
1. INTRODUCCIÓNucv.altavoz.net/prontus_unidacad/site/artic/...Otro factor de huerto que incide en...
69
1. INTRODUCCIÓN Actualmente en nuestro país hay alrededor de 7.663 hectáreas (ha) de limonero, las que se encuentran distribuidas entre la III y VIII región, siendo la región Metropolitana la que posee la mayor superficie plantada en el país con 3.065 ha, seguido de la VI con 1580 ha, y en tercer lugar se encuentra la V con 1.570 ha (ODEPA, 2004). El limón (Citrus limon) es uno de los frutos más importantes en Chile, comercializándose en el mercado interno y externo. La exportación se realiza en la época invernal, siendo Estados Unidos el principal país de destino, sin embargo China se presenta hoy en día como el segundo socio comercial de Chile, ya que su economía posee perspectivas de crecimiento cada vez más altas, representando para los productores chilenos grandes posibilidades, lo que debe ser asumido como un gran desafío, esto significa no sólo llegar con una fruta de calidad al país de destino, sino también conservar su condición durante el proceso de comercialización, y satisfacer de este modo las expectativas del mercado. Para definir la calidad de los limones, los países que importan y exportan cítricos han dictado normas oficiales con fines comerciales. Éstas se basan en las características externas del fruto, tales como color, tamaño, ausencia de defectos en la cáscara, relación sólidos solubles/acidez y contenido de jugo mínimo (MAGDAHL, 1998) Uno de los principales problemas que se presenta es el desorden fisiológico peteca, la que se desarrolla principalmente en la época invernal, dañando la apariencia del fruto, lo que provoca una disminución de su valor comercial y un rechazo a nivel de exportación (RAZETO, 2000).
Transcript of 1. INTRODUCCIÓNucv.altavoz.net/prontus_unidacad/site/artic/...Otro factor de huerto que incide en...

1. INTRODUCCIÓN
Actualmente en nuestro país hay alrededor de 7.663 hectáreas (ha) de limonero, las
que se encuentran distribuidas entre la III y VIII región, siendo la región
Metropolitana la que posee la mayor superficie plantada en el país con 3.065 ha,
seguido de la VI con 1580 ha, y en tercer lugar se encuentra la V con 1.570 ha
(ODEPA, 2004).
El limón (Citrus limon) es uno de los frutos más importantes en Chile,
comercializándose en el mercado interno y externo. La exportación se realiza en la
época invernal, siendo Estados Unidos el principal país de destino, sin embargo
China se presenta hoy en día como el segundo socio comercial de Chile, ya que su
economía posee perspectivas de crecimiento cada vez más altas, representando
para los productores chilenos grandes posibilidades, lo que debe ser asumido como
un gran desafío, esto significa no sólo llegar con una fruta de calidad al país de
destino, sino también conservar su condición durante el proceso de
comercialización, y satisfacer de este modo las expectativas del mercado.
Para definir la calidad de los limones, los países que importan y exportan cítricos
han dictado normas oficiales con fines comerciales. Éstas se basan en las
características externas del fruto, tales como color, tamaño, ausencia de defectos en
la cáscara, relación sólidos solubles/acidez y contenido de jugo mínimo (MAGDAHL,
1998)
Uno de los principales problemas que se presenta es el desorden fisiológico peteca,
la que se desarrolla principalmente en la época invernal, dañando la apariencia del
fruto, lo que provoca una disminución de su valor comercial y un rechazo a nivel de
exportación (RAZETO, 2000).

2
La peteca corresponde a una alteración que se manifiesta principalmente en
limones, en forma de manchas circulares y deprimidas, ubicadas dispersamente
sobre la superficie del fruto. El albedo (parte blanca esponjosa) bajo cada mancha
se deseca pero la pulpa no se ve dañada, afectando solamente la presentación
(LATORRE, 1992; KLOTZ, 1973; KHALIDY, JAMALI y BOLKAN, 1969).
El problema se produce normalmente en el huerto, pero se manifiesta
mayoritariamente tiempo después que la fruta ha sido cosechada (RAZETO, 2000).
La peteca afecta normalmente al limón que se cosecha en invierno y no al de
verano. La causa exacta de este desorden no se conoce; aparentemente está
asociada a condiciones de baja temperatura y tal vez elevada humedad del aire
durante el periodo de maduración (RAZETO, 1987).
Gómez (1984), observó que la mayor incidencia de peteca se presentó en frutos
cosechados al principio de la temporada. Además, señala que el desorden se
presentó con mayor frecuencia en los árboles ubicados en sectores sombríos. De
igual modo, observó que generalmente los frutos ubicados en la periferia y
preferentemente aquellos ubicados en la parte sur del árbol, se afectan más. Esto
puede ser causado porque permanecen expuestos por un mayor tiempo a una
menor cantidad de radiación, temperatura más baja y mayor humedad relativa que
el resto de la fruta.
Hipótesis:
De acuerdo con los antecedentes señalados se puede establecer que, las
condiciones ambientales de temperatura y humedad relativa, así como también la
posición de la fruta en el árbol y la fecha de cosecha, inciden en el desarrollo del
desorden fisiológico peteca.

3
● Objetivo general:
Evaluar el efecto de las condiciones ambientales, temperatura (ºC) y humedad
relativa (%), sobre el desarrollo de peteca en limonero cv. Eureka, relacionándolas
con la posición del fruto en la copa del árbol, así como también la fecha de cosecha
en que se recolecten dichos frutos.
● Objetivos específicos:
- Evaluar el efecto de la fecha de cosecha de limones cv. Eureka, sobre el desarrollo
de peteca, en poscosecha.
- Evaluar la incidencia de la temperatura y humedad relativa, durante el periodo
invernal, sobre la aparición de peteca.
- Evaluar el efecto de la ubicación del fruto en el árbol, proveniente de distintas
posiciones de la copa, sobre los parámetros de calidad y el desarrollo de Peteca en
limones cv. Eureka, antes y después de un almacenaje refrigerado.

4
2. REVISION BIBLIGRAFICA
2.1. Características del fruto:
El fruto del limonero corresponde a una baya llamada hesperidio, originado a partir
del desarrollo y crecimiento del ovario (SCHNEIDER, 1968). El pericarpio (cáscara),
esta compuesta por el flavedo o epicarpio (parte coloreada) y albedo o mesocarpo,
que corresponde a la parte blanca esponjosa. El pericarpio envuelve al endocarpo
que es la pulpa, comprendida por ocho o más segmentos. Estos segmentos son los
que contienen a las vesículas de jugo, desarrollando células altamente vacuoladas
(GIL, 2000).
2.2. Antecedentes de la especie y variedad:
La variedad Eureka es originaria de California, es una de las variedades más
plantada los últimos años en Chile. Es productiva y de precoz entrada en
producción. Se caracteriza fundamentalmente, porque se puede cosechar fruto
durante todo el año, siendo su principal producción durante el periodo invernal
(70%), y el resto se cosecha durante verano-otoño (ORTÚZAR et al., 2002).
El árbol es de tamaño medio, vigoroso, con hábito de crecimiento abierto
(ORTÚZAR et al., 2002). Con tendencia a fructificar en los extremos de las ramas,
es sensible al frío (AGUSTÍ, 2000).
Los frutos son de buen tamaño, forma elíptica y presentan un mamelón apical poco
acentuado, presenta buen contenido de jugo, con elevada acidez y escaso número
de semillas (ORTÚZAR et al., 2002).

5
2.3. Antecedentes sobre peteca:
La peteca corresponde a una alteración que se presenta en limones, como manchas
circulares y deprimidas, ubicadas en la superficie del fruto. Estas lesiones se
localizan en el albedo, pero no llegan a dañar la pulpa. Células ubicadas bajo la
mancha se desecan y contraen, pudiendo observarse manchas de color blanco o
ligeramente incoloro (LATORRE, 1992; RAZETO, 1987; KHALIDY, JAMILI and
BOLKAN, 1969).
KLOTZ (1973), señala que la peteca es una depresión de la superficie de la
cáscara, que ocurre después que el fruto se ha cosechado.
GÓMEZ (1984), concluye que este desorden se desarrolla con mayor frecuencia en
sectores con una alta fertilidad natural y que además se les adiciona fertilización
nitrogenada. Otro factor de huerto que incide en su desarrollo, es el sombreamiento
ocurrido durante gran parte del día, sumado a condiciones de alta humedad (como
resultado del roció e inviernos lluviosos). BONELLI (1998), señala que a medida que
la cosecha se realiza distanciada de una lluvia, el desarrollo de peteca es menor, lo
que esta directamente relacionado a elevada humedad relativa y bajas
temperaturas.
Su presencia también se ha relacionado a bajas temperaturas, alta humedad
atmosférica, condiciones de alta humedad y rocío (SCHULTZE, 2000; RAZETO,
1987; GÓMEZ, 1984; KLOTZ, 1978).
SCHULTZE (2000) y PASTEN (2004) observaron que el mayor número de frutos
dañados, se cuantificaron a fines de junio, disminuyendo o siendo casi nulos a fines
de agosto, esto directamente relacionado a las condiciones atmosféricas.
La posición del fruto en la copa del árbol, es otro factor influyente en el desarrollo de
peteca, observándose un mayor desarrollo en la fruta ubicada en el sector sur de la

6
copa del árbol. Cabe señalar que frutos ubicados en la parte exterior del árbol,
desarrollan más peteca, que aquellos ubicados al interior de la copa (SCHULTZ,
2000; WILD 1991).
El estado de latencia del árbol es otro factor que estaría influyendo en la
manifestación del desorden. Ya que en el mes de junio los limoneros experimentan
una fuerte desfoliación, pudiendo aumentan su sensibilidad frente a este desorden,
el que se expresará principalmente cuando la fruta sea cosechada (RAZETO, 2000).
BLOKHINA (2003), indica que bajo condiciones de estrés, las plantas generan
reactivos al oxigeno, tales como, peróxido de hidrógeno, anión súper oxido y radical
hidroxilo, las cuales aumentarían la capacidad de lignificación, y reacción con
proteínas y lípidos de membrana, alterando su estructura y función.
LATORRE (1992) determinó que una de las medidas que favorece la disminución de
la incidencia de peteca en la fruta que será exportada, es postergar el embalaje por
dos o cuatro días, de este modo los frutos que revelen dicho daño, podrán ser
descartados antes de ser embalados. Otra medida es la práctica realizada en
limones de invierno, denominada curado de la fruta, esto es mantener la fruta
durante un par de días a temperatura ambiente, para deshidratar la epidermis,
descartando de este modo fruta que manifieste los síntomas (RAZETO, 2000).
Temperaturas de almacenaje de 13ºC pueden evitar el desarrollo de desordenes
fisiológicos como: peteca, membranosis, pitting y oleoselosis (ARTES et al., 1993).
La peteca surge por un desbalance del equilibrio de calcio (LATORRE, 1992). Se ha
determinado que suelos delgados, con déficit de humedad durante los meses
cálidos, influyen en la disponibilidad de calcio para el desarrollo del fruto,
imposibilitando su disponibilidad. Cabe destacar que durante el momento crítico de
desarrollo del fruto, en los tejidos afectados, surge oxalato de calcio, fijando el calcio
que estaba disponible (KHALIDY, JAMILI and BOLKAN, 1969).

7
2.4. Variables determinantes en la calidad de los frutos cítricos:
2.4.1. Temperatura
La temperatura es el factor más importante en el cultivo de los cítricos, influyendo en
el desarrollo vegetativo, floración, cuajado, desarrollo de los frutos y calidad de
estos. AGUSTÍ (2000) señala que temperaturas de 25 a 30ºC son consideradas
óptimas para la actividad fotosintética, bajas temperaturas y estrés hídrico con
llevan a una detención del desarrollo radicular, restringiendo el desarrollo de
gliberelinas a la copa del árbol. Temperaturas inferiores a 13ºC estimulan el cambio
de color de la fruta, por el contrario temperaturas elevadas producen
reverdecimiento de estas. Días calidos y noches frías inducen la degradación de
clorofila y posterior síntesis de carotenoides. Temperaturas inferiores a 0ºC afectan
el desarrollo del árbol y la calidad de los frutos.
2.4.2. Humedad relativa
Según AGUSTÍ (2000), los cítricos se adaptan a una amplitud de humedad relativa,
siendo posibles cultivarlos en regiones desérticas, subtropicales y tropicales. La
humedad relativa es uno de los factores determinantes en la calidad de los frutos.
Lluvias durante el periodo otoñal en climas templados mejoran el tamaño final de la
fruta y el contenido de jugo. El crecimiento y desarrollo del fruto coinciden con
estaciones secas y calurosas. Un déficit durante este periodo puede afectar el
tamaño final del fruto en forma irreparable.
Fruta cosechada bajo condiciones de elevada humedad ambiental afectan la calidad
de la fruta, favoreciendo la aparición de desordenes fisiológicos, tales como
oleocelosis, provocado por ruptura de las glándulas de aceites presentes en el
flavedo (MAZZUZ, 1996).

8
2.5. Calcio:
El calcio es un elemento esencial en el desarrollo y crecimiento de las plantas.
Desempeña una función estructural al conceder estabilidad a los tejidos vegetales
de paredes celulares (CRISTOFFANINI, 1998; DERMARTY et al., 1984). Actúa
como puente intermolecular, uniendo moléculas de pectinas con polisacáridos y
proteínas, formando estructuras complejas de tipo pecticocelulósico, otorgando
rigidez a los tejidos vegetales (CASERO, 1995). Es por esto, que bajos niveles de
calcio en esa zona reducen su resistencia, provocando la desorganización de las
paredes celulares y el colapso de los tejidos afectados (DUARTE y GUARDIOLA,
1995).
2.6. Oxalato de calcio:
El oxalato de calcio es un producto del ión calcio obtenido del suelo y del ión oxalato
formado metabólicamente. En plantas, la presencia de pequeñas cantidades de
oxalato de calcio en células y paredes celulares es casi normal, no afectando su
desarrollo y función (ILARSLAN et al., 1997).
Los cristales de oxalato de calcio son compuestos elaborados por la planta en forma
natural para neutralizar elementos dañinos para ella, como lo es el ácido oxálico. La
formación de estos cristales se realizan en base al calcio, siendo la manera más
eficiente de realizarlos, ya que este es un proceso físico que no requiere gasto de
energía. Esto se asemeja a lo sucedido en bitter pit en manzanas, donde
disminuciones en concentraciones de calcio en paredes celulares y en membranas
provocan desorganización y ruptura celular (CARVALHAO, 1997; BANGERTH,
1974).
El desarrollo de cristales de oxalato de calcio al interior de las vacuolas, es una
característica propia en la mayoría de las células formadoras de cristales, aun

9
cuando las características de estructuras celulares varían entre células según la
morfología del cristal (HORNER y WARNER, 1995; FRANCESCHI y HORNER,
1980).
ARNOTT (1965), señala que los cristales presentan diversas formas, denominadas,
rafidios, drusas, prisma y gránulos. Los que pueden encontrarse como
monohidratado o díhidratado (formas de hidratación), las cuales pueden ser
identificadas mediante el uso de difracción de rayos X, espectroscopia, microscópica
de contraste y microscopia polarizada (FREY-WYSSLING, 1981). El monohibrido es
más estable y se encuentra comúnmente en plantas.
Se ha observado que la presencia de oxalato de calcio, oxalatos e ión calcio,
estarían involucrados en el desorden fisiológico peteca (FRANCESCHI y LOEWUS,
1995; FRANCESCHI y HORNER, 1980).
2.7. Peróxido de hidrógeno:
Diversos autores señalan que el peróxido de hidrógeno en plantas, puede ser
producido por oxalato oxidasa (XU et al., 2003).
Existe una cierta interacción entre los peróxidos de hidrógeno intracelulares y el
calcio del citosol en respuesta al estrés abiótico y biótico. Estudios indican que un
aumento en el calcio citosólico promueve la generación de peróxidos de hidrógeno
(YANG y POOVAIAH, 2002).
Experimentos han demostrado, que el peroxido de hidrógeno es una señal difusible
en la inducción de genes de defensa de las plantas, tales como la glutatión-S-
transferasa y la glutatión peroxidasa (LEVINE et al., 1994).

10
2.8. Glutatión peroxidasa:
La glutatión peroxidasa es una de las enzimas que participan en las
transformaciones de especies reactivas del oxígeno, catalizando la reducción del
peróxido o lipoperóxido, para lo cual utiliza como agente reductor al glutatión
reducido. Esta enzima desempeña un importante papel en la defensa antioxidante
por su localización en todos los órganos y tejidos, como parte del sistema
antioxidante del glutatión (CISNEROS et al., 1997; GACHOTE y BENVENISTE,
1995).
Estrés oxidativo puede generar radicales libres, lo que gatilla la síntesis de glutatión
peroxidasa (HOLLAND et al., 1994).
2.9. Polifenol oxidasa:
La polifenol oxidasa es una proteína que contiene cobre como grupo prostético y a
diferencia de la mayoría de las enzimas, puede catalizar dos tipos de reacciones
diferentes. Estas reacciones incluyen la hidroxilación de monofenoles para producir
o-difenoles y la remoción de hidrógenos de los o-difenoles pare producir quinonas
(GUADARRAMA y RIVAS, 1995).
La PPO es una oxireductasa de gran importancia en vegetales, debido a su
participación en la síntesis de lignina, oxidación del ácido indol acético (AIA) y en
aspectos fitopatológicos relacionado con resistencia a enfermedades
(GUADARRAMA y RIVAS, 1995). Está presente en todas las plantas pero se
encuentra particularmente en altas concentraciones en champiñones, tubérculos de
papa, duraznos, manzana, cambur, aguacate, hojas de té, café y hojas de tabaco.
Es de gran importancia en la determinación de los atributos de calidad de frutas y
hortalizas ya que a través de sus reacciones puede producir cambios de color, sabor

11
y valor nutritivo en productos vegetales frescos, enlatados y congelados (MATHEW
y PARPIA, 1971 citado por GUADARRAMA y RIVAS, 1995).
Frutos estresados mecánicamente manifiestan degeneraciones visibles en el
mesocarpo y endocarpo durante el almacenaje, produciendo un incremento de la
actividad de las enzimas poligalacturonasa y polifenoloxidasa, así como también,
cambios en el metabolismo y la expresión genética (MARTÍNEZ-ROMERO et al.,
1999).
2.10. Peroxidasas totales:
Diversos autores señalan que existe una producción de genes de defensa en fluidos
intercelulares, entre los que destacan las peroxidasas (BERNARDINI, PENEL and
GREPPIN, 1986).
La actividad de las peroxidasas se manifiesta con la aparición de un color parduzco,
al ser puestas en contacto con el producto guayacol y peróxido de hidrógeno
(CHEFTEL y CHEFTAL, 1992).
Las peroxidasas totales actúan en los entrecruzamientos oxidativo, tienen como
función dirigir la formación de lignina, están involucradas en la construcción de la
pared celular y en el control de su plasticidad. También participan en las reacciones
de entrecruzamiento que ocurren en la pared celular, cuando se producen
infecciones de patógenos (MOERSCHBACHER, 1992). Para estas reacciones
utiliza peróxido de hidrógeno como electrón aceptor (BARCELÓ et al., 1998).
Las enzimas peroxidasas (POXs) están ampliamente distribuidas en animales,
plantas y microorganismos. Éstas presentan múltiples formas isoenzimáticas que
difieren tanto en la secuencia de sus aminoácidos como en sus propiedades
químicas. Los resultados de numerosos estudios realizados con diferentes especies,
determinaron que las POXs están involucradas en procesos fisiológicos relevantes

12
de las plantas superiores tales como lignificación, catabolismo de auxinas,
resistencia a patógenos, así como también en diversos mecanismos de respuesta
que éstas presentan a diferentes situaciones de estrés (TARRAGÓ et al., 2003)
3. MATERIALES Y MÉTODOS
3.1. Descripción del ensayo:
El ensayo se realizó en un huerto de limones cv. Eureka, perteneciente a la
Pontificia Universidad Católica de Valparaíso, en El Fundo La Palma, sector La
Palma, Provincia de Quillota, V Región (32º 50` latitud sur, 71º 13` longitud oeste),
Chile. Este huerto fue elegido por tener un historial de incidencia de peteca.
Los árboles fueron plantados en 1999, a una distancia de 6x3 m (555 árboles/ha),
injertados sobre Citrus macrophylla.
El ensayo se realizó durante el periodo comprendido entre los meses de abril y
agosto de 2004. Para ello durante el mes de abril se seleccionaron tres árboles, en
base a su homogeneidad, en cuanto a desarrollo, vigor, carga frutal y estado
sanitario. En cada árbol se instalaron cinco sensores de temperatura y humedad
relativa (marca HOBO y Dickson, modelo TP120,) los cuales se ubicaron en el
interior y periferia de la copa del árbol, quedando de esta forma dividido en: sur-
superior, sur-inferior, norte-superior, norte-inferior e interior del árbol.
Los sensores fueron programados para registrar temperatura (ºC) y humedad
relativa (%) cada una hora, la que fue descargada semanalmente. Cabe señalar que
estos fueron protegidos con macetas plásticas, para evitar daño provocado por
lluvia, llovizna o rocío, con la consiguiente pérdida de información y daño de los
mismos.

13
Se cosecharon un total de 160 limones (16 frutos/tratamiento) con índice de color
amarillo y calibre homogéneo (115-160 gramos) el día 23 de julio y 6 de agosto,
ambas dos días después de una lluvia de 15 y 25 milímetros (sumatoria de tres días
seguidos de lluvia) respectivamente.
La fruta después de cosechada, se trasladó al Laboratorio de Poscosecha de la
Facultad de Agronomía de la Pontificia Universidad Católica de Valparaíso, donde
fue sometida a curado durante dos días y posteriormente almacenada en cámara
frigorífica a 3º C con 90 % de humedad relativa (H.R.) durante 50 días.
3.2. Variables evaluadas:
3.2.1. Presencia y ausencia de peteca
La presencia y/o ausencia de peteca, se evaluó en forma visual utilizando 10 frutos
por tratamiento, cuantificando los limones con peteca, lo cual se realizo cada 10
días a partir del día 0 hasta el día 50 de almacenaje refrigerado, determinando el
número de frutos afectados por tratamiento, los frutos evaluados fueron devueltos a
la cámara. Se consideraron dos niveles, fruto sin peteca (0), y fruto con peteca (1).
Los valores se expresaron en porcentaje de frutos que manifestaron daño.
Cabe señalar que la evaluación de peteca subepidermal se realizó a la salida de
cámara (50 días), ya que se requirió de la destrucción de los frutos.
3.2.2. Parámetros de calidad
La cuantificación de los parámetros de calidad: acidez titulable, deshidratación,
grosor de cáscara, porcentaje de jugo, pH, y sólidos solubles, se realizó para la
primera y segunda cosecha, a los 0 y 50 días de almacenaje refrigerado, utilizando
tres frutos/tratamiento.

14
3.2.2.1. Acidez titulable
La acidez del jugo de los limones se determinó mediante la titulación de 2 ml de jugo
con NaOH 0,5 N.
La acidez se expresó en gramos de ácido cítrico por 100 ml de jugo de limón
(KIMBALL, D. 1991), según la siguiente fórmula:
Acidez = Gasto NaOH (ml) * Normalidad NaOH * 6,41
Volumen Muestra (ml)
3.2.2.2. Deshidratación
Su determinación se realizó por diferencia de peso. Se identificaron los limones
mediante una marca y se midió el peso de cada limón antes de almacenarlo en
cámara y a los 50 días de almacenaje. Se expresó en porcentaje.
3.2.2.3. Grosor de cáscara
Para determinar el grosor de cáscara se cortaron los frutos por la mitad y se
determinó mediante el uso de un pie de metro digital marca Mitutoyo, en partes
opuestas del fruto, promediando ambos valores. Los valores se expresaron en mm.
3.2.2.4. Porcentaje de jugo
Para determinar el porcentaje de jugo de limón, se pesaron los limones, luego se
exprimieron en su totalidad y se filtró el jugo para excluir la pulpa. Posteriormente se
pesó el jugo extraído y correlaciono con el peso del fruto. Los valores se expresaron
en porcentaje.

15
3.2.2.5. pH
En éste parámetro se utilizó un pHímetro electrónico portátil, marca SCHOTT,
modelo Handilab 1, el cual midió el pH directamente del jugo de limón filtrado.
.
3.2.2.6. Sólidos solubles
Se determinó usando un refractómetro termocompensado, marca ATAGO modelo
ATC-1, calibrado de 0 – 32 ºBrix. La medición se expresó en grados Brix.
3.2.2.7. Cuantificación de calcio en el albedo y flavedo (%)
La determinación del contenido de calcio en el albedo y flavedo, se efectuó en el
Laboratorio de Suelo de la Facultad de Agronomía de la Pontificia Universidad
Católica de Valparaíso, con muestras provenientes de la segunda cosecha (tres
frutos/tratamiento), antes y después del almacenaje refrigerado.
Se utilizó el método de digestión ácida, donde la concentración del elemento se
determinó utilizando espectroscopía de absorción atómica en un equipo GBC 902, a
una longitud de onda de 420 y 422 nm respectivamente (A.O.A.C., 1984).
3.2.3. Variables bioquímicas
La evaluación de las variables bioquímica: peróxido de hidrógeno, Polifenol oxidasa,
peroxidasas totales, glutatión peroxidasa y oxalato de calcio, se realizaron en el
Instituto de Química de la Pontificia Universidad Católica de Valparaíso, para la
primera cosecha, el día 0 y el día 50 de almacenaje refrigerado. Utilizando tres
frutos por tratamiento.

16
3.2.3.1. Actividad Peroxidasas totales
Los reactivos utilizados fueron: ABTS 2*10-3 M (donador de hidrogeno), en tampón
fosfato 0,067 M., peróxido de hidrógeno 10-2 M estandarizado con Kmno4, agua de
alto grado de pureza analítica.
Se agregaron 0,2 ml de ABTS, 0,2 ml de peróxido de hidrógeno y 2 ml de tampón
fosfato pH 6 y finalmente se agregaron 0,2 ml de muestra. En esta mezcla se leyó la
absorbancia a 420 nm desde tiempo cero a seis minutos de reacción (A.O.A.C.,
1984).
U Actividad/ mg proteína total = 32,26 x delta de absorbancia /delta tiempo
mg proteína total
3.2.3.2. Actividad de Polifenol oxidasa
La PPO oxida tirosina a dihidroxifenilalanina, la que a su vez es oxidada a O-
Quinona. Esto va acompañado por un aumento de la absorbancia a 280 nm. El
aumento de la velocidad catalítica, es proporcional a la concentración de enzima y
durante un período lineal de 5 a 10 minutos después de un período inicial no lineal
(A.O.A.C., 1984).
Se utilizaron 0,5M Buffer Fosfato pH 6,5, 0,001M l-Tirosina.
Se ajustó el espectrofotómetro a 280 nm y 25ºC. Se agregaron 1 ml de solución
buffer, 1 ml de tirosina y 0,9 ml de agua de alto grado de pureza. Registrando la
absorbancia y añadieron 0,1 ml de homogenado que contenga la enzima. Se
registró la absorbancia por 10 a 12 minutos.
Unidad de Actividad/mg proteína Total = Delta de Absorbancia *1000
mg proteína total

17
Unidad de actividad: corresponde a un cambio de absorbancia a 280 nm por minuto
a 25 ºC y bajo condiciones específicas (A.O.A.C., 1984).
3.2.3.3. Actividad de Glutatión peroxidasa
La medición se basó en la oxidación de glutatión reducido por la glutatión
peroxidasa acoplada a la desaparición de NADPH inducida por su oxidación por la
glutatión reductasa (BOVERIS, 1997).
Se utilizó Buffer fosfato 50 mM pH 7,0 0,5 mM EDTA, 8 mM, NADPH, 0,1M GSH,
50U/ml Glutatión reductasa, 30 mM t- BOOH (terbutil- hidroperóxido).
Actividad: U GSHPx / mg proteina = OD x Vt x 2 x 1000 x 1
Mg proteína total
Vt: volumen total
Vs: volumen muestra
OD: Delta absorbancia por minuto.
3.2.3.4. Actividad Peróxido de hidrógeno
Se determinó por espectroscopía de fluorescencia utilizando diacetato-2’,7’-
diclorofluoresceina (DCF-DA), para medir el estrés oxidativo originado por peróxido
de hidrógeno en células vivas. Las muestras se lavaron con una solución
amortiguadora (4 ml) y posteriormente se adicionó DCF-D (10 µM) y el antioxidante.
Luego se incubaron durante dos horas a 37 ºC. Sobre este material filtrado se
determinó la emisión de fluorescencia excitando a 503 nm y emitiendo a 529 nm
(RAMARKRISHNAN et al., 1996).

18
3.2.3.5. Contenido de Oxalato de calcio
● Extracción de oxalatos solubles
A los frutos de limón se les sacó el albedo, que corresponde a la parte blanca de la
piel, se molieron en trozos de aproximadamente 0,4 cm2 y se secaron en estufa a 60
ºC hasta eliminar completamente la humedad, lo que fue verificado por la variación
de la masa de cada muestra en función del tiempo hasta valor constante (app. 20
horas). Posteriormente al albedo seco de cada muestra se le adiciono 25 ml de
etanol al 99.8% durante 32 horas para eliminar compuestos solubles orgánicos e
inorgánicos polares que forman interacciones de puente de hidrógeno con las
moléculas de alcohol, principalmente colorantes naturales del limón. Transcurrido
este tiempo se lavaron las muestras con agua desionizada y se mezclaron con 25
ml de una solución acuosa de hipoclorito de sodio (NaClO) al 2,5% durante 16 horas
a fin de eliminar compuestos residuales solubles en agua, impurezas adsorbidas en
los tejidos de las muestras, y destruir la materia orgánica alrededor de los cristales
de oxalato de calcio que se encuentren presentes en las muestras. Finalmente se
lavaron las muestras, repetidas veces, con agua destilada (cinco porciones de 10
ml) y se secaron a 60ºC hasta completa eliminación de la humedad (app. 20 horas).
● Extracción de oxalatos insolubles
Para obtener el oxalato insoluble se aplicó el procedimiento descrito por CHARLES,
SHAW y KNIGHT (1982), las muestras secas se pulverizaron en un mortero y se
colectó la fracción correspondiente a malla #60 (250 micrones). Posteriormente se
pesaron de 0,3000-0,4500 gramos de albedo (dos por cada muestra) en una
balanza analítica marca Denver Instrument Company, modelo AA-200 (±0,0001 g) y
se adicionó una solución de 6 ml de ácido clorhídrico (HCl) 3N a temperatura
ambiente, manteniéndose el sistema con agitación a 200 rpm durante 60 minutos
(tiempo óptimo determinado por los autores, cuantificando la concentración de
oxalato en el extracto a 1, 5, 10,15, 20, 45 y 60, minutos hasta concentración

19
constante) en un agitador mecánico “orbit shaker, lab-line”, modelo número 3521-
240. Ello permitió asegurar la extracción del 100% de oxalato insoluble presente en
las muestras de albedo, y evitar la conversión de carbohidratos y otros componentes
orgánicos en ácido oxálico, lo cual se produce al utilizar bajas concentraciones de
ácido clorhídrico (0,5-1,5 N) o haciendo el tratamiento con ácido en caliente
(PIOMBO et al., 1996).
Para purificar el extracto obtenido previo a su determinación espectrofotométrica se
aplicó la siguiente metodología:
- Se filtraron los extractos con un filtro miliporo MFS catalogo número A020A013A,
de tamaño de poro de 0,2 µm.
- Se agregó al filtrado 200 µL de reactivo tungstico-fosfórico (ácido tungsténico,
ácido fosfórico, molibdato de sodio, sulfato de litio) para remoción de proteínas, se
dejó reposar una hora y se centrifugó a 2500 rpm por cinco minutos en una
centrífuga Heraeus Christ omnifuge modelo número 97695.
● Determinación de oxalato en solución por espectrofotometría
Este procedimiento comprende la determinación de oxalato por un método cinético
en el cual la especie analizada (oxalato) cataliza la oxidación de Mn(II) en presencia
de periodato, aumentando cuantitativamente la absorbancia de Mn(VII), la que es
registrada mediante un equipo de Espectrofotometría Molecular.
El procedimiento realizado comprendió las siguientes etapas:
- En un matraz aforado de 10 ml se adicionó 1 ml de solución Mn(II) (200 ppm), 0,70
ml de H3PO4 (0,2M), 1,3 ml de acetato de sodio (0,1M), 1 ml de la solución a

20
analizar, se realizó una dilución del extracto proveniente de la hidrólisis ácida de
albedo en 1000 veces. (100µl/10ml→1ml/10ml).
- Se llevó la mezcla reaccionante a un baño termostatizado a 35±1°C durante
10min, luego del cual se adicionó 2ml de solución de periodato de sodio 0,003 M.
- Se mantuvo el matraz con la mezcla reactiva durante 18 min a 35°C, hasta
completar la reacción.
- Finalmente, se adicionó 0,1 g de molibdato de sodio (Na2MoO4*H2O) (apagador de
la reacción) y se completó el volumen de aforo con agua desionizada, midiéndose la
absorbancia de la solución a una longitud de onda de 525 nm con un
espectrofotómetro UV-VISIBLE SHIMADSU modelo 1603 equipado con software y
celdas de vidrio de 1 cm de ancho. Simultáneamente, se registró el espectro de
absorción molecular correspondiente a Mn(VII).
• Reactivos
Todo el material de vidrio utilizado en la determinación fue lavado exhaustivamente
con solución de HNO3 0,2M y dejado durante toda la noche en esta solución, se
enjuagó con agua destilada y posteriormente, con agua desionizada previamente a
su utilización. Todos los reactivos utilizados son de grado analítico (Merck), y para la
preparación de las soluciones sólo se utilizó agua desionizada.
- Patrones de oxalato (664,4ppm): se preparó disolviendo 1,0112 g (±0,001mg) de
oxalato de sodio (Merck) en 1000 ml de agua, desde este se prepararon patrones de
66,44 ppm y 6,644 ppm.
- Mn (II) (1mg/ml) en H2SO4 0,1M: Se preparó a partir de MnSO4*H2O (Merck), se
pesó 0,600 g del reactivo, se adicionó 5,6 ml en ácido sulfúrico concentrado 18 M y
se completó un volumen de 1 litro.

21
- H3PO4 (0,2M) (MercK, 85%, d=1,7g/cm3): se vertió 10ml del ácido concentrado a
un matraz de aforo de 1l y se completo el volumen con agua desionizada.
- KIO4 (0,003M): se utilizan soluciones frescas preparadas desde cristales KIO4
(Merck), manteniendo la solución en la oscuridad. La preparación consistió en tomar
una masa de 0,64g del reactivo, y disolver en 1litro de agua desionizada.
- Acetato de sodio (NaOOCCH3) (0,1M): desde reactivo (Merck), se midió una masa
de 8,2 g y se llevó a un litro de solución.
• Equipamiento.
- Espectrofotómetro Shimadzu UV 1603-PC, equipado con celdas de cuarzo, vidrio
de 1,0cm, provisto de software UV-1603.
- Baño termostatizado y sonicador TRANSSONIC 570/H.
- Termómetro Brannan 0-60°C.
- Cronómetro CONTROL COMPANY.
- Micropipeta HIRSCHMANN 50 a 200µl.
- Balanza analítica DENVER INSTRUMENT COMPANY modelo AA200.
- Balanza granataria CG.
• Preparación de la curva de calibrado.
La curva de calibrado fue preparada utilizando los patrones de oxalato de 66,44 ppm
y 6,644 ppm, desde ellos se prepararon patrones de: 0,0166; 0,024; 0,033; 0,040;
0,050; 0,056 y 0,066 ppm de oxalato. Estos fueron diluidos 10 veces, y medida su
absorbancia según procedimiento a 525 nm (Figura 1).

22
0,001 0,002 0,003 0,004 0,005 0,006 0,0070,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,45
0,50 Absorbancia = 0.06697 + 55.252xppm
Y = A + B * XParametror Valor Error A 0,066966 0,00277 B 55,25211 0,631314 R SD0,999674 0,002728
Abs
orba
ncia
(u.a
)
Oxalato (ppm)
.
Figura 1. Curva calibrado para la determinación de oxalato en solución.

23
• Preparación de las muestras.
De los extractos ácidos de cada muestra pre-tratada se tomó 1 ml y se aforó hasta
un volumen de 10 ml con agua desionizada (factor de dilución 10). De esta muestra
se tomaron 100 uL y se aforaron a 10 ml con agua desionizada (factor de dilución
100). Finalmente de esta solución se tomó 1 ml y se determino
espectrofotométricamente el oxalato en un volumen total de 10 ml (factor de dilución
10).
3.3. Diseño experimental
El ensayo fue dispuesto siguiendo un diseño completamente al azar con arreglo
factorial (5x2x2), es decir 20 tratamientos con tres repeticiones cada uno. La unidad
experimental es la fruta.
Las variables son:
- Posición del sensor en el árbol
P1: sur-superior
P2: sur-inferior
P3: norte-superior
P4: norte- inferior
P5: interior
- Fecha de cosecha
F1: 23 de julio
F2: 6 de agosto
- Almacenaje
A1: 0 días
A2: 50 días

24
CUADRO 1. Tratamientos
Primera cosecha Segunda cosecha
Antes de
almacenaje
Después de
Almacenaje
Antes de
almacenaje
Después de
Almacenaje
Sur-superior P1-F1-A1 P1-F1-A2 P1-F2-A1 P1-F2-A2
Sur-Inferior P2-F1-A1 P2-F1-A2 P2-F2-A1 P2-F2-A2
Norte-Superior P3-F1-A1 P3-F1-A2 P3-F2-A1 P3-F2-A2
Norte- Inferior P4-F1-A1 P4-F1-A2 P4-F2-A1 P4-F2-A2
Interior P5-F1-A1 P5-F1-A2 P5-F2-A1 P5-F2-A2
Donde P: Posición F: Fecha de cosecha A: Tiempo de almacenaje
Para el análisis estadístico de las variables evaluadas, se utilizó un Modelo
Completamente al Azar con arreglo factorial de 5x2x2 de la siguiente forma:
Yjkl = µ+Pj + Ak + Fl + PAjk + PFil + AFkj + APjk + APFjkl + �ijk
Donde
Yjkl : Variables en estudio
µ : Media general
Pj : j-ésima posición
Ak : k-ésimo periodo de almacenaje. Fl : l-ésima fecha de cosecha
PAij : Interacción entre posición y periodo de almacenaje.
PFik : Interacción entre posición y fecha de cosecha. AFki : Interacción entre Periodo de almacenaje y fecha de cosecha
APkj : Interacción entre Periodo de almacenaje y Posición
PAFjkl : Interacción entre posición, periodo de almacenaje y fecha de cosecha
�jkl : Error aleatorio ~ N(0;σ²)

25
En los casos que existió efecto significativo de las interacciones de los factores, se
compararon las medias con el Test de Tukey al 5%.

26
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Presencia de peteca:
De los resultados obtenidos en relación al daño por peteca epidermal (Anexo 1), se
determinó que existe un efecto significativo de la interacción doble entre la fecha de
cosecha (23 de julio y 6 de agosto) y almacenaje refrigerado a 3ºC hasta 50 días
(Cuadro 2). No así de la posición del fruto en la copa del árbol (Cuadro 3).
La peteca se manifiesta en limones que han sido almacenados a 3ºC y 90 % H.R.,
durante un periodo de 50 días. Se observó además que el porcentaje de peteca
disminuye conforme avanza la fecha de cosecha.
CUADRO 2. Efecto de la interacción entre la fecha de cosecha y tiempo de almacenaje, sobre el desarrollo de peteca epidermal en limones cv. Eureka.
*Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Diversos autores señalan que el tiempo de almacenaje para que se produzca daño
por frío varía desde tres a doce semanas, las que están directamente relacionadas
con la época de cosecha (PURVIS , KAMADA and GRIESON, 1979; PURVIS y
GRIERSON, 1982).
Fecha de Cosecha Días de almacenaje Peteca epidermal (%)
0 0 a 23 julio
50 28 b
0 0 a 6 agosto
50 10 a

27
Los incrementos del daño observados al final del período de almacenaje (50 días),
se pueden atribuir a la temperatura a la cual fueron expuestos los limones.
LUCHSINGER (1997), señala que frutos de origen tropical o subtropical, son
susceptibles a los daños por frío al ser almacenados por períodos prolongados a
bajas temperaturas, siendo estas superiores a 0ºC. Se recomiendan temperaturas
de almacenamiento de 14 ºC en limones, de manera de evitar daños por frío
(MARTINEZ-JÁVEGA y CUQUERELLA, 1995).
En cuanto a la fecha de cosecha, el mayor porcentaje de peteca, se manifestó en
frutos provenientes de la primera cosecha (23 julio), lo que se puede atribuir a las
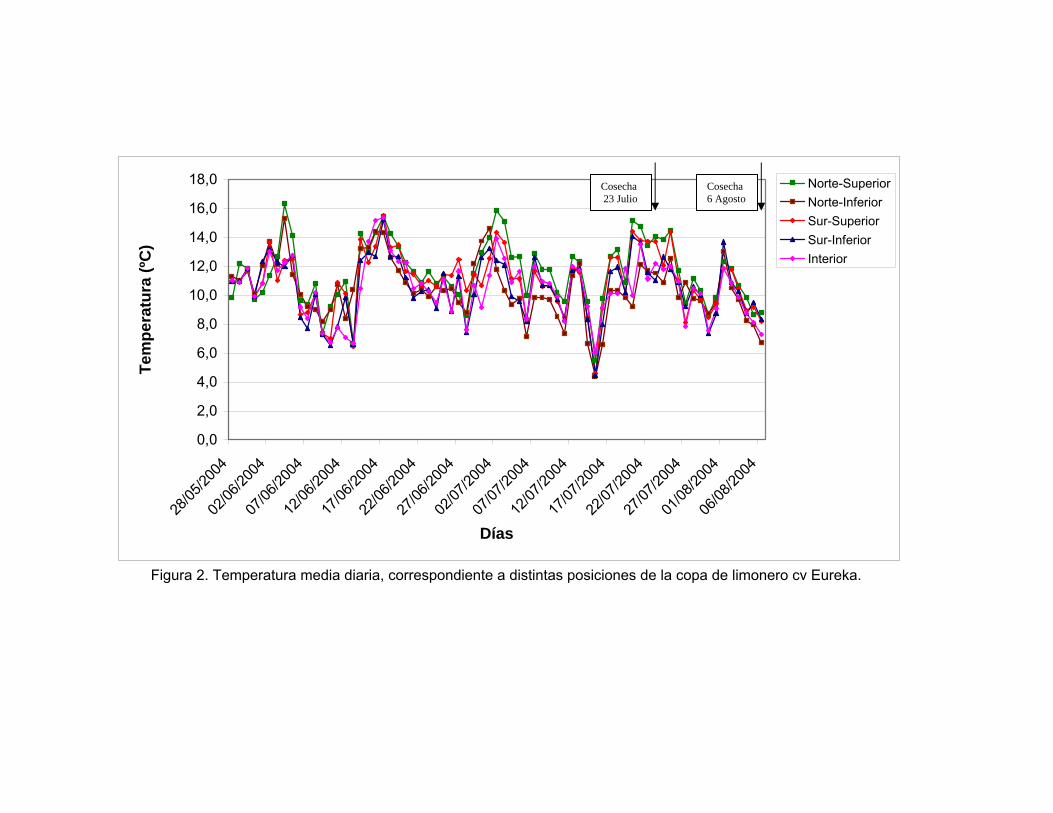
temperaturas y H.R registradas, donde es posible observar (Figura 2), una
tendencia de las temperaturas medias diarias a disminuir durante el periodo de
medición (fines de mayo hasta la primera quincena de julio), luego del cual se
observa un leve aumento de estas. Es importante destacar, la presencia de heladas
en la primera quincena de junio y segunda de julio (Figura 3), lo que incide
directamente en el proceso de desarrollo y maduración del fruto, siendo considerado
un factor de estrés. (AGUSTÍ, 2000). Esto concuerda con lo señalado por
SCHULTZE (2000) y PASTEN (2004) quienes observaron que los % de peteca llega
a un máximo a fines de junio y disminuye en agosto, coincidiendo con el aumento de
temperaturas y disminución de la H.R. Esto se ratifica en el presente trabajo, ya que
el porcentaje de peteca disminuye de 28% a 10%.
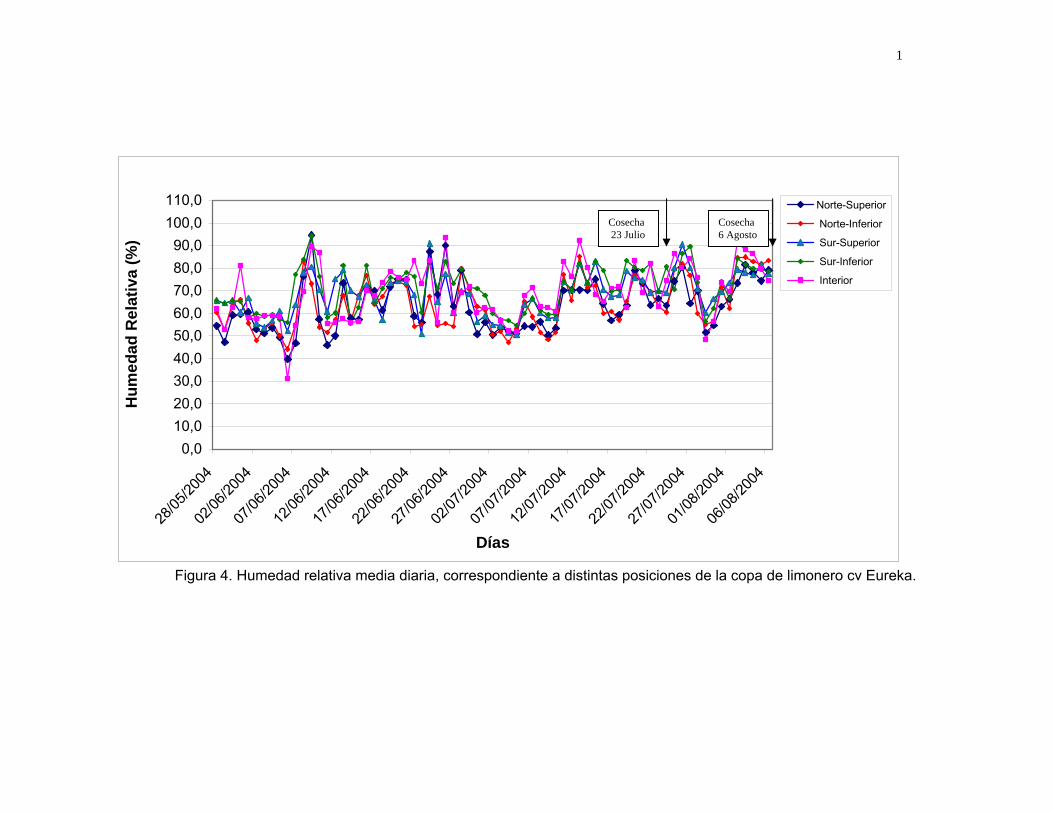
Con respecto a la H.R., se observa un alza luego de las precipitaciones (Figura 4),
lo que sumado al descenso de temperaturas, se ve reflejado en un aumento en el %
de peteca en las siguientes cosechas. Lo anterior concuerda con lo expresado por
SCHULTZE (2000); SNOWDON (1990); RAZETO (1987) y KLOTZ (1973). Quienes
indican que el desarrollo de peteca se ve favorecido por condiciones ambientales de
baja temperatura y elevada H.R. LIZANA y RIVEROS (1983), encontraron a
principios de julio un 13,6 % de daño, lo que disminuyó a 1,2%, a fines de agosto.
También las cosechas realizadas después de una lluvia aumentan la incidencia de
peteca en la fruta (BONELLI, 1998).

28
RAZETO (2000), señala que la peteca se presenta principalmente en limones que
han pasado por condiciones climáticas adversas para la planta, tales como baja
temperatura ambiental, alta H.R. e inicio de las precipitaciones.
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
16,0
18,0
28/05
/2004
02/06
/2004
07/06
/2004
12/06
/2004
17/06
/2004
22/06
/2004
27/06
/2004
02/07
/2004
07/07
/2004
12/07
/2004
17/07
/2004
22/07
/2004
27/07
/2004
01/08
/2004
06/08
/2004
Días
Tem
pera
tura
(ºC
)
Norte-SuperiorNorte-InferiorSur-SuperiorSur-InferiorInterior
Figura 2. Temperatura media diaria, correspondiente a distintas posiciones de la copa de limonero cv Eureka.
Cosecha 6 Agosto
Cosecha 23 Julio

-1,0
1,0
3,0
5,0
7,0
9,0
11,0
13,0
28/05/2004
02/06/2004
07/06/2004
12/06/2004
17/06/2004
22/06/2004
27/06/2004
02/07/2004
07/07/2004
12/07/2004
17/07/2004
22/07/2004
27/07/2004
01/08/2004
06/08/2004
Días
Tem
pera
tura
(ºC
)
Norte-SuperiorNorte-InferiorSur-SuperiorSur-InferiorInterior
Figura 3. Temperatura mínima diaria, correspondiente a distintas posiciones de la copa de limonero cv Eureka.
Cosecha 23 Julio
Cosecha 6 Agosto
1
0,010,020,030,040,050,060,070,080,090,0
100,0110,0
28/05
/2004
02/06
/2004
07/06
/2004
12/06
/2004
17/06
/2004
22/06
/2004
27/06
/2004
02/07
/2004
07/07
/2004
12/07
/2004
17/07
/2004
22/07
/2004
27/07
/2004
01/08
/2004
06/08
/2004
Días
Hum
edad
Rel
ativ
a (%
)
Norte-Superior
Norte-Inferior
Sur-Superior
Sur-Inferior
Interior
Figura 4. Humedad relativa media diaria, correspondiente a distintas posiciones de la copa de limonero cv Eureka.
Cosecha 23 Julio
Cosecha 6 Agosto

32
CUADRO 3. Efecto de la posición del fruto en la copa del árbol sobre el desarrollo de peteca epidermal en limones cv. Eureka.
Posición Peteca epidermal (%)
Sur-Inferior 10 a
Sur-Superior 12.5 a
Norte-Inferior 10 a
Norte-Superior 15 a
Interior 2 a *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
No hay efecto significativo de la posición del fruto en el árbol sobre el desarrollo de
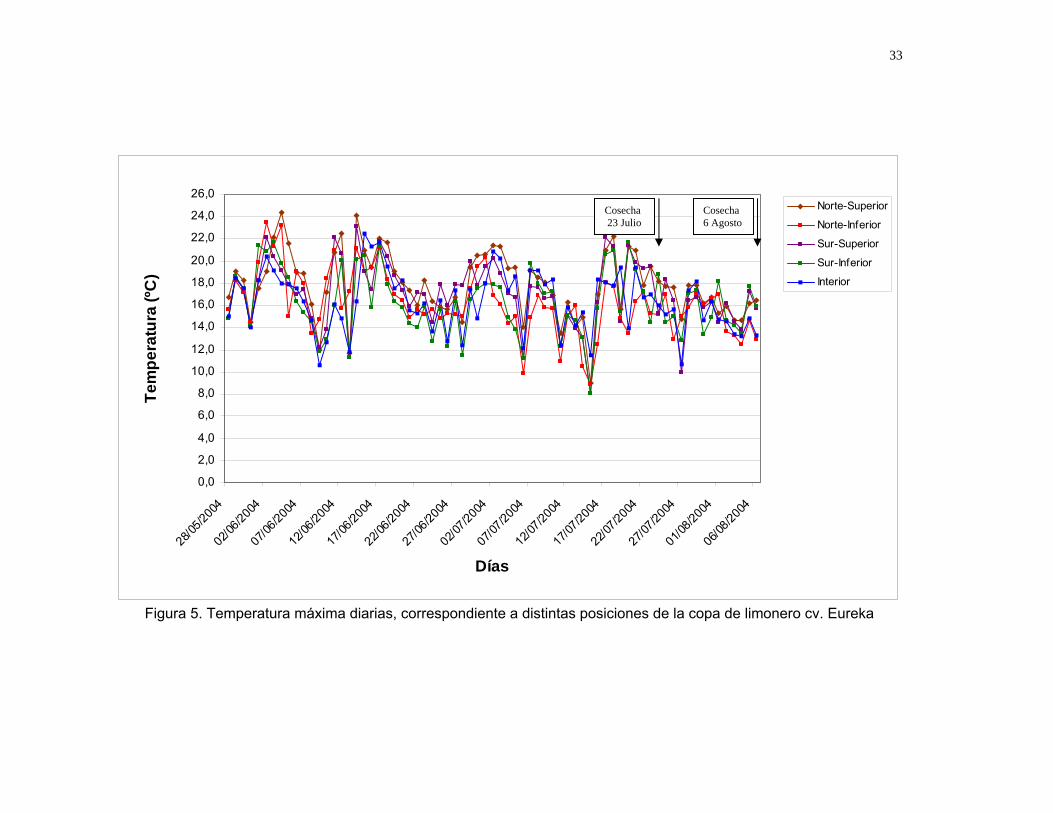
peteca epidermal. Esto se puede atribuir a que las temperaturas registradas no
presentan grandes diferencias en las posiciones (Figura 3 y 5). Los resultados
presentados anteriormente concuerdan con lo observado por SCHULTZE (2000),
quien señala que no existe una clara tendencia del desarrollo de peteca con
respecto a la posición del fruto en el árbol. Sin embargo, los frutos ubicados en el
interior manifestaron un menor daño (2%) con respecto a frutos ubicados en la
posición exterior (10 a 15%). Diversos autores han señalado la existencia de una
mayor influencia de las bajas temperaturas en los frutos externos respecto a los
internos durante su desarrollo (PURVIS, 1980; MCDONALD, McCOLLUM and
NORDBODY, 1993; NORDBY y MCDONALD; 1995; MAZZUZ, 1996). Esto
probablemente debido a que frutos ubicados en el interior se encuentran más
protegidos de las condiciones climáticas adversas (GÓMEZ, 1984; SCHULTZE,
2000).
Del análisis de peteca subepidermal de limones (Anexo 2), se determinó que existe
efecto del tiempo de almacenaje (0 y 50 días), sobre la manifestación del desorden
(Cuadro 4).
33
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
16,0
18,0
20,0
22,0
24,0
26,0
28/05
/2004
02/06
/2004
07/06
/2004
12/06
/2004
17/06
/2004
22/06
/2004
27/06
/2004
02/07
/2004
07/07
/2004
12/07
/2004
17/07
/2004
22/07
/2004
27/07
/2004
01/08
/2004
06/08
/2004
Días
Tem
pera
tura
(ºC
)
Norte-Superior
Norte-Inferior
Sur-Superior
Sur-Inferior
Interior
Figura 5. Temperatura máxima diarias, correspondiente a distintas posiciones de la copa de limonero cv. Eureka
Cosecha 23 Julio
Cosecha 6 Agosto

34
Se observó un claro aumento en el porcentaje de peteca subepidermal pasando de
0 a 19% al término del periodo.
CUADRO 4. Efecto del tiempo de almacenaje a 3º C por 50 días sobre el desarrollo de peteca subepidermal en limones cv. Eureka.
*Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación múltiple de Tuckey (p= 0.05)
El incremento en el desarrollo del desorden, al igual que en el caso de peteca
epidermal pude atribuirse a la temperatura y largo período de almacenaje al que fue
sometida la fruta.
Al analizar la evolución del daño por peteca epidermal y subepidermal (Figura 6 y 7),
se observa en ambos casos que en la posición norte-superior se manifiesta en
mayor proporción el desorden, esto puede deberse a que las lluvias en Chile
ocurren en dirección norte-sur, lo que puede considerarse como un factor de estrés,
debido al efecto del golpe de las gotas de agua sobre el fruto; la lluvia es agua fría,
por lo tanto, la disminución de temperatura provocado después de una lluvia se
refleja con mayor severidad en el sector norte del árbol, principalmente en la parte
superior de este, lo que trae como consecuencia un aumento de la humedad relativa
en dicho sector.
Es importante destacar que el porcentaje de peteca subepidermal observado en
ambas cosecha, en la posición interior de la copa del árbol, presentan mayores
niveles de daño con respecto a la peteca epidermal en dicho sector, esto puede
deberse a que el desorden se produce inicialmente en el albedo del fruto, la que se
manifestaría posteriormente en el flavedo, lo anterior también puede explicarse en
base a las condiciones agroclimáticas registradas durante el periodo, las cuales
Días de almacenaje Peteca subepidermal (%)
0 0 a
50 19 b

35
pueden no haber sido suficientemente adversas para que el desorden se
manifestará en el flavedo.

36
05
1015202530354045
NorteSuperior
NorteInferior
SurSuperior
SurInferior
Interior
Posición
% P
etec
a
Primera cosechaSegunda cosecha
Figuras 6. Presencia de peteca epidermal en limones cv Eureka cosechados en dos
fechas de cosecha (23 de julio y 6 de agosto), medida a los 50 días de almacenaje refrigerado (3ºC).
05
1015202530354045
NorteSuperior
NorteInferior
SurSuperior
SurInferior
Interior
% P
etec
a Su
bepi
derm
al
Primera cosechaSegunda cosecha
Figuras 7. Presencia de peteca subepidermal en limones cv. Eureka cosechados en
dos fechas de cosecha (23 de julio y 6 de agosto), medida a los 50 días de almacenaje refrigerado (3ºC).
37
4.2. Variables Bioquímicas:
4.2.1. Peroxidasas totales
Del análisis de esta variable, se puede observar que existe una interacción entre la
posición de la fruta en la copa del árbol y los días de almacenaje refrigerado (Anexo
3) para la fruta cosechada el 23 de julio (Cuadro 5).
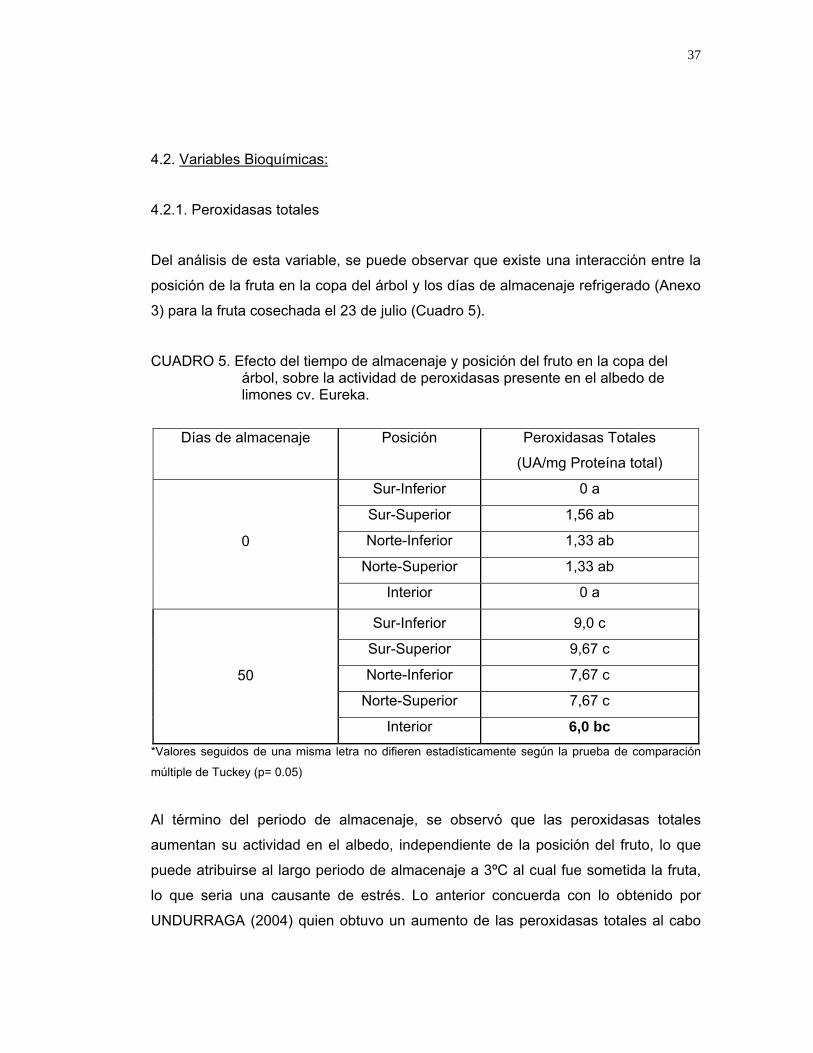
CUADRO 5. Efecto del tiempo de almacenaje y posición del fruto en la copa del árbol, sobre la actividad de peroxidasas presente en el albedo de limones cv. Eureka.
Días de almacenaje
Posición
Peroxidasas Totales
(UA/mg Proteína total)
Sur-Inferior 0 a
Sur-Superior 1,56 ab
Norte-Inferior 1,33 ab
Norte-Superior 1,33 ab
0
Interior 0 a
Sur-Inferior 9,0 c
Sur-Superior 9,67 c
Norte-Inferior 7,67 c
Norte-Superior 7,67 c
50
Interior 6,0 bc *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05)
Al término del periodo de almacenaje, se observó que las peroxidasas totales
aumentan su actividad en el albedo, independiente de la posición del fruto, lo que
puede atribuirse al largo periodo de almacenaje a 3ºC al cual fue sometida la fruta,
lo que seria una causante de estrés. Lo anterior concuerda con lo obtenido por
UNDURRAGA (2004) quien obtuvo un aumento de las peroxidasas totales al cabo

38
de 45 días a 3ºC, ratificando el efecto de estrés de largos periodos de almacenaje a
bajas temperaturas.
Cabe señalar que la menor actividad de las peroxidasas totales medidas a los 50
días de almacenaje en UA/mg proteína total, se obtuvieron en frutos provenientes
del interior del árbol, lo que coincide con los datos entregados anteriormente en
relación al daño por peteca epidermal, ratificando que los frutos ubicados en dicho
sector se encuentran mas protegidos de las condiciones ambientales estresantes.
DESMARCHELIER (1996) señala que las peroxidasas son enzimas que actúan
como defensa antioxidantes de las plantas.
4.2.2. Glutatión peroxidasa
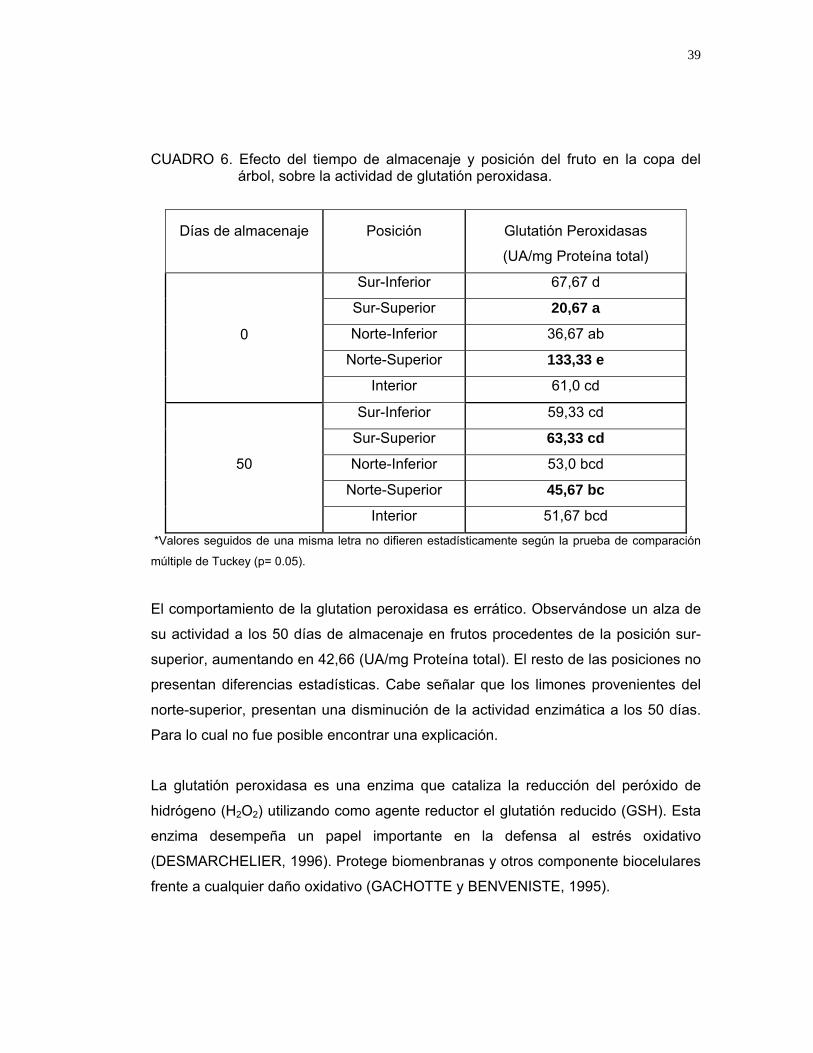
En el Cuadro 6, se puede observar que existe efecto significativo de la interacción
entre días de almacenaje a 3ºC y posición de la fruta en la copa del árbol, sobre la
actividad de la enzima glutatión peroxidasa (Anexo 4), evaluadas en la cosecha
realizada el día 23 de julio en albedo de limones cv. Eureka.
39
CUADRO 6. Efecto del tiempo de almacenaje y posición del fruto en la copa del árbol, sobre la actividad de glutatión peroxidasa.
Días de almacenaje
Posición
Glutatión Peroxidasas
(UA/mg Proteína total)
Sur-Inferior 67,67 d
Sur-Superior 20,67 a
Norte-Inferior 36,67 ab
Norte-Superior 133,33 e
0
Interior 61,0 cd
Sur-Inferior 59,33 cd
Sur-Superior 63,33 cd
Norte-Inferior 53,0 bcd
Norte-Superior 45,67 bc
50
Interior 51,67 bcd *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
El comportamiento de la glutation peroxidasa es errático. Observándose un alza de
su actividad a los 50 días de almacenaje en frutos procedentes de la posición sur-
superior, aumentando en 42,66 (UA/mg Proteína total). El resto de las posiciones no
presentan diferencias estadísticas. Cabe señalar que los limones provenientes del
norte-superior, presentan una disminución de la actividad enzimática a los 50 días.
Para lo cual no fue posible encontrar una explicación.
La glutatión peroxidasa es una enzima que cataliza la reducción del peróxido de
hidrógeno (H2O2) utilizando como agente reductor el glutatión reducido (GSH). Esta
enzima desempeña un papel importante en la defensa al estrés oxidativo
(DESMARCHELIER, 1996). Protege biomenbranas y otros componente biocelulares
frente a cualquier daño oxidativo (GACHOTTE y BENVENISTE, 1995).
40
4.2.3. Polifenol oxidasa
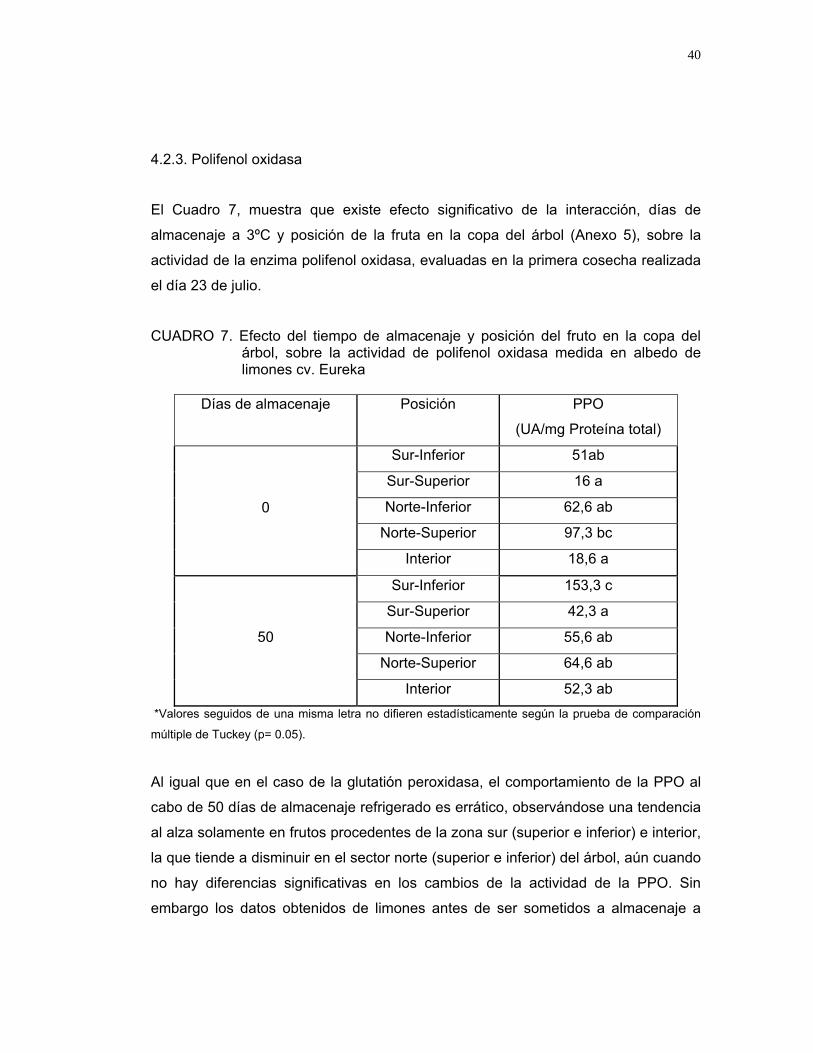
El Cuadro 7, muestra que existe efecto significativo de la interacción, días de
almacenaje a 3ºC y posición de la fruta en la copa del árbol (Anexo 5), sobre la
actividad de la enzima polifenol oxidasa, evaluadas en la primera cosecha realizada
el día 23 de julio.
CUADRO 7. Efecto del tiempo de almacenaje y posición del fruto en la copa del árbol, sobre la actividad de polifenol oxidasa medida en albedo de limones cv. Eureka
Días de almacenaje
Posición
PPO
(UA/mg Proteína total)
Sur-Inferior 51ab
Sur-Superior 16 a
Norte-Inferior 62,6 ab
Norte-Superior 97,3 bc
0
Interior 18,6 a
Sur-Inferior 153,3 c
Sur-Superior 42,3 a
Norte-Inferior 55,6 ab
Norte-Superior 64,6 ab
50
Interior 52,3 ab *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Al igual que en el caso de la glutatión peroxidasa, el comportamiento de la PPO al
cabo de 50 días de almacenaje refrigerado es errático, observándose una tendencia
al alza solamente en frutos procedentes de la zona sur (superior e inferior) e interior,
la que tiende a disminuir en el sector norte (superior e inferior) del árbol, aún cuando
no hay diferencias significativas en los cambios de la actividad de la PPO. Sin
embargo los datos obtenidos de limones antes de ser sometidos a almacenaje a

41
3ºC, muestra que la mayor actividad de la polifenol oxidasa se presentó en limones
pertenecientes al sector norte-superior (Cuadro 7), donde se manifestó el mayor
daño de peteca epidermal (Cuadro 3), quedando de manifiesto que dicho sector se
encontraba bajo condiciones más estresantes que el resto de las posiciones, lo que
llevó a una activación de los mecanismos que generan un aumento de los genes de
defensa que originan moléculas tales como polifenol oxidasa (VAMOS y VIGYAZO,
1981).
GUADARRAMA y RIVAS (1997), señalan que bajas temperaturas de
almacenamiento disminuye la actividad de la polifenol oxidasa, lo que podría
controlar el oscurecimiento enzimático. De lo anterior se extrae, que la disminución
de la actividad enzimática cuantificada para frutos del sector norte (superior e
inferior) a los 50 días, pudo haber sido causa del almacenaje a 3ºC.
MARTÍNEZ-ROMERO et al., (1998) postula que frutos que han sido estresados
mecánicamente, presentan degeneraciones en el mesocarpo y endocarpo durante
el periodo de almacenaje, lo que se suma a un incremento de la actividad de las
enzimas poligalactouronasa y polifenoloxidasa.
4.2.4. Oxalato de calcio
En el Cuadro 8, se puede observar que existe un efecto significativo de la
interacción entre días de almacenaje refrigerado y posición del fruto en la copa del
árbol, sobre la concentración de oxalato de calcio en el albedo de limones cv.
Eureka. evaluados en la primera cosecha (Anexo 6).

42
CUADRO 8. Efecto del tiempo de almacenaje y posición del fruto en la copa del árbol, sobre la concentración de oxalato de calcio.
Días de almacenaje
Posición
Oxalato de Calcio
(mg / 100 gr de tejido)
Sur-Inferior 102,78 de
Sur-Superior 65,76 bcd
Norte-Inferior 80,62 cd
Norte-Superior 138,63 e
0
Interior 65,68 bcd
Sur-Inferior 14,40 a
Sur-Superior 34,62 ab
Norte-Inferior 14,78 a
Norte-Superior 40,71 abc
50
Interior 52,84 abc *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05)
Diversos estudios han demostraron la presencia de formaciones de oxalato de
calcio, en el albedo de limones que presentaban la sintomatología de peteca. Por
otra parte, los cristales de oxalato de calcio son compuestos que la planta elabora
en forma natural para poder neutralizar elementos nocivos para ella como el ácido
oxálico. (CARVALHAO, 1997).
Lo anterior no concuerda con los resultados obtenidos en el presente trabajo, ya que
se observa una disminución de la concentración de oxalato de calcio en el albedo en
frutos que han sido sometidos a almacenaje refrigerado durante 50 días, donde se
manifestó el daño de peteca. (Cuadro 2). ARTES et al. (1993), señalan que aún no
está claro si el desarrollo de la peteca, durante el almacenaje refrigerado, está
directamente relacionado con la formación o la propagación de cristales de oxalato
de calcio.
43
4.3. Variables de calidad:
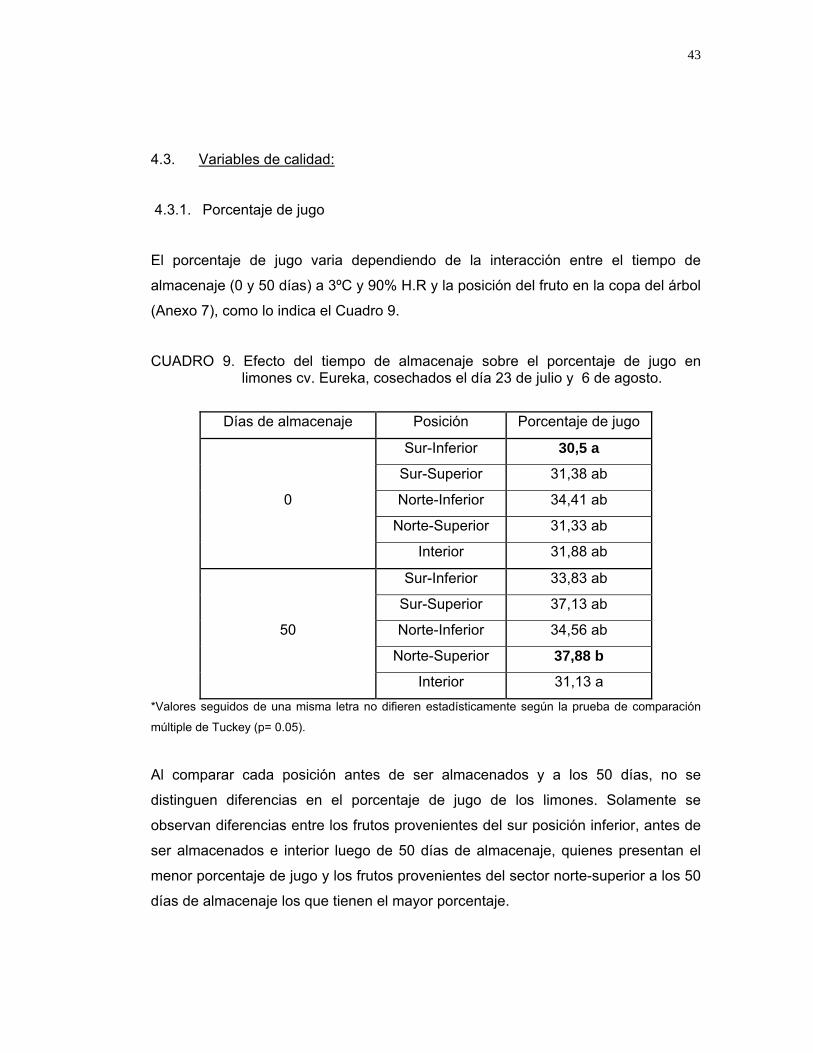
4.3.1. Porcentaje de jugo
El porcentaje de jugo varia dependiendo de la interacción entre el tiempo de
almacenaje (0 y 50 días) a 3ºC y 90% H.R y la posición del fruto en la copa del árbol
(Anexo 7), como lo indica el Cuadro 9.
CUADRO 9. Efecto del tiempo de almacenaje sobre el porcentaje de jugo en limones cv. Eureka, cosechados el día 23 de julio y 6 de agosto.
Días de almacenaje Posición Porcentaje de jugo
Sur-Inferior 30,5 a
Sur-Superior 31,38 ab
0 Norte-Inferior 34,41 ab
Norte-Superior 31,33 ab
Interior 31,88 ab
Sur-Inferior 33,83 ab
Sur-Superior 37,13 ab
50 Norte-Inferior 34,56 ab
Norte-Superior 37,88 b
Interior 31,13 a *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Al comparar cada posición antes de ser almacenados y a los 50 días, no se
distinguen diferencias en el porcentaje de jugo de los limones. Solamente se
observan diferencias entre los frutos provenientes del sur posición inferior, antes de
ser almacenados e interior luego de 50 días de almacenaje, quienes presentan el
menor porcentaje de jugo y los frutos provenientes del sector norte-superior a los 50
días de almacenaje los que tienen el mayor porcentaje.

44
Diversos autores señalan que no existe diferencia entre la fruta cosechada desde
distintos puntos de la copa del árbol sobre el porcentaje de jugo (LEGUIZAMÓN,
1998; SCHULTZE, 2000).
Cabe señalar que existe una tendencia de los limones a aumentar el contenido de
jugo con el almacenaje refrigerado aún cuando estadísticamente esta no sea
significativa, EAKS (1961) señala que gran parte del aumento del contenido de jugo
en la fruta, se debe a cambios dentro del tejido de la pulpa.
4.3.2. Deshidratación
Del análisis de deshidratación de limones cv. Eureka (Anexo 8), se determinó que
existe efecto de dos interacciones dobles, producidas entre: días de almacenaje y
fecha de cosecha y días de almacenaje y posición del fruto en la copa del árbol.
En el Cuadro 10, se observa que limones cosechados el día 6 de agosto, presentan
un mayor porcentaje de deshidratación a la salida de cámara, que los cosechados el
23 de julio.
CUADRO 10. Efecto de la interacción entre los días de almacenaje a 3ºC y 90% de H.R y la fecha de cosecha, sobre la deshidratación de limones cv. Eureka.
*Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación múltiple de Tuckey (p= 0.05). Lo anterior puede deberse a las condiciones ambientales y regimenes hídricos
imperantes antes de la cosecha., o perdida de agua después de ser cosechados.
Días de almacenaje Fecha de Cosecha Deshidratación (%)
23 julio 0 a 0
6 agosto 0 a
23 julio 4,58 b 50
6 agosto 5,31 c

45
COHEN et al., (1994) determinaron que las perdidas de agua del fruto es un
indicador del daño por frío en limones y pomelos, las que están asociadas a
microheridas producidas en la cutícula de frutos visiblemente sanos. Almacenajes a
temperaturas inferiores a la crítica, producen habitualmente perdidas del agua del
fruto (WILLS, 1989).
En el Cuadro 11, se observa que la deshidratación se manifiesta en limones cv.
Eureka almacenados durante 50 días.
CUADRO 11. Efecto del tiempo de almacenaje a 3º C por 50 días y posición del fruto en el árbol, sobre la deshidratación en limones cv Eureka
Días de almacenaje Posición Deshidratación (%)
Sur-Inferior 0 a
Sur-Superior 0 a
0 Norte-Inferior 0 a
Norte-Superior 0 a
Interior 0 a
Sur-Inferior 5,04 b
Sur-Superior 6,12 b
50 Norte-Inferior 6,21 b
Norte-Superior 4,13 b
Interior 4,67 b *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Del análisis estadístico, se extrae que existe efecto de la interacción entre días de
almacenaje y posición del fruto en la copa del árbol, sin embargo es posible señalar
que el efecto de la interacción esta influenciada por el tiempo de almacenaje, ya que
la deshidratación se manifiesta a los 50 y no hay diferencias entre las diferentes
posiciones, mientras que al día cero no hay deshidratación.

46
4.3.3. Grosor de cáscara
En el Cuadro 12, se observa que existe efecto significativo de la interacción triple
producida por las variables: días de almacenaje, época de cosecha y posición del
fruto en la copa del árbol sobre el grosor de cáscara en limones cv. Eureka (Anexo
9).
CUADRO 12. Efecto del tiempo de almacenaje a 3º C por 50 días, fecha de cosecha y posición del fruto en la copa del árbol, sobre el grosor de cáscara.
Días de Fecha almacenaje de cosecha
Posición Grosor de cáscara (mm)
Sur-Inferior 0,59 a Sur-Superior 0,77 ab 23 julio Norte-Inferior 0,76 ab Norte-Superior 0,84 b 0 Interior 0,73 ab Sur-Inferior 0,73 ab Sur-Superior 0,63 ab 6 agosto Norte-Inferior 0,66 ab Norte-Superior 0,74 ab Interior 0,79 ab Sur-Inferior 0,67 ab Sur-Superior 0,62 ab 23 julio Norte-Inferior 0,60 ab
50 Norte-Superior 0,72 ab Interior 0,73 ab Sur-Inferior 0,66 ab Sur-Superior 0,70 ab 6 agosto Norte-Inferior 0,72 ab
Norte-Superior 0,70 ab Interior 0,65 ab
*Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05)
Al evaluar el grosor de cáscara se determinó que no existe una tendencia clara de
este parámetro. Solamente es posible observar que los frutos provenientes de la

47
posición sur-inferior presentan el menor grosor de cáscara (mm), lo contrario de lo
que sucede en frutos provenientes de la posición norte-superior cosechados el día
23 de julio y antes de entrar a cámara, quienes presentan los mayores valores. Esto
probablemente se deba a la ubicación de la fruta, ya que en la zona norte del árbol,
principalmente parte superior de este, está expuesta por mayor tiempo a la luz solar,
lo que produce una mayor actividad metabólica en dicho sector, mayor
transpiración, favoreciendo así la capacidad de sink de los limones por los
fotoasimilados y elementos minerales, obteniendo frutos de mayor tamaño
(AGUSTI, 2000), presentando un mayor desarrollo de la cáscara con respecto a los
limones ubicados en la zona sur. BALDWIN (1993) reportó que el grosor de cáscara
está asociado a condiciones climáticas.
BERGER (1994), señala que limones de tamaño grande, se asocian con mayor
grosor de cáscara.
4.3.4. pH
Al evaluar el pH, se determinó que existe efecto de dos interacciones dobles,
producidas entre la fecha de cosecha (23 julio y 6 agosto) y posición del fruto en la
copa del árbol, y entre almacenaje refrigerado y posición del fruto en la copa del
árbol, sobre el pH de limones cv. Eureka (Anexo 10).
El Cuadro 13 indica que existe un efecto significativo de la interacción, entre la
época de cosecha y posición del fruto en la copa del árbol. Sobre el parámetro de
calidad pH.

48
CUADRO 13. Efecto de la fecha de cosecha y posición del fruto en el árbol, sobre el pH del fruto.
Fecha Cosecha Posición pH
Sur-Inferior 2,39 ab
Sur-Superior 2,41 ab
Norte-Inferior 2,39 ab
Norte-Superior 2,46 a
23 julio
Interior 2,38 ab
Sur-Inferior 2,34 ab
Sur-Superior 2,34 ab
Norte-Inferior 2,34 ab
Norte-Superior 2,30 b
6 agosto
Interior 2,40 ab *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Solo la posición norte-superior presenta diferencias significativas entre la primera y
segunda cosecha, el resto de los tratamientos no presentan diferencias estadísticas
entre ellos. Es importante señalar que el mayor valor se presenta en la primera
cosecha norte-superior, en donde se obtuvo la mayor manifestación de daño por
peteca.
GUADARRAMA y RIVAS (1995) indicaron que al desplazarse el pH hacia el lado
alcalino o hacia el lado ácido, se produce una disminución de la actividad
enzimática, probablemente debido a modificaciones en el centro activo al alterarse
los aminoácidos catalíticos que conforman el mismo. Lo que estaría relacionado con
la modificación de los parámetros bioquímicas evaluados anteriormente y la
manifestación del desorden.

49
Del Cuadro 14, se extrae que existe un efecto significativo entre las variables días
de almacenaje refrigerado y posición del fruto en la copa del árbol sobre el pH del
fruto.
Se puede determinar que los valores mas bajos se obtuvieron al igual que en el
caso anterior de frutos cosechados de la posición norte-superior antes de ser
almacenados. El resto de los tratamientos no presentaron diferencias significativas
entre ellos, de acuerdo a la separación de medias.
CUADRO 14. Efecto del tiempo de almacenaje a 3º C por 50 días y posición del fruto en el árbol, sobre el pH de limones cv. Eureka.
Días de almacenaje Posición pH
Sur-Inferior 2,37 ab
Sur-Superior 2,38 ab
Norte-Inferior 2,37 ab
Norte-Superior 2,45 b
0
Interior 2,38 ab
Sur-Inferior 2,35 ab
Sur-Superior 2,38 ab
Norte-Inferior 2,36 ab
Norte-Superior 2,31 a
50
Interior 2,39 ab *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Los resultados concuerdan con lo establecido por COOK (1983), LUTTGES (2000) y
UNDURRAGA (2004), quienes observaron una tendencia del pH a disminuir
después de permanecer en almacenaje refrigerado, debido probablemente a una
descomposición de los ácidos débiles, lo que habría incrementado el nivel de H+,

50
generando la disminución del pH. Este cambia poco o nada en almacenaje
refrigerado (CORFO-ENAFRI, 1970).
El Cuadro 13 y 14 muestran solo una diferencia estadística para los frutos obtenidos
de la zona norte del árbol parte superior. Cabe recordar lo señalado anteriormente
donde la mayor manifestación de peteca epidermal y cambios a nivel bioquímico, se
producen en dicho lugar, lo que puede atribuirse a que cambios en el pH del fruto
puede ser un indicador de la desorganización celular como consecuencia de un
estrés, ya que estos cambios afectan las funciones de las proteínas reguladoras de
procesos metabólicos, asociadas a las membranas (KURKDJIAN y GUERN, 1989).
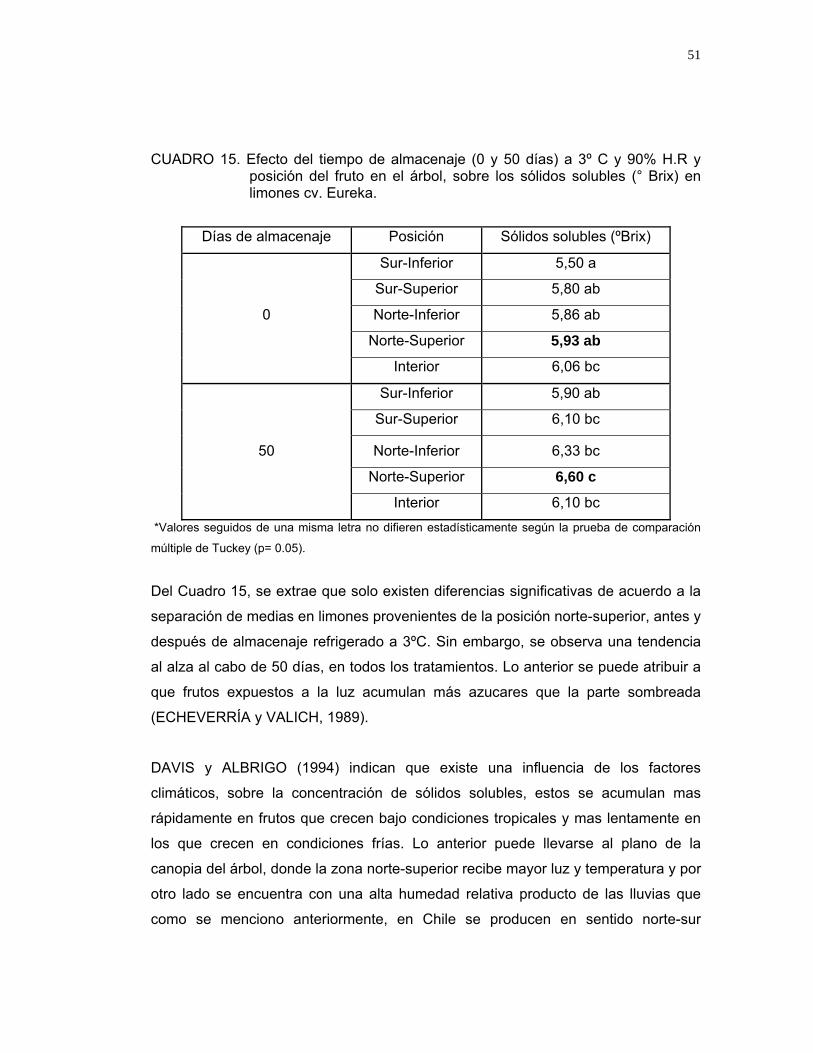
4.3.5. Sólidos solubles
Del análisis estadístico de sólidos solubles realizado en limones cv. Eureka (Anexo
11), se determinó que existe efecto de la variable época de cosecha y de la
interacción entre el tiempo de almacenaje y la posición del fruto en la copa del árbol,
sobre el contenido de sólidos solubles (Cuadro 15).
51
CUADRO 15. Efecto del tiempo de almacenaje (0 y 50 días) a 3º C y 90% H.R y posición del fruto en el árbol, sobre los sólidos solubles (° Brix) en limones cv. Eureka.
Días de almacenaje Posición Sólidos solubles (ºBrix)
Sur-Inferior 5,50 a
Sur-Superior 5,80 ab
Norte-Inferior 5,86 ab
Norte-Superior 5,93 ab
0
Interior 6,06 bc
Sur-Inferior 5,90 ab
Sur-Superior 6,10 bc
Norte-Inferior 6,33 bc
Norte-Superior 6,60 c
50
Interior 6,10 bc *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Del Cuadro 15, se extrae que solo existen diferencias significativas de acuerdo a la
separación de medias en limones provenientes de la posición norte-superior, antes y
después de almacenaje refrigerado a 3ºC. Sin embargo, se observa una tendencia
al alza al cabo de 50 días, en todos los tratamientos. Lo anterior se puede atribuir a
que frutos expuestos a la luz acumulan más azucares que la parte sombreada
(ECHEVERRÍA y VALICH, 1989).
DAVIS y ALBRIGO (1994) indican que existe una influencia de los factores
climáticos, sobre la concentración de sólidos solubles, estos se acumulan mas
rápidamente en frutos que crecen bajo condiciones tropicales y mas lentamente en
los que crecen en condiciones frías. Lo anterior puede llevarse al plano de la
canopia del árbol, donde la zona norte-superior recibe mayor luz y temperatura y por
otro lado se encuentra con una alta humedad relativa producto de las lluvias que
como se menciono anteriormente, en Chile se producen en sentido norte-sur

52
aumentando la humedad relativa en dicha sector inmediatamente después de una
lluvia. Es importante recordar que fue esta posición la que manifestó mayor daño de
peteca.
Los sólidos solubles tienden a aumentar en los frutos almacenados durante 50 días.
Lo que concuerda con los resultados obtenidos por LUTTGES (2000) y
UNDURRAGA (2004). El aumento de los azucares se produce debido a la
degradación de los constituyentes de las paredes celulares, hemicelulosa y
pectinas, BARTHOLOMEW y SINCLAIR (1951) citado por LUTTGES (2000), otro
factor importante de destacar, es el efecto de la deshidratación sobre la
concentración de sólidos solubles, ya que frutos luego de un período de almacenaje,
aumentan los ºBrix debido probablemente a la perdida de humedad de los tejidos, lo
que produciría un aumento en la concentración de sólidos solubles de la fruta.
(LUTTGES, 2000).
En el Cuadro 16, se puede apreciar que existe efecto de la época de cosecha sobre
los sólidos solubles (ºBrix).
CUADRO 16. Efecto de la época de cosecha, sobre sólidos solubles (ºBrix) en limones cv Eureka.
Época Cosecha Sólidos solubles (ºBrix)
23 julio 6,11 a
6 agosto 5,90 b *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05)
Limones provenientes de la cosecha realizada el 23 de julio, estuvieron expuestos a
condiciones climáticas más adversas en comparación con los frutos cosechados el 6
de agosto. Condiciones de estrés unida a la transpiración producida por medio de
los estomas, producen una deshidratación de las células, lo que conlleva a la
acumulación de azúcares (YAKUSHIJI, 1996).

53
4.3.6. Acidez titulable
Según los resultados obtenidos en el análisis estadístico (Anexo 12), se puede
concluir que existe un efecto significativo de la posición del fruto en el árbol y de la
interacción entre días de almacenaje refrigerado y fecha de cosecha (23 julio y 6
agosto), sobre la acidez titulable.
En el Cuadro 17, se observa que el menor nivel de acidez titulable se cuantifico en
limones procedentes de la cosecha realizada el día 23 de julio almacenados hasta
50 días. El resto de los limones no presentan diferencias estadísticas de acuerdo a
la separación de medias.
CUADRO 17. Efecto del tiempo de almacenaje a 3º C por 50 días y época de cosecha, sobre la acidez en limones cv. Eureka.
*Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Limones cosechados el día 23 de julio y almacenados durante un periodo de 50 días
a 3ºC y 90% de H.R presenta la menor acidez titulable, esto se puede relacionar con
el Cuadro 2, donde se observa que la mayor manifestación de daño por peteca
ocurre en dichos frutos. Lo anterior ratifica los resultados obtenidos por GÓMEZ
(1984), quien observó una menor acidez en frutos con peteca al compararlos con
frutos sanos.
Fecha de Cosecha Días de almacenaje Acidez titulable
0 8,37 a 23 julio
50 6,82 b
0 7,93 a 6 agosto
50 8,11 a

54
En el Cuadro 18 se observa el efecto de la posición del fruto en el árbol sobre la
acidez titulable.
CUADRO 18. Efecto de la posición del fruto en la copa del árbol, sobre la acidez en limones cv. Eureka.
Posición Acidez titulable
Sur-Inferior 7,86 ab
Sur-Superior 7,74 ab
Norte-Inferior 7,71 ab
Norte-Superior 7,44 a
Interior 8,26 b *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Existe diferencias estadísticas en los limones provenientes de la posición norte-
superior e interior, lo que se relaciona a con la presencia de peteca, ya que limones
ubicados en el interior del árbol presentaron menor manifestación del daño en
comparación con frutos pertenecientes a la zona norte parte superior, estos últimos
alcanzaron la menor acidez.
La respiración en cítricos es generalmente baja, esta es la responsable que
compuestos azucarados se reduzcan dando paso a un mayor contenido de acidez
(UNDURRAGA, 1998).
4.3.7. Cuantificación de calcio (%) en el albedo del fruto.
En el Cuadro 19, se observa que existe un efecto significativo de la interacción de
los factores, días de almacenaje refrigerado y posición del fruto en la copa del árbol,
sobre el porcentaje de calcio presente en el albedo de limones cv. Eureka (Anexo
13), evaluados en la cosecha realizada el día 6 de agosto.

55
CUADRO 19. Efecto del tiempo de almacenaje a 3º C hasta 50 días y posición del fruto en la copa del árbol, sobre el porcentaje de calcio en el albedo del fruto.
Días de almacenaje Posición % calcio albedo
Sur-Inferior 0,38 b
Sur-Superior 0,36 ab
Norte-Inferior 0,45 d
Norte-Superior 0,40c
0
Interior 0,35 a
Sur-Inferior 0,38 b
Sur-Superior 0,43 c
Norte-Inferior 0,42 c
Norte-Superior 0,41 c
50
Interior 0,43 c *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
Existe una tendencia al alza en el porcentaje de calcio evaluado a los 50 días de
almacenaje a 3ºC, principalmente en limones proveniente de la posición sur-
superior, norte-inferior e interior. Frutos procedentes de la zona norte del árbol antes
de ser sometidos a almacenaje refrigerado, presentan las mayores concentraciones
de calcio (0,45 y 0,40%). Esto se puede atribuir a que limones ubicados en dicha
zona, principalmente sector superior de la copa del árbol reciben durante mayor
parte del día la influencia de los rayos solares en comparación al resto de los frutos
ubicados en la zona sur, presentando de esta forma una mayor actividad
metabólica, mayor respiración y transpiración. HANSON (1995) señala que aquellos
órganos que presentan una alta tasa de transpiración, tienden a acumular mayores
concentraciones de calcio en desmedro de otros que transpiran menos.
El calcio se mueve dentro de la planta vía xilema, siendo la humedad uno de los
factores que afectan la transpiración. Frutos que crecen con una baja humedad

56
durante el día evidencian menor contenido de calcio al compararlo con frutos que
crecen bajo condiciones de elevada humedad (SILVA, H y RODRIGUEZ, J. 1995).
Clima, riego, poda, fertilización, madurez de cosecha, tamaño del fruto y tipo de
fruta, además de su posición en el árbol, influyen sobre los niveles de calcio en la
fruta (KUPFERMAN, 1988 citado por UNDURRAGA, 2004).
Del presente análisis se obtiene una relación entre el % de calcio en el albedo y la
manifestación del daño por peteca, lo cual nos permite tener una idea de los niveles
de calcio registrados en frutos al cabo de 50 días de almacenaje, donde fue
precisamente cuando se manifestó el daño, si bien la posición del fruto en el árbol
no tuvo en efecto significativo estadísticamente, si fue posible observar la
manifestación del desorden a los 50 días de almacenaje. De lo anterior se extrae
que el desarrollo de peteca coincide con un aumento en el contenido de calcio en el
albedo, luego de un largo período de almacenaje refrigerado.
4.3.8. Cuantificación de calcio (%) en el flavedo del fruto.
Según los resultados obtenidos (Anexo 14) se puede concluir que existe un efecto
significativo de la interacción entre días de almacenaje y posición del fruto en la
copa del árbol sobre el contenido de calcio en el flavedo de limones cv. Eureka,
evaluado en la cosecha realizada el día 6 de agosto (Cuadro 20).

57
CUADRO 20. Efecto del tiempo de almacenaje a 3º C por 50 días y posición del fruto en la canopia del árbol, sobre el porcentaje de calcio.
Días de almacenaje Posición % calcio flavedo
Sur-Inferior 0,76 cd
Sur-Superior 0,81 d
Norte-Inferior 0,99 e
Norte-Superior 0,98e
0
Interior 0,69 b
Sur-Inferior 0,55 a
Sur-Superior 0,76 cd
Norte-Inferior 0,74 bc
Norte-Superior 0,77 cd
50
Interior 0,76 cd *Valores seguidos de una misma letra no difieren estadísticamente según la prueba de comparación
múltiple de Tuckey (p= 0.05).
El % de calcio en el flavedo disminuye en frutos almacenados durante 50 días a
3ºC. Lo anterior puede ser efecto de las lluvias originadas días antes de la cosecha,
lo que causaría una dilución del contenido de calcio en dicho lugar.

58
5. CONCLUSIONES
Limones cv. Eureka cosechados en pleno invierno (julio), presentan mayor
porcentaje de peteca epidermal y subepidermal, que limones cosechados a la salida
de este (agosto).
La posición del fruto en la copa del árbol no tiene un efecto significativo sobre el
desarrollo de peteca epidermal y subepidermal, en limones cv. Eureka.
El almacenaje refrigerado influye en el desarrollo de peteca epidermal y
subepidermal, manifestándose el desorden a partir de los 30 días a 3ºC y 90 % de
H.R. aumentando la severidad del daño a los 50 días.
Existe efecto de la posición de la fruta en la copa del limonero en conjunto con
almacenaje refrigerado sobre los parámetros de calidad: porcentaje de jugo,
deshidratación, pH y sólidos solubles. Limones evaluados al término del almacenaje
y ubicados en la zona norte posición superior, presentan un mayor contenido de
jugo, menor pH y mayor concentración de sólidos solubles, en comparación con el
resto de las posiciones evaluadas en un mismo periodo.
Limones cv. Eureka provenientes de la posición norte-superior cosechados pleno
invierno, antes de ser almacenados, presentan un mayor grosor de cáscara que
limones cosechados de la posición sur-inferior a las mismas condiciones.
Limones cv. Eureka cosechados a la salida de invierno, evaluados al término de un
almacenaje a 3ºC por 50 días, presentan un mayor porcentaje de deshidratación
que aquellos cosechados en pleno invierno a igual almacenaje.
Existe efecto de la posición de la fruta en la copa del limonero en conjunto con el
almacenaje a 3ºC y 90% H.R por 50 días sobre los parámetros bioquímicos:

59
glutatión peroxidasa, peroxidasas totales y oxalato de calcio, cuantificándose una
menor actividad al término del almacenaje, en los frutos provenientes del interior de
la copa del árbol. La concentración de oxalato de calcio disminuye independiente de
la posición de la fruta.

60
6. RESUMEN
El siguiente ensayo se realizó con el objetivo de determinar el efecto de la fecha de cosecha y ubicación del fruto en la copa del árbol sobre la incidencia de peteca en limonero cv. Eureka, evaluados a los 0 y 50 días de almacenaje refrigerado a 3ºC y 90% de humedad relativa. Se utilizaron limones de calibre homogéneo (95), cosechados de distintos puntos de la copa del árbol: sur-superior, sur-inferior, norte-superior, norte-inferior e interior. Ubicando en cada una de estos sectores sensores de temperatura y humedad relativas, relacionándolas con sus respectivas posiciones y con el desarrollo de peteca en dichos sectores. Se evaluó la presencia y ausencia de peteca, parámetros de calidad como grosor de cáscara, sólidos solubles, pH, acidez titulable, porcentaje de jugo y deshidratación y variables bioquímicas como peróxido de hidrógeno, peroxidasas totales, oxalato de calcio, glutatión peroxidasa, polifenol oxidasas y porcentaje de calcio. Las mediciones se realizaron al día 0 y 50 de almacenaje refrigerado. Al analizar la intensidad del daño causado por peteca, se observó que los limones provenientes de la primera cosecha (23 de julio), desarrollaron un mayor porcentaje de peteca en comparición con la fruta cosechada el día 6 de agosto, lo que estaría relacionado a condiciones de menor temperatura y mayor H.R. El porcentaje de peteca epidermal y subepidermal, aumenta cuando los frutos son almacenados durante 50 días a 3ºC y 90% H.R. La variable posición del fruto en la copa del árbol no tuvo efecto sobre la manifestación del desorden. En cuanto a la evolución de los parámetros de calidad de la fruta, se observaron cambios significativos a los 50 días de almacenaje refrigerado, provocando una disminución del grosor de cáscara, aumento del contenido de jugo (%), acidez, sólidos solubles (ºBrix). El pH se mantuvo casi inalterable. Del ensayo se desprende que el almacenaje refrigerado a 3ºC durante 50 días provoca un incremento de la actividad enzimática, siendo ésta mayor en el lado sur del árbol, para el caso de glutatión peroxidasa, polifenol oxidasa y peroxidasas totales.

61
7. SUMMARY The following test was made with the objective to determine the effect of the date of harvest and location of the fruit in the crown of the tree on the incidence of peteca in lemon tree cv. Eureka, evaluated to the 0 and 50 days of storage cooled to 3ºC and 90% of relative humidity. Lemons of homogenous caliber (95) were used, harvested of different points from the crown of the tree: south-superior, south-inferior, north-superior, north-inferior and inner. Locating in each one of these sensorial sectors of temperature and relative humidity, relating them to its respective positions and the development of peteca in these sectors. One evaluated the presence and absence of peteca, quality parameters like rind thickness, solubles solids, pH titleable, percentage of juice and dehydration and biochemical variables like hydrogen peroxide, total peroxidasas, oxalato of calcium, glutatión peroxidasa, polifenol oxidasas and percentage of calcium. The measurements were made to day 0 and 50 of cooled storage. When analyzing the intensity of the damage caused by peteca, was observed that the originating lemons of the first harvest (23 of July), developed to a greater percentage of peteca in comparisson with the harvested fruit day 6 of August, which would be related to conditions of smaller temperature and greater H.R. The percentage of epidermal and subepidermal peteca, increases when the fruits are stored during 50 days 3ºC and 90% H.R. The position variable of the fruit in the crown of the tree did not have effect on the manifestation of the disorder. As far as the evolution of the parameters of quality of the fruit, significant changes were observed the 50 days of cooled storage, causing a diminution of the rind thickness, increase of the juice content (%),acidity, soluble solids (ºBrix). pH stayed almost unalterable. The test it is come off that the storage cooled to 3ºC during 50 days causes an increase of the enzymatic activity, being the this one greater one in the South side of the tree, for the case of glutathione peroxidase, polyphenol oxidase and total peroxidases.

62
8. LITERATURA CITADA AGUSTÍ, M. 2000. Citricultura. Madrid, MundiPrensa. 416 p. ARNOTT, H; PAUTARD, F and STEINFINK, H. 1965. Structure of calcium oxalate
monohydrate. Nature 208(1):197-1198. ARTES, F. ESCRICHE, A. and MARÍN, J. 1993. Treating Primifori lemons incold
storage with intermittent warming and carbon dioxide. HortScience 28 (8): 819-821.
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS (A.O.A.C.). 1984. Official
methods of analysis. 15th ed. Washintong, D.C. Assn. Offic. Anal. Chemists. 1018 p.
BALDWIN, E. 1993. Citrus Fruit, In: Symour, G; Taylor, J. and Tucker, G. eds.
Biotchemistry of Fruit Ripening, London, Chapman & Hall. pp 107-150. BANGERTH, F. 1974. The funtion of calcium inthe cell and in the subcellular units
of apple fruit. Acta Hortocultirae 45: 43-52. BARCELO, R. MORALES, M. and PEDRENO, M. 1998. Specific
compartmentalization of peroxidase isoenzymes in relation to lignin biosynthesis in the plant cell. In: Lewis, N. and Sarkanen, S. eds. Lignin and Lignan Biosynthesis. Washington, DC: American Chemical Society. pp. 84–95.
BERGER, H. 1994. Indice de madurez de cosecha en cítricos. Publ. Tec. Soc. Agr.
Chile. 3 : 113-114. BERNARDINI, N. PENEL, C. and GREPPIN, H. 1986. Preliminar immunological
studies on horseradish and spinach peroxidase isoenzymes. In: Greppin, H. Penel, C. and Gaspar. Eds. Th. Molecular and physiological aspects of plant peroxidases. Geneva. University of Geneva. pp. 97 – 103.

63
BONELLI, C. 1998. Efecto de aplicaciones de calcio en poscosecha de limones cv. Génova, cosechados en invierno con dos estados de madurez, sobre la aparición de peteca. Taller de licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 73 p.
BOVERIS, A. 1997. Biochemistry of free radicals: from electrons to tissues.
Medicine 58: 350-356. BLOKHINA, O.; VIROLAINEN, E.; FAGERSTEDT, K. 2003. Antioxidant, Oxidative
Damage and Oxigen Deprivation Stress. Annals of Botany 91: 179-194. CARVALAHO, F. 1997. El bitter pit de las manzanas, desarrollo y control.
Fruticultura Profesional N° 86: 12-20. CASERO, T. 1995. La nutrición cálcica en frutales. Fruticultura Profesional 71: 45-
55. CHARLES, W. SHAW, P. and KNIGHT, R. 1982. Analysis of Oxalic Acid in
Carambola (Averrhoa carambola L.) and Spinach by High-Performance Liquid Chromatography. Journal of Agricult. Food Chemistry 30 (6): 1106-1108.
CHEFTEL, C. y CHEFTAL., H. 1992. Introducción a la Bioquímica y Tecnología
de los Alimentos. Madrid, Acribia. 310 p. (Vol 1). CISNEROS, P.; PUPO, J.; CÉSPEDES, E. 1997. Enzimas que participan como
barreras fisiológicas para eliminar los radicales libres: III. Glutatión peroxidasa. Rev Cubana Invest Biomed 1997; 16(1):10-15
COHEN, E. SHAPIRO, B. SHALOM, and KLEIN, J.1994. Water Loss: A
Nondestructive Indicator of Enhanced (Cell Membrane permeability of Chilling injured Citrus Fruit). HortScience 119 (5): 983-986.
COOK, R. 1983. Quality of citrus juices as related to composition and processing
practices. Journal of Food Science and Technology. 37(6): 68-71 p.

64
CORPORACIÓN DE FOMENTO A LA PRODUCCIÓN – EMPRESA NACIONAL FRIGORÍFICA. 1970. Grado de madurez, tamaño y aditivos de almacenaje de limones. Santiago, Corfonafri. 106p.
CRISTOFFANINI, L. 1998. Calcio en frutales persistentes. Empresa y Avance
Agrícola. 57: 14-145 DAVIS, F and ALBRIGO, L. 1994. Citrus. Wallingford. CAB Internacional. 254 p. DERMARTY, M; MORVAN, C. and THELLIER, M. 1984. Calcium and the cell wall
Plant Cell Environ. 7: 441-448. DESMARCHELIER, C; CICCI, G. 1996. Antioxidantes de origen vegetal. Ciencia
Hoy. 34: 32-43 p. DUARTE, A. Y GUARDIOLA, J. 1995. Aplicaciones del calcio en citricultura.
Levante Agrícola 34 (331): 11-14. EAKS, I. L. 1961. Effect of temperature and holding periods on physical and
chemical characteristics of lemon fruit. Journal Food Science. 26: 253-599.
ECHEVERRIA, E and VALICH, J. 1989. Enzymes of sugar and acid metabolism in
stored Valencia oranges. J. Amer. Soc. Hort. Sci. 114: 445-496. FRANCESCHI, V. and LOEWUS, F. 1995. Oxalate Biosynthesis and function in
plants and fungi. In: Khan, S.R. ed. Calcium oxalate in biological systems. Florida. Ed. Boca Raton. CRC Press. pp. 113-130
____________. and HORNER, H. T. 1980. Calcium oxalate crystals in plants. Bot.
Rev. 46:361-427 FREY-WYSSLING, A. 1981. Crystallography of the two hydrates of crystalline
calcium oxalate in plants. Am. J. Bot. 68:130-141.

65
GACHOTTE, D. and BENVENISTE, P. 1995. Cloning and sequesncing of a glutathione peroxidase homologue from Arabidpsis thaliana (Accession No. X89866). Plant Physiol 110:713
GIL, G. 2000. La Producción de Fruta. Santiago, Ediciones Universidad Católica
de Chile. 583 p. GOMEZ, P. 1984. Características de los limones y factores de huerto que incidirían
en el desarrollo de peteca. Tesis Ing. Agr. Santiago, Universidad de Chile. Escuela de Agronomía. 76 p.
GUADARRAMA, A. y RIVAS, N. 1995. Purificación y caracterización cinética de la
enzima polifenol oxidasa del ocumo (Xanthosoma sagittifolium). Rev Fac. Agron. (Maracay) 16:65-86.
HANSON, E. 1995. Translocación distribución del calcio en la plantas. Universidad
de Talca. Simposio Internacional Calcio en fruticultura. Talca, 17-18 octubre 1995. pp. 37-45.
HOLLAND, D. FALTIN, Z. PERL, A. BEN-HAYYIM, G. and ESHDAT, Y. 1994. A
novel plnt glutathione peroxidase-like protein provides tolerance to oxygen radicals generated by paraquat in E. Coli. FEBS Letters. 337: 52-55.
HORNER, H. and WAGNER, B. 1995. The association of druse crystals with the
developing stomium of Capsicum annuum (Solanaceae) anthers. Am. J. Bot. 67:1347-1360.
ILARSLAN, H.; PALMER, G.; IMSADE, J. and HORNER, T. 1997. Quantitative
determination ofcalcium oxalate in developing seeds of soya. American Journal of Botany. 84:1024-1046.
KHALIDY, R. JAMALI, A. and BOLKAN, H. 1969. Causes of the peteca disease of
lemons as occurring in Lebanon. Proceeding of the First International Citrus Symposium, 2: 1253-1261.
KIMBAL, D. 1991. Citrus Processing: Quality Control and Tecnology. New York,
CHAPMAN and HALL. 473 p.

66
KLOTZ, L. 1973. Color handbook of citrus diseases. Berkeley, ed. California University, Division of agricultural science. pp. 108-119
KLOTZ, L. 1978. Color Handbook of Citrus Diseases. Riverside, University of
California. 55- 57. KURKDJIAN, A and GUERN, J. 1989. Intracelullar Ph: Measurement and
Importance In Cell Activity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40: 271-303.
LATORRE, B. 1992. Enfermedades de las plantas cultivadas. Santiago,
Universidad Católica de Chile. 626 p. LEVINE, A. TENHAKEN, R. DIXON, R. and LAMB, C. 1994. H2O2 from the
oxidative Bursa orchestrates the plant hypersensitive disease resistance response. Cell 79: 583-593.
LIZANA, A., y RIVEROS, J. 1983. Evaluación de la calidad y naturaleza del
desecho de limones para exportación. Simiente, Vol. 53: 3-4. LUCHSINGER, L. and Walsh, C. 1997. Problemática de la exportación de
duraznos, nectarines y ciruelas. Aconex 55: 5-10. LUTTGES, C. 2000. Efecto del calibre y estado de madurez de limones cv. Fino
49, sobre el desarrollo de peteca en almacenaje refrigerado. Taller de licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 53 p.
MAGDAHL, C. 1998. Situación actual de la citricultura chilena. Sociedad
Gardiazabal y Magdahl Ltda. Seminario Internacional de Cítricos. Viña del Mar, 13-15 mayo 1998. pp. 1-16.
MARTÍNEZ-JAVEGA, J. y CUQUERELLA, J 1995. Alteraciones fisiológicas en la
pos-recolección de frutos cítricos. Fruticultura Profesional Nº 69: 57-67. (2ª parte).

67
MARTINEZ-ROMERO, D. VALERO, D. SERRANO, M. MARTINEZ-SANCHEZ, F. and RIQUELME, F. 1999. Effects of postharvest putrescine and calcium treatments on reducing mechanical damage and poluamines and abscisic acid levels during lemon storage. J. Sci. Food Agric. 79: 1589-1585.
MAZZUZ, C. 1996. Calidad de frutos cítricos. Manual para su gestión desde la
recolección hasta la expedición. España. Reus, Ed. Tecnidex. Ediciones de Horticultura. 317 p.
McDONAL, R. McCOLLUM, T. and NORDBODY, H. 1993. Temperature
conditioning and surface treatments of grapefruit affect expression of chilling injury and gas difussion. J. Amer. Soc. Hort. Sci. 118(4): 490-496.
MOERSCHBACHER, B. 1989. Lignin biosynthesis in stem rust infected wheat.
ACS Symposium Series – American Chemical Society. Nº 399: 370-382 p. NORDBY, H and McDONALD, R. 1995. Variations in chilling injury and
epicuticular wax composition of white grapefruit with Canopo position and fruti development during the season. Journal of Agricultural and Foodd Chemistry 43: 1128-1133.
OFICINA DE ESTUDIOS y POLITICAS AGRARIA. 2003. Superficies de huertos
frutales, (on line). http://www.cffa.org ORTUZAR, J.; RAFA, V.; MARTIZ, J.; QUINTEROS, J.; ARENAS, M. 2002.
Descripción de variedades de cítricos. Ficha técnica Nº4. Limonero Eureka. Aconex. 77: 31-32.
PASTEN, M. 2004. Efecto de la fecha de cosecha, estado de madurez y ubicación
del fruto en la copa del árbol sobre la incidencia de peteca en limoneros cv. Eureka. Taller de licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 80p.
PIOMBO, G. BABRE, D. FALLAVIER, P. CAZEVIEILLE, P. ARVIEU, J. and
CALLOT, G. 1996. Oxalate extraction and determination by ionic chromatography in calcareous soils and in mycorrhized roots environment. Commun. Soil Sci. and Plant Anal. 27(5/8): 1663-1677

68
PURVIS, A.; KAWANDA, K. and GRIESON, W. 1979. Relationship between midseason resistance to chilling injury and reducing sugar level in grapefruit peel. HortScience 14(3):227-229.
PURVIS, A. 1980. Influence of canopy depth on susceptibility of Marshgrapefruit to
chilling injury. HortScience 15(6):731-733 PURVIS, A. and GRIESON, W. 1982. Acumulation of reducing sugar and
resistance of grapefruit peel to chilling injury as relatted to winter temperaturas. Jornal of the American Society for Horticultural Science. 107(1): 139-142.
RAMARKRISHNAN, N. KALINICH, J. and MCCLAIN, D. 1996. Ebselen inhibition
of apoptosis by reduction of peroxides. Biochem. Pharmacol. 51: 1443-1451.
RAZETO, B. 2000. La peteca del limón y su relación con el clima. Aconex Nº 66:
pp 23-24. ________. 1987. Desórdenes fisiológicos en cítricos. Aconex 17: 13-17. SCHNEIDER, H. 1968. The anatomy of citrus. In: Reuther, W.; Batchelor, L. Y
Webber, H. ed. The Citrus Industry. Riberside. University of California. pp 1-22.
SCHULTZE, J. 2000. Influencia de la ubicación geográfica del huerto y del fruto en
el árbol sobre el desarrollo de peteca. Tesis Ing. Agr. Santiago, Universidad de Chile. Escuela de Agronomía. 76 p.
SILVA, H. y RODRIGUEZ, J. 1995. Fertilización en plantas frutales. Santiago.
Pontificia Universidad Católica de Chile. 519p. SNOWDON, A. 1990. A color atlas of post-harvest Diseases and Disorders of fruti
and Vegetables General introducction and fruit. London, Wolfe Scientific. 302 p. (Volumen 2).

69
TARRAGÓ, J.; FALOCI, M.; LUNA, C.; SANSBERRO, P., MROGINSKI, L. 2003. Determinación de enzimas peroxidasas en tejidos caulinares de Ilex dumosa R. e Ilex paraguariensis St. Hil. (on line). http://www1. unne.edu.ar/cyt/2003/comunicaciones/05-Agrarias/A-042.pdf.
. UNDURRAGA, P. 1998. Manejo de cosecha y poscosecha de frutos cítricos.
Sociedad Gardiazabal y Magdahl Ltda. Seminario Internacional de Cítricos. Viña del Mar, 13-15 mayo 1998. pp: 93-117.
UNDURRAGA, R. 2004. Efecto de las aplicaciones de calcio en pre y poscosecha
sobre la aparición de peteca y calidad final. En almacenaje refrigerado a 3 ºC de limones cv. Eureka provenientes de árboles con y sin raleo. Taller de licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaíso. Facultad de Agronomía. 80p.
VAMOS, L.; VIGYAZO, L. 1981. Polyphenol oxidase and peroxidase in fruits and
vegetables. Crit Rev Food Sci Nutr. 15(1): 49-127. WILD, B.L. 1991. Postharvest factors governing the development of peteca rind
pitting on “Meyer” lemons. Hortscience 26(3):287-289.
WILLS, H; LEE, H; McGLASSON, B; HALL, G; GRAHAM, D. 1989. Fisiología y Manipulación de Frutas y Hortalizas Posrecolección. Zaragoza, Acribia, 195 p.
XU, H. BIDNEY, D. YALPANI, N. DUVICK, J. CRASTA, O. FOLKERTS, O. and
LU, G. 2003. Overexpression of a gene encoding hydrogen peroxide-genereating oxalate oxidase evokes defense responses in sunflower. Plant Physiol. 133:170-181.
YAKUSHIJI, H.; NOMANI, H; FUKUYAMA, T: ONO, S; TAKAGI, N and
HASHIMOTO, Y. 1996. Sugar accumulation enhanced by osmoregulation in satsuma mandarin fruit. J. Amer. Soc. Hort. Sci. 121(3): 466-472.
YANG, T. and POOVAIAH, B. 2002. Hydrogen peroxide homeostasis: Activation of
plant catalase by calcium/calmodulin. Plant Biology 99(6): 4097-4102.


















