4 membrana y citoplasma
17
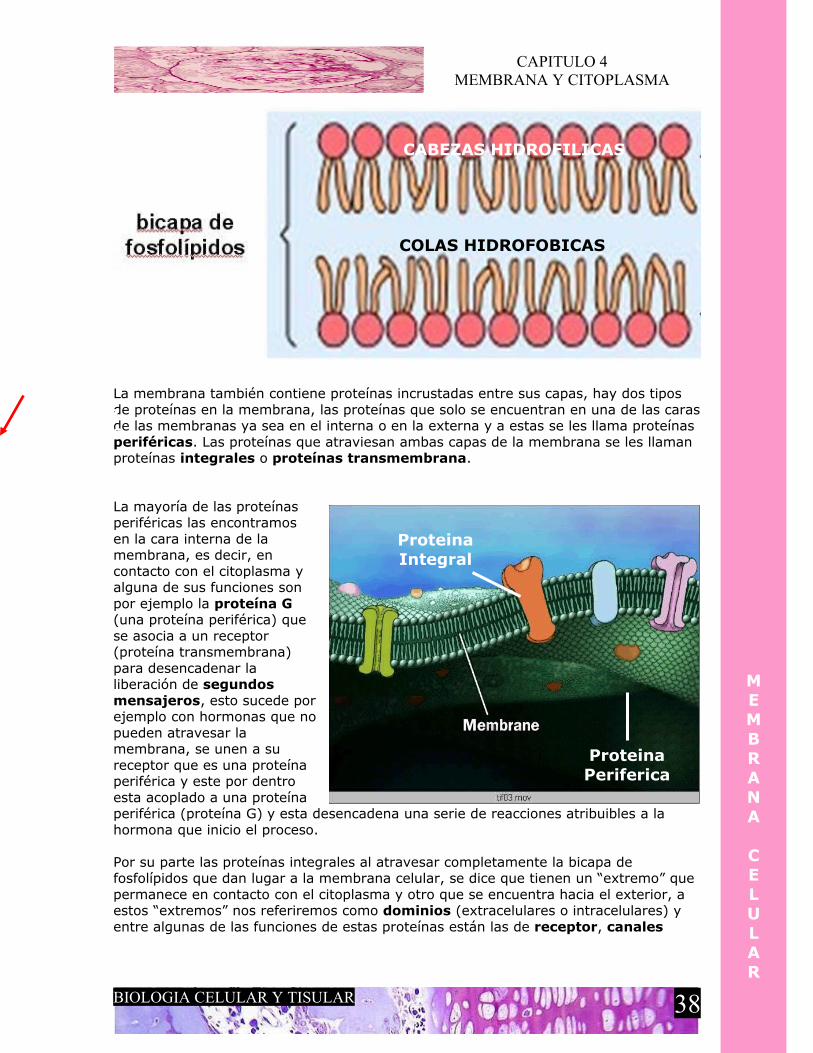
CAPITULO 4 MEMBRANA Y CITOPLASMA MEMBRANA CELULAR La membrana celular es una capa que delimita la célula, esta membrana adopta la forma que el citoesqueleto le proporcione a la célula, por lo tanto, encontramos células cubicas, cilíndricas, esferoidales, alargadas y estrelladas. Es una capa doble de fosfolípidos pero al microscopio se ve como una membrana trilaminar. La capa externa y la capa interna se ven electrodensas y la capa intermedia se ve electrolucida porque coincide con las colas de los fosfolipidos que constituyen ambas capas. La membrana esta formada por dos capas de fosfolípidos, un fosfolípidos es una molécula anfipática, es decir, que tiene propiedades tanto hidrofóbicas como hidrofílica, por eso, un fosfolípidos consta de dos partes, una cabeza y una cola. La cabeza de la molécula del fosfolípidos corresponde a la porción hidrofílica y la cola corresponde a la porción hidrofóbica, esto es comprensible ya que la cabeza de las moléculas fosfolipídicas de la capa externa dan hacia el liquido extracelular, que en su mayoría esta formado de agua, de la misma manera, las cabezas de las moléculas fosfolipídicas de la capa interna dan hacia el citoplasma, también un componente acuoso hacia el cual tienen afinidad los extremos hidrofílicos de estas moléculas, quedando de esta manera las colas hidrofóbicas protegidas y constituyendo la capa intermedia. Esto le permite a la membrana tener selectividad de acuerdo a lo que puede entrar o no al citoplasma, en este caso solo pueden atravesar la membrana celular algunos gases y moléculas pequeñas sin carga y con la propiedad de que sean hidrofóbicas, para que así puedan atravesar esta bicapa lipídica BIOLOGIA CELULAR Y TISULAR 37 Capa Externa Capa Interna Capa intermedia M E M B R A N A C E L U L A R
-
Upload
uatmatamoros -
Category
Education
-
view
797 -
download
3
Transcript of 4 membrana y citoplasma

CAPITULO 4MEMBRANA Y CITOPLASMA
MEMBRANA CELULAR
La membrana celular es una capa que delimita la célula, esta membrana adopta la forma que el citoesqueleto le proporcione a la célula, por lo tanto, encontramos células cubicas, cilíndricas, esferoidales, alargadas y estrelladas.
Es una capa doble de fosfolípidos pero al microscopio se ve como una membrana trilaminar.La capa externa y la capa interna se ven electrodensas y la capa intermedia se ve electrolucida porque coincide con las colas de los fosfolipidos que constituyen ambas capas.
La membrana esta formada por dos capas de fosfolípidos, un fosfolípidos es una molécula anfipática, es decir, que tiene propiedades tanto hidrofóbicas como hidrofílica, por eso, un fosfolípidos consta de dos partes, una cabeza y una cola.
La cabeza de la molécula del fosfolípidos corresponde a la porción hidrofílica y la cola corresponde a la porción hidrofóbica, esto es comprensible ya que la cabeza de las moléculas fosfolipídicas de la capa externa dan hacia el liquido extracelular, que en su mayoría esta formado de agua, de la misma manera, las cabezas de las moléculas fosfolipídicas de la capa interna dan hacia el citoplasma, también un componente acuoso hacia el cual tienen afinidad los extremos hidrofílicos de estas moléculas, quedando de esta manera las colas hidrofóbicas protegidas y constituyendo la capa intermedia.
Esto le permite a la membrana tener selectividad de acuerdo a lo que puede entrar o no al citoplasma, en este caso solo pueden atravesar la membrana celular algunos gases y moléculas pequeñas sin carga y con la propiedad de que sean hidrofóbicas, para que así puedan atravesar esta bicapa lipídica
BIOLOGIA CELULAR Y TISULAR 37
CapaExterna
CapaInterna
Capa intermedia
MEMBRANA
CELULAR
CAPITULO 4MEMBRANA Y CITOPLASMA
CABEZAS HIDROFILICAS
COLAS HIDROFOBICAS
ProteinaIntegral
ProteinaPeriferica
La membrana también contiene proteínas incrustadas entre sus capas, hay dos tipos de proteínas en la membrana, las proteínas que solo se encuentran en una de las caras de las membranas ya sea en el interna o en la externa y a estas se les llama proteínas periféricas. Las proteínas que atraviesan ambas capas de la membrana se les llaman proteínas integrales o proteínas transmembrana.
La mayoría de las proteínas periféricas las encontramos en la cara interna de la membrana, es decir, en contacto con el citoplasma y alguna de sus funciones son por ejemplo la proteína G (una proteína periférica) que se asocia a un receptor (proteína transmembrana) para desencadenar la liberación de segundos mensajeros, esto sucede por ejemplo con hormonas que no pueden atravesar la membrana, se unen a su receptor que es una proteína periférica y este por dentro esta acoplado a una proteína periférica (proteína G) y esta desencadena una serie de reacciones atribuibles a la hormona que inicio el proceso.
Por su parte las proteínas integrales al atravesar completamente la bicapa de fosfolípidos que dan lugar a la membrana celular, se dice que tienen un “extremo” que permanece en contacto con el citoplasma y otro que se encuentra hacia el exterior, a estos “extremos” nos referiremos como dominios (extracelulares o intracelulares) y entre algunas de las funciones de estas proteínas están las de receptor, canales
BIOLOGIA CELULAR Y TISULAR 38
ProteinaPeriferica
MEMBRANA
CELULAR
CAPITULO 4MEMBRANA Y CITOPLASMA
iónicos o complejos de unión, por ejemplo en las uniones intercelulares de los epitelios que se estudian mas adelante.
Por ultimo, la membrana celular también esta compuesta, en menor medida por carbohidratos, estos carbohidratos se pegan a las proteínas periféricas externas de la membrana y constituyen lo que se le llama glucocalix, una sustancia en estado de gel que le sirve a la célula como medio de adherencia y para atraer agua a ella.
POTENCIAL DE MEMBRANA
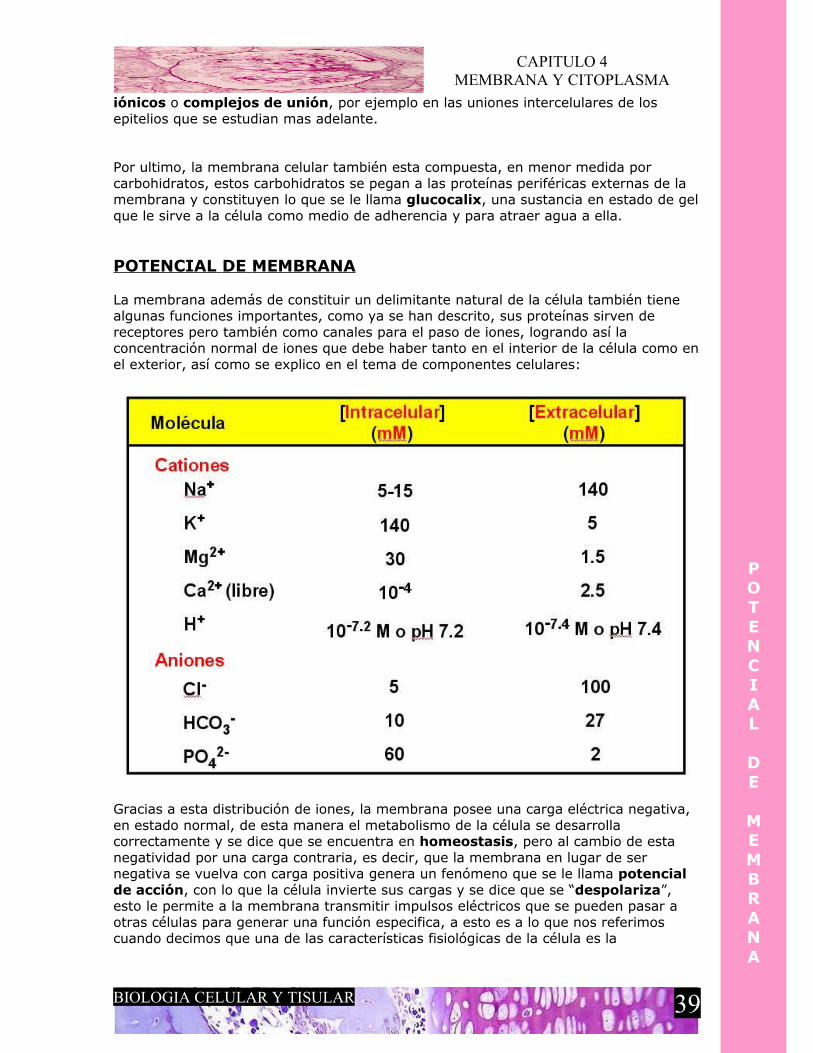
La membrana además de constituir un delimitante natural de la célula también tiene algunas funciones importantes, como ya se han descrito, sus proteínas sirven de receptores pero también como canales para el paso de iones, logrando así la concentración normal de iones que debe haber tanto en el interior de la célula como en el exterior, así como se explico en el tema de componentes celulares:
Gracias a esta distribución de iones, la membrana posee una carga eléctrica negativa, en estado normal, de esta manera el metabolismo de la célula se desarrolla correctamente y se dice que se encuentra en homeostasis, pero al cambio de esta negatividad por una carga contraria, es decir, que la membrana en lugar de ser negativa se vuelva con carga positiva genera un fenómeno que se le llama potencial de acción, con lo que la célula invierte sus cargas y se dice que se “despolariza”, esto le permite a la membrana transmitir impulsos eléctricos que se pueden pasar a otras células para generar una función especifica, a esto es a lo que nos referimos cuando decimos que una de las características fisiológicas de la célula es la
BIOLOGIA CELULAR Y TISULAR 39
POTENCIAL
DE
MEMBRANA

CAPITULO 4MEMBRANA Y CITOPLASMA
excitabilidad, aunque esta propiedad se limite solo a células musculares y neuronas.
Cuando se cambia esta carga negativa y la membrana se vuelve positiva se dice que sufrió una despolarización y esto es lo que causa que se transmita el impulso. La despolarización se da por la entrada excesiva de los iones que normalmente abundan en el espacio extracelular, en este caso es el sodio, cuando por medio de un impulso nervioso o mecánico comienza a entrar sodio a la célula, la membrana de esta se despolariza y cambia su carga que era originalmente negativa a positiva, esta despolarización que inicia en un punto de la membrana se disemina por toda esta hasta alcanzar otra y otra célula, la despolarización se lleva a cabo por ejemplo, en la neurona para transmitir el impulso nervioso y en el músculo para iniciar la contracción.
Una vez que se cumplió con el objetivo, la membrana debe regresar a su estado original para estar lista para un nuevo potencial de acción, es decir, debe repolarizarse, la repolarizacion se consigue gracias a que la célula permite la salida excesiva de potasio, para tratar de disminuir la carga positiva que se dio el entrar el sodio, una vez que la membrana vuelve a su estado original con una carga negativa, por medio de una proteína llamada “bomba de sodio/potasio” las concentraciones de estos dos iones se estabilizan, mas sodio en el liquido extracelular y mas potasio en el citoplasma, de esta manera la membrana vuelve a estar cargada negativamente y lista para nuevamente sufrir una despolarización.
El termino hiperpolarizacion se menciona como una consecuencia que resulta al haber una salida excesiva de potasio en la etapa de repolarizacion, cuando sale demasiado potasio de la célula esta se vuelve mas negativa de lo que originalmente debería de ser, esto quiere decir que esta célula, que ahora esta hiperpolarizada, tardara mas tiempo en volver a despolarizarse otra vez, que una célula que regresa a su estado normal en la etapa de repolarizacion, por lo tanto, una célula al hiperpolarizarse quedara inhibida por algún tiempo.
El potencial de acción lo podemos resumir en los siguientes pasos:
La membrana de una célula tiene una carga negativa, en su interior hay grandes concentraciones de potasio y en el líquido extracelular hay grandes concentraciones de sodio.
Cuando se abren los canales de sodio (ya sea por impulso nervioso, mecánico o eléctrico) el sodio entra en grandes cantidades a la célula
Al mismo tiempo que se abrieron los canales de sodio, los canales de potasio se cierran para que este no pueda salir de la célula
Una vez que hay una concentración considerable de sodio en el interior de la membrana, esta se vuelve positiva
Al cambio de carga de negativa a positiva en la membrana se le llama despolarización y ocurre en toda la membrana
Una vez que la despolarización esta ocurriendo se generan cambios que pueden llevar a cabo una función especifica, por ejemplo, en una neurona debe despolarizarse su membrana para que se libere el neurotransmisor que actuara en el tejido a “inervar”, o bien, en el músculo esquelético es necesaria una despolarización para que se libere calcio en el citoplasma de la célula muscular para que esta se pueda contraer.
BIOLOGIA CELULAR Y TISULAR 40
POTENCIAL
DE
MEMBRANA

CAPITULO 4MEMBRANA Y CITOPLASMA
Mientras se esta llevando a cabo una despolarización no puede haber otra despolarización sino hasta que esta célula se repolarize, al tiempo que tarda en hacerlo se le llama periodo refractario.
En el estado de repolarizacion, se abren los canales de potasio y este sale fácilmente de la célula para que esta pierda su carga positiva
Con este estado de repolarizacion quedo un numero considerable de iones sodio en el interior de la célula y una parte del potasio que abundaba en el interior de la célula se escapo hacia el liquido intracelular, por lo tanto es necesario estabilizar estos dos iones como en su estado original, es decir, mas potasio dentro de la célula y mas sodio fuera de ella, para que esta estabilización suceda entra en acción una proteína llamada bomba de sodio/potasio que saca el sodio de la célula hacia el liquido extracelular al mismo tiempo que mete potasio del liquido extracelular al interior de la célula.
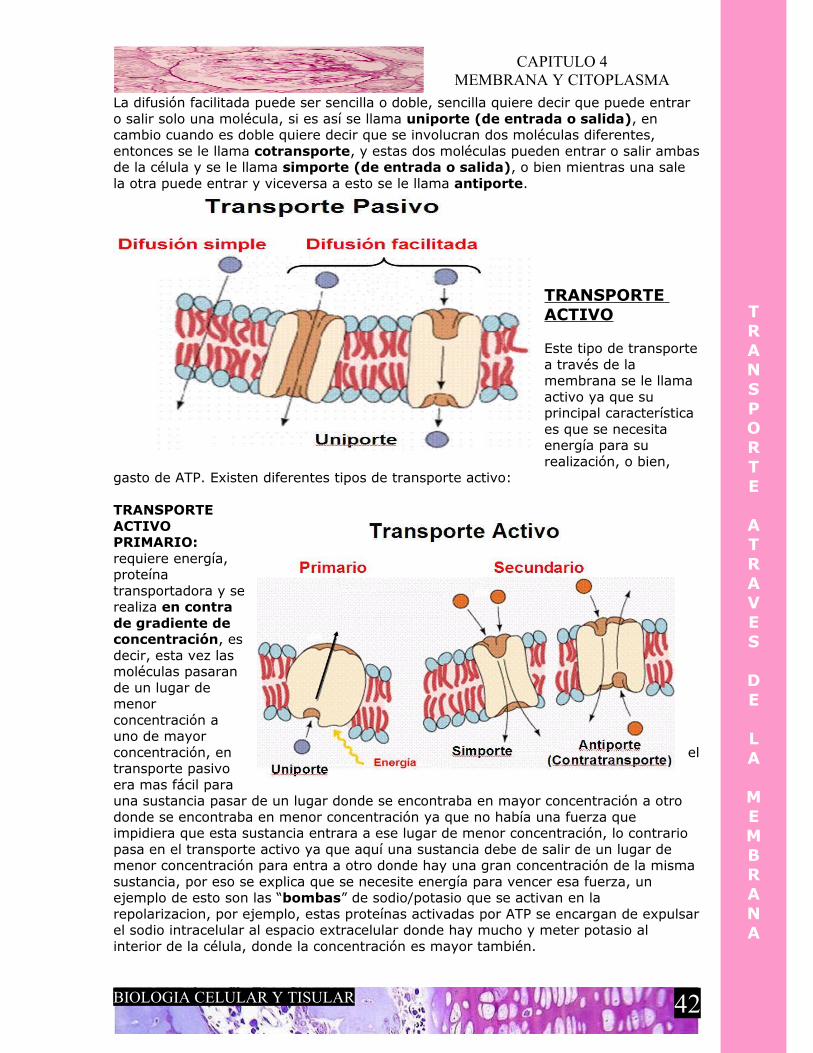
En esto se basa el potencial de membrana, esta regulado por la entrada y salida de iones, pero si ya vimos que la membrana solo deja pasar gases y moléculas pequeñas sin carga como es que se dan estos fenómenos de despolarización, repolarizacion e hiperpolarizacion, pues bien, los iones implicados en estos procesos entran y salen de la célula por medio de un mecanismo llamado transporte através de la membrana y este se divide en transporte pasivo y transporte activo.
TRANSPORTE PASIVO
Este se refiere al tipo de transporte que puede presentar la membrana celular (plasmalema) en cual no es necesario algún tipo de energía (ATP), es decir, las moléculas que deben entrar o salir simple y sencillamente lo hacen, existen varios tipos de transporte pasivo:
DIFUSION SIMPLE: Este transporte se da a favor de gradiente de concentración, no necesita energía (ATP) ni requiere proteínas transportadoras, por ejemplo, en los pulmones, el O2 solo atraviesa las paredes del alveolo para entra al capilar, y el CO2 sale del capilar para penetrar al alveolo y así ser expulsado en la expiración, todo esto por una diferencia de concentraciones (a favor de gradiente de concentración), la sustancia que se va a transportar se moverá de un lugar donde esta en mayor concentración a otro lugar donde hay menor concentración de la misma , no se necesita energía ni ayuda de alguna proteína integral que funcione como canal o como proteína transportadora.
DIFUSION FACILITADA: Este transporte también es a favor de gradiente de concentración, no requiere energía y si requiere de una proteína transportadora, al igual que la difusión simple, la sustancia que se vaya a transportar, se moverá de un lugar de donde se encuentre en mayor concertación a un lugar donde haya menor concentración de la misma. La única diferencia es que ahora se requiere de una proteína transportadora que no es mas que una proteína integral que sirve de canal para iones, pero estos canales normalmente están cerrados y solo se abrirán por dos mecanismos, o por cambio de voltaje (despolarización) o bien por medio de un ligando, por ejemplo, en una despolarización se abren los canales de sodio para que este pueda entrar, y un ejemplo de ligando seria la insulina que es necesario que se pegue a un receptor para que se abra un canal por donde puede entrar la glucosa a las células para que sea utilizada como energía.
BIOLOGIA CELULAR Y TISULAR 41
TRANSPORTE
ATRAVES
DE
LA
MEMBRANA
CAPITULO 4MEMBRANA Y CITOPLASMA
La difusión facilitada puede ser sencilla o doble, sencilla quiere decir que puede entrar o salir solo una molécula, si es así se llama uniporte (de entrada o salida), en cambio cuando es doble quiere decir que se involucran dos moléculas diferentes, entonces se le llama cotransporte, y estas dos moléculas pueden entrar o salir ambas de la célula y se le llama simporte (de entrada o salida), o bien mientras una sale la otra puede entrar y viceversa a esto se le llama antiporte.
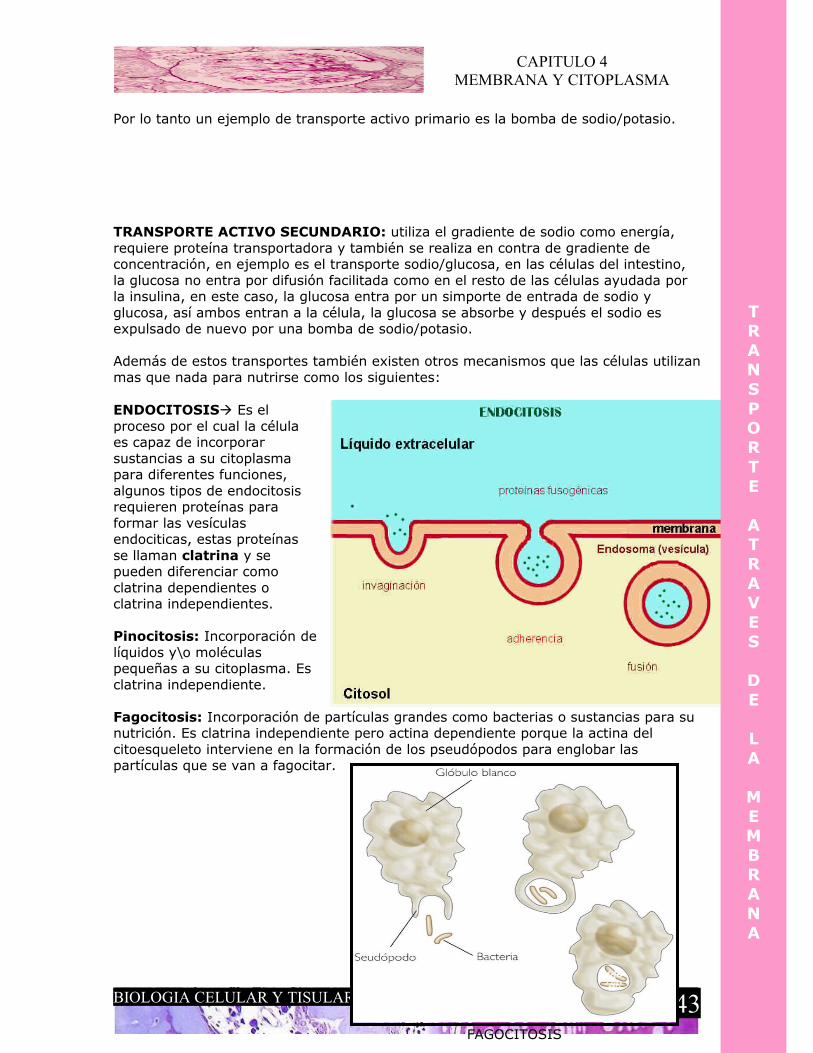
TRANSPORTE ACTIVO
Este tipo de transporte a través de la membrana se le llama activo ya que su principal característica es que se necesita energía para su realización, o bien,
gasto de ATP. Existen diferentes tipos de transporte activo:
TRANSPORTE ACTIVO PRIMARIO: requiere energía, proteína transportadora y se realiza en contra de gradiente de concentración, es decir, esta vez las moléculas pasaran de un lugar de menor concentración a uno de mayor concentración, en el transporte pasivo era mas fácil para una sustancia pasar de un lugar donde se encontraba en mayor concentración a otro donde se encontraba en menor concentración ya que no había una fuerza que impidiera que esta sustancia entrara a ese lugar de menor concentración, lo contrario pasa en el transporte activo ya que aquí una sustancia debe de salir de un lugar de menor concentración para entra a otro donde hay una gran concentración de la misma sustancia, por eso se explica que se necesite energía para vencer esa fuerza, un ejemplo de esto son las “bombas” de sodio/potasio que se activan en la repolarizacion, por ejemplo, estas proteínas activadas por ATP se encargan de expulsar el sodio intracelular al espacio extracelular donde hay mucho y meter potasio al interior de la célula, donde la concentración es mayor también.
BIOLOGIA CELULAR Y TISULAR 42
TRANSPORTE
ATRAVES
DE
LA
MEMBRANA
CAPITULO 4MEMBRANA Y CITOPLASMA
Por lo tanto un ejemplo de transporte activo primario es la bomba de sodio/potasio.
TRANSPORTE ACTIVO SECUNDARIO: utiliza el gradiente de sodio como energía, requiere proteína transportadora y también se realiza en contra de gradiente de concentración, en ejemplo es el transporte sodio/glucosa, en las células del intestino, la glucosa no entra por difusión facilitada como en el resto de las células ayudada por la insulina, en este caso, la glucosa entra por un simporte de entrada de sodio y glucosa, así ambos entran a la célula, la glucosa se absorbe y después el sodio es expulsado de nuevo por una bomba de sodio/potasio.
Además de estos transportes también existen otros mecanismos que las células utilizan mas que nada para nutrirse como los siguientes:
ENDOCITOSIS Es el proceso por el cual la célula es capaz de incorporar sustancias a su citoplasma para diferentes funciones, algunos tipos de endocitosis requieren proteínas para formar las vesículas endociticas, estas proteínas se llaman clatrina y se pueden diferenciar como clatrina dependientes o clatrina independientes.
Pinocitosis: Incorporación de líquidos y\o moléculas pequeñas a su citoplasma. Es clatrina independiente.
Fagocitosis: Incorporación de partículas grandes como bacterias o sustancias para su nutrición. Es clatrina independiente pero actina dependiente porque la actina del citoesqueleto interviene en la formación de los pseudópodos para englobar las partículas que se van a fagocitar.
BIOLOGIA CELULAR Y TISULAR 43FAGOCITOSIS
TRANSPORTE
ATRAVES
DE
LA
MEMBRANA

CAPITULO 4MEMBRANA Y CITOPLASMA
Endocitosis mediada por receptores: Permite la entrada de moléculas especificas a la célula, cuando estas moléculas se pegan a su receptor, (proteína transmembrana) se activan las clatrinas que forman las vesículas desde el interior de la célula y así es posible que entren las moléculas al citoplasma, por lo tanto es clatrina dependiente.
EXOCITOSIS Es el proceso mediante el cual una vesícula de desecho se mueve desde el citoplasma hacia la membrana citoplasmática desde donde vierte su contenido en el espacio extracelular.
CITOPLASMA
Es la parte acuosa de la célula, en el se encuentra flotando el núcleo y los organelos que llevan a cabo las funciones de la célula. La porción del citoplasma periférica que da hacia el plasmalema se le llama ectoplasma el cual es más claro, hialino y no tiene organelas. La porción del citoplasma que da hacia la membrana nuclear se le llama endoplasma esta porción es mas granular por la presencia de los organelos, sin embargo en las células humanas estas dos porciones del citoplasma no se delimitan con gran nitidez.
Los organelos se dividen en dos grandes grupos, los organelos que están delimitados por membrana (organelos membranosos) y los que no están delimitados por membrana (organelos no membranosos).
BIOLOGIA CELULAR Y TISULAR 44
ENDOCITOSIS MEDIADA POR RECEPTORES
CITOPLASMA
CELULAR

CAPITULO 4MEMBRANA Y CITOPLASMA
ORGANELOS MEMBRANOSOS: Mitocondrias, RER, REL, Ap. Golgi, Lisosomas, Peroxisomas y laminillas anulares.
ORGANELOS NO MEMBRANOSOS: Ribosomas, Proteasomas, Filamentos y Microtubulos,
MITOCONDRIAS
Organelas membranosa, alargada, con una doble membrana interna y externa. También se le conoce como condriosomas.
Las mitocondrias tienen su propio DNA e incluso puede sintetizar algunas de sus propias proteínas funcionales y además se pueden dividir por fisión binaria, por eso mismo se cree que las mitocondrias provienen de una bacteria capaz de sintetizar energía en forma de ATP, esta bacteria para obtener protección empezó a vivir dentro de una célula, así esta le brindaba protección y la bacteria le brindaba energía en forma de ATP. Después con el tiempo esta bacteria siguió viviendo dentro de las células originando así a lo que hoy conocemos como la mitocondria.
Las mitocondrias son muy abundantes en células de tejidos muy activos.
Esta formada por una doble membrana (interna y externa).
La membrana interna mitocondrial está formada por fosfolipidos, se pliega sobre sí misma para formar crestas. En estas cresta se encuentran las partículas elementales o partículas F1. En estas partículas se lleva a cabo la fosforilación oxidativa donde se liberan moléculas de ATP como energía.
La matriz mitocondrial, se encuentra dentro de la membrana interna, contiene agua, iones, ácidos grasos, enzimas, ribosomas y su propio ADN, en la matriz se lleva a cabo la beta-oxidación de los ácidos grasos que libera energía en forma de calor.
La membrana externa solo es de sostén, formadas por proteínas y es rígida.
BIOLOGIA CELULAR Y TISULAR 45
ORGANELOS
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
RETICULO ENDOPLASMICO RUGOSO
Es una agrupación de cisternas de membrana, estas cisternas tiene ribosomas en su superficie, la función de este organelo está ligada con la síntesis,
transporte y exportación de proteínas.
RETICULO ENDOPLASMICO LISO
Es una agrupación de cisternas de membrana, estas cisternas no tienen ribosomas en su superficie, la función de este
organelo está ligada con la síntesis, metabolismo, y transporte de lípidos y esteroides. Además se encarga de la desintoxicación y almacena calcio.
APARATO DE GOLGI
Organelo membranoso que recibe, almacena y secreta sustancias para la célula. También excretan desechos al espacio extracelular. Es una pila de cisternas con una cara interna que da hacia al nucleolema que es la cara de formación (cara cis) y una cara externa que da hacia el plasmalema que es la cara de maduración (cara trans).
VESICULAS DE TRANSPORTE
BIOLOGIA CELULAR Y TISULAR 46
ORGANELOS
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
El retículo endoplasmico rugoso o liso, sintetiza sustancias y las manda hacia el aparato de Golgi para que este las almacene hasta que sean necesarias. O bien el aparato de Golgi regresa sustancias que vienen del exterior y las envía hacia el retículo rugoso o liso según sea necesario.
En ambos tipos de movimiento intervienen diferentes partículas denominadas COP, estas intervienen dependiendo de si el movimiento es hacia el aparato de golgi (anterogrado), si es así, entonces son las partículas cop II las que actuaran, de lo contrario, si el movimiento es desde el aparato de Golgi (retrogrado) entonces serán las partículas cop I las que estarán involucradas.
LISOSOMAS
Organela membranosa redonda que contiene enzimas hidroliticas para llevar a cabo la digestión de las células, los lisosomas se fusionan con las vesículas endociticas mezclándose así la sustancia fagocitada con las enzimas del lisosoma. Los lisosomas se pueden clasificar en:
Lisosomas primarios: Los que aun no llevan a cabo un proceso de digestión.
BIOLOGIA CELULAR Y TISULAR 47
RER
REL
AP.
GOLGI
Movimiento anterogrado. Particulas COP II
Movimiento retrogrado. Particulas COP I
ORGANELOS
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
Lisosomas secundario: Cuando el lisosoma se une a un fagosoma (vesícula de endocitocis).Lisosoma terciario: Cuando el material endocitado no pudo ser desnaturalizado por las enzimas del lisosoma este se convierte en un cuerpo residual y su fin puede ser por dos vías, o se excreta por exocitosis o permanece en el interior de la célula en forma de lipofucsina (pigmento residual del metabolismo de lípidos).
PEROXISOMA
Organela membranosa que contiene enzimas oxidativas como la catalasa.
Su principal función es llevar a cabo la beta-oxidación de los ácidos grasos y liberan peróxido de hidrogeno (H2O2).
El peróxido de hidrogeno es degradado en agua y oxigeno.
Encontramos Peroxisomas en células con actividad notable como en el hígado y riñón.
LAMINILLAS ANULARES
Pila de cisternas parecidas a la membrana nuclear, presentan pequeños orificios o anillos con características de los poros nucleares, se ven en células de crecimiento rápido como en las células embrionarias o tumorales.
La función de las laminillas anulares es desconocida.
ORGANELOS NO MEMBRANOSOS
RIBOSOMAS
Organela no membranosa que se encarga de “leer” las cadenas de ARN mensajero para así sintetizar proteínas a partir de aminoácidos. Los ribosomas los encontramos de las siguientes maneras:
Ribosomas libres: Flotan en el citoplasma, sintetizan proteínas para el uso de la célula.Polirribosomas (polisomas): Conjunto de ribosomas que leen una misma cadena de RNA mensajero.Ribosomas sobre le RER: Sintetizan proteínas directamente en el interior del RER para que se puedan exportar.
BIOLOGIA CELULAR Y TISULAR 48
ORGANELOS
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
PROTEASOMAS
Organelas no membranosas, cilíndricas, multiproteicas formadas por subunidades con un canal central, complejas. Contienen abundantes proteasas (enzimas que degradan proteínas).
Marcan las proteínas defectuosas o con vida media cumplida con una molécula llamada ubiquitina. El proteosoma capta las proteínas marcadas con ubiquitina y las degrada con sus proteasas.
CITOESQUELETO
El citoesqueleto esta formados por diversas estructuras proteicas filamentosas, proteínas asociadas y otros filamentos que le dan soporte y forma a las células, entre otras funciones, por ejemplo en el eritrocito el citoesqueleto es indispensable para su “flexibilidad”. Todas las estructuras del citoesqueleto se consideran organelas no membranosas.
FILAMENTOS: proteínas filamentosas, en forma de fibras. Los filamentos se clasifican en tres tipos: filamentos finos (filamentos de actina, microfilamentos), filamentos intermedios (tonofilamentos) y filamentos gruesos (filamentos de miosina).
MICROFILAMENTOS Son delgados, miden entre 5 y 7 nm de grueso. La actina es una proteína globular, por eso se le llama actina G y esta es la forma libre de la actina en el citoplasma, sin embargo, la actina se puede polimerizar y formar el filamento de actina o actina F (actina filamentosa). Los filamentos de actina tienen entre otras funciones:
• Anclaje y movimiento de proteínas de la membrana.
• Formación del “esqueleto” de las microvellosidades.
• Locomoción celular y emisión de pseudopodos.
La actina tiene algunas proteínas asociadas a ella que le ayudan a cumplir sus funciones como la:
BIOLOGIA CELULAR Y TISULAR 49
ORGANELOS
NO
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
• Fascina y fimbrina: establecen enlaces entre los microfilamentos para brindar una mejor organización, como en las microvellosidades, establecen fascículos. (véase tejido epitelial)
• Gelsolina: cortan los filamentos de actina• Tropomodulina: forman casquetes, regulan la longitud del microfilamento
• Proteínas formadoras de enlace: al igual que la fascina y fimbrina forman enlaces, pero estas no agrupan los microfilamentos en fascículos, por ejemplo la espectrina, en el eritrocito.
FILAMENTOS INTERMEDIOS
También llamados tonofilamentos, tienen función de sostén o como estructura general, los podemos encontrar en todas las células, son más fuertes que los microfilamentos. Se les llama “intermedios” por su tamaño, miden entre 8 y 10 nm y están formados por diferentes proteínas fibrilares que dependen de cada célula:
• Queratina: células epiteliales• Vimentina: células del tejido conjuntivo• Desmina: células musculares• GFAP (gliofibrilar acid protein): células de la neuroglia• Lamina nuclear: en los núcleos celulares
FILAMENTOS GRUESOS
Son los filamentos de miosina, miden más de 10 nm. El filamento de miosina esta formado por dos cadenas pesadas que constituyen la “cola” del filamento de miosina y cuatro cadenas ligeras, mismas que constituyen las cabezas (2) del filamento de miosina, este tipo de filamentos tiene gran importancia en las células musculares ya que en ellas la miosina se asocia con la actina para así brindar el fenómeno de la contracción que se estudiara mas adelante. (Véase tejido muscular).
MICROTUBULOS
Además de los filamentos, los microtúbulos también forman parte del citoesqueleto.
Son estructuras alargadas huecas, no se ramifican, rígidas, compuestas por moléculas de alfa y beta tubulina.
Inician en una proteína de casquete y un anillo de gamma tubulina, a partir de este se comienzan a polimerizar los microtubulos añadiéndose moléculas de alfa y beta tubulina. La polimerización se da hacia el extremo plus (+), es decir crecen. Si por el contrario,
BIOLOGIA CELULAR Y TISULAR 50
ORGANELOS
NO
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
las moléculas de tubulina se disocian se dice que se despolimeriza hacia el extremo minus (-) es decir hacia el anillo de gamma tubulina y se reduce la longitud del microtúbulo.
Su función principal es la de transporte, existen unas proteína que permiten que ciertas estructuras se puedan mover sobre los microtubulos, la cinesina que avanza hacia el extremo plus (+), hacia la periferia de la célula y la dineina que avanza hacia el extremo minus (-), hacia el centro de la célula.
Además del transporte, el microtúbulos es la unidad estructural de: centriolos, cilios, flagelos y cuerpos basales además el huso mitótico que funciona durante la división celular esta formado por microtúbulos.
ORGANELOS FORMADOS POR MICROTUBULOS
Centriolo s:
Formados por 9 tripletes de microtubulos organizados en forma helicoidal.
Intervienen en la organización del huso mitótico durante la división celular.
Cilios y flagelos:
Inician con un cuerpo basal que es de igual estructura a un centriolo y después el cuerpo del cilio (axonema) esta formado por 9 dobletes de microtubulos y un par central. Cada doblete esta unido a otro por medio de “brazos” de dineina lo que les permite el movimiento, los cilios brindan la capacidad de arrastrar sustancias como por ejemplo en el epitelio respiratorio para expulsar el moco. El flagelo tiene exactamente la misma estructura, lo único que varía es la longitud. (Véase tejido epitelial)
INCLUSIONES CITOPLASMATICAS
BIOLOGIA CELULAR Y TISULAR 51
ORGANELOS
NO
MEMBRANOSOS

CAPITULO 4MEMBRANA Y CITOPLASMA
Son depósitos de material que contiene productos del metabolismo celular, se les menciona como gránulos, gotitas o como pigmentos. Pueden estar rodeadas de membrana o no, por ejemplo la lipofucsina el pigmento en que se transforma el lisosoma terciario, tendrá como delimitante membrana ya que el lisosoma es una organela membranosa.
• Lipofuscina: pigmento que se observa en las células que no se dividen como las neuronas y musculares esqueléticas y cardiacas. Contienen restos no digeridos del metabolismo celular. Lisosomas terciarios.
• Hemosiderina: complejo de hierro que queda de la incompleta digestión de la hemoglobina. Estos gránulos son muy notables en células del bazo, lugar de degradación de los eritrocitos.
• Glucógeno: no se tiñe con H-E, el glucógeno es almacenado como fuente de energía y se ven bien en hepatocitos y músculo estriado.
• Inclusiones lipídicas (gotitas de lípido): restos del metabolismo que se almacenan como fuente alternativa de energía, el depósito más grande de lípidos lo tiene los adipocitos, pero igual pueden estar brevemente en otras células como los enterocitos (intestinales).
DIFERENCIACION CELULAR
La diferenciación de una célula es la capacidad que tiene la misma de poder dar origen a una o más líneas de células a partir de ella, es decir, cuando una sola célula puede dar origen a varias líneas celulares se dice que es mas indiferenciada. Cuando una célula esta mas diferenciada es porque poco a poco va perdiendo la capacidad de dar origen a mas células diferentes a ella.
Esto ocurre normalmente en el ser humano al ser un cigoto. El cigoto es una célula totalmente indiferenciada, porque a partir de ella se originan numerosas líneas celulares diferentes, tantas como para formar a un ser humano.
Ahora, sabiendo que un tejido es un conjunto de células que se agrupan para llevar a cabo un función específica podemos comenzar por explicar que la primera célula de un ser humano es el cigoto aquella célula que se forma al fusionarse el ovulo con el espermatozoide, el cigoto es una célula totipotencial, es decir que da origen a todo un ser completo, después de la formación del cigoto, este se divide inmediatamente dando lugar a 2 células idénticas llamadas blastomeras y estas 2 blastomeras dan origen cada una a otras 2 blastomeras para formar así un total de 4 blastomeras, hasta entonces estas 4 blastomeras siguen conservando su poder totipotencial, es decir, si se llegara a aislar una de estas 4 blastomeras, estas podrían dar lugar a un nuevo ser completo cada una.
Al cabo de 8 a 9 días, el embrión se convierte en un embrión trilaminar, formado como su nombre lo dice por 3 capas: ectodermo, mesodermo y endodermo y cada una de estas capas dan lugar a ciertos tejidos diferentes.
A partir de las capas germinativas (ecto, meso y endodermo) las células se van diferenciando y pasan de ser totipotenciales a pluripotenciales como la célula madre sanguínea que da origen a todas las células sanguíneas, así se va diferenciando cada
BIOLOGIA CELULAR Y TISULAR 52
DIFERENCIACION
CELULAR

CAPITULO 4MEMBRANA Y CITOPLASMA
vez mas, pasa a ser multipotencial, bipotencial y unipotencial cuando al dividirse solo de origen a células iguales a ella.
DESARROLLO DE LOS TEJIDOS BASICOS Y ORIGEN EMBRIONARIO.
Derivados de las hojas germinativas:
ECTODERMO Epidermis, pelo, unas, oído interno, cristalino, sistema nervioso, melanocitos, células de Schwann.
MESODERMO Algunos músculos, aparato urogenital, gónadas, tejido conjuntivo, sangre, tejido óseo, cartílago.
ENDODERMO Epitelio de traquea, bronquios, pulmones, tubo digestivo, higado, páncreas, uraco, faringe, glándula tiroides, cavidad timpánica, trompa de Eustaquio, amígdalas.
VIDEOS EN INTERNET
Lisosomashttp://highered.mcgraw-hill.com/olcweb/cgi/pluginpop.cgi?it=swf::535::535::/sites/dl/free/0072437316/120067/bio01.swf::Lysosomes
Cilios y flageloshttp://programs.northlandcollege.edu/biology/Biology1111/animations/flagellum.html
BIOLOGIA CELULAR Y TISULAR 53
DERIVADOS
GERMINATIVOS