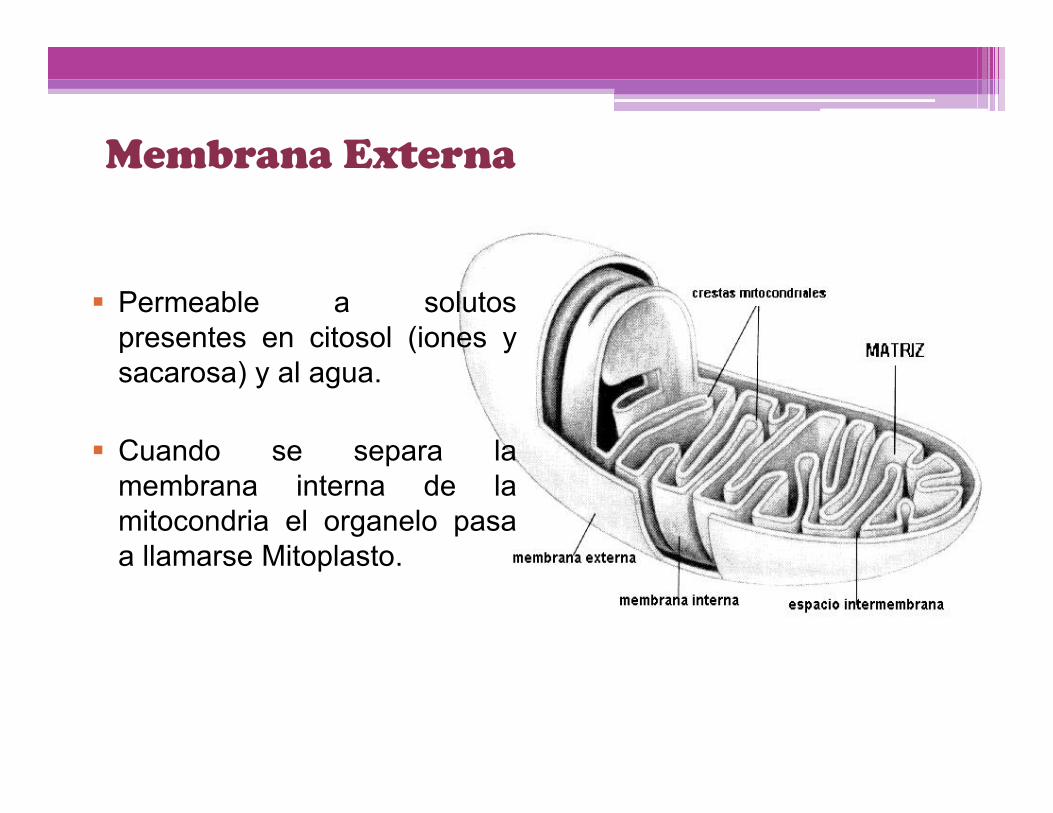
Membrana Externa
Permeable a solutospresentes en citosol (iones ysacarosa) y al agua.
Cuando se separa lamembrana interna de lamitocondria el organelo pasaa llamarse Mitoplasto.
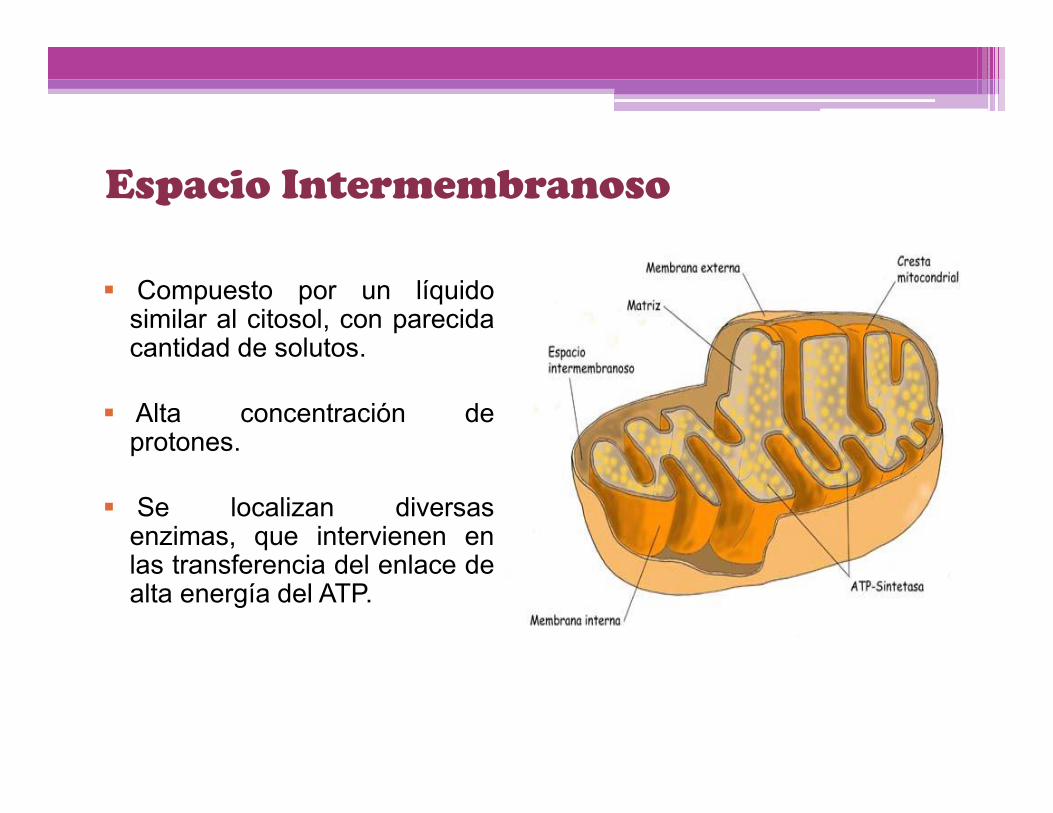
Espacio Intermembranoso
Compuesto por un líquidosimilar al citosol, con parecidacantidad de solutos.
Alta concentración deprotones.
Se localizan diversasenzimas, que intervienen enlas transferencia del enlace dealta energía del ATP.

Matriz Mitocondrial
Parte interna que conforma la mitocondria.
En la Matriz se cumplen ciclos fundamentales para el desarrollo dela Mitocondria, como el Ciclo de Krebs, la beta oxidativa de losácidos grasos, se oxida aminoácidos, entre otros.
Componentes de la Matriz:- Ribosomas- Enzimas- ADN mitocondrial- Coenzimas A (CoA) y NAD+- O2, ADP y fosfato

Las únicas moléculas que cruzan lamembrana interna son aquellas paralas que hay proteínas transportadorasespecíficas. La membrana internaposee transportadores de metabolitosesenciales (ADP, ATP, ácidoscarboxílicos, Ca2+, aminoácidos, etc.)
La membrana interna aloja a lasproteínas pertenecientes de loscomponentes de la cadenarespiratoria y el complejo enzimáticoresponsable de la síntesis de ATP(ATP sintasa)

Transportadores universales de electrones- Nucleótidos de piridina (NAD y NADP)- Nucleótidos de flavina (FMN y FAD)
Otros grupos transportadores de electrones- Ubiquinona ó Coenzima Q (benzoquinona)- Citocromos- Proteínas ferro-sulfuradas
Cadena respiratoria mitocondrial ócadena de transporte de electrones
proteínas

Sustrato reducido + NAD+ Sustrato oxidado + NADH + H+
Sustrato reducido + NADP+ Sustrato oxidado + NADPH + H+
Los electrones son canalizados haciaaceptores universales de electrones
La mayor parte de los e- que entran en la cadena respiratoriaproviene de la acción de deshidrogenasas que capturan e-provenientes de reacciones catabólicas, canalizándolos en formade pares de e- hacia aceptores universales
Aceptores universales de e-: NAD, NADP, FMN ó FAD
Las deshidrogenasas ligadas a nucleótidos de piridina, catalizan:
Las deshidrogenasas ligadas al NAD eliminan dos átomos dehidrógeno de sus sustratos (:H- y H+)

El NADH y el NADPH son transportadores electrónicoshidrosolubles, asociados reversiblemente con deshidrogenasas
El NADH actúa como transportador de difusión, llevando e- queprovienen de reacciones catabólicas a su punto de entrada en lacadena respiratoria, el complejo NADH deshidrogenasa (Complejo I)
La flavoproteínas contienen nucleótidos de flavina (FAD o FMN)fuertemente unido, a veces covalentemente. El nucleótido oxidadopuede aceptar un e- (dando la forma semiquinona) o dos e- dandoFADH2 o FMNH2
NADPH + NAD+ NADP+ + NADH
El NAD+ puede colectar equivalentes de reducción a partir del NADPH.Esta reacción es catalizada por las enzimas transhidrogenasas:
Los electrones son canalizados haciaaceptores universales de electrones

La cadena respiratoria consta de una serie de transportadoreselectrónicos, la mayoría proteínas integrales de membrana, congrupos prostéticos capaces de aceptar y donar 1 ó 2 e-
Cada componente de la cadena acepta e- del transportadorprecedente y se los transfiere al siguiente en una secuenciaespecífica
Tipos de transferencia de e- (equivalentes de reducción) en lacadena respiratoria:
(1) transferencia directa de e- (Fe3+/Fe2+)(2) transferencia de un H+ (1 e-)(3) transferencia de un hidruro (:H-) portador de 2 e-
Cadena respiratoria mitocondrial ócadena de transporte de electrones

Otros grupos transportadores de electronesUbiquinona o Coenzima Q
La reducción completa (UQH2)requiere 2 e- y 2 H+ y se produceen 2 pasos sucesivos
La UQ puede aceptar 1 e-formándose radical semiquinona(UQH)
Capaz de actuar como uniónentre un dador de 2 e- y unaceptor de 1 e-
Debido a que es pequeña ehidrofóbica difunde a través de lamembrana interna, actuando delanzadera de equivalentes dereducción entre otrostransportadores electrónicos de lamembrana, menos móviles
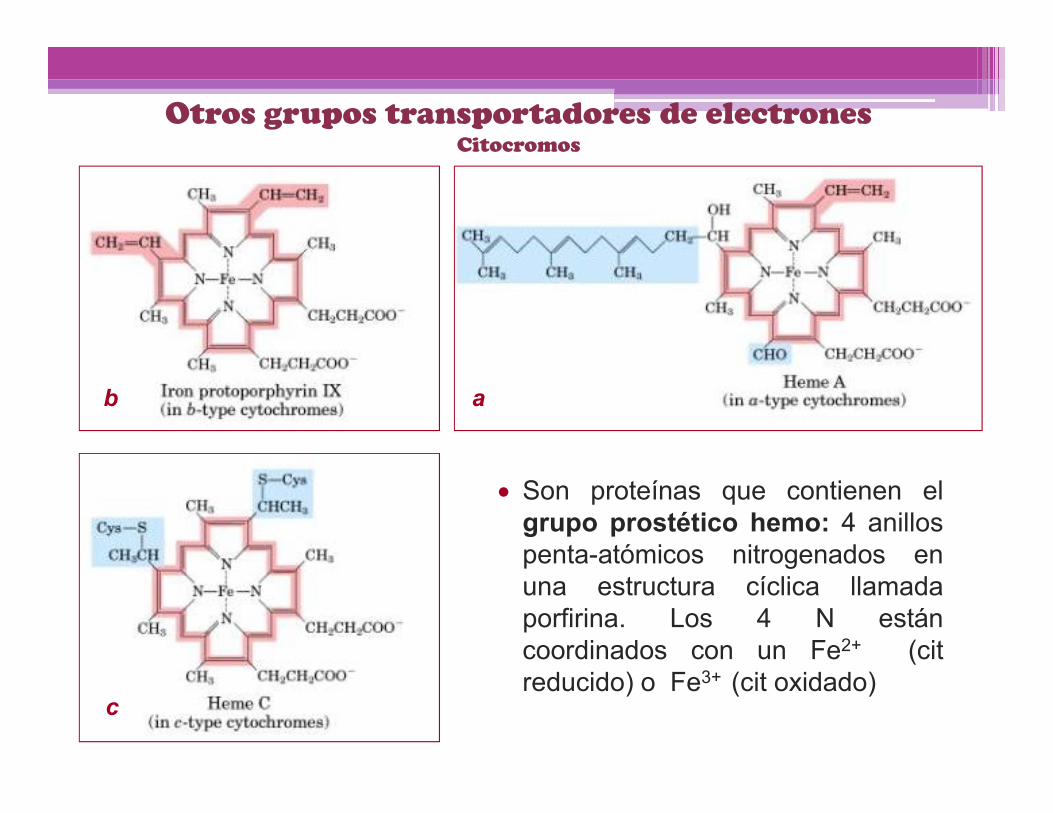
Otros grupos transportadores de electronesCitocromos
Son proteínas que contienen elgrupo prostético hemo: 4 anillospenta-atómicos nitrogenados enuna estructura cíclica llamadaporfirina. Los 4 N estáncoordinados con un Fe2+ (citreducido) o Fe3+ (cit oxidado)
ab
c

Otros grupos transportadores de electronesProteínas ferro-sulfuradas
En las proteínas ferro-sulfuradas, el hierro está presente no en forma de hemo,sino en asociación con átomos de azufre inorgánico o con azufre de residuosCys de la proteína, o con los dos simultáneamente
Centros (Fe-S): estructuras sencillas a complejas
Participan en reacciones de transferencia de 1 e- en la que se oxida o reduceuno de los átomos de Fe
Al menos, 8 proteínas Fe-S intervienen en la cadena e transporte de e-
Potenciales de reducción: -0.65V a + 0.45 V

E’o (mV)
NADH - 320FMNH - 290Fe-S (Complejo I) - 270FADH - 10Fe-S (Complejo II) 20Ubiquinol (UQH2) 50Citocromo bk 77Citocromo bT 190Citocromo c (+c1) 230Citocromo a+a3 380Oxigeno 816
Potenciales de reducción estándar de los transportadores de la cadenarespiratoria mitocondrial
Los e- fluyen espontáneamente desde los transportadores de E’omás bajo hacia los transportadores con E’o más elevado. En la tabla,los de más arriba reducen a los de mas abajo

Una serie de moléculas con grupos prostéticos redox con potencialesde reducción estándar crecientes desde el NADH (-320 mV) alO2 (+ 816 mV)
Cadena respiratoria mitocondrial ócadena de transporte de electrones
Transporte de e- mitocondrial: reacción global
NADH Fp(FMN) UQ cit bKbT cit c,c1 cit aa3 O2

Los electrones son canalizados hacia trasportadoresuniversales de electrones
• La mayor parte de los electrones que entran en la cadenarespiratoria mitocondrial provienen de la acción de deshidrogenasasque captan electrones de las reacciones oxidativas del complejo dela piruvato deshidrogenasa, del ciclo de krebs, de la ruta de la b-oxidación, canalizándolos en forma de pares electrónicos hacia lacadena respiratoria.
• Utilizan nucleótidos de piridina (NAD o NAPH) o nucleótidos deflavina (FMN o FAD) como aceptores electrónicos.


La CTE mitocondrial consta de una serie de transportadoreselectrónicos, la mayoría proteínas integrales de membrana, congrupos prostéticos capaces de aceptar y donar uno o dos electrones.Cada componente de la cadena puede aceptar electrones deltrasportador precedente y transferirlos al siguiente en una secuenciaespecífica.
Algunas de las reacciones de esta secuencia son transferencias deun electrón mientras que otras son transferencias de pares deelectrones.
Además del NAD y de las flavoproteínas, hay otros tres tipos detransportadores de electrones que funcionan en la cadenarespiratoria: una benzoquinona hidrofóbica (ubiquinona) y dos tiposdiferentes de proteínas con hierro (citocromos y proteínas ferro-sulfuradas)

COENZIMA Q, O UBIQUINONA(UQ)
• Larga cadena lateralisoprenoide.
• Puede aceptar un electróntransformándose en el radicalsemiquinona o dos electronesformando ubiquinol (UQH2)

LOS CITOCROMOS
• Proteínas transportadorasde electrones que
contienen hierro situadasen la membrana interna
mitocondial



Componentes de la cadena respiratoria mitocondrialpH = 7.0Espacio intermembranas (Lado P)
pH = 7.8Matriz (Lado N)
Reacciones de transferenciade e- se realiza a altavelocidad por estar lostransportadores de e-incorporados en lamembrana interna(restringidos en susmovimientos) y asociadosentre sí (unionesintermoleculares)
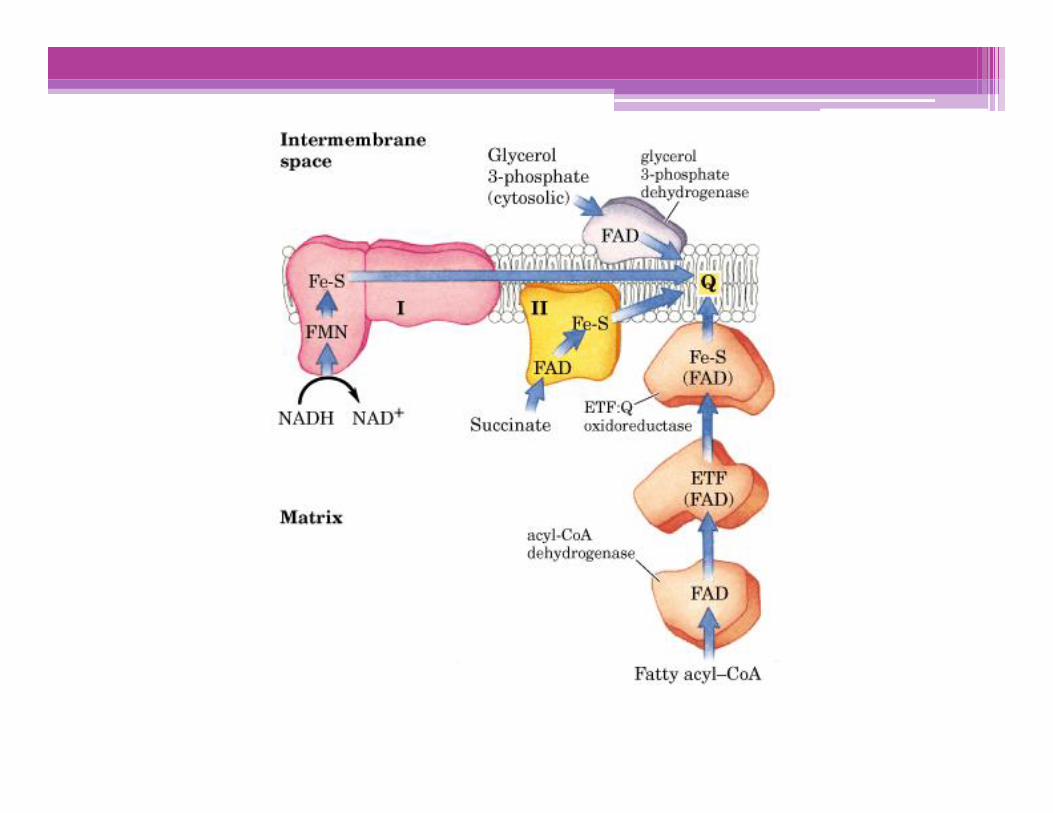

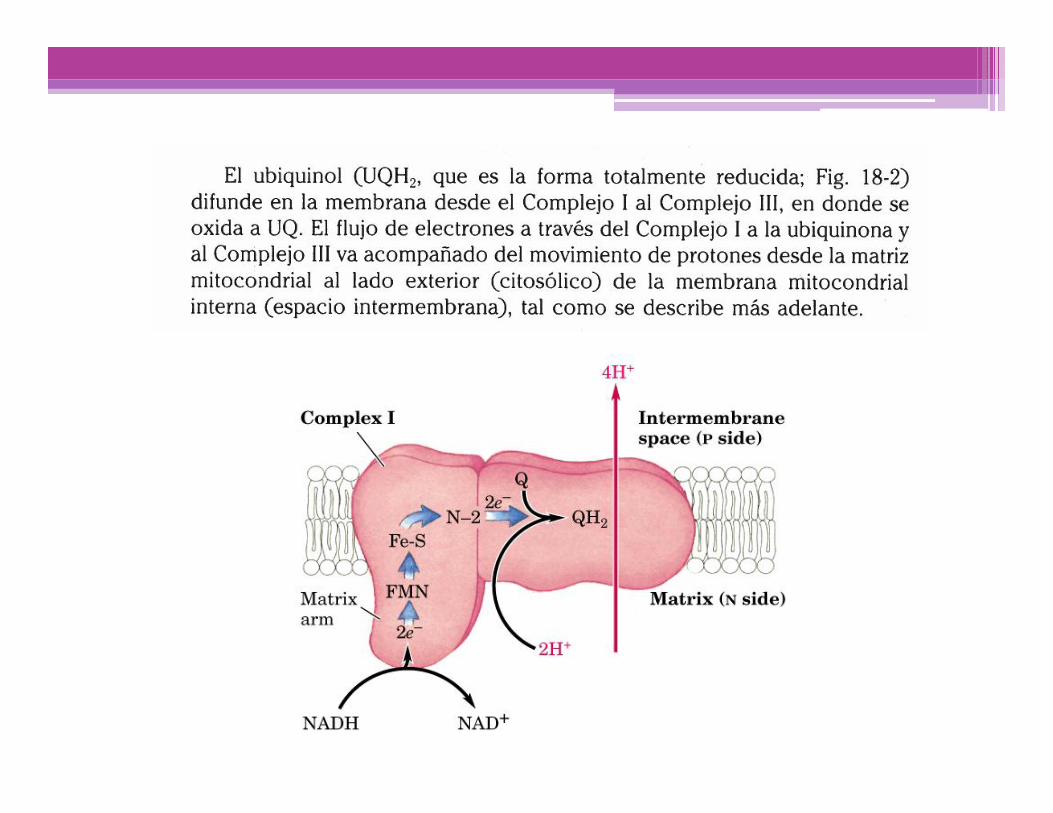


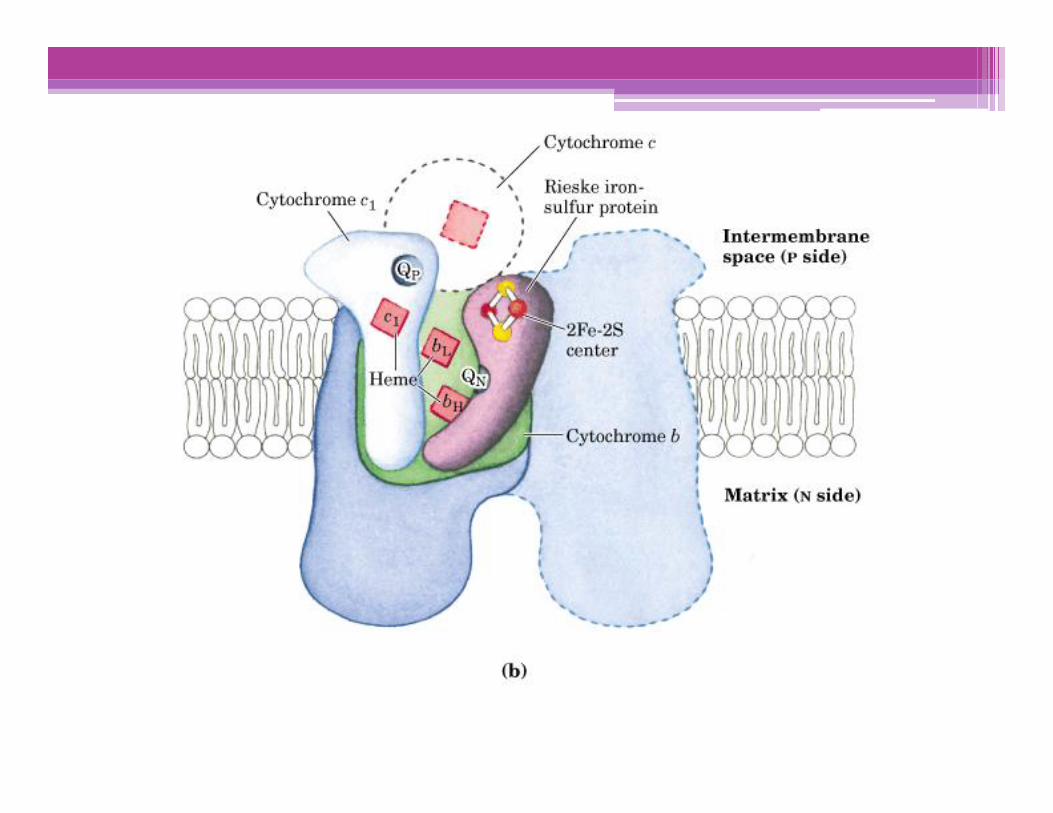


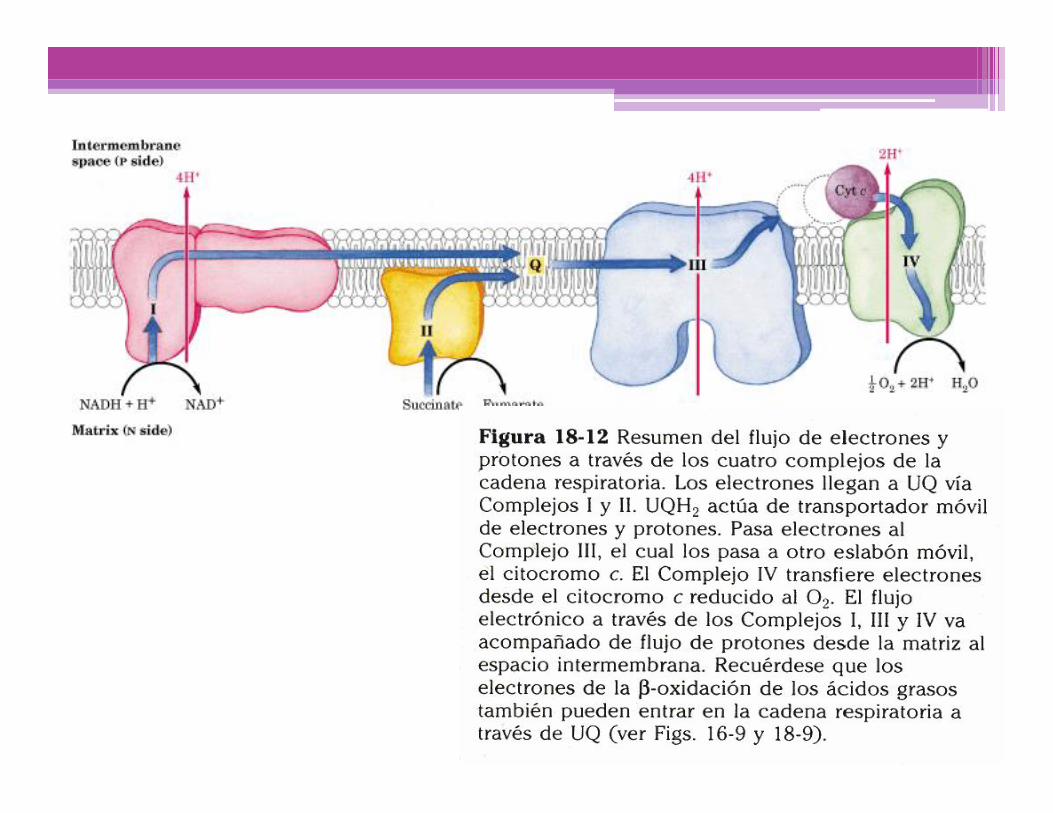

Translocación de H+ asociada al flujo de electrones
pH = 7.0Espacio intermembranas (Lado P)
pH = 7.8Matriz (Lado N)
Por cada par de e- transferidos al O2, 4 H+ son bombeados por elComplejo I, 4 H+ por el Complejo III y 2 H+ por el complejo IV; todosellos desde la matriz mitocondrial (Lado N), hacia el espaciointermembranas (Lado P)
NADH + 11 H+(N) + 1/2 O2 H2O + NAD+ + 10 H+
(P)
En las condiciones celulares, la oxidación mitocondrial del NADH o del succinato libera unaenergía superior a la necesaria para la síntesis de ATP
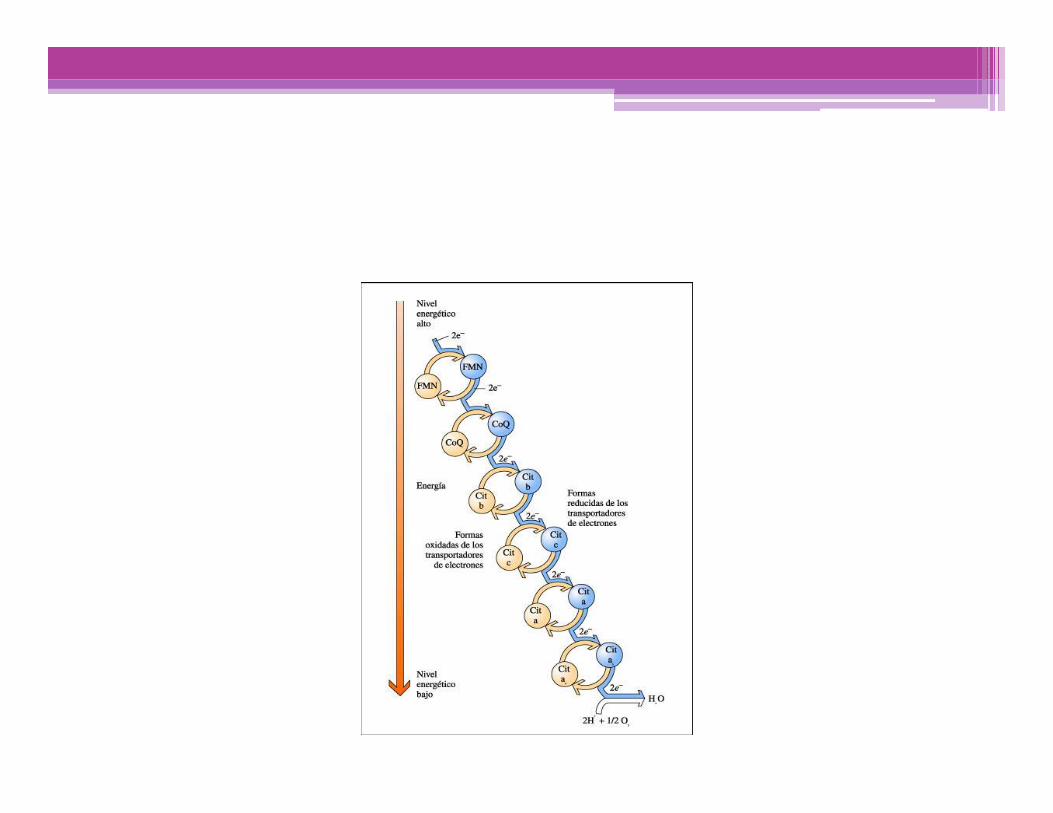


• La transferencia de electrones a través de la cadena respiratorialibera energía más que suficiente para formar ATP.
• La enzima ATP sintasa, es el equivalente de una bomba de protonesdependiente de ATP, donde el flujo de protones a favor de sugradiente electroquímico a través de esta bomba impulsa lacondensación de Pi y ADP.
• La fosforilación oxidativa se refiere a la síntesis química de ATPimpulsada por el proceso exergónico de transferencia de electronesdesde el NADH al O2
• La fosforilación oxidativa comienza con la entrada de e- en la cadenarespiratoria
FOSFORILACIÓN OXIDATIVA
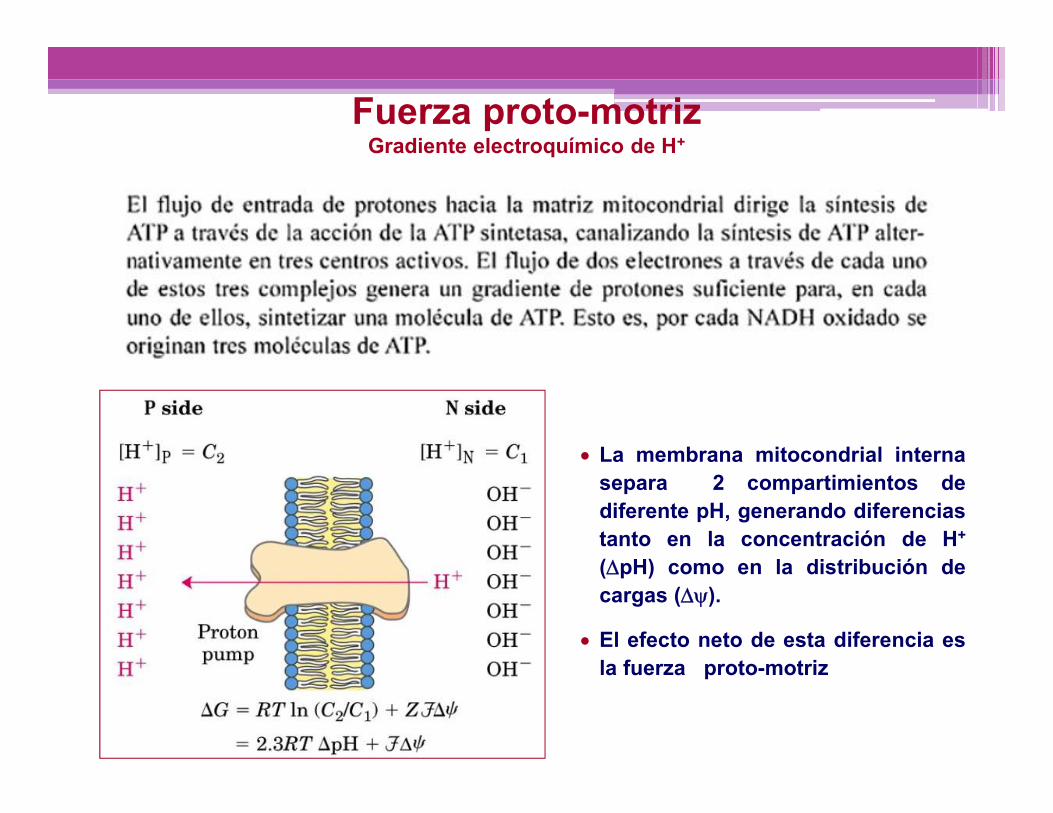
La membrana mitocondrial internasepara 2 compartimientos dediferente pH, generando diferenciastanto en la concentración de H+
(pH) como en la distribución decargas ().
El efecto neto de esta diferencia esla fuerza proto-motriz
Fuerza proto-motrizGradiente electroquímico de H+
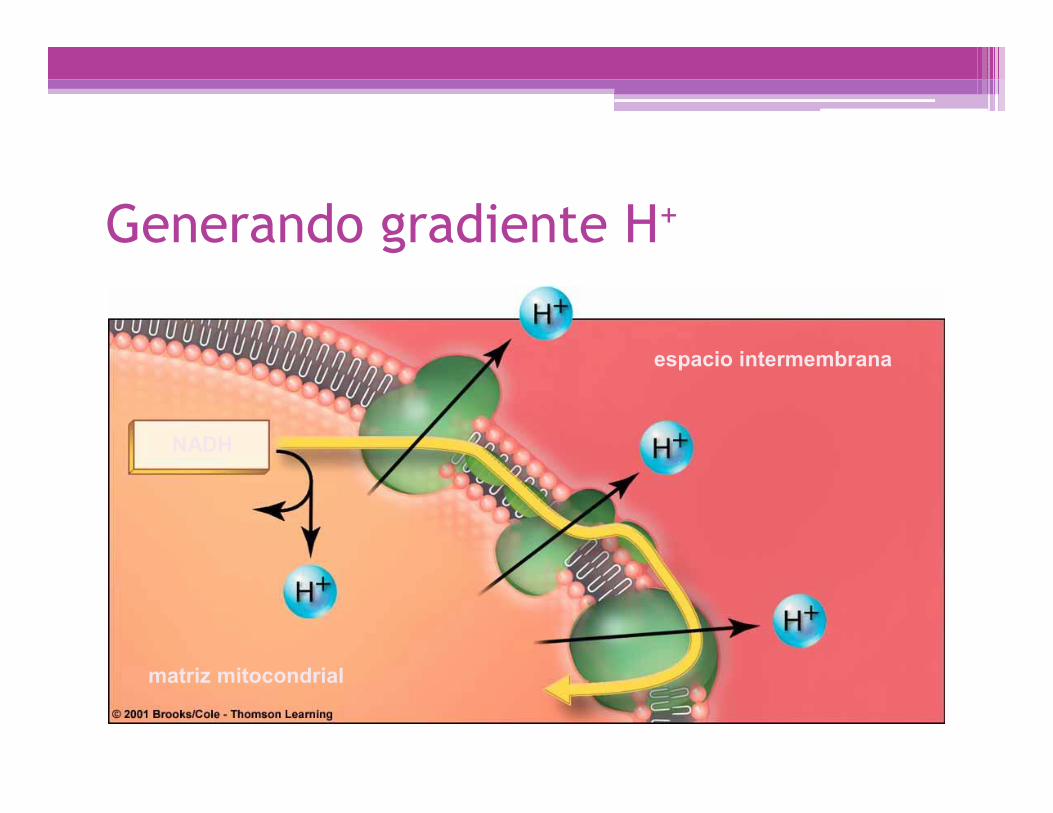
Generando gradiente H+
NADH
espacio intermembrana
matriz mitocondrial

La teoría quimiosmótica mitocondrial(P. Mitchell, 1961)
Peter Mitchell1920-1992
The Nobel Prize in Chemistry 1978"for his contribution to the understandingof biological energy transfer through theformulation of the chemiosmotic theory"
El gradiente electroquímico de protones a través de lamembrana interna mitocondrial (fuerza proto-motriz)
es crucial para los procesos de transducción de energíay síntesis de ATP

Membranamitocondrialinterna
Matrizmitocondrial (N)
Espaciointermembrana (P)
ADP + Pi ATP
O2
FP b c a a3
H+ H+ H+ pH = 7.0
pH = 7.8
ATPasa
H+
La teoría quimiosmótica mitocondrial(P. Mitchell, 1961)
La fuerza proto-motriz, lleva a la síntesis de ATP, comoconsecuencia del flujo pasivo de H+ hacia la matriz mitocondrial, através de un poro de H+ (F0) asociado a la ATP sintasa (Complejo V)
ADP + Pi + nH+(p) ATP + H2O + nH+
(N)

Produccción ATP:Modelo Quimiosmótico
ATP
ADP+Pi
MATRIZ MITOCONDRIAL

Cadena respiratoria y Fosforilación oxidativa

Fosforilación oxidativaSíntesis de ATP acoplada al flujo de electrones
Cuando se suspenden mitocondrias en una soluciónamortiguadora, que contiene un sustrato oxidable, ADP yPi, ocurren 3 procesos:
(1) se oxida el sustrato(2) se consume O2 (hay respiración)(3) se sintetiza ATP
Si el dador de e- es el NADH, las mitocondrias sintetizan 3ATP por cada par de e- pasados al O2
Si el dador es succinato, se sintetizan 2 ATP
El consumo de O2 y la síntesis de ATP son dependientesde la presencia de un sustrato oxidable y de ADP y Pi

Postulados de la teoría quimiosmótica mitocondrial(P. Mitchell, 1961)
El transporte de e- a través de la cadena respiratoria está asociado altransporte de H+ desde la matriz hacia el espacio intermembranas
La membrana interna es impermeable a los H+
Se conserva la energía de oxidación de los procesos metabólicos enforma de potencial electroquímico, ya que se genera un gradienteelectroquímico de H+
La cadena respiratoria está acoplada a la síntesis de ATP
El flujo de H+ a favor de su gradiente electroquímico proporciona laenergía libre para la síntesis de ATP. Las concentraciones de H+ enlas 2 fases acuosas (EIM y M) separadas por la membrana interna,constituyen la fuerza responsable (fuerza proto-motriz) de la formaciónde ATP a partir de ADP y de Pi, por acción de la F1-ATPasa de lamembrana mitocondrial

TEORÍA DE MITCHELL•El flujo de electrones producido por la cadena de electrones esbombeado al espacio intermembrana de la mitocondria
• Esto crea un gradiente electroquímico gracias al cual los protonestienden a volver al interior del orgánulo.
•Los protones atraviesan la ATP-Sintetasa ingresando a la matrízmitocondrial.
•Este paso conlleva un desprendimiento de energía que se utiliza paraque un ADP se una a un grupo fosfato, dando lugar al ATP.

Síntesis de ATP por ATP sintasa
John Walker
The Nobel Prize in Chemistry 1997"for their elucidation of the enzymaticmechanism underlying the synthesis ofadenosine triphosphate (ATP)"
Paul Boyer
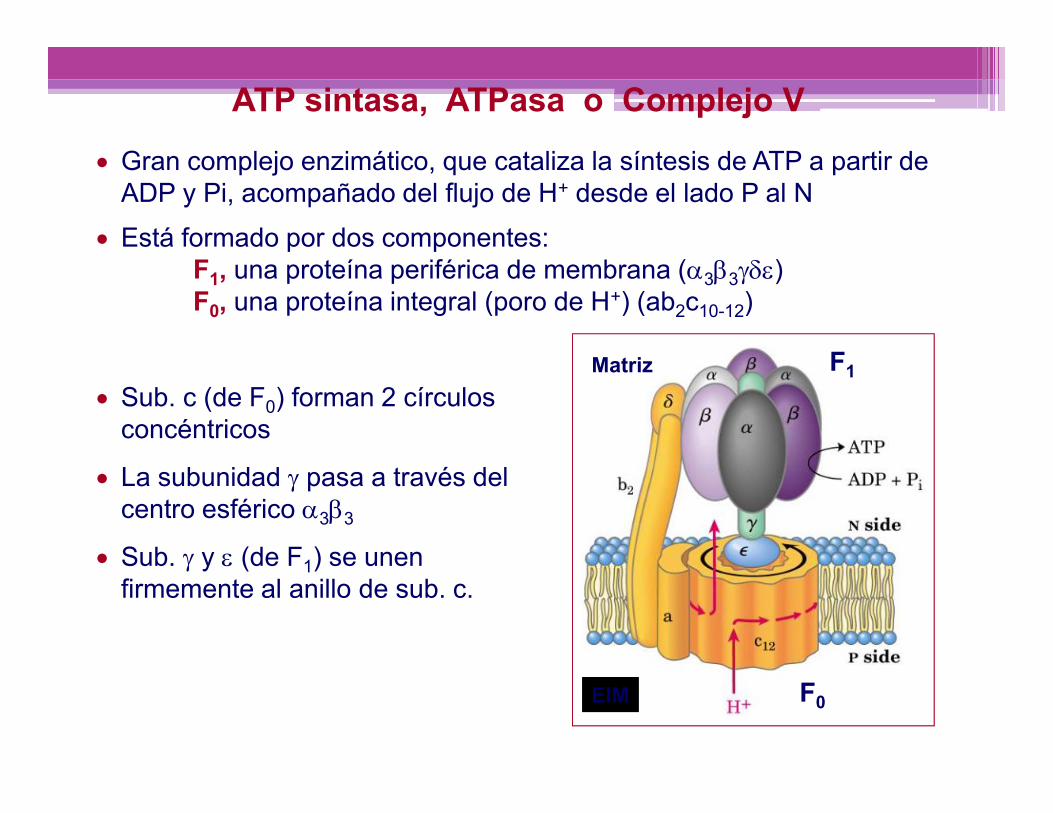
ATP sintasa, ATPasa o Complejo V Gran complejo enzimático, que cataliza la síntesis de ATP a partir de
ADP y Pi, acompañado del flujo de H+ desde el lado P al N Está formado por dos componentes:
F1, una proteína periférica de membrana (33)F0, una proteína integral (poro de H+) (ab2c10-12)
Sub. c (de F0) forman 2 círculosconcéntricos
La subunidad pasa a través delcentro esférico 33
Sub. y (de F1) se unenfirmemente al anillo de sub. c.
F1
F0
Matriz
EIM

Las translocación de H+ a través del poro F0 provoca que el cilindro desub. c y la subunidad adjunta, roten alrededor del eje de g(perpendicular al plano de la membrana)
ATP sintasa, ATPasa o Complejo V
El pasaje de H+ a través de F0 llevan acambios conformacionales de lasubunidad de la F1-ATPasa El proceso endergónico de rotación de la
sub. es impulsado por el procesoexergónico de la translocación de H+ dellado P al N
F1-ATPasa = Rotor molecular
El acoplamiento no es químico,sino electroquímico y físico

La fosforilación oxidativa se refiere a la síntesis químicade ATP impulsada por el proceso exergónico detransferencia de electrones desde el NADH al O2
FOSFORILACIÓN OXIDATIVA
• Ocurre en la mitocondria• La fosforilación oxidativa comienza con la entrada de e- en la cadena
respiratoria• Los e- pasan a través de una serie de transportadores incluidos en la membrana
interna mitocondrial• Los transportadores electrónicos mitocondriales funcionan dentro de complejos
proteicos ordenados en serie• La cadena de transporte de e- es un proceso exergónico, que libera energía
suficiente para la síntesis de ATP• Existe una translocación de H+ desde la matriz hacia el EIM (fuerza protomotriz)• Síntesis de ATP por ATP sintasa



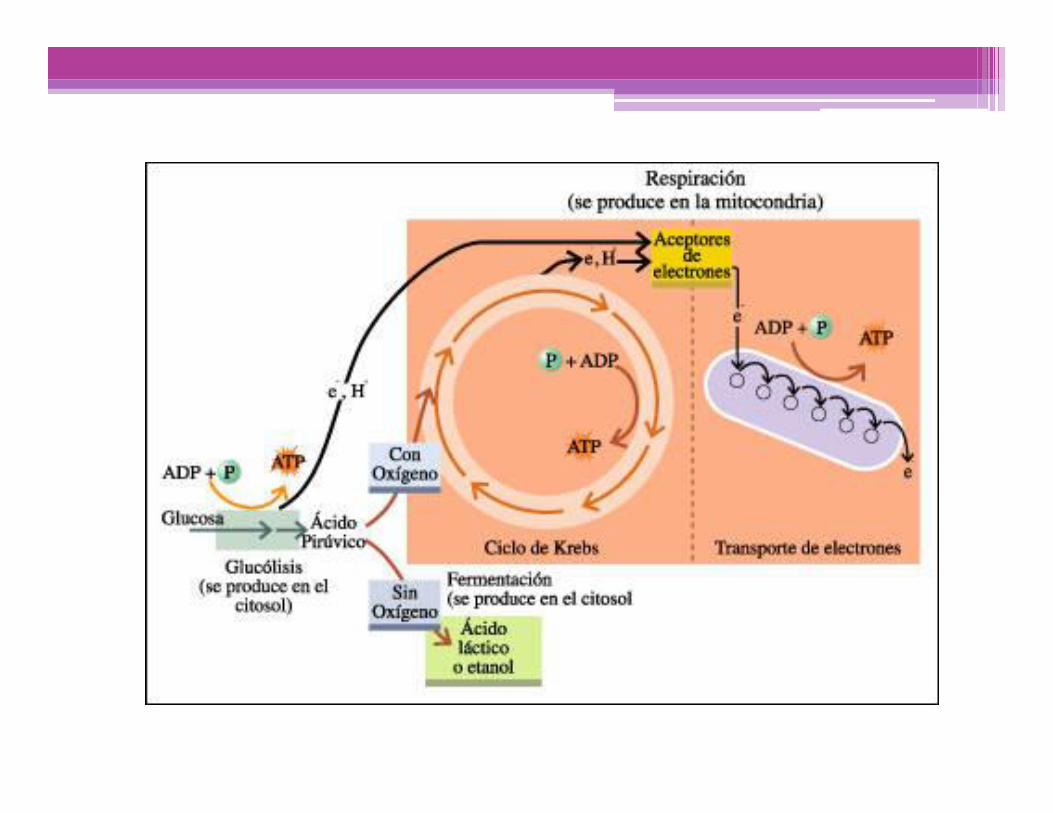




ESPACIO INTERMEMBRANA
MATRIZ MITOCONDRIAL
Cadena transportadora Electrones ATP Sintetasa
ATP
H+
H+H+ H+
H+
H+H+ H+
H+H+
H+
H+H+
H+
H+
H+
H+
H+H+
H+
H+
H+
H+
H+
NADH + H+ NAD+ + 2H+ FAD + 2H+FADH2 2H+ + 1/2 02 H2O ADP + Pi
e-e- e-
Fosforilización
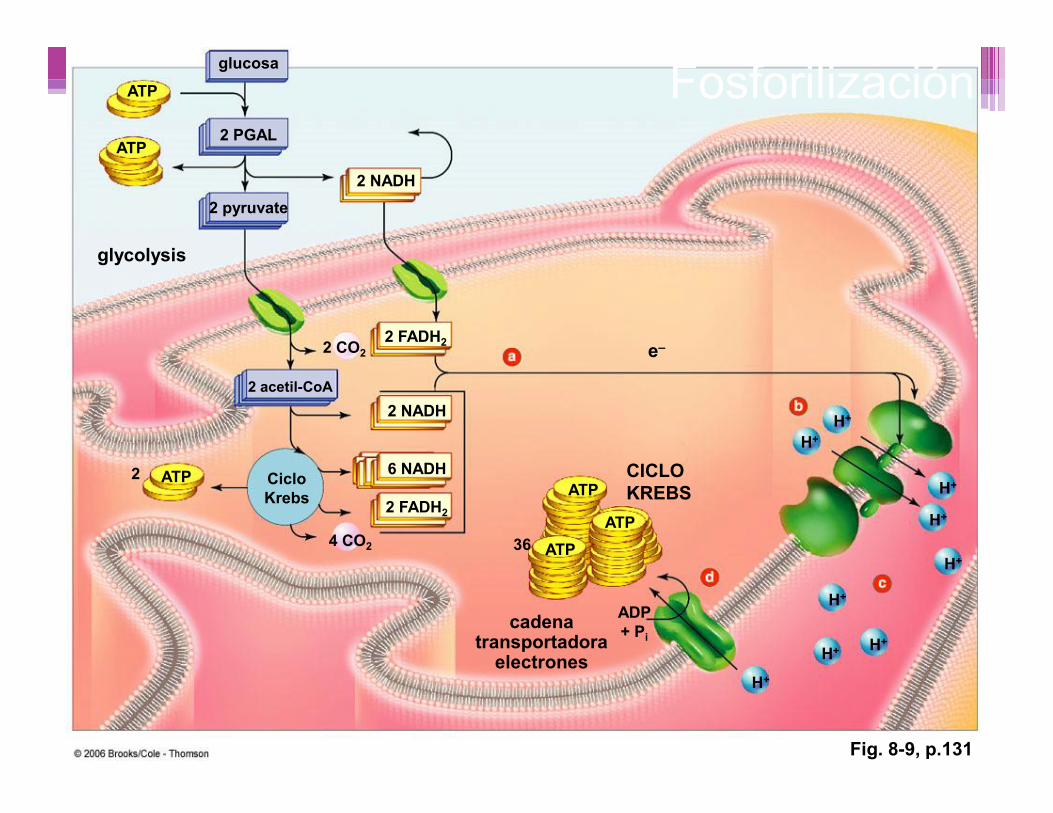
glucosa
glycolysis
e–
CICLOKREBS
cadenatransportadora
electrones
2 PGAL
2 pyruvate2 NADH
2 CO2
ATP
ATP
2 FADH2
H+
2 NADH
6 NADH
2 FADH2
2 acetil-CoA
ATP2 CicloKrebs
4 CO2
ATPATP
ATP
36
ADP+ Pi
H+
H+
H+
H+
H+
H+H+
H+
Fig. 8-9, p.131
Fosforilización

Importancia Oxígeno• Cadena transportadora electrones requiere
oxígeno
• Oxígeno recoge electrones de la cadena y loscombina con H+ para formar agua

Resumen Ganancia de Energía(por molécula glucos)• Glucólisis
▫ 2 ATP
• Ciclo Krebs y reacciones preparación▫ 2 ATP
• Cadena transportadora electrones▫ 32 ATP

• 686 kcal de energía se liberan
• 7.5 kcal se conservan en cada ATP
• Cuando se forman 36 ATP, 270 kcal (36 X 7.5) se
capturan en ATP
• Eficiencia = 270 / 686 X 100 = 39%
• Mayoría energía se pierde en calor
Eficiencia de respiración aeróbica

ATPasa

VOLVERComp.

AcCoA
CO2
CO2

Fosforilación Oxidativa y Cadena Transportadora de Electrones.














![FOSFORILACIÓN OXIDATIVA - [DePa] Departamento de ...depa.fquim.unam.mx/amyd/archivero/FosforilacionOxidativa_35344.pdfy aloja a los componentes de la cadena la fosforilaciÓn oxidativa](https://static.fdocuments.es/doc/165x107/5eb606cf0882961e336d88ce/fosforilacin-oxidativa-depa-departamento-de-depafquimunammxamydarchiverofosforilacionoxidativa35344pdf.jpg)


